
Evolution of the hominoid vertebral column: the long and the short of it
18
ARTICLE Evolution of the Hominoid Vertebral Column: The Long and the Short of It SCOTT A. WILLIAMS AND GABRIELLE A. RUSSO The postcranial axial skeleton exhibits considerable morphological and func- tional diversity among living primates. Particularly striking are the derived features in hominoids that distinguish them from most other primates and mammals. In contrast to the primitive catarrhine morphotype, which presumably possessed an external (protruding) tail and emphasized more pronograde trunk posture, all living hominoids are characterized by the absence of an external tail and adaptations to orthograde trunk posture. Moreover, modern humans evolved unique vertebral features that satisfy the demands of balancing an upright torso over the hind limbs during habitual terrestrial bipedalism. Our ability to identify the evolutionary timing and understand the functional and phylogenetic significance of these fun- damental changes in postcranial axial skeletal anatomy in the hominoid fossil record is key to reconstructing ancestral hominoid patterns and retracing the evo- lutionary pathways that led to living apes and modern humans. Here, we provide an overview of what is known about evolution of the hominoid vertebral column, focusing on the currently available anatomical evidence of three major transitions: tail loss and adaptations to orthograde posture and bipedal locomotion. TAIL LOSS The logical evolutionary event with which to begin this review is the loss of an external tail, since taillessness is one of the earliest identifiable modern ape-like features of the post- cranial axial skeleton. The number of postsacral vertebrae in primates is highly variable and can reach over 30 in long-tailed taxa. 1 However hominoids typically exhibit only two to four postsacral elements. Because all living hominoids lack tails, it is assumed that all extinct hominoids also lacked tails. Proconsul hesloni, the earliest known hominoid (the closest to the estimated divergence date of cercopi- thecoids and hominoids at 25-30 Ma) 6 with adequate postcranial material for inferring tail length, indi- cates that taillessness evolved by at least 18.5 Ma. 2–5 The morphology of a last sacral vertebra – which distally articulates with the first postsacral (caudal or coccygeal) vertebra – belonging to a subadult Proconsul heseloni (KNM-KPS V42) individual is similar to that of last sacral verte- brae in living hominoids, and differs from those of tail-bearing prima- tes. 2–4,7,8 These differences are illus- trated in Figure 1 (A-C), which compares the distal sacra of a long- tailed primate (Macaca fascicularis), an extant hominoid (Pan troglodytes), and the Proconsul sacral specimen (see also Ward and colleagues 3 for a comparison with a subadult Pan indi- vidual). Although the Proconsul speci- men suffers from some distortion and breakage (particularly on the left side), its caudal articular surface and right lateral side are well preserved. Compared to long-tailed primates, liv- ing hominoids and Proconsul exhibit last sacral vertebral bodies that are mediolaterally narrower at their cau- dal ends than at their cranial ends, have dorsoventrally narrower distal articular surfaces with reduced sur- face areas, and less laterally expanded transverse processes (right side in Proconsul); they also lack postzygapophyses, which would oth- erwise articulate with the prezyga- pophyses of the first tail vertebra (Fig. 1D). 2–4,7,8 Comparative research confirms the association of these fea- tures with tail loss, demonstrating that tailless nonprimate mammals converge with, or trend toward, the ape distal sacral condition. 4 Nacholapithecus kerioi (KNM-BG 40949) provides later evidence of taillessness in the Middle Miocene (15 Ma). 9 Compared to the first postsacral vertebrae of primates and mammals with long tails, those of living hominoids (Fig. 1E) and Nacholapithecus (Fig. 1F) have more elliptically shaped (mediolaterally wide, dorsoventrally narrow) and more caudally tapered bodies that bear shallow dorsal grooves (instead of neural arches), as well as reduced Scott Williams is an Assistant Professor in the Department of Anthropology at New York University. He studies the ver- tebral column and other skeletal systems and is primarily interested in the evolu- tion of upright posture and bipedalism. [email protected]. Gabrielle Russo is an Assistant Professor in the Department of Anthropology at Stony Brook University. She studies the primate locomotor skeleton. Her current work focuses on the evolution of the axial skeleton. She is particularly inter- ested in questions concerning tail func- tion and loss, and adaptations to orthogrady and bipedalism. Gabrielle. [email protected]. Key words: spine; tail; orthogrady; hominin; bipedalism V C 2015 Wiley Periodicals, Inc. DOI: 10.1002/evan.21437 Published online in Wiley Online Library (wileyonlinelibrary.com). Evolutionary Anthropology 24:15–32 (2015)
Transcript of Evolution of the hominoid vertebral column: the long and the short of it

ARTICLE
Evolution of the Hominoid Vertebral Column:The Long and the Short of ItSCOTT A. WILLIAMS AND GABRIELLE A. RUSSO
The postcranial axial skeleton exhibits considerable morphological and func-tional diversity among living primates. Particularly striking are the derived featuresin hominoids that distinguish them from most other primates and mammals. Incontrast to the primitive catarrhine morphotype, which presumably possessed anexternal (protruding) tail and emphasized more pronograde trunk posture, all livinghominoids are characterized by the absence of an external tail and adaptations toorthograde trunk posture. Moreover, modern humans evolved unique vertebralfeatures that satisfy the demands of balancing an upright torso over the hindlimbs during habitual terrestrial bipedalism. Our ability to identify the evolutionarytiming and understand the functional and phylogenetic significance of these fun-damental changes in postcranial axial skeletal anatomy in the hominoid fossilrecord is key to reconstructing ancestral hominoid patterns and retracing the evo-lutionary pathways that led to living apes and modern humans. Here, we providean overview of what is known about evolution of the hominoid vertebral column,focusing on the currently available anatomical evidence of three major transitions:tail loss and adaptations to orthograde posture and bipedal locomotion.
TAIL LOSS
The logical evolutionary event withwhich to begin this review is the loss
of an external tail, since taillessnessis one of the earliest identifiablemodern ape-like features of the post-cranial axial skeleton. The number ofpostsacral vertebrae in primates ishighly variable and can reach over30 in long-tailed taxa.1 Howeverhominoids typically exhibit only twoto four postsacral elements. Becauseall living hominoids lack tails, it isassumed that all extinct hominoidsalso lacked tails.
Proconsul hesloni, the earliestknown hominoid (the closest to theestimated divergence date of cercopi-thecoids and hominoids at �25-30Ma)6 with adequate postcranialmaterial for inferring tail length, indi-cates that taillessness evolved by atleast 18.5 Ma.2–5 The morphology ofa last sacral vertebra – which distallyarticulates with the first postsacral(caudal or coccygeal) vertebra –belonging to a subadult Proconsulheseloni (KNM-KPS V42) individualis similar to that of last sacral verte-brae in living hominoids, and differs
from those of tail-bearing prima-tes.2–4,7,8 These differences are illus-trated in Figure 1 (A-C), whichcompares the distal sacra of a long-tailed primate (Macaca fascicularis),an extant hominoid (Pan troglodytes),and the Proconsul sacral specimen(see also Ward and colleagues3 for acomparison with a subadult Pan indi-vidual). Although the Proconsul speci-men suffers from some distortionand breakage (particularly on the leftside), its caudal articular surface andright lateral side are well preserved.Compared to long-tailed primates, liv-ing hominoids and Proconsul exhibitlast sacral vertebral bodies that aremediolaterally narrower at their cau-dal ends than at their cranial ends,have dorsoventrally narrower distalarticular surfaces with reduced sur-face areas, and less laterallyexpanded transverse processes (rightside in Proconsul); they also lackpostzygapophyses, which would oth-erwise articulate with the prezyga-pophyses of the first tail vertebra(Fig. 1D).2–4,7,8 Comparative researchconfirms the association of these fea-tures with tail loss, demonstratingthat tailless nonprimate mammalsconverge with, or trend toward, theape distal sacral condition.4
Nacholapithecus kerioi (KNM-BG40949) provides later evidence oftaillessness in the Middle Miocene(�15 Ma).9 Compared to the firstpostsacral vertebrae of primates andmammals with long tails, those ofliving hominoids (Fig. 1E) andNacholapithecus (Fig. 1F) have moreelliptically shaped (mediolaterallywide, dorsoventrally narrow) andmore caudally tapered bodies thatbear shallow dorsal grooves (insteadof neural arches), as well as reduced
Scott Williams is an Assistant Professorin the Department of Anthropology atNew York University. He studies the ver-tebral column and other skeletal systemsand is primarily interested in the evolu-tion of upright posture and [email protected].
Gabrielle Russo is an Assistant Professorin the Department of Anthropology atStony Brook University. She studies theprimate locomotor skeleton. Her currentwork focuses on the evolution of theaxial skeleton. She is particularly inter-ested in questions concerning tail func-tion and loss, and adaptations toorthogrady and bipedalism. [email protected].
Key words: spine; tail; orthogrady; hominin;
bipedalism
VC 2015 Wiley Periodicals, Inc.DOI: 10.1002/evan.21437Published online in Wiley Online Library(wileyonlinelibrary.com).
Evolutionary Anthropology 24:15–32 (2015)

transverse processes; they also lackzygapophyseal articular proc-esses.2,4,9–11 In contrast, first postsac-ral vertebrae in primates andmammals with long tails exhibitmore circularly shaped articularsurfaces, well-developed transverseprocesses, neural arches bearing dor-sally projecting spinous processes,and prezygapophyses that articulatewith the postzygapophyses of thedistal sacrum to form a mobile sac-rocaudal joint (Fig. 1D).4,11
Tail loss is also reflected in themorphology of presacral vertebrae.Primates and other mammals withlong tails bear well-developed ana-pophyses (styloid processes) thatproject caudally from the inferiorpedicle ventrolateral to the postzyga-pophyses of lower thoracic andupper lumbar vertebrae, anchoringlongissimus and a common tendonfor extensor caudae lateralis (a basaltail extensor muscle) and intertrans-versarius (Fig. 2). Well-developed sty-loid processes may extend to orbeyond the caudal extent of the post-zygapophyses (Fig. 2A) and canimpinge on the subjacent prezyga-pophyses, limiting extension and lat-eral rotation of lower thoracic andupper lumbar vertebrae.12,13 Because
anapophysis morphology has astrong relationship with the lumbartransverse processes, the more dor-sally positioned transverse processesand associated erector spinae mus-
culature of hominoids, especiallyhominids, prohibit the existence oflong, functional styloid processes.14
Their remnants, sometimes referredto as accessory processes, are oftenobserved on the bases of the trans-verse processes of hominoid upperlumbar vertebrae and anchor onlylongissimus (Fig. 2B, C).12,14
Although styloid processes are largeon a last thoracic vertebra and firstlumbar vertebra of P. nyanzae, thoseon middle and lower lumbar levelsare weakly developed.14 In P. hese-loni and N. kerioi, none of the lower
thoracic or lumbar vertebrae bearfunctional styloid processes and onlysmall anapophyses are present.10,15
Similarly, Morotopithecus bishopiand Pierolapithecus catalaunicus lackstyloid processes, but may possessremnants of anapophyses.14,16–19
It is widely recognized that tailless-ness appears early in hominoid evo-lution, but questions about why tailloss evolved remain unresolved.Early researchers suggested a directlink between the evolution of tailless-ness and orthogrady, hypothesizingthat tail loss was associated with areduction and reorganization of thepelvicaudal musculature in order tosupport the abdominal and pelvicviscera during upright positionalbehaviors.20 However, although fea-tures of the P. heseloni distal sacrumand N. kerioi first postsacral vertebraindicate a reduction in mobility atthe sacrococcygeal joint anddecreased bony leverage of the pelvi-caudal musculature otherwiseresponsible for moving a tail, fossilevidence indicates decoupling of theevolution of tail loss and orthog-rady.4 Taillessness appeared first inthe context of pronograde trunk pos-ture—both P. nyanzae and N. kerioiexhibit pronograde-adapted axial
Figure 1. Comparison of distal sacra and first postsacral vertebrae in catarrhines. Top row shows sacra for A) an extant long-tailed cerco-pithecoid (Macaca fascicularis), B) an extant hominoid (Pan troglodytes), and C) Proconsul heseloni (image modified from Figure 1 inWard, Walker and Teaford3). Bottom row shows first sacral vertebrae for A) Macaca fascicularis, B) Pan troglodytes, and C) Nacholapi-thecus (modified from Nakatsukasa and coworkers9). Note that the distal sacrum of Macaca fascicularis has laterally-expanded trans-verse processes and well-developed postzygapophyses for articulation for the prezygapophyses of a robust first postsacral vertebra,whereas the distal sacra of Pan and Proconsul bear reduced transverse processes and vestigial sacral cornuae which approach, but donot make articular contact with, the proximal cornuae (if present) of first postsacral vertebrae. Distal sacrum for extant taxa is croppedat mid-height of the last sacral foramina. The Proconsul element is an isolated last sacral vertebra. Black bars approximately 1 cm.
