
An Architecture to Stimulate Behavioral Development of Academic Cloud Users
Upload
independentCategory
view
3download
0
Matrix Biology 29 (2010) 690–700
Contents lists available at ScienceDirect
Matrix Biology
j ourna l homepage: www.e lsev ie r.com/ locate /matb io
Extracellular matrix-derived products modulate endothelial and progenitor cellmigration and proliferation in vitro and stimulate regenerative healing in vivo
Ekaterina Vorotnikova a, Donna McIntosh a, Abiche Dewilde a, Jianping Zhang a, Janet E. Reing b, Li Zhang b,Kevin Cordero b, Khamilia Bedelbaeva c, Dimitri Gourevitch c, Ellen Heber-Katz c,Stephen F. Badylak b, Susan J. Braunhut a,⁎a Department of Biological Sciences, University of Massachusetts, Lowell, MA 01854, USAb Department of Surgery, McGowan Institute for Regenerative Medicine, University of Pittsburgh, 100 Technology Dr., Suite 200, Pittsburgh, PA 15219, USAc The Wistar Institute, 3601 Spruce St., Philadelphia, PA 19104, USA
⁎ Corresponding author. Olsen Building, 415C, 1 UnivUSA. Tel.: +1 978 934 2876; fax: +1 978 934 3044.
E-mail address: [email protected] (S.J. Brau
0945-053X/$ – see front matter © 2010 Elsevier B.V. Adoi:10.1016/j.matbio.2010.08.007
a b s t r a c t
a r t i c l e i n f oArticle history:Received 21 December 2009Received in revised form 5 August 2010Accepted 17 August 2010
Keywords:Extracellular matrixBlastemaWound healingTissue regeneration
Most adult mammals heal without restorative replacement of lost tissue and instead form scar tissue at aninjury site. One exception is the adult MRL/MpJ mouse that can regenerate ear and cardiac tissue afterwounding with little evidence of scar tissue formation. Following production of a MRL mouse ear hole, 2 mmin diameter, a structure rapidly forms at the injury site that resembles the amphibian blastema at a limbamputation site during limb regeneration. We have isolated MRL blastemal cells (MRL-B) from this structureand adapted them to culture. We demonstrate by RT-PCR that even after continuous culturing of these cellsthey maintain expression of several progenitor cell markers, including DLK (Pref-1), and Msx-1. We haveisolated the underlying extracellular matrix (ECM) produced by these MRL-B cells using a new non-proteolytic method and studied the biological activities of this cell-free ECM. Multiplex microELISA analysisof MRL-B cell-free ECM vs. cells revealed selective enrichment of growth factors such as bFGF, HGF and KGFin the matrix compartment. The cell-free ECM, degraded by mild enzyme treatment, was active in promotingmigration and proliferation of progenitor cells in vitro and accelerating wound closure in a mouse fullthickness cutaneous wound assay in vivo. In vivo, a single application of MRL-B cell matrix-derived productsto full thickness cutaneous wounds in non-regenerative mice, B6, induced re-growth of pigmented hair,dermis and epidermis at the wound site whereas scar tissue replaced these tissues at wound sites in micetreated with vehicle alone. These studies suggest that matrix-derived products can stimulate regenerativehealing and avert scar tissue formation in adult mammals.
ersity Ave., Lowell, MA 01854,
nhut).
ll rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
The MRL/MpJ wild type strain of mice (MRL) was originallygenerated by interbreeding C57BL/6 J (0.3%), C3H/HeDi (12.1%), AKR/J(12.6%) and LG/J (75%) strains (Theofilopoulos & Dixon, 1985; Clarket al., 1998). MRL mice at particular sites of the body exhibit increasedwound healing capacity as compared to other inbred strains (Heber-Katz, 1999; Samulewicz et al., 2002). For example, ~4 weeks afterreceiving a 2 mm ear hole injury, MRL mice showed ~80% reduction inear hole size whereas C57BL/6 mice only exhibited a 36% closure(Samulewicz et al., 2002). Histological examination revealed that theprocess of healing in MRLmice was faster andmore complete than thatof the C57BL/6 mice and exhibited parallels to amphibian regeneration,including the formationof a blastemal structure, a dynamic extracellularmatrix, re-appearance of cartilage and hair follicles, healing without
scarring, and up-regulation of tenascin, Pref-1 and down-regulation ofMSX-2 in cells present within the blastema (Clark et al., 1998;Gourevitch et al., 2003). MRL mice also have the ability to heal burn-induced infarct of the ventricle with restoration of normal structuresand function of the heart and reduced cardiac scarring, a finding thatdoes not occur in other adult mice such as C57BL/6 mice (Leferovich etal., 2001; Leferovich & Heber-Katz, 2002). During ear hole healing inMRLmice, a blastemal-like structure forms as early as day 4 post-injuryin the form of a swelling along the circumference of thewound (Heber-Katz, 1999). We have isolated cells from this MRL blastemal structure(MRL-B) on day 18 post injury, adapted the cells to culture andcharacterized the cells. The present work focused on the isolated cell-free matrix of these cells and its ability to reduce scar tissue formationand stimulate regenerative healing.
An intact ECM is present in all tissues and organs and it is secreted bythe tissue cells to provide a physical bioscaffold for cellular support. TheECM is a complex mixture of molecules comprised of collagen, elasticfibers, heparin, proteoglycans, as well as associated non-matrixmolecules such as growth factors (Brown et al., 2006; Badylak, 2007;

Fig. 1. After receiving a 2 mm diameter ear punch, the wound closure of ear holes wascompared in MRL (A–D) and B6 (E–H) mice at time zero (A,E), 11 days (B,F) and18 days (C,G) post wounding. During earhole closure in MRL mice, a swelling occurs atthe wound edge and a blastema like structure forms (*). This does not occur in B6 mice.The region shown (dotted line) was extracted on day 18 and MRL blastemal cells wereisolated from this structure in an explant culture. By day 25, MRL mouse ear holes are90% closed (D) whereas B6 mouse ear holes remain unclosed at 64 days (H). Therestored tissue in MRL mouse ears has normal histological architecture with nerves,muscle and hair follicles and is not scar tissue (scale bar=1 mm).
691E. Vorotnikova et al. / Matrix Biology 29 (2010) 690–700
Paige et al., 1991). The ECM is known to serve as a natural reservoir ofthese growth factors and other signaling molecules. Thus, the ECMfunctions as a physical support for cells; but it is also in a state ofcontinuous remodeling catalyzed by the rate limiting degradationenzymes, the matrix metalloproteinases (MMPs). Remodeling of theECM is involved in survival, migration and proliferation of many celltypes and has been implicated in the wound healing response. Recentlythe ECM has been shown to have important roles in the normal andabnormal response of cells and tissues to injury (Chen et al., 1999;Armour et al., 2006). ECMs from such decellularized tissues and organsas the small intestine (Badylak et al., 1995; Kropp et al., 1995), liver(Brown et al., 2006), urinary bladder (Chen et al., 1999), arterialvasculature, heart valves (Cebotari et al., 2006), heart, and dermis(Armour et al., 2006) have been used as biological scaffolds and havealso been shown to promote reconstruction of injured tissue andfacilitate tissue remodeling and healing processes (Sclamberg et al.,2004; Badylak et al., 2005; Badylak, 2007). It has been noticed thatscaffold degradation is necessary for the realization of these acceleratedprocesses (Badylak, 2007). The heparin found within ECMs has beenshown tobind to andprotect growth factors fromdegradation causedbyheat, trypsin or chymotrypsin digestion, as well as highly basic or acidpH changes in thewound environment (Onda et al., 1990). Thus, duringmatrix remodeling, numerous stored matrix-associated molecules aremobilized. Previous studies have shown that matrix-derived moleculesproduced in the course of acid, heat and enzymatic treatments of ECMretain bioactivity and can go on to affect angiogenesis (Li et al., 2004),cellmigration (Li et al., 2004; Badylak et al., 2001; Reing et al., 2009) andproliferation (Reing et al., 2009) and possess antimicrobial activity(Brennan et al., 2006; Sarikaya et al., 2002). The present study focusedon products derived from an ECM produced by cells isolated from theMRL blastema and their ability to evoke tissue regeneration.
2. Materials and methods
All animal study protocols were in compliance with the Universityof Massachusetts guidelines and the National Research Council'scriteria for the humane care of animals and were conducted usingapproved IACUC procedures.
