
Expression, purification and functional characterization of Wnt signaling co-receptors LRP5 and LRP6
8
Please cite this article in press as: Sundararaman B, et al. Expression, purification and functional characterization of AmiA of acetamidase operon of Mycobacterium smegmatis. Microbiol Res (2014), http://dx.doi.org/10.1016/j.micres.2014.02.011 ARTICLE IN PRESS G Model MICRES-25663; No. of Pages 8 Microbiological Research xxx (2014) xxx–xxx Contents lists available at ScienceDirect Microbiological Research j ourna l h om epage: www.elsevier.com/locate/micres Expression, purification and functional characterization of AmiA of acetamidase operon of Mycobacterium smegmatis Balaji Sundararaman 1 , Kannan Palaniyandi 1 , Arunkumar Venkatesan, Sujatha Narayanan ∗ Department of Immunology, National Institute for Research in Tuberculosis, Mayor V.R. Ramanathan Road, Chetput, Chennai 600031, India a r t i c l e i n f o Article history: Received 7 August 2013 Received in revised form 19 December 2013 Accepted 14 February 2014 Available online xxx Keywords: Acetamidase operon Mycobacterium smegmatis AmiA Operator Gene regulation a b s t r a c t Regulation of gene expression is one of the mechanisms of virulence in pathogenic organisms. In this context, we would like to understand the gene regulation of acetamidase enzyme of Mycobacterium smeg- matis, which is the first reported inducible enzyme in mycobacteria. The acetamidase is highly inducible and the expression of this enzyme is increased 100-fold when the substrate acetamide is added. The acetamidase structural gene (amiE) is found immediately downstream of three predicted open reading frames (ORFs). Three of these genes along with a divergently expressed ORF are predicted to form an operon and involved in the regulation of acetamidase enzyme. Here we report expression, purification and functional characterization of AmiA which is one of these predicted ORFs. Electrophoretic mobil- ity shift assays showed that AmiA binds to the region between the amiA and amiD near the predicted promoter (P2). Over-expression of AmiA significantly lowered the expression of acetamidase compared to the wild type as demonstrated by qRT-PCR and SDS-PAGE. We conclude that AmiA binds near P2 promoter and acts as a repressor in the regulation of acetamidase operon. The described work is a fur- ther step forward toward broadening the knowledge on understanding of the complex gene regulatory mechanism of Mycobacterium sp. © 2014 Elsevier GmbH. All rights reserved. 1. Introduction An understanding of the regulation of gene expression is fun- damental to our comprehension of any infection process and can identify potential targets for disease prevention and therapy. A number of central gene regulatory functions govern the life and growth of pathogenic organisms, and these have been studied for decades. The acetamidase enzyme of Mycobacterium smegmatis is a highly inducible enzyme which enables the organism to uti- lize several amides, including acetamide and formamide, as sole carbon source (Draper, 1967). It is expressed at basal level in non- induced conditions or bacteria grown in nutrient rich medium. This enzyme can be induced 100-fold in the presence of simple chemical inducers like acetamide that results in increase of its pro- tein concentration up to 10% of total proteins (Mahenthiralingam et al., 1993; Parish et al., 1997). The study of the expression of gene encoding this enzyme should provide a better understanding of how mycobacterial gene expression is regulated. Inducible gene ∗ Corresponding author. Tel.: +91 44 28369727; fax: +91 44 28362528. E-mail address: [email protected] (S. Narayanan). 1 These authors contributed equally to this study. expression systems that allows turning mycobacterial genes on and off during infection studies are powerful tools to studying the gene function and validation of drug targets. Amidase is expressed by many bacteria including Mycobac- terium tuberculosis, Pseudomonas aeruginosa and fungi. Amidase operon of P. aeruginosa is well characterized and it shares simi- larity with the acetamidase operon of M. smegmatis (Wilson et al., 1996; Norman et al., 2000). Although there are similarities in the DNA/protein sequence and their function, the organization of genes and the regulation of amidase operon in P. aeruginosa is entirely different compared to acetamidase operon of M. smegmatis. In M. smegmatis the genes are organized as amiADS and E and the gene amiC is divergently transcribed (Parish et al., 1997; Narayanan et al., 2000). P. aeruginosa amidase operon has amiEBCRS genes in which amiE, amiC and amiS exhibit a percentage of sequence identity and significant structural similarity to their homologues in M. smegma- tis operon (Parish et al., 1997). The structural gene, acetamidase (amiE) along with 1.5 kb upstream to amiE region was sequenced and predicted to have three open reading frames (ORFs) named amiA, amiD and amiS (Mahenthiralingam et al., 1993). Parish et al., 1997 further sequenced 1.5 kb upstream to amiA and showed the presence of a divergently transcribed gene, amiC (Fig. 1). These predicted ORFs http://dx.doi.org/10.1016/j.micres.2014.02.011 0944-5013/© 2014 Elsevier GmbH. All rights reserved.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Expression, purification and functional characterization of Wnt signaling co-receptors LRP5 and LRP6

M
Ea
BSD
a
ARR1AA
KAMAOG
1
dingdalciTcteeh
h0
ARTICLE IN PRESSG ModelICRES-25663; No. of Pages 8
Microbiological Research xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
Microbiological Research
j ourna l h om epage: www.elsev ier .com/ locate /micres
xpression, purification and functional characterization of AmiA ofcetamidase operon of Mycobacterium smegmatis
alaji Sundararaman1, Kannan Palaniyandi1, Arunkumar Venkatesan,ujatha Narayanan ∗
epartment of Immunology, National Institute for Research in Tuberculosis, Mayor V.R. Ramanathan Road, Chetput, Chennai 600031, India
r t i c l e i n f o
rticle history:eceived 7 August 2013eceived in revised form9 December 2013ccepted 14 February 2014vailable online xxx
eywords:cetamidase operonycobacterium smegmatis
a b s t r a c t
Regulation of gene expression is one of the mechanisms of virulence in pathogenic organisms. In thiscontext, we would like to understand the gene regulation of acetamidase enzyme of Mycobacterium smeg-matis, which is the first reported inducible enzyme in mycobacteria. The acetamidase is highly inducibleand the expression of this enzyme is increased 100-fold when the substrate acetamide is added. Theacetamidase structural gene (amiE) is found immediately downstream of three predicted open readingframes (ORFs). Three of these genes along with a divergently expressed ORF are predicted to form anoperon and involved in the regulation of acetamidase enzyme. Here we report expression, purificationand functional characterization of AmiA which is one of these predicted ORFs. Electrophoretic mobil-ity shift assays showed that AmiA binds to the region between the amiA and amiD near the predicted
miAperatorene regulation
promoter (P2). Over-expression of AmiA significantly lowered the expression of acetamidase comparedto the wild type as demonstrated by qRT-PCR and SDS-PAGE. We conclude that AmiA binds near P2promoter and acts as a repressor in the regulation of acetamidase operon. The described work is a fur-ther step forward toward broadening the knowledge on understanding of the complex gene regulatorymechanism of Mycobacterium sp.
