
Environment, glucocorticoids, and the timing of reproduction
7
General and Comparative Endocrinology xxx (2008) xxx–xxx 0016-6480/$ - see front matter © 2008 Elsevier Inc. All rights reserved. doi:10.1016/j.ygcen.2008.09.009 ARTICLE IN PRESS Please cite this article in press as: Schoech, S.J. et al., Environment, glucocorticoids, and the timing of reproduction..., Gen. Comp. Endocrinol. (2008), doi:10.1016/j.ygcen.2008.09.009 Contents lists available at ScienceDirect General and Comparative Endocrinology journal homepage: www.elsevier.com/locate/ygcen 1. Introduction We have learned a considerable amount about the effects of environmental perturbations on the hypothalamo–pituitary–adre- nal (HPA) axis’ production and secretion of glucocorticoids over the past several decades. We have also gained insight into the means by which glucocorticoids can negatively impact the reproductive axis (hypothalamo–pituitary–gonadal [HPG] axis) at multiple lev- els (for review, see Breuner et al., 2008). Despite our knowledge of these relationships, relatively little is known about the interaction between environment, glucocorticoids, and timing of breeding in free-living animals. Although some studies provide insight into this matter, most fail to track animals to the time when they repro- duce, usually because experiments elucidating the mechanisms whereby glucocorticoids impact the central nervous system (CNS) or gonads necessitate sacrificing the animals. Despite having a myriad of effects upon multiple systems, the primary function of corticosterone (CORT), the primary avian glu- cocorticoid, is to facilitate glucose release for utilization during varied challenges. In response to an acute stressor, such as attack by a predator, the adrenal medulla releases epinephrine (EPI) and norepinephrine (NE) which facilitate the ‘immediate’ response to the stimulus (e.g., increases of heart and respiration rates, glucose availability, and blood flow to muscles used in the fight-or-flight response). CORT is subsequently released by the adrenal cortex. If the stimulus is short-lived, there will be a transitory spike in CORT levels a few minutes after the stimulus, though should the stimulus be sustained, CORT levels will remain elevated for a considerable time before returning to pre-stress levels upon adrenal exhaustion, negative feedback of the CORT signal, or cessation of the stimulus. Use of the capture stress paradigm of Wingfield et al. (1992) has demonstrated the nature of the CORT profile for time periods that vary from 30 to 180 min in a number of species, although few stud- ies prolong capture stress long enough to characterize the adrenal exhaustion phase. A number of factors may contribute to variance among baseline CORT levels, the CORT response to a stressor, or both. (1) Body mass is often inversely related to CORT levels (Smith et al., 1994; Schoech et al., 1997, 1999). (2) CORT levels of individ- uals or populations often differ between-seasons or life-history stages (Goymann et al., 2006; Sorato and Kotrschal, 2006; Newman et al. 2008). (3) Birds that live in severe environments in which the opportunity to re-nest, should a nest fail or be abandoned, often Environment, glucocorticoids, and the timing of reproduction Stephan J. Schoech a, * , Michelle A. Rensel a , Eli S. Bridge a,1 , Raoul K. Boughton a,b , Travis E. Wilcoxen a a Department of Biology, University of Memphis, 3774 Walker Avenue, Memphis, TN 38152, USA b Archbold Biological Station, 123 Main Drive, Venus, FL 33860, USA article info abstract Article history: Received 5 August 2008 Revised 10 September 2008 Accepted 26 September 2008 Available online xxxx Glucocorticoids mediate glucose availability under stressful and non-stressful conditions and, therefore, are essential for life. However, data across taxa demonstrate that chronic or elevated secretion of cor- ticosterone or cortisol (CORT) can have negative effects at many levels and can trigger physiological or behavioral responses that may delay or, even halt reproduction. We present a brief overview of the effects that glucocorticoids, primarily the avian form, corticosterone, can have on the reproductive axis. Consid- erable data have demonstrated that environmental perturbations can result in elevated CORT levels that alter a bird’s investment in current reproduction. Studies in our laboratory have shown a link between CORT and timing of reproduction in Florida scrub-jays: in “bad” years, clutch initiation dates are posi- tively correlated with baseline CORT levels of female breeders. Also, population-level differences in CORT levels may explain timing of reproduction as lower CORT levels in suburban-dwelling jays are coupled with early breeding. Most research on stress and CORT concentrates on transient effects of CORT secretion. However, devel- opmental CORT exposure, either from the yolk or embryo, may have long-term effects upon adult phe- notype. For example, CORT levels in nestling scrub-jays predicts later ‘personality,’ as levels were highly correlated (r 2 = 0.84) with fearfulness at 7 months of age. One can imagine that such ‘personality’ traits might also translate into differential success in gaining a territory or a mate. While speculative, it may be that early CORT exposure effectively programs adult behaviors that have wide ranging effects, including upon reproduction. © 2008 Elsevier Inc. All rights reserved. Keywords: Corticosterone Stress Timing of breeding Environmental perturbation * Corresponding author. Fax: +1 901 678 4746. E-mail address: [email protected] (S.J. Schoech). 1 Present address: University of Oklahoma, Oklahoma Biological Survey, 111 E. Chesapeake, Norman, OK 73019, USA.
Transcript of Environment, glucocorticoids, and the timing of reproduction

General and Comparative Endocrinology xxx (2008) xxx–xxx
ARTICLE IN PRESS
Contents lists available at ScienceDirect
General and Comparative Endocrinology
journal homepage: www.elsevier.com/ locate /ygcen
Environment, glucocorticoids, and the timing of reproduction
Stephan J. Schoech a,*, Michelle A. Rensel a, Eli S. Bridge a,1, Raoul K. Boughton a,b, Travis E. Wilcoxen a
a Depart ment of Biol ogy, Uni ver sity of Mem phis, 3774 Walker Ave nue, Mem phis, TN 38152, USAb Arch bold Bio log i cal Sta tion, 123 Main Drive, Venus, FL 33860, USA
a r t i c l e i n f o a b s t r a c t
Article history:
Received 5 August 2008
Revised 10 September 2008
Accepted 26 September 2008
Available online xxxx
Glu co cor ti coids medi ate glu cose avail abil ity under stress ful and non-stress ful con di tions and, there fore,
are essen tial for life. How ever, data across taxa dem on strate that chronic or ele vated secre tion of cor-
ti co ste rone or cor ti sol (CORT) can have neg a tive effects at many lev els and can trig ger phys i o log i cal or
behav ioral responses that may delay or, even halt repro duc tion. We pres ent a brief over view of the effects
that glu co cor ti coids, pri mar ily the avian form, cor ti co ste rone, can have on the repro duc tive axis. Con sid-
er able data have dem on strated that envi ron men tal per tur ba tions can result in ele vated CORT lev els that
alter a bird’s invest ment in cur rent repro duc tion. Stud ies in our lab o ra tory have shown a link between
CORT and tim ing of repro duc tion in Flor ida scrub-jays: in “bad” years, clutch ini ti a tion dates are pos i-
tively cor re lated with base line CORT lev els of female breed ers. Also, pop u la tion-level dif fer ences in CORT
lev els may explain tim ing of repro duc tion as lower CORT lev els in sub ur ban-dwell ing jays are cou pled
with early breed ing.
Most research on stress and CORT con cen trates on tran sient effects of CORT secre tion. How ever, devel-
op men tal CORT expo sure, either from the yolk or embryo, may have long-term effects upon adult phe-
no type. For exam ple, CORT lev els in nest ling scrub-jays pre dicts later ‘per son al ity,’ as lev els were highly
cor re lated (r2 = 0.84) with fear ful ness at 7 months of age. One can imag ine that such ‘per son al ity’ traits
might also trans late into dif fer en tial suc cess in gain ing a ter ri tory or a mate. While spec u la tive, it may be
that early CORT expo sure effec tively pro grams adult behav iors that have wide rang ing effects, includ ing
upon repro duc tion.
© 2008 Else vier Inc. All rights reserved.
Key words:
Cor ti co ste rone
Stress
Tim ing of breed ing
Envi ron men tal per tur ba tion
1. Intro duc tion
We have learned a con sid er able amount about the effects of
envi ron men tal per tur ba tions on the hy po thal amo–pitu i tary–adre-
nal (HPA) axis’ pro duc tion and secre tion of glu co cor ti coids over the
past sev eral decades. We have also gained insight into the means
by which glu co cor ti coids can neg a tively impact the repro duc tive
axis (hy po thal amo–pitu i tary–gonadal [HPG] axis) at multiple lev-
els (for review, see Breu ner et al., 2008). Despite our knowl edge of
these rela tion ships, rel a tively little is known about the inter ac tion
between envi ron ment, glu co cor ti coids, and tim ing of breed ing in
free-living ani mals. Although some stud ies pro vide insight into
this mat ter, most fail to track ani mals to the time when they repro-
duce, usu ally because exper i ments elu ci dat ing the mech a nisms
whereby glu co cor ti coids impact the cen tral ner vous sys tem (CNS)
or gonads neces si tate sac ri fic ing the ani mals.
Despite hav ing a myr iad of effects upon multiple sys tems, the
primary func tion of cor ti co ste rone (CORT), the primary avian glu-
co cor ti coid, is to facil i tate glu cose release for uti li za tion dur ing
0016-6480/$ - see front matter © 2008 Else vier Inc. All rights reserved.
doi:10.1016/j.ygcen.2008.09.009
Please cite this article in press as: Scho ech, S.J. et al., Environment, gluc
(2008), doi:10.1016/j.ygcen.2008.09.009
* Cor re spond ing author. Fax: +1 901 678 4746.
