
Epizootic hemorrhagic disease virus serotype 7 in European cattle and sheep: Diagnostic...
9
Epizootic hemorrhagic disease virus serotype 7 in European cattle and sheep: Diagnostic considerations and effect of previous BTV exposure Michael Eschbaumer a,1, *, Kerstin Wernike a,1 , Carrie A. Batten b , Giovanni Savini c , Lorraine Edwards b , Annapia Di Gennaro c , Liana Teodori c , Chris A.L. Oura b , Martin Beer a , Bernd Hoffmann a a Institute of Diagnostic Virology, Friedrich-Loeffler-Institut, Suedufer 10, 17493 Greifswald – Insel Riems, Germany b Institute for Animal Health, Pirbright Laboratory, Ash Road, Pirbright, Woking, Surrey GU24 0NF, United Kingdom c Istituto ‘‘G. Caporale’’, Via Campo Boario, 64100 Teramo, Italy 1. Introduction Epizootic hemorrhagic disease virus (EHDV), an arthro- pod-borne orbivirus (family Reoviridae), is an emerging pathogen of wild and domestic ruminants with at least seven distinct serotypes (Anthony et al., 2009; Savini et al., 2011). It is transmitted by blood-feeding Culicoides midges (Diptera: Ceratopogonidae). EHDV had first been described in the United States in the middle of the 20th century, but has since been found to occur in many parts of the world. It most severely affects deer; sheep and goats are not considered to be susceptible, while cattle are potential reservoir hosts (reviewed in Savini et al., 2011). Of particular concern to the livestock industry is Ibaraki virus, an EHDV serotype 2 (EHDV-2) strain from Japan that is known to cause disease in cattle. While the Ibaraki strain had long been considered unique in this regard, cattle were affected by outbreaks of diverse strains of EHDV on the island of La Re ´ union (Bre ´ ard et al., 2004) and more recently in Northern Africa, the Near East and Asia Minor (reviewed in Yadin et al., 2008). Whether this was precipitated by an Veterinary Microbiology 159 (2012) 298–306 A R T I C L E I N F O Article history: Received 14 January 2012 Received in revised form 30 March 2012 Accepted 12 April 2012 Keywords: Epizootic hemorrhagic disease virus (EHDV) Bluetongue virus (BTV) Experimental infection Viremia Interference Sheep Cattle A B S T R A C T Epizootic hemorrhagic disease virus (EHDV), an arthropod-borne orbivirus (family Reoviridae), is an emerging pathogen of wild and domestic ruminants that is closely related to bluetongue virus (BTV). The present study examines the outcome of an experimental EHDV-7 infection of Holstein cattle and East Frisian sheep. Apart from naı¨ve animals that had not been exposed to BTV, it included animals that had been experimentally infected with either BTV-6 or BTV-8 two months earlier. In addition, EHDV-infected cattle were subsequently challenged with BTV-8. Samples were tested with commercially available ELISA and real-time RT-PCR kits and a custom NS3-specific real-time RT-PCR assay. Virus isolation was attempted in Vero, C6/ 36 and KC cells (from Culicoides variipennis), embryonated chicken eggs and type I interferon receptor-deficient IFNAR / mice. EHDV-7 productively infected Holstein cattle, but caused no clinical signs. The inoculation of East Frisian sheep, on the other hand, apparently did not lead to a productive infection. The commercial diagnostic kits performed adequately. KC cells proved to be the most sensitive means of virus isolation, but viremia was shorter than 2 weeks in most animals. No interference between EHDV and BTV infection was observed; therefore the pre-existing immunity to some BTV serotypes in Europe is not expected to protect against a possible introduction of EHDV, in spite of the close relation between the viruses. ß 2012 Elsevier B.V. All rights reserved. * Corresponding author. Tel.: +49 38351 71280. E-mail address: michael.eschbaumer@fli.bund.de (M. Eschbaumer). 1 Both authors contributed equally. Contents lists available at SciVerse ScienceDirect Veterinary Microbiology jou r nal h o mep ag e: w ww .els evier .co m/lo c ate/vetm ic 0378-1135/$ – see front matter ß 2012 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.vetmic.2012.04.020
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Epizootic hemorrhagic disease virus serotype 7 in European cattle and sheep: Diagnostic...

Esh
MLoMa Inb Inc Is
1.
popase20(D
Veterinary Microbiology 159 (2012) 298–306
A
Art
Re
Re
Ac
Ke
Ep
Blu
Ex
Vir
Int
Sh
Ca
*
1
03
htt
pizootic hemorrhagic disease virus serotype 7 in European cattle andeep: Diagnostic considerations and effect of previous BTV exposure
ichael Eschbaumer a,1,*, Kerstin Wernike a,1, Carrie A. Batten b, Giovanni Savini c,rraine Edwards b, Annapia Di Gennaro c, Liana Teodori c, Chris A.L. Oura b,artin Beer a, Bernd Hoffmann a
stitute of Diagnostic Virology, Friedrich-Loeffler-Institut, Suedufer 10, 17493 Greifswald – Insel Riems, Germany
stitute for Animal Health, Pirbright Laboratory, Ash Road, Pirbright, Woking, Surrey GU24 0NF, United Kingdom
tituto ‘‘G. Caporale’’, Via Campo Boario, 64100 Teramo, Italy
Introduction
Epizootic hemorrhagic disease virus (EHDV), an arthro-d-borne orbivirus (family Reoviridae), is an emergingthogen of wild and domestic ruminants with at leastven distinct serotypes (Anthony et al., 2009; Savini et al.,11). It is transmitted by blood-feeding Culicoides midgesiptera: Ceratopogonidae). EHDV had first been described
in the United States in the middle of the 20th century, buthas since been found to occur in many parts of the world. Itmost severely affects deer; sheep and goats are notconsidered to be susceptible, while cattle are potentialreservoir hosts (reviewed in Savini et al., 2011). Ofparticular concern to the livestock industry is Ibarakivirus, an EHDV serotype 2 (EHDV-2) strain from Japan thatis known to cause disease in cattle. While the Ibaraki strainhad long been considered unique in this regard, cattle wereaffected by outbreaks of diverse strains of EHDV on theisland of La Reunion (Breard et al., 2004) and more recentlyin Northern Africa, the Near East and Asia Minor (reviewedin Yadin et al., 2008). Whether this was precipitated by an
R T I C L E I N F O
icle history:
ceived 14 January 2012
ceived in revised form 30 March 2012
cepted 12 April 2012
ywords:
izootic hemorrhagic disease virus (EHDV)
etongue virus (BTV)
perimental infection
emia
erference
eep
ttle
A B S T R A C T
Epizootic hemorrhagic disease virus (EHDV), an arthropod-borne orbivirus (family
Reoviridae), is an emerging pathogen of wild and domestic ruminants that is closely related
to bluetongue virus (BTV). The present study examines the outcome of an experimental
EHDV-7 infection of Holstein cattle and East Frisian sheep. Apart from naıve animals that
had not been exposed to BTV, it included animals that had been experimentally infected
with either BTV-6 or BTV-8 two months earlier. In addition, EHDV-infected cattle were
subsequently challenged with BTV-8.
