
Evolution of H9N2 influenza viruses from domestic poultry in Mainland China
14
Evolution of H9N2 influenza viruses from domestic poultry in Mainland China Chengjun Li, Kangzhen Yu, Guobin Tian, Dandan Yu, Liling Liu, Bo Jing, Jihui Ping, Hualan Chen * Animal Influenza Laboratory of the Ministry of Agriculture and National Key Laboratory of Veterinary Biotechnology, Harbin Veterinary Research Institute, Chinese Academy of Agricultural Sciences, 427 Maduan Street, Harbin 150001, People’s Republic of China Received 2 April 2005; returned to author for revision 6 May 2005; accepted 13 June 2005 Available online 18 July 2005 Abstract H9N2 viruses have circulated in domestic poultry in Mainland China since 1994, and an inactivated vaccine has been used in chickens to control the disease since 1998. The present study analyzed 27 H9N2 avian influenza viruses that were isolated from chickens and ducks from 1996 to 2002. Infection studies indicated that most of the viruses replicate efficiently but none of them is lethal for SPF chickens. However, these viruses exhibit different phenotypes of replication in a mouse model. Five viruses, including 4 early isolates and one 2000 isolate, are not able to replicate in mice; 14 viruses replicate to moderate titers in mouse lungs and cause less than 5% weight loss, while other 8 viruses could replicate to high titers in the lungs and 7 of them induce 10 – 20% weight loss of the mice on day 5 after inoculation. Most of the viruses isolated after 1996 are antigenically different from the vaccine strain that is currently used in China. Three viruses isolated in central China in 1998 are resistant to adamantanes. Phylogenetic analysis revealed that all of the viruses originated from CK/BJ/1/94-like virus and formed multiple genotypes through complicated reassortment with QA/HK/G1/97-, CK/HK/G9/97-, CK/SH/F/98-, and TY/WI/66-like viruses. This study is a description of the previously uncharacterized H9N2 avian influenza viruses recently circulating in chickens and ducks in Mainland China. Our findings suggest that urgent attention should be paid to the control of H9N2 influenza viruses in animals and to the human’s influenza pandemic preparedness. D 2005 Elsevier Inc. All rights reserved. Keywords: Avian influenza viruses; H9N2; Evolutionary analysis Introduction Wild aquatic birds are the natural hosts of influenza A viruses (Hinshaw et al., 1980; Sharp et al., 1993; Webster et al., 1992). Currently, 16 HA and 9 NA subtype viruses have been identified from avian species (Baker et al., 1987; Fouchier et al., 2004), but only a few of them transmitted and are widespread in the domestic poultry and caused mild respiratory or severe systemic disease (Campitelli et al., 2002; Guo et al., 2000; Hall, 2004; Webster et al., 2002). In the last century, four human influenza pandemics occurred and resulted in significant mortality and morbidity; genetic evidences indicate that these pandemic strains were partially or entirely derived from avian influenza viruses (Kawaoka et al., 1989; Taubenberger et al., 1997; Webster et al., 1992). In 2003 and 2004, H5N1 highly pathogenic influenza viruses spread across Southeast Asia countries and caused disastrous avian influenza outbreaks, and were also trans- mitted into humans, resulting in 32 of 42 infected cases fatal (Hien et al., 2004). Therefore, the existing avian influenza viruses in the poultry also bear big public health threats. H9N2 subtype avian influenza viruses (AIV) have widely circulated in the world since its first detection from turkeys in Wisconsin in 1966 (Homme and Easterday, 1970). In North America, H9N2 viruses were mainly found in shore birds and wild ducks (Kawaoka et al., 1988; Shaw et al., 2002) and no H9-associated disease has been reported in chickens (Perez et 0042-6822/$ - see front matter D 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.virol.2005.06.025 * Corresponding author. Fax: +86 451 82733132. E-mail address: [email protected] (H. Chen). Virology 340 (2005) 70 – 83 www.elsevier.com/locate/yviro
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Evolution of H9N2 influenza viruses from domestic poultry in Mainland China

www.elsevier.com/locate/yviro
Virology 340 (2
Evolution of H9N2 influenza viruses from domestic
poultry in Mainland China
Chengjun Li, Kangzhen Yu, Guobin Tian, Dandan Yu, Liling Liu, Bo Jing,
Jihui Ping, Hualan Chen*
Animal Influenza Laboratory of the Ministry of Agriculture and National Key Laboratory of Veterinary Biotechnology,
Harbin Veterinary Research Institute, Chinese Academy of Agricultural Sciences,
427 Maduan Street, Harbin 150001, People’s Republic of China
Received 2 April 2005; returned to author for revision 6 May 2005; accepted 13 June 2005
Available online 18 July 2005
Abstract
H9N2 viruses have circulated in domestic poultry in Mainland China since 1994, and an inactivated vaccine has been used in chickens to
control the disease since 1998. The present study analyzed 27 H9N2 avian influenza viruses that were isolated from chickens and ducks from
1996 to 2002. Infection studies indicated that most of the viruses replicate efficiently but none of them is lethal for SPF chickens. However,
these viruses exhibit different phenotypes of replication in a mouse model. Five viruses, including 4 early isolates and one 2000 isolate, are
not able to replicate in mice; 14 viruses replicate to moderate titers in mouse lungs and cause less than 5% weight loss, while other 8 viruses
could replicate to high titers in the lungs and 7 of them induce 10–20% weight loss of the mice on day 5 after inoculation. Most of the viruses
isolated after 1996 are antigenically different from the vaccine strain that is currently used in China. Three viruses isolated in central China in
1998 are resistant to adamantanes. Phylogenetic analysis revealed that all of the viruses originated from CK/BJ/1/94-like virus and formed
multiple genotypes through complicated reassortment with QA/HK/G1/97-, CK/HK/G9/97-, CK/SH/F/98-, and TY/WI/66-like viruses. This
study is a description of the previously uncharacterized H9N2 avian influenza viruses recently circulating in chickens and ducks in Mainland
China. Our findings suggest that urgent attention should be paid to the control of H9N2 influenza viruses in animals and to the human’s
influenza pandemic preparedness.
D 2005 Elsevier Inc. All rights reserved.
Keywords: Avian influenza viruses; H9N2; Evolutionary analysis
Introduction
Wild aquatic birds are the natural hosts of influenza A
viruses (Hinshaw et al., 1980; Sharp et al., 1993; Webster et
al., 1992). Currently, 16 HA and 9 NA subtype viruses have
been identified from avian species (Baker et al., 1987;
Fouchier et al., 2004), but only a few of them transmitted
and are widespread in the domestic poultry and caused mild
respiratory or severe systemic disease (Campitelli et al.,
2002; Guo et al., 2000; Hall, 2004; Webster et al., 2002). In
the last century, four human influenza pandemics occurred
and resulted in significant mortality and morbidity; genetic
0042-6822/$ - see front matter D 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.virol.2005.06.025
* Corresponding author. Fax: +86 451 82733132.
E-mail address: [email protected] (H. Chen).
evidences indicate that these pandemic strains were partially
or entirely derived from avian influenza viruses (Kawaoka
et al., 1989; Taubenberger et al., 1997; Webster et al., 1992).
In 2003 and 2004, H5N1 highly pathogenic influenza
viruses spread across Southeast Asia countries and caused
disastrous avian influenza outbreaks, and were also trans-
mitted into humans, resulting in 32 of 42 infected cases fatal
(Hien et al., 2004). Therefore, the existing avian influenza
viruses in the poultry also bear big public health threats.
H9N2 subtype avian influenza viruses (AIV) have widely
circulated in the world since its first detection from turkeys in
Wisconsin in 1966 (Homme and Easterday, 1970). In North
America, H9N2 viruses were mainly found in shore birds and
wild ducks (Kawaoka et al., 1988; Shaw et al., 2002) and no
H9-associated disease has been reported in chickens (Perez et
005) 70 – 83
C. Li et al. / Virology 340 (2005) 70–83 71
al., 2003). In Asia, H9N2 viruses were isolated regularly
from ducks before the 1990s (Shortridge, 1992). However,
infections of H9 subtype AIV in chickens have been reported
in many Asian countries in chickens since later 1990s
(Alexander, 2000; Guo et al., 2000; Naeem et al., 1999).
