
Potential Occurrence of MTBE and BTEX in Groundwater Resources of Amman-Zarqa Basin, Jordan
Upload
independentCategory
view
4download
0
MINI-REVIEW
Enzymes and genes involved in the aerobic biodegradationof methyl tert-butyl ether (MTBE)
Nicolas Lopes Ferreira & Cédric Malandain &
Françoise Fayolle-Guichard
Received: 3 April 2006 /Revised: 4 May 2006 /Accepted: 4 May 2006 /Published online: 28 June 2006# Springer-Verlag 2006
Abstract Fuel oxygenates, mainly methyl tert-butyl ether(MTBE) but also ethyl tert-butyl ether (ETBE), are addedto gasoline in replacement of lead tetraethyl to enhance itsoctane index. Their addition also improves the combustionefficiency and therefore decreases the emission of pollu-tants (CO and hydrocarbons). On the other hand, MTBE,being highly soluble in water and recalcitrant to biodegra-dation, is a major pollutant of water in aquifers contami-nated by MTBE-supplemented gasoline during accidentalrelease. MTBE was shown to be degraded throughcometabolic oxidation or to be used as a carbon and energysource by a few microorganisms. We have summarized thepresent state of knowledge about the microorganismsinvolved in MTBE degradation and the MTBE catabolicpathways. The role of the different enzymes is discussed aswell as the rare and recent data concerning the genesencoding the enzymes involved in the MTBE pathway. Thephylogeny of the microorganisms isolated for their capacityto grow on MTBE is also described.
Introduction
Petroleum compounds are the most widely used chemicalsworldwide and their use requires numerous transportationand storage facilities. Such an intensive and disseminateduse leads to numerous cases of spillage resulting ingroundwater contamination (Johnson et al. 2000). Petro-leum compounds are actually the most frequently found
contaminants of groundwater and among them is gasoline(world consumption in 2003–2004 was 11,675 Mt). Gaso-line is a complex mixture of numerous individual nativechemicals. Other chemicals are also added to gasoline tomeet specific requirements. For instance, lead tetraethyl hasbeen added to gasoline for 50 years to reach thespecifications regarding the octane index. In the 1970s,lead was banned from gasoline formulation because of itsdetrimental effect on catalytic converters fitted on exhaustpipes. Oxygenates were then chosen by refiners inreplacement of lead because of their high octane index.Moreover, the use of methyl tert-butyl ether (MTBE), ethyltert-butyl ether (ETBE), or tert-amyl methyl ether (TAME)was also of interest because they reduced the emission ofpollutants (CO and hydrocarbons) in exhaust gases. MTBE,added to gasoline at a maximal concentration of 15% (on avolume basis), was mostly used, with ETBE being alsoused in Europe (for details, see Fayolle and Monot 2005).
The high solubility of these compounds (ca. 40 and10 g l−1 for MTBE and ETBE, respectively) makes them themost water-soluble compounds of reformulated gasoline.The environmental impact of ether fuels on groundwaterwas not estimated before their utilization. Studies carriedout years after the beginning of their current use showedthat MTBE was detected at various concentrations innumerous aquifers in the USA (Johnson et al. 2000). Whenan MTBE-supplemented gasoline is spilled, the plumesgenerated by MTBE are indeed much larger than thosegenerated by benzene (water solubility=1.755 g l−1).According to Wilson (2003), the retardation factor (ratio ofwater speed to pollutant speed) was 1.8 for benzene and 1.1for MTBE, which means that MTBE migrates practically asrapidly as the aquifer. This is the consequence of (1) its lowadsorption onto organic matter, (2) its high solubility, and(3) its recalcitrance to biodegradation. Thus, the half-life of
Appl Microbiol Biotechnol (2006) 72:252–262DOI 10.1007/s00253-006-0494-3
N. Lopes Ferreira :C. Malandain : F. Fayolle-Guichard (*)Biotechnology and Biomass Chemistry Department,Institut Français du Pétrole,1-4 avenue de Bois-Préau,92852 Rueil-Malmaison, Francee-mail: [email protected]

MTBE in groundwater is estimated at 2–3 years (Wilson2003), whereas that of benzene is 2–3 months under similarconditions.
The isolation of microorganisms exhibiting degradationcapacities towards ether fuels was crucial because a study onthem could help in determining limitations to their biodegra-dation. Moreover, the characterization of the enzymesinvolved in MTBE biodegradation and the isolation of relatedgenes can provide useful tools for assessing the potential fornatural attenuation in ether-fuel-contaminated sites.
MTBE was shown to be degraded either by cometabolismor by being used as a sole carbon and energy source by afew microorganisms (Fayolle and Monot 2005). We sum-marize in this paper the present knowledge on the micro-organisms following these two ways of biodegradation andon the enzymes and genes involved in MTBE degradation.
Cometabolic MTBE oxidation
Several microorganisms were able to degrade MTBEcometabolically (for an exhaustive list, see Fayolle andMonot 2005). In aerobiosis, the initial attack of the MTBE
molecule is performed by a monooxygenase whose naturewas only rarely demonstrated. However, well-characterizedmonooxygenases, both non-hemic and hemic, were shownto be able to oxidize MTBE after growth on differentsubstrates. These enzymes catalyze the following reaction.
Cytochrome P450 monooxygenases
Two different cytochrome P450 monooxygenases wereshown to be able to catalyse the oxidation of MTBE.
– The cytochrome P450 encoded by the eth genes inseveral strains able to grow on ETBE, Rhodococcusruber IFP 2001 (Chauvaux et al. 2001; Hernandez-Perez et al. 2001) and also Rhodococcus zopfii IFP
Fig. 2 Comparison of the structure of the eth locus in R. ruber IFP2001, R. zopfii IFP 2005, and Mycobacterium sp. IFP 2009 (takenfrom Béguin et al. 2003). The structure of eth genes in R. ruber IFP2001 is detailed. The insert shows that the rightmost extremity of the
right transposon is deleted in R. zopfii IFP 2005. An IS3 insertionsequence, which is not linked to the eth locus, is present within thegenome of Mycobacterium sp. IFP 2009
CH3
CH3 CH3C O
Methyl tert-butyl ether
(MTBE)
CH3
CH3
CH3 CH2C O
CH3
O2 H2O
Tert-butoxy methanol
OH
2 [H]
Ferredoxinreductase(ethA)
cytochromeP-450 (ethB)
ROH
2R-H
O
H2O
Ferredoxin(ethC)
Fe3+
Fe3+
Fe2+
FADH2
NADH + H+
NAD+
FAD+Fig. 1 Electron chain transferduring oxidation of MTBE by acytochrome P450 monooxyge-nase
Appl Microbiol Biotechnol (2006) 72:252–262 253

2005 and Mycobacterium sp. IFP 2009 (Béguin et al.2003).
