
Establishing an in vivo p48ZnF bioluminescence mouse brain imaging model
6
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/authorsrights
Transcript of Establishing an in vivo p48ZnF bioluminescence mouse brain imaging model

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/authorsrights

Author's personal copy
Neuroscience Letters 542 (2013) 97– 101
Contents lists available at SciVerse ScienceDirect
Neuroscience Letters
jou rn al hom epage: www.elsev ier .com/ locate /neule t
Establishing an in vivo p48ZnF bioluminescence mouse brain imaging model
Klaus Heese ∗
Department of Biomedical Engineering, College of Engineering, Hanyang University, 222 Wangsimni-ro, Seongdong-gu, Seoul 133-791, Republic of Korea
h i g h l i g h t s
� In vivo p48ZnF mouse brain bioluminescence imaging.� p48ZnF interacts with Drg1 and Pcbp1.� p48ZnF regulates gene transcription and translation.
a r t i c l e i n f o
Article history:Received 12 January 2013Received in revised form 20 February 2013Accepted 23 February 2013
Keywords:p48ZnFDfrp1Drg1Zinc fingerGene transcription
a b s t r a c t
p48ZnF is a C3H1 zinc finger domain-containing protein that is involved in the control of gene tran-scription and translation. In the present study a novel transgenic p48ZnF mouse model is described thatis useful for in vivo brain imaging using luciferase as bioluminescence-mediating reporter gene. Yeasttwo-hybrid screening and western blot analyses revealed Drg1 (developmentally regulated GTP bindingprotein 1) and Pcbp1 (poly (rC)-binding protein 1) as p48ZnF-associated proteins. Interestingly, p48ZnF’cellular location of action depends on the cell’s differentiation status: nuclear in proliferating cells andcytoplasmic in differentiated neurons.
© 2013 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
Many transcription factors or DNA regulatory proteins con-tain zinc-finger domains which enable these proteins to bind toDNA and regulate DNA expression [11]. The recently identifiednovel protein p48ZnF is glutamate/lysine-rich and contains twocharacteristic C-x8-C-x5-C-x3-H (CCCH) zinc finger domains [3](Fig. 1).
C3H1-type domains are relatively rare – only 58 discrete C3H1-type domains can be found in the human and murine genomes andmost of these domains have yet to be characterized [10]. Thus, to getfurther insights into the neuronal function of p48ZnF, in the presentstudy a transgenic mouse model has been established where brainp48ZnF protein levels could be observed in vivo in a living mousevia a bioluminescence-mediated luciferase reporter gene expres-sion. Alongside, p48ZnF’s signaling pathways have been exploredby in vitro molecular and cellular methods as well as by bio-computational analyses.
∗ Tel.: +82 2 2220 0398; fax: +82 2 2296 5943.E-mail addresses: [email protected], [email protected]
2. Materials and methods
2.1. Reagents
All reagents used for experiments were purchased fromSigma–Aldrich (Milwaukee, WI, USA) unless otherwise stated.
2.2. Antibodies
Anti-�-tubulin: 1:1000, mouse monoclonal (Santa Cruz Biotech-nology Inc., Santa Cruz, CA, USA), anti-beta-actin (Actb, 1:3000, goatpolyclonal; Santa Cruz), anti-p48ZnF (rabbit polyclonal, 1:3000;raised against peptide aa cSGGRAENGERSDLEE), anti-Drg1 (devel-opmentally regulated GTP binding protein 1, rabbit polyclonal,1:400; kindly provided by Inuoe et al. [8]), anti-Gapdh (glyceralde-hyde 3-phosphate dehydrogenase, goat polyclonal, 1:5000; SantaCruz), anti-Pcbp1 (poly (rC)-binding protein 1, mouse polyclonal,1:1000; Abnova, Taiwan), and anti luciferase (rabbit polyclonal,1:1000; Abcam, Cambridge, UK).