. Introduction
An understanding of the regulation of gene expression is fun-amental to our comprehension of any infection process and can
dentify potential targets for disease prevention and therapy. Aumber of central gene regulatory functions govern the life androwth of pathogenic organisms, and these have been studied forecades. The acetamidase enzyme of Mycobacterium smegmatis is
highly inducible enzyme which enables the organism to uti-ize several amides, including acetamide and formamide, as solearbon source (Draper, 1967). It is expressed at basal level in non-nduced conditions or bacteria grown in nutrient rich medium.his enzyme can be induced 100-fold in the presence of simplehemical inducers like acetamide that results in increase of its pro-ein concentration up to 10% of total proteins (Mahenthiralingam
Please cite this article in press as: Sundararaman B, et al. Expression, puoperon of Mycobacterium smegmatis. Microbiol Res (2014), http://dx.d
t al., 1993; Parish et al., 1997). The study of the expression of genencoding this enzyme should provide a better understanding ofow mycobacterial gene expression is regulated. Inducible gene
∗ Corresponding author. Tel.: +91 44 28369727; fax: +91 44 28362528.E-mail address: [email protected] (S. Narayanan).
1 These authors contributed equally to this study.
ttp://dx.doi.org/10.1016/j.micres.2014.02.011944-5013/© 2014 Elsevier GmbH. All rights reserved.
© 2014 Elsevier GmbH. All rights reserved.
expression systems that allows turning mycobacterial genes on andoff during infection studies are powerful tools to studying the genefunction and validation of drug targets.
Amidase is expressed by many bacteria including Mycobac-terium tuberculosis, Pseudomonas aeruginosa and fungi. Amidaseoperon of P. aeruginosa is well characterized and it shares simi-larity with the acetamidase operon of M. smegmatis (Wilson et al.,1996; Norman et al., 2000). Although there are similarities in theDNA/protein sequence and their function, the organization of genesand the regulation of amidase operon in P. aeruginosa is entirelydifferent compared to acetamidase operon of M. smegmatis. In M.smegmatis the genes are organized as amiADS and E and the geneamiC is divergently transcribed (Parish et al., 1997; Narayanan et al.,2000). P. aeruginosa amidase operon has amiEBCRS genes in whichamiE, amiC and amiS exhibit a percentage of sequence identity andsignificant structural similarity to their homologues in M. smegma-tis operon (Parish et al., 1997).
The structural gene, acetamidase (amiE) along with 1.5 kbupstream to amiE region was sequenced and predicted to have
rification and functional characterization of AmiA of acetamidaseoi.org/10.1016/j.micres.2014.02.011
three open reading frames (ORFs) named amiA, amiD and amiS(Mahenthiralingam et al., 1993). Parish et al., 1997 furthersequenced 1.5 kb upstream to amiA and showed the presence of adivergently transcribed gene, amiC (Fig. 1). These predicted ORFs

ARTICLE IN PRESSG ModelMICRES-25663; No. of Pages 8
2 B. Sundararaman et al. / Microbiological Research xxx (2014) xxx–xxx
F etamid miD ag nscrip
etCmcopocpbaipPtrd
tes
sopwi
2
2
flwe0mM8W
TP
ig. 1. Picture depicts the order of genes and positions of the promoters in the acirection of transcription. amiA negative regulator of amidase operon. amiC and aene. Promoter regions are indicated by bars with arrows indicating direction of tra
xhibit considerable homology with the regulatory and amideransporter proteins of other bacteria (Wilson and Drew, 1995;hebrou et al., 1996). Previously, we had shown the presence ofultiple transcripts of 3.0, 1.8 and 1.2 kb and mapped two trans-
riptional start sites at 1868 and 51 bp upstream to the start codonf acetamidase and hypothesized the involvement of either RNArocessing or multiple promoters in the regulation of acetamidaseperon (Narayanan et al., 2000). Subsequently, by promoter probeloning and �-galactosidase assay, Parish et al., 2001 showed theresence of three promoters (Fig. 1). The first promoter is locatedetween amiC and amiA (P1), the second is flanked by amiA andmiD (P2), which is amide inducible and the third one is presentmmediately upstream to amiE (P3). They have also identified aromoter oriented in opposite direction to other promoters, calledc which expresses amiC. Studies with AmiA deletion mutants illus-rated the negative role played by amiA and knockout of this geneesulted in amide independent, constitutive expression of acetami-ase (Parish et al., 2001).
Further knockouts of other genes revealed the presence of mul-iple regulatory elements in acetamidase expression (Raghunandt al., 2006). By multiple gene knockout strategy, the authorshowed that amiC and amiD act as positive regulators.
The DNA binding activities of these regulatory proteins are nottudied before to identify the operator regions within this complexperon. Hence our lab was interested to study one of regulatoryrotein AmiA and its operator regions in the operon. In this study,e show for the first time the recombinant expression of AmiA and
t’s binding in P2 promoter region.
. Materials and methods
.1. Bacterial strains and media
Escherichia coli DH5� and E. coli BL21 (DE3) were obtainedrom Novagen. E. coli strains were grown in Luria Bertani (LB)iquid and solid (with 1.5% agar) medium at 37 ◦C. Kanamycin
as supplemented at 40 �g/ml. M. smegmatis mc2 155 was grownither in LB or Kohn–Haris minimal medium (NaCl 4 g, MgSO4.2 g, KH2PO4 2 g, (NH4)3PO4 2 g/l) containing 5 ml of trace ele-
Please cite this article in press as: Sundararaman B, et al. Expression, puoperon of Mycobacterium smegmatis. Microbiol Res (2014), http://dx.d
ents (ZnCl2 40 mg, Fe(NH4)2(SO4)·6H2O 200 mg, Cu2SO4 10 mg,nCl2 10 mg, Borax 10 mg, (NH4)2MoO4 10 mg/l) with 0.05% Tween
0 at 37 ◦C with shaking (Kohn and Harris, 1941; Hopwood andright, 1978). Kanamycin was supplemented whenever needed
able 1lasmids constructed in this study.
Plasmid name Description
pET30 E. coli expression vector, Kanr, Lac promoter
pCR2.1 TOPO cloning vector, Kanr, Ampr,
pMV261 E. coli mycobacterial shuttle vector, Kanr, hsppETOrf1 Coding region of AmiA in pET30(c)
pMVOrf1 Coding region of AmiA in E. coli MycobacteriapMVOrf2 Coding region of AmiD in E. coli MycobacteriapMVOrfC Coding region of AmiC in E. coli MycobacteriapCML 1776–1975 bp fragment in pCR2.1 vector
dase operon. ORFs – are indicated by thick arrows and thin arrows depicting there proposed regulators. amiS is putative ABC transporter. amiE is the acetamidasetion.
at 20 �g/ml. Inducer (acetamide) was added whenever needed atappropriate concentrations as indicated elsewhere.
2.2. Plasmids and DNA manipulation
PCR product of 553 bp (coding region of amiA) was cloned usingspecific primers (Supplementary Table S1) with restriction enzymesite overhangs into the BamHI-EcoRI site of pET30(c) and in E. colimycobacterial shuttle vector pMV261, and transformed into E. coliDH5� (Table 1). The resulting constructs were sequenced to con-firm the fidelity of the sequence.
