
Protective Efficacy of N-(2-Hydroxyphenyl) Acetamide against Adjuvant-Induced Arthritis in Rats
Upload
independentCategory
view
0download
0
Vaccine 26 (2008) 3362–3370
Contents lists available at ScienceDirect
Vaccine
journa l homepage: www.e lsev ier .com/ locate /vacc ine
Enhanced immunogenicity of HPV16E7 accompanied by Gp96 as an adjuvant intwo vaccination strategies
Azam Bolhassania,b, Farnaz Zahedifarda, Mohammad Taghikhanib, Sima Rafati a,∗
a Molecular Immunology and Vaccine Research Laboratory, Pasteur Institute of Iran, Tehran, Iranb Department of Clinical Biochemistry, Tarbiat Modarres University, Tehran, Iran
articuc prouiress) likin hu
juvanGp9immand GGp96
ions.d a mytokig E7 aas ims DNA
a r t i c l e i n f o
Article history:Received 3 February 2008Received in revised form 16 March 2008Accepted 19 March 2008Available online 18 April 2008
Keywords:Human papillomavirus (HPV)Cervical cancerE7Heat shock protein (HSP)Glycoprotein 96 (Gp96)DNA vaccinationPrime-boost vaccination
a b s t r a c t
Human papillomavirus, pE7 is the major oncogeniagainst viral infection reqHeat shock proteins (HSPand are currently studiedalong with Gp96 as an adimmune responses by E7 +strategies. In prime-boostopen-reading frame of E7(N-terminal extension of720 in different formulatforms of rGp96 suggestelymphoproliferative and cDNA vaccination includinDNA E7 + Gp96 plasmid wco-delivery of E7 + Gp96 a
induce E7-specific immune res1. Introduction
Human papillomaviruses (HPVs) infections including skin infec-tions, cervical and anal cancers are common all over the worldespecially in Iran. Therefore, an efficient vaccination are highlydemanded to control these infections. The HPV type16 is expressedin the majority of human squamous cell carcinoma. The HPV16E6/E7 oncogene products represent a target of choice for the ther-apeutic vaccination [1,2]. The product of the E7 gene is a relativelysmall phosphoprotein consisting of 98 amino acids. The transform-ing activity of the E7 protein is linked to its interaction with hostregulatory proteins such as the retinoblastoma tumor suppressorprotein. E7 protein can be divided into three main domains CR1, aa1–15; CR2, aa 16–37; CR3, aa 38–98. All three domains are essen-tial for the manifestation of the biological properties of E7 [3,4].E7 elicits special humoral and cellular immune responses due to
∗ Corresponding author. Tel.: +98 21 66953311; fax: +98 21 66465132.E-mail addresses: s [email protected], [email protected]
(S. Rafati).
0264-410X/$ – see front matter © 2008 Elsevier Ltd. All rights reserved.doi:10.1016/j.vaccine.2008.03.082
larly type 16 (HPV16) is present in more than 99% of cervical cancers.tein produced in cervical cancer-associated HPV16. An efficient vaccineinduction of strong humoral and cellular responses against viral proteins.e Gp96 have been described as potent tumor vaccines in animal modelsman clinical trials. In this study, we investigated the utility of HPV16 E7t in C57BL/6 mice model. We compared the level of humoral and cellular6 co-injection as DNA/DNA and prime-boost (DNA/protein) immunizationunization strategies, we first immunized C57BL/6 mice with the complete
p96 (pcDNA-E7 and pcDNA-Gp96) and then boosted with rE7, rNT-gp96) and rCT-gp96 (C-terminal extension of Gp96) mixed with Montanide
The humoral immune responses against rE7 and the different truncatedixed Th1/Th2 response with high intensity toward Th2. Assessment of
ne responses against rE7 and the different fragments of Gp96, showed thatnd Gp96 induced Th1 response. We concluded that co-delivery of nakedmunologically more effective than E7 alone. Our study demonstrated that/DNA and E7 + CT-gp96 as DNA/protein could be an effective approach to
ponses as a potential vaccine candidate for cervical cancer.© 2008 Elsevier Ltd. All rights reserved.
its persistence of expression from the initial stages of infectionthrough transformation and malignant progression. However, E7 isa potential target antigen for cytotoxic T-cell-mediated killing andan ideal antigen for incorporation into a therapeutic vaccine. Theimmunogenicity of E7 has been established in animal models [5].
Since the immunogenicity of most antigens is highly limitedwhen they are applied as pure substances, it is often necessary tomake a fine formulation of antigens with other immune-enhancingsubstances or adjuvant in order to achieve the desired responses forthe control of disease progression [6]. Heat shock proteins (HSPs)have been proposed as the effective adjuvants. HSP preparationsderived from cancer cells and virus-infected cells have been shownpreviously to elicit cancer-specific or virus-specific immunity. Theimmunogenicity of HSP preparations has been attributed to pep-tides associated with the HSPs. These studies demonstrated thatimmunogenic HSP–peptide complexes can also be reconstitutedin vitro. Different studies showed that (a) complexes of HSP70 orGp96 (GRP94) with a variety of synthetic peptides can be gen-erated in vitro; (b) the binding of HSPs with peptides is specificin that a number of other proteins tested do not bind syntheticpeptides under the conditions in which Gp96 molecules do; (c)

ccine
A. Bolhassani et al. / VaHSP–peptide complexes reconstituted in vitro are immunologicallyactive, as tested by their ability to elicit antitumor immunity andspecific CD8+ cytolytic T lymphocyte response; and (d) syntheticpeptides reconstituted in vitro with Gp96 are capable of beingtaken up and re-presented by macrophage in the same manneras Gp96-peptides complexes generated in vivo. These observationsdemonstrated that HSPs are CD8+ T-cell response eliciting adju-vants [7–11]. The Gp96 (GRP94), a 96-kDa glycoprotein, is theendoplasmic reticulum (ER) resident member of the HSP90 fam-ily. The protein possesses an N-terminal signal peptide of 21 aminoacids which is cleaved cotranslationally. The C-terminal end con-tains an ER-retention sequence, KDEL. It has five potential N-linkedglycosylation sites. It contains four high affinity binding sites and11 low affinity binding sites for calcium. Gp96 is a phosphopro-tein in some cell types and is a substrate for casein kinase II. Ithas long been used to augment peptide presentation to T cells.This chaperone binds antigenic peptides, binds to receptors on pro-fessional antigen-presenting cells (APCs), activates these cells andafter internalization, transfers the peptides to MHC class I for acti-vation of T cells [12,13]. It has been shown that the Gp96 fragmentsas mini-chaperones are better choice for immunization [14–16].
The main interest of our investigation is to utilize Gp96 asan adjuvant with viral oncogen HPV16E7 as vaccine candidate inmice model. We evaluated the use of Gp96 fragments aiming toenhance and improve the desired immune response in C57BL/6mice model using both DNA immunization and prime-boost strate-gies. The protective role of the different formulations was evaluatedby measuring antibody production and cytokine release of isolatedsplenocytes at different time intervals.
2. Materials and methods
2.1. Cloning of HPV16E7 and Xenopus Gp96 genes in eukaryoticexpression vector and large scale preparation of endotoxin-freeplasmids
The coding sequence of E7 protein from human papillomavirustype16 (accession number K02718, 294 bp, kindly provided by Prof.T.C. Wu, John Hopkins Medical Institutions, USA) had been clonedinto the BamHI and HindIII cloning sites of the pcDNA3.1(−) vectordownstream of the cytomegalovirus promoter (Invitrogen, Carls-bad, CA). The pcDNA-E7 plasmid was transformed into the DH5�strain of Escherichia coli. Plasmid DNA was purified from recombi-nant colonies by an alkaline lysis method (QIAGEN Plasmid Midi
Kit) verified by restriction enzyme digestion and sequenced usingthe dideoxy chain termination method on an automated sequencer.pBluescript-Gp96 is a plasmid containing the Gp96 gene andwas kindly provided by Dr. Jacques Robert (University of RochesterMedical Center, USA). For the generation of pcDNA-Gp96, XenopusGp96 DNA (accession number AY187545, 2552 bp) was amplifiedby PCR using the primer set designed as follows:
Forward primer: 5′-GGGGATCCACCATGAGGAAGTTTTGGGCA-3′
Reverse primer: 5′-ATGAGCTCGGTACCCGGCCGCGAAAGGTTTTA-3′
Kozak sequence has been shown in bold. Also, the BamHI in for-ward primer and SacI and KpnI restriction sites in reverse primerwere in italics.
