
Expression of genes and processing of enzymes for the biosynthesis of penicillins and cephalosporins
17
AntonievanLeeuwenhoek 65: 227-243, 1994. 227 Q 1994 Kluwer Academic Publishers.Printed in the Netherlands. Expression of genes and processing of enzymes for the biosynthesis of penicillins and cephalosporins Juan F. Martin, Santiago Guti6rrez, Francisco J. Fermindez, Javier Velasco, Francisco Fierro, Ana T. Marcos & Katarina Kosalkova Area of Microbiology, Department of Ecology, Genetics and Microbiology, Faculty of Biology, University of Ledn, 24071 Le6n, Spain Key words: Penicillin and cephalosporin biosynthesis, pcbAB genes, pcbC genes, penDE genes, ceflEF genes, cefG genes, transcription analysis, protein processing, regulation Abstract The genes pcbAB, pcbC and penDE encoding the enzymes (c~-aminoadipyl-cysteinyl-valinesynthetase, isopeni- cillin N synthase and isopenicillin N acyltransferase, respectively) involved in the biosynthesis of penicillin have been cloned from Penicillium chrysogenum and Aspergillus nidulans. They are clustered in chromosome I (10.4 Mb) of P. chrysogenum, in chromosome II of Penicillium notatum (9.6 Mb) and in chromosome VI (3.0 Mb) of A. nidulans. Each gene is expressed as a single transcript from separate promoters. Enzyme regulation studies and gene expression analysis have provided useful information to understand the control of genes involved in penicillin biosynthesis. The enzyme isopenicillin N acyltransferase encoded by the penDE gene is synthesized as a 40 kDa protein that is (self)processed into two subunits of 29 and 1 l kDa. Both subunits appear to be required for acyl- CoA 6-APA acyltransferase activity. The isopenicillin N acyltransferase was shown to be located in microbodies, whereas the isopenicillin N synthase has been reported to be present in vesicles of the Golgi body and in the cell wall. A mutant in the carboxyl-terminal region of the isopenicillin N acyltransferase lacking the three final amino acids of the enzymes was not properly located in the microbodies and failed to synthesize penicillin in vivo. In C. acremonium the genes involved in cephalosporin biosynthesis are separated in at least two clusters. Cluster I (pcbAB-pcbC) encodes the first two enzymes (~-aminoadipyl-cysteinyl valine synthetase and isopenicillin N synthase) of the cephalosporin pathway which are very similar to those involved in penicillin biosynthesis. Cluster II (ceJEF-cefG), encodes the last three enzymatic activities (deacetoxycephalosporin C synthetase/hydroxylaseand deacetylcephalosporin C acetyltransferase) of the cephalosporin pathway. It is unknown, at this time, if the cey'D gene encoding isopenicillin epimerase is linked to any of these two clusters. Methionine stimulates cephalosporin biosynthesis in cultures of three different strains of A. chrysogenum. Methionine increases the levels of enzymes (isopenicillin N synthase and deacetylcephalosporin C acetyltransferase) expressed from genes (pcbC and cefG respectively) which are separated in the two different clusters of cephalosporin biosynthesis genes. This result suggests that both clusters of genes have regulatory elements which are activated by methionine. Methionine- supplemented cells showed higher levels of transcripts of the pcbAB, pcbC, cejEF genes and to a lesser extent of cefG than cells grown in absence of methionine. The levels of the cefG transcript were very low as compared to those ofpcbAB, pcbC and cefEE The induction by methionine of transcription of the four cephalosporin biosynthesis genes and the known effect of this amino acid on differentiation ofA. chrysogenum indicates that methionine exerts a pleiotropic effect that regulates coordinately cephalosporin biosynthesis and differentiation.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Expression of genes and processing of enzymes for the biosynthesis of penicillins and cephalosporins

Antonie van Leeuwenhoek 65: 227-243, 1994. 227 Q 1994 Kluwer Academic Publishers. Printed in the Netherlands.
Express ion of genes and processing of enzymes for the biosynthesis of penicill ins and cephalosporins
Juan F. Martin, Santiago Guti6rrez, Francisco J. Fermindez, Javier Velasco, Francisco Fierro, Ana T. Marcos & Katarina Kosalkova Area of Microbiology, Department of Ecology, Genetics and Microbiology, Faculty of Biology, University of Ledn, 24071 Le6n, Spain
Key words: Penicillin and cephalosporin biosynthesis, pcbAB genes, pcbC genes, penDE genes, ceflEF genes, cefG genes, transcription analysis, protein processing, regulation
Abstract
The genes pcbAB, pcbC and penDE encoding the enzymes (c~-aminoadipyl-cysteinyl-valine synthetase, isopeni- cillin N synthase and isopenicillin N acyltransferase, respectively) involved in the biosynthesis of penicillin have been cloned from Penicillium chrysogenum and Aspergillus nidulans. They are clustered in chromosome I (10.4 Mb) of P. chrysogenum, in chromosome II of Penicillium notatum (9.6 Mb) and in chromosome VI (3.0 Mb) of A. nidulans. Each gene is expressed as a single transcript from separate promoters. Enzyme regulation studies and gene expression analysis have provided useful information to understand the control of genes involved in penicillin biosynthesis. The enzyme isopenicillin N acyltransferase encoded by the penDE gene is synthesized as a 40 kDa protein that is (self)processed into two subunits of 29 and 1 l kDa. Both subunits appear to be required for acyl- CoA 6-APA acyltransferase activity. The isopenicillin N acyltransferase was shown to be located in microbodies, whereas the isopenicillin N synthase has been reported to be present in vesicles of the Golgi body and in the cell wall. A mutant in the carboxyl-terminal region of the isopenicillin N acyltransferase lacking the three final amino acids of the enzymes was not properly located in the microbodies and failed to synthesize penicillin in vivo. In C. acremonium the genes involved in cephalosporin biosynthesis are separated in at least two clusters. Cluster I (pcbAB-pcbC) encodes the first two enzymes (~-aminoadipyl-cysteinyl valine synthetase and isopenicillin N synthase) of the cephalosporin pathway which are very similar to those involved in penicillin biosynthesis. Cluster II (ceJEF-cefG), encodes the last three enzymatic activities (deacetoxycephalosporin C synthetase/hydroxylase and deacetylcephalosporin C acetyltransferase) of the cephalosporin pathway. It is unknown, at this time, if the cey'D gene encoding isopenicillin epimerase is linked to any of these two clusters. Methionine stimulates cephalosporin biosynthesis in cultures of three different strains of A. chrysogenum. Methionine increases the levels of enzymes (isopenicillin N synthase and deacetylcephalosporin C acetyltransferase) expressed from genes (pcbC and cefG respectively) which are separated in the two different clusters of cephalosporin biosynthesis genes. This result suggests that both clusters of genes have regulatory elements which are activated by methionine. Methionine- supplemented cells showed higher levels of transcripts of the pcbAB, pcbC, cejEF genes and to a lesser extent of cefG than cells grown in absence of methionine. The levels of the cefG transcript were very low as compared to those ofpcbAB, pcbC and cefEE The induction by methionine of transcription of the four cephalosporin biosynthesis genes and the known effect of this amino acid on differentiation ofA. chrysogenum indicates that methionine exerts a pleiotropic effect that regulates coordinately cephalosporin biosynthesis and differentiation.

228
Introduction
Penicillins and cephalosporins are synthesized by the condensation of the three precursor amino acids L-s- aminoadipic acid, L-cysteine and L-valine. Pools of these three amino acids specific for/3-1actarn antibiot- ic biosynthesis (separate from the main pool of each amino acid) have been proposed (Htinlinger & Kubicek 1989b) but the evidence in support of this proposal is very scanty. ~-Aminoadipic acid-requiring lysine auxotrophs are blocked in penicillin biosynthesis, thus confirming the role of this precursor amino acid in the biosynthesis of fl-lactams (Somerson et al. 1961; Luengo et al. 1980). However, the production of fl- lactam antibiotics in prototrophic strains is not usually stimulated by addition of exogenous o~-aminoadipic acid, cysteine or valine. Only when the cultures were limited by the availability of a nitrogen source was penicillin biosynthesis stimulated by the addition of ~-aminoadipic acid (Revilla et al. 1986; H6nlinger & Kubicek 1989a).
The three amino acids are activated and linked together to form the tripeptide 6-(L-o~-aminoadipyl)- L-cysteinyl-D-valine (ACV) by the enzyme ACV syn- thetase, a multifunctional enzyme that carries out the activation of the three amino acids with ATP (presum- ably as aminoacyl adenylates) and the epimerization of L- to D-valine before forming the tripeptide. The ACV synthetase is encoded by the pcbAB gene, an unusu- ally large gene that was cloned from P. chrysogenum by Diez et al. (1990) and Smith et al. (1990) and from Acremonium chrysogenum by Guti6rrez et al. (1991). The deduced molecular weight of the ACV synthetases of P chrysogenum and A. chrysogenum were 425971 and 414791 respectively.
The ACV tripeptide is cyclized by the enzyme isopenicillin N synthase (cyclase) to form isopenicillin N (Fig. 1), an intermediate that shows a weak peni- cillin activity. The gene pcbC encoding isopenicillin N synthase was the first cloned gene of the penicillin and cephalosporin pathways (Samson et al. 1985; Barredo et al. 1989) and a very similar gene has been also cloned from A. nidulans (Ram6n et al. 1987) and from six dif- ferent bacteria which produce fl-lactam antibiotics (see reviews by Martin et al. (1992) and Aharonowitz et al. (1993)).
In the last step of the penicillin biosynthetic path- way, the c~-aminoadipyl side chain of isopenicillin N is removed and exchanged by a phenylacetyl or phe- noxyacetyl side chain in the penicillin G or penicillin V fermentations, respectively. The biochemistry of this
reaction is complex since five-related enzyme activi- ties are encoded by the same gene (penDE) (Alvarez et al. 1993). The penDE gene was cloned from P. chrysogenum (Barredo et al. 1989) and A. nidulans (Montenegro et al, 1990). This gene is absent from the cephalosporin producer A. chrysogenum (Gutirrrez et al. 1991a).
The first two genes of the penicillin pathway con- tain no introns and are very similar to those found in a variety of/3-1actam producing bacteria whereas the last gene (penDE) contains three introns and appears to be of fungal origin (Aharonowitz et al. 1993).
The first two steps of the cephalosporin pathway in A. chrysogenum are identical to those of the penicillin pathway in P. chrysogenum. In cephalosporin produc- ers isopenicillin N is later isomerized to penicillin N by the enzyme isopenicillin N epimerase encod- ed by the ceJD gene. Although the ceflD gene has been cloned from Streptomyces clavuligerus (Kovace- vick et al. 1990) and Nocardia lactamdurans (Coque et al. 1993b), repeated attempts to clone the cor- responding gene from A. chrysogenum have failed. The five-membered ring of penicillin N is expand- ed to a six-membered dihydrothiazine ring of deace- toxycephalosporin C by the deacetoxycephalosporin C synthase (so-called expandase) and the deacetoxy- cephalosporin C is later hydroxylated at the C3 ~ exo- cyclic methyl group to give deacetylcephalosporin C by the enzyme deacetoxycephalosporin C hydroxylase. In A. chrysogenum both the deacetoxycephalosporin C synthase and hydroxylase activities are encoded by a single gene ceflEF whereas in S. clavuligerus and N. lactamdurans they are encoded by two separate genes (ceflE and cefl?) which strongly ressemble each other (see reviews by Martfn (1992) and Aharonowtiz et al. (1993)).
