
Evoluci¶on de la atm¶osfera y evoluci¶on temprana de la vida
10
Evoluci¶ on de la atm¶ osfera y evoluci¶ on temprana de la vida L o z a n o { R a m ¶ ³r e z C.,A lva r e z { H e r n ¶a n d e z , S . yD e L a r a { Is a s s i, G. D e p a r t a m e n t o d e H id r o b o lo g ¶³a ,D ivis i¶on d e CB S ,U A M{ I. En su magn¶ ³¯co libro GAIA: A new look at life on Earth (GAIA: una visi¶ on de la vida en la Tierra) James D. Lovelock se~ nal¶ o en 1987 la similitud que existe entre el funcionamiento del planeta y el de un organismo. A pesar de la fuerza de esta comparaci¶ on, la Tierra no es un macroorganismo; sin embargo, es innegable que los elementos planetarios han sido decisivos en la ruta que la evoluci¶ on biol¶ ogica ha seguido. Probablemente m¶ as de lo que pensabamos hasta hace poco. En los sistemas ecol¶ ogicos las diferentes especies in- teractuan unas con otras, si la interacci¶ on de dos o m¶ as especies es muy ¶ ³ntima, tanto que la evolu- ci¶ on de una in°uya sobre la de otra, entonces esta- mos hablando de coevoluci¶ on. Existen diferentes y numerosos ejemplos de coevoluci¶ on, los m¶ as conoci- dos y espectaculares son los de parasitismo. Sin em- bargo ¶ este no es el ¶ unico ¯nal posible de los proce- sos coevolutivos, el mutualismo, donde ambas par- tes obtienen una ventaja es otra posibilidad (Car- ton, 1988). Los ejemplos mencionados se re¯eren a la interacci¶ on de dos entidades vivientes, pero, >podr¶ ³amos hablar de coevoluci¶ on entre elementos \inertes"como la cor- teza terrestre o la atm¶ osfera, con la biosfera?,>acaso su relaci¶ on es tan ¶ ³ntima como para decir que la evo- luci¶ on de una ha afectado de forma determinante a la otra?. Seguramente, la atm¶ osfera primordial de la Tierra fue adquirida durante su paso por su ¶ orbita donde el material circundante fue siendo atrapado gravitacio- nalmente por el planeta. Rasool (1972) cita a Urey, el cual menciona que a una temperatura de 100 a 300 ± K ({173 a 27 ± C) la composici¶ on de la nebulosa pla- netaria debi¶ o ser hidr¶ ogeno (H), helio (He), carbo- no (C), nitr¶ ogeno (N) y ne¶ on (Ne). Elementos co- mo el silicio (Si), magnesio (Mg) y ¯erro (Fe) que son los constituyentes m¶ as importantes de la corte- za terrestre, conformaban menos del 1% de la nebu- losa original; la capa gaseosa pudo ser hasta 400 ve- ces m¶ as densa que la atm¶ osfera actual. La edad de la Tierra se ha calculado por medios isot¶ opicos en aproximadamente 4:5 £ 10 9 a~ nos. Las rocas m¶ as antiguas que se conocen son granitos y gneises localizados en el occidente de Groenlan- dia, fechadas por el m¶ etodo de rubidio{estroncio (Rb ¡ Sr) con una edad de 3:75 £ 10 9 a~ nos (Nagy y Nagy, 1982). Algunos estratos de esta edad pre- sentan zonas de cantos rodados, rocas erosionadas por la acci¶ on del agua. Por otra parte los yacimien- tos m¶ as antiguos que contienen pirita (FeS 2 sulfu- ro de ¯erro), compuesto mineral inestable en el aire h¶ umedo y en presencia de ox¶ ³geno, no se encuentran oxidados, esto apoya fuertemente la idea de un pla- neta con una atm¶ osfera reductora en ese tiempo. Di- cha atm¶ osfera se constitu¶ ³a de hidr¶ ogeno (H 2 ), helio (He), di¶ oxido de carbono (CO 2 ), amoniaco (NH 3 ) y metano (CH 4 ) b¶ asicamente. El hidr¶ ogeno y el he- lio son ligeros y el poderoso viento solar del joven Sol los debi¶ o expulsar al espacio. En una atm¶ osfera sin hidr¶ ogeno, el amoniaco y el metano son inestables y tienden a disociarse, lo cual libera m¶ as hidr¶ ogeno adicional que escapa al espacio. (Rhodes, 1982). A lo largo del tiempo geol¶ ogico la atm¶ osfera gan¶ o ga- ses liberados por actividad volc¶ anica y cristalizaci¶ on de magmas, principalmente agua y di¶ oxido de car- bono con algunas trazas de ¶ acido clorh¶ ³drico (HCl), ¶ acido °uorh¶ ³drico (HF ), ¶ acido sulfh¶ ³drico (H 2 S); ox¶ ³geno por fotodisociaci¶ on del agua, ox¶ ³geno pro- ducido por fotos¶ ³ntesis, He (helio) producido por rompimiento radiactivo de uranio (U ), torio (Th)y arg¶ on (Ar) producido por rompimiento de potasio 40 (K 40 ) as¶ ³ como adiciones debidas al viento solar (Ra- sool, 1972). La Tierra que tiene el tama~ no y la densi- dad correcta para impedir que toda su atm¶ osfera ha- ya escapado al espacio, retuvo los compuestos m¶ as pesados. Durante la formaci¶ on de la Tierra, el agua hab¶ ³a quedado secuestrada en los enlaces de los mi- nerales hidratados, el calor producido por la dife- renciaci¶ on en estratos de la Tierra durante el proce- so de acreci¶ on rompe estos enlaces y el agua se libe- ra por actividad volc¶ anica. Una de las fuentes de ox¶ ³geno en esta atm¶ osfera era la fotodisociaci¶ on del agua en las capas altas de la atm¶ osfera, con el consiguiente escape de hidr¶ ogeno al espacio, este mecanismo era la ¶ unica fuente de ox¶ ³geno en la etapa prebiol¶ ogica, las estimaciones de la cantidad de ox¶ ³geno producido de esta mane- ra son discordantes pero debieron estar entre 10 ¡3 18
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of Evoluci¶on de la atm¶osfera y evoluci¶on temprana de la vida

Evoluci¶on de la atm¶osfera y evoluci¶on temprana de la vida
L o z a n o { R a m ¶ ³r e z C., A lva r e z { H e r n ¶a n d e z , S . y D e L a r a { Is a s s i, G.D e p a r t a m e n t o d e H id r o b o lo g ¶ ³a , D ivis i¶o n d e CB S , U A M{ I.
En su magn¶³¯co libro GAIA: A new look at life onEarth (GAIA: una visi¶on de la vida en la Tierra)James D. Lovelock se~nal¶o en 1987 la similitud queexiste entre el funcionamiento del planeta y el de unorganismo. A pesar de la fuerza de esta comparaci¶on,la Tierra no es un macroorganismo; sin embargo,es innegable que los elementos planetarios han sidodecisivos en la ruta que la evoluci¶on biol¶ogica haseguido. Probablemente m¶as de lo que pensabamoshasta hace poco.
En los sistemas ecol¶ogicos las diferentes especies in-teractuan unas con otras, si la interacci¶on de doso m¶as especies es muy ¶³ntima, tanto que la evolu-ci¶on de una in°uya sobre la de otra, entonces esta-mos hablando de coevoluci¶on. Existen diferentes ynumerosos ejemplos de coevoluci¶on, los m¶as conoci-dos y espectaculares son los de parasitismo. Sin em-bargo ¶este no es el ¶unico ¯nal posible de los proce-sos coevolutivos, el mutualismo, donde ambas par-tes obtienen una ventaja es otra posibilidad (Car-ton, 1988).
Los ejemplos mencionados se re¯eren a la interacci¶onde dos entidades vivientes, pero, >podr¶³amos hablarde coevoluci¶on entre elementos \inertes"como la cor-teza terrestre o la atm¶osfera, con la biosfera?,>acasosu relaci¶on es tan ¶³ntima como para decir que la evo-luci¶on de una ha afectado de forma determinante ala otra?.