The conditions for PCR were as follows: a 50-�l PCR mix-ture comprised of pBluescript-Gp96 as the template (100 ng);deoxynucleoside triphosphates (200 �M of each, Roche); PWO DNApolymerase as a thermostable polymerase (5 U/�l, Roche); 2 �M ofeach primer in the presence of 2 mM MgSO4. The following ampli-fication program was used: 94 ◦C, 1 min; 62 ◦C, 2 min; 72 ◦C for1.5 min for a total of 30 cycles. The amplified Gp96 DNA was then
26 (2008) 3362–3370 3363
cloned into the BamHI/KpnI site of the pcDNA3.1(−) vector. ThepcDNA-Gp96 plasmid was transformed in E.coli XL1-Blue strain.Plasmid DNA was purified by an alkaline lysis method (QIAGENPlasmid Midi Kit). The presence of the inserted Gp96 fragment wasconfirmed by restriction enzyme digestion and gel electrophoresis.The accuracy of this construct was confirmed by DNA sequencing.
Large-scale purification of plasmid DNA samples (pcDNA-E7 andpcDNA-Gp96) was conducted by ion-exchange chromatographywith an Endofree plasmid Giga kit (QIAGEN, Valencia, CA) accord-ing to the manufacturer’s instructions. Plasmid DNA was analyzedby agarose gel electrophoresis and quantified by spectrophotome-try. The 260: 280 UV absorption ratios ranged from 1.8 to 2.0. Thenplasmids were stored in endotoxin-free PBS1X at −20 ◦C until used.
2.2. Expression and purification of the recombinant E7, NT-gp96and CT-gp96 proteins [rE7, rNT-gp96 and rCT-gp96]
The E7 gene was digested from the pcDNA-E7 using BamHI andHindIII and the resulting DNA fragment was gel-purified (QIAGEN).The digested product was cloned into the expression vector pQE30(QIAGEN) which encodes the N-terminal 6XHis-tag. The resultingplasmid is referred to as pQE-E7. The E.coli M15 strain was trans-formed with pQE-E7 and grown at 37 ◦C in LB broth supplementedwith 100 �g/ml ampicillin and 25 �g/ml kanamycin. For theproduction of rE7, the cultures were grown to an optical densityof 0.6–0.8 at 600 nm (OD600 = 0.6–0.8), and protein expression wasinduced with 1 mM IPTG at 37 ◦C. The cell pellets were collectedand stored at −20 ◦C until used for protein purification.
Due to lack of Gp96 expression as tested in different bacterialexpression vectors, attempts were made to clone and expressN-terminal and C-terminal fragments of Gp96 (NT-gp96 andCT-gp96). To reach this end, the NT- and CT-gp96 genes wereobtained from the pBluescript-Gp96 vector. The DNA encodingthe N-terminal and C-terminal fragments of Gp96 consisted of64–1077 and 1645–2552 nucleotide sequences, respectively, werecloned into the BamHI and KpnI sites of expression vector pQE30.For obtaining the NT-gp96 and CT-gp96, four pairs of primers weredesigned (the BamHI, SacI and KpnI restriction sites have beenshown in bold). The forward and reverse primers for amplifyingthe NT-gp96 (NT-1 and NT-2) and CT-gp96 (CT-1 and CT-2) weredesigned as following, respectively:
NT-1: 5′-CGGGGATCCGAAGATGACGTTGAA-3′
NT-2: 5′-ATGAGCTCGGTACCTTTGTAGAAGGCTTTGTA-3′.
CT-1: 5′-GGGGATCCGAGTCCTCTCCATTTGTTGAA-3′CT-2: 5′-ATGAGCTCGGTACCCGGCCGCGAAAGGTTTTA-3′
PCRs were performed under standard conditions (94 ◦C for 1 min;62 ◦C for 2 min and 72 ◦C for 1.5 min for 30 cycles) and the productswere separated on a 0.8% agarose gel. The band correspondingto each product was gel purified (QIAGEN), digested with BamHIand KpnI and ligated into a similarly digested expression plasmidpQE30. The ligation mixture was used to transform E.coli M15strain. The resulting clones were confirmed by sequencing (thedideoxy chain termination method on an automated sequencer).The NT-gp96 and CT-gp96 were induced individually using 1 mMIPTG (as mentioned above).
Protein samples were analyzed by SDS-PAGE in a gel containing15% (w/v) polyacrylamide for HPV16E7 and 12.5% (w/v) poly-acrylamide for NT-gp96 and CT-gp96, followed by staining withCoomassie Brilliant Blue. For western blot analysis, the proteinsresolved on the gel were transferred onto protran nitrocellulosetransfer membrane (Schleicher and Schuell Bioscience, Dassel, Ger-many). The membrane was pre-equilibrated with TBST solution(10 mM Tris–HCl (pH7.4), 150 mM NaCl, and 0.1% Tween 20) con-

ccine
T
trol) and incubated for 5 days at 37 ◦C in an atmosphere of 5% CO2.After supernatant extraction, 1 �Ci of [3H]-thymidine (Amersham,UK) was added to each well and incubated overnight. Cells werecollected on filters with a harvestor and [3H]-thymidine incorpo-ration was determined by liquid scintillation counting. All testswere performed in triplicate, and proliferative responses wereexpressed as their stimulation indices (SI), which represent theratio of mean proliferation after stimulation to the mean prolif-eration of medium controls. IFN-� and IL-5 were measured in thecollected supernatants by commercially available, sandwich-basedELISA kits (R&D, Minneapolis, MN, USA) as described in the kit man-ual. The lower detection limits of IFN-� and IL-5 were 2 and 7 pg/ml,respectively. All tests were performed in duplicate and the medianwas recorded for each set of samples.
3364 A. Bolhassani et al. / Va
taining 2.5% bovin serum albumin (BSA) for overnight and thenreacted with anti-His antibody (QIAGEN) for 2 h at room tempera-ture. After three washes with TBST, the membrane was incubatedwith anti-mouse IgG-HRP (1:2000, Sigma) for 1.5 h at room tem-perature. The immunoreactive protein bands were visualized usingperoxidase substrate named 3, 3′-diaminobenzidine (DAB, Sigma).
Recombinant proteins were purified by affinity chromatographyon Ni-NTA resin using 6XHis-tag. Fast protein liquid chromatogra-phy (FPLC) purification of 6XHis-tagged E7 using Ni-NTA column(Amersham Pharmacia Biotech) was performed under denaturingcondition. The recombinant NT-gp96 and CT-gp96 proteins werealso purified by Ni-NTA chromatography under native conditionsaccording to the manufacturer’s protocol (QIAGEN). The purifiedprotein fractions were concentrated by ultrafiltration (Amicon) anddialyzed against PBS. The protein concentrations were measured bythe BCA procedure (Pierce, Rockford, USA) using BSA as a standardand stored at −70 ◦C, until used.
2.3. PEI-mediated DNA delivery in vitro
For the generation of E7- and Gp96-expressing plasmids(pEGFP-E7 and pEGFP-Gp96), the E7 and Gp96 was subcloned frompcDNA3.1(−) into the BglII–HindIII and BglII–KpnI cloning sites ofthe pEGFP-N1 expression vector, respectively (Clontech, MountainView, CA). The pEGFP-E7 and pEGFP-Gp96 plasmids were trans-formed into the DH5� strain of E.coli. Plasmid DNA were purifiedfrom recombinant colonies by an alkaline lysis method (QIAGENPlasmid Midi Kit) and confirmed by PCR.
COS-7 cells were seeded at 37 ◦C in the presence of 5% CO2into the 6-well plates (Greiner) in complete RPMI 10%. PEI/DNAcomplexes were generated by mixing 10 �M LINPEI 25 kDa (Nr. ofEquivalents (NrE): 5, 7 and 10, Polysciences, Europe) with 5 �g ofeach pEGFP, pEGFP-E7 and pEGFP-Gp96 in a final volume of 200 �land incubating at room temperature for 15 min. After washing cellswith serum-free media, PEI/DNA complexes were added to eachwell. Plates were incubated at 37 ◦C for 6 h. Complete RPMI 10% wasadded to each well and the level of GFP expression of each constructwas assessed by fluorescence microscopy at different time intervals(24 and 48 h).
