
Evidence for Temperature Elevation in the Aerobic Swimming Musculature of the Common Thresher Shark,...
6
q 2005 by the American Society of Ichthyologists and Herpetologists SHORTER CONTRIBUTIONS Copeia, 2005(1), pp. 146–151 Evidence for Temperature Elevation in the Aerobic Swimming Musculature of the Common Thresher Shark, Alopias vulpinus DIEGO BERNAL AND CHUGEY A. SEPULVEDA This paper reports on the in vivo temperature measurements for slow-twitch, red aerobic muscle (RM) and fast twitch, white muscle (WM) of 24 Common Thresher Sharks, Alopias vulpinus, captured off the coast of Southern California. Mean (6 SE) RM temperature elevation was 2.33 6 0.30 C warmer than the ambient sea surface temperature (SST, 19.13 6 0.22 C) and 2.25 6 0.35 C warmer than the coldest WM temperature measured (19.23 6 0.30 C). The maximum RM tempera- ture elevation was 5.4 C above SST and up to 7.6 C above the WM. Nine Common Thresher Sharks exhibited RM temperatures that exceeded 3.0 C above SST. No significant relationship was found between fish total length and RM temperature elevation. Although there was individual variability in RM temperature elevation, this study documents that the Common Thresher Shark is capable of significantly elevating its RM temperature above that of the ambient conditions. T HE Common Thresher Shark, Alopias vul- pinus, is a large (maximum total length 760 cm, body mass 348 kg) actively swimming pelagic species classified in the Family Alopiidae (Hart, 1973; Compagno, 2001; R. Froese and D. Pauly, unpubl., www.fishbase.org). The internal anatomy of the swimming musculature of A. vul- pinus closely resembles that of the endothermic sharks (e.g., Shortfin Mako Shark, Isurus oxyrin- chus) in the Family Lamnidae (Carey and Teal, 1969; Carey et al., 1971; Carey et al., 1985) in having the red aerobic muscle (RM) located in an axial position (i.e., closer to the vertebral col- umn) and the circulation to this tissue is through a putative counter-current heat ex- changer (retia; Bone and Chubb, 1983; Bernal et al., 2003; pers. obs.). Although morphologi- cal similarities between the Common Thresher Shark and the lamnids have been used to sug- gest that threshers are capable of regional en- dothermy, no in vivo temperature measure- ments exist for this species. The only two in vivo RM temperature measurements that exist for any alopiid shark were taken from two Bigeye Thresher Sharks (Alopias superciliosus; Carey et al., 1971), for which the RM temperature was warmer than ambient, in one specimen, and cooler than ambient in the other. Collectively, the inconclusive RM temperature data for the Bigeye Thresher (Carey et al., 1971) and the lack of temperature measurements for the other two thresher species (i.e., Common Thresher and Pelagic Thresher, Alopias pelagicus) presents the question as to whether this group is capable of RM endothermy as are the lamnid sharks. The objective of this communication is to pre- sent in vivo temperature measurements for the RM and white muscle (WM) of the Common Thresher Shark and to test whether this species is capable of elevating its RM temperature ele- vation above ambient, indicating regional en- dothermy. MATERIALS AND METHODS Specimen collection.—Twenty-four Common Thresh- er Sharks were captured off the coast of Southern California from 1999–2003. Seven sharks were captured using rod and reel techniques at depths from the surface (0 m) to 30 m, whereas 17 sharks were captured using a 4 km longline (O’Brien and Sunada, 1994) during the 1999 National Marine Fisheries Service (NMFS) shark indexing survey. Longline hook-depths ranged from 35–50 m and the thermal profile of the water column was recorded using man- ually launched expendable bathythermographs (XBT, Sippican, Inc.; Marion, MA). When pos- sible, after recording morphological data (e.g., total length, estimated mass, sex) and the phys- ical condition of the animal, the sharks were tagged with NMFS conventional spaghetti tags and released. Heavy fishing tackle (24 kg) was used to collect all sharks by rod and reel in an
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Evidence for Temperature Elevation in the Aerobic Swimming Musculature of the Common Thresher Shark,...