Seguramente, la atm¶osfera primordial de la Tierrafue adquirida durante su paso por su ¶orbita donde elmaterial circundante fue siendo atrapado gravitacio-nalmente por el planeta. Rasool (1972) cita a Urey, elcual menciona que a una temperatura de 100 a 300±
K ({173 a 27± C) la composici¶on de la nebulosa pla-netaria debi¶o ser hidr¶ogeno (H), helio (He), carbo-no (C), nitr¶ogeno (N) y ne¶on (Ne). Elementos co-mo el silicio (Si), magnesio (Mg) y ¯erro (Fe) queson los constituyentes m¶as importantes de la corte-za terrestre, conformaban menos del 1% de la nebu-losa original; la capa gaseosa pudo ser hasta 400 ve-ces m¶as densa que la atm¶osfera actual.
La edad de la Tierra se ha calculado por mediosisot¶opicos en aproximadamente 4:5 £ 109 a~nos. Lasrocas m¶as antiguas que se conocen son granitos
y gneises localizados en el occidente de Groenlan-dia, fechadas por el m¶etodo de rubidio{estroncio(Rb ¡ Sr) con una edad de 3:75 £ 109 a~nos (Nagyy Nagy, 1982). Algunos estratos de esta edad pre-sentan zonas de cantos rodados, rocas erosionadaspor la acci¶on del agua. Por otra parte los yacimien-tos m¶as antiguos que contienen pirita (FeS2 sulfu-ro de ¯erro), compuesto mineral inestable en el aireh¶umedo y en presencia de ox¶³geno, no se encuentranoxidados, esto apoya fuertemente la idea de un pla-neta con una atm¶osfera reductora en ese tiempo. Di-cha atm¶osfera se constitu¶³a de hidr¶ogeno (H2), helio(He), di¶oxido de carbono (CO2), amoniaco (NH3)y metano (CH4) b¶asicamente. El hidr¶ogeno y el he-lio son ligeros y el poderoso viento solar del joven Sollos debi¶o expulsar al espacio. En una atm¶osfera sinhidr¶ogeno, el amoniaco y el metano son inestablesy tienden a disociarse, lo cual libera m¶as hidr¶ogenoadicional que escapa al espacio. (Rhodes, 1982).
A lo largo del tiempo geol¶ogico la atm¶osfera gan¶o ga-ses liberados por actividad volc¶anica y cristalizaci¶onde magmas, principalmente agua y di¶oxido de car-bono con algunas trazas de ¶acido clorh¶³drico (HCl),¶acido °uorh¶³drico (HF ), ¶acido sulfh¶³drico (H2S);ox¶³geno por fotodisociaci¶on del agua, ox¶³geno pro-ducido por fotos¶³ntesis, He (helio) producido porrompimiento radiactivo de uranio (U), torio (Th) yarg¶on (Ar) producido por rompimiento de potasio 40(K40) as¶³ como adiciones debidas al viento solar (Ra-sool, 1972). La Tierra que tiene el tama~no y la densi-dad correcta para impedir que toda su atm¶osfera ha-ya escapado al espacio, retuvo los compuestos m¶aspesados. Durante la formaci¶on de la Tierra, el aguahab¶³a quedado secuestrada en los enlaces de los mi-nerales hidratados, el calor producido por la dife-renciaci¶on en estratos de la Tierra durante el proce-so de acreci¶on rompe estos enlaces y el agua se libe-ra por actividad volc¶anica.
Una de las fuentes de ox¶³geno en esta atm¶osfera erala fotodisociaci¶on del agua en las capas altas de laatm¶osfera, con el consiguiente escape de hidr¶ogenoal espacio, este mecanismo era la ¶unica fuente deox¶³geno en la etapa prebiol¶ogica, las estimacionesde la cantidad de ox¶³geno producido de esta mane-ra son discordantes pero debieron estar entre 10¡3
18

Evoluci¶on de la atm¶osfera y. . . Lozano{Ram¶³rez C., Alvarez{Hern¶andez, S. y De Lara{Isassi, G. 19
y 10¡1 de la cantidad actual. Conocer la concentra-ci¶on de ox¶³geno es importante porque una concen-traci¶on de 10¡1 podr¶³a haber oxidado r¶apidamenteal metano y al amoniaco a di¶oxido de carbono ynitr¶ogeno, lo cual podr¶³a indicar que en la Tierraprimitiva la existencia de amoniaco era dif¶³cil o de-bi¶o encontrarse en cantidades muy peque~nas, aun-que es posible que cantidades peque~nas de amonia-co hayan podido ¯ltrar una gran cantidad de radia-cion ultravioleta (radiaci¶on UV) y as¶³ evitar la fo-todisociaci¶on del agua, lo cual podr¶³a haber evita-do la oxidaci¶on de la Tierra por varios millones dea~nos.
Para que aparezca la vida deben cumplirse tres con-diciones fundamentales:
1. Presencia de materia org¶anica, constituyenteprincipal en cuanto a la estructura y funci¶on pa-ra los sistemas biol¶ogicos.
2. Temperatura, la cual debe situarse en un inter-valo que permita la no desnaturalizaci¶on de loscompuestos org¶anicos y la existencia de agua enestado l¶³quido.
3. Agua, matriz b¶asica en la cual se efectuan lasreacciones qu¶³micas que permiten el funciona-miento de los sistemas biol¶ogicos.
Revisemos cada factor. La materia org¶anica, es decir,compuestos formados por uno o m¶as ¶atomos de car-bono, pudo haberse formado espont¶aneamente en lascondiciones de la Tierra primitiva. Alexander I. Opa-rin public¶o en la ex Uni¶on Sovi¶etica en 1924 un tra-bajo donde se se~nalan las condiciones atmosf¶ericasde la Tierra en esos tiempos y establec¶³a la presen-cia de metano, amoniaco, hidr¶ogeno y agua que ba-jo los efectos de fuentes de energ¶³a libre como ra-diaci¶on ultravioleta (radiaci¶on UV) del sol, rayosc¶osmicos y descargas el¶ectricas podr¶³an haber for-mado compuestos org¶anicos m¶as complejos (Opa-rin, 1978). Posteriormente, Haldane en 1929 en In-glaterra, de forma totalmente independiente lleg¶o ala misma conclusi¶on. En 1953 Miller y Urey en unelegante experimento prueban que es factible ob-tener compuestos org¶anicos relativamente comple-jos como amino¶acidos bajo estas condiciones (Mi-ller, 1953). Posteriormente otros investigadores hanlogrado obtener una gran cantidad de compues-tos org¶anicos de complejidad bastante m¶as eleva-da, incluso ¶acidos nucl¶eicos autocatal¶³ticos (Dan-chin, 1988; Orgel, 1994).
En la Tierra primitiva se debieron haber forma-do una gran cantidad de compuestos org¶anicos, sinembargo tambi¶en debieron desnaturalizarse con fa-cilidad, conforme la temperatura disminuy¶o la ta-sa de producci¶on de moleculas org¶anicas debi¶o de-caer en cierto grado, sin detenerse totalmente debi-do a la presencia de otras fuentes de energ¶³a libre co-mo las descargas el ¶ectricas en la atm¶osfera y el im-pacto de cometas. As¶³, la materia org¶anica fue unanecesidad que no fue dif¶³cil cubrir en las etapas tem-pranas de la evoluci¶on planetaria.
La temperatura es un factor que depende de va-rias condiciones, la distancia del planeta a la estre-lla que orbita, la composici¶on de la atm¶osfera, elgrado de inclinaci¶on del eje de rotaci¶on, la veloci-dad de la rotaci¶on, la excentricidad de la ¶orbita e in-cluso la topograf¶³a del planeta in°uyen en la tempe-ratura super¯cial.
La Tierra se ecuentra a una distancia media de1:5 £ 108 Km del Sol (distancia llamada unidad as-tron¶omica o UA), ¶este produce una cantidad enormede energ¶³a que es su¯ciente para mantener una tem-peratura super¯cial promedio en Venus de 460± C,en Marte de ¡60± C y en la Tierra de 15± C, las dis-tancias de cada uno de estos planetas al Sol no sonla explicaci¶on para tales diferencias. La composici¶onde la atm¶osfera es determinante (Kargel y Strom,1996). Siendo el CO2 es el principal gas de efec-to invernadero en la Tierra. La atm¶osfera es transpa-rente para la mayor parte del espectro de la radia-ci¶on solar, la radiaci¶on ultarvioleta queda atrapa-da casi en su totalidad en la capa de ozono de la al-ta atm¶osfera, la radiaci¶on que logra llegar a la super-¯cie se ve re°ejada nuevamente a la atm¶osfera, peroha perdido energ¶³a calentando a la super¯cie, su lon-gitud de onda se ha hecho larga y no puede salir denuevo al espacio, as¶³ queda rebotando repetidamen-te entre el suelo y las capas bajas de la atm¶osfera. Es-to es el fundamento del efecto invernadero; as¶³, aun-que la luz llega desde arriba, la atm¶osfera se calien-ta desde abajo.