2.3. Expression and purification of AmiA
E. coli BL21 (DE3) harboring pETorf1 (amiA cloned into pET30(c))was grown in LB medium containing 40 �g/ml of kanamycin at37 ◦C to 0.6 OD at 600 nm and induced with 1 mM IPTG and contin-ued to grow for further 2 h at 37 ◦C. The cells were placed in ice for15 min, pelleted down by centrifugation at 6000 × g for 15 min andstored at −80 ◦C. The cells were thawed on ice, suspended in bufferA (50 mM NaH2PO4 pH 8.0, 300 mM NaCl, 10 mM imidazole, 20%glycerol, and 0.5 mM phenyl methyl sulfonyl fluoride) and sonica-ted on ice using SoniPrep 150 for 1 min with an interval of 5 minon ice for seven cycles. The lysate was centrifuged at 15000 × g for15 min at 4 ◦C. The crude lysate was bound with ProBond Nickelaffinity resin (Invitrogen) packed column by end-to-end shaking at8 ◦C for 1 h. Column was washed with 60 column volumes of bufferB (50 mM NaH2PO4 pH 8.0, 300 mM NaCl, 40 mM imidazole, 20%glycerol, and 0.5 mM phenyl methyl sulfonyl fluoride) and recom-binant AmiA was eluted with buffer C (50 mM NaH2PO4 pH 8.0,300 mM NaCl, 200 mM imidazole, 20% glycerol, and 0.5 mM phenylmethyl sulfonyl fluoride). The resulting fractions were pooled anddialyzed against 1 l of 0.5× buffer A, concentrated on sucrose andstored at −80 ◦C.
2.4. Glutaraldehyde cross-linking of AmiA
Glutaraldehyde (1.5%) was added to AmiA protein in 0.5× bufferA to a final concentration of 0.05%, incubated at 37 ◦C for 15 minand cross-linking reaction was quenched immediately by addition
rification and functional characterization of AmiA of acetamidaseoi.org/10.1016/j.micres.2014.02.011
of 6X SDS dye. The cross-linked products were analyzed in 12%SDS PAGE and visualized by Coomassie staining. The cross-linkedproducts were subjected to western blotting and identified withanti-His antibody (Novagen).
Reference/source
NovagenInvitrogen
60 promoter Stover et al., 1991In this study
l shuttle vector pMV261 In this studyl shuttle vector pMV261 In this studyl shuttle vector pMV261 In this study
In this study

IN PRESSG ModelM
iological Research xxx (2014) xxx–xxx 3
2
dducosp1Tbg1leEa(cpig
2
pcpaamwrtao2ua
2
w8wamCatat−
2
awbNP
Table 2Primers used for quantitative real-time PCR (qRT-PCR).
Primer name Sequence (5′–3′) Use
Ace F ACCAACCCCATCTTCATGC qRT-PCRAce R TGTAGGCGTTGCGATTCTC qRT-PCRAce Probe AACGTGAGCCATTCGGAGTAGAGC qRT-PCR16s rRNA F GGGAGCGAACAGGATTAGATAC qRT-PCR16s rRNA R CCTTTGAGTTTTAGCCTTGCG qRT-PCR16s rRNA Probe CGGTGGGTACTAGGTGTGGGTTTC qRT-PCRAmiA F GGTCCGACTCTCACCAGAAC qRT-PCRAmiA R CAGTCCGGTAGATGATCCCT qRT-PCRAmiA Probe CGTCGATCGCCTTGTGTCGC qRT-PCRAmiD F CCGACCTACACATTCCGTTG qRT-PCRAmiD R GTGAATGTCGTCAGCCCTAC qRT-PCRAmiD Probe TTCGATCTCACCTGCGCGATCTC qRT-PCRAmiC F TAACAGGATTCGGCCATCG qRT-PCR
ARTICLEICRES-25663; No. of Pages 8
B. Sundararaman et al. / Microb
.5. Electrophoretic mobility shift assay (EMSA)
Primers were designed to amplify approximately 300 bp byesigning overlapping primers for every 150 bp of the acetami-ase operon (Supplementary Table S1) and labeled with [�32P]dCTPsing RediPrimeII random primer labeling kit (Amersham Pharma-ia) as per kit protocol. Binding reaction was set-up with 1.2 ngf labeled DNA in 20 �l reaction mix containing 50 �g/ml doubletrand poly dI:dC, 50 �g/ml BSA, 1× binding buffer (25 mM HEPES,H 7.5, 0.1 mM ZnCl2, 1 mM DTT, 40 mM KCl and 5% glycerol) and0% glycerol and incubated at room temperature (25–27 ◦C) for 1 h.his protein–DNA complex was resolved at 150 V in 0.25× Tris-orate-EDTA (TBE) for 4 h on 6% poly acrylamide gel cast with 2.5%lycerol in 0.5× Tris-acetate-EDTA (TAE). The gel was pre run at00 V for 30 min; wells were thoroughly washed with buffer before
oading the samples. After that run gel was dried at 80 ◦C for 10 min,xposed to photographic films and developed by autoradiography.MSA experiments were also titrated with different DNA, proteinnd acetamide concentrations. Double-stranded oligonucleotidesATTTCGGGTGAATGGAAAGGGGTGACCATG) and mutated oligonu-leotides (ATTTCGCCTGAATGGAAAGGGTTGACCATG) were pre-ared by mixing complementary single-stranded DNA oligomers
n Tris-EDTA buffer, heating at 95 ◦C for 15 min and annealed byradually cooling to 20 ◦C.
.6. Foot printing analysis
The fragment which gave the band shifts in the EMSA, com-rising 1776–1975 bp (Supplementary Table S1) of the operon wasloned into pCR2.1 vector using TOPO Cloning kit (Invitrogen). Thelasmid was cut with BamHI-XbaI, the fragment was gel purified,nd end filled by Klenow fragment, dephosphorylated with CIAPnd then ends labeled with [�-32P] ATP by T4 PNK. Labeled frag-ent was cut with XhoI to ensure only one end was labeled. Probeas bound with 200 and 300 nM of purified AmiA using binding
eaction conditions similar to EMSA. Protein DNA complex wasreated with DNase I (0.15 U) as per kit protocol, phenol extracted,nd ethanol precipitated and washed. Pellet was suspended in 8 �lf loading dye and 2 �l of 1 M urea, vortexed, heated to 95 ◦C for
min, snap cooled on ice and loaded on 8% sequencing gel (7 Mrea, 40% formamide, 1× TBE) along with DNA sequencing ladders per the kit protocol (USB PCR product sequencing kit).
.7. In vivo over-expression studies
Wild type and recombinant M. smegmatis were transformedith the appropriate plasmids cultured in Luria Bertani Tween-
0 (LBT) media supplemented with 20 �g/ml of kanamycin andere grown at 37 ◦C on shaking for 48 h. Cells were pelleted down
t 4000 × g for 10 min, washed and suspended in Kohn-Harrisedium with 20 �g/ml of kanamycin and grown at 37 ◦C overnight.
ells were exposed to heat shock for induction of hsp60 promotert 45 ◦C for 2 h and continued to grow further at 37 ◦C for 3 h andhen induced with various concentration of acetamide as indicatednd allowed to grow at 37 ◦C. The culture was harvested by cen-rifugation at 6800 × g for 15 min and the pellet was preserved at20 ◦C for further use.