2.4. Mice immunization
Inbred C57BL/6 female mice, 6–8 week old, were obtained fromthe breeding stocks maintained at the Pasteur Institute of Iran.
Eleven groups of 10 mice were selected. For priming, all mice wereimmunized subcutaneously at the footpad with 100 �g of plasmidDNA encoding either E7, Gp96 or the combination of E7 and Gp96.For boosting (3 weeks after priming), groups 1–3 received DNAconstructs similar to priming, but groups 4–8 immunized withcorresponding recombinant E7, NT-gp96 and CT-gp96 proteinsemulsified with Montanide ISA720 (M720) at the ratio of 7:3(oil:aqueous phase). The control groups 9–11 were injected PBS,empty pcDNA3.1(−) plasmid and PBS + Montanide, respectively.Table 1 shows the immunization regimens of each group.2.5. Monitoring humoral immune response by ELISA
Mice were bled from retro-orbital and sera were prepared fromthe whole blood samples of each mouse at different time inter-vals before, 3, 6 and 9 weeks after the booster injection. Pooledsera of each group were stored at −20 ◦C. ELISA was performedwith rE7 and rE7 + rNT-gp96 + rCT-gp96 for the identification ofantigen-specific total IgG, IgG1 and IgG2a. Briefly, 96-microwellplates (NUNC) were coated overnight at 4 ◦C with 100 �l of eitherrE7 (5 �g/ml) or the combination of rE7 (5 �g/ml) + rNT-gp96
26 (2008) 3362–3370
(2.5 �g/ml) + rCT-gp96 (10 �g/ml) diluted in PBS (pH 7.2). Afterwashing with PBS containing 0.05% (v/v) Tween 20 (PBS-Tween)and blocking with 1% BSA in PBS for 2 h at 37 ◦C, 100 �l of micesera (diluted 1:50 in 1% BSA/PBS-Tween) was added and the plateswere incubated at 37 ◦C for 2 h. After washing, 100 �l of eitherbiotinylated goat anti-mouse IgG (diluted 1:10,000 in 1% BSA/PBS-ween, Cedarlane Laboratories, Hornby, Ontario, Canada), or rat
anti-mouse IgG1 or IgG2a conjugated with Biotin (diluted 1:1000in 1% BSA/PBS-Tween, Cedarlane Laboratories, Hornby, Ontario,Canada) was added and the plates were incubated at 37 ◦C for 2 h.Then the plates were washed and incubated with 1:600 dilutionof streptavidin-horseradish peroxidase (Cedarlane Laboratories,Hornby, Ontario, Canada) at 37 ◦C for 1 h. Detection was done with100 �l of O-Phenylenediamine (OPD, Sigma) as the substrate in cit-rate phosphate buffer (citric acid 0.1 M, Na2HPO4 0.2 M, pH 4.5).After 30 min, the enzyme reaction was stopped by 1 M sulfuric acidand the absorbance was measured at 492 nm.
2.6. Lymphoproliferative responses and cytokine detection
Three and six weeks after booster, two mice from each groupwere scarificed and the spleens were removed. A single cell sus-pension of splenocytes was seeded in complete RPMI medium1640 supplemented with 5% FCS, 2mM glutamine, 5 × 10−5 mM 2-mercaptoethanol (2-ME), 10 mM HEPES and 40 �g/ml gentamycin.Cells were incubated in U-bottomed, 96-well microtiter plates(Costar, Cambridge, MA) at a density of 2 × 106 cells/ml in the pres-ence of 10 �g/ml of all recombinant proteins (rE7, rNT-gp96 andrCT-gp96) and 5 �g/ml of concanavalinA (ConA, as positive con-
2.7. Statistical analysis
The differences in the level of cellular and antibody responseswere determined by one-way ANOVA. A value less than 0.05(p < 0.05) was used for statistical significance.
Table 1DNA and prime-boost immunization in C57BL/6 mice by using E7 in the differentformulations
Group First week Third week
1 DNA [E7] DNA [E7]2 DNA [Gp96] DNA [Gp96]3 DNA [E7 + Gp96] DNA [E7 + Gp96]4 DNA [E7] Protein [rE7 + Montanide]5 DNA [Gp96] Protein [rNT-gp96 + rCT-gp96 + Montanide]6 DNA [E7 + Gp96] Protein [rE7 + rNT-gp96 + Montanide]7 DNA [E7 + Gp96] Protein [rE7 + rCT-gp96 + Montanide]8 DNA [E7 + Gp96] Protein [rE7 + rNT-gp96 + rCT-gp96 + Montanide]9 Control [PBS] Control [PBS]10 Control [empty vector] Control [empty vector]11 Control [empty vector] Control [PBS + Montanide]
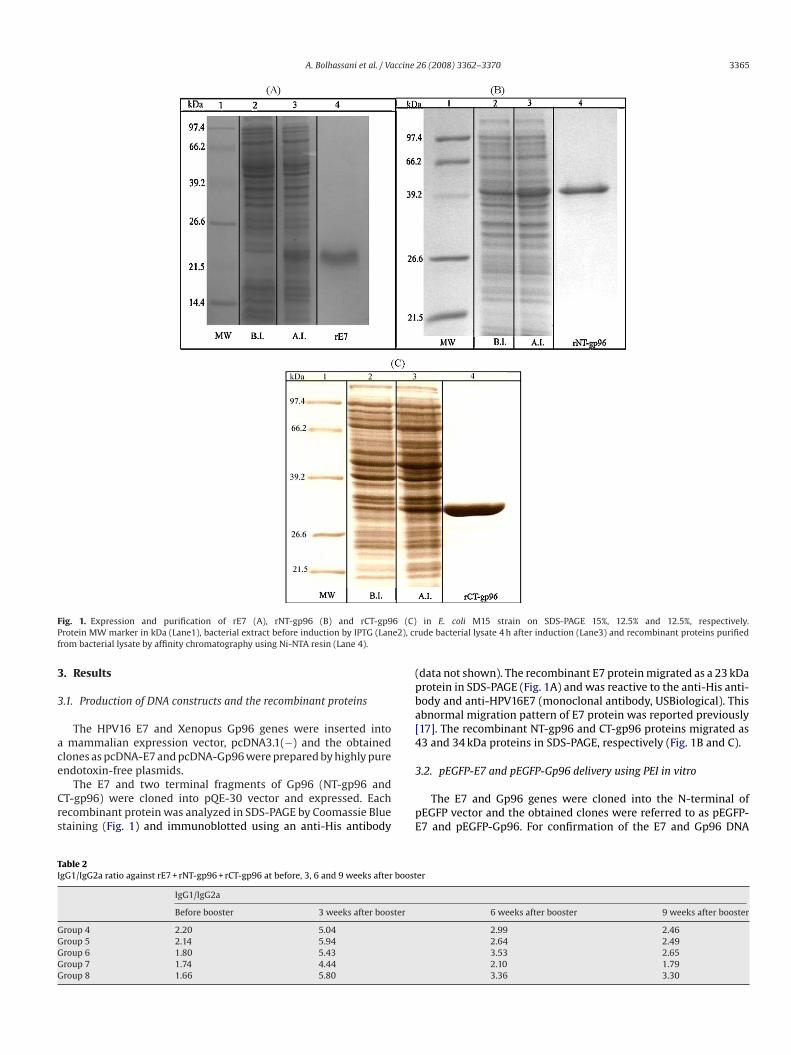
A. Bolhassani et al. / Vaccine 26 (2008) 3362–3370 3365
Fig. 1. Expression and purification of rE7 (A), rNT-gp96 (B) and rCT-gp96 (C)Protein MW marker in kDa (Lane1), bacterial extract before induction by IPTG (Lane2), crfrom bacterial lysate by affinity chromatography using Ni-NTA resin (Lane 4).
3. Results
3.1. Production of DNA constructs and the recombinant proteins
The HPV16 E7 and Xenopus Gp96 genes were inserted intoa mammalian expression vector, pcDNA3.1(−) and the obtainedclones as pcDNA-E7 and pcDNA-Gp96 were prepared by highly pureendotoxin-free plasmids.