El CO2 abunda en la Tierra en las rocas carbonata-das, al nivel del mar alcanza una presi¶on de 0.0003bares (un bar equivale a 1.0197 atm¶osferas), si fue-ra liberado de las rocas alcanzar¶³a unos 60 bares depresi¶on, lo cual producir¶³a un enorme efecto de in-vernadero. Kasting y sus colaboradores (1988) en-contraron que las variaciones en el CO2 no son alea-torias sino que °uctuan en respuesta a determina-das condiciones del ambiente relacionadas a la tem-peratura super¯cial. Cuando la super¯cie se enfria-ba aumentaba la concentraci¶on de di¶oxido de car-bono atmosf¶erico y el planeta se calentaba, cuan-do se calentaba disminu¶³a la liberaci¶on de di¶oxido

20 ContactoS 29, 18{27 (1998)
F ig ura 1 . Esq ue ma de lo s dife re nte s e sta do s de e v o lu-c i¶o n po r lo s c ua le s de bi¶o pa sa r nue stra a tm¶o sfe ra . a )A tm¶o sfe ra primig e nia m¶a s c a lie nte y de nsa q ue la a c -tua l c a ra c te riz a da po r lo s g a se s mo stra do s e n e l e sq ue may ma te ria l de la ne bulo sa o rig ina l. b) A tm¶o sfe ra m¶a s fr¶ ³ay pro ba ble me nte c o nstitu¶ ³da po r CO2;N2, v a po r de a g uay mo l¶ e c ula s o rg ¶a nic a s fo rma da s a bi¶o tic a me nte . c ) C ua n-do la a tm¶o sfe ra se e nfri¶o lo su c̄ ie nte e l a g ua se c o nde n-so fo rma ndo lo s o c ¶ e a no s primig e nio s. d) A tm¶o sfe ra a c -tua l, a lta me nte o x ida nte de bido a l O2 libe ra do po r la fo -to s¶ ³nte sis. T o ma do de La ndsbe rg , 1 9 5 3 .
de carbono a la atm¶osfera y la super¯cie se enfria-ba. Este ciclo de retroalimentaci¶on negativa es el ci-clo del carbono{silicato donde el carbonato hace unrecorrido entre la atm¶osfera y la corteza aproxima-damente en 5 £ 105 a~nos. Este ciclo comienza cuan-do el di¶oxido de carbono atm¶osferico se disuelve en elagua y forma ¶acido carb¶onico (H2CO3), la lluvia ero-siona rocas con carbonatos y silicatos, en tal proce-so el ¶acido carb¶onico reacciona con las rocas y libe-ra iones calcio (Ca+ 2) y bicarbonatos (HCO3
¡) quellegan al agua y ¯nalmente a los oc¶eanos. Este ci-clo debi¶o establecerse poco despu¶es del enfriamien-to de la atm¶osfera y de la formaci¶on de los oc¶eanos.
Actualmente estos iones llegan al mar y son incorpo-rados a las conchas de los organismos como carbona-to c¶alcico (CaCO3), estas conchas se hunden y even-tualmente subducen junto con los sedimentos que lasacogen, siendo sometidas a gran presi¶on y tempera-tura, los carbonatos reaccionan con los silicatos vol-viendo a formar rocas, este proceso es llamado me-tamor¯smo de los carbonatos. Finalmente gracias ala actividad volc¶anica el CO2 regresa a la atm¶osferaen forma de gas (Zaburunov, 1992).
Sup¶ongase que el sol disminuye la energ¶³a que irra-dia y por lo tanto la Tierra comienza a enfriarse,se evapora menos agua del oc¶eano y por lo tan-to llueve menos, as¶³ que tambi¶en la actividad ero-sionadora de la lluvia disminuye, bajo tales condicio-
Fig. 2 Ciclo biogeoqu¶³mico del carbonato{silicato.Con una duraci¶on superior a los 500,000 a~nos pasael carbono de la atm¶osfera a las rocas carbonatadaspara eventualmente regresar a la atm¶osfera. N¶oteseque el papel de los organismos es vital ya que usan losiones calcio y bicarbonato para producir sus conchasy otras estructuras de soporte. Tomado de Kastinget al. 1988.
nes disminuye la tasa con la cual el di¶oxido de carbo-no abandona la atm¶osfera, pero no disminuye la ta-sa a la cual °uye a la atm¶osfera debida a la activi-dad tect¶onica. As¶³, el di¶oxido de carbono se acumu-la en la atm¶osfera y se incrementa el efecto de inver-nadero, con lo cual la temperatura super¯cial no dis-minuye de forma importante. De otro modo, si latemperatura aumentara la evaporaci¶on en los cuer-pos de agua, se producir¶³an m¶as lluvias, aumentar¶³atambi¶en la erosi¶on sobre las rocas y la tasa de aban-dono del di¶oxido de carbono de la atm¶osfera se in-crementar¶³a, as¶³, el efecto de invernadero disminu-ye y la temperatura tambi¶en. Incluso, si los oc¶eanosse congelaran la lluvia cesar¶³a, pero el di¶oxido de car-bono se seguir¶³a acumulando y a la velocidad ac-tual en 20 £ 106 a~nos aumentar¶³a su presi¶on a unbar, lo que elevar¶³a la temperatura de super¯cie aunos 50± C, m¶as que su¯ciente para fundir los hie-los y restablecer temperaturas compatibles con la vi-da. Hoy d¶³a sabemos que aun bajo condiciones ex-tremas de fr¶³o o calor los microorganismos son ca-paces de proliferar (Waters, 1994; Madigan y Marrs,1997).
En otros planetas rocosos como el caso de Martey Venus las cosas han sido diferentes pero ha sidoel efecto de invernadero lo que principalmente hadeterminado el porque de sus condiciones actualesde extremo calor en Venus y de fr¶³o glacial en Marte.(Prinn, 1990; Regis, et al. 1992).

Evoluci¶on de la atm¶osfera y. . . Lozano{Ram¶³rez C., Alvarez{Hern¶andez, S. y De Lara{Isassi, G. 21
Aparici¶on de las principales rutas metab¶olicaspara la obtenci¶on de energ¶³a.
Gluc¶olisisLa evoluci¶on geoqu¶³mica del planeta di¶o las ba-ses para el surgimiento de los sistemas biol¶ogicos,¶estos no tardaron mucho en aparecer. Las prime-ras c¶elulas debieron emerger en un ambiente reduc-tor, sin ox¶³geno libre, rico en compuestos org¶anicosformados abi¶oticamente por concurso de las fuen-tes de energ¶³a libre y de los elementos presentes enla atm¶osfera.
Todos los sistemas biol¶ogicos necesitan energ¶³a pa-ra mantener su actividad. Actualmente, la mol¶eculaenerg¶etica que usan todos los organismos es el trifos-fato de adenosina (ATP), es evidente que en la Tie-rra primitiva no pod¶³a existir ATP biog¶enico, sin em-bargo en experimentos que simulan las condicionesatmosf¶ericas primitivas se ha logrado obtener ATP,as¶³ como muchos otros compuestos energ¶eticos po-tencialmente ¶utiles para estos sistemas, las c¶elulasprimigenias debieron de haber utilizado directamen-te estas mol¶eculas como alimento (Ponnamperuma,et al. 1963). Estos compuestos energ¶eticos fuerondisminuyendo por consumo y porque las condicio-nes de la Tierra fueron siendo cada vez menos favo-rables para su producci¶on. Los organismos tuvieronque empezar a producirlos. La fermentaci¶on es unode los mecanismos m¶as antiguos conocidos para ob-tener energ¶³a, pr¶acticamente todos los organismos lopresentan en alguna etapa de su metabolismo. LouisPasteur la descubri¶o y le llam¶o la vie sans l'air (la vi-da sin aire). El principio de que lo com¶un es lo an-cestral (De la Sota, 1982) nos sugiere de que los an-cestros m¶as antiguos fermentaban y que esa carac-ter¶³stica ha sido heredada a los descendientes. La fer-mentaci¶on se lleva a cabo en ausencia de ox¶³geno yen esencia se re¯ere al rompimiento de una mol¶eculaorg¶anica, casi siempre un az¶ucar. La energ¶³a liberadapor este rompimiento produce ATP a partir de pre-cursores como difosfato de adenosina (ADP) y fos-fato. La fermentaci¶on la encontramos en una grancantidad de v¶³as metab¶olicas, sin embargo la ma-yor¶³a la usan en una ruta llamada gluc¶olisis o v¶³ade Embden{Meyerhof, en honor de Gustav Emb-den y Otto Meyerhof que junto con otros investi-gadores dilucidaron la ruta bioqu¶³mica en 1940, lacual consiste en que a partir de una mol¶ecula de glu-cosa (6 carbonos) obtenemos dos mol¶eculas de pi-ruvato (con 3 carbonos cada una) m¶as una ganan-cia de 2 ATP. En algunas c¶elulas animales, comolas musculares, en presencia de ox¶³geno el piruva-to pasa a ¶acido l¶actico. En algunos microorganis-mos como las levaduras y algunas bacterias el piru-vato o ¶acido pir¶uvico se transforma bajo condicionesmicroaerof¶³licas a CO2 y etanol (C2H5OH). Tam-
bi¶en existe la posibilidad de que en otros microorga-nismos el piruvato se convierta en otro producto ¯-nal, por ejemplo, un ¶acido org¶anico, como el ac¶eticoo vinagre (Lehninger, 1986).