q 2005 by the American Society of Ichthyologists and Herpetologists
SHORTER CONTRIBUTIONS
Copeia, 2005(1), pp. 146–151
Evidence for Temperature Elevation in the Aerobic SwimmingMusculature of the Common Thresher Shark, Alopias vulpinus
DIEGO BERNAL AND CHUGEY A. SEPULVEDA
This paper reports on the in vivo temperature measurements for slow-twitch, redaerobic muscle (RM) and fast twitch, white muscle (WM) of 24 Common ThresherSharks, Alopias vulpinus, captured off the coast of Southern California. Mean (6SE) RM temperature elevation was 2.33 6 0.30 C warmer than the ambient seasurface temperature (SST, 19.13 6 0.22 C) and 2.25 6 0.35 C warmer than thecoldest WM temperature measured (19.23 6 0.30 C). The maximum RM tempera-ture elevation was 5.4 C above SST and up to 7.6 C above the WM. Nine CommonThresher Sharks exhibited RM temperatures that exceeded 3.0 C above SST. Nosignificant relationship was found between fish total length and RM temperatureelevation. Although there was individual variability in RM temperature elevation,this study documents that the Common Thresher Shark is capable of significantlyelevating its RM temperature above that of the ambient conditions.
THE Common Thresher Shark, Alopias vul-pinus, is a large (maximum total length
760 cm, body mass 348 kg) actively swimmingpelagic species classified in the Family Alopiidae(Hart, 1973; Compagno, 2001; R. Froese and D.Pauly, unpubl., www.fishbase.org). The internalanatomy of the swimming musculature of A. vul-pinus closely resembles that of the endothermicsharks (e.g., Shortfin Mako Shark, Isurus oxyrin-chus) in the Family Lamnidae (Carey and Teal,1969; Carey et al., 1971; Carey et al., 1985) inhaving the red aerobic muscle (RM) located inan axial position (i.e., closer to the vertebral col-umn) and the circulation to this tissue isthrough a putative counter-current heat ex-changer (retia; Bone and Chubb, 1983; Bernalet al., 2003; pers. obs.). Although morphologi-cal similarities between the Common ThresherShark and the lamnids have been used to sug-gest that threshers are capable of regional en-dothermy, no in vivo temperature measure-ments exist for this species. The only two in vivoRM temperature measurements that exist forany alopiid shark were taken from two BigeyeThresher Sharks (Alopias superciliosus; Carey etal., 1971), for which the RM temperature waswarmer than ambient, in one specimen, andcooler than ambient in the other. Collectively,the inconclusive RM temperature data for theBigeye Thresher (Carey et al., 1971) and thelack of temperature measurements for the othertwo thresher species (i.e., Common Thresher
and Pelagic Thresher, Alopias pelagicus) presentsthe question as to whether this group is capableof RM endothermy as are the lamnid sharks.The objective of this communication is to pre-sent in vivo temperature measurements for theRM and white muscle (WM) of the CommonThresher Shark and to test whether this speciesis capable of elevating its RM temperature ele-vation above ambient, indicating regional en-dothermy.
MATERIALS AND METHODS
Specimen collection.—Twenty-four Common Thresh-er Sharks were captured off the coast of SouthernCalifornia from 1999–2003. Seven sharks werecaptured using rod and reel techniques at depthsfrom the surface (0 m) to 30 m, whereas 17sharks were captured using a 4 km longline(O’Brien and Sunada, 1994) during the 1999National Marine Fisheries Service (NMFS)shark indexing survey. Longline hook-depthsranged from 35–50 m and the thermal profileof the water column was recorded using man-ually launched expendable bathythermographs(XBT, Sippican, Inc.; Marion, MA). When pos-sible, after recording morphological data (e.g.,total length, estimated mass, sex) and the phys-ical condition of the animal, the sharks weretagged with NMFS conventional spaghetti tagsand released. Heavy fishing tackle (24 kg) wasused to collect all sharks by rod and reel in an

147BERNAL AND SEPULVEDA—THRESHER MUSCLE TEMPERATURE
Fig. 1. Cross-sectional slice of Common ThresherShark #20 taken midway along the body (under thefirst dorsal fin) showing the transverse trajectory inwhich red and white muscle temperature measure-ment were obtained using a thermocouple thermom-eter.
attempt to minimize the struggle and fighttimes associated with capture and once alongside the boat. Rapid handling of the sharks al-lowed for the measurement of muscle temper-atures within 1 min of the shark reaching thefishing vessel. Struggle time for longline cap-tured fishes is unknown and may have rangedfrom a maximum of approximately 240 min (to-tal gear soak time) to a minimum of 15 min(immediately prior to gear haul back).