In the final step of cephalosporin biosynthesis, the intermediate deacetylcephalosporin C is acetylated to cephalosporin C by the enzyme deacetylcephalosporin C acetyltransferase (DAC-AT) that uses acetyl-CoA as donor of the acetyl group. This enzyme is encoded by the cefG gene (Gutirrrez et al. 1992; Matsuda et al. 1992).
Very similar genes occur in cephamycin-producing bacteria (Coque et al. 1993a). None of the cephal- osporin biosynthetic genes except the last one (cefG) contain introns. The cefG gene contains two introns and is in fact very similar to the met2 of filamentous fungi, what suggest that it has evolved separatedly from the other genes of the pathway. Indeed, no coun- terpart of the cefG is known in the cephamycin pro-
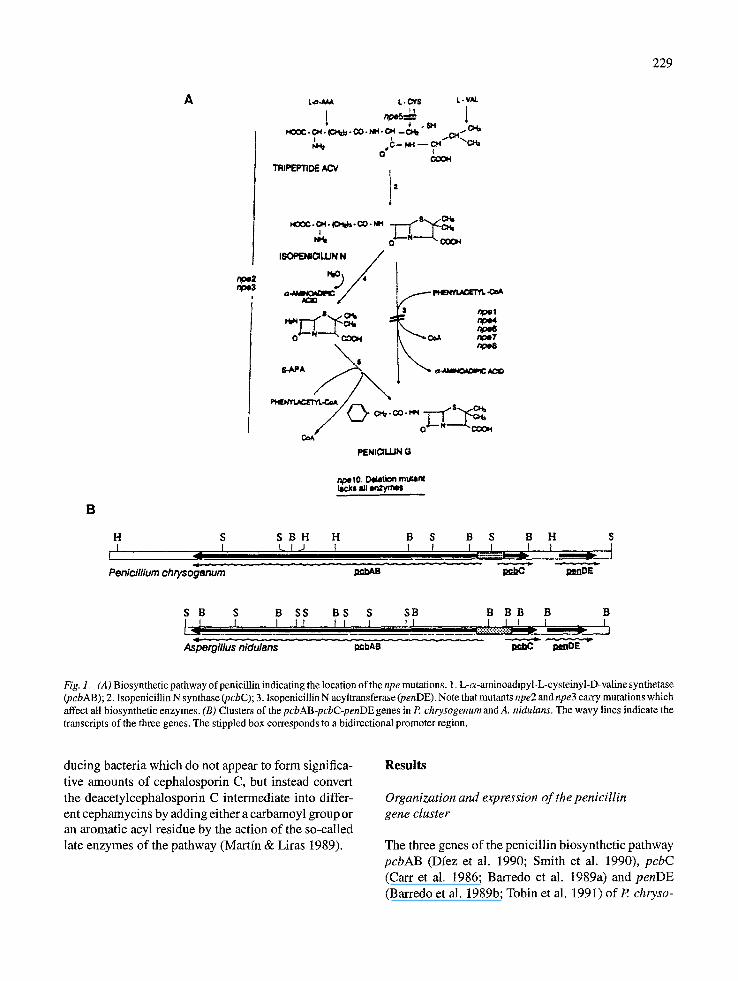
229
A
B
H S I i
I 11 Penicillium chtysogenum
~ 2 ¢~e3
L.a J , ~ k L • C'~fS L • V N .
HOOC- C,,H- ~ . CO- NH- CH --0'~ ~.i~ CI'~ i ~ i _ __ CH ~" "CH~
o,C NH I GOOH
TRIPEFTIDE ACV t
1' N~ 0 J ' ' N - - - - ~ COOH
ISOPENIGILLIN N /
= npel =- " ripe4
~ 7
PHINY1JCf1YI.-CaA / ~ - S C~, /
PENICILUN G
~ l O . ~ M ~ m ~ ~s¢l lmzymes
S B H H B S B S B
l~ l~e l~Jac
H S t , , {
01roDE
S B S B SS B S S SB B B B B B L ~ v I tT I r i ~f I I I L , - L_J .., ...... :.:.. - . P I
Aspergillus nidulans i ~AB ~bC I~IIIlDE
Fig. 1. (A)Bi~synthe~cpathway~fpenici~inindicatingthe~cati~n~fthenpemuta~ns.~.L-~-amin~adipy~-L-cysteiny~-D-va~inesynt~tase (pcbAB); 2. Isopenicillin N synthase (pcbC); 3. Isopenicillin N acyltransferase (penDE). Note that mutants npe2 and npe3 carry mutations which affect all biosynthetic enzymes. (B) Clusters of the pcbAB-pcbC-penDE genes in P chrysogenum and A. nidulans. The wavy lines indicate the transcripts of the three genes. The stippled box corresponds to a bidirectional promoter region.
ducing bacteria which do not appear to form significa- tive amounts of cephalosporin C, but instead convert the deacetylcephalosporin C intermediate into differ- ent cephamycins by adding either a carbamoyl group or an aromatic acyl residue by the action of the so-called late enzymes of the pathway (Martfn & Liras 1989).
Resul ts
Organization and expression of the penicillin gene cluster
The three genes of the penici l l in biosynthet ic pathway pcbAB (D/ez et al. 1990; Smith et al. 1990), pcbC (Carr et al, 1986; Barredo et al. 1989a) and penDE (Barredo et al. 1989b; Tobin et al. 1991) ofP. chryso-

230
genum have been cloned and shown to be linked in a single cluster (Diez et al. 1989; Dfez et al. 1990; Martin 1992). Similarly, the penicillin biosynthetic genes of A. nidulans were found to be linked in a single cluster (MacCabe et al. 1990; Montenegro et al. 1990, 1992). When the cluster of penicillin biosynthetic genes of P. chrysogenum was introduced in Neurospora crassa or Aspergillus niger (two species which do not form penicillin in nature) small levels of penicillin were syn- thesized (Smith et al. 1990b). This result indicates that all genetic information required for penicillin biosyn- thesis is contained in the cloned R chrysogenum DNA fragment that carries the penicillin gene cluster. How- ever, the low level of penicillin produced in A. niger and N. crassa suggest that these fungi lack activator pro- teins or enhancer sequences that may not be present in the cloned DNA fragment.
The gene encoding ACVS was designated pcbAB because two genetic loci were supposed to be respon- sible for synthesis of the c~-aminoadipyl-cysteine (AC) dipeptide and the ACV tripeptide. However, genet- ic evidence indicates that a single gene encodes all the activities needed to form ACV. Diez et al. (1990) cloned the pcbAB gene encoding ACVS by comple- mentation of the ACVS defective mutant npe5 and npe 10. The pcbAB gene restored penicillin production to mutant npe5 but not to npelO (a deletion mutant that lacks also the pcbC and penDE genes). Transcrip- tional mapping studies revealed a long transcript of about 11.5 kb. The approximate transcription initia- tion and termination sites were identified and the entire DNA region covering them sequenced. An unusually large open reading frame (ORF) was found containing 11,376 bp that encodes a protein of 3792 amino acids with a deduced molecular weight of 425,971. Similar findings were reported independently from a different P. chrysogenum strain (Smith et al. 1990a; Smith et al. 1990b). No introns were found in the P. chrysogenum gene (or other fungal pcbAB genes).
A conspicuous feature of the ACVS protein sequence is the presence of three repeated regions, or domains, each containing more than 500 amino acids. They show extensive sequence similarity with the Bacillus brevis peptide synthetases, gramicidin S synthetase I and tyrocidin synthetase I, and two other enzymes that carry out ATP-pyrophosphate exchange reactions, luciferase and 4-coumerate-CoA ligase (Tur- gay et al. 1992; Aharonowitz et al. 1993). All of these enzymes activate carboxyl groups present in amino acids, hydroxy acids, or simple acids. A multien- zyme thiotemplate mechanism has been proposed for
the ACVS reaction (Dfez et al. 1990; Gutirrrez et al. 1991; Coque et al. 1991; Aharonowitz et al. 1993). 'The three domains of ACVS probalby specify, there- fore, the amino acid-activating centers.
The P. chrysogenum pcbAB gene is separated from the two remaining genes needed for penicillin pro- duction, pcbC and penDE, by about 1100 bp and is transcribed in opposite orientation to them (Dfez et al. 1989) (Fig. 1). Transcriptional mapping studies showed the presence of one long transcript of about 11.5 kb, which hybridized with several probes internal to the pcbAB gene, and two small transcripts of 1.15 kb, which hybridized with the pcbC or thepenDE gene, respectively (Fig. 2). Probes internal to the pcbAB gene hybridized with an 11.5 kb transcript and also with bands of smaller size (Fig. 2). Two hypothesis may explain this result. Either the large 11.5 kb tran- script is broken down during the isolation procedure or transcription of the pcbAB may terminate prematurily, thus forming incomplete proteins. Proteins of smaller size than that of ACVS that react with anti-ACVS anti- bodies are often found in P. chrysogenum cultures (J. Velasco & J.F. Martfn, unpublished). These proteins are probably unable to synthesize the complete ACV molecule.
Similar pcbAB genes encoding ACV synthetases have been cloned fromA, chrysogenum (Gutirrrez et al. 1991a), A. nidulans (MacCabe et al. 1991; Montene- gro et al. 1993) and Nocardia lactamdurans (Coque et al. 1991a). A 24 kb region ofA. chrysogenum C10 DNA was cloned by hybridization with the pcbAB and pcbC genes of P. chrysogenum. A functional c~- aminoadipyl-cysteinyl-valine (ACV) synthetase was encoded by a 15.6 kb EcoRI-BamHI DNA fragment, as shown by complementation of the ACV synthetase- deficient mutant npe5 of R chrysogenum. The pcbAB was found to be closely linked to the pcbC gene form- ing a cluster of early cephalosporin biosynthetic genes (Gutirrrez et al. 1991b) (Fig. 3). A 3.2 kb BamHI frag- ment of this region complemented the mutation in the structuralpcbC gene oftheA, chrysogenum N2 mutant, resulting in cephalosporin production. Two transcripts of 11.4 and 1.15 kb were found by Northern hybridiza- tion ofA. chrysogenum RNA with probes internal to the pcbAB and pcbC genes, respectively (Fig. 4). The A. nidulanspcbAB gene encodes a protein of 3770 amino acids with a predicted molecular weight of 422,486. The pcbAB and pcbC genes are tightly linked in A. nidulans (as in A. chrysogenum) and are transcribed from an intergenic region of 872 bp in A. nidulans (as compared to 1233 bp in A. chrysogenum).