La fermentaci¶on es un proceso que resulta poco e¯-ciente pues las mol¶eculas desechadas todav¶³a alma-cenan cantidades apreciables de energ¶³a, de hecho al-gunos microorganismos usan estas mol¶eculas de de-secho para obtener \energ¶³a de segunda generaci¶on".La reacci¶on neta de la transformaci¶on de glucosa apiruvato es:
Glucosa + 2Pi + 2ADP + 2NAD + ¡!
2 piruvato + 2 ATP + 2 NADH + 2H2O + 2H +
Fijaci¶on del nitr¶ogenoEl nitr¶ogeno participa en la constituci¶on de los ami-no¶acidos, purinas, pirimidinas y otras biomol¶eculas,en forma reducida como NH4. Los eucariontes nopueden convertir el N2 en esta forma. El enlaceN ¡N tiene una energ¶³a de enlace de 225 kcal/mol,lo cual lo hace extraordinariamente resistente al ata-que qu¶³mico. As¶³ el proceso biol¶ogico de ¯jaci¶on re-quiere de un complejo enzim¶atico, el cual consta dedos prote¶³nas, una reductasa que suministra electro-nes con gran poder reductor y una nitrogenasa queusa estos electrones para reducir el N2 a ion amo-nio (NH4
+ ). La conversi¶on del N2 en NH4+ por
este complejo enzim¶atico necesita energ¶³a en for-ma de ATP y un gran poder reductor. En la ma-yor¶³a de los ¯jadores de nitr¶ogeno este poder reduc-tor se obtiene de la ferrodoxina reducida que se ge-nera en el fotosistema I de la fotos¶³ntesis. La este-quiometr¶³a de la reacci¶on catalizada por el comple-jo nitrogenasa es:
N2 + 6e¡ + 12 ATP + 12 H2O ¡!2NH4
+ + 12 ADP + 12 Pi + 4H + :
Como puede observarse este proceso requiere de unagran cantidad de energ¶³a, sin embargo el proceso seve limitado por la presencia de ox¶³geno ya que ¶este seune a la leghemoglobina. El NH4
+ es recogido porotras v¶³as metab¶olicas las cuales lo incorporan a las¶³ntesis de amino¶acidos, ¶estos formar¶an b¶asicamenteprote¶³nas y ¶acidos nucleicos.
La ¯jaci¶on del nitr¶ogeno se ve fuertemente reduci-da a¶un con cantidades muy peque~nas de ox¶³geno, yaque las enzimas nitrogenasas se desactivan en pre-sencia de ¶este. Esto hace suponer que esta ruta me-tab¶olica se desarroll¶o antes de que las concentracio-nes de ox¶³geno en la atm¶osfera se incrementaran.

22 ContactoS 29, 18{27 (1998)
Figura 3. Gr¶a¯ca donde se muestra como el ox¶³genoinhibe la ¯jaci¶on del nitr¶ogeno y fotos¶³ntesis en cia-nobacterias. Tomado de Schopf, J. W. 1988.
Fijaci¶on del carbonoCasi la totalidad de la energ¶³a de la biosfera procededel Sol, la radiaci¶on que emite excita a la cloro¯la delos aut¶otrofos, ¶esta es una por¯rina con un ¶atomo demagnesio coordinado. Diversas mol¶eculas de cloro¯lallevan a los electrones excitados por la luz hasta uncentro de reacci¶on donde la energ¶³a de los electronesse convierte en energ¶³a qu¶³mica que eventualmenteservir¶a para reducir ¶atomos de carbono en el procesode la fotos¶³ntesis.
Los vegetales poseen dos fotosistemas acoplados, enel fotosistema I se obtiene poder reductor como tri-fosfopirid¶³n{nucle¶otido reducido (NADPH), en tan-to que en el fotosistema II se transportan los electro-nes provenientes del agua a una mol¶ecula de quino-na al tiempo que se libera ox¶³geno molecular. El gra-diente de electrones que se genera entre ambos fo-tosistemas crea la energ¶³a su¯ciente para producirATP.
H2O + CO2luz¡! (CH2O) + O2
Ecuaci¶on general de la fotos¶³ntesis.
El NADPH formado por el fotosistema I y el ATPson usados para ¯jar el CO2 y producir az¶ucares, ge-neralmente a partir del 3{fosfoglicerato a trav¶es delas reacciones obscuras en el Ciclo de Calvin, don-de a partir de 2 mol¶eculas de 3 carbonos (3{ fosfo-glicerato) se genera un az¶ucar de 6 carbonos.
Se ha observado que la fotos¶³ntesis de las bacteriasverde{sulfurosas es muy similar a la de las plantasverdes:
CO2 + H2Sluz¡! (CH2O) + 2S + H2O;
donde se libera azufre como producto de desecho,similar al ox¶³geno liberado cuando se utiliza agua.Para formar una hexosa (az¶ucar de seis carbonos) serequieren 6 vueltas al ciclo, ya que en cada vueltase reduce un ¶atomo de carbono. As¶³ la reacci¶on netadel ciclo es:
6CO2 + 18 ATP + 12 NADPH + 12 H2O ¡!
C6H12O6 + 18 ADP + 18 Pi + 12 NADP + + 6H + :
Esta no es la ¶unica forma de ¯jar carbono, exis-ten cuando menos 3 grupos de bacterias que ¯janCO2. La ruta del acetato, usada por el g¶enero Clos-tridium que incluye a especies como C. tetani y C.botulinum causantes del t¶etanos y del botulismo res-pectivamente, presentan quiz¶a la ruta biosint¶eticam¶as antigua de este tipo; la ruta que usa la fosfoe-nol piruvato carboxilasa que se encuentra en bac-terias anaerobias fotosint¶eticas y ¯nalmente la ru-ta de la ribulosa bifosfocarboxilasa que se encuentraen la mayor¶³a de los organismos fotosint¶eticos y qui-mioaut¶otrofos, que debe ser la m¶as reciente.
En todos los organismos fotosint¶eticos oxig¶enicos elfotosistema II complementa al fotosistema I. No seha observado a alg¶un organismo donde el fotosistemaII substutuya al I, esto hace pensar que la fotos¶³ntesisno oxig¶enica se desarroll¶o antes que la fotos¶³ntesisproductora de ox¶³geno.
Hasta mediados de los a~nos 70's se supon¶³a quelos procariontes fotosint¶eticos s¶olo pose¶³an cloro¯-la a, en tanto que las plantas verdes pose¶³an cloro¯-las a y b. El descubrimiento de un procarionte foto-sint¶etico, Prochloron, normalmente encontrado co-mo simbionte de ascidias (organismos marinos ¯l-tradores s¶esiles en estado adulto), posee ambos ti-pos de cloro¯la, as¶³ como fotosistemas I y II indi-ca que la cloro¯la b ya estaba en procariontes an-tes de que aparecieran los fotosint¶eticos superiores yque la fotos¶³ntesis oxig¶enica es un invento de los pro-cariontes en la Tierra primitiva. (Lewin, 1976; 1977).
Respiraci¶onConforme las cianobacterias comenzaron a poblar lasuper¯cie terrestre y a liberar ox¶³geno en cantida-des cada vez mayores, gran cantidad de microbiosincapaces de tolerar el ox¶³geno fueron desaparecien-do o refugi¶andose en h¶abitats marginales sin ox¶³geno.