E-mail address: sscho ech@mem phis.edu (S.J. Schoech).
1 Pres ent address: Uni ver sity of Okla homa, Okla homa Bio log i cal Sur vey, 111 E.
Ches a peake, Nor man, OK 73019, USA.
var ied chal lenges. In response to an acute stressor, such as attack
by a pred a tor, the adre nal medulla releases epi neph rine (EPI) and
nor epi neph rine (NE) which facil i tate the ‘imme di ate’ response to
the stim u lus (e.g., increases of heart and res pi ra tion rates, glu cose
avail abil ity, and blood flow to mus cles used in the fight-or-flight
response). CORT is sub se quently released by the adre nal cor tex. If
the stim u lus is short-lived, there will be a tran si tory spike in CORT
lev els a few min utes after the stim u lus, though should the stim u lus
be sus tained, CORT lev els will remain ele vated for a con sid er able
time before return ing to pre-stress lev els upon adre nal exhaus tion,
neg a tive feed back of the CORT sig nal, or ces sa tion of the stim u lus.
Use of the cap ture stress par a digm of Wing field et al. (1992) has
dem on strated the nature of the CORT pro file for time peri ods that
vary from 30 to 180 min in a num ber of spe cies, although few stud-
ies pro long cap ture stress long enough to char ac ter ize the adre nal
exhaus tion phase. A num ber of fac tors may con trib ute to var i ance
among base line CORT lev els, the CORT response to a stressor, or
both. (1) Body mass is often inversely related to CORT lev els (Smith
et al., 1994; Scho ech et al., 1997, 1999). (2) CORT lev els of indi vid-
u als or pop u la tions often dif fer between-sea sons or life-his tory
stages (Goy mann et al., 2006; Sor a to and Ko trs chal, 2006; New man
et al. 2008). (3) Birds that live in severe envi ron ments in which the
oppor tu nity to re-nest, should a nest fail or be aban doned, often
ocorticoids, and the timing of reproduction..., Gen. Comp. Endocrinol.

2 S.J. Scho ech et al. / General and Comparative Endocrinology xxx (2008) xxx–xxx
ARTICLE IN PRESS
exhibit a damp ened CORT response dur ing the breed ing sea son
(e. g., Wing field et al., 1994a,b). (4) Sim i larly, birds with exper i-
men tally enlarged broods dis played damp ened CORT responses to
stress (Lend vai et al., 2007). (5) The length of the breed ing sea son
or lat i tude, two fac tors that are often inter-cor re lated, may affect
the HPA axis (see Goy mann et al., 2006).
In this paper, we first briefly review some of the evi dence that
glu co cor ti coids impinge on the avian HPG axis. Sub se quently, we
pres ent a bit of back ground on envi ron men tal effects upon the
HPA axis: a sec tion that will pro vide a ‘mini-review’ with exam-
ples of how an ani mal’s envi ron ment can stim u late the HPA axis.
Finally, we pres ent data on the inter ac tion between envi ron ment,
glu co cor ti coids, and repro duc tion in our study spe cies, the Flor ida
scrub-jay (Ap he lo co ma coe rules cens). Because this paper is based
upon an oral pre sen ta tion at the qua dren nial Inter na tional Sym po-
sium on Avian Endo cri nol ogy, the vast major ity of the exam ples we
con sider through out are avian. Thus, the glu co cor ti coid of inter est
is cor ti co ste rone (but see Schmidt and Soma, 2008), although in
mam ma lian sys tems it may be cor ti co ste rone, cor ti sol, or both.
1.1. Glu co cor ti coid effects on the avian HPG axis
One of the first avian stud ies to exper i men tally link CORT
with the repro duc tive axis was con ducted by Wil son and Fol lett
(1975) who implanted CORT within the basal hypo thal a mus of
tree spar rows (Spiz el la ar bo rea) and found a marked reduc tion in
both plasma lutein iz ing hor mone (LH) lev els and gonadal growth
rates. Some what par a dox i cally, tree spar rows that received CORT
implants in the field failed to express neg a tive effects upon lev els
of repro duc tive hor mones (e.g., tes tos ter one, T; dihy dro tes tos ter-
one, DHT; and LH: Ast hei mer et al., 2000). Subsequent research has
shown that CORT not only acts via numer ous cen tral sites to down-
reg u late the gon a do tro pin-releas ing hor mone (GnRH) sys tem, but
CORT bind ing sites within the gonad facil i tate down-reg u la tion of
enzy matic sys tems that reg u late T pro duc tion, as well as increase
the like li hood of Ley dig cell apop to sis (Moore and Zo el ler, 1985;
review in Wing field and Far ner, 1993). Sal van tes and Wil liams
(2003) admin is tered exog e nous CORT to female zebra finches and
found (1) decreased vitel lo genin pro duc tion, (2) decreased num-
ber of pairs ini ti at ing repro duc tion (56% vs. 100% of con trols), and
(3) for those CORT-treated females that did lay, an 8-day delay in
clutch ini ti a tion.
Gon a do tro pin-inhib it ing hor mone (GnIH), a recently dis cov-
ered pep tide that plays a role in down-reg u la tion of the HPG axis
(Bent ley et al., 2006; Ubuka et al., 2006; Grei ves et al., 2008), may
medi ate CORT’s effects upon the GnRH sys tem. Cal is i et al. (2008)
exposed house spar rows to a stan dard 1 h cap ture and han dling
stress pro to col and found that these birds had sig nifi cantly more
cells that expressed GnIH immu no re ac tiv ity within the para ven-
tric u lar nucleus (PVN) than did unstressed con trol birds. Con sis-
tent with an anti-gonadal and anti-repro duc tion role for GnIH in
this instance is their find ing that there was no effect dur ing the
fall when all birds had rel a tively high num bers of cells express-
ing GnIH immu no re ac tiv ity. Although work is ongo ing to iso late
CORT recep tors on GnIH cells in birds (Cal is i, per sonal com mu ni-
ca tion), to date recep tors have been iden ti fied on GnIH neu rons of
rats (Kir by et al., 2007).
1.2. Envi ron men tal effects on glu co cor ti coid secre tion
Wing field (1985a) first pub lished an exam ple of an envi ron-
men tal per tur ba tion that was directly linked to increased plasma
CORT lev els in a study of song spar rows (Me losp iza melo dia). A late
spring snow storm led to mark edly increased plasma CORT lev els
along with decreased estra diol lev els in females. While CORT lev-
els returned to pre-storm lev els within a week and a half, plasma
Please cite this article in press as: Scho ech, S.J. et al., Environment, gluco
(2008), doi:10.1016/j.ygcen.2008.09.009
estra diol remained depressed for con sid er ably longer and, when
com pared to the pre vi ous year, the mean lay date was delayed by
1 week. It is likely that the snow cover led to decreased food avail-
abil ity which drove the CORT increase, as food restric tion gen er-
ally results in ele vated CORT lev els (Har vey et al., 1980; Lynn et al.,
2003). Given the above evi dence of direct effects of CORT upon the
HPG axis, it seems likely that the increased CORT lev els were a key
fac tor in the decreased estra diol lev els and delayed breed ing in the
Wing field (1985a) study. How ever, one can not rule out cen tral or
periph e ral effects that may be med i ated by other endo crine or neu-
ro crine secre tions. There has been a reg u lar pro ces sion of newly
dis cov ered met a bolic hor mones for which a full range of func tions
is only revealed some time after the ini tial char ac ter iza tion. For
instance, there is some evi dence that ghre lin and lep tin (which
may not exist in birds, see Sharp et al., 2008), in addi tion to their
roles in food intake and metab o lism, mod ify repro duc tive func-
tion. Tena-Sem pere et al. (2007) posit that these two hor mones,
as well as numer ous other neu ro pep tides of cen tral or periph e ral
ori gin (see Grei ves et al., 2008), “may jointly coop er ate to mod u-
late a wide set of repro duc tive func tions, thereby con trib ut ing to
the phys i o logic inte gra tion of energy bal ance and repro duc tion.”
These ideas are based pri mar ily upon mam ma lian stud ies and it
must be noted that study of these and other pep tides that have
been impli cated in mam ma lian repro duc tion is some what lag ging
in birds.
While most think of cli matic events when con sid er ing envi ron-
ment, in the broad est sense an ani mal’s envi ron ment encom passes
far more than just weather. For exam ple, sev eral stud ies have doc-
u mented that the pres ence of a pred a tor can affect plasma CORT
lev els. Silv er in (1998) found that pied fly catcher males (Fice du la
hyp ol eu ca) exposed to a live-mount of a weasel dur ing the nest
build ing period had ele vated lev els of CORT fol low ing 10 min of
expo sure. Sim i larly, great tits (Pa rus major) in an avi ary that were
exposed to a stuffed owl responded with ele vated CORT lev els
(Coc krem and Silv er in, 2002). In the same pub li ca tion, CORT lev els
in free-living tits so treated exhib ited a trend toward an increase.
