
ERP assessment of semantic memory in Alzheimer's disease
14
ELSEVIER INTERNATIONAL JOURNAL OF PSYCHOPHYSIOLOGY International Journal of Psychophysiology 27 (1997) 201-214 ERP assessment of semantic memory in Alzheimer’s disease Marisol Castaiiedaa, Feggy Ostrosky-Solisa’*, Martha PCrez”, Maria Antonieta Bobesb, Luz Esther Rangel’ a Departamento de Psicofisiologia, Far&ad de Psicologia, Universidad National Autdnoma de M&co, Riwra a’e Cupia 11 O- 71, Lomas de Reforma, I 1930 M&co DF, Mexico hlnstituto de Neurociencias, La Habana, Cuba ‘Centro de Atencih Diumo ‘Francisco Espinozn Figueroa i Franz Hab 119, Col Alfonso xiii, Del Alvaro Obregon, Mako DF, Mexico Received 11 February 1997; revised 1 July 1997; accepted 1 July 1997 Abstract This study used the technique of Event-Related Potentials (ERPs), specifically the component N400, to evaluate semantic functions in Alzheimer’s disease (AD). ERPs of 10 patients with probable AD and 10 asymptomatic elderly control subjects were recorded during a semantic categorization task using pictures as stimuli, 50% of the stimuli were congruent and the other 50% were incongruent. Significant differences for the incongruent ERPs, with a diminished N400 component, was observed in the patients group whereas the amplitude of the congruent ERPs was similar in both groups. The characteristics of the N400 in the patients with probable AD suggest that there is a lack of efficiency in accessing information and shows that there are deficient associative connections within the semantic network. 0 1997 Elsevier Science B.V. Keywor&: Event-related potentials; N400; Picture categorization; Semantic memory; Alzheimer’s disease *Corresponding author. Tel.: + 52 5 2517656; fax: + 52 5 5502560; e-mail: [email protected] 0167-8760/97/$17.00 0 1997 Elsevier Science B.V. All rights reserved. PII SO167-8760(97)00064-O
-
Upload
desdeelharem -
Category
Documents
-
view
6 -
download
0
Transcript of ERP assessment of semantic memory in Alzheimer's disease

ELSEVIER
INTERNATIONAL JOURNAL OF PSYCHOPHYSIOLOGY
International Journal of Psychophysiology 27 (1997) 201-214
ERP assessment of semantic memory in Alzheimer’s disease
Marisol Castaiiedaa, Feggy Ostrosky-Solisa’*, Martha PCrez”, Maria Antonieta Bobesb, Luz Esther Rangel’
a Departamento de Psicofisiologia, Far&ad de Psicologia, Universidad National Autdnoma de M&co, Riwra a’e Cupia 11 O- 71, Lomas de Reforma, I 1930 M&co DF, Mexico
hlnstituto de Neurociencias, La Habana, Cuba
‘Centro de Atencih Diumo ‘Francisco Espinozn Figueroa i Franz Hab 119, Col Alfonso xiii, Del Alvaro Obregon, Mako DF, Mexico
Received 11 February 1997; revised 1 July 1997; accepted 1 July 1997
Abstract
This study used the technique of Event-Related Potentials (ERPs), specifically the component N400, to evaluate
semantic functions in Alzheimer’s disease (AD). ERPs of 10 patients with probable AD and 10 asymptomatic elderly control subjects were recorded during a semantic categorization task using pictures as stimuli, 50% of the stimuli were congruent and the other 50% were incongruent. Significant differences for the incongruent ERPs, with a diminished N400 component, was observed in the patients group whereas the amplitude of the congruent ERPs was similar in both groups. The characteristics of the N400 in the patients with probable AD suggest that there is a lack of efficiency in accessing information and shows that there are deficient associative connections within the semantic network. 0 1997 Elsevier Science B.V.
Keywor&: Event-related potentials; N400; Picture categorization; Semantic memory; Alzheimer’s disease
*Corresponding author. Tel.: + 52 5 2517656; fax: + 52 5 5502560; e-mail: [email protected]
0167-8760/97/$17.00 0 1997 Elsevier Science B.V. All rights reserved.
PII SO167-8760(97)00064-O

202 M. Castaiieda et al. /International Journal of Psychophysiology 27 (1997) 201-214
1. Introduction.
Dementia is among the highest occurrences of illnesses of the Central Nervous System in old age, and the most frequent cause of this syn- drome is Alzheimer’s disease (AD) (Cummings and Benson, 1992). AD is a progressive and de- generative disorder of difficult diagnosis, charac- terized by a gradual loss of various cognitive functions, the most characteristic of which are the disruptions in semantic memory (Nebes, 1989).
Memory implies a biological function which allows for the registry, retention and the evoca- tion of information stored at a previous time. Recently, there has been a variety of theoretical models that have divided memory into multiple systems that differ in the nature of the informa- tion they store, how they represent this informa- tion and in the types of operations carried out within each system. One approach to partitioning memory that has had a major effect on the field is the distinction between semantic memory and episodic memory (Tulving, 1984, 1987). Semantic memory consists of an organized body of knowledge which includes words, concepts, mean- ings, associations and the rules for handling these symbols and concepts, whereas episodic memory provides the additional capacity to acquire and retain the knowledge of unique and personal ex- periences or events occurring at a specific time and within a specific context.
According to several studies, it has been stated that episodic memory is impaired both in AD and in normal aging (although to a very different degree), while semantic memory is severely dis- rupted in AD and remains essentially intact in the normal elderly population (see Nebes, 1989 for a review; Ostrosky-Solis et al., 1994). The evidence that AD patients have semantic memory impair- ments not found in the normal elderly comes from the word-finding problems evident in the patient’s spontaneous speech as well as in their performance on tests of verbal fluency and object naming (Weingartner et al., 1981 Bayles and To- moeda, 1983; Martin and Fedio, 1983; Grober et al., 1985; Nicholas et al., 1985; Huff et al., 1986; Ober et al., 1986; Butters et al., 1987; Kempler et
al., 1987). Two hypothesis have been offered to explain these semantic deficits. The first one sug- gests that patients have a procedural deficit and somehow this deficit prevents accessing the con- tents of semantic memory, whereas the second explanation supports the existence of a degraded semantic store where the actual contents of se- mantic memory are degraded (see Chenery, 1995 for a review).
One way to solve the issue of procedural vs. storage deficits, is to examine the performance of patients with Dementia of Alzheimer’s Type (DAT) on semantic priming tasks. With this methodology, it is possible to study the effect that a context has on the processing of a stimulus. Semantic priming may be obtained in normal subjects with tasks using pictures as primes (Irwin and Lupker, 19831, on lexical decision tasks (Meyer et al., 1975) and on word naming tasks (Warren, 1977).