– The cytochrome P450 induced in Pseudomonas putidaCAM ATCC 17453 during growth on camphor (Steffanet al. 1997)
Microorganisms growing on ETBE
R. ruber IFP 2001 was isolated for its capacity to grow onETBE with a concomitant accumulation of tert-butylalcohol (TBA). R. ruber IFP 2001 was also able to degradeMTBE and TAME after growth on ETBE (Hernandez-Perez et al. 2001). An ETBE− mutant, R. ruber IFP 2006,was obtained which had lost both the capacity to grow onETBE and to degrade MTBE (Chauvaux et al. 2001; Urioset al. 2002). A cluster of genes (eth genes) was isolated andcharacterized by Chauvaux et al. (2001). The putativeelectron transfer chain leading to the oxidation of MTBE byeth genes is presented in Fig. 1.
The different genes, ethRABCD, encode a putativeregulator (similar to the AraC/XylS family positive regula-tor), a ferredoxin reductase, a cytochrome P450 (43 kDa), aferredoxin, and an unknown protein (10 kDa), respectively(Fig. 2).The product of ethD gene was shown to be essentialfor the activity as the transfer of only ethRABC to R. ruberIFP 2006 (ETBE−) did not restore the wild-type phenotype.In contrast, the transfer of the whole ethRABCD operon toR. ruber IFP 2006 restored the wild-type phenotype. The ethcluster is highly similar to the thc cluster of Rhodococcussp. NI86/21 responsible for N-dealkylation of S-ethyldipropylthiocarbamate (Nagy et al. 1995).
The eth gene cluster in R. ruber IFP 2001 is locatedbetween two identical repeats (5.6 kb) including class II
transposons (Fig. 2). A unique recombination event occur-ring between the two identical repeats can lead to the lossof the eth genes, which occurs at a high frequency (Urios etal. 2002).
A new intermediate of the ETBE pathway, tert-butylacetate (TBAc), was recently characterized during growthon ETBE of R. ruber IFP 2001 and its constitutivederivative IFP 2007. Furthermore, R. ruber IFP 2006(ETBE−) was also shown to be able to grow on TBAc,with concomitant accumulation of TBA. All these strainswere able to growth on acetate (Malandain, unpublishedresults). These results demonstrated that the reactionscheme involves (1) a sub-terminal oxidation of the ETBEethoxy moiety, via the tert-butoxy ethanol and (2) hydro-lysis by an esterase activity (Fig. 3). The production of tert-butyl formate (TBF), the homolog of TBAc, was notreported during MTBE degradation, and this point woulddeserve further investigation. However, formate was tran-siently detected (Hernandez-Perez et al. 2001), mostprobably as the product of TBF hydrolysis.
Béguin et al. (2003), using both Southern hybridizationand specific polymerase chain reaction amplifications,showed that genes highly similar to eth genes were alsofound in two other bacteria isolated for their capacity to growon ETBE, R. zopfii IFP 2005 and Mycobacterium sp. IFP2009. The rate of mutations did not exceed 0.1% throughoutthe locus by comparison to R. ruber IFP 2001 eth genes. Thecapacity to grow on ETBE was also unstable in both strains.The context of insertion in the genome was different amongthe three strains (Fig. 2). R. ruber IFP 2001 and R. zopfii IFP2005 were isolated from the same sludge whereas Mycobac-terium sp. IFP 2009 was from another origin. The presenceof extremely conserved eth loci located amongst otherwise
H3C
CH3
CH3
CH3CH2C O H3C
CH3
CH3
CH3 CH2 C O
OH
H3C
CH3
CH3
CH3 C C O
O
H3C
CH3
CH3
C OH
CH3 C - O
O CO2
2 [H]
O2 H2O
2 [H]
Biomass
Ethyl tert-butyl ether Tert-butoxy ethanol
Tert-butyl acetate
Not metabolized Acetate
eth genes
Cyt P450
Esterase
Dehydrogenase?
Tert-butyl alcohol
Fig. 3 ETBE catabolic pathwayin R. ruber IFP 2001. Theintermediates and enzymes ac-tivities identified are in bold
254 Appl Microbiol Biotechnol (2006) 72:252–262

unrelated sequences in R. zopfii IFP 2005 and Mycobacte-rium sp. IFP 2009 indeed strongly suggests that interspecificgene transfer does occur, allowing the dissemination of theeth genes through horizontal gene transfer.
It is interesting to note that R. ruber IFP 2007, a mutantconstitutively expressing the eth genes, was easily obtainedfrom R. ruber IFP 2001 by successive transfer on ETBE(Hernandez-Perez et al. 2001). If such deregulated ethgenes inserted next to transposons happen to be transferredto strains that are able to grow on TBA, very efficientstrains that are able to grow on MTBE might emerge underselective pressure in MTBE-contaminated environments.Strains able to growth on TBA are probably not scarce(Fayolle, unpublished results; Piveteau et al. 2001;Rohwerder 2006).
It would be interesting to compare the degradationcapacity and the genetic content of R. ruber IFP 2001 tothose of the Rhodococcus species isolated from MTBE-degrading culture BC-1 (Salanitro et al. 1994), Bc663, and10bc312T, recently identified by Goodfellow et al. (2004)as R. aetherivorans sp. nov., the 16S rRNA genes of whichshowed a strong similarity with that of R. ruber.
It is also interesting to note that a gene analog to ethA(encoding the ferredoxin reductase in R. ruber IFP 2001)was found in the annotated genome of Methylibiumpetroleiphilum PM1 (Gene MpeB0597) with a significantlyhigh identity of 49% (e value 1e−96). This strain is one ofthe few strains able to grow on MTBE. Nevertheless, noanalogs of the other genes of the eth cluster were detectedin the vicinity.
Microorganism growing on camphor
Resting cells of P. putida CAM (ATCC 17453), which hasthe capacity to oxidize camphor, thanks to a cytochromeP450cam (Aramaki et al. 1993), were shown able to oxidizeMTBE (Steffan et al. 1997) after growth on camphor butnot after growth on a rich medium. Cytochrome P450 wasresponsible for the initial oxidation during growth oncamphor (Aramaki et al. 1993; Unger et al. 1986).