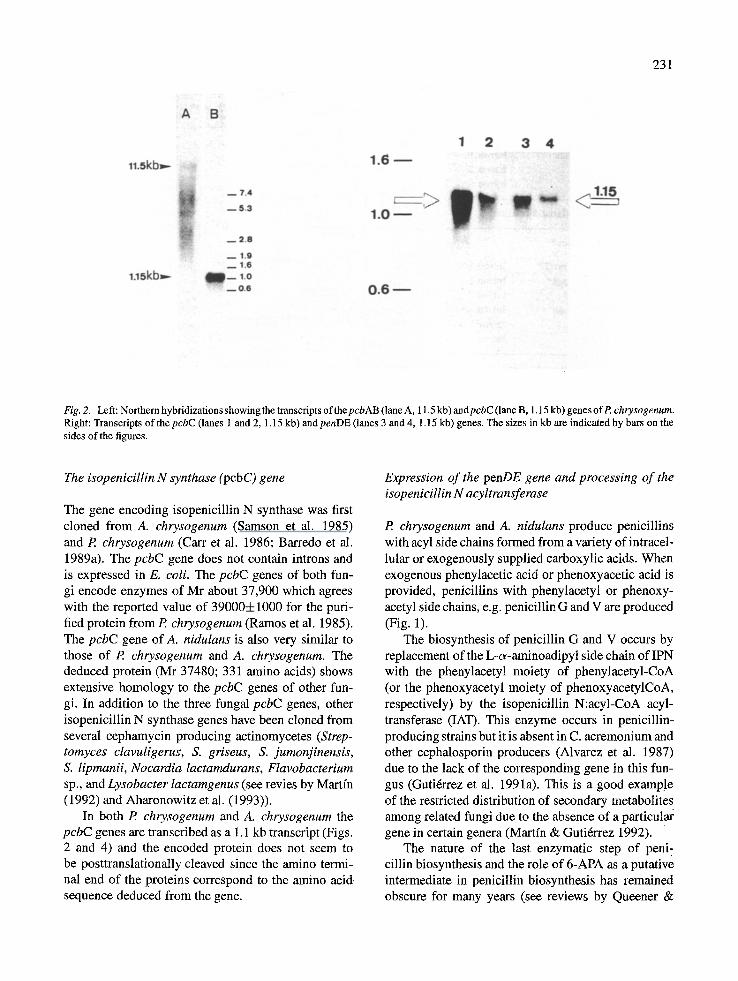
231
A B
11.5kb~,- ~,~iiii~
1.15kb~--
1 . 6 I
D 7 . 4 ~ - - , ~ _ 5a 1 . 0 1
m 2 . 8
_ _ 1 , 9
_ _ 1 . 6
0 . 6 1
1 2 3 4
Fig. 2. Left: Northern hybridizations showing the transcripts of the pcbAB (lane A, 11.5 kb) andpcbC (lane B, 1.15 kb) genes ofP. chrysogenum. Right: Transcripts of the pcbC (lanes 1 and 2, 1.15 kb) and penDE (lanes 3 and 4, 1.15 kb) genes. The sizes in kb are indicated by bars on the sides of the figures.
The isopenicillin N synthase (pcbC) gene
The gene encoding isopenicillin N synthase was first cloned from A. chrysogenum (Samson et al. 1985) and P. chrysogenum (Carret al. 1986; Barredo et al. 1989a). The pcbC gene does not contain introns and is expressed in E. coli. The pcbC genes of both fun- gi encode enzymes of Mr about 37,900 which agrees with the reported value of 39000-t-1000 for the puri- fied protein from P. chrysogenum (Ramos et al. 1985). The pcbC gene of A. nidulans is also very similar to those of P. chrysogenum and A. chrysogenum. The deduced protein (Mr 37480; 331 amino acids) shows extensive homology to the pcbC genes of other fun- gi. In addition to the three fungal pcbC genes, other isopenicillin N synthase genes have been cloned from several cephamycin producing actinomycetes (Strep- tomyces clavuligerus, S. griseus, S. jumonjinensis, S. lipmanii, Nocardia lactamdurans, Flavobacterium sp., and Lysobacter lactamgenus (see revies by Martfn (1992) and Aharonowitz et al. (1993)).
In both P. chrysogenum and A. chrysogenum the pcbC genes are transcribed as a 1.1 kb transcript (Figs. 2 and 4) and the encoded protein does not seem to be posttranslationally cleaved since the amino termi- nal end of the proteins correspond to the amino acid sequence deduced from the gene.
Expression of the penDE gene and processing of the isopenicillin N acyltransferase
P. chrysogenum and A. nidulans produce penicillins with acyl side chains formed from a variety of intracel- lular or exogenously supplied carboxylic acids. When exogenous phenylacetic acid or phenoxyacetic acid is provided, penicillins with phenylacetyl or phenoxy- acetyl side chains, e.g. penicillin G and V are produced (Fig. 1).
The biosynthesis of penicillin G and V occurs by replacement of the L-c~-aminoadipyl side chain of lPN with the phenylacetyl moiety of phenylacetyl-CoA (or the phenoxyacetyl moiety of phenoxyacetylCoA, respectively) by the isopenicillin N:acyl-CoA acyl- transferase (IAT). This enzyme occurs in penicillin- producing strains but it is absent in C. acremonium and other cephalosporin producers (Alvarez et al. 1987) due to the lack of the corresponding gene in this fun- gus (Gutirrrez et al. 1991a). This is a good example of the restricted distribution of secondary metabolites among related fungi due to the absence of a particular gene in certain genera (Martin & Gutirrrez 1992).
The nature of the last enzymatic step of peni- cillin biosynthesis and the role of 6-APA as a putative intermediate in penicillin biosynthesis has remained obscure for many years (see reviews by Queener &
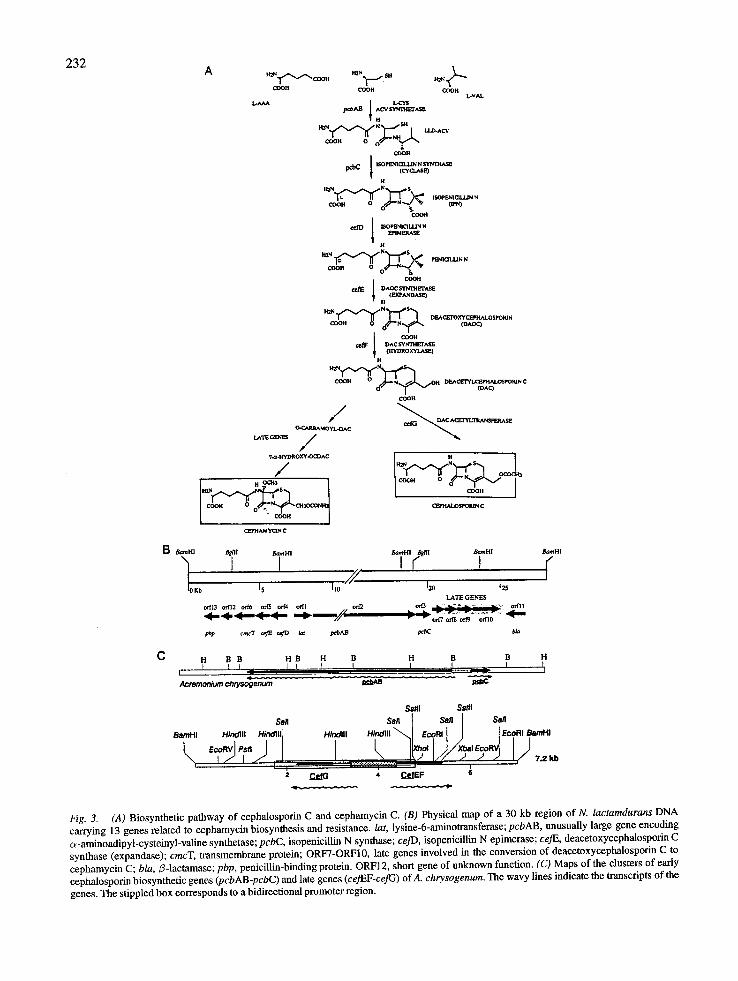
232
B ~ . . ,
C
COOH COOH ~ t l I~VAL
H N SH
COOH
I ISOM~NICILLIN N
H It~ N $
o ~.. 0 ~ ) COOll
I: o ~3OH
Q~E I DAOC ~IN~IL~T'4~E g a~X~rDASC~
H
, ~ f - v ~ % - f - - 1
~ g
~.a .14YDROXY.OCDAC II
II OC1t3 I ~ t
Jlg/I I BamH1 BamHI tBg/ll BamHI BamHl
OKb 15 ll0 I:0 lz~ LATE GENES
orfl3 orfl2 orf6 od5 off4 orfl od2 ~ ~ ~ ~ . ~ < - 4" < " 4 1 " ~ " " 1 ~ - - " ~ ~" " - o , ~ , ~ , n 9 o~10 pbp cmcT ct.JE ceil) /m pcbAB pcbC b/~
H B B H B H B H B B H I I I I I I I I I -.-,, i I t i i il
A.cremonium chrysogenum
BamHI
\
Sslll Ssfll
Sa~ Sail Satl I B
San
Hindlll H/ndlll I Hinolll Hindl,l ~ 1 E c o R I / l / EcoRI BamHI
I / I / i I, . ........ . ) : , ~ . / V I / / / ~ ~ i / 7 = k ~ i ~ 4 C~fEF 6
Fig. 3. (A) Biosynthetic pathway of cephalosporin C and cephamycin C. (B) Physical map of a 30 kb region of N. lactamdurans DNA carrying 13 genes related to cephamycin biosynthesis and resistance, lat, lysine-6-aminotransferase; pcbAB, unusually large gene encoding c~-aminoadipyl-cysteinyl-valine synthetase; pcbC, isopenicillin N synthase; ceJD, isopenicillin N epimerase; ceJE, deacetoxycephalosporin C synthase (expandase); cmcT, transmembrane protein; ORF7-ORF10, late genes involved in the conversion of deacetoxycephalosporin C to cephamycin C; bla,/3-1actamase; pbp, penicillin-binding protein. ORF12, short gene of unknown function. (C) Maps of the clusters of early cephalosporin biosynthetic genes (pcbAB-pcbC) and late genes (cej'<EF-ceJG) ofA. chrysogenum. The wavy lines indicate the transcripts of the genes. The stippled box corresponds to a bidirectional promoter region.
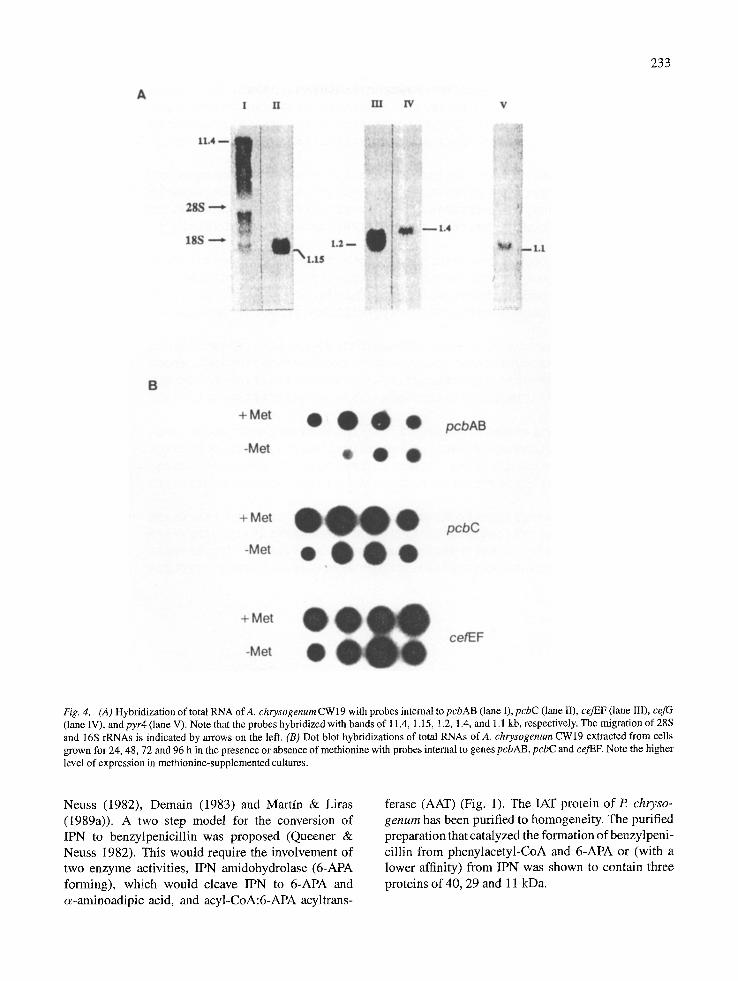
A I I I 11.1 I V
i u .4 - ....
t I
2 8 S -'*" : I
- - 1 .4
18S ~ 1 . 2 -
:: "~1.t5
i ; }
V
} /
233
B
+Me • O 0 • -Met 0 O O
pcbAB
+ Met
-Met o O O o pcbC
+ Met
-Met O cefEF
Fig. 4. (A ) Hybridizati•n •f t•ta• RNA •f A. chrys•genum CW • 9 with pr•bes interna• t• pcbAB (•ane •), pcbC (•ane I•), ceJEF (•ane ••I), cefG (lane IV), and pyr4 (lane V). Note that the probes hybridized with bands of 11.4, 1.15, 1.2, 1.4, and 1.1 kb, respectively. The migration of 28S and 16S rRNAs is indicated by arrows on the left. (B) Dot blot hybridizations of total RNAs ofA. chrysogenum CW19 extracted from cells grown for 24, 48, 72 and 96 h in the presence or absence of methionine with probes internal to genes pcbAB, pcbC and cej'EF. Note the higher level of expression in methionine-supplemented cultures.
