
Einfluß anthropogener Störreize auf Verhalten ... - Wildlife
104
Aus dem Forschungsinstitut für Wildtierkunde und Okologie der Veterinärmedizinischen Universität Wien Leiter: o.Univ.Prof. Dr. K. Onderscheka A-1160 Wien, Savoyenstraße 1 Einfluß anthropogener Störreize auf Verhalten, Aktivitätsmuster und Herzfrequenz von Reh- und Rotwild I AUGURAL-DISSERTATION zur Erlangung des Grades eines DOCTOR IEDICI AE VETERINARIAE der Veterinärmedizinischen Universität Wien vorgelegt von Dipl.Tzt. Susanne Wagner Wien, im September 1992
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of Einfluß anthropogener Störreize auf Verhalten ... - Wildlife

Aus dem
Forschungsinstitut für Wildtierkunde und Okologie
der Veterinärmedizinischen Universität Wien
Leiter: o.Univ.Prof. Dr. K. Onderscheka
A-1160 Wien, Savoyenstraße 1
Einfluß anthropogener Störreize
auf Verhalten,
Aktivitätsmuster und Herzfrequenz
von Reh- und Rotwild
I AUGURAL-DISSERTATION
zur Erlangung des Grades eines
DOCTOR i\lIEDICI AE VETERINARIAE
der Veterinärmedizinischen Universität Wien
vorgelegt von
Dipl.Tzt. Susanne Wagner
Wien, im September 1992

Z9
TT
68
8Z
LZ
LZ
LZ
97,
TZ
6I
6I
LI
9I
gI
9T
pll.4alou s"P rnJ essruqä8rg
asfll?uPzuenbe.IlzreH t'1'8
' esdlt?ul]suallBrlrr^ t'I'8
ralsnurslsll^l?{Y z'I't
sezleldeSelT saP Iq?^ . I'1't
PII.r\qeu seP rru essruqe8rg I'tassruqa8rg I
' esfl"u"suellEqre^ t'v'z
uaBunuqf,IalzyßItezztny Jap Eunparrrsny T,'?'7,
uaBunuqctazlneltaz8uel rep Eunparv'sny TT'7,
ualeO rap 8un1ra.\\sn\-: puu BunssuJJeuJleO alfl T'Z
' ellf,t1s.Ir,\rols eICI t'z
' ulneJllezs8unqrrs.ralui) puneJatlsqtnsre1 Z'Z
apuela8sqrnsra1 srzC TZ
uapoqlal I pun IBITaIUIAI 7,
7,'t
T,I snutq].it1.IslBll^I1{V t'I
0I ' Sunratstllqrsurs Pun uol]tsn1|clu11 Z'I
8 ' [lr.I]S uo.\ 8unssatrl ?'I'I
q ' raII rulaq Bunqtnsralungarls rap Eunlnaprg alg 8'1'I
1 . . iser?s lsr sr?^\ z.I.I
t ' ' 18unr91§ eura lsl sI?^A I'I'I
I ' uaratlPlllA Ieq Sunqrsro;gerls rnz saureuraSlly I'i
t Eunyalurg I
sruqclazro^s+IBt{uI

INHALTSVERZEICHNIS
3.2.7 Wahl des Liegeplatzes
3.2.2 Aktivitätsmuster
3.2.3 Verhaltensanalyse
3.2.4 Herzfrequenzanalyse
4 Diskussion
5 Zusammenfassung
6 Literaturverzeichnis
7 Anhang
77
86
89
100
52
53
63
69

1 Einleitung
Mit der Zunahme der Freizeitaktivitäten des Menschen in den ohnehin schorr durch
Siedlungen und Verkehrswege zerschnittenen und flächenmäßig verringerten Lebensräu-
men für die freilebenden Tiere kommt es bei diesen Tieren immer häufiger zu Störungen
der normalen Lebensabläufe. Sowohl den Naturschützer als auch den Jäger beschäftigt
vor allem die Frage, ob und in welchem Ausmaß das Wild durch Erholungsuchende
oder andere Waldnutzer (2.8. forstwirtschaftliches Personal, Jäger) beunruhigt wirdund welche Auswirkungen das auf das Wohlbefinden und unter Umständen sogar auf
die Gesundheit des Wildes hat. Vor allem Naturschutzgebiete haben die Aufgabe,
einerseits als Schongebiete für die heimische Tier- und Pflanzenwelt nt fungieren, an-
dererseits sollen sie aber auch den Menschen einen Kontakt zur Natur ermöglichen.
Beides zu vereinbaren ist eine der schwierigen Aufgaben der Ökologie und Wildbiolo-
gie (BuncH 1988, Bovr,p u. SRIvrsoN 1985). Ziel dieser Arbeit ist, die Auswirkung
verschiedener von Menschen verursachter Störreize auf das Verhalten und die Herzfre-
quenz bei Rehwild (Capreolus capreolzs) und Rotwild (Ceruus elaphus) zu erfassen.
Außerdem wird geprüft, inwieweit sich die untersuchten Tiere an mehr oder weniger
häufig auftretende Störreize gewöhnen können.
1.1 Allgemeines zur Streßforschung bei Wildtieren
1.1.1 Was ist eine Störung?
In der Literatur wird der Begriff ,,Störung" verschieden interpretiert, entweder als
Ursache eines Geschehens, als deren Folge oder beides gemeinsam (Rvxrnl u. Eowano1985). In der vorliegenden Arbeit wird der Begriff,,Störung" als die Reaktion auf einen
Störfaktor bzw. Störreiz definiert.
Eine Störung kann nach RBrcHHoLF u. ScneltEt (1988) durch ihre Intensitrit, d.h.
die Wirkung des Störreizes auf das Tier, ihre Dauer und die Hriufigkeit beschrieben
werden. RvxIor, u. EowanD (1985) charakterisieren die Störung nach der Richtung
ihrer Wirkung, dem Umfang und der Dauer der Störung. Sie kann vorübergehend (2.B.
eine kurzfristig erhöhte Herzfrequenz) oder auch dauerhaft sein (2.8. Veränderung imAktivitätsrhythmus eines Tieres). CHnTsIENSEN (1988) beschreibt fiinf Parameter fürStörreizei die Hriufigkeit, d.h. wie oft der Störreiz in einem Gebiet auftritt, die Wie-
derkehrzeil an einer bestimmten Stelle, die Intensifal unabhängig von seiner Wirkung,

1 EINLEITUNG
die Heftigkeit, d.h. die Wirkung auf den Organismus (dies ist jedoch ein Parameter
der Störung und nicht des Störreizes) und die Vorhersehbarkeit (predictability).
ZuuNrBR (1991) betrachtet den Begriff,,Störung" als wertneutral: Das Tier vergleicht
seine momentane Situation (,,Ist-Wert") mit dem ihm angeborenen und auch durch
Erfahrungen modulierten ,,Soll-Wert". Weichen beide voneinander ab, versucht das
Tier durch eine verhaltensmäßige oder eine physiologische Anpassung den angestrebten
Sol1-Wert wieder zu erreichen. Erst bei Vorliegen einer meßbaren Reaktion kann von
eiler Störung gesprochen werden" Die Störung muß jedoch nicht unbedingt negative
Auswirkungen haben. Erst eine über längere Zeit anhaltende Störung wird für das
Tier pathogen.
Honnrst (1976) unterscheidet zwischen aktiver und passiver Störung des Tieres. Er-
steres wirkt sich in offensichtlichen Verhaltensreaktionen, wie Sicherungsverhalten und
Flucht aus. Dagegeri läßt eine passive Störung keine auf den ersten Blick augenfälli-
gen Reaktionen des Tieres erkennen, sondern nur unscheinbare Veränderungen seines
Aktivitätsmusters.
L.1.2 Was ist Streß?
Allgernein wird Streß als ,,starke körperliche oder seelische Anspannung, Belastung" be-
zeichlet, im englischen Sprachraum auch als die ,,Reaktion des Körpers auf von aul3en
einwirkende Belastungen". Splyp (1936, 1952) versteht unter Streß eine generalisierte,
unspezifische Reaktion des Körpers auf schädliche Reize (sogenannte Stressoren). Diese
Stressoren können gänziich verschiedener Natur sein, z.B. Kälte, Verletzungen, Hunger
oder Toxine. Bei trinwirkung derartiger Stressoren kommt es zu einem ,,allgemeinen
Adaptionssyndrom", das sich in drei Phasen gliedern läßt, die aber nur bei längeran-
haltenden Streßsituationen a1le durchlaufen werden:
1. Alarmreaktion: Freisetzung von ACTH aus der Hypophyse" Die Nebennieren-
rinde wird vermehrt zur Ausschüttung von Gluko- und Mineralocorticoiden ak-
tiviert.
2. Widerstandphase: Der Körper hat sich der erhöhten Streßbelastung weitgehenci
angepaßt.
3. Erschöpfungsphase: Die Körperreserven sind erschöpft. Die Symptome wie bei
der Alarmreaktion kehren zurück, es entstehen jedoch diesrnal irreversible Schäden
(Nierenschäden, Polyarthritis, Myokarditis,. . . ), die bis zum Tod führen.
4

1.1 Allgemeines zur Streßforschung bei Wildtieren
Inzwischen hat sich jedoch herausgestellt, daß die Streßreaktion nicht immer unspezi-
fisch abläuft, sondern daß verschiedene Stressoren sich beim ,ib"r, Individuum unter-
schiedlich auswirken können (Monrnc 1985a).
JnNxrns u. KnucnR (1973) unterscheiden zwischen psychischen Stressoren (2.8. angst-
auslösende Momente) url.d somatischen Stressoren (2.8. Verletzungen, Umgebungstem-
peratur). PoncBs (1985) weist darauf hin, daß die Streßreaktion abhängig von der Re-
aktionslage des Tieres ist, wenn es dem Reiz ausgesetzt wird. Verschiedene Individuen
können auf den gleichen Störreiz unterschiedlich reagieren und sogar dasselbe Indivi-
duum kann zu verschiedenen Zeitpunkten ein anderes Verhaiten zeigen. Die Intensität
der Streßreaktion auf einen bestimmten Auslöser wird durch viele Faktoren modifiziert,
z.B. Alter, Geschlecht, Kondition, Temperament des Tieres, vorhergehende Erfahrun-
gen oder gleichzeitig herrschende Umwelteinflüsse (Juxxrms u. KnucpR 1973).
Caumox (1929) hat als erster die Bedeutung des vegetativen Nervensystems im Streß-
geschehen erkannt. In dem von ihm bezeichneten ,,Emergency state" kommt es durch
Einwirkung von Schmerz oder emotionale Erregung zur Ausschüttung von Adrena-
lin, Herzfrequenzerhöhung und Blutdrucksteigerung. Der Organismus wird dadurch zu
Höchstleistungen befähigt.
Zwätzlich zu dieser kurzfristigen Wirkung des autonomen Nervensystems und der zu-
erst erläuterten neuroendokrinen Reaktion spielt das Verhalten des Tieres eine wichtige
Rolle im Streßgeschehen. Verhaltensreaktionen sind beim Tier oft die einfachste und
ökonomischste Art, auf einen Stressor zu reagieren, z.B. entzieht sich ein Tier zu großer
Hitze, indem es Schatten aufsucht (Monunc 1985a).
1.1.3 Die Bedeutung der Streßuntersuchung beim Tier
Im allgemeinen ist die Streßreaktion als eine Anpassung des Körpers an geänderte Um-
weltbedingungen zu interpretieren. Mitunter kann sich aber eine reizarme Umgebung
auch negativ auf den Organismus auswirken (Fn.o.snn et al. 1975). Generell wirkt sich
jedoch, wie im vorigen Abschnitt schon erwähnt, ein Übermaß an Stimulierung nach-
teilig auf das Tier aus, ja es kann sogar lebensgefährdend sein (Gursr 1971). Tiere,
die einem Stressor ausgesetzt sind, zeigten eine erhöhte Resistenz gegenüber weiteren
schädlichen Noxen (Suvn 1952),jedoch kommt es bei Tieren, die sich an einen Stres-
sor stark adaptierten) zt7 einer Sensibilisierung gegenüber anderen Noxen. Dies gilt
v.a., wenn mehrere verschiedene Streßfaktoren zur selben Zeit avf das Tier einwirken,
z.B. kalte Witterung trei gleichzeitiger Beunruhigung des Tieres.

1 EINLEITUNG
Durch die vermehrte Adrenalinausschüttung wird der gesamte Grundumsatz des Kör-
pers gesteigert, was zu einem erhöhten Energieverbrauch des Tieres führt. Außerdem
wird bei häufigen Störungen des Tieres dieZeit, die es zur Nahrungsaufnahme aufwen-
det, herabgesetzt, wie es z.B. bei Gemsen (Rupri,capra rupricapra) festgestellt wurde
(Cnounua u. Lovr.nr 1985).
Mitunter wandern die Tiere in Gebiete mit einem ungünstiger Arrngrr,ngebot ab, z.B.
Gemsen von offenen Grasflächen in den Wald (Rlurn-GRoss et al. 1988). Vor allem
in der Wintersaison kann dies zu einer negativen Energiebiianz führen, d.h. das Tier
verbraucht mehr Energie pro Zeiteinheit, als es Energie aufnimmt (Ou»rnscHEKA
1935). Dadurch wird das Überleben im Winter in Frage gestellt (Grrsr 1971). Ebenso
ist der Verdauungprozeß der Wiederkäuer bei häufigen Beunruhigungen gestört, da
ausreichende Ruhephasen {ür die Tätigkeit des Wiederkäuens nötig sind. Bei Beun-
ruhigung des Tieres wird durch den überwiegenden Sympathikotonus die Produktion
der Verdauungssäfte vermindert. Durch die fehlende Kautätigkeit wird auch weniger
Speichel produziert, der jedoch durch seinen alkalischen pH-Wert einer Pansenüber-
säuerung entgegenwirkt. Durch solche Verdauungsstörungen wird die Kondition des
Tieres herabgesetzt (B,Lnru 1984).
Nicht nur das Wild leidet an übermäßigen Störungen, auch der Wald wird indirekt
durch das verstärkte Schälen der Baumrinden und durch den Verbiß von Jungtrieben
geschädigt " Zwei Ursachen sind in diesem Zusammenhang von besonderer Bedeutung:
Einerseits sucht das Wild vermehrt die Deckung des Waldes auf (Sichtschutz), an-
dererseits findet es hier nicht die entsprechende Asung. Das Schälen beugt zudem
einer Pansenazidose vor, weil hierbei die entsprechende Speichelmenge produziert wird
(ON»onscHEKA 1979). Rotwild überbrückt die Wartezeit bis zur Abenddämmerung,
in der es wieder ungestört äsen kann, mit der Aufnahme von Fichtenrinde (Puurrnnu. HRnTTIEL 1984). Im Winter wirken sich durch die mangelnde Deckung und den
Nahrungsengpaß Störreize verstärkt auf Schälschäden aus (Rrurnosnn et al. 1987).
Weitere negative Auswirkungen für das Tier sind verminderte Fruchtbarkeit und Abor-
tus. Seurss et al. (1986) steliten bei Wapitis (Ceraus elaphus) einen abrupten Ab-
fall des Befruchtungserfolges mit dem Beginn der Jagdsaison fest. Sie vermuten als
Ursache eine Unterbrechung des Brunftzyklus oder frühen embryonalen Tod. Nach
Moennc (1985b) kommt eine streßbedingte Fruchtresorption vor der Nidation des
befruchteten Eies oft vor, nachher jedoch nur noch selten. Bei hochträchtigen Rentie-
ren (Rangi,fer taranilus tarandus), die im tiefen, verharschten Schnee flüchten müssen,
6

1.1 Allgemeines zur Streßforcchung bei Wildtieren
kann durch Anschlagen des Bauches an der Schneedecke ein Abortus ausgelöst werden
(Grtst 1971). Ein Ausweichen in ungünstige Abkalbegebiete hat für Karibus (Rangifer
tarandus caribou) eine erhöhte Jungtiersterblichkeit nn Folge (Wutrrox et al. 1992).
Werden sie während der Laktation ntsätzlich durch Insekten belästigt, können Kari-
bus durch erhöhte Bewegungsaktivität sogar in eine negative Energiebilanz gelangen,
wodurch das Wachstum der Jungtiere beeinträchtigt wird (Walsu et al. 1992).
Abwanderungen des Wildes in andere Gebiete infolge menschlicher Störeinflüsse können
vorkommen, z.B.ließen sich Wapitis durch Brennholzsammler eher aus ihrem Streif-
gebiet vertreiben als die weniger empfindlich reagierenden Maultierhirsche (Odocoilus
hemionus) (W,t no 1984). Ebenso mieden Wapitis eine neu ausgebaute Straße (Czacu
1991). Auch Weißwedelhirsche (Odocoileus uirginianus) ließen sich durch starke Jagd-
aktivität aus ihrem Einstandsgebiet vertreiben (Sr,a.nnowr u. SpntxcEn 1970) und
traditionelle Wintereinstände des Rotwildes blieben nach hohem Jagddruck ungenutzt
(Scuirnmr 1992).
Im allgemeinen sind Cerviden aber recht ortstreu und kehren nach Beendigung der
Störeinflüsse in ihr ursprüngliches Streifgebiet (home range) zurück. Nach Swnprqpv
et al" (1971) kehrten von Hunden gehetzte Weißwedelhirsche spätestens nach einem
Tag in ihr Stammgebiet zurück. Während mäinnliche Weißwedelhirsche nach häufigen
Störungen durch Viehtrieb ihr Streifgebiet verlagerten, blieben weibliche Tiere relativ
ortstreu (Hooo u. INcLIS 1974). Selbst durch mehrfach gezielt ausgelöste Störungen
ließen sich Rothirsche nicht aus ihrem Streifgebiet vertreiben (GooncII 1980). Rehe,
die durch Orientierungsläufer und Treibjagden aus ihrem Tagesaktivitätsraum vertrie-
ben wurden, kehrten bald wieder zurück (JaeensnN 1984). Nach einem großflächi-
gen Waldbrand kehrte das Rotwild trotz völlig veränderter Landschaft und fehlender
Aruog in seinen Lebensraum zurück (Ransnor,D u. REULECKE 1988). Auch Karibus,
die häufig von Hubschraubern verfolgt und belästigt wurden, verließen das Gebiet nicht
(BnncnnuD et al. 1984) und zeigten auch keine Einbußen in ihrer Reproduktionslei-
stung.
L.1.4 Messung von Streß
trs gibt zahlreiche verschiedene physiologische Parameter, die als Indikator für Streß
und Erregtheit in Frage kommen (Lovtxu 1985, Prarr 1982, Snaxony 1989). Viele
von ihnen sind allerdings für die Untersuchung gezielter Störungsversuche am freiie-
benden Tier ungeeignet. Technische Schwierigkeiten ergeben sich bei der Messung von

1 EINLEITUNG
Blutdruck, Hauttemperatur, elektrodermaler Aktivität, Muskelspannung, EEG und
O2-Verbrauch. Die Konzentration von Katecholamin, freien Fettsäuren und Cortico-
steroiden im Blut ändert sich bei Einwirkung von Streßfaktoren, scheiden jedoch als
spezifische Meßindikatoren aus, weil das Tier durch die Beunruhigung beim Fangen
und bei der Blutabnahme stark belastet wird, wodurch sich seine Blutwerte verändern.
Außerdem ist die Interpretation der Corticosteroidkonzentration im Blutplasma äußerst
schwierig, da sie, je nachdem, ob ein akutes oder ein chronisches Geschehen vorliegt,
erhöht oder erniedrigt sein kann (SrorT 1981, RUsHBN 1991). Das Isoenzym LDH-5
erwies sich beim Damwild (Dama dama) als ein gutgeeigneter Meßparameter für Streß
(JoNrs u. Pntco 1990), ist jedoch auch mit der obenerwähnten Problematik der Blut-
abnahme verbunden.
Ein weiteres Merkmal im Zusammenhang mit der Streßbelastung ist die Vergröße-
rung der Nebennieren. WpnmER (1976) konnte z.B. bei einem während Störversuchen
verendeten Rehbock als einzige pathologische Veränderung erheblich vergrößerte Ne-
bennieren feststellen. Jedoch können mit der Messung der Nebennierengewichte nicht
die unterschiedliche Wirkung verschiedener Störreize auf das gleiche Tier und die Aus-
wirkung kurzdauernder Belastungen untersucht werden. Außerdem ist die individuelle
Variabilität der Nebennierengewichte beim Reh sehr groß (GnoRGII 1976).
Die Herzfrequenz ist dagegen ein sehr gut geeigneter Parameter für diese Untersuchung.
Durch eine telemetrische Erfassung der Meßwerte wird das Verhalten des Tieres nicht
zusätzlich beeinflußt. Vor allem können arch kuzzeitige Reaktionen miterfaßt wer-
den. Bedeutend ist auch die Korrelation von Herzfrequenz und Energieverbrauch des
Tieres, wie es etwa beim Rotschenkel (Tringa totanus) (Frnus et al. 1979), beim
Karibu (Fancv u. Wnrrn 1985) und beim Weißwedelhirsch (Holron et al. 1976,
Maurz u. FatR 1980) nachgewiesen wurde. Nach Fnptty (1979) gibt die Herzfre-
quenz auch Auskunft über psychische Belastungen beim Tier ohne äußere erkennbare
Verhaltensänderungen. FpRNs et al. (1979) stellten eine Verdopplung der Herzfrequenz
beim schlafenden Rotschenkel fest, sobald eine Person das Labor betrat, ohne daß der
Vogel seine Körperhaitung änderte. Zu ähnlichen Ergebnissen kamen auch B.q.r,r, u.
Avrramnn (1979) mit Siibermöwen (Larus argentatus). JlconsEN (1979b) maß eine
Erhöhung der Herzfrequenz bei liegenden Weißwedelhirschkälbern um 50%, wenn sie
beunruhigt waren. RosncssvsKll et al. (1975) fanden einen Anstieg der Herzfreqlterrz
um das 2.4fache beim El"ch (Alces alces) und um das 2.7fache beim Rentier, wenn sie
erregt waren, rind FnnoDY (1979) konnte bei stehenden Maultierhirschen eine Herzfre-
quenzerhöhung messen, wenn Menschen vorbeigingen.
8