H9N2 subtype viruses were isolated from pigs in 1998 (Peiris
et al., 1999a) and, subsequently, were isolated from humans
with influenza-like illness in Hong Kong and Mainland
China (Guo et al., 1999; Peiris et al., 1999b). Detailed
antigenic and genetic analysis of the H9N2 influenza viruses
from avian species in Hong Kong live bird markets revealed
the presence of three groups, represented by A/Quail/Hong
Kong/G1/97 (G1), A/Duck/Hong Kong/Y280/97(Y280),
and A/Duck/Hong Kong/Y439/97(Y439) (Guan et al.,
1999). The six internal genes of the G1 group viruses and
the PB2 and PB1 genes of A/chicken/Hong Kong/G9/97(G9)
are closely related to the corresponding gene segments of the
1997 Hong Kong H5N1 influenza viruses (Lin et al., 2000;
Shaw et al., 2002). The two human H9N2 isolates from Hong
Kong belong to the G1 group, while at least 1 human isolate
from China and 2 isolates from pigs in Hong Kong are G9-
like viruses (Guo et al., 1999; Lin et al., 2000).
In Mainland China, H9N2 AIV was firstly isolated from
chickens in Guangdong province in 1994 (Chen et al., 1994;
Li et al., 2003; Liu et al., 2003a, 2003d), and then the
viruses spread to several other southern provinces and
resulted in severe economic losses for poultry industries. An
inactivated vaccine derived from an early isolate, A/chicken/
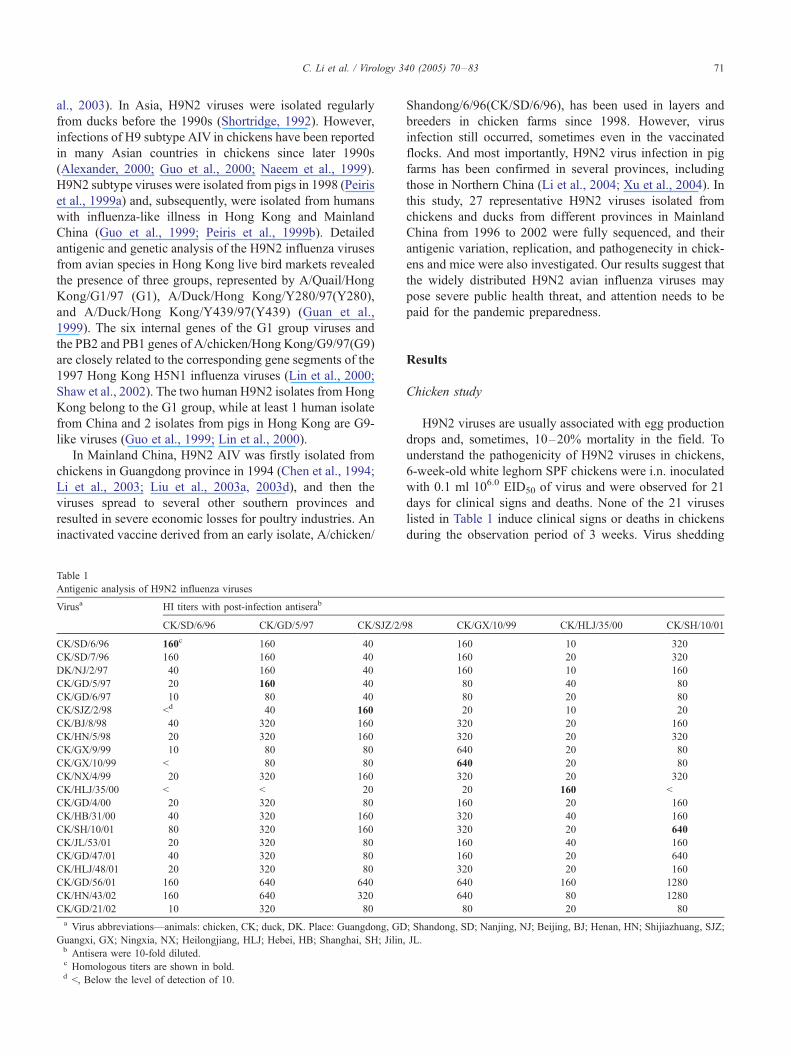
Table 1
Antigenic analysis of H9N2 influenza viruses
Virusa HI titers with post-infection antiserab
CK/SD/6/96 CK/GD/5/97 CK/SJZ/2/9
CK/SD/6/96 160c 160 40
CK/SD/7/96 160 160 40
DK/NJ/2/97 40 160 40
CK/GD/5/97 20 160 40
CK/GD/6/97 10 80 40
CK/SJZ/2/98 <d 40 160
CK/BJ/8/98 40 320 160
CK/HN/5/98 20 320 160
CK/GX/9/99 10 80 80
CK/GX/10/99 < 80 80
CK/NX/4/99 20 320 160
CK/HLJ/35/00 < < 20
CK/GD/4/00 20 320 80
CK/HB/31/00 40 320 160
CK/SH/10/01 80 320 160
CK/JL/53/01 20 320 80
CK/GD/47/01 40 320 80
CK/HLJ/48/01 20 320 80
CK/GD/56/01 160 640 640
CK/HN/43/02 160 640 320
CK/GD/21/02 10 320 80
a Virus abbreviations—animals: chicken, CK; duck, DK. Place: Guangdong, GD
Guangxi, GX; Ningxia, NX; Heilongjiang, HLJ; Hebei, HB; Shanghai, SH; Jilin,b Antisera were 10-fold diluted.c Homologous titers are shown in bold.d <, Below the level of detection of 10.
Shandong/6/96(CK/SD/6/96), has been used in layers and
breeders in chicken farms since 1998. However, virus
infection still occurred, sometimes even in the vaccinated
flocks. And most importantly, H9N2 virus infection in pig
farms has been confirmed in several provinces, including
those in Northern China (Li et al., 2004; Xu et al., 2004). In
this study, 27 representative H9N2 viruses isolated from
chickens and ducks from different provinces in Mainland
China from 1996 to 2002 were fully sequenced, and their
antigenic variation, replication, and pathogenecity in chick-
ens and mice were also investigated. Our results suggest that
the widely distributed H9N2 avian influenza viruses may
pose severe public health threat, and attention needs to be
paid for the pandemic preparedness.
Results
Chicken study
H9N2 viruses are usually associated with egg production
drops and, sometimes, 10–20% mortality in the field. To
understand the pathogenicity of H9N2 viruses in chickens,
6-week-old white leghorn SPF chickens were i.n. inoculated
with 0.1 ml 106.0 EID50 of virus and were observed for 21
days for clinical signs and deaths. None of the 21 viruses
listed in Table 1 induce clinical signs or deaths in chickens
during the observation period of 3 weeks. Virus shedding
8 CK/GX/10/99 CK/HLJ/35/00 CK/SH/10/01
160 10 320
160 20 320
160 10 160
80 40 80
80 20 80
20 10 20
320 20 160
320 20 320
640 20 80
640 20 80
320 20 320
20 160 <
160 20 160
320 40 160
320 20 640
160 40 160
160 20 640
320 20 160
640 160 1280
640 80 1280
80 20 80
; Shandong, SD; Nanjing, NJ; Beijing, BJ; Henan, HN; Shijiazhuang, SJZ;
JL.

C. Li et al. / Virology 340 (2005) 70–8372
was detected from the tracheas of inoculated chickens, with
the titers ranging from 1.2 to 6.5 log10EID50/ml on day 3
after infection. However, only a small proportion of
inoculated chickens shed detectable viruses in the cloacal
swabs, indicating that the H9N2 influenza viruses in
Mainland China are mainly spread via the respiratory route.
All of the chickens were seroconverted on day 21 after
inoculation. These results implied that the disease or death
in the poultry farms where these viruses were isolated may
not be caused by the H9N2 avian influenza viruses alone
and may be a result of co-infection with other pathogens
(Kishida et al., 2004).