The E7 and two terminal fragments of Gp96 (NT-gp96 andCT-gp96) were cloned into pQE-30 vector and expressed. Eachrecombinant protein was analyzed in SDS-PAGE by Coomassie Bluestaining (Fig. 1) and immunoblotted using an anti-His antibody
Table 2IgG1/IgG2a ratio against rE7 + rNT-gp96 + rCT-gp96 at before, 3, 6 and 9 weeks after boost
IgG1/IgG2a
Before booster 3 weeks after booster
Group 4 2.20 5.04Group 5 2.14 5.94Group 6 1.80 5.43Group 7 1.74 4.44Group 8 1.66 5.80
in E. coli M15 strain on SDS-PAGE 15%, 12.5% and 12.5%, respectively.ude bacterial lysate 4 h after induction (Lane3) and recombinant proteins purified
(data not shown). The recombinant E7 protein migrated as a 23 kDaprotein in SDS-PAGE (Fig. 1A) and was reactive to the anti-His anti-body and anti-HPV16E7 (monoclonal antibody, USBiological). Thisabnormal migration pattern of E7 protein was reported previously[17]. The recombinant NT-gp96 and CT-gp96 proteins migrated as43 and 34 kDa proteins in SDS-PAGE, respectively (Fig. 1B and C).
3.2. pEGFP-E7 and pEGFP-Gp96 delivery using PEI in vitro
The E7 and Gp96 genes were cloned into the N-terminal ofpEGFP vector and the obtained clones were referred to as pEGFP-E7 and pEGFP-Gp96. For confirmation of the E7 and Gp96 DNA
er
6 weeks after booster 9 weeks after booster
2.99 2.462.64 2.493.53 2.652.10 1.793.36 3.30
ccine
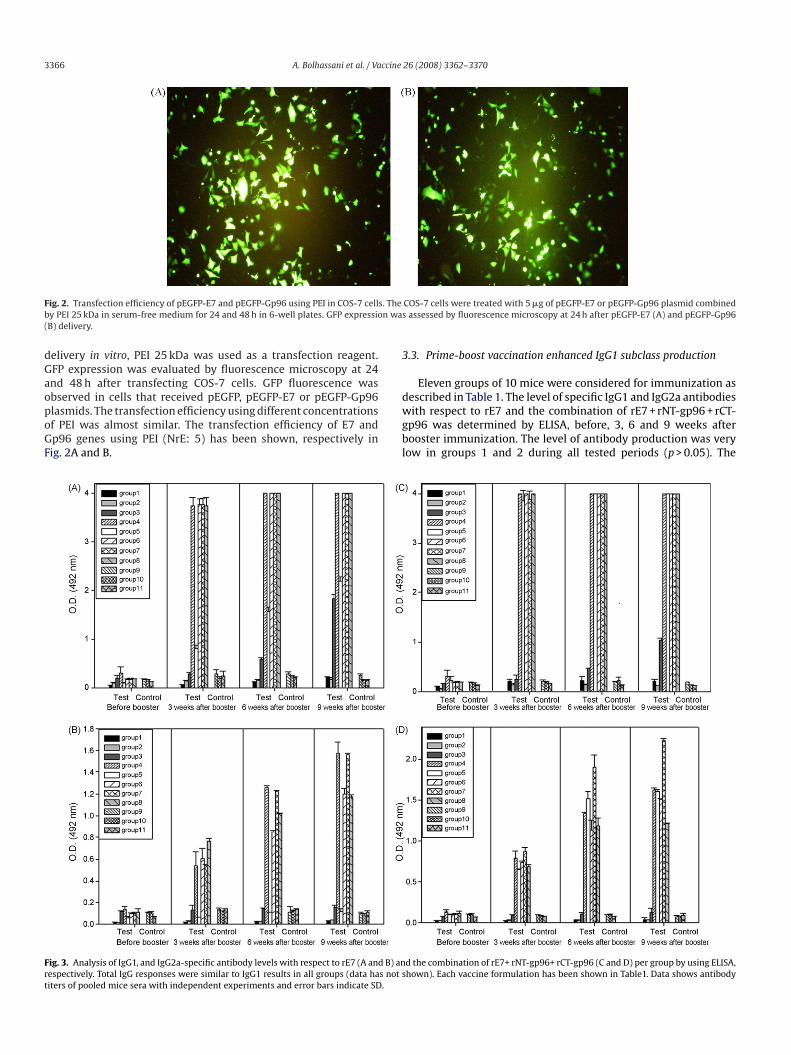
3366 A. Bolhassani et al. / VaFig. 2. Transfection efficiency of pEGFP-E7 and pEGFP-Gp96 using PEI in COS-7 cells. Theby PEI 25 kDa in serum-free medium for 24 and 48 h in 6-well plates. GFP expression wa(B) delivery.
delivery in vitro, PEI 25 kDa was used as a transfection reagent.GFP expression was evaluated by fluorescence microscopy at 24and 48 h after transfecting COS-7 cells. GFP fluorescence wasobserved in cells that received pEGFP, pEGFP-E7 or pEGFP-Gp96plasmids. The transfection efficiency using different concentrationsof PEI was almost similar. The transfection efficiency of E7 andGp96 genes using PEI (NrE: 5) has been shown, respectively inFig. 2A and B.
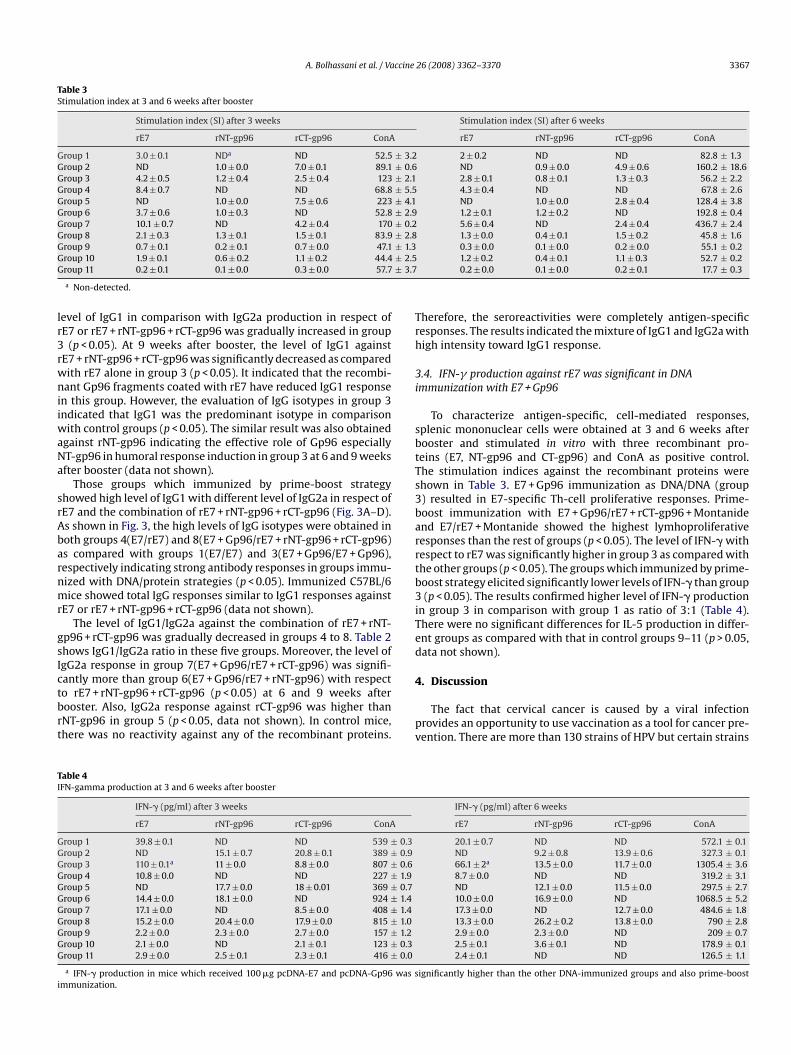
Fig. 3. Analysis of IgG1, and IgG2a-specific antibody levels with respect to rE7 (A and B) anrespectively. Total IgG responses were similar to IgG1 results in all groups (data has nottiters of pooled mice sera with independent experiments and error bars indicate SD.