Samples were tested with commercially available ELISA and real-time RT-PCR kits and
a custom NS3-specific real-time RT-PCR assay. Virus isolation was attempted in Vero, C6/
36 and KC cells (from Culicoides variipennis), embryonated chicken eggs and type I
interferon receptor-deficient IFNAR�/� mice.
EHDV-7 productively infected Holstein cattle, but caused no clinical signs. The
inoculation of East Frisian sheep, on the other hand, apparently did not lead to a productive
infection. The commercial diagnostic kits performed adequately. KC cells proved to be the
most sensitive means of virus isolation, but viremia was shorter than 2 weeks in most
animals. No interference between EHDV and BTV infection was observed; therefore the
pre-existing immunity to some BTV serotypes in Europe is not expected to protect against
a possible introduction of EHDV, in spite of the close relation between the viruses.
� 2012 Elsevier B.V. All rights reserved.
Corresponding author. Tel.: +49 38351 71280.
E-mail address: [email protected] (M. Eschbaumer).
Both authors contributed equally.
Contents lists available at SciVerse ScienceDirect
Veterinary Microbiology
jou r nal h o mep ag e: w ww .e ls evier . co m/lo c ate /vetm i c
78-1135/$ – see front matter � 2012 Elsevier B.V. All rights reserved.
p://dx.doi.org/10.1016/j.vetmic.2012.04.020

incs
indeaurCvv
Epinaininclivc
2
2
(YVetwV2is
M. Eschbaumer et al. / Veterinary Microbiology 159 (2012) 298–306 299
creased susceptibility of the hosts, changes in vectorompetency or an inherently different pathogenicity of thetrains is unclear.
The close proximity of these outbreaks caused concern Europe, where EHDV is now seen as a potential threat to
omestic cattle. The recent bluetongue virus (BTV)pizootic has demonstrated that orbiviruses can suddenlyppear, spread quickly and cause severe disease innprotected livestock (Maclachlan, 2011). There is cur-ently widespread immunity to BTV serotype 8 (BTV-8) inentral Europe as a result of natural infection and massaccination campaigns (Zientara et al., 2010), but no EHDVaccines are available in the European Union.
The susceptibility of Holstein cattle to infection withHDV-6 has recently been shown (Batten et al., 2011). Theresent study examines the outcome of EHDV-7 infection
Holstein cattle and East Frisian sheep. Apart from naıvenimals that had not been exposed to BTV, animals werecluded that had been infected with either BTV-6 or BTV-8 a preliminary experiment. This was done to mirror the
urrent prevalence of immunity to BTV in the Europeanvestock population. Additionally, cattle that had pre-iously been infected with EHDV were subsequentlyhallenged with BTV-8.
. Materials and methods
.1. Viruses
An isolate of EHDV-7 from the 2006 outbreak in Israeladin et al., 2008) was obtained from the Kimron
eterinary Institute. The virus had been isolated in anmbryonated chicken egg (ECE) and was further passaged
ice on Vero cells (L0015, Collection of Cell Lines ineterinary Medicine, Friedrich-Loeffler-Institut [FLI]). A920 bp fragment of genome segment 2 of the passaged
Nucleotide collection,2 revealing over 99.9% agreementwith the published segment 2 sequence of Israeli EHDV-7(GenBank3 Accession No. HM156731). Partial sequences ofall 10 segments were uploaded to GenBank (JQ070177–JQ070186).
BTV-6 and BTV-8 used for the study were derived fromviruses originally isolated in Germany in 2008 (Eschbau-mer et al., 2009, 2010). In both cases, Holstein calves hadbeen inoculated with pooled BTV-positive blood samplesthat had been submitted to the national reference lab atthe FLI. The viruses were then isolated from the inoculatedcalves and passaged twice on Vero cells.
2.2. Animals and samples
An overview of the experimental setup is shown inFig. 1. Two groups of four Holstein cattle were infectedwith 5 � 105 50% tissue culture infectious doses (TCID50) ofeither BTV-6 or BTV-8 by subcutaneous (s.c.) injection. Twomonths after the BTV infection, all were inoculated s.c.with 5 � 105 TCID50 of EHDV-7. Four BTV-naıve cattle wereincluded as controls. The cattle that had previously beeninfected with BTV were removed from the experiment 2months after the EHDV infection. The BTV-naıve butEHDV-infected controls, on the other hand, were inocu-lated s.c. with 5 � 105 TCID50 of BTV-8 a month later andkept for a further 4 weeks. A second control group of naıveanimals was included. Whole blood and serum sampleswere taken throughout the experiments (Fig. 1).
Two groups of four East Frisian sheep were included inthe first round of BTV infections and later inoculated withEHDV-7 as described above. A control group of four BTV-naıve sheep were inoculated with EHDV-7 at the same
Fig. 1. Overview of the experiment.