The genes responsible for these reaction scheme arelocated on the CAM plasmid (Aramaki et al. 1993). Theyencode a repressor, a dehydrogenase (see above), acytochrome P450, a putidaredoxin reductase, and a putidar-edoxin (Fig. 4). In this case, the involvement of the camgenes in MTBE oxidation was not genetically proven bycomparison to CAM plasmid-cured cells.
The three-dimensional structure of the cytochrome P450CamC was determined by Poulos et al. (1985). We havecompared the sequences of cytochromes P450 EthB andCamC which both oxidize MTBE with that of cytochromeP450soy of Streptomyces griseus (ATCC13273) which isunable to degrade MTBE (Liu et al. 2001). Invariant aminoacids found among all cytochrome P450 are conserved inthe three enzymes. The comparison of the secondarystructures revealed a strong conservation of motifs involvedin the active site. The only exception is a valine residue atposition 295, well conserved in CAM C and EthB butwhich was not detected in cytochrome P450soy. This amino
acid is involved in the accessibility to the substrate and thisdifference could potentially explain the incapacity ofMTBE to have an access to the active site of P450soy.
CH3
CH3 CH2C O
CH3
Tert-butoxy methanol
OH CH3
CH3 CC O
CH3
Tert-butyl formate
(TBF)
NAD(P)+
NAD(P)H + H+
O
H
camR camD camC camA camB
CamR Dehydrogenase Cytochrome Putidaredoxin Putidaredoxin(repressor) P450 reductase
(20.4 kDa) (38.4 kDa) (46.5 kDa) (45.6 kDa) (11.4 kDa)
Fig. 4 Structure of the camgenes (cytochrome P450cam) inP. putida CAM (ATCC 17453)(modified from Aramaki et al.1993)
Cytochrome P450 cam Dehydrogenas
O
O2
NADH
O
OH
D-camphor 5-exo-hydroxycamphor
NADNADH
Cytochrome P450 cam Dehydrogenase
O
O2
NADH
O O
OH
O
D-camphor 5-exo-hydroxycamphor 2,5-diketocamphane
NADNADH
Appl Microbiol Biotechnol (2006) 72:252–262 255

It is interesting to note that the CAM operon includes adehydrogenase responsible for the second step, generating2,5-diketocamphane from 5-exo-hydroxycamphor. A simi-lar enzyme may account for the oxidation of tert-butox-yethanol to TBAc during ETBE degradation and, possibly,of tert-butoxymethanol to tert-butyl formate during MTBEdegradation.
Using P. putida CAM, Steffan et al. (1997) showed that25% of the [14C]-MTBE was converted to CO2, which wasdue to the mineralization of the C1-compound formedeither from hydrolysis of TBF (formate) or by directdismutation (formaldehyde).
Alkane hydroxylase
P. putida GPo1 was isolated for its capacity to grow onvarious n-alkanes (C5 to C12) (Baptist et al. 1963). It wasoriginally reported that this strain was unable to degradeMTBE (Steffan et al. 1997). However, Smith and Hyman(2004) clearly demonstrated more recently that P. putidaGPo1 could degrade MTBE after growth on octane. Nodegradation of TBA was observed and the Ks value forMTBE was high (20–40 mM). TBF was not detected as anintermediate. The low affinity for MTBE could explain whyMTBE degradation failed to be detected previously. Theinduction of MTBE oxidation activity by octane suggestedthat, in P. putida GPo1, MTBE oxidation may be performedby enzymes involved in alkane degradation. This wasconfirmed by showing that an OCT plasmid-cured variant,P. putida GPo12, previously isolated (Chen et al. 1996) nolonger oxidized MTBE (Smith and Hyman 2004).
The alkane degradation genes of P. putida GPo1 areorganized in two operons, alkBFGHJKL operon (Eggink etal. 1987) and the alkST region (Eggink et al. 1988) whichare on the plasmid OCT. The non-heme monooxygenase(AlkB), located in the membrane, is involved in an electrontransfer chain, including also a rubredoxin with a non-hemeiron atom and a FAD-dependent rubredoxin reductase
which transfer electron from the dioxygen to the substrate(Fig. 5). This enzyme system is able to oxidize a widerange of substrates (van Beilen et al. 1994). The specificityof the enzyme toward n-alkanes of different chain lengthscould be dramatically affected by a transition at the position55 or 58 from tryptophane to serine or cysteine (van Beilenet al. 2005). It was even shown, very recently, that P. putidaGPo1 was able to grow on propane and butane due to theactivity of AlkB (Johnson and Hyman 2006).
The capacity of alkane hydroxylase to oxidize MTBEcould also be inferred from the study of Pseudomonasmendocina KR-1 by Smith et al. (2003b). The growth ofP. mendocina KR-1 on n-alkanes (C5 to C8), but not ontoluene (Steffan et al. 1997), was found to promote theability to oxidize MTBE, suggesting that the reaction maybe performed by an alkane hydroxylase even if, to ourknowledge, the presence of an alkB gene has not beendemonstrated in this strain. In this case, the transientproduction of tert-butyl formate was not detected. TBAaccumulated and was not further oxidized. The possibilitythat TBF was produced but very rapidly hydrolyzed by anefficient esterase was ruled out because the resting cells ofP. mendocina KR-1 were not able to hydrolyze TBF. Thisscheme necessarily involves the production of formalde-hyde, which was detected at a low level during MTBEoxidation. Moreover, the strain was able to degradeformaldehyde.
The involvement of an alkane monooxygenase in MTBEoxidation by Mycobacterium vaccae JOB5 (recently re-classified Mycobacterium austroafricanum ATCC 29678)was proposed by Smith et al. (2003a). The role of such analkane monooxygenase with oxidation capacity towardsMTBE would not be negligible in MTBE-contaminatedenvironments as Johnson et al. (2004) demonstrated thatMTBE itself was an inducer of the MTBE oxidationcapacity in strain JOB5. An investigation on the capacityof various alkane hydroxylases to degrade MTBE iscertainly of interest in view of these results.
