
Evidence for a Cysteine-Histidine Thioether Bridge in Functional Units of Molluscan Haemocyanins and...
10
Eur. J. Biochem. 248, 879-888 (1997) 0 FEBS 1997 Evidence for a cysteine-histidine thioether bridge in functional units of molluscan haemocyanins and location of the disulfide bridges in functional units d and g of the P,-haemocyanin of Helix pomatia Constant GIELENS I, Natalie DE GEEST’, Xue-Qin XIN’, Bart DEVREESE’, Jozef VAN BEEUMEN’ and Cis& PREAUX’ I Laboratorium voor Biochemie, Katholieke Universiteit Leuven, Leuven, Belgium Laboratorium voor Eiwitbiochemie en Eiwitengineering, Universiteit Gent, Gent, Belgium (Received 15 May 1997) - EJB 97 0688/3 In functional units d and g from the P,-haemocyanin of the gastropod Helix pornatia, aminoacid analysis in the presence of dimethyl sulfoxide showed the occurrence of seven cysteine residues. Titration with 5,5‘-dithiobis(2-nitrobenzoic acid), however, did not reveal any free thiol group. Pepsinolysis at pH 2.0 followed by amino acid analysis and partial sequencing of the cysteine-containing peptides showed that six cysteine residues are involved in the formation of three disulfide bridges at corresponding posi- tions. The results indicated that the remaining cysteine residue is linked by a thioether bridge to a histidine residue located two positions further in the sequence (H. pomatia d m i s 6 2 ; H. pomatiu g mHis68). This residue corresponds to one of the three histidine residues considered to be involved in the coordination of the copper A atom of the dinuclear copper group of the functional units. The presence of the thioether bond was further evidenced by an absorption band at 255 nm and by molecular mass determinations with electrospray mass spectrometry on a peptic peptide from H. pomatiu d and on peptides obtained by proteolysis of carboxymethylated H. pomutiu d with trypsin and pronase. Upon sequence analysis the cysteine-histidine thioether bridge was cleaved into didehydroalanine (poly- mers) and 2-thiolhistidine. A peptide with a cysteine-histidine thioether bridge was isolated from pepsinolysates of functional units c, e, ,f, g and h of Sepia officinalis and unit g of Octopus vulgaris, obtained from haemocyanin of the cephalopods S. ofiicinalis and 0. vulgaris. A cysteine-histidine thioether bridge, which was reported previously for tyrosinase of Neumsporu crassa, thus seems to be a general feature for functional units of molluscan haemocyanins. Keywords: haemocyanin ; functional unit; thioether bridge ; disulfide bridge; electrospray mass spec- trometry. The &haeinocyanin is one of the three copper-containing dioxygen-binding glycoproteins found freely dissolved in the haemolymph of the gastropodan mollusc Helix pomatia [I]. It differs from the two a components (a,,- and a,-haemocyanin) by its ability to precipitate or crystallize during dialysis against sodium acetate, pH 5.3, at low ionic strength (10 mM), and by its subunit composition, consisting of only one type of polypep- tide chain compared with two types in each of the two a-haemo- cyanins [2]. In the gastropod haemocyanin molecules (Mr 9 000 000) the subunits (Mr 450000) are associated into dimers. Five of these dimers form a hollow cylinder with a collar at one side, and two such decamers are further associated into a didecamer [I, 3, 41. The subunits themselves are comprised of eight covalently linked globular entities, as deduced from electron microscopy Correspondence fo G. Preaux, Laboratorium voor Biochemie, Celes- tijnenlaan 200G, B-3001 Leuven-Heverlee, Belgium Abbreviations. CM, carboxymethyl; DabNCS, 44,N-dimethyl- aminoazobenzene 4’-isothiocyanate; Dabth, 4-N,N-dimethylaminoazo- benzene 4’-thiohydantoin ; E/S, enzymehubstrate ratio ; Me,SO, di- methyl sulfoxide; Nbs,, 5,5’-dithiobis(2-nitrobenzoic acid) ; PE, pyridyl- ethyl; PhNCS, phenylisothiocyanate; TosPheCh,CI, I.-I -tosylamido-2- phenylalanyl chloromethdne. Enzymes. Endoproteinase Glu-C (EC 3.4.21.19); pepsin (EC 3.4.23.1); subtilisin (EC 3.4.21.62); trypsin (EC 3.4.21.4). [5] and, more accurately, from limited-proteolysis experiments [6]. The latter method, moreover, allowed the liberation and the isolation of each of the eight globular parts from a molluscan haemocyanin subunit [7]. These entities, called a-h from the N to the C terminus of the subunit, are also referred to as func- tional units as they each contain two copper ions able to bind one dioxygen molecule [8]. The haemocyanins of the Decubruchiu cephalopod Sepia ojl ,ficinalis and the Octabrachia cephalopod Octopus vulgaris seem to consist of only one type of component. The molecules, with M, of 3900000 [9] and 3500000, respectively 1101 have cylin- drical decameric shapes [3, 11, 121. Their subunits, according to the cDNA sequencing data of the S. ofSicinaZi.s haemocyanin (De Geest, N., unpublished results) and of the haemocyanin of Octo- pus dofleini [13], appear to be of two types. The subunits of the S. oficinalis haemocyanin [9] are, like those of the H. pornatia &haemocyanin [6, 71, built up of eight covalently linked func- tional units, those of 0. vulgaris [lo] and 0. dofleini haemocya- nin [14] of only seven. The primary structures of the functional units d [15] and g [16] of H. pomatia /&haemocyanin and of part of functional unit f of S. ofSicinalis haemocyanin 117, 181 were determined through amino acid sequencing, while those of S. oficinalis c, e (De Geest, N., unpublished results), h and part of g [19], and of 0. dofleini e-g [13], and parts of 6, c and d [20] were deter-
-
Upload
universiteitgent -
Category
Documents
-
view
4 -
download
0
Transcript of Evidence for a Cysteine-Histidine Thioether Bridge in Functional Units of Molluscan Haemocyanins and...

Eur. J. Biochem. 248, 879-888 (1997) 0 FEBS 1997
Evidence for a cysteine-histidine thioether bridge in functional units of molluscan haemocyanins and location of the disulfide bridges in functional units d and g of the P,-haemocyanin of Helix pomatia Constant GIELENS I, Natalie DE GEEST’, Xue-Qin XIN’, Bart DEVREESE’, Jozef VAN BEEUMEN’ and Cis& PREAUX’ I Laboratorium voor Biochemie, Katholieke Universiteit Leuven, Leuven, Belgium
Laboratorium voor Eiwitbiochemie en Eiwitengineering, Universiteit Gent, Gent, Belgium
(Received 15 May 1997) - EJB 97 0688/3
In functional units d and g from the P,-haemocyanin of the gastropod Helix pornatia, aminoacid analysis in the presence of dimethyl sulfoxide showed the occurrence of seven cysteine residues. Titration with 5,5‘-dithiobis(2-nitrobenzoic acid), however, did not reveal any free thiol group. Pepsinolysis at pH 2.0 followed by amino acid analysis and partial sequencing of the cysteine-containing peptides showed that six cysteine residues are involved in the formation of three disulfide bridges at corresponding posi- tions. The results indicated that the remaining cysteine residue is linked by a thioether bridge to a histidine residue located two positions further in the sequence (H. pomatia d m i s 6 2 ; H. pomatiu g m H i s 6 8 ) . This residue corresponds to one of the three histidine residues considered to be involved in the coordination of the copper A atom of the dinuclear copper group of the functional units.
The presence of the thioether bond was further evidenced by an absorption band at 255 nm and by molecular mass determinations with electrospray mass spectrometry on a peptic peptide from H. pomatiu d and on peptides obtained by proteolysis of carboxymethylated H. pomutiu d with trypsin and pronase. Upon sequence analysis the cysteine-histidine thioether bridge was cleaved into didehydroalanine (poly- mers) and 2-thiolhistidine.
A peptide with a cysteine-histidine thioether bridge was isolated from pepsinolysates of functional units c, e, ,f, g and h of Sepia officinalis and unit g of Octopus vulgaris, obtained from haemocyanin of the cephalopods S. ofiicinalis and 0. vulgaris. A cysteine-histidine thioether bridge, which was reported previously for tyrosinase of Neumsporu crassa, thus seems to be a general feature for functional units of molluscan haemocyanins.
Keywords: haemocyanin ; functional unit; thioether bridge ; disulfide bridge; electrospray mass spec- trometry.
The &haeinocyanin is one of the three copper-containing dioxygen-binding glycoproteins found freely dissolved in the haemolymph of the gastropodan mollusc Helix pomatia [I]. It differs from the two a components (a,,- and a,-haemocyanin) by its ability to precipitate or crystallize during dialysis against sodium acetate, pH 5.3, at low ionic strength (10 mM), and by its subunit composition, consisting of only one type of polypep- tide chain compared with two types in each of the two a-haemo- cyanins [2].
In the gastropod haemocyanin molecules (Mr 9 000 000) the subunits (Mr 450000) are associated into dimers. Five of these dimers form a hollow cylinder with a collar at one side, and two such decamers are further associated into a didecamer [ I , 3, 41.
The subunits themselves are comprised of eight covalently linked globular entities, as deduced from electron microscopy
Correspondence fo G. Preaux, Laboratorium voor Biochemie, Celes- tijnenlaan 200G, B-3001 Leuven-Heverlee, Belgium
Abbreviations. CM, carboxymethyl; DabNCS, 44,N-dimethyl- aminoazobenzene 4’-isothiocyanate; Dabth, 4-N,N-dimethylaminoazo- benzene 4’-thiohydantoin ; E/S, enzymehubstrate ratio ; Me,SO, di- methyl sulfoxide; Nbs,, 5,5’-dithiobis(2-nitrobenzoic acid) ; PE, pyridyl- ethyl; PhNCS, phenylisothiocyanate; TosPheCh,CI, I.-I -tosylamido-2- phenylalanyl chloromethdne.