1.1 Allgemeines zur Streßforschung bei Wildtieren
Eine Erregtheit des Tieres muß nicht unbedingt eine Erhöhung der Herzfrequeuz zlu
Folge haben, unter Umständen kann es auch zu einer Absenkung der Herzfrequenz
kommen, der sogenannten ,,Alarmbradykardie". Sowohl Rotwildkälber (Esruanxu. LaNcvlrN 1979), Kälber von Weißwedelhirschen (MonN et al. 1977, JtcoBSEN
1979a) als auch Florida-Waldkaninchen (Sylai,lagus fl,oridanus) (Srvuur u. Wonrn1979) wiesen eine Alarmbradykardie auf. Es handelt sich in diesen Fällen um Tiere,
die bei Gefahr das Verhalten des ,,Sich-Drückens" zeigen. Dabei liegen sie mit erhöhter
Aufmerksamkeit flach und bewegungslos, um möglichst unerkannt zrbleiben (,,freezing
and hiding" im Ggs. zu ,,fight and flight"). Srönn (1982) fand bei Tupajas (Tupaia
belangeri,) Individuen, die gegenüber ein und demselben Störreiz rnit Tachykardie rea-
gierten und andere, die eine ausgeprägte Bradykardie zeigten. Es kann sogar ein Tier,
das normalerweise eine tachykarde Reaktion durch Stimuli zeigt, bei besonders starken
Reizen mit Bradykardie und ,,Sich-Totste1len" reagieren. Mrr,r,pn (1982) interpretiert
diese beiden gegensätzlichen Reaktionen auf Störreize als zwei verschiedene angeborene
Verhaltensmuster von Angst.
Kapucon (1990) hat bei Wöifen (Canis lupus) die Herzfrequenz mittels Telemetrie ge-
messen. Bei Annäherung von Menschen stieg zuerst die Herzfrequenz von durchschnitt-
lich 131 auf durchschnittlich 184 Schlägef rrlir-, sank aber bei weiterer Annäherung unter
den ursprünglichen Wert bis auf 109 Schläge/min. Es handelt sich zwar beim Wolf nicht
um eine ,,sich drückende" Tierart, doch andere Autoren konnten ebenfalls bei Tieren,
die erhöht aufmerksam warer, eine Herzfrequenzabsenkung feststellen,, wie z.B. SrBIrnp
(1983) bei Dickhornschafen (Oui,s canadensis canadensis). Reaktionszeitversuche beim
Menschen bekräftigen die Annahme, daß es bei verstärkter Aufmerksamkeit und Erwar-
tungshaltung durch vagale Erregung zu einer Verlangsamung der Herzschlagfrequenz
kommt (Zrrr.tuon et al. 1989).
Neben diesen bisher erwähnten Beispielen von kurzfristigen (,,phasischen") Herzfre-
quenzänderungen durch Störreize gibt es aber auch längerfristige (,,tonische") Ande-
rungen der Herzfrequenz nach Störurlgen, wie Srönn (1982) es z.B. bei Tupajas nach-
weisen konnte.
Neben der Analyse der Herzschlagfrequenz eignet sich auch die Verhaltensbeobachtung
für die Untersuchung der Streßbelastung freilebender Tiere. Sowohl kurzfristige Reak-
tionen auf Störreize als auch Veränderungen des gesamten Aktivitätsrhythmus geben
Aufschluß über die Beunruhigung von Wildtieren. Insbesondere, weil die Herzfre-
quenz nicht nur psychische Zustände des Tieres widerspiegelt, sondern sehr stark von

10 1 EINLEITUNG
der Aktivität des Tieres beeinflußt wird und daher nicht als alleiniges Maß für Streß
ausreichend ist (SroMP 1983), ist es von Bedeutung, die jeweilige Verhaltenszusam-
mensetzung und die allgemeine Aktivität des Tieres vor und während der Störversuche
mit in die Untersuchung einzubeziehen.
Um Aussagen über das Wohlbefinden eines Tieres machen zu können, müssen zahlrei-
che Parameter aus den verschiedensten biologischen Funktionskreisen miteinbezogen
werden, wie Gesundheit, Fruchtbarkeit, physiologische Meßwerte und das Verhalten
(BlacxsnAw 1986). Die Herzfrequenz ist nur ein Teilaspekt, gibt jedoch zusammen
mit der Verhaltensbeobachtung einen guten Hinweis auf das Vorliegen einer Beunruhi-
gung beim freilebenden Wildtier.
L.2 Habituation und Sensibilisierung
Bei wiederholtem Setzen eines identischen Reizes nimmt die Reaktion eines Individu-
ums auf diesen Reiz ab, es kommt nn Habituation (IUuELMANN 1982). Im Gegensatz
zur Adaptation, die durch eine Ermüdung des peripheren Nervensystems zustande
kommt, ist die Habituation vom ZNS ausgehend. Durch einen Fremdreiz kann der
Habituationsverlauf unterbrochen werden, die Reaktion auf den ursprünglichen Reiz
nimmt dann wiedet ^t. Man spricht in diesem Fall von Dishabituatioz (Sctt,tNDRY
1989). Jedoch ist die Habituation nicht immer der gewöhnliche Ablauf, manchmal
kommt es bei einer wiederholten Reizdarbietung anfangs zu einer verstärkten Reaktion
(besonders wenn der Reiz einen aversiven Charakter hat), das Individuum wird sensibi-
lisiert" Es können auch physiologische Reaktionen des Körpers langsamer habituieren
als die beobachteten Verhaltensreaktionen (LnvtNE 1985, Sruup 1983).
Habituation und Dishabituation in Bezug auf Alarmbradykardie gegenüber einem sich
nähernden Menschen, Steinwürfen und Hundegebell konnten Espuenx u. Lamcvann
(1985) in ihren Versuchen mit Rotwildkälbern feststellen. GAtssA.uBR (1988) stellte
bei wiederholter Annäherung von Menschen eine Abnahme der Fluchtreaktion von
Rehen fest. Srour u. ScHwaB (1979) zeigten in ihren Versuchen, daß sich Dela-
warenmöwen (Larus ilelawarensfs) durch Abnahme der Herzfreqretz rasch an vom
Kassettenrekorder abgespielte Alarmrufe habituieren. Ar,snscHr (1988) unterschei-
det zwischen unspezifischer und personenspezifischer Gewöhnung. Er konnte sich nach
einer Gewöhnüngsdauer von drei Monaten einem Gamsrudel durchschnittlich arf 27
Meter nähern, während die mittlere Fluchtdistanz gegenüber anderen Testpersonen

1.2 Habituation und Sensibilisierung
noch 46 Meter betrug. Ar,ruawx (1958) beobachtete bei Elchen eine geringere Flucht-
distanz gegenüber in der Gegend häufig auftretenden Anglern als gegenüber anderen
Personen. Auch Bürzr,nn (1986) beschreibt eine selektive Vertrautheit von Rotwild
gegenüber bestimmten Personen, z.B. Waldarbeitern und Personen, die die Fütterung
betreuen.
Jedoch auch die räumliche Anordnung von Versuchen spielt eine entscheidende Rolle
bei der Habituation von Tieren gegenüber Stimuli. Scnr,rror u. Su.q.r,trn (1983)
ließen Attrappen an einem Segelflosser (Pterophyllum eimi,cki) vorbeiziehen, wobei die
zuerst starke Fiuchtreaktion des Fisches nach baldiger Habituation nachließ. Wurde
jedoch dieselbe Attrappe von einer anderen Seite vorbeigezogen, war wieder die ur-
sprüngliche Fluchtreaktion vorhanden. In Versuchen mit Hühnern (Gallus spec.) kam
SHar,tpn (1984) zu sehr ähnlichen Ergebnissen. Er zog daraus den Schluß, daß sich
Tiere in natürlicher Umgebung, im Gegensatz zu den künstlichen Versuchsanordnun-
gen, nicht an Feinde gewöhnen, weil diese sich den Beutetieren stets von verschiedenen
ungewohnten Richtungen nähern.
MIllnn u. GuNN (1934) berichten, daß einzelne Moschusochsen (Oaibos moschatus)
gegenüber wiederholten Hubschrauberüberflügen Habituation zeigten, während andere
Individuen sensibilisiert wurden, und folgern daraus, daß die Toleranzgrenze gegenüber
Störreizen individuell verschieden hoch sein kann. Keine Habituation konnten MoBu
et aI. (1982) bei Weißwedelhirschen gegenüber Motorschlitten erkennen, und MecAn-THUR et al. (1982) bemerkten bei Dickhornschafen sogar eine erhöhte Empfindlich-
keit der Herzfrequenzreaktion bei wiederholter Annäherung durch Menschen" Dies
stimmt mit den Beobachtungen von Fnpnoy et al. (1986) überein, als sie bei Maul-
tierhirschen eine von der ersten bis zur dritten Annäherung fortschreitend verstärkte
Herzfrequenzreaktion messen konnten. Entgegen seinen Erwartungen hat auch StpIr,lp
(1983) bei Dickhornschafen eine Sensibilisierung der Herzfrequenz gegenüber wieder-
holten menschlichen Annäherungen feststellen können, obwohl sie sich im Verhalten
offensichtlich angepaßt hatten. Er schließt daraus, daß es zu nachhaltigen Herzfre-
quenzreaktionen bei häufigen Störungen kommen kann.
Zusammenfassend kann gesagt werden, daß Tiere sich gegenüber regelmäßig wied.erkeh-
renden Störeinflüssen unterschiedlich verhalten. Meist kommt es zu einer allmählichen
Gewöhnung des Tieres an den Störreiz, es kann aber auch eine erhöhte Empfindlichkeit
gegenüber dem Störfaktor erfolgen.
11

12 1 EINLEITUNG
1.3 Aktivitätsrhythmus
Der Aktivitätsrhythmus von Tieren wird von mehreren Faktoren beeinflußt. Einerseits
spielen endogene Faktoren, wie die momentane Stoffwechsellage des Tieres und erbli-
che Verhaltensprogramme, eine Rolle, andererseits exogene Faktoren, wie Witterung,
Tageslänge oder auch anthropogene Störeinflüsse. Stlvrn et al. (1969) untersuchten
den Energiegrundumsatz bei hungernden Weißwedelhirschen in Respirationskammern.
Trotz gleichbleibender Temperatur in der Kammer war im Sommerfell (Mai bis August)
der Grundumsatz weit höher als im Winterfell. Diese Anderung des Energiehaushaltes
nach dem Fellwechsel führen sie auf einen durch die sich ändernde Tageslänge aus-
gelösten Wechsel in der endokrinen Aktivität zurück.
J ahre s z eitli che Aktiaitätsu er än d erun g en
Die Bewegungsaktivität des Tieres wird von Jahreszeit und Tageszeit stark beeinflußt.
In den meisten Untersuchungen erwiesen sich Cerviden im Winter weit weniger aktiv als
im Sommer (Groncrr u. Scunöonn 1978, Lras l-981, Gnnnu u. Bpan 1990)" hrdem sind bei Rehen im Winter meist die Aktiv- bzw. Passivschübe verlängert gegenüber
denjenigen im Sommer, wodurch ein häufigerer Aktivitätswechsel in der warmen Jah-
reszeit erfolgt (CroonluND 1981, JpppBspN 1989). Nach Coopnr,uxo (1931) ist
dies durch das Asungsangebot bedingt. Im Sommer ist reichlich Asung vorhanden,
der Pansen ist schnell angefüllt und das Tier legt sich bald wieder zum Wiederkäuen
nieder. Im Winter ist der Zeitarfwand zur Nahrungsaufnahme dagegen viel höher, was
sich in einem längeren Aktivitätsschub äußert (,,rumen fill theorie"). Zrdernwird imSommer die energiereiche, leichtverdauliche Nahrung schnell umgesetzt, der Pansen ist
bald leer und das Tier geht wieder zum Ar"n über. Im Winter wiederum ist, bedingt
durch die rohfaserreiche Nahrung, die für das Wiederkäuen aufgewendete Zeit höher.
Dies drückt sich in einem längeren Passivschub aus.
Entgegen den obigen Ergebnissen stehen Untersuchungen bei Rehen unter konstanten
Haltungsbedingungen und konstanter Futterzusammensetzung. Auch hier kommt es zu
einer Verringerung der Nahrungsaufnahme und der Aktivität im Winter (ELlnwnnnc1978, Ozoca u. Vpnur 1970), was als Hinweis auf eine genetische Fixierung dieses
saisonalen Verhaltens gelten kann.
Von GBoncII (1981) untersuchte weibliche Rothirsche zeigten, anders als beim Reh, imWinter kürzere, aber häufigere Aktivschübe als im Sommer. Die von GnooN u. BBan
(1990) untersuchten Wapitis waren im Winter eher nachtaktiv als im Sommer, während
Lran (1981) bei der gleichen Tierart im Winter stärkere Tagesaktivität feststellte.

1.3 Aktivitätsrhythmus
C i,rc a di an er Aktiui,t ätsrhythmus
Weit verbreitet bei sehr vielen Tieren ist ein zweigipfeliger Tagesrhythmus, d.h. ver-
stärkte Aktivität während der Morgen- und Abenddämmerung und eine weitgehend
inaktive Phase am frühen Nachmittag (Ascnorp 1966). Dieser Rhythmus ist einer-
seits durch eine endogene, selbsterhaltende Oszillation bedingt, wie AscnoFF (1966)
es bei Grünfinken (Chloris chloris) nachweisen konnte, andererseits durch sogenannte
"Zeitgeber* (Licht-Dunkel) modifiziert. Nach Ozoca u. Vpalts (1970) können Um-
weltbedingungen Teile des Aktivitätsmusters unterdrücken oder betonen, sie können
aber keinen Grundrhythmus erzeugen. Fast alle untersuchten Cerviden zeigten dieses
zweigipfelige Muster (Bü:rzlun L974, Gnonctt u. ScuaöpER 1978, Gpoacrr 1981
beim Rotwild; Collrivs et al. 1978, GnoEN 11. BBan 1990, LrBu 1981, Lroa u. LBs
Mlncurt 1979 bei Wapitis; K.q.uunnuEyER u. MlncuINToN 1977 bei Weißwedel-
hirschen; Cpoenr,uNo 1981, JeernsnN 1989, Mpnrpus 1984 beim Rehwild). Nach
Jppppspn (1989) ist dieser zweigipfelige Rhythmus beim Reh im Winter ausgeprägter,
das gieiche gilt für Wapitis (GnunN u. Bnen 1990) und Rothirsche (Gnonctt 1934).
TunmBn (1980) führt diese verstärkte Dämmerungsaktivität auf eine erhöhte Synchro-
nisation des Verhaltens der Tiere untereinander zurück, da hier der Lichtwechsel als
,,Zeitgeber" wirkt. Durch die individuell variable Verdauungszeit wird tagsüber das
Verhalten mehr oder weniger desynchronisiert.
Potentielle Beutetiere können sich vor Feinden schützen, indem sie stets wachsam (vigi-
lant) sind. Die aufgewendete Zeit fiu aktive Vigilanz (Sicherungsverhalten) geht jedoch
auf Kosten anderer Verhaltenselemente, die für die Erhaltung des Tieres wichtig sind,
wie z.B. das Asen. Eine Möglichkeit, die Sicherzeit für das einzelne Tier zu red,uzieren,
ohne die individuelle Sicherheit zu vermindern, ist das Zusammenschließen mehrerer
Tiere zu einer Gruppe, wie das z.B. beim Feldreh gegenüber dem Waldreh geschieht.
Im Winter steigt die Anzahl der Rehe im Rudel (,,Sprung") drastisch an, und zwar
im offenen Feld viel ausgeprägter ais im Waid (Rucnnor,F 1980). Laut Mür,r,Bn-
IJsrncpn u. Scnr,oBTH (1967) können in ebenen deckungslosen Kulturflächen Rehru-
del mit bis zu 100 Stück entstehen. Ahn["h"r berichten auch SrusBE u. Passanco
(1979), v.a. in Gegenden, wo viele Beunruhigungen vorkommen. Nach Pror,owsxr
(1983) ist das Feidreh mit der Neigung zur großen Sprungbildung ein schon genetisch
fixierter Ökotyp. TunNpn (1983, zit. nach Kunr 1991) verglich die Zeitbudgetierung
von einzelnen Feldrehen mit Rehen in Gruppen. Das einzelne Tier wendet im Schnitt
26 Minut'en pro Stunde für Sichern auf, das in größeren Rudeln lebende Tier sichert nur
rund 12 Minuten. Auch UNoonwooD (1982) beobachtete bei verschiedenen Antilopen-
13

74 1 EINLEITUNG
arten eine Abnahme der Sicherungszeit für das einzelne Individuum mit zunehmender
Gruppengröße. BürtNER (1980) konnte beim Rehwild mit zunehmender Sprunggröße
eine Abnahme der Sicherungszeit verbunden mit einer Zunahme des Zeitaufwandes fürdas Asen beobachten. Dennoch war der gesamte Zeitaufwand für das Sichern aller imSprung lebenden Tiere positiv mit der Gruppengröße korreliert.
Eine weitere Möglichkeit, den Zeitaufwand für das Sichern zrt redrzieren, ist die Nut-zung von deckungsreichem Gelände (Druoxo u. Lezanus 1974). Ist jedoch wenig
Deckung vorhanden, wie z.B. im Winter, besteht für das Tier noch die Möglichkeit,
seine Aktivität vermehrt in die Nacht zu verlegen. Diese Annahme stimmt gut überein
mit Untersuchungen von Gurttönl (1987) und JBpprsEN (1989) für das Reh, Borea u.
McCulr.oucH (1990) und KluurnuEyER u. MancurNroN (L977) für den Weißwe-
delhirsch und JTppESEN (1987) frir das Rotwild. Die von Gnoncrr u. Scunöopn(1981) untersuchten Rothirsche hatten in deckungsreichem Gelände gleichmäßig über
den Tag verteilte Aktivitätsschübe, dagegen waren Hirsche in deckungsarmen Regio-
nen vorwiegend nachtaktiv. Sie leiten daraus ab, daß das Aktivitätsmuster eines Tieres
auch abhängig von der Habitatstruktur ist.

15
2 Material und Methoden
2.L Das Versuchsgelände
Die Untersuchungen wurden in einem ca. 0.3 ha großen, gut einsehbaren Versuchsgehege
am Forschungsinstitut für Wildtierkunde und Okologie durchgeführt (siehe Abb. 1).
Dabei handelt es sich um eine nach Norden hin leicht geneigte Wiese am Rand eines
Buchen-Fichten-Eichen-Mischwaldes in unmittelbarer Nachbarschaft zu den Instituts-
gebäuden. Einzelne Flächen des Geheges wurden mit verschiedenen Futterpflanzen
(Luzerne, Mais, Raps, Markstammkohl) eingesät bzw. von der zweimaligen Mahd aus-
gespart. Sicht- und Witterungsschutzwar ntsätzlichzum hohen Gras auchdurch einige
Büsche, Fichten und Thujen gegeben. Fast sämtliche für die Untersuchung benötigten
Videoaufnahmen wurden vom Fenster des Futterlagers aus gemacht, nur einige wenige
erfolgten von einer Beobachtungskanzel. Das für die Flugdrachenversuche notwendige
Seil wurde am Nachmittag des 24J,A.90 in ca. 4*5 m Höhe quer über das Gehege
gespannt.
:iii!ii!+
,2,
..*N\
il1ilil
=
ungemähteWiese
v e r unkra ute teFläche
Fichtongruppe
Thujengruppe
Sträucher
Mais
ffi stron
o Malbaum
16"17'E 48'15'Nca. 360m Seehöhe
Abbildung 1: Versuchsgehege
Beobachtungskanzel
..i.titit,+.:t,
ffi.o
:
=lr rllllllr
S

16 2 MATERIAL UND METHODEN
2.2 Versuchstiere und LJntersuchungszeitraum
In die Störversuchel wurden drei Stück Rehwild und zwei Stück Rotwild einbezogen.
Die Tiere stammten aus Gehegehaltungen, waren jedoch nicht handzahm. Um mögli-
che gegenseitige Beeinflussungen der beiden Tierarten zu vermeiden, wurden die Un-
tersuchungen von Reh- und Rotwild zeitlich getrennt vorgenommen (vgl. Tab. 1). Kli-madaten während des Untersuchungszeitraumes sind im Anhang (Tabelle 23 tnd 24)
angegeben.
Tabelle 1: Allgemeine Daten zu den Versuchstieren
Alle Versuchstiere waren mit implantierten Aktivitäts- und Herzfrequenzsendern ver-
sehen (frir technische Details der Sendereinheiten siehe Scuosun et al. (1982)). Die
Sender waren im linken Abdominalbereich zwischen dem Peritoneum und der Bauch-
wandmuskulatur implantiert und mit zwei EKG-Elektroden subkutan im Brustbereich
und einem subkutan im Nackenbereich befindlichen Quecksilber-Neigungsschalter ver-
bunden" Auch die Leitungen zu den Elektroden und zum Neigungsschalter sowie
die Antenne verliefen subkutan (weitere Details zur Implantationsmethode sind bei
Wacmpn (1991) angegeben). Durch die Implantation der gesamten Sendereinheit
wurde eine langfristige Übertragung der Meßwerte, ohne stärkere Behinderungen der
Tiere, ermöglicht. Außerdem entfiel dabei die Gefahr einer Zerstörung der Senderein-
heit infolge bestimmter Verhaltensweisen der Tiere (2.B. Suhlen beim Rotwild).
Vom Sender wird die Herzfrequenz übertragen, und zwar pro QRS-Komplex des EKGs
ein Impuls von mehreren Millisekunden. Je nach Stellung des Kopfes (erhoben oder
gesenkt) wird durch den Neigungsschalter die Dauer des Impulses moduliert.
lDie vom Bundesministerium für Wissenschaft und Forschung erteilte Genehmigung für den Tier-
versuch gemäß §8 Tierversuchsgesetz 1988 liegt unter der Nummer GZ 68 2051293-L2190 vor.
Tier-
Nr.
Tier-
art
Geschl. Alter
(Jahre)
Ohr-
marke
Gewicht
(ks)
Sender
irnplantiert
Versuchs-
zeit
1
2
o
4
5
Reh
Reh
Reh
Roth.
Roth.
Id
Id
I
2
3
3
1+
I
bf)
23
3416e
070
073
20
25
20
75
45
05.06.90
i8.07.90
16.10.90
28.11.90
03.12.90
10.09.90-18.11.90
16.09.90-18.11.90
26.10.90-18.11.90
10.12.90-30.03.91
11.12.90-30.03.91

2.3 Die Störversuche
Die, mit geringfügigen Ausfallszeiten, über die gesamte Versuchszeit empfangenen Da-
ten über Herzfrequenz, Kopfstellung und Empfangsfeldstärke wurden durchgehend von
zwei PCs aufgezeichnet. Die Empfangsfeldstärke ist von der Position der implantier-
ten Senderantennen zur Empfängerantenne abhängig und gibt Aufschluß über Orts-
veränderungen des Tieres. Die Software wurde von ScuogER et al. (1988) entwickelt.
Technische Angaben zu den Empfangsgeräten finden sich bei ScHosnn (1986).
2.3 Die Störversuche
Mit zumindest halbstündigen Abständen zwischen den einzelnen Versuchen wurden un-
terschiedliche standardisierte Störreize in unwillkürlicher Reihenfolge gesetzt. Außer-
dem wurde darauf geachtet, daß jede Störversuchsart zu verschiedenen Tageszeiterr
vorkam. Nachstehende Störreize wurden gesetzt, dabei ist die Anzahl der Versuche für
Reh-/Rotwild in Klammern angegeben:
r Optische Störversuche
Rundgang einer Persoz.' Eine Person geht innerhalb des Geheges am Zaun
entlang (Dauer 3-4 Minut€ri rz : 29132).
Rundgang einer Person mit Hund: Eine Person geht mit angeleintem Kurz-
haardackel innerhalb des Geheges am Zaun entlang (Dauer 3-4 Minuten;
n:77 lt7).
Rundgang einer Personengruppe; Mindestens drei Personen gehen, sich in
normaler Lautstärke unterhaltend, innerhalb des Geheges antt Zaun entlang
(Dauer 4-5 Minut€oi ?? : 12114).
Rundritt auf einem Pferd: Abwechselnd auf einem schwarzen oder weißen
Isländer wird innerhalb des Geheges im Schrittempo oder Trab und Galopp
arn Zaun entlanggeritten (Dauer 2-4 Minuten; n : 79131).
Rundritt in, einer Pferdegruppe: Mit drei Islandpferden wird innerhalb des
Geheges im Schrittempo am Zaun entlanggeritten, dabei unterhalten sich
die Reiter in nortnaler Lautstärke (Dauer 4-5 Minuteni r?. : 5/10).
Flugdracheu Ein Drachenmodell (Spannweite 1,5m) gleitet in einer Höhe
von 4-5 m über dem Erdboden an einem quer über das Gehege gespannten
Führungsseil entlang (Dauer 2-3 Minut€rli 12 : 10114).
17

18 2 MATERIAL UND METHODEN
Traktor: Mit einem Kleintraktor wird außerhalb des Zaunes entlanggefahren
(Dauer 5 6 Minuten, n : 8/5).
- Iianzelaufsti,eg: Eine Person besteigt über eine Leiter die Beobachtungskan-
zel (Dauer 1-2 Minut€r, r? : 12l-).
- Futteruorlage: Täglich wird zur selben ZeiL (15 h) die Fütterung durch
dieselbe Person beschickt (Dauer 3-5 Minuteri ?2 : 31129).
o Optischer und olfaktorischer Störversuch
- Feuer: Ein Strohballen wird innerhalb des Geheges in Brand gesetzt (Dauer
8-l0Minuten; n:-15)"
o Akustische Störversuche
- RadiomtLsik: Yon einem am Zaun befestigten Lautsprecher mit einer Spiel-
dauer von drei Minuten (n : 12112).
- Hundegebell: Abspielen von einern Kassettenrekorder in der Folge 20 Se-
kunden Gebell, 30 Sekunden Pause und erneut 40 Sekunden Gebell (n :6112).
- Motorsägen: Arbeit mit einer Motorsäge in der Nähe cies Geheges (Ent-
fernung vom Gehegezaun ca. 20 m) für die Dauer von drei Minuten (n :11/11).
- SchuJlabgabe: Zünden eines Knallkörpers in der Nähe des Geheges (n :24117).
Bei allen akustischen Störversuchen wurde darauf geachtet, daß die durchführenden
Personen von den Versuchstieren optisch nicht registriert werden konnten.