Evoluci¶on de la atm¶osfera y. . . Lozano{Ram¶³rez C., Alvarez{Hern¶andez, S. y De Lara{Isassi, G. 23
Los organismos que pueden tolerar el ox¶³geno po-seen enzimas protectoras como las catalasas, las pe-roxidasas o las super ¶oxido dismutasas, que al reac-cionar con los radicales producidos por el ox¶³genolo convierten en compuestos org¶anicos inofensivosy agua. Es de suponerse que a lo largo del tiem-po los microorganismos no s¶olo desarrollaron meca-nismos que los protegieran del nuevo gas, sino que loincorporaron en sus rutas metab¶olicas a ¯n de obte-ner una cantidad de energ¶³a mayor, es decir, apren-dieron a respirar.
Cuando la nicotinamida{adenin{dinucle¶otido redu-cida (NADH) y el °av¶³n{aden¶³n{ dinucle¶otido redu-cido (FADH2) formados en la gluc¶olisis, en la oxida-ci¶on de los ¶acidos grasos y en el ciclo de Krebs trans-¯eren sus electrones al ox¶³geno se produce una grancantidad de energ¶³a con la cual se produce ATP. Co-mo parte de la respiraci¶on tenemos el proceso de lafosforilaci¶on oxidativa donde se generan 32 de los36 ATP que se obtienen por oxidar una hexosa has-ta CO2 y H2O.
El ciclo del ¶acido c¶³trico, de los ¶acidos tricarbox¶³licoso de Krebs constituye la fase inicial de la respiraci¶ondonde los carbohidratos, grasas y amino¶acidos ter-minan su ciclo de oxidaci¶on.
La ecuaci¶on general del ciclo de Krebs se presentaen la ecuaci¶on 1.
Posteriormente los NADH y el FADH2 son oxida-dos en la cadena respiratoria, donde se genera ATPal transferir estos electrones hasta el ox¶³geno.
Es importante notar que el ciclo de Krebs es estric-tamente aerobio ya que las mitocondrias s¶olo pue-den regenerar la nicotinamida{aden¶³n{dinucle¶otido(NAD + ) y el °av¶³n{aden¶³n{ dinucle¶otido (FAD)trans¯riendo electrones hasta el ox¶³geno. La fosfo-rilaci¶on oxidativa es el proceso mediante el cual seforma el ATP cuando el NADH y el FADH2 pro-ducidos en el ciclo de Krebs pasan por un jue-go de mol¶eculas transportadoras de electrones queconducen a ¶estos hasta el ox¶³geno. El ox¶³geno mo-lecular es un aceptor de electrones muy e¯ciente,su gran a¯nidad por ¶estos genera una gran energ¶³aen la cadena de transporte electr¶onico. A partir de2 pares electr¶onicos normalmente se obtiene agua(2 mol¶eculas); sin embargo, la reducci¶on parcialgenera al ani¶on super¶oxido, el cual es sumamen-te t¶oxico. Las super¶oxido dismutasas convierten aestos radicales en per¶oxido de hidr¶ogeno (H2O2) yox¶³geno molecular, este per¶oxido es captado por unacatalasa la cual rompe al per¶oxido de hidr¶ogeno enagua y ox¶³geno. Las peroxidasas son otras prote¶³nasque realizan funciones similares con el per¶oxido dehidr¶ogeno, lo cual protege a la c¶elula del ox¶³geno(Trainer, et al. 1983).
As¶³, el balance energ¶etico que da la respiraci¶on aloxidar a una mol¶ecula de glucosa es:
1. C6H12O6 + 2 NAD + + 2 ADP + 2 Pi ¡!2 piruvato + 2 NADH + 2 H2O + 2 ATP , re-cordemos que en las c¶elulas anaer¶obicas en lu-gar de obtener 2 mol¶eculas de piruvato se ob-tienen 2 de lactato. Las 2 mol¶eculas de NADHpasan posteriormente a la cadena respiratoria,donde por cada par de electrones se generan 2mol¶eculas de ATP.
2. 2 NADH + 4 Pi + 4 ADP + O2 ¡!2 NAD + + 6 H2O + 4 ATP . N¶otese que aqu¶³el ox¶³geno ya interviene en las reacciones, hastaeste punto todas las reacciones se han efectuadofuera de las mitocondrias. A partir de la acetilCoA formada en el ciclo de Krebs y para oxidara 2 NADH tenemos:
3. 2 piruvato + 2 NAD + CoA¡!2 acetil CoA + 2 CO2 + 2 NADH
4. 2 NADH + 6 Pi + 6 ADP + O2 ¡!2 NAD + + 8 H2O + 6 ATP . Entonces, de laoxidaci¶on de dos mol¶eculas de acetato efectua-da durante dos vueltas al ciclo de Krebs, don-de en cada vuelta al ciclo ocho electrones seunen a los ox¶³genos. Generalmente obtenemostres mol¶eculas de ATP por cada par de electro-nes, esto da entonces un total de 24 ATP's.
5. 2 acetato + 24 Pi + 24 ADP + 4 O2 ¡!4 CO2 + 28 H2O + 24 ATP . Considerando es-tas ¶ultimas 5 ecuaciones tenemos 36 mol¶eculasde ATP. Lo cual constituye un incremento enor-me comparado con los 2 ATP obtenidos en lafermentaci¶on.
Geo y Bioevoluci¶on>Por qu¶e apareci¶o un proceso como el ciclo del ¶acidoc¶³trico en la evoluci¶on de la vida?. Es muy cla-ro que este mecanismo provee de una gran canti-dad de energ¶³a v¶³a el traslado de electrones hasta elox¶³geno. En este proceso es muy importante notarque el oxalacetato es vital en virtud de que ¶este fun-ciona como acarreador directo hasta el ox¶³geno y tie-ne la caracter¶³stica de ser regenerado constantemen-te por el ciclo, lo cual elimina la necesidad de produ-cirlo de novo constantemente, de otra manera el cos-to energ¶etico de su producci¶on exceder¶³a al produci-do por el ciclo (Baldwin y Krebs, 1981).
Una diferencia importante entre procariontes y eu-cariontes es su capacidad para tolerar el ox¶³geno. Al-gunas bacterias son estrictamente anaerobias y elox¶³geno resulta letal para ellas, otras lo toleran s¶olo

24 ContactoS 29, 18{27 (1998)
Acetil Co A + 3 NAD + + FAD + GDP + Pi + 2 H2O ¡!2 CO2 + 3 NADH + FADH2 + GTP + 2H + + Co A:
Ecuaci¶on 1.
en bajas concentraciones y ¯nalmente otras son to-talmente aerobias, en tanto que los eucariontes sontotalmente aer¶obicos. Los f¶osiles m¶as antiguos iden-ti¯cados son procariontes y alcanzan una edad apro-ximada de 3:7£109 a~nos en el grupo Fig Tree, forma-ci¶on geol¶ogica en Barberton, Africa del Sur (Knolly Barghoorn, 1977; Folger, 1994). Otros investiga-dores se~nalaron f¶osiles de comunidades bacterianascomplejas, tambi¶en con una edad de 3:7£ 109 a~nos.Esto indica que para esta ¶epoca dichas comunida-des ten¶³an un tiempo previo de evoluci¶on (Dunlopet al. 1978; Mestel, 1993).
Los comunidades bacterianas fotosint¶eticas pudieronhaber sido abundantes desde el Proterozoico inferior.Algunos yacimientos que contienen ¯erro y otros mi-nerales oxidados apoyan la idea de que el ox¶³genomolecular ya se encontraba en cantidades importan-tes desde hace unos 2£109 a~nos (Cloud, 1978). Estoimplica un tiempo considerable de producci¶on pre-via de ox¶³geno, tal que estos yacimientos se oxidaran,de hecho este tipo de minerales caracterizan al Ar-queano tard¶³o y Proterozoico temprano, es decir, en-tre 3 £ 109 y 2:6 £ 109 a~nos atr¶as. Se han detecta-do restos de carb¶on ¯jado biol¶ogicamente a las mis-mas tasas que en rocas m¶as jovenes, incluso en es-tratos del Arqueano hace 3:4 £ 109 a~nos, lo que su-giere comunidades fotosint¶eticas (no necesariamenteoxig¶enicas) bien desarrolladas. (Reimer, et al. 1979;Svitil, 1994).