Neuss (1982), Demain (1983) and Martin & Liras (1989a)). A two step model for the conversion of IPN to benzylpenicillin was proposed (Queener & Neuss 1982). This would require the involvement of two enzyme activities, IPN amidohydrolase (6-APA forming), which would cleave IPN to 6-APA and c~-aminoadipic acid, and acyl-CoA:6-APA acyltrans-
ferase (AAT) (Fig. 1). The IAT protein of P. chryso- genum has been purified to homogeneity. The purified preparation that catalyzed the formation of benzylpeni- cillin from phenylacetyl-CoA and 6-APA or (with a lower affinity) from IPN was shown to contain three proteins of 40, 29 and 11 kDa.

234
~T-~6 MLHILCQGTPFEIGYEHGSAAKAVIARSIDFAVDLIRGKTKKTDE ~T-111~7/8 MLHILCQGTPFEIGYEHGSAAKAVIARSIDFAVDLIRGKTKKTDE
45 ~T-AS-P.7$ MLHILCQGTPFEIGYEHGSAA-KAVIARSIDFAVDLIRGKTKKTDE ~T-~ ~dula~ MLHVTCQGTPSEIGYHHGSAAKGEIAKAIDFATGLIHGKTKKTQA
I ELKQVLSQLGRVIEERWPKYYEEIRGIAKGAERDVSEIVMLNTRTEFAYGLKAARDGCTT ELKQVLSQLGRVIEERWPKYYEEIRGIAKGAERDVSEIVMLNTRTEFAYGLKA2%RDGCTT
105 ELKQVLSQLGRVIEERWPKYYEEIRGIAKGAERDVSEIVMLNTRTEFAYGLKA3%RDGCTT ELEQLLRELEQVMKQRWPRYYEEICGIAKGAEREVSEIVMLNTRTEFAYGLVEARDGCTT
AYCQLPNGALQGQNWDFFSATKENLIRLTIRQAGLPTIKFITE~IGKVGFNSAGVAVN AYCQLPNGALQGQNWDFFSATKENLIRLTIRQAGLPTIKFITE~iIGKVGFNSAGVAVN AYCQLPNGALQGQNWDFFSATKENLIRLTIRQAGLPTIKFITE~IGKVGFNSAGVAVN VYCKTPNGALQGQNWDFFTATKENLIQLTICQPGLPTIKMITE~IGKVGFNSAGVAVN
165
YNALHLQGLRPTGVPSHIALRIALESTSPSQAYDRIVEQGGMA2%SAFIMVGNGHEAFGLE YNALHLQGLILPTGVPSHIALRIALESTSPSQAYDRIVEQGGMAASAFIMVGNGHEAFGLE YNALHLQGLRPTGVPSHIALRIALESTSPSQAYDRIVEQGGMAASAFIMVGNGHEAFGLE YNALHLHGLRPTGLPSHLALRMALESTSPSEAYEKIVSQGGMAASAFIMVGNAHEAYGLE
215
FSPTSIRKQVLDANGRMVHTNHCLLQH~NEK~LPDSWNRHQR_MEFLLDGFDGTKQA FSPTSIRKQVLDANGRMVHTNHCLLQHG~NEK~D~LPDSWNRHQRMEFLLDGFDGTKQA
FSPTSIRKQVLDANGRMVHTNHCLLQH~NE~LPDSWNRHQR24.EFLLDGFDGTKQA FSPISLCKQVADTNGRIVHTNHCLLNH ~P SAQ~:.~.~P LPDSWSRHGRMEHLLSGFDGTKEA
275
FAQLWADEDNYPFSICRAYEEGKSRGATLFNIIYDHARREATVP.LGRPTNPDEMFVM_~FD FAQLWADEDNYPFSICRAYEEGKSRGATLFNIIYDHARREATVRLGRPTNPDEMFVMRFD FAQLWADEDNYPFSICRAYEEGKSRGATLFNIIYDHARREATVRLGRPTNPDEMFVM_RFD FAKLWEDEDNYPLSICRAYKEGKSRGSTLFNIVFDHVGR_KATVRLGRPNNPDETFVMTFS
335
EEDERSAL~ EEDERSAL~ARL] 347 EEDERSAL~ NLDTKSAI QANI
Fig. 5. Deducedamin•acidsequences•ftheis•penici••inNacy•transferases•fP.chrys0genumAS•P•78•then•n•pr•ducermutantsnPe6•nPe7 or npe8, and A. nidulans showing the processing site (vertical arrow), and the amino acid changes (Gly ~5° to Val or Glu 258 to Lys) in the npe mutants (shadowed). Note that Gly 15° and Glu 258 are conserved in A. nidulans. GlylS° is nested in a highly conserved region in P. chrysogenum and A. nidulans whereas Glu 258 is the first conserved residue following a region of poorly conserved amino acids in P. chrysogenum and A. nidulans (boxed). The mutations do not affect the conserved region around the processing site (underlined) or the microbody targeting signal (ARL) at the C-terminal end of the protein (boxed).
The N-terminal sequence of the 29 and 40 kDa pro- teins were used to clone a D N A fragment (containing t hepenDE gene) of P. chrysogenum that complemented the deficiency of IAT in npe6, a mutant blocked in peni- cillin biosynthesis. T h e p e n D E gene contained an open reading frame with three introns (Barredo et al. 1989b). The deduced amino acid sequence of the open read- ing frame encodes a 39 kDa protein (Fig. 5). A DNA
sequence in the gene was found that corresponded to the N-terminal sequence of the 40 kDa protein, and downstream of this sequence the nucleotide sequence matched the N-terminal sequence o f the 29-kDa pro- tein (Barredo et al. 1989b). The large protein (40-kDa) corresponded to a heterodimer formed from the 11- and 29-kDa subunits (Whiteman et al. 1990). The 11- and 29-subunits are formed by (self)cleavage of the
S CH.~
|SOPENICILLJN N
~kl4tlN OJ~J~Pt¢ ACID
P H E ~ - C O A
1
0.~----- A -.----,--~ COOH
~ID ~ 3 a'AMINOADtpI~ ACiD
PtCEInl"AC EIYL'Cc~ ~ O . M H S CH3
o~---,-- coo . oj~,-- ~ ..--.--LCOO.
PHE N OXYMETHYLPENICILLIN 6-APA
235
\
5
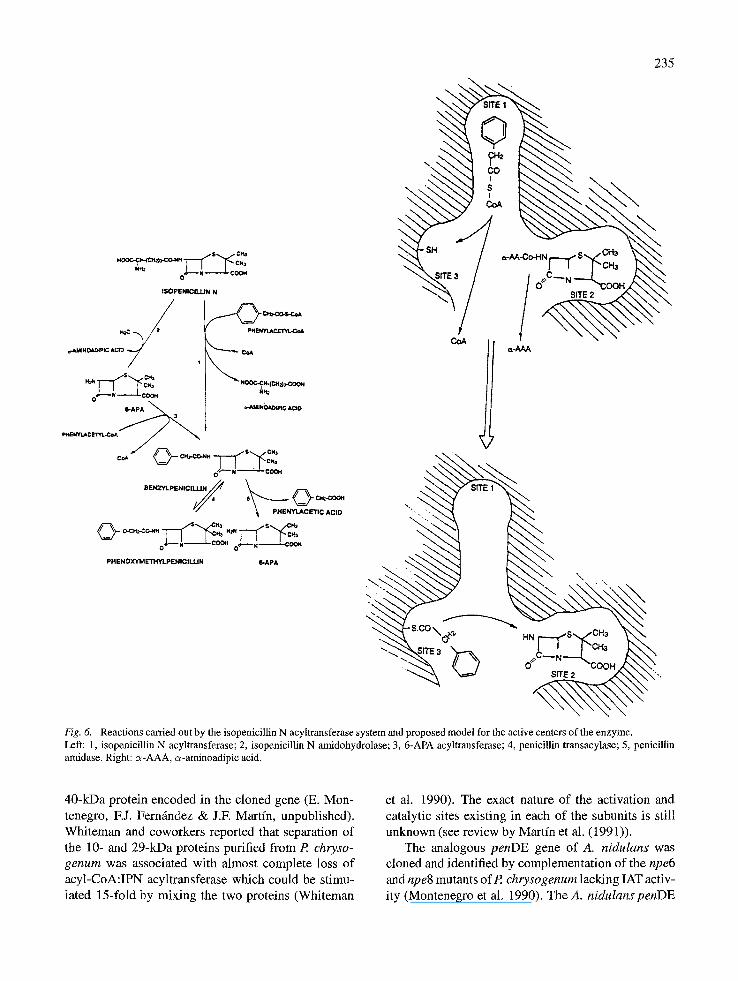
Fig. 6. Reactions carried out by the isopenicillin N acyltransferase system and proposed model for the active centers of the enzyme. Left: 1, isopenicillin N acyltransferase; 2, isopenicillin N amidohydrolase; 3, 6-APA acyltransferase; 4, penicillin transacylase; 5, penicillin amidase. Right: c~-AAA, e~-aminoadipic acid.
40-kDa protein encoded in the cloned gene (E. Mon- tenegro, EJ. Fernfindez & J.F. Martin, unpublished). Whiteman and coworkers reported that separation of the 10- and 29-kDa proteins purified from P. chryso- genum was associated with almost complete loss of acyl-CoA:IPN acyltransferase which could be stimu- lated 15-fold by mixing the two proteins (Whiteman
et al. 1990). The exact nature of the activation and catalytic sites existing in each of the subunits is still unknown (see review by Martfn et al. (1991)).
The analogous penDE gene of A. nidulans was cloned and identified by complementation of the npe6 and npe8 mutants ofP. chrysogenum lacking IAT activ- ity (Montenegro et al. 1990). The A. nidulanspenDE

236
gene is very similar to the previously cloned penDE gene ofP. chrysogenum (Barredo et al. 1989b) (Fig. 5). Both genes contain three introns in similar positions that were identified by comparison with the fungal consensus intron/exon splicing sequences (Ballance 1986) and confirmed by RNA hybridization exper- iments using three oligonucleotides internal to each intron.