Enzymes. Endoproteinase Glu-C (EC 3.4.21.19); pepsin (EC 3.4.23.1); subtilisin (EC 3.4.21.62); trypsin (EC 3.4.21.4).
[5] and, more accurately, from limited-proteolysis experiments [6]. The latter method, moreover, allowed the liberation and the isolation of each of the eight globular parts from a molluscan haemocyanin subunit [7]. These entities, called a-h from the N to the C terminus of the subunit, are also referred to as func- tional units as they each contain two copper ions able to bind one dioxygen molecule [8].
The haemocyanins of the Decubruchiu cephalopod Sepia ojl ,ficinalis and the Octabrachia cephalopod Octopus vulgaris seem to consist of only one type of component. The molecules, with M , of 3900000 [9] and 3500000, respectively 1101 have cylin- drical decameric shapes [3, 11, 121. Their subunits, according to the cDNA sequencing data of the S. ofSicinaZi.s haemocyanin (De Geest, N., unpublished results) and of the haemocyanin of Octo- pus dofleini [13], appear to be of two types. The subunits of the S. oficinalis haemocyanin [9] are, like those of the H. pornatia &haemocyanin [6, 71, built up of eight covalently linked func- tional units, those of 0. vulgaris [ lo] and 0. dofleini haemocya- nin [14] of only seven.
The primary structures of the functional units d [15] and g [16] of H. pomatia /&haemocyanin and of part of functional unit f of S. ofSicinalis haemocyanin 117, 181 were determined through amino acid sequencing, while those of S. oficinalis c, e (De Geest, N., unpublished results), h and part of g [19], and of 0. dofleini e-g [13], and parts of 6, c and d [20] were deter-

880 Gielens et al. (Eur: J . Biochem. 248)
mined via cDNA sequencing. The carbohydrate contents of haemocyanins [21] and of functional units [22] were determined. For H. ponzatia d [23] and g [24] the carbohydrate chains were localized, and heterogeneity was demonstrated at several sites.
Comparison of the sequences obtained via direct protein chemistry with those deduced via cDNA technology showed that the five cysteine residues in H. yomatia d (positions S O , 59, 232, 321 and 332) are conserved in the other functional units, but the cysteine residue found at position 166 in H. pomatia g is absent. The comparison of cDNA sequences, however, revealed the presence of another conserved cysteine residue, at a position (Cys60) immediately following the second conserved cysteine residue, and of a histidine residue two positions further on (His62). In the sequences derived by amino-acid-sequence analysis a conserved residue was identified at the first of these two positions but it was a serine instead of a cysteine residue
The peculiar difference in residue identity, seemingly being a function of the method of sequence determination, prompted us, during a study of the localization of the disulfide groups in H. pomatia d and g, to reinvestigate the residues at those two positions.
Notes on part of this work have appeared in abstract form (26, 271. In this paper, additional evidence, including electro- spray-mass-spectrometry data, is provided for the existence of an unusual cysteine-histidine thioether bridge in H. pomatiu d and g. Moreover, the study was extended to functional units c, e, .f; g and h of S. ofiicinali.7 haemocyanin and to functional unit g of 0. vulgaris haemocyanin to determine whether the non- peptidic cysteine-histidine linkage may be considered as a gene- ral feature for molluscan haemocyanin functional units.
~ 5 1 .
MATERIALS AND METHODS Reagents. SDS (specially purified for electrophoresis), phe-
nol (special for chromatography), trisodiuin citrate and EDTA were from BDH. Ammonium acetate, disodium hydrogen phos- phate, sodium dihydrogen phosphate, dimethyl sulfoxide (Me,SO), guanidinium chloride and potassium cyanide were ob- tained from Merck. Methane sulfonic acid and 5,5’-dithiobis(2- nitrobenzoic acid) (Nbs,) were from Aldrich. 2-Mercaptoethanol was from Merck-Schuchardt, 1,4-dithiothreitol from Boehringer, iodoacetic acid from Fluka, and 4-vinylpyridine from Sigma.
Acetic acid, ammonium hydrogen carbonate, formic acid, hydrogen chloride and sulfuric acid were purchased from UCB.
4-N,N-Dimethylaminoazobenzene 4’-isothiocyanate (DabNCS) was from Pierce. Phenylisothiocyanate (PhNCS ; sequencer grade), tritluoroacetic acid (sequencer grade), and acetonitrile (HPLC grade) were obtained from Rathburn. L-2-Thiolhistidine was from USB.
Reagents for automated sequence analysis were obtained from the supplier of the instrument.
Protein aiid peptide preparations. Bovine trypsin, treated with L- 1 -tosyIamido-2-phenylalanyl chloromethane (TosPhe- Ch,CI), was purchased from Sigma. Pepsin and endoproteinase Glu-C were from Boehringer. Pronase was obtained from Cal- biochem, subtilisin (type Carlsberg) from Novo.
&Haemucyanin of H. pomatia was isolated according to [81 from an ammonium sulfate precipitate obtained from the haemo- lymph, collected and treated as described [28]. Functional units d and g were liberated from the dimers of subunits by limited proteolysis with TosPheCh,CI-treated trypsin and isolated by ion-exchange chromatography on DEAE-Sepharose as reported
The apoderivatives were prepared by means of KCN [30] for the removal of the copper ions. To about 100 mg protein (H .
1291.
pomutia d or g), solubilized in 2.8 ml 0.1 M NH,HCO,, about 1.5 ml of 1 M KCN, brought to pH 8.5 with aceti’c acid, was added dropwise until the protein was decoloured. The solution was dialysed overnight against 50 mM KCN, made 10 mM in EDTA, brought to pH 8.5 with acetic acid, and dialysed three times overnight against 0.1 M NH,HCO, before being lyophi- lized.
The reduction and carboxymethylation of H. pomatia d [yielding d (CM)] were performed according to [31] with slight modifications. To about 90 mg d, solubilized in 5.0 ml 0.1 M Tris/HCI, pH 8.5, 6 M guanidinium chloride, and 3 mM EDTA, dithiothreitol was added to 5 mM. The solution was flushed with nitrogen and kept for 1 h at 37°C. 0.5 M 1CH2C001H, adjusted to pH 7.0 with NaOH, was added to 15 mM. The solution was again flushed with nitrogen and kept for 1 h at 37°C. After the addition of S O p1 2-mercaptoethanol to block the excess of ICHZCOOH, the solution was dialysed extensively against 50 mM NH,HCO,.
The haemocyanin of S. oficinalis was isolated as described [32], except that the preparative ultracentrifugation performed to sediment the haemocyanin molecules was not preceded by chromatography on Ultrogel A4. Fragment a-f was isolated by limited proteolysis according to [9]. Limited trypsinolysis of a-5 followed by chromatography on Ultrogel AcA34, allowed the isolation of functional unit f in addition to fragments a-c and de [9]. Functional unit c was liberated from fragment u-c (20 mg/ml) by limited treatment with subtilisin at an enzyme/ substrate (E/S) ratio of 1 :300 (by mass) for 24 h at 25°C in 0.1 M NH,HCO,, 0.02% NaN,, and separated by chromatogra- phy on a column (150 cmX7 cm) of Ultrogel AcA34. Functional unit e was obtained from fragment de as described [33]. Func- tional units g and h were isolated according to [34].
The haemocyanin of 0. vulgaris and its functional unit g were prepared as reported [lo, 121.
Pepsinolysis of the functional units and of the apoderivatives of H. pomatiu d and g was carried out for 7 h at 20°C at an E/ S of 1 :40 (by mass) on 50- 100 mg protein solubilized in 5 ml 0.2 M formic acid, pH 2.0. The trypsinolysis of d(CM) (15 mg/ ml in 50 mM NH,HCO,) was performed for 8 h at 37°C at an E/S of 1 :35 (by mass).
The fractionation on Sephadex G-25 (Pharmacia) of the pep- tic digests of the functional units was performed on a column (170 cmX2.4 cm) equilibrated with 0.5 % acetic acid (flow rate 19 ml/h, fractions of 2 ml), that of the tryptic hydrolysate of d(CM) on a column (190 cmXl .8 cm) equilibrated with SO mM NH,HCO, (flow rate 18.2 ml/h, fractions of 2 ml).
The further fractionations by reverse-phase HPLC were car- ried out with a SP8700 or SP8800 HPLC system (Spectra-Phys- ics). The column (25 cmX4.6 mm) was a Zorbax Protein Plus or a Hypersil 5 ODS, both obtained from Chrompack (Middel- burg). For the elution (flow rate 0.8 ml/min), a linear gr a d’ lent of acetonitrile, from 0 to 6 4 % (by vol.) over 60 nilin i n 0.1 % (by vol.) aqueous trifluoroacetic acid or in 50 mM ammonium acetate (for fractions H. pomatia gII and gIII) was applied. The eluate was monitored at 220 nm and/or at 255 nm with a vari- able-wavelength absorbance detector (V4, ISCO ; or Spectra 200, Spectra-Physics). The fractions were dried by vacuum cen- trifugation (Speed Vac concentrator, Savant).
The reduction of peptides H. pomatia d1-5 and ,gI-3, and S. o~iicinalisfl1-1, and the carboxymethylation of reduced gl-3 and JI-1 to gI-3(CM) andJ1-l(CM), were carried out as for H. po- rnatia d but on smaller amounts of material (50-150 nmol dried peptide). The pyridylethylation of peptide H. ponmtia dII-5, yielding dII-S(PE), was performed by overnight incubation at room temperature with 1.5 mM 4-vinylpyridine[351. The car-

Gielens et al. (Eur: J . Biochern. 248) 88 1
boxymethylated and pyridylethylated peptides were recovered by HPLC and dried on a vacuum centrifuge.