Existe un grupo de sedimentos muy peculiares a loscuales se les llama formaciones bandeadas de ¯erro,¶estos est¶an constituidos por bandas alternadas de ¯e-rro oxidado y no oxidado, as¶³ como de estratos po-bres en ¯erro. Estos sedimentos son t¶³picos del Ar-queano temprano. De hecho estos yacimientos se hanrelacionado a una pobre presencia de ox¶³geno ya quela depositaci¶on de ¯erro no puede darse bajo condi-ciones ricas en ox¶³geno, esto se toma como una prue-ba de una atm¶osfera reductora en ese tiempo. Los ya-cimientos del Proterozoico m¶as temprano empiezana ser abundantes en sedimentos oxidados, donde va-rios minerales de ¯erro altamente oxidados, como lamagnetita, son frecuentes. Para el Proterozoico se-perior, hace unos 750£106 a~nos la atm¶osfera era de-cididamente oxidante.
Al ¯nal del Hadeano, hace 3:9 £ 109 a~nos se carac-teriz¶o por una gran actividad volc¶anica, es en es-ta ¶epoca donde se establece la estructura b¶asica de
la Tierra y es donde debieron tambi¶en haberse gene-rado los primeros sistemas vivientes. No es f¶acil lo-calizar ni identi¯car f¶osiles de esta antigÄuedad yaque las rocas con esta edad han sufrido metamor¯s-mo severo en muchas localidades. La vida surgi¶o po-co despu¶es de que la evoluci¶on terrestre hab¶³a alcan-zado cierto grado de \estabilidad", despu¶es de la for-maci¶on de los oc¶eanos y del establecimiento del sis-tema Tierra{Luna. (Hargraves, 1976).
Esto implica que despu¶es de haberse agotado lasmol¶eculas org¶anicas, formadas espont¶aneamente enbase a procesos abi¶oticos, que sirvieron de alimento alos primeros seres vivos mediante la fermentaci¶on sedebieron innovar nuevas rutas metab¶olicas que poruna parte fueran m¶as e¯cientes y por otra fueran ca-paces de utilizar los elementos disponibles. Una fuen-te energ¶etica practicamente inagotable es la luz, lacual fue usada en la fotos¶³ntesis no oxig¶enica. Poste-riormente los aut¶otrofos incorporaron un segundo fo-tosistema al ya existente, logrando m¶as energ¶³a parala ¯jaci¶on del carbono, esta innovaci¶on ¯siol¶ogica li-beradora de O2 tuvo un impacto fundamental en laevoluci¶on de la vida.
F¶osiles de fotosintetizadores muy parecidos a ciano-bacterias actuales se han encontrado en la regi¶on delTransvaal, en Africa del Sur, que datan aproximada-mente de 2:3 £ 109 a~nos, estos f¶osiles presentan es-tructuras sumamente parecidas a acinetos y hete-rocistos, estructuras especializadas para la ¯jaci¶ondel nitr¶ogeno hoy d¶³a. No es dif¶³cil suponer que es-tas estructuras ya tuvieran esa funci¶on desde en-tonces. (Nagy et al. 1977). La antigÄuedad de es-tos f¶osiles as¶³ como las caracter¶³sticas metab¶olicasde la ruta de ¯jaci¶on del nitr¶ogeno sugieren que ¶estay la fotos¶³ntesis no oxig¶enica surgieron con poca di-ferencia en el tiempo. Con alto grado de probabili-dad la mayor¶³a de las rutas anaerobias para la ob-tenci¶on de energ¶³a debieron surgir en esta ¶epoca.
Pudi¶endose ya obtener nitr¶ogeno y carbono en abun-dancia de la atm¶osfera, los organismos escaparonde la posibilidad de extinguirse al agotarse lasmol¶eculas formadas abioticamente. El cambio pos-terior de una atm¶osfera reductora a una oxidantepropici¶o el proceso de coevoluci¶on de diversos mi-croorganismos, lo que culmin¶o con la aparici¶on delos eucariontes, organismos con c¶elulas nucleadas,con organelos altamente especializados para la ob-tenci¶on de energ¶³a, los cloroplastos y mitocondrias,

Evoluci¶on de la atm¶osfera y. . . Lozano{Ram¶³rez C., Alvarez{Hern¶andez, S. y De Lara{Isassi, G. 25
Figura 4. a) Fotograf¶³a de estromatolitos vivos, co-munidades bacterianas en Shark Bay, Australia. b)F¶osil de un microorganismo tomado de un estroma-tolito de aproximadamente 3:69 a~nos. c) Esquemadel f¶osil del inciso b. Tomado de Orgel, 1994.
ambos estrictamente aerobios. Los procariontes \in-ventan"las rutas metab¶olicas b¶asicas que heredan alos eucariontes, ¶estos hacen sus propias innovacio-nes para producir metabolitos que los procariontesno pueden sintetizar, tal es el caso de los esteroi-des, componentes importantes de la membrana celu-lar, de los n¶ucleos de gran cantidad de hormonas ycuya bios¶³ntesis es aer¶obica (se presume que algunosprocariontes si pueden sintetizar esteroides). Los eu-cariontes estamos tan comprometidos con el ox¶³genoque incluso la mitosis celular no es posible sin ¶el.
La evoluci¶on de la atm¶osfera y la corteza terres-tre ha dirigido a la evoluci¶on biol¶ogica, los organis-mos tambi¶en han hecho lo propio. El oc¶eano inter-cambi¶o gran cantidad de CO2 con la atm¶osfera esta-bleci¶endose un equilibrio entre ambos. Actualmentela cantidad de carbono disuelto en el mar es aproxi-madamente 50 veces mayor que en la atm¶osfera y es-tos dos reservorios lo intercambian a un ritmo tanelevado como de 90£109 toneladas, tal que la canti-dad total de carbono en la atm¶osfera se renueva cada8 a~nos. El oc¶eano tiene en sus sedimentos una canti-dad colosal de carbono en forma de carbonato de cal-cio. As¶³ el oc¶eano ha regulado la cantidad de carbo-no en la atm¶osfera a trav¶es del tiempo con una e¯-ciencia muy alta. (Minster y Merlivat, 1992). Es-ta capacidad de disoluci¶on del CO2 en los oc¶eanosdurante la formaci¶on de ¶estos debi¶o regular el efec-to invernadero y por lo tanto la temperatura super-¯cial de la Tierra. Al estar la mayor parte del car-bono secuestrado en los sedimentos marinos y en ro-cas calc¶areas, la Tierra no se dirigi¶o a un efecto in-vernadero incontrolable como ocurri¶o en Venus. Seestima que las ¶areas terrestres del planeta ¯jan de20 £ 109 a 30 £ 109 toneladas de carb¶on por a~no,en el mar el ¯toplancton ¯ja unas 40 £ 109 tonela-das por a~no (Bolin, 1970). Los bosques y selvas con-tienen entre 40£ 109 y 50£ 109 toneladas de carbo-no, lo que equivale a dos tercios de la cantidad de car-bono en la atm¶osfera. En los oc¶eanos primitivos lascomunidades bacterianas debieron ¯jar carbono en
cantidades equivalentes a los oc¶eanos de hoy, es evi-dente que la cobertura vegetal terrestre no exist¶³ay por lo tanto no participaba en el proceso. Los or-ganismos debido a su ciclo de vida tan corto reci-clan muy rapidamente la materia org¶anica, al mo-rir y sedimentarse en el mar se disuelven, una par-te de esta materia org¶anica no se oxida y llega al fon-do donde forma sedimentos ricos en carbono. A es-tas profundidades la tasa de reciclamiento entre el se-dimento y el agua es muy lenta y recordemos que en-tre m¶as grande sea la profundidad menor ser¶a la can-tidad de ox¶³geno, adem¶as las corrientes marinas tar-dan aproximadamente unos 1,000 a~nos en recircu-lar el agua de los oc¶eanos, por la tanto ese carbo-no tarda mucho tiempo en regresar a aguas super-¯ciales, donde potencialmente puede intercambiar-se con la atm¶osfera a tasas elevadas. Los sedimen-tos marinos ricos en carbono pueden subducir ba-jo una placa continental debido al proceso de derivacontinental y entonces el carbono puede ser devuel-to a la atm¶osfera v¶³a actividad volc¶anica. Sin em-bargo, la mayor cantidad de carbono est¶a atrapa-do en dep¶ositos carbonatados inorg¶anicos que alcan-zan 20,000 billones de toneladas. Este ciclo en el queinicialmente no hab¶³a vida, permiti¶o al planeta teneruna atm¶osfera con una temperatura capaz de permi-tir la formaci¶on y no desnaturalizaci¶on de mol¶eculasorg¶anicas as¶³ como la existencia de agua en esta-do l¶³quido. La vida logr¶o cambiar toda la atm¶osferade un planeta, generando dep¶ositos de minerales queno se habr¶³an podido formar de otra manera, as¶³ co-mo ser elemento esencial en la regulaci¶on del ciclodel carbono y dando nuevos rumbos a su propia evo-luci¶on (Allµegre y Schneider, 1994).