Some studies that have assessed the ability of DAT subjects on semantic priming tasks have reported intact priming (Hartman, 1991; Ober et al., 19911, some no priming (Huff et al., 1988; Ober and Shenaut, 1988; Salmon et al., 19881, some excessive or hyperpriming (Chertkow et al., 1989; Nebes et al., 1989; Balota and Duchek, 1991), some a mixture of both no priming and intact facilitation (Albert and Milberg, 1989) and some a dissociation between intact associative- level priming and disturbed semantic priming (Glosser and Friedman, 1991).
Additional information on the semantic pro- cessing in DAT patients can be obtained by study- ing a component of the Event-Related Potentials (ERPs) called N400. In recent years it has been shown that ERPs are a valuable non-invasive tool for evaluating the neural function of the nervous system linked to sensorial, perceptual and/or cognitive processes. In contrast to behavioral measurements (such as reaction time) the ERPs give a continuous monitoring of cerebral activity during the processing of information. Described by Kutas and Hillyard 1980, 1984), the N400 is a negative deflection produced around 400 ms post-stimulus which has been observed to occur with semantic processing or detection of semantic

M. Castarieda et al. /International Journal of Psychophysiology 27 (1997) 201-214 203
anomalies in sentences with incongruent endings (i.e. ‘I drank coffee with milk and HAM’), while it is not observed in sentences with congruent end- ings (i.e. ‘I drank coffee with milk and SUGAR’). This component has also been presented by using as stimuli pairs of written words (Harbin et al., 19841, spoken words (Neville, 1985) and drawings (Barrett and Rugg, 1990) generating a broader N400 in incongruent situations than in congruent ones. In contrast to semantic anomalies, physi- cally deviant words (e.g. printed in boldface type) elicited a positive potential rather than a negativ- ity. Other non-semantic deviations, such as musi- cal or grammatical violations also failed to elicit the N400 (Besson and Macar, 1987; Osterhout and Holcomb, 1992; Hagoort et al., 1993).
The exact mental operation that is indexed by the N400 is still a matter of controversy. Some authors have viewed the N400 as an index of the ease of lexical access, with more negative respon- ses when less contextual pre-activation of the eliciting word is present (Kutas and Hillyard, 1984). Several psycholinguistic experiments have demonstrated that it is associated with some as- pects of lexical processing especially in relation to the eliciting word meaning. The N400 amplitude has proven to be sensitive to factors that are known to influence the speed of lexical access and word recognition, such as frequency, repetition, semantic priming and semantic context (see Ku- tas and Van Petten, 1994). Another interpretation of the N400 views this waveform as a reflection of post-lexical processes, such as the integration of the products of lexical access with the context (Rugg et al., 1988 Halgren, 1990; Rugg, 1990; Brown and Hagoort, 1993; Holcomb, 1993; Chwilla et al., 1995). This implies that an N400 effect will occur when the lexical integration process is hampered as a result of a mismatch between the semantic specifications of a particu- lar word and the semantic specifications of its preceding word or sentence context.
Up until now, very few studies have used the N400 to assess the semantic abilities in DAT patients. Hamberger et al. (1995) used a lexical decision task with sentences as stimuli and found that the N400 amplitude was identically respon-
sive to semantic relatedness in young normal and DAT groups although DAT patients produced significantly greater number of errors, therefore they concluded that the disruption in semantic processing occurs at some point between the elic- itation of the N400 and the generation of the reaction time response. According to Hamberger et al. (1995) these results reflect both an index of the adequate access and organization of informa- tion in the semantic memory. Thus the alteration must be at the level of the behavioral-response organization.
On another study, Margolin 0987) measured the reaction time of patients with DAT and observed that in contrast to normal elderly sub- jects, the patients did not show semantic priming when the context was the picture of an object, but did show priming when the context was a word rather than a picture. It is known that there are differences between the processing of words and drawings (Potter and Faulconer, 1975), by using pictures the linguistic process of decoding printed words is bypassed, thus the follow-up of these data could contribute with significant information on the limitations of the semantic functioning in DAT patients.
In the present study a group of patients with probable DAT was compared to a group of nor- mal elderly control subjects in a semantic catego- rization task that used pictures of common ob- jects and animals as stimuli. The ERPs associated with picture semantic processing were examined in order to determine if the N400 component was altered in the patients. If alterations in the ampli- tude and/or latency of the N400 were observed, then these abnormalities would be a direct re- flection of the procedural routines in the patients and the situation in the semantic storage. Special attention was directed to the amplitude character- istics of N400. The comparison of N400 amplitude was carried out separately for congruent and in- congruent trials. Specific contrasts were per- formed to establish if the N400 amplitude found in the DAT patients during the semantic catego- rization task, was due to an enhanced negativity for congruent trials, or an enhanced positivity for incongruent trials.

204 M. Castafieda et al. /International Journal of Psychophysiology 27 (1997) 201-214
2. Method
2.1. Subjects
Ten patients with probable DAT (four men and six women; 59-89 years old, mean age 75.4, mean years of education 9.9) who attended the Center for Alzheimer’s Disease ‘Francisco Espinoza Figueroa’ in Mexico City, and 10 normal elderly subjects (five men and five women; 54-82 years old, mean age 67.8, mean years of education 8.6) participated in the study. All subjects were right handed, had normal to corrected vision and none of the patients were taken psychoactive medica- tion. The inclusion criteria for the normal group were: (1) absence of dementia according to the criteria of DSM-IV (American Psychiatric Associ- ation, 1995); (2) a score of at least 23 in the Mini-Mental State Examination (MMS) (Folstein et al., 1975); (3) absence of psychiatric or neuro- logical diseases; and (4) scored within normal range on a neuropsychological battery, that was developed, standardized and validated in a Span- ish speaking population (Ostrosky-Solis et al., 1997). The battery assessed the domains of atten- tion, memory, language, visual perception, visuos- patial process and executive functions. The bat- tery was administered by a trained certified neu- ropsychologist.
The diagnosis of probable DAT was made ac- cording to the criteria of the National Institute of Neurological and Communicative Disorders and Stroke and Alzheimer’s Disease and Related Dis- orders Association (McKhann et al., 1984), on the neuropsychological test results and on the patient’s level of functioning in daily activities. Patients were mildly to moderately impaired, MMS scores were between 14 and 22, and dura- tion of illness was between 6 months and 3 years. On the neuropsychological battery patients scored more than two standard deviations below stan- dardized norms on measures of delay recall of verbal and visual material, on confrontation naming, verbal fluency and on executive func- tions. Other possible sources of dementia were excluded by appropriate laboratory evaluations, such as computed tomography, electroen- cephalography, blood count, serum B12, hepatic,
renal and thyroid function test. Their scores on the Hachinski Ischemic Rating Scale (Hachinski et al., 1975) were used to aid in excluding multin- farct dementia.