Antigenic analysis
Post-infection chicken antisera to 6 selected viruses of
each year were used to analyze the antigenic diversity of 21
Table 2
Replication of H9N2 influenza viruses in micea
Viruses and their stock titers Amino acids of the RBS in
Namee TiterHA gene (H3 number)
(logEID50/ml) 183 190 226
CK/SD/6/96 9.2 N A Q
CK/SD/7/96 9.2 N A Q
DK/NJ/1/97 8.5 N T Q
DK/NJ/2/97 9.5 N T Q
CK/GD/5/97 8.8 N V Q
CK/GD/6/97 9.2 N V Q
CK/SZ/9/97 8.6 N V Q
CK/BJ/8/98 9.4 N V L
CK/SJZ/2/98 8.8 N V Q
CK/HN/5/98 9.6 N V L
CK/GX/9/99 10.0 N T Q
CK/GX/10/99 9.9 N T L
CK/NX/4/99 9.4 N V L
CK/HLJ/35/00 9.5 H E Q
CK/GD/4/00 9.2 N T L
CK/HB/31/00 8.5 N V L
CK/FJ/25/00 9.4 N V L
CK/GD/10/00 9.8 N T L
CK/HN/26/00 8.8 N V Q
CK/JS/1/00 9.5 N T L
CK/SH/10/01 9.0 N A Q
CK/GD/56/01 9.5 N A L
CK/JL/53/01 9.0 N V L
CK/GD/47/01 9.4 N A Q
CK/HLJ/48/01 10.0 N A L
CK/HN/43/02 10.4 N A L
CK/GD/21/02 8.7 N V Q
a 6-week old female BALB/c mice were used in all experiments and were obseb Mice were inoculated i.n. with 106.0 EID50 of each virus in a 50-Al volume. O
infectivity in 10-day-old embryonated chicken eggs. Virus titers were given in units
in the undiluted samples. The infection experiments were repeated one more time
mice. Six mice per group were used; three were killed on day 3 and day 5, respe
described above.c The phenotype groups of the H9N2 avian influenza viruses were divided on th
the text.d Genotypes were determined on the basis of the diversity of the nucleotide seqe Virus abbreviation: Fujian, FJ; Jiangsu, JS.f ND: not done.
H9N2 isolates (Table 1). CK/SD/6/96 was one of the early
isolates and was used as a vaccine strain; however, the HI
test showed that 16 of 21 viruses were antigenic heterolo-
gous with this virus (the HI titers were fourfold lower than
the homologous HI titers). The antisera to CK/SJZ/2/98
could cross-react well with all of the viruses isolated after
1998, except the CK/HLJ/35/00, but not with those viruses
isolated before 1998. The cross-reaction HI titers of the
antisera of CK/HLJ/35/00 with 18 viruses were fourfold
lower than the homologous titer, indicating that CK/HLJ/35/
00 was antigenically quite different with others. CK/SH/10/
01 and CK/GX/10/99 were antigenically heterologous;
however, antisera to CK/SH/10/01 and CK/GX/10/99
reacted equally well with 19 of the 21 viruses. CK/SJZ/2/
98 and CK/HLJ/35/00 could only react well with the
homologous antisera but poorly with other antisera. This
analysis indicated that the H9N2 viruses circulated in China
Virus isolation in mouseb Phenotypec Genotyped
lungs after inoculation group
Day3 Day5
< < I A
< < I A
< < I A
< < I A
6.1 T 0.3 NDf III A
6.6 T 0.1 ND III B
6.0 T 0.7 ND III B
4.5 T 0.3 ND II A
6.4 T 0.1 ND III A
5.2 T 0.6 ND II A
5.3 T 0.4 ND II D
5.7 T 0.5 ND II C
2.8 T 0.7 ND II A
< < I E
5.2 T 0.4 ND II A
6.8 T 0.4 ND III A
5.5 T 1.3 ND II A
7.0 T 0.4 ND III F
6.8 T 0.4 ND III G
4.3 T 0.5 ND II F
5.2 T 1.4 ND II H
< 1.2 T 2.0 II I
4.8 T 1.2 ND II F
5.8 T 1.5 ND II F
< 1.6 T 1.4 II H
< 2.0 T 1.9 II F
7.3 T 0.5 ND III F
rved for 14 days post-infection.
rgans were collected on day 3 p.i., and homogenates were titrated for virus
of log10EID50 per 1 ml T standard deviation (SD). <, virus was not detectedif the virus was not detected in the undiluted lung samples of the inoculated
ctively. The virus titers in the mouse lungs were determined by the method
e basis of their ability to replicate and cause disease in mice, as described in
uences of all the eight gene segments, as described in Fig. 2.

Fig. 1.
C. Li et al. / Virology 340 (2005) 70–83 73

Fig. 1 (continued).
C. Li et al. / Virology 340 (2005) 70–8374
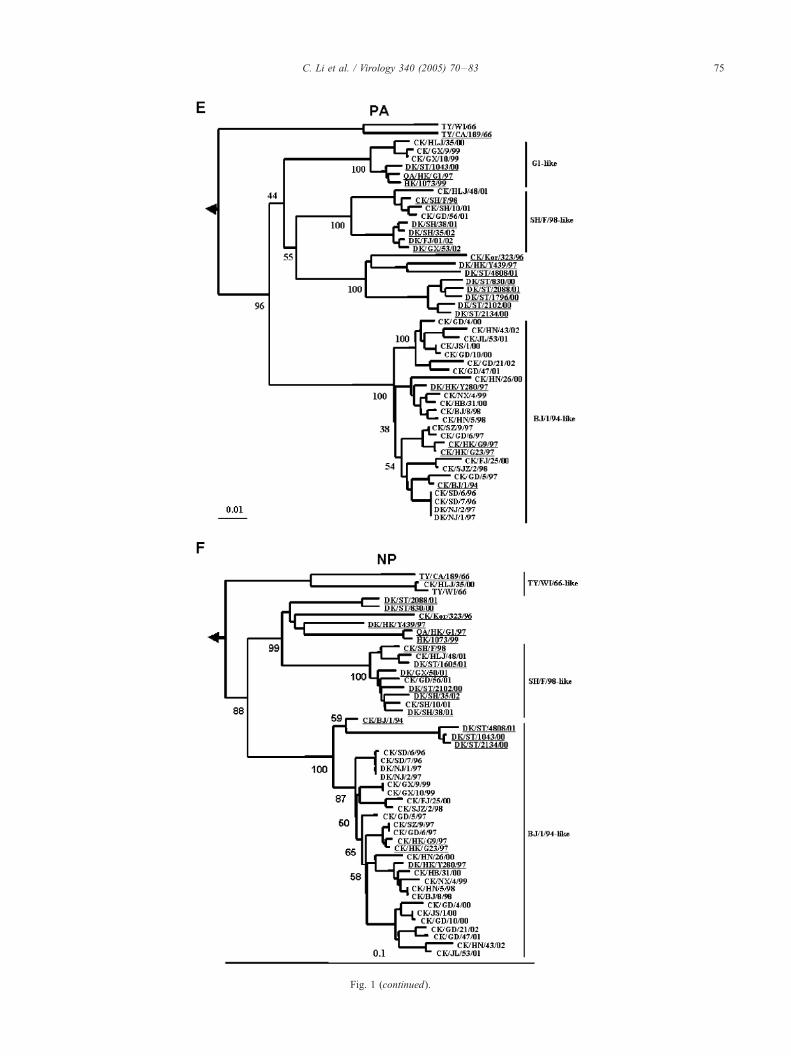
Fig. 1 (continued).
C. Li et al. / Virology 340 (2005) 70–83 75

Fig. 1 (continued).
C. Li et al. / Virology 340 (2005) 70–8376

C. Li et al. / Virology 340 (2005) 70–83 77
are quite different antigenically. Our previous challenge
study indicated that chickens inoculated with the CK/SD/6/
96 inactivated vaccine were poorly protected from shedding
virus in the trachea after challenge with the heterologous
viruses of CK/GX/10/99 or CK/GD/21/02 (authors unpub-
lished data). These results suggested that to maintain
optimal protection by vaccination, the prevailing strains of
influenza virus need to be included in each year’s influenza
vaccine, requiring yearly reevaluation and frequent changes
to the vaccine formulation (Lee et al., 2004).