.8. RNA isolation and qRT-PCR
Total RNA was isolated from the over-expressed cultures usingn RNeasy kit (QIAGEN, Inc.). The RNA was subsequently treated
Please cite this article in press as: Sundararaman B, et al. Expression, puoperon of Mycobacterium smegmatis. Microbiol Res (2014), http://dx.d
ith DNaseI at 37 ◦C for 45 min. The DNase was then inactivatedy incubation at 75 ◦C for 10 min. RNA was quantified by using aD-1000 Nanodrop spectrophotometer (Nanodrop Technologies).urified RNA was stored at −80 ◦C. For the determination of relative
AmiC R CTCCACCGAAAACCTCTATGTC qRT-PCRAmiC Probe CGTAGCTGCCCATCAGGTCCAT qRT-PCR
The probes were labeled with 5′ FAM and 3′ TAMARA.
mRNA concentrations by quantitative reverse transcription-PCR(qRT-PCR), cDNA was synthesized with 2 �g of RNA using Super-script III first strand synthesis system with random hexamers(Invitrogen), according to the manufacturer’s instructions. qRT-PCRs were carried out using Taqman PCR master mix (AppliedBiosystems) according to manufacture’s instruction using theApplied Biosystems 7300 real-time PCR system (Applied Biosys-tems). The qRT-PCR primers used in this study were listed in Table 2.To check for DNA contamination, control reactions for each samplewere carried out in the absence of reverse transcriptase. The ampli-fication conditions for all reactions were 1 cycle of 50 ◦C for 2 min,95 ◦C for 10 min, followed by 40 cycles of 95 ◦C for 15 s and 60 ◦Cfor 1 min.
Analysis of qRT-PCR data were carried out using the compar-ative CT method. For each qRT-PCR run, the calculated thresholdcycle (CT) was normalized to the CT of the internal control 16SrRNA gene amplified from the corresponding sample. Statisticalanalysis was carried out using GraphPad Prism software. The datapresented are averages of three independent experiments and errorbars represent standard deviations.
2.9. SDS-PAGE and protein densitometry
Pellet generated from 30 ml of culture of over-expressed cellswere lysed in 3 ml of lysis buffer (50 mM Tris-HCl, pH 7.6,10 mM MgCl2, 60 mM NaCl, 10% glycerol, 1 mM PMSF, 10 mM �-mercaptoethanol) by sonication for 5 min with an interval of 5 minon ice for a total of seven cycles. The cell-free lysate was collectedby centrifugation at 15,000 × g, 4 ◦C for 15 min and stored in −80 ◦C.Protein concentration was determined by BCA kit (Pierce) as per thesupplier’s protocol. Cell-free lysates with equal protein concentra-tion were resolved in 18.3 cm × 20 cm 10% SDS-PAGE at 30 V for 6 h,stained with Coomassie blue and gels (triplicate) were subjected todensitometer scanning and acetamidase concentration was deter-mined by measuring the intensity of 47 kDa band using QuantityOne software (Bio-Rad).
2.10. Estimation of amidase activity
To estimate intracellular amidase production, the acetamideinduced cells were harvested and pelleted. The cells were resus-pended in 20 mM PO4 buffer (pH 7.4) and lysozyme was addedto the cells (2 mg/ml) and stirred gently for 1 h at 4 ◦C. The cell-free lysates were collected by centrifugation at 20,000 × g, 4 ◦C
rification and functional characterization of AmiA of acetamidaseoi.org/10.1016/j.micres.2014.02.011
for 20 min. This clear supernatant was used to estimate amidaseactivity after incubation with acetamide (100 mM). The intracel-lular acetamidase activity and release of ammonia was estimatedby Nesselerization (Imada et al., 1973). All the experiments were
ARTICLE IN PRESSG ModelMICRES-25663; No. of Pages 8
4 B. Sundararaman et al. / Microbiological Research xxx (2014) xxx–xxx
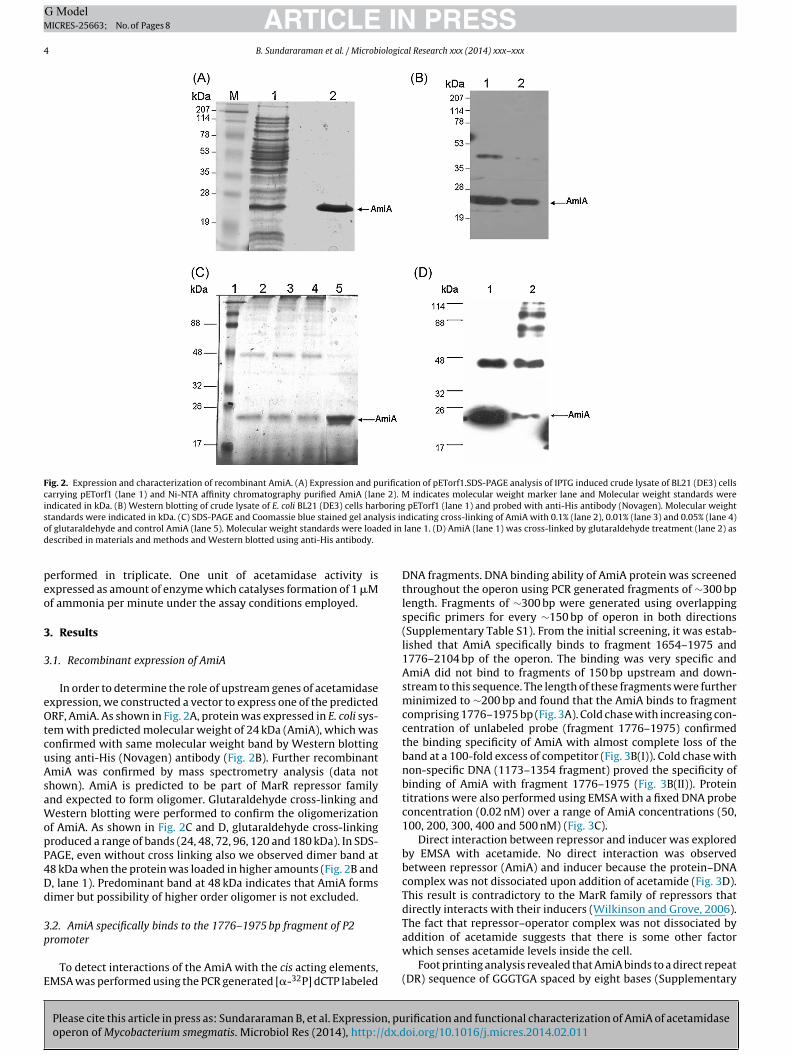
Fig. 2. Expression and characterization of recombinant AmiA. (A) Expression and purification of pETorf1.SDS-PAGE analysis of IPTG induced crude lysate of BL21 (DE3) cellscarrying pETorf1 (lane 1) and Ni-NTA affinity chromatography purified AmiA (lane 2). M indicates molecular weight marker lane and Molecular weight standards wereindicated in kDa. (B) Western blotting of crude lysate of E. coli BL21 (DE3) cells harboring pETorf1 (lane 1) and probed with anti-His antibody (Novagen). Molecular weights lysis ino ed ind
peo
3
3
eOtcuAsaWopP4Dd
3p
E
tandards were indicated in kDa. (C) SDS-PAGE and Coomassie blue stained gel anaf glutaraldehyde and control AmiA (lane 5). Molecular weight standards were loadescribed in materials and methods and Western blotted using anti-His antibody.
erformed in triplicate. One unit of acetamidase activity isxpressed as amount of enzyme which catalyses formation of 1 �Mf ammonia per minute under the assay conditions employed.