The treatment of peptides H. pomatia dI-S(PE) and S. offici- nalisfI1-1 (CM) with TosPheCh,Cl-treated trypsin and of peptide H. pomatia gI-3(CM) with endoproteinase Glu-C was carried out at a peptide concentration of about 3 pg/pl in 0.1 M NH,HCO, for 2 h at 37°C and at an E/S of 1 :25 (by mass). The proteolysis of H. pomatia dV-I(CM) with pronase was per- formed on 50 nmol peptide, solubilized in 300 pl 0.1 M NH,HCO,, for 90 h at 37 "C and at an E/S of 1 : 4 (by mass).
Analyses. The purity of the samples was checked by analyti- cal ultracentrifugation in a Spinco Model E analytical ultra- centrifuge (Beckman), by SDS/PAGE on 7.5% or 10% poly- acrylamide gels according to the method of Laemmli [36] after treatment in the presence of 2-mercaptoethanol, and by crossed immunoelectrophoresis [37] against antisera prepared in rabbits [38].
The concentration of the haemocyanins and of the functional units was determined spectrophotometrically at 278 nm and 20°C in 50mM borax by means of the following specific ab- sorption coefficients : (H . pomatia &haemocyanin, 1.416 1 . g 1 . cm-1 [28]; H. pomatia apod, 1.2221 . g-' . cm-' [ 2 2 ] ; H. pornatia apog, 1.242 1 . g-' . cm-' [ 2 2 ] ; 0. vulgaris haemocy- anin, 1.45 1 . gg' . cm-' [lo]; 0. vulgaris g, 1.45 1 . g' . cm-' [lo]. The values for S. ojfiicinalis haemocyanin (1.35), functional units c (1.33), e (1.28),f(1.44) and gh (1.31) were deduced in this work by spectrophotometry coupled to differential refrac- tometry at 20°C with a Rayleigh Interference Refractometer M447 (Hilger and Watts) on the basis of a specific refraction increment dnldc of 1.944X lo-' dl/g at 546 nm [22]. The absorp- tion coefficients for S. oflcinalis g and h were assumed to be the same as that for gh. The ultraviolet absorption spectra of the thioether-containing peptides were recorded with a Lambda 14 spectrophotometer (Perkin-Elmer).
The presence of glycopeptides in the Sephadex fractions was examined with the carbohydrate test according to Dubois et al. [39]. To samples of 0.25 ml, 50 pl 80% phenol (by mass) in water and 4.5 ml sulfuric acid were added. After 30 min at room temperature the absorbance at 485 nm was measured.
The residual copper content of the apoprotein preparations was determined with a 372 atomic absorption spectrophotometer (Perkin-Elmer) at 324.7 nm.
Free thiol groups were estimated by the method of Habeeb [40], making use of the reagent of Ellman [41]. To a series of protein solutions (30nrnol each; =1.5 ml) in 80 mM sodium phosphate, pH 8.0, 2 % SDS, 0.5 % EDTA, increasing volumes of a 1 mM Nbs, solution in 80 mM sodium phosphate, pH 8.0 were added, to molar ratios of NbsJprotein of between 0.2:l and 2:2 . The volume was brought to 2 ml with 80 mM sodium phosphate, pH 8.0. After 2 h the absorbances were measured at 412 nm against the buffer.
The amino acid analyses were carried out on a 4150 Alpha amino acid analyser (LKB), equipped with an Ultropac 11 cation exchanger [42]. The hydrolyses were performed with azeotropic HCl, 0.1 % phenol, in evacuated sealed tubes at l l0"C for 24 h. For the determination of the total cysteine/cystine content the hydrolysis was carried out in the presence of 0.21 M Me,SO [43]. The acid was removed by vacuum centrifugation on a Speed Vac and the residue dissolved in 150 pl trisodium &ate/ HCI, pH 2.2. For the determination of tryptophan the hydrolysis was performed with 4 M methane sulfonic acid (1 % tryptamine) 1441.
The manual sequencing was performed by the DabNCS/ PhNCS double-coupling method [45] followed by identification of the Dabth-amino acids by a two-dimensional TLC. The auto- mated sequence analysis [46] was carried out on a 477A pulsed-
1'50 F a
120 200 280 360 440 Tube number
Fig. 1. Chromatography on Sephadex G-25 of a peptic digest of H. pomutiu functional units d (a) and g (b). The column (170 cmX2.4 cm), developed with 0.5% acetic acid, was loaded with the pepsinolysate of about 100 mg H. pomatia d (after removal of the core dC) or g, and the eluate collected in 2-ml fractions. The peptides were detected by absorbance at 230nm (- , 5-mm cell) and the glycopeptides by the method of Dubois et al. [39] (---. absorbance at 485 nm, I-cm cell). The fractions pooled, as indicated, were those with cysteine-residue-rich peptides.
liquid protein sequencer with on-line analysis of the phenylthio- hydantoin derivatives on a 120A analyser (Perkin-Elmer, Ap- plied Biosystems Division).
Mass analysis was performed on a VG Bio-Q-triple quadru- pole electrospray-ionization instrument (Micromass), using the first quadrupole as the mass analyser. Typically 100 pmol of sample was dissolved in 10 pl MeOH/HOAc/H,O (50: 1 :49, by vol.). The sample was injected into the electrospray source at a flow rate of 4 pl/min, pumped by a 140A solvent delivery pump (Applied Biosystems). The voltage at the capillary tip was set at 4 kV and the sample-cone voltage was 40 V. Scans between masdcharge values from 400 to 1300, each lasting 9 s, were accuinulated over 3 min. Calibration was performed with 20 pmol horse heart myoglobin. This technique is known to have an error of 0.01 -0.05 % [47].
RESULTS
Gastropod haemocyanin. Cysteinekystine content. Analysis of functional units d and g of H. pomati~l by amino acid analysis after hydrolysis in the presence of Me,SO showed seven cyste- ine residues in each unit, although only five and six, respective- ly, were expected from the amino-acid-sequence data [15, 161. Titration of both functional units with Nbs, at pH 8.0 in the pres- ence of SDS did not reveal any thiol group, indicating that none of the cysteine residues occurred freely.
Identification of a thioether bridge and localization of disul- fide bridges in H. pomatia d. Pepsinolysis at pH 2.0 of H. po- matia d, glycosylated at residues 253 and 387 115 231 led to formation of an insoluble core. Chromatography of the soluble part of the hydrolysate on Sephadex G-25 in 0.5% acetic acid yielded several fractions absorbing at 230 nm with the glycopep- tides eluting at the void volume and the cysteine-containing pep-

882 Gielens et al. ( E m J. Binchem. 248)
Table 1. Amino acid composition of cysteine-containing peptides from H. pomatia d. Peptides dII-2, &I-5 and dC-10 are peptic peptides of H. pomatia d, T1 and T2 are the tryptic subfragments of reduced and pyridylethylated H p &I-5 [dlI-S(PE)]. Integral values between parenthe- ses are the number of residues calculated from the sequence of H. PO-
rnatia d 1151. Total cysteinekystine was determined as cysteic acid by hydrolysis in the presence of Me2S0. For dII-5 and T2, some 2-thiolhis- tidine and components eluting between phenylalanine and histidine were observed. n.d., not determined.
Amino acid dll-2 dII-5 dC-I0 d11-5 (PE)
TI T2
Residuedpcptide molecule
cys 2.4 (2) 3.2 (2) 2.0 (1) n.d. n.d. Asp 3.0 (3) 0.4 (0) 4.9 (6) 0.1 (0) 0.0 (0) Thr 0.1 (0) 1.0 (1) 0.1 (0) 0.0 (0) 1.0 ( I ) Ser 1.0 ( I ) 0.2 (2) 0.3 (0) 0.1 (0) 0.1 (2) Glu 0.3 (0) 2.0 (2) 3.3 (3) 2.2 (2) 0.1 (0) Pro 2.2 (2) 2.1 (2) 0.5 (0) 1.2 (1) 1.3 (1) GlY 2.9 (3) 3.9 (4) 0.4 (0) 3.0 (3) 1.1 ( 1 ) Ala 1.1 (1) 1.2 ( I ) 2.6 (3) 0.1 (0) 1.1 (1) Val 2.9 (3) 1.9 (2) 0.1 (0) 0.1 (0) 1.6 (2) Met 0.0 (0) 0.9 (1) 0.0 (0) 0.0 (0) 0.9 ( I ) Ile 0.0 (0) 0.1 (0) 1.0 ( 1 ) 0.0 (0) 0.0 (0) Leu 0.2 (0) 1.1 ( I ) 1.9 (2) 1.0 ( I ) 0.1 (0) S r 0.9 ( I ) 0.1 (0) 0.9 ( 1 ) 0.0 (0) 0.0 (0) Phe 1.0 (1) 1.0 (1) 2.3 (2) 0.0 (0) 1.0 ( I ) His 1.0 (1) 2.8 (3) 0.1 (0) 2.7 (3) 0.1 (0) Lys 3.1 (3) 2.1 (2) 0.1 (0) 1.8 (2) 0.1 (0) Cys(PE) n.d. n.d. n.d. 0.6 ( I ) 0.6 (1) A% 0.1 (0) 0.0 (0) 0.3 (0) 0.0 (0) 0.0 (0)
tides in fraction 11 (Fig. 1 a). This latter fraction was fractionated by reverse-phase HPLC on Zorbax Protein Plus and the cyste- ine-containing peptides were localized by amino acid analyses after hydrolysis in the absence and presence of Me,SO. The core of the pepsinolysate (dC) was solubilized in water by the addi- tion of guanidinium chloride and fractionated by HPLC on a Zorbax Protein Plus column.