In an obser va tional study, male trop i cal ston ech ats (Saxi co la tor qu-
at a ax il lar is) whose ter ri to ries were shared with pred a tory fis cal
shrikes (La nius col lar is) had ele vated base line CORT lev els, sug gest-
ing chronic stress (Sche uer lein et al., 2001). Although a direct link
between CORT and repro duc tion was not estab lished, the research-
ers note that stone chat pairs shar ing their ter ri tory with shrikes
delay ini ti at ing, and are less likely to ini ti ate, a sec ond clutch than
are nearby birds with out shrikes.
Inter ac tions with con spe cifi cs can also be ‘stress ful.’ Sim u lated
ter ri to rial intru sions (STI), in which either a live or a stuffed bird
is pre sented to a ter ri tory holder along with con spe cific song play-
back, are valu able in elu ci dat ing the nature of the tes tos ter one
response to a per ceived con spe cific chal lenge (Wing field, 1985b).
How ever, in some spe cies this pro to col has been found to result
in ele vated lev els of CORT. Male pied fly catch ers responded to a
ter ri to rial intru sion by ele vat ing both T and CORT lev els (Silv er in,
1998). In con trast, blue tits (Cy an istes caeru le us) responded to STIs
with ele vated CORT lev els while exhib it ing decreased T lev els
(Lan dys et al., 2007). Fur ther, Lan dys et al. (2007) used a meta-
anal y sis of the stud ies to date that have used STIs and found that
sin gle-brooded spe cies, like the blue tit, con sis tently ele vate CORT
while exhib it ing either no change or decreased T lev els.
The social inter ac tions that are a part of an ani mal’s envi ron-
ment are not lim ited to those com ing from outside of the social
group. For instance, Ange lier et al. (2007) note that in newly formed
pairs of black-leg ged kit tiwa kes (Ris sa tri dac tyla), both males and
females had higher base line CORT lev els than estab lished pairs,
perhaps result ing from poor cohe sion in shared duties or gen-
eral anx i ety with a new mate. Re mage-Hea ley et al. (2003) noted
sim i lar pat terns in a study with cap tive zebra finches (Ta e ni o py-
corticoids, and the timing of reproduction..., Gen. Comp. Endocrinol.

S.J. Scho ech et al. / General and Comparative Endocrinology xxx (2008) xxx–xxx 3
ARTICLE IN PRESS
gia gut ta ta). Of spe cial inter est to our research group are stud ies
of social groups, spe cifi cally coop er a tive breed ers and espe cially
those spe cies for which there is only one breed ing pair in a group.
To explain repro duc tive qui es cence, it has been pos tu lated that the
non breeder help ers in a group, which are invari ably sub or di nate
to the breed ers, are ‘psy cho log i cally cas trated’ (see Brown, 1978).
This hypoth e sis pre dicts that nonb ree ders are repro duc tively sup-
pressed through dom i nant/sub or di nate inter ac tions that act to
ele vate CORT lev els, thereby down-reg u lat ing HPG axis func tion.
Of the too few stud ies that have addressed this issue in avian
coop er a tive breed ers, there is little evi dence to sug gest that this
hypoth e sis has merit (Scho ech et al., 1991, 1997; Mays et al., 1991;
Wing field et al., 1991; but see Ru ben stein, 2007). How ever, this is a
com mon and recur ring theme in coop er a tively breed ing mam mals
in which it is not uncom mon to find repro duc tive skew enforced
via active social sup pres sion accom pa nied with increased lev els of
CORT and decreased HPG axis func tion of help ers (see Young et al.,
2006; Young, in press and cita tions within).
1.3. Humans and their effects upon the envi ron ment
An increas ingly prom i nent ele ment of all spe cies’ envi ron ment
is human pres ence and activ ity. The influ ence of humans can range
from indi rect effects, such as expo sure to con tam i nants, which may
alter HPA axis func tion, to occa sional direct con tact with humans,
to con sis tent “cohab i ta tion.” There is con sid er able evi dence that
all of the above types of encoun ter can be ‘stress ful.’ For exam-
ple, blood lev els of lead in nest ling white storks (Ci co nia ci co nia)
were pos i tively cor re lated with max i mal CORT lev els (Baos et al.,
2006). Par a dox i cally, over all high est lev els were found in the ref-
er ence pop u la tion rather than in the con tam i nant-exposed col ony.
Sev eral research ers have exam ined the effects of human vis i ta tion
at pen guin col o nies with find ings that vary by spe cies. Mag el lanic
pen guins (Sphe nis cus mag ell an i cus) in tour ist areas exhibit a damp-
ened respon sive ness to stress, sug gest ing habit u a tion (Walker et
al., 2006). Con versely, yel low-eyed pen guins (Meg a dyp tes antip o-
des) appear to be sen si tized to tour ists as birds in tour ist areas have
higher stress-induced CORT lev els and lower repro duc tive suc cess
than birds rarely vis ited (El len berg et al., 2007).
Although many avian spe cies can not coex ist with humans and
rap idly dis ap pear as we encroach on their hab i tat, there are a
num ber of spe cies that seem ingly thrive in cit ies; e.g., house spar-
rows (Passer do mes ti cus) and Euro pean star lings (Stur nus vul ga ris).
While there is a grow ing inter est in urban ecol ogy and demog ra-
phy (see Marz luff et al., 2001), there have been few inves ti ga tions
into the stress phys i ol ogy asso ci ated with urban life. Com par i sons
of city (Munich) and nearby for est-dwell ing Euro pean black birds
(Tur dus mer u la) found that the former had func tional gonads from
3 (males) to 4 (females) weeks ear lier than birds in nat u ral hab i tat
(Parte cke et al., 2005). A fol low-up study with hand-reared birds
found that city birds also had a damp ened CORT response to cap-
ture and han dling (Parte cke et al., 2006). The authors inter pret this
find ing as a micro-evo lu tion ary change allow ing urban-dwell ing
birds to cope with a stress ful envi ron ment. Con versely, Bon ier et al.
(2007) found that male white-crowned spar rows (Zonotrichia leu-
coph rys), but not females, in cit ies had higher base line CORT than
con spe cifi cs in nat u ral hab i tat.
Our study group has exam ined numer ous envi ron men tal vari-
ables to address our obser va tion that Flor ida scrub-jays in a sub-
ur ban devel op ment con sis tently breed ear lier than jays in nat u ral
hab i tat at nearby (10 km) Arch bold Bio log i cal Sta tion (ABS) (Scho-
ech and Bow man, 2001, 2003). Sub ur ban jays’ base line CORT lev-
els are less than one half those of ‘wild land’ jays (2.16 ± 0.28 vs.
4.81 ± 0.33 ng/ml), and Scho ech et al. (2004) spec u late that the
higher CORT in the wild lands might act as a ‘brake’ on the HPG
axis, thereby offer ing a partial expla na tion for their later breed ing.
Please cite this article in press as: Scho ech, S.J. et al., Environment, gluco
(2008), doi:10.1016/j.ygcen.2008.09.009
How ever, a fol low-up study in which wild land jays received exog-
e nous CORT failed to sup port this expla na tion as CORT-dosed birds
did not delay clutch ini ti a tion (Scho ech et al., 2007a). It should be
noted that the CORT doses admin is tered increased CORT lev els
mark edly over the short-term (rep re sen ta tive of an “acute” stress
response), thus fail ing to chron i cally ele vate CORT and con found-
ing inter pre ta tion. Subsequent research to deter mine whether the
low lev els in sub ur ban jays reflected habit u a tion or a com pro mised
HPA axis due to chem i cal con tam i nant expo sure sug gests that nei-
ther of these pos tu lated under ly ing causes has merit, as there was
no between-pop u la tion dif fer ence in the CORT response to cap-
ture stress, whereas both pre dict a damp ened CORT response to a
stressor (Scho ech et al., 2007b). Inter est ingly, sub ur ban jays tended
to have a more robust response as was indi cated by a more rapid
rate of increase over the ini tial 5 min of cap ture.
1.4. Manip u lat ing the envi ron ment with sup ple men tal food: les sons
from Flor ida scrub-jays
In the deci sion to repro duce, resource avail abil ity in the form of
food can oper ate at both the prox i mate and ulti mate lev els (Lack,
1968; Per rins, 1970): food sup ple men ta tion stud ies have been used
to address ques tions across lev els (see Scho ech and Hahn, 2008;
Scho ech et al., 2008 and cita tions within). Con cep tu ally, such a
tech nique can be viewed as a method of alter ing an ani mal’s envi-
ron ment. In a series of exper i ments begin ning in 1993, Scho ech
and col leagues have used food sup ple men ta tion of coop er a tively
breed ing Flor ida scrub-jays to inves ti gate both the phys i o log i cal
mech a nisms link ing food avail abil ity and tim ing of repro duc tion
(Scho ech, 1996; Scho ech and Bow man, 2001, 2003; Scho ech et al.,
2004, 2007a,b) as well as whether there are fit ness ben e fits (Scho-
ech et al., 2008).