Pathogenicity of the H9N2 isolates in mice
Avian influenza viruses were regarded to be restricted to
replicate in avian species, but H9N2 avian influenza viruses
have been isolated from pigs and humans previously (Guo
et al., 1999; Peiris et al., 1999a, 1999b; Xu et al., 2004). We
use BALB/c mice as a model to evaluate the ability of
replication in the mammalian hosts of the H9N2 viruses
isolated from avian species in China. Groups of 6-week-old
female BALB/c mice were inoculated i.n. with 106.0 EID50
of the H9N2 viruses, organs from 3 mice were collected on
day 3 or 5 p.i. for virus titration, and 5 mice from each
group were observed for 2 weeks. On the basis of their
ability to replicate and cause disease in mice, these viruses
could be divided into 3 groups (Table 2). Group I consists of
5 viruses, including 4 early isolates, CK/SD/6/96, CK/SD/7/
96, DK/NJ/1/97, and DK/NJ/2/97, and one 2000 isolate,
CK/HLJ/35/00. These 5 viruses could not be recovered from
any of the organs of the inoculated mice on day 3 or day 5
after inoculation, and the inoculated mice stayed healthy and
kept gaining weight during the observation period. Group II
includes 14 viruses. Eleven viruses in this group could
replicate well in the lungs with titers of 2.8–5.8 log10EID50
of inoculated mice on day 3 after inoculation, while 3 other
viruses, CK/GD/56/01, CK/HLJ/48/01, and CK/HN/43/02,
replicate slowly and could only be detected in the mouse
lungs with titers of 1.2–2.0 log10EID50 on day 5 after
inoculation. The weight loss of the mice induced by this
group of viruses is less than 5%. Group III contains 8
viruses that replicate very well in the lungs on day 3 after
inoculation, and the titers reach 6.0–7.3 log10EID50. The
mice showed disease signs as ruffled fur and 10–20%
weight loss. The only exception in group III is the CK/GD/
10/00 virus, which could replicate in the mouse lungs with a
high titer of 7.0 log10EID50, but did not induce any weight
loss. None of the viruses was recovered from spleens and
Fig. 1. Phylogenetic trees for the HA (A), NA (B), PB2 (C), PB1 (D), PA (E),
generated with the PHYLIP program of the CLASTALX software package (versio
(916 bp) of HA; (B) 88–1271 (1184 bp ) of NA; (C) 1036–2289 (1254 bp) of PB
999 (933 bp) of NP; (G) 26–984 (959 bp) of M; and (H) 41–859 (819 bp) of NS.
phylogenetic trees of PB2, NP, NA, M, and NS are rooted to A/Equine/Praque/1/
London/1416/73 (H7N7). Horizontal distances are proportional to the minimum nu
for spacing branches and labels. Internal branching probabilities were determined b
at the key branches in the trees. The sequences obtained from GenBank are under
Hong Kong, HK; Korea, Kor; Hokkaido, Hok; Wisconsin, WI.
brains and no deaths were observed from the inoculated
mice.
Genetic and phylogenetic analysis
To understand the molecular evolution of the H9N2
viruses in Mainland China, the entire genomes of the 27
Chinese isolates and the North American strain, TY/WI/66,
were completely sequenced. The sequences were compared
with representative H9N2 viruses, including CK/BJ/1/94,
G1, G9, Y-280, CK/SH/F/98, and some duck viruses
isolated in Hokkaido and Shantou, reported previously by
other researchers (Guan et al., 1999; Li et al., 2003; Liu et
al., 2003b, 2003c, 2004; Lu et al., 2003).
All of the HA genes contain 1742 nucleotides, except the
DK/NJ/2/97, which has a 21-nucleotide deletion from 1267
to 1287 encoding 7 amino acids. Twenty-six HA genes
shared 94.3% to 97.7% nucleotide homology with the HA
gene of CK/BJ/1/94, whereas the HA gene of CK/HLJ/35/
00 shared a lower than 79.9% homology with the HA genes
of other 26 isolates but 99.3% homology with TY/WI/66,
the prototype virus of North American lineage. Twenty-five
viruses share the same amino acid sequence of PARSSR,Gat the cleavage site (arrow) between HA1 and HA2, while
the sequences are PAVSSR,G and PARLSR,G in the HA of
CK/HLJ/35/00 and CK/GD/56/01, respectively. The
sequences of the cleavage site of these viruses are of the
characters of the low pathogenic avian influenza virus
(Steinhauer, 1999; Taubenberger, 1998). Five potential
glycosylation sites (PGS) in HA1 (11, 123, 200, 280, and
287) and two PGS in HA2 (154 and 213) are conserved in
most of the viruses, while the amino acid changes of N to S
at position 200 of CK/HLJ/35/00, N to D at position 200 of
CK/SD/6/96, CK/SD/7/96, DK/NJ/1/97, and DK/NJ/2/97, S
to G at position 289 of CK/GD/4/00, and S to P at position
215 (HA2) of CK/BJ/8/98 led to the loss of the related PGS.
However, the amino acid S to N change at position 127 led
to the addition of one PGS in CK/GX/9/99, CK/GX/10/99,
and CK/HB/31/00 viruses. These viruses also have different
amino acid substitutions in the receptor binding sites of the
HA gene at positions 183, 190, and 226 (H3 numbers)
(Table 2).
Twenty-six of 27 HA genes of the viruses from this study
belong to the CK/BJ/1/94 sub-lineage and the gradual evo-
lution of these HA genes are observed over the last several
years. The HA gene of CK/HLJ/35/00 is quite different from
others and goes to the North American lineage (Fig. 1A). No
NP (F), M (G), and NS (H) genes of the H9N2 influenza A viruses were
n 1.81) by using the neighbor-joining algorithm. (A) Nucleotides 162–1077
2; (D) 40–1473 (1434 bp) of PB1; (E) 754–2151 (1398 bp) of PA; (F) 67–
The HA phylogenetic tree is rooted to A/Duck/Alberta/60/76 (H12N5). The
56 (H7N7). The phylogenetic trees of PB1 and PA are rooted to A/Equine/
mber of nucleotide differences required to join nodes. The vertical lines are
y bootstrap analysis using 1000 replications and are indicated as percentages
lined. Turkey, TY; Liaoning, LN; Shantou, ST; Tianjin, TJ; California, CA;

Table 3
Effect of 1-adamantylamine on the replication of H9N2 influenza viruses
Viruses Virus yield in the
supernatant with
1-adamantylaminea
Residue at
31 of M2
Phenotype
Absence Presence
CK/SD/6/96 128 0 S Sensitive
CK/GD/6/96 256 0 S Sensitive
CK/SJZ/2/98 64 64 N Resistant
CK/HN/5/98 256 512 N Resistant
CK/BJ/8/98 128 128 N Resistant
CK/GX/10/99 128 0 S Sensitive
CK/HN/43/02 128 0 S Sensitive
a The HA titer of the supernatant from MDCK cells was determined 48 h
after infection with 100 TCID50 infectious dose of the indicated virus
strains in the absence or presence of 2 Ag/ml of 1-adamantylamine in the
cultures.
C. Li et al. / Virology 340 (2005) 70–8378
G1- or Y439-like HA genes are found in this study. The 8 HA
genes of Shantou duck viruses cluster to the clades of the CK/
BJ/1/94 sub-lineage, while the HA genes from the duck
viruses isolated in Hokkaido are closely related with the
reported Y439 virus.
The NA gene of all the H9N2 viruses in this study
belongs to the Eurasian lineage and forms two sister sub-
lineages (Fig. 1B), with CK/BJ/1/94 at the base of one sub-
lineage and with G9 at the base of another one. Analysis of
the deduced amino acid sequence reveals that G9 sub-
lineage viruses have full-length NA gene, while our 16 NA
genes derived from the CK/BJ/1/94 virus have deletions, 15
of them have a 9-nt deletion at positions 206–214 in the
stalk region as reported by Liu et al. (2003b), and the NA of
CK/FJ/25/00 has a deletion from nucleotide 190 to 228,
resulting in a loss of 13 amino acids, whereas CK/BJ/1/94
itself does not. Despite the deletion, the homology of the
viruses between the two sub-lineages is less than 95%,
suggesting that they evolved independently in nature. The
NA genes of the viruses isolated from ducks in Shantou
cluster to the G9 or CK/BJ/1/94 sub-lineages, while the
genes of the viruses from ducks in Hokkaido form a distinct
sub-lineage.