It is widely recognizedthat taillessness appearsearly in hominoid evolu-tion, but questions aboutwhy tail loss evolvedremain unresolved.
16 WILLIAMS AND RUSSO ARTICLE

morphology, including longer lum-bar vertebral regions (6-7 lumbarvertebrae) and primitive morpholo-gies of the vertebral body and neuralarch (see below)—and well beforethe adoption of adaptations toorthograde trunk postures and loco-motor behaviors. Moreover, N. kerioiprobably retained a sacrum with theprimitive primate number of sacralvertebrae, and although sacral verte-brae are not known well enough toaccurately determine sacral numbersin Proconsul,21 the P. nyanzae partialfirst sacral vertebra articulated witha pelvis had blades oriented in theparasagittal (parallel to the midsagit-talplane) plane, as in many monkeys.
The combination of taillessnessand pronograde-adapted axial mor-phology in early hominoids is animportant consideration because themammalian tail is often recruited formaintaining balance during arborealquadrupedal progression or forassisting in feeding postures.22–24
Given the body masses estimated forProconsul (for example, P. heseloni,10-20 kg; P. nyanzae, 20-50 kg), the
presence of a tail for maintainingstability on small branches (relativeto body size) should confer an adapt-ive advantage. It is possible that thetail would have to be very large forits mass to effectively stabilize ananimal having a body size similar tothat of P. heseloni or P. nyanzae dur-ing quadrupedalism. Therefore, thefunctional demand for dynamic sta-bilization during arboreal quadrupe-dal locomotion may have been takenover by enhanced grasping abilitiesand limb mobility that permittedcareful maneuvering of the bodyover secure handholds and foot-holds25 (but see MacLatchy26). Theuse of slow, deliberate locomotion,as suggested for Proconsul, mightalso be viewed as a compensatorymechanism for the combination oflarge body size and tail loss.25,27–29
Greater insight into why tail lossevolved in hominoids will probably begained only by exploring the occur-rence of tail length reduction and lossacross different primate and mamma-lian lineages, and by greater consider-ation of its various functional roles.
Among nonhominoid primates, sev-eral taxa, including Macaca sylvanus(the “Barbary ape”) and some lorisidssuch as Nycticebus lack external tails.Reduced tail lengths also appear tohave characterized some extinct pri-mates. Based on sacral and postsacralvertebral evidence, Palaeopropithecusand Megaladapis probably had shortertails than did Archaeolemur or Hadro-pithecus4,30,31; also, at least one pliopi-thecoid (Epipliopithecusvindobonensis) probably possessed atail less than half its total head andbody length (but see Ankel7). It is pos-sible that each primate lineageevolved tail loss or length reductionfor a different reason. Some of thehypotheses put forth include terres-triality, increased body size, and cli-mate (Allen’s rule).32 Reasons whytails were reduced in length or lostwithin multiple primate lineages thusawait further exploration.
NUMBERS OF VERTEBRAE
In conjunction with absence of anexternal tail, hominoids also exhibit
GLOSSARY
Homeotic — the transformationof one segment into a different seg-ment. Homeotic changes in verte-bral identity occur at vertebralboundaries, such as the lumbo-sacral border, and are associatedwith Homeobox (Hox) gene expres-sion. The vertebral precursors, orsomites, take on cervical, thoracic,lumbar, sacral, or coccygeal fea-tures during early development,depending on their anteroposterioraxis of the embryo and Hox geneexpression. Homeotic shifts in ver-tebral boundaries can be classifiedas cranially-directed or caudallydirected, depending on the direc-tion of change in regional identity.
Meristic — the addition or sub-traction of segments. Meristicchanges in vertebrae occur by theproduction of extra somites orreduction in the number of somitesthat are produced. Meristicchanges result in an increased or
decreased total number ofvertebrae.
Orthograde/orthogrady — uprightposture, usually in reference tothe body’s torso. In orthogrady,the long axis of the torso is paral-lel to the line of gravity.
Pronograde/pronogrady — hori-zontal posture, usually in referenceto the body’s torso. In pronogrady,the long axis of the torso is perpen-dicular to the line of gravity.
Sacralization — incorporationof vertebrae into the sacrum.Sacralization can occur pathologi-cally (during an individual’s life) aslumbar or caudal vertebrae fuse tothe sacrum, but it occurs evolutio-narily via cranial homeotic shifts insomite identity at the lumbo-sacralborder and caudally-directedchange at the sacro-caudal border.Hominoid primates have experi-enced evolutionary sacralization,either by the former or the latter
(contrast Figures 3 and 4), or by acombination of both processes.
Somites — a series of paired,blocklike segments of paraxial mes-oderm that flank the neural tube inthe developing embryo. Somitesare the unsegmented precursors ofvertebrae and give rise to othermesenchymal tissues.
Zygapophyses — cranially andcaudally directed processes thatrise from the neural arch and bearthe articular facets of vertebrae.Prezygapophyses are situated onthe cranial aspect of the neuralarch and bear the superior articu-lar facets, which articulate with thesupradjacent vertebrae (or occipitalcondyles if C1). Postzygapophysesarise from the caudal aspect of theneural arch and bear the inferiorarticular facets, which articulatewith subjacent vertebrae (or thesuperior articular processes of thesacrum).
ARTICLE Evolution of the Hominoid Vertebral Column 17

Figure 2. Comparison of upper-level lumbar vertebrae in dorsal (left column), lateral (middle column), and caudal (right column) views.Species shown are Macaca mulatta (A), Hylobates lar (B), Pan troglodytes (C), and Homo sapiens (D). Arrows identify styloid (in A) orremnant anapophyses (B-D) in all views. Note that the styloid processes in Macaca are long, approximating the length of the postzyga-pophyses, arise from the caudal aspects of the pedicles, and would act as a bony stop during particular intervertebral movements;the anapophyses of hominoids are associated with (B) and arise directly from (C-D) the inferior aspects of the lumbar transverse proc-esses. Styloid processes anchor longissimus and a common tendon for extensor caudae lateralis and intertransversarius, whereas longis-simus attaches along the inferior aspects of the lumbar transverse processes in hominoids.
18 WILLIAMS AND RUSSO ARTICLE

reduced numbers of lumbar (andtotal thoracolumbar) vertebrae com-pared to the primitive catarrhinecondition. Thus, a brief discussionof the developmental processes thatcontrol the formation and identifi-cation of vertebrae is warranted.Segmentation, or the determinationof total vertebral count, occursthrough somitogenesis, the produc-tion of somites from presomiticmesoderm, the speed of which isdetermined by a “segmentationclock.”33 The speed of that clockdetermines the number and size ofthe somites that are produced; thefaster the clock, the smaller andmore numerous are the resultingsomites.34 Somites are produced atthe rate of the segmentation clockuntil the presomitic mesoderm isexhausted. Vertebrae are thenderived from the somites through aprocess called resegmentation.33,35
While changes in segmentationinvolve meristic changes, or changesin the total number of somites pro-duced, changes in specification, theregionalization of vertebrae into oneof several types,35,36 are homeotic innature and involve change in theidentity of a somite (for example, ashift from thoracic to lumbar iden-tity). In this way, both cranially-directed and caudally-directed shiftsin identity can occur, contributing toa decreased number of resulting ver-tebrae in the region toward whichthe shift is directed and a concomi-tant increase in the region awayfrom which the shift occurs. Home-otic changes are associated withalterations in the expression of Hoxgenes.36 Mutations of this nature cancause homeotic transformations, ashave occurred numerous times invertebrate evolution to produceregionalization of the mammalianspine. Rib-bearing, thoracic-like ver-tebrae represent the developmentaland evolutionary “ground state”(default condition) for vertebral pat-terning, suggested by both the fossilrecord37 and experimental data onHox mutant mice.38 Thus, vertebralregionalization in mammals islargely associated with the presenceor suppression of lateral costal struc-tures such as ribs and sacral alae.Accordingly, we adhere to a costal
definition of vertebral boundaries,wherein cervical and lumbar verte-brae lack moveable ribs, thoracicvertebrae possess moveable ribs, andsacral alae contribute to sacralforamina.1 We will discuss other ver-tebral morphologies, such as the ori-entation of zygapophyses, usingdifferent terminology.39
Most primates have presacral ver-tebral formulae of 7C:13T:6L (where“C” is cervical, “T” is thoracic, and“L” is lumbar) or 7C:12T:7L, coupledwith a sacrum composed of threefused vertebrae.4,40 Hominoids areunusual among primates (and othermammals) in having reduced num-bers of lumbar vertebrae by an over-all reduction of the thoracolumbarregion.20,40–43 Numerical variationexists within as well as between spe-cies (Table 1): hylobatids typicallyhave 17 or 18 thoracolumbar verte-brae; western gorillas, humans, andchimpanzees and bonobos typicallyhave 17; and orangutans and easterngorillas typically have just 16.Accordingly, hylobatids generallypossess four to five sacral vertebraeand extant hominids have five to six.The mechanisms and evolutionarytiming of these changes in hominoidevolution are debated.20,21,40,44–48
Keith20 and others40–42,48,49 arguethat reduced numbers of lumbar ver-tebrae and increased numbers ofsacral vertebrae occurred via cranialhomeotic shifts at the lumbosacralborder (that is, sacralization, orKeith’s “posterior transmutation”) inan ancestral hominoid. Further, thisprocess occurred in a stepwise evolu-tionary fashion, with lumbar reduc-tion occurring first in the lastcommon ancestor (LCA) of hyloba-tids and hominids, then again in theLCA of hominids, so that a “short-backed” ancestry is likely for homi-nins (Fig. 3).
In contrast to this scenario,McCollum and coworkers45 contendthat increased sacral numbersoccurred via a caudally directedsacro-caudal shift that was associ-ated with tail loss in an ancestralhominoid; lumbar reductionoccurred later via both meristic lossand homeotic change at the lumbo-sacral border (Fig. 4; see also Figure4 in McCollum and coworkers45).
McCollum and colleagues45:102 havefurther challenged the hypothesis ofhomology for a reduced lumbarregion in hominoids by arguing that“lumbar column reduction occurredindependently in chimpanzees, bono-bos, gorillas, and hominins” (Fig. 4).Their hypothesis of homoplasy for areduced lumbar region in hominoidshas since been cited50 as demonstrat-ing that “The ‘long-backed’ lumbo-sacral vertebral formula . . . is theprobable primitive condition forhumans and the great apes”.50:574
An earlier version of the “long-backed” model was put forth by Lat-imer and Ward,51 who argued thatsimilarity in the number of lumbarvertebrae (�6) in Proconsul nyanzaeand early hominins suggests thatAfrican apes, orangutans, and laterhominins, including modernhumans, are convergent with respectto reduced numbers of thoracolum-bar vertebrae. In its entirety, the“long-backed” scenario envisioned byLovejoy and colleagues implies thathomoplasy is more rampant thanwas previously thought, to the extentthat chimpanzees and bonoboswould have reduced lumbar regionsindependently. However, similarityin western gorilla and chimpanzeevertebral formulae is remarkablyhigh (86%, as compared to 78% simi-larity between siamangs and white-handed gibbons).48 Thus, Africanapes either represent the primitivehominine condition or have evolvednearly identical frequencies of verte-bral formulae in parallel despite dif-ferences in body mass, locomotion,and ecology. While these two scenar-ios (“long-” and “short-backed”) havebeen at the center of recent discus-sion concerning hominin vertebralcolumn evolution, they represent justtwo possible evolutionary scenariosfor the reduction of hominoid trunks(see Figs. 3–5). For example, Haeus-ler, Martelli, and Boeni52 argued thatearly hominins retained five lumbarvertebrae from an ancestor with ahylobatid-like number of vertebrae, ascenario that requires the parallelreduction of lumbar numbers inorangutans, gorillas, and panins(chimpanzees and bonobos) (Fig. 5).