The presence of three introns in the penDE genes of P. chrysogenum and A. nidulans suggests that these genes have not originated from bacteria in a recent transfer event as proposed for the other genes of the penicillin and cephalosporin pathways. These genes appear to have evolved in P chrysogenum and A. nidu- lans from ancestral fungal genes.
The IAT activity co-purified with the acyl-CoA:6- APA acyltransferase (AAT) which transfers an acyl- group from acyl-CoA derivatives (e.g. phenylacetyl- CoA, phenoxyacetyl-CoA) to 6-APA (Alvarez et al. 1993). Other thioesters of phenylacetic acid were also used as substrates. An amino acid sequence similar to that of the active site of thioesterases was found in the isopenicillin N acyltransferase, suggesting that this site is involved in the transfer of phenylacetyl residues from phenylacetyl thioesters. Purified isopenicillin N acyltransferase also showed isopenicillin N amidohy- drolase, penicillin transacylase and penicillin amidase activities. The isopenicillin N amidohydrolase (releas- ing 6-APA) showed a much lower specific activity than the isopenicillin N acyltransferase of the same enzyme preparation suggesting that in the isopenicillin N acyltransferase reaction the 6-APA is not released and is directly converted into benzylpenicillin. Peni- cillin transacylase exchanged side chains between two hydrophobic penicillins or between one penicillin and 6-APA. The penicillin amidase activity is probably the reverse of the biosynthetic acyl-CoA:6-APA acyltrans- ferase. Four P chrysogenum mutants deficient in acyl- CoA:6-APA acyltransferase lacked the other four relat- ed activities. Transformation of these mutants with the penDE gene restored all five enzyme activities (Alvarez et al. 1993).
The IAT may contain three binding sites (Martfn et al. 1991)
- an acyl-binding site which interacts with activated forms of phenylacetyl, phenoxyacetyl and other acyl moieties (site 1 in Fig. 6);
- a binding site for IPN (site 2); and
- a binding site for 6-APA that may be the same as the one for IPN.
The first step in the conversion of IPN into benzylpeni- cillin most likely involves formation of an IPN-enzyme complex (site 2), followed by hydrolysis of IPN with the release of L-c~-aminoadipic acid. The conversion of IPN to 6-APA and ~-aminoadipic acid by the IPN amidohydrolase was proposed by Queener and Neuss (Queener & Neuss 1982) but no clear evidence of the existence of this enzyme was available until recent- ly (Alvarez et al. 1993). The IPN amidohydrolase activity in purified IAT preparations showed the same optimal pH and temperature as IAT. Genetic evidence indicated that both enzyme activities are located in a single gene product. However, the IPN amidohydro- lase activity was much lower than the IAT activity of the same enzyme preparation, suggesting that the enzyme activity is modified when the second substrate phenylacetyl-CoA interacts with the acyl-CoA bind- ing site (site 1). This hypothesis is supported by the considerably higher affinity of the IAT for IPN when phenylacetyl-CoA has been added (apparent Km 23 #M) than when phenylacetyl-CoA is absent (apparent Km 4.0 mM). The release of 6-APA by IPN amido- hydrolase activity seems to be a residual activity of IAT when no acyl-CoA precursor of the side chain is available (Vmax, 50 pkatals/mg of protein). During the complete IAT reaction (Vmax, 1380 pkatals/mg of protein), 6-APA appears to remain enzyme-bound and is efficiently converted into hydrophobic penicillins.
The second step in the reaction catalyzed by IAT is better known. An AAT activity catalyzes the transfer of the acyl-moiety of phenylacetyl-CoA (or phenoxyacetyl-CoA) to form benzylpenicillin (or phe- noxymethylpenicillin) with release of CoA. Queener and Neuss (Queener & Neuss 1982) suggested that an acyl-CoA binding site exists on the enzyme and that it interacts primarily with the acyl moiety of acyl- CoA substrates. Our results support this hypothesis and provide evidence that this active site (site 1) also recognizes other acylthioesters, such as phenylacetyl- glutathione or phenylacetyl-ACV. In particular, pheny- lacetylglutathione may be used as a natural activated form of phenylacetic acid for penicillin biosynthesis. The AAT activity is inhibited by PheMeSO2F presum- ably by interfering with the binding of phenylacetyl- CoA (or phenylacetylglutathione), which agrees with previous reports on the effect of this inhibitor on peni- cillin amidases. PheMeSO2F binds covalently to the penicillin amidase of E. coli (Siewinski et al. 1984).
The amino acid sequence GKSRGA, similar to the consensus sequence of the thioesterase active site of a variety of polyketide synthetases (Martin & Guti6rrez

1992), fatty acid synthetases (Naggert et al. 1988; Yuan et al. 1988; Hardie et al. 1985; Poulose et al. 1981) and ACV synthetases (Guti6rrez et al. 1991; Coque et al. 1991), has been found in the carboxyl terminal region of the IAT (Fig. 5). This sequence contains the consen- sus motif GXSXG found in several serine active site esterases (Dayhoffet al. 1976). The serine in this motif has been found to be the reactive residue of thioesteras- es (Randhawa et al. 1987). This finding suggests that the transfer of acyl-groups from phenylacetyl-CoA or other acyl-thioesters may work by a mechanism sim- ilar to those of thioesterases in which the activated acyl group is transferred from acyl-thioesters to an SH group on the enzyme (site 3 in Fig. 1). Four cys- teine residues are conserved in the deduced amino acid sequences of the acyltransferases of P. chrysogenum and A. nidulans (Montenegro et al. 1990). This would explain the need for reducing agents in the reaction mixture.
The studies with strains npe6, npe8 and npelO (Martin et al. 1987; Cantoral et al. 1993) showed that all the enzyme activities of the IAT are lost simulta- neously. Transformants of the three mutants recovered all five activities after complementation with plasmid pULB 101, which carries the penDE gene, i.e. the five activities are encoded by a single gene.
The role of the subunit of the IAT on recognition of the different substrates in still obscure. Overexpression of an intron-less penDE gene obtained by the rDNA technique has resulted in the production of an a-/3- heterodimer containing subunits of 11 kDa and 29 kDa (Aplin et al. 1993; E. Montenegro, EJ. Fern~indez & J.F. Martfn, unpublished). This configuration is created by cleaving the 40 kDa precursor polypeptide between Gly 1°2 and Lys 1°3, as observed initially by Barredo et al. (1989) and Veenstra and coworkers (1989).
Tobin and coworkers (1993) have obtained separat- edly the 29 and 11 kDa subunits in E. coli and have made reconstitution experiments. Production of either subunit in the absence of the other did not form active AAT. Co-transformation of E. coli with two plasmids, each encoding a different IAT subunit, produced AAT activity. Mutation ofpenDE replacing Thr 1°5 (located at the cleavage site) with Asn resulted in inactive and uncleaved IAT. In vitro transcription translation experi- ments suggested that IAT hydrolysis is an autocatalytic event (Tobin et al. 1993). However it is unclear whether selfcleavage occurs in vitro using purified preparations of the enzyme.
237
Involvement of microbodies in penicillin biosynthesis
The IAT of P. chrysogenum was shown to be locat- ed in organelles which show a size, shape and cellu- lar distribution characteristic for fungal microbodies (Mtiller et al. 1991). In constract the isopenicillin N synthase (cyclase) was reported to be located in vesi- cles belonging to the Golgi body and in the cell wall (Kurz~tkowski et al. 1991). The amino terminal end of the IAT of P. chrysogenum showed the presence of a putative targeting signal for microbodies (Ala- Arg-Leu). To verify the hypothesis that the acyltrans- ferase is directed to microbodies by the ARL termi- nal sequence (MOiler and coworkers 1992) construct- ed mutants lacking the ARL sequence. The mutant and wild typepenDE genes were introduced into P. chryso- genum npe6, a non-penicillin producing strain which lacks IAT activity (Cantoral et al. 1993). Penicillin production was restored in transformants containing the wild type penDE gene but not in transformants containing the mutant gene. The ARL mutant gene was expressed in transformants although at reduced levels as revealed by immunoblotting. However cau- tion should be taken when interpreting this result since strain npe6 itself produces some IAT protein which may be detected by immunoblotting (EJ. Fernfindez & J.E Martfn, unpublished). Using immunoelectron microcopy the wild type IAT was found in specific organelles surrounded by a single membrane where- as the mutated enzyme was located in vacuoles and in neighbouring cytoplasm. These results indicate that the targeting signal Ala-Arg-Leu plays a role in direct- ing the IAT to microbodies for penicillin biosynthesis. However the A. nidulans IAT does not contain the C- terminal Ala-Arg-Leu sequence and is able to synthe- size penicillin.
Resolution of the chromosomes of A. nidulans, E chrysogenum and E notatum: Mapping of the pen gene cluster
The genome ofA. nidulans ATCC 28901 was resolved into seven chromosomal bands by pulsed field gel elec- trophoresis (Montenegro et al. 1992). The three genes of the penicillin biosynthetic pathway pcbAB, pcbC and penDE are clustered together on a chromosome of 3.0 Mb, corresponding to linkage group VI, whereas the argB gene was located on a chromosome of 3.4 Mh corresponding to linkage group III (Montenegro et al. 1992).

238
Four chromosomes were resolved by pulse field gel electrophoresis in P. notatum (sizes of 10.8, 9.6, 6.3 and 5.4 Mb) and in five different strains of P. chryso- genum (10.4, 9.6, 7.3 and 6.8 Mb in the wild type) (Fierro et al. 1993). Small differences in sizes were found between the four chromosomes of five different P. chrysogenum strains. The penicillin gene cluster was located by hybridization with a pcbAB probe in chro- mosome II of P. notatum and in chromosome ! of all P. chrysogenum strains except in the deletion mutant P. chrysogenum npelO that lacks this DNA region. The pyr G gene was located in chromosome I in P. notatum and in chromosome II in all P. chrysogenum strains except in mutant AS-P-78 where it hybridized with chromosome III. A major chromosomal rearrangement seems to have occurred in the high penicillin-producing strain AS-P-78. A fast moving DNA band observed in all gels corresponds to mitochondrial DNA. The total genome size has been calculated as 32.1 Mb in P. nota- turn and 34.1 Mb for the P. chrysogenum strains (Fierro et al. 1993).
The cephalosporin genes are organized in at least two clusters
The cluster of early genes The genes (pcbAB and pcbC) encoding the first two steps of the cephalosporin pathway have been cloned from the cephalosporin-producer A. chrysogenum and the cephamycin producers N. lactamdurans and S. clavuligerus. They have been described above since they are analogous to the genes involved in penicillin biosynthesis. The pcbAB was found to be closely linked to the pcbC gene forming a cluster of early cephalosporin-biosynthetic genes. These two genes in A. chrysogenum are transcribed in opposite orienta- tions, as in P. chrysogenum (Gutirrrez et al. 1991a) (Figs. 1 and 3). However, in N. lactamdurans and S. clavuligerus this organization is different since pcbAB and pcbC are clustered head to tail (Fig. 3).