2.1. MRL blastema cell isolation and cell culture
MRL-B cells were isolated from the ears of MRL/MpJ mice (JacksonLaboratories, Bar Harbor, Maine) as previously described (Reing et al.,2009; Bedelbaeva et al., 2009). Briefly, each of the ears of six MRL micereceived a 2.0 mm hole punch. B6 mice receiving ear hole punches inparallel did not form a blastema nor close the wounds even 54 daysafter ear punching whereas MRL mice after constructing a blastemalstructure closed a 2 mm wound by 35–40 days (Fig. 1). On day 18(MRL-B d.18) post-wounding, when a blastema-like structure wasmaximal, a donut shaped tissue from the edge of each earhole fromthree mice was cut (Fig. 1) and pooled and placed in a Petri dish withDMEM supplemented with 2 mM L-glutamine, 1× non-essential aminoacids, 20 U/mL Penicillin–20 ug/mL Streptomycin, 0.25 ug/mL fungi-zone, 0.1 mM B-mercaptoethanol and 10% fetal calf serum (FCS). Thetissue was minced and then incubated in a 37 °C/5%CO2 incubator forthree hours. The particulate tissue was then removed and mediareplaced every day for three days to remove dead or dying cells. Cellsthat grew out of these explants were then grown in DMEM (Cat. No.D6429, Sigma-Aldrich, St. Louis, MO) supplemented with 10% FCS,supplemented with 2 mM glutamine, 100 U/mL penicillin and 100 μg/mL streptomycin at 37 °C under a humidified atmosphere in 95% air/5%CO2 with media changes every three days and characterized.
Human aortic endothelial cells (HAEC) were purchased from Lonza(Walkersville, MD). HAEC were cultured in EGM2 medium andcorresponding supplements (Lonza, Walkersville, MD) under a humid-ified atmosphere in 95% air/5% CO2 and were used in migration assays.
Bovine capillary endothelial cells (BCE cells) were purchased fromClonetics and cultured in DMEM (GIBCO) and a final concentration of5% FCS and were used in proliferation assays. Human multilineageprogenitor(MLPC) cells were obtained from BioE (St. Paul, MN) andcultured in mesenchymal stem growth media (Cambrex, EastRutherford, NJ) under humidified 5% CO2 at 37 °C.
2.2. RNA isolation and analysis
RT-PCR was used to detect and characterize gene expression of theday 18 MRL-B cells and compared to expression patterns in humanMLPC cells.We evaluated the expression of themarkers Tenascin C, Thy-1, Dlk (Pref-1), Msx-1, Thrombospondin (THBS), and Tbx-5. Inpreparation for RT-PCR analysis, MRL-B d.18 cells and MLPC cells weregrown to at least 50% confluence in 75 cm2
flasks, collected and lysed.

692 E. Vorotnikova et al. / Matrix Biology 29 (2010) 690–700
Total RNA was extracted using the Qiagen RNA easy Kit (Qiagen,Valencia, CA) via manufacturer's instructions with the recommendedDNase treatment for removal of genomic DNA. RNA concentrationswere determined by measuring absorbance at 260 nm on a BioMate3spectrophotometer (Thermo Spectronic, Rochester, NY). First strandcDNA was synthesized from 1 μg of RNA using the Superscript First-Strand Synthesis System (Invitrogen, Carlsbad, CA) according to themanufacturer's instructions. PCR was performed with primers specificfor Tbx-5, Msx-1, Pref-1 (DLK-1), Thy1, Tenascin C (TnC), Thrombos-pondin (THBS) and glyceraldehyde 3-phosphate dehydrogenase(GAPDH) as previously described by us (Reing et al., 2009). cDNAswere amplified in EasyStart Micro100 tubes (Molecular BioProducts,San Diego, CA) with 100 nM of primer in a Mastercycler (Eppendorf,Westbury, NY). All PCR runs were 35 cycles. PCR products wereseparated on 2% agarose gels, stained with ethidium bromide, andvisualized on Kodak Image Station 2000R (Rochester, NY). For eachdistinct marker, mouse positive and negative control tissues or cell lineRNA was processed in parallel. For MRL blastemal cell studies (mouse):positive and negative controls, respectively, were for tenascin, mouseNIH3T3 cell RNA vs. mouse spleen tissue RNA; for Thy-1, NIH3T3 cell vs.mouse lung tissue RNA; for DLK and THBS, NIH3T3 vs. mouse hearttissue; for MSX-1, mouse brain tissue vs. NIH3T3 cells; and for Tbx5,mouse heart tissue vs. brain tissue RNA were used. For humanmultilineage progenitor cells for tenascin C, Thy-1 and DLK, positiveand negative controls, respectively, were human liver RNA vs. MCF-7,the humanbreast cancer cell lineRNA; forMsx-1,MCF-7vs. human liverRNA was used; for THBS, RNA from human liver vs. human heart wasused; and finally, for TBX5, human heart vs. human liver RNAwas used.
2.3. MRL blastemal cell matrix isolation
MRL-B cells were grown in DMEM (Sigma, D6429) with 10% FCS,1% fungizone and 1% GPS (200 mM glutamine, 100 U/mL penicillin,100 μg/mL streptomycin) till confluency in T-150 flasks to maximizeECM production by the cells. Media was changed after 24 h, and everythree days until the flask bottom was completely covered by cells.
To remove cells non-proteolytically, we used a method developedin our laboratory (Braunhut et al., 1992; Paige et al., 1991; Braunhut &Palomares, 1991) using EGTA (glycol-bis(2-aminoethylether)-N,N,N′,N′-tetraacetic acid, 5×10−4 M, pH=7.4), a calcium chelator. Thismethod exploits the calcium dependence of integrin binding and byremoving all extracellular calcium, the cells release passively andintact from their underlying matrix, leaving on the dish anuncontaminated and non-degraded ECM. At confluency, cells werewashedwith calcium free PBS and then removed from thematrix withfour successive 1-hour EGTA washes on a velocity table, 220 rpm, at4 °C. After each wash, EGTA with the floating cells was removed, afresh EGTA solution was added to the flask, and the flask was put onthe shaker for the next 1-hour wash. Fetal bovine serumwas added tothe removed cells in each EGTA solution and cells centrifuged. Thecells in the pellet were treated with trypsin for 10 min and countedelectronically using an electronic Coulter Counter (Beckman CoulterZ1). This procedure was repeated 4 times until all the cells were liftedfrom the ECM. Cells in 8-chamber Labteks were treated in parallel, andat 24 h the cell free ECM in labteks was fixed with methanol, rinsedwith PBS, blocked with normal serum for 30 min and then visualizedusing indirect immunohistochemistry with rabbit IgG raised tohuman fibronectin (Zymed Laboratories, San Franciso, CA) visualizedusing FITC-labeled anti-rabbit antibody conjugates (1:100). The cell-free ECMs were also stained after methanol fixation using 4′6-diamidino-2-phenlindole to detect DNA, as described previously by us(Vorotnikova et al., 2006). The unfixed flask derived cell-free matrixand the cell preparations derived from MRL-B cultures in the T-150flasks, were normalized for cell numbers and either incubated in amammalian cell lysis and extraction RIPA buffer consisting of 25 mMTris–HCl pH 7.6, 150 mM NaCl, 1% NP-40, 1% Sodium deoxycholate
and 0.1% SDS from Pierce Products (Thermo Fisher Scientific, Rock-ford, IL.) for multiplexmicroELISA analysis, or thematrix was digestedwith enzymes for extraction of ECM derived products.
2.4. Protein multiplex array analysis of cell and ECM associated growthfactors
In three separate experiments, the cell-free ECM derived fromMRL-B cells and the cells from these cultures, were collected. The cellswere counted and the matrix or cell lysates were normalized for thesame number of cells (matrix synthesized by the same number of cellsas cells themselves), and solubilized in the above RIPA buffer andanalyzed using the SearchLight Multiplex chemiluminescent sand-wich-ELISA systems based on arrays Human angiogenesis array 2 (cat# 84694) and mouse cytokine array 1 (cat # 84649) obtained fromPierce Products (Thermo Fisher Scientific, Rockford, IL.). A standardcurve of pure growth factor representing each cytokine was analyzedin parallel and each sample was analyzed in triplicate at threedifferent dilutions in each experiment to confirm quantification.Precise quantification of chemiluminescent was performed using theCCD imaging and array software provided by Pierce Biotechnology(Thermo Fisher Scientific, Rockford, IL).
2.5. Enzymatic digestion of ECM
For the enzymatic derivation of bioactive ECM products, the cellfree ECM was digested with 1 mg/ml of pepsin in 0.01 N HCl on theshaker at 4 °C overnight. Then the pepsin digest was neutralized with1:10 of 0.01 N NaOH to pH 7.0 frozen and stored at −70 °C until use.ECM derived products were assayed for in vitro chemotactic andmitogenic activity, and in vivo wound healing activities as follows. Inthe chemotactic assays, doses of pepsin digest were tested (withpepsin protein still present) ranging from 39 to 115 μg dry weight/mL,representing approximately 1.25–3.75 μg/mL of matrix material. Thepepsin is inactive once returned to neutral pH. Pepsin solution of0.01 N HCl, without MRL-B matrix, was prepared and neutralized inparallel, and used as a control in all assays (vehicle alone).