Support for the “long-backed” sce-narios, in particular, hinges on fossil
ARTICLE Evolution of the Hominoid Vertebral Column 19
evidence. Numbers of vertebrae inthe stem catarrhine Epipliopithecusvindobonensis53 are consistent withthe primitive anthropoid and catar-rhine condition of 12 or 13 thoracicvertebrae, 6 or 7 lumbar vertebrae,and 3 sacral vertebrae.21,40,54 Extantcercopithecoids probably retainthis formula, with variants ofeither 7:12:7:3 (most common) or7:13:6:3 (less common, but modallycharacterizing the Papio-Lophocebus-Theropithecus clade).48 No Miocenehominoid preserves a complete verte-bral column, but Proconsul heseloni(KNM-RU 2036) and P. nyanzae(KNM-MW 13142) preserve partialcolumns, the former with three lum-bar vertebrae28 and the latter with fivethoracolumbar vertebrae (one lowerthoracic, four lumbar) and the firstsacral vertebra.14 Ward14 argued thatP. nyanzae probably had two addi-tional lumbar vertebrae, totaling six.
Early Miocene Morotopithecusbishopi is known from several lum-
bar vertebrae16,18,55 and Middle Mio-cene Pierolapithecus catalaunicusfrom a partial skeleton (IPS21350)with two lumbar vertebrae17,19; how-ever, their vertebral columns are tooincomplete to reconstruct the totalnumber of lumbar vertebrae. Nacho-lapithecus kerioi is represented by apartial skeleton (KNM-BG 35250),which has the most complete verte-bral column known for a Miocenehominoid, consisting of 19 elementsranging from atlas to the last lumbarvertebra.15 Three other isolated ver-tebral elements are known, repre-senting two lumbar vertebrae and afirst sacral element.15,56 Heavy dis-tortion and missing elements preventdefinitive assessment of the regionalnumbers of vertebrae, but six lumbarvertebrae are preserved, which prob-ably either represent the completelumbar column or one lacking onlyone element (6-7 lumbar verte-brae).10,15 Hispanopithecus laietanusis known from a partial skeleton
(CLI-18800) that preserves variousthoracic and lumbar fragments.57,58
On the basis of preserved morphol-ogy and similarities to extant taxa,Susanna and colleagues58 proposethat the H. laietanus lumbar columnprobably consisted of four or fiveelements. Oreopithecus bambolii isknown from several partial vertebralsegments (IGF 11778, Bac. 72, Bac.170; Bac. 50) that preserve aspects ofthe thoracic, lumbar, and sacral col-umns.59–63 Schultz59 and subsequentworkers have reported five lumbarvertebrae in the IGF 11778 (“1958”)partial skeleton; a partial sacrum(Bac. 50) has six sacral elements.
As with Miocene hominoids, noextinct hominin preserves a completevertebral column. Although the inter-pretation of Ardipithecus ramidus con-tributes to Lovejoy and colleagues’long-backed” model,44–46 the partialskeleton (ARA-VP-6/500) itself pre-serves very few vertebrae and no lum-bar vertebrae.64 The A.L. 288-1 partial
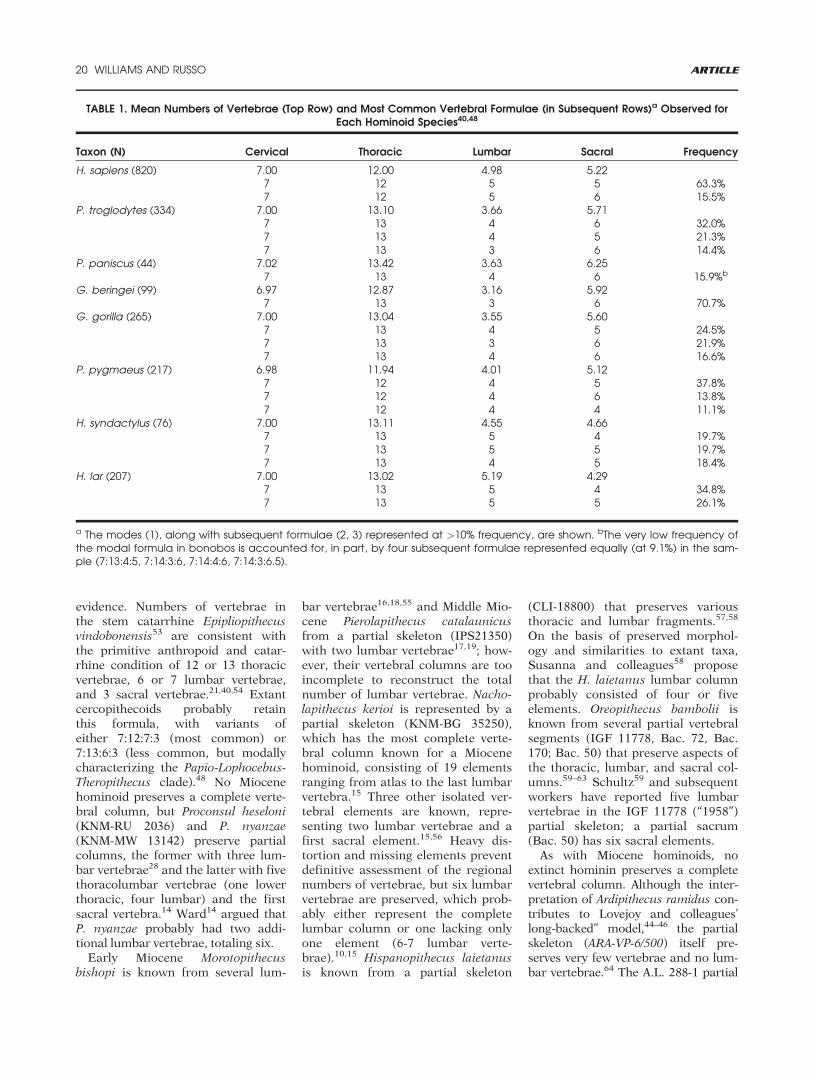
TABLE 1. Mean Numbers of Vertebrae (Top Row) and Most Common Vertebral Formulae (in Subsequent Rows)a Observed forEach Hominoid Species40,48
Taxon (N) Cervical Thoracic Lumbar Sacral Frequency
H. sapiens (820) 7.00 12.00 4.98 5.227 12 5 5 63.3%7 12 5 6 15.5%
P. troglodytes (334) 7.00 13.10 3.66 5.717 13 4 6 32.0%7 13 4 5 21.3%7 13 3 6 14.4%
P. paniscus (44) 7.02 13.42 3.63 6.257 13 4 6 15.9%b
G. beringei (99) 6.97 12.87 3.16 5.927 13 3 6 70.7%
G. gorilla (265) 7.00 13.04 3.55 5.607 13 4 5 24.5%7 13 3 6 21.9%7 13 4 6 16.6%
P. pygmaeus (217) 6.98 11.94 4.01 5.127 12 4 5 37.8%7 12 4 6 13.8%7 12 4 4 11.1%
H. syndactylus (76) 7.00 13.11 4.55 4.667 13 5 4 19.7%7 13 5 5 19.7%7 13 4 5 18.4%
H. lar (207) 7.00 13.02 5.19 4.297 13 5 4 34.8%7 13 5 5 26.1%
a The modes (1), along with subsequent formulae (2, 3) represented at >10% frequency, are shown. bThe very low frequency ofthe modal formula in bonobos is accounted for, in part, by four subsequent formulae represented equally (at 9.1%) in the sam-ple (7:13:4:5, 7:14:3:6, 7:14:4:6, 7:14:3:6.5).
20 WILLIAMS AND RUSSO ARTICLE

Australopithecus afarensis skeletonpreserves seven thoracic vertebrae ofvariable condition and two lumbarvertebrae, along with a nearly com-plete five-element sacrum.47,65 Nineadditional vertebrae are associatedwith A.L. 333 and A.L. 444, includingcervical, thoracic, and lumbar verte-brae.66,67 The partial skeleton KSD-VP-1/1 has five cervical vertebrae, apartial sacrum, and a number ofother vertebral fragments.68 There-
fore, regional numbers are not cur-rently known in A. ramidus and A.afarensis, although full descriptions ofthe infant A. afarensis partial skeleton(DIK-1-1) may clarify thoracic num-ber as full cervical and thoracicregions are preserved, as well as thefirst two lumbar vertebrae,69 once fulldescriptions are published.
Austalopithecus africanus is repre-sented by three partial vertebral col-umns, StW 8/41, StW 431, and Sts
14. StW 8/41 consists of two articu-lated lower thoracic vertebrae (StW41) that articulate with four articu-lated lumbar vertebrae (StW 8).70,71
StW 431 is known from a number ofthoracolumbar vertebrae, includingfive thoracic and five lumbar verte-brae, as well as a partial sacrum52,70;however, many of the neural archesare broken away from the vertebralbodies, making their direct associa-tions difficult to assess and resulting
Figure 3. Schematic of the “short-backed” model from Williams.40,48 Vertebral formulae represented at >10% frequency in extant taxaare shown on the right. Hypothesized ancestral vertebral profiles are listed at nodes, with the proposed modal formula listed first (at thetop of each set). Illustrations show the proposed homeotic (indicated by arrows) changes from ancestral conditions (shown at the pre-vious node) or the proposed modal ancestral configuration (when no change is shown) for (from left to right) Catarrhini, Hominoidea,Hominidae, and H. sapiens (far right, in bold). Note that “African ape-like” vertebral profiles are proposed to be primitive for hominids,hominines, gorillas, panins, and the hominin-panin LCA. This implies that reduced lumbar regions are homologous in orangutans, goril-las, and panins. Early fossil hominins (shown above the human branch), then, evolved the modal human formula, which was retainedin Australopithecus (above the human branch, right). Therefore, hominins evolved five lumbar vertebrae from a short-backed ancestor;modern humans simply retain this modal number. Upward-facing arrows indicate cranially directed homeotic shifts at the lumbo-sacraland thoraco-lumbar borders. [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
ARTICLE Evolution of the Hominoid Vertebral Column 21
in reports of different numbers ofvertebrae.52,70,72 Sts 14 is the mostcomplete South African early homi-nin vertebral column, preserving 15consecutive thoracolumbar vertebraeand a partial sacrum.52,70,73 A verte-bra at the thoracolumbar border (Sts14f) is asymmetrical in that it bearsa fully functional rib facet on oneside (with an associated last rib) anda short, lumbar transverse process-
like structure on the left side.52,70,73,74
The latter process is unusual becauseit is not fully fused to the vertebraand bears a transverse foramen,which are morphologies often associ-ated with short, ankylosed ribs. Rob-inson73:103 made this observation inhis original description: “It seemsprobable that the transverse processwas not firmly attached but articu-lated with the rest of the vertebra in
much the manner of a rib.” In combi-nation with an unknown number ofthoracic elements (since only the lastnine are preserved), the hybridappearance of the vertebra at thethoracolumbar transition has led todisagreement about how many lum-bar vertebrae are present in thisspecimen.
