
Extraction and application of antioxidants from black glutinous rice
Upload
independentCategory
view
1download
0
Original Contribution
EXERCISE, ANTIOXIDANTS, AND HSP72: PROTECTION AGAINSTMYOCARDIAL ISCHEMIA/REPERFUSION
KARYN L. HAMILTON ,* JESSICA L. STAIB,* TRACEY PHILLIPS,* A NDREA HESS,* SHANNON L. LENNON,* and
SCOTT K. POWERS*†
*Department of Exercise and Sport Science and†Department of Physiology, Center for Exercise Science,University of Florida, Gainesville, FL, USA
(Received 2 July 2002;Revised 11 December 2002;Accepted 12 December 2002)
Abstract—Endurance exercise is associated with protection against myocardial ischemia/reperfusion (I/R) injury andhas been shown to increase heat shock protein 72 (HSP72). Dietary antioxidants have also been reported to decreaseI/R-induced injury. Because exercise and antioxidants may provide cardioprotection via different mechanisms, com-bining these countermeasures could provide additive protection. Alternatively, because exercise-induced oxidantproduction may promote expression of HSP72, antioxidants could attenuate exercise-induced HSP72 expression anddecrease exercise-related cardioprotection. These experiments examined the individual and combined effects of exerciseand antioxidants on myocardial I/R injury (in vivo). Rats receiving a mixed antioxidant diet or control diet were assignedto exercise or sedentary groups and randomized to receive: (i) short I/R (myocardial stunning), (ii) long I/R (myocardialinfarction), or (iii) sham surgery. Antioxidants significantly increased total antioxidant capacity and attenuated exercise-related HSP72 accumulation. Nonetheless, during short I/R, exercise-trained animals demonstrated improved leftventricular developed pressure (LVDP), independent of diet. Further, antioxidants alone resulted in improved LVDP.Finally, compared to control diet/sedentary animals, both exercise groups (control and antioxidant diets) and theantioxidant diet/sedentary group sustained smaller infarctions. We conclude that exercise and antioxidants can inde-pendently provide protection against myocardial contractile dysfunction and infarction, and the combination of thesetwo strategies does not enhance or inhibit the protection observed with each individual countermeasure. © 2003Elsevier Science Inc.
Keywords—Heat shock proteins, Reactive oxygen species, Lipid peroxidation, Stunning, Infarction, Free radicals
INTRODUCTION
Myocardial ischemia/reperfusion (I/R) can lead to ionicimbalances, activation of proteases, oxidative injury, andultimately cardiac cell death. Hence, developing coun-termeasures to protect the myocardium from I/R injury isimportant. In this regard, evidence indicates that severalmembers of the heat shock protein (HSP) family canprovide myocardial protection against cellular stress.Specifically, HSP72 has been shown to protect cells fromI/R injury [1–3]. Perhaps the most convincing evidencethat HSP72 provides cardioprotection is provided bytransgenic mice overexpressing HSP72 with improved
postischemic contractile function and decreased infarc-tion size [4,5].
Endurance exercise is a pragmatic means of elevatingmyocardial levels of HSP72 and, importantly, exercisetraining is also associated with cardioprotection during I/R[1–4]. While the exercise-induced cellular signals respon-sible for increased expression of myocardial HSP72 con-tinue to be investigated, an increased production of reactiveoxygen species (ROS) has been postulated to play a prom-inent role [6–13]. Although ROS regulate several redox-signaling pathways in healthy cells, it is also clear that highlevels of ROS contribute to numerous pathologies includingI/R injury [14]. Therefore, dietary antioxidants could act asanother countermeasure to protect the myocardium againstI/R injury [15,16].
While the specific mechanism(s) by which exerciseprovides cardioprotection continues to be investigated
Address correspondence to: Dr. Karyn L. Hamilton, University ofFlorida, Center for Exercise Science, P.O. Box 118206, Gainesville, FL32611, USA; Tel: (352) 392-9575; Fax: (352) 392-0316; E-Mail:[email protected].
Free Radical Biology & Medicine, Vol. 34, No. 7, pp. 800–809, 2003Copyright © 2003 Elsevier Science Inc.Printed in the USA. All rights reserved
0891-5849/03/$–see front matter
doi:10.1016/S0891-5849(02)01431-4
800

[17–22], it has been argued that exercise-induced in-creases in myocardial HSP72 and enzymatic antioxidantsplay important and distinct roles in exercise-related car-dioprotection [2,3,22]. HSP72 has been proposed to pro-vide cellular protection via its ability to orchestrateproper folding of nascent proteins, assist in protein re-pair, and facilitate degradation of irreparably damagedproteins. Antioxidants, alternatively, have been shown toprevent I/R-associated oxidative damage to lipids, pro-teins, and DNA. It is plausible, therefore, that the com-bination of exercise and antioxidants could provide ad-ditive protection against I/R-induced myocardial injury.However, if ROS are important signals to increaseHSP72 expression or antioxidant enzyme activity, anti-oxidant supplementation could attenuate the exercise-induced elevation in myocardial HSP72 and potentiallyeliminate the cardioprotection associated with exercise.
Whether the combination of exercise and dietary an-tioxidants would provide additive cardioprotection orwhether increased antioxidant capacity might attenuateexercise-induced cardioprotection via attenuation of HSPinduction is not known and forms the basis for thecurrent experiments. We hypothesized that supplemen-tation with antioxidants would attenuate exercise-in-duced expression of myocardial HSP72 and decrease thecardioprotection associated with exercise. We tested thishypothesis by determining the individual and combinedeffects of exercise training and supplementation withantioxidants on myocardial expression of HSP72, anti-oxidant enzyme activity, and protection against I/R-in-duced myocardial injury in vivo.
MATERIALS AND METHODS
Experimental design
All experiments were approved by the University ofFlorida animal care and use committee and were con-ducted in accordance with institutional guidelines. Fe-male Sprague-Dawley rats (3 months old) were dividedinto two dietary groups: (i) control diet, AIN93M (Amer-ican Institute of Nutrition 1993 diet formulation for ma-ture rodents); and, (ii) antioxidant diet, AIN93M plusantioxidants.
After completing a 6 week feeding protocol, ratswithin each group were further randomized to receive 5consecutive days of treadmill training or to remain sed-entary. Each of these groups was further randomized intoone of three surgical treatment groups (Fig. 1): (i) shamsurgery, (ii) in vivo myocardial stunning (20 min isch-emia/30 min reperfusion), or (iii) in vivo myocardialinfarction (60 min ischemia/120 min reperfusion). Shamsurgery animals, which were used for assessment ofantioxidant capacity, underwent all surgical proceduresexcept coronary artery occlusion and reperfusion. Theinclusion of experimental groups that were exposed toeither a 20 min or a 60 min period of ischemia provideda rigorous test of our hypothesis at two different levels ofI/R-induced myocardial injury. Specifically, the 20 minduration of regional ischemia results in myocardial stun-ning without a significant infarction. In contrast, the 60min duration of regional ischemia results in a frankmyocardial infarction with approximately 60% of therisk area being infarcted.