R CH3
RubredoxinFe3+
H2O
R CH2OH
O2
RubredoxinFe2+
Reductase- FADH2
Reductase- FAD
NAD+
NADH + H+
AlkB AlkG AlkT
Alkane hydroxylase RubredoxinRubredoxinreductase
Fig. 5 Electron chain transferduring oxidation of MTBE byalkane hydroxylase of P. putidaGPo1
256 Appl Microbiol Biotechnol (2006) 72:252–262
2-Hydroxyisobutyric acid (HIBA)
Tert-butyl alcohol (TBA)
2-Methyl 1,2 propanediol(2-M1,2-PD)
CO2
HydroxyisobutyraldehydeAcetone
CH 3
C H 3 CC O
CH 3
O
H
CH3
CH3 C OH
CH3
O2
CH3
CH3
CH2
C OH
OH
Tert-butoxy methanolMethyl tert-butyl ether(MTBE)
NAD(P)+
CH3
CH3 CH3C O
CH3
CH3
CH3 CH2C O
CH3
OH
H2O
Formate
CO2
Tert-butyl formate (TBF)
NAD(P) +NAD(P)H+ H+
CO
OHH
NAD+
NAD(P)H + H+
H2O
NAD(P)H + H+
NAD+
CH3
CH3 C
OH
CO
H
NAD(P)+NAD(P)H
+ H+
H20
CH3
CH3 CC
OH
O
OH
CH3
C O
CH3
CO
HH
Formaldehyde
CO2
CH3
H2C C OHC
O
CoA-S
H3-Hydroxybutyryl-CoA
CoAS _ HATP
CO2
BIOMASS
NAD(P)H+ H+
O2 H2O
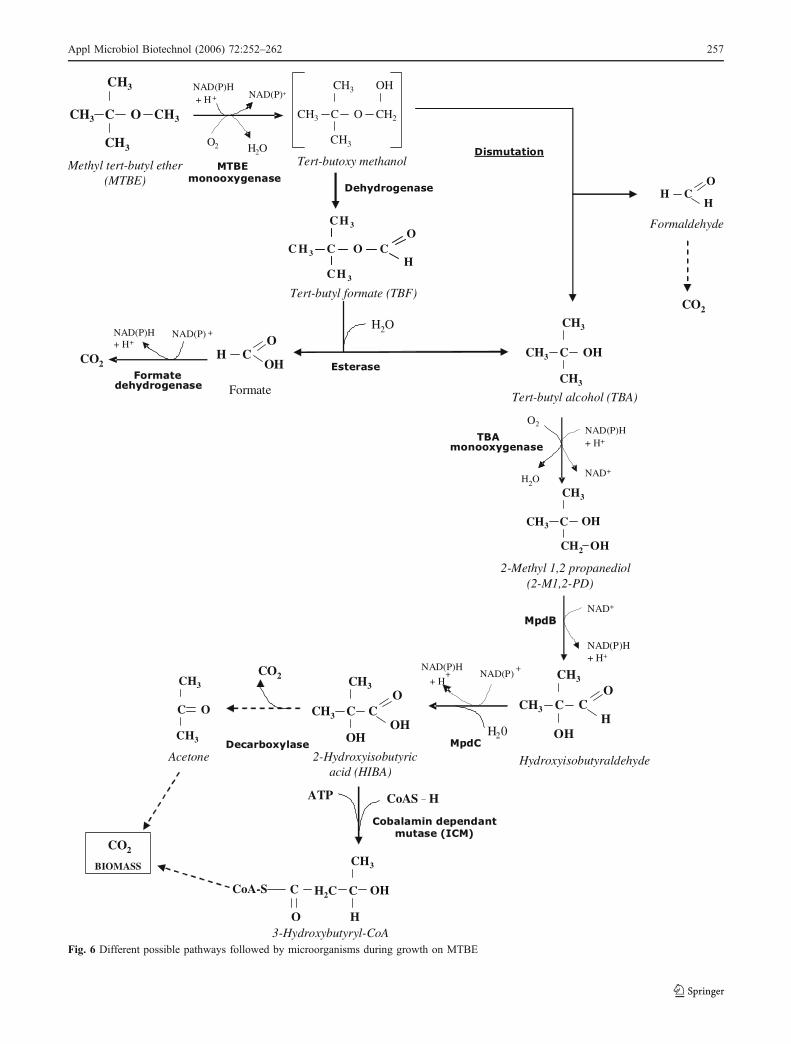
Fig. 6 Different possible pathways followed by microorganisms during growth on MTBE
Appl Microbiol Biotechnol (2006) 72:252–262 257

MTBE as a growth substrate
We will present the different possible metabolic pathwaysthat were demonstrated by the isolation of several MTBEdegradation intermediates during its metabolism. We willthen try to build a phylogenetic tree of the different knownmicroorganisms able to grow on MTBE. Finally, we willfocus on the present knowledge on enzymes and genesinvolved in the MTBE metabolic pathway.
Metabolic pathways and phylogeny of the microorganisms
The different MTBE degradation pathways proposed aresummarized in Fig. 6. Hanson et al. (1999) isolated aGram-negative bacterium, PM1, from an MTBE-fed bio-filter. PM1 is able to grow on MTBE with a low biomassyield (0.18 g dry weight g−1 MTBE). The genome of thisstrain has been sequenced (http://genome.jgi-psf.org/finished_microbes/metpe/metpe.home.html) and strainPM1 was recently identified as M. petroleiphilum PM1T
gen. nov., sp. nov. It is a facultative methylotroph whichbelongs to the beta-Protobacteria in the Sphaerotilus–Leptothrix group (Nakatsu et al. 2006). No TBA accumu-lated during growth on MTBE, even when it was providedat a concentration of 500 mg l−1 (Hanson et al. 1999). Thestrain was able to grow on TBF, TBA, and hydroxyisobu-tyric acid (HIBA) (Deeb et al. 2000).
Hydrogenophaga flava ENV735, another Gram-negativebacterium belonging to the beta-Proteobacteria, was alsoisolated for its capacity to grow on MTBE (Steffan et al.2000) and was characterized by Hatzinger et al. (2001). TBAand HIBAwere detected as degradation intermediates duringgrowth on MTBE, whereas no TBF could be detected, whichwould suggest that tert-butoxy methanol undergoes a directdismutation to TBA and formaldehyde (Fig. 6).
Mycobacterium austroafricanum IFP 2012 was isolatedfrom an activated sludge for its capacity to grow on TBAand was then shown to grow on MTBE (François et al.2002). M. austroafricanum IFP 2015 was isolated fromthe drain water of an MTBE-supplemented gasoline storagetank for its capacity to grow on MTBE (Lopes Ferreira etal. 2006b). In these strains, several intermediates of MTBEdegradation, TBF, TBA, 2-methyl 1,2-propanediol(2-M1,2-PD), HIBA, and acetone, were detected (Fran-çois et al. 2002, 2003; Lopes Ferreira et al. 2006b).