Two partial skeletons of Australopi-thecus sediba are associated with 23
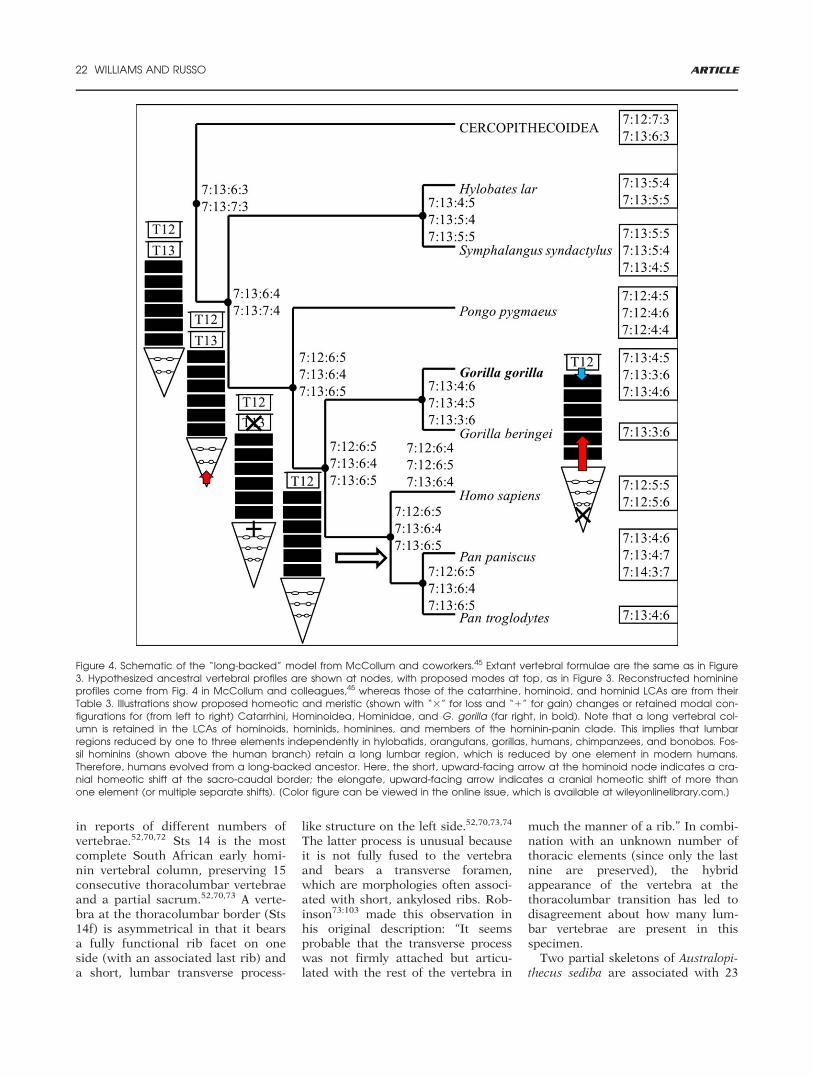
Figure 4. Schematic of the “long-backed” model from McCollum and coworkers.45 Extant vertebral formulae are the same as in Figure3. Hypothesized ancestral vertebral profiles are shown at nodes, with proposed modes at top, as in Figure 3. Reconstructed hominineprofiles come from Fig. 4 in McCollum and colleagues,45 whereas those of the catarrhine, hominoid, and hominid LCAs are from theirTable 3. Illustrations show proposed homeotic and meristic (shown with “3” for loss and “1” for gain) changes or retained modal con-figurations for (from left to right) Catarrhini, Hominoidea, Hominidae, and G. gorilla (far right, in bold). Note that a long vertebral col-umn is retained in the LCAs of hominoids, hominids, hominines, and members of the hominin-panin clade. This implies that lumbarregions reduced by one to three elements independently in hylobatids, orangutans, gorillas, humans, chimpanzees, and bonobos. Fos-sil hominins (shown above the human branch) retain a long lumbar region, which is reduced by one element in modern humans.Therefore, humans evolved from a long-backed ancestor. Here, the short, upward-facing arrow at the hominoid node indicates a cra-nial homeotic shift at the sacro-caudal border; the elongate, upward-facing arrow indicates a cranial homeotic shift of more thanone element (or multiple separate shifts). [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
22 WILLIAMS AND RUSSO ARTICLE

vertebrae: 12 presacral vertebrae witha juvenile male (MH1) and 11 plus afive-element sacrum with an adultfemale (MH2).39 Two lumbar verte-brae are preserved per individual, butno full region in either individual iscomplete. Isolated elements are alsoknown for A. africanus (for example,Sts 73, StW 458, StW 572), and Para-nthropus robustus (SK 3981a, b; Skx53; Skx 41692).70,73,75,76 The “Littlefoot” skeleton (StW 573), which mayrepresent a new species,77 is associ-ated with at least two lumbar verte-
brae, “and there is every indicationthat the rest of the vertebral columnwill follow.”78:525
Vertebral remains of early Homoare rare. Only a partial sacral bodyassociated with KNM-ER 3733(Homo habilis) is known.79 Homoerectus is represented by five suba-dult vertebrae from Dmanisi80 and anearly complete vertebral columnfrom Nariokotome.51,52,81,82 TheDmanisi vertebrae consist of a singlelumbar vertebra (D2672), two cervi-cal vertebrae, and an upper and a
lower thoracic vertebra.80 The“Turkana Boy” (KNM-WT 15000) isalso subadult, but is represented bythe largest number of vertebrae inthe hominin fossil record before theMiddle Pleistocene. These sixteenvertebrae span from the last cervicalthrough the last lumbar, with justtwo mid-thoracic vertebrae (T5 andT8)81 missing in between. Along witha sacrum probably composed of fiveelements,47 the KNM-WT 15000 verte-brae comprise a nearly complete verte-bral column. However, as with Sts 14,
Figure 5. Schematic of an intermediate model from Haeusler, Martelli, and Boeni52 The phylogeny these authors presented, which includeda Pan-Gorilla clade to the exclusion of Homo, has been adjusted to a Pan-Homo clade with the exclusion of Gorilla. Following the authors,only modal formulae are shown as hypothesized ancestral configurations (at nodes); however, vertebral formulae of extant taxa are shownin full as before. The human modal formula is proposed to be primitive for hominids. As such, lumbar regions are reduced from five to four ele-ments independently in orangutans, gorillas, and panins. Fossil hominins (shown above the human branch) experience no change in thisprimitive formula, which modern humans simply retain. Upward-facing red arrows indicate cranial homeotic shifts at the lumbo-sacral andthoraco-lumbar borders; the downward-facing arrow indicates a caudal homeotic shift at the thoraco-lumbar border in many extant taxa,exemplified by G. gorilla (in bold). [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
ARTICLE Evolution of the Hominoid Vertebral Column 23
the number of thoracic and lumbarvertebrae associated with KNM-WT15000 is debated,51,52,81,82 in partbecause of missing elements and, aswith StW 431, uncertainty of associa-tion between broken neural arches andvertebral bodies. Moreover, as with Sts14 and StW 431, there is a dissociationbetween the last rib-bearing (5 tho-racic) vertebra and the transitional (ordiaphragmatic) vertebra, which bearsflat, coronally oriented prezygapophy-ses and curved, sagittally orientedpostzygapophyses.52,81,83 (See Wil-liams and coworkers39 and Haeusler,Martelli, and Boeni52 for justificationof the use of “transitional” over“diaphragmatic”).
In all extant hominoids, the transi-tional vertebra is modally located at
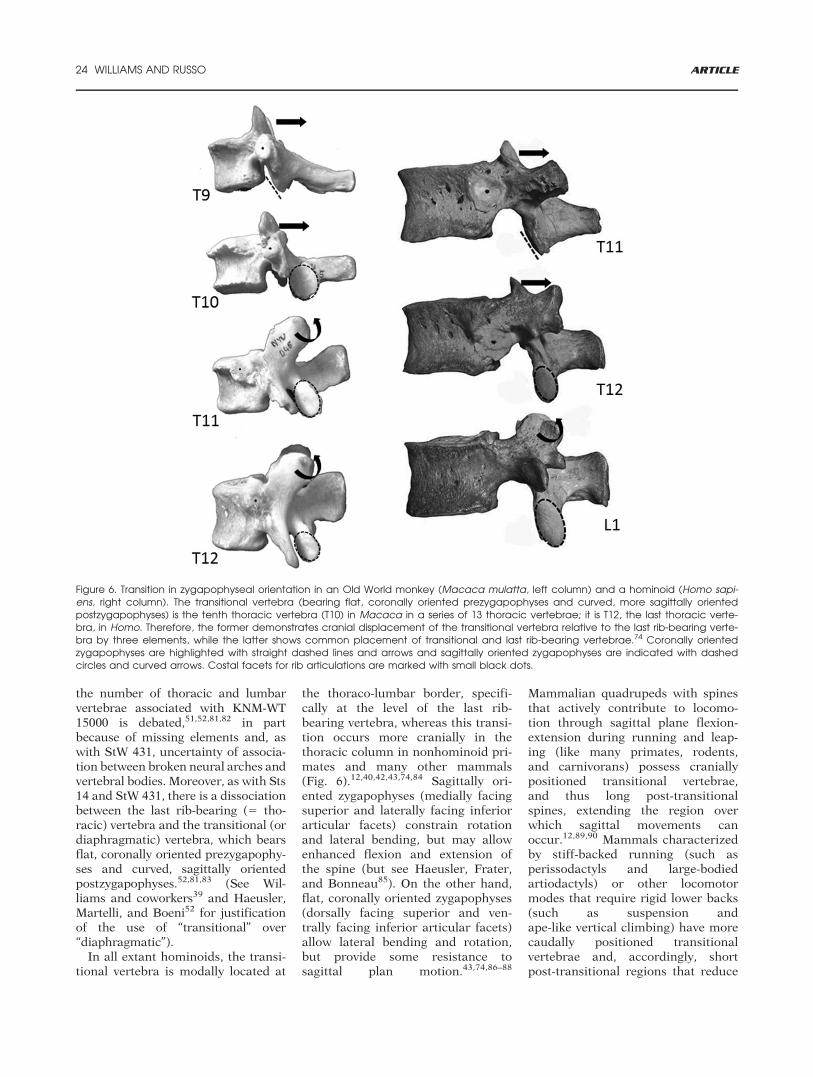
the thoraco-lumbar border, specifi-cally at the level of the last rib-bearing vertebra, whereas this transi-tion occurs more cranially in thethoracic column in nonhominoid pri-mates and many other mammals(Fig. 6).12,40,42,43,74,84 Sagittally ori-ented zygapophyses (medially facingsuperior and laterally facing inferiorarticular facets) constrain rotationand lateral bending, but may allowenhanced flexion and extension ofthe spine (but see Haeusler, Frater,and Bonneau85). On the other hand,flat, coronally oriented zygapophyses(dorsally facing superior and ven-trally facing inferior articular facets)allow lateral bending and rotation,but provide some resistance tosagittal plan motion.43,74,86–88
Mammalian quadrupeds with spinesthat actively contribute to locomo-tion through sagittal plane flexion-extension during running and leap-ing (like many primates, rodents,and carnivorans) possess craniallypositioned transitional vertebrae,and thus long post-transitionalspines, extending the region overwhich sagittal movements canoccur.12,89,90 Mammals characterizedby stiff-backed running (such asperissodactyls and large-bodiedartiodactyls) or other locomotormodes that require rigid lower backs(such as suspension andape-like vertical climbing) have morecaudally positioned transitionalvertebrae and, accordingly, shortpost-transitional regions that reduce
Figure 6. Transition in zygapophyseal orientation in an Old World monkey (Macaca mulatta, left column) and a hominoid (Homo sapi-ens, right column). The transitional vertebra (bearing flat, coronally oriented prezygapophyses and curved, more sagittally orientedpostzygapophyses) is the tenth thoracic vertebra (T10) in Macaca in a series of 13 thoracic vertebrae; it is T12, the last thoracic verte-bra, in Homo. Therefore, the former demonstrates cranial displacement of the transitional vertebra relative to the last rib-bearing verte-bra by three elements, while the latter shows common placement of transitional and last rib-bearing vertebrae.74 Coronally orientedzygapophyses are highlighted with straight dashed lines and arrows and sagittally oriented zygapophyses are indicated with dashedcircles and curved arrows. Costal facets for rib articulations are marked with small black dots.