Temperature measurements.—Muscle temperaturemeasurements were taken from sharks as theywere brought alongside the fishing vessel butwhile still partially submerged in the water. A0.5 cm incision was made using a surgical bladeat 20–30% total length (TL; i.e., below the firstdorsal fin) where the relative amount of RM ishighest (Bernal et al., 2003). A thermocouplethermometer (Barnant model 600) was insertedtransversely into the swimming musculature, ap-proximately 2–4 cm beneath the skin and a tem-perature measurement of WM was recorded.Thermocouple trajectory is shown in Figure 1and illustrates the path taken by temperatureprobe from the WM until reaching the RM. Im-mediately after the muscle temperatures weretaken the thermocouple probe was immersed inthe surrounding water to measure the SST dur-ing rod and reel collection and to confirm theSST obtained by the XBT during the longlineoperations.
Statistical analyses.—The mean RM and WM tem-perature elevations above SST were comparedusing a Student’s Paired t-test and comparisonsof mean temperature elevations for RM andWM between the different collection tech-niques were done with a Student’s two samplet-test. A least-squares linear regression analysiswas used to determine the presence of a signif-icant relationship between RM temperature andfish length. All tests were performed using an aof 0.05.
RESULTS
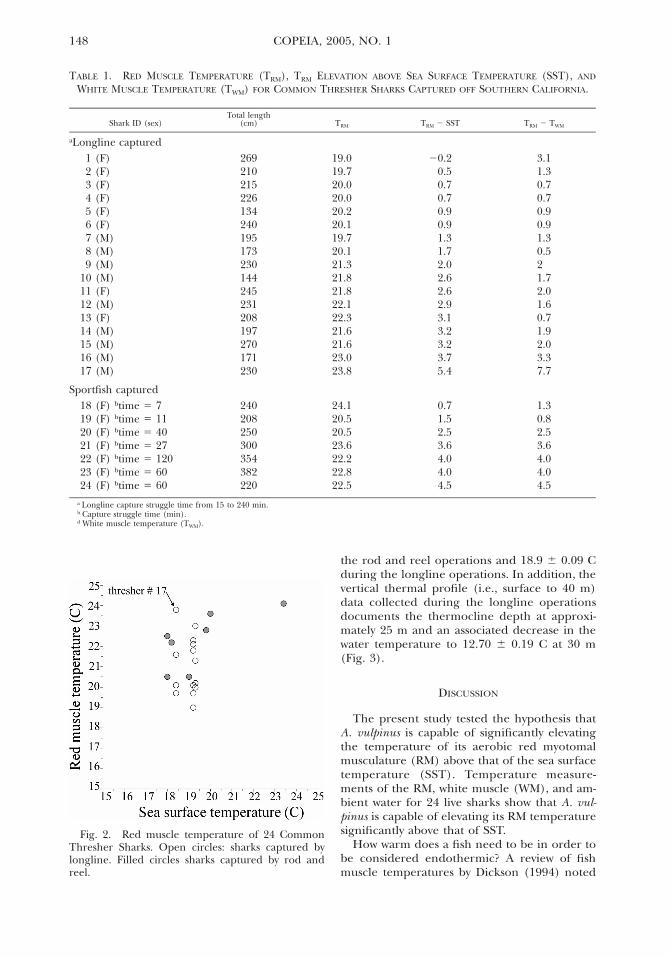
Red muscle temperature data were obtainedfrom 24 Common Thresher Sharks ranging inTL from 134–382 cm (80–215 cm fork length,FL) and with an estimated body mass rangingfrom 3–150 kg. There was no significant rela-tionship between body size (i.e., FL) and RMtemperature elevation above SST (r2 5 0.09, F5 2.24, P 5 0.14, df 5 1,22), but there was asignificant relationship (r2 5 0.23, F 5 6.61, P5 0.017, df 5 1,22) between FL and RM tem-perature elevation above WM (described by TRM-
WM 5 0.0265 3 FL2 1.28, where TRM-WM is theRM temperature elevation above WM and FL isin cm). All but one of the Common Threshershad a RM temperature that was warmer thanthe ambient SST and all RM temperatures werewarmer than those of the WM (Table 1). Themean (6 SE) RM temperature for all threshers(21.47 6 0.31 C) was significantly higher (t 57.76, P , 0.001, df 5 23) than the mean SST(19.13 6 0.22 C), but there was no significantdifference (t 5 0.36, P 5 0.71, df 5 23) betweenthe mean SST and the mean WM temperature(19.22 6 0.30 C). The mean RM temperatureelevation for all threshers was 2.33 6 0.30 Cabove SST and 2.25 6 0.35 C above WM, withCommon Thresher #17 having the warmest rel-ative RM temperature (i.e., 5.4 C above SST and7.6 C above WM, Table 1, Fig. 2). There was nosignificant difference between the mean RMtemperature elevation above SST (t 5 1.38, P 50.17, df 5 22) for threshers caught by rod andreel (2.97 6 0.54 C) and longline (2.07 6 0.35C) and no significant difference (t 5 1.61, P 50.11, df 5 22) was determined between the RMtemperature elevation above WM for sharkscaught by rod and reel (3.10 6 0.61 C) or bylongline (1.89 6 0.41 C). For seven rod and reelcaught sharks, there was no significant relation-ship between the RM temperature elevationabove SST and time spent struggling on the line(r2 5 0.52, F 5 5.59, P 5 0.064, df 5 1,5).