26 (2008) 3362–3370
COS-7 cells were treated with 5 �g of pEGFP-E7 or pEGFP-Gp96 plasmid combineds assessed by fluorescence microscopy at 24 h after pEGFP-E7 (A) and pEGFP-Gp96
3.3. Prime-boost vaccination enhanced IgG1 subclass production
Eleven groups of 10 mice were considered for immunization asdescribed in Table 1. The level of specific IgG1 and IgG2a antibodieswith respect to rE7 and the combination of rE7 + rNT-gp96 + rCT-gp96 was determined by ELISA, before, 3, 6 and 9 weeks afterbooster immunization. The level of antibody production was verylow in groups 1 and 2 during all tested periods (p > 0.05). The
d the combination of rE7+ rNT-gp96+ rCT-gp96 (C and D) per group by using ELISA,shown). Each vaccine formulation has been shown in Table1. Data shows antibody
ccine
± 3.2± 0.6± 2.1± 5.5± 4.1± 2.9± 0.2± 2.8± 1.3± 2.5± 3.7
A. Bolhassani et al. / Va
Table 3Stimulation index at 3 and 6 weeks after booster
Stimulation index (SI) after 3 weeks
rE7 rNT-gp96 rCT-gp96 ConA
Group 1 3.0 ± 0.1 NDa ND 52.5Group 2 ND 1.0 ± 0.0 7.0 ± 0.1 89.1Group 3 4.2 ± 0.5 1.2 ± 0.4 2.5 ± 0.4 123Group 4 8.4 ± 0.7 ND ND 68.8Group 5 ND 1.0 ± 0.0 7.5 ± 0.6 223Group 6 3.7 ± 0.6 1.0 ± 0.3 ND 52.8Group 7 10.1 ± 0.7 ND 4.2 ± 0.4 170Group 8 2.1 ± 0.3 1.3 ± 0.1 1.5 ± 0.1 83.9Group 9 0.7 ± 0.1 0.2 ± 0.1 0.7 ± 0.0 47.1Group 10 1.9 ± 0.1 0.6 ± 0.2 1.1 ± 0.2 44.4Group 11 0.2 ± 0.1 0.1 ± 0.0 0.3 ± 0.0 57.7
a Non-detected.
level of IgG1 in comparison with IgG2a production in respect ofrE7 or rE7 + rNT-gp96 + rCT-gp96 was gradually increased in group3 (p < 0.05). At 9 weeks after booster, the level of IgG1 againstrE7 + rNT-gp96 + rCT-gp96 was significantly decreased as comparedwith rE7 alone in group 3 (p < 0.05). It indicated that the recombi-nant Gp96 fragments coated with rE7 have reduced IgG1 responsein this group. However, the evaluation of IgG isotypes in group 3indicated that IgG1 was the predominant isotype in comparisonwith control groups (p < 0.05). The similar result was also obtainedagainst rNT-gp96 indicating the effective role of Gp96 especially
NT-gp96 in humoral response induction in group 3 at 6 and 9 weeksafter booster (data not shown).Those groups which immunized by prime-boost strategyshowed high level of IgG1 with different level of IgG2a in respect ofrE7 and the combination of rE7 + rNT-gp96 + rCT-gp96 (Fig. 3A–D).As shown in Fig. 3, the high levels of IgG isotypes were obtained inboth groups 4(E7/rE7) and 8(E7 + Gp96/rE7 + rNT-gp96 + rCT-gp96)as compared with groups 1(E7/E7) and 3(E7 + Gp96/E7 + Gp96),respectively indicating strong antibody responses in groups immu-nized with DNA/protein strategies (p < 0.05). Immunized C57BL/6mice showed total IgG responses similar to IgG1 responses againstrE7 or rE7 + rNT-gp96 + rCT-gp96 (data not shown).
The level of IgG1/IgG2a against the combination of rE7 + rNT-gp96 + rCT-gp96 was gradually decreased in groups 4 to 8. Table 2shows IgG1/IgG2a ratio in these five groups. Moreover, the level ofIgG2a response in group 7(E7 + Gp96/rE7 + rCT-gp96) was signifi-cantly more than group 6(E7 + Gp96/rE7 + rNT-gp96) with respectto rE7 + rNT-gp96 + rCT-gp96 (p < 0.05) at 6 and 9 weeks afterbooster. Also, IgG2a response against rCT-gp96 was higher thanrNT-gp96 in group 5 (p < 0.05, data not shown). In control mice,there was no reactivity against any of the recombinant proteins.
Table 4IFN-gamma production at 3 and 6 weeks after booster
IFN-� (pg/ml) after 3 weeks
rE7 rNT-gp96 rCT-gp96 ConA
Group 1 39.8 ± 0.1 ND ND 539 ± 0.3Group 2 ND 15.1 ± 0.7 20.8 ± 0.1 389 ± 0.9Group 3 110 ± 0.1a 11 ± 0.0 8.8 ± 0.0 807 ± 0.6Group 4 10.8 ± 0.0 ND ND 227 ± 1.9Group 5 ND 17.7 ± 0.0 18 ± 0.01 369 ± 0.7Group 6 14.4 ± 0.0 18.1 ± 0.0 ND 924 ± 1.4Group 7 17.1 ± 0.0 ND 8.5 ± 0.0 408 ± 1.4Group 8 15.2 ± 0.0 20.4 ± 0.0 17.9 ± 0.0 815 ± 1.0Group 9 2.2 ± 0.0 2.3 ± 0.0 2.7 ± 0.0 157 ± 1.2Group 10 2.1 ± 0.0 ND 2.1 ± 0.1 123 ± 0.3Group 11 2.9 ± 0.0 2.5 ± 0.1 2.3 ± 0.1 416 ± 0.0
a IFN-� production in mice which received 100 �g pcDNA-E7 and pcDNA-Gp96 was simmunization.
26 (2008) 3362–3370 3367
Stimulation index (SI) after 6 weeks
rE7 rNT-gp96 rCT-gp96 ConA
2 ± 0.2 ND ND 82.8 ± 1.3ND 0.9 ± 0.0 4.9 ± 0.6 160.2 ± 18.62.8 ± 0.1 0.8 ± 0.1 1.3 ± 0.3 56.2 ± 2.24.3 ± 0.4 ND ND 67.8 ± 2.6ND 1.0 ± 0.0 2.8 ± 0.4 128.4 ± 3.81.2 ± 0.1 1.2 ± 0.2 ND 192.8 ± 0.45.6 ± 0.4 ND 2.4 ± 0.4 436.7 ± 2.41.3 ± 0.0 0.4 ± 0.1 1.5 ± 0.2 45.8 ± 1.60.3 ± 0.0 0.1 ± 0.0 0.2 ± 0.0 55.1 ± 0.21.2 ± 0.2 0.4 ± 0.1 1.1 ± 0.3 52.7 ± 0.20.2 ± 0.0 0.1 ± 0.0 0.2 ± 0.1 17.7 ± 0.3
Therefore, the seroreactivities were completely antigen-specificresponses. The results indicated the mixture of IgG1 and IgG2a withhigh intensity toward IgG1 response.
3.4. IFN-� production against rE7 was significant in DNAimmunization with E7 + Gp96
To characterize antigen-specific, cell-mediated responses,splenic mononuclear cells were obtained at 3 and 6 weeks afterbooster and stimulated in vitro with three recombinant pro-
teins (E7, NT-gp96 and CT-gp96) and ConA as positive control.The stimulation indices against the recombinant proteins wereshown in Table 3. E7 + Gp96 immunization as DNA/DNA (group3) resulted in E7-specific Th-cell proliferative responses. Prime-boost immunization with E7 + Gp96/rE7 + rCT-gp96 + Montanideand E7/rE7 + Montanide showed the highest lymhoproliferativeresponses than the rest of groups (p < 0.05). The level of IFN-� withrespect to rE7 was significantly higher in group 3 as compared withthe other groups (p < 0.05). The groups which immunized by prime-boost strategy elicited significantly lower levels of IFN-� than group3 (p < 0.05). The results confirmed higher level of IFN-� productionin group 3 in comparison with group 1 as ratio of 3:1 (Table 4).There were no significant differences for IL-5 production in differ-ent groups as compared with that in control groups 9–11 (p > 0.05,data not shown).4. Discussion
The fact that cervical cancer is caused by a viral infectionprovides an opportunity to use vaccination as a tool for cancer pre-vention. There are more than 130 strains of HPV but certain strains
IFN-� (pg/ml) after 6 weeks
rE7 rNT-gp96 rCT-gp96 ConA
20.1 ± 0.7 ND ND 572.1 ± 0.1ND 9.2 ± 0.8 13.9 ± 0.6 327.3 ± 0.166.1 ± 2a 13.5 ± 0.0 11.7 ± 0.0 1305.4 ± 3.68.7 ± 0.0 ND ND 319.2 ± 3.1ND 12.1 ± 0.0 11.5 ± 0.0 297.5 ± 2.710.0 ± 0.0 16.9 ± 0.0 ND 1068.5 ± 5.217.3 ± 0.0 ND 12.7 ± 0.0 484.6 ± 1.813.3 ± 0.0 26.2 ± 0.2 13.8 ± 0.0 790 ± 2.82.9 ± 0.0 2.3 ± 0.0 ND 209 ± 0.72.5 ± 0.1 3.6 ± 0.1 ND 178.9 ± 0.12.4 ± 0.1 ND ND 126.5 ± 1.1
ignificantly higher than the other DNA-immunized groups and also prime-boost

ccine
3368 A. Bolhassani et al. / Vaare considered more oncogenic than the others [18–20]. Approxi-mately 50–60% of these cancers contain HPV16 and another 10–20%contains HPV18. Therefore, these two HPV genotypes have beenthe focus of most vaccine development efforts. The HPV oncopro-teins E6 and E7 represent ideal targets for HPV vaccine development[18–21].