24 WILLIAMS AND RUSSO ARTICLE

the length of the column overwhich bending moments canoccur.12,16,40,43,74 Running and leap-ing mammals may also benefit fromthe rotational stability that a crani-ally positioned transitional vertebraprovides over a long lower back,whereas rotation and lateral bendingof the spine occur during and mayfacilitate forelimb-dominated loco-motor behaviors.85
P. nyanzae and N. kerioi, the fossilMiocene hominoids that preservelower thoracic vertebrae, are charac-terized by at least one lower thoracicvertebra with sagittally oriented pre-zygapophyses, thus signaling a crani-ally placed transitional vertebrarelative to the last thoracic vertebraby at least one element.10,14,15,40,74
All fossil hominin partial skeletonsthat preserve aspects of the thoraco-lumbar transition (Sts 14, StW 431,MH2, and KNM-WT 15000) are char-acterized by cranial placement of thetransitional vertebra at the level ofthe penultimate rib-bearing vertebra.39,40,52,74,81,83 Conflation of these twomorphologies (presence or absenceof unfused ribs and articular facetorientation) has led to the miscon-ception that early hominins had lon-ger vertebral columns than domodern humans.44–46 In fact, regard-less of whether vertebrae arecounted by lack of rib facets or sagit-tally oriented zygapophyses, no earlyhominin is known from more than17 thoracolumbar vertebrae (12T:5Lversus 11T:6L), the same numberthat characterizes modern humansand is found modally in chimpan-zees and western gorillas.40,48,74
Because it preserves a nearly com-plete vertebral column, KNM-WT15000 nicely demonstrates this point,having 12 thoracic and five lumbarvertebrae and 11 thoracolumbar ver-tebrae with coronally oriented supe-rior articular facets and six withsagittally oriented superior articularfacets.74,81,83,91 This configurationprobably also characterizes earlierhominins, but only the recovery andstudy of additional, more completespecimens, such as DIK1-1, will tell.The functional implications of a lon-ger post-transitional spine is notclear. Some researchers39,74,92 explic-itly argue that it would facilitate the
evolution of lordosis, whereas othersquestion the functional significanceof this difference.81,91 Recently,Haeusler, Frater, and Bonneau85 sug-gested that a transitional vertebra atthe penultimate thoracic level mayhave evolved in early hominins tofacilitate rotational stability forclimbing, but this hypothesis awaitsrigorous testing.
ORTHOGRADY
Adaptations to orthogrady prob-ably evolved in a mosaic fashion andinvolved the reduction of lumbar ver-tebrae discussed in the last section.In general, extant hominoids demon-strate a suite of trunk morphologiesthat distinguish them from mostother primates. These include a dor-soventrally shallower and mediolat-erally broader thorax, thoracicvertebrae invaginated ventrally intothe ribcage, a reduced lumbar col-umn with craniocaudally short verte-brae, and a numerically longersacrum.1,14,27,42,93–97 A broad, shal-low thorax is achieved by increasedrib curvature, transverse broadeningof the sternum, and ventral invagina-tion of the vertebral column into thethorax, which occurs via the dorsalorientation of the transverse proc-esses of thoracic vertebrae coupledwith increased costal angles.1,92,98
The broad thorax and elongatedclavicles position the scapulae dor-sally with craniolaterally oriented gle-noid fossae, allowing increasedshoulder mobility.99 Extant homi-noids stabilize the lower back by twointerrelated mechanisms, reductionin the number and length of lumbarvertebrae and reorganization of epax-ial musculature that deemphasizesspinal flexibility. Here, we outlinesome of these features and discusstheir presence in fossil hominoids.
Extant hominoids have craniocau-dally short but mediolaterally wideand dorsoventrally tall lumbar verte-bral centra.14,16,100,101 They lack“pinched” ventral keels like those incercopithecoids, although they areoccasionally characterized by araised ventral median bulge; theyalso have more columnar, ratherthan laterally hollowed vertebralbodies.16 The lumbar vertebral
bodies of nonhominoid primates aremore craniocaudally elongated thanthose of apes and have “heart-shaped” (cordiform) articular surfa-ces reflecting the presence of anotch-like dorsal concavity and aventral keel (Fig. 7A).14,16 Extanthominoid lumbar vertebral bodiesare transversely wider, giving theirarticular surfaces a kidney-shaped(reniform) or elliptical outline15,16
(Fig. 7D-F) and an increased surfacearea.14,101 Few, short centra contrib-ute to an overall short lumbar regionand promote stability by resistingbending and buckling,14,16,41,101 thusprotecting the integrity of the lowerback during forelimb-dominatedpositional behavior.14,27,41,97,102
Among extinct hominoids, allknown species have relatively long(craniocaudal) vertebral bodiesexcept Oreopithecus bambolii and, toa lesser extent, Pierolapithecus cata-launicus and Hispanopithecus laieta-nus, especially at lower vertebrallevels.17,19,58 Although O. bambolii isthe only species with body propor-tions like those of modern homi-noids,60,62 Morotopithecus bishopi,Nacholapithecus kerioi, and especiallyP. catalaunicus and H. laietanus havemediolaterally wider vertebral bodiesthan do cercopithecoids and Procon-sul nyanzae.14,16–19,58
Other aspects of vertebral bodyshape follow similar “mosaic” pat-terns. In cranial view, P. nyanzae hascercopithecoid-like, heart-shapedarticular surface areas with ventralkeeling and some lateral hollowingof the “spool-shaped” vertebral body(Fig. 7B) N. kerioi lumbar vertebraeare described as having “triangular”articular surface areas in cranialview56 due to their mediolateralwidth posteriorly and the presence ofa ventral keel.10,15 M. bishopi lumbarvertebrae are somewhat heart-shaped in cranial view, but expandmediolaterally at the transverse mid-line and lack a ventral keel, insteadhaving a slight median bulge. In lat-eral view, M. bishopi exhibit ventralconcavity of the body, as do modernhominoids (Fig. 7C).16,18 P. catalau-nicus also lacks a ventral keel and issimilar to M. bishopi in its cranialand caudal articular surface shapes(somewhat heart-shaped cranially,
ARTICLE Evolution of the Hominoid Vertebral Column 25

Figure 7. Middle lumbar vertebrae in comparative perspective: cercopithecoid (A), Proconsul nyanzae (B), Morotopithecus bishopi (C),Hylobates lar (D), Pan troglodytes (E), Homo sapiens (F) in ventral (left column), lateral (middle), and cranial (right column) views. Notethe ventral keel in A and B and lack thereof in C-F in anterior and lateral views, position of the transverse process in cranial view, andorientation of the transverse and spinous processes in all views.
26 WILLIAMS AND RUSSO ARTICLE

more kidney-shaped caudally), but iswider ventrally.17,19 In both P. cata-launicus and H. laietanus, the bodyis less spool-shaped than that of P.nyanzae and M. bishopi (that is, thereis “waisting” of the vertebral body inventral view); the body also is morecolumnar, as in extant hominoids,lacking the lateral hollowing thatcharacterizes P. nyanzae and cercopi-thecoids.17,58 The lower lumbar ver-tebra of H. laietanus is elliptical,resembling the lower lumbar verte-brae of extant hominoids.58 The O.bambolii partial vertebral columnsare severely crushed, but evinceshort, wide vertebral bodies, as inmodern hominoids.59–63 A raisedventral structure that some workershave characterized as a prominentventral keel60 is present in IGF11778, although it may be more sim-ilar to the ventral median bulge pres-ent in M. bishopi and occasionally inmodern hominoids (personal com-munication, T. Harrison).
Long ago, Keith93,103 showed thatthe weight of erector spinae in a gib-bon was less than half that in asimilar-sized langur. Reduction andrearrangement of the sacrospinalisgroup of the erector spinae (namely,iliocostalis and longissimus) andenlargement and reorganization ofthe transversospinalis group, espe-cially multifidus, and quadratus lum-borum,16,42,94,103 help resist flexionand stabilize the trunk duringorthograde postures and forelimb-dominated activities such as suspen-sion.16 Where the lumbar transverseprocesses arise on a vertebra relatesto the size of the erector spinae,which is situated dorsal to the lum-bar transverse processes and lateralto the spinous processes. Specifi-cally, a more ventral placement (onthe centrum) is associated with alarge erector spinae mass, while amore dorsal placement (on the pedi-cle-body junction or pedicle) is asso-ciated with a smaller mass.42,92
Therefore, the dorsal position of thelumbar transverse processes in homi-noids (in hylobatids, on the pedicle-body junction and, in great apes, onthe pedicle) reduces area for theerector spinae relative to the moreventral position in cercopithecoidsand most other mammals. A reposi-
tioning of the transverse processes(from more ventral to more dorsal),along with increasing dorsal and lat-eral inclinations of their apices, isprobably related to a de-emphasis onrunning and leaping.12–14 Dorsalpositioning of the erector spinaemuscles increases their leverage forextension and resisting flexion bymoving them further from the axisof rotation,14,42,43 thereby assistingin stabilizing and controlling thelower back during forelimb-dominated locomotor behavior suchas brachiation other forms of sus-pensory locomotion and African-ape-like vertical climbing.12,14 In addi-tion, reorganization of the erectorspinae and other muscles (for exam-ple, quadratus lumborum) is appa-rent in hominoids and especiallyhominids, in which dorsal lumbartransverse processes are associatedwith widely flared iliac blades of thepelvis and a wide lower rib cage(Fig. 8). Dorsally positioned lumbartransverse processes provide a con-sistent line of action for the erectorspinae from the iliac blade along thevertebral column.14,93,104
As with the vertebral body, fossilhominoids show morphology of thelumbar transverse process that isintermediate between that of cerco-pithecoids and extant hominids. Thetransverse processes of P. nyanzaelumbar vertebrae are positionedmore dorsally on the vertebral bodythan in cercopithecoids and otheranthropoids (except atelids), but donot approximate the body-pedicleborder as they do in gibbons (Fig. 7),and instead are nearly horizontal inthe coronal plane.14,16 Pedicles in P.nyanzae are craniocaudally tall andmediolaterally narrow, as in cercopi-thecoids,16 contrasting with theshort, robust pedicles generallyobserved in extant hominoids.Ward14:319 argued that, despite themuch smaller size of Proconsul hese-loni,, “In all visible morphologies,Proconsul heseloni vertebrae resem-ble those of P. nyanzae,” However,Kelley105 suggested that the lumbartransverse processes of P. heseloniare hylobatid-like in their locationand orientation.