2
http://blast.ncbi.nlm.nih.gov3 http://www.ncbi.nlm.nih.gov/genbank/ olate was sequenced and matched with the NCBI
timafBT
inGethan
ag9.6
2.3
a
getimkicuseNoprTAGG7ICAbecoRTdaanvipr
2.4
inwthIn‘‘G4 8
inPiVe50secewscteincew
4
Vo
M. Eschbaumer et al. / Veterinary Microbiology 159 (2012) 298–306300
e. All sheep were removed from the experiment 35 dayster EHDV infection and did not participate in the secondV infection.The experimental protocol was reviewed by an
dependent ethics commission pursuant to §15 of therman Animal Welfare Act, and it has been approved bye competent authority.4 Throughout the experiment, allimals were examined by veterinarians daily.On the day of the EHDV infection (day 0), the average
e of the cattle was 11.9 � 2.1 months; the sheep were � 0.5 months old.
. Real-time RT-PCR
Total RNA from whole blood samples was extracted withBioSprint 96 robot (Qiagen, Germany). At the FLI, BTVnome in samples was detected using a segment 5 real
e RT-PCR (Toussaint et al., 2007) with the AgPath-IDTM
t (Ambion, USA) in a LightCycler1480 (Roche, Germany). Astom real-time RT-PCR assay based on the segment 10quence of the Israeli EHDV-7 isolate (GenBank Accession. FJ896143.1) was used for EHDV detection (forward
imer EHDV-7Isr-NS3-49F: 50-GCAAGTGATGAAGTGAGTT-GTG-30, reverse primer EHDV-7Isr-NS3-126R: 50-GTATTGGAGCTGACGGTAC-30, hydrolysis probe EHDV-
sr-NS3-78FAM: 50-6-FAM-CCAAGAACAGGTGAGGCCGCC-G-30-BHQ-1). For the purposes of this experiment, it haden verified that this assay was as sensitive as themmercial ‘‘pan-EHDV’’ TaqVetTM EHDV Duplex real-time-PCR kit (Laboratoire Service International [LSI], France;ta not shown). Serial dilutions of in-vitro transcribed BTVd EHDV RNA were used for absolute quantification of the
ral RNA content in samples (B. Hoffmann, unpublished;otocol available on request).
. Virus isolation
An aliquot of each blood sample taken after EHDVfection was washed with PBS (OIE, 2009) and blood cellsere ultrasonically disrupted. Each lysate was divided intoree identical aliquots, to be analyzed at the FLI, thestitute for Animal Health (IAH), Pirbright, and the Istituto. Caporale’’, Teramo (ICT). The samples were shipped atC and kept refrigerated until analysis.At the FLI, virus isolation was attempted on Vero cells and
type I interferon receptor-deficient IFNAR�/�mice (Calvo-nilla et al., 2009). Briefly, 90% confluent monolayers ofro cells in 12-well plates were incubated at 37 8C with0 ml of sample (see above) diluted in an equal amount ofrum-free MEM. After 1 h, the inoculum was removed, thells were washed twice with fresh MEM, and incubatedith 1 ml of MEM at 37 8C for 7 days. The cell layer wasored for cytopathic effect (CPE), the supernatant wassted for viral RNA by real-time RT-PCR, and the presence offectious virus was confirmed by another passage on Verolls. For the mouse inoculations, pairs of IFNAR�/� miceere intraperitoneally injected with 250 ml of sample
diluted in an equal amount of serum-free MEM and keptfor 2 weeks. Surviving mice were euthanized; their spleenswere harvested and tested for EHDV RNA by real-time RT-PCR, as were the spleens of mice that had died over the 2weeks.
At the IAH, KC cells (derived from Culicoides variipennis
midges) were used for virus isolation. Confluent KC cells in12-well plates were inoculated with 500 ml of blood celllysate and incubated at room temperature for 1 h. Theinoculum was removed, the cell layer was washed twicewith fresh Schneider’s medium (Sigma, UK) and incubatedwith 2 ml of medium at 26 8C for 7 days. The supernatantwas tested by real-time RT-PCR for the presence of BTV(Shaw et al., 2007) or EHDV RNA (Batten et al., 2011).
At the ICT, the blood samples were used for inoculation ofVero cells (including quantification by end-point titration),ECE and C6/36 cells from the Asian tiger mosquito Aedes
albopictus. Embryonated chicken eggs were inoculated asdescribed in the Manual of Diagnostic Tests and Vaccines forTerrestrial Animals of the World Organisation for AnimalHealth (OIE, 2009). For the virus isolation in C6/36 cells,confluent monolayers in 24-well plates were covered with200 ml of either neat blood lysate or 1/5 dilutions andincubated at 28 8C for 1 h. The inoculum was removed andafter being washed once with fresh Leibovitz medium(GIBCO1, New Zealand), 1 ml of medium with 10% fetalbovine serum (FBS) and 1% HEPES5 was added to each welland the plates were incubated at 28 8C for 7 days. Thesupernatant was then blindly passaged on Vero cells and thepresence of EHDV confirmed by CPE and/or real time RT-PCR.
For the virus titration, blood lysates were diluted inMEM. Four tenfold dilutions of each sample (from 1/10 to1/10,000) were inoculated into flat-bottomed 96-wellmicrotiter plates, following the method described by OIE(2009). Four replicates were made for each dilution.Approximately 104 Vero cells in a volume of 100 ml ofMEM plus antibiotics and 3% FCS were added per well andthe plates were incubated at 37 8C with 5% CO2. They wereexamined after 7 days and the TCID50 calculated by theReed and Muench (1938) formula.
2.5. Serology
Serum samples were tested with a commercially availableEHDV blocking ELISA (LSI, France) according to the manu-facturer’s instructions. Samples (days 0, 13, 28, 56) from naıvecattle and cattle that had previously been infected with BTV-8were evaluated for neutralizing activity against EHDV-7 in aserum neutralization test (SNT) as described previously(Batten et al., 2011). Samples from sheep S07 and S08 takenon days 0 and 35 were tested in the same manner.
3. Results
3.1. Clinical signs
No increase in body temperature was observed in any ofthe EHDV-infected animals. Isolated lesions of the oral
State Office for Agriculture, Food Safety and Fisheries Mecklenburg-
rpommern, ref. LALLF M-V TSD/7221.3-1.1-058/10. 5 2-[4-(2-Hydroxyethyl)piperazin-1-yl]ethanesulfonic acid

minw
3
ainthwbawo(Cgto(F
sEd
3
cpb
F
P
re
fo
M. Eschbaumer et al. / Veterinary Microbiology 159 (2012) 298–306 301
ucosa were found in three cattle (two in cattle previouslyfected with BTV-8, one in a heifer previously infectedith BTV-6) between 5 and 10 days post infection (dpi).