2.3. Animal experiments
Experimental methods, including the killing of animals, wereperformed in accordance with the International Guiding Principles
0304-3940/$ – see front matter © 2013 Elsevier Ireland Ltd. All rights reserved.http://dx.doi.org/10.1016/j.neulet.2013.02.044

Author's personal copy
98 K. Heese / Neuroscience Letters 542 (2013) 97– 101
Fig. 1. Overexpression of p48ZnF in transgenic mice brain using a bioluminescencereporter gene. (A) Schematic diagram of the p48ZnF (also known as Zc3h15, Dfrp1,Lerepo4 or Tma46; locus: AAR24540 and NP 001010963) amino acid sequence.The two zinc finger domains (aa100–aa125, aa175–aa211) depicted are of theC3H1-type with a DNA-binding domain at aa147–aa188. In addition to the lysine-rich region (aa4–aa175), two bipartite nuclear localization signals (NLS, aa4–aa36,aa262–aa279), one nuclear export domain (NES, aa415–aa424) and a DFRP domain(aa234–aa295) have been identified [3,8]. (B) Schematic diagram of the transgenevector insertion construct used to establish the transgenic p48ZnF mouse line. Themouse synapsin-I promotor (Syn1P) with its natural intron was introduced behindthe rabbit �-globulin 5′ UTR (untranslated region) followed by the mouse p48ZnFgene, an internal ribosome entry site (IRES), the luciferase gene, the woodchuckhepatitis virus post-transcriptional regulatory element (WPRE), and the rabbit �-globulin 3′ UTR containing its own polyadenylation signal (pA). (C) In vivo mousebrain bioluminescence luciferase activity imaging in alive mice showing p48ZnFexpression in the brain of a transgenic (tg) mouse compared with a correspondingwild-type littermate (top: female, 1 week old, wild type; bottom: female, 1 week old,tg p48ZnF strain 37.1013). Small inserted panel: ex vivo mouse brain biolumines-cence luciferase activity imaging (tg p48ZnF strain 37.1013 (female, 1 week old)). (D)IHC analysis of transgenic (tg) mouse brain using anti-luciferase (tg-luc) and anti-p48ZnF antibodies confirmed neuronal expression of p48ZnF in the hippocampusand dentate gyrus (same mouse brain but different slices). Wild-type (wt) mice alsoexpressed endogenous neuronal p48ZnF in the hippocampal area. Tg- and wt-mice:both female, 3 months old. Scale bar represents 100 �m.
for Animal Research (WHO) and were approved by the local Institu-tional Animal Care and Use Committee. Mouse tissues were isolatedfrom C57BL/6J mice after the humane killing of the animals usingapproved anesthetic methods to isolate brain tissue and to ana-lyze p48ZnF expression. All efforts were made to minimize animalsuffering and to reduce the number of animals used [1,14,15,17].
2.4. Establishment of the transgene and the transgenic p48ZnFmouse lines
The p48ZnF mouse lines were generated analogously to apreviously described transgenic ‘synapsin-I-p60TRP’ mouse [14].Fig. 1 shows a schematic diagram of the insertion construct usedto establish the transgenic p48ZnF mouse line. The transgene
vector was constructed as indicated (Fig. 1B). The transgene vectorwas injected in different sessions into zygotes of C57BL/6N femalemice obtained from Charles River Wiga GmbH (Sulzfeld, Germany),superovulated and mated with C57BL/6N males. Injected zygoteswere cultivated overnight and transferred into pseudo-pregnantB6CBAF1 females (Charles River) and the animals were kept inindividually ventilated cages (IVG). Transgenic founder mice result-ing from second-round pronucleus injections were identified bypolymerase chain reaction (PCR). From the founders the F1 gen-erations were produced and bred to maintain as heterozygouslines (PolyGene AG, Switzerland). Expression of p48ZnF/luciferasein transgenic mice was confirmed by western blot (data not shown)and bioluminescence luciferase activity-based imaging via theNightOWL LB983 Imager (Berthold Technologies GmbH & Co. KG,Bad Wildbad, Germany). Brain tissue luciferase assay was per-formed as described previously. Adult mice were anesthetized viaintraperitoneal injection of ketamine/xylazine (RBI, Natick, MA,USA) and brain tissue was taken and subjected to luciferase assayusing reagents obtained from Promega (Madison, WI, USA) [14].Representative pictures are shown. Mouse brain immunohisto-chemistry (IHC) was performed as described previously [13,14,20].