Food sup ple mented Flor ida scrub-jays invari ably advance lay-
ing, although the degree is less ened in ‘good’ years and increased
in ‘bad’ years (see Rey nolds et al., 2003; Scho ech et al., 2007b,
2008; see below). In some years, jays pro vided with high qual ity
sup ple men tal food (i.e., high in fat and pro tein) not only advanced
lay ing, but had lower base line CORT lev els than con trol jays and
birds that were pro vided a high fat but low pro tein sup ple ment
(Scho ech et al., 2004). How ever, whether or not food sup ple men ta-
tion affects plasma CORT lev els also appears to vary between years,
assum edly with vary ing con di tions and resource avail abil ity (see
Scho ech et al., 2007b). Although defin i tively link ing envi ron men tal
con di tions, plasma CORT, and tim ing of repro duc tion in this spe-
cies is dif cult (in part due to its threatened sta tus that rules out
some manip u la tive study), the responses to sup ple men tal food
and the nat u ral var i a tion in these three vari ables addressed below
are sug ges tive of causal links.
Over the last 8 years (2001–2008), we have tracked all of the
demo graphic aspects of the study pop u la tion of Flor ida scrub-jays
that occupy the south ern part of ABS, and col lected hun dreds of
blood sam ples from which we’ve deter mined base line lev els of
CORT. For all jays in our pop u la tion, we: (1) track fates from the egg
through death; (2) deter mine sex, sta tus (breeder or non-breed-
ing helper), and age; (3) locate all nests and deter mine lay ing and
hatch ing dates and order, as well as fledg ing dates; and (4) mon i tor
sur vi vor ship to inde pen dence (»70 days post-hatch) and beyond,
includ ing recruit ment into the breed ing pop u la tion. Jays were
trapped in con tin u ously mon i tored Potter traps, thereby assur ing
that an ini tial blood sam ple to mea sure base line CORT was col-
lected within 2–3 min (see Scho ech et al., 1991, 1997, 1999; Ro mer o
and Ro mer o, 2002). Sam ples were later assayed in the Scho ech lab
at the Uni ver sity of Mem phis. While we con ducted food sup ple-
men ta tion stud ies dur ing this period, the find ings pre sented here
are only for non-sup ple mented jays, although some indi vid u als
may have been sup ple mented in pre vi ous years.
corticoids, and the timing of reproduction..., Gen. Comp. Endocrinol.
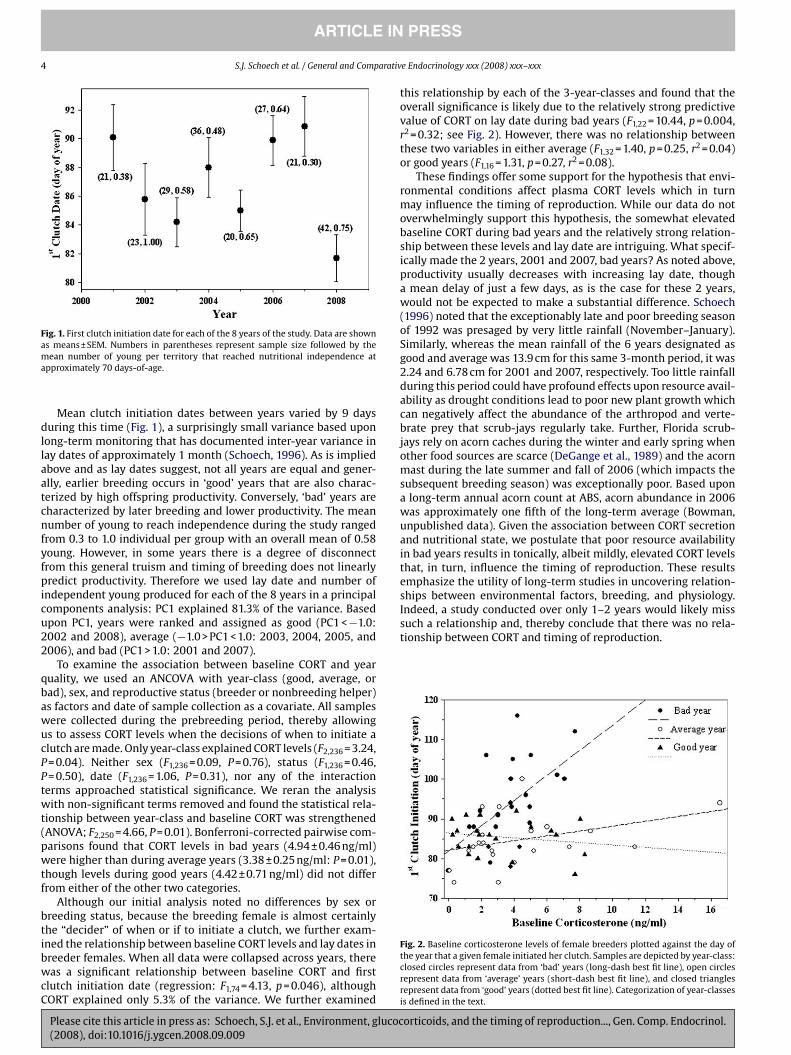
4 S.J. Scho ech et al. / General and Comparative Endocrinology xxx (2008) xxx–xxx
ARTICLE IN PRESS
Fig. 1. First clutch ini ti a tion date for each of the 8 years of the study. Data are shown
as means ± SEM. Num bers in paren the ses rep re sent sam ple size fol lowed by the
mean num ber of young per ter ri tory that reached nutri tional inde pen dence at
approx i mately 70 days-of-age.
Fig. 2. Base line cor ti co ste rone lev els of female breed ers plot ted against the day of
the year that a given female ini ti ated her clutch. Sam ples are depicted by year-class:
closed cir cles rep re sent data from ‘bad’ years (long-dash best fit line), open cir cles
rep re sent data from ‘aver age’ years (short-dash best fit line), and closed tri an gles
rep re sent data from ‘good’ years (dot ted best fit line). Cat e go ri za tion of year-clas ses
is defined in the text.
Mean clutch ini ti a tion dates between years var ied by 9 days
dur ing this time (Fig. 1), a sur pris ingly small var i ance based upon
long-term mon i tor ing that has doc u mented inter-year var i ance in
lay dates of approx i mately 1 month (Scho ech, 1996). As is implied
above and as lay dates sug gest, not all years are equal and gen er-
ally, ear lier breed ing occurs in ‘good’ years that are also char ac-
ter ized by high off spring pro duc tiv ity. Con versely, ‘bad’ years are
char ac ter ized by later breed ing and lower pro duc tiv ity. The mean
num ber of young to reach inde pen dence dur ing the study ranged
from 0.3 to 1.0 indi vid ual per group with an over all mean of 0.58
young. How ever, in some years there is a degree of dis con nect
from this gen eral tru ism and tim ing of breed ing does not lin e arly
pre dict pro duc tiv ity. There fore we used lay date and num ber of
inde pen dent young pro duced for each of the 8 years in a prin ci pal
com po nents anal y sis: PC1 explained 81.3% of the var i ance. Based
upon PC1, years were ranked and assigned as good (PC1 < ¡1.0:
2002 and 2008), aver age (¡1.0 > PC1 < 1.0: 2003, 2004, 2005, and
2006), and bad (PC1 > 1.0: 2001 and 2007).
To exam ine the asso ci a tion between base line CORT and year
qual ity, we used an ANCOVA with year-class (good, aver age, or
bad), sex, and repro duc tive sta tus (breeder or non breed ing helper)
as fac tors and date of sam ple col lec tion as a covar i ate. All sam ples
were col lected dur ing the pre breed ing period, thereby allow ing
us to assess CORT lev els when the deci sions of when to ini ti ate a
clutch are made. Only year-class explained CORT lev els (F2,236 = 3.24,
P = 0.04). Nei ther sex (F1,236 = 0.09, P = 0.76), sta tus (F1,236 = 0.46,
P = 0.50), date (F1,236 = 1.06, P = 0.31), nor any of the inter ac tion
terms approached sta tis ti cal sig nifi cance. We reran the anal y sis
with non-sig nifi cant terms removed and found the sta tis ti cal rela-
tion ship between year-class and base line CORT was strength ened
(ANOVA; F2,250 = 4.66, P = 0.01). Bon fer ron i-cor rected pair wise com-
par i sons found that CORT lev els in bad years (4.94 ± 0.46 ng/ml)
were higher than dur ing aver age years (3.38 ± 0.25 ng/ml: P = 0.01),
though lev els dur ing good years (4.42 ± 0.71 ng/ml) did not dif fer
from either of the other two cat e go ries.
Although our ini tial anal y sis noted no dif fer ences by sex or
breed ing sta tus, because the breed ing female is almost cer tainly
the “decider” of when or if to ini ti ate a clutch, we fur ther exam-
ined the rela tion ship between base line CORT lev els and lay dates in
breeder females. When all data were col lapsed across years, there
was a sig nifi cant rela tion ship between base line CORT and first
clutch ini ti a tion date (regres sion: F1,74 = 4.13, p = 0.046), although
CORT explained only 5.3% of the var i ance. We fur ther exam ined
Please cite this article in press as: Scho ech, S.J. et al., Environment, gluc
(2008), doi:10.1016/j.ygcen.2008.09.009
this rela tion ship by each of the 3-year-clas ses and found that the
over all sig nifi cance is likely due to the rel a tively strong pre dic tive
value of CORT on lay date dur ing bad years (F1,22 = 10.44, p = 0.004,
r2 = 0.32; see Fig. 2). How ever, there was no rela tion ship between
these two vari ables in either aver age (F1,32 = 1.40, p = 0.25, r2 = 0.04)
or good years (F1,16 = 1.31, p = 0.27, r2 = 0.08).