Although the N. kerioi lumbartransverse processes are broken dis-
tally, their bases indicate that theywere craniocaudally long, as in cer-copithecoids and P. nyanzae; how-ever, they arise more dorsally on thebody near the body-pedicle border.15
Pedicles are moderately tall dorso-ventrally, but somewhat robust com-pared to those of cercopithecoidsand Proconsul.15 In an M. bishopiupper lumbar vertebra (UMP 67.06),the base of the left transverse pro-cess originates cranially from a dor-sal position on the vertebral body,whereas in a lower lumbar vertebra(UMP 67.28) it arises cranially fromthe body-pedicle border, as in hylo-batids, and caudally from the pedi-cle, as in all extant hominoids.16,18
The transverse process is dorsallyoriented, as in hominids and somehylobatids, and may orient caudallyto some degree (Fig. 7C).16 The pedi-cles are robust and anchor dorsoven-trally thick transverse processes.16 InP. catalaunicus, the pedicles are alsoshort and robust; the transverseprocesses are dorsoventrallythick,17,19 as are those in M. bishopi,but not P. nyanzae. Similarly, thetransverse processes, although bro-ken distally, originate at the body-pedicle border, as in the former andnot the latter.17 Preserved transverseprocess bases appear to have a slightdorsal orientation, as in M. bish-opi.17,19 In H. laietanus, the pre-served roots of lumbar transverseprocesses arise dorsally from thepedicle and are oriented dorsally;pedicles are robust, as in modernhominoids.58
Together, the spinous and trans-verse processes of lumbar vertebraeact as biomechanical levers for epax-ial musculature, notably the erectorspinae. Caudally oriented spinousprocesses on lumbar vertebrae maysignal emphasis on multifidus to sta-bilize the lower back12,43 or on theprovision of stability through closeapproximation of adjacent vertebrae,which can serve as bony blocks toextension, especially when spinousprocesses are craniocaudally tall.15
Ward14 identified caudal spinousprocess orientation as the onlyderived lumbar vertebral feature Pro-consul shares with extant hominoids.Unlike cercopithecoids, P. nyanzaeand P. heseloni both possess caudally
ARTICLE Evolution of the Hominoid Vertebral Column 27
oriented spinous processes (Fig 7B).In N. kerioi, many lumbar vertebralspinous processes are missing orbroken at their bases. However,some preserved aspects project cau-dally between the notch that sepa-rates the postzygapophyses, as inliving hominoids, suggesting that N.kerioi also had caudally directed spi-nous processes.10,15 This configura-tion between the spinous processand postzygapophyses contrasts withthat observed in cercopithecoidsand, to some degree, in other Mio-cene hominoids (P. nyanzae, M. bish-
opi, and P. catalaunicus), where theconcave notch that separates thepostzygapophyses is unobstructed(Fig. 7). P. catalaunicus has a nearlyhorizontal spinous process with avery slight caudal orientation.19 Twoneural arch fragments from H. laieta-nus preserve the base of the spinousprocess, which suggests a caudalorientation.58
On the basis of evidence from theappendicular skeleton of fossil homi-noids (for example, Sivapithecus,Ardipithecus, Pierolapithecus, and His-panopithecus) some researchers have
suggested that orthogrady,27,44,106 orat least suspensory behavior,17,107,108
evolved independently in multipleextant hominoid lineages. While thisdegree of homoplasy is not antici-pated by studies of extant speciesalone,40,109,110 homoplasy is a com-mon evolutionary phenomenon111–113
that clearly has played a role in homi-noid evolution.10,106,112–115 Yet,whether homoplasy is so prevalentthat “we can no longer rely onhomologies with African apes foraccounts of our origins” 116:74e1
remains to be sufficiently
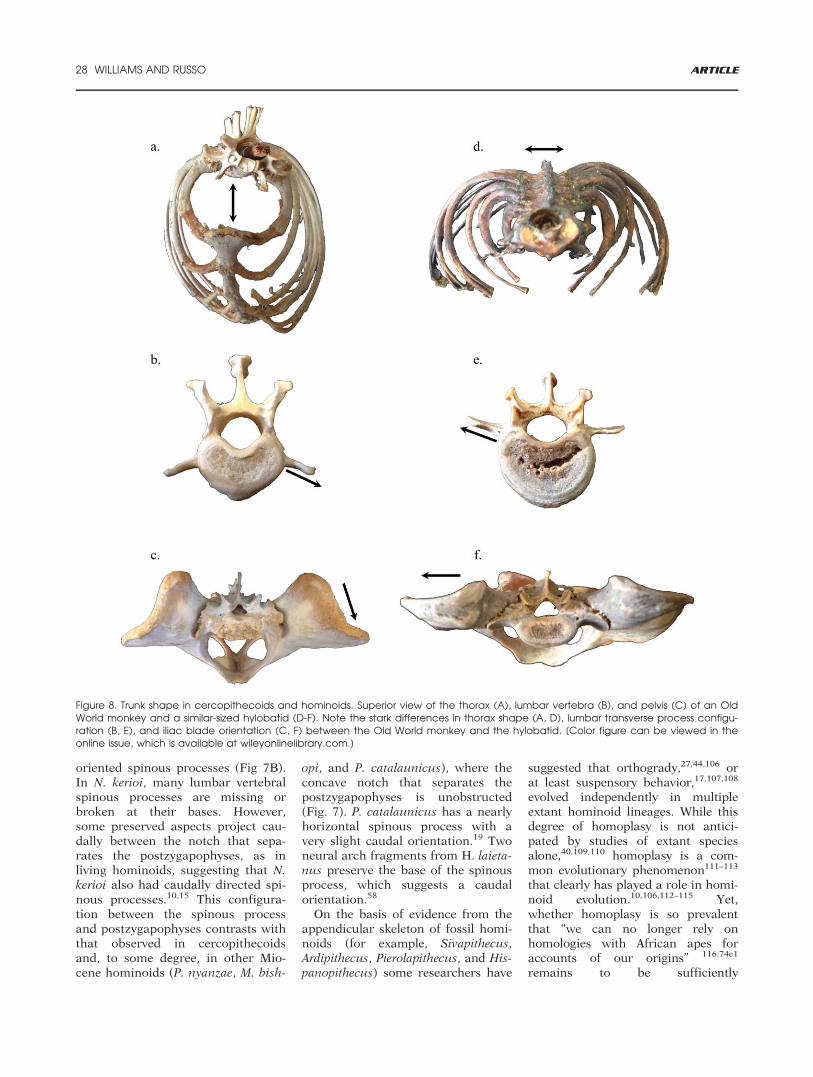
Figure 8. Trunk shape in cercopithecoids and hominoids. Superior view of the thorax (A), lumbar vertebra (B), and pelvis (C) of an OldWorld monkey and a similar-sized hylobatid (D-F). Note the stark differences in thorax shape (A, D), lumbar transverse process configu-ration (B, E), and iliac blade orientation (C, F) between the Old World monkey and the hylobatid. [Color figure can be viewed in theonline issue, which is available at wileyonlinelibrary.com.]
28 WILLIAMS AND RUSSO ARTICLE

demonstrated. The question ofwhether the orthograde-adapted fea-tures characteristic of living apesevolved once or multiple times duringthe course of hominoid evolution notonly presents us with contrastingideas about the tempo and mode ofhominoid evolution, but also presentsdrastically different baupl€ane fromwhich hominins (and therefore biped-alism) evolved. Because the shortlumbar column of extant apes con-tributes to stiffening an upright torsoduring forelimb-dominated activitiessuch as suspension, by reducingbending moments about the interver-tebral discs, a “short-backed” Africanape-like LCA would have limited flexi-bility to lordose a lumbar columnand might have required flexion ofthe hip and knee to reposition thecenter of mass over a narrow base ofsupport.46,92 In contrast, because anumerically long lumbar column per-mits sagittal plane flexibility,12,42,43
proponents of the “long-backed” sce-nario contend that an Old Worldmonkey-like LCA44,46 could arguablyachieve at least partial lordosis dur-ing facultative bipedalism and miti-gate vertical moments of the kneeand hip. This possibility suggests thatearly hominins likely never relied ona bent-hip, bent-knee bipedalgait.46:3297
BIPEDALISM
The distinct role of the homininvertebral column in force transmis-sion and weight-bearing duringbipedalism can be linked with speci-alized traits not seen in other mam-mals.39,51,80,92,104,117–120 Whereasmost primates are characterizedsolely by a primary, kyphotic curveof the spine, hominins have evolved asecondary lordotic curve, achieved bydorsal wedging of lower lumbar ver-tebrae, as well as different degrees ofthickness of the intervertebraldiscs.39,51,92,118,119 In modern humansand fossil hominins, lumbar lordosisis related to sacral slope or angle,pelvic (or sacral) incidence, and pel-vic tilt,39,51,120–124 which place thecenter of mass in line with the sacro-iliac and hip joints. Increased ventralinvagination relative to that in otherhominoids places the vertebral col-
umn in the middle of the rib cage,improving the moment arm of theepaxial musculature for trunk stabil-ity and minimizing bending momentsand shear stresses on vertebrae andintervertebral discs during eccentricloading.51,98 The lumbar transverseprocesses of hominins, which aregenerally more dorsally placed thanthose of other hominids, arise fromthe pedicle-lamina juncture on allbut the last two lumbar vertebrae,where the transverse process basesare broad, arising ventrally from thebody and dorsally from the pedicle-lamina juncture. This is because themovements of iliocostalis and longis-simus are increased when they arelocated more dorsal to their center of
rotation, giving them strong resist-ance to spine flexion.14,16,43 In con-junction with a ventrodorsallyoriented iliac crest, a mediolaterallybroad and an anteriorly tilted humansacrum provide expanded surfacearea and increased leverage for erec-tor spinae musculature to maintainupright trunk posture and stabilizethe lower back.63,125
Women are generally more lor-dotic than men, being characterizedby lower degrees of kyphosis (ventral
wedging) in lower thoracic andupper lumbar vertebrae and higherdegrees of lordosis (achieved in partby strong dorsal wedging of verte-brae) across a greater number oflumbar vertebral levels.118,126 Inaddition to more widely spaced zyga-pophyseal articular facets withgreater surface areas, which rein-force the dorsal components of verte-brae for increased load-bearingduring lordosis (and especiallyhyperlordosis), an increased lordoticcurve of the lower back observed inwomen is hypothesized to be anadaptation to the ventral shift in thecenter of mass that accompaniespregnancy.118 Sexual dimorphism inbony wedging has been documentedin Australopithecus africanus,118 andadult female A. sediba (MH2) is char-acterized by a highly dorsallywedged last lumbar vertebra.39
Although the mid-lumbar vertebraeof the Nariokotome skeleton aredamaged, precluding the ability toobtain body angle measurements,the ultimate and penultimate lumbarvertebrae of this juvenile male arealso strongly dorsally wedged,51,82
highlighting the extensive overlapbetween male and female homininsin this trait (Williams, unpublisheddata). Upright trunk posture andbipedalism also place the dorsalcomponents (zygapophyses, laminae,and pedicles) of the vertebral columnin a weight-bearing role, especiallyduring prolonged extension andhyperextension. The so-called dorsalpillar (zygapophyses and pars inter-articularis) bears significant com-pressive loads; the pedicles transmitbending forces and tension betweenthe vertebral body and the dorsal pil-lar.104,118,127 Accordingly, humanshave expanded lumbar and sacralarticular surface areas (body andzygapophyseal,63 and females arecharacterized by significantly largersurfaces areas than those in malesrelative to overall vertebral size.118
Hominins are distinct from otherhominoids in two other features ofthe lumbar and sacral articular fac-ets. In nonhominin hominoids, thearticular facets become more sagit-talized from the upper to lower lum-bar column and either decrease inmediolateral distance from one
The question of whetherthe orthograde-adaptedfeatures characteristic ofliving apes evolvedonce or multiple timesduring the course ofhominoid evolution notonly presents us withcontrasting ideas aboutthe tempo and mode ofhominoid evolution, butalso presents drasticallydifferent baupl €ane fromwhich hominins (andtherefore bipedalism)evolved.
ARTICLE Evolution of the Hominoid Vertebral Column 29

another or maintain relatively equiv-alent distances. In contrast, in homi-nins the lower lumbar and sacralarticular facets are more obliquelyoriented and set wider apart thanthose of upper and mid-lumbar ver-tebrae.51,63,104,117,128 A progressiveincrease in the mediolateral distancebetween lumbosacral articular facetsresults in a pyramidal configuration,which, together with correspondinglywider laminae and vertebral bodies,facilitates load transmission at thelumbosacral (LS) joint. This configu-ration also prevents spondylolysis, orseparation of the inferior neural archat the pars interarticularis, which iscaused by repeated impingement ofthe postzygapophyses of the superja-cent vertebra and prezygapophysesof the subjacent vertebra into thelamina during hyperextension associ-ated with lordosis.117,126,128 Homi-nins develop laminar fossae, or“imbrication pockets,” mechanicallyinduced nonarticular hollows thatform via bone remodeling when thedistal tips of the postzygapophysesimpinge on the pars interarticularisportion of the laminae during hyper-extension (see Fig. 2D).39,51,92,117
Early hominin lumbar vertebraeand sacra do not evince the full suiteof bipedal adaptations exhibited bymodern humans.71,73,76,104,129,130 Inparticular, lumbosacral body articu-lar surface areas are relativelysmaller in early hominins than inmodern humans,71,73,104,129,130 sothat when the area of the lumbosac-ral joint is used to predict bodymass, it produces much smaller esti-mates than other (e.g., limb) joints,especially for A. africanus.129 Sand-ers71 proposed that early homininvertebral columns were either loadeddifferently than those of modernhumans (for example, more throughthe dorsal pillar39,104,118,127), or wereloaded less frequently, or withreduced loads relative to modernhumans. These factors have beenhypothesized to explain differencesbetween A. afarensis and A. africa-nus130 (but see Haeusler131). Someresearchers argue that A. afarensiswas a fully committed biped with ahuman-like gait132; others maintainthat it retained arboreal abilitiesand/or practiced a form of bipedal-
ism different from that of modernhumans.133 We may expect morestrongly expressed features related toweight-bearing through the vertebralbodies in hominins that are moreterrestrial than arboreal, especiallywith the transition to obligate terres-trial bipedalism. Detecting andexploring the full suite of vertebralscaling differences may help revealhow the locomotor repertoires ofearly hominins differed amongstthemselves and from later hominins,including modern humans, a long-debated topic in paleoanthropology.