Fig. 1. Experimental design with treatments as follows: 20I/30R � 20 min ischemia/30 min reperfusion, and 60I/120R � 60 minischemia/120 min reperfusion. Methods used for analysis of the eight surgical groups are indicated by dotted, dashed, or solid boxes.
801Exercise, antioxidants, I/R

Antioxidant diet and treadmill exercise training
The antioxidant diet (Bioserv, Frenchtown, NJ, USA)used in this study was chosen to provide global antiox-idant protection in both aqueous and lipid phases of thecell. Specific doses of the selected antioxidants (Table 1)in our experimental diet were selected based on a de-tailed series of antioxidant dose-response studies indicat-ing that the current diet results in significant improve-ments in antioxidant capacity in rodent tissues withouttoxic side effects [23,24].
Exercise training included 5 d of habituation to thetreadmill beginning with 10 min of running at 30 m/minand 0% grade, with daily increases of 10 min until 50min was achieved. Habituation was followed by 5 con-secutive d of continuous running (60 min/d) at 30 m/minand 0% grade or approximately 60–70% of VO2max. Thisprotocol has been shown to improve cardioprotectionduring in vivo I/R and result in accumulation of myo-cardial HSP72 in adult rats [18,20].
In vivo myocardial ischemia/reperfusion
At the completion of the feeding and exercise proto-cols (24 h postexercise in trained animals), rats wereanesthetized (40 mg/kg�1 sodium pentobarbital, ip), in-tubated via tracheostomy, and ventilated. Following aleft thoracotomy, a ligature was placed around the leftanterior descending coronary artery close to its origin.Coronary occlusion was maintained for either 20 or 60min followed by reperfusion for either 30 or 120 min formyocardial stunning or myocardial infarction, respec-tively. Colonic temperature and electrocardiographic ac-tivity were continuously monitored. Rats with more thanthree episodes of ventricular fibrillation (VF), or an ep-isode of VF persisting longer than 30 s, were eliminated.In rats exhibiting VF, cardioversion was attempted firstby gentle massage of the noncyanotic area and then bybrief electrical pacing (3–5 v/0.1 ms duration/300 bpm).All experimental procedures were executed in a blindedfashion.
Measurement of cardiac contractile function
Cardiac function measurements, monitored through-out the myocardial stunning protocol, included left ven-tricular developed pressure (LVDP) and rate of ventric-ular pressure development and relaxation (�dP/dt).Ventricular pressures were measured by placing a cath-eter filled with heparinized saline into the left ventricularcavity via the carotid artery. The catheter was attached toa pressure transducer interfaced with a computerizedheart performance analyzer (Digi-Med, Louisville, KY,USA).
Tissue storage/assessment of myocardial infarction size
At the conclusion of the myocardial stunning andsham surgical protocols, hearts were removed, rinsed ina cold antioxidant buffer (50 mM NaHPO4, 0.1 mMbutylated hydroxytoluene, and 0.1 mM diethylenetri-aminepentaacetic acid, pH 7.4), and rapidly frozen inliquid nitrogen.
Following completion of the myocardial infarctionprotocol, the coronary artery was reoccluded and 1 ml ofEvans blue dye was injected into the left ventricle. Theheart was sliced into 1.5 mm cross-sections, incubated in1% triphenyl tetrazolium chloride at 37°C for 20 min,placed in 10% formalin for 24 h, and photographed.Image analysis (SigmaScan, Jandel Scientific, CorteMadera, CA, USA) was executed in a blinded fashionand infarct size (portions within the risk area not stainedbrick red by the triphenyl tetrazolium chloride formazanprecipitates) was expressed as a percentage of the area atrisk (areas not stained blue by the Evans dye) in eachslice [25].
Preparation of protein extract/Western blotting
Left ventricular samples from sham surgeries werehomogenized and proteins were separated on a 12%polyacrylamide gel and transferred to nitrocellulosemembranes. Membranes were then incubated withHSP72 (SPA810) monoclonal antibodies (StressGen,Victoria, British Columbia, Canada) and secondary an-tibodies conjugated to horseradish peroxidase for chemi-luminescent detection. Following densitometric analyses(SigmaGel, Jandel Scientific), values were expressed as apercentage of control untrained.
Assessment of antioxidant enzyme activity, totalantioxidant status, and oxidative damage
To determine if treatments altered enzymatic anti-oxidant activity, left ventricles from sham surgerieswere assayed for the activities of manganese superox-ide dismutase (MnSOD), copper-zinc superoxide dis-mutase (Cu,ZnSOD), glutathione peroxidase (GPX),and catalase (CAT). Samples were assayed for Mn-
Table 1. Antioxidant Diet Composition
Antioxidant added to AIN93MaAmount/kg diet
(mg unless noted)
Dl-� tocopherol acetate 1000 IUSelenium 0.3Ascorbic acid 1000Beta-carotene (20%) 225Coenzyme Q10 30N-acetyl cysteine 200Catechin 100
a AIN93M � American Institute of Nutrition 1993 diet formulationfor mature rodents.
802 K. L. HAMILTON et al.
SOD and Cu,ZnSOD using the cytochrome c reductiontechnique of McCord and Fridovich [26]. SeleniumGPX activity was assayed as described by Flohe andGunzler [27]. CAT was assayed using the protocoldescribed by Aebi [28]. Total antioxidant status (TAS)of plasma and supernatant from left ventricular tissuewas measured using a kit purchased from RandoxLaboratory (Antrim, Northern Ireland). The resultsfrom this kit do not distinguish between lipid- andwater-soluble antioxidants but rather provide an esti-mate of global antioxidant capacity. Finally, as amarker of lipid peroxidation, total 8-isoprostane wasquantified in left ventricle samples from the area atrisk during myocardial stunning surgeries by compet-itive enzyme immunoassay (Cayman Chemical, AnnArbor, MI, USA). These and other biochemical anal-yses were performed in triplicate at 25°C and samplesfrom all experimental animals were assayed on thesame day to avoid interassay variation.
Statistical analyses
Values reported are means � SEM. Data werecompared by one-way ANOVA followed by a studentNewman-Keuls posthoc test. Where normalized valuesare compared with control values (Western blots), datawere analyzed using an ANOVA on ranks (Kruskal-Wallis) followed by a Dunn’ s test; if data passed testsof normality and equal variance, one-way ANOVAwas performed. Level of significance was set a prioriat p � .05.
RESULTS
Myocardial stunning and infarction
Across all experimental groups, a total of 20 rats wereeliminated from the study based on the exclusion criteriafor VF. The control diet/untrained and antioxidant diet/trained groups each had five eliminated. Four were elim-inated from the control diet/trained group and six wereeliminated from the antioxidant diet/untrained group.Hence, the number of rats eliminated based on the ex-clusion criteria for VF set a priori was similar acrossexperimental groups.