.2. Real-time RT-PCR
Real-time RT-PCR results of cattle after EHDV infectionre shown in Fig. 2. All animals were negative beforefection. One heifer (C07) was positive from 1 dpi, four ofe twelve cattle were positive from 3 dpi, and all cattleere positive from 5 to 12 dpi. No viral RNA was detected
y real-time RT-PCR in heifer C04 after 12 dpi, in heifer C11fter 13 dpi and in heifer C10 after 40 dpi. All other animalsere positive in the EHDV real-time RT-PCR until the end
f the experiment at 56 dpi (C01–C03, C05–C08) or 112 dpi09, C12). Following the subsequent BTV-8 infection, BTV
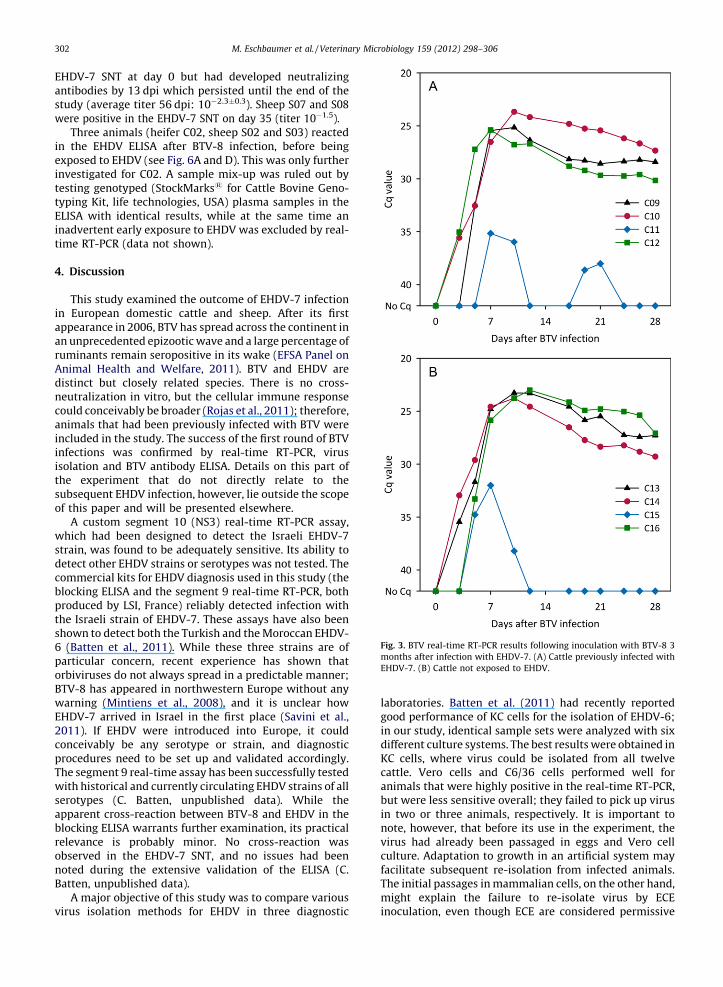
enome was detected in both the cattle previously exposed EHDV (C09–C12) and in the naıve cattle (C13–C16)ig. 3).
No EHDV genome was found in ten of twelve sheep. Inamples from two sheep (S07 and S08), small amounts ofHDV RNA (quantification cycle [Cq] values over 35) wereetected during the second week only (data not shown).
.3. Virus isolation
At the beginning of the EHDV experiment (day 0) allattle previously infected with BTV-6 or BTV-8 wereositive for BTV by real-time RT-PCR (except heifer C04),ut virus could no longer be isolated (data not shown).
Virus isolation results from samples taken after EHDVinfection are shown in Fig. 4. No virus could be isolated in ECE.Using KC cells, EHDV was re-isolated from all twelve cattle,while Vero and C6/36 cells failed to pick up virus in samplesfrom heifers C10, C11 and C04 (C6/36 only). The period inwhich virus could be isolated was longest in KC cells as well. Agroup-level comparison of the virus titers as determined byend-point titration on Vero cells is shown in Fig. 5.
Twelve pairs of IFNAR�/� mice were inoculated withblood samples taken on day 7. Three mice (one of two miceinoculated with blood from C05, and both mice inoculatedwith blood from C12) displayed clinical signs of EHDVinfection (ruffled fur, apathy) (Eschbaumer et al., 2012),but all but one (inoculated with blood from C12) survived.Upon real-time RT-PCR analysis of spleen samples, all miceexcept those inoculated with blood from C04 and C11 werefound to be infected. Real-time RT-PCR results generallymatched gross pathological changes recorded at necropsy;enlarged spleens were positive, while spleens thatappeared to be normal were not (data not shown).
Only the ICT attempted virus isolation from sheepsamples. No virus could be recovered using Vero or C6/36cells or ECE.
3.4. Serology
As determined by blocking ELISA, cattle that had beenpreviously exposed to BTV seroconverted to EHDV earlierthan naıve cattle. All cattle were positive in the ELISAby 10 dpi (Fig. 6). All tested cattle were negative in the
ig. 2. Real-time RT-PCR results after EHDV-7 infection. (A) EHDV real-time RT-PCR results of cattle previously infected with BTV-6. (B) EHDV real-time RT-
CR results of cattle previously infected with BTV-8. (C) EHDV real-time RT-PCR results of previously naıve cattle. (D) Comparison of EHDV real-time RT-PCR
sults between groups of cattle. The average of the BTV-8 group does not include C04, and the average of the naıve group does not include C11. See Section 4
r details.