2.4 Die Datenerfassung und Auswertung der Daten
2.4 Die Datenerfassung und Auswertung der Daten
Für diese Untersuchung wurden sowohl während der Störversuche gemachte Video-
aufnahmen herangezogen als auch die vom PC gespeicherten Telemetriesignale über
die Empfängerfeldstärke (,,Pegel"), die Kopfstellung und die Herzfrequenz der Tiere
(Online- und Offline-Daten). Die vom Tier empfangener Biotelemetriesignale wur-
den als minutenweise verdichtete Datensätze atl Magnetband gespeichert (,,Offline-
Dateien") und stellten die Grundlage für langfristige Untersuchungen dar. Gleichzei-
tig waren unverdichtete Einzeldaten von Herzschlag nt Herzschlag jederzeit am PC-
Bildschirm ersichtiich und wurden ebenfalls abgespeichert (,,Online-Dateien"). Diese
für kurzfristige Untersuchungen gut geeigneten Online-Dateien konnten im Playback
analysiert werden und ermöglichten durch die hohe zeitliche Auflösung die Rekonstruk-
tion jedes einzelnen Herzschlages.
2.4.1 Auswertung der Langzeitaufzeichnungen
Die Offline-Daten geben Aufschluß über die Gesamtaktivität eines Tieres über einen
längeren Zeitraum, und zwar, ob das Tier sich zu einer gegebenen Zeit, aktiv oder in-
aktiv verhielt. Als passiv gilt das liegende Tier, alle anderen Verhaltensformen gelten
als aktiv. Der Offline-Auswertung nach war das Tier aktiv, wenn es zu ständigen Pe-
geländerungen kam, da sich die Stellung der Antennen beim sich bewegenden Tier lau-
fend verändert und damit auch die Stärke des Empfanges. Zusätzlich war die Herzfre-
qtteßz gegenüber dem passiven Zustand erhöht, und es kam zu häufigem Wechsel der
Kopfstellung (Abb. 2 u. 3). Der Übergarrg z17rr_ aktiven Zustand war gekennzeichnet
durch Pegelregelungerr, Kopfstellungswechsel und einen deutlichen Anstieg der mittle-
ren Herzfrequenz. Die minimale Herzfreqttetz stieg erst in der darauffolgenden Minute
an. Die Herzfrequenz sank meist wieder auf ein bestimmtes Niveau herab, das aber
höher war als in der vorhergehenden Liegephase (Abb. 4). Beim Üb"rgung zum in-
aktiven Zustand war die Absenkung der Herzfrequenz allmäihlich über einige Minuten
hingezogen (Abb. 5). Wenn das Tier inaktiv war, d.h. gelegen ist, kam es zu keinen
Pegelregdlungen, und die Herzfrequenz'war gleichmäßig niedrig. Wenige Kopfstellungs-
wechsel konnten vorkommen, z.B. beim Putzen oder Asen im Liegen (Abb. 6 u. 7).
19

20
Tag: 26. tStunde: 22
llinute
Herzf requenz : gemitteltHerzfrequenz: maximalHerzfrequenz: minimal
Anzah1 d. f,opfrechself,opfuntenzeit [sec]
max. ununt€rb. f,U-Zeitmax. ununterb. f,U-Zeit
Anzahl d. Pe6elregel,Pegel: gemitteltPegel: maximalPegel: minimaL
Ausfallzeit [sec]
Tag: 27. 1
Stunde: 2t
Hinute
Herzf requenz : gemitteltHerzfrequenz: maximalHerzfrequenz: minimal
Anzahl d. f,opfnechself,opfuntenzeit [sec]
max, ununterb, f,U-Zeitmax. ununterb. tro-Zeit
Anzahl d. Pege1regel.Pegel: gemitteltPegel: maximalPegel: minimal
Ausfallzeit [sec]
Abbildung 3: Beispiel für eine
motorischer Aktivität. Die hohen
typisch für Fortbewegung.
9
829074
6060
5;5454
10
a29274
,494411
545454
11
8611076
4421810
I555844
t2
96LL286
44t2A13
t2556247
13
9810688
60u:
2t466031
L4
98110a4
,5947
1
6535748
15
9210880
60u:
535353
16
941087a
60u:
545454
,.7
9611088
60u:
4565752
18
9410880
60u:
5
54564A
19
9210486
60u:
6545547
20
92106
a2
,594A
1
18525847
21
9411080
60u:
11
485739
22
92LL476
44A26I
t7555946
43
98130a2
9t24
19
26579842
44
142308
86
88,
26
32326844
u
45
120150100
2II
46
101040oo
46
t76374106
662
26
3636964A
a
47
138L72116
11
L75
13
31
319028
48
160zooL24
9104
19
35359940
4
49
t72368t20
t219
20
2L
2t5439
)
50
118136102
4252319
11
LL5339
51
981L290
4
9
8
2L
66
4739
52
116L46
90
10104
37379738
53
t62410104
931
32
2a2A6238
4
54
166308146
11
73
22
3939974L
2
7
7
52
2
55
176304130
16113
30
56
t24206t02
5
5
324
9I
4A31
2 MATERIAL LIND METHODEN
Offlinedatei eines Tieres (Rothirsch Nr. ) bei loko-
Herzfrequenzen und häufigen Pegeländerunger sind
Abbildung 2: Beispiel für eine Offline-Datei eines Tieres (Rothirsch Nr. 4) im aktiven
Zustand. Die lange Kopfuntenzeit und mittlere Pegelregelungen sind typisch für Asen
oder olfaktorische Kontrolle des Bodens.

2.4 Die Datenerfassung und Auswertung der Daten
Tag: 28. 1
Stunde: 19
llinute
Herzf requenz : gemitteltHerzfrequenz: maximaLHerzfrequenz: minimal
Anzahl d. f,.opfwechself,opfuntenzeit [sec]
max. ununterb. f,U-Zeitmax. ununterb. f,0-Zeit
Anzahl d. Pegelregel.Pe6e1: gemitteltPegel: maximalPegel: minimal
Ausfallzeit [sec]
Abbildung 4: Offlinedatei eines Tieres (Rothirsch Nr. 4), das um 19.34 Uhr aufsteht.
27
Iag: 26. L
Stunde: L7
llinute
Herzf requenz : gemitteltHerzfrequenz: maximaLHerzfrequenz: nininal-
Anzah1 d, f,opfrechself,opfuntenzeit [sec]
max. ununterb. f,U-Zeitmax. ununterb. f,o-Zeit
Anzah1 d. Pege1regel.P€ge1: gemitteltPegel: maximalPegel: minimal
Ausfallzeit [sec]
Abbildungniederiegt.
t42
94t9280
8261021.
25586251
43
98LL2a4
73925
11
o
586051
44
10482
4
23
2026
b
585950
45
110TO
2
2323
26
16
515843
46
768272
bU
4;4242
47
74
70
60
2
36100
48
74tö68
6;
32
32
49
72/b
68
6;
353535
50
748666
:60
353535
51
727668
-60
353535
52
727670
60
3;3535
53
727A66
60
3;3535
54
727670
60
3;3535
55
727670
u;
353535
Offlinedatei eines Tieres (Rothirsch Nr. 4), das sich um 17.45 Uhr
7A
9668
6;
535353
8o9670
6;
535353
76a268
6;
505050
28
748470
6;
51
5151
,o
728268
6;
5151
51
31
a210870
6;
505050
34
8813674
1
I9
51
9536253
35
10013668
3109
30
1055s748
36
104L2474
6301618
21
545946
37
106L2492
332192t
1
4610046
38
L04LL494
1
3838,,
54651
37
39
88L7476
4201025
,333432
40
88L0274
92LL4t2
3323530
4t
94LL284
3117
42
16314tL7

22 2 MATERIAL UND METHODEN
Tag: 7. IStunde: 3
llinute
Herzf requenz : gemitteltHerzfrequenz: maximalHerzfrequenz: minimal
Anzahl d. (opfrechself,opfuntenzeit [sec]
max. ununterb. f,U-Zeitmax. ununterb. f,0-Zeit
Anzahl d. Pege1regel,Pegel: gemitteLtPegeL: maximalPegel: minimal.
Ausfallzeit [sec]
Abbildung 6: Offlinedatei
den Kopf auf den Boden legt
eines ruhenden
(Schlaf).
Tieres (Rehgeiß Nr. 1), das um 3.5o Uhr
Tag: 5.9Stunde: 15
I{inute
Herzf requenz : gemitt€Itllerzfrequenz: maximaLHerzfrequenz: minimal-
AnzahL d. f,opfrechself,opfuntenzeit [sec]
max, ununterb. f,U-Zeitmax. ununterb. f,o-Zeit
Anzah1 d, PegeJ.regel.Pegel: gemitteltPegel: maximalPegel: minimal
Ausfallzeit [sec]
Abbildung 7: Offlinedatei eines ruhenden Tieres (Rehgeiß Nr. 1). Ab 15.46 Uhr
kommt es zu einigen kleinen Pegeländerungen und Kopfbewegungen. Die Herzfrequenz
steigt nicht sonderlich an. Dies kann als Asen oder Putzen im Liegen interpretiert
werden.
Beim Rehbock funktionierte der Neigungsschalter jedoch nur bis zum 7"10.1990, beim
Hirschspießer bis z:uirrr 4.2.L991 und beim Rotwildkalb von Beginn an nicht. In die-
sen Fällen konnte aber trotzdem weitgehend zwischen aktiv und inaktiv unterschieden
werden. Zur Kontroiie wurden die Offline-Auswertungen mit den synchron gemachten
Videoaufnahmen verglicher. Der Anteil der falsch interpretierten Minuten betrug bei
4t
929684
uo
555555
42
869476
ui
555555
43
a2887A
uo
555555
7A
8468
uo
555555
45
a28676
uo
55
46
a29274
:60
55
55
55
47
809070
uo
575757
48
80o,74
:60
545454
L
4646t4
55§555
51
7A
8668
60u:
55
5555
52
768072
60u:
555555
53
768670
60u:
555555
54
749866
60u:
555555
60
52s252
42
667264
60
545454
44
647260
uo
5252s2
45
646862
46
6672
uo
1
57
'-n
47
68L20
58
1
?
58
5
535345
48
6266
1
50
5010
535353
49
606854
2EE
5
545454
50
607052
4
46zo10
545454
51
627A58
2
342622
545454
52
749660
4ö
6
25
545454
53
606856
4444
8
545454
49
808676
uo
555555
4t
647056
ui
51
51
51
43
687662
uo
51
51
51

2.4 Die Datenertassung und Auswertung der Daten
den Rehgeißen zwischen 1,470 :und 4Yo" Die Fehler traten fast ausschiießlich bei kurz-
fristigen Aktiv- bzw. Inaktivschüben unter fünf Minuten auf (vergl. JnppBsoN 1989).
Fälschlicherweise als aktiv bewertet wurden Situationen, in denen ein Tier sich nur
einige Minuten hinlegte und darau{hin wieder aufstand, fälschlich als passiv, wenn ein
Tier einige Minuten regungslos stillstand, z.B. beim Stehen und Sichern, da sich hier
keine Pegelregelungen zeigten, der Kopf ununterbrochen erhoben und die Herzfrequenz
oft erstaunlicherweise niedrig war.
Der Rehbock ist o{t lange Zeit stillgestanden, statt sich hinzulegen, zeigte dabei aber
alle Anzeichen inaktiven Verhaltens und verbrachte teilweise auch längere Zeit mit
Wiederkäuen im Stehen. In 13,2% der nach der Offline als inaktiv bewerteten Mi-
nuten ist der Bock gestanden, davon zr 40,2% mit Wiederkäuen und zu 40.2% rrrit
Sicherverhalten.
Bezogen auf die Anzahl der Aktivitätswechsel (von aktiv zu inaktiv und vice versa) lag
dieFehlerquotebeill,T%fürReh1und16,7%beiReh2. DiesistdurchkurzeAktiv-bzw. Inaktiv-Phasen unter 5 Minuten erklärbar, die oft nicht erkannt wurden. Auf
die Gesamttagesaktivität bezogen ist der Einfluß solcher Fehlinterpretationen zu ver-
nachlässigen, jedoch werden andere Kenngrößen der allgemeinen Aktivität, wie Anzahl
und durchschnittliche Dauer der Aktivphasen, dadurch verändert.
Zurn Vergleich zwischen Tag- und Nachtaktivität wurde die Zeitspanne von einer hal-
ben Stunde vor Sonnenaufgang bis zu einer halbe Stunde nach Sonnenuntergang als
Lichttag bezeichnet" Kamen an einem Tag bzw. an einer Licht- oder Dunkelphase mehr
als 5% Datenausfälle oder nicht interpretierbare Offline-Daten vor, so wurde der ge-
samte Tag bzw. der jeweilige Lichtiag oder die entsprechende Nacht für die Berechnung
des prozentualen Inaktivanteiles nicht herangezogen. Für die Auswertung der Dauer
der Liegephasen wurden ausschließlich jene Inaktivphasen, denen nicht ein Ausfall vor-
anging oder nachfolgte, einbezogen. Zur Auswertung der Anzahl der Liegephasen am
Lichttag wurden nur Tage verwendet, an denen keine Ausfälle vorhanden wa er].
2.4.2 Auswertung der Kurzzeitaufzeichnungen
Die Online-Daten eignen sich gut, um Aussagen über kurzfristige Veränderungen der
Herzfrequetz nach einer Störreizsetzung zu treffen. Sowohl das Ausmaß der Herzfre-
quenzveränderungen ais auch deren Dauer bis zum Erreichen der Ruhewerte könnet zlJr
Beurteilung der Intensität der Beunruhigung herangezoget werden. Wenn die Herzfre-
23

24 2 MATERIAL UND METHODEN
qlueuz infolge einer Störreizsetzung um mehr als die doppelte Standardabweichung von
der durchschnittlichen Herzfrequenz in der Minute vor Beginn des Störversuches an-
stieg, wurde dies als Beginn der Störung angesehen. Als Ende der Störung wurde
jener Zeitpunkt gewählt, ab dem die Herzfrequenz mindestens eine Minute wieder
innerhalb der durch die doppelte Standardabweichung bestimmten Grenzen des vor
Störreizsetzung liegenden Zeitraumes verlief. Von besonderem Interesse ist die Anzahl
der Herzschläge, die als Auswirkung eines Störreizes zusätzlich zu denen, die während
des unmittelbar vor Störreizsetz;.lrr.g gezeigten ,,Normalverhaltens" gezählt wurden, ge-
leistet wurden, da sie in einem engen Zusammenhang mit dem durch die Störung
verursachten vermehrten Energieaufwand stehen. Diese entspricht der Störungsinten-
sität. Um diese zusätzlichen Herzschläge zu ermitteln, wurde das Integral der Fläche
zwischen der Herzfrequenzkurve und dem mittleren Herzfrequenzniveau vor Störreiz-
setzung zwischen Beginn und Ende der Störung belechnet (siehe Abb. 8).
300
250
200
150
100
Zeit Imin]Abbildung 8: Modell einer Störung unter Verwendung der Herzfrequenz als Kri-terium: Verlauf der Herzfrequenz vor, während. und nach einer Störung des Tieresl
B: Beginn der Störreizsetz;t;orrg; E: Ende der Störreizsetzrngl schattierte Fläche:Anzahl der ntsätzlich geleisteten Herzschläge.
tr
o)ä0
:CB
.'l(.)
V)
Ntro)
orotiqiNk(l)
50
1510
Dauer der Herzfrequenzerhöhung
Störungs-inten sität
i; t=

2.4 Die Datenerfassung und Auswertung der Daten
2.4.3 Verhaltensanalyse
Das Verhalten der Tiere wurde während eines Störversuches sowie jeweils zehn Minu-
ten (bei den Rehen fünf Minuten) vor und nach einem Versuch mit einem 8mm Sony-
Camcorder aufgezeichnet. Es wurde anhand der Aufnahmen ein Verhaltenskatalog
(Ethogramm) erstellt, der als Auswertungsgrundlage für die Videoaufzeichnungen diente
(siehe Abb. 49 im Anhang). Der Beginn jeder Verhaitensänderung wurde protokolliert
und die Dauer in Sekunden bis zur nächsten Verhaltensänderung errechnet (Abb" 50 imAnhang). Da das laufende Uhrwerk der Videokamera täglich neu mit dem PC für die
Herzfrequenzarfzeichnungen synchronisiert wurde, war es möglich, die Herzfrequenz
mit dem Verhaiten zu korrelieren.
Es wurden 48 Verhaltensweisen ermittelt, die sich zu 7 Verhaltenskategorien zusam-
menfassen ließen. Folgende Verhaltenskategorien wurclen bei cler Auswertung berück-
sichtigt:
Liegen: In diese Kategorie wurden alle Verhaltensweisen eingeordnet, bei denen
das Tier lag. Dazu zählte das Liegen mit und ohne Wiederkäuen, das Liegen mit
dem Kopf flach auf dem Boden, das eingerollte Liegen, sowie das Putzen, das
Asen und die olfaktorische Kontrolle im Liegen.
Stehen: Das Tier steiit ohne zu sichern. Sowohl Stehen mit und ohne Wie-
derkäuen wurde hineingeordnet.
Stehen und Sichern.' Das vorhergehende Verhalten (2.B. A."rr, Putzen,.. " ) wird
unterbrochen und der Kopf gehoben. Kennzeichnendes Merkmal sind die gestei-
gerte Aufmerksamkeit und das Prüfen der lJmgebung mit den drei Sinnesorganen
Geruch, Gehör und Clesichtssinn. Es liegt eine erhöhte Fluchtbereitschaft vor.
Fortbewegung: In diese Klasse gehören das Ziehen (Schritt), der Troll (Trab)
und die Flucht (Galopp), aber ebenso das Sich-Drehen auf der Stelle und das
Rückwärtsschreiten.
Olfaktorische Kontrolle; Bei der olfaktorischen Kontrolle des Bodens ist der Kopf
gesenkt, der Nasenspiegel gleitet nur wenige Zentimeter über den Boden. Diese
Verhaltensweise ist leicht zu verwechseln mit dem Asen, doch fehlen die Kaube-
we§ungen, der Mund ist geschlossen. Auch das Beriechen höherer Gräser wurde
hier zugeordnet; in diesem Fall ist der Kopf allerding etwas höher gehoben. Das
25

26 2 MATERIAL LTND METHODEN
Tier kann während der olfaktorischen Kontrolle auch ziehen oder traben. Bei
der olfaktorischen Kontrolle der Luft wird der Nasenspiegel schräg nach oben
gerichtet, manchmal unter Verdrehen des Halses.
Nahrungsaufnahme: Beim Asen ist der Kopf gesenkt, es sei denn, höhere Gräser
oder Aste werden aufgenommen. Im Gegensatz zur olfaktorischen Bodenkontrolle
sind beim Asen die Kopfbewegungen, bedingt durch das Rupfen des Grases,
ruckartig. Außerdem erkennt man Kaubewegungen. Beim plötzlichen Sichern
hängen oft noch Grashalme aus dem Mund. Als Nahrungsaufnahme zählt auch
das Fressen an der Fütterung (Langheu, pelletiertes Ergänzungsfutter). Dem
Rotwild wurden auch Rüben vorgelegt.
Sonstige Verhaltensforn'Len: Zum Komfortverhalten zählt das Putzen des Fel-
les mit der Zunge, den Zähnen,, den Klauen oder dem Geweih, aber auch das
Körper- und Kopfschütteln, das Strecken von Hals und Extremitäten und das
Suhlen. Weitere Verhaltensformen, die erfaßt wurden, sind Scharren, Harnab-
satz, Schälen, Kopfreiben, Trinken, Bocken, Kämpfen, Springen, Flehmen und
Gähnen.

27
3 Ergebnisse
Aufgrund der verschiedenen Biologie von Reh- und Rotwild und des unterschiedlichen
Zeitraumes der Untersuchung beider Tierarten werden die Darlegung der Ergebnisse
und die dazugehörenden Erläuterungen für das Reh- und Rotwild getrennt vorgenom-
men.
3.1 Ergebnisse für das Rehwild
3.1.1 Wahl des Liegeplatzes
Aus den Videoaufnahmen wurde die Wahl des Liegeplatzes der Tiere ermittelt. Die
einzelnen Liegeplätze wulden zusarnmen mit einer Daturn- und Uhrzeitangabe auf
einem Gehegeplan eingezeichnet.
Abgesehen von einer Ausnahme, sind die Rehe vom 20.09.90 bis zl:r'n 24.10.90 mittags
ausschließlich im großen Grasfeld gelegen (Abb. 9). Ab dem Nachmittag desselben
Tages haben die Rehe jedoch ihre Ruheplätze, wiederum bis auf wenige Ausnahmen,
nicht mehr in dern großen Feld aufgesucht, sondern bevorzugten entweder den kleinen
Fichtenbestand oder die nordöstliche Ecke des Geheges als Liegeplätze (Abb. 10).
Abbildring 9: Prozentuelle Verteilung der Liegeplätze beim Rehwild bis zum 24.10.90
um 12 Uhr (n: 53 Liegeplätze).
n"**,u
o,*.HolunderQ

28 3 ERGEBAIISSE
Abbildung 1O: Prozentuelie Verteilung der Liegeplätze beim Rehwild ab dem 24.L0.90
um 15 Uhr (n : 102 Liegeplätze).
Wechsel des Liegeplatzes nach Störuersuchen"
Im frühen Herbst suchten die Rehe immer das große Feld als Liegeplatz auf und änder-
ten dieses Verhalten auch nicht durch Störungen. Dagegen wechselten sie im späten
Herbst nach einem Störversuch, durch den sie aktiv wurden, in 35,3% der Fälle auf
einen anderen Liegeplatz als vor der Störung.
3.1.2 Aktivitätsmuster
Für jedes Tier wurde anhand der Aktiv/Inaktiv-Werte aus den Offline-Dateien ein
Aktogramm erstellt. Die für das Rehwild ausgewertete Aufzeichnungszeit aus den
Offline-Dateien betrug etwa 227 500 Minuten. Es zeigten sich keine deutlichen indi-
viduellen Unterschiede in den Aktogrammen" Allerdings änderte sich das Aktivitäts-
muster der Rehe im Verlauf des Herbstes: Etwa um den 21.10.90 kam es zu einer
deutlichen Erhöhung der Aktivität während des Lichttages, wohingegen die Tiere imvorangegangenen Zeitraurn des frühen Herbstes während des Lichttages überwiegend
inaktiv (gelegen) waren (vergl. Abb. 11-13). Aus diesem Grunde wurden alle weiteren
Auswertungen der Aktivität getrennt in ,,frühe Herbstphase" (10.09"90-20.10"90) und
,,späte Herbstphase" (21.10.90-18.11.90) vorgenommen. Entsprechende Klirnadaten
siehe Tabelle 23 irn Anhang.