The cluster of late genes A separate cluster of genes (ceflEF-cefG) in the genome of A. chrysogenum encodes the bifunc- tional deacetoxycephalosporin C (DAOC) syn- thetase/hydroxylase (Samson et al. 1987b) and the acetyl-CoA:deacetylcephalosporin C acetyltransferase (Gutirrrez et al. 1992; Matsuda et al. 1992). The ceil) gene of C. acremonium encoding the isopenicillin N
epimerase, has not yet been cloned and it is not known whether it is linked to any of the two clusters.
The ceflEF-cefG cluster is located in chromosome II whereas the pcbC gene (and also the pcbAB which is closely linked) was found in chromosome VI (Ska- trud & Queener 1989). Chromosome rearrangements have been found in improved cephalosporin C produc- ing strains of C. acremonium. In one improved strain the chromosomal changes altered the size of the chro- mosome on which the pcbC was located (Smith et al. 1991).
The ce3'EF gene coding for a bifunctional peni- cillin N expandase/DAOC 3~-hydroxylase was cloned by Samson et al. (1987b), using DNA probes based on the amino acid sequence of internal peptide frag- ments of the purified bifunctional protein. A protein of Mr 36461 was encoded by the open reading frame. The gene was expressed in E. coli and the protein formed contained both penicilin expandase and DAOC 3'-hydroxylase activities demonstrating the bifunction- al nature of the polypeptide encoded by the gene.
The gene (cefG) encoding the acetyl-CoA:deacetyl- cephalosporin C acetyltransferase Which catalyzes the last step of the cephalosporin pathway of A. chryso- genum C 10 has been cloned recently in our laboratory (Gutirrrez et al. 1992) and simultaneously by Matsu- da et al. (1992), and later by Mathison et al. (1993). It contains two introns and encodes a protein of 444 amino acids with a Mr of 49269 that correlates well with the Mr deduced by gel filtration. The cefG gene is linked to the cejEF gene but it is expressed in the opposite orientation to the ceflEF gene. Two transcripts of 1.2 and 1.4 kb were found in A. chrysogenum that correspond to the ceflEF and cefG genes respectively; the degree of expression of the cefG gene was clear- ly smaller than that of the cejEF gene in 48 h cul- tures. The cloned cefG complemented the deficiency of deacetylcephalosporin acetyltransferase in the non- producer mutant A. chrysogenum ATCC 20371 and restored cephalosporin biosynthesis in this strain. Het- erologous expression of the cefG genes took place in P. chrysogenum. The deacetylcephalosporin acetyltrans- ferase showed a much higher degree of homology with the O-acetylhomoserine acetyltransferases of Saccha- romyces cerevisiae and Ascobolous immersus than with other O-acetyltransferases. It has been reported that the DAC acetyltransferase is processed into two subunits (Matsuyama et al. 1992) but the availble information is confusing since the molecular weight of the pro- posed subunits does not agree with that of the deduced molecular weight of the protein encoded by the gene.

The ceJEF-cefG cluster of genes encodes, therefore, the enzymes that carry out the three late steps of the cephalosporin biosynthetic pathway.
The missing gene: CefD encoding isopenicillin N epimerase After the synthesis of isopenicillin N, the biosynthet- ic route to penicillins and cephalosporins diverges into two branches (Figs. 1 and 3). In P. chryso- genum, the a-aminoadipyl side chain of isopeni- cillin N is exchanged for one of several hydropho- bic side chains derived from the corresponding acyl- CoA derivatives. In cephalosporin-producing organ- isms, the L-a-arninoadipyl side chain is first isomer- ized to the D-enantiomer to produce penicillin N; peni- cillin N and not isopenicillin N is the substrate for the ring-expansion enzyme. Isopenicillin N epimerase activity was first shown in cell-free extracts of A. chrysogenum by Knomi et al. (1979), who detected trace amounts of cephalosporin (i.e. a penicillinase- resistant antibiotic) in reaction mixtures containing ACV. Cephalosporin was found only if penicillin N was produced in the reaction. This observation was confirmed by Baldwin et al. (1981), who showed conversion of isopenicillin N into the penicillinase- resistant deacetoxycephalosporin C. They proposed thatA, chrysogenum possess an epimerase activity that was highly labile in cell-flee preparations.
The gene (ceil)) encoding isopenicillin N epimerase has been cloned from the DNA of the cephamycin producing bacteria S. clavuIigerus (Kovacevick et al. 1990), Nocardia lactamdurans (Coque et al. 1993b) and Lysobacter lactamgenus (Japanese Patent application by Takeda Industria no. 2-291274, 1990). However repeated attempts to clone the ceil) gene ofA. chrysogenum by hybridization with the bacterial ce3ff) genes have failed, presumably due to the very different G+C content and codon usage.
Methionine increases the levels of pcbAB, pcbC, cefEF and cefG genes in a. chrysogenum
Methionine, particularly the D-isomer is known to stimulate the synthesis of cephalosporin by A. chrysogenum (Demain & Wolfe 1987; Martfn & Aharonowitz 1983; Niiesch et at. 1987). Methion- ine stimulates the biosynthesis of penicillin N and cephalosporin C (Komatsu et al. 1975; Zanca & Martfn 1983) and at the same time causes fragmen- tation of the mycelium (Drew et ai. 1976; Nash &
239
Huber 1971). Methionine increases the formation of the isopenicillin N synthetase (cyclase), deacetoxy- cephalosporin C synthetase (expandase) (Sawada et al. 1980), a-aminoadipyl-cysteinyl-valine (ACV) syn- thetase (Zhang et al. 1987) and deacetylcephalosporin C acetyltransferase (Velasco et al. 1994) activities in A. chrysogenum.
Methionine stimulates cephalosporin production in cultures of three different strains of A. chrysogenum (the wild type isolate of Brotzu and the two improved strains CW19 and C10) when added either at inocula- tion time, or at 72 h, to cells grown previously in the absence of methionine. When methionine was added at 72 h the stimulation of cephalosporin biosynthesis was observed 12 h later and required de novo protein synthesis. Methionine increases the levels of enzymes (isopenicillin N synthase and deacetylcephalosporin C acetyltransferase) expressed from genes (pcbC and cef G respectively) which are separated in the two dif- ferent clusters of cephalosporin biosynthesis genes. This result suggests that both clusters of genes have regulatory elements which are activated by methio- nine. Methionine-supplemented cells showed higher levels of transcripts of the four known genes (pcbAB, pcbC, ceJEF and cefG) of the cephalosporin biosyn- thetic pathway than cells grown in absence of methio- nine. The levels of the cefG transcript were very low as compared to those of pcbAB, pcbC and ce]EF and transcription of cefG is induced by methionine to a lower extent than the other three genes (Velasco et al. 1994).
The degree of stimulation of cephalosporin produc- tion by methionine is specific for each industrial strain and even methionine-independent strains have been reported (Komatsu et al. 1975). Our findings indicate that the mutations introduced during strain improve- ment have resulted in a higher degree of expression of the cephalosporin biosynthetic genes in the CW19 and particularly in the C 10 strains. Methionine induces higher levels of transcription of the pcbAB, pcbC and ceCEF genes in the three strains of A. chrysogenum, although the induction effect is more clear in the high producing strain C-10. It is important to note that each transcript is formed as an independent unit from a separate promoter; pcbAB and pcbC, and also ceJEF and cefG, are expressed from bidirectional promoter regions (Fig. 3). This suggests a coordinated mech- anism of control affecting pcbAB, pcbC and cefEF but probably not cefG. This coordinated mechanism of control has been suggested previously by Ramos et al. (1986) and by Ramsdem and coworkers (1989). Tran-

240
scription of the cefG gene may be the limiting step in the pathway. It is important that cefG is the only cephalosporin gene that contains introns (Guti6rrez et al. 1992) and is not present in the bacterial cluster of cephamycin biosynthetic genes (Martin 1992). It is unclear at this time whether cefG is regulated in a different manner or whether it is inherently less stable than pcbAB,pcbC and ceflEF mRNAS or if its transport from the nucleus is less efficient.
We can not distinguish at this time whether the higher levels of transcripts in cells grown in methio- nine may be also favoured by a lower rate of RNA degradation, although it seems unlikely that degra- dation by ribonucleases would specifically affect the cephalosporin transcripts but not the pyrG transcript.
Methionine also enhances cell differentiation resulting in the formation of fragmented hyphae with swollen cells known as arthrospores (Drew et al. 1976; Nash & Huber 1971). Arthrospores have been reported to show a high specific rate of cephalosporin biosynthe- sis and it is likely that these differentiated cells have a high content of biosynthetic enzymes. The stimu- latory effect of methionine on differentiation together with the inducing effect on the four cephalosporin tran- scripts suggest that methionine may act by changing the level of a high-hierarchy regulatory protein (perhaps a methionine-receptor) that interacts with the promoters or regulatory signals of several genes, i.e. methion- ine acts as a pleiotropic regulatory effector; pleiotropic effectors are well known in antibiotic-producing acti- nomycetes (Martin & Liras 1989). An indirect effect of methionine allowing a better aeration of the cul- ture by decreasing the broth viscosity (due to the for- mation of arthrospores) may also contribute to higher cephalosporin biosynthesis.
Several genes in yeast and other eucaryotic organ- isms are known to be regulated by methionine at the transcription level. The MET14 gene of S. cere- visiae encoding APS kinase (ATP: adenylylsulfate-3'- phosphotransferase, EC 2.7.1.25) (Korck et al. 1991) contains an upstream activating region 'UASmet' iden- tical to that recognized by the yeast centromere and promoter factor CF1 (Baker & Masison 1990; Cai & Davis 1990; Mellor et al. 1990), and also to the yeast acid-phosphatase regulator PHO4 (Vogel et al. 1989) and the protein encoded by the human onco- gene c-myc (Murre et al. 1989). In the MET14 gene deletion or modification of these sequences destroys the methionine effect (Korck et al. 1991). A similar UASmet has been found in several other MET genes of S. cerevisiae (MET2, MET3, MET16, MET25 and
SAM2) (Thomas et al. 1989). The consensus octameric sequence TCACGTGA has been deduced. These UAS are recognized by proteins which belong to the bHLH family; this group of proteins, which include several regulatory factors, contain a basic domain adjacent to a helix-loop-helix motif (Murre et al. 1989) that rec- ognize the consensus sequence CANNTG.
Several sequences similar to CANNTG are present in the upstream region of the pcbAB, pcbC, ce2EF and cefG genes. The presence of these sequences which are recognized by several eucaryotic regulatory factors is intriguing. It is likely that those motifs are recognized by regulatory proteins in A. chrysogenum. Deletion and modification of these putative UASmet is in progress to establish whether the methionine effect at the tran- scriptional level is similar to that observed on the MET genes ofS. cerevisiae.
Stimulation of transcription of pcbAB, pcbC, ce3'EF and cefG by direct interaction of methionine with CANNTG sequences found upstream of these genes in A. chrysogenum is less likely than methionine regu- lation of a single master regulatory protein that inter- acts with those sequences to increase transcription. In summary, the induction by methionine of tran- scription of the four cephalosporin biosynthesis genes and the known effect of this amino acid on differen- tiation of A. chrysogenum indicates that methionine exerts a pleiotropic effect that regulates coordinately cephalosporin biosynthesis and differentiation.