SUMMARY AND CONCLUSIONS
Current anatomical evidence indi-cates that Miocene hominoids are char-acterized by mosaic sets of features thatindicate a stepwise accumulation (forexample, tail loss occurred in the con-text of pronogrady, followed by theacquisition of adaptations for orthog-rade postures), and perhaps independ-ent evolution in some lineages, ofpostcranial traits that otherwise appearin conjunction in living apes. Accord-ingly, though it is common practiceamong researchers, the sole use ofextant apes as analogs for extinct spe-cies limits the range of morphologiesand behaviors from which to makeinferences. The problem lies in inferringthe phylogenetic position and interpret-ing the functional implications of fos-sils. In the absence of completepresacral vertebral sequences, research-ers also struggle with determining the
vertebral composition of extinct homi-noids, particularly the hominin lastcommon ancestor with panins. As aresult, researchers have put forth multi-ple, often contrasting, presumed axialmorphologies that evince drasticallydifferent functional contexts in whichbipedalism evolved. Clearly, additionalfossil material is needed to resolve thesecomplicated issues in hominoid evolu-tion. However, there may be more toglean from the currently available fossilevidence with the help of new compara-tive perspectives and analytical meth-ods. New technologies such as 3D laserand computed tomography scanningand geometric morphometric techni-ques, along with advances in our under-standing of the developmental andevolutionary processes that underlie thegenetic and phenotypic integration andmodularity of traits,134 will no doubtcontribute to the resolution of thesephylogenetic and functionalcontroversies.
ACKNOWLEDGMENTS
We thank S. Alm�ecija, I. Susanna, D.Alba, and S. Moy�a-Sol�a for sharing a3D scan of the Pierolapithecus lumbarvertebra. We also thank J. Fleagle andthree anonymous reviewers for helpfulcomments and constructive critiquesthat improved the manuscript.
REFERENCES
1 Schultz AH. 1961. Vertebral column andthorax. Primatologia 4:1–66.
2 Nakatsukasa M, Ward CV, Walker A, et al.2004. Tail loss in Proconsul heseloni. J HumEvol 46:777–784.
3 Ward CV, Walker A, Teaford MF. 1991. Procon-sul did not have a tail. J Hum Evol 21:215–220.
4 Russo GA. 2013. Functional morphology ofmammalian sacra and caudal vertebrae: impli-cations for tail loss and positional behaviors inextinct primates. PhD dissertation, Universityof Texas at Austin.
5 Harrison T. 2002. Late Oligocene to middleMiocene cataarrhines from Afro-Arabia. In:Hartwig WC, editor. The primate fossil record.Cambridge: Cambridge University Press. p 311–338.
6 Stevens NJ, Seiffert ER, O’Connor PM, et al.2013. Palaeontological evidence for an Oligo-cene divergence between Old World monkeysand apes. Nature 497:611–614.
7 Ankel F. 1972. Vertebral morphology of fossiland extant primates. In: Tuttle R, editor. Func-tional and evolutionary biology of primates.Chicago: Adline Press. p 223–240.
8 Russo GA, Shapiro LJ. 2011. Morphologicalcorrelates of tail length in the catarrhine sac-rum. J Hum Evol 61:223–232.
Detecting and exploringthe full suite of vertebralscaling differences maysolve a long-debatedtopic in paleoanthropol-ogy, which is how thegaits of early homininsdiffered amongst them-selves and from laterhominins, includingmodern humans.
30 WILLIAMS AND RUSSO ARTICLE

9 Nakatsukasa M, Tsujikawa H, Shimizu D,et al. 2003. Definitive evidence for tail loss inNacholapithecus, an East African Miocene hom-inoid. J Hum Evol 45:179–186.
10 Nakatsukasa M, Kunimatsu Y. 2009. Nacho-lapithecus and its importance for understandinghominoid evolution. Evol Anthropol 18:103–119.
11 Russo GA. 2015. Postsacral vertebral mor-phology in relation to tail length among prima-tes and other mammals. Anat Rec 298:354–375.
12 Slijper EJ. 1946. Comparative biologic-anatomical investigations on the vertebral col-umn and spinal musculature of mammals. VerhKon Ned Akad Wet (Tweede Sectie) 42:1–128.
13 Ankel-Simons F. 2007. Primate anatomy: anintroduction. Amsterdam: Academic Press.
14 Ward CV. 1993. Torso morphology and loco-motion in Proconsul nyanzae. Am J PhysAnthropol 92:291–328.
15 Nakatsukasa M, Kunimatsu Y, Nakano Y,et al. 2007. Vertebral morphology of Nacholapi-thecus kerioi based on KNM-BG 35250. J HumEvol 52:347–369.
16 Sanders WJ, Bodenbender BE. 1994. Mor-phometric analysis of lumbar vertebra UMP 67-28: implications for spinal function and phylog-eny of the Miocene Moroto hominoid. J HumEvol 26:203–237.
17 Moy�a-Sol�a S, K€ohler M, Alba DM, et al.2004. Pierolapithecus catalaunicus, a new Mid-dle Miocene great ape from Spain. Science 306:1339–1344.
18 Nakatsukasa M. 2008. Comparative study ofMoroto vertebral specimens. J Hum Evol 55:581–588.
19 Susanna I, Alba DM, Alm�ecija S, et al. 2010.The lumbar vertebrae of the Middle Miocenestem great ape Pierolapithecus catalaunicus(Primates: Hominidae). Cidaris 30:311–316.
20 Keith A. 1903. The extent to which the pos-terior segments of the body have been trans-muted and suppressed in the evolution of manand allied primates. J Anat Physiol 37:18–40.
21 Pilbeam D. 2004. The anthropoid postcra-nial axial skeleton: comments on development,variation, and evolution. J Exp Zool B Part B302:241–267.
22 Grand T. 1977. Body weight: its relation totissue composition, segment distribution, andmotor function. I. Interspecific comparisons.Am J Phys Anthropol 47:211–239.
23 Hickman G. 1979. The mammalian tail: areview of functions. Mammal Rev 9:143–157.
24 Dunbar DC, Badam GL. 2000. Locomotionand posture during terminal branch feeding.Int J Primatol 21:649–669.
25 Kelley J. 1997. Paleobiological and phyloge-netic significance of life history in MioceneHominoidea. In: Begun DR, Ward CV, RoseMD, editors. Function, phylogeny, and fossils:Miocene hominoid evoluion and adaptations.New York: Plenum Press. p 173–208.
26 MacLatchy L. 2004. The oldest ape. EvolAnthropol 13:90–103.
27 Cartmill M, Milton K. 1977. The lorisiformwrist joint and the evolution of “brachiating”adaptations in the Hominoidea. Am J PhysAnthropol 47:249–272.
28 Walker AC, Pickford M. 1983. New postcra-nial fossils of Proconsul africanus and Procon-sul nyanzae. In: Ciochon RL, Corruccini RS,editors. New interpretations of ape and humanancestry. New York: Plenum Press. p 325–351.
29 Begun D, Teaford M, Walker A. 1994. Com-parative and functional anatomy of Proconsulphalanges from the Kaswanga primate site,Rusinga Island, Kenya. J Hum Evol 26:89–165.
30 Godfrey LR, Jungers WL, Burney DA, et al.2006. New discoveries of skeletal elements ofHadropithecus stenognathus from Andraho-mana Cave, southeastern Madagascar. J HumEvol 51:395–410.
31 Russo GA, Sayre MK. 2012. Functional mor-phology of proximal caudal vertebrae innonprehensile-tailed primates. Am J PhysAnthropol S56:237–238.
32 Fooden J. 1980. Classification and distribu-tion of living macaques (Macaca Lacepded,1799). In: Lindburg DG, editor. The Macaques:Studies in Ecology, Behavior, and Evolution:Van Nostrand Reinhold Co. p. 1–9.
33 Dequeant ML, Pourqui�e O. 2008. Segmentalpatterning of the vertebrate embryonic axis.Nat Rev Genet 9:370.
34 Gomez C, Ozbudak EM, Wunderlich J, et al.2008. Control of segment number in vertebrateembryos. Nature 454:335–339.
35 Iimura T, Denans N, Pourqui�e O. 2009.Establishment of Hox vertebral identities in theembryonic spine precursors. Curr Top Dev Biol88:201–234.
36 Mallo M, Wellik DM, Deschamps J. 2010.Hox genes and regional patterning of the verte-brate body plan. Dev Biol 344:7–15.
37 Hildebrand M. 1995. Analysis of vertebratestructure, 4th ed. New York: Wiley.
38 Carapuco M, N�ovoa A, Bobola N, et al. 2005.Hox genes specify vertebral types in the preso-mitic mesoderm. Genes Dev 19:2116–2121.
39 Williams SA, Ostrofsky KR, Frater N, et al.2013. The vertebral column of Australopithecussediba. Science 340:1232996.
40 Williams SA. 2011. Evolution of the homi-noid vertebral column. PhD dissertation, Uni-versity of Illinois, Urbana-Champaign.
41 Jungers WL. 1984. Scaling of the hominoidlocomotor skeleton with special reference tolesser apes. In: Preuschoft H, Chivers DJ,Brockelman WY, et al., editors. The lesser apes:evolutionary and behavioral biology. Ebin-burgh: Edinburgh University Press. p 146–169.
42 Benton RS. 1967. Morphological evidencefor adaptations within the exaxial region of theprimates. In: van der Hoeven F, editor. Thebaboon in medical research, Vol II. Austin: Uni-versity of Texas Press. p 201–216.
43 Shapiro L. 1993. Functional morphology ofthe vertebral column in primates. In: Gebo DL,editor. Postcranial adaptation in nonhumanprimates. Dekalb, IL: Northern Illinois Univer-sity Press. p 121–149.
44 Lovejoy CO, Suwa G, Simpson SW, et al.2009. The great divides: Ardipithecus ramidusreveals the postcrania of our last common ances-tors with African apes. Science 326:100–106.
45 McCollum MA, Rosenman BA, Suwa G,et al. 2010. The vertebral formula of the lastcommon ancestor of African apes and humans.J Exp Zool Part B 314B:123–134.
46 Lovejoy CO, McCollum MA. 2010. Spinopel-vic pathways to bipedality: why no hominidsever relied on a bent-hip-bent-knee gait. PhilosTrans R Soc B 365:3289–3299.
47 Russo GA, Williams SA. 2015. “Lucy” (A.L.288-1) had five sacral vertebrae. Am J PhysAnthropol 156:295–303.
48 Williams SA. 2012. Variation in anthropoidvertebral formulae: implications for homologyand homoplasy in hominoid evolution. J ExpZool Part B 318:134–147.
49 Abitbol MM. 1987. Evolution of the sacrumin hominoids. Am J Phys Anthropol 74:65–81.
50 Kimbel WH. 2013. Hesitation on homininhistory. Nature 497:573–574.
51 Latimer B, Ward CV. 1993. The thoracicand lumbar vertebrae. In: Walker A, Leakey R,editors. Cambridge, MA: Harvard UniversityPress. p 266–293.
52 Haeusler M, Martelli SA, Boeni T. 2002.Vertebrae numbers of the early hominid lum-bar spine. J Hum Evol 43:621–643.
53 Zapfe H. 1958. The skeleton of Pliopithecus(Epipliopithecus) vindobonensis Zapfe andH€urzeler. Am J Phys Anthropol 16:441–458.
54 Schultz AH, Straus WL. 1945. The numbersof vertebrae in primates. Proc Am Philos Soc89:601–626.
55 Walker A, Rose MD. 1968. Fossil hominoidvertebra from the Miocene of Uganda. Nature271:980–981.
56 Rose MD, Nakano Y, Ishida H. 1996. Kenya-pithecus postcranial specimens from Nachola,Kenya. Afr Stud Monogr 24:3–56.
57 Moy�a-Sol�a S, K€ohler M. 1996. A Dryopithe-cus skeleton and the origins of great-ape loco-motion. Nature 379:156–159.
58 Susanna I, Alba DM, Alm�ecija S, et al. 2014.The vertebral remains of the late Miocene greatape Hispanopithecus laietanus from Can Lloba-teres 2 (Valles-Penedes Basin, NE Iberian Pen-insula). J Hum Evol 73:15–34.
59 Schultz AH. 1960. Einige Beobachtungenund Maße am Skelett von Oreopithecus im Ver-gleich mit anderen catarrhinen Primaten. ZMorphol Anthropol 50:136–149.
60 Straus WL. 1963. The classification of Oreo-pithecus. In: Washburn SL, editor. Classifica-tion and human evolution. Chicago: Aldine. p146–177.
61 H€urzeler J. 1968. Questions et r�eflexions surl’histoire des Anthropomorphes. Ann Pal�eontol54:195–233.
62 Harrison T. 1987. A reassessment of thephylogenetic relationships of Oreopithecus bab-molii Gervais. J Hum Evol 15:541–583.