Dos entidades interdependientes han generado unabiosfera con relaciones tan complejas y sutiles quehacen de la Tierra un planeta ¶unico y maravilloso,tratemos de entenderlo para conservar ese delicadoequilibrio que hace posible a la vida.
B ib liogra f¶³a
1 . A llµ e g re , C . J. a nd Sc hne ide r, S. H . 1 9 9 4 . T he Ev o -lutio n o f the Ea rth. Scienti¯c A merican. V o l. 2 7 1 .N o . 4 . 4 4 { 5 1 .
2 . Ba ldw in, J. E. a nd K re bs, H . 1 9 8 1 . T he Ev o lutio no f M e ta bo lic C y c le s. N atu re 291: 381{382.
3 . Ba rtho lo me w N a g y y Lo uis A nne N a g y . 1 9 8 2 . Lo sprime ro s o rg a nismo s v iv ie nte s. E l Redes cu brimientode la T ierra. 1 0 3 { 1 1 2 . C ON A C y T . M ¶ e x ic o .
4 . Bo lin, B. 1 9 7 0 . El C ic lo de l c a rbo no . En E colog¶³a,E volu ci¶on y Biolog¶³a de las P oblaciones . 1 9 7 8 Ed.Ome g a S. A ., Ba rc e lo na , Espa ~na .
5 . C a rto n, Y v e s. 1 9 8 8 . La c o ¶ e v o lutio n. L a Recherche1 0 2 2 { 1 0 3 1 .
6 . C lo ud, P. E. Jr. 1 9 7 8 . Ev o lutio n o f Ec o sy ste ms. Sci.A m. 6 2 : 5 4 { 6 6
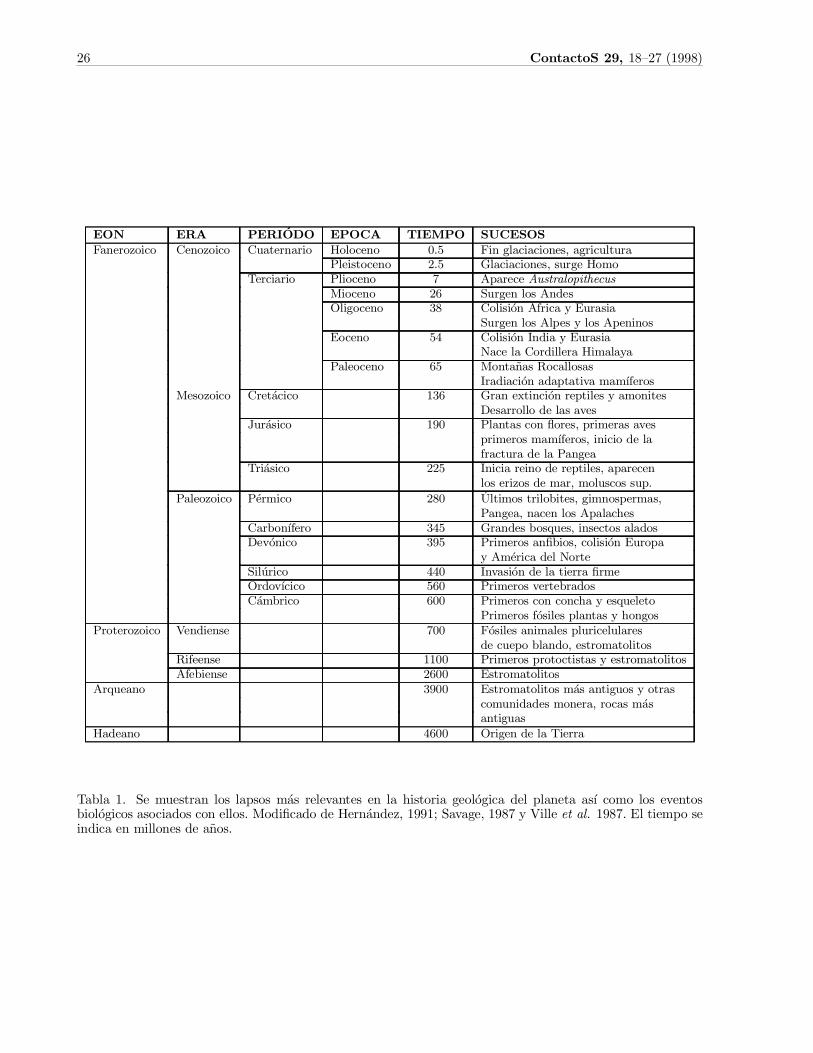
26 ContactoS 29, 18{27 (1998)
EON ERA P ERI¶ODO EP OC A TIEM P O S UC ES OSF a ne ro z o ic o C e no z o ic o C ua te rna rio H o lo c e no 0 .5 F in g la c ia c io ne s, a g ric ultura
Ple isto c e no 2 .5 Gla c ia c io ne s, surg e H o moT e rc ia rio Plio c e no 7 A pa re c e A u s tralopithecu s
M io c e no 2 6 Surg e n lo s A nde sOlig o c e no 3 8 C o lisi¶o n A fric a y Eura sia
Surg e n lo s A lpe s y lo s A pe nino sEo c e no 5 4 C o lisi¶o n India y Eura sia
N a c e la C o rdille ra H ima la y aPa le o c e no 6 5 M o nta ~na s Ro c a llo sa s
Ira dia c i¶o n a da pta tiv a ma m¶ ³fe ro sM e so z o ic o C re t¶a c ic o 1 3 6 Gra n e x tinc i¶o n re ptile s y a mo nite s
D e sa rro llo de la s a v e sJur¶a sic o 1 9 0 Pla nta s c o n °o re s, prime ra s a v e s
prime ro s ma m¶ ³fe ro s, inic io de lafra c tura de la Pa ng e a
T ri¶a sic o 2 2 5 Inic ia re ino de re ptile s, a pa re c e nlo s e riz o s de ma r, mo lusc o s sup.
Pa le o z o ic o P¶e rmic o 2 8 0 ¶U ltimo s trilo bite s, g imno spe rma s,Pa ng e a , na c e n lo s A pa la c he s
C a rbo n¶ ³fe ro 3 4 5 Gra nde s bo sq ue s, inse c to s a la do sD e v ¶o nic o 3 9 5 Prime ro s a n¯bio s, c o lisi¶o n Euro pa
y A m¶e ric a de l N o rteSil¶uric o 4 4 0 Inv a si¶o n de la tie rra ¯rmeOrdo v ¶ ³c ic o 5 6 0 Prime ro s v e rte bra do sC ¶a mbric o 6 0 0 Prime ro s c o n c o nc ha y e sq ue le to
Prime ro s f¶o sile s pla nta s y ho ng o sPro te ro z o ic o V e ndie nse 7 0 0 F ¶o sile s a nima le s pluric e lula re s
de c ue po bla ndo , e stro ma to lito sRife e nse 1 1 0 0 Prime ro s pro to c tista s y e stro ma to lito sA fe bie nse 2 6 0 0 Estro ma to lito s
A rq ue a no 3 9 0 0 Estro ma to lito s m¶a s a ntig uo s y o tra sc o munida de s mo ne ra , ro c a s m¶a sa ntig ua s
H a de a no 4 6 0 0 Orig e n de la T ie rra
Tabla 1. Se muestran los lapsos m¶as relevantes en la historia geol¶ogica del planeta as¶³ como los eventosbiol¶ogicos asociados con ellos. Modi¯cado de Hern¶andez, 1991; Savage, 1987 y Ville et al. 1987. El tiempo seindica en millones de a~nos.

Evoluci¶on de la atm¶osfera y. . . Lozano{Ram¶³rez C., Alvarez{Hern¶andez, S. y De Lara{Isassi, G. 27
7 . C o urtin, R., M c k a y , C . P. y Po lla c k , J. 1 9 8 1 . ElEfe c to Inv e rna de ro e n e l Siste ma So la r. M u ndoC ient¶³¯co. V o l. 1 2 , N o . 1 2 6 . 6 3 2 { 6 3 8 .