2.2. Material
The stimuli consisted of 118 pairs of drawings of objects and animals (Snodgrass and Vander- wart, 1980) which had been adapted in denomina- tion, familiarity and imaginability in the Mexican population (Aveleyra et al., 1996). The figures, which were 8 cm’, were presented in a computer monitor. Since subjects sat approximately 1 m away from the monitor, the drawings sustained a vertical and horizontal angle of 4.6”. Fifty percent of the pairs were semantically related and consti- tuted the congruent condition (they could be cat- egorized in a supraordinated group, i.e. zebra-deer), while the other 50% of the pairs had no semantic relation and formed the incongruent condition (i.e. trumpet-shoe).
2.3. Procedure
The task of the subjects was to discriminate between the congruent and incongruent pairs of pictures. During the experiment, the subjects sat in a comfortable chair in front of the computer monitor and were asked to minimize body and eye movements. They were instructed about the task with a training phase which consisted of 10 pairs of drawings not present in the task of ERPs recordings. The first figure of the pair served as the context for the second figure, which was the stimulus on which ERPs were registered. The electroencephalographic recording was synchro- nized with the presentation of the second drawing and the following sequence of events composed each test: the letter string XXXX was presented as a warning stimuli, the first picture (Sl) was presented for 1 s, followed by a 180-ms blackout, following the blank interval a second picture (S2) was presented (1 s duration) and the screen was again blanked. After the blank screen, the word ‘RESPONDA’ (answer) appeared and the sub- jects responded verbally (yes or no> if the stimu- lus (S2) was related to the previous one (Sl). The

M. Castaieda et al. / IntemationalJournal of Psychophysiology 27 (1997) 201-214 205
Stimulus Onset Asynchrony (SOA) between onset of Sl and onset of S2 was therefore equal to 1180 ms. The response was delayed until EEG record- ing was finished in order to reduce artifacts, thus reaction time was not measured. Average ERPs were obtained only for stimuli S2 in both congru- ent and incongruent conditions.
2.4. Electrophysiological recording
Electrophysiological data acquisition and analy- sis were carried out on a Neuroscan System. Scalp electrical activity was recorded from 32 monopolar derivations according to the lo-20 International System. An electrode cap (Electra-Cap Interna- tional) was used. All electrodes were referenced to linked ear lobes and an additional bipolar derivation recorded EOG placed at the inner and outer canthi of the right eye. Electrode impedance was always below 5 Kfl. The signal was filtered between 0.5 and 30 Hz (3 dB down). In each trial 256 points of digitized EEG (12 bit resolution) were recorded at a sampling rate of 256 Hz, totalling 1 s and stored on a magnetic disk for off-line analysis. A pre-stimulus baseline of 100 ms was obtained in each trial and data acquisition continued 900 ms after stimulus onset. Only trials on which a correct behavioral response was made were used to form the averages. Also trials on which EOG activity exceeded a peak-to-peak am- plitude of +50 to -50 pV, or on which satura- tion of one or more channels of the analog-dig- ital converter occurred, were also excluded (ap- prox. 15% of trials were lost due to such artifacts).
For every subject, averaged ERPs for each recording site were obtained for each stimulus condition. Average ERPs at the congruent condi- tion and incongruent condition were 29 trials for both groups. These ERPs were submitted to low- pass digital (zero phase distortion) filtering with an upper cut of 5.5 Hz. All data points were corrected (prior to plotting or measurement) by subtracting the average pre-stimulus amplitude value. In other words, all amplitude values were measured with respect to the average amplitude of the pre-stimulus value corresponding to each ERP.
2.5. Statistical analysis
Discrimination behavior was analyzed using the Signal Detection Theory measures d’ and the log,, of /3 @wets, 1964). The use of d’ permits an assessment of the accuracy of performance with- out contamination from variations in criterion (response bias), which could be important between controls and patients. Both behavioral and ERPs measured were submitted to analysis of variance (ANOVA). For the ERPs measured, 11 electrode sites where the negativity was largest were se- lected (Fz, Cz, Pz, Fc3, Cp3, T3, Tp7, Fc4, Cp4, T4 and Tp8). For some analysis, homologous left and right electrodes were collapsed as Anterior to Posterior sites (frontal, central, parietal) and Coronal sites (left, midline, right). Amplitude and latency values were subjected to repeated mea- sures ANOVA using Group (normal elderly and DAT patients) as between subject variable and two within subject variable conditions (congruent vs. incongruent) and Anterior to Posterior (fron- tal, central, parietal) or Coronal (left, midline, right). The Greenhouse-Geisser procedure was used, when appropriate, to mitigate violations of sphericity assumptions in the repeated measures design (Keselman and Rogan, 1980). When neces- sary significant interactions were reassessed with repeated measures ANOVAs of normalized am- plitude as outlined by McCarthy and Wood (1985); only interactions that were significant on the nor- malized amplitude measures are reported.
3. Results
3.1. Behavior
Both groups were able to discriminate between congruent and incongruent pictures, as reflected by the mean d’ (Table 1) which was significantly different from zero (t-tests, P < 0.001). The mean d’ differed between groups, as confirmed by an
ANOVA, Fi i&l = 10.04, P < 0.0005. The mean percent of hits was very high for the normal elderly group and lower for the DAT group but always above 50%, these data differed between groups as confirmed by an ANOVA, Fl.18 = 14.43,

206 M. Castaiieda et al. /International Journal of Psychophysiology 27 (1997) 201-214
Table 1 Behavioral results (d’, Hits % and log,, p) obtained in the semantic matching task in both groups studied
d’
Hits (% Congr.) Hits (% Incon.)
IJJg,o P
Elderly
4.32 ( f 0.47)
94.3 ( * 7.9)
95.4 (k6.1)
- 2.43 (k4.69)
DAT
2.55 (* 1.21)
78.7 (+ 10.85)
84.2 (* 15.1)
0.21 tf3.6)
P < 0.0005. Response bias (log,, p > is also shown in Table 1, and was not statistically different among the two groups.