UHRZEIT ThI
f,,1,,17;,lt,,1
I l:rI:a::::l:i::.:.': .:::"'::l t r.rll::. ll::.:::,1 l::: ,:l. lt I:t::
lrrlrrlttltrt I I t-,:::, t-i lll,:.,rt
-
lI:+::- illl r::.]I::- ::rI I::t:. Ilr::t rII I t :::,:::l :,,.. a,lt'. t:,r II I l: rI :a::::l:i ::.:.:: ,:."',,1 t,r.rll
3.1 Ergebnisse für das Rehwild 29
2l , ,9::: :::::::::::I :::::::
:::'::l:::-:::::::l :t.::::::::::-::::::::-l::::-l:::j,,,,....I: t,-:
I l..r it..::. 1:::.:i I,':'::Irr.l
- - II:
i: -::::::::::::.::
I',::,.taa|-ii
-:i::[l I
I I :i.irr;rl, I,iI ':::::::-xI i:.:::-l:x.r:I:::l::i f
t:, ,:::.. I::, x
-rr:l:Ix
I'::::: I X:r:'lI X!:::;:rjjt Ix,i:ri:Iiiliiirllll X
I I ':::-.,.1::r'I.::::': f rrr:r.,r,. Il:i.,:: .:.1 I l,r,.l ; :t::'.:::: ,tt:. ,,:::a, ii::r}r.l li:, Ii:, |:,itii I
i': :hI':., I:::l tr::r::':. I,::::.1'r.: II,:i:,rI:.. II|i r.l::l [I:::,;.,' ,l
I,:ir:!' ::rri:r-XI'r,,. l.,.,triIrx
llrtti: :I :Ix:,:Itr ,.,t:x
I,:: l:I:.1I I I::I :..
i: iIi:i'-Xi.: i Ii:'r-XIi ir:',1i11:IX
Il: x,IrxI -
,,.:::: lx'Il::I
I :.-
-:,:r::I
ll
-
:,:r::IX,:,-..::::.,. I
20.10:I.,,i,r-
22-10- ll :, r,,
It,::ro7.1'l-r,, r i
IIO9.'l 1. I t: r r::j
l::I | :,. : :::l::[:::]::l::::::::!rr rr I
-.
a::-:::::.::::j,:a::::!::.!.:Llt:j::i:i:::.:rr..iüäit:l ä:äitaäääia|i|
-:::j:1
r:l:::::::::::::::::l::::l::::::!;r;.rll:!r:l:li:liiliiiiiiiiliil:l111i1:iJl111ä!!äää:.i)ri:)ä:;L:r i:t::l;lt;:I!:::i:::.:1;t:;;ri:;l:iala:]l'tirllll:
.:: .::..- irijj:il tj::::! j:1.:
l:r:r:::::::"I ::::::l:::- titii'IiiiiiiI
--::::::::::I:
| .r,Iiii I:::I: ltl :i::
.II,,.x.IiIx:::I.,.::::: x
Il.:::r,rr-I,:::::l::I,:, Ir'l:::i:::::I:I
I:,i I24-1O. a ' ,,: I,'': I [:,] .::::::I:,r.r I ':''f :l :::II:::. I:t:t a I ,li:::,::iiII:.:i,ir: ::'r I i
26,1 O; irrri:'l::::ll::r'::.' ::: : II::::::::::::r:::I,''' I
28.1 o -r:::::::i:':;:::'-" r:,.::Il ,,.,,1 :,:,1 I:ti,, t
I I I .I ,-:I,''::Il::::'. I :::::II :::,:;i:'i::,Ii ii, iiil
30.1 O- rlr::r:::.:r:.:,I
Il:,ti;itr:rtririilitii:rtr IIII,l'1a11,,11!,11a,1!.14 | | i.i:i:i:i:ttlila
a::aa::a. :a::::::.a:-:::::t:,1' a::.a:::::t':1 |:i:::::::::::::i::-] j,::.:::::::::::l
:::::::::::I()1-'l 1r -:,t::::::::::,
rI,.. ', l-:r,i:I:I Il::l:l:.,
I I I::1.:::::::::::I:.,:I
- lx
::::::rI::::,::r::::I,, I,fI :::ri::::::I I
O3.l 1; I .:-I I 1, I t:i::i-:.:l.. I I I I:.'-I
I::::::::::::J::I:']: r::. x:::r::.I::ri:,:r:rr- :r,, x.TI. I Io5.11.r j r. I
a:,i';a t:ti i:.at taiijt',.-,r.,.-,1;,:::r j l,Ii'+:::. xililll:iii:ij:ii .,!,,::, t:::::..t.:,I, X:.:::l:' ::: . I.
:I:::[::::::::.']::::I I::::::::r:l x::I:::t:::::::::::r, I,::::::I x
. I I:::X
I,t!ri:iri:::l::Iii:::I::fu,tll iil:t:,:I x
-
:;::;::::::::I:::t:::::::::It::::::::::: x'ri.iii.1iti,:!.1:1it1i1tat.:t1a1,.a,1:ti.alrrll l,, Il X
l::l::1:rir:l:r::j::::1:|:::l:111n::1uu,I:::::ut x
| ..i::..I:::i..!:..:
-I.'.,11 i,:'la1::,r-x
f r.i I rjt:lIr il: XI " :,:::,I::::: ::.lxIlI,.::..:, IxIljtxIII:l::::::, Ix
Iiiiiiii r iiiiiiiiiiiiilt;t::t::::::::::t::i::1 3.1 1 .a:;;:;:;I,riri!i iriri!nr;;'
ll).111;a1.a,- ;. : :, aa:aa:a:aaaLaaLLall
1 5. 1 1 .fjijiliriilr:. !::: ! ::t-:a::::::::::::::,::.:,
Jlii:ajiili I ii'I:i:::i:a::::r:::17 -'l 1 - ::l:::::'::::: .;i'l
l,: I:..::...:.: I
lnaktiv I Aktiv
Abbildung 11: Aktogramm von Reh Nr. 1 (Geiß, 2 Jahre). Tage, an denen Störver-
suche durchgeführt wurden, sind an der rechten Seite mit ,,x" markiert. Ausfallzeitett
und nichtauswertbare Daten sind in der Abbildung weiß dargestellt. Die schräg nach
unter verlaufenden Linien markieren Sonnenauf- und -untergang.
02.1()04.10()6.1
o4.10't o.112.114.1'I 6.1 ()

30 3 ERGEBAIISSE
9,,?,'l 6.O9- riiirinir
I-IIOgrr
-
lI= I I
-II:
-''.r:,,','l'I I
r:ili i I
:ti:::a:::::::::::,I:::,.j, III't':. r , t
Iiilljljlil:llil::i::ii:r:lr::i.i:i1r::i:::ri:r:'r|:;::l::rljiiiiliiiill.::l:rrrt:';,-...iaiii..jj1äjirtliliäitl iIiliiliiiiiii:ii:i;iliiiiiiiiiiiiill;iii;!.ti:t:i:titi::,a..a,:..::.;::.'.jll!:,1.':a.!.n:!'aar::aalta:a::,t::,1:i',..aiA -,i,itiiä;tiir:':|tji:i)tltJ xI:J:II X
:al::r:l:.::::::::l:::l::::l:rii:rt.ä,a:ä4.,1 a,,,a,,'a:):aiaa;j,a:! :! |: ::::,::;::::i.1::;::i::::::::::::: :::t :::::::!J::i X
I ar.' ':: , ::il:, I l': lI,,:::::' 't. t.,: .: lt I:i:::tI r::: I::.r..
,i;!iill::i t,,.,,1i.,,,a4:aa: x,lü,trr1La-,:ii: x
-.r:::::t:::x-ii,::i,:
IIlt: f.l:: x:,.::::::I :1,:i;::::lil I lu
,:::::':': I::-:
:I.,:i:rI::II :::::I .:::rrl:::.,iI,l::,1'I:f':,1 l'r:l
,tt?. :.rltt al:.,a:.-:aaaaa.:;,a, x
I I il:l:!:::l::r llll:i::l:::li1:i:i. ' ,,i:::::::::::i:.:l::::::::a:.: :.:i:::::Il:;,::;::::::::::::::t:il:t::::::,aaa:'.ata.,aaa:,a':aaaaa::,1:',: t:ajaaaa!:a'jLitt.;ili ) ):41 :a :a': +a-:la::a : : :aaaaaa::::: i:: l'ir;ii::::;::.:r: :r1 ::- !:,,i I:::f rl:::r:[:::::]:rr1:;::::r::r:::::::'iilll:
Ii :,i.:, ::,. ::-:.... :
l,,l ,, I'j:: .:i:lll,:: ll I f:::rr:::l 'r:::l:lt,.
-
j:::::::'
I -:,:j
xI.f.x
I :I,'l :r:-1 r.:',ll!I .r..:.r: I.::::::.i lIlI I rf r' lx
I r
-:j:i::xI l'. I: I'lil'rl':'rll x
xx
lnaktiv I Aktiv
Abbildung L2: Aktogramm von Reh Nr. 2 (Bock, 3 Jahre). Tage, an denen Störver-
suche durchgeführt wurden, sind an der rechten Seite mit ,,x" markiert. Die schräg
nach unten verlaufenden Linien markieren Sonnenauf- und -untergang.
?,,1,,2 6 .1 O......,!..-a-ääääiääin
UHRZEIT ThI
, ? ,01?;','F-:
rf'24.1O-I I .::i:::::::;::::::::: I
l:;r:::-:::aliriilr iii iliiilljjiijiijjii:jlliilil lrlilli:ilill;lrliir ,,,iI
I
-:;:i::i::3 O. 1 O -::::::::::- iiiiiiiiiiii:i::I
:I:
-:::::,:.,1
jj-I ,.:Irrl
l::::::::il:lilttl tI,ri:ri :r,r,r:
O 1 - l 1 .:irallir li riliriiilltiitail!-.:,::lI f
I:::.I I I I]
aiiiiiiiijiii:l-,:::;::r::::t::,()3 -l I I ;ijiit;ii;;1:- :,t:t: t:iut::,
:ir,if .::,:,,1 I1..:r,:l
-:..1:::I.,I I
I l:ir I,::::ttIII:l-::l:r::.:l lI I
I lr.::::l:l I .:::.1If ::.r' , :lI l'. tIl,:
ll r:::ll I,:::::,, I , f ,j.j:,f:t:.,_Ir
-r,
-i,
Ili ,
-
i,::I','-,::l'I
xxxxxxxxxx
xxxxxxx
:-:I::::::::::-Iiiiiiiiiiiill:aili:iliir:ili
I 5. 1 1 -I:::iir: iiiii ri:iiI:rtl,::r,I I
-
,.:l::I ,::ll
l-r IlI::::::l I::,: I17.11.-, :.:.: r ,tl, :::ilr fi,.,t l,
l-r IlI:I,Il, II
. .l:::::It. lI
I:l I f l Il I ll-rl.::l I I::::, I.,lII-. l.
- .tI
II!,
-:::::
:.:,. Il-:l:rll l:r::-l:: :...:::II IIll tll,:::::lrl i I'IllIllI I
:::,, lnaktiv
-
Aktiv
Abbildung lb: Aktogramm von Reh Nr. 3 (Geiß, 3 Jahre). (vergl. Abb. 11.)
,,,1'Fä'IN', ,
()2.10.04.10.o6.oa.1().12-14.16.1a.1().20.22.24-26-2A.90.o1.1 Io3.1 'l
o5.1 1
07 -11o9.1 I11.11ts.1 1
15.1 1
17.11

3.1 Ergebnisse für das Rehwild
Tageszeitlicher Verlauf der Aktivität
Die Aktivität war bei allen Rehen zu den Dämmerungszeiten erhöht, sowohl im Früh-
als auch im Spätherbst. In Abb. 14 ist die tageszeitliche Abhängigkeit der Aktivität von
Reh Nr. 1 an ungestörten Tagen, in Abb. 15 an Tagen mit Störversuchen dargestellt. Es
ist zwischen Tagen mit und ohne Störversuche kein Unterschied in der tageszeitlichen
Aktivit ät festzustellen (Kruskal-Wallis- Test ) .
Abbildung 14: Tageszeitliche Abhängigkeit der Aktivität von Reh Nr. 1 an Tagen
ohne Störversuche im Zeitraum vom 8.10. bis 25.10.90. Zur Glättung der Kurve sind die
Aktivitätswerte über 5 aufeinanderfolgenden Minuten gemittelt und der Durchschnitt
von zehn Tagen errechnet. Grau schattierte Flächen entsprechen der Nacht.
Abbildung 15: Tageszeitliche Abhängigkeit der Aktivität von Reh Nr. 1 an Tagen
mit Störversuchen im Zeitraum vom 8.10. bis 25"10.90.
31
=rcoN
EEEooc,
äco!:e
€ 'lo
F|(zot]-yo
ErcoN
EEooo(!,
bco!§E10F{20FtFyo

32 3 ERGEBNISSE
Gesamtaktivitätszeit
Sowohl Reh Nr" 1 als auch Reh Nr. 2 waren im frühen Herbst überwiegend nachtaktiv,
unabhängig davon, ob es sich um Tage mit oder ohne Störversuche handelte (Tab. 2
u. 3). Der Rehbock (Nr. 2) zeigte sich weniger aktiv als die Geiß Nr. I (Mann-Whitney-
U-Test, p < 0.05), sowohl am Lichttag als auch auf clen 24-Stunden-Tag bezogen" Ab
dem 21.10.90 war bei beiden Tieren eine deutliche Zunahrne der Aktivität während
des Lichttages zu erkennen (p < 0.05). Auch das Reh Nr" 3, welches am 25.10.90
in das gleiche Gehege überstellt wurde. war vermehrt tagaktiv" Im Gegensatz z:urlrl
Rehbock (p < 0.001) änderte sich bei Reh Nr. 1 nach dem Wechsei in den späten
Herbst die Gesamtaktivitätszeit nicht. Die höchste Gesamtaktivität war bei Reh Nr. 3
zu verzeichnen, es unterschied sich in seiner 24-Stunden-Aktivität signifikant von den
beiden anderen Rehen (p < 0.01). Sowohl die Gesamtaktivitätszeit a1s auch die auf
den Lichttag bezogene Aktivitätszeit änderte sich infolge von Störversuchen weder im
frühen noch irn späten Herbst bei keinem der drei Rehe.
Tabelle 2: Yo-Anteil aktiv während des 24-Stunden-Tages beim Rehwild. (n : Anzahl
der Tage; med : meclian; r : arithm. Mittelwert; s : Standardabweichung; ntin
mex : Extremwerte)
24-Stunden-Tag
Saison/ | Tage ohne Störversuche I Tage rnit Störversuchen
Tier-Nr.in med .r s tnin-marl n med r s min-mar
Frühh./1
l2
12 42.4 40.2 7.5 39.2-53.2
9 29.7 28.4 4.6 19.9*35.9
15 39.8 40.7 5.9 32.5-52.6
7 28.0 28.2 4.5 27.0-36.5
Späth./1
l2l3t-
7 43.7 42.4 4.3 33.8 47.4
6 36.6 40.9 5.9 35.6-50.0
4 53.0 51.6 3.5 46_5-54.t
14 41.6 43.0 5.2 33.9-53.0
t7 43.3 43.6 7.0 31.8 {2.9
15 54.5 54.9 3.9 54.2 d0.8

3.1 Ergebnisse für das Rehwtld
Tabelle 3: %-Anteil aktiv während des Lichttages beim Rehwild.
Lichttag
Saison/
Tier-Nr.
Tage ohne Störversuche
nmed,rsmin-marTage mit Störversuchen
nmedrsmin-marFrühh./1
l2
12 25.9 29.8 14.5 11.2-60.5
I 17.5 16.5 5.9 0.5-26.8
15 28.3 28.1 8.8 14.7-50.t
8 12.3 14.0 6.3 7.7-24.7
Späth./1
l2
l3
7 52.8 49.9 8.3 35.2-59.5
6 44.7 *5.2 7.4 36.0-53.3
4 57.6 56.4 9.5 46.3-64.8
74 48.3 49.9 7.7 63.5-37.6
17 50.6 49.5 8.0 31.242.0
15 63.5 62.0 7.5 5t.9-7t.7
Ä.nzahl und Dauer der Liegephasen
Die Dauer der Liegephasen am Lichttag war an ungestörten Tagen individuell verschie-
den. Beim Rehbock (Nr. 2) waren die Liegephasen rund 20 Minuten länger als bei den
Rehgeißen (Tab. a). Reh Nr. 1 und Nr. 2 verringerten im frühen Herbst an Tagen
mit Störversuchen nur geringfügig ihre mittlere Liegephasendauer, im späten Herbst
wird jedoch bei allen drei Rehen die durchschnittliche Liegedauer durch Störversuche
deutlich reduziert (Kruskal-Wa11is-Test).
Tabelle 4: Dauer der Liegephasen (in Minuten) bei Rehen an Lichttagen ohne, mit
wenigen (2* ) und mit vielen (5-10) Störversuchen. (n - Anzahl der Liegephasen; r :arithm. Mittelwertl s = Standardabweichung; min*mar: Extremwerte)
Mit steigender Anzahl der Störversuche am Tag sank die Länge der Liegephasen.
Ebenso erhöhte sich die Anzahl der Liegephasen pro Lichttag mit steigender Anzahl
der Störungen, ein signifikanter Unterschied (Mann-Whitney-U-Test) bestand aber nur
in der zWeiten Herbsthälfte (Tab. 5). Zwischen Früh- und Spätherbst änderte sich an
ungestörten Tagen die Anzahl der Liegephasen nicht signifikant.
oqJi)
Saison/
Tier-Nr.
ohne Störversuche
n i s rnin-rrtur
wenige Störversuche
n t s min-rrtar
viele Störversuche
n r s min-mar
Frühh./1
l2
2L 86.3 64.5 4*276
35 107.6 114.1 6-408
33 70.t 50.5 3-202
lt 732.7 12L.5 10-415
69 65.3 47.0 4-207
53 85.4 88.4 3-367
Späth./1
l2l3
25 78.0 40.8 5-151
31 86.5 51.6 6-767
14 56.1 37.2 15-139
19 51.3 39.1 7-108
46 42.7 34.5 5-138
15 39.1 38.6 4-110
63 42.5 38.7 1*155
ß4 $.4 26.7 2-108
63 31.0 26.6 3-91
qAö+ 3 ERGEBAIISSE
Tabelle 5: Anzahl der Liegephasen bei Rehen an Lichttagen ohne, mit wenigen (2-a)
und mit vielen (5-10) Störversuchen. (n : Arzahl der Liegephasenl r, : arithm.
Mittelwertl s : Standardabweichung; min-mar : Extremwerte)
Saison/
Tier-Nr.
ohne Störversuche
n r s min-mac
wenige Störversuche
n z .s tnin-mar
viele Störversuche
n i s min-mar
Frühh./1/,
7 6.0 2.8 3-9
9 7.6 3.4 3-18
4 8.3 1.3 7-70
26.0 0 6
6-1 1
5-1 1
7 8.0 2.0
7 7.6 2.0
Späth./1
l2l3
6 4.5 0.8 4-6
6 5.3 2.0 2-8
3 4.7 t.2 4-6
3 6.3 1.5 5-8
7 7.:3 2.2 5-11
2 7.5 0.7 7-8
7 8.3 2.4 6-72
11 9.6 3.4 6-18
8 7.4 2.1 6-10
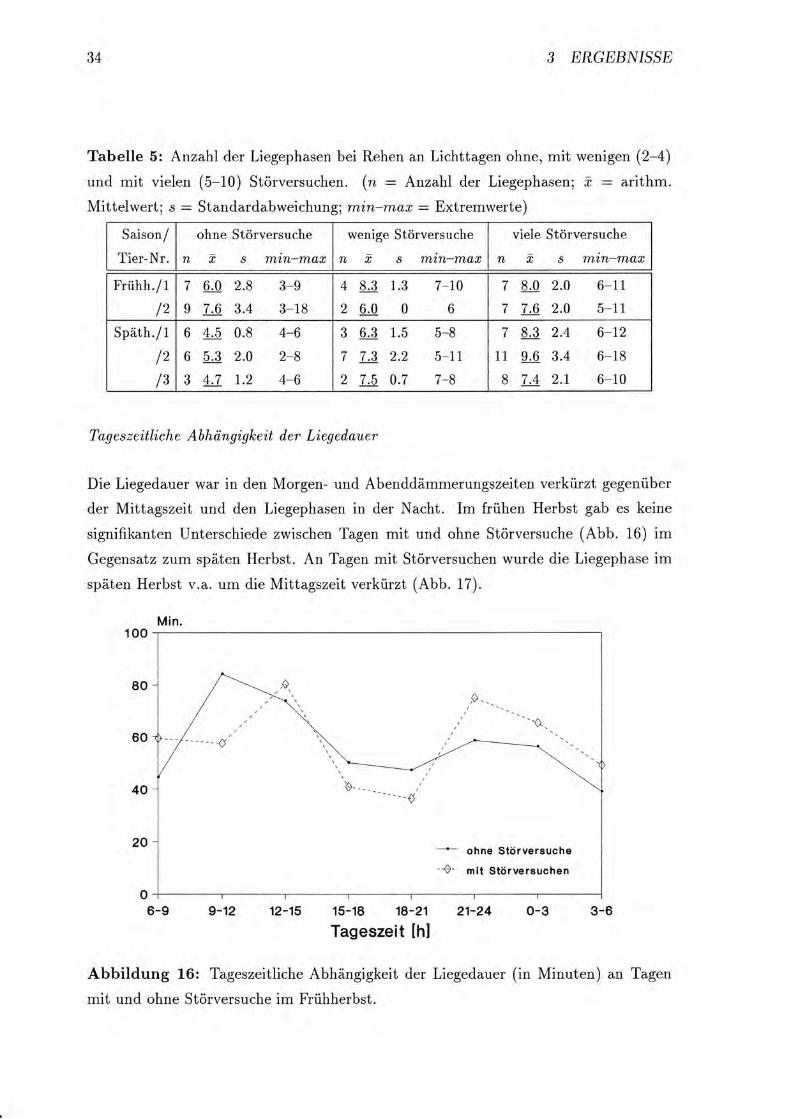
Tageszeitliche Abhtingigkeit der Liegedauer
Die Liegedauer war in clen Morgen- und Abenddämmerungszeiten verkürzt gegenüber
der Mittagszeit und den Liegephasen in der Nacht. Im frühen Herbst gab es keine
signifikanten Unterschiede zwischen Tagen mit und ohne Störversuche (Abb" 16) im
Gegensatz zum späten Herbst. An Tagen mit Störversuchen wurde die Liegephase im
späten Herbst v.a. um die Mittagszeit verkürzt (Abb. 17).
15-18 18-21 21-24
Tageszeit lhl
Abbildung 16: Tageszeitliche Abhängigkeit der Liegedauer (in Minuten) an Tagen
mit und ohne Störversuche im Frühherbst.
Min.
§
------s
-.....- ohne Störversuche-$- mit Störversuchen
o-F6-9

3.1 Ergebnisse für das Rehwild
100
80
60
40
20
15-18 18-21
Tageszeit lhl
Abbildung 17: Tageszeitliche Abhängigkeit der Liegedauer (in Minuten) an Tagen
mit und ohne Störversuche im Spätherbst.
Aktivierbarkeit durch Störreize
Als ,,Aktivierung" werden jene Fälle bezeichnet, bei denen liegende Tiere infolge eines
Störreizes ihre Liegephasen beendeten und in ein aktives Verhalten übergingen" Als
,,Aktivierungsdauer" wurde die Länge der Aktivphase eines durch einen Störreiz akti-
vierten Tieres ab dern Störversuchsencle bis zum Wiederniederlegen bezeichnet. Auch
in Versuchen, bei denen keine Videoaufnahmen vorhanden warerl) war die Aktivierung
sehr gut aus der Offline-Datei ersichtlich. Störversuche, bei denen die Tiere notwen-
digerweise flüchten mußten (wenn z.B" ein Tier auf der vorgegebenen Route für den
Rundgang gelegen ist, v.a. im späten Herbst der Fa11), blieben bei der Analyse der
Aktivierbarkeit unberücksi chtigt.
Die Assoziation der Aktivierung der Rehe untereinander war eher gering im frühen
Herbst (Assoziationskoeffizient nach Cole : +0.38 + 0"11 zwischen Reh Nr. 1 und 2).
Das gleiche gilt für den späten Herbst, hier lag der Assoziationskoeffizient zwischen
+0.22 + 0.15 (Reh Nr. 2 und 3) und +0.56 t 0.18 (Reh Nr. 1 und 3).
Abhringigkeit der Aktiaierbat*ei,t uon Tages- und Jahreszeit
Sowohl im frühen als auch im späten Herbst war die Aktivierbarkeit signifikant von
35
Min.