2.6. Heparin protection of FGF from pepsin degradation
The form of heparin found in ECMwas previously shown to protectheparin-binding growth factors including FGF from denaturation byheat (60, 90 °C), extremepH (pH2 or 9) or enzymatic digestion (trypsinor chymotrypsin) (Onda et al., 1990). Hog intestinal matrix derivedunfractionated sodium heparin (Lot.80788) was a generous gift fromPfizer (Franklin Operations), and was reconstituted in double distilledwater (20 mg/ml). bFGF was obtained from Sigma and was recon-stituted in 50 mM Tris–HCl pH7.5 (50 μg/ml). Pepsin treatment of FGFwas as above for ECM preparations and trypsin digest was carried outusing trypsin/EDTA solution from Gibco (Carlsbad, CA) (0.5 mg/ml) aspreviously reported (Onda et al., 1990). To perform degradationprocedures, 1 μg aliquots of bFGF were equilibrated at 37 °C for 5 minand incubated with 30 μg or 100 μg of heparin or with PBS alone(without Heparin). One half hour later, solutions containing pepsin,trypsin, or HCL were added. The solutions with trypsin or HCL wereincubated at 37 °C for 3 h and the solutionswith pepsin were incubatedat4 °Covernight inparallelwithbFGFalone. Sampleswere thenbroughtto neutral pH and analyzed in three day proliferation assays usingcapillary endothelial cells or by western blotting using an antibody tobFGF and visualized using a secondary antibody labeled with amplifiedalkaline phosphatase immuno-blot kit (BioRad 170-1643).
2.7. Neuro probe chamber chemotaxis assays using MRL-B and ECs
Migration of MRL-B and ECs to matrix derived products wereevaluated utilizing the Neuro Probe 48-well micro chemotaxis

693E. Vorotnikova et al. / Matrix Biology 29 (2010) 690–700
chamber (Neuro Probe Inc., Gaithersburg, MD). Cells to be assayed formigration responses were starved for 14 to 17 h in media with noadded growth factors but containing 0.5% heat-inactivated FCS. Thestarved cells were then harvested with trypsin, resuspended in serumfree media at a concentration of 6×105 cells/mL, and pre-incubatedfor 1 h in a humidified 95% air/5% CO2 37 °C incubator. PolycarbonatePFB chemotaxis filters (Neuro Probe, Gaithersburg, MD), 8 μm poresize filters for ECs and 12 μm pore size filters for MRL-B cells, werecoated with 0.05 mg/mL collagen I (BD Biosciences, San Jose, CA). TheECM derived products collected from the enzyme digestion at varyingconcentrations were added to the bottom chamber wells, the filterwas put in place over the bottom chamber, and the apparatus wasassembled according to manufacturer's instructions. Approximately30,000 cells were then added to each upper chamber well of theapparatus and the chamber was incubated for 3 h at 37 °C under ahumidified atmosphere in 95% air/5% CO2. In some experiments, ECMproducts were added to the top chamber as a control. No increase inthe appearance of cells to the lower chamber was observed,eliminating haptotaxis effects. Cells remaining on the topside of themembrane (i.e., non-migrated cells) were removed and then cells onthe bottom side of the membrane (i.e., migrated cells) were stainedwith Diff Quik (Dade AG Liederbach, Germany). Three pre-determinedfields were counted from each well at 20× magnification. Fields werepre-determined by location in the well: cells in the top left, top right,and bottomcenterfields in eachwellwere counted. Each experimentalconditionwas tested in quadruplicatewells and the averagenumber ofmigrated cells was determined for each condition. The Student's t-testwas used to test the null hypothesis that there was no differencebetween the results of cellmigration toward ECMdegradation productand the appropriate control buffer. P values≤0.05 were consideredsignificant.
2.8. Proliferation assays using MRL-B and ECs
Quantitative measurement of MRL-B and EC proliferation wasdetermined in a 3 day proliferation assay as previously described(Braunhut et al., 1992). Cells were plated in their media withoutgrowth factor additions at 1000 cells per well in a 48-well plate. 24 hlater, media was removed, the cells were washed, and media wasreplenished. At the time of media replenishment, samples or vehiclecontrols were added at varying doses to wells in triplicate. Time zerocell counts were determined in three of the wells using trypsinizationand electronic cell counting (Beckman Coulter Counter Z1). Threedays after sample additions, cells in triplicate wells for buffer aloneand each treatment group were trypsinized and counted electroni-cally using a Coulter counter. Experiments were repeated at leastthree times and data were pooled and analyzed. The mean value andstandard error were determined for each condition. The Student's t-test was used to test the null hypothesis that there was no differencebetween the results of cell growth for cells grown in the presence ofmatrix products and those that were grown in the presence ofneutralized pepsin alone; or when comparing samples of bFGF, beforeand after degradation, with or without heparin protection.
2.9. Full-thickness excisional skin wound assays in mice
For each experiment, thirty C57BL/6 J female mice, 7 weeks oldand weighing 20–25 g, were obtained from Jackson Laboratory (BarHarbor, ME). The mice were anesthetized with IP injections of avertinat a dose of 250 mg/kg. The fur of the dorsum of each animal wasshaved with a small cordless rechargeable trimmer (Wahl ClipperCorporation, Sterling, IL), and the skin of the entire body cleaned anddecontaminated by wiping the whole body with 10% Povidone iodinetopical solution (Maxi Drug, Inc.,Warwick, RI) and 70% ethyl alcohol.Two full thickness stage IV excisional wounds were inflicted, one oneach dorsal side of the mice, with a skin punch specifically made for
this purpose. Placement of the wounds was as follows. From the baseof the skull to a point 3.5 cm down the spinal cord, a mark was madealong the midline. Two additional points were marked, 1 cm on eitherside of this point to delineate the centers of two excisional wounds.Then the punch was used to pierce the epidermis, dermis, andpanniculus carnosus muscle. Oval sections of dorsal skin andunderlying paniculous carnosus muscle (with average diameterequal to approximately 9 mm)were removed (Tomlinson & Ferguson,2003; Birch et al., 2005). The mice were divided into: control andtreatment groups, 15 animals per group. The control group received asingle topical application of neutralized pepsin (80 μl) on bothwounds; the treatment group received a single topical application ofMRL blastema cell ECM pepsin digest (80 μl, corresponding to thematrix produced by 8.8×103cells/wound) on each wound at the timeof wounding. Each wounded animal was then placed on heating padsuntil they awakened and then housed separately in sterile isolationcages until being euthanized.
2.10. Wound closure rates and histology
The mouse wounds were photographed and measured using anelectronic vernier caliper on day 0, 3, 8, 13, and 28 post-wounding bybriefly anesthetizing animals under a vapor isoflurane anesthesiadevice. For wound closure rates, each wound was measured acrossfour different diagonals and the diameter of 2 wounds per animal wasaveraged at each time point, all animals in each treatment group weremeasured. Thus, on day 0, 3, 8, 13 and 28, the diameters of 30, 24, 18,12, and 6 wounds, respectively, were measured per treatment group.After wound diameter measurements, on day 1, 3, 8, 13 and 28 postwounding, 3 animals per each group were euthanized by CO2
asphyxiation and 2 cross-sectional full-thickness wounds from eachanimal were biopsied to include a margin of naïve skin and collectedfor histological examination. The samples were fixed in 10% bufferedformalin, passed through different grades of alcohol for dehydrationand embedded in paraffin wax. Medial samples were sectioned(5 mm) perpendicular to the surface and stained with Hemotoxylin-Eosin (H&E) andMasson's trichrome stain. Sectionswere assessed in ablinded fashion under the light microscope (Olympus BX41, OlympusAmerica Inc., Center Valley, PA) and digitized images captured. Forstandard histology using H & E and Masson's trichrome stains, tissueswere scored in a blinded study for the degree of inflammation, re-epithelialization, connective tissue organization and granulation.
2.11. Statistical analysis
All experiments were repeated at least three times and the datawere pooled. Statistical analysis was carried out using unpairedStudent's t test for independent samples and P value less than 0.05was considered significant. All error bars shown on graphs representSE. Data representing a minimum of three and up to nine separateexperiments were then pooled and plotted as a percent change of theexperimental value relative to the vehicle alone treated groupsperformed in parallel.
3. Results
3.1. Ear hole closure and isolation of blastemal cells from MRL wounds
It has previously been reported that MRL mice will close a 2.0 mmear hole punch over a 30–40 day period (Clark et al., 1998;Samulewicz et al., 2002). During the course of ear hole closure, wenoted the formation of an avascular structure that others havedescribed as a blastema-like structure. We produced ear holes in MRL(panel A–D) vs. B6 (Panel E–H)mice andmonitored the formation of ablastemal like structure (*) and ear hole closure over a 64 day timecourse (Fig. 1). We noted the formation of a avascular zone at the edge
694 E. Vorotnikova et al. / Matrix Biology 29 (2010) 690–700
of a 2 mm ear hole in MRL mice approximately 4 days after ear punchand this avascular zone was well developed by day 11 (B) and day 18(C) in these mice (*). On day 18, the blastemal area shown wasexcised (dotted line, panel C) and cells were extracted from the tissue(Fig. 1). In our studies of MRL female mice, all the 2 mm ear holeswere 90% closed by day 25 (D). In contrast, B6 mice receivingequivalent ear hole punches at time zero (E) do not close the woundsby 25 days (H) and the majority of B6 mice have ear holes that persistbeyond 64 days. Cells were isolated from the day 18 MRL blastemalstructures, and we characterized the day 18 cells after continuouspassage in vitro prior to our matrix isolation procedures.