3.2. Event related potentials (ERPs)
The morphology of the waveform was charac- terized by an early negativity, with a peak latency of approximately 100 ms, followed by a positive deflection peaking at approximately 200 ms. This deflection was followed by a second negative de- flection, broadly distributed across the scalp, peaking at approximately 300-570 ms (N400), that was larger for incongruent than congruent pic- tures and finally a positive deflection at approxi- mately 600 ms.
No significant effects in the amplitude or la- tency of ERPs were found before 300 ms or at 600 ms for any of the factors, thus a direct expla- nation will be given of what was observed in the N400. In the normal elderly group, after 300 ms, the ERPs related to the incongruent trials gener- ated a substantial negativity which was not observed in the congruent trials. The N400 observed was generated with greater amplitude in centro-parietal regions, with greater activity in the right hemisphere than in the left (Fig. 1). In the DAT group, a substantial reduction in the amplitude of this component was observed and they showed no gradient of significant activity (Fig. 2).
The N400 effect (the mean amplitude of the difference waveforms obtained by subtracting the incongruent trials from the congruent trials) was calculated in the 300~580-ms time window and
adjusted for each group in its respective N400 peak. That is, between 333 and 571 ms for the normal elderly group and between 353 and 531 ms in the DAT group. The negativity of the N400 appears in these different waveforms as a defined peak in the normal elderly group and as a dimin- ished peak in the DAT group as can be seen in Fig. 3.
These data were analyzed applying an ANOVA with Group (normal elderly and DAT patients) as between subject variable and two within subject variable Anterior to Posterior (frontal, central, parietal) and Coronal (left, midline, right). A sig- nificant main effect of Group (Fols) = 4.95, P < O.OS>, Anterior to Posterior (F2,36 = 7.71, P < 0.005) and Coronal (FZ,36 = 11.04, P < 0.005) fac- tors were found reflecting the fact that the N400 was largest in the normal elderly group than in DAT group in all the leads analyzed. Statistical analysis also revealed significant differences between the normal elderly group and the DAT group in the mean peak latency of the N400 effect (Fi,is = 3.89, P < 0.05). In the normal el- derly group, the mean peak latency was 472 f 56.4 ms whereas in the DAT group the mean peak latency was at 528 + 42 ms.
The analysis of the scalp distribution of the N400 effect for each group showed striking dif- ferences in distribution between groups. For the normal elderly group, the maximum negativity was more posterior (main effect of Anterior to Posterior: F2,36 = 15.75, P < 0.001) involving the parietal region, and it was clearly displaced to- wards the right hemisphere (main effect of Coro-
nal (E;O,,O = 8.42, P < O.OOS), while the electroen- cephalographic activity of DAT group did not reflect significant topographic effects.
The mean amplitude of the congruent and in- congruent recordings for the time window from 300 to 580 ms is shown in Table 2 for both groups studied.
As exemplified in Fig. 4, wave morphology on the N400 amplitude values corresponding to the congruent trials were similar in both groups, whereas the wave morphology on the N400 ampli- tude values corresponding to the incongruent tri- als for the DAT group were clearly more positive than those of the normal elderly group. Thus the
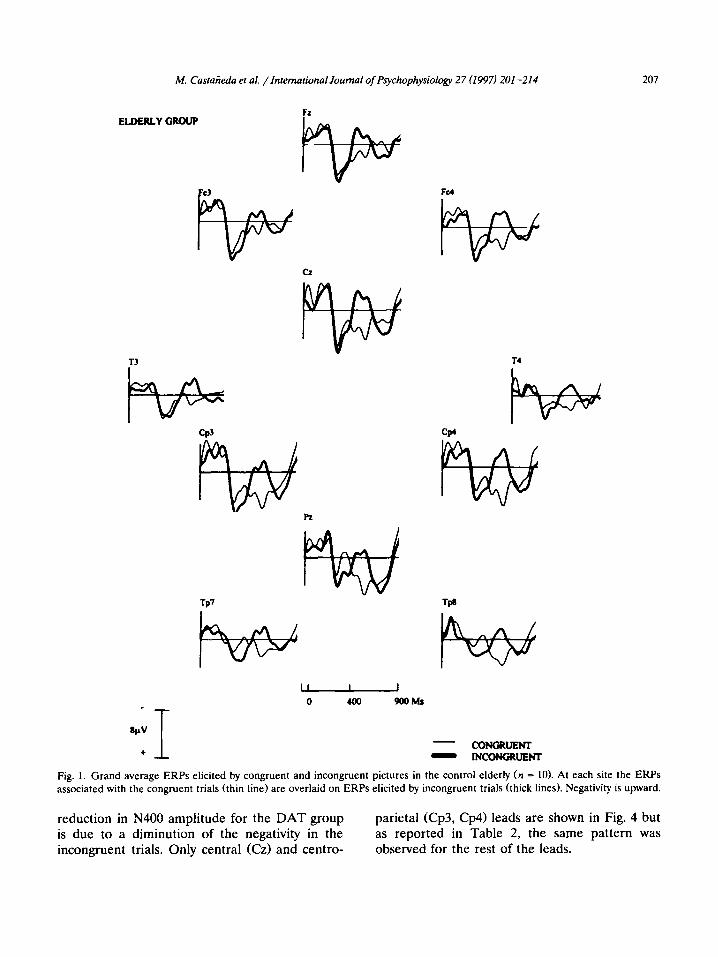
M. Castarieda et al. /International Journal of Pychophysiology 27 (1997) 201-214 207
TP’
W
+ I - coNauJENr - IN-UENT
Fig. 1. Grand average ERPs elicited by congruent and incongruent pictures in the control elderly (n = 10). At each site the ERPs associated with the congruent trials (thin line) are overlaid on ERPs elicited by incongruent trials (thick lines). Negativity is upward.
reduction in N400 amplitude for the DAT group is due to a diminution of the negativity in the incongruent trials. Only central (Cz) and centro-
parietal (Cp3, Cp4) leads are shown in Fig. 4 but as reported in Table 2, the same pattern was observed for the rest of the leads.