36 3 ERGEBNISSE
der Tageszeit abhängig (X2-Test, p 10.05), und zwar waren die Rehe insbesondere
am Vormittag und am Abend leichter aktivierbar als um die Mittagszeit, dies ließ
sich jedoch nicht statistisch absichern (Abb. 18). Die Rehe Nr. 1 und 2 wurden im
späten Herbst durch Störreize signifikant häufiger (55.4%) als imfrühen Herbst (27 "3%)
aktiviert (X2-Test, p < 0.001)"
100
Fruhherbst
--E- Spätherbst
8-10 1A-12 i2 14 141A t6-r8Tageszeit lhl
Abbildung 18: Tageszeitliche Abhängigkeit der Aktivierbarkeit im frühen und im
späten Herbst. Angegeben ist die relative Häufigkeit der Aktivierung bei 2 Rehen im
frühen Herbst und 3 Rehen im späten Herbst.
Abhringigkeit der Aktiuierung uorn Störreiztyp
Im frühen Herbst bestand wohl ein llnterschied in der Aktivierbarkeit zwischen opti-
sclren (31.4%) und akustischen Störreizen (18.8%), doch ist diese scheinbar beachtliche
Differenz statistisch nicht gesichert. Im späten Herbst waren die Rehe jedoch durch op-
tische Reize signifikant häufiger (68.0%) aktivierbar als durch akustische Reize (2\.4%).
Außerdem war die Aktivierbarkeit durch optische Störreize im späten Herbst höher als
im frülren Herbst, während bei akustischen Störreizen in diesem Zeitrarrt kein signifi-
kanter {Jnterschied feststellbar war (X2-Test).
Sowohl irn frühen als auch im späten Herbst hatten die einzelnen Störreiztypen eine
unterschiedliche Aktivierbarkeit zur Folge gehabt (x2-Test). Generell war die Aktivier-
80
60
4a
2a
%

3-1 Ergebnisse für das Rehwild
barkeit jeweils bei denselben Störversuchstypen imHerbst (Tab. 6).
späten Herbst höher als im frühen
Tabelle 6: Aktivierung in Abhängigkeit von Störversuchstyp und Saison. In Klam-
mern ist die Stichprobenzahl angegeben.
*entsprechende Versuche wurden nicht durchgeführt
Dauer der Aktiai,erung
Weder im frühen noch im späten Herbst waren individuelle Unterschiede in der Akti-
vierungsdauer zwischen den Rehen vorhanden (Kruskal-Wallis-Test).
Im frühen Herbst war die Aktivierungsdauer im Durchschnitt 2.7 Minuten (med:1 min) lang, im späten Herbst rund 26 Minuten (med: 16 min). Der Unterschied war
hoch signifikant (Kruskal-Wallis-Test, p < 0.0001).
Die Aktivierungsdauer war unabhängig von der Dauer der unmittelbar vorangegange-
nen Liegephase^ Es war auch kein Zusammenhang zwischen der Art des Störreizes und
der Dauer des Aktivierungsschubes zu erkennen.
Jedoch bestand eine tageszeitliche Abhängigkeit der Aktivierungsdauer im Spätherbst
(Abb. 1e).
Die Aktivschübe im Anschluß an eine erfolgte Aktivierung waren bei den drei Re-
hen im späten Herbst, insbesondere am frühen Vormittag und am späten Nachmittag,
verlängert und um die Mittagszeit verkürzt. Dies 1ieß sich jedoch statistisch nicht
absichern, wahrscheinlich aufgrund der zu geringen Stichprobenzahl" Die durch die
Futtervorlage ausgelöste Aktivierung dürfte für den Anstieg der durchschnittlichen
Aktivierungsdauer in der Zeit von 14-16 Uhr nicht verantwortlich sein. da die Akti-
vierungsdauer bei Futtervorlage und die Aktivierungsdauer bei anderen Störreizen in
t)l
Störtyp (n) Frühh. Späth. Störtyp (rz) Frühh. Späth.
Einzelperson (21/6) 79.0% 66.7%
Person mit Hund (11/8) 54.5% 100.0%
Personengruppe (l4l-) 92.9% -*Fferderitt (-/6) -+ 66J%
Flugdraclien (-/13) -* 76.9%
Traktor (5/10) 20.0% 40.0%
Kanzelaufstieg (1215) 0.0% 40.0%
Futtervorlage (23128) 13.0% 71.4%
Radiomusik (9/11) 22.2% 9.1.%
Hundegebell (21lrr) 0.0% 0.0%
Motorsägen (11/9) 9J% 0.0%
Schuß (17110) 23.5% 80.0%

38 3 ERGEBNISSE
dieser Tageszeitklasse sich nicht signifikant unterschieden haben (Mann-Whitney-U-
Test).
Im frühen Herbst betrug die Aktivierungsdauer jeweils nur einige Minuten, es bestand
hier keine tageszeitliche Abhängigkeit.
t=29 I x=18 1 t=20 0 t=30 1 i=32 6n=21 n-24 n=23 n=33 n=14
kt ivierungsdauer(in Minuten)
fI sehr iang (>OO)
ffi lans (3r-60)
VV7T/lt mttter (r1-30)
I kurz (t 10)
B-10 10-12 12-14 14*16 t6 18
Tageszeit Ih]
Abbildung 19: Tageszeitliche Unterschiede der Aktivierungsdauer bei 3 Rehen imspäten Herbst. Über den Säulen sind <lie Anzahl der Stichproben und die mittlere
Dauer der Aktivierung (in Minuten) für jede Tageszeitklasse angegeben.
Es wurde auch verglichen, wie sehr sich die Aktivierungsdauer zweier gleichzeitig akti-vierter Rehe unterschied, um Rückschlüsse auf eine Verhaltenssynchronis ation zt zie-
hen. Es zeigte sich eine relativ hohe Differenz zwischen der Aktiviemngsdauer jeweils
zweier Rehe (Tab. 7).
Tabelle 7: Differenz in der Aktivierungsdauer (in Minuten) zwischen je zwei Tieren.
Tier-Nr. nmedismin-marrl2113
213
28 5 17.2 26.4 0-87
17 t4 25.8 29"9 1-89
2t 15 33.0 40.0 1-162

3.1 Ergebnisse für das Rehwtld
Meist hat sich der Rehbock (Nr. 2) als erster wieder niedergelegt, und zwar in 68% der
Fälle vor Reh 1 und in 76,2% cler Fälle vor Reh 3. Reh 3 hat sich zumeist als letztes
niedergetan, nur in29,4% der Fälle legte es sich früher als das Reh 1 nieder"
Äbhcirugigkeit der Aktiuierungsdauer aom Störreiztyp
Beim Rehwild waren keine signifikanten Unterschiede in der Länge der Aktivierungs-
dauer bei verschiedenen Störreiztypen erkennbar. Generell bestanden auch zwischen
optischen und akustischen Störreizen keine Unterschiede in der Aktivierungsdauer.
3.1.3 Verhaltensanalyse
Das Verhaltensmuster der Rehe im Anschluß an Störreize wurde minutenweise aufge-
gliedert und rnit dem Verhalten vor dem Setzen der Störreize verglichen. Llngestörtes
(norrnales) Verhalten nach dem Setzen der Störreize lag wieder ab jenem Zeitpunkt
(Minute) vor. in der sich das Verhaltensmuster nicht mehr von demjenigen vor der
Störreizsetzung unterschied. Dabei wurde der relative Zeitanteil jeder einzelnen Ver-
haltenskategorie für jede Minute nach Störreizende berechnet und den entsprechenden
Werten des ungestörten Verhaltens gegenübergestellt. Die Berechnung der Verhal-
tensanteile erfolgte auf Sekunclenbasis. 250 Störversuche wurden ausgewertet. Für die
Phase vor Störreizsetntng gingen 1065 Minuten Beobachtungsdauer in die Bewertung
ein, für den Zeitraum nach den Störversuchen 1 142 Minuten"
39

40 3 ERGEBATISSE
Im Frühherbst änderte sich das Verhalten der Rehe durch die Störreizsetzung nur
geringfügig (Abb. 20).
a.)
100%
75%
50%
25%
o%
I Liegen
N Nahrungsaufn.
ITITTTTI stehen
LiIIII Sichern
ffi Pe;1lsvysgung
[--l Sonstiges
vw12345678910
Abbildung 20: Verhaltensmuster (relative Zeitanteile) der Rehe Nr. 1 u. 2 im frühen
Herbst vor (V), während (W) und in den einzeinen Minuten nach (1,2,3,. .. ) a.) opti-
schen Störversuchen (insgesamt 504 Minuten Beobachtungsdauer) und b.) akustischen
Störversuchen (insgesamt 3 1 5 Minuten Beobachtungsdauer).
b.)
100%
75%
50%
25%
o%

3.1 Ergebnisse für das Rehwild
Im späten Herbst war das Verhalten der Rehe nach optischen Störversuchen stärker
und länger beeinflußt als im frühen Herbst (Abb. 21a). Sie lagen im Anschluß an
Störreize weniger und der Zeitaufwand für das Sichern war erhöht. Nach akustischen
Versuchen zeigten die Rehe dagegen ein nur gering gestörtes Verhalten (Abb. 21b).
a.)
100%
75%
50%
25%
o%
47
I Liegen
Nl Nahrungsaufn.
[[TlITIl srehen
lIL.l ,l slchern
ffi Fortbewegung
[--l Sonstiges
75%
50%
25%
0%
Abbildung 21: Verhaltensmuster (relative Zeitanteile) der Rehe Nr. 1 bis 3 im späten
Herbst vor (V), während (W) und in den einzelnen Minuten nach (1,2,3,...) u.) op-
tischen Störversuchen (insgesami 1275 Minuten Beobachtungsdauer) und b.) akusti-
schen Störversuchen (insgesamt 317 Minuten Beobachtungsdauer).

42 3 ERGEBNISSE
Um die Dauer der Störung an einern Tier, das ,,aktiviert" wurde, sich aber nicht wie-
der hinlegte, zl bestimmen, wurde das Verhalten der durch die Störreizsetzrng aktiv
gewordenen Tiere mit dern Verhaltensmuster aktiver Tiere, die jedoch keinem Störreiz
ausgesetzt waren, verglichen. Im Frühherbst reagierten die Rehe nach Störreizen zr-
meist mit Bewegungsaktivität und Sichern, so sie sich nicht wieder hingelegt haben
(Abb. 22).
1O0o/o
7 5o/o
50%
25%
o%
N Nahrungsaufn.
[TlTllTl srehen
llifl] sichern
ffi Fortbewegung
l-_l Sonstiges
Abbildung 222 Verhaltensmuster (relative Zeitanteile) aktiver Rehe vor (V), während
(W) und in den einzelnen Minuten nach (1,2,3,. . . ) optischen und akustischen Störver-
suchen im frühen Herbst (insgesamt 101 Minuten Beobachtungsdauer).
ryI:]l
]
I
*J
ruru

3.1 Ergebnisse für das Rehwild
Im Spätherbst ästen die Rehe nach optischen Störversuchen nach 7 Minuten analog
wie ein ungestörtes Reh (Abb. 23a). Nach akustischen Störreizen war bereits in der 5.
Minute nach dem Störreiz die Asungsaktivität wieder auf dem normalen Niveau. Der
Zeitaufwand für das Sichern war jedoch noch deutlich erhöht (Abb. 23b).
a.)
100%
75%
50%
25%
o%
43
10 N\\\\
mm
E
E
Nahrungsaufn.
Stehen
Sichern
Fortbewegung
Sonstiges
500Ä
25o/o
Abbildung 23: Verhaltensmuster (relative Zeitanteile) aktiver Rehe im Spätherbst
vor (V), während (W) und in den einzelnen Minuten nach (1,2,3,. ..) ".) opti-
schen Störversuchen (insgesamt 906 Minuten Beobachtungsdauer) und b.) akustischen
Störversuchen (insgesamt 49 Minuten Beobachtungsdauer).
2345678

44 3 ERGEBNISSE
3.L.4 Herzfrequenzanalyse
Einfluß der Jahreszeit auf die lferzfrequenz
Die Tagesherzfrequenz entspricht dem arithmetischen Mittel aller Herzfrequenzwerte
über 24 Stunden. Sie war im Oktober und November bei beiden Rehgeißen etwa
gleichbleibend. Beim Rehbock lag sie generell auf einem höheren Niveau (Abb. 24).
Tler Nr.2
Abbildung 24: trinfluß der Jahreszeit auf die Herzfrequenz. Tagesherzfrequenz (jede
auswertbare Minute gernittelt). Tage mit Störversuchen sind in der Grafik grau schat-
tiert.
E 1zoEog 110
s(,o 1oo
Nz5eooITJ
tr80NE,uJ-70
Tler Nr.3
Tier Nr.l

3.1 Ergebnisse tür das Rehwild
Tageszeitlicher Verlauf der Herzfrequenz
Die mittlere Herzfrequenz zeigte im Verlauf des Tages regelmäßige Unterschiede. Mor-
gens war sie am höchsten und am Nachmittag am tiefsten (Abb. 25). Wurden Störver-
suche durchgeführt, änderte sich die Herzfrequenzkurve bezogen auf den Tagesverlauf
im Vergleich nt Tagen ohne Störversuche nicht (Abb. 26).
Abbildun g 252 Tageszeitlicher Verlauf der Herzfrequenz bei Reh Nr. 1 an Tagen
ohne Störversuche im Zeitraurn vorn 8.10. bis 25.10.90. Dazu wurden die Werte von
zehn Tagen gemittelt. Zur Glättung der Kurve sind die Herzfrequenzen von jeweils
fünf aufeinanderfolgenden Minuten gemittelt" Grau schattierte Flächen entsprechen
der Nacht.
Abbildung 26: Tageszeitlicher Verlauf der Herzfrequenz bei Reh Nr. 1 an Tagen mit
Störversuchen im Zeitraum vom 8.10. bis 25.10.90.
45
110
too
co
80
70
co
50
tß
EEoED
Eo9,N-IJJ
=oIJIEltNElrJ
110
too
eo
ao
70
co
50
ß
EEooil(,g.
N-UI
oUJcltNEIU

46 3 ERGEBNISSE
Ruheherzfrequenz
Für jede Liegephase wurde das arithmetische Mittel aus den Minutenherzfreqrenz-
werten gebildet. In der Berechnung für das Rehwild wurden 7262 Liegephasen mitden entsprechenden Ruheherzfrequenzen berücksichtigt. Die statistische Überprüfung
der Herzfrequenzen erfolgte mittels Kruskal-Wallis-Test. Zwischen den einzelnen In-
dividuen bestanden signifikante Unterschiede in der Liegeherzfreqrenz. Sie war am
höchsten beim Rehbock (Nr. 2) mit im Mittel 100.8 + 18.8 Schl./min, während sie bei
den Rehgeißen zwischen 71.0 * 6.4 Schl./min (Reh Nr. 1) und 80.3 + 5.0 Schl./min
(Reh Nr. 3) lag (Abtr. 27).
Hf n=115 n=134 n '138 't 128 n.122150
140
130
120
110
100
90
80
70
60
50Reh 1 Reh 2
FrühherbstReh 1 Reh 2 Reh 3
Spätherbst
Abbildung2T: Arithm. Mittelwert, Standardabweichung und Extremwerte der Ruhe-
herzfrequenz (Schläge/min) bei den verschiedenen Individuen im Früh- und Spätherbst.
Abhringigkeit der Ruheherzfrequenz aon iler Tages- und Jahreszeit
Während der Liegephasen war die Herzfrequenzim Frühherbst signifikant von der Ta-
geszeit abhängig" Nachts war sie erhöht und um die Mittagszeit und am frühen Nach-
mittag am niedrigsten (Abb. 28). Im Spätherbst waren die tageszeitlichen Schwankun-
gen nicht signifikant (Abb. 29). Die Ruheherzfrequenz war im frühen Herbst tagsüber
signifikant niedriger als im späten Herbst. Weder im Früh- noch im Spätherbst konnten
zwischen Tageh mit und ohne Störversuche signifikante Unterschiede bei der Herzfre-
quenz ruhender Tieren festgestellt werden.
II T+TTtT
1
I+!

3.1 Ergebnisse {ür das Rehwild
100
Hf
12-15 15-18 18-21 21-24 0-3 3-6Tageszeit [hl
Abbildung 28: Abhängigkeit der Ruheherzfrequenz (Schläge/min) von der Tageszeit
an Tagen mit und ohne Störversuche im Frühherbst. Dazu wurde der Tag in 8 Zeitab-
schnitte unterteilt und die durchschnittlichen Herzfrequenzen des jeweiligen Zeitraumes
wurden gemittelt.
47
110
90
80
70
60 +-6-9
9-12 12-15 15-18 18-21
Tageszeit [hl
Abbitdun g 29t Abhängigkeit der Ruheherzfrequenz (Schläge/min) von der Tageszeit
an Tagen mit und ohne Störversuche im Spätherbst.
21-24 0-3
100
Hf
* ohne Störversuche-$- mlt Störversuchen
110
90
70
60 l-6-9

48 3 ERGEBI\IISSE
Abhcingigkeit der Ruheherzfrequenz uon der Anzahl der Störuersuche am Tag
Zwischen der Anzahl der Störungen am Tag und der Ruheherzfrequenz bestand beim
Rehwild kein Zusammenhang (Kruskal-Wallis-Test).
Abhringigkeit der Ru,heherzfrequenz uon der Umgebungstemperatur
Bei Berücksicirtigung aller Daten gab es einen signifikanten? wenn auch nur geringen,
negativen Zusammenhang zwischen Umgebungstemperatur und Ruheherzfrequenz. Da
die Außentemperatur mit der Tageszeit eng korreliert ist und die Herzfrequenz starke
tageszeitliche Schwankungen zeigte, wurde nochmals die Abhängigkeit zwischen Ru-
heherzfrequenz und Temperatur überprüft, in diesem Falle allerdings nur für eine be-
schränkte Tageszeit (12-18 tlhr), in der keine tlnterschiede der Herzfrequenz festge-
stellt werden konnten. Wiederum ergab sich eine signifikante, wenngleich nur geringe
negative Korrelation zur Umgebungstemperatur. Wurden diese Daten jedoch nochmals
getrennt für den Früh- und Spätherbst betrachtet, war im Frühherbst nur ein Zusarn-
menhang für Reh Nr. 1 vorhanden (geringgradige positive Korrelation, r : +0.42),
während er für den Spätherbst sowohl für Reh Nr. 1 als auch Reh 2 (geringgradige
negative Korrelatiorl, ?': -0.27), jedoch nicht für Reh Nr.3 nachgewiesen werden
konnte.
Abhöngigkeit der Ruheherzfrequenz aon der Liegedauer
Es bestand ein sehr geringer negativer Zusammenhang zwischen Ruheherzfrequenz und
Liegedauer (r : -0.14), der aber statistisch signifikant war.