The topology of the phylogenetic trees of PB2, PB1 and
PA are very similar (Figs. 1C, D, E). All of the polymerase
genes of the viruses in the present study belong to the
Eurasian lineage. They form three sub-lineages represented
by the CK/BJ/1/94, G1, and CK/SH/F/98 viruses, respec-
tively. The genes share 94–100% homology within the sub-
lineages, but 84–92% homology between the sub-lineages.
The PB2 gene of the duck viruses isolated from Shantou
cluster to all of the 3 sub-lineages, while some PB1 and PA
genes of these viruses form a separate sub-lineage in the
phylogenetic trees. The NP gene is different from the
polymerase (Fig. 1F); twenty-three of the NP genes obtained
from this study share very high homology and form a large
sub-lineage rooted to CK/BJ/1/94, and NP genes of CK/
HLJ/48/01, CK/SH/10/01, and CK/GD/56/01 share a 97–
99% homology with CK/SH/F/98 and form a distinct sub-
lineage. The NP gene of CK/HLJ/35/00 shares less than
90% homology with other Chinese isolates analyzed in this
study but a 99.6% homology with the NP gene of TY/WI/66
and belongs to the North American lineage.
The M genes of the 27 viruses analyzed in the present
study share 96–100% homology; phylogenetically, they all
belong to the CK/BJ/1/94 sub-lineage (Fig. 1G). Detail
analysis indicated that 3 viruses, CK/BJ/8/98, CK/HN/5/98,
and CK/SJZ/2/98, have an S to N substitution at residue 31
in the M2, which is related to the resistance phenotype of
adamantanes of influenza A viruses (Takeda et al., 2002),
and the resistance to adamantanes of these three viruses was
experimentally confirmed, as shown in Table 3. The NS
genes of 25 viruses in this study are highly related and
belonged to the CK/BJ/1/94 sub-lineage, while the NS
genes of CK/GX/9/99 and CK/GX/10/99 are identical and
belong to the G1 sub-lineage (Fig. 1H).
The genes in the same sub-lineage in Fig. 1 share over
94% homology and are considered as one genotypic group.
On the basis of the genomic diversity, the viruses in the
present study are divided into 9 genotypes, whose evolution
and relationship are shown in Fig. 2. Twelve of the 27
analyzed viruses are classified as genotype A virus and they
are genetically close to the reported CK/BJ/1/94 virus, and
the two genotype B viruses and the two genotype H viruses
are similar to the previously reported G9 or CK/SH/F/98
viruses, respectively (Guan et al., 1999; Lu et al., 2003); the
other 6 genotypes are newly discovered in the present study.
G1-like polymerase genes and NS genes are found in 6
viruses of genotype B, C, D, E, and G. The CK/HLJ/35/00
is likely a reassortant virus that derived the genes from CK/
BJ/1/94-, G1-, G9-, and the North American TY/WI/66-like
viruses. Most of the duck H9N2 viruses isolated in Shantou
described by Li et al. (2003) have different polymerase and
NP genes from the viruses in the present study and form 8
more different genotypes (Li et al., 2003). These results
suggest that complicated reassortments of avian influenza
viruses occurred in the poultry in China. It is worthy to note
that there are no strong links between the genotypes and the
phenotypes of replication in mice of these H9N2 viruses.
Detailed analysis and comparison reveal that 34 amino acids
of the entire genome correlate with the replication pheno-
types in mice (Table 4), although the residue or residues
contributed to the difference of this phenotype remain to be
pinpointed.
Discussion
H9N2 avian influenza viruses always cause disease
outbreaks in the field. Guo et al. reported that the CK/BJ/1/
94 virus was lethal for chickens in field and also under the
experimental condition (Guo et al., 2000). Kida et al. reported
that co-infection with Staphylococcus or Haemophilus
increases the virulence of H9N2 viruses in chickens (Kishida
et al., 2004). Our study indicated that none of the analyzed

Fig 2. The genotypic evolution of the H9N2 viruses isolated from chickens and ducks in Mainland China from 1996 through 2002. The eight gene segments in
each schematic virus particle are (top to bottom) the PB2, PB1, PA, HA, NP, NA, M, and NS genes. Genes of the same sub-lineage are shown in the same color.
The capital letters indicate the genotypes. Table 2 lists the viruses of each genotype.
C. Li et al. / Virology 340 (2005) 70–83 79
viruses isolated from 1996 to 2002 induce disease signs or
deaths in SPF chickens during the observation period,
implying that co-infections with other pathogens might be
the cause for these H9N2 viruses to be lethal in the field.
An oil-emulsion, formalin-inactivated vaccine derived
from an early isolate, CK/SD/6/96, has been used for
chickens to control H9N2 virus infection, since 1998, in
China. The antigenic analysis of 21 representative viruses
performed in this study indicated a large extent of antigenic
diversity among the H9N2 viruses isolated over the years,
and the antisera induced by the vaccine strain could not react
efficiently with most of the viruses isolated after 1996,
although it reacted well to the CK/SH/10/01, CK/GD/56/01,
and CK/HN/43/02 viruses. The previous studies also
demonstrated that the currently used CK/SD/6/96 vaccine
could not completely prevent the replication and shedding of
antigenically different viruses (authors’ unpublished data),
which is similar to the observation in the H5N2 avian
influenza virus in Mexico, where the commercial vaccine
was not able to prevent virus shedding when chickens were
challenged with antigenically different isolates (Lee et al.,
2004). Human’s H1N1 and H3N2 viruses easily subject to
antigenic drift since the vaccines could not completely
prevent the virus replication (Kilbourne et al., 1990, 2002).
About half a billion doses of the inactivated H9N2 vaccines
have been used in chickens every year in China since 1998,
and the antigenic drift of the H9N2 viruses that emerged in
China may also be due to the incomplete protection induced
by the vaccination. The optimal strategy for control of
pandemic influenza is early intervention with a vaccine,
produced, ideally, from the actual pandemic strain or at least
from an antigenically well-matched strain (Lu et al., 2001).
Therefore, it is necessary to evaluate how well the prevailing
isolates match the present vaccine and update the vaccine
formulation accordingly (Lee et al., 2004).
Most of the avian influenza viruses are restricted in the
avian species. Our previous studies indicated that the H5N1
viruses that circulated in ducks in southern China have
gradually acquired the ability to replicate and kill mice (Chen
et al., 2004). Previous studies reported that G9 and G1
viruses and H9N2 viruses isolated in Hong Kong in 2003
replicate systemically and are lethal for mice (Choi et al.,
2004; Guo et al., 2000), while Lu et al. reported that the G9-
and G1-like H9N2 viruses only replicate in the respiratory
system of mice and are not lethal for mice (Lu et al., 2001).
The present study demonstrated that the H9N2 viruses
isolated in Mainland China exhibited diverse replication
phenotype in mice. The 4 early isolates and CK/HLJ/35/00
could not replicate in mice; all others are able to replicate in
the mouse lungs efficiently without any prior adaptation, but
none of the viruses are lethal, although some viruses could
induce a 10–20% weight loss of the inoculated mice.

Table 4
Amino acid residues of the H9N2 viruses that correlate with mouse
replication phenotype
Genes Position of
amino acids
Viruses that
Cannot replicate in micea Replicate in miceb
PB2 105 I T
156 V A
176 M I
285 N H
319 V I
448 D/E N
472 D E/G
PB1 157 I T/A
PA 27 Y D
129 M I
181 D E
388 N G/S
465 V I
672 F L
723 H Y
HA 4 T V/I/L
12 L V/A
15 T V/A
38 A T
218 D N
256 D N
393 M I
537 V I
NP 239 V M
398 K Q
NA 72 I T
370 T S/L
392 K I/T
427 V I
M1 195 A S
M2 14 E G
NS1 25 R Q
225 A T
NS2 14 T M/I/K
a Including the CK/SD/6/96, CK/SD/7/96, DK/NJ/1/97, and DK/NJ/2/97
viruses.b Including 22 viruses that can replicate in mice. CK/HLJ/35/00 contains
an HA gene of North American lineage and was not included in the
comparison.