References
Aharonowitz Y, Bergmeyer J, Cantoral JM, Cohen G, Demain AL, Fink U, Kinghom J, Kleinkauf H, MacCabe A, Palissa H, Pfeifer E, Schwecke T, Van Liempt H, yon D0hren H, Wolfe S & Zhang J (1993) 6-(L-c~-aminoadipyl)-L-cysteinyl-D-valine synthetase, the multienzyme integrating the four primary reactions in /3- lactam biosynthesis, as a model peptide synthetase. Biotechnol. 11:807-810
Alvarez E, Cantoral JM, Barredo JL, Dfez B & Martin JF (1987) Purification to homegeneity and characterization of acyl coen- zyme A:6-aminopenicillanic acid aeyltransferase of P. chryso- genum. Antimicrob. Agents Chemother. 31:1675-1682
Alvarez E, Meesschaert B, Montenegro E, Guti6rrez S, Diez B, Barredo JL & Martin JF (1993) The isopenicillin N acyltrans- ferase of P. chrysogenum has isopenicillin N amidohydrolase, 6-aminopenicillanic acid acyltransferase and penicillin amidase activities, all of which are encoded by the single penDE gene. Eur. J. Biochem. 215:323-332
Aplin RT, Baldwin JE, Cole SCJ, Sutherland JD & Tobin MB (1993) On the production of c¢,/3-hetrodimeric acyl-coenzyme A:isopenieillin N acyltransferase of Penicillium chrysogenum: Studies using a recombinant source. FEBS Lett. 319:166-170

Baker RE & Masison DC (1990) Isolation of the gene encoding the Saccharomyces cerevisiae centromere-binding protein CP 1. Mol. Cell Biol. 10:2458-2467
Baldwin JE, Keeping JW, Singh PD & Vallejo CA (1981)Cell-free conversion of isopenicillin N into deacetoxycephalosporin C by Cephalosporium acremonium mutant M-0198. Biochem. J. 194: 649-651
Baldwin JE, Bird JW, Field RA, O'Callaghan NM & Schofield CJ (1990) Isolation and partial characterisation of ACV syn- thetase from Cephalosporium acremonium and Streptomyces clavuligerus. J. Antibiot. 43:1055-1057
Ballance DJ (1986) Sequences important for gene expression in filamentous fungi. Yeast 2:229-236
Barredo JL, Cantoral JM, Alvarez E, Dfez B & Martin JF (1989a) Cloning, sequence analysis and transcriptional study of the isopenicillin N synthase of Penicillium chrysogenum AS-P-78. Mol. Gen. Genet. 216:91-98
Barredo JL, Van Solingen P, Diez B, Alvarez E, Cantoral JM, Kat- tevilder A, Smaal EB, Groenen MAM, Veenstra AE & Martin JF (1989b) Cloning and characterization of acyI-CoA:6-APA acyl- transferase gene of Penicillium chrysogenum. Gene 83:291-300
Cai M & Davis RW (1990) Yeast centromere binding protein CBF1, of the helix-loop-loop protein family, is required for chromosome stability and methionine prototrophy. Cell 61:437-446
Cantoral JM, Guti6rrez S, Fierro E Gil-Espinosa S, Van Liempt H & Martin JF (1993) Biochemical characterization and molecular genetics of nine mutants of Penicillium chrysogenumimpaired in penicillin biosynthesis. J. Biol. Chem. 268:737-744
Carr LG, Skatrud PL, Scheetz ME, Queener SW & Ingolia TD (1986) Cloning and expression of isopenicillin N synthetase gene from Penicillium chrysogenum. Gene 48:257-266
Castro JM, Liras P, L~iiz L & Martin JF (1988) Purification and characterization of the isopenicillin N synthase of Streptomyces lactamdurans. J. Gen. Microbiol. 134:133-141
Coque JJR, Martin JE Calzada JG & Liras P (1991a) The cephamycin biosynthetic genes pcbAB, encoding a large mul- tidomain peptide synthetase, and pcbC of Nocardia lactamdu- rans are clustered together in an organization different from the same genes in Acremonium chrysogenum and Penicillium chryso- genum. Mol. Microbiol. 5:1125-1133
Coque J JR, Liras P & Martin JF (1993a) Characterization and expres- sion in Streptomyces lividans of ceil) and cefE gene from Nocar- dia lactamdurans: The organization of the cephamycin gene clus- ter differs from that in Streptomyces clavuligerus. Mol. Gen. Genet. 236:453-458
Coque JJR, Liras P & Martin JF (1993b) Gene for a/3-1actamase, a penicillin-binding protein and a transmembrane protein are clus- tered with the cephamycin biosynthetic genes in Nocardia lac- tamdurans. EMBO J. 12:631-639
Dayhoff MO, Barker WC & Hardman JK (1976) Atlas of protein sequencing and structure. In: Demain AL (Ed) Atlas of Protein Sequencing and Structure, Vol 5 (pp 53-66). National Biomedical Research Foundation, Washington DC
Demain AL (1983) Biosynthesis of/3-1actam antibiotics, In: Demain AL & Salomon NA (Eds) Antibiotics Containing the/3-Lactam Structure, Vol I (pp 189-228). Springer-Verlag, Berlin
Demain AL & Wolfe S (1987) Biosynthesis of cephalosporins. Dev. Ind. Microbiol. 27:175-182
Diez B, Barredo JL, Alvarez E, Cantoral JM, Van Solingen P, Groe- nen MAM, Veenstra AE & Martin JF (1989) Two genes involved in penicillin biosynthesis are linked in a 5.1 kb SalI fragment in the genome of Penicillium chrysogenum. Mol. Gen. Genet. 218: 572-576
241
Dfez B, Guti6rrez S, Barredo JL, Van Solingen P, Van der Voort LHM & Martin JF (1990) The cluster of penicillin biosynthetic genes. Identification and characterization of the pcbAB gene encoding the c~-aminoadipyl-cysteinyl-valine synthetase and linkage to the pcbC and penDE genes. J. Biol. Chem. 265:16358-16365
Drew SW, Winstanley DJ & Demain AL (1976) Effect of nor- leucine on mycelial fragmentation in Cephalosporium acremoni- urn. Appl. Environ. Microbiol. 31:143-145
Fierro E Guti6rrez S, Diez B & Martin JF (1993) Resolution of four large chromosomes in penicillin-producing filamentous fungi: the penicillin gene cluster is located on chromosome II (9.6 mb) in PeniciUium notatum and chromosome I (10.4 mb) in Penicillium chrysogenum. Mol. Gen. Genet. 241:573-579
Guti6rrez S, Dfez B, Alvarez E, Barredo JL & Martin JF (1991a) Expression of the penDE gene of Penicillium chrysogenum encod- ing isopenicillin N acyltransferase in Cephalosporium acremo- nium: Production of benzylpenicillin by the transformants. Mol. Gen. Genet. 225:56-64
Guti6rrez S, Dfez B, MontenegroE & Martin JF (1991b) Characteri- zation of the CephalosporiumacremoniumpcbAB gene encoding oe-aminoadipyl-cysteinyl-valine synthetase, a large multidomain peptide synthetase: Linkage to the pcbC gene as a cluster of early cephalosporin-biosynthetic genes and evidence of multiple functional domains. J. Bacteriol. 173:2354-2365
Guti6rrez S, Velasco J, Feru~indez FJ & Martin JF (1992) The ceJG gene of Cephalosporium acremonium is linked to the ce3'EF gene and encodes a deacetylcephalosporin C acetyltransferase close- ly related to homoserine O-acetyltransferase. J. Bacteriol. 174: 3056-3064
Hardie DG, Dewart KB, Aitken A & McCarthy AD (1985) Amino acid sequence around the reactive serine residue of the thioesterase domain of rabbit fatty acid synthase. Biochem. Bin- phys. Acta 828:380-382
HSnlinger C & Kubicek CP (1989a) Metabolism and compartmen- tation of ~-aminoadipic acid in penicillin-producing strains of Penicillium chrysogenum. Biochim. Biophys. Acta 993:204-211
H0nlinger C & Kubicek CP (1989b) Regulation of 6-(L-a- aminoadipyl)-L-cysteinyl-D-valine and isopenicillin N biosyn- thesis in PeniciUium chrysogenum by the cr-aminoadipate pool size. FEMS Microbiol. Lett. 65:71-76
Komatsu K-I, Mizuno M & Kodaira R (1975) Effect of methionine on cephalosporin C and penicillin N production by a mutant of Cephalosporiumacremonium. J. Antibiot. 28:881-888
Korck C, Mountain HA & Bystr6m AS (1991) Cloning, nucleotide sequence, and regulation of MET14, the gene encoding the APS kinase of Saccharomyces cerevisiae. Mol. Gen. Genet. 229: 96- 108
Kovacevic S, Tobin MB and Miller JR (1990) The /3-1actam biosynthesis genes for isopenicillin N epimerase and deacetoxy- cephalosporin C synthetase are expressed from a single transcript in Streptomyces clavuligerus. J. Bacteriol. 172:3952-3958
KurzhtkowskiW, Palissa H, Van Liempt H, von D6hren H, Kleinkauf H, Wolf WP & Kurylowicz W (1991 ) Localization ofisopenicillin N synthase in PeniciUium chrysogenum PQ-96. Appl. Microbiol. Biotechnol. 35:517-520
Luengo JM, Revilla G, Villanueva JR & Martin JF (1980) Inhibition and repression of homocitrate synthase by lysine in Penicillium chrysogenum. J. Bacteriol. 144:869-876
MacCabe AP, Riach MBR, Unkles SE & Kinghorn JR (1990) The Aspergillus nidulans npeA locus consists of the three contiguous genes required for penicillin biosynthesis. EMBO J. 9:279-287
MacCabe AE Van Liempt H, Palissa H, Unkles SE, Riach MBR, Pfeifer E, von D6hren H & Kinghorn JR (1991) 6-(L-c~-

242
Aminoadipyl)-L-cysteinyl-D-valine synthetase from Aspergillus nidulans. J. Biol. Chem. 266:12646-12654
Martin JF (1992) Clusters of genes for the biosynthesis of antibiotics: Regulatory genes and overproduction of pharmaceuticals. J. Ind. Microbiol. 9:73-90
Martin JF & Aharonowitz Y (1983) Regulation of biosynthesis of/3-1actam antibiotics. In: Demain AL & Solomon NA (Eds) Antibiotics Containing the Beta-Lactam Structure I (pp 229- 254). Springer-Verlag, Berlin, Heidelberg
Martin JF & Guti6rrez S (1992) Molecular genetics of fungal sec- ondary metabolites. In: Kinghorn JR & Turner G (Eds) Applied Molecular Genetics of Filamentous Fungi (pp 214-252). Blackie and Son Ltd, Glasgow
Martin JF & Liras P (1989a) Enzymes involved in penicillin, cephalosporin and cephamycin biosynthesis. In: Fiechter A (Ed) Advances Biochemical Engineering/Biotechnology, Vol 39 (pp 153-187). Springer Verlag, Berlin, Heidelberg
Martin JF & Liras P (1989b) Organization and expression of genes involved in the biosynthesis of antibiotics and other secondary metabolites. Ann. Rev. Microbiol. 43:173-206
Martin JF, Dfez B, Alvarez E, Barredo JL & Cantoral JM (1987) Development of a transformation system in Penicillium chryso- genum. Cloning of genes involved in penicillin biosynthesis. In: Alacevic M, Hranueli D & Toman Z (Eds) Genetics of Industrial Microorganisms (pp 297-308). Pliva, Zagreb