. Results
.1. Recombinant expression of AmiA
In order to determine the role of upstream genes of acetamidasexpression, we constructed a vector to express one of the predictedRF, AmiA. As shown in Fig. 2A, protein was expressed in E. coli sys-
em with predicted molecular weight of 24 kDa (AmiA), which wasonfirmed with same molecular weight band by Western blottingsing anti-His (Novagen) antibody (Fig. 2B). Further recombinantmiA was confirmed by mass spectrometry analysis (data nothown). AmiA is predicted to be part of MarR repressor familynd expected to form oligomer. Glutaraldehyde cross-linking andestern blotting were performed to confirm the oligomerization
f AmiA. As shown in Fig. 2C and D, glutaraldehyde cross-linkingroduced a range of bands (24, 48, 72, 96, 120 and 180 kDa). In SDS-AGE, even without cross linking also we observed dimer band at8 kDa when the protein was loaded in higher amounts (Fig. 2B and, lane 1). Predominant band at 48 kDa indicates that AmiA formsimer but possibility of higher order oligomer is not excluded.
.2. AmiA specifically binds to the 1776–1975 bp fragment of P2
Please cite this article in press as: Sundararaman B, et al. Expression, puoperon of Mycobacterium smegmatis. Microbiol Res (2014), http://dx.d
romoter
To detect interactions of the AmiA with the cis acting elements,MSA was performed using the PCR generated [�-32P] dCTP labeled
dicating cross-linking of AmiA with 0.1% (lane 2), 0.01% (lane 3) and 0.05% (lane 4) lane 1. (D) AmiA (lane 1) was cross-linked by glutaraldehyde treatment (lane 2) as
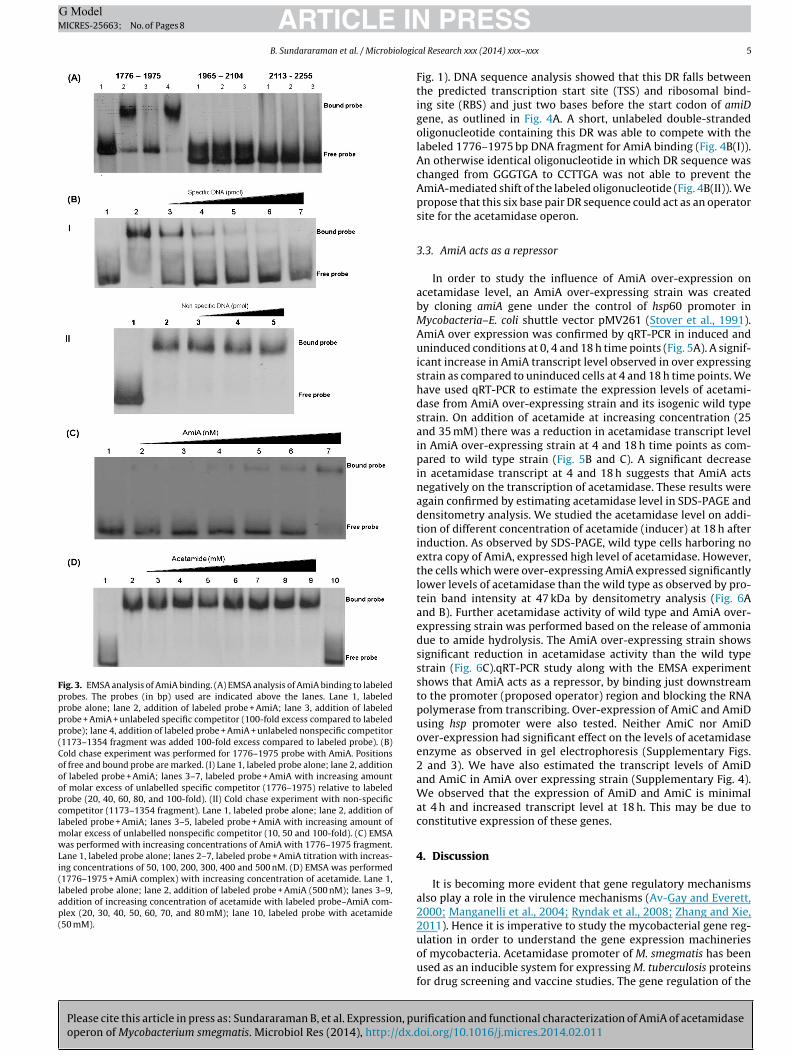
DNA fragments. DNA binding ability of AmiA protein was screenedthroughout the operon using PCR generated fragments of ∼300 bplength. Fragments of ∼300 bp were generated using overlappingspecific primers for every ∼150 bp of operon in both directions(Supplementary Table S1). From the initial screening, it was estab-lished that AmiA specifically binds to fragment 1654–1975 and1776–2104 bp of the operon. The binding was very specific andAmiA did not bind to fragments of 150 bp upstream and down-stream to this sequence. The length of these fragments were furtherminimized to ∼200 bp and found that the AmiA binds to fragmentcomprising 1776–1975 bp (Fig. 3A). Cold chase with increasing con-centration of unlabeled probe (fragment 1776–1975) confirmedthe binding specificity of AmiA with almost complete loss of theband at a 100-fold excess of competitor (Fig. 3B(I)). Cold chase withnon-specific DNA (1173–1354 fragment) proved the specificity ofbinding of AmiA with fragment 1776–1975 (Fig. 3B(II)). Proteintitrations were also performed using EMSA with a fixed DNA probeconcentration (0.02 nM) over a range of AmiA concentrations (50,100, 200, 300, 400 and 500 nM) (Fig. 3C).
Direct interaction between repressor and inducer was exploredby EMSA with acetamide. No direct interaction was observedbetween repressor (AmiA) and inducer because the protein–DNAcomplex was not dissociated upon addition of acetamide (Fig. 3D).This result is contradictory to the MarR family of repressors thatdirectly interacts with their inducers (Wilkinson and Grove, 2006).The fact that repressor–operator complex was not dissociated by
rification and functional characterization of AmiA of acetamidaseoi.org/10.1016/j.micres.2014.02.011
addition of acetamide suggests that there is some other factorwhich senses acetamide levels inside the cell.
Foot printing analysis revealed that AmiA binds to a direct repeat(DR) sequence of GGGTGA spaced by eight bases (Supplementary
Please cite this article in press as: Sundararaman B, et al. Expression, puoperon of Mycobacterium smegmatis. Microbiol Res (2014), http://dx.d
ARTICLE ING ModelMICRES-25663; No. of Pages 8
B. Sundararaman et al. / Microbiologic
Fig. 3. EMSA analysis of AmiA binding. (A) EMSA analysis of AmiA binding to labeledprobes. The probes (in bp) used are indicated above the lanes. Lane 1, labeledprobe alone; lane 2, addition of labeled probe + AmiA; lane 3, addition of labeledprobe + AmiA + unlabeled specific competitor (100-fold excess compared to labeledprobe); lane 4, addition of labeled probe + AmiA + unlabeled nonspecific competitor(1173–1354 fragment was added 100-fold excess compared to labeled probe). (B)Cold chase experiment was performed for 1776–1975 probe with AmiA. Positionsof free and bound probe are marked. (I) Lane 1, labeled probe alone; lane 2, additionof labeled probe + AmiA; lanes 3–7, labeled probe + AmiA with increasing amountof molar excess of unlabelled specific competitor (1776–1975) relative to labeledprobe (20, 40, 60, 80, and 100-fold). (II) Cold chase experiment with non-specificcompetitor (1173–1354 fragment). Lane 1, labeled probe alone; lane 2, addition oflabeled probe + AmiA; lanes 3–5, labeled probe + AmiA with increasing amount ofmolar excess of unlabelled nonspecific competitor (10, 50 and 100-fold). (C) EMSAwas performed with increasing concentrations of AmiA with 1776–1975 fragment.Lane 1, labeled probe alone; lanes 2–7, labeled probe + AmiA titration with increas-ing concentrations of 50, 100, 200, 300, 400 and 500 nM. (D) EMSA was performed(1776–1975 + AmiA complex) with increasing concentration of acetamide. Lane 1,labeled probe alone; lane 2, addition of labeled probe + AmiA (500 nM); lanes 3–9,addition of increasing concentration of acetamide with labeled probe–AmiA com-plex (20, 30, 40, 50, 60, 70, and 80 mM); lane 10, labeled probe with acetamide(50 mM).