Three cysteine-containing fractions were revealed : &I-2 and dII-5, from the soluble part, eluting at 24.0% and 28.5% aceto- nitrile, respectively, and dC-10, from the core, eluting at 31.4% acetonitrile.
Partial amino acid sequencing coupled to amino-acid- composition analyses allowed us to localize the fragments in the total sequence of H. pomatia d [ 151 respectively as residues 319-339 (dI1-2), 44-67 (&I-5) and 157-168 and 230-236 (dC-10).
For peptide &I-2 there was almost perfect agreement be- tween the amino acid analysis and the values expected from the sequence of H. pomutia d (Table 1). The Edman degradation (DabNCS/PhNCS method) indicated that this peptide consisted of two peptides (residues 319-327 and 328-339; Fig. 2), with the cleavage next to Lys327 originating from an internal proteol- ysis of H. pornatia d during the liberation of the functional unit from /&-haemocyanin by trypsin [8]. These data indicate the presence of a disulfide bridge between Cys321 and Cys332.
Amino acid analysis of peptide &I-5 (Table 1 ) did not reveal any serine residue, although two are expected from the sequence [15], but it did show some unusual peaks: a minor peak just after aspartic acid, which eluted with 2-thiolhistidine; and a peak, with some side peaks, in thc region between phenylalanine and histidine. Amino acid analysis after acid hydrolysis in the presence of Me2S0 indicated three cysteine residues; this is one
H . pamatia d
dII-5
44 1- 67 HGKPGLCQHEGHKVACCVHGMPTF
(S S)
dv-1 ( CM )
57 n 72 VACCVHGMPTFPSWHR
A
dC-10
157 (D) 168 YALEQDNFCDFE
I 230 1 236 ADCAINL
dII-2
319 339 WCVPSGPKGKNDCNHKAGVF
H . pomatia g
gI-3
50 I- r, 72 HGSPPGCEHENHSVACCIHGMAN
* (S G)
q11-8
161 167 EQTDYCD
218 I234 QALQKFRGLPYNSANCA
qIII-3
318 328 DLCDSHDHCEF
Fig. 2. Location of the thioether bond and of disulfide bridges in H. pomatia d and g. The peptides were obtained with pepsin, except dV-l(CM) which was liberated with trypsin. The numbers next to their sequence indicate the positions (without deletions or insertions for align- ment) of the residues in the amino acid sequences of H. ponzutia d [15] and g [16]. In parentheses the residues are given from the formerly reported total sequences [15, 161. A, carboxymethylated residue in dV-l(CM); *, the carbohydrate-attachment site [24] in gI-3.
residue more than deduced from the sequence. Peptide dl-5, moreover, strongly absorbed at 255 nm, which is similar to the observations of a peptide of tyrosinase of N. crussu [48, 491. This latter peptide yielded 2-thiolhistidine upon acid hydrolysis and was found to contain a thioether bridge between a cysteine and a histidine residue. The likely presence of the same type of posttranslational modification in 41-5 was further supported by the odd number of cysteine residues found in the peptide itself and in H. pornatia d (3 and 7, respectively) where no free thiol groups had been found. This implies the occurrence of a disul- fide bridge between the two other cysteine residues found in
With the aim to identify the cysteine residue implicated in the thioether bridge and to locate the disulfide bridge, peptide dII-5 was reduced, pyridylethylated, and split at residue 56 with trypsin. The fragments T I and T2 were separated by HPLC on a Zorbax Protein Plus column. Amino acid analysis (Table 1) revealed for peptide T2 (residues 57-67) the same peculiar peaks as observed for dII-5. The sequencing (DabNCWPhNCS method) of the fragments revealed an S-(4-pyridylethyl)cysteine in each of the peptides, i.e. at positions 50 and 59, indicating that these are the two residues involved in the disulfide bridge. At positions 60 and 62 we identified didehydroalanine, usually indicative of a serine residue [45, 501, and 2-thiolhistidine, re- spectively. They should be considered to result from the cleav- age of the cysteine-histidine thioether bridge by the highly acidic conditions used in the Edman degradation.
The proposed structure for peptide dII-5 with a disulfide bridge and a cysteine-histidine thioether bridge (Fig. 2) was con- firmed by electrospray mass spectrometry (Fig. 3 a ; Table 2). The experimental mass of 2569.3 Da is 4.3 Da less than the mass expected from the peptide in the absence of posttranslational modifications (2573.6 Da).
For peptide dC-10, consisting of two covalently linked pep- tides, the amino acid analysis (Table 1) indicated one aspartic
&I-5.

Gielens et al. (Eur: J. Biockern. 248) 883
2569.3
I
230 250 270 290 Wavelength [ nm I
1883.8 Fig. 4. Ultraviolet absorption spectrum of the tetrapeptide Cys(CM)- Cys-Val-His62 from carboxymethylated H. pomutiu d. The peptide (11.4 pM) was dissolved in water and measured in a I-cm cell.
c 1600 1800 2000 2200m/z
Fig.3. Transformed electrospray mass spectrum of &I-5 (a) and 0 - l ( C M ) (b). The peptides were dissolved in 50% methanolll % acetic acid.
Table 2. Theoretical and experimental mass data for cysteine-con- taining peptides with thioether bridges. All theoretical mass values were calculated from the mean mass values for the amino acid residues without posttranslational modifications.
Organism Func- Sequence Theo- Experi- Mass tional position retical mental differ- unit mass mass ence
Da
H . ponzutici d 44 - 61 2573.6 2569.3 4.3 d(CM) 57-72 1885.9.' 1883.8 2.1
59-62 518.6,' 516.3 2.3
e 44 - 69 2739.3 2735.1 4.2 S. officinulis L' 46-70 2687.6 2683.8 3.8
i 45-66 2399.4 2395.6 3.8
Presence of one carboxymethylated cysteine residue assumed.
acid residue less than expected from the sequence of H. pomatia d [I51 and, after hydrolysis in the presence of Me,SO, the pres- ence of a second cysteine residue. The wrongly presumed aspai- tate at position 165 should be replaced by this cysteine residue, as a cysteine residue is always found in other functional units at the equivalent position. This cysteine residue in peptide 157- 168 of dC-10 was inferred to form a disulfide bridge with Cys232 from the other peptide (Fig. 2) and thus explained why the two peptides eluted together upon HPLC.
Thioetlzer peptides isolated from carboxymethylated H. po- matia d. To further confirm the presence of a thioether bridge between residues 60 and 62, H . pomatia functional unit d was reduced and, to avoid interfering absorbances at 255 nm, was carboxymethylated instead of pyridylethylated, and subse- quently treated with trypsin. While the cysteine residues in- volved in the formation of disulfide bridges were expected to be
carboxymethylated, the cysteine residue involved in the thio- ether bridge should be unaffected.
The trypsinolysate of d(CM) was subjected to chromatogra- phy on Sephadex G-25 in 50 mM NH,HCO,. Fraction V, having the highest A,,, nln/A220 nm ratio, indicative for the thioether bond, was subfractionated by HPLC (Hypersil 5 ODS).
Amino acid analyses, after hydrolysis both in the absence and presence of Me,SO, of peptide dV-1(CM) (Fig. 2), eluting at 40.5 % (by vol.) acetonitrile and absorbing mostly at 25.5 nm, indicated that it covered the sequence 57-72 with one of the two cysteine residues being carboxymethylated. 2-Thiolhistidine and the components eluting between phenylalanine and histidine were observed. Automated and manual sequence analysis showed that the cysteine residue at position 59 was carboxy- methylated. At position 60 only didehydroalanine, which is re- presentative for serine, was observed. Automated Edman degra- dation revealed the presence of glutamine at position 62. Since no glutamic acid was found upon amino acid analysis, the increase of the phenylthiohydantoin derivative was assigned to the presence of 2-thiolhistidine. This assignment was confirmed by submitting purified 2-thiolhistidine to one cycle of automated Edman degradation. This conclusion was supported by the man- ual method where Dabth-2-thiolhistidine was identified from its migration on TLC.
Electrospray mass spectrometry of peptide dV-1 (CM) (Fig. 3 b ; Table 2) revealed a mass of 1883.8 Da. The calculated value with a carboxymethylated cysteine residue at position 59 and cysteine and histidine residues at positions 60 and 62, re- spectively, amounted to 1885.9 Da. The observed mass differ- ence of 2.1 Da supports the proposal of the formation of the thioether bridge.
Peptide dV-1(CM) was cleaved with pronase and the hydro- lysate was fractionated by HPLC (Hypersil 5 ODS). A fraction, strongly absorbing at 255 nm and eluting at 14.1 7i (by vol.) acetonitrile, was isolated. Amino acid analysis showed, in addi- tion to S-carboxymethylcysteine and valine, some 2-thiolhisti- dine, and peaks eluting between phenylalanine and histidine. Electrospray mass spectrometry revealed a mass of 516.3 Da (Table 2), indicating a tetrapeptide Cys(CM)-Cys-Val-His62, having a thioether bond.
Fig. 4 shows the ultraviolet absorption spectrum of the tetrapeptide. The molar absorption coefficient at 2.55 nm, de- duced from the concentration determined by amino acid analysis, was 1.5600M~' . cm I . Nearly the same value (IS 800M-' . cm I) was observed for fragment dV-l(CM). These values are very similar to those for 2-thiolhistidine and the tripeptide Cys94-Thr-His96 from tyrosinase of Neurospora cra.s.sa which also contains a cysteine-histidine thioether bond [48, 491.