These find ings offer some sup port for the hypoth e sis that envi-
ron men tal con di tions affect plasma CORT lev els which in turn
may influ ence the tim ing of repro duc tion. While our data do not
over whelm ingly sup port this hypoth e sis, the some what ele vated
base line CORT dur ing bad years and the rel a tively strong rela tion-
ship between these lev els and lay date are intrigu ing. What spe cif-
i cally made the 2 years, 2001 and 2007, bad years? As noted above,
pro duc tiv ity usu ally decreases with increas ing lay date, though
a mean delay of just a few days, as is the case for these 2 years,
would not be expected to make a substantial dif fer ence. Scho ech
(1996) noted that the excep tion ably late and poor breed ing sea son
of 1992 was pre saged by very little rain fall (Novem ber–Jan u ary).
Sim i larly, whereas the mean rain fall of the 6 years des ig nated as
good and aver age was 13.9 cm for this same 3-month period, it was
2.24 and 6.78 cm for 2001 and 2007, respec tively. Too little rain fall
dur ing this period could have pro found effects upon resource avail-
abil ity as drought con di tions lead to poor new plant growth which
can neg a tively affect the abun dance of the arthro pod and ver te-
brate prey that scrub-jays reg u larly take. Fur ther, Flor ida scrub-
jays rely on acorn caches dur ing the win ter and early spring when
other food sources are scarce (DeG ange et al., 1989) and the acorn
mast dur ing the late sum mer and fall of 2006 (which impacts the
subsequent breed ing sea son) was excep tion ally poor. Based upon
a long-term annual acorn count at ABS, acorn abun dance in 2006
was approx i mately one fifth of the long-term aver age (Bow man,
unpub lished data). Given the asso ci a tion between CORT secre tion
and nutri tional state, we pos tu late that poor resource avail abil ity
in bad years results in ton i cally, albeit mildly, ele vated CORT lev els
that, in turn, influ ence the tim ing of repro duc tion. These results
empha size the util ity of long-term stud ies in uncov er ing rela tion-
ships between envi ron men tal fac tors, breed ing, and phys i ol ogy.
Indeed, a study con ducted over only 1–2 years would likely miss
such a rela tion ship and, thereby con clude that there was no rela-
tion ship between CORT and tim ing of repro duc tion.
ocorticoids, and the timing of reproduction..., Gen. Comp. Endocrinol.

S.J. Scho ech et al. / General and Comparative Endocrinology xxx (2008) xxx–xxx 5
ARTICLE IN PRESS
Fig. 3. Base line cor ti co ste rone lev els (power trans formed data, see Scho ech et al.,
2007b) of day 11 nes tlings plot ted against indi vid u als’ over all fear ful ness rank ing at
approx i mately 7 months of age. Lower ranks are bolder than higher ranks. Shared
sym bols rep re sent indi vid u als that are sib lings.
1.5. ‘Orga ni za tional’ effects of CORT
The orga ni za tional–ac ti va tion al dichot omy of hor mone
actions has elu ci dated how hor mones can shape an organ ism’s
phys i o log i cal and behav ioral responses (Phoe nix et al., 1959; Goy
and Phoe nix, 1972; Arnold and Breed love, 1985; Moore, 1991).
Devel op men tal effects of hor mones are usu ally con sid ered with
ref er ence to sex ste roids. How ever, it has long been known that
devel op ing male embryos of moth ers that are stressed dur ing
preg nancy are some what fem i nized, with a num ber of male-typ-
i cal behav ior pat terns affected upon reach ing adult hood (Nel son,
2005). Sim i larly, post-natal paren tal care can have crit i cal effects
on off spring devel op ment that can influ ence an indi vid ual’s
adult phe no type and per son al ity (Mea ney, 2001; Kim and Dia-
mond, 2002; Re pet ti et al., 2002; Dinge manse et al., 2004; Zhang
et al., 2006). For exam ple, in rats, off spring reared by more atten-
tive dams dis played more explor atory behav ior and recov ered
more rap idly fol low ing expo sure to a stressor (Liu et al., 1997;
Weaver et al., 2002; Szyf et al., 2005). Con versely, off spring from
less atten tive moth ers were more fear ful and had greater stress
responses (i.e., increased lev els of ACTH and CORT). Weaver et al.
(2006) found that the “high-stress” devel op men tal path way in
rats was estab lished early in life by deac ti va tion of DNA regions
that encode glu co cor ti coid recep tors, thereby affect ing adult
responses to stress ors. Thus, dif fer ences in mater nal care can
direct a per ma nent change in DNA expres sion that can shape an
indi vid ual’s per son al ity by for ever alter ing how it responds to
envi ron men tal chal lenges.
Sim i lar links between devel op men tal stress and adult phe no-
type have been observed in birds. For exam ple, in black-leg ged
kit tiwa kes, nutri tional stress dur ing devel op ment results in cog-
ni tively and phys i o log i cally com pro mised adults (Ki tay sky et al.,
2006). Also, western scrub-jays (Ap he lo co ma cal i for ni ca), a Flor-
ida scrub-jay con ge ner, that were food-restricted as nes tlings had
higher base line CORT lev els and more pro nounced stress responses
at 1year-of-age than ad libi tum fed con trols (Prav osudov and Ki tay-
sky, 2006). Sev eral recent stud ies of pas ser ine and non-pas ser ine
spe cies have noted links between phe no type (e.g., ‘per son al ity’ and
stress respon sive ness) and CORT expo sure (dur ing devel op ment or
as an adult) that par al lel those found in the above described mam-
ma lian research (e.g., Euro pean star ling, Love and Wil liams, 2008;
great tit, Ca re re et al., 2003; Jap a nese quail [Co tur nix co tur nix];
Hay ward and Wing field, 2004; and see Coc krem, 2007 for a review
that cov ers sev eral spe cies).
It has been hypoth e sized that such ‘pro gram ming’ of an indi-
vid ual’s phe no type is adap tive, and that in a fluc tu at ing envi ron-
ment, there is no sin gle opti mal phe no type. The qual ity or quan-
tity of paren tal care, there fore, may serve as a sig nal that directs
off spring devel op ment down the path that best ensures sur vival
dur ing the vul ner a ble, early stages of life (for review, see Ko o lhaas
et al., 1999; Wells, 2003; Zhang et al., 2006). For exam ple, in harsh
con di tions, a high-CORT phe no type may increase an indi vid ual’s
chances of sur viv ing, espe cially if fear ful ness trans lates into anti-
pred a tor behav ior and if the abil ity to metab o lize stored nutri ents
(facil i tated through CORT secre tion) is at a premium.
We rea soned that var i ance in per son al ity, pri mar ily bold ness
or timid ity, could have a strong effect on the like li hood that an
indi vid ual gains a breed ing ter ri tory or a mate. In Flor ida scrub-
jays there is con sid er able var i ance in the time that a young bird
remains as a helper in its natal ter ri tory before becom ing a breeder
(see Wool fen den and Fitz pa trick, 1984, 1990, 1996). An excit-
ing new line of research from our group addresses the rela tion-
ship between a nest ling’s envi ron ment, its CORT lev els, and adult
phe no type; and although it is too early to empir i cally deter mine
impacts on the tim ing of breed ing, we pres ent these find ings as an
illus tra tion of the orga ni za tional effects of CORT.
Please cite this article in press as: Scho ech, S.J. et al., Environment, gluco
(2008), doi:10.1016/j.ygcen.2008.09.009
All nes tlings are banded, mea sured (mass and lin ear size mea-
sures), and a small blood sam ple is col lected on day 11 post-hatch.
Each nest ling is removed from the nest, bled within 3 min, and
mea sured before being returned to the nest; the pro ce dure is then
repeated for each subsequent nest ling. This allows base line CORT
lev els of nes tlings to be mea sured (the HPA axis of 11-day-old nes-
tlings is capa ble of mount ing a stress response, Ren sel and Bough-
ton, unpub lished data). In the bad year of 2007 (see above), of the
55 nes tlings banded and sam pled at day 11 only 17 sur vived to
inde pen dence.
We con ducted three behav ioral tests on 10 of the 17 sur vi vors
when they were approx i mately 7 months of age. Prior to test ing,
the ‘naive’ young were trained to come to a pile of pea nuts and, as
is their wont; once jays dis cover pea nuts they return to the source
until it’s depleted, eat ing until sati ated and cach ing the rest. The
tests exploited this trait and were video-taped to facil i tate indi vid-
ual iden ti fi ca tion and accu rate time keep ing. Test 1, the ring test,
used a bright orange ring 50 cm in diam e ter and 3 cm high which
was placed around a peanut pile; the time a jay spent within 1 m
of the ring before cross ing to take a peanut was then mea sured.
For tests 2 and 3, a hid den buzzer beneath the pile or a mov ing leaf
attached to a hid den motor within the pile, respec tively, star tled
sub jects; return times were then assessed. The ring test yielded
hes i tancy times that ranged from 2 to 192 s; how ever, two of the
10 birds failed to cross the ring dur ing the 1 h allot ted test period.
For the sound and motion tests, return times ranged from 1 to 57 s
and 10 s to 32 min, respec tively.