3.1 Ergebnisse für das Rehwild
Erhöhung der Herzfrequenz bzw. zusätzliche Herzschläge bei Störungen
Im allgemeinen zeigten die während der Störversuche liegengebliebenen Rehe geringere
Herzfrequenzreaktionen als aktive Rehe. Die Intensität der Reaktion auf eine Störung
nahm jedoch im Spätherbst insbesondere bei den liegenden Tieren zu (Tab. 8 u. 9).
Tabelle 8: Durchschnittliche Herzfrequenzreaktionen nach Störreizen bei liegenden
und aktiven Tieren im Frühherbst (2 Rehe).
VersuchsartZusätzl.
Heru-schläge
Dauer derHerzfrequenz-
erhöhung
Herzfrequenz-erhöhung
(Schläee/min)n
liegende Tiere
aktive Tiere
50
181
265 sek
378 sek
11
29
70
59
Tabelle 9: Durchschnittliche Herzfrequenzreaktionen nach Störreizen bei liegenden
und aktiven Tieren im Spätherbst (3 Rehe).
VersuchsartZwät21.
Herz-schläge
Dauer derHerzfrequenz-
erhöhung
Herzfrequenz-erhöhung
(Schlägelmin)n
liegende Tiere
aktive Tiere
137
316
263 sek
680 sek
31
28
o
95
Abhringigkeit der Herzfrequenzreaktioru uon der Art des Störreizes
Im frühen Herbst wurden die stärksten Herzfrequenzreaktionen durch Schüsse aus-
gelöst, hier kam es beim Rehbock (Nr. 2) zu durchschnittlich 281 zrsätzlichen Heru-
schlägen" Andere akustische Störreize hatten, ebenso wie die Futtervorlage, einen
relativ geringen Einfluß auf die Herzfrequenz, während Personenrundgänge eine Mit-
telstellung eingenommen haben (Tab. 10).
hn Spätherbst war die Herzfrequenzreaktion stärker ausgeprägt als im Frühherbst.
Der Rehbock (Nr. 2) zeigte nach einem Schuß wiederum hohe Werte, während das Reh
Nr. 3 kaum eine Erhöhung der Herzfrequenz zeigte. Personenrundgänge und Pferderitte
lösten in diesem Zeitraum die stärkste Herzfrequenzreaktion aus mit durchschnittlich
384-446 zusätzlichen Herzschlägen (Tab. 11).
49

3 ERGEBAIISSE50
o;.i6lo
lia)gan
()-tatiC)
lio)
Ao
atiq)
t{3c)€aq)
J-r6\
od,lio
HotHoۤotrN(,)
q)L(+iNt{(,
H
o
.lJ
0
'-r)
OOr-l oJOJO
3*(E(atr{ 'ä
O)
Fl
<iÖT
I
ol
o6ü
qa
-tatr0)
r-i
a)ti
d
crt
oFA
0i,"zl+rfr]E-r
rnH
OaHIJ.I
r-l+C)aEr
ölöl
rnHH(J
alJitrl
fr]H
C)aFa
.<FFl I-
t^N-HbD;u-
H,r\
ö# EoH 'V .Jk-.!v
S!-Hr\-ovuH CI)t+r
@N f-
INb E s^
-iHuY-r*
b E# ätH.vi(tsitliA §89
ov
<.t
c00lt'- IO
C\l öl
-jo)N .r. b0.J ! 'rvmvd
=iJl uNa
F<@t-t
t-IA
F-i@ÖI
tr
Ua1-1
F4zfrlFi
Lr'-{eö t--'l
ölr--l
F{
t-l
I/^NHE 9.EiH* 5 -::-
E€ b(+i - .5v§!tU OE
HA
!-lÖl I- I.- +
Nb E p^'o
=.E*!u;rlU
---HdN-1 .da b 6-
H-
f-Fico Ci
<.1f-!-lön
___iN-\bn
., l.i .rvav4rH*,J+iU
NaF-l
T\0l
ri-+
cö.+r"{
!v)
ati(.)
obo
bo
tr-lc)d
alro
l+r
bc
troFro
h
obo
:«3aO
q)rFl
a
z-!
Qa
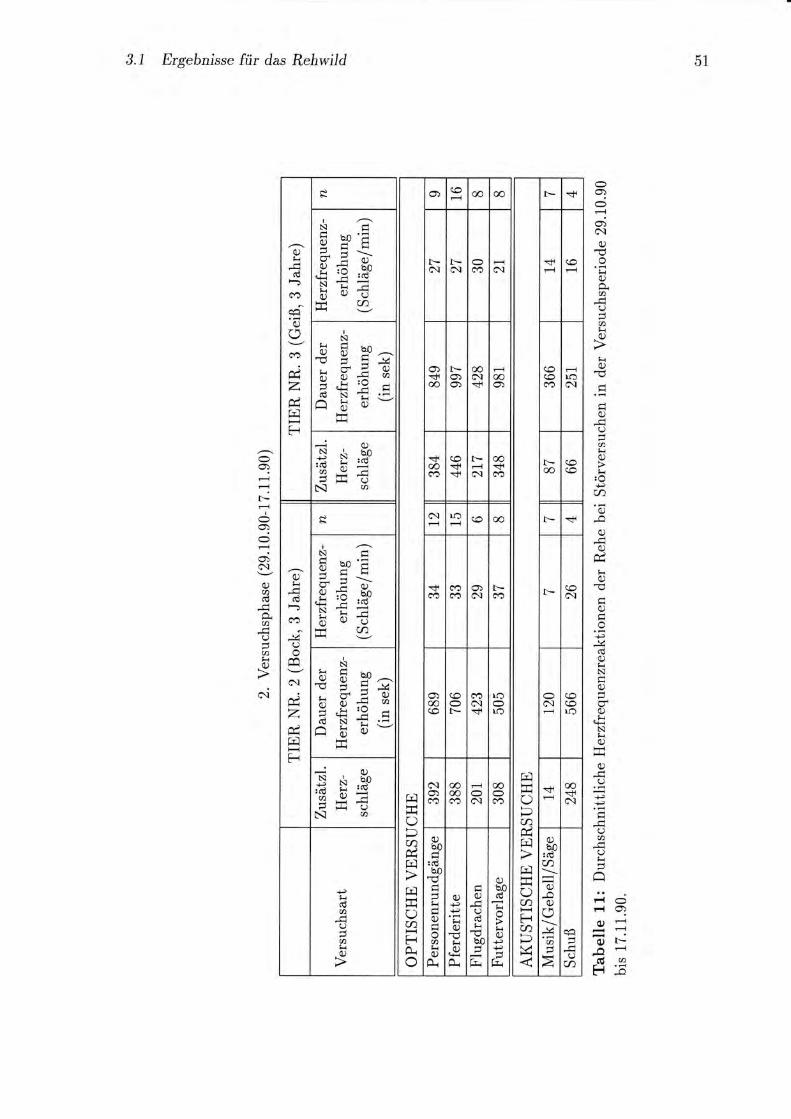
513.1 Ergebnisse für das Rehwild
O!-i
^löla)
"d
lioa
U)ti.q)
tio
no
-tafiq.)
li
a0-)
q.)
C)
fia)
a.)
q.)l-iN
O-a
(.)trNfiq)
H
c,
U
€4
a-,i
e
oJ-oI._(ga3E
.j!--l
I-r-i
I
-l0]a)aäga
aIO
Öl
q)tr6Ot-
cö
caiorrl
F4zFdrnE<
E,lHHOaH
frlH
Qa3Iric
@ @
r.fg#H(-)
aür--l
?frlHHOaE-la
4
t- +t,^NiF bo'=Va- H
cr= aiq)= boH IV
"J*-'rvL\LdLli*:i U
H C')ui
t\c-n
t-öl ea
r--'lC\l
+Fl r-
I
NLn-A, U :JJ^-<!J-?--'i,H;-d äUY.^!(+-licd N il .i
L7vl-''l oJ uH
<j@
t: @öl<r
@(o r--'l
Ic)ö]
.-j cJs §.9.l L '(vAv.-rH-;J*TU
No
<.r@
I++t-t-l0l
@<r t-
@
otrd
F-)
C];+rzlrrrI]Fi
ö]r-l
r(l @ t\ +r
t^!itr|^. bo ;dHH1a"\u# 8,H 'V .J(ri
- ..v
Nt=b ötHU)
.$öl
f.- t- (oöl
I
N
b E s^fr r H u- 15-ib u* ä!rv,qitH(dN-1 :-o b 6-
H
@(ot- N
$LO
IOö]Fl Io
__: ,l§,\.F! ..v!.v ;i -av-rH-
-FrrUNa
CNO)
@@
Fi
öl@ <i
r-i@.<f0i
€tiaOFl
atiOJ
,2
()bo
bo
fili0)tioalrq)
tu
o€€tiq)
9r.o0r
oU
trho
Obo
troti
€tr
ubo
a
U
U
a
z-1
(.)a

52 3 ERGEBAIISSE
3.2 Ergebnisse für das Rotwild
3.2.1 Wahl des Liegeplatzes
Beim Rotwild wurde keine besondere Bevorzugung bestimmter Liegeplätze festgestellt
(Abb. 30). Im Gegensatz zu den Rehen lagen sie oft im höhergelegenen Teil des Ge-
heges, der auch näher den durch Menschen gesetzten Störquellen lag. Am häufigsten
lagen sie unter einem Holunderbusch und auf einem kleinen, mit Gras bewachsenen
Hügel. Im Gegens atz za den Rehen lagen sie nie im Grasfeld.
Abbildung 30: Prozentuelle Verteilung der
Versuchszeit (n. : 173 Liegeplätze).
Wechsel d,es Li,egeplatzes nach Störuer.suchen
Liegeplätze beim Rotwiid während der
In 53,60/o der Fälle suchten Tiere, die durch Störversuche aktiviert wurden, einen an-
deren Liegeplatz auf als vor dem Versuch.
Auffallend war nur! daß an bestirnmten Tagen ein bevorzugter Liegeplatz, auch nach
Störungen, für den Rest dieses Tages immer wieder aufgesuclit wurde, am darauffol-
genden Tag wurde der Liegeplatz jedoch an einer ganz anderen Stelle gewählt (Abb" 31
und 32).
Il'*""uFichten
c)oo*,

3.2 Ergebnisse für das Rotwild
Abbildung 31: Liegeplätze beim
diesen Tagen jeweils mindestens 4
Rotwild zwischen
Störversuche statt.
53
.10.1.91+ll.l.9lo l5.l.9l* 16.1.91
10.01.91-12.01.91. Es fanden
.13.2.91+ 14.2.9ro 15.2.91* 26.2.91
Abbildung 32: Liegeplätze beim Rotwild zwischen 13.02.91-26.02.91. Es fanden an
diesen Tagen jeweils mindestens 4 Störversuche statt.
3.2.2 Aktivitätsmuster
Für das Rotwild wurden etwa 311000 Minuten aus der Offline ausgewertet. Die Akto-gramme beider Tiere zeigten einen über die gesamte Versuchsperiode einheitlichen Ver-
lauf. Während der Dämmerungszeiten waren die Tiere, gleich wie die Rehe, vermehrt
aktiv. Diese Periode der verstärkten Aktivität verschob sich mit der zunehmenden
Tageslänge entsprechend den Sonnenauf- und -untergängen (Abb" 33 und 34).

54 J ERGEBNISSE
22-12t l rIII24-12-
-
ii::iiJ:ii:::iu:;tii,:t:;:26 -12iääüä!-.1!ir,:'::24.'12- ,r r
O3.Ol-Ir::::::- I ,: aI
-
o5.o1- r r I'::i'- r,l:'ll,l
07.O1:,
-
:,: l::r':I:Il:l::::::ri' -::,'''r''''
IO9.O1.- , r,
-,':,:::
;iil;riliiii,i11-O1-- iI ,,i, ,trii
;:l::I::l'':''I':'i:r::raltl;illiliiüttitil!u:l!
II II'T-.,.r,::::::l',-:.,1::::: .jll
-
I ::": :"'':I
l::::I I30.12r
-
:I
-1,:rI
I I II::j"-:::, I
- I ,irl :,r,I
o1.o1.:::!
I:,,.
-. Iir:::::ilru':II ,,,..:l I I.:.,,', I I::.', ,rI r::,::I
t-
-'',::- -
IIII
-
-r::::::
:.,,', I I::.., ,rI l:iI::i.:;-:: I::::iIi::: I:r I I::::.: I:
r::t:::!:t:t-i::ataia:::
:::::lIl:::l::l:::::rl I
-Il-:t'I:::t' I ::u:':Il:-
-
rII IEII:J II.
13.O1I I II :i'i:a::::i:ll:::al-:::
15.O 1 .Jiriiiil- i , ::l::::::::::::::lI
li i JriI : r Iaii il,,tiillill I I i:r,r.ri Ix
IIIIIal: I.l,l I
' ,l
-
x. il'::ll,l
-x
17.01 t l, r ;Ir I I'I 9-ol-I-::'::::'.. I!.!] fr tr- | r:::: I
-
I II I
r ..,,r Irx,l:.r':r.:t;-
::::I::':il ,-I l:rr:':l'r-'::'
2 1 .O 1 .I-::::::::..:.::::.::::.:..::..::..:.::.,::
-.::::::::::::t::::::tr:r::r:::r::I -l::: I::1
I I
-x
:lr.:.:i ',:l Ix29.()1--L:l;,,:::,:::,:,,1.,1:,.,i:: E I
-
li..Iil::lrlI'iJ:: :: :j:l, Il
- -::::::
I!:.rl25-01 ,:, lI:::::l:.:::i,. l-- I
-I
I ::r:
Fl::t::r :tl::::::::::I:.t:::::1.,1
-t;,i
I lr I I t tl27.O1J,: ...:::t:;:.::.:..:.:.:i..,,i.r.IlI r . r IIrjl
lllrlll:lrll:rl1;r::::::::::::::::::::::I:I I
-,L
.-)ii29.Ol j,Iil',::::i:::::l:r:::t::::::::r,I. I f I I.. Iuiljii!|
rr
-I
'I Il. :]]:II .: :rj::: :l xI 'l r""1 xl'r r I xt::Ji.::::"i:.1
-
x
l::r::r :ll::::::::::I:.r:::::1.27.O1 J,: . ..:::t:;::::.::.:::::i..::i.l
I,.',r:'.i,.,1
-
x| '.,: I:::l:.
I 1,,: l,':',-,r Il
"-lxIt'::'fIx:i:: .i::
-
xI r.'-. ,:,1 ::l:: x
i::::::r:::jll:.:::-:::r: lx::t:r:t:l::::'-:::r: lx:,::. .',.j .] lI:::::[
xxi-
:::il::l::l:::::l:::::::::'::- X:-,:::l::', :,;::::::::.: I X
:::::aa.: :aa::a,:: :. :a,,aa::::
- x
::::::::l-.','....,. :
-.',.',:
:,'I
-.I,::r:::,,:Ix
l.l:,:" ;:': ' I:'.,-':.'::;':".;'i:I , I'i::ir:].,:.:li.':I
!t:.:.1I. I .,:, II- I- I
l:l::iil I :i:::::':
-
,:'::-::. ,-I:i.::l:::: aI'' ,:.:::::' ':::l:..jll Il
22.02-,, I
'-1a.o3.r-
r| ::,it::I20.o3.r r::.
Il I I"--M l ::j r': I I.l
II-ITII.I-'TII
:illriiiI i:iii::i:iu,i:-I:::l:i:::rr::::::::::[:I:::IO4.O3. l-ri:i::r::,
!lliiil Iiir:r:r::r::::::::::lr:.'i:: 'I llt I Ia,:rlIi'I I,t
-,jr!
:]:::::::::::::-:::::::::l.I xIlI:::::::::::::::::::::::::::r x
-
I llll ,:i: I :ra-1().03.- r t-, :.i::::r ..r r12-O3-:,:: '
-
I l-r,:::r.., ItIf lllI'::'::,. I:,' f
-r-14.O3; i
-
l,::'::,. I:,' f f:'l ! ,l lr ::. I ::::'
:':.::r:.:::::-.::rl :-::l::':l:::.
-',
:,l:l:i i.::.li-r xI ,ir- xI ,ir- xI ..:r- x. l.:r,,r.II- , x: .l :t.,,.,:::EIx
f I :r:: j['j:j:
-
l-':ll::p:I::IlI
-::::l:
:l r:r"rl I
]I::Ilrr I1l I. I
,II,
-
II::, I
22.O3::::''::::::': I l:il:i:illilllliiii;::lI Ili::::i:1:l)llliillii I::.r-i: .l .ll,. I-I:, I.',,4..1.r....:...
-aa,r,.at:,,/!::.):26.O3--,:::: I : :t::i:r::j:r.,:,rI :I lI:.,.: r
I.:t:,::::i.-,::::i'-:::::-::':''I''''":"'' Il::I' ,II
iit:ili,1i:11*i lnaktlv I Aktiv
Abbildung 33: Aktogramm von Rothirsch Nr. 4 (Spießer). Tage, an denen Störver-
suche durchgeführt wurden, sind an der rechten Seite mit ,,x" markiert. Die schräg
nach unten verlaufenden Linien markieren Sonnenauf- und -untergang.

3.2 Ergebnisse für das Rotwild 55
'l tI l'rjrl
-,,l
i,,,iiii.ii:iil iiitirlI I aI ijj:,lrr-:l
r:;rii-i:i:IiI::::::::I,I:::::I I I i
::Il:-,r:::::::::rr',-::, l
-l:i.iiri
' : :. :aaa:aa.:.::-aa::.. ::t- .::::::-Jlli:r::i::ii,-r::;, x
-
f''.::::rr':' .,:,:- "::+::I,'::::::::l
I Irlr Il,:f I:::::fI
-I'lI r.
-l
r,'rl:'-xI iIr,r..-rri lx
t,, r
If I ::'Ii:I-..::::I
I,,i :rt II :,trfIxIrr.,:::r::.,,-
-::I.
::::rl.:ll,:.::,-,,::'..'::::.
-
::::::: I lx
:::::tu;rt:;:t:r:I:,, I I::, I:::::::,ll::ti'lj;:.II ,r .r::11 t I
l-,:::i-:::l
t:::t.:::l:r,::-:::tri l x:l:.:.:::::ri.:::::::::iil
-
x,l ff ::1:::lI''::'- l:::::':.,
E::r:r:'I': ,lr ::j:l rx
I I :i:.:t .:::::tx
:::I::::::::::I
I'.'.:,., I Ix
:'i :l::::ai .,,',::,Jrl:i, ,:II,,l
I' ::'I I '::: I
-lI
l::,rxI t. :...,: I:rra
III ': It I III -]IIIIIIIIII:']III!II
I',:l l' I I
:: :Il::: x.-,xl-lx
I,-x- r-
l:t:::t:rl a:r:::::::l II:::::ll:::r:r f lllIIr::'-'::::::r:'I::l::::1. i-
,-::.::::-
:.:::lx'l' IxIX
xx
-
il,.:I I'l
-"I
-t
a 'a:a:. :.:.: :
'I::::::-:.::'l:rt:rI::,x
I :, ,rt::: xIl I I
-tIII:I
I II' I:::::l::ll::.:
--I
-III
! . :::::::l 'l:, : ,,:l:ll.l-...a, ll:::iliI
I-llr:1.: :'ltlllrt :::::t-::t:r :::.:.El
-
:aa1
I ::l::rIf.:[:l :.:,:]:rll Il:"1
-:'! ::rl::'I
I I:::::., II ::::,4
!
-:'.':r,:I r'.::::-: ,::l:: I,;r t:!II II
iivnillllilllli lnaktiv I Aktiv
Abbildung 34: Aktogramm von Rothirsch Nr. 5 (weibl. Kalb). Tage, an denen
Störversuche durchgeführt wurden, sind ar der rechten Seite mit ,,x" markiert. Die
schräg näch unten verlaufenden Linien markieren Sonnenauf- und -untergang.
II I IIIIt t.:. r r rrl.j:: II:: l:: I II I_r t-t t I !
I I] I II
-I l,:':1 , I
I II lt-,. I l: II I..rI l ,,tI
- IIl,,tI-
Lt lrt
Ilrlr11.13.15.17.19.
25.25.27.29.31.
21 .12.
02.o104.o 1
o6.o 1
o8.o 1
1().o1'12.o1
'14.o1
16.O1
l a.ol20.o 1
22.O124.O126.O128.O'l30.o 1
o1
1-l
1
1
1
17
2527olo9o5o7
11
131517

bf) 3 ERGEBNISSE
Tageszeitlicher Verlauf der Aktivität
An ungestörten Tagen war die Aktivität besonders am Morgen und am Abend erhöht.
Ein kleiner Aktivitätsgipfel war auch am frühen Nachmittag vorhanden (Abb. 35).
Wurden Störversuche durchgeführt, erhöhte sich die Tagesaktivität. Auch in der Nacht
waren Unterschiede im Aktivitätsrhythmus zu erkennen (Abb. 36).
Abbildung 35: Tageszeitliche Abhängigkeit der Aktivität von Tier l§r. 4 an Tagen
ohne Störversuche im Zeitraum vom 28.12.90 bis 22.01.91. Zur Glättung der Kurve
sind die Aktivitätswerte über jeweils fünf aufeinanderfolgenden Minuten gemittelt und
der Durchschnitt von zehn Tagen errechnet. Grau scha,ttierte Flächen rnarkieren die
Nacht.
=E 10oN
E§oooo5coEic.E .o
Ff( 20
=FYo
=6 looN
E3ooo(J
bcoE>e
.EßFT( 20
tt-yo
Abbildung 36: Tageszeitliche Abhängigkeit
mit Störversuchen im Zeitraum vom 28.12.90
der Aktivität von Tier Nr. 4
bis 22.01.91.
an Tagen
UHRZEIT
UHRZEIT
3.2 Ergebnisse für das Rotwild
Gesamtaktivitätszeit
In den Tabellen 12 und 13 sind die prozentuellen Anteile für aktives Verhalten innerhalb
von 24 Stunden bzw. beschränkt auf den Liclittag angegeben. Dabei wird zwischen
Tagen mit und ohne Störversuche unterschieden"
Tabelle 12 %o-Anteil aktiv während des 24-stunden-Tages beim Rotwild. (n : An-
zahl der Tage; med : median; r : arithm. Mittelwert; s : Standardabweichung;
min mar: Extremwerte)
24-Stunden-Tag
Monat/
Tier-Nr.
ohne Störversuche
rr med i s min-mat
mit Störversuchen
n med J s min mar
Dez. 14
l5
18 51.6 48.7 6.4 35.0-55.6
13 48.6 48.2 6.7 32.5-61.1
2 50.6 50.6 1i"9 42.2-59.0
Jau 14
l5
13 46.4 47.2 4.0 38.6-53.6
9 50.0 49.8 4.7 42.9-56.2
18 48.5 49.4 5.9 40.243.4
11 55.9 55.1 6.6 44.445.5
Feb. lal5
14 47.9 47.t 5.0 39.6-55.3
9 53.4 52.9 3.8 46.9-57.7
14 50.2 54.7 7 "1 45.245.6
9 57.5 56.9 7.0 47.347.3
Mä,rz 14
l5
4 54.6 53.5 3.6 48.5 56.5
4 58.1 58.0 1.5 56.1-59.6
5 49.7 56.1 7.6 49.247.8,1 52.9 61.8 6"9 57.4 72.1
Tabelle 13: %-Anteil aktiv während des Lichttages beim Rotwild.
Lichttag
57
Monat/
Tier-Nr.
ohne Störversuche
nm,edrsmin-marmit Störversuchen
n med i s min mar
Dez" 14
l5
19 56.8 59.1 8.6 39.3-74.4
t4 53.3 52.1 7.4 45.2-62.8
2 72.5 72.5 2.1 77.0 74.0
I 59.8 59.8
Jan. 14
l5
12 47.t 49.1. 7.3 4L.7-64.2
9 53.1 52.1 9.5 40.3-69.2
18 61.1 62.2 9.7 48.0-84.0
15 68.4 6t.2 7.9 54.2-77.7
Feb. /4
l5
72 51.7 51.1 7.6 40.4-66.3
12 56.9 57.0 7.9 50.3-72.3
L2 62.6 60.7 11.5 43.0-78.9
8 68.5 67.5 9.8 50.6-80.5
März f 4
l5
5 55.3 49.3 4.1 54.2-42.8
5 59.0 54.5 4.3 49.8-59.7
5 58.2 56.0 5.2 50.0*60.3
4 67.7 60.3 4.5 53.9-63.1

58 3 ERGEBNISSE
Hinsichtlich der Gesamtaktivitätszeit war bei Störungsfreiheit während 24 Stunden
zwischen beiden Tieren nur im Februar und März ein signifikanter Unterschied zu er-
kennen (Mann-Whitney-Ll-Test, p ( 0.05). Die Aktivität des Rothirschkalbes (Tier
Nr" 5) zeigte bezogen auf jeweils 24 Stunden von Dezeinber bis März eine steigende
Tendenz, doch ist nur der Anstieg von Februar zum März hin signifikant (p < 0.05)"
Der Hirschspießer (Tier Nr. 4) begann hingegen erst ab März seine Aktivität zu stei-
gern (p < 0.05). Bezogen auf den Lichttag ist der Aktivitätsanteil des Rothirschkal-
bes an ungestörten Tagen während des Beobachtungszeitraumes etwa gleichbleibend.
Der Hirschspießer wies hinsichtlich seiner Aktivität bezogen auf den Lichttag zwischen
Dezember und Januar signifikante tlnterschiede auf. Ein wesentlicher lJnterschied zwi-
schen der Tagesaktivität dieser beiden Tiere ist nur im Dezember gegeben. An Tagen
mit Störversuchen bestand cler Trend, die Aktivität während 24 Stunden geringfügig zu
erhöhen. Dabei wenden die Tiere insgesamt ungefähr eine halbe Stunde weniger für Lie-
gen auf als an den störungsfreien Tagen. Die Unterschiede waren aber, mit Ausnahme
von Tier Nr. 5 im Januar, bezogen auf die Gesamttagesaktivität nicht signifikant" An
Tagen mit Störversuchen reduzierten die Tiere während cles Lichttages ihre Liegean-
teile um etwa 10% (fär beide Tiere nur im Januar signifikant; p < 0.01). Daraus folgt,
daß die Verringerung der Gesarntruhezeitbezogen auf 24 Stunden praktisch ausschließ-
lich während cles Lichttages erfolgte und in der anschließenden Dunkelphase kaum eine
Kompensation der reduzierten Liegezeit stattfand.