The development of an improved, effective vaccine againstviral disease is highly demanded. Ideal therapeutic vaccines forinfectious diseases and cancer should elicit not only the humoralresponse, but also the cellular response [22]. It has been reportedthat without suitable adjuvants, a small synthetic peptide or arecombinant protein can elicit only a weak CTL response. Thus,the antigens need to be combined with an efficient adjuvant ordelivery system with potential for use in humans. In this context,heat shock proteins have emerged as an attractive immunostimu-latory component for vaccines. Several heat shock proteins (HSPs)like Gp96 act as potent adjuvants for eliciting anti-tumor immu-nity. The immunomodulatory properties of Gp96 may be usedfor antigen-specific activation or suppression of cellular immuneresponses [23–30]. We investigated the immunogenic potential ofGp96 for utilization in immunization strategies in C57BL/6 micemodel.
Our major goals of this study include (1) to determine thepotency of DNA and prime-boost immunization strategies usingHPV16E7 as antigenic model which is associated with cervical can-cer and Gp96 as an adjuvant and (2) to clarify Gp96 effects as anadjuvant on the immune responses without covalent binding to E7.
In the present study, naked DNA constructs of E7 and Gp96were prepared. The recombinant E7 and Gp96 (N-terminal andC-terminal of Gp96) proteins were expressed in E. coli as His-tagged proteins and purified by affinity chromatography usingFPLC system. The DNA delivery was confirmed using the GFPreporter construct and LINPEI 25 kDa, in vitro. The cationic poly-mer polyethylenimines (PEIs) have been widely used as nonviralgene-delivery vehicles. PEIs with a molecular weight (MW) of25,000 Da or greater have shown high transfection efficiency bothin vitro and in vivo [31,32]. In this experiment, the COS-7 eukaryoticcells transfected with pEGFP-E7 and pEGFP-Gp96 exhibited a fairlyhigh GFP fluorescence. Furthermore, we evaluated foot pad DNAimmunization using the E7, Gp96 and E7 + Gp96 genes in C57BL/6mice (groups 1–3). Also, we simultaneously evaluated the vaccinecandidate potential of E7 using Gp96 as an adjuvant by a prime-boost strategy in C57BL/6 mice (groups 4–8). Several papers haverecently highlighted the power of prime-boost strategies in elicit-
ing protective cellular immunity to a variety of pathogens and havedemonstrated efficacy in humans [33–36].The entire ORF of Xenopus Gp96 was used for priming as nakedDNA, since there was a great homology between Xenopus, mouseand human. In vertebrates from man to frogs, the heat shock proteinGp96 elicits T-cell responses against antigenic peptides that it chap-erones. In addition, at least in Xenopus, Gp96 can prime CD8+ T-celleffectors that are not MHC restricted. The high degree of structuralconservation of HSP and its potential to cross-present antigen hasbeen published by the other studies [37–40]. This strongly sug-gests that the HSP–APC interaction is phylogenetically conserved,including the surface receptor that binds HSPs [40].
For a booster injection, we applied the recombinant E7 proteinas well as the truncated Gp96 in different formulations emulsifiedwith Montanide 720. To understand the immunological mecha-nisms underlying the results, we analyzed the humoral immuneresponses at four different periods. A criterion for vaccine designis the ability of the antigen to modulate the immune response,specifically the T helper (Th) subtype. IgG isotypes are often usedas indicators for the type of Th immune responses in vivo. Produc-tion of the IgG2a is normally associated with IFN-� secretion and
26 (2008) 3362–3370
the development of a Th1 immune response. However, in contrast,production of the IgG1 is normally associated with IL-5 secretionand the development of Th2 immune responses.
DNA vaccines with high potency are an attractive approach forgenerating antigen-specific immunotherapy. In this study, nakedDNA E7 revealed a low immunogenicity. The mice immunized withE7/E7 as DNA/DNA did not show a strong antibody response andin fact, it is a very poor inducer of an antibody response, whenbeing used as antigen in DNA immunization. Most of the proteingenerated after DNA immunization in cells is not capable of prim-ing an immune response, then we need to enhance the potencyof genetic vaccination by facilitating transport of these antigensinto the MHC class I antigen presentation pathway of DCs. Manystudies have shown that DNA vaccines can stimulate both humoraland cellular immune responses. However, the cellular uptake ofinjected plasmid DNA is very inefficient. In published reports bythe others, naked DNA E7 has been shown as a very poor inducerof a cytotoxic T-cell response. The immunogenicity of E7 has beenimproved in DNA vaccination by fusion to the gene of a heat shockprotein or to sequences coding for an export signal. Linkage of anti-gens to HSPs (such as HSP70, HSP60, Calreticulin and Gp96) hasbeen shown as a potential approach for increasing the potencyof DNA vaccines. It has been demonstrated that DNA vaccinationwith this construct strongly enhanced an antigen-specific CD8+T–cell response as well as a specific B cell response in mice. Forexample, vaccines containing E7-HSP70 fusion genes increased thefrequency of E7-specific CD8+ T cells by at least 30-fold relativeto vaccines containing the wild-type E7 gene. Also, DNA vaccina-tion using Gp96-peptide fusion proteins against an intracellularbacterial pathogen revealed profound epitope-specific IFN-� andcytotoxic T-cell responses [41–54].
However, DNA vaccines have been widely used to develop vac-cines against various pathogens as well as for cancer. In currentstudy, naked DNA E7 + Gp96 immunization gradually enhancedIgG1 response against rE7 or rE7 + rNT-gp96 + rCT-gp96 signif-icantly higher than the control groups. This result was alsoconfirmed against rNT-gp96 indicating that Gp96 drives humoralimmune responses in vivo. Prime-boost immunizations withthe recombinant proteins in different formulations (groups 4–8)showed significantly elevated systemic IgG production in compar-ison with group 3 and also control groups. Furthermore, therewere also significantly enhanced levels of IgG1 and IgG2a incombination with or without rNT-gp96 and rCT-gp96. IgG2a pro-duction using the different formulations was significantly higher
as compared with DNA immunization versus the control groups.However, induction of antibody titer was not significantly observedin Montanide 720-injected group as negative control. IgG1/IgG2aratios gradually decreased in groups 4–8. Also, coinjection of E7with CT-gp96 (group 7) indicated higher IgG2a response thanthe other formulations with respect to rE7 + rNT-gp96 + rCT-gp96,especially at 9 weeks after booster. Here, we have shown thatthree recombinant proteins are highly immunogenic in prime-boost strategy. Surprisingly, our results indicate considerable levelof IgG1 response against rE7 in group 5 which did not receive E7as part of vaccine component and probably demonstrate the Gp96conformational changes [55]. All together, our data suggest a mixedTh1/Th2 response more towards Th2 were switched by the differ-ent formulations for E7 as prime-boost immunization. It has beenclearly demonstrated that immune responses observed in the micewere antigen-specific and adjuvant-dependent. According to thepublished data especially about DNA vaccines, assays to determineTh1/Th2 bias are useful in comparing different regimens, but theydo not always reflect the extent of cellular or humoral responsesthat have been elicited; both antibody and CTL responses has beenfound in an animal with bias to Th1 or Th2 [56,57]. The previ-
ccine
[
[
[
[
[
[
A. Bolhassani et al. / Va
ous studies have indicated that an established Th2 response to atumor-specific antigens (TSA) does not prevent the developmentof TSA-specific tumor protective CTL. This result has been obtainedby mice immunization with E7/Algammulin and GF001/Quil-A. Infact, Th2-type CD4+ cells neither enhance nor suppress antitumorCTL activity in a mouse tumor model [58].