3.2. Characterization of MRL blastemal cells as possessing progenitor cellmarkers
RT-PCR studies demonstrated that day 18 MRL-B cells (Fig. 2B), evenafter continuous culture, express several well accepted progenitor cellmarkers including Tenascin C, Thy 1, DLK (Pref-1), MSX-1 , thrombos-pondin (THBS), and Tbx5 (Abdallah et al., 2004; Masson et al., 2006; vonHolst, 2007) andwhen compared to humanmultilineage progenitor cells(Fig. 2A). InpanelB, thefirst lane represents theMRLblastemacells (MRL-B d.18 cells), the second lane represents the appropriate negative controlcell line or tissue for that marker and lane 3 represents the appropriatepositive control cell line or tissue in the mouse for the mRNA beinganalyzed. The expression pattern of cultured MRL Blastema cells iscomparable to their expression of these markers reported in situ duringearhole closure (Naviaux et al., 2009).
3.3. Isolation of the matrix and cell fractions from cultures of MRL-B cells
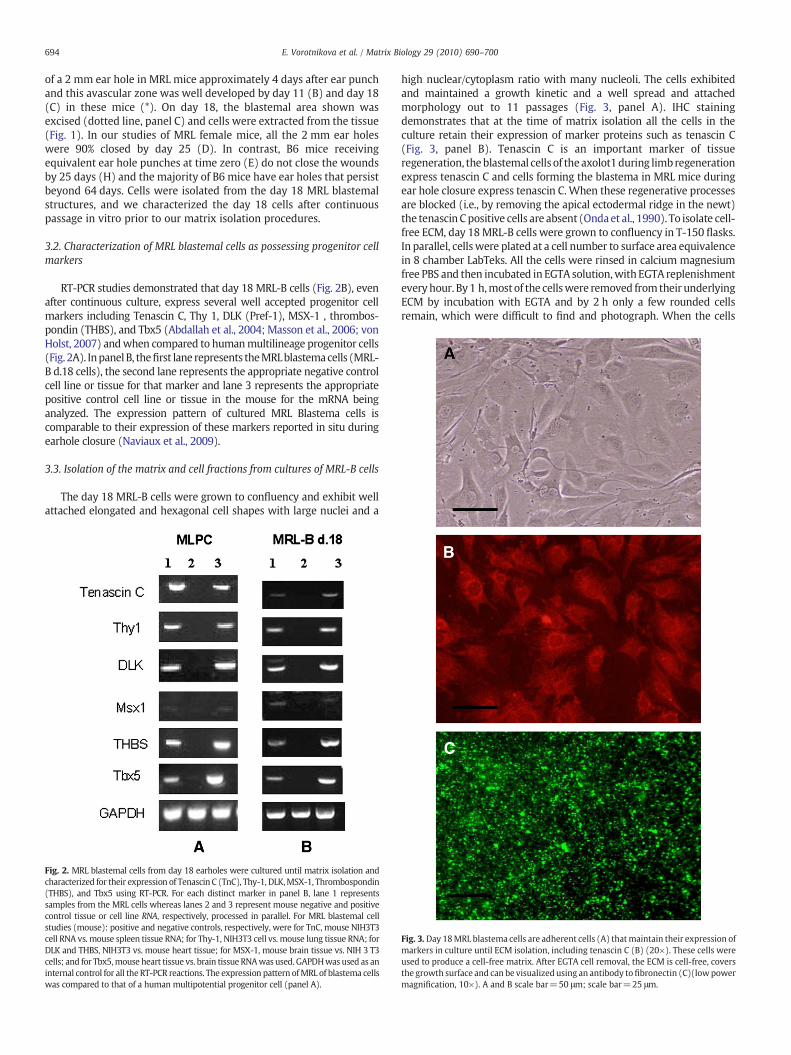
The day 18 MRL-B cells were grown to confluency and exhibit wellattached elongated and hexagonal cell shapes with large nuclei and a
Fig. 2. MRL blastemal cells from day 18 earholes were cultured until matrix isolation andcharacterized for their expression of Tenascin C (TnC), Thy-1, DLK,MSX-1, Thrombospondin(THBS), and Tbx5 using RT-PCR. For each distinct marker in panel B, lane 1 representssamples from the MRL cells whereas lanes 2 and 3 represent mouse negative and positivecontrol tissue or cell line RNA, respectively, processed in parallel. For MRL blastemal cellstudies (mouse): positive and negative controls, respectively, were for TnC, mouse NIH3T3cell RNA vs. mouse spleen tissue RNA; for Thy-1, NIH3T3 cell vs. mouse lung tissue RNA; forDLK and THBS, NIH3T3 vs. mouse heart tissue; for MSX-1, mouse brain tissue vs. NIH 3 T3cells; and for Tbx5,mouse heart tissue vs. brain tissue RNAwas used. GAPDHwas used as aninternal control for all theRT-PCR reactions. The expression pattern ofMRL of blastema cellswas compared to that of a human multipotential progenitor cell (panel A).
high nuclear/cytoplasm ratio with many nucleoli. The cells exhibitedand maintained a growth kinetic and a well spread and attachedmorphology out to 11 passages (Fig. 3, panel A). IHC stainingdemonstrates that at the time of matrix isolation all the cells in theculture retain their expression of marker proteins such as tenascin C(Fig. 3, panel B). Tenascin C is an important marker of tissueregeneration, theblastemal cells of theaxolot1during limbregenerationexpress tenascin C and cells forming the blastema in MRL mice duringear hole closure express tenascin C.When these regenerative processesare blocked (i.e., by removing the apical ectodermal ridge in the newt)the tenascin Cpositive cells are absent (Ondaet al., 1990). To isolate cell-free ECM, day 18MRL-B cells were grown to confluency in T-150 flasks.In parallel, cellswere plated at a cell number to surface area equivalencein 8 chamber LabTeks. All the cells were rinsed in calcium magnesiumfree PBS and then incubated in EGTA solution,with EGTA replenishmenteveryhour. By1 h,most of the cellswere removed fromtheir underlyingECM by incubation with EGTA and by 2 h only a few rounded cellsremain, which were difficult to find and photograph. When the cells
Fig. 3.Day 18MRL blastema cells are adherent cells (A) thatmaintain their expression ofmarkers in culture until ECM isolation, including tenascin C (B) (20×). These cells wereused to produce a cell-free matrix. After EGTA cell removal, the ECM is cell-free, coversthe growth surface and can be visualized using an antibody tofibronectin (C)(lowpowermagnification, 10×). A and B scale bar=50 μm; scale bar=25 μm.
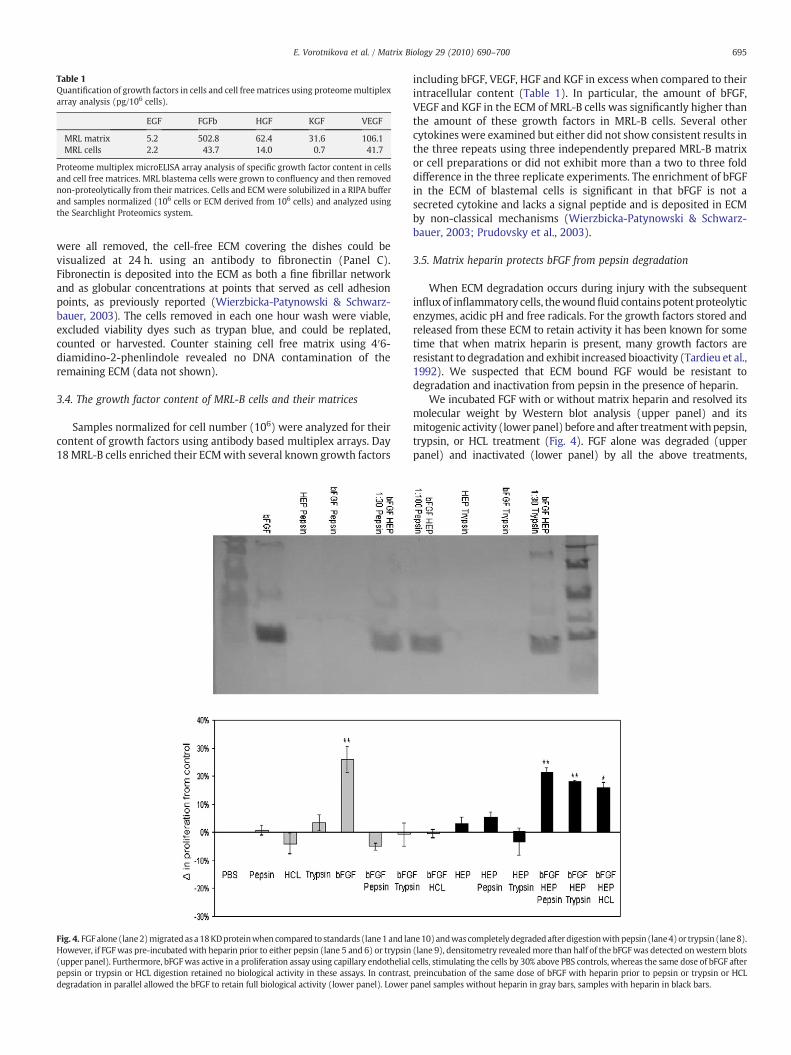
Table 1Quantification of growth factors in cells and cell freematrices using proteomemultiplexarray analysis (pg/106 cells).