PRESSal Research xxx (2014) xxx–xxx 5
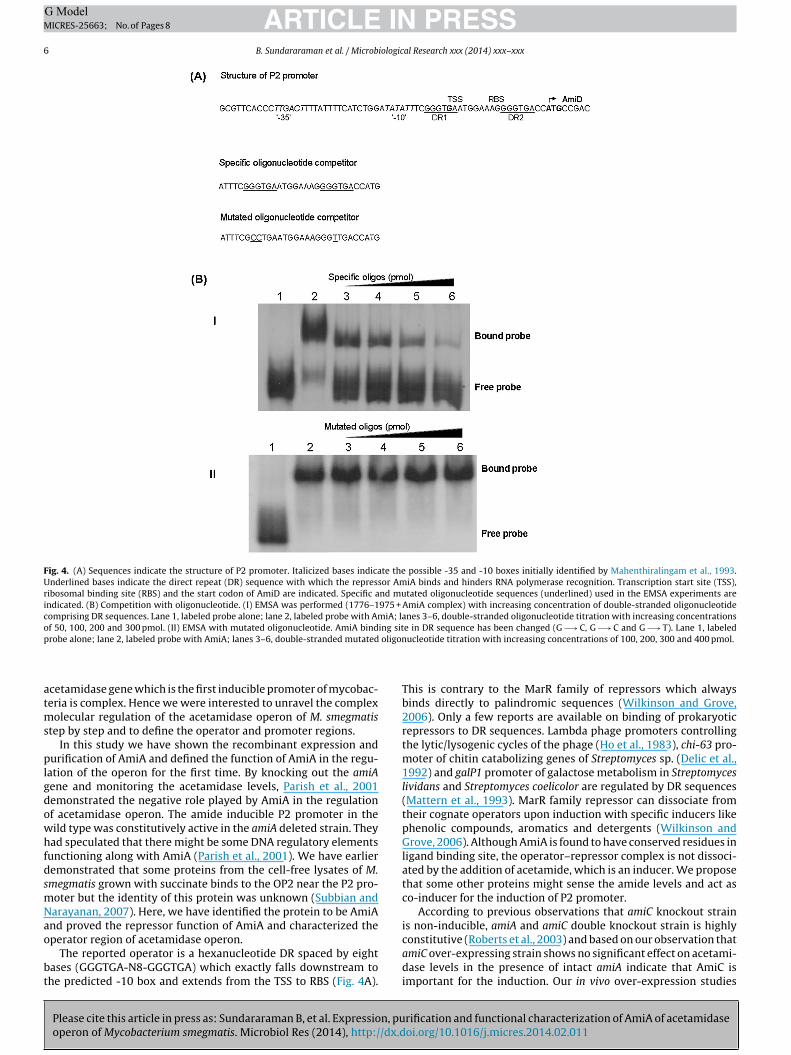
Fig. 1). DNA sequence analysis showed that this DR falls betweenthe predicted transcription start site (TSS) and ribosomal bind-ing site (RBS) and just two bases before the start codon of amiDgene, as outlined in Fig. 4A. A short, unlabeled double-strandedoligonucleotide containing this DR was able to compete with thelabeled 1776–1975 bp DNA fragment for AmiA binding (Fig. 4B(I)).An otherwise identical oligonucleotide in which DR sequence waschanged from GGGTGA to CCTTGA was not able to prevent theAmiA-mediated shift of the labeled oligonucleotide (Fig. 4B(II)). Wepropose that this six base pair DR sequence could act as an operatorsite for the acetamidase operon.
3.3. AmiA acts as a repressor
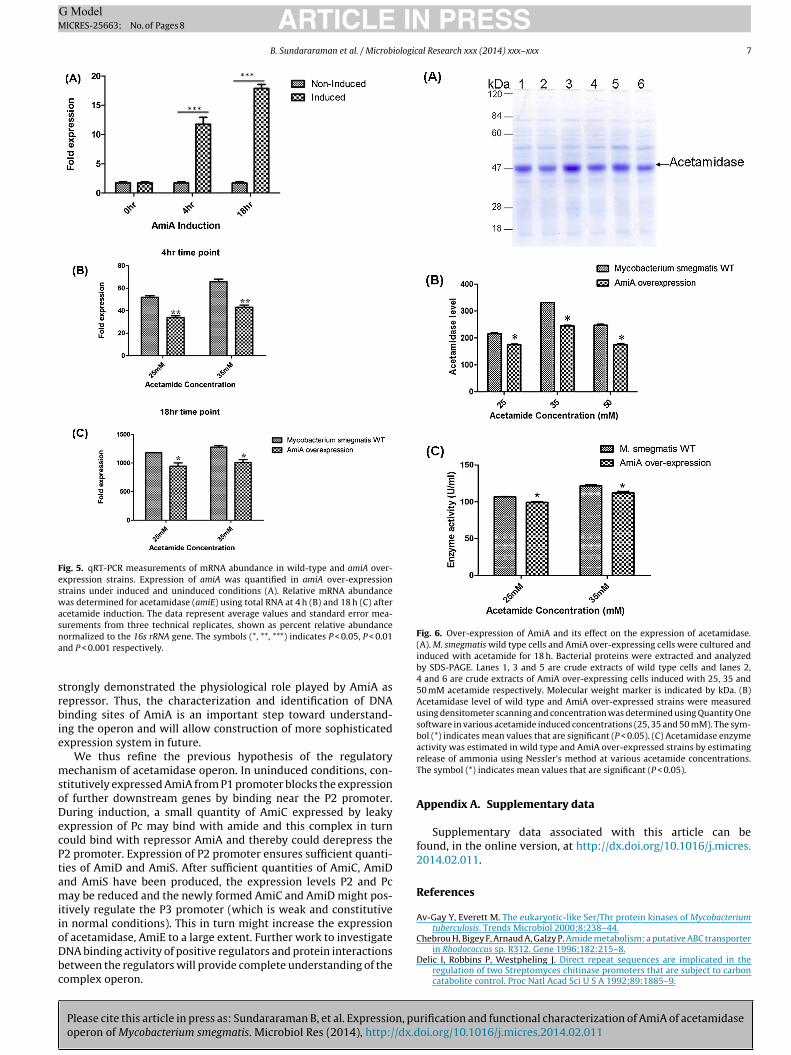
In order to study the influence of AmiA over-expression onacetamidase level, an AmiA over-expressing strain was createdby cloning amiA gene under the control of hsp60 promoter inMycobacteria–E. coli shuttle vector pMV261 (Stover et al., 1991).AmiA over expression was confirmed by qRT-PCR in induced anduninduced conditions at 0, 4 and 18 h time points (Fig. 5A). A signif-icant increase in AmiA transcript level observed in over expressingstrain as compared to uninduced cells at 4 and 18 h time points. Wehave used qRT-PCR to estimate the expression levels of acetami-dase from AmiA over-expressing strain and its isogenic wild typestrain. On addition of acetamide at increasing concentration (25and 35 mM) there was a reduction in acetamidase transcript levelin AmiA over-expressing strain at 4 and 18 h time points as com-pared to wild type strain (Fig. 5B and C). A significant decreasein acetamidase transcript at 4 and 18 h suggests that AmiA actsnegatively on the transcription of acetamidase. These results wereagain confirmed by estimating acetamidase level in SDS-PAGE anddensitometry analysis. We studied the acetamidase level on addi-tion of different concentration of acetamide (inducer) at 18 h afterinduction. As observed by SDS-PAGE, wild type cells harboring noextra copy of AmiA, expressed high level of acetamidase. However,the cells which were over-expressing AmiA expressed significantlylower levels of acetamidase than the wild type as observed by pro-tein band intensity at 47 kDa by densitometry analysis (Fig. 6Aand B). Further acetamidase activity of wild type and AmiA over-expressing strain was performed based on the release of ammoniadue to amide hydrolysis. The AmiA over-expressing strain showssignificant reduction in acetamidase activity than the wild typestrain (Fig. 6C).qRT-PCR study along with the EMSA experimentshows that AmiA acts as a repressor, by binding just downstreamto the promoter (proposed operator) region and blocking the RNApolymerase from transcribing. Over-expression of AmiC and AmiDusing hsp promoter were also tested. Neither AmiC nor AmiDover-expression had significant effect on the levels of acetamidaseenzyme as observed in gel electrophoresis (Supplementary Figs.2 and 3). We have also estimated the transcript levels of AmiDand AmiC in AmiA over expressing strain (Supplementary Fig. 4).We observed that the expression of AmiD and AmiC is minimalat 4 h and increased transcript level at 18 h. This may be due toconstitutive expression of these genes.