Analysis of the cellular immune response exhibited significantdifferences between groups. To evaluate the production level of IFN-�, cells were stimulated with rE7, rNT-gp96 and rCT-gp96 proteins.IFN-� levels in the supernatant of splenocytes from mice injectedwith E7 + Gp96 as DNA/DNA (group 3) were significantly higherthan the other groups (p < 0.05). This illustrates that E7 in the pres-ence of Gp96 drives T-cell responses towards a Th1-type fashion.This data is in agreement with other reports which indicated thatimmunization of mice with the E7 antigen fused to Hsp65 markedlyalters the E7 recall response from IL-5 to IFN-� production, suggest-ing a shift toward a type-1 response [59].
In addition, Th-cell proliferative responses with respect to rE7were enhanced and significantly higher in groups 4 and 7 than theother groups with fairly more response in group 7 in a manner sim-ilar to antibody responses. Furthermore, the mice immunized withdifferent formulations of E7 as prime-boosting indicated high anti-body titers and low cytokine (IFN-�) responses in comparison withDNA immunization. In groups 4–8, IFN-� responses were signifi-cantly increased as compared with the control groups (p < 0.05).
Heat shock proteins are involved in eliciting a potent, specific,cellular adaptive immune response, which was suggested to bedependent on their ability to chaperone a large variety of pep-tides. Immunotherapies using HSPs to generate specific antitumorresponses have been evaluated in clinical studies. Gp96, as one ofthe most abundant intracellular HSPs, possesses multiple functions.Among these functions, its ability to bridge the innate and adaptiveimmune systems has attracted extensive interest. Different inves-tigations have shown that Gp96 and its N-terminal fragment couldserve as potential adjuvants to enhance the peptide-specific CTLresponse in mice. Also, the findings implied that the N-terminal partof Gp96 might contain the motif to stimulate the innate immunesystem. These results have clearly demonstrated the potential ofusing Gp96 or its N-terminal fragment as a possible adjuvant to aug-ment CTL response against HBV infection and HCC [14–16]. But withregard to our data, C-terminal fragment of Gp96 is also effectiveand potent in production of humoral (IgG1 and IgG2a) and T-cell-mediated immune responses. On the other hand, co-delivery of E7with Gp96 without covalent linkage can produce specific immune
response to E7 as an antigen.In summary, this study indicates the potential value of DNAvaccination using co-delivery of E7 + Gp96. DNA vaccination usingE7 and Gp96 revealed E7-specific IFN-� response. Our datashows that such a combined DNA vaccine is successful in elic-iting the proper immune response. However, the co-delivery ofE7 + Gp96 as DNA/DNA and E7 + CT-gp96 as DNA/protein could bean effective approach to induce E7-specific immune responsesas a potential vaccine candidate for cervical cancer. Gp96 stim-ulates cell-mediated immunity and modulates antibody isotypeinduction. This study likely provides an additional option for induc-ing E7-specific T-cell-mediated immune responses as a possibleimmune therapy modality against HPV-associated cervical cancer.However, more research on Gp96 and its fragments are required inorder to provide more potent vaccine candidate.
Acknowledgments
The financial support by the Research Council of the RepublicPresidentship is gratefully acknowledged. We would like to thank
[
[
[
[
[
[
[
[
[
[
[
[
26 (2008) 3362–3370 3369
A. Javadi (Pasteur Institute of Iran, Department of Immunology), Dr.Hoorieh Soleimanjahi (Tarbiat Modarres University, Departmentof Virology) and N. Moradin, E. Gholami, and S. Alizadeh (PasteurInstitute of Iran, Molecular Immunology and Vaccine Research Lab-oratory) for their technical assistance.
References
[1] Bubenik J. Animal models for development of therapeutic HPV16 vaccines. IntJ Oncol 2002;20:207–12.
[2] Kotecha M, Afghan R, Vasilikopoulou E, Wilson E, Marsh P, Davies DH.Enhanced tumour growth after DNA vaccination against human papillomavirusE7 oncoprotein: evidence for tumour-induced immune deviation. Vaccine2003;21:2506–15.
[3] Vousden K, Jat PS. Functional similarity between HPV16E7 SV40 large T andadenovirus E1A proteins. Oncogene 1989;4:153–8.
[4] Dyson N, Howley PM, Munger K, Harlow E. The human papillomavirus-16E7 oncoprotein is able to bind to the retinoblastoma gene product. Science1989;243:934–7.
[5] Mirecka EA, Rudolph R, Hey T. Expression and purification of His-taggedHPV16E7 protein active in pRb binding. Prot Expr Purif 2006;48:281–91.
[6] Kaufmann SHE, Hilf N, Radsak M, Schild H. Host-derived adjuvants. Novel Vac-cination Strategies 2004:129–45.
[7] Segal B, Wang XY, Dennis C, Youn R, Repasky E, Manjili M, et al. Heatshock proteins as vaccine adjuvants infections and cancer. Drug Discov Today2006;11:534–40.
[8] More S, Breloer M, Fleischer B, Bonin AV. Activation of cytotoxic T cells in vitro byrecombinant gp96 fusion proteins irrespective of the “fused” antigenic peptidesequence. Immunol Lett 1999;69(2):275–82.
[9] Menoret A, Bell G. Purification of multiple heat shock proteins from a singletumor sample. J Immunol Methods 2000;237:119–30.
10] Binder RJ, Vatner R, Srivastava P. The heat shock protein receptors: someanswers and more questions. Tissue Antigens 2004;64:442–51.
[11] Enomoto Y, Bharti A, Khaleque AA, Song B, Liu C, Apostolopoulos V. Enhancedimmunogenicity of heat shock protein 70 peptide complexes from dendriticcell-tumor fusion cells. J Immunol 2006;177:5946–55.
12] Yang Y, Li Z. Roles of heat shock protein gp96 in the ER quality control: redun-dant or unique function? Molecules Cells 2005;20(2):173–82.
13] Facciponte J, Wang XY, MacDonald I, Park J, Arnouk H, Manjili M. Heat shockproteins HSP70 and Gp96: structural insights. Cancer Immunol Immunother2006;55:339–46.
14] Li H, Zhou M, Han J, Zhu X, Dong T, Gao GF, et al. Generation of murine CTLby a Hepatitis B virus-specific peptide and evaluation of the adjuvant effectof heat shock protein glycoprotein 96 and its terminal fragments. J Immunol2005;174:195–204.
15] Biswas C, Sriram U, Ciric B, Ostrovsky O, Gallucci S, Argon Y. The N-terminalfragment of GRP94 is sufficient for peptide presentation via professionalantigen-presenting cells. Int Immunol 2006;18(7):1147–57.
16] Warger T, Hilf N, Rechtsteiner G, Haselmayer P, Carrick DM, Jonuleit H,et al. Interaction of TLR2 and TLR4 ligands with the N-terminal domainof Gp96 amplifies innate and adaptive immune responses. J Biol Chem2006;281(32):22545–53.
[17] Armstrong DJ, Roman A. The anomalous electrophoretic behavior of the humanpapillomavirus type 16 E7 protein is due to the high content of acidic amino
acid residues. Biochem Biophys Res Commun 1993;192(3):1380–7.18] Schiller J, Davies P. Delivering on the promise: HPV vaccines and cervical cancer.Nat Rev 2004;2:343–7.
19] Damania B. DNA tumor viruses and human cancer. Trends Microbiol2006;15(1):38–44.
20] Hausen HZ, Villiers EMD. Human papillomaviruses. Ann Rev Microbiol1994;48:427–47.
21] Yan Q, Cheung YK, Cheng SC, Xian-Hua W, Yong X. A DNA vaccine constructedwith human papillomavirus type 16 (HPV16) E7 and E6 genes induced specificimmune responses. Gynecol Oncol 2007;104(1):199–206.
22] Arbyn M, Dillner J. Review of current knowledge on HPV vaccination: anappendix to the European guidelines for quality assurance in cervical cancerscreening. J Clin Virol 2007:1–9.
23] Robert J. Evolution of heat shock protein and immunity. Develop CompImmunol 2003;27:449–64.
24] Zugel U, Kaufmann S. Role of heat shock proteins in protection from and patho-genesis of infectious disease. Clin Microbiol Rev 1999:19–39.