C. Li et al. / Virology 340 (2005) 70–8380
Most of the viruses contain G1-like genes are able to
replicate to high titers in the mouse lungs without prior
adaptation; however, the CK/HLJ/35/00 virus cannot
replicate in mice, although it contains 3 polymerase genes
of G1-like virus. The CK/HLJ/35/00 virus contains the HA
and NP genes of the North American H9N2 viruses;
however, there is no evidence to indicate that the North
American lineage viruses have been stably established in
Mainland China. How the CK/HLJ/35/00 virus emerged has
not been determined.
Many factors are related to the host range of influenza A
viruses. Subbarao et al. reported that the amino acid at
position 627 of PB2 plays an important role in the host
range of the influenza A viruses (Subbarao et al., 1993). We
recently demonstrated that a single amino acid aspartic acid
to asparagine substitution at 701 of PB2 enables a duck
H5N1 virus to cross the host-barrier to infect and replicate in
mice (Li et al., in press). The receptor specificity of the HA
is important in determining host range, and changes in
preference for SAa2, 3Gal to SAa2, and 6Gal moieties have
been observed to accompany the establishment of avian
viruses (or their HAs) in human and porcine hosts
(Matrosovich et al., 2000). All of the viruses in this study
have conserved glutamine at position of 627 and aspartic
acid at position of 701 in PB2, suggesting that these amino
acids may not be the determinant for the host range
deference of these H9N2 viruses. A total of 34 amino acids
of the entire genome are observed to correlate with the
replication phenotypes of these viruses in mouse, and the
use of plasmid-based reverse genetics will enable an
evaluation of the contribution of the specific amino acid
residues to the host range phenotype of these H9N2 viruses.
Phylogenetic analysis indicated that complicated geno-
types of H9N2 viruses existed in the avian species in China,
and the viruses in the same clades appeared to reflect
temporal parameter rather than geographical parameter,
implying that the viruses were not restricted in the areas
where they appeared, but were spread very quickly to the
neighbor provinces or the provinces far away. Most of the
viruses are genetically closely related to the reported CK/BJ/
1/94 virus, although some of them contain a different NA
gene. A previous report described that the G1-like H9N2
viruses were mainly maintained in the quail and no
reassortant between the G1 and Y280 sub-lineages was
detected despite their co-circulation in the Hong Kong
poultry markets (Guan et al., 2000). G1-like virus was also
not identified in this study, but viruses that contain one or
more internal genes from G1-like viruses were found
primarily in Guangxi and Guangdong provinces, and it
seems that the reassortments happened independently,
implying that the viruses that contain the internal genes of
G1-like viruses may still circulate in those areas. The three
polymerase genes and NP gene of CK/SH/10/01, CK/GD/
56/01, and CK/HLJ/48/01 are closely related with the CK/
SH/F/98 virus and form a distinct sub-lineage in the present
study. These genes are also found in Shantou duck viruses
by Li et al. and in the H5N1 viruses that were isolated from
healthy ducks in Shanghai, Guangxi, and Fujian, including
the DK/SH/35/02, DK/SH/38/01, DK/FJ/01/02, DK/GX/50/
01, and DK/GX/53/02 viruses (Chen et al., 2004). Liu et al.
reported that 4 genotypes of H9N2 viruses were identified
from a live bird market in southern China; one of the
genotype is similar to our genotype A, while viruses in other
3 genotypes contain different NP and polymerase genes
from H3N2, H3N6, or H4N6 viruses (Liu et al., 2003a,
2003d) and those genotypes are different from the ones
detected in the present study. We cannot compare the
genotypes of the viruses described by Li et al. and Liu et al.,
since only partial sequences are available from their reports,
and these sequences are not overlapped; therefore, it is
difficult to confirm the relationship of those viruses. Based
on available information, at least 17 genotypes of H9N2
viruses have been detected in poultry in Mainland China

C. Li et al. / Virology 340 (2005) 70–83 81
and the complicated reassortment occurred not only within
H9N2 avian influenza viruses, but also among the H9N2,
H5N1, H3N2 and H4N6 viruses.
In addition to vaccines, two classes of antiviral agents
have been used to treat influenza—the M2 ion channel
inhibitors and the neuraminidase inhibitors (Kiso et al.,
2004). In comparison to neuraminidase inhibitors, adaman-
tane has a low cost and is frequently used for the influenza
treatment in humans and also for H9N2 avian influenza
treatment in some areas in China. The S to N substitution at
residue 31 in M2 of the influenza virus may occur
spontaneously or as a result of the treatment of adaman-
tanes. The three viruses bearing the S to N substitution at
residue 31 in M2, which confers resistance to adamantanes,
were all isolated in 1998 from central China, supporting the
idea that this mutation may be a result of the application of
adamantanes for avian influenza treatment in some regions.
In summary, the present study systemically analyzed 27
previously uncharacterized H9N2 representative viruses
isolated from poultry in China and revealed that these viruses
posed multiple genotypes and big diversity of the biological
properties. The low pathogenic properties to poultry make
these viruses easy to be widespread and transmitted to a new
host; their gradually acquired ability to infect and replicate in
mammalian and the fact that some genotype viruses are
widespread and established in the pig population in China
rank H9N2 viruses on the top of the list to cause human
influenza pandemics in the future. The antigenic diversity and
the resistance to the adamantanes of these viruses will pose
great difficulties for future human influenza control caused
by H9N2 viruses. Therefore, urgent attention should be paid
to the control of H9N2 influenza viruses in animals and to
human’s influenza pandemic preparedness.
Materials and methods
Viruses and facilities
The H9N2 viruses used in this study were isolated from
the lung samples of sick chickens or ducks sent to the
National Avian Influenza Reference Laboratory for disease
diagnosis. In brief, the lungs were collected under sterile
conditions and stored under �70 -C before inoculation into
embryonated eggs. The lung samples were ground, and a
10% suspension with cold phosphate-buffered saline (PBS),
containing 1000 U/ml of penicillin and 1000 Ag/ml of
streptomycin, was made. Solid debris was pelleted by
centrifugation at 4000�g for 5 min, and the homogenates
were inoculated into the allantoic cavity of 10-day-old
embryonated chicken eggs and incubated for 48–72 h at
37 -C. The isolated viruses were conducted with a three-
round limited dilution purification in 10-day-old embryo-
nated, specific-pathogenic-free (SPF) chicken eggs and were
identified by hemagglutination inhibition (HI) and neura-
minidase inhibition (NI) tests with a panel of antisera
provided by the Office International Des Epizooties (OIE)
Reference Laboratory, Veterinary Laboratory Agency, Sur-
rey, United Kingdom. The virus stocks were grown in the
allantoic cavity of 10-day-old embryonated SPF chicken
eggs at 37 -C for 48 h and aliquoted and stored at �70 -Cfor use in all the experiments described herein. The 50% egg
infection dose (EID50) of each virus stock was determined in
10-day-old embryonated eggs and calculated by the method
of Reed and Muench as described previously (Hoffmann
et al., 2000). All animal experiments were conducted in
HEPA-filtered isolators.
Chicken study
6-week-old white leghorn SPF chickens were intra-
nasally inoculated with 0.1 ml 106.0 EID50 of virus and
were observed for 21 days for clinical signs and deaths.
Tracheal and cloacal swabs were collected on days 3 and 5
post-inoculation (p.i.) for virus titration in eggs as described
(Chen et al., 2003). Sera were collected on day 21 p.i. for
seroconversion confirmation and antigenic analysis.
Antigenic analysis
Antisera to 6 selected H9N2 viruses generated in 6-week-
old white leghorn SPF chickens were included for the
antigenic analysis of 21 H9N2 avian influenza viruses by
hemagglutination inhibition (HI) assays with 0.5% chicken
erythrocytes.
Mouse study
Groups of eight 6-week-old female BALB/c mice (Beijing
Experimental Animal Center, Beijing) were lightly anesthe-
tized with CO2 and inoculated i.n. with 106.0 EID50 of H9N2
influenza virus in a volume of 50 Al. Control mice were
inoculated with PBS. On day 3, three mice were killed and
their lungs, spleens, and brains were collected under sterile
conditions. The organs were ground and homogenized in 1ml
of cold phosphate-buffered saline (PBS), solid debris was
pelleted by centrifugation at 4000�g for 5 min, and the
homogenates were used to determine for virus titration in 10-
day-old embryonated chicken eggs. Virus titers are given in
units of log10EID50 per 1 ml T standard deviation (SD). The
remaining five mice were monitored daily for weight loss and
mortality. For those viruses that could not be recovered from
any organs of inoculated mice on day 3 p.i., a second
experiment was conducted by inoculating groups of 6 mice
i.n. with 106.0 EID50 in a volume of 50 Al. Three mice were
killed on day 3 and day 5, respectively, and the organs were
collected for virus titration.