Sea surface temperatures (SST) ranged from18–23 C with a mean of 19.5 6 0.72 C during
148 COPEIA, 2005, NO. 1
TABLE 1. RED MUSCLE TEMPERATURE (TRM), TRM ELEVATION ABOVE SEA SURFACE TEMPERATURE (SST), AND
WHITE MUSCLE TEMPERATURE (TWM) FOR COMMON THRESHER SHARKS CAPTURED OFF SOUTHERN CALIFORNIA.
Shark ID (sex)Total length
(cm) TRM TRM 2 SST TRM 2 TWM
aLongline captured1 (F) 269 19.0 20.2 3.12 (F) 210 19.7 0.5 1.33 (F) 215 20.0 0.7 0.74 (F) 226 20.0 0.7 0.75 (F) 134 20.2 0.9 0.96 (F) 240 20.1 0.9 0.97 (M) 195 19.7 1.3 1.38 (M) 173 20.1 1.7 0.59 (M) 230 21.3 2.0 2
10 (M) 144 21.8 2.6 1.711 (F) 245 21.8 2.6 2.012 (M) 231 22.1 2.9 1.613 (F) 208 22.3 3.1 0.714 (M) 197 21.6 3.2 1.915 (M) 270 21.6 3.2 2.016 (M) 171 23.0 3.7 3.317 (M) 230 23.8 5.4 7.7
Sportfish captured18 (F) btime 5 7 240 24.1 0.7 1.319 (F) btime 5 11 208 20.5 1.5 0.820 (F) btime 5 40 250 20.5 2.5 2.521 (F) btime 5 27 300 23.6 3.6 3.622 (F) btime 5 120 354 22.2 4.0 4.023 (F) btime 5 60 382 22.8 4.0 4.024 (F) btime 5 60 220 22.5 4.5 4.5
a Longline capture struggle time from 15 to 240 min.b Capture struggle time (min).d White muscle temperature (TWM).
Fig. 2. Red muscle temperature of 24 CommonThresher Sharks. Open circles: sharks captured bylongline. Filled circles sharks captured by rod andreel.