EHanstuwe
inexintetyELintim
4.
inapanruAndinecoanininisothsuof
wstrdecoblprthsh6
paorBTwEH20coprThwseapblreobnoBa
vi
M. Eschbaumer et al. / Veterinary Microbiology 159 (2012) 298–306302
DV-7 SNT at day 0 but had developed neutralizingtibodies by 13 dpi which persisted until the end of thedy (average titer 56 dpi: 10�2.3�0.3). Sheep S07 and S08re positive in the EHDV-7 SNT on day 35 (titer 10�1.5).Three animals (heifer C02, sheep S02 and S03) reacted
the EHDV ELISA after BTV-8 infection, before beingposed to EHDV (see Fig. 6A and D). This was only furthervestigated for C02. A sample mix-up was ruled out bysting genotyped (StockMarks1 for Cattle Bovine Geno-ping Kit, life technologies, USA) plasma samples in theISA with identical results, while at the same time anadvertent early exposure to EHDV was excluded by real-
e RT-PCR (data not shown).
Discussion
This study examined the outcome of EHDV-7 infection European domestic cattle and sheep. After its firstpearance in 2006, BTV has spread across the continent in unprecedented epizootic wave and a large percentage ofminants remain seropositive in its wake (EFSA Panel onimal Health and Welfare, 2011). BTV and EHDV are
stinct but closely related species. There is no cross-utralization in vitro, but the cellular immune responseuld conceivably be broader (Rojas et al., 2011); therefore,imals that had been previously infected with BTV werecluded in the study. The success of the first round of BTVfections was confirmed by real-time RT-PCR, viruslation and BTV antibody ELISA. Details on this part of
e experiment that do not directly relate to thebsequent EHDV infection, however, lie outside the scope
this paper and will be presented elsewhere.A custom segment 10 (NS3) real-time RT-PCR assay,
hich had been designed to detect the Israeli EHDV-7ain, was found to be adequately sensitive. Its ability totect other EHDV strains or serotypes was not tested. Themmercial kits for EHDV diagnosis used in this study (theocking ELISA and the segment 9 real-time RT-PCR, bothoduced by LSI, France) reliably detected infection withe Israeli strain of EHDV-7. These assays have also beenown to detect both the Turkish and the Moroccan EHDV-(Batten et al., 2011). While these three strains are ofrticular concern, recent experience has shown thatbiviruses do not always spread in a predictable manner;V-8 has appeared in northwestern Europe without any
arning (Mintiens et al., 2008), and it is unclear howDV-7 arrived in Israel in the first place (Savini et al.,11). If EHDV were introduced into Europe, it couldnceivably be any serotype or strain, and diagnosticocedures need to be set up and validated accordingly.e segment 9 real-time assay has been successfully tested
ith historical and currently circulating EHDV strains of allrotypes (C. Batten, unpublished data). While theparent cross-reaction between BTV-8 and EHDV in theocking ELISA warrants further examination, its practicallevance is probably minor. No cross-reaction wasserved in the EHDV-7 SNT, and no issues had beented during the extensive validation of the ELISA (C.tten, unpublished data).A major objective of this study was to compare various
laboratories. Batten et al. (2011) had recently reportedgood performance of KC cells for the isolation of EHDV-6;in our study, identical sample sets were analyzed with sixdifferent culture systems. The best results were obtained inKC cells, where virus could be isolated from all twelvecattle. Vero cells and C6/36 cells performed well foranimals that were highly positive in the real-time RT-PCR,but were less sensitive overall; they failed to pick up virusin two or three animals, respectively. It is important tonote, however, that before its use in the experiment, thevirus had already been passaged in eggs and Vero cellculture. Adaptation to growth in an artificial system mayfacilitate subsequent re-isolation from infected animals.The initial passages in mammalian cells, on the other hand,might explain the failure to re-isolate virus by ECE
Fig. 3. BTV real-time RT-PCR results following inoculation with BTV-8 3
months after infection with EHDV-7. (A) Cattle previously infected with
EHDV-7. (B) Cattle not exposed to EHDV.
oculation, even though ECE are considered permissive
rus isolation methods for EHDV in three diagnostic in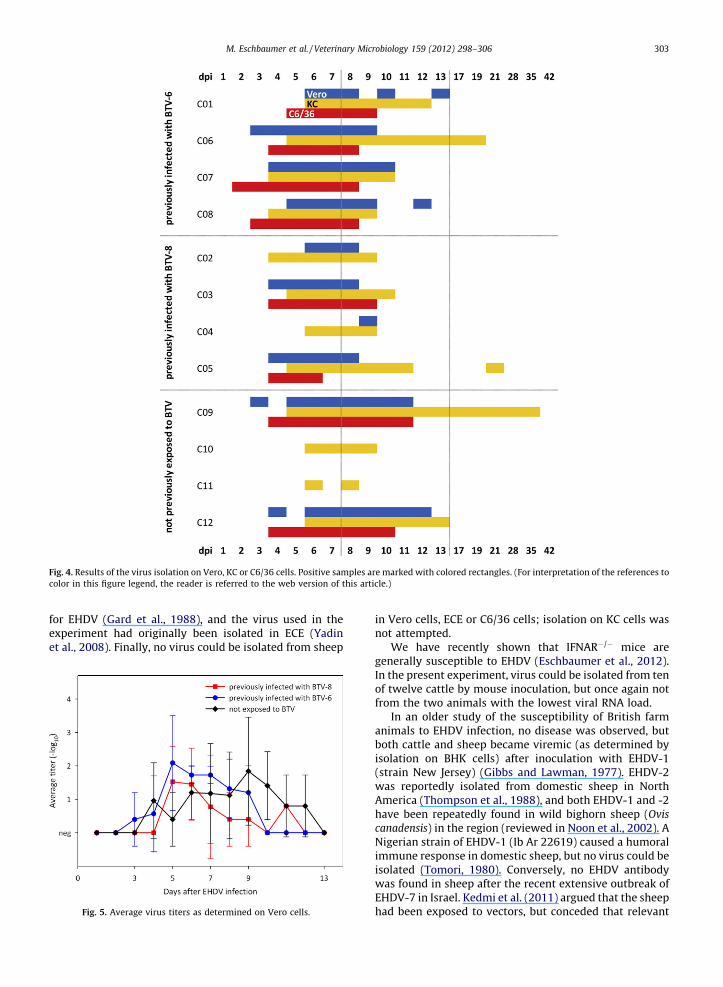
foee
F
c
M. Eschbaumer et al. / Veterinary Microbiology 159 (2012) 298–306 303
r EHDV (Gard et al., 1988), and the virus used in thexperiment had originally been isolated in ECE (Yadint al., 2008). Finally, no virus could be isolated from sheep
in Vero cells, ECE or C6/36 cells; isolation on KC cells wasnot attempted.