2.5. Cell culture of PC12 cells
Rat pheochromocytoma PC12 cells (American Type Culture Col-lection, ATCC, Manassas, VA, USA) were propagated at 37 ◦C inhumidified 5% CO2/95% air, in Dulbecco’s Modified Eagle’s Medium(DMEM, GlutaMaxTM; Invitrogen, Carlsbad, CA, USA) supplementedwith 15% fetal bovine serum (FBS, Invitrogen), non-essential aminoacids (Invitrogen), and antibiotic–antimycotic (Invitrogen). Allmedia used for cell culture were from Gibco unless otherwisestated. Cells were cultured at 37 ◦C in humidified 5% CO2/95% air. Forneuronal differentiation PC12 cells were seeded in poly-d-lysine(PDL)-coated six-well plates (Becton Dickinson, San Jose, CA, USA)at 1.5 × 104 cells per well. The cells were allowed to attach anddivide for 20 h, after which 100 ng/ml NGF (nerve growth factor)was added for 1 week. Images were captured via an inverted micro-scope (Nikon Eclipse TE2000, Tokyo, Japan). Vehicle controls weretreated with the solvent only. Experiments were performed twice,with each set repeated in triplicates and representative data areshown [3,15].
2.6. Cell lysis and protein extraction
Adherent cells were washed in the dish using ice-coldCa2+/Mg2+-free saline phosphate buffer (PBS), lysed using a dis-posable cell scraper (Greiner Bio-One GmbH, Frickenhausen,Germany). Lysed cells were centrifuged at 10,000 × g at 4 ◦C for10 min. Lysis buffer (20 mM Tris–HCl (pH 7.4), 150 mM NaCl,40 mM NaF, 5 mM EDTA, 0.5 mM dithiothreitol (DTT), 1% Triton-X-100, 1 mM sodium orthovanadate, 1% (vol/vol) Nonidet P-40, 0.1%(wt/vol) sodium deoxycholate, 0.1% (wt/vol) sodium dodecyl sul-fate (SDS), 1 mM phenylmethylsulfonyl fluoride (PMSF), 10 ng/mlof aprotinin and 10 ng/ml leupeptin plus a EDTA-containing(serine- and cysteine-) protease inhibitor (Roche) as well as ser-ine/threonine/tyrosine phosphatase inhibitor cocktails 1 and 2(Sigma)) was added and incubated on ice for 5–10 min before apply-ing for SDS–polyacrylamide gel electrophoresis (SDS–PAGE) andwestern blot analysis [13].
2.7. Co-immunoprecipitation
For immunoprecipitation [13], PC12 cells were lysed in lysisbuffer (20 mM Tris–HCl (pH 7.4), 150 mM NaCl, 40 mM NaF,1 mM sodium orthovanadate, 1% (vol/vol) Nonidet P-40, 0.1%(wt/vol) sodium deoxycholate, 1% (vol/vol) Triton-X-100, 1 mM

Author's personal copy
K. Heese / Neuroscience Letters 542 (2013) 97– 101 99
PMSF, 20 ng/ml of aprotinin and 20 ng/ml leupeptin), followedby centrifugation at 14,000 rpm at 4 ◦C for 15 min. After pro-tein quantification, equal amounts of the protein supernatantswere incubated with polyclonal rabbit anti-p48ZnF antibody-loaded protein A/G-sepharose at 4 ◦C overnight. As a control,the lysates were incubated with a non-specific rabbit polyclonalantibody-loaded protein A/G-sepharose. The immunoprecipitateswere then washed five times in lysis buffer, and the bound pro-teins were recovered by boiling the beads in 2× SDS sample bufferand separated by SDS–PAGE, followed by western blot with aspecific antibody (anti-Pcbp1 or anti-Drg1) as indicated. Experi-ments were performed three times and representative data areshown.