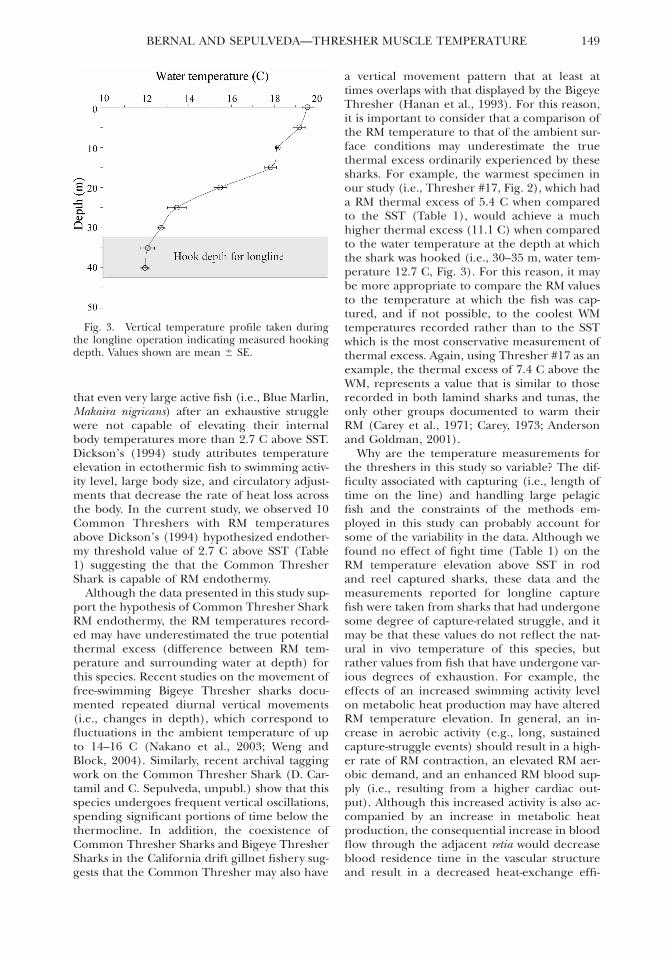
the rod and reel operations and 18.9 6 0.09 Cduring the longline operations. In addition, thevertical thermal profile (i.e., surface to 40 m)data collected during the longline operationsdocuments the thermocline depth at approxi-mately 25 m and an associated decrease in thewater temperature to 12.70 6 0.19 C at 30 m(Fig. 3).
DISCUSSION
The present study tested the hypothesis thatA. vulpinus is capable of significantly elevatingthe temperature of its aerobic red myotomalmusculature (RM) above that of the sea surfacetemperature (SST). Temperature measure-ments of the RM, white muscle (WM), and am-bient water for 24 live sharks show that A. vul-pinus is capable of elevating its RM temperaturesignificantly above that of SST.
How warm does a fish need to be in order tobe considered endothermic? A review of fishmuscle temperatures by Dickson (1994) noted
149BERNAL AND SEPULVEDA—THRESHER MUSCLE TEMPERATURE
Fig. 3. Vertical temperature profile taken duringthe longline operation indicating measured hookingdepth. Values shown are mean 6 SE.
that even very large active fish (i.e., Blue Marlin,Makaira nigricans) after an exhaustive strugglewere not capable of elevating their internalbody temperatures more than 2.7 C above SST.Dickson’s (1994) study attributes temperatureelevation in ectothermic fish to swimming activ-ity level, large body size, and circulatory adjust-ments that decrease the rate of heat loss acrossthe body. In the current study, we observed 10Common Threshers with RM temperaturesabove Dickson’s (1994) hypothesized endother-my threshold value of 2.7 C above SST (Table1) suggesting the that the Common ThresherShark is capable of RM endothermy.
Although the data presented in this study sup-port the hypothesis of Common Thresher SharkRM endothermy, the RM temperatures record-ed may have underestimated the true potentialthermal excess (difference between RM tem-perature and surrounding water at depth) forthis species. Recent studies on the movement offree-swimming Bigeye Thresher sharks docu-mented repeated diurnal vertical movements(i.e., changes in depth), which correspond tofluctuations in the ambient temperature of upto 14–16 C (Nakano et al., 2003; Weng andBlock, 2004). Similarly, recent archival taggingwork on the Common Thresher Shark (D. Car-tamil and C. Sepulveda, unpubl.) show that thisspecies undergoes frequent vertical oscillations,spending significant portions of time below thethermocline. In addition, the coexistence ofCommon Thresher Sharks and Bigeye ThresherSharks in the California drift gillnet fishery sug-gests that the Common Thresher may also have
a vertical movement pattern that at least attimes overlaps with that displayed by the BigeyeThresher (Hanan et al., 1993). For this reason,it is important to consider that a comparison ofthe RM temperature to that of the ambient sur-face conditions may underestimate the truethermal excess ordinarily experienced by thesesharks. For example, the warmest specimen inour study (i.e., Thresher #17, Fig. 2), which hada RM thermal excess of 5.4 C when comparedto the SST (Table 1), would achieve a muchhigher thermal excess (11.1 C) when comparedto the water temperature at the depth at whichthe shark was hooked (i.e., 30–35 m, water tem-perature 12.7 C, Fig. 3). For this reason, it maybe more appropriate to compare the RM valuesto the temperature at which the fish was cap-tured, and if not possible, to the coolest WMtemperatures recorded rather than to the SSTwhich is the most conservative measurement ofthermal excess. Again, using Thresher #17 as anexample, the thermal excess of 7.4 C above theWM, represents a value that is similar to thoserecorded in both lamind sharks and tunas, theonly other groups documented to warm theirRM (Carey et al., 1971; Carey, 1973; Andersonand Goldman, 2001).