We have recently shown that IFNAR�/� mice aregenerally susceptible to EHDV (Eschbaumer et al., 2012).In the present experiment, virus could be isolated from tenof twelve cattle by mouse inoculation, but once again notfrom the two animals with the lowest viral RNA load.
In an older study of the susceptibility of British farmanimals to EHDV infection, no disease was observed, butboth cattle and sheep became viremic (as determined byisolation on BHK cells) after inoculation with EHDV-1(strain New Jersey) (Gibbs and Lawman, 1977). EHDV-2was reportedly isolated from domestic sheep in NorthAmerica (Thompson et al., 1988), and both EHDV-1 and -2have been repeatedly found in wild bighorn sheep (Ovis
canadensis) in the region (reviewed in Noon et al., 2002). ANigerian strain of EHDV-1 (Ib Ar 22619) caused a humoralimmune response in domestic sheep, but no virus could beisolated (Tomori, 1980). Conversely, no EHDV antibodywas found in sheep after the recent extensive outbreak ofEHDV-7 in Israel. Kedmi et al. (2011) argued that the sheephad been exposed to vectors, but conceded that relevant
ig. 4. Results of the virus isolation on Vero, KC or C6/36 cells. Positive samples are marked with colored rectangles. (For interpretation of the references to
olor in this figure legend, the reader is referred to the web version of this article.)
Fig. 5. Average virus titers as determined on Vero cells.

Fig. 6. Results of the EHDV blocking ELISA. The dotted vertical lines mark the BTV (-56) and EHDV infections (0), respectively, and the dotted horizontal line
shows the positive cut-off value of the test (60% blocking). (A) ELISA results for cattle previously infected with BTV-8. (B) ELISA results for cattle previously
infected with BTV-6. (C) ELISA results for previously naıve cattle. (D) ELISA results for sheep previously infected with BTV-8. (E) ELISA results for sheep
previously infected with BTV-6. (F) ELISA results for previously naıve sheep.
M. Eschbaumer et al. / Veterinary Microbiology 159 (2012) 298–306304

CmwnRptoEor
estheuthmswcouEin(Yeaisedhasevp
rthebocbdpEwinmpinsEw(Anocth
M. Eschbaumer et al. / Veterinary Microbiology 159 (2012) 298–306 305
ulicoides spp. often prefer cattle. In the present experi-ent, the antibody response after inoculation of sheepith virus from that outbreak was inconsistent, there was
o detectable viremia, and only a small amount of viralNA was found in the blood of two sheep. Both sheep withositive RT-PCR results developed neutralizing antibodies
EHDV-7, and sheep S08 clearly seroconverted in theLISA. Nevertheless, our results corroborate the conclusionf Kedmi et al. (2011) that sheep do not play a meaningfulole in the epidemiology of the Israeli strain of EHDV-7.
In the outbreak in Israel, EHDV-infected cattle report-dly displayed diverse clinical signs that were in factimilar to bluetongue disease (Yadin et al., 2008). Since
ere was no uninfected control group in the presentxperiment, the significance of the lesions we observed isncertain. The lesions were very mild and it is likely thatey would not have been noticed outside of an experi-ental setting. In a recent challenge study using two
trains of EHDV-6 (Batten et al., 2011), no clinical signsere observed in cattle, even though these strains had
aused disease in the field in Turkey and Morocco. Thebserved viremia in otherwise healthy animals, however,nderscores the relevance of cattle as reservoir hosts forHDV. In any case, the within-herd morbidity was variable
all three outbreaks; in Israel, it lay between 5% and 80%adin et al., 2008). The absence of disease in the present
xperiment, therefore, could be due to the small number ofnimals together with the artificial environment in anolation unit (Batten et al., 2011). Relatedly, intensexposure to sunlight is said to exacerbate bluetongueisease (Erasmus, 1990). Experimental animals are in goodealth and well cared for, which is not necessarily true fornimals in the field. Independent of its cause, immuneuppression can exacerbate orbivirus infection (Brodiet al., 1998). Finally, the possible culture adaptation of theirus which has already been mentioned could affect thehenotype of orbiviruses (Kirkland and Hawkes, 2004).
In this study, most cattle remained positive in the EHDVeal-time RT-PCR until the end of the experiment. For BTV,
e long persistence of virus and viral RNA in blood isxplained by the close association of virus particles withovine blood cells (Brewer and MacLachlan, 1994). Similarbservations were made for EHDV and cervine erythro-ytes (Stallknecht et al., 1997), but no data on EHDV andlood cells of cattle are available. In spite of the persistentetection of viral RNA, only three of twelve heifers wereositive in the virus isolation for longer than 2 weeks afterHDV infection. Only one animal was still positive after 3eeks. Two studies where cattle were experimentallyfected with EHDV reported longer viremia. After 1onth, 50% of animals infected with EHDV-6 were still
ositive (Batten et al., 2011), as were 75% of animalsfected with EHDV-1 (Gibbs and Lawman, 1977). In a
mall study with only two animals, the one infected withHDV-1 was positive in the virus isolation until day 23,hile EHDV-2 was isolated from the other for only 16 daysradaib et al., 1994). The duration of viremia observed in
aturally infected sentinel cattle (in Australia), on thether hand, was shorter: 2 weeks or less in 86% of EHDV-2ases and 94% of EHDV-5 cases. There was only one animalat was found positive for EHDV-7; its viremic period was
also less than 2 weeks (Gard and Melville, 1992, as cited inSavini et al., 2011).