884 Gielens et al. (Eur: J . Biochem. 248)
Table 3. Amino acid composition of cysteine-containing peptides ob- tained by pepsinolysis of H. pornatia g. Integral values between paren- theses are the residues calculated from the sequence of H. pornatia g (161. For gI-3, some 2-thioihistidine and components eluting between phenylalanine and histidine were observed.
Amino acid gI-3 gII-8 ~111-3
Residuedpeptide molecule
CYS Asp Thr Ser Glu Pro
Ala Val Met Ile Leu TYr Phe His Lys Arg
GlY
2.5 (2) 2.5 (2) 0.0 (0) 2.2 (3) 2.3 (2) 1.9 (2) 3.1 (4) 2.3 (2) 1.0 ( 1 ) 0.9 (1) 0.8 (1) 0.0 (0) 0.0 (0) 0.0 (0) 2.9 (3) 0.0 (0) 0.5 (0)
2.4 (2) 3.9 (4) 1.0 (1) 1.0 (1) 3.9 (4) 1.2 ( 1 ) 1.2 ( I ) 2.7 (3) 0.0 (0) 0.0 (0) 0.0 (0) 1.9 (2) 1.7 (2) 1.1 (1) 0.0 (0) 1.0 (1) 1.0 (1)
2.1 (2) 2.8 (3) 0.3 (0) 1.1 ( 1 ) 1.4 (1) 0.3 (0) 0.0 (0) 0.3 (0) 0.3 (0) 0.0 (0) 0.0 (0) 1.3 (1) 0.0 (0) 1.0 (1) 2.1 (2) 0.0 (0) 0.3 (0)
so c 4 6 11~1 70 HGSTKWCPSPDAAQKYACCHHGMAT
SO e 4 4 IIT, 69 HGLPAQCPNADGTMVHTCCLHGMPTF
so f 45 1- 1-166 HALPPLCPNPTAKHRYACCLHG
( s Q )
so 9 S l y l 69 CPTPDAPQYACCLHGMF'TF
So h 46 1-1 1765 HGMPLSCHYPNGTAFACCQH *
0" 9 4 6 1- ~ 1 6 6 HGIPLSCHXENGTAXACCQHG *
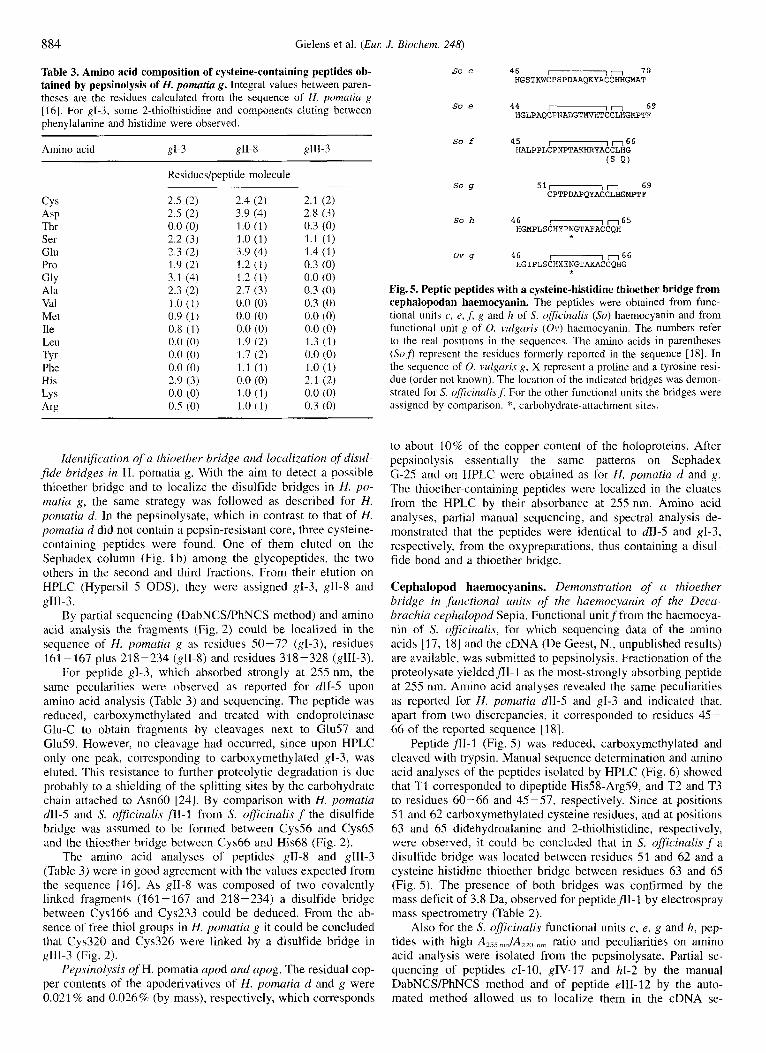
Fig. 5. Peptic peptides with a cysteine-histidine thioether bridge from cephalopodan haemoeyanin. The peptides were obtained from func- tional units c, e, .fi g and h of S. officinalis ( S o ) haemocyanin and from functional unit g of 0. vulgLiris (Ov) haemocyanin. The numbers refer to the real positions in the sequences. The amino acids in parentheses (S0.f) represent the residues formerly reported in the sequence [18]. In the sequence of 0. vulgaris g, X represent a proline and a tyrosine resi- due (order not known). The location of the indicated bridges was demon- strated for S. ofSicina/is,f. For the other functional units the bridges were assigned by comparison. *, carbohydrate-attachment sites.
Identification of a thioether bridge and localizatioiz of disul- fide bridges in H. pomatia g. With the aim to detect a possible thioether bridge and to localize the disulfide bridges in H. po- matia g, the same strategy was followed as described for H. pomatiu d. In the pepsinolysate, which in contrast to that of H. pomatiu d did not contain a pepsin-resistant core, three cysteine- containing peptides were found. One of them eluted on the Sephadex column (Fig. 1 b) among the glycopeptides, the two others in the second and third fractions. From their elution on HPLC (Hypersil 5 ODS), they were assigned gI-3, gII-8 and gI11-3.
By partial sequencing (DabNCS/PhNCS method) and amino acid analysis the fragments (Fig. 2) could be localized in the sequence of H. poinutia g as residues 50-72 (gI-3), residues 161 -167 plus 218-234 (gII-8) and residues 318-328 (gIII-3).
For peptide gl-3, which absorbed strongly at 255 nm, the same pecularities were observed as reported for dl-5 upon amino acid analysis (Table 3) and sequencing. The peptide was reduced, carboxymethylated and treated with endoproteinase Glu-C to obtain fragments by cleavages next to Glu57 and Glu59. However, no cleavage had occurred, since upon HPLC only one peak, corresponding to carboxymethylated gI-3, was eluted. This resistance to further proteolytic degradation is due probably to a shielding of the splitting sites by the carbohydrate chain attached to Am60 [24]. By comparison with H. pomatia &I-5 and S. oflicinalis ,frI-I from S. oficinalis f the disulfide bridge was assumed to be formed between Cys.56 and Cys65 and the thioether bridge between Cys66 and His68 (Fig. 2).
The amino acid analyses of peptides gIJ-8 and gIII-3 (Table 3) were i n good agreement with the values expected from the sequence (161. As gII-8 was composed of two covalently linked fragments (161-167 and 218-234) a disulfide bridge between Cys166 and Cys233 could be deduced. From the ab- sence of free thiol groups in H. pomatia g it could be concluded that Cys320 and Cys326 were linked by a disulfide bridge in gIII-3 (Fig. 2).
Prpsinolysis of H. pomatia apod and apog. The residual cop- per contents of the apoderivatives of H. pomutia d and g were 0.021 % and 0.026% (by mass), respectively, which corresponds
to about 10% of the copper content of the holoproteins. After pepsinolysis essentially the same patterns on Sephadex G-25 and on HPLC were obtained as for H. pomatia d and g. The thioether-containing peptides were localized in the eluates from the HPLC by their absorbance at 255 nm. Amino acid analyses, partial manual sequencing, and spectral analysis de- monstrated that the peptides were identical to dl-5 and gI-3, respectively, from the oxypreparations, thus containing a disul- fide bond and a thioether bridge.
Cephalopod haemocyanins. Demonstration of a thioether bridge in functional units of the huemocyanin of the Decu- hrachia cephalopod Sepia. Functional unitf from the haemocya- nin of S. officinalis, for which sequencing data of the amino acids [17, 181 and the cDNA (De Geest, N., unpublished results) are available, was submitted to pepsinolysis. Fractionation of the proteolysate yielded fll-I as the most-strongly absorbing peptide at 255 nm. Amino acid analyses revealed the same peculiarities as reported for H. pomatia dII-5 and gI-3 and indicated that, apart from two discrepancies, it corresponded to residues 45- 66 of the reported sequence [ 181.
Peptide fr1-1 (Fig. 5 ) was reduced, carboxymethylated and cleaved with trypsin. Manual sequence determination and amino acid analyses of the peptides isolated by HPLC (Fig. 6) showed that T1 corresponded to dipeptide His.58-Arg59, and T2 and T3 to residues 60-66 and 45-57, respectively. Since at positions 51 and 62 carboxymethylated cysteine residues, and at positions 63 and 65 didehydroalanine and 2-thiolhistidine, respectively, were observed, it could be concluded that in S. ?ficinal~s f a disulfide bridge was located between residues 51 and 62 and a cysteine-histidine thioether bridge between residues 63 and 65 (Fig. 5) . The presence of both bridges was confirmed by the mass deficit of 3.8 Da, observed for peptideflI-I by electrospray mass spectrometry (Table 2).