2.8. SDS–PAGE and western blot analysis
Twenty micrograms of cell lysates were resolved by 8–12%SDS–PAGE at 0.02 A of constant current and transferred to apolyvinylidine fluoride (PVDF) membrane (0.22 �m; Amersham,Piscataway, NJ, USA) using the ‘semi-dry’-transfer method (BioRad,Singapore) for 60 min at 0.12 A in buffer containing 25 mM Tris,192 mM glycine, 20% methanol, and 0.01% (wt/vol) SDS. The mem-brane was blocked with 5% bovine serum albumin (BSA) (BioRad)in Tris-buffered saline (TBS) solution plus 0.1% Tween-20 (TBS-T) orPBS-T for 2 h at room temperature (RT), washed three times in PBS-T for 10 min each, and incubated with primary antibody (diluted in2% BSA in PBS-T) for overnight at 4 ◦C. The membranes were washedas described above, incubated with a horseradish peroxidase (HRP)-conjugated secondary antibody for 1 h at RT, and developed usingthe ECL plus western-blot detection reagent (Amersham). X-rayfilms (Konica Minolta Inc., Tokyo, Japan) were exposed to the mem-branes before film development in a Kodak X-OMAT 2000 processor(Kodak, Ontario, Canada). For equal sample loading, protein quan-tification was performed using a ‘2D Quant’ kit (Amersham) withtwo independent replicates. BSA was used as a standard for pro-tein quantification. To re-probe the same membrane with anotherprimary antibody, Pierce’s (Pierce Biotechnology, Inc., Rockford, IL,USA) ‘stripping solution’ was used to strip the membranes. In addi-tion, equal sample loading was confirmed using Gapdh or Actb asa reference protein (data not shown). Western blot experimentswere performed three times and representative blots are shown[13].
2.9. Immunocytochemistry (ICC)
Cells seeded on poly-l-lysine (PLL; Sigma)-coated coverslipswere fixed with 4% paraformaldehyde in PBS for 10 min at RT,washed twice with PBS and then permeabilized with 0.5% TritonX-100 (Sigma) in PBS for 5 min at RT. The cells were washed thricewith PBS and incubated with primary antibody diluted in TBSTwith 4% BSA (AppliChem GmbH, Denmark) for 1 h at RT. Follow-ing incubation, cells were washed thrice with TBST and incubatedfor 30 min with a secondary antibody (labeled with FITC (Alex-aFluor 568, goat polyclonal anti-rabbit, 1:400, or AlexaFluor 488,goat monoclonal anti-mouse, 1:400; Molecular Probes, Eugene,OR, USA)) in TBST with 4% BSA for 1 h in the dark at RT. Afterthree final washes with TBST, the coverslips were mounted ontomicroscope glass slides using anti-fade solution containing DAPI(4′,6-diamidino-2-phenylindole; Chemicon, Temecula, CA, USA).Slides were visualized using an Axiovert 200 M inverted florescencecompound microscope (Carl Zeiss, Germany) and images were ana-lyzed with Axiovision software. Experiments were performed threetimes and representative figures are shown [16,20].
2.10. ProQuestTM two-hybrid-system with GatewayTM
technology
The two-hybrid system is an in vivo yeast-based system thatidentifies the interaction between two proteins (for instanceX = p48ZnF and Y = cDNA library, Pcbp1 or Drg1) by reconstitutingan active transcription factor. Analyses were performed accordingto the manufacturer’s protocol (Invitrogen’s brain ProQuestTM two-hybrid-system, Singapore) using p48ZnF as bait. In the ProQuestTM
two-hybrid-system, in comparison to standard two-hybrid sys-tems, false positives are reduced because three independenttranscription events (from distinct promoters) must occur atindependent chromosomal loci. Positive clones were confirmedby retransformation assays and co-immuneprecipitation analyses[4,13].
3. Results
3.1. In vivo mouse brain imaging of neuronal p48ZnF expression
A novel transgenic mouse model has been developed in whichthe expression of p48ZnF was driven by the synapsin-1 promoter
Fig. 2. p48ZnF interacts with Pcbp1 and Drg1. Co-immune precipitation (Co-IP) performed with the anti-p48ZnF antibody. (A) Western blot membrane probed with theanti-Drg1 antibody: lane 1: Co-IP product obtained with the anti-p48ZnF antibody; lane 2: total cell lysate. (B) Western blot membrane probed with the anti-Pcbp1 antibody:lane 1: Co-IP product obtained with the anti-p48ZnF antibody; lane 2: total cell lysate. (C) Western blot membrane probed with the anti-p48ZnF antibody: lane 1: Co-IPproduct obtained with the anti-p48ZnF antibody; lane 2: total cell lysate.