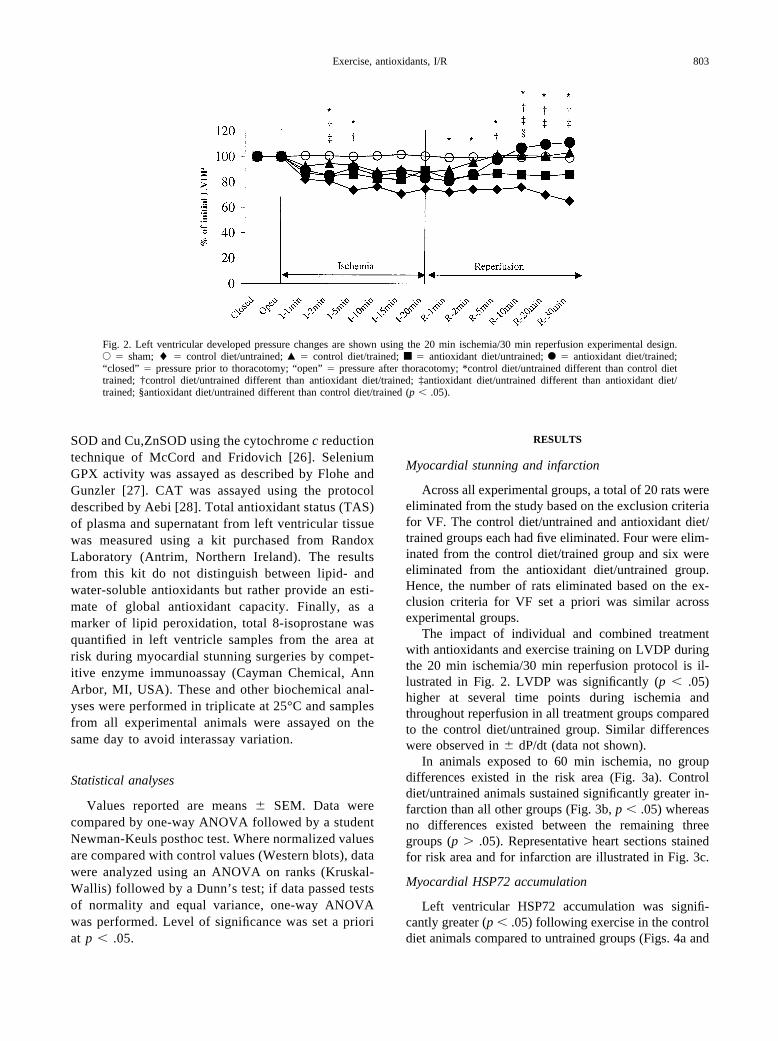
The impact of individual and combined treatmentwith antioxidants and exercise training on LVDP duringthe 20 min ischemia/30 min reperfusion protocol is il-lustrated in Fig. 2. LVDP was significantly (p � .05)higher at several time points during ischemia andthroughout reperfusion in all treatment groups comparedto the control diet/untrained group. Similar differenceswere observed in � dP/dt (data not shown).
In animals exposed to 60 min ischemia, no groupdifferences existed in the risk area (Fig. 3a). Controldiet/untrained animals sustained significantly greater in-farction than all other groups (Fig. 3b, p � .05) whereasno differences existed between the remaining threegroups (p � .05). Representative heart sections stainedfor risk area and for infarction are illustrated in Fig. 3c.
Myocardial HSP72 accumulation
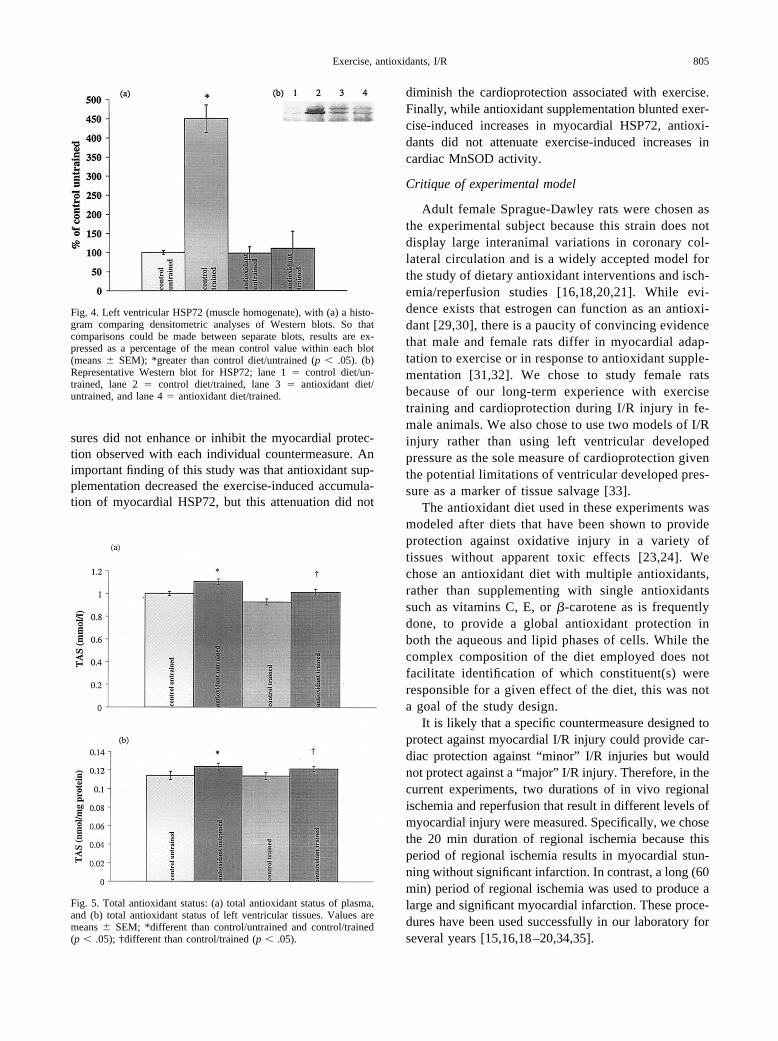
Left ventricular HSP72 accumulation was signifi-cantly greater (p � .05) following exercise in the controldiet animals compared to untrained groups (Figs. 4a and
Fig. 2. Left ventricular developed pressure changes are shown using the 20 min ischemia/30 min reperfusion experimental design.E � sham; � � control diet/untrained; Œ � control diet/trained; ■ � antioxidant diet/untrained; F � antioxidant diet/trained;“closed” � pressure prior to thoracotomy; “open” � pressure after thoracotomy; *control diet/untrained different than control diettrained; †control diet/untrained different than antioxidant diet/trained; ‡antioxidant diet/untrained different than antioxidant diet/trained; §antioxidant diet/untrained different than control diet/trained (p � .05).