Genetic and phylogenic analysis
Viral RNA was extracted from allantoic fluid by using
the RNeasy Mini Kit (Qiagen, Valencia, CA) and was

C. Li et al. / Virology 340 (2005) 70–8382
reverse-transcribed using a 12-nucleotide primer, 5VAGCAQAAAGCAGG 3V, with the M-MLV reverse transcriptase
(Life Technologies, Invitrogen) for 1 h at 37 -C. PCR
amplification was performed by using fragment-specific
primers (primer sequences available on request). The PCR
products were purified with the QIAquick PCR purification
kit (Qiagen) and sequenced by using the CEQ DTCS-Quick
Start Kit on a CEQ 8000 DNA sequencer (Beckman
Coulter). Sequence data were compiled with the SEQMAN
program (DNASTAR, Madison, WI), and the phylogenetic
analysis was carried out with the PHYLIP program of the
CLASTALX software package (version 1.81) to implement
a neighbor-joining algorithm.
Sensitivity to adamantanes
Biological susceptibility to 1-adamantylamine was deter-
mined by infecting monolayers of MDCK cells with a 100
TCID50 infectious dose, in OPTI-MEM (Invitrogen Corp.,
Carlsbad, CA) containing 2 Ag/ml trypsin-TPCK (Sigma,
Fluka), in the absence or presence of 2 Ag/ml of 1-
adamantylamine (Sigma, Fluka). Relative virus replication
was determined by measuring the HA titer of the super-
natant after 48-h incubation at 37 -C with 5% CO2.
Nucleotide sequence accession numbers
The nucleotide sequences obtained in this study are
available from GenBank under the access numbers
DQ064354–DQ064569 and DQ067437–DQ067444.
Acknowledgments
This study was supported by Chinese National Key Basic
Research Program (973 Program) Grants G199901190, by
the Chinese National Natural Science Foundation
30440008, and by the Chinese Science and Technology
Development Program (863) 2004AA3071. We thank Dr.
Denis Alexander (Office International Des E’pizooties
Reference Laboratory) for providing the avian influenza
virus reference strains and antisera. The manuscript has
been reviewed by Dr. Paul Dowling from ProofItRight.
References
Alexander, D.J., 2000. A review of avian influenza in different bird species.
Vet. Microbiol. 74, 3–13.
Baker, A.T., Varghese, J.N., Laver, W.G., Air, G.M., Colman, P.M., 1987.
Three-dimensional structure of neuraminidase of subtypes N9 from an
avian influenza virus. Proteins 2, 111–117.
Campitelli, L., Fabiani, C., Puzelli, S., Fioretti, A., Foni, E., Marco, A.D.,
Krauss, S., Webster, R.G., Donatelli, I., 2002. H3N2 influenza viruses
from domestic chickens in Italy: an increasing role for chickens in the
ecology of influenza. J. Gen. Virol. 83, 413–420.
Chen, B.L., Zhang, Z.J., Chen, W.B., 1994. Isolation and preliminary
serological characterization of type A influenza viruses from chickens.
Chin. J. Vet. Med. (Chinese) 22, 3–5.
Chen, H., Matsuoka, Y., Swayne, D., Chen, Q., Cox, N.J., Murphy, B.R.,
Subbarao, K., 2003. Generation and characterization of a cold-adapted
influenza A H9N2 reassortant as a live pandemic influenza virus
vaccine candidate. Vaccine 21, 4430–4436.
Chen, H., Deng, G., Li, Z., Tian, G., Li, Y., Jiao, P., Zhang, L., Liu,
Z., Webster, R.G., Yu, K., 2004. The evolution of H5N1 influenza
viruses in ducks in southern China. Proc. Natl. Acad. Sci. U.S.A.
101, 10452–10457.
Choi, Y.K., Ozaki, H., Webby, R.J., Webster, R.G., Peiris, J.S., Poon,
L., Butt, C., Leung, Y.H.C., Guan, Y., 2004. Continuing evolution
of H9N2 influenza viruses in southeastern China. J. Virol. 78,
8609–8614.
Fouchier, R.A.M., Munster, V., Wallensten, A., Bestebroer, T.M.,
Herfst, S., Smith, D., Rimmelzwaan, G.F., Olsen, B., Osterhaus,
A.D., 2004. Characterization of a novel influenza A virus
hemagglutinin subtype (H16) obtained from black-headed gulls. J.
Virol. 79, 2814–2822.
Guan, Y., Shortridge, K.F., Krauss, S., Webster, R.G., 1999. Molecular
characterization of H9N2 influenza viruses; were they the donors of the
‘‘internal’’ genes of H5N1 viruses in Hong Kong? Proc. Natl. Acad. Sci.
U.S.A. 96, 9363–9367.
Guan, Y., Shortridge, K.F., Krauss, S., Chin, P.S., Dyrting, K.C., Ellis,
T.M., Webster, R.G., Peiris, M., 2000. H9N2 influenza viruses
possessing H5N1-like internal genomes continue to circulate in poultry
in southeastern China. J. Virol. 74, 9372–9380.
Guo, Y.J., Li, J.W., Cheng, I., 1999. Discovery of humans infected
by avian influenza A (H9N2) virus. Chin. J. Exp. Clin. Virol. 15,
105–108.
Guo, Y.J., Krauss, S., Senne, D.A., Mo, I.P., Lo, K.S., Xiong, X.P.,
Norwood, M., Shortridge, K.F., Webster, R.G., Guan, Y., 2000.
Characterization of the pathogenicity of members of the newly
established H9N2 influenza virus lineages in Asia. Virology 267,
279–288.
Hall, C., 2004. Impact of avian influenza on U.S. poultry trade relations—
2002: H5 or H7 low pathogenic avian influenza. Ann. N. Y. Acad. Sci.
1026, 47–53.
Hien, T.T., Jong, M.D., Farrar, J., 2004. Avian influenza—A challenge to
global heath care structures. N. Engl. J. Med. 351, 2363–2365.
Hinshaw, V.G., Webster, R.G., Turner, B., 1980. The perpetuation of
orthomyxoviruses and paramyxoviruses in Canadian waterfowl. Can. J.
Microbiol. 26, 622–629.
Hoffmann, E., Stech, J., Leneva, I., Krauss, S., Scholtissek, C., Chin,
P.S., Peiris, M., Shortridge, K.F., Webster, R.G., 2000. Character-
ization of the influenza A virus gene pool in avian species in
southern China: was H6N1 a derivative or a precursor of H5N1?
J. Virol. 74, 6309–6315.
Homme, P.J., Easterday, B.C., 1970. Avian influenza virus infections: I.
Characteristics of influenza A-turkey-Wisconsin-1966 virus. Avian Dis.
14, 66–74.
Kawaoka, Y., Chambers, T.M., Sladen, W.L., Webster, R.G., 1988. Is the
gene pool of influenza viruses in shorebirds and gulls different from that
in wild ducks? Virology 163, 247–250.
Kawaoka, Y., Krauss, S., Webster, R.G., 1989. Avian-to-human trans-
mission of the PB1 gene of influenza A viruses in the 1957 and 1968
pandemics. J. Virol. 63, 4603–4608.
Kilbourne, E.D., Johansson, B.E., Grajower, B., 1990. Independent and
disparate evolution in nature of influenza A virus hemagglutinin and
neuraminidase glycoproteins. Proc. Natl. Acad. Sci. U.S.A. 87,
786–790.
Kilbourne, E.D., Smith, C., Brett, I., Pokorny, B.A., Johansson, B.,
Cox, N., 2002. The total influenza vaccine failure of 1947 revisited:
major intrasubtypic antigenic change can explain failure of vaccine
in a post-World-War II epidemic. Proc. Natl. Acad. Sci. U.S.A. 99,
10748–10752.