Also for the S. qficinalis functional units c, e, g and h, pep- tides with high A,,, n,JA220 "," ratio and peculiarities on amino acid analysis were isolated from the pepsinolysate. Partial se- quencing of peptides cI-10, gIV-17 and hI-2 by the manual DabNCS/PhNCS method and of peptide eIII-12 by the auto- mated method allowed us to localize them in the cDNA se-

Gielens et al. (Eul: J . Biochern. 248) 885
Fig.6. Fractionation by HPLC on Hypersil 5 ODS of the tryptic hydrolysate of reduced and carboxymethylated peptidefl1-1. Elution with a linear gradient of acetonitrile from 0 to 64% over 60 min in 0.1 % aqueous trifluoroacetic acid. Absorbance at 220 nm (a, full-scale A = 1.0) and at 255 nni (b, full-scale A = 0.2).
quence of the S. oficinalis functional units c, e , g (De Geest, N., unpublished results) and h [I91 as residues 46-70, 44-69, 51 -69 and 46-65, respectively (Fig. 5). Electrospray mass spectrometry performed on cI-10 and eIII-12 (Table 2) revealed mass deficits of 3.8 and 4.2 Da, respectively, indicating that, in addition to the cysteine-histidine thioether bond, there must be a disulfide bridge between the other two cysteine residues in these peptides. By analogy the two brigdes were assumed to exist at the same sites in S. officinalis g and h.
Demonstration of a thioether bridge in functional crnit g of the haemocyanin qf the Octabrachia cephalopod Octopus. To investigate the presence of a cysteine-histidine thioether bridge in the haemocyanin of an octabrachian cephalopod, the same strategy of pepsinolysis, subfractionation and analysis was ap- plied to functional unit g from the haemocyanin of 0. vulgaris. A peptide (g1-l), strongly absorbing at 255 nm and showing upon amino acid analysis peaks characteristic for the presence of a cysteine-histidine thioether bond, was isolated. It eluted in the V,, fraction from the Sephadex G-25 column, i.e. among the glycopeptides. As the sequence of g has not been determined, the peptide was located in the sequence on the basis of the cDNA sequence of functional unit g of the haemocyanin of 0. dojkini [13], a species very related to 0. vulgaris. Amino acid analysis indicated that the peptide from 0. vulgciris g (Fig. 5 ) corresponds to residues 46-66 of 0. dofleini g with one of the two tyrosine residues being substituted by a proline residue.
DISCUSSION
Although an odd number of cysteic acid residues was always deduced after oxidation, no free thiol groups could be titrated in H. pomatia d and in g despite that the experiments were per- formed in the presence of SDS, to detect hidden groups, and of
EDTA, to prevent oxidation by the copper ions that may be lib- erated from the functional units under the denaturing conditions. This indicates that all cysteine side chains must be involved i n bonds. One of these, however, had to be quite peculiar as the detection of this cysteine residue was only possible as cysteic acid and not as a carboxymethylated or pyridylethylated residue, i.e. after oxidation of the functional unit but not after its reduc- tion.
To localize the C'ysCys and Cys Xaa non-peptidic bonds in the known sequence of H. pomatia d [I51 and g [16], and the possible same C T X a a bond in functional units of other mollus- can haemocyanins, pepsin was chosen for the proteolysis to avoid thiol-disulfide exchanges. This allowed us to isolate three Cys-rich peptides from H. pomatia d and g, two of them having a disulfide bridge, and the other consisting of two peptides linked by a disulfide bridge (Fig. 2). These bridges occurred in three distinct parts of the functional units and at corresponding positions in H. pomatia d and g, which indicates that, as in ar- thropodan haemocyanins [51], the functional units in molluscan haemocyanins may have quite similar conformations. This pro- posal is supported by the hydrophilicity pattern [52] of H. po- matiu g , which was very similar to that of H. pomatia d [15]. From the study of these peptides it is clear that Asp165 in the sequence of H. pomatia d [I51 was wrongly attributed. As in all sequences of molluscan functional units determined, it is a cyste- ine residue (Fig. 2).
The peptides dII-5 and gI-3 (Fig. 2), each having the first disulfide bridge of the functional units, also contained the pecu- liar cysteine residue, as deduced from cysteic acid determin- ations. Their characteristically strong absorption at 255 nm and the liberation of 2-thiolhistidine on acid hydrolysis indicated a plausible C T H i s bridge. This could be confirmed from a more thorough study of peptides harbouring the peculiar cysteine resi- due. These peptides were liberated from reduced and pyridyleth- ylated &I-5 or carboxymethylated H. pomatia d by proteolysis with trypsin (undecapeptide) and with trypsin followed by pro- nase (tetrapeptide), respectively. The amino acid sequencing of the undecapeptide revealed Dabth derivatives of polymerized forms of didehydroalanine at the fourth position, corresponding to residue 60 in H. pornatia d , and of 2-thiolhistidine at the sixth position.
As no serine residue was detected by amino acid analysis of the undecapeptide, and no traces of Dabth-serine were found on TLC, the didehydroalanine deduced on sequencing must be considered to originate, together with the 2-thiolhistidine, from the bridge and not from serine [50] as wrongly deduced in the earlier sequencing work [15]. Moreover, when one is not aware of the presence of 2-thiolhistidine, the TLC spot may be con- fused with that of Dabth-glutamine [IS] or of Dabth-serine 1151.
The absorption spectrum of the tetrapeptide (Fig. 4) was quite similar to that of 2-thiolhistidine [48, 491, in favour of the existence of the bridge. The amino acid analysis yielded, i n addi- tion to S-carboxymethylcysteine, valine and some 2-thiolhisti- dine, the same supplementary peaks eluting between phenylala- nine and histidine as found for &I-5 and its undecapeptide, for gI-3, and for the peptides with the peculiar cysteine residue from functional units of other molluscan haemocyanins. The unex- pected peaks thus originate from the bridge and are tentatively ascribed to products derived from polymerized forms of didehy- droalanine, as deduced from the sequencing, and possibly also to some oxidized 2-thiolhistidine.
The conversion to cysteic acid upon acid hydrolysis in the presence of Me,SO, observed for the cysteine involved in the thioether bridge with histidine, also occurs on oxidation of cyste- ine residues linked by a thioether bond to bilins or to heme [53].
-

886 Gielens et al. (Eur: J. Biochern. 248)
The formation of didehydroalanine and 2-thiolhistidine from a C T H i s bond is quite compatible with an acid cleavage of this bond occurring under the conditions required for the total hydrolysis of the protein or for the liberation of the N-terminal residues upon Edman degradation. That a non-peptidic bridge exists in the functional unit between a cysteine and a histidine residue was furthermore corroborated by the electrospray-mass- spectrometry measurement of the tetrapeptide, whereby two pro- tons were deduced to be inissing compared with the calculated mass for the sequence Cys(CM)-Cys-Val-His. The residues at position 60 and 62 in H. pomatia d were thus wrongly attributed in the previous amino acid sequencing 1151 and must, like the equivalent ones in H. pomatia g [I61 and S. officinalis f [18], be replaced by Cys and His, respectively (Figs. 2, 5) .
The hydrophilicity pattern of H. pomatia d and g indicates that the thioether bridge is located in a hydrophobic region of the sequence. Application of a prediction program [52] indicates that it is buried in the protein structure, being part of a p turn.
The thioether bridge seems to be a general feature for func- tional units of molluscan haemocyanins since, in addition to H. pomnriu d and g , it was demonstrated in the functional units c, e, ,f, g and h of S. oficinulis haemocyanin and in functional unit g of 0. vulgaris haemocyanin (Fig. 5) . Cysteine and histidine residues were always deduced at the corresponding positions when the sequence of the functional units was determined from the cDNA (Fig. 5) . These units include S. officinalisf, for which serine and glutamine residues were deduced upon amino acid sequencing 1181. Functional units LI , b and d of S. ofiicinnlis haemocyanin, whose cDNAs have been sequenced recently (De Geest, N., unpublished results), and functional units b, c, d, e , f’ and g of 0. dqfkini haemocyanin, for which sequence data are available [13, 201, show the same typical Cys-Xaa-His sequence.
A thioether bridge linking a cysteine to a histidine residue has been reported for the tyrosinase of N . crassa [48, 491. This cysteine residue is the only one in the sequence [48]. The possi- bility of the occurrence of a same type of bond in the tyrosinase of Aspergillus oryzue was also assumed from the presence of a similar conserved sequence Cys-Xaa-His at an equivalent posi- tion 1541. By physicochemical analysis of peptides obtained from the enzyme isolated from a Neurospora culture grown on a medium supplemented with [”S]sulfate, [2,5-’H]histidine, and [5-’H]histidine, i t was demonstrated that the sulfur of the cyste- ine side group was linked to C2 of the imidazole group 1481. In the same study the presence of 2-thiolhistidine in an acid hydro- lysate of a tripeptide containing the thioether bridge was consid- ered as further evidence for this kind of linkage. As 2-thiolhis- tidine appeared on acid hydrolysis of the thioether-containing peptides studied here, it can be assumed that, as for Neurospora tyrosinase, the sulfur is bound at position 2 of the imidazole nucleus (Fig. 7). Moreover, in ergothioneine, a N-trimethylated and thiolated derivative of histidine 1551 that is synthesized from histidine, methionine and cysteine in fungi and in mycobacteria and accumulates in higher organisms by ingestion [56], thiol- ation occurs OR position 2 of the imidazole ring. Studies of the biosynthetic pathways in N. crussa [55, 571 and Mycobacterium smegmatis [58] indicated that cysteine is the principal sulfur source and the first step is the formation of a thioether bond with the histidine 155, 581 or the sulfoxide of this bond 1571. A thioether bridge between a cysteine residue and a tyrosine resi- due (sulfur bound at C3 atom) was identified in galactose oxi- dase from the fungus Dactylium dendroides [59].