Author's personal copy
100 K. Heese / Neuroscience Letters 542 (2013) 97– 101
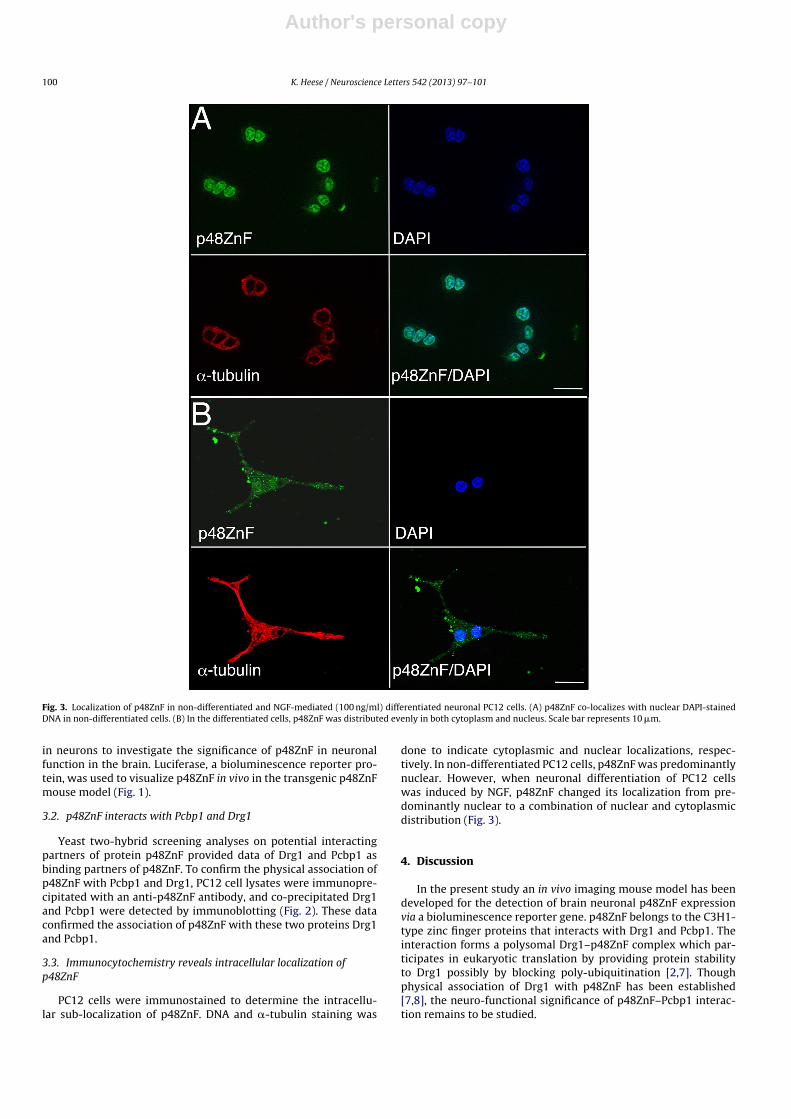
Fig. 3. Localization of p48ZnF in non-differentiated and NGF-mediated (100 ng/ml) differentiated neuronal PC12 cells. (A) p48ZnF co-localizes with nuclear DAPI-stainedDNA in non-differentiated cells. (B) In the differentiated cells, p48ZnF was distributed evenly in both cytoplasm and nucleus. Scale bar represents 10 �m.
in neurons to investigate the significance of p48ZnF in neuronalfunction in the brain. Luciferase, a bioluminescence reporter pro-tein, was used to visualize p48ZnF in vivo in the transgenic p48ZnFmouse model (Fig. 1).
3.2. p48ZnF interacts with Pcbp1 and Drg1
Yeast two-hybrid screening analyses on potential interactingpartners of protein p48ZnF provided data of Drg1 and Pcbp1 asbinding partners of p48ZnF. To confirm the physical association ofp48ZnF with Pcbp1 and Drg1, PC12 cell lysates were immunopre-cipitated with an anti-p48ZnF antibody, and co-precipitated Drg1and Pcbp1 were detected by immunoblotting (Fig. 2). These dataconfirmed the association of p48ZnF with these two proteins Drg1and Pcbp1.
3.3. Immunocytochemistry reveals intracellular localization ofp48ZnF
PC12 cells were immunostained to determine the intracellu-lar sub-localization of p48ZnF. DNA and �-tubulin staining was
done to indicate cytoplasmic and nuclear localizations, respec-tively. In non-differentiated PC12 cells, p48ZnF was predominantlynuclear. However, when neuronal differentiation of PC12 cellswas induced by NGF, p48ZnF changed its localization from pre-dominantly nuclear to a combination of nuclear and cytoplasmicdistribution (Fig. 3).