EGF FGFb HGF KGF VEGF
MRL matrix 5.2 502.8 62.4 31.6 106.1MRL cells 2.2 43.7 14.0 0.7 41.7
Proteome multiplex microELISA array analysis of specific growth factor content in cellsand cell free matrices. MRL blastema cells were grown to confluency and then removednon-proteolytically from their matrices. Cells and ECMwere solubilized in a RIPA bufferand samples normalized (106 cells or ECM derived from 106 cells) and analyzed usingthe Searchlight Proteomics system.
695E. Vorotnikova et al. / Matrix Biology 29 (2010) 690–700
were all removed, the cell-free ECM covering the dishes could bevisualized at 24 h. using an antibody to fibronectin (Panel C).Fibronectin is deposited into the ECM as both a fine fibrillar networkand as globular concentrations at points that served as cell adhesionpoints, as previously reported (Wierzbicka-Patynowski & Schwarz-bauer, 2003). The cells removed in each one hour wash were viable,excluded viability dyes such as trypan blue, and could be replated,counted or harvested. Counter staining cell free matrix using 4′6-diamidino-2-phenlindole revealed no DNA contamination of theremaining ECM (data not shown).
3.4. The growth factor content of MRL-B cells and their matrices
Samples normalized for cell number (106) were analyzed for theircontent of growth factors using antibody based multiplex arrays. Day18MRL-B cells enriched their ECMwith several known growth factors
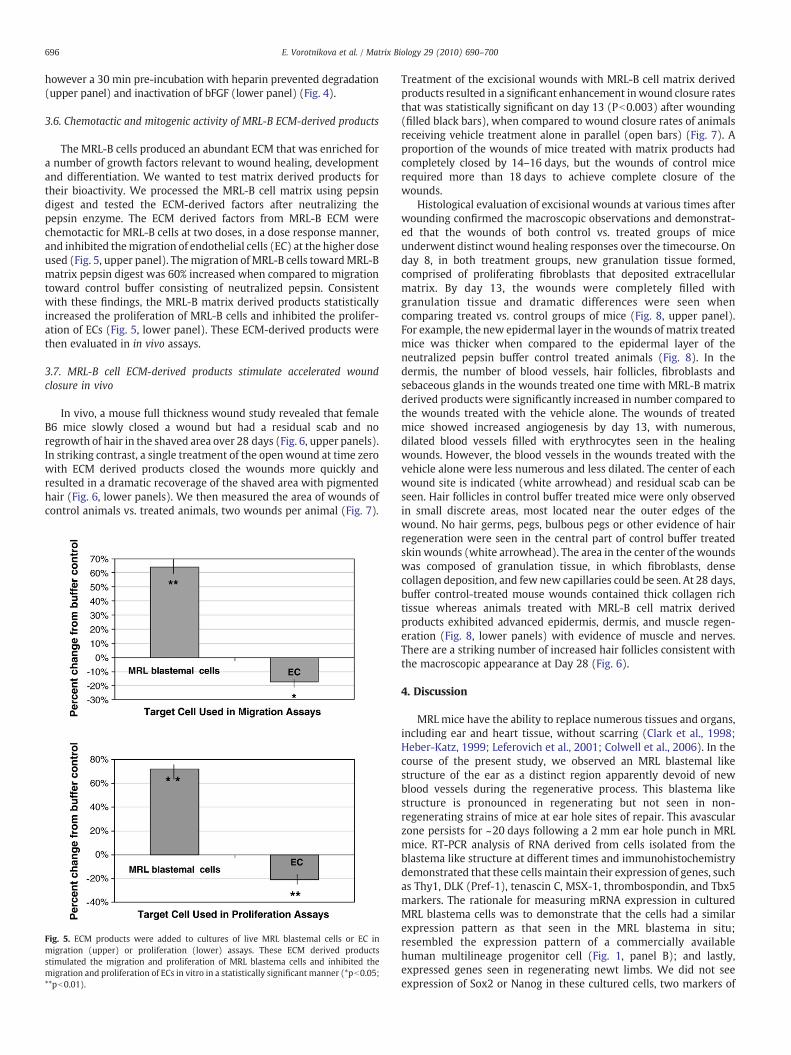
Fig. 4. FGFalone (lane2)migratedasa18KDproteinwhen compared to standards (lane1and laHowever, if FGFwas pre-incubatedwith heparin prior to either pepsin (lane 5 and 6) or trypsin(upper panel). Furthermore, bFGFwas active in a proliferation assay using capillary endothelialpepsin or trypsin or HCL digestion retained no biological activity in these assays. In contrast,degradation in parallel allowed the bFGF to retain full biological activity (lower panel). Lower
including bFGF, VEGF, HGF and KGF in excess when compared to theirintracellular content (Table 1). In particular, the amount of bFGF,VEGF and KGF in the ECM of MRL-B cells was significantly higher thanthe amount of these growth factors in MRL-B cells. Several othercytokines were examined but either did not show consistent results inthe three repeats using three independently prepared MRL-B matrixor cell preparations or did not exhibit more than a two to three folddifference in the three replicate experiments. The enrichment of bFGFin the ECM of blastemal cells is significant in that bFGF is not asecreted cytokine and lacks a signal peptide and is deposited in ECMby non-classical mechanisms (Wierzbicka-Patynowski & Schwarz-bauer, 2003; Prudovsky et al., 2003).
3.5. Matrix heparin protects bFGF from pepsin degradation
When ECM degradation occurs during injury with the subsequentinfluxof inflammatory cells, thewoundfluid containspotentproteolyticenzymes, acidic pH and free radicals. For the growth factors stored andreleased from these ECM to retain activity it has been known for sometime that when matrix heparin is present, many growth factors areresistant to degradation and exhibit increased bioactivity (Tardieu et al.,1992). We suspected that ECM bound FGF would be resistant todegradation and inactivation from pepsin in the presence of heparin.
We incubated FGF with or without matrix heparin and resolved itsmolecular weight by Western blot analysis (upper panel) and itsmitogenic activity (lower panel) before and after treatmentwithpepsin,trypsin, or HCL treatment (Fig. 4). FGF alone was degraded (upperpanel) and inactivated (lower panel) by all the above treatments,
ne10)andwascompletelydegradedafter digestionwithpepsin (lane4)or trypsin (lane8).(lane 9), densitometry revealedmore than half of the bFGFwas detected onwestern blotscells, stimulating the cells by 30% above PBS controls, whereas the same dose of bFGF afterpreincubation of the same dose of bFGF with heparin prior to pepsin or trypsin or HCLpanel samples without heparin in gray bars, samples with heparin in black bars.
696 E. Vorotnikova et al. / Matrix Biology 29 (2010) 690–700
however a 30 min pre-incubation with heparin prevented degradation(upper panel) and inactivation of bFGF (lower panel) (Fig. 4).
3.6. Chemotactic and mitogenic activity of MRL-B ECM-derived products
The MRL-B cells produced an abundant ECM that was enriched fora number of growth factors relevant to wound healing, developmentand differentiation. We wanted to test matrix derived products fortheir bioactivity. We processed the MRL-B cell matrix using pepsindigest and tested the ECM-derived factors after neutralizing thepepsin enzyme. The ECM derived factors from MRL-B ECM werechemotactic for MRL-B cells at two doses, in a dose response manner,and inhibited themigration of endothelial cells (EC) at the higher doseused (Fig. 5, upper panel). Themigration of MRL-B cells towardMRL-Bmatrix pepsin digest was 60% increased when compared to migrationtoward control buffer consisting of neutralized pepsin. Consistentwith these findings, the MRL-B matrix derived products statisticallyincreased the proliferation of MRL-B cells and inhibited the prolifer-ation of ECs (Fig. 5, lower panel). These ECM-derived products werethen evaluated in in vivo assays.
3.7. MRL-B cell ECM-derived products stimulate accelerated woundclosure in vivo
In vivo, a mouse full thickness wound study revealed that femaleB6 mice slowly closed a wound but had a residual scab and noregrowth of hair in the shaved area over 28 days (Fig. 6, upper panels).In striking contrast, a single treatment of the open wound at time zerowith ECM derived products closed the wounds more quickly andresulted in a dramatic recoverage of the shaved area with pigmentedhair (Fig. 6, lower panels). We then measured the area of wounds ofcontrol animals vs. treated animals, two wounds per animal (Fig. 7).