25] Binder R, Srivastava P. Essential role of CD91 in representation of gp96 chaper-oned peptides. PNAS 2004;101(16):6128–33.
26] Li Z, Menoret A, Srivastava P. Roles of heat shock proteins in antigen presenta-tion and cross presentation. Curr Opin Immunol 2002;14:45–51.
27] Demine R, Walden P. Testing the role of gp96 as peptide chaperone in antigenprocessing. J Biol Chem 2005;280(18):17573–8.
28] Chandawarkar R, Wagh M, Kovalchin J, Srivastava P. Immune modulation withhigh-dose heat shock protein gp96: therapy of murine autoimmune diabetesand encephalomyelitis. Int Immunol 2004;16(4):615–24.
29] Pockley AG. Heat shock proteins as regulators of the immune response. Lancet2003:1–8.

ccine
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
3370 A. Bolhassani et al. / Va
30] Nicchitta CV, Carrick DM, Baker-Lepain JC. The messenger and the message:gp96 (GRP94)-peptide interactions in cellular immunity. Cell Stress Chaperones2004;9(4):325–31.
31] Boussiff O, Lezoualch F, Zanta MA, Mergny MD, Scherman D, DemeneixB, et al. A versatile vector for gene and oligonucleotide transfer into cellsin culture and in vivo: polyethylenimine. Proc Natl Acad Sci 1995;92:7297–301.
32] Ferrari S, Moro E, Pettenazzo A, Behr JP, Zacchello F, Scarpa M. ExGen 500 isan efficient vector for gene delivery to lung epithelial cells in vitro and in vivo.Gene Ther 1997;4:1100–6.
33] Chen CH, Wang TL, Hung CF, Pardoll DM, Wu TC. Boosting with recombinantvaccinia increases HPV16 E7 specific T cell precursor frequencies of HPV16 E7expressing DNA vaccines. Vaccine 2000;18:2015–22.
34] Woodland D. Jump-starting the immune system: prime-boosting comes of age.Trends Immunol 2004;25(2):1–7.
35] Lin CT, Hung CF, Juang J, He L, Lin KY, Kim TW, et al. Boosting with recombinantvaccinia increases HPV16 E7 specific T cell precursor frequencies and antitu-mor effects of HPV16 E7 expressing sindbis virus replicon particles. Mol Ther
2003;8(4):559–66.36] Ramshaw I, Ramsay A. The prime-boost strategy: exciting prospects forimproved vaccination. Trends Immunol Today 2000;21(4):163–5.
37] Maniero GD, Robert J. Phylogenetic conservation of gp96-mediated antigen-specific cellular immunity: new evidence from adoptive cell transfer inXenopus. Transplantation 2004;78(10):1415–21.
38] Goyos A, Cohen N, Gantress J, Robert J. Anti-tumor MHC class Ia-unrestrictedCD8 T cell cytotoxicity elicited by the heat shock protein gp96. Eur J Immunol2004;34(9):2449–58.
39] Robert J, Menoret A, Cohen N. Cell surface expression of the endoplasmicreticular heat shock protein gp96 is phylogenetically conserved. J Immunol1999;163(8):4133–9.
40] Robert J, Cohen N, Maniero GD, Goyos A, Morales H, Gantress J. Evolution ofthe immunomodulatory role of the heat shock protein gp96. Cell Mol Biol2003;49(2):263–75.
[41] Chen CH, Wang TL, Hung CF, Yang Y, Young R, Pardoll D, et al. Enhancement ofDNA vaccine potency by linkage of antigen gene to an HSP70 gene. Cancer Res2000;60:1035–42.
42] Michel N, Osen W, Gissmann L, Schumacher TNM, Zentgraf H, Muller M.Enhanced immunogenicity of HPV16 E7 fusion proteins in DNA vaccination.Virology 2002;294:47–59.
43] Hauser H, Shen L, Gu QL, Krueger S, Chen SY. Secretory heat-shock protein asa dendritic cell-targeting molecule: a new strategy to enhance the potency ofgenetic vaccines. Gene Ther 2004;11(11):924–32.
44] Li H, Ou X, Xiong J. Modified HPV16 E7/HSP70 DNA vaccine with high safetyand enhanced cellular immunity represses murine lung metastatic tumorswith downregulated expression of MHC class I molecules. Gynecol Oncol2007;104(3):564–71.
[
[
[
[
[
[
[
[
26 (2008) 3362–3370
45] Kim TY, Myoung HJ, Kim JH, Moon IS, Kim TG, Ahn WS, et al. Both E7 and CpGoligodeoxynuclotide are required for protective immunity against challengewith human papillomavirus 16 (E6/E7) immortalized tumor cells: involvementof CD4+ and CD8+ T cells in protection. Cancer Res 2002;62:7234–40.
46] Rapp U, Kaufmann S. DNA vaccination with gp96-peptide fusion proteinsinduces protection against an intracellular bacterial pathogen. Int Immunol2004;16(4):597–605.
47] Zugel U, Sponaas AM, Neckermann J, Schoel B, Kaufmann SHE. Gp96-peptide vaccination of mice against Intracellular bacteria. Infect Immun2001;69(6):4164–7.
48] Lewis J. Therapeutic cancer vaccines: using unique antigens. PNAS2004;101(2):14653–6.
49] Hauser H, Chen SY. Augmentation of DNA vaccine potency through secretoryheat shock protein-mediated antigen targeting. Methods 2003;31:225–31.
50] Suzue K, Zhou X, Eisen HN, Young RA. Heat shock fusion proteins as vehicles forantigen delivery into the major histocompatibility complex class I presentationpathway. PNAS 1997;94:13146–51.
[51] Suzue K, Young RA. Adjuvant-free hsp70 fusion protein system elicits humoral
and cellular immune responses to HIV-1p24. J Immunol 1996;156:873–9.52] Huang CY, Chen CA, Lee CN, Chang MC, Cheng WF. DNA vaccine encoding heatshock protein 60 co-linked to HPV16 E6 and E7 tumor antigens generates morepotent immunotherapeutic effects than respective E6 or E7 tumor antigens.Gynecol Oncol 2007;107(3):404–12.
53] Kim D, Gambhira R, Karanam B, Monie A, Hung CF, Roden R, et al. Generationand characterization of a preventive and therapeutic HPV DNA vaccine. Vaccine2008;26(3):351–60.
54] Kuck D, Leder C, Kern A, Muller M, Piuko K, Gissmann L, et al. Efficiency ofHPV16L1/E7 DNA immunization: influence of cellular localization and capsidassembly. Vaccine 2006;24:2952–65.
55] Fan H, Kashi RS, Middaugh CR. Conformational lability of two molecular chap-erones Hsc70 and Gp96: effects of pH and temperature. Arch Biochem Biophys2006;447(1):34–45.
56] Doria-Rose NA, Haigwood NL. DNA vaccine strategies: candidates for immunemodulation and immunization regimens. Methods 2003;31:207–16.
57] Tindle RW, Croft S, Herd K, Malcolm K, Geczy AF, Stewart T, et al. A vaccineconjugate of ‘ISCAR’ immunocarrier and peptide epitopes of the E7 cervicalcancer-associated protein of human papillomavirus type 16 elicits specific Th1-and Th2-type responses in immunized mice in the absence of oil-based adju-vants. Clin Exp Immunol 1995;101(2):265–71.
58] Fernando G, Stewart T, Tindle R, Frazer I. Th2-type CD4+ cells neither enhancenor suppress antitumor CTL activity in a mouse tumor model. J Immunol1998;161:2421–7.
59] Chu NR, Wu HB, Wu TC, Boux LJ, Siegel MI, Mizzen LA. Immunotherapy of ahuman papillomavirus (HPV) type 16 E7-expressing tumour by administrationof fusion protein comprising Mycobacterium bovis bacille Calmette–Guerin(BCG) Hsp65 and HPV16 E7. Clin Exp Immunol 2000;121:216–25.
Copyright © 2022 FDOKUMEN




















![Poly[di(carboxylatophenoxy)phosphazene] is a potent adjuvant for intradermal immunization](https://static.fdokumen.com/doc/165x107/6335c6c4a1ced1126c0af097/polydicarboxylatophenoxyphosphazene-is-a-potent-adjuvant-for-intradermal-immunization.jpg)