In the first part of the experiment, BTV-8 replication inheifer C04 was much reduced compared to the otheranimals in its group (data not shown); a similar reductionin the level of BTV RNA in the circulation was againobserved in heifer C15 in the second round of BTV-8infections (Fig. 3B). Individual differences in susceptibilityto BTV have been observed before, but no satisfyingexplanation is forthcoming (Wackerlin et al., 2010).Interestingly, when C04 was later inoculated withEHDV-7, this infection took a similar course, with a lower,shorter viremia and less viral RNA in blood (Fig. 2B). HeiferC11, where exposure to EHDV preceded BTV infection,showed the same pattern of diminished susceptibility toboth EHDV and BTV (Figs. 2C and 3A). Given the equivalentreaction to two separate inoculations, it is unlikely thatthis was caused by inadequate injection technique.Hypothetically, individual variations in receptor morphol-ogy or the innate immune response could lead todifferences in the progression of infection (Vos et al.,2009). Further investigations are ongoing.
Experiments with white-tailed deer found cross-protection between EHDV serotypes (Gaydos et al.,2002), but immunity to EHDV did not protect againstsubsequent infection with BTV and vice versa (Hoff andTrainer, 1974; Quist et al., 1997). Even though theseviruses are closely related, they are antigenically distinctand there is no cross-neutralization in vitro either.Fittingly, the present experiment found no meaningfuldifference in the course of EHDV-7 infection betweencattle that had been infected with BTV-6 or BTV-8 andthose that had not been exposed to BTV. The average viralloads in the groups are shown in Fig. 5. Heifers C04 and C11were not included in the analysis because they are notconsidered representative; it is important to note, how-ever, that this level of variability is often observed inexperimental orbivirus infections.
Finally, previous exposure to EHDV-7 did not interferewith subsequent BTV-8 infection (Fig. 3).
5. Conclusion
The 2006 Israeli strain of EHDV-7 productively infectedHolstein cattle, but caused no clinical signs. The inocula-tion of East Frisian sheep, on the other hand, apparently didnot lead to a productive infection. The commerciallyavailable diagnostic kits (real-time RT-PCR and blockingELISA) performed adequately. KC cells (from C. variipennis)proved to be the most sensitive means of virus isolation,but viremia was shorter than 2 weeks in most animals. Nointerference between EHDV and BTV infection was found;therefore, the pre-existing immunity to some BTV ser-otypes in Europe is not expected to protect against apossible introduction of EHDV.
Acknowledgments
The authors thank Valerie Chaignat for critically readingthe manuscript. Fanny Breski, Patrick Zitzow and Christian

KodethwNeCoFr24
Re
An
Ar
Ba
Bre
Bre
Bro
Ca
EF
Era
Esc
Esc
Esc
Ga
Ga
Ga
M. Eschbaumer et al. / Veterinary Microbiology 159 (2012) 298–306306
rthase provided excellent technical assistance, and thedicated animal care by the staff of the isolation unit ofe FLI is gratefully acknowledged. Michael Eschbaumeras supported by EPIZONE, the European Union (EU)twork of Excellence for Epizootic Disease Diagnosis andntrol (FOOD-CT-2006-016236) and the EU Seventh
amework Programme project ORBIVAC (reference5266).
ferences
thony, S.J., Maan, S., Maan, N., Kgosana, L., Bachanek-Bankowska, K.,Batten, C., Darpel, K.E., Sutton, G., Attoui, H., Mertens, P.P.C., 2009.Genetic and phylogenetic analysis of the outer-coat proteins VP2 andVP5 of epizootic haemorrhagic disease virus (EHDV): comparison ofgenetic and serological data to characterise the EHDV serogroup.Virus Res. 145, 200–210.
adaib, I.E., Sawyer, M.M., Osburn, B.I., 1994. Experimental epizootichemorrhagic disease virus infection in calves: virologic and serologicstudies. J. Vet. Diagn. Invest. 6, 489–492.
tten, C.A., Edwards, L., Bin-Tarif, A., Henstock, M.R., Oura, C.A., 2011.Infection kinetics of epizootic haemorrhagic disease virus serotype 6in Holstein-Friesian cattle. Vet. Microbiol. 154, 23–28.
´ard, E., Sailleau, C., Hamblin, C., Graham, S.D., Gourreau, J.M., Zientara,S., 2004. Outbreak of epizootic haemorrhagic disease on the island ofReunion. Vet. Rec. 155, 422–423.
wer, A.W., MacLachlan, N.J., 1994. The pathogenesis of bluetonguevirus infection of bovine blood cells in vitro: ultrastructural char-acterization. Arch. Virol. 136, 287–298.die, S.J., Wilson, W.C., O’Hearn, P.M., Muthui, D., Diem, K., Pearson, L.D.,1998. The effects of pharmacological and lentivirus-induced immunesuppression on orbivirus pathogenesis: assessment of virus burden inblood monocytes and tissues by reverse transcription-in situ PCR. J.Virol. 72, 5599–5609.
lvo-Pinilla, E., Rodrıguez-Calvo, T., Anguita, J., Sevilla, N., Ortego, J.,2009. Establishment of a bluetongue virus infection model in micethat are deficient in the alpha/beta interferon receptor. PLoS One 4,e5171.
SA Panel on Animal Health and Welfare, 2011. Scientific opinion onbluetongue monitoring and surveillance. EFSA J. 9, 2192–2253.
smus, B.J., 1990. Bluetongue. In: Dinter, Z., Morein, B. (Eds.), VirusInfections Of Ruminants. Elsevier Science Publishers B.V., Amsterdam,The Netherlands.
hbaumer, M., Hoffmann, B., Konig, P., Teifke, J.P., Gethmann, J.M.,Conraths, F.J., Probst, C., Mettenleiter, T.C., Beer, M., 2009. Efficacyof three inactivated vaccines against bluetongue virus serotype 8 insheep. Vaccine 27, 4169–4175.
hbaumer, M., Hoffmann, B., Moss, A., Savini, G., Leone, A., Konig, P.,Zemke, J., Conraths, F., Beer, M., 2010. Emergence of bluetongue virusserotype 6 in Europe–German field data and experimental infection ofcattle. Vet. Microbiol. 143, 189–195.
hbaumer, M., Keller, M., Beer, M., Hoffmann, B., 2012. Epizootichemorrhagic disease virus infection of type I interferon receptordeficient mice. Vet. Microbiol. 155, 417–419.
rd, G.P., Melville, L.F., 1992. Results of a decade’s monitoring fororbiviruses in sentinel cattle pastured in an area of regular arbovirusactivity in Northern Australia. In: Bluetongue, African Horse Sicknessand Related Orbiviruses: Proceedings of the Second InternationalSymposium, pp. 85–89.
rd, G.P., Weir, R.P., Walsh, S.J., 1988. Arboviruses recovered fromsentinel cattle using several virus isolation methods. Vet. Microbiol.18, 119–125.
ydos, J.K., Davidson, W.R., Elvinger, F., Howerth, E.W., Murphy, M.,Stallknecht, D.E., 2002. Cross-protection between epizootic hemor-rhagic disease virus serotypes 1 and 2 in white-tailed deer. J. Wildl.Dis. 38, 720–728.