M. vaccae JOB5 (M. austroafricanum ATCC 29678),isolated long ago on 2-methylbutane (Ooyama and Foster1965), is able to oxidize MTBE and TBA (Hyman andO’Reilly 1999; Johnson et al. 2004; Smith et al. 2003a).2-M1,2-PD, HIBA, and hydroxyacetone (Hyman et al.2004) were detected as intermediates in MTBE degrada-tion. Nevertheless, Smith et al. (2003a) stated that neitherMTBE nor TBF nor TBA could sustain the growth of thisstrain.
Strain L108, which grows on MTBE, was isolatedfrom a MTBE-contaminated site in Germany (Rohwerderet al. 2004) and was further characterized as a beta-Proteobacteria (Rohwerder et al. 2006). It is interesting tonote that this strain showed the highest 16S rRNA genesimilarity (99.9% identity) with strain I-2052 (or IFP2003), a methylotrophic organism isolated from activatedsludge that grow on TBA but not on MTBE (Piveteau et al.2001).
Other strains able to grow on MTBE, UC1, UC2, andUC3, were isolated from a MTBE-degrading biofilter byPruden and Suidan (2004). Strains ENV472 and ENV736,able to grow on MTBE, were also isolated (R Steffan,P. Hatzinger, S. Vainberg, personal communication).
From the 16S rRNA gene sequences of these differentstrains, a phylogenetic tree was built to determine their
UC1
UC2
ENV 472
PM1
ENV 736
L10 / L 108
I-2052 (IFP 2003)
ENV735
UC3
IFP2015
JOB5
IFP20120.05
0.038
0.033
0.063
0.2820.031
0.031
L
-
.
Fig. 7 Phylogenetic tree of thedifferent strains able to grow onMTBE (or TBA). The phyloge-netic tree of 16S rRNA genesequences is represented usingthe “midpoint rooting” option.The tree was built using theneighbor-joining method ofSaitou and Nei (1987) and thedistance method of Tamura andNei (1993). Boostrap values of>70% are indicated at branchnodes (Felsenstein 1985)
258 Appl Microbiol Biotechnol (2006) 72:252–262

relationship (Fig. 7). This tree was built on partial 16SrRNA gene sequences as the whole sequences were notavailable for all the strains.
From this tree, three groups of strains appear. Twodistinct clusters belong to the beta-Proteobacteria phylum(order Burkholderiales): one cluster containing strain L108and related strains and the other one includingM. petroleiphilum PM1 and related strains. A third clusterof Gram-positive bacteria belongs to the Actinobacteria (orderActinomycetales) and includes M. austroafricanum species.H. flava ENV735 is separate from these three groups.
These observations, though based on a relatively lownumber of strains, are potentially interesting as they couldallow the design of a range of primers dedicated to theamplification of the different specific 16S rRNA genes tohelp in predicting the potential for natural attenuation inMTBE-contaminated environments (Hristova et al. 2003;Lopes Ferreira et al. 2006b).
Enzymes and genes involved in the MTBE degradationpathway
MTBE and TBA monooxygenases
In M. petroleiphilum PM1, Deeb et al. (2001) showed thattwo different monooxygenases are probably involved in theoxidation of MTBE and TBA.
Using different inhibitors or inactivators of monooxyge-nases, Hatzinger et al. (2001) showed that two differentmonooxygenases were involved in MTBE and TBA
oxygenation in H. flava ENV735. Moreover, the authorsshowed that the MTBE monooxygenase was constitutivelyexpressed, which made its identification challenging.
Our results, at a physiological level, indicated that aunique monooxygenase was probably responsible for theoxidation of both MTBE and TBA in M. austroafricanumIFP 2012 (Fayolle et al. 2003) but with a different affinityfor MTBE and TBA (Ks for MTBE <0.2 mM and Ks forTBA=1.1 mM).
In strain L108, two different monooxygenases wereclearly involved in MTBE and TBA oxidations as a mutant,L10, unable to grow on MTBE but still able to grow onTBA was easily isolated from L108 by successive transferson a rich medium (Rohwerder et al. 2006).
However, to date and to our knowledge, none of themonooxygenases enabling growth on MTBE and/or TBAhave been isolated and genetically characterized, even if, asdiscussed above, cytochromes P450 and AlkB monoox-ygenases are plausible candidates.
Esterase
The role of an esterase hydrolyzing TBF to TBA andformate is essential in the early steps of the MTBEmetabolic pathway if the pathway is through TBF. Theabiotic hydrolysis of TBF at neutral pH is actually very low.At an initial TBF concentration of 230 μM, the rate ofabiotic TBF degradation is 2 μM h−1, whereas the rateincreases to 50 μM h−1 in the presence of M. austro-africanum IFP 2012 resting cells (François et al. 2003),
Fig. 8 Structure and function ofmpd genes of M. austroafrica-num IFP 2012. The intermedi-ates identified are in bold
Appl Microbiol Biotechnol (2006) 72:252–262 259

which clearly shows the role of the microorganism in TBFhydrolysis. Smith et al. (2003a) demonstrated the activity ofa heat-stable esterase in M. vaccae JOB5 resting cells. Theinitial TBF degradation rates were in the same range for M.austroafricanum IFP 2012 and M. vaccae JOB5, 12 and5 nmol min−1 mg−1 protein, respectively.
As said above, such an esterase activity was alsodetected in R. ruber IFP 2001 which was able to grow onTBAc.
TBF was not always detected as an intermediate with allstrains able to grow on MTBE, which could mean (1) that itwas very rapidly degraded by an efficient esterase and, as aconsequence, was difficult to identify during MTBEdegradation or (2) that the pathway is through dismutationrather than by dehydrogenation of tert-butoxy methanol.Whether the strains can use TBF as a growth substrate ornot may help in discriminating between these two options.
Dehydrogenases (mpd genes of M. austroafricanumIFP 2012)
A cluster of mpd genes (Fig. 8) involved in the conversionof 2,M1,2PD to HIBA was isolated from M. austro-africanum IFP 2012, cloned, and expressed in M. smegma-tis mc2 155 (Lopes Ferreira et al. 2006a). The transformantobtained, M. smegmatis mc2 155 (p4D2), stoichiometricallyconverted 2M1,2PD to HIBA. This requires a de novoprotein synthesis as no conversion occurred in the presenceof chloramphenicol.