803Exercise, antioxidants, I/R

4b). However, exercise did not result in significant in-creases in HSP72 accumulation in antioxidant-supple-mented animals.
Total antioxidant status, antioxidant enzyme activity,and oxidative injury
Antioxidant diet animals had significantly higherplasma and myocardial (p � .05) TAS compared tocontrol diet rats (Figs. 5a and 5b).
Table 2 contains mean (� SEM) activities of GPX,SOD, MnSOD, Cu,ZnSOD, and CAT in the left ventri-cle. Note that antioxidant enzyme activities were assayedin left ventricular tissues from sham surgery animals.Total and MnSOD activity was higher in both exercise-trained groups, regardless of diet, compared to sedentarygroups (p � .05). While no differences in CAT activityexisted between groups, GPX activity was significantlyhigher in the antioxidant/trained group compared to allother groups (p � .05).
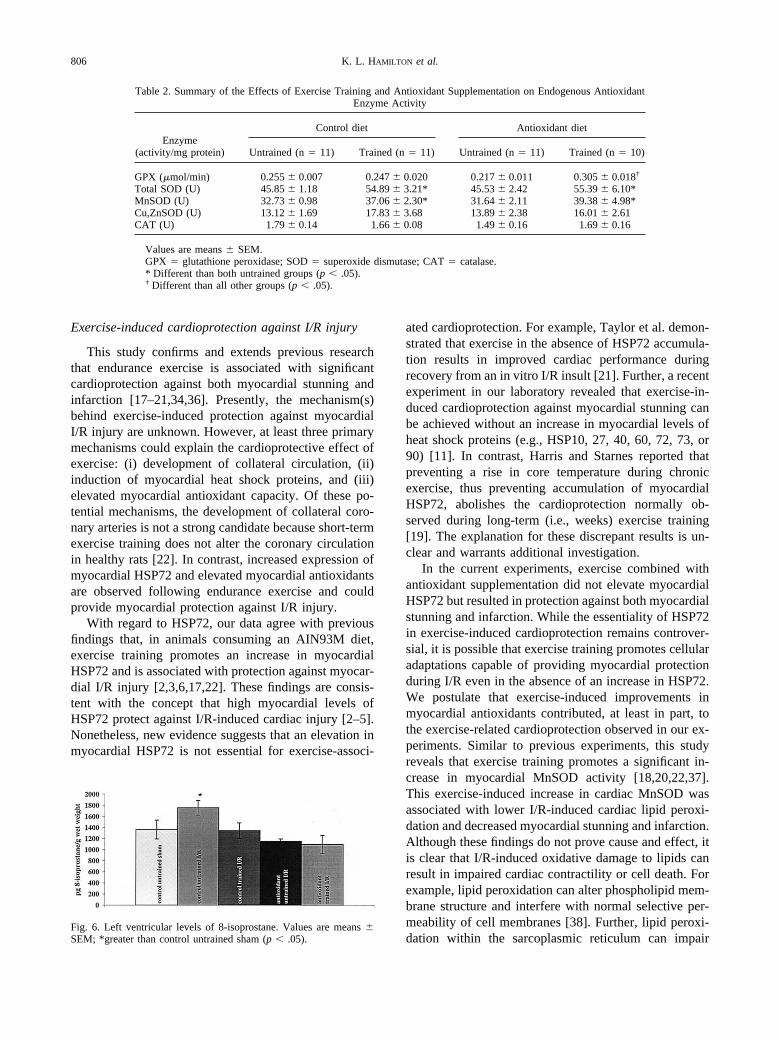
Figure 6 contains the mean (� SEM) values formyocardial 8-isoprostanes. I/R resulted in higher levelsof 8-isoprostanes in hearts from control diet/untrainedanimals compared to all other groups. Exercise and an-tioxidant supplementation resulted in protection of themyocardium from this I/R-induced lipid peroxidation (p� .05).
DISCUSSION
Overview of principal findings
This is the first experiment to investigate the com-bined effects of exercise training and antioxidant supple-mentation on in vivo cardioprotection against I/R-in-duced myocardial stunning and infarction. Our resultsindicate that while exercise and antioxidant supplemen-tation each provided independent cardioprotectionagainst I/R injury, the combination of these countermea-
Fig. 3. Analysis of long-duration I/R (60 min ischemia/120 min reperfusion) is presented. (a) Risk area expressed as a percentage ofthe heart; values are means � SEM; no differences among groups (p � .05). (b) Infarction area expressed as a percentage of the areaat risk; values are means � SEM; *less than control diet/untrained (p � .05). (c) Representative heart sections stained for infarctionfollowing long-term I/R. Blue-stained areas represent tissues that were well perfused during coronary occlusion. Brick red areasrepresent tissue that, despite ischemia, was not infarcted. Yellowish-white areas represent infarcted tissue (indicated by arrows). A �control diet/untrained, B � antioxidant diet/untrained, C � control diet/trained, and D � antioxidant diet/trained.
804 K. L. HAMILTON et al.
sures did not enhance or inhibit the myocardial protec-tion observed with each individual countermeasure. Animportant finding of this study was that antioxidant sup-plementation decreased the exercise-induced accumula-tion of myocardial HSP72, but this attenuation did not
diminish the cardioprotection associated with exercise.Finally, while antioxidant supplementation blunted exer-cise-induced increases in myocardial HSP72, antioxi-dants did not attenuate exercise-induced increases incardiac MnSOD activity.
Critique of experimental model
Adult female Sprague-Dawley rats were chosen asthe experimental subject because this strain does notdisplay large interanimal variations in coronary col-lateral circulation and is a widely accepted model forthe study of dietary antioxidant interventions and isch-emia/reperfusion studies [16,18,20,21]. While evi-dence exists that estrogen can function as an antioxi-dant [29,30], there is a paucity of convincing evidencethat male and female rats differ in myocardial adap-tation to exercise or in response to antioxidant supple-mentation [31,32]. We chose to study female ratsbecause of our long-term experience with exercisetraining and cardioprotection during I/R injury in fe-male animals. We also chose to use two models of I/Rinjury rather than using left ventricular developedpressure as the sole measure of cardioprotection giventhe potential limitations of ventricular developed pres-sure as a marker of tissue salvage [33].
The antioxidant diet used in these experiments wasmodeled after diets that have been shown to provideprotection against oxidative injury in a variety oftissues without apparent toxic effects [23,24]. Wechose an antioxidant diet with multiple antioxidants,rather than supplementing with single antioxidantssuch as vitamins C, E, or �-carotene as is frequentlydone, to provide a global antioxidant protection inboth the aqueous and lipid phases of cells. While thecomplex composition of the diet employed does notfacilitate identification of which constituent(s) wereresponsible for a given effect of the diet, this was nota goal of the study design.