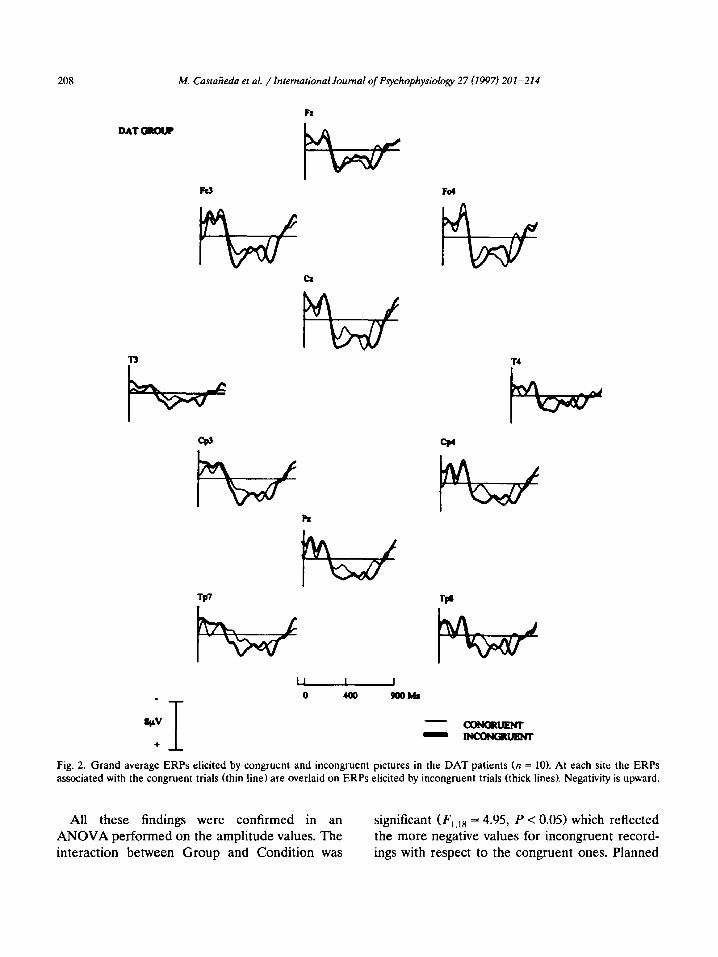
208 M. Castatieda et al. / IntemationaiJoumnl of Psychophysiology 27 (1997) 201-214
Fd
11 I I
* 0 400 9ooh4
W -- -INcoN(IRuEM
+ I
Fig. 2. Grand average ERPs elicited by congruent and incongruent pictures in the DAT patients (n = 10). At each site the ERPs associated with the congruent trials (thin line) are overlaid on ERPs elicited by incongruent trials (thick lines). Negativity is upward.
All these findings were confirmed in an significant (F,,,, = 4.95, P < 0.05) which reflected
ANOVA performed on the amplitude values. The the more negative values for incongruent record- interaction between Group and Condition was ings with respect to the congruent ones. Planned
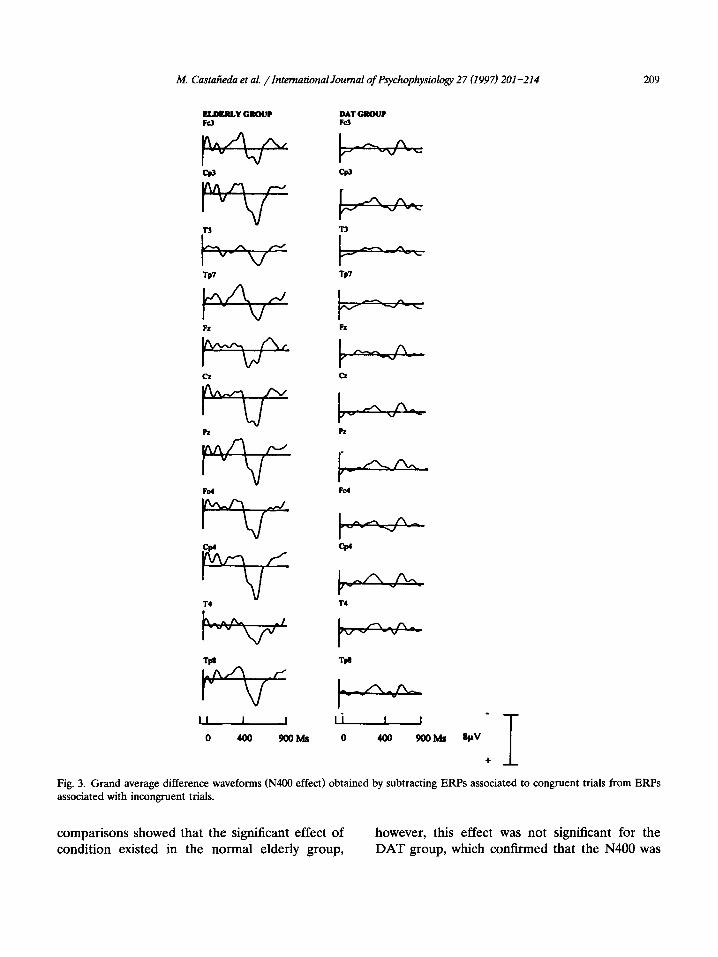
M. Castaiieda et al. /International Journal of Pqvchophysiology 27 (1997) 201-214 209
Fig. 3. Grand average difference waveforms (N400 effect) obtained by subtracting ERPs associated to congruent trials from ERPs associated with incongruent trials.
comparisons showed that the significant effect of condition existed in the normal elderly group,
however, this effect was not significant for the DAT group, which confirmed that the N400 was

210 M. Castaiieda et al. /International Journal of Psychophysiology 27 (1997) 201-214
Table 2
Mean amplitude and standard deviation (in parenthesis) of
the congruent and incongruent recordings for both groups
studied at the 11 electrodes sites selected
Fc3
Fc4
Fz
CP3
CP4 cz
T3
T4
Pz
TP~
TP8
Congruent Incongruent
Elderly DAT Elderly DAT
0.7 (2.1) 2.7 (1.2) 0.1 (1.3) 2.6 (1.7)
1.2 (2.3) 2.5 (1.5) - 0.34 (1.2) 2.2 (1.8)
0.8 (2.1) 2.2 (1.3) - 0.63 (1.6) 2.0 (1.5)
1.9 (2.2) 1.9 (1.3) - 0.03 (1.9) 1.5 (2.0)
2.0 (3.0) 2.3 (1.5) - 0.68 (1.6) l.l(l.8)
1.4 (2.6) 1.9 (1.2) - 0.51(2.1) 1.2 (2.2)
0.3 (1.4) 2.5 (1.6) - 0.05 (1.4) 2.7 (1.7)
0.9 (2.1) 0.9 (0.7) -0.40 (1.1) 1.2 (1.6)
1.7 (2.4) 1.1 (1.1) - 0.28 (1.5) 1.1 (1.3)
0.6 (1.9) 1.2 (1.2) - 0.27 (1.3) 1.2 (1.1)
1.5 (2.7) 1.4 (0.8) -0.17 (1.7) 1.3 (1.0)
diminished in this group. Comparisons done a posteriori confirmed that there were no signifi- cant differences in the congruent trials between the normal elderly group and the DAT group but there were significant differences for the incon- gruent trials (Fr,rs = 9.33, P < 0.05).