The cluster encodes an alcohol dehydrogenase (MpdB),an aldehyde dehydrogenase (MpdC), and a putativeregulator presumably controlling the synthesis of MpdBand MpdC. MpdB (552 residues, 64 kDa) is highly similar(42 and 56%, respectively) to a glucose–methanol–cholineoxidoreductase of Bradyrhizobium japonicum; MpdB isalso similar (33% identical and 48% similar residues) to anamino acid sequence of a similar size (522 amino acids)deduced from the annotated genome of M. petroleiphilumPM1 (gene MpeA0945). The latter may be an enzymeplaying a role similar to MpdB in the MTBE pathway ofM. petroleiphilum PM1. MpdC (55 kDa) is highly similar(45% identical and 61% similar residues) with a nicotin-amide adenine dinucleotide (NAD)-dependent aldehydedehydrogenase of Burkholderia fungorum. Both MpdBand MpdC were strongly induced during the growth ofM. austroafricanum IFP 2012 on MTBE as shown by theprotein profile on sodium dodecyl sulfate-polyacrylamidegel electrophoresis.
The step converting 2M1,2PD to HIBA (Fig. 6) allowsthe production of reduced coenzyme equivalents which islinked to the production of ATP and it has been proposedthat the energy loss during growth in the presence of MTBEcould be a key limiting factor in its catabolism (Fayolle et
al. 2003; Fortin et al. 2001; Salanitro 1995). Therefore, ahighly efficient conversion of 2M1,2PD to HIBA iscertainly beneficial to the MTBE pathway.
Mutase (icm gene of strain L108)
The growth of strain L108 on HIBA is cobalamindependent (Rohwerder et al. 2006) and cobalt cations arerequired for growth on TBA of strain I-2052 (IFP 2003)(Piveteau et al. 2001) which is phylogenetically closelyrelated to L108 (Fig. 7). Rohwerder et al. (2006) identifiedan isobutyryl-CoA mutase (ICM, EC 5.4.99.13) subunit instrain L108 (wild type) and in its variant L10, which wasunable to grow on MTBE but still able to grow on TBA.
A sequence identical to that of icm in strain L108 was alsofound completely conserved (nearly 100% identity in thededuced peptide sequences) in the sequenced genome ofM. petroleiphilum PM1 (MpeB0541 and MpeA1014 genes).
The authors showed the presence of a correspondingenzymatic activity in crude extracts of strain L108 bydemonstrating the conversion of HIBA into 3-hydroxybu-tyrate in the presence of ATP and CoA. The authorsconsequently proposed a pathway for the assimilation ofHIBA in the strains possessing such an icm gene (Fig. 6).
The conservation of this gene among microorganism,from Archae to Bacteria, shown in Blast alignment isremarkable and could be due to its crucial role in the centralmetabolism of microorganisms.
Enzymes of the methylotrophic pathway
To improve the efficiency of the MTBE degradationpathway, the regeneration of cofactor via the oxidation ofthe C1 compound formed (formate or formaldehyde) toCO2 is certainly an advantage in microorganisms able togrow on MTBE. Moreover, if formaldehyde is producedinstead of formate (Fig. 6), the capacity to degradeformaldehyde may be a prerequisite because of its toxicity.
M. austroafricanum IFP 2012 and IFP 2015 are methylo-trophs that are able to mineralize formate to CO2 (François etal. 2002; Lopes Ferreira, unpublished result). Methylotrophyis indeed a property very common in mycobacteria (Park etal. 2003). For example, a NAD-dependent formate dehydro-genase was cloned and characterized inM. vaccae N10 whichis very close to M. austroafricanum (Galkin et al. 1995).
H. flava ENV735 is likewise able to grow on formaldehyde(Hatzinger et al. 2001) and M. petroleiphilum PM1 is afacultative methylotroph that uses methanol as a solecarbon and energy source (Nakatsu et al. 2006).
260 Appl Microbiol Biotechnol (2006) 72:252–262

Conclusions
Investigating microorganisms that are able to grow onMTBE remains a challenge due to: (1) the low biomassproduced from MTBE by microorganisms (Fayolle et al.2003; Fortin et al. 2001) and (2) the lack of efficient genetictools for working with environmental bacteria. Neverthe-less, a few genes involved in the MTBE biodegradationpathway have been isolated and their DNA sequences couldbe used as tools for studying other microorganisms thatdegrade MTBE. In this regard, the results obtained bycomparing the sequences of eth, mpd, and icm genes to thesequenced genome of M. petroleiphilum PM1 are reallyinteresting and could help to understand the MTBEpathway of this strain. The isolation of the MTBE/TBAmonooxygenases in the strains growing on MTBE has stillto be performed.
Concerning the remediation of MTBE-contaminatedaquifers, the monitored natural attenuation could benefitvery efficiently from the use of genetic tools for thecharacterization of the biodegradation potential towardsMTBE in contaminated environments. Microarrays couldbe built including genes involved in the MTBE biodegra-dation pathways as well as sequences specifically designedfrom the 16S rRNA genes from the different groups ofstrains involved in the biodegradation. Their use could helpin directing the remediation process in such contaminatedsites.
Acknowledgements Nicolas Lopes Ferreira and Cédric Malandainwere supported by a Convention Industrielle de Formation par laRecherche (C.I.F.R.E.) fellowship provided by both the InstitutFrançais du Pétrole (I.F.P.) and the Association Nationale de laRecherche Technique (A.N.R.T.). We thank Charles W. Greer fornumerous helpful suggestions and discussions. We thank Diane Labbé(BRI, Montréal) for her help in building the phylogenetic tree. Wethank Robert Steffan, Paul Hatzinger, and Simon Vainberg (ShawGroup) for communicating the 16S rDNA sequences of strainsENV735, ENV736, and ENV472. We thank Thore Rohwerder forthe early communication of his manuscript in press. We are reallygrateful to Pierre Béguin (Pasteur Institute) and Frédéric Monot (IFP)for their comments and corrections on the manuscript.