Why are the temperature measurements forthe threshers in this study so variable? The dif-ficulty associated with capturing (i.e., length oftime on the line) and handling large pelagicfish and the constraints of the methods em-ployed in this study can probably account forsome of the variability in the data. Although wefound no effect of fight time (Table 1) on theRM temperature elevation above SST in rodand reel captured sharks, these data and themeasurements reported for longline capturefish were taken from sharks that had undergonesome degree of capture-related struggle, and itmay be that these values do not reflect the nat-ural in vivo temperature of this species, butrather values from fish that have undergone var-ious degrees of exhaustion. For example, theeffects of an increased swimming activity levelon metabolic heat production may have alteredRM temperature elevation. In general, an in-crease in aerobic activity (e.g., long, sustainedcapture-struggle events) should result in a high-er rate of RM contraction, an elevated RM aer-obic demand, and an enhanced RM blood sup-ply (i.e., resulting from a higher cardiac out-put). Although this increased activity is also ac-companied by an increase in metabolic heatproduction, the consequential increase in bloodflow through the adjacent retia would decreaseblood residence time in the vascular structureand result in a decreased heat-exchange effi-

150 COPEIA, 2005, NO. 1
ciency because heat flux (i.e., heat exchange) isinversely correlated to the rate of blood flow(Graham, 1983; Brill et al., 1994). Hence, theaerobic demand forced upon a struggling fish(capture struggle) may translate into a decreasein RM temperature elevation, further suggest-ing that the temperature data presented in thisstudy (all of which underwent some degree ofcapture-related struggle) may be lower than val-ues for nonstressed fish. This hypothesis is sup-ported by Carey at al. (1981) and Goldman(1997) who found that lamnid sharks that un-derwent the longest capture-related struggle ex-hibited the lowest visceral thermal excess (theviscera of lamnids is also served by a heat-ex-changing retia). Resolving questions related toRM temperature elevation and swimming activ-ity level requires future studies to focus on col-lecting RM temperature data from free-swim-ming sharks that undergo natural diving move-ments.
Although this work documents that the Com-mon Thresher Shark is capable of RM endo-thermy, the degree to which the temperature ismaintained and the possibility of physiologicaland behavioral thermoregulation remain un-known. In addition, the assumption that allmembers of the Alopiidae are capable of RMendothermy may be premature. Little is knownabout the other two thresher species (Alopias su-perciliosus and Alopias pelagicus) and although re-cent work has focused on the movements offree-swimming Bigeye Thresher Sharks (Nakanoet al., 2003; Weng and Block, 2004), there is noinformation on their RM morphology and thepresence or absence of a heat exchange system(retia) serving the RM. This precludes the gen-eralization that RM endothermy is a synapo-morphy of the Family Alopiidae and warrantsfuture investigations aimed at describing themuscle morphology, vascular anatomy, and en-dothermic capacity of all three thresher sharkspecies. Understanding the physiological andbehavioral differences among the three thresh-er shark species may provide valuable informa-tion regarding their geographic distribution,habitat use, and possible niche overlap.
ACKNOWLEDGMENTS
We would like to thank J. Graham for review-ing early versions of this manuscript and threeanonymous reviewers for their helpful com-ments. We express our deepest appreciation toC. Chan, J. Donley, H. Dowis, J. Lyon, and N.Sepulveda for their assistance in specimen col-lection and sea-going temperature measure-ments. We thank the captain and crew of the
R/V STARR JORDAN, R/V SPLUNKY I, and D.Holts and D. Ramon from NMFS-SWFSC for as-sistance during the longline operations. The au-thors give special recognition to J. Valdez andthe crew of the F/V KOREAN LIMITS for muchlogistical support during this project. Sharkswere collected under CDFG permit 803054-04,and the study was approved by UCSD IACUCprotocol 106509 and by the WSU ACUC.