Kishida, N., Sakoda, Y., Eto, M., Sunaga, Y., Kida, H., 2004. Co-

C. Li et al. / Virology 340 (2005) 70–83 83
infection of Staphylococcus aureus or Haemophilus paragallinarum
exacerbates H9N2 influenza A virus infection in chickens. Arch
Virol. 149, 2095–2104.
Kiso, M., Mitamura, K., Tagawa, Y.S., Shiraishi, K., Kawakami, C.,
Kimura, K., Hayden, F.G., Sugaya, N., Kawaoka, Y., 2004. Resistant
influenza A viruses in children treated with oseltamivir: descriptive
study. Lancet 364, 759–765.
Lee, C.W., Senne, D.A., Suarez, D.L., 2004. Effect of vaccine use in the
evolution of Mexican lineage H5N2 avian influenza virus. J. Virol. 78,
8372–8381.
Li, K.S., Xu, K.M., Peiris, J.S.M., Poon, L.L.M., Yu, K.Z., Yuen, K.Y.,
Shortridge, K.F., Webster, R.G., Guan, Y., 2003. Characterization of
H9 subtype influenza viruses from the ducks of southern China: a
candidate for the next influenza pandemic in humans? J. Virol. 77,
6988–6994.
Li, H., Yu, K., Yang, H., Xin, X., Chen, J., Zhao, P., Bi, Y., Chen, H., 2004.
Isolation and characterization of H5N1 and H9N2 influenza viruses
from pigs in China. Chin. J. Prev. Vet. Med. (Chinese) 26, 1–6.
Li, Z., Chen H., Jiao P., Deng G., Tian G., Li Y., Hoffmann E., Webster
R.G., Matsuoka Y., Yu, K., in press. Molecular basis associated with
replication of duck H5N1 influenza viruses in a mammalian mouse
model. J. Virol.
Lin, Y.P., Shaw, M., Gregory, V., Cameron, K., Lim, W., Klimov, A.,
Subbarao, K., Guan, Y.L., Krauss, S., Shortridge, K., Webster, R., Cox,
N., Hay, A., 2000. Avian-to-human transmission of H9N2 subtype
influenza A viruses: relationship between H9N2 and H5N1 human
isolates. Proc. Natl. Acad. Sci. U.S.A. 97, 9654–9658.
Liu, H.Q., Liu, X.F., Cheng, J., Peng, D.X., Jia, L.J., Huang, Y., 2003a.
Phylogenetic analysis of the hemagglutinin genes of twenty-six avian
influenza viruses of subtype H9N2 isolated from chickens in China
during 1996–2001. Avian Dis. 47, 116–127.
Liu, J.H., Okazaki, K., Shi, W.M., Wu, Q.M., Mweene, A.S., Kida, H.,
2003b. Phylogenetic analysis of neuraminidase gene of H9N2 influenza
viruses prevalent in chickens in China during 1995–2002. Virus Genes
27, 197–202.
Liu, J.H., Okazaki, K., Shi, W.M., Kida, H., 2003c. Phylogenetic analysis
of hemagglutinin and neuraminidase genes of H9N2 viruses isolated
from migratory ducks. Virus Genes 27, 291–296.
Liu, M., He, S.Q., Walker, D., Zhou, N.N., Perez, D.R., Mo, B., Li, F.,
Huang, X.T., Webster, R.G., Webby, R.J., 2003d. The influenza
gene pool in a poultry market in south central China. Virology 305,
267–275.
Liu, J.H., Okazaki, K., Mweene, A., Shi, W.M., Wu, Q.M., Su, J.L., Zhang,
G.Z., Bai, G.R., Kida, H., 2004. Genetic conservation of hemaggluinin
gene of H9 influenza virus in chicken population in Mainland China.
Virus Genes 29, 329–334.
Lu, X.H., Renshaw, M., Tumpey, T.M., Kelly, G.D., Primmer, J.H., Katz,
J.M., 2001. Immunity to influenza A H9N2 viruses induced by infection
and vaccination. J. Virol. 75, 4896–4901.
Lu, J.H., Liu, X.F., Shao, W.X., Zhang, P.H., Wei, D.P., 2003. Genetic
characterization of the entire genome of an H9N2 Avian influenza
virus A/Chicken/Shanghai/F/98. Acta Microbiol. Sin. (Chinese) 43,
434–441.
Matrosovich, M., Tuzikov, A., Bovin, N., Gmbaryan, A., Klimov, A.,
Castrucci, M.R., Donatelli, I., Kawaoka, Y., 2000. Early altercations of
the receptor-binding properties of H1, H2, and H3 avian influenza virus
hemagglutinins after their introduction into mammals. J. Virol. 74,
8502–8512.
Naeem, K., Ullah, A., Manvell, R.J., Alexander, D.J., 1999. Avian influenza
A subtype H9N2 in poultry in Pakistan. Vet. Rec. 145, 560.
Peiris, M., Yam, W.C., Chan, K.H., Ghose, P., Shortridge, K.F., 1999a.
Influenza A H9N2: aspects of laboratory diagnosis. J. Clin. Microbiol.
37, 3426–3427.
Peiris, M., Yuen, K.Y., Leung, C.W., Chan, K.H., Ip, P.L., Lai, R.W., Orr,
W.K., Shortridge, K.F., 1999b. Human infection with influenza H9N2.
Lancet 354, 916–917.
Perez, D.R., Lim, W., Seiler, J.P., Guan, Y., Peiris, M., Shortridge,
K.F., Webster, R.G., 2003. Role of quail in the interspecies
transmission of H9 influenza A viruses: molecular changes on HA
that correspond to adaptation from ducks to chickens. J. Virol. 77,
3148–3156.
Sharp, G.B., Kawaoka, Y., Wright, S.M., Turner, B., Hinshaw, V., Webster,
R.G., 1993. Wild ducks are the reservoir for only a limited number of
influenza A subtypes. Epidemiol. Infect. 110, 161–176.
Shaw, M., Cooper, L., Xu, X., Thompson, W., Krauss, S., Guan, Y., Zhou,
N., Klimov, A., Cox, N., Webster, R., Lim, W., Shortridge, K.,
Subbarao, K., 2002. Molecular changes associated with the trans-
mission of avian influenza a H5N1 and H9N2 viruses to humans.
J. Med. Virol. 66, 107–114.
Shortridge, K.F., 1992. Pandemic influenza: a zoonosis? Semin. Respir.
Infect. 7, 11–25.
Steinhauer, D.A., 1999. Role of hemagglutinin cleavage for the pathoge-
nicity of influenza virus. Virology 258, 1–20.
Subbarao, E.K., London, W., Murphy, B.R., 1993. A single amino acid in
the PB2 gene of influenza A virus is a determinant of host range.
J. Virol. 67, 1761–1764.
Takeda, M., Pekosz, A., Shuck, K., Pinto, L.H., Lamb, R.A., 2002.
Influenza A virus M2 ion channel activity is essential for efficient
replication in tissue culture. J. Virol. 76, 1391–1399.
Taubenberger, J.K., 1998. Influenza virus hemagglutinin cleavage into
HA1, HA2: no laughing matter. Proc. Natl. Acad. Sci. U.S.A. 95,
9713–9715.
Taubenberger, J.K., Reid, A.H., Krafft, A.E., Bijwaard, K.E., Fanning, T.G.,
1997. Initial genetic characterization of the 1918 ‘‘Spanish’’ influenza
virus. Science 275, 1793–1796.
Webster, R.G., Bean, W.J., Gorman, O.T., Chambers, T.M., Kawaoka, Y.,
1992. Evolution and ecology of influenza A viruses. Microbiol. Rev. 56,
152–179.
Webster, R.G., Guan, Y., Peiris, M., Walker, D., Krauss, S., Zhou,
N.N., Govorkova, E.A., Ellis, T.M., Dyrting, K.C., Sit, T., Rerez,
D.R., Shortridge, K.F., 2002. Characterization of H5N1 influenza
viruses that continue to circulate in Geese in Southeastern China.
J. Virol. 76, 118–126.
Xu, C.T., Fan, W.X., Rong, W., Zhao, H.K., 2004. Isolation and
identification of swine influenza recombinant A/Swine/Shan-
dong/1/2003(H9N2) virus. Microbes Infect. 6, 919–925.




