Because the times to com plete the test were not nor mally dis-
trib uted, for each test birds were ranked from one to 10 (note, the
two birds that did not return to cross the ring shared a score of
nine) with the low est rank cor re spond ing to the short est time
(i.e., the least fear ful indi vid ual). An indi vid ual’s three ranks were
summed and this total was used to gen er ate an over all fear ful-
ness rank across the 10 test jays. To test the degree to which early
CORT expo sure influ enced the jays’ ‘per son al ity,’ we regressed the
over all fear ful ness rank against nest ling CORT lev els and found an
extremely strong rela tion ship (F1,8 = 41.48, p < 0.001, r2 = 0.84; Fig.
3). Fur ther, seven of the tested jays sur vived the remain der of the
win ter and were again tested with the ring test at 1 year of age. We
found that this mea sure of fear ful ness tended to per sist (r = 0.68,
p = 0.066, n = 7). While the small sam ple sizes ren der draw ing gen-
eral con clu sions prob lem atic, we also observed indi ca tions of per-
sis tence in stress phys i ol ogy. For exam ple, (1) base line CORT lev els
corticoids, and the timing of reproduction..., Gen. Comp. Endocrinol.

6 S.J. Scho ech et al. / General and Comparative Endocrinology xxx (2008) xxx–xxx
ARTICLE IN PRESS
of 11-day-old nes tlings tended to be cor re lated with lev els at 2.5
months (r = 0.64, p = 0.12, n = 7); (2) as were base line CORT lev els at
approx i mately 2.5 months and 1 year (r = 0.72, p = 0.069, n = 7); and
(3) as were max i mum CORT lev els at approx i mately 2.5 months
and 1 year (r = 0.81, p = 0.052, n = 6). We look for ward to gath er ing
fur ther data to deter mine whether these trends per sist.
2. Con clu sions
We have pre sented a brief over view of how the envi ron men tal
con di tions an ani mal expe ri ences can affect its CORT lev els and, in
some cases how this can then play a role in deter min ing the tim ing
of repro duc tion. While there is con sid er able evi dence link ing the
envi ron ment and CORT and CORT and the repro duc tive axis, there
are fewer stud ies that inte grate these three vari ables. Regard less,
it seems clear that ele vated CORT lev els in response to a pleth ora
of envi ron men tal con di tions can slow the up-reg u la tion of the
HPG axis, thereby delay ing the onset of repro duc tion. How ever, it
is equally clear that CORT lev els are not the sole deter mi nant of
when the onset of repro duc tion occurs.
Acknowl edg ments
S.J.S. thanks John Coc krem, Pierre De vi che, and Wolf gang Goy-
mann for invit ing him to speak in the Ecol ogy and Evo lu tion sym po-
sium they con vened at the July 2008 Inter na tional Sym po sium on
Avian Endo cri nol ogy in Leu ven, Bel gium: this paper is based upon
S.J.S.’s talk at that meet ing. Dur ing the col lec tion of data described
herein, we were par tially sup ported by NSF fund ing to S.J.S. (IBN-
9983201 and IOS-0346328). M.A.R. has also been funded in part by
Sigma Xi, the Amer i can Orni thol o gists’ Union, and the Depart ment
of Biol ogy at the Uni ver sity of Mem phis: the lat ter has pro vided
sup port for all of the co-authors. Spe cial thanks to S. James Rey-
nolds and Gina Mor gan for help with data col lec tion and multiple
aspects of the jay pro ject. Fur ther assis tance in the field came from
Jon a than At well, Tim Har ri son, and numer ous field assis tants. The
research was greatly aided by S.J.S.’ col lab o ra tion with R. Bow man
at ABS. We thank Dave Free man for help ful com ments on an ear lier
draft of this paper.
Ref er ences
Ange lier, F., Moe, B., Clem ent-Cha stel, C., Bech, C., Cha stel, O., 2007. Cor ti co ste rone lev els in rela tion to change of mate in black-leg ged kit tiwa kes. Con dor 109, 668–674.
Arnold, A.P., Breed love, S.M., 1985. Orga ni za tional and ac ti va tion al effects of sex ste roids on brain and behav ior: a reanal y sis. Hor mones and Behav ior 19, 469–498.
Ast hei mer, L.B., But te mer, W.A., Wing field, J.C., 2000. Cor ti co ste rone treat ment has no effect on repro duc tive hor mones or aggres sive behav ior in free-living male tree spar rows, Spiz el la ar bo rea. Hor mones and Behav ior 37, 31–39.
Baos, R., Blas, J., Bor tolotti, G.R., Mar chant, T.A., Hir aldo, F., 2006. Adre no cor ti cal response to stress and thy roid hor mone sta tus in free-living nest ling white storks (Ci co nia ci co nia) exposed to heavy metal and arsenic con tam i na tion. Envi ron men tal Health Per spec tives 114, 1497–1501.
Bent ley, G.E., Jen sen, J.P., Kaur, G.J., Wac ker, D.W., Tsut sui, K., Wing field, J.C., 2006. Rapid inhi bi tion of female sex ual behav ior by gon a do tro pin-inhib i tory hor-mone (GnIH). Hor mones and Behav ior 49, 550–555.
Bon ier, F., Mar tin, P.R., Shel don, K.S., Jen sen, J.P., Fol tz, S.L., Wing field, J.C., 2007. Sex-spe cific con se quences of life in the city. Behav ioral Ecol ogy 18, 121–129.
Brown, J.L., 1978. Avian com mu nal breed ing sys tems. Annual Review of Ecol ogy and Sys tem at ics 9, 123–155.
Breu ner, C.W., Patt er son, S.H., Hahn, T.P., 2008. In search of rela tion ships between the acute adre no cor ti cal response and fit ness. Gen eral and Com par a tive Endo-cri nol ogy 157, 288–295.
Cal is i, R.M., Riz zo, N.O., Bent ley, G.E., 2008. Sea sonal dif fer ences in hypo tha lamic EGR-1 and GnIH expres sion fol low ing cap ture-han dling stress in house spar-rows (Passer do mes ti cus). Gen eral and Com par a tive Endo cri nol ogy 157, 278–283.
Ca re re, C., Gro othuis, T.G.G., Möstl, E., Daan, S., Ko o lhaas, J.M., 2003. Fecal cor-ti co ste roids in a ter ri to rial bird selected for dif fer ent per son al i ties: daily rhythm and the response to social stress. Hor mones and Behav ior 43, 540–548.
Please cite this article in press as: Scho ech, S.J. et al., Environment, gluco
(2008), doi:10.1016/j.ygcen.2008.09.009
Coc krem, J.F. 2007. Stress, cor ti co ste rone responses and avian per son al i ties. Jour nal of Orni thol ogy 148 (Sup pl. 2). doi:10.1007/s10336-007-0175-8.
Coc krem, J.F., Silv er in, B., 2002. Sight of a pred a tor can stim u late a cor ti co ste rone response in the great tit (Pa rus major). Gen eral and Com par a tive Endo cri nol ogy 125, 248–255.
DeG ange, A.R., Fitz pa trick, J.W., Lay ne, J.N., Wool fen den, G.E., 1989. Acorn har vest-ing by Flor ida scrub jay. Ecol ogy 70, 348–356.
Dinge manse, N.J., Both, C., Drent, P.J., Tin ber gen, J.M., 2004. Fit ness con se quences of avian per son al i ties in a fluc tu at ing envi ron ment. Pro ceed ings of the Royal Soci ety of Lon don. Series B: Bio log i cal Sci ences 271, 847–852.
El len berg, U., Seti a wan, A.N., Cree, A., Hous ton, D.M., Sed don, P.J., 2007. Ele vated hor monal stress response and reduced repro duc tive output in yel low-eyed pen guins exposed to unreg u lated tour ism. Gen eral and Com par a tive Endo cri-nol ogy 152, 54–63.
Goy, R.W., Phoe nix, C.H., 1972. The effects of tes tos ter one pro pi o nate admin is tered before birth on the devel op ment of behav ior in genetic female rhe sus mon keys. In: Saw yer, C., Gor ski, R. (Eds.), Ste roid Hor mones and Brain Func tion. Uni ver-sity of Cal i for nia Press, Berke ley, pp. 193–201.
Goy mann, W., Geue, D., Schwabl, I., Flinks, H., Sch midl, D., Schwabl, H., Gwin ner, E., 2006. Tes tos ter one and cor ti co ste rone dur ing the breed ing cycle of equa to-rial and Euro pean ston ech ats (Saxi co la tor qu at a ax il lar is and S. t. ru bi co la). Hor-mones and Behav ior 50, 779–785.
Grei ves, T.J., Kriegs feld, L.J., Bent ley, G.E., Tsut sui, K., Demas, G.E., 2008. Recent advances in repro duc tive neu ro en do cri nol ogy: a role for RFa mide pep tides
in sea sonal repro duc tion? Pro ceed ings of the Royal Soci ety, B 275, 1943–1951.
Har vey, W.N., Merry, B.J., Phil lips, J.G., 1980. Influ ence of stress on the secre tion of
cor ti co ste rone in the duck (Anas platy rhyn chos). Jour nal of Endo cri nol ogy 87, 161–171.
Hay ward, L.S., Wing field, J.C., 2004. Mater nal cor ti co ste rone is trans ferred to avian yolk and may alter off spring growth and adult phe no type. Gen eral and Com par-a tive Endo cri nol ogy 135, 365–371.
Kim, J.J., Dia mond, D.M., 2002. The stressed hip po cam pus, syn ap tic plas tic ity and lost mem o ries. Nature Reviews Neu ro sci ence 3, 453–462.