4. Discussion
In the present study an in vivo imaging mouse model has beendeveloped for the detection of brain neuronal p48ZnF expressionvia a bioluminescence reporter gene. p48ZnF belongs to the C3H1-type zinc finger proteins that interacts with Drg1 and Pcbp1. Theinteraction forms a polysomal Drg1–p48ZnF complex which par-ticipates in eukaryotic translation by providing protein stabilityto Drg1 possibly by blocking poly-ubiquitination [2,7]. Thoughphysical association of Drg1 with p48ZnF has been established[7,8], the neuro-functional significance of p48ZnF–Pcbp1 interac-tion remains to be studied.

Author's personal copy
K. Heese / Neuroscience Letters 542 (2013) 97– 101 101
Fig. 4. STRING-9.0 analysis (at http://string-db.org/; default mode) of humanp48ZnF’s potential interactive signaling pathways. The neuro-functional signifi-cance of p48ZnF within the PCBP1 and DRG1 networks remains to be elucidated, inparticular in view of its potential function in terms of neuronal development, axonguidance and synaptic plasticity. HNRNPA2B1, heterogeneous nuclear ribonucleo-protein A2/B1; HNRNPD, HNRNP D (AU-rich element RNA binding protein 1); LMNA,lamin A/C; PTBP1, polypyrimidine tract binding protein 1; SF3A2, splicing factor 3a,subunit 2; SRSF2, serine/arginine-rich splicing factor 2; SRSF9, serine/arginine-richsplicing factor 9; YBX1, Y box binding protein 1.
Pcbp1 belongs to the heterogeneous nuclear ribonucleoproteinfamily (hnRNP) and participates in transcriptional and translationalregulation. Pcbps are generally known as RNA-binding proteinsthat interact in a sequence-specific fashion with single-strandedpoly(C). Although Pcbp1 is known to affect the stability of geneexpression through post-translational regulation, its exact functionin neuronal axonal navigation and connectivity is still unclear [9].
The presence of Pcbp1 and p48ZnF interaction suggests aclose transcriptional–translational pathway with p48ZnF regu-lating processes on the DNA and protein levels while Pcbp1 isinvolved in a diverse set of post-translational control pathwayssuch as mRNA stabilization, translational activation and transla-tional silencing [12]. Meanwhile, coupled with Drg1, which is acytoplasmic developmentally regulated GTP-binding protein thatregulates the transcriptional–translational process by modulatingGTP availability, the Drg1–p48ZnF–PcbpP1 complex may governcytoplasmic and nuclear transcriptional/translational processespotentially involved in neuronal differentiation, axon guidance andsynaptic plasticity (Fig. 4).
Besides, Pcbp1 is an iron chaperone that binds iron and deliversit to ferritin, a cytosolic iron storage protein. Pcbp1 is also requiredfor the iron incorporation into iron-dependent prolyl hydroxylases(PHDs). Such data suggests a broad role for Pcbp1 in deliveringiron to cytosolic non-heme iron enzymes [19,21]. Since Pcbp1may control the expression or activity of Wnt-, TGF-� and mu-opioid receptor (MOR) signaling pathway genes too [5,6,18], thep48ZnF mouse model could be useful to study these pivotal neuro-developmental signaling cascades as it still remains a challengingresearch area.
Acknowledgements
This study was supported by the Hanyang University. I wouldlike to thank Ms. B. Jiao, Ms. Y.P. Lee, Ms. M. Mishra, Dr. O. Islamand Mr. B. Cher for technical assistance.
References
[1] L. Feng, A. Manavalan, M. Mishra, S.K. Sze, J.M. Hu, K. Heese, Tianma modulatesblood vessel tonicity, Open Biochem. J. 6 (2012) 56–65.
[2] S.M. Francis, M.E. Gas, M.C. Daugeron, J. Bravo, B. Seraphin, Rbg1–Tma46 dimerstructure reveals new functional domains and their role in polysome recruit-ment, Nucleic Acids Res. 40 (2012) 11100–11114.
[3] K. Heese, Y. Nagai, T. Sawada, Nerve growth factor (NGF) induces mRNA expres-sion of the new transcription factor protein p48ZnF, Exp. Mol. Med. 36 (2004)130–134.