Fig. 5. ECM products were added to cultures of live MRL blastemal cells or EC inmigration (upper) or proliferation (lower) assays. These ECM derived productsstimulated the migration and proliferation of MRL blastema cells and inhibited themigration and proliferation of ECs in vitro in a statistically significant manner (*pb0.05;**pb0.01).
Treatment of the excisional wounds with MRL-B cell matrix derivedproducts resulted in a significant enhancement inwound closure ratesthat was statistically significant on day 13 (Pb0.003) after wounding(filled black bars), when compared to wound closure rates of animalsreceiving vehicle treatment alone in parallel (open bars) (Fig. 7). Aproportion of the wounds of mice treated with matrix products hadcompletely closed by 14–16 days, but the wounds of control micerequired more than 18 days to achieve complete closure of thewounds.
Histological evaluation of excisional wounds at various times afterwounding confirmed the macroscopic observations and demonstrat-ed that the wounds of both control vs. treated groups of miceunderwent distinct wound healing responses over the timecourse. Onday 8, in both treatment groups, new granulation tissue formed,comprised of proliferating fibroblasts that deposited extracellularmatrix. By day 13, the wounds were completely filled withgranulation tissue and dramatic differences were seen whencomparing treated vs. control groups of mice (Fig. 8, upper panel).For example, the new epidermal layer in the wounds of matrix treatedmice was thicker when compared to the epidermal layer of theneutralized pepsin buffer control treated animals (Fig. 8). In thedermis, the number of blood vessels, hair follicles, fibroblasts andsebaceous glands in the wounds treated one time with MRL-B matrixderived products were significantly increased in number compared tothe wounds treated with the vehicle alone. The wounds of treatedmice showed increased angiogenesis by day 13, with numerous,dilated blood vessels filled with erythrocytes seen in the healingwounds. However, the blood vessels in the wounds treated with thevehicle alone were less numerous and less dilated. The center of eachwound site is indicated (white arrowhead) and residual scab can beseen. Hair follicles in control buffer treated mice were only observedin small discrete areas, most located near the outer edges of thewound. No hair germs, pegs, bulbous pegs or other evidence of hairregeneration were seen in the central part of control buffer treatedskin wounds (white arrowhead). The area in the center of the woundswas composed of granulation tissue, in which fibroblasts, densecollagen deposition, and few new capillaries could be seen. At 28 days,buffer control-treated mouse wounds contained thick collagen richtissue whereas animals treated with MRL-B cell matrix derivedproducts exhibited advanced epidermis, dermis, and muscle regen-eration (Fig. 8, lower panels) with evidence of muscle and nerves.There are a striking number of increased hair follicles consistent withthe macroscopic appearance at Day 28 (Fig. 6).
4. Discussion
MRL mice have the ability to replace numerous tissues and organs,including ear and heart tissue, without scarring (Clark et al., 1998;Heber-Katz, 1999; Leferovich et al., 2001; Colwell et al., 2006). In thecourse of the present study, we observed an MRL blastemal likestructure of the ear as a distinct region apparently devoid of newblood vessels during the regenerative process. This blastema likestructure is pronounced in regenerating but not seen in non-regenerating strains of mice at ear hole sites of repair. This avascularzone persists for ~20 days following a 2 mm ear hole punch in MRLmice. RT-PCR analysis of RNA derived from cells isolated from theblastema like structure at different times and immunohistochemistrydemonstrated that these cells maintain their expression of genes, suchas Thy1, DLK (Pref-1), tenascin C, MSX-1, thrombospondin, and Tbx5markers. The rationale for measuring mRNA expression in culturedMRL blastema cells was to demonstrate that the cells had a similarexpression pattern as that seen in the MRL blastema in situ;resembled the expression pattern of a commercially availablehuman multilineage progenitor cell (Fig. 1, panel B); and lastly,expressed genes seen in regenerating newt limbs. We did not seeexpression of Sox2 or Nanog in these cultured cells, two markers of
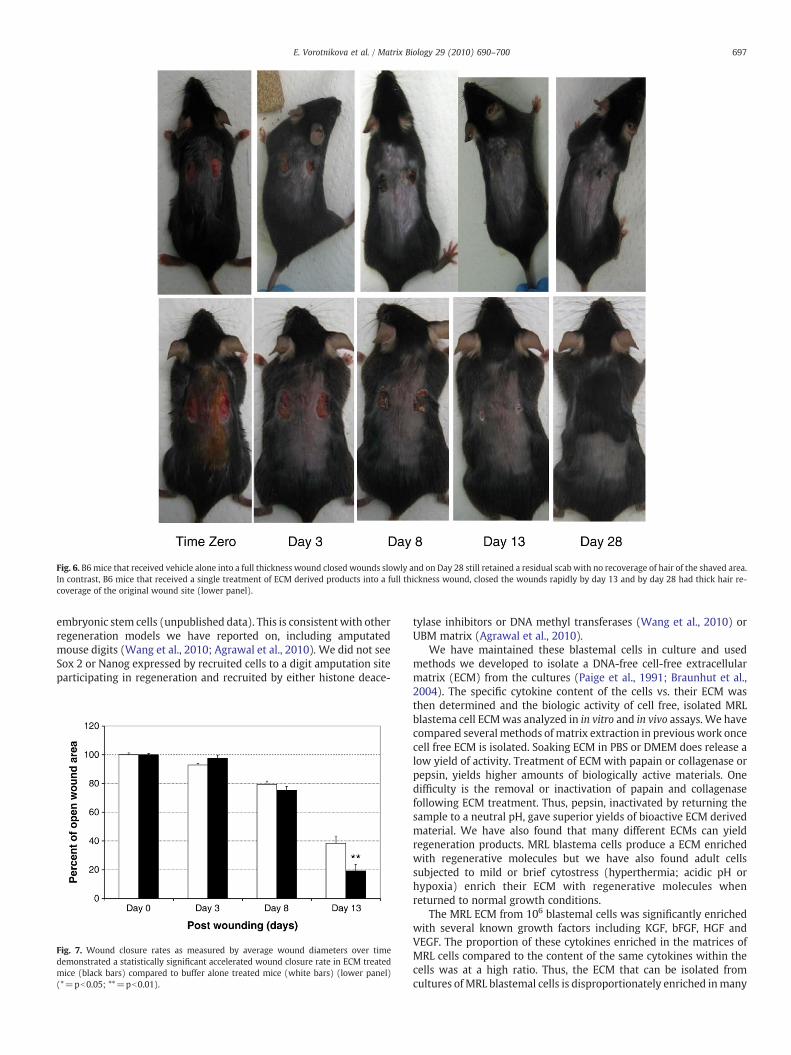
Fig. 6. B6mice that received vehicle alone into a full thickness wound closed wounds slowly and on Day 28 still retained a residual scab with no recoverage of hair of the shaved area.In contrast, B6 mice that received a single treatment of ECM derived products into a full thickness wound, closed the wounds rapidly by day 13 and by day 28 had thick hair re-coverage of the original wound site (lower panel).
697E. Vorotnikova et al. / Matrix Biology 29 (2010) 690–700
embryonic stem cells (unpublished data). This is consistent with otherregeneration models we have reported on, including amputatedmouse digits (Wang et al., 2010; Agrawal et al., 2010). We did not seeSox 2 or Nanog expressed by recruited cells to a digit amputation siteparticipating in regeneration and recruited by either histone deace-
Fig. 7. Wound closure rates as measured by average wound diameters over timedemonstrated a statistically significant accelerated wound closure rate in ECM treatedmice (black bars) compared to buffer alone treated mice (white bars) (lower panel)(*=pb0.05; **=pb0.01).
tylase inhibitors or DNA methyl transferases (Wang et al., 2010) orUBM matrix (Agrawal et al., 2010).
We have maintained these blastemal cells in culture and usedmethods we developed to isolate a DNA-free cell-free extracellularmatrix (ECM) from the cultures (Paige et al., 1991; Braunhut et al.,2004). The specific cytokine content of the cells vs. their ECM wasthen determined and the biologic activity of cell free, isolated MRLblastema cell ECMwas analyzed in in vitro and in vivo assays. We havecompared several methods of matrix extraction in previouswork oncecell free ECM is isolated. Soaking ECM in PBS or DMEM does release alow yield of activity. Treatment of ECM with papain or collagenase orpepsin, yields higher amounts of biologically active materials. Onedifficulty is the removal or inactivation of papain and collagenasefollowing ECM treatment. Thus, pepsin, inactivated by returning thesample to a neutral pH, gave superior yields of bioactive ECM derivedmaterial. We have also found that many different ECMs can yieldregeneration products. MRL blastema cells produce a ECM enrichedwith regenerative molecules but we have also found adult cellssubjected to mild or brief cytostress (hyperthermia; acidic pH orhypoxia) enrich their ECM with regenerative molecules whenreturned to normal growth conditions.