References
Aramaki H, Sagara Y, Hosoi M, Horiuchi T (1993) Evidence forautoregulation of camR, which encodes a repressor for thecytochrome P-450cam hydroxylase operon on the Pseudomonasputida CAM plasmid. J Bacteriol 175:7828–7833
Baptist JN, Gholson RK, Coon MJ (1963) Hydrocarbon oxidation bya bacterial enzyme system. I. Products of octane oxidation.Biochim Biophys Acta 69:40–47
Béguin P, Chauvaux S, Miras I, François A, Fayolle F, Monot F(2003) Genes involved in the degradation of ether fuels bybacteria of the Mycobacterium–Rhodococcus complex. Oil GasSci Technol 58:489–497
Chauvaux S, Chevalier F, Le Dantec C, Fayolle F, Miras I, Kunst F,Béguin P (2001) Cloning of a genetically unstable cytochrome P-450 gene cluster involved in degradation of the pollutant ethyltert-butyl ether by Rhodococcusruber. J Bacteriol 183:6551–6557
Chen Q, Janssen DB, Witholt B (1996) Physiological changes and alkgene instability in Pseudomonas oleovorans during induction andexpression of alk genes. J Bacteriol 178:5508–5512
Deeb RA, Nishino S, Spain J, Hu H-Y, Scow K, Alvarez-Cohen L(2000) MTBE and benzene biodegradation by a bacterial isolatevia two independent monooxygenase-initiated pathways. Pre-prints of extended abstracts, ACS national meeting. AM ChemSoc Div Environ Chem 40: 280–282
Deeb RA, Hu HY, Hanson JR, Scow KM, Alvarez-Cohen L (2001)Substrate interactions in BTEX and MTBE mixtures by anMTBE-degrading isolate. Environ Sci Technol 35:312–317
Eggink G, van Lelyveld PH, Arnberg A, Arfman N, Witteveen C,Witholt B (1987) Structure of the Pseudomonas putida alkBAC.Identification of transcription and translation products. J BiolChem 262:6400–6406
Eggink G, Engel H, Meijer WG, Otten J, Kingma J, Witholt B (1988)Alkane utilization in Pseudomonas oleovorans: structure andfunction of the regulatory locus alkR. J Biol Chem 263:13400–13405
Fayolle F, Monot F (2005) Biodegradation of fuel ethers. In: MagotM, Ollivier B (eds) Petroleum microbiology. ASM, Washington,DC, USA, pp 301–316
Fayolle F, François A, Garnier L, Godefroy D, Mathis H, Piveteau P,Monot F (2003) Limitations in MTBE biodegradation. Oil GasSci Technol 58:497–504
Felsenstein J (1985) Confidence limits on phylogenies: an approachusing the bootstrap. Evolution 39:783–791
Fortin N, Morales M, Nakagawa Y, Focht DD, Deshusses MA (2001)Methyl tert-butyl ether (MTBE) degradation by a microbialconsortia. Environ Microbiol 3:407–416
François A, Mathis H, Godefroy D, Piveteau P, Fayolle F, Monot F(2002) Biodegradation of methyl tert-butyl ether and other fueloxygenates by a new strain, Mycobacterium austroafricanum IFP2012. Appl Environ Microbiol 68:2754–2762
François A, Garnier L, Mathis H, Fayolle F, Monot F (2003) Roles oftert-butyl formate, tert-butyl alcohol and acetone in the regula-tion of methyl tert-butyl ether degradation by Mycobacteriumaustroafricanum IFP 2012. Appl Microbiol Biotechnol 62:256–262
Galkin A, Kulakova L, Tishkov V, Esaki N, Soda K (1995) Cloning offormate dehydrogenase gene from a methanol-utilizing bacteriumMycobacterium vaccae N10. Appl Microbiol Biotechnol 44:479–483
Goodfellow M, Jones LJ, Maldonado LA, Salanitro J (2004) Rhodo-coccus aetherivorans sp. nov., a new species that contains methylt-butyl ether-degrading actinomycetes. Syst Appl Microbiol27:61–65
Hanson JR, Ackerman CE, Scow KM (1999) Biodegradation ofmethyl tert-butyl ether by a bacterial pure culture. Appl EnvironMicrobiol 65:4788–4792
Hatzinger PB, Mc Clay K, Vainberg S, Tugusheva M, Condee CW,Steffan RJ (2001) Biodegradation of methyl tert-butyl ether by apure bacterial culture. Appl Environ Microbiol 67:5601–5607
Hernandez-Perez G, Fayolle F, Vandecasteele JP (2001) Biodegrada-tion of ethyl t-butyl ether (ETBE), methyl t-butyl ether (MTBE)and t-amyl methyl ether (TAME) by Gordoniaterrae. ApplMicrobiol Biotechnol 55:117–121
Hristova K, Gebreyesus B, Mackay D, Scow KM (2003) Naturallyoccurring bacteria similar to the methyl tert-butyl ether (MTBE)-degrading strain PM1 are present in MTBE-contaminatedgroundwater. Appl Environ Microbiol 69:2616–2623
Appl Microbiol Biotechnol (2006) 72:252–262 261

Hyman M, O’Reilly K (1999) Physiological and enzymatic features ofMTBE-degrading bacteria. In: Alleman BC, Leeson A (eds) Insitu bioremediation of petroleum hydrocarbons and other organiccompounds. Battelle, Columbus, Ohio, pp 7–12
Hyman M, Glover K, House A, Johnson E, Smith C (2004)Physiological and enzymatic diversity of aerobic MTBE biodeg-radation processes. In: Barcelo D, Petrovic M (eds) Proceedingsof the second European conference on MTBE. C.S.I.C.,Barcelona, Spain, pp 39–43
Johnson EL, Hyman MR (2006) Propane and n-butane oxidation byPseudomonas putida GPo1. Appl Environ Microbiol 72:950–952
Johnson R, Pankow J, Bender D, Price C, Zogorsky J (2000)MTBE. To what extent will past releases contaminatecommunity water supply wells? Environ Sci Technol34:210A–217A
Johnson EL, Smith CA, O’Reilly KT, Hyman MR (2004) Induction ofmethyl tert-butyl ether (MTBE)-oxidizing activity in Mycobac-teriumvaccae JOB5. Appl Environ Microbiol 70:1023–1030
Liu CY, Speitel GE, Georgiou G (2001) Kinetics of methyl t-butylether cometabolism at low concentrations by pure cultures ofbutane-degrading bacteria. Appl Environ Microbiol 67:2197–2201
Lopes Ferreira N, Labbé D, Monot F, Fayolle-Guichard F, Greer CW(2006a) Genes involved in the methyl tert-butyl ether (MTBE)metabolic pathway of Mycobacterium austroafricanum IFP 2012.Microbiology 152:1361–1374