[4] K. Heese, T. Yamada, H. Akatsu, T. Yamamoto, K. Kosaka, Y. Nagai, T. Sawada,Characterizing the new transcription regulator protein p60TRP, J. Cell. Biochem.91 (2004) 1030–1042.
[5] L.R. Huo, J.T. Liang, J.H. Zou, L.Y. Wang, Q. Li, X.M. Wang, Possible novel roles ofpoly(rC)-binding protein 1 in SH-SY5Y neurocytes: an analysis using a dynamicBayesian network, Neurosci. Bull. 28 (2012) 282–290.
[6] L.R. Huo, C. Shen, W.N. Ju, J.H. Zou, W. Yan, W.T. Brown, N. Zhong, Identificationof novel partner proteins of PCBP1, Beijing da xue xue bao Yi xue ban 41 (2009)402–408.
[7] K. Ishikawa, T. Akiyama, K. Ito, K. Semba, J. Inoue, Independent stabi-lizations of polysomal Drg1/Dfrp1 complex and non-polysomal Drg2/Dfrp2complex in mammalian cells, Biochem. Biophys. Res. Commun. 390 (2009)552–556.
[8] K. Ishikawa, S. Azuma, S. Ikawa, K. Semba, J. Inoue, Identification of DRG familyregulatory proteins (DFRPs): specific regulation of DRG1 and DRG2, Genes Cells10 (2005) 139–150.
[9] S. Layalle, E. Coessens, A. Ghysen, C. Dambly-Chaudiere, Smooth, a hnRNPencoding gene, controls axonal navigation in Drosophila, Genes Cells 10 (2005)119–125.
[10] J. Liang, J. Wang, A. Azfer, W. Song, G. Tromp, P.E. Kolattukudy, M. Fu, Anovel CCCH-zinc finger protein family regulates proinflammatory activationof macrophages, J. Biol. Chem. 283 (2008) 6337–6346.
[11] N.M. Luscombe, S.E. Austin, H.M. Berman, J.M. Thornton, An overview of thestructures of protein–DNA complexes, Genome Biol. 1 (2000), REVIEWS001.
[12] A.V. Makeyev, S.A. Liebhaber, The poly(C)-binding proteins: a multiplicity offunctions and a search for mechanisms, RNA 8 (2002) 265–278.
[13] M. Mishra, H. Akatsu, K. Heese, The novel protein MANI modulates neurogenesisand neurite-cone growth, J. Cell. Mol. Med. 15 (2011) 1713–1725.
[14] M. Mishra, K. Heese, P60TRP interferes with the GPCR/secretase pathway tomediate neuronal survival and synaptogenesis, J. Cell. Mol. Med. 15 (2011)2462–2477.
[15] M. Mishra, J. Huang, Y.Y. Lee, D.S. Chua, X. Lin, J.M. Hu, K. Heese, Gastrodia elatamodulates amyloid precursor protein cleavage and cognitive functions in mice,Biosci. Trends 5 (2011) 129–138.
[16] M. Mishra, N. Inoue, K. Heese, Characterizing the novel protein p33MONOX,Mol. Cell. Biochem. 350 (2011) 127–134.
[17] M. Mishra, A. Manavalan, S.K. Sze, K. Heese, Neuronal p60TRP expression mod-ulates cardiac capacity, J. Proteomics 75 (2012) 1600–1617.
[18] P. Nahar-Gohad, H. Sultan, Y. Esteban, A. Stabile, J.L. Ko, RACK1 Identified asthe PCBP1-interacting protein with a novel functional role on the regulation ofhuman MOR gene expression, J. Neurochem. 124 (2013) 466–477.
[19] A. Nandal, J.C. Ruiz, P. Subramanian, S. Ghimire-Rijal, R.A. Sinnamon, T.L.Stemmler, R.K. Bruick, C.C. Philpott, Activation of the HIF prolyl hydrox-ylase by the iron chaperones PCBP1 and PCBP2, Cell Metab. 14 (2011)647–657.
[20] S. Nehar, M. Mishra, K. Heese, Identification and characterisation of thenovel amyloid-beta peptide-induced protein p17, FEBS Lett. 583 (2009)3247–3253.
[21] H. Shi, K.Z. Bencze, T.L. Stemmler, C.C. Philpott, A cytosolic iron chaperone thatdelivers iron to ferritin, Science 320 (2008) 1207–1210.




