4. Discussion
It is becoming more evident that gene regulatory mechanismsalso play a role in the virulence mechanisms (Av-Gay and Everett,2000; Manganelli et al., 2004; Ryndak et al., 2008; Zhang and Xie,2011). Hence it is imperative to study the mycobacterial gene reg-
rification and functional characterization of AmiA of acetamidaseoi.org/10.1016/j.micres.2014.02.011
ulation in order to understand the gene expression machineriesof mycobacteria. Acetamidase promoter of M. smegmatis has beenused as an inducible system for expressing M. tuberculosis proteinsfor drug screening and vaccine studies. The gene regulation of the
ARTICLE IN PRESSG ModelMICRES-25663; No. of Pages 8
6 B. Sundararaman et al. / Microbiological Research xxx (2014) xxx–xxx
Fig. 4. (A) Sequences indicate the structure of P2 promoter. Italicized bases indicate the possible -35 and -10 boxes initially identified by Mahenthiralingam et al., 1993.Underlined bases indicate the direct repeat (DR) sequence with which the repressor AmiA binds and hinders RNA polymerase recognition. Transcription start site (TSS),ribosomal binding site (RBS) and the start codon of AmiD are indicated. Specific and mutated oligonucleotide sequences (underlined) used in the EMSA experiments areindicated. (B) Competition with oligonucleotide. (I) EMSA was performed (1776–1975 + AmiA complex) with increasing concentration of double-stranded oligonucleotidecomprising DR sequences. Lane 1, labeled probe alone; lane 2, labeled probe with AmiA; lanes 3–6, double-stranded oligonucleotide titration with increasing concentrationsof 50, 100, 200 and 300 pmol. (II) EMSA with mutated oligonucleotide. AmiA binding site in DR sequence has been changed (G −→ C, G −→ C and G −→ T). Lane 1, labeledp oligon
atms
plgdowhfdsmNao
bt
robe alone; lane 2, labeled probe with AmiA; lanes 3–6, double-stranded mutated
cetamidase gene which is the first inducible promoter of mycobac-eria is complex. Hence we were interested to unravel the complex
olecular regulation of the acetamidase operon of M. smegmatistep by step and to define the operator and promoter regions.
In this study we have shown the recombinant expression andurification of AmiA and defined the function of AmiA in the regu-
ation of the operon for the first time. By knocking out the amiAene and monitoring the acetamidase levels, Parish et al., 2001emonstrated the negative role played by AmiA in the regulationf acetamidase operon. The amide inducible P2 promoter in theild type was constitutively active in the amiA deleted strain. Theyad speculated that there might be some DNA regulatory elements
unctioning along with AmiA (Parish et al., 2001). We have earlieremonstrated that some proteins from the cell-free lysates of M.megmatis grown with succinate binds to the OP2 near the P2 pro-oter but the identity of this protein was unknown (Subbian andarayanan, 2007). Here, we have identified the protein to be AmiAnd proved the repressor function of AmiA and characterized the
Please cite this article in press as: Sundararaman B, et al. Expression, puoperon of Mycobacterium smegmatis. Microbiol Res (2014), http://dx.d
perator region of acetamidase operon.The reported operator is a hexanucleotide DR spaced by eight
ases (GGGTGA-N8-GGGTGA) which exactly falls downstream tohe predicted -10 box and extends from the TSS to RBS (Fig. 4A).
ucleotide titration with increasing concentrations of 100, 200, 300 and 400 pmol.
This is contrary to the MarR family of repressors which alwaysbinds directly to palindromic sequences (Wilkinson and Grove,2006). Only a few reports are available on binding of prokaryoticrepressors to DR sequences. Lambda phage promoters controllingthe lytic/lysogenic cycles of the phage (Ho et al., 1983), chi-63 pro-moter of chitin catabolizing genes of Streptomyces sp. (Delic et al.,1992) and galP1 promoter of galactose metabolism in Streptomyceslividans and Streptomyces coelicolor are regulated by DR sequences(Mattern et al., 1993). MarR family repressor can dissociate fromtheir cognate operators upon induction with specific inducers likephenolic compounds, aromatics and detergents (Wilkinson andGrove, 2006). Although AmiA is found to have conserved residues inligand binding site, the operator–repressor complex is not dissoci-ated by the addition of acetamide, which is an inducer. We proposethat some other proteins might sense the amide levels and act asco-inducer for the induction of P2 promoter.
According to previous observations that amiC knockout strainis non-inducible, amiA and amiC double knockout strain is highly
rification and functional characterization of AmiA of acetamidaseoi.org/10.1016/j.micres.2014.02.011
constitutive (Roberts et al., 2003) and based on our observation thatamiC over-expressing strain shows no significant effect on acetami-dase levels in the presence of intact amiA indicate that AmiC isimportant for the induction. Our in vivo over-expression studies
ARTICLE IN PRESSG ModelMICRES-25663; No. of Pages 8
B. Sundararaman et al. / Microbiological Research xxx (2014) xxx–xxx 7
Fig. 5. qRT-PCR measurements of mRNA abundance in wild-type and amiA over-expression strains. Expression of amiA was quantified in amiA over-expressionstrains under induced and uninduced conditions (A). Relative mRNA abundancewas determined for acetamidase (amiE) using total RNA at 4 h (B) and 18 h (C) afteracetamide induction. The data represent average values and standard error mea-surements from three technical replicates, shown as percent relative abundancena
srbie
msoDecPtamiioDbc
Fig. 6. Over-expression of AmiA and its effect on the expression of acetamidase.(A). M. smegmatis wild type cells and AmiA over-expressing cells were cultured andinduced with acetamide for 18 h. Bacterial proteins were extracted and analyzedby SDS-PAGE. Lanes 1, 3 and 5 are crude extracts of wild type cells and lanes 2,4 and 6 are crude extracts of AmiA over-expressing cells induced with 25, 35 and50 mM acetamide respectively. Molecular weight marker is indicated by kDa. (B)Acetamidase level of wild type and AmiA over-expressed strains were measuredusing densitometer scanning and concentration was determined using Quantity Onesoftware in various acetamide induced concentrations (25, 35 and 50 mM). The sym-bol (*) indicates mean values that are significant (P < 0.05). (C) Acetamidase enzyme
Chebrou H, Bigey F, Arnaud A, Galzy P. Amide metabolism: a putative ABC transporter
ormalized to the 16s rRNA gene. The symbols (*, **, ***) indicates P < 0.05, P < 0.01nd P < 0.001 respectively.
trongly demonstrated the physiological role played by AmiA asepressor. Thus, the characterization and identification of DNAinding sites of AmiA is an important step toward understand-
ng the operon and will allow construction of more sophisticatedxpression system in future.