4. Discussion
The behavioral results indicated that the DAT group committed more errors than the normal elderly group in the categorization task. It has also been observed in various studies that patients with DAT are less efficient than the control sub- jects in semantic tasks using both words and pictures (Bayles and Tomoeda, 1983; Huff et al., 1986; Ober et al., 1986; Nebes, 1989). Picture matching is assumed to share with word matching the same mechanisms involving semantic memory and response organization. The fact that catego- rization is faster with pictures than with words has led to the hypothesis that pictures are more directly connected to semantic memory (Potter and Faulconer, 1975). Thus a possible explanation of the impairments observed in semantic match- ing for both words and pictures is obtained under the hypothesis that the underlying deficiency is located either in the use of the semantic memory or in the response organization, discarding the theory that the abnormal pairing of pictures arise
from difficulties in identifying the stimulus. This pattern of results has been confirmed by other studies suggesting that the mistakes made by the patients probably arise from deficiencies subse- quent to accessing the information stored in the memory and/or in integrating the information evoked by the context.
It has been theorized that the N400 is sensitive to semantic processing and that the amplitude of this component varies inversely according to the quantity of activation offered by the semantic context (Kutas and Hillyard, 1980, 1984). In the present study a reduction of the N400 component in the DAT group was observed. This is a result that conforms to prior studies of the N400 com- ponent which have used semantic matching tasks in other types of pathologies, such as schizophre- nia and aphasia (Babes et al., 1996; Swaab et al., 1997), in which the reduced amplitude and de- layed latency of this component indicates alter- ations at the level of the semantic storage. An aspect that was not sufficiently discussed in previ- ous papers is the basis of this reduction, which could be due to either an increased positivity in incongruent trials (smaller N400 in incongruent trials), an increased negativity in congruent trials (larger N400 in congruent trials) or both. In this study the amplitude measurements in the N400 time window were similar in the normal elderly group and the DAT group for congruent trials, however, at incongruent trials the amplitude was significantly more positive for the DAT group relative to the normal elderly group. Thus, the results of this study indicate that the DAT group reflected both normal and abnormal patterns in ERPs. On one hand, they showed normal ERPs for congruent trials, but on the other, they de- monstrated abnormal ERPs for incongruent tri- als.
The interpretation of this pattern of results depends on the type of hypothesis about the functional meaning of N400 that is accepted. If N400 is considered to reflect pre-lexical priming (Kutas and Hillyard, 1984), then these results suggest that in patients with DAT, less priming is produced by incongruent stimuli, since the amount of negativity should be inversely related to the amount of contextual preactivation. This would
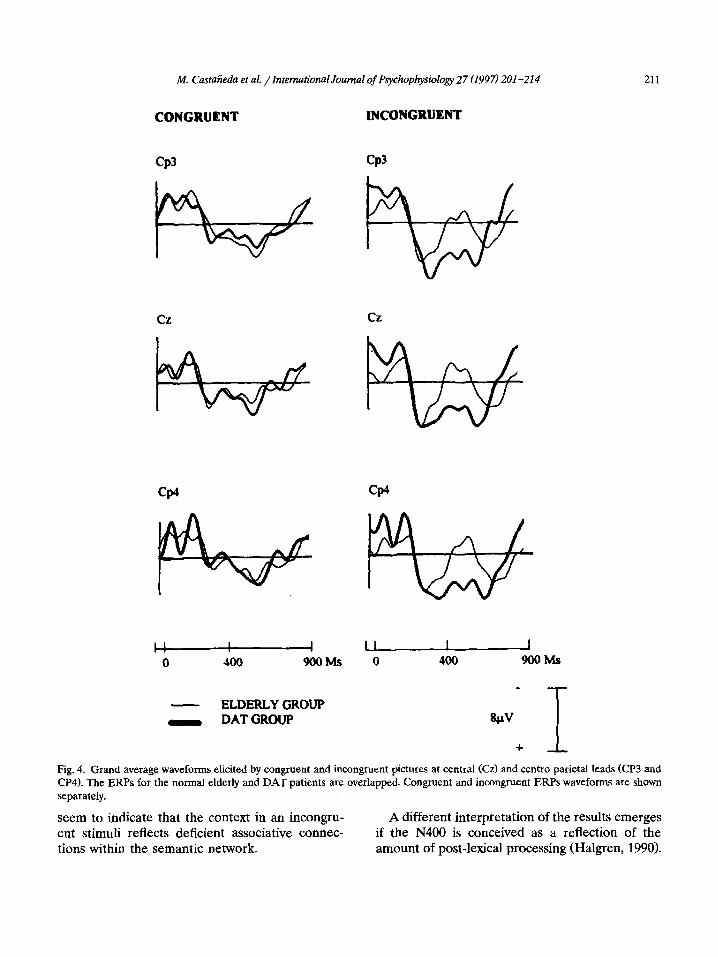
M. Castafieda et al. /International Journal of Psychophysiology 27 (1997) 201-214 211
CONGRUENT
CP3
iNCONGRUENT
CP3
w
I ! I I 0 400 9OoMs
- ELDERLY GROUP - DATGROUP
11 I I 0 400 9ooMs
Fig. 4. Grand average waveforms elicited by congruent and incongruent pictures at central (Cz) and centro parietal leads (CP3 and CP4). The ERPs for the normal elderly and DAT patients are overlapped. Congruent and incongruent ERPs waveforms are shown separately.
seem to indicate that the context in an incongru- ent stimuli reflects deficient associative connec- tions within the semantic network.
A different interpretation of the results emerges if the N400 is conceived as a reflection of the amount of post-lexical processing (Halgren, 1990).

212 M. Castaiieda et al. /International Journal of Psychophysiology 27 (1997) 201-214
Under this hypothesis, the results obtained in the DAT group suggest that the incongruent stimuli required equivalent amounts of post-lexical pro- cessing as in the congruent stimuli, since the amount of amplitude in both cases is similar. In contrast, the normal elderly group seemed to require less processing for the congruent stimuli since the N400 associated with this condition is smaller than the N400 associated with the incon- gruent stimuli.