The MRL ECM from 106 blastemal cells was significantly enrichedwith several known growth factors including KGF, bFGF, HGF andVEGF. The proportion of these cytokines enriched in the matrices ofMRL cells compared to the content of the same cytokines within thecells was at a high ratio. Thus, the ECM that can be isolated fromcultures of MRL blastemal cells is disproportionately enriched inmany

Fig. 8. B6 mice that received a single treatment of ECM derived products (upper right panel) into a full thickness wound had increased number of hair follicles, new blood vessels,sebaceous glands, and a well developed dermis and epidermis by day 13 compared to wound sites of animals that had received buffer treatments (upper left panel). The center ofeach wound is indicated (white arrow head) and remnants of the clot can been seen demarcating the midline of each original wound. By day 28, buffer-treated animals had thickaligned collagen fibers and the absence of normal skin architecture in contrast to a restored dermis and epidermis at the wound site in ECM-treated mouse wounds (left vs. rightlower panel)(original magnification 100×).
698 E. Vorotnikova et al. / Matrix Biology 29 (2010) 690–700
growth factors that play a significant role in migration, proliferationand differentiation of progenitor and stem cells, and in wound repair.
Many investigators have shown that ECM produced by differenttypes of cells can serve as an ideal biological scaffold material.Bioscaffolds composed of naturally occurring ECM and processed forclinical purposes are widely used in tissue reconstruction. Forexample, ECM bioscaffolds have been shown to accelerate theconstructive remodeling of tissues in many organs such as heartvalves, urinary bladder and musculoskeletal tissues (Badylak et al.,1995; Kropp et al., 1995; Brown et al., 2006; Chen et al., 1999;Armour et al., 2006; Sclamberg et al., 2004; Badylak et al., 2005;Badylak, 2007). The mechanisms of ECM induced acceleratedhealing are not fully understood. It is known that the ECM biologicscaffolds start to degrade rapidly after implantation and that, forECM to have an impact on tissue repair, the ECM must remodelcontinuously for days to weeks (Gilbert et al., 2007). It is known thatthis degradation process is required because ECM scaffolds that are
experimentally rendered resistant to degradation produce a chronicinflammatory response and fibrous connective tissue deposition,not healing (Gilbert et al., 2007). It has been shown that growthfactors such as VEGF and bFGF are released during bioscaffolddegradation from bioscaffolds derived from pig small intestine orurinary bladder (McDevitt et al., 2003; Hodde & Hiles, 2001; Hoddeet al., 2006). These growth factors are known to promote mitogen-esis and cellular differentiation in the course of the remodelingprocess (McDevitt et al., 2003). In recent years it has becomeincreasingly apparent that it is not only known growth factorsembedded in ECM that are responsible for the biological activities ofcell free ECM. Many large ECM molecules can be processed to yieldsmall biologically active peptide fragments termed matricryptinsthat have not been purified or characterized completely, yet play animportant role in distal recruitment and mobilization of progenitorcells to a wound site (Li et al., 2004; Badylak et al., 2001; Gargett,2006).

699E. Vorotnikova et al. / Matrix Biology 29 (2010) 690–700
For example, cryptic peptides released by the ECM have beenshown to initiate and sustain the recruitment of circulating, bone-marrow-derived cells that actively participate in tissue remodeling(Sottile, 2004) and may be key in averting scar tissue formation atwound sites of non-regenerating mammals. Some of these recruitedcells have been shown to express markers of an undifferentiatedprogenitor population of cells (Badylak et al., 2001). The peptidesproduced by ECM degradation have also been demonstrated topossess potent antimicrobial activity (Brennan et al., 2006), capable ofkeeping wounds sterile during the slower regeneration process; anadded benefit of ECM derived products. We now report that thematrix produced by MRL-B cells and isolated in our laboratorycontains many important growth factors and remodeling moleculesactive in effecting migration and proliferation of cells in vitro andwound healing in vivo.
Following pepsin digestion of MRL-B cell matrices and then pepsininactivation, matrix-derived products were shown to statisticallyincrease migration and proliferation of MRL-B cells, and decreasemigration and proliferation of endothelial cells. We have also shownthat heparin found in ECM protects growth factors such as FGF frompepsin inactivation. It has long been known that bFGF when bound toheparin, a molecule enriched in most ECM, protects this growth factorfrom heat, pH, or proteolytic degradation by trypsin or chymotrypsin.Furthermore, the addition of heparin to samples of bFGF that have lostbiological activity in solution or during storage, can restore its activity.Thus bFGF and other heparin binding growth factors (VEGF, TGF-B,HB-EGF) are likely intact and in the matrix derived preparation andmay participate in the outcomes observed. The steps required forangiogenesis, namely a proliferative phase followed by a differenti-ation phase (non-proliferative), are regulated by the composition ofactive growth factors. VEG-F and FGF are chemotactic and mitogenicwhen alone, but in the presence of TGFB1, a potent antimitogen of EC,yet potent pro-angiogenic cytokine, will support cessation of ECdivision, tube formation and the differentiation of the capillary plexus.We have detected a TGF-B like activity in the pepsin-digest of MRLblastema cell matrix, that is partially neutralized when usingneutralizing antibodies to TGFB (unpublished data). TGFB can act asa differentiating factor for endothelial cells promoting vesselformation. Thus, the combination of these factors in the matrixmaterials would inhibit migration and proliferation of ECs in vitrowhile being angiogenic in vivo.
Alternatively, it is known that progenitor cells in their multi-potential state exhibit increased proliferation and resistance toapoptosis under hypoxic conditions (Csete, 2005), and one hypothesisis that in the course of early stages of tissue regeneration, blood vesselformation may be purposely delayed to give time for progenitor cellsto migrate, proliferate and accumulate at the site of injury prior todifferentiation. The inhibition of endothelial cell migration andproliferation by MRL-B cell matrix degradation products observed inthe present study may be a reflection of a possible mechanism ofaction of ECM in being permissive for regenerative repair and avertingscar tissue formation in adult animals. It may be one of the attributesof the MRL ear hole blastema remaining avascular that allows it toregenerate. Additional experimentswill need to be conducted using invivomodels to verify this hypothesis and elucidate the role of differentcell compartments, including endothelial and progenitor cells, duringthe early and later stages of regenerative tissue repair. We arecurrently using preparatory HPLC to identify and purify regenerativemolecules from ECM of a variety of embryonic and adult cell sources,grown in the presence or absence of physiologic stressors during theECM production period.
Our wound healing studies support the idea that matrix derivedproducts facilitate tissue restoration processes and avert scarformation in vivo. We demonstrate here that matrix derived productsaccelerate wound closure rates of full thickness cutaneous wounds inC57Bl/6 J mice, increase hair follicle number and restore normal
dermis and epidermal structures while at the same time reducing scartissue formation. Further in vitro and in vivo experiments are neededto characterize the MRL-B cell matrix derived products and elucidatethemechanisms of their influence on critical wound healing processesand tissue regeneration.
Acknowledgements
The work described was funded by an award from the DefenseAdvanced Research Program Agency, award # W911NF-06-1-0067.
References
Abdallah, B.M., Jensen, C.H., Gutierrez, G., 2004. Regulation of human skeletal stem cellsdifferentiation by Dlk1/Pref-1. J. Bone Miner. Res. 19, 841–852.
Agrawal, V., Johnson, S., Reing, J., Zhang, L., Tottey, S., Wang, G., Hirschi, K., Braunhut, S.J.,Gudas, L., Badylak, S., 2010. An epimorphic regeneration approach to tissuereplacement in adult mammals Feb 23 PNAS 107 (8), 3351–3355 Dec. 4 [Epubahead of print].
Armour, A.D., Fish, J.S., Woodhouse, K.A., Semple, J.L., 2006. A comparison of human andporcine acellularized dermis: interactions with human fibroblasts in vitro. Plast.Reconstr. Surg. 117, 845.
Badylak, S.F., 2007. The extracellular matrix as a biologic scaffold material. Biomaterials28, 3587–3593.
Badylak, S.F., Tullius, R., Kokini, K., Shelbourne, K.D., Klootwyk, T., Voytik, S.L., Kraine, M.R.,Simmons, C., 1995. The use of xenogeneic small intestinal submucosa as a biomaterialfor Achilles tendon repair in a dog model. J. Biomed. Mater. Res. 29, 977.
Badylak, S.F., Park, K., Peppas, N., McCabe, G., Yoder, M., 2001. Marrow-derived cellspopulate scaffolds composed of xenogeneic extracellular matrix. Exp. Hematol. 29,1310.
Badylak, S.F., Vorp, D.A., Spievack, A.R., Simmons-Byrd, A., Hanke, J., Freytes, D.O., Thapa,A., Gilbert, T.W., Nieponice, A., 2005. Esophageal reconstruction with ECM andmuscle tissue in a dog model. J. Surg. Res. 128, 87.
Bedelbaeva, K., Gourevitch, D., Heber-Katz, E., 2009. Characterization of blastemamarkers in the regenerative cells of the MRL ear in vivo and in vitro. Manuscript inpreparation.
Birch, M., Tomlinson, A., Ferguson, M.W., 2005. Animal models for adult dermal woundhealing. Meth. Mol. Med. 117, 223–235.