As copper is liberated from the functional units at the low pH needed for pepsinolysis, and under the denaturing conditions used for the carboxymethylation, the question can be raised whether the thioether bridge is really present in the native func- tional units or whether it is formed as the result of a Cu’+-
H
CH, CH, I I
... - NH - CH - CO - NH - CH - CO - NH - CH-CO - ... R
Fig. 7. Chemical structure of the thioether bond in the consensus sequence Cys-Xaa-His found to occur in the functional units of mol- luscan haemocyanins. The consensus sequence 15 also found in the tyro- Sinase of N c m s w [48, 491
I
catalysed oxidation during or after the proteolytic treatment of the protein. As essentially the same thioether-containing pep- tides were obtained from a pepsinolysate of the apopreparations of H. pnmatia d and g as from the oxypreparations, it can be concluded that the cysteine-histidine thioether bridge is present in the native protein and that it is formed by a posttranslational modification.
Crystallographic data obtained on functional unit g of 0. dojZeini haemocyanin indicate that, as in tyrosinase, the histidine residue making part of the thioether bridge is probably one of the ligands of the first copper atom (Cu,) of the dinuclear copper group 1601. A similar conclusion was derived from resonance- Raman-spectroscopy data, which revealed lower frequencies for one of the copper-bound histidines 1611. In D. dendroides galac- tose oxidase the tyrosine involved in the thioether bridge func- tions as a ligand for the active-site copper atom [59].
The formation of the intermediary thioether bond or of its sulfoxide in the biosynthetic pathway of ergothioneine was found to require oxygen but may also need some metal and/or an enzyme [57, 581. In the posttranslational induction of the cysteine-histidine thioether bond in the functional units of the molluscan haemocyanins, where a copper able to participate i n the binding of dioxygen is in the vicinity, an autocatalytic oxida- tion reaction cannot be excluded.
The thioether bridge equivalent to that in Neuro.ypora tyrosi- nase fits with the hypothesis that molluscan haemocyanin evolved from tyrosinase 1621. The absence of a thioether bridge in the haemocyanin of the arthropod Cancer pagurus [48] is a further indication for the different origin of molluscan and arthropodan haemocyanins, as concluded from the sequence data 120, 25, 621.
We wish to thank the Fund fiir J o i r z t Basic Research, for research grants, the Katholirke Universitrit Leuven, for graduate fellowships (X.-Q. X.), the Station Marine at Wimereux, France, for providing S. officinalis, Dr S. v. Boletzky, Laboratoire Arago, Banyuls-sur-Mer, France, for collecting the haemolymph of 0. vulgaris, and Mrs J. Lem- mens and Mr J . Ryckeboer, for their skilful assistance. J . V. B. is in- debted to the Flemish Government for a Concerted Research Action (12052293).
REFERENCES 1. PrCaux, G. & Gielens, C. (1984) Hemocyanins, in Copper proteins
and copper enzyn2e.c. (Lontie. R., ed.) vol. 11, pp. 159-205, CRC Press, Boca Raton FL.
2. Lontie, R. (1983) Components, functional units, and active sites of Helixponzuria haemocyanin, Life Chem. Rep. Suppl. I , 109- 120.
3. Van Bruggen, E. F. J., Schutter, W. G., Van Breemen, J. F. L., Bijl- holt, M. M. C. & Wichertjes T. (1981) Arthropodan and mollus-

Gielens et al. (Eur: J. Biochem. 248) 887
can hernocyanins. in Electron microscopy of proteins (Harris, J , R., ed.) vol. 1 , pp. 1-38, Academic Press, London.
4. Lambert, 0.. Boisset, N., Tavedu J.-C., Preaux, G. & Lamy, J. N. (1 995) Three-dimensional reconstruction of the a, and P,.-hemo- cyanins of Helix pomatia from frozen-hydrated specimens, ,/. Mid. Biol. 248, 431-448.
5. Van Breemen, J. F. L., Schuurhuis, G. J . & Van Bruggen, E. F. J. (1 977) Morphology of Helix pornatia hemocyanin and its sub- units, in Structure andfiuzctiorz of haemocyanins (Bannister, J. V., ed.) pp. 122- 127, Springer-Verlag, Berlin.
6. Gielens, C., Prkaux, G. & Lontie, R. (1977) Structural investigations on b-haemocyanin of Helix pornaria by limited proteolysis, in Structure and ,function of haernocyanin (Bannister, J. V., ed.) pp. 85 -94, Springer-Verlag, Berlin.
7. Gielens, C., Verschueren, L. J., Prtaux, G. & Lontie, R. (1981) Lo- calization of the domains in the polypeptide chain of p, hemocya- nin of Helix pomatia, in Invertebrate oxygen-binding proteins: structure, active site, and function (Lamy, J. & Lamy, J., eds) pp. 295-303, Marcel Dekker, New York.
8. Gielens, C., Verschueren, L. J., PrCaux, G. & Lontie, R. (1980) Frag-
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
mentation of crystalline p-haemocyanin of Helix pornatia with plasmin and trypsin. Location of the fragments in the polypeptide chain, Eur: J. Biochern. 103, 463-470.
Gielens, C., Bosman, F., Prtaux, G. & Lontie, R. (1983) Structural studies by limited proteolysis of the haemocyanin of Sepia offci- nalis, Lifi Chem. Rep. Suppl. I , 12 1 - 124.
Gielens, C., Benoy, C., Prtaux, G. & Lontie, R. (1986) Presence of only seven functional units in the polypeptide chain of the haelnocyanin of' the cephalopod Octopus vulguris, in lnvertehrute oxygen ccirriers (Linzen, B., ed.) pp. 223 -226, Springer-Verlag, Berlin.
Wichertjes, T.. Gielens, C., Loncke, P., PrCaux, G. & Van Bruggen, E. F. J. (1990) Comparative electron microscopy of the haemocy- anins of the cephalopods Sepia oflicinulis and Octopus vulgaris, in Invertebrate dioxygen carriers (PrCaux, G. & Lontie, R.. eds) pp. 289-294, Leuven University Press, Louvain.
Lamy, J., Gielens, C., Lambert, O., Taveau, J.-C., Motta, G., Loncke, P., De Geest, N., Prtaux, G. & Lamy, J. (1993) Further approaches to the quaternary structure of Ocropus hemocyanin: a model based on immunoelectron microscopy and image processing, Arch. Biochem. Biophys. 305, 17-29.
Lang, W. H. & Van Holde, K. E. (1990) cDNA cloning and sequenc- ing of Octopus dofleini hemocyanin, in Invertebrate dioxygen carriers (Prtaux, G. & Lontie, R., eds) pp. 135-140, Leuven University Press, Louvain.
Lamy, J., Leclerc, M., Sizaret, P.-Y., Lamy, J. N., Miller, K. I., McParland, R. & Van Holde, K. E. (1986) Octopus dofleini hemo- cyanin : structure of the sevendomain polypeptide chain, Bio- chemistry 26, 3509-3518.
Drexel, R., Siegmund, S., Schneider, H.-J., Linzen, B ., Gielens, C., Prtaux, G., Lontie, R., Kellermann, J. & Lottspeich, F. (1987) Complete amino-acid sequence of a functional unit from a mol- luscan hemocyanin (Helix pomatia), Biol. Chem. Hoppe-Seyler 368, 617-635.
Xin, X.-Q., Gielens, C., Witters, R. & Prtaux, G. (1990) Amino- acid sequence of functional unit g from the P,-haemocyanin of Helix pomutia, in Invertebrate dioxygen carriers (Prtaux, G. & Lontie, R., eds) pp. 113-1 17, Leuven University Press, Louvain.
Top, A,, Witters, R., Gielens, C., Van Beeumen, J. & Prtaux, G. (1 988) Amino-acid sequence determination of an N-terminal frag- ment from functional unit f of Sepia oflicinalis haemocyanin, Arch. Intern. Physiol. Biochim. 96, B187.
Top, A,, Gielens, C., Witters, R., Van Beeumen, J. & PrCaux, G. (1990) Partial amino-acid sequence and location of the carbohy- drate chain in functional unit f of Sepia officinalis haemocyanin, in Invertebrate dioxygen carriers (Prtaux, G. & Lontie, R., eds) pp. 119- 123, Leuven University Press, Louvain.
Declercq, L., Witters, R. & Preaux, G. (1990) Partial sequence deter- mination of Sepia oficinalis haemocyanin via cDNA, in Inverte- brate dioxygen carriers (Prtaux, G. & Lontie, R., eds) pp. 131- 134, Leuven University Press, Louvain.
Van Holde, K. E. & Miller, K. I. (1995) Hemocyanins, Adv. Protein Ckem. 47, 1-81.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
Van Kuik. J. A., Kamerling, J. P. & Vlicgcnthart, J. F. G. (1990) Carbohydrate analysis of hernocyanins, in /nwrtebrotc dioxJgen carrier.s (Prtaux, G. & Lontie, R., eds) pp. 157-163, Leuven University Press, Louvain.
Wood, E. J., Chaplin, M. F., Gielens, C., De Sadeleer, J., PrCaux, G. & Lontie, R. (1985) Relative molecular mass of the polypep- tide chain of P,.-haemocyanin of Helix pomatia and carbohydrate composition of the functional units, Cornp. Biochern. Physiol.
Vanhoegaerden, R., Gielens, C. & PrCatix, G. (1988) Location of the carbohydrate in the fuiictional unit d of' the &haemocyanin of Helix poniutia, Arch. Intern. Physiol. Biochem. 96, B64.