Gibbs, E.P., Lawman, M.J., 1977. Infection of British deer and farm animalswith epizootic haemorrhagic disease of deer virus. J. Comp. Pathol. 87,335–343.
Hoff, G.L., Trainer, D.O., 1974. Observations on bluetongue and epizootichemorrhagic disease viruses in white-tailed deer: (1) distribution ofvirus in the blood (2) cross-challenge. J. Wildl. Dis. 10, 25–31.
Kedmi, M., Levi, S., Galon, N., Bomborov, V., Yadin, H., Batten, C., Klement,E., 2011. No evidence for involvement of sheep in the epidemiology ofcattle virulent epizootic hemorrhagic disease virus. Vet. Microbiol.148, 408–412.
Kirkland, P.D., Hawkes, R.A., 2004. A comparison of laboratory and ‘wild’strains of bluetongue virus—is there any difference and does itmatter? Vet. Ital. 40, 448–455.
Maclachlan, N.J., 2011. Bluetongue: history, global epidemiology, andpathogenesis. Prev. Vet. Med. 102, 107–111.
Mintiens, K., Meroc, E., Mellor, P.S., Staubach, C., Gerbier, G., Elbers, A.R.,Hendrickx, G., De Clercq, K., 2008. Possible routes of introduction ofbluetongue virus serotype 8 into the epicentre of the 2006 epidemicin north-western Europe. Prev. Vet. Med. 87, 131–144.
Noon, T.H., Wesche, S.L., Cagle, D., Mead, D.G., Bicknell, E.J., Bradley, G.A.,Riplog-Peterson, S., Edsall, D., Reggiardo, C., 2002. Hemorrhagic dis-ease in bighorn sheep in Arizona. J. Wildl. Dis. 38, 172–176.
OIE, 2009. Bluetongue. In: Manual of Diagnostic Tests and Vaccines forTerrestrial Animals, Paris, France, Office International des Epizooties.
Quist, C.F., Howerth, E.W., Stallknecht, D.E., Brown, J., Pisell, T., Nettles,V.F., 1997. Host defense responses associated with experimentalhemorrhagic disease in white-tailed deer. J. Wildl. Dis. 33, 584–599.
Reed, L.J., Muench, H., 1938. A simple method for estimating fifty percentendpoints. Am. J. Hyg. 27, 493–497.
Rojas, J.M., Rodrıguez-Calvo, T., Pena, L., Sevilla, N., 2011. T cell responsesto bluetongue virus are directed against multiple and identical CD4+and CD8+ T cell epitopes from the VP7 core protein in mouse andsheep. Vaccine 29, 6848–6857.
Savini, G., Afonso, A., Mellor, P., Aradaib, I., Yadin, H., Sanaa, M., Wilson,W., Monaco, F., Domingo, M., 2011. Epizootic heamorragic disease.Res. Vet. Sci. 91, 1–17.
Shaw, A.E., Monaghan, P., Alpar, H.O., Anthony, S., Darpel, K.E., Batten, C.A.,Guercio, A., Alimena, G., Vitale, M., Bankowska, K., Carpenter, S., Jones,H., Oura, C.A.L., King, D.P., Elliott, H., Mellor, P.S., Mertens, P.P.C., 2007.Development and initial evaluation of a real-time RT-PCR assay todetect Bluetongue virus genome segment 1. J. Virol. Methods 145,115–126.
Stallknecht, D.E., Howerth, E.W., Kellogg, M.L., Quist, C.F., Pisell, T., 1997.In vitro replication of epizootic hemorrhagic disease and bluetongueviruses in white-tailed deer peripheral blood mononuclear cells andvirus-cell association during in vivo infections. J. Wildl. Dis. 33, 574–583.
Thompson, L.H., Mecham, J.O., Holbrook, F.R., 1988. Isolation and char-acterization of epizootic hemorrhagic disease virus from sheep andcattle in Colorado. Am. J. Vet. Res. 49, 1050–1052.
Tomori, O., 1980. Bluetongue and related viruses in Nigeria: experimentalinfection of West African dwarf sheep with Nigeria strains of theviruses of epizootic haemorrhagic disease of deer and bluetongue.Vet. Microbiol. 5, 177–185.
Toussaint, J.F., Sailleau, C., Breard, E., Zientara, S., De Clercq, K., 2007.Bluetongue virus detection by two real-time RT-qPCRs targeting twodifferent genomic segments. J. Virol. Methods 140, 115–123.
Vos, S.M., Yabsley, M.J., Howerth, E.W., 2009. Initial sequencing and tissuedistribution of Toll-like receptor 3 mRNA in white-tailed deer (Odo-coileus virginianus). J. Wildl. Dis. 45, 785–790.
Wackerlin, R., Eschbaumer, M., Konig, P., Hoffmann, B., Beer, M., 2010.Evaluation of humoral response and protective efficacy of threeinactivated vaccines against bluetongue virus serotype 8 one yearafter vaccination of sheep and cattle. Vaccine 28, 4348–4355.
Yadin, H., Brenner, J., Bumbrov, V., Oved, Z., Stram, Y., Klement, E., Perl, S.,Anthony, S., Maan, S., Batten, C., Mertens, P.P., 2008. Epizootic hae-morrhagic disease virus type 7 infection in cattle in Israel. Vet. Rec.162, 53–56.
Zientara, S., MacLachlan, N.J., Calistri, P., Sanchez-Vizcaino, J.M., Savini, G.,2010. Bluetongue vaccination in Europe. Expert Rev. Vaccines 9,989–991.




