Martin JE lngolia TD & Queener SW (1991) Molecular genetics of penicillin and cephalosporin antibiotic biosynthesis. In: Leong S & Berka RM (Eds) Molecular Industrial Mycology. Systems and Applications for Filamentous Fungi (pp 149-196). Marcel Dekker, Inc., New York
Martin JF, Gutitrrez S, Montenegro E, Coque JJR, Fern~Jldez FJ, Velasco J, Gil S, Fierro F, Calzada JG, Cardoza RE & Liras P (1992) Genes, enzymes and control of the production of/3- lactam antibiotics. In: Ladisch MR & Bose A (Eds) Harnessing Biotechnology for the 21st Century. Proceedings of the Ninth International Biotechnology Symposium (pp 131-137). Ameri- can Chemiccal Society, Washington DC
Mathison L, Soliday C, Stepan T, Aldrich T & Rambosek J (1993) Cloning, characterization, and use in strain improvement of the Cephalosporium acremonium gene cejG encoding acetyltrans- ferase. Curr. Genet. 23:33-41
Matsuda A, Sugiura H, Matsuyama K, Matsumoto H, Ichikawa S & Komatsu K-I (1992) Molecular cloning of aceylt coen- zyme A:deacetylcephalosporin C O-acetyltransferase cDNA from Acremonium chrysogenum: Sequence and expression of catalytic activity in yeast. Biochem. Biophys. Res. Comm. 182: 995-1001
Matsuyama K, Matsumoto H, Matsuda A, Sugiura H, Komat- su K-I & Ichikawa S (1992) Purification of acetyl coenzyme A:deacetylacephalosporin C O-acetyltransferase from Acremo- nium chrysogenum. Biosci. Biotech. Biochem. 56:1410-1412
Mellor J, Jiang W, Funk M, Rathjen J, Barnes CA, Hinz T, Hegemann JH & Philippsen P (1990) CPF1, a yeast protein which functions in centromeres and promoters. EMBO J. 9:4017-4026
Montenegro E, Barredo JL, Gutitrrez S, Dfez B, Alvarez E & Martin JF (1990) Cloning, characterization of the acyl-CoA:6-amino penicillanic acid acyltransferase gene ofAspergillus nidulans and linkage to the isopenicillin N synthase gene. Mol. Gen. Genet. 221:322-330
Montenegro E, Fierro F, Fernandez FJ, Gutitrrez S & Martin JF (1992) Resolution of chromosomes III and VI of Aspergillus nidulans by pulsed-field gel electrophoresis shows hat the peni- cillin biosynthetic pathway genes pcbAB, pcbC, and penDE are
clustered on chromosome VI (3.0 megabases). J. Bacteriol. 174: 7063-7067
Miiller WH, Van der Krft TP, Krouwer AJJ, Wtsten HAB, Van der Voort LHM, Smaal EB & Verkleij AJ (1991) Localization of the pathway of the penicillin biosynthesis in PeniciUium chryso- genum. EMBO J. 10:489-495
Miiller WH, Bovenberg RAL, Groothuis MH, Kattevilder F, Smaal EB, Van der Voort LHM & Verkleij AJ (1992) Involvement of microbodies in penicillin biosynthesis, Biochim. Biophys. Acta 1116:210-213
Murre C, McCaw PS & Baltimore D (1989) A new DNA bind- ing and dimerization motif in immunoglobulin enhancer binding, daughterless, MyoD, and myc proteins, Cell 56:777-783
Naggert J, Witkowski A, Mikkelsen J & Smith S (1988) Molecular cloning and sequencing of a cDNA encoding the thioesterase domain of the rat fatty acid synthetase. J. Biol. Chem. 263:1146- 1150
Nash CH & Huber FM ( 1971) Antibiotic synthesis and morphologi- cal differentiation of Cephalosporiumacremonium. Appl. Micro- biol. 22:6-10
Niiesch J, Helm J & Treichler H-J (1987) The biosynthesis of sulfur- containing/3-1attain antibiotics. Ann. Rev. Microbiol. 41: 51-57
Poulose AJ, Rogers L & Kolattukudy PE (1981) Primary structure of a chymotyptic peptide containing the 'active serine' of the thioesterase domain of fatty acid synthetase. Biochem. Biophys. Res. Comm. 103:377-383
Queener SW & Neuss N (1982) The biosynthesis of beta-lactam antibiotics. In: Morin RB & Gorman M (Eds) Chemistry and Biology of Beta-Lactam Antibiotics, Vol 3 (pp 1-82). Academic Press, New York
Ram6n D, Carramolino L, Patino C, Stlnchez F & Pefialva MA (1987) Cloning and characterization of the isopenicillin N synthetase gen mediating the formation of the/3-1actam ring in Aspergillus nidulans. Gene 57:171-181
Ramos FR, L6pez-Nieto MJ & Martin JF (1985) Isopenicillin N synthetase of Penicillium chrysogenum, an enzyme that con- verts 6-(L-c~-aminoadipyl)-L-cysteinyl-D-valine to isopenicillin N. Antimicrob. Agents Chemother. 27:380-387
Ramos FR, Ltpez-Nieto MJ & Martin JF (1986) Coordinate increase of isopenicillin N synthetase, isopenicillin N epimerase and deacetoxycephalosporin C synthetase in a high cephalosporin- producing mutant of Acremonium chrysogenum and simultane- ous loss of the three enzymes in a non-producing mutant. FEMS Microbiol. Lett. 35:123-127
Ramsdem M, McQuade BA, Saunders K, Turner MK & Hafford S (1989) Characterization of a loss-of-function mutation in the isopenicillin N synthase gene ofAcremonium ehrysogenum. Gene 85:267-273
Randhawa ZI, Naggert J, Blacher RW & Smith S (1987) Amino acid sequence of the serine active-site region of the medium-chain S- acyl fatty acid synthetase thioester hydrolase from rat mammary gland. Eur. J. Biochem. 162:577-581
Revilla G, Ramos FR, Ltpez-Nieto MJ, Alvarez E & Martin JF (1986) Glucose represses formation of 6-(L-c~-aminoadipyl)-L- cysteinyl-D-valine and isopenicillin N synthase but not penicillin acyltransferase in Penicillium ehrysogenurn. J. Bacteriol. 168: 947-952
Samson SM, Belagaje R, Blankenship DT, Champman JL, Perry D, Skatrud PL, Van Frank RM, Abraham EP, Baldwin JE, Queen- er SW & Ingolia TD (1985) Isolation, sequence determination and expression in E. coli of the isopenicillin N synthetase from Cephalosporiumacremonium. Nature 318:191-194
Samson SM, Chapman JL, Belagaje P, Queener SW & Ingolia TD (1987a) Analysis of the role of cysteine residues in isopenicillin

N synthetasc activity by site-directed mutagenesis. Proc. Natl. Acad. Sci. USA 84:5705-5709
Samson SM, Dotzlaf JE, Slisz ML, Becker GW, Van Frank RM, Veal LE, Yeh WK, Miller JR, Queener SW & lngolia TD (1987b) Cloning and expression of the fungal expandase/hydroxylase gene involved in cephalosporin biosynthesis. Biotechnol. 5: 1207-1216
Sawada Y, KonomiT, Solomon NA & Demain AL (1980) Increase in activity of/3-1actam synthetases after growth of Cephalosporium acremonium with methionine or norleucine. FEMS Microbiol. Lett. 9:281-284
Siewinski M, Kuropatwa M & Szewczuk A (1984) Phenylalkylsul- fonyl derivatives as covalent inhibitors of penicillin amidase. Z. Physiol. Chem. 365:829-837
Skatrud PL & Queener SW (1989) An electrophoretic molecular karyotype for an industrial strain of Cephalosporium acremoni- urn. Gene 78:331-338
Smith D J, Burnham MKR, Bull JH, Hodgson JE, Ward JM, Browne P, Brown J, Barton B, Earl AJ & Turner G (1990a)/3-Lactam antibiotic biosynthetic genes have been conserved in clusters in prokaryotes and eukaryotes. EMBO J. 9:741-747
Smith DJ, Burnham MKR, Edwards J, Earl AJ & Turner G (1990b) Cloning and heterologous expression of the penicillin biosyn- thetic gene cluster from Penicillium chrysogenum. Biotechnol. 8: 39-41
Smith AW, Collis K, Ramsden M, Fox HM & Peberdy JE (1991) Chromosome rearrangements in improved cephalosporin C pro- ducing strains of Acremonium chrysogenum. Curr. Genet. 19: 235-237
Somerson NL, Demain AL & Nunheimer TD (1961) Reversal of lysine inhibition of penicillin production by a-aminoadipic acid. Arch. Biochem. Biophys. 93:238-241
Thomas D, Cherest H & Surdin-Kerjan Y (1989) Elements involved in S-adenosylmethionine-mediated regulation of the Saccha- romyces cerevisiae MET25 gene. Mol. Cell Biol. 9:3292-3298
Tobin MB, Kovacevic S, Madduri K, Hoskins JA, Skatrud PL, Vin- ing LC, Stuttard C & Miller JR (1991) Localization of lysine e-aminotransferase (lat) and 6-(L-a-aminoadipyl)-L-cysteinyl- D-valine (ACV) synthetase (pcbAB) genes in Streptomyces
243
clavuligerus and production of lysine e-aminotransferase activity in Escherichia coli. J. BacterioL 173:6223-6229
Tobin MB, Baldwin JE, Cole SCJ, Miller JR, Skatrud PL & Sutherland D (1993) The requirement for subunit interaction in the production of Pencillium chrysogenum acyl-coenzyme A:isopenicillin N acyltransferase in Escherichia coil Gene 132: 199-206
Turgay K, Krause M & Marahiel MA (1992) Four homologous domains in the primary structure of GrsB are related to domains in a superfamily of adenylate forming enzymes. Mol. Microbiol. 6:529-546
Veenstra AE, Van Solingen P, Huninga-Muurling H, Koekman BE Groenen MAM, Smaal EB, Kattevilder A, Alvarez E, Barredo JL & Martin JF (I 989) Cloning of penicillin biosynthetic genes. In: Hershberger CL, Queener SW & Hegeman G (Eds) Genetics and Molecular Biology of Industrial Microoganisms (pp 262-269). American Society for Microbiology, Washington DC
Velasco J, Gutirrrez S, Fernfindez FJ, Marcos AT, Aren6s C & Martfn JF (l 994) Exogenous methionine increases levels ofmRNAs tran- scribed from pcbAB, pcbC, and ceyEF gene, encoding enzymes of the cephalosporin biosynthetic pathway, in Acremonium chryso- genum. J. Bacteriol. 176, in press
Vogel K, H6rz W & Hinnen A (1989) The two positively acting regulatory proteins PHO2 and PHO4 physically interact with PHO5 upstream activation regions. Mol. Cell Biol. 9:2050--2057
Whiteman PA, Abraham EE Baldwin JW, Fleming MD, Schofield CJ, Sutherland JD & Willis AC (1990) Acyl-coenzyme A:6- aminopenicillanic acid acyltransferase from Penicillium chryso- genum and Aspergillus nidulans. FEMS Lett. 262:342-344
Yuan Z, Liu W & Hammes GG (1988) Molecular cloning and sequencing of DNA complementary to chicken liver fatty acid synthase mRNA. Proc. Natl. Acad. Sci. USA 85:6328-6331
Zanca DM & Martin JF (1983) Carbon catabolite regulation of the conversion of penicillin N into cephalosporin C. J. Antibiot. 36: 700-708
Zhang J-Y, Banko G, Wolfe S & Demain AL (1987) Methionine induction of ACV synthetase in Cephalosporium acremonium. J. Ind. Microbiol. 2:251-255




