63 Russo GA, Shapiro LJ. 2013. Reevaluationof the lumbosacral region of Oreopithecus bam-bolii. J Hum Evol 65:253–265.
64 White TD, Asfaw B, Beyene Y, et al. 2009.Ardipithecus ramidus and the paleobiology ofearly hominids. Science 326:75–86.
65 Johanson DC, Lovejoy CO, Kimbel WH,et al. 1982. Morphology of the Pliocene partialhominid skeleton (A.L. 288-1) from the HadarFormation, Ethiopia. Am J Phys Anthropol 57:403–452.
66 Lovejoy CO, Johanson DC, Coppens Y.1982. Elements of the axial skeleton recoveredfrom the Hadar Formation: 1974-1977 collec-tions. Am J Phys Anthropol 57:631–635.
67 Ward CV, Kimbel WH, Harmon EH, et al.2012. New postcranial fossils of Australopithe-cus afarensis from Hadar, Ethiopia (1990-2007). J Hum Evol 63:1–51.
68 Haile-Selassie Y, Latimer BM, Alene M,et al. 2010. An early Australopithecus afarensispostcranium from Woranso-Mille, Ethiopia.Proc Natl Acad Sci USA 107:12121–12126.
69 Alemseged Z, Spoor F, Kimbel WH, et al.2006. A juvenile early hominin skeleton fromDikika, Ethiopia. Nature 443:296–301.
70 Benade MM. 1990. Thoracic and lumbarvertebrae of African hominids ancient andrecent: morphological and functional aspectswith special reference to upright posture. Mas-ter’s thesis, University of the Witwatersrand.
71 Sanders WJ. 1998. Comparative morpho-metric study of the australopithecine vertebralseries Stw-H8/H41. J Hum Evol 34:249–302.
72 Toussaint M, Macho GA, Tobias PV, et al.2003. The third partial skeleton of a late
ARTICLE Evolution of the Hominoid Vertebral Column 31

Pliocene hominin (Stw 431) from Sterkfontein,South Africa. S Afr J Sci 99:215–223.
73 Robinson J. 1972. Early hominid postureand locomotion. Chicago: University of ChicagoPress.
74 Williams SA. 2012. Placement of the dia-phragmatic vertebra in catarrhines: implica-tions for the evolution of dorsostability inhominoids and bipedalism in hominins. Am JPhys Anthropol 148:111–122.
75 Robinson JT. 1970. New finds at theSwartkrans australopithecine site. Nature 225:1217–1219.
76 McHenry HM. 1991. Petite bodies of the“robust” australopithecines. Am J Phys Anthro-pol 86:445–454.
77 Clarke RJ. 2008. Latest information onSterkfontein’s Australopithecus skeleton and anew look at Australopithecus. S Afr J Sci 104:443–449.
78 Clarke RJ. 2002. Newly revealed informationon the Sterkfontein Member 2 Australopithecusskeleton. S Afr J Sci 98:523–526.
79 Leakey RE, Walker A, Ward CV, et al. 1989.A partial skeleton of a gracile hominid from theUpper Burgi Member of the Koobi Fora Forma-tion, East Lake Turkana, Kenya. Hominidae:proceedings of the 2nd International Congressof Human Paleontology. Milan: Editoriale JacaBook. p 167–173.
80 Meyer MR. 2005. Functional biology of theHomo erectus axial skeleton from Dmanisi,Georgia. PhD dissertation, University ofPennsylvania.
81 Haeusler M, Schiess R, Boeni T. 2011. Newvertebral material points to modern bauplan ofthe Nariokotome Homo erectus skeleton. J HumEvol 61:575–582.
82 Walker A, Leakey R. 1993. The postcranialbones. In: Walker AC, Leakey R, editors. TheNariokotome Homo erectus skeleton. Cam-bridge, MA: Harvard University Press. p 95–160.
83 Williams SA. 2012. Modern or distinct axialbauplan in early hominins? Comments onHaeusler et al. (2011). J Hum Evol 63:552–556.
84 Erikson G. 1963. Brachiation in New Worldmonkeys and in anthropoid apes. Symp ZoolSoc Lond 10:135–163.
85 Haeusler M, Frater N, Bonneau N. 2014.The transition from thoracic to lumbar facetjoint orientation at T11: functional implicationsof a more cranially positioned transitional ver-tebra in early hominids. Am J Phys AnthropolS58:133.
86 Rockwell H, Evans FG, Pheasant HC. 1938.The comparative morphology of the vertebratespinal column: its form as related to function.J Morphol 63:87–117.
87 Russo GA. 2010. Prezygapophyseal articularfacet shape in the catarrhine thoracolumbarvertebral column. Am J Phys Anthropol 142:600–612.
88 Shapiro L. 1995. Functional morphology ofindrid lumbar vertebrae. Am J Phys Anthropol98:323–342.
89 Hildebrand M. 1959. Motions of the runningcheetah and horse. J Mammal 40:481–495.
90 Hurov JR. 1987. Terrestrial locomotion andback anatomy in vervets (Cercopithecusaethiops) and patas monkeys (Erythrocebuspatas). Am J Primatol 13:297–311.
91 Haeusler M, Regula S, Boeni T. 2012. Mod-ern or distinct axial bauplan in early hominins?A reply to Williams (2012). J Hum Evol 63:557–559.
92 Lovejoy CO. 2005. The natural history ofhuman gait and posture Part 1. Spine and pel-vis. Gait Posture 21:95–112.
93 Keith A. 1923. Man’s posture: its evolutionand disorders. Br Med J 1:451–454, 499-502,545-548, 587-590, 624-626, 669–672.
94 Benton RS. 1974. Structural patterns in thePongidae and Cercopithecidae. Yearbk PhysAnthropol 18:65–88.
95 Sarmiento EE. 1995. Cautious climbing andfolivory: a model of hominoid differentiation.Hum Evol 10:289–321.
96 Gebo DL. 1996. Climbing, brachiation, andterrestrial quadrupedalism: historical precur-sors of hominid bipedalism. Am J Phys Anthro-pol 101:55–92.
97 Ward CV. 2014. Postcranial and locomotoradaptations of hominoids. In: Henke W, Tatter-sal I, editors. Handbook of paleoanthropology,Vol 2. Berlin: Springer-Verlag. p 1–22.
98 Jellema LM, Latimer B, Walker A. 1993. Therib cage. In: Walker AC, Leakey R, editors. TheNariokotome Homo erectus skeleton. Cambridge,MA: Harvard University Press. p 294–325.
99 Chan LK. 2007. Scapular position in prima-tes. Folia Primatol 78:19–35.
100 Schultz AH. 1938. The relative lengths ofthe regions of the spinal column in Old Worldprimates. Am J Phys Anthropol 24:1–22.
101 Rose MD. 1975. Functional proportions ofprimate lumbar vertebral bodies. J Hum Evol4:21–38.
102 Waterman HC. 1929. Studies on the evolu-tion of the pelves of man and other primates.Am Mus Nat Hist Bull 58:585–642.
103 Keith A. 1940. Fifty years ago. Am J PhysAnthropol 26:251–267.
104 Shapiro L. 1993. Evaluation of “unique”aspects of human verterbal bodies and pedicleswith a consideration of Australopithecus africa-nus. J Hum Evol 25:433–470.
105 Kelley JJ. 1986. Paleobiology of Miocenehominoids. Ph.D. dissertation, Yale University.
106 Larson SG. 1998. Parallel evolution in thehominoid trunk and forelimb. Evol Anthropol6:87–99.
107 Alm�ecija S, Alba DM, Moy�a-Sol�a S, et al.2007. Orang-like manual adaptations in the fos-sil hominoid Hispanopithecus laietanus: firststeps towards great ape suspensory behaviours.Proc R Soc B 274:2375–2384.
108 Alm�ecija S, Alba DM, Moy�a-Sol�a S. 2009.Pierolapithecus and the functional morphologyof Miocene ape hand phalanges: paleobiologicaland evolutionary implications. J Hum Evol 57:284–297.
109 Young NM. 2003. A reassessment of livinghominoid postcranial variability: implicationsfor ape evolution. J Hum Evol 45:441–464.
110 Pilbeam D, Young N. 2004. Hominoid evo-lution: synthesizing disparate data. ComptesRendus Palevol 3:305–321.
111 Wake DB, Wake MH, Specht CD. 2011.Homoplasy: from detecting pattern to deter-mining process and mechanism of evolution.Science 331:1032–1035.
112 Reno PL. 2014. Genetic and developmentalbasis for parallel evolution and its significance forhominoid evolution. Evol Anthropol 23:188–200.
113 Wood B, Harrison T. 2011. The evolution-ary context of the first hominins. 470:347–352.
114 Alba DM. 2012. Fossil apes from the Valles-Penedes Basin. Evol Anthropol 21:254–269.
115 Alm�ecija S, Tallman M, Alba DM, et al.2013. The femur of Orrorin tugenensis exhibitsmorphometric affinities with both Mioceneapes and later hominins. Nat Comm 4:2888.
116 Lovejoy CO. 2009. Reexamining humanorigins in light of Ardipithecus ramidus. Science326:74.
117 Ward CV, Latimer B. 2005. Human evolu-tion and the development of spondylolysis.SPINE 30:1808–1814.
118 Whitcome KK, Shapiro LJ, Lieberman DE.2007. Fetal load and the evolution of lumbar lor-dosis in bipedal hominins. Nature 450:1075–1078.
119 Been E, G�omez-Olivencia A, Kramer PA.2012. Lumbar lordosis of extinct hominins. AmJ Phys Anthropol 147:64–77.
120 Been E, G�omez-Olivencia A, Kramer PA.2014. Lumbar lordosis in extinct hominins:implications for the pelvic incidence. Am JPhys Anthropol 154:307–314.
121 Tague RG, Lovejoy CO. 1986. The obstetricpelvis of A.L. 288-1 (Lucy). J Hum Evol 15:237–255.
122 Abitbol MM. 1987. Evolution of thelumbosacral angle. Am J Phys Anthropol 72:361–372.
123 Abitbol MM. 1995. Lateral view of Austral-opithecus afarensis: primitive aspects of bipedalpositional behavior in the earliest hominids.J Hum Evol 28:211–229.
124 Tardieu C, Bonneau N, Hecquet J, et al.2013. How is sagittal balance acquired duringbipedal gait acquisition? Comparison of neona-tal and adult pelves in three dimensions. Evolu-tionary implications. J Hum Evol 65:209–222.
125 Gregory WK. 1928. The upright posture ofman: a review of its origin and evolution. ProcAm Philos Soc 67:339–377.
126 Masharawi Y, Dar G, Peleg S, et al. 2010. Amorphological adaptation of the thoracic andlumbar vetebrae to lumbar hyperlordosis inyoung and adult females. Eur Spine J 19:768–773.
127 Pal GP, Routal RV. 1987. Transmission ofweight through the lower thoracic and lumbarregions of the vertebral column in man. J Anat152:93–105.
128 Masharawi Y, Alperovitch-Najenson D,Steinberg N, et al. 2008. Lumbar facet orienta-tion in spondylolysis: a skeletal study. SPINE32:E176–E180.
129 McHenry HM. 1992. Body size and propor-tions in early hominids. Am J Phys Anthropol87:407–431.
130 Dobson SD. 2005. Are the differencesbetween Stw 431 (Australopithecus africanus)and A.L. 288-1 (A. afarensis) significant? J HumEvol 49:143–154.
131 Haeusler M. 2002. New insights into thelocomotion of Australopithecus africanus basedon the pelvis. Evol Anthropol 11:53–57.
132 Lovejoy CO, Meindl RS, Ohman JC, et al.2002. The Maka femur and its bearing on theantiquity of human walking: applying contem-porary concepts of morphogenesis to thehuman fossil record. Am J Phys Anthropol 119:97–133.
133 Stern JT, Susman R. 1983. The locomotoranatomy of Australopithecus afarensis. Am JPhys Anthropol 60:279–317.
134 Rolian C. 2014. Genes, development, andevolvability in primate evolution. Evol Anthro-pol 23:93–104.
VC 2015 Wiley Periodicals, Inc.
32 WILLIAMS AND RUSSO ARTICLE




