Vanhoegaerden, R., Gielens, C. & PrCaux, G. (1990) Location of the carbohydrate chains in functional unit g of the /&haemocyanin of Helix pomatiu, in Invertebrate dioxygen curriers (Preaux. G. & Lontie, R., eds) pp. 153-156, Leuven University Press, Louvain.
Lontie, R., Witters, R., Gielens, C. & Preaux, G. (1990) Comparison of the amino-acid sequences of molluscan haemocyanin5 and of tyrosinases, in Invertebrate d ioxypn urriers (Prtaux, G. & Lon- tie, R., eds) pp. 142 - 146, Leuven University Press, Louvain.
Gielens, C., De Geest, N., Xin, X.-Q. & Preaux, G. (1992) Localiza- tion of disulfide bridges and identification of a thioether bond in functional units d and g of &haemocyanin of Helix pomatia, International congress on invertebrute dioxygen carriers, 12 - 1 7 April 1992, Lunteren, The Netherlands, abstract no. 11-L2.
Gielens, C., De Geest, N., Xin, X.-Q. & Prtaux, G. (1994) Identifi- cation of a thioether bond and of disulfide bridges in functional units d and g of bc-haemocyanin of Helix pomatia, Arch. Intern. Physiol. Biochim. Biophys. 102, B1 1.
Heirwegh, K., Borginon, H. & Lontie, R. (1961) Separation and absorption spectra of a- and 1-haemocyanin of Heli,~ pomatia, Biochim. Biophys. Acta 48, 517-526.
PrCaux, G., Gielens, C., Verschueren, L. J. & Lontie, R. (1981) Im- munological comparison of the hernocyanins of He1i.x pomutici and of the proteolytic fragments of the b,. component, in Inverte- brate oxygen-binding proteins : structure, active site, and,fhction (Lamy, J. & Lamy, J., eds) pp. 197-205, Marcel Dekker, New York.
Kubowitz, F. (1938) Spaltung und Resynthese der Polyphenoloxi- dase und des Hamocyanins, Biochem. Z. 2YY, 32-57.
Perham, R. N. (1978) Techniques for determining the amino-acid composition and sequence of proteins, in Techniques in protein and enzyme biochemistq (Kornberg H. L., Metcalfe, J . C., North- cote, D. H., Pogson, C. I. & Tipton, eds) pp. 1-39, Elsevier/ North-Holland, County Clare.
Wichertjes, T., Gielens, C., Schutter, W. G., Prtaux, G., Lontie, R. & Van Bruggen, E. E J . (1986) The quaternary structure of Sepia officinalis haemocyanin, Biochini. Biophys. Acta 872, 183 - 194.
Loncke, P., Vanderzande, M., Gielens, C. & PrCaux, G. (1990) lden- tification of the missing functional unit in Ocropus vulgaris haemocyanin from a comparison with Sepia oflicinalis haemocya- nin, in Invertebrate dioxygen carriers (PrCaux, G. & Lontie, R., eds) pp. 295 -298, Leuven University Press, Louvain.
Vanderzande, M., Gielens, C. & PrCaux, G. (1990) Isolation of func- tional units g and h from the haemocyanin of Sepiu officinalis and partial amino-acid sequence of functional unit h, in lnvertrhrutr dioxygen curriers (PrCaux, G. & Lontie, R., eds) pp. 125-129, Leuven University Press, Louvain.
Friedman, M., Krull, L. H. & Cavins, J. F. (1970) The chromato- graphic determination of cystine and cysteine residues in proteins as S-P-(4-pyridylethyl)cysteine, J. B id . Chem. 245, 3868 -3871.
Laemmli, U. K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4, Nature 227, 680-685.
Weeke, B. (1973) Crossed immunoelectrophoresis, in A munuul of quuntitutive imrnurioelectrophoresis. Methods and applications (Axelsen, N. H., Krflll, J. & Weeke, B., eds) pp. 47-56, Universi- tetsforlaget, Oslo.
Gielens, C., Verschueren, L. J. , PrCaux, G. & Lontie, R. (1981) Gel chromatographic separation of the haemocyanins of Helix po- matia. Further electrophoretic and immunological characteriza- tion of the components, Comp. Biochem. Physiol. 69B, 455 -462.
Dubois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A. & Smitt, F. (1956) Colorimetric method for determination of sugars and related substances, Anal. Chew. 28, 350-356.
82B, 179-186.

888 Gielens et al. (Eur: J. Biochern. 248)
40. Habeeb, A. F. S. A. (1972) Reaction of protein sulfhydryl groups
41. Ellman, G. L. (1959) Tissue sulthydryl groups, Arch. Biocheni. Bio-
42. Andrews, R. P. & Condon, G. D. (1983) The analysis of protein hydrolysates on the LKB 4150 alpha amino acid analyser, Protein Chemistry Note No. 26, LKB Biochrom, Cambridge.
43. Spencer, R. L. & Wold, F. (1969) A new convenient method for estimation of total cystine-cysteine in proteins, Antrl. Biochern.
44. Inglis, A. S. (1983) Single hydrolysis method for all amino acids, including cysteine and tryptophan. Methods En~yniol. 91, 26-36.
45. Chang, J. Y., Brauer, D. & Wittniann-Liebold, B. (1978) Micro- sequence analysis of peptides and proteins using 4-NN-dimethyl- aminoazobenzene 4’-isothiocyanate/phenylisothiocyanate double coupling method, FEBS Leu. 93, 205-214.
46. Hunkapiller, M. W. & Hood, L.. E. (1989) Protein sequence analy- sis: automated microsequencing. Science 219, 650-659.
47. Fenn. J. B., Mann, M., Meng, C. K., Wong, S. F., Whitehouse. C. M. (1989) Electrospray ionization for inass spectrometry of large biomolecules, Science 246, 64-71.
48. Lerch, K . (1982) Primary structure of tyrosinase from Neuro.spora cru,ssu. 11. Complete amino acid sequence and chemical structure of a tripeptide containing an unusual thioether, J . Bid . Chenz.
49. Lerch, K. (1984) S[’-(2-Histidyl)cysteine: properties, assay, and oc- currence, Methods Enzy/nnol. 106, 355-359.
50. Chang, J. Y. (1979) The destruction of serine and threonine thiohy- dantoins during the sequence determination of peptides by 4-N,N- dimethylaminoazohenxene 4’-isothiocyanate, Biochim. Biophys.
51. Linzen, B., Soeter, N., Riggs, A. F., Schneider. H.-J., Schartau, W., Moore, M. D., Yokota, E., Behrens, P. Q., Nakashima, H., Takagi, T., Nemoto, T., Vereijken, J. M., Bak, H. J., Beintema, J . J., Volbeda, A,, Gaykema, W. P. J . & Hol, W. G. J. (1985) The struc- ture of arthropod hemocyanins, Science 229, 519-524.
52. Carmenas, R. S., Freije. J. P., Molina, M. M. & Martin, J. M. (1989) PREDICT7, a program for protein structure prediction, Biochem. Biophyy. Ke.r. Commun. 159, 687-693.
with Ellman’s reagent, Methods Enzymol. 25, 457-464.
phys. 82, 70-77.
32, 185- 190.
257, 6414-6419.
Acfu 57X, 175-187.
53. Glazer, A. N. (1984) Sp-(Bilin)cysteine derivatives: structures, spec- troscopic properties, and quantitation, Metho& Enzyrnol. 106, 359 -364.
54. Fujita, Y., Uraga, Y. & Ichisima, E. (1995) Molecular cloning and nucleotide sequence of the protyrosinase gene, rnelO, from Asper- gillus o9zue and expression of the gene in yeast cells, Biochim.
55. Melville. D. B., Eich, S. & Ludwig, M. L. (1957) The biosynthesis
56. Hartman, P. E. (1990) Ergothioneine as antioxidant, Methods Enzy-
57. Ishikawa, Y.. Israel, S. E. & Melville. D. B. (1974) Participation of an intermediate sulfoxide in the enzymatic thiolation of the imid- azole ring of hercynine to form ergothioneine, J. Biol. Chem. 249,
58. Genghof, D. S. & Van Damme, 0. (1968) Biosynthesis of ergothio- neine from endogenous hercynine in Mycobacteriurn srneynzaris, J , Racteriol. 95, 340-344.
59. Ito, N., Phillips, S. E. V., Stevens, C., Ogel, 2. B., McPherson, M. J. , Keen, J. N., Yadav, K. D. S. & Knowles, P. F. (1991) Novel thioether bond revealed by a 1.7 A crystal structure of galactose oxidase, Nature 350, 87-90,
60. Cuff, M. & Hendrickson, W. A. (1996) The crystal structure of a molluscan heinocyanin, in Infernational conference on suprarno- leculmr organization and functionul regulution of invertebrate di- oxygen binding proteins, 1-5 July 1996, Padova, Abstract no. L-7.
61. Ling, J., Nestor, L. P., Czernuszewicz, R. S., Spiro, T. G., Fraczkie- wicz, R., Sharma, K. D., Loehr, T. M. & Sanders-Loehr, J. (1994) Common oxygen binding site in hemocyanins from arthropods and mollusks. Evidence from Raman spectroscopy and normal coordinate analysis, .I. Am. Chem. Soc. 116, 7682-7691.
62. PrCaux, G., Gielens, C., Witters, R. & Lontie, R. (1988) The struc- ture of molluscan haemocyanins and their homology with tyrosi- nases, Bull. Soc. Chim. Belg. 97, 1037- 1044.
Biophys. Acts 1261, 151-154.
of ergothioneine, J. Biol. Chern. 224, 871 -877.
mol. 186, 310-318.
4420-4427.




