Braunhut, S.J., Palomares, M., 1991. Modulation of endothelial cell shape and growth byretinoids. Microvasc. Res. 41, 47–62.
Braunhut, S.J., D'Amore, P.A., Gudas, L.J., 1992. The location and expression of fibroblastgrowth factor (FGF) in F9 visceral and parietal embryonic cells after retinoic acid-induced differentiation. Differentiation 50, 141–152.
Braunhut, S.J., McIntosh, D., Vorotnikova, E., Zhou, T., Marx, K.A., 2004. Development ofa smart bandage: applying electrical potential to selectively release wound healinggrowth factors from cell free extracellular matrices. Mater. Res. Soc. Symp. Proc. 3,1–3.
Brennan, E.P., Reing, J., Chew, D., Myers-Irvin, J.M., Young, E.J., Badylak, S.F., 2006.Antibacterial activity within degradation products of biological scaffolds composedof extracellular matrix. Tissue Eng. 12, 2949.
Brown, B., Lindberg, K., Reing, J., Stolz, D.B., Badylak, S.F., 2006. The basementmembrane component of biologic scaffolds derived from extracellular matrix.Tissue Eng. 12, 519.
Cebotari, S., Lichtenberg, A., Tudorache, I., Hilfiker, A., Mertsching, H., Leyh, R.,Breymann, T., Kallenbach, K., Maniuc, L., Batrinac, A., Repin, O., Maliga, O.,Ciubotaru, A., Haverich, A., 2006. Clinical application of tissue engineered humanheart valves using autologous progenitor cells. Circulation 114, I132.
Chen, F., Yoo, J.J., Atala, A., 1999. Acellular collagen matrix as a possible “off the shelf”biomaterial for urethral repair. Urology 54, 407.
Clark, L.D., Clark, R.K., Heber-Katz, E., 1998. A new murine model for mammalianwound repair and regeneration. Clin. Immunol. Immunopath. 88, 35–45.
Colwell, A.S., Krummel, T.M., Kong, W., Longaker, M.T., Lorenz, H.P., 2006. Skin woundsin the MRL/MpJ mouse heal with scar. Wound Repair Regen. 81–90.
Csete, M., 2005. Oxygen in the cultivation of stem cells. Ann. NY Acad. Sci. 1049, 1.Gargett, C.E., 2006. Identification and characterisation of human endometrial stem/
progenitor cells. Aust. N. Z. J. Obstet. Gynaecol. 46, 250–253.Gilbert, T.W., Stewart-Akers, A.M., Simmons-Byrd, A., Badylak, S.F., 2007. Degradation
and remodeling of small intestinal submucosa in canine Achilles tendon repair.J. Bone Joint Surg. Am. 89, 621.
Gourevitch, D., Clark, L., Chen, P., Seitz, A., Samulewicz, S.J., Heber-Katz, E., 2003. Matrixmetalloproteinase activity correlates with blastema formation in the regeneratingMRL mouse ear hole model. Dev. Dyn. 226, 377–387.
Heber-Katz, E., 1999. The regenerating mouse ear. Semin. Cell Dev. Biol. 10, 415–419.Hodde, J., Hiles, M., 2001. Bioactive FGF-2 in sterilized extracellular matrix. Wounds 13,
195.Hodde, J.P., Record, R.D., Liang, H.A., Badylak, S.F., 2006. Vascular endothelial growth
factor in porcine-derived extracellular matrix. Endothelium 8, 11–24.Kropp, B.P., Eppley, B.L., Prevel, C.D., Rippy, M.K., Harruff, R.C., Badylak, S.F., Adams, M.C.,
Rink, R.C., Keating, M.A., 1995. Experimental assessment of small intestinalsubmucosa as a bladder wall substitute. Urology 46, 396.
Leferovich, J.M., Heber-Katz, E., 2002. The scarless heart. Semin. Cell Dev. Biol. 13,327–333.

700 E. Vorotnikova et al. / Matrix Biology 29 (2010) 690–700
Leferovich, J.M., Bedelbaeva, K., Samulewicz, S., Zhang, X.M., Zwas, D., Lankford, E.B.,Heber-Katz, E., 2001. Heart regeneration in adult MRL mice. Proc. Natl Acad. Sci.USA 98, 9830–9835.
Li, F., Li, W., Johnson, S., Ingram, D., Yoder, M., Badylak, S.F., 2004. Low-molecular-weight peptides derived from extracellular matrix as chemoattractants for primaryendothelial cells. Endothelium 11, 199.
Masson, N.M., Currie, I.S., Terrace, J.D., Garden, O.J., Parks, R.W., Ross, J.A., 2006. Hepaticprogenitor cells in human fetal liver express the oval cell marker Thy-1. Am. J.Physiol. Gastrointest. Liver Physiol. 291, G45–G54.
McDevitt, C.A., Wildey, G.M., Cutrone, R.M., 2003. Transforming growth factor-beta1 ina sterilized tissue derived from the pig small intestine submucosa. J. Biomed. Mater.Res. 67A, 637–640.
Naviaux, R.K., Le, T.P., Bedelbaeva, K., Leferovich, J., Gourevitch, D., Sachadyn, P., Zhang,X.M., Clark, L., Heber-Katz, E., 2009. Retained features of embryonic metabolism inthe adult MRL mouse. Mol. Genet. Metab. 96, 133–144.
Onda, H., Goldhamer, D.J., Tassava, R.A., 1990. An extracellular matrix molecule of newtand axolotl regenerating limb blastemas and embryonic limb buds: immunologicalrelationship of MT1 antigen with tenascin. Development 108, 657–668.
Paige, K., Palomares, M., D'Amore, P.A., Braunhut, S.J., 1991. Retinol-inducedmodification of the extracellular matrix of endothelial cells: its role in growthcontrol. In Vitro Cell. Dev. Biol. 27, 151–157.
Prudovsky, I., Mandinova, A., Soldi, R., Bagala, C., Graziani, I., Landriscina, M., Tarantini,F., Duarte, M., Bellum, S., Doherty, H., Maciag, T., 2003. The non-classical exportroutes: FGF1 and IL-1alpha point the way. J. Cell Sci. 116, 4871–4881.
Reing, J.E., Zhang, L., Myers-Irvin, J., Cordero, K., Freytes, D., Heber-Katz, E., Bedelbaeva,K., McIntosh, D., Dewilde, A., Braunhut, S.J., Badylak, S.F., 2009. Degradationproducts of extracellular matrix affect cell migration and proliferation. Tissue Eng.A 15 (3), 605–614.
Samulewicz, S.J., Seitz, A., Clark, L., Heber-Katz, E., 2002. Expression of preadipocytefactor-1(Pref-1), a delta-like protein, in healing mouse ears. Wound Repair Regen.10, 215–221.
Sarikaya, A., Record, R., Wu, C.C., Tullius, B., Badylak, S.F., Ladisch, M., 2002.Antimicrobial activity associated with extracellular matrices. Tissue Eng. 8, 63–71.
Sclamberg, S.G., Tibone, J.E., Itamura, J.M., Kasraeian, S., 2004. Six-month magneticresonance imaging follow-up of large and massive rotator cuff repairs reinforcedwith porcine small intestinal submucosa. J. Shoulder Elbow Surg. 13 5OFF 538.
Sottile, J., 2004. Regulation of angiogenesis by extracellular matrix. Biochim. Biophys.Acta 1654, 13.
Tardieu, M., Gamby, C., Avramoglou, T., Jozefonvicz, J., Barritault, D., 1992. Derivatizeddextrans mimic heparin as stabilizers, potentiators, and protectors of acidic or basicFGF. J. Cell. Physiol. 150, 194–203.
Theofilopoulos, A.N., Dixon, F.J., 1985. Murine models of systemic lupus erythematosus.Adv. Immunol. 37, 260–390.
Tomlinson, A., Ferguson, M.W., 2003. Wound healing: a model of dermal wound repair.Meth. Mol. Biol. 225, 249–260.
von Holst, A., 2007. Tenascin C in stem cell niches: redundant, permissive orinstructive? Cells Tissues Organs 188 (1-2), 170–177 Epub.
Vorotnikova, E., Ivkov, R., Foreman, A., Tries, M., Braunhut, S.J., 2006. Themagnitude andtime-dependence of the apoptotic response of normal and malignant cellssubjected to varying treatments of ionizing and thermal radiation. Int. J. Radiat.Biol. 82, 549–559.
Wang, G., Badylak, S., Braunhut, S.J., Heber-Katz, E., Odelberg, S., Gudas, L., 2010. Theeffects of DNA methyltransferase inhibitors and histone deacetylase inhibitors ondigit regeneration in the mouse. Regenerative Medicine Mar 5 (2), 201–220.
Wierzbicka-Patynowski, I., Schwarzbauer, J., 2003. The ins and outs of fibronectinmatrix assembly. J. Cell Sci. 116, 3269–3276.
Copyright © 2022 FDOKUMEN




