It is likely that a specific countermeasure designed toprotect against myocardial I/R injury could provide car-diac protection against “minor” I/R injuries but wouldnot protect against a “major” I/R injury. Therefore, in thecurrent experiments, two durations of in vivo regionalischemia and reperfusion that result in different levels ofmyocardial injury were measured. Specifically, we chosethe 20 min duration of regional ischemia because thisperiod of regional ischemia results in myocardial stun-ning without significant infarction. In contrast, a long (60min) period of regional ischemia was used to produce alarge and significant myocardial infarction. These proce-dures have been used successfully in our laboratory forseveral years [15,16,18–20,34,35].
Fig. 4. Left ventricular HSP72 (muscle homogenate), with (a) a histo-gram comparing densitometric analyses of Western blots. So thatcomparisons could be made between separate blots, results are ex-pressed as a percentage of the mean control value within each blot(means � SEM); *greater than control diet/untrained (p � .05). (b)Representative Western blot for HSP72; lane 1 � control diet/un-trained, lane 2 � control diet/trained, lane 3 � antioxidant diet/untrained, and lane 4 � antioxidant diet/trained.
Fig. 5. Total antioxidant status: (a) total antioxidant status of plasma,and (b) total antioxidant status of left ventricular tissues. Values aremeans � SEM; *different than control/untrained and control/trained(p � .05); †different than control/trained (p � .05).
805Exercise, antioxidants, I/R
Exercise-induced cardioprotection against I/R injury
This study confirms and extends previous researchthat endurance exercise is associated with significantcardioprotection against both myocardial stunning andinfarction [17–21,34,36]. Presently, the mechanism(s)behind exercise-induced protection against myocardialI/R injury are unknown. However, at least three primarymechanisms could explain the cardioprotective effect ofexercise: (i) development of collateral circulation, (ii)induction of myocardial heat shock proteins, and (iii)elevated myocardial antioxidant capacity. Of these po-tential mechanisms, the development of collateral coro-nary arteries is not a strong candidate because short-termexercise training does not alter the coronary circulationin healthy rats [22]. In contrast, increased expression ofmyocardial HSP72 and elevated myocardial antioxidantsare observed following endurance exercise and couldprovide myocardial protection against I/R injury.
With regard to HSP72, our data agree with previousfindings that, in animals consuming an AIN93M diet,exercise training promotes an increase in myocardialHSP72 and is associated with protection against myocar-dial I/R injury [2,3,6,17,22]. These findings are consis-tent with the concept that high myocardial levels ofHSP72 protect against I/R-induced cardiac injury [2–5].Nonetheless, new evidence suggests that an elevation inmyocardial HSP72 is not essential for exercise-associ-
ated cardioprotection. For example, Taylor et al. demon-strated that exercise in the absence of HSP72 accumula-tion results in improved cardiac performance duringrecovery from an in vitro I/R insult [21]. Further, a recentexperiment in our laboratory revealed that exercise-in-duced cardioprotection against myocardial stunning canbe achieved without an increase in myocardial levels ofheat shock proteins (e.g., HSP10, 27, 40, 60, 72, 73, or90) [11]. In contrast, Harris and Starnes reported thatpreventing a rise in core temperature during chronicexercise, thus preventing accumulation of myocardialHSP72, abolishes the cardioprotection normally ob-served during long-term (i.e., weeks) exercise training[19]. The explanation for these discrepant results is un-clear and warrants additional investigation.
In the current experiments, exercise combined withantioxidant supplementation did not elevate myocardialHSP72 but resulted in protection against both myocardialstunning and infarction. While the essentiality of HSP72in exercise-induced cardioprotection remains controver-sial, it is possible that exercise training promotes cellularadaptations capable of providing myocardial protectionduring I/R even in the absence of an increase in HSP72.We postulate that exercise-induced improvements inmyocardial antioxidants contributed, at least in part, tothe exercise-related cardioprotection observed in our ex-periments. Similar to previous experiments, this studyreveals that exercise training promotes a significant in-crease in myocardial MnSOD activity [18,20,22,37].This exercise-induced increase in cardiac MnSOD wasassociated with lower I/R-induced cardiac lipid peroxi-dation and decreased myocardial stunning and infarction.Although these findings do not prove cause and effect, itis clear that I/R-induced oxidative damage to lipids canresult in impaired cardiac contractility or cell death. Forexample, lipid peroxidation can alter phospholipid mem-brane structure and interfere with normal selective per-meability of cell membranes [38]. Further, lipid peroxi-dation within the sarcoplasmic reticulum can impair
Table 2. Summary of the Effects of Exercise Training and Antioxidant Supplementation on Endogenous AntioxidantEnzyme Activity
Enzyme(activity/mg protein)
Control diet Antioxidant diet
Untrained (n � 11) Trained (n � 11) Untrained (n � 11) Trained (n � 10)
GPX (�mol/min) 0.255 � 0.007 0.247 � 0.020 0.217 � 0.011 0.305 � 0.018†
Total SOD (U) 45.85 � 1.18 54.89 � 3.21* 45.53 � 2.42 55.39 � 6.10*MnSOD (U) 32.73 � 0.98 37.06 � 2.30* 31.64 � 2.11 39.38 � 4.98*Cu,ZnSOD (U) 13.12 � 1.69 17.83 � 3.68 13.89 � 2.38 16.01 � 2.61CAT (U) 1.79 � 0.14 1.66 � 0.08 1.49 � 0.16 1.69 � 0.16
Values are means � SEM.GPX � glutathione peroxidase; SOD � superoxide dismutase; CAT � catalase.* Different than both untrained groups (p � .05).† Different than all other groups (p � .05).
Fig. 6. Left ventricular levels of 8-isoprostane. Values are means �SEM; *greater than control untrained sham (p � .05).
806 K. L. HAMILTON et al.

calcium handling and contribute to contractile dysfunc-tion during I/R [39]. Lipid peroxidation may also causedisturbances in mitochondrial function leading to dimin-ished oxidative phosphorylation and apoptosis [40,41].Collectively, these oxidant-mediated myocardial injuriescan contribute to both myocardial stunning and infarc-tion.
Nutritional antioxidants and protection against I/Rinjury
Another important finding in these experiments wasthat dietary antioxidants diminished cardiac injury asso-ciated with both short- and long-duration myocardialischemia. The fact that antioxidant supplementation at-tenuated both I/R-induced myocardial stunning and in-farction supports the notion that production of ROSduring I/R plays an important mechanistic role in boththe depressed myocardial contractility observed in stun-ning and cellular death in infarcted hearts; similar con-clusions have been reached by others [14,42].