3.2 Ergebnisse für das Rotwild
Anzahl und Dauer der Liegephasen
Mit zunehmender Anzahl der Störversuche während des Tages nahm bei beiden Tieren
die Liegedauer am Lichttag ab, die Dauer der Liegephasen wurde jedoch im Vergleich
zu wenigen Störversuchen durch viele Störversuche nicht wesentlich reduziert (Tab. 1a).
Die Anzahl der Liegephasen am Lichttag btieb etwa gleich (Tab. 15). Die statistischen
Berechnungen erfolgten mittels Mann-Whitney U-Test.
Tabelle 14: Dauer der Liegephasen (in Minuten) beim Rotwild bezogen auf den Licht-
tag, getrennt nach Tagen ohne, mit wenigen (1-3) uncl mit vielen (5-9) Storversuchen.
(n: Dauer der Liegephasen; f : arithm. Mittelwert; s: Standardabweichung;min
mar : Extremwerte)
Tier-Nr.
ohne Störversuche
n i s min-rruu:r:
wenige Störversuche
n i s min-mar
viele Störversuche
n r s min-mar4
5
218 61.8 37.3 3-L74
208 52.6 37.0 6-200
34 42.5 29.2 7-116
33 39.5 24.0 10-95
15t 42.4 30.3 5-144
t54 37.4 24.6 2-122
Tabelle 15: Anzahl der Liegephasen bezogen auf den Lichttag im Zeitraum Jan.-
März, getrennt nach Tagen ohne, mit wenigen (1-3) und mit vielen (5-9) Störversuchen.
(n: Anzahl der Liegephasen; i: arithm. Mittelwert; s : Standardabweichung; rnin
?n0,t : Extremwerte)
Tier-Nr.
ohne Störversuche
n i s min-ntar
wenige Störversuche
n r s min-marviele Störversuche
n i s minnto,:t
4
5
30 4.9 1.6 3-8
35 5.0 1.5 2-8
B 5.0 1.1
6 4.7 1.6
4-7
3-7
27 5.4 1.8 1-8
17 5.7 2.7 3-10
Tageszei,tliche Abhringi,gkeit der Liegedauer
Die durchschnittliche Liegedauer war tagsüber etwas kürzer als in der Nacht (Abb. 37).
Durch Störversuche wurden die Liegephasen v.a. während des Lichttages verkürzt (si-
gnifikant nach Kruskal-Wallis-Test).
59

60 3 ERGEBI\IISSE
Min.
§-- - ----g
.+--"-'-"'o'
6-9 9-12 12-15 15-18 18-21 21-24 0-3 3-6
Tageszeit lhl
Abbildung 37: Tageszeitliche Abhängigkeit der durchschnittlichen Liegedauer (in
Minuten) an Tagen mit und ohne Störversuche.
Aktivierbarkeit durch Störreize
Die wechselseitige Abhängigkeit der Aktivierbarkeit der beiden Tiere war sehr hoch
(Assoziationskoeffizient nach Cole : +0.915 + 0.14).
Bei beiden Tieren war im Verlauf der gesamten Versuchsperiode (Dez.-März) keine
Änderung in der Aktivierbarkeit zu erkennen. Tendenziell war das Rothirschkalb clurch
optische und akustische Störreize etwas häufiger (76.7%) aktivierbar ais der Hirsch-
spießer (65.9%), der l.lnterschied war jedoch statistisch nicht signifikant (12-Test).
Ta g e s z e itli ch e Ab h cin q i qk e it d e r Aktiui e rb ark e it
Die tageszeitlichen llnterschiede in der Aktivierbarkeit lassen sich bei beiden Tieren
statistisch nicht absichem. Die niedrigste Aktivierbarkeit war zwischen 12 und 14 tlhrzu beobachten, danach stieg sie wieder an und fiel zum Abend hin wieder ab (Abb. 38)"
50

3.2 Ergebnisse {ür das Rotwild
- Tier 4
---f-- I ter 5
8-10 10-12 12-14 14-16 16-18
Tageszeit [hl
Abbildung 38: Tageszeitliche Abhängigkeit der Aktivierbarkeit. Angegeben ist die
relative Häufigkeit der Aktivierung zu verschiedenen Tageszeitklassen "
Abhringigkeit der Aktiuie.rung uom Störreiztyp
Die Aktivierbarkeit durch einzelne Störreize ist in Tab. 16 dargestellt. Beide Tiere wa-
ren durch optische Störreize signifikant (X2-Test, p < 0.01) häufiger aktivierbar (etwa
87Yo) als durch akustische Reize (ca.28%). Dies gilt für den von zwangsläufig erfolgten
Aktivierungen (wenn z.B. das Tier auf der vorgegebenen Route des Personenrundgan-
Tabelle 16: Aktivierung der Rothirsche durch verschiedene Störreize in Prozent (n :Anzahl der Störversuche).
Störtyp n % aktiviert Störtyp n % aktiviert
Einzelperson 27 100.0
Person mit Hund 14 100.0
Personengruppe 14 100.0
Pferderitt 25 92.0
Flugdrachen 12 33.3
Traktor 7 28.6
Feuer 10 100.0
Futtervorlage 1i 90^9
Radiomusik 8 12.5
Hundegebell 8 25.0
Motorsägen 11 9.1
Schuß 15 53.3
61
Yo

62 3 ERGEBATISSE
ges lag) bereinigten Datensatz. Bei allen optischen Reizen lag die Aktivierbarkeit zwi-
scherr 90 und I00%, außer beim Traktor (28.6%) und Drachengleitflug (33.3%). Bei
akustischen Reizen war die Aktivierbarkeit nur beirn Schuß relativ hoch (53.3%). Auf-
grund der geringen Stichprobenzahl rvar eine statistische Absicherung nicht möglich.
Aktiaierungsdauer
lrlach einem Störreiz betrug die Aktivierungsdauer im Mittel 38.5 Minuten. Indivi-
duelle Unterschiede waren nicht vorhanden (Kruskal-Wallis-Test). Auch zwischen den
einzelnen Monaten bestand kein signifikanter Unterschied. Es deutete sich aber ein
leichter Trend zur Abnahme der Aktivierungsdauer zum Spätwinter hin an.
Die tageszeitlichen Unterschiede in der Aktivierungsdauer waren signifikant (Kruskal-
Wallis-Test); und zwar war die Dauer der Aktivierung am frühen Vormittag eher gering,
stieg bis zum Mittag etwas an, fiel wieder am frühen Nachmittag und stieg zum Abend
hin sehr stark an (Abb" 39).
t=28 O t=37 6 ,.-25 5 i=34 0 t=76 5n=2ö n=4O n=26 n=31 n=15
Aktivierungs dauer(in Mlnuten)
= lang (>60)
tffig m'rte t3t-6Or
N kurz (lr-30)
I sehr kurz (l-10)
8-1C 10-12 12 14 1,4-18 16-18
Tageszelt [h]
Abbildung 39: Tageszeitliche Unterschiede in der Aktivierungsdauer beim Rotwild.
Über den Säulen sind die Anzahl der Stichproben und die mittlere Dauer der Aktivie-
rung (in Minuten) für jede Tageszeitklasse angegeben.

3.2 Ergebnisse für das Rotwild
Bei jeweils ein und demselben Störversuch, der eine Aktivierung zur Folge hatte, waren
die Unterschiede in der Aktivierungdauer zwischen beiden Tieren eher gering (z : 10.5
rrtin, med,: 5 min, 0-57 min). In gleich vielen Fällen war jeweils ein Tier länger aktivgeblieben als das andere Tier.
Abhängigkeit. der Aktiuierungsd,auer aom Störrei,ztyp
Grundsätzlich haben optische oder akustische Störreize keinen Unterschied in der Länge
der Aktivierungsdauer bewirkt. Zwischen einzelnen Störversuchstypen war die Akti-
vierungsdauer jedoch hochsignifikant unterschiedlich (Tab. 17) (Kruskal-Wallis-Test).
Tabelle 17: Aktivierungsdauer in Minuten in Abhängigkeit vom Störreiztyp (n :Stichprobenzahl, med, : Median).
Störtyp n med Störtyp n rned. Störtyp n med
Personenrundgang 24 34.5
Person mit Hund 15 51.0
Gruppenrundgang I 55.0
Einzelpferd 24 21.0
Pferdegruppe 11 12.0
Flugdrachen 4 32.5
Traktor 5 19.0
Feuer 6 35.5
Futtervorlage 9 42.0
Radiomusik 1 3.0
Hundegebell 2 18.5
Motorsägen 1 3.0
Schuß 3 68.0
3.2.3 Verhaltensanalyse
Die Verhaltenselemente nach Störversuchen wurden wie beim Rehwild minutenweise
aufgegliedert und mit der Zeit vor der Störreizsetzung verglichen. Ungestörtes Verhal-
ten war ab dem Zeitpunkt (Minute) vorhanden, in dem sich das Verhalten der Tiere
nicht mehr signifikant von jenem vor dem Störversuch unterschied. Es wurden 244
Störversuche auf Sekundenbasis ausgewertet. Für die Zeitspanne vor der Störreizset-
zung gingen 2 484 Minuten in die Bewertung ein, für den Zeitraum nach Störversuchen
1869 Minuten.
63

64 3 ERGEBNISSE
In der ersten Versuchswoche war das Rotwild durch optische Störreize mindestens 10
Minuten nach Absetzen des Störreizes in seinem Verhalten gestört (Abb. 40a), hinge-
gen waren nach akustischer Störreizsetzung bereits ab der 6. Minute keine signifikan-
ten Unterschiede im Vergleich zu den Verhaltensanteilen vor der Störreizsetzung mehr
nachweisbar (Kruskal-Wa11is-Test) (Abb. a0b).
a.)
100%
75%
50%
25%
o%
I Liegen
N Nahrungsaufn.
II3 g;s6.r.
ffi Fortbewegung
ffi oltakt. Kontr.
[.-l Sonstiges
b.)100%
75%
50%
25%
Abbildung 40: Verhaltensmuster (relative Zeitanteile) der Rothirsche in der ersten
Versuchswoche vor (V), während (W) und in den einzelnen Minuten nach (7,2,,3,. ".)a.) optischen Störversuchen (insgesamt 642 Minuten Beobachtungsdauer) und b.) aku-
stischen Störversuchen (insgesamt 4 1 0 Minuten Beobachtungs dauer).

3.2 Ergebnisse {ür das Rotwild
In der letzten Versuchswoche zeigten die Tiere nach optischen Versuchen ab der 10.
Minute wieder ungestörtes Verhalten (Abb. 1a). Nach akustischen Versuchen hatte
sich ihr Verhalten bereits in der ersten Minute nach Absetzen des Störreizes normalisiert
(Kruskal-Wallis-Test) (Abb. 41b).
a.)
100%
75%
50%
25%
o%
65
I Liegen
N Nahrungsaufn.
l7r----Ti Sichern
ffi Fortbewegung
ffi otfakt. Kontr.
l--l Sonstiges
Abbildung 41: Verhaltensmuster (relative Zeitanteile) der Rothirsche in der letzten
Versuchswoche vor (V), während (W) und in den einzelnen Minuten nach (1,2,3,. . . )a.) optischen Störversuchen (insgesamt 476 Minuten Beobachtungsdauer) und b.) aku-
stischen Störversuchen (insgesamt 172 Minuten Beobachtungsdauer).
b.)
100%
75%
50%
25%
o%

bf) 3 ERGEBNISSE
Wurde das Verhalten von nach Störversuchen aktiven Tieren mit jenem von ungestörten
aktiven Tieren verglichen, dann war in der ersten Woche, sowohl nach optischen als
auch nach akustischen Störversuchen, das Verhalten über den gesamten Zeil,;raum der
Videoaufzeichnung, d.h. mindestens 10 Minuten, gestört (Abb. 42).
a.)
l OOo/o
50%
25%
o%10N
El
ffiE
Nahrungsaufn.
S iche rn
Fortbewegung
Olfakt. Kontr.
S onstigesb.)100%
75%
50%
25%
o%
Abbildung 422 Verhaltenmuster (relative Zeitanteile) aktiver Rothirsche in der ersten
Versuchswoche vor (V), während (W) und in den einzelnen Minuten nach (1,2,3,. . . )a.) optischen Störversuchen (insgesamt 466 Minuten Beobachtungsdauer) und b.) aku-
stischen Störversuchen (insgesamt 201 Minuten Beobachtungsdauer).

3.2 Ergebnisse für das Rotwild
In der letzten Untersuchungswoche zeigten aktive Tiere nach optischen Versuchen min-
destens 8 Minuten, nach akustischen Versuchen mindestens 6 Minuten gestörtes Ver-
halten (Abb. 43). Für die Auswertung der 9. und 10. bzw. der 7. bis 10. Minute war
das Datenmaterial nicht ausreichend.
a.)
100%
75%
50%
25%
67
N Nahrungsaufn.
lllll Sichern
ffi Fortbewegung
ffi g;9s11. 146n1r.
[-l Sonstiges
b.)100%
75%
50%
25%
o%
Abbildung 43: Verhaltensmuster (relative Zeitanteile) aktiver Rothirsche in der letz-
ten Versuchswoche vor (V), während (W) und in den einzelnen Minuten nach (1,2,3,. . . )optischen Störversuchen (insgesamt 173 Minuten Beobachtungsdauer) und b.) akusti-
schen Stgrversuchen (insgesamt 66 Minuten Beobachtungsdauer).
68 3 E,RGEBNISSE
Bei der Betrachtung einzelner Störfaktoren dauerte es unterschiedlich lang, bis in allen
analysierten Verhaltenskategorien keine signifikanten Unterschiede mehr festzustellen
waren (Tab. 18). Bei Personenrundgängen und Schuß waren nach 10 Minuten (maxi-
male Beobachtungsdauer) noch signifikante Unterschiede in einzelnen Verhaltenskate-
gorien, verglichen mit dem Verhalten vor dem Setzen des Störreizes, vorhanden. Bei
den anderen akustischen Reizen, aber auch bei Flugdrachen, Traktorfahrten, Pferderit-
ten und Feuer gab es nach Beendigung des Störversuches praktisch keine Unterschiede
im Verhalten im Vergleich zu jenem vor dem Störversuch.
Tabelle 18: Zeitraum (in Minuten) nach Beendigung des Störversuches, in dem noch
signifikante Unterschiede in mindestens einer Verhaltenskategorie im Vergleich zum
Verhalten vor der Störreizsetzung bestanden.
Störversuch Liegen Fortbew. Ar"r, Sichern
10
10
i
SonstigesOlfakt. Kontr.
9
9
9
Einzelperson IPerson mit Hund IPersonengru ppe
Pferderitt
Flugdrachen -Traktor
Feuer
Musik/Gebel1/Säge
Schu ß
10
8
4
4
I
410

3.2 Ergebnisse {ür das Rotwild
3.2.4 Herzfrequenzanalyse
Einfluß der Jahreszeit auf die Herzfrequenz
Die mittlere Tagesherzfreqrenz nahm bei beiden Tieren von Dezember bis Mitte März
kontinuierlich ab, Mitte März begann sie wieder anzusteigen. Beim Rothirschspießer
war die Herzfrequenz durchwegs höher als beim Rotwildkalb (Abb" 44).
90
Tier Nr.4
Tier Nr.5
Abbildung 44: Jahreszeitlicher Verlauf der mittleren Tagesherzfrequenz beim Rot-
wild. Die mittleren Tagesherzfrequenzen wurden aus allen auswertbaren Minuten ge-
mittelt. Tage mit Störversuchen sind grau schattiert.
Tageszeitlicher Verlauf der Herzfrequenz
Die Herzfrequenz zeigte beim Rotwild tageszeitliche Unterschiede. Vor Sonnenaufgang
lag sie am tiefsten. Im Verlauf des Tages stieg sie allmählich an und erreichte den
höchsten Wert am späten Nachmittag (Abb. a5). An Tagen mit Störversuchen lag die
Herzfrequenz erheblich niedriger, vornehmlich zwischen 10 und 14 Uhr. In der Nacht
kam es dtrrch Störversuche zu keinen nennenswerten Unterschieden in der Herzfrequenz
(Abb. 46).
69
110
70
50
EEoBil
=o9.NzIJJfotuE,lJ.NE,UJ

70 3 ERGEBNISSE
Abbildung 45: Tageszeitlicher Verlauf der Herzfrequenz des Rothirschspießers (Nr. 4)
an Tagen ohne Störversuche im Zeitraum vom 28.L2.90 bis 22.01.91. Dazlt wurden die
Werte von zehn Tagen ermittelt. Zur Glättung der Kurve sind die Herzfrequenzen von
jeweils fünf aufeinanderfolgenden Minuten gemittelt. Die grau schattierten F1ächen
entsprechen der Nacht.
EEoo5E(,oNzUI
=oLrJE!rNEIJI
00
80
to
GO
flo
100
eo
80
70
=EooEsooN2lu:)outG,ILNG,UJ-
Abbildung 46: Tageszeitlicher
an Tagen mit Störversuchen im
Verlauf der Herzfrequenz des Rothirschspießers (Nr. 4)
Zeitraum vom 28.12.90 bis 22.0L.91.
UHRZEIT
UHRZEIT

3.2 Ergebnisse {ür das Rotwild
Ruheherzfrequenz
Die Ruheherzfrequenz wurde wie beim Rehwild ausgewertet, dabei gingen 1569 Liege-
phasen in die Auswertung ein. Die Herzfrequenzen wurden mittels Kruskal-Wallis-Test
statistisch überprüft.
Indiai,duelle [Jnterschiede in der Liegeherzfrequenz
Die mittlere Ruheherzfrequenz war beim Hirschspießer (Nr. 4) mit 73.9+10.7 bedeutend
höher als beim Rothirschkalb (Nr. 5) (58.1 + 7.6).
J ah re s z eitli ch e Abh rin g i gk e it d er Ruh eh, er zf re qu e n, z
Es war ein allmählicher Abfall der Ruheherzfrequenzyonr Dezember (73.2+11.6) über
Januar und Februar (63.4t 10.7) bis März (53.7+8.1) vorhanden (Abb.47).
110
100
90
80
70
60
50
40Dezember Januar Februar
Abbildung 472 Mittelwert, Standardabweichung und Extremwerte der Ruheherzfre-
quenz (Schläge/min) in den einzelnen Monaten (beide Tiere zusammen).
7L

72 3 ERGEBNISSE
Ta g e s z e itli ch e A bh cin g i gk e it d e r Ruh eh e r zfreq u en z
Die Ruheherzfrequenz war nachts und vormittags eher niedrig, am Nachmittag stieg
sie zum Abend hin an. Die tageszeitlichen Schwankungen waren an Ruhetagen imGegensatz zu Tagen mit Störversuchen nicht signifikant. Allgernein war der Tagesver-
lauf der Ruheherzfrequenz zwischen Ruhetagen und solchen rnit Störversuchen ähnlich,
obwohl insgesamt an den Tagen mit Störversuchen niedrigere Werte gemessen wurden
(Abb. 48).
Hf
§._
s-':-ö--- ----+----------
'--9 - ---- '4
..-...- ohne Störversuche
+- mit Störversuchen
9-12 15-18 18-21 21-24 0-3 3-6Tageszeit [hl
Abbildung 48: Tageszeitlicher Verlauf der Ruheherzfrequenz (Schläge/min) beim
Rotwild an Tagen mit und ohne Störversuche (2 Stück Rotwild im Januar und Februar).
Abhängigkeit der Ruheherzfrequenz uon Störuersuchen
An Tagen rnit Störversuchen war die mittiere Herzfrequenz rnit 60"6 t 9.1 signifikant
niedriger als an Tagen ohne Störversuche (66.0 + l'2.2)"
Abhringigkeit der Ruh,eherzfre(luenz uon der Umgebungstemperatur
Ein auf Januar und Februar von 6-15 tihr eingeschränkter Datensatz wurde herange-
zogetl) da in dieser Zeitperiode keine tages- und jahreszeitliciren Herzfrequenzschwan-
kungen auftraten. Es konnte kein Zusammenhang zwischen der Ruhelierzfrequenz und
der Außentempera,tu, (-11 bis *9"C) festgestellt werden"
75
70
65
60
55
50 +*6-9

3.2 Ergebnisse für das Rotwild
Erhöhung der Herzfrequenz bzw. zusätzliche Herzschläge durch Störungen
Tiere, die bei Störversuchen liegengeblieben sind, zeigten eindeutig geringereHerzfre-
quenzreaktionen im Vergleich mit während des Störversuches aktiven Tieren (Tab. 19).
Tabelle 19: Durchschnittliche Herzfrequenzreaktionen nach Störreizen bei liegenden
und aktiven Tieren (2 Stück Rotwild).
VersuchsartZrcätzl,
Heru-schläge
Dauer derHerzfrequenz-
erhöhung
Herzfrequenz-erhöhung
(Schläee/min)n
liegende Tiere
aktive Tiere
I257
102 sek
760 sek
5
20
25
207
Abhtingigkeit der l{erzfrequenzreaktion uon der Art des Störreizes
Beim Hirschspießer (Nr. 4) wurde die stärkste Herzfrequenzänderung durch Feuer aus-
gelöst mit durchschnittlich 622 zusätzlichen Herzschlägen. Das weibliche Rothirschkalb
(Nr" 5) wies mit durchschnittlich 530 ntsätzlichen Herzschlägen die höchsten Herzfre-
quenzänderungen bei der Futtervorlage auf. Die geringsten Reaktionen ergaben sich bei
den Versuchen mit dem Traktor und der Drachengleiterattrappe sowie bei akustischen
Störreizen (Tab. 20).
In der letzten Versuchswoche waren die Zahl der zusätzlichen Herzschläge und die
durchschnittliche Herzfrequenzerliöhung sowohl bei den optischen als auch bei den
akustischen Versuchen geringer als in der ersten Versuchswoche (Tab. 21).
Dennoch war lediglich beim Störreiz ,,Pferderitt" mit fortschreitender Versuchsnummer
eine signifikante Abnahme der Reaktion der Herzfrequertz zLr erkennen (Regressions-
gleiclrung ... y :793 - 3.5r; Korrelationskoeffizient ... 12 :0.304).
IJ