Whatever explanation of the N400 is accepted, the N400 pattern observed in the DAT group shows that the processing of the incongruent sti- muli is not facilitated within the semantic net- work as in the normal elderly group. The dif- ficulty of patients with probable DAT in the pro- cessing of incongruent stimuli could be due to the damage in the cortical association areas. Neuro- pathological studies have shown that DAT patients showed marked pathology in temporal, parietal and frontal association cortices. The se- nile plaques and neurofibrillary tangles of DAT prominently involve the origins and termination of long cortico-cortical associative fibers of the brain (Brun, 1988; Morrison et al., 1986; Lewis et al., 1987) and neurophysiological studies have found that there are changes in brain functional connectivity in DAT and that EEG decrease between parietal and frontal areas due to degen- eration of fibers that connect these two areas (Leuchter et al., 1992). It has been suggested that DAT is a ‘neocortical disconnection syndrome’, where associative areas lose afferent and efferent connections through white matter tracts. This dis- connection occurs at the cortical level and is associated with the death of pyramidal neurons that provide corticocortical projections (Pearson et al., 1985; Lewis et al., 1987; Leuchter et al., 1992). These neuropathological characteristics can lead to the deterioration in the associative net- work, which forms the structural skeleton of the semantic memory. The findings of the present study are consistent with a lack of efficiency in accessing information and show that there are deficient associative connections within the se- mantic network. The results of this study indicate that there is an alteration in the structure and
organization of the semantic memory in patients with DAT.
Acknowledgements
This research was partially supported by a grant given to Dr. Ostrosky-Solis by DGAPA (IN- 2019941, National University of Mexico.
References
Albert, M., Milberg, W., 1989. Semantic processing in patients with Alzheimer’s disease. Brain Lang. 37, 163-171.
American Psychiatric Association, 1995. Diagnostic and Statis- tical Manual of Mental Disorders (DSM-IV). Mason, Barcelona.
Aveleyra, E., Gbmez, C., OstrosI@olis, F., Rigalt, C., Cruz, F., 1996. Adaptation de 10s estimulos no verbales de Snod- grass y Vanderwart en poblaci6n hispanohablante: criterios para la denominackjn, concordancia de la imagen, famil- iaridad y complejidad visual. Rev. Mex. Psicol. 13, 5-19.
Balota, D.A., Duchek, J.M., 1991. Semantic priming effects, lexical repetition effects and contextual disambiguation ef- fects in healthy aged individuals and individuals with senile dementia of the Alzheimer type. Brain Lang. 40, 181-201.
Barrett, S., Rugg, M., 1990. Event-related potentials and the semantic matching of pictures. Brain Cogn. 14, 201-212.
Bayles, D., Tomoeda, C., 1983. Confrontation naming in de- mentia. Brain Lang. 19, 98-114.
Besson, M., Macar, F., 1987. An event-related potential analy- sis of incongruity in music and other nonlinguistic contexts. Psychophysiology 24, 14-25.
Brown, C., Hagoort, P., 1993. The processing nature of the N400: evidence from masked priming. J. Cogn. Neurosci. 5, 34-44.
Butters, N., Granholm, E., Salmon, D., Grant, E., Wolfe, J., 1987. Episodic and semantic memory: a comparison of amnesic and demented patients. J. Clin. Exp. Neuropsy- chol. 9,479-497.
Bobes, M.A., Lei, Z.X., Ibaiiez, S., Yi, Hou., Valdes-Sosa, Mitchell., 1996. Semantic matching of pictures in schizophrenia: a cross-cultural ERPS study. Biol. Psychol. 37, 1-14.
Brun, A., 1988. White matter disease in the elderly demented. Bull. Clin. Neurosci. 53, 125-127.
Chenery, H., 1995. Semantic priming in Alzheimer’s dementia. Aphasiology 10, l-20.
Chertkow, H., Bub, D., Seidenberg, M., 1989. Priming and semantic memory loss in Alzheimer’s disease. Brain Lang. 36,420~446.
Chwilla, D., Brown, C., Hagoort, P., 1995. The N400 as a function of the level or processing. Psychophysiology 32, 274-285.
Cummings, J., Benson, F., 1992. Dementia: A Clinical Ap- proach. Plenum Press, USA.

M. Castai’iedu et al. /International Journal of Psychophysiology 27 (1997) 201-214 213
Folstein, M., Folstein, S., McHugh, P., 1975. ‘Mini mental Rosenberg-Thompson, S., Lachenbrunch, P.A., 1992. state’. A practical method for grading the cognitive state of Changes in brain functional connectivity in Alzheimer-type patients for the clinician. J. Psychiatr. Res. 12, 189-198. and multi-infarct dementia. Brain 115, 1543-1561.
Glosser, G., Friedman, R., 1991. Lexical but not semantic Lewis, D.A., Campbell, M.J., Terry, R.D., Morrison, J.H., priming in Alzheimer’s disease. Psychol. Aging 6, 522-527. 1987. I-aminar and regional distributions of neurofibrillary
Grober, E., Buschke, H., Kawas, C., Fuld, P., 1985. Impaired tangles and neuritic plaques in Alzheimefs disease: a quan- ranking of semantic attributes in dementia. Brain Lang. 26, titative study of visual and auditory cortices. J. Neurosci. 7, 276-286. 1799-1808.
Hachinski, V., Illif, L., Phil, M., et al., 1975. Cerebral blood flow in dementia. Arch. Neurol. 32, 632-637.
Hagoort, P., Brown, C., Groothusen, J., 1993. The syntactic positive shift (SPS) as an ERPS measure of syntactic pro- cessing. Lang. Cognit. Process. 8, 439-483.
Halgren, E., 1990. Insights from evoked potentials into the neuropsychological mechanisms of reading. In: Schiebel, A.B., Wechsler, A.F. (Eds.), Neurobiology of Higher Cogni- tive Function, Guilford, New York, pp. 103-150.
Hamberger, M., Friedman, D., Ritter, W., Rosen, J., 1995. Event-related potential and behavioral correlates of se- mantic processing in Alzheimer’s patients and normal con- trols. Brain Lang. 45, 33-68.
Martin, A, Fedio, P., 1983. Word production and comprehen- sion in Alzheimer’s disease: the breakdown of semantic knowledge. Brain Lang. 19, 124-141.
Margolin, D.I., 1987. Lexical Priming by Pictures and Words in Aging, Stroke and Dementia (Doctoral dissertation, LJni- versity of Oregon). Dissertation Abstracts International, 49, 1416B.
McCarthy, G., Wood, C.C., 1985. Scalp distributions of event related potentials: an ambiguity associated with analysis of variance models. Electroencephalogr. Clin. Neurophysiol. 62,203-208.
Harbin, T., Marsh, G., Harvey, M., 1984. Differences in the late components of the event related potentials due to age and to semantic and non-semantic tasks. Electroen- cephalogr. Clin. Neurophysiol. 59, 489-496.
Hartman, M., 1991. The use of semantic knowledge in AIzheimer’s disease: evidence for impairments of attention. Neuropsychologia 29,213-228.