Compared to control diet/untrained animals, antioxi-dant supplementation in sedentary animals protected theheart against myocardial stunning as evidenced by thehigher LVDP during reperfusion following ischemia.Further, our results suggest that the combination of ex-ercise and dietary antioxidants may be superior to dietaryantioxidants alone in the protection against myocardialstunning, particularly during reperfusion. The observa-tion that the level of myocardial protection against stun-ning did not differ between the control diet/exercisegroup and the antioxidant diet/exercise group indicatesthat exercise is a key component in providing protectionagainst myocardial stunning.
The antioxidant diet used in these experiments hasbeen shown to provide protection against oxidative in-jury in a variety of tissues [23,24]. Our dietary supple-mentation protocol resulted in enhanced total antioxidantcapacity of both plasma and left ventricular tissue. Im-portantly, our results also indicate that antioxidant sup-plementation does not alter the activity of any of theprimary antioxidant enzymes; similar results have beenreported by others [8,42].
We measured 8-isoprostane as a marker of lipid per-oxidation to determine if antioxidant supplementationdecreased left ventricular oxidative injury following I/R.Our data indicate that dietary antioxidant supplementa-tion decreases I/R-induced lipid peroxidation in vivo. Inreference to specific antioxidants and cardioprotection,several important “networking” antioxidants such asascorbic acid, vitamin E, glutathione, and ubiquinonesare depleted during I/R-induced myocardial oxidation[43]. Therefore, antioxidant supplementation may helpprevent this selective depletion of these antioxidants and
provide protection against I/R-induced lipid peroxida-tion.
Interaction between exercise and antioxidants
Again, our data indicate that nutritional antioxidantsor exercise provide independent protection against I/R-induced myocardial injury. Interestingly, the combina-tion of exercise and antioxidants did not impair or pro-vide an additive protection against infarction resultingfrom I/R. A potential explanation for this finding is thateach countermeasure (i.e., exercise or antioxidant sup-plementation) employed in our experiments providednear optimal protection against I/R injury (Figs. 2 and 3).Hence, additive effects of these countermeasures werenot possible because of the limited room for additionalprotection.
An important new finding in these experiments wasthat dietary supplementation with nutritional antioxi-dants resulted in the retardation of exercise-induced ac-cumulation of HSP72 in the heart. These results areconsistent with studies indicating that antioxidants caninhibit heat stress-induced expression of HSP72 in var-ious tissues [7,12,13]. Data exist, however, suggestingthat effects of antioxidants on HSP regulation are likelydependent on antioxidant selection, the redox sensitivityof each HSP isoform, the cell stress used, and the celltype [9–11,44]. To our knowledge, this is the first reportof the effect of an antioxidant diet on exercise-relatedinduction of stress proteins and cardioprotection.
The fact that antioxidant supplementation attenuatedthe exercise-associated accumulation of HSP72 is con-sistent with the notion that production of ROS duringexercise plays an important role in the exercise-associ-ated induction of HSP72. While the specific stress re-sponsible for HSP induction remains unknown, it hasbeen speculated that a host of physiochemical stressesassociated with exercise, including increases in temper-ature or calcium concentration, decreases in pH or oxy-gen content, exposure to oxidants, or any other stressescapable of damaging protein, may contribute to the ex-ercise-induced HSP72 accumulation [3]. Indeed, ROSplay an important role in the activation of heat shockfactor 1 (HSF1) and the elevation of HSP72 mRNA[8,45]. Furthermore, two recent reports indicate that anincrease in body temperature during exercise is manda-tory to achieve exercise-induced increases in myocardialHSP72 [18,21]. However, the current experiments indi-cate that an increase in body temperature alone duringexercise does not promote HSP72 induction. That is,enhancing myocardial antioxidant capacity via dietaryantioxidants attenuated the exercise-induced HSP72 ac-cumulation despite an increase in body temperature. It ispossible that the exercise-induced heat production is, in
807Exercise, antioxidants, I/R

part, responsible for production of ROS, which in turnserve as a signal for HSF activation [46]. Further studiesare needed to better understand the complex cellularevents resulting from exercise stress.
Another interesting observation in these experimentswas that, while dietary antioxidants prevented the exer-cise-induced increase in HSP72, antioxidants did notprevent an exercise-associated rise in myocardial Mn-SOD activity. Because MnSOD expression is thought tobe regulated at the transcriptional level via activation ofredox-sensitive transcription factors such as AP-1 andNF-�B [47–49], it was plausible that enhancing antiox-idant capacity would decrease MnSOD gene expression.Nonetheless, our results indicate that our antioxidantsupplementation does not impair the exercise-inducedincrease in myocardial MnSOD activity normally ob-served in animals maintained on a standard AIN93 diet.A potential explanation for this finding is that the limit-ing factor in exercise-enhanced MnSOD expression isnot at the transcriptional level but may be under post-transcriptional control. In support of this notion, recentstudies indicate that the stability of MnSOD transcriptscan be altered in both the basal and stimulated expressionof this gene [50]. It follows that increases in MnSODmRNA half-life could provide a mechanism for regulat-ing cellular protein and activity levels of MnSOD inresponse to exercise training. Furthermore, a specificMnSOD RNA-binding protein has been identified and isthought to be required for optimal translation [51,52].Hence, these collective data suggest that regulation ofMnSOD gene expression is complex and may occur atboth transcriptional and post-transcriptional levels.
In summary, these experiments demonstrate that ex-ercise training or antioxidant supplementation can inde-pendently provide protection against myocardial I/R in-jury. While antioxidant supplementation decreased theexercise-induced accumulation of myocardial HSP72,this attenuation did not diminish the cardioprotectionassociated with exercise. An exercise-related increase inMnSOD activity persisted despite enhanced antioxidantcapacity. The complexity of the mechanisms related toexercise-induced cardioprotection warrant further inves-tigation into the regulation of gene expression followingendurance exercise.
Acknowledgements — This work was supported by a grant-in-aid(S.K.P.) from the American Heart Association, Florida Affiliate.
REFERENCES
[1] Benjamin, I. J.; McMillan, D. R. Stress (heat shock) proteins:molecular chaperones in cardiovascular biology and disease. Circ.Res. 83:117–132; 1998.
[2] Locke, M. The cellular stress response to exercise: role of stressproteins. Exerc. Sport Sci. Rev. 25:105–136; 1997.
[3] Powers, S. K.; Locke, M.; Demirel, H. A. Exercise, heat shockproteins, and myocardial protection from I/R injury. Med. Sci.Sports Exerc. 33:386–392; 2001.
[4] Hutter, J. J.; Mestril, R.; Tam, E. K.; Sievers, R. E.; Dillmann,W. H.; Wolfe, C. L. Overexpression of heat shock protein 72 intransgenic mice decreases infarct size in vivo. Circulation 94:1408–1411; 1996.