Kir by, E.D., Ubuka, T., Kau fer, D. Bent ley, G.E., 2007. Acute immo bi li za tion stress increases puta tive gon a do tro pin inhib i tory hor mone expres sion in the rat hypo thal a mus. The 37th Annual Meet ing for the Soci ety of Neu ro sci ence, San Diego, Cal i for nia (Abstract).
Ki tay sky, A.S., Kit ais kaia, E.V., Platt, J.F., Wing field, J.C., 2006. A mech a nis tic link between chick diet and decline in sea birds? Pro ceed ings of the Royal Soci ety of Lon don. Series B: Bio log i cal Sci ences 273, 445–450.
Ko o lhaas, J.M., Kor te, S.M., de Boer, S.F., van der Vegt, B.J., van Ree nen, C.G., Hop ster, H., de Jong, I.C., Ruis, M.A.W., Blokhuis, H.J., 1999. Cop ing styles in ani mals: cur-rent sta tus in behav ior and stress-phys i ol ogy. Neu ro sci ence and Bio be hav ior Reviews 23, 925–935.
Lack, D., 1968. Eco log i cal Adap ta tions for Breed ing in Birds. Chap man and Hall, Lon-don.
Lan dys, M.M., Goy mann, W., Ra ess, M., Sla gsv old, T., 2007. Hor monal responses to male–male social chal lenge in the blue tit, Cy an istes ca ru le us: sin gle-brood-ed ness as an explan a tory var i able. Phys i o log i cal and Bio chem i cal Zool ogy 80, 228–240.
Lend vai, A.Z., Gi rau deau, M., Cha stel, O., 2007. Repro duc tion and mod u la tion of the stress response: an exper i men tal test in the house spar row. Pro ceed ings of the Royal Soci ety, B 274, 391–397.
Liu, D., Dio rio, J., Tan nen baum, B., Cal dji, C., Fran cis, D., Freed man, A., Shar ma, S.,
Pear son, D., Plot sky, P.M., Mea ney, M.J., 1997. Mater nal care, hip po cam pal glu-co cor ti coid recep tors, and hypo tha lamic–pitu i tary–adre nal responses to stress. Sci ence 277, 1659–1662.
Love, O.P., Wil liams, T.D., 2008. Plas tic ity in the adre no cor ti cal response of a free-living ver te brate: The role of pre- and post-natal devel op men tal stress. Hor-mones and Behav ior 54, 496–505.
Lynn, S.E., Breu ner, C.W., Wing field, J.C., 2003. Short-term fast ing affects loco mo-tor activ ity, cor ti co ste rone, and cor ti co ste rone bind ing glob u lin in a migra tory song bird. Hor mones and Behav ior 43, 150–157.
Marz luff, J.M., Bow man, R., Don nel ly, R., 2001. Avian Ecol ogy and Con ser va tion in an Urban iz ing World. Klu wer Aca demic Press, Nor well, Mas sa chu setts.
Mays, N.A., Vleck, C.M., Daw son, J., 1991. Plasma lutein iz ing hor mone, ste roid hor-mones, behav ioral role, and nest stage in coop er a tively breed ing Har ris’ hawks (Pa rabu teo uni cinc tus). Auk 108, 619–637.
Mea ney, M.J., 2001. Mater nal care, gene expres sion, and the trans mis sion of indi vid-ual dif fer ences in stress reac tiv ity across gen er a tions. Annual Review of Neu ro-sci ence 24, 1161–1192.
Moore, F.L., Zo el ler, R.T., 1985. Stress-induced inhi bi tion of repro duc tion: evi dence of sup pressed secre tion of LH-RH in an amphib ian. Gen eral and Com par a tive Endo cri nol ogy 60, 252–258.
Moore, M.C., 1991. Appli ca tion of orga ni za tion–acti va tion the ory to alter na-tive male repro duc tive strat e gies: a review. Hor mones and Behav ior 25, 154–179.
Nel son, R.J., 2005. An Intro duc tion to Behav ioral Endo cri nol ogy. Si na uer Asso ci ates, Inc., Sun der land, MA.
New man, A.E.M., Prad han, D.S., Soma, K.K., 2008. Dehy dro epi an dros ter one and cor-ti co ste rone are reg u lated by sea son and acute stress in a wild song bird: jug u lar ver sus brachial plasma. Endo cri nol ogy 149, 2537–2545.
Parte cke, J., Schwabl, I., Gwin ner, E., 2006. Stress and the city: urban i za tion and its effects on the stress phys i ol ogy of Euro pean black birds. Ecol ogy 87, 1945–1952.
corticoids, and the timing of reproduction..., Gen. Comp. Endocrinol.

S.J. Scho ech et al. / General and Comparative Endocrinology xxx (2008) xxx–xxx 7
ARTICLE IN PRESS
Parte cke, J., Van’t Hof, T.J., Gwin ner, E., 2005. Under ly ing phys i o log i cal con trol of repro duc tion in urban and for est-dwell ing Euro pean black birds Tur dus mer u la. Jour nal of Avian Biol ogy 36, 295–305.
Per rins, C., 1970. The tim ing of birds’ breed ing sea sons. Ibis 112, 242–255.Phoe nix, C.H., Goy, R.W., Ger all, A.A., Young, W.C., 1959. Orga niz ing action of pre-
na tally admin is tered tes tos ter one pro pi o nate on the tis sues medi at ing mat ing behav ior in the female guinea pig. Endo cri nol ogy 65, 369–382.
Prav osudov, V.V., Ki tay sky, A.S., 2006. Effects of nutri tional restric tions dur ing post-hatch ing devel op ment on adre no cor ti cal func tion in western scrub-jays (Ap he-lo co ma cal i for ni ca). Gen eral and Com par a tive Endo cri nol ogy 145, 25–31.
Re mage-Hea ley, L., Ad kins-Re gan, E., Ro mer o, L.M., 2003. Behav ioral and adre no-cor ti cal responses to mate sep a ra tion and reunion in the zebra finch. Hor mones and Behav ior 43, 108–114.
Re pet ti, R.L., Tay lor, S.E., Se eman, T.E., 2002. Risky fam i lies: fam ily social envi ron-ments and the men tal and phys i cal health of off spring. Psy cho log i cal Bul le tin 128, 330–366.
Rey nolds, S.J., Scho ech, S.J., Bow man, R., 2003. Nutri tional qual ity of pre breed ing diet influ ences breed ing per for mance of the Flor ida Scrub-Jay. Oec o lo gia 134, 308–316.
Ro mer o, L.M., Ro mer o, R.C., 2002. Cor ti co ste rone responses in wild birds: the impor tance of rapid ini tial sam pling. Con dor 104, 129–135.
Ru ben stein, D.R., 2007. Stress hor mones and soci al ity: inte grat ing social and envi-ron men tal stress ors. Pro ceed ings of the Royal Soci ety, Lon don. Series B: Bio log-i cal Sci ences 274, 967–975.
Sal van tes, K.G., Wil liams, T.D., 2003. Effects of cor ti co ste rone on the pro por tion of breed ing females, repro duc tive output and yolk pre cur sor lev els. Gen eral and Com par a tive Endo cri nol ogy 130, 205–214.
Sche uer lein, A., Van’t Hof, T.J., Gwin ner, E., 2001. Pre da tors as stress ors? Phys i o-log i cal and repro duc tive con se quences of pre da tion risk in trop i cal ston ech ats (Saxi co la tor qu at a ax il lar is). Pro ceed ings of the Royal Soci ety, Lon don. Series B: Bio log i cal Sci ences 268, 1575–1582.
Schmidt, K.L., Soma, K.K., 2008. Cor ti sol and cor ti co ste rone in the song bird immune and ner vous sys tem: local vs. sys temic lev els dur ing devel op ment. Amer i can Jour nal of Phys i ol ogy. Reg u la tory, Inte gra tive and Com par a tive Phys i ol ogy 295, R103–R110.
Scho ech, S.J., 1996. The effect of sup ple men tal food on body con di tion and the tim-ing of repro duc tion in a coop er a tive breeder, the Flor ida scrub-jay (Ap he lo co ma coe rules cens). Con dor 98, 234–244.
Scho ech, S.J., Bow man, R., 2001. Var i a tion in the tim ing of breed ing in two Flor-ida Scrub-Jay (Ap he lo co ma coe rules cens) pop u la tions: do phys i o logic mea sures reflect dif fer ent envi ron ments? In: Marz luff, J.M., Bow man, R., Don nel ly, R. (Eds.), Avian Ecol ogy and Con ser va tion in an Urban iz ing World. Klu wer Aca-demic Press, Nor well, Mas sa chu setts, pp. 289–306.
Scho ech, S.J., Bow man, R., 2003. Does dif fer en tial access to pro tein influ ence dif fer-ences in tim ing of breed ing of Flor ida Scrub-Jays (Ap he lo co ma coe rules cens) in sub ur ban and wild land hab i tats? Auk 120, 1114–1127.
Scho ech, S.J., Bow man, R., Rey nolds, S.J., 2004. Food sup ple men ta tion and pos si ble mech a nisms under ly ing early breed ing in the Flor ida Scrub-Jay (Ap he lo co ma coe rules cens). Hor mones and Behav ior 46, 565–573.