We thus refine the previous hypothesis of the regulatoryechanism of acetamidase operon. In uninduced conditions, con-
titutively expressed AmiA from P1 promoter blocks the expressionf further downstream genes by binding near the P2 promoter.uring induction, a small quantity of AmiC expressed by leakyxpression of Pc may bind with amide and this complex in turnould bind with repressor AmiA and thereby could derepress the2 promoter. Expression of P2 promoter ensures sufficient quanti-ies of AmiD and AmiS. After sufficient quantities of AmiC, AmiDnd AmiS have been produced, the expression levels P2 and Pcay be reduced and the newly formed AmiC and AmiD might pos-
tively regulate the P3 promoter (which is weak and constitutiven normal conditions). This in turn might increase the expressionf acetamidase, AmiE to a large extent. Further work to investigate
Please cite this article in press as: Sundararaman B, et al. Expression, puoperon of Mycobacterium smegmatis. Microbiol Res (2014), http://dx.d
NA binding activity of positive regulators and protein interactionsetween the regulators will provide complete understanding of theomplex operon.
activity was estimated in wild type and AmiA over-expressed strains by estimatingrelease of ammonia using Nessler’s method at various acetamide concentrations.The symbol (*) indicates mean values that are significant (P < 0.05).
Appendix A. Supplementary data
Supplementary data associated with this article can befound, in the online version, at http://dx.doi.org/10.1016/j.micres.2014.02.011.
References
Av-Gay Y, Everett M. The eukaryotic-like Ser/Thr protein kinases of Mycobacteriumtuberculosis. Trends Microbiol 2000;8:238–44.
rification and functional characterization of AmiA of acetamidaseoi.org/10.1016/j.micres.2014.02.011
in Rhodococcus sp. R312. Gene 1996;182:215–8.Delic I, Robbins P, Westpheling J. Direct repeat sequences are implicated in the
regulation of two Streptomyces chitinase promoters that are subject to carboncatabolite control. Proc Natl Acad Sci U S A 1992;89:1885–9.

ING ModelM
8 iologic
D
H
H
I
K
M
M
M
N
N
ARTICLEICRES-25663; No. of Pages 8
B. Sundararaman et al. / Microb
raper P. The aliphatic acrylamide amidohydrolase of Mycobacterium smegmatis: itsinducible nature and relation to acyl-transfer to hydroxylamine. J Gen Microbiol1967;46:111–23.
o YS, Wulff DL, Rosenberg M. Bacteriophage lambda protein cII binds promo-ters on the opposite face of the DNA helix from RNA polymerase. Nature1983;304:703–8.
opwood DA, Wright HM. Bacterial protoplast fusion: recombination in fused pro-toplasts of Streptomyces coelicolor. Mol Gen Genet 1978;162:307–17.
mada A, Igarasi S, Nakahama K, Isono M. Asparaginase and glutaminase activitiesof micro-organisms. J Gen Microbiol 1973;76:85–99.
ohn HI, Harris JS. On the mode of action of sulphonamides. I. Action on E. coli. JPharmacol Exp Ther 1941;73:343–8.
ahenthiralingam E, Draper P, Davis EO, Colston MJ. Cloning and sequencing ofthe gene which encodes the highly inducible acetamidase of Mycobacteriumsmegmatis. J Gen Microbiol 1993;139:575–83.
anganelli R, Provvedi R, Rodrigue S, Beaucher J, Gaudreau L, Smith I. Sigmafactors and global gene regulation in Mycobacterium tuberculosis. J Bacteriol2004;186:895–902.
attern SG, Brawner ME, Westpheling J. Identification of a complex operator forgalP1, the glucose-sensitive, galactose-dependent promoter of the Strepto-myces galactose operon. J Bacteriol 1993;175:1213–20.
Please cite this article in press as: Sundararaman B, et al. Expression, puoperon of Mycobacterium smegmatis. Microbiol Res (2014), http://dx.d
arayanan S, Selvakumar S, Aarati R, Vasan SK, Narayanan PR. Transcriptional analy-sis of inducible acetamidase gene of Mycobacterium smegmatis. FEMS MicrobiolLett 2000;192:263–8.
orman RA, Poh CL, Pearl LH, O’Hara BP, Drew RE. Steric hindrance regulation of thePseudomonas aeruginosa amidase operon. J Biol Chem 2000;275:30660–7.
PRESSal Research xxx (2014) xxx–xxx
Parish T, Turner J, Stoker NG. amiA is a negative regulator of acetamidase expressionin Mycobacterium smegmatis. BMC Microbiol 2001;1:19.
Parish T, Mahenthiralingam E, Draper P, Davis EO, Colston MJ. Regulation ofthe inducible acetamidase gene of Mycobacterium smegmatis. Microbiology1997;143:2267–76.
Raghunand TR, Bishai WR, Chen P. Towards establishing a method to screen forinhibitors of essential genes in mycobacteria: evaluation of the acetamidasepromoter. Int J Antimicrob Agents 2006;28:36–41.
Roberts G, Muttucumaru DG, Parish T. Control of the acetamidase gene of Mycobac-terium smegmatis by multiple regulators. FEMS Microbiol Lett 2003;221:131–6.
Ryndak M, Wang S, Smith I. PhoP, a key player in Mycobacterium tuberculosis viru-lence. Trends Microbiol 2008;16:528–34.
Stover CK, de la Cruz VF, Fuerst TR, Burlein JE, Benson LA, Bennett LT, et al. New useof BCG for recombinant vaccines. Nature 1991;351:456–60.
Subbian S, Narayanan S. Identification and characterization of the regulatory ele-ments of the inducible acetamidase operon from Mycobacterium smegmatis. CanJ Microbiol 2007;53:599–606.
Wilkinson SP, Grove A. Ligand-responsive transcriptional regulation by members ofthe MarR family of winged helix proteins. Curr Issues Mol Biol 2006;8:51–62.
Wilson SA, Drew RE. Transcriptional analysis of the amidase operon from Pseu-domonas aeruginosa. J Bacteriol 1995;177:3052–7.
rification and functional characterization of AmiA of acetamidaseoi.org/10.1016/j.micres.2014.02.011
Wilson SA, Wachira SJ, Norman RA, Pearl LH, Drew RE. Transcription antiter-mination regulation of the Pseudomonas aeruginosa amidase operon. EMBO J1996;15:5907–16.
Zhang F, Xie JP. Mammalian cell entry gene family of Mycobacterium tuberculosis.Mol Cell Biochem 2011;352:1–10.




