LITERATURE CITED
ANDERSON, S. A., AND K. J. GOLDMAN. 2001. Temper-ature measurements from Salmon Sharks, Lamnaditropis, in Alaskan waters. Copeia 2001:794–796.
BERNAL, D., C. SEPULVEDA, O. MATHIEU-COSTELLO,AND J. B. GRAHAM. 2003. Comparative studies ofhigh performance swimming in sharks. I. Red mus-cle morphometrics, vascularization, and ultrastruc-ture. J. Exp. Biol. 206:2831–2843.
BONE, Q., AND A. D. CHUBB. 1983. The retial systemof the locomotor muscle in the thresher shark. J.Mar. Biol. Assoc. U.K. 63:239–241.
BRILL, R. W., H. DEWAR, AND J. B. GRAHAM. 1994. Basicconcepts relevant to heat transfer in fishes andtheir use in measuring physiological thermoregu-latory abilities in tuna. Environ. Biol. Fish. 40:109–124.
CAREY, F. G. 1973. Fishes with warm bodies. Sci. Am.228:36–44.
———, AND J. M. TEAL. 1969. Mako and porbeagle:warm bodied sharks. Comp. Biochem. Physiol. 28:199–204.
———, J. M. TEAL, J. W. KANWISHER, AND K. D. LAW-SON. 1971. Warm bodied fish. Am. Zool. 11:135–145.
———, ———, AND ———. 1981. The visceral tem-peratures of Mackerel Sharks (Lamnidae). Physiol.Zool. 54:334–344.
———, J. G. CASEY, H. L. PRATT, D. URQUHART, AND
J. E. MCCOSKER. 1985. Temperature, heat produc-tion and heat exchange in lamnid sharks. Mem. So.Cal. Acad. Sci. 9:92–108.
COMPAGNO, L. J. V. 2001. Bullhead, Mackerel, andCarpet Sharks (Heterodontiformes, Lamniformesand Orectolobiformes), p. 78–88. In: Sharks of theworld. An annotated and illustrated catalogue ofshark species known to date. FAO Species Cata-logue for Fishery Purposes. No. 1, Vol. 2,FAO,Rome.
DICKSON, K. A. 1994. Tunas as small as 207 mm forklength can elevate muscle temperatures significant-ly above ambient water temperature. J. Exp. Biol.190:79–93.
GOLDMAN, K. J. 1997. Regulation of body temperaturein the White Shark, Carcharodon carcharias. J. Comp.Physiol. 167:423–429.
GRAHAM, J. B. 1983. Heat transfer, p. 248–278. In: Fishbiomechanics. P. W. Webb and D. Weihs (eds.).Praeger, New York.
HANAN, D. A., D. B. HOLTS, AND A. L. COAN JR. 1993.The California drift gill net fishery for sharks and

151BERNAL AND SEPULVEDA—THRESHER MUSCLE TEMPERATURE
swordfish, 1981–82 through 1990–91. Fish. Bull.175:1–90.
HART, J. L. 1973. Pacific fishes of Canada. Fish. Res.Bd. Can. Bull. 180:1–740.
NAKANO, H., H. MATSUNAGA, H. OKAMOTO, AND M.OKAZAKI. 2003. Acoustic tracking of Bigeye Thresh-er Shark Alopias superciliosus in the eastern PacificOcean. Mar. Ecol. Prog. Ser. 265:255–261.
O’BRIEN, J. W., AND J. S. SUNADA. 1994. A review ofthe southern California experimental drift longlinefishery for sharks, 1988–1991. CalCOFI Rep. 35:222–229.
WENG, K. C., AND B. A. BLOCK. 2004. Diel vertical mi-
gration of the Bigeye Thresher Shark (Alopias su-perciliosus), a species possessing orbital retia mira-bilia. Fish. Bull. 102:221–229.
(DB) DEPARTMENT OF ZOOLOGY, WEBER STATE
UNIVERSITY, OGDEN, UTAH 84408; AND (CS)CENTER FOR MARINE BIOTECHNOLOGY AND
BIOMEDICINE, SCRIPPS INSTITUTION OF OCEAN-OGRAPHY, LA JOLLA, CALIFORNIA 92093-0204.E-mail (DB) [email protected]. Send re-print requests to DB. Submitted: 21 June2004. Accepted: 4 Oct. 2004. Section editor:S. J. Beaupre.




