Scho ech, S.J., Bow man, R., Bridge, E.S., Mor gan, G.M., Ren sel, M.A., Wil coxen, T.E., Bough ton, R.K., 2007a. Cor ti co ste rone admin is tra tion does not affect tim ing of breed ing in Flor ida Scrub-Jays (Ap he lo co ma coe rules cens). Hor mones and Behav ior 52, 191–196.
Scho ech, S.J., Bow man, R., Bridge, E.S., Bough ton, R.K., 2007b. Base line and acute lev els of cor ti co ste rone in Flor ida Scrub-Jays (Ap he lo co ma coe rules cens): effects
of food sup ple men ta tion, sub ur ban hab i tat, and year. Gen eral and Com par a tive Endo cri nol ogy 154, 150–160.
Scho ech, S.J., Bridge, E.S., Bough ton, R.K., Rey nolds, S.J., At well, J.W., Bow man, R., 2008. Food sup ple men ta tion: a tool to increase repro duc tive output? A case study in the Threatened Flor ida Scrub-Jay. Bio log i cal Con ser va tion 141, 162–173.
Scho ech, S.J., Hahn, T.P., 2008. Lat i tude affects degree of advance ment in lay ing by birds in response to food sup ple men ta tion: a meta-anal y sis. Oec o lo gia 157, 369–376.
Scho ech, S.J., Mum me, R.L., Moore, M.C., 1991. Repro duc tive endo cri nol ogy and mech a nisms of breed ing inhi bi tion in coop er a tively breed ing Flor ida scrub jays (Ap he lo co ma c. coe rules cens). Con dor 93, 354–364.
Scho ech, S.J., Mum me, R.L., Wing field, J.C., 1997. Breed ing sta tus, cor ti co ste rone, and body mass in the coop er a tively breed ing Flor ida scrub-jay (Ap he lo co ma coe rules cens). Phys i o log i cal Zool ogy 70, 68–73.
Scho ech, S.J., Kett er son, E.D., No lan Jr., V., 1999. Exog e nous tes tos ter one and the adre no cor ti cal response in Dark-eyed Jun cos. Auk 116, 64–72.
Sharp, P.J., Dunn, I.C., Wadd ing ton, D., 2008. Chicken lep tin. Gen eral and Com par a-tive Endo cri nol ogy 158, 2–4.
Silv er in, B., 1998. Behav ioral and hor monal responses of the pied fly catcher to envi-ron men tal stress ors. Ani mal Behav iour 55, 1411–1420.
Please cite this article in press as: Scho ech, S.J. et al., Environment, glucoc
(2008), doi:10.1016/j.ygcen.2008.09.009
Smith, G.T., Wing field, J.C., Veit, R.R., 1994. Adre no cor ti cal response to stress in the com mon div ing-petrel, Pele can o ides urin a trix. Phys i o log i cal Zool ogy 67, 526–537.
Sor a to, E., Ko trs chal, K., 2006. Hor monal and behav ioral sym me tries between the sexes in the North ern bald ibis. Gen eral and Com par a tive Endo cri nol ogy 146, 265–274.
Szyf, M., Weaver, I.C.G., Cham pagne, F.A., Dio rio, J., Mea ney, M.J., 2005. Mater nal pro gram ming of ste roid recep tor expres sion and phe no type through DNA meth yl a tion in the rat. Fron tiers in Neu ro en do cri nol ogy 26, 139–162.
Tena-Sem pere, M., Bar ri ero, M.L., Lage, M., Die guez, C., Ca sanu eva, F.F., 2007. Role of lep tin and ghre lin in the reg u la tion of gonadal func tion. Expert Review of Endo-cri nol ogy and Metab o lism 2, 239–249.
Ubuka, T., Uk e na, K., Sharp, P.J., Bent ley, G.E., Tsut sui, K., 2006. Gon a do tro pin-inhib-i tory hor mone inhib its gonadal devel op ment and main te nance by decreas ing gon a do tro pin syn the sis and release in male quail. Endo cri nol ogy 147, 1187–1194.
Walker, B.G., Bo ersma, P.D., Wing field, J.C., 2006. Habit u a tion of adult Mag el lanic pen guins to human vis i ta tion as expressed through behav ior and cor ti co ste-rone secre tion. Con ser va tion Biol ogy 20, 146–154.
Weaver, I.C.G., Mea ney, M.J., Szyf, M., 2006. Mater nal care effects on the hip po cam-pal tran scrip tome and anx i ety-med i ated behav iors in the off spring that are revers ible in adult hood. Pro ceed ings of the National Acad emy of Sci ences of the United States of Amer ica 103, 3480–3485.
Weaver, I.C.G., Szyf, M., Mea ney, M.J., 2002. From mater nal care to gene expres sion:
DNA meth yl a tion and the mater nal pro gram ming of stress responses. Endo-crine Research 28, 699.
Wells, J.C.K., 2003. The thrifty phe no type hypoth e sis: thrifty off spring or thrifty
mother? Jour nal of The o ret i cal Biol ogy 221, 143–161.Wil son, F.E., Fol lett, B.K., 1975. Cor ti co ste rone-induced go nado sup pres sion in phot-
osti mu lat ed tree spar rows. Life Sci ences 17, 1451–1456.Wing field, J.C., 1985a. Influ ences of weather on repro duc tive func tion in female
song spar rows, Me losp iza melo dia. Jour nal of Zool ogy, Lon don 205, 545–558.
Wing field, J.C., 1985b. Short term changes in plasma lev els of hor mones dur-ing estab lish ment and defense of a breed ing ter ri tory in male song spar rows, Me losp iza melo dia. Hor mones and Behav ior 19, 174–187.
Wing field, J.C., De vi che, P., Sharb augh, S., Ast hei mer, L.B., Hol ber ton, R., Suy dam, R., Hunt, K., 1994a. Sea sonal changes of the adre no cor ti cal responses to stress in red polls, Acan this flam mea, in Alaska. Jour nal of Exper i men tal Zool ogy 270, 372–380.
Wing field, J.C., Suy dam, R., Hunt, K., 1994b. The adre no cor ti cal responses to stress in snow bun tings (Plec troph e nax ni va lis) and Lap land long spurs (Calca ri us lap-po ni cus) at Bar row, Alaska. Jour nal of Com par a tive Bio chem i cal Phys i ol ogy C 108, 299–306.
Wing field, J.C., Far ner, D.S., 1993. Endo cri nol ogy of repro duc tion in wild spe cies. In: Far ner, D.S., King, J.R., Parks, K.C. (Eds.), In: Avian Biol ogy, Vol. IX. Aca demic Press, New York, pp. 163–327.
Wing field, J.C., Heg ner, R.E., Lewis, D., 1991. Cir cu lat ing lev els of lutein iz ing hor-mone and ste roid hor mones in rela tion to social sta tus in the coop er a tively breed ing white-browed spar row weaver, Ploce pass er ma hal i. Jour nal of Zool ogy, Lon don 225, 43–58.
Wing field, J.C., Vleck, C.M., Moore, M.C., 1992. Sea sonal changes in adre no cor ti cal response to stress in birds of the Son o ran Des ert. Jour nal of Exper i men tal Zool-ogy 264, 419–428.
Wool fen den, G.E., Fitz pa trick, J.W., 1984. The Flor ida Scrub Jay: Demog ra phy of a
Coop er a tively-Breed ing Bird. Prince ton Uni ver sity Press, Prince ton, NJ.Wool fen den, G.E., Fitz pa trick, J.W., 1990. Flor ida Scrub-Jays: a syn op sis after 18
years of study. In: Sta cey, P.B., Ko enig, W.D. (Eds.), Coop er a tive Breed ing in Birds: Long-Term Stud ies of Ecol ogy and Behav ior. Cam bridge Uni ver sity Press, New York, pp. 239–266.
Wool fen den, G.E., Fitz pa trick, J.W.. 1996. Flor ida Scrub Jay Ap he lo co ma coe rules cens. In: Poole, A., Gill, F. (Eds.), The Birds of North Amer ica, No. 228. The Acad emy of Nat u ral Sci ences, Phil a del phia, PA, and the Amer i can Orni thol o gists’ Union, Wash ing ton, DC, pp. 1–28.
Young, A.J., in press. The causes of phys i o log i cal sup pres sion in ver te brate soci e-ties: a syn the sis. In: Hag er R., Jones, C.B. (Eds.), Repro duc tive Skew in Ver te-brate Soci e ties: Prox i mate and Ulti mate Causes. Cam bridge Uni ver sity Press, Cam bridge.
Young, A.J., Carl son, A.A., Mont fort, S.L., Rus sell, A.F., Ben nett, N.C., Clut ton-Brock, T., 2006. Stress and the sup pres sion of sub or di nate repro duc tion in coop er a-tively breed ing meerk ats. Pro ceed ings of the National Acad emy of Sci ences of the United States of Amer ica 103, 12005–12010.
Zhang, T.Y., Ba got, R., Par ent, C., Nes bitt, C., Bre dy, T.W., Cal dji, C., Fish, E., An is-man, H., Szyf, M., Mea ney, M.J., 2006. Mater nal pro gram ming of defen sive responses through sus tained effects on gene expres sion. Bio log i cal Psy chol-ogy 73, 72–89.
orticoids, and the timing of reproduction..., Gen. Comp. Endocrinol.



