8 . D a nc hin, A . 1 9 8 8 . L'Orig ine de la V ie . L a Recher-che. V o l. 1 9 N o . 2 0 1 . 8 7 8 { 8 8 6
9 . D e la So ta , E. R. 1 9 8 2 . La ta x o no mia y la Re v o -luc i¶o n de la s C ie nc ia s Bio l¶o g ic a s. Sr¶ ³a . Ge ne ra l dela Org a niz a c i¶o n de lo s Esta do s A me ric a no s. Pro g ra -ma Re g io na l de D e sa rro llo C ie nt¶ ³¯c o y T e c no l¶o g ic o .M ¶ e x ic o D . F . pp. 9 0
1 0 . D unlo p, J. S. R., M . D . M uir, V . A . M ilne , a ndD . I. Glo v e s. 1 9 7 8 . A N e w M ic ro fo ssil A sse mbla -g e fro m the A rc ha e a n o f We ste rn A ustra lia . N atu -re 274: 6 7 6 { 6 7 8 .
1 1 . F o lg e r, T . 1 9 9 4 . Primo rdia l La ndlubbe rs. Dis cover.V o l. 1 5 . N o . 4 . 2 8 { 2 9 .
1 2 . H a rg ra v e s, R. B. 1 9 7 6 . Pre c a mbria n Ge o lo g ic H is-to ry . Science. 193: 3 6 3 { 3 7 1 .
1 3 . H e rn¶a nde z La sc a re s, D e l¯no . 1 9 9 1 . L a es cala deltiempo geologico, la deriva de los continentes y latectonica de placas . Ed. U niv e rsida d A uto no ma M e -tro po lita na U . Iz ta pa la pa .
1 4 . K a rg e l, J. S. a nd Stro m, R. G. 1 9 9 6 . Glo ba l C lima -tic C ha ng e o n M a rs. Scienti¯c A merican. V o l. 2 7 5 .N o . 5 . 6 0 { 6 8 .
1 5 . K a sting , Ja me s F ., Ow e n B. T o o n y Ja me s B. Po -lla c k . 1 9 8 8 . Ev o luc i¶o n de l C lima e n lo s Pla ne ta s T e -rre stre s. Inves tigaci¶on y C iencia. N o . 1 3 9 . 4 8 { 5 7
1 6 . K no ll, A . H ., a nd E. S. Ba rg ho o rn. 1 9 7 7 . A rc ha e a nM ic ro fo ssils sho w ing C e ll D iv isio n fro m the sw a z i-la nd Sy ste m o f So uth A fric a . Science 198: 3 9 6 { 3 9 8 .
1 7 . Le hning e r, A . L. 1 9 8 6 . Bioenerg¶etica. Siste ma s T ¶ e c -nic o s de Edic i¶o n, S.A de C .V
1 8 . Le w in, R. A . 1 9 7 6 Pro c hlo ro phy ta a s a pro po se dne w div isio n o f a lg a e . N atu re 261: 6 9 7 { 6 9 8 .
1 9 . Le w in, R. A . 1 9 7 7 . Pro c hlo ro n, tipe g e nus o f thePro c hlo ro phy ta . P hycologya 16: 2 1 7 .
2 0 . Lo v e lo c k , J. 1 9 8 7 . Ga ia : A N ew L ook of L ife onE arth. Ox fo rd U niv e rsity Pre ss. U .S.A .
2 1 . M a dig a n, M . T . A nd M a rrs, B. L. 1 9 9 7 . Ex tre mo p-hile s. Scienti¯c A merican. V o l. 2 7 6 . N o . 4 . 6 6 { 7 1 .
2 2 . M e ste l, R. 1 9 9 3 . Pushing Ba c k Life 's Orig in. E arth.V o l. 2 . N o . 6 . 1 6 .
2 3 . M ille r, S.L. 1 9 5 3 . A pro duc tio n o f a mino a c idsunde r po ssible primitiv e Ea rth c o nditio ns. Scien-ce 117: 5 2 8 { 5 2 9 .
2 4 . M inste r, Je a n{ fra n»c o ise y Lilio ne M e rliv a t 1 9 9 2 .>A D o nde V a e l Ga s C a rb¶o nic o ? El Pa pe l de lo sOc ¶ e a no s. M u ndo C ient¶³¯co N o . 1 2 6 . V o l. 1 2 : 6 8 2 {6 8 6 .
2 5 . N a g y , B., L. A . N a g y , J. E. Zumbe rg e , D . S. Sk la -re w a nd P. H . A nde rso n. 1 9 7 7 . Indic a tio n o f a Bio -lo g ic a l a nd Bio c he mic a l Ev o lutio na ry T re nd D uringthe A rc ha e a n a nd Ea rly Pro te ro z o ic . P recambrianRes . 5: 1 0 9 { 2 0 1 .
2 6 . Org e l, L. E. 1 9 9 4 . T he Orig in o f Life o n the Ea rth.Scienti¯c A merican V o l. 2 7 1 . N o , 4 . 5 2 { 6 1 .
2 7 . Opa rin, A .I. Ed. 1 9 7 8 . E l O rigen de la V ida. Edito -re s M e x ic a no s U nido s. M ¶ e x ic o .
2 8 . Po nna mpe ruma , C ., C . Sa g a n, a nd R. M a rine r.1 9 6 3 . Sy nthe sis o f A T P unde r po ssible primitiv eEa rth c o nditio ns. N atu re 199: 2 2 2 { 2 2 6 .
2 9 . Prinn, Ro na ld G. 1 9 9 0 . T he V o lc a no e s a nd C lo udso f V e nus. Scienti¯c A merican. V o l. 2 . N o . 1 . 9 4 { 1 0 1 .
3 0 . Ra so o l. S.I. 1 9 7 2 . P lanetary atmos pheres in E x obio-logy. 3 6 9 { 3 9 9 e d. by C y ril Po nna mpe ruma . A me ri-c a n Else v ie r Publishing C o mpa ny , Inc . N .Y . U :S:A
3 1 . Re g is C o urtin, C hristo phe r P. M c k a y y Ja me s Po -lla c k . 1 9 9 2 . El Efe c to Inv e rna de ro e n e l Siste ma So -la r. M u ndo C ient¶³¯co. V o l. 1 2 . N o . 1 2 6 . 6 3 2 { 6 3 8 .
3 2 . Re ime r, T ., E. S. Ba rg ho o rn, a nd L. M a rg ulis. 1 9 7 9 .Prima ry Pro duc tiv ity in a n Ea rly A rc ha e a n M ic ro -bia l Ec o sy ste m. P recambrian Res . 9: 9 3 { 1 0 4 .
3 3 . Rho de s W. F a irbridg e . 1 9 8 2 . H isto ria de l C lima e nla T ie rra .2 3 3 { 2 5 0 . En E l Redes cu brimiento de laT ierra. C ON A C y T . M ¶ e x ic o .
3 4 . Sa v a g e , J. M . 1 9 8 7 . E volu cion. Ed. C .E.C .S.A . M e -x ic o , D .F .
3 5 . Sc ho pf, J. Willia m. 1 9 7 8 . La Ev o luc i¶o n de la sC ¶ e lula s Primitiv a s. Inves tigaci¶on y C iencia. N o . 2 6 .5 8 { 7 5 .
3 6 . Sv itil, K . A . 1 9 9 4 . M ig hty M ic ro be s. Dis cover. V o l.1 5 . N o . 2 . 2 5 .
3 7 . T ra ine r, J. A ., Ge tz o ®, E. D ., Ric ha rdso n, j. S. a ndRic ha rdso n, D . C . 1 9 8 3 . Struc ture a nd M e c ha nismo f C u, Zn supe ro x ide dismuta se . N atu re 306: 2 8 4 {2 8 7 .
3 8 . V ille , C . A .,So lo mo n, E. P., y D a v is P. W. 1 9 8 7 .Biologia. Ed. Inte ra me ric a na S. A . de C . V .
3 9 . Wa te rs, T . 1 9 9 4 . Life in a Ro c k . E arth. V o l. 3 . N o .4 . 2 0 { 2 3 .
4 0 . Za buruno v , S. A . 1 9 9 2 . A s the Wo rld Bre a the s. T heC a rbo n D io x ide C y c le . E arth. V o l.1 . N o . 1 . 2 6 { 3 3 .
cs




