[5] Radford, N. B.; Fina, M.; Benjamin, I. J.; Moreadith, R. W.;Graves, K. H.; Zhao, P.; Gavva, S.; Wiethoff, A.; Sherry, A. D.;Malloy, C. R.; Williams, R. S. Cardioprotective effects of 70-kDaheat shock protein in transgenic mice. Proc. Natl. Acad. Sci. USA93:2339–2342; 1996.
[6] Locke, M.; Noble, E. G.; Tanguay, R. M.; Feild, M. R.; Ianuzzo,S. E.; Ianuzzo, C. D. Activation of heat-shock transcription factorin rat heart after heat shock and exercise. Am. J. Physiol. 268:C1387–C1394; 1995.
[7] Gorman, A. M.; Heavey, B.; Creagh, E.; Cotter, T. G.; Samali, A.Antioxidant-mediated inhibition of the heat shock response leadsto apoptosis. FEBS Lett. 445:98–102; 1999.
[8] Nishizawa, J.; Nakai, A.; Matsuda, K.; Komeda, M.; Ban, T.;Nagata, K. Reactive oxygen species play an important role in theactivation of heat shock factor 1 in ischemic-reperfused heart.Circulation 99:934–941; 1999.
[9] Peng, J.; Jones, G. L.; Watson, K. Stress proteins as biomarkers ofoxidative stress: effects of antioxidant supplements. Free Radic.Biol. Med. 28:1598–1606; 2000.
[10] Pinot, F.; el Yaagoubi, A.; Christie, P.; Dinh-Xuan, A. T.; Polla,B. S. Induction of stress proteins by tobacco smoke in humanmonocytes: modulation by antioxidants. Cell Stress Chaperones2:156–161; 1997.
[11] Morino, M.; Tsuzuki, T.; Ishikawa, Y.; Shirakami, T.; Yoshimura,M.; Kiyosuke, Y.; Matsunaga, K.; Yoshikumi, C.; Saijo, N. Spe-cific regulation of HSPs in human tumor cell lines by flavonoids.In Vivo 11:265–270; 1997.
[12] Westman, J.; Drieu, K.; Sharma, H. S. Antioxidant compoundsEGB-761 and BN-520 21 attenuate heat shock protein (HSP 72kD) response, edema and cell changes following hyperthermicbrain injury. An experimental study using immunohistochemistryin the rat. Amino Acids 19:339–350; 2000.
[13] Ushakova, T.; Melkonyan, H.; Nikonova, L.; Mudrik, N.; Gog-vadze, V.; Zhukova, A.; Gaziev, A. I.; Bradbury, R. The effect ofdietary supplements on gene expression in mice tissues. FreeRadic. Biol. Med. 20:279–284; 1996.
[14] Bolli, R.; Marban, E. Molecular and cellular mechanisms ofmyocardial stunning. Physiol. Rev. 79:609–634; 1999.
[15] Coombes, J. S.; Powers, S. K.; Demirel, H. A.; Hamilton, K. L.;Jessup, J.; Vincent, H. K.; Shanely, R. A. Vitamin E deficiencyfails to affect myocardial performance during in vivo ischemia/reperfusion. Int. J. Vitam. Nutr. Res. 70:293–300; 2000.
[16] Coombes, J. S.; Powers, S. K.; Demirel, H. A.; Jessup, J.; Vin-cent, H. K.; Hamilton, K. L.; Naito, H.; Shanely, R. A.; Sen,C. K.; Packer, L.; Ji, L. L. Effect of combined supplementationwith vitamin E and �-lipoic acid on myocardial performanceduring in vivo ischemia/reperfusion. Acta Physiol. Scand. 169:261–269; 2000.
[17] Locke, M.; Tanguay, R. M.; Klabunde, R. E.; Ianuzzo, C. D.Enhanced postischemic myocardial recovery following exerciseinduction of HSP 72. Am. J. Physiol. 269:H320–H325; 1995.
[18] Hamilton, K. L.; Powers, S. K.; Sugiura, T.; Kim, S.; Lennon, S.;Tumer, N.; Mehta, J. L. Short-term exercise training can improvemyocardial tolerance to I/R without elevation in heat shock pro-teins. Am. J. Physiol. Heart Circ. Physiol. 281:H1346–H1352;2001.
[19] Harris, M. B.; Starnes, J. W. Effects of body temperature duringexercise training on myocardial adaptations. Am. J. Physiol. HeartCirc. Physiol. 280:H2271–H2280; 2001.
[20] Powers, S. K.; Demirel, H. A.; Vincent, H. K.; Coombes, J. S.;Naito, H.; Hamilton, K. L.; Shanely, R. A.; Jessup, J. Exercisetraining improves myocardial tolerance to in vivo ischemia/reper-fusion in the rat. Am. J. Physiol. 275:R1468–R1477; 1998.
[21] Taylor, R. P.; Harris, M. B.; Starnes, J. W. Acute exercise can
808 K. L. HAMILTON et al.

improve cardioprotection without increasing heat shock proteincontent. Am. J. Physiol. 276:H1098–H1102; 1999.
[22] Yamashita, N.; Hoshida, S.; Otsu, K.; Asahi, M.; Kuzuya, T.;Hori, M. Exercise provides direct biphasic cardioprotection viamanganese superoxide dismutase activation. J. Exp. Med. 189:1699–1706; 1999.
[23] North, J. A.; Tappel, A. L. Measuring the oxidation of hemecompounds in heart homogenates from rats supplemented withdietary antioxidants. Free Radic. Biol. Med. 22:175–184; 1997.
[24] Chen, H.; Tappel, A. L. Protection of vitamin E, selenium, troloxC, ascorbic acid palmitate, acetylcysteine, coenzyme Q0, coen-zyme Q10, �-carotene, canthaxanthin, and (�)-catechin againstoxidative damage to rat blood and tissues in vivo. Free Radic.Biol. Med. 18:949–953; 1995.
[25] Fishbein, M. C.; Meerbaum, S.; Rit, J.; Lando, U.; Kanmatsuse,K.; Mercier, J. C.; Corday, E.; Ganz, W. Early phase acutemyocardial infarct size quantification: validation of the triphenyltetrazolium chloride tissue enzyme staining technique. Am.Heart J. 101:593–600; 1981.
[26] McCord, J. M.; Fridovich, I. The reduction of cytochrome c bymilk xanthine oxidase. J. Biol. Chem. 243:5753–5760; 1968.
[27] Flohe, L.; Gunzler, W. A. Assays of glutathione peroxidase.Methods Enzymol. 105:114–121; 1984.
[28] Aebi, H. Catalase in vitro. Methods Enzymol. 105:121–126; 1984.[29] Barp, J.; Araujo, A. S.; Fernandes, T. R.; Rigatto, K. V.; Llesuy,
S.; Bello-Klein, A.; Singal, P. Myocardial antioxidant and oxida-tive stress changes due to sex hormones. Braz. J. Med. Biol. Res.35:1075–1081; 2002.