Lopes Ferreira N, Maciel H, Mathis H, Monot F, Fayolle-Guichard F,Greer CW (2006b) Isolation of a new Mycobacterium austro-africanum, IFP 2015, growing on MTBE. Appl MicrobiolBiotechnol 70:358–365
Nagy I, Scoofs G, Compernolle F, Proost P, Vanderleyden J, De Mot R(1995) Degradation of the thiocarbamate herbicide EPTC S-ethyldipropylcarbamothioate) and biosafing by Rhodococcus sp.strain NI86/21 involve an inducible cytochrome P-450 systemand aldehyde dehydrogenase. J Bacteriol 177:676–687
Nakatsu CH, Hrsitova K, Hanada S, Meng X-Y, Hanson J, Scow KM,Kagamata Y (2006) Methylibium petroleiphilum PM1T gen. nov.,sp. nov., a new methyl tert-butyl ether (MTBE) degradingmethylotroph of the beta-Proteobacteria. Int J Syst Evol Microbiol(DOI 10.1099/ijs.0.63524-0)
Ooyama J, Foster JW (1965) Bacterial oxidation of cycloparaffinichydrocarbons. Antonie van Leeuwenhoek 31:45–65
Park SW, Hwang EH, Park H, Kim JA, Heo J, Lee KH, Song T, KimE, Ro YT, Kim SW, Kim YM (2003) Growth of mycobacteria oncarbon monoxide and methanol. J Bacteriol 185:142–147
Piveteau P, Fayolle F, Vandecasteele JP, Monot F (2001) Biodegrada-tion of tert-butyl alcohol and related xenobiotics by a methylo-trophic bacterial isolate. Appl Microbiol Biotechnol 55:369–373
Poulos TL, Finzel BC, Gunsalus IC, Wagner GC, Kraut J (1985) The2.6-A crystal structure of Pseudomonas putida cytochromeP-450. J Biol Chem 30:16122–16130
Pruden A, Suidan M (2004) Effect of benzene, toluene, ethylbenzeneand p-xylene (BTEX) mixture on biodegradation of methyl tert-butyl ether (MTBE) and tert-butyl alcohol by pure culture UC1.Biodegradation 15:213–227
Rohwerder T, Cenini V, Held C, Martienssen M, Lechner U, MüllerRH (2004) Novel MTBE-degrading bacterial isolate from Leunagroundwater (Germany): characterization of the degradationpathway with focus on HIBA oxidase. In: Barcelo D, Petrovic
M (eds) Proceedings of the second European conference onMTBE. C.S.I.C., Barcelona, Spain, pp 47–50
Rohwerder T, Breuer U, Benndorf D, Lechner U, Müller RH (2006)The alkyl tertiary butyl ether intermediate 2-hydroxyisobutyrateis degraded via a novel cobalamin-dependent mutase pathway.Appl Environ Microbiol. DOI 10.1128/AEM.00080-06
Saitou N, Nei M (1987) The neighbour-joining method: a newmethod for reconstructing phylogenetic trees. Mol Biol Evol4:406–425
Salanitro JP (1995) Understanding the limitations of microbialmetabolism of ethers used as fuel octane enhancers. Curr OpinBiotechnol 6:337–340
Salanitro JP, Diaz LA, Williams MP, Wisniewski HL (1994) Isolationof a bacterial culture that degrades methyl t-butyl ether. ApplEnviron Microbiol 60:2593–2596
Smith CA, Hyman MR (2004) Oxidation of methyl tert-butyl ether bypropane-grown Pseudomonas putida GPo1. Appl Environ Microbiol70:4544–4550
Smith CA, O’Reilly KT, Hyman MR (2003a) Characterization of theinitial reactions during the cometabolic oxidation of methyl tert-butyl ether by propane-grown Mycobacterium vaccae JOB5.Appl Environ Microbiol 69:796–804
Smith CA, O’Reilly KT, Hyman MR (2003b) Cometabolism ofmethyl tert-butyl ether and gaseous n-alkanes by Pseudomonasmendocina KR-1 grown on C5 to C8n-alkanes. Appl EnvironMicrobiol 69:7385–7394
Steffan RJ, McClay K, Vainberg S, Condee CW, Zhang D (1997)Biodegradation of the gasoline oxygenates methyl tert-butyl ether,ethyl tert-butyl ether, and tert-amyl methyl ether by propane-oxidizing bacteria. Appl Environ Microbiol 63:4216–4222
Steffan RJ, Vainberg S, Condee CW, McClay K, Hatzinger PB(2000) Biotreatment of MTBE with a new bacterial isolate.In: Wickramanayake GB, Gavaskar AR, Alleman BC, MagarVS (eds) Bioremediation and phytoremediation of chlorinatedand recalcitrant compounds. Battelle, Columbus, Ohio, pp165–173
Tamura K, Nei M (1993) Estimation of the number of nucleotidesubstitutions in the control region of mitochondrial DNA inhumans and chimpanzees. Mol Biol Evol 10:512–526
Unger BP, Gunsalus IC, Sligar SG (1986) Nucleotide sequence of thePseudomonas putida P-450cam gene and its expression inEscherichia coli. J Biol Chem 261:1158–1163
Urios A, Fayolle F, Monot F, Chauvaux S, Béguin P (2002)Physiological comparison of an ETBE (MTBE)-degrading strainand some derived mutants. In: Gavaskar AR, Chen ASC (eds)Remediation of chlorinated and recalcitrant compounds—2002.ISBN 1-57477-132-9. Battelle, Columbus, Ohio. http://www.battelle.org/bookstore
Van Beilen JB, Wubbolts MG, Witholt B (1994) Genetics of alkaneoxidation by Pseudomonas oleovorans. Biodegradation 5:161–174
Van Beilen JB, Smits THM, Roos FF, Brunner T, Balada SB,Röthlisberger M, Witholt B (2005) Identification of an aminoacid position that determines the substrate range of integralmembrane alkane hydroxylase. J Bacteriol 187:85–91
Wilson JT (2003) Fate and transport of MTBE and other gasolinecomponents. In: Moyer EE, Kostecki PT (eds) MTBE remedia-tion handbook. Amherst Scientific, Amherst, MA, USA,pp 19–61
262 Appl Microbiol Biotechnol (2006) 72:252–262
Copyright © 2022 FDOKUMEN














![Multinuclear NMR Studies and Reaction with tert-Butyl Isocyanide of Dinuclear Tungsten- or Molybdenum-Palladium .mu.-Alkylidene Complexes. X-ray Structure of [PdI[.mu.-C(p-tolyl)dmba][.mu.-CO]Mo(Cp)(CN-t-Bu)2]](https://static.fdokumen.com/doc/165x107/63358a2364d291d2a3028408/multinuclear-nmr-studies-and-reaction-with-tert-butyl-isocyanide-of-dinuclear-tungsten-.jpg)