Holcomb, P., 1993. Semantic priming and stimulus degrada- tion: Implications for the role of the N400 in language processing. Psychophysiology 30, 47-61.
Huff, F., Corkin, S., Growden, J., 1986. Semantic impairment and anomia in Alzheimer’s disease. Brain Lang. 28, 235-249.
McKhann, G., Dracchman, ,.D$, Folstein, M., Katzman, R., Price, D., Stadlan, E., 1984. Clinical diagnosis of Alzheimer’s disease: report of the NINCDS-ADRDA WORK group under the auspices of the department of health and human services task force on Alzheimer’s disease. Neurology 34, 939-1139.
Meyer, D., Schvaneveldt, R., Ruddy, M., 1975. Loci of contex- tual effects on visual word recognition. In: Rabbit, P.M.A., Domic, S. (Eds.), Attention and Performance V. Academic Press, NY.
Huff, F., Mack, L., Mahlmann, J., Greenberg, S., 1988. A comparison of lexical-semantic impairments in left-hemi- sphere stroke and Alzheimer’s disease. Brain Lang. 34, 262-278.
Irwin, D., Lupker, S., 1983. Semantic priming of pictures and words: a levels of processing approach. J. Verb. Learn. Verb. Behav. 22,45-60.
Kempler, D., Curtiss, S., Jackson, C., 1987. Syntactic preserva- tion in Alzheimer’s disease. J. Speech Hear. Res. 30, 343-350.
Morrison, J., Scherr, S., Lewis, D., Campbell, M., Bloom, F., Rogers, J., 1986. The laminar and regional distribution of neocortical somatostatin and neuritic plaques: implications for Alzheimer’s disease as a global neocortical disconnec- tion syndrome. In: Scheibel, A., Wechsler, A., Brazier, M. (Eds.), The Biological Substrates of Alzheimer’s Disease. Academic Press, Orlando, FL, pp. 115-131.
Nicholas, M., Obler, L., Albert, M., Helm-Estrabrooks, N., 1985. Empty speech in Alzheimer’s disease and fluent aphasia. J. Speech Hear. Res. 28, 405-410.
Nebes, R., 1989. Semantic memory in Alzheimer’s disease. Psychol. Bull. 106, 377-394.
Kesehnan, H.J., Rogan, J.C., 1980. Repeated measure F test and psychophysiological research: controlling the number of false positives. Psychophysiology 17, 499-503.
Kutas, M., Hillyard, S., 1980. Reading senseless sentences: brain potentials reflect semantic anomaly. Science 225, 1168-1170.
Nebes, R., Brady, C., Huff, F., 1989. Automatic and attentio- nal mechanisms of semantic priming in Alzheimer’s dis- ease. J. Clin. Exp. Neuropsychol. 11, 219-230.
Neville, H., 1985. Biological constrains on semantic process- ing: a comparison of spoken and signed languages. Psy- chophysiology 22, 576.
Kutas, M., Hillyard, S., 1984. Brain potentials during reading reflect word expectancy and semantic association. Nature 307, 161-163.
Kutas, M., Van Petten, C., 1994. Psycholinguistics Electrified. Event-related Brain Potential Investigations. Handbook of Psycholinguistics. Academic Press, pp. 83-141.
Leuchter, A.F., Newton, T.F., Cook, IA., Walter, D.O.,
Ober, B., Dronkers, N., Koss, E., Delis, D., Friedland, R., 1986. Retrieval from semantic memory in Alzheimer-type dementia. J. Clin. Exp. Neuropsychol. 8, 75-92.
Ober, A., Shenaut, G., 1988. Lexical decision and priming in Alzheimer’s disease. Neuropsychologia 26, 600-647.
Ober, B., Shenaut, G., Jagust, W., Stillman, R., 1991. Auto- matic semantic priming with various category relations in Alzheimer’s disease and normal aging. Psychol. Aging 6, 647-660.

214 M. Castaiieda et al. /International Journal of Psychophysiology 27 (1997) 201-214
Ostethout, L., Holcomb, P., 1992. Event-related brain poten- Lexical and semantic priming deficits in patients with tials elicited by syntactic anomaly. J. Mem. Lang. 31, Alzheimer’s disease. J. Clin. Exp. Neutopsychol. 10, 78.5-806. 177-194.
Ostrosky-Solis, F., Ardila, A., Rosselli, M., 1997. Evaluation Neuropsicologica Breve en Espafrol: NEUROPSI. BAYER, Mexico.
Ostrosky-Solis, F., Rodriguez, Y., Garcia de la Cadena, C., et al., 1994. Marcadores mntsicos de1 envejecimiento normal y patologico. Rev. Mex. Pensamiento Lenguaje. 1, 367-375.
Pearson, R., Esiri, M., Hiorns, R., Wilcock, G., Powell, T., 1985. Anatomical correlates of the distribution of the pathological changes in the neocortex in Alzheimet dis- ease. Proceedings of the National Academy of Sciences of the USA, 82, pp. 4531-4534.
Snodgrass, J., Vandetwart, M., 1980. A standardized set of 260 pictures: norms for name agreement, image agreement, familiarity and visual complexity. J. Exp. Psychol. Hum. Learn. Mem. 6, 174-215.
Swaab, T., Brown, C., Hagoort, P., 1997. Spoken sentence comprehension in Aphasia: Event-related potential evi- dence for a lexical integration deficit. J. Cognit. Neurosci. 9, 36-66.
Potter, M., Faulconer, B., 1975. Time to understand pictures and words. Nature 253, 437-438.
Rugg, M., 1990. Event related potentials dissociate repetition effects of high and low frequency words. Mem. Cognit. 18, 367-379.
Swets, J.A., 1964. Signal Detection and Recognition by Hu- man Observers. Wiley, New York.
Tulving, E., 1984. How many memory systems are there?. Am. Psychol. 40, 385-398.
Tulving, E., 1987. Multiple memory systems and conscious- ness. Hum. Neurobiol. 6, 67-80.
Rugg, M., Furda, J., Lorist, M., 1988. The effects of task on the modulation of event-related potentials by word repeti- tion. Psychophysiology 25, 55-63.
Salmon, D., Shimamuta, A., Butters, N., Smith, S., 1988.
Warren, R., 1977. Time and the spread of activation in me- mory. J. Exp. Psychol. Hum. Learn. Mem. 3,458-466.
Weingartnet, H., Kaye, W., Smallberg, S., Ebert, M., Gillin, J., Sitaram, N., 1981. Memory failures in progressive idio- pathic dementia. J. Abnorm. Psychol. 90, 187-196.




