[30] Thibodeau, P.; Kachadourian, R.; Lemay, R.; Bisson, M.; Day,B.; Paquette, B. In vitro pro- and antioxidant properties of estro-gens. J. Steroid Biochem. Mol. Biol. 81:227; 2002.
[31] Paroo, Z.; Haist, J. V.; Karmazyn, M.; Noble, E. G. Exerciseimproves postischemic cardiac function in males but not females:consequences of a novel sex-specific heat shock protein 70 re-sponse. Circ. Res. 90:911–917; 2002.
[32] Yamamoto, T.; Ohkuwa, T.; Itoh, H.; Sato, Y.; Naoi, M. Effect ofgender differences and voluntary exercise on antioxidant capacityin rats. Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 132:437;2002.
[33] Gelpi, R. J.; Morales, C.; Cohen, M. V.; Downey, J. M. Xanthineoxidase contributes to preconditioning’s preservation of left ven-tricular developed pressure in isolated rat heart: developed pres-sure may not be an appropriate end-point for studies of precon-ditioning. Basic Res. Cardiol. 97:40–46; 2002.
[34] Demirel, H. A.; Powers, S. K.; Zergeroglu, M. A.; Shanely, R. A.;Hamilton, K.; Coombes, J.; Naito, H. Short-term exercise im-proves myocardial tolerance to in vivo ischemia/reperfusion in therat. J. Appl. Physiol. 91:2205–2212; 2001.
[35] Demirel, H. A.; Powers, S. K.; Caillaud, C.; Coombes, J. S.;Naito, H.; Fletcher, L. A.; Vrabas, I.; Jessup, J. V.; Ji, L. L.Exercise training reduces myocardial lipid peroxidation followingshort-term ischemia/reperfusion. Med. Sci. Sports Exerc.30:1211–1216; 1998.
[36] Yamashita, N.; Baxter, G. F.; Yellon, D. M. Exercise directlyenhances myocardial tolerance to ischemia/reperfusion injury inthe rat through a protein kinase C mediated mechanism. Heart85:331–336; 2001.
[37] Powers, S. K.; Criswell, D.; Lawler, J.; Martin, D.; Lieu, F. K.; Ji,L. L.; Herb, R. A. Rigorous exercise training increases superoxidedismutase activity in ventricular myocardium. Am. J. Physiol.265:H2094–H2098; 1993.
[38] Massey, K. D.; Burton, K. P. Alpha-tocopherol attenuates myo-cardial membrane-related alterations resulting from ischemia andreperfusion. Am. J. Physiol. 256:H1192–H1199; 1989.
[39] Romaschin, A. D.; Wilson, G. J.; Thomas, U.; Feitler, D. A.;Tumiati, L.; Mickle, D. A. Subcellular distribution of peroxidized
lipids in myocardial reperfusion injury. Am. J. Physiol. 259:H116–H123; 1990.
[40] Pepe, S. Mitochondrial function in ischemia and reperfusion ofthe aging heart. Clin. Exp. Pharmacol. Physiol. 27:745–750;2000.
[41] Paradies, G.; Petrosillo, G.; Pistolese, M.; Di Venosa, N.; Serena,D.; Ruggiero, F. M. Lipid peroxidation and alterations to oxida-tive metabolism in mitochondria isolated from rat heart subjectedto ischemia and reperfusion. Free Radic. Biol. Med. 27:42–50;1999.
[42] Castenmiller, J. J.; Lauridsen, S. T.; Dragsted, L. O.; van het Hof,K. H.; Linssen, J. P.; West, C. E. Beta-carotene does not changemarkers of enzymatic and nonenzymatic antioxidant activity inhuman blood. J. Nutr. 129:2162–2169; 1999.
[43] Haramaki, N.; Stewart, D. B.; Aggarwal, S.; Ikeda, H.; Reznick,A. Z.; Packer, L. Networking antioxidants in the isolated rat heartare selectively depleted by ischemia/reperfusion. Free Radic.Biol. Med. 25:329–339; 1998.
[44] Arnaud, C.; Joyeux, M.; Garrel, C.; Godin-Ribuot, D.; Demenge,P.; Ribuot, C. Free radical production triggered by hyperthermiacontributes to heat stress-induced cardioprotection in isolated rathearts. Br. J. Pharmacol. 135:1776–1782; 2002.
[45] Kukreja, R. C.; Kontos, M. C.; Loesser, K. E.; Batra, S. K.; Qian,Y. Z.; Gbur, C. J. Jr.; Naseem, S. A.; Jesse, R. L.; Hess, M. L.Oxidant stress increases heat shock protein 70 mRNA in isolatedperfused rat heart. Am. J. Physiol. 267:H2213–H2219; 1994.
[46] Zuo, L.; Christofi, F. L.; Wright, V. P.; Liu, C. Y.; Merola, A. J.;Berliner, L. J.; Clanton, T. L. Intra- and extracellular measure-ment of reactive oxygen species produced during heat stress indiaphragm muscle. Am. J. Physiol. Cell Physiol. 279:C1058–C1066; 2000.
[47] Sun, Y.; Oberley, L. W. Redox regulation of transcriptionalactivators. Free Radic. Biol. Med. 21:335–348; 1996.
[48] Kiningham, K. K.; Xu, Y.; Daosukho, C.; Popova, B.; St. Clair,D. K. Nuclear factor �B-dependent mechanisms coordinate thesynergistic effect of PMA and cytokines on the induction ofsuperoxide dismutase 2. Biochem. J. 353:147–156; 2001.
[49] Fujii, J.; Taniguchi, N. Phorbol ester induces manganese-super-oxide dismutase in tumor necrosis factor-resistant cells. J. Biol.Chem. 266:23142–23146; 1991.
[50] Davis, C. A.; Monnier, J. M.; Nick, H. S. A coding regiondeterminant of instability regulates levels of manganese superox-ide dismutase mRNA. J. Biol. Chem. 276:37317–37326; 2001.
[51] Knirsch, L.; Clerch, L. B. Tyrosine phosphorylation regulatesmanganese superoxide dismutase (MnSOD) RNA-binding proteinactivity and MnSOD protein expression. Biochemistry 40:7890–7895; 2001.
[52] Chung, D. J.; Wright, A. E.; Clerch, L. B. The 3' untranslatedregion of manganese superoxide dismutase RNA contains a trans-lational enhancer element. Biochemistry 37:16298–16306; 1998.
ABBREVIATIONS
CAT—catalaseCu,ZnSOD—copper-zinc superoxide dismutaseGPX—glutathione peroxidaseHSP—heat shock proteinI/R—ischemia/reperfusionLVDP—left ventricular developed pressureMnSOD—manganese superoxide dismutaseROS—reactive oxygen speciesVF—ventricular fibrillation
809Exercise, antioxidants, I/R
Copyright © 2022 FDOKUMEN




