3 ERGEBNISSE
Itr-o
o
aobotio
--otr
U
(,
,ot+
€ao
ao
4
l.4i
0o
U
dOtiN(,
otiNti
#
o
(.)(a
U,4
oc-t oo?.odl.) ---
Fta
74
.X
€H
tO
;zHr-'l
Er
frl*rOapdr-l
frlH
OaFui
0löi oo roF-l
E,lHFri(-)
aHr-lH
filH
OaEra
Fl
I,^NlE 9.E
H/i\r# sH 'V .Ja : .rv[n+1 =d oE
HU)
F.löl
r!?F-l
TO Ci tOFl
I-c-n @ ö]
IN
b E 9!^fr H uYPi*
b rE sH.V,ed!
(§Nt:ia b 6-H
Flt- <i@t--
rr?
r-i<ir..l
@öi$r'-'l
C.lt-Fir-.{
*iöl
-s§-
§ *Sa v d;J rrr U
N tf)
c§
Ic)
r-'l@+ ro
F-l
rllN t-I-
frq)
ol-la4
B.P
d.+r
)l TzütrlFr
c.l 0lN I- TO FtTOt-l
FlFi
t^Ntr-.l..OIJ-vt-
H-* 5 -:>ö* EoH .V .i
Hd.Lv
§Lr-oeQ
HA
roc§
FlF-l Fi
#öl ö.1
t-_Fi
IN
b E P^'rJ = =-&Ldl!2it a\ * v)UV-^ !.v,qaidlj(§N='Ha b 6-
*r
rC)@
-i@ <,i
t- f-§-Fl @
Flroöl
IOIO0l
-_-.: a \§ .\ .qo.i ! .Lvmvt;*rU
NcA
r'{11)
t.-f.-0l
@§-
F-I-Fl
c-n0] c-n
=F C\l*it-
tra
atsro
bo
bo
'-rooatro
lJr
O€€t-i
ti.o0r
O
dtibo
l+t
tio€d
-tiH
lra)
_oEi
a)bodtrofro€ta:
l+t
oho
36a
.oq)
(Jw.
a
z-{t
a

753.2 Ergebnisse für das Rotwild
bo
:OULr-
N;J4AoL'-{ lY-taP:O
.H€X crrtiAHHqJl4r .iLa:{ a
'O +ra
o=b0 .,:d*
v)Ndi-r c)Uti-IJ ()
ad.-Oi.
NXaY§tEBUA>to:i.- a(x)Hä#.:ö=
'§i
tss
Or{.oUh
+00)Zbo9LU)
*O 'F.§t-,t 0)N5o:oH-o9FI
rJ]e
l+rfrll+{aH+iO(tHH
r!FzatrI
LO-$Fl
ö.1
t4{HFli(-)caH
Q
alJ,l
HFatl
tOCN
I-ri
E]Hi+i(-)
aH(-)
al+rfrl
frlFNFHFl
f.- oÖil '-r
NAo
U c,! ':'
= i§.=
€LrdirOtr'=F-\l-i-o+,rb0!?o:dq)a- §P boL)= ta)tv
d
trtrl
Oöl
r-iH
LOc\l r-i
ro I-
boH
ci=.?30(Ed
-t oON^
.!H-ooa)
.E U.EH.v
HCH-.d NHÜ
f--§-
@öl
I-oIli
I1a
Nö.1 (oI: r."r
obo
(nNt-{o
H+r
N+,a)
N
TO
roöl
€I,o
6]
Öl
!'-'l
Lo
aI\trr:
€trU)
cnlro
OA(J
(At{d)
q)tQa
g
o4
aFi0)>
aa
d
atrOJ
oUrn
0)tQ-tatr
U
a
a
o_-
OL)O@F<Uö-O 'U
-qaa, -ä

77
4 Diskussion
Das Ziel der vorliegenden Arbeit war, die Wirkung anthropogener Störreize auf das
Verhalten und die Herzschlagfrequenz beim Reh- und Rotwild unter kontrollierten Be-
dingungen kontinuierlich zu erfassen und zu analysieren.
Eine wesentliche Voraussetzung dazu ist die l(enntnis der natürlichen tages- und jah-
reszeitlichen Aktivitätsrhythmen und der entsprechenden Herzfrequenzen beim un-
gestörten Tier"
Bei den Rehen, fand ein auffallender Aktivitätswechsel um den 2l.Oktober statt. Vor
diesern Zeitpunkt hatten die Rehe tagsüber offensichtlich ein starkes Ruhebedürfnis rrnd
verbrachten die meiste Zeit liegend im großen Grasfeld" Im späten Herbst waren sie
hingegen tagsüber wesentlich aktiver. Dieser Wechsel ging ziemlich plötzlich vonstat-
ten, und es wurden zunächst mehrere mögliche Ursachen in Erwägung gezogen, die aber
dennoch keine eindeutige zeitliche Übereinstimmung mit dem Wechsel in der Tagesak-
tivität zwischen frühem und spätern Herbst zeigten: Sowohl das Spannen eines Seiles
für die Drachengleitversuche und die Liegeplatzwechsel ab dem 24.Oktober als auch das
Hinzukornmen eines dritten Rehes arn 25.Oktober fanden erst einige Tage nach der vor-
erwähnten markanten Veränderung im Aktivitätsmuster statt. Das gleiche gilt für die
Versuche rnit der Drachengleiterattrappe und den Pferderitten, die zum erstenmal am
29.Oktober durchgeführt wurden. Als einziges zeitlich eindeutig übereinstimmendes
Ereignis ist der plötzliche Temperatursturz am 2l.Oktober anzuführen, wo es von den
ursprünglichen Temperaturschwankungen zwischen 9'C und 19"C auf nachher -3"Cbis 8"C gekommen ist. Wahrscheinlich liegt der Anderung im Aktivitätsmuster eine
endogene Umstirnmung der Rehe zugrunde, die durch die vorgenannte plötzliche Wet-
terveränderung ausgelöst wurde. Ein entsprechender Anstieg der Aktivität von R.ehen
im Oktober wurde bisher nur von Et t BNsBnG (1978) erwähnt"
Bei den Rehen waren irn frühen Herbst deutliche Tag-Nacht-Unterschiede in der Herzfre-
quenz zu erkennen. Die Schwankungen scheinen z.T" endogen bedingt zu sein, da
der Zusammenliang zwischen Herzfrequenz und Umgebungstemperatur nur sehr ge-
ring war.
Die ausgeprägten Llnterschiede im frühen Herbst deuten auf eine eventuelle in die-
ser Jahreszeit besonders hohe Sensibilität des Kreislaufes hin. Dies kann zumindest
teilweise mit dem Wechsel des Sornmerhaarkleides und mit der Umstellung auf den
Winterstoffwechsel im Zusammenhang stehen. Sowohl die starke Absenkung der Ru-

78 4 DISI(USSION
heherzfrequenz als auch die ausgeprägte Inaktivität während des Lichttages im frühen
Herbst deuten auf ein stark ausgeprägtes Ruhebedürfnis der Rehe in diesem Zeitraum
hin. Im späten Herbst ist dieses Ruhebedürfnis offensichtlich nicht so sehr ausgeprägt.
Das Rotwild zeigte im Zeitraum zwischen Dezember und März keine auffällige Verände-
rung des täglichen Aktivitätsr-nusters. hn Winter war es etwas mehr nachtaktiv als zum
Frühjahr lrin. Die Gesamtaktivität schwankte bezogen auf 24 Stunden zwischen 47To
und 58% und war somit deutlich höher als die von GEoRGII (1981) im Winter gernes-
senen Werte, die ca. 29Trt betntgen (aus den angegebenen Stundenwerten errechnet).
Wapitis wendeten im Winter 45To iht'er Zeil z;rtm Asen und lediglich 11% fnr die Fort-
bewegung auf (Gnnnm u. Bo.tR 1990). Sie waren jedoch ausgeprägt nachtaktiv, denn
am Liclrttag ästen sie nur während 11% der Zeit. Die im Vergleich zu den in der
vorliegenden lJntersuchung errnittelten Werten (a9% bis 59% Aktivität während des
Lichttages) niedrigen Angaben von GREEtrt u. BB.qR (1990) kommen höchstwahrschein-
lich dadurch zustande, daß letztgenannte Autoren den Lichttag von zwei Stunden nach
Sonnenaufgang bis zwei Stunden vor Sonnenuntergang einstuften. während in dieser
Untersuchung die Dämmerungsphasen rniteingeschlossen sind.
Die Tagesherzfrequenz sank beim Rotwild von Dezember bis März deutlich ab" Sai-
sonale Rhythmen der Herzfrequenz wurden schon oft beschrieben. FRpooy (1984)
maß bei Maultierhirschen die niedrigsten Herzfrequenzen Ende Januar, und Monu u.
Cuovar,ton (1977) fanclen bei Weißwedelhirschen die tiefsten Werte im Februar. Lrps
u" Lps \4aRcutvt (1979) fanden eine negative Korrelation der Herzfrequenz und der
Umgebungstemperatur bei Wapitis, möglicherweise liegt dem auch ein jahreszeitlicher
endogener Rhythmus zugrunde"
Die Ruheherzfrequenz war beim Hirschspießer generell höher als beim weiblichen Rot-
wildkalb, was besonders erstaunlich ist, da bei den untersuchten Säugetieren im all-
gemeinen die Herzfrequenz bei älteren Tieren und bei männlichen Tieren niedriger ist
als bei Jungtieren oder weiblichen Tieren (ScunuNnRT u. TRaurltaNN 1987). Doch
auch bei den Rehen verhielt es sich so, daß die weiblichen Tiere, und insbesondere die
Schmalgeiß, niedrige Ruheherzfrequenzen aufwiesen.
Zwischen den beiden Rothirschen konnte eine stärkere Verhaltenssynchronisation fest-
gestellt werden als zwischen den Rehen. Eine mögliche Erklärung liegt in dem starken
Sozialverhalten des Rotwildes, das eher zu gleichartigen Reaktionen führt. Es muß
allerdings darauf hingewiesen werden, daß diese zwei Wildarten zu unterschiedlichen
Jahreszeiten untersucht wurden, schon deshalb ist ein direkter Vergleich nicht zulässig.

79
Verschiedene Untersuchungen an Rehen ergaben eine stärkere Synchronisation des Ver-
haltens während der Winterrnonate (Er,loNnERG 1978, Mpnrpxs 1984, JBpprsBN
1989). Das gleiche trifft auch für Rotwild zu (GnonGII u. Scunöoon 1981). Jrppp-snu (1989) führt diese Tatsache auf die stärkere photoperiodische Kontrolle des Ver-
haltens im Winter zurück. Daher ist bei Rehen die Verhaltenssynchronisation in den
Dämrnerungsphasen auch höher als tagsüber (TunNER 1980). Nach Bürzlpn (1974)
sind bestimmte Verhaltensweisen, wie Flucht oder Sichern, stärker synchronisiert als
andere (2.8. Komfortverhalten)"
Auffällig war der plötzliche Wechsel des Liegeplatzes beim Rehwild am 24.Oktober.
Als mögliche Ursaclie komrnt die Anderung in ihrem tageszeitlichen Aktivitätsmuster,
die schon einige Tage vorher stattfand, in Frage" Das Grasfeld, das den Rehen bis
dahin als Deckung und als nahezu ausschließlicher Liegeplatz gedient hatte, war in der
letzten Oktoberwoche zwar aufgelockert, das Gras stand aber dennoch hoch. Somit ist
die allmähliche Abnahme des Sichtschutzes durch das Grasfeld für den sehr plötzlichen
Liegeplatzwechsel kaurn verantwortlich. Das Einbringen des dritten Rehes erfolgte
erst nachher und war daher keinesfalls der auslösende Faktor. Sehr wahrscheinlich
für die Anclerung des Liegeplatzes ist, da cler Zeitpunkt genau übereinstimmt, jedoch
das Spanneri eines Seiles quer über das Grasfeld für die Drachengleitversuche. Ein
Wetterweclisel hat an diesem Tag nicht stattgefunden.
Beirn Rotwild war hinsichtlich cler Liegeplatzwahl keine Regelmäßigkeit zu erkennen.
Auffallend war nur, daß ein an einern bestirnmten Tag gewählter Liegeplaiz oft für
clen Rest des Tages immer wieder aufgesucht wurde, hingegen am darauffolgenden Tag
nicht rnehr" Möglicherweise war die Wahl abhängig von den jeweiligen Witterungsbe-
dingungen. Nach Hr:torltlwN (1973) ist bei Darnwild der Windschrtz ausschlaggebend
für die Liegeplatzwahl. Die von FrscuBa u. Gossow (1985) untersuchten Rothirsche
bevorzugten Liegeplätze im Schutz von Bäumen. Diese Tendenz ist bei den in dieser
Arbeit untersuchten Tieren nicht zu erkennen, irnmerhin lagen sie in fast der Hälfte a11er
Beobachtungen frei auf der Wiese, z.T. sogar auf dern kleinen, exponierten Grashüge1.
Reh- und Rotwild verhielten sich auf anthropogene Einflüsse oftrnals verschieden.
So war beim Rehwildim frähen Herbst ein starker en,dogener Aktiuitritsrhythmusvor-
handen, der auch durch eine oftmalige Stimulierung kaum beeinflußbar war. In diesern
Zeitraurn hatten die Rehe offensichtlich ein starkes Ruhebedürfnis und ließen sich kaum
durch Störreize aktivieren. Kam es dennoch zu einer Flucht, war sie nur von kurzer
Dauer, und die Tiere legten sich meist innerlialb weniger Minuten wieder nieder. Durch

80 4 DIS.KUSSIO^r
solche kurzen Unterbrechungen wurde eine Liegephase wohl geteilt, wodurch sich die
Anzahl der Liegephasen erhöhte, die gesamte Dauer der Liegephase sich aber verkürzte.
Das Gesamtaktivitätsbudget der Rehe blieb jedoch durch die Störungen unbeeinflußt.
Die Verhaltensanalyse zeigte, daß die Rehe während des kurzen Aktivschubes fast aus-
schließlich sicherten bzw. sich fortbewegten. Demnach kann man das Reh so lange als
gestört betrachten, bis es sich wieder niederlegte.
Im späten Herbst reagierten die Tiere vermehrt auf Störreize, was jedoch nicht als
Sensibilisierung durch den Störreiz gedeutet werden soll, da die Rehe im allgemeinen,
auch an Tagen ohne Störversuche, zu hoher Aktivität neigten. Wurde ein liegendes
Reh durch eine Störung aktiviert, brauchte es länger, bevor es sich wieder hinlegte,
manchmal blieb das Tier über Stunden aktiv (durchschnittlich jedoch nur 26 min).
Trotzdem zeigte das Reh nicht die ganze Zeit über ein gestörtes Verhalten. Der Störreiz
schien als Stimulans fiir einen Aktivitätswechsel zu wirken, wie dies auch HBIoEMANN
(1973) schon erwähnte. Es erwies sich als sinnvoll, nach dem Störversuch aktive Tiere
mit solchen mit ungestörter Aktivität zu vergleichen und zu messen, wie lange es
dauerte, bis das Tier ein normales Verhaltensmuster zeigte. Dazt wurden besonders
Asen und Sichern als Kriterium herangezogefi. Dies vor allem deshalb, weil einerseits
die Nahrungsaufnahme für das Energiebudget und somit für das Überleben wichtig
ist, andererseits vermehrtes Sichern mit Beunruhigungen des Tieres verbunden ist.
Dabei ist der prozentuelle Anteil der für das Sichern aufgewendeten Zeit eir besserer
Hinweis auf Beunruhigung des Tieres als die Anzahl der Sicherungsphasen. Diesen
Kriterien entsprechend zeigten die Rehe bei optischen Störreizen im späten Herbst ab
der 7. Minute nach dem Ende der Störreizsetzrng wieder normales Verhalten, während
sie nach akustischen Reizen zwar bereits ab der 5. Minute genauso lange ästen wie
ungestörte Tiere, zwischendurch aber vermehrt sicherten.
Sowohl im frühen als auch im späten Herbst karn es bei Rehen durch Störungen zu
keiner Veränderung der Gesamtaktivität. Dies trifft gleichennaßen für den ,,Lichttag"
zu als auch für den 24-Stunden-Zeitraum"
Wurde das Rotwild durch Störreize aktiviert, dauerte es im Schnitt knapp 40 Minuten,
bis es sich wieder hinlegte. Als Folge der Aktivierungen wurden wie beim Rehwild die
Liegephasen verkürzt, aber statt daß es zum Ausgleich dieses Zeitverlustes die Anzahl
der Liegephasen erhöht hätte, blieb es vermehrt aktiv. Damit wurde durch Störun-
gen sein Aktivitätsbudget verändert, und zwar lag es an Tagen mit Störversuchen
(unabhängig von der Anzahl der Versuche) im Schnitt am ,,Lichttag" um eine halbe

81
Stunde geringer und kompensierte diesen Verlust auch nicht während der Nacht.
Zwischen dem Verhalten des Reh- und Rotwildes können auch einige Parallelen gezogen
werden. Optische Störreize hatten, sowohl beim Reh- als auch beim Rotwild, einen
weitaus stärkeren Einfluß auf die Herzfrequenz und das Verhalten als akustische Reize.
Dieses Ergebnis stimmt auch mit den von HorrMsrsrEn (1979) an Reh- und Damwild
durchgeführten Störversuchen überein. Eine Ausnahme bildet lediglich der Schuß, der
von allen geprüften akustischen Reizen besonders starke Reaktionen auslöste, obwohl
die Störung nur einen Bruchteil einer Sekunde andauerte, verglichen mit einer bzw.
drei Minuten bei den anderen akustischen Reizen. Auch Hpnsol» (1990) stellte bei
Rehen nach Abgabe eines Schusses höhere Fluchtdist arrzer fest als bei Beunruhigungen
durch Forstarbeiten oder durch Erholungsuchende.
Auf optische Störreize reagierten die Rehe v.a. mit Stehen und Sichern, während die
Rothirsche mit vermehrter Ortsveränderung und Sichern reagierten. Bei den opti-
schen Versuchen waren zwischen Personenrundgängen mit oder ohne Hund keine Un-
terschiede festzustellen. Auch anderen Untersuchungen entsprechend stellte ein Hund
in Begleitung von Menschen für das Wild keine besondere Bedrohung dar, im Gegen-
satz nt freilaufenden Caniden, durch die oft maximale Herzfrequenzen und hochgradige
Fluchtreaktionen ausgelöst werden (M,tcAntHUR et al. 1979, Sr.q.or,sn 1985, Gars-
BAUER 1988). Nach Personenrundgängen mit oder ohne Hund reagierte das Rotwild
mit ausgiebigem Beriechen des Bodens. Häufig zog es dabei entlang der abgegangenen
Route. Interessanterweise zeigte es dieses Verhalten nach Pferderitten kaum. Es ist
denkbar, daß die Pferdehufe weniger Duftspuren hinterlassen als die Sohlen der Hunde
oder das Schuhwerk von Personen.
Die Reaktion auf Rundritte war der auf Personenrundgänge ähnlich. Während das
Rehwild auf die erste Begegnung mit starker Flucht reagierte und sich erst allmählich
an die neue Erscheinung gewöhnte, ließ sich das Rotwild auch bei der ersten Begegnung
mit Pferden erstaunlicherweise wenig stören. Dabei erwies es sich als gleichgültig, ob
mit einem braunen Pferd oder einem Schimmel geritten wurde.
Gegenüber dem Traktor zeigten die Tiere im Vergleich zrt anderen Störreizen eine
relativ geringe Reaktion. Dieser Befund deckt sich mit Untersuchungen von Scnur,rz
u. Betlrv (1978) und MIcARTHUR et al. (1979). Nur Mnr,tr (1937) konnte beim Reh
hohe Fluchtdistanzen gegenüber Traktoren und anderen Kraftfahrzeugen beobachten.
Bemerkeriswert ist aber auch die geringe Störung des Rotwildes durch den Flugdrachen.
Dies steht im krassen Gegensatz zu den Rehen. Einmal flog zufällig ein Heißluftballon

82 4 DISKUSSIOAI
mit niedriger Hölte iiber das Versuchsgehege, worauf das Rotwild nur mit Sichern imLiegen reagierte. Auch Duap (zit. nach HotouultvN 1957) erwähnt beim Rothirsch
das totale Desinteresse gegenüber Gefahren von oben. Der Grund scheint darin zu
liegen, daß für Rothirsche Greifvögel keine Bedrohung darstellen. Die Rehe zeigten da-
gegen höhere Herzfrequenzreaktionen bei Beunruhigung mit dem Flugdrachen, obwohl
nur deren Kitze durch Cireifvögel gefährdet sind (RansFELD et al" 1985). Gemsen,
bei denen der Adler einen Feind darstellt, sind durch Segelflugzeuge, Drachenflieger
und Adlerattrappen sehr beunruhigt (HaMR 1988). Dickhornschafe flüchten vor Hub-
schraubern panikartig (Honnrsr 1976).
Überraschend sind ciie hohen Herzfrequenzwerte beirn Rotwildkalb zur Zeit der Fut-
tervorlage. Dies rührt vennutlich daher, daß es während der Futieraufnahme,, wenn
die zugeworfenen Futterrüben knapp beieinander lagen. vom Hirschspießer vertrieben
wurde" Das Kalb trollte darauflrin einigemale hin und her, bevor es sich wieder zur
Futterrübe wagte. Praxisbezogen becleutet das, daß bei der Fütterung von Rotwild
ausreichend Futterstellen rnit genügendem Abstand zueinander vorliegen müssen, um
soziale Auseinandersetzungen zu vermeiden. Rangniedere Tiere, die an zu kleinen
Fütterungen warten rnüssen, bis sie von den ranghöheren Tieren freigegeben werden,
neigen auch zu vermehrtem Schälen (ONonnscHEKA 1986).
Sowohl die Rehe als auch die Rothirsche waren am Vormittag und am Nachmittag
leichter zu aktivieren und blieben anschljeßend länger aktiv als in der Mittagszeit.
Das Rotwild zeigte in seinem Verhalten gegen Ende der Versuchsperiode eine verrin-
gerte l)auer der Störung, was für eine gewisse Gewöhnung an optische und akustische
Störeinflüsse spricht. Jedoch ausschließlich bei Pferderitten war mit fortschreitender
Versuchsnummer eine signifikante Abnahrne der Herzfrequenzreaktion festzustellen.
Die Empfindlichkeit der Rehe gegenüber Störungen scheint stark von der Jahreszeit
abhängig zu sein. Das Rehwild war in der zweiten Versuchsperiode (Spätherbst) deut-
lich störanfälliger als zu Beginn der Versuche. Es konnte daher keine Habituation der
Rehe gegenüber Störreizen in diesem Zeitraum festgestellt werden"
Im Gegens atz zu den kurzfristigen starken Herzfrequenzreaktionen nach Störreizen kam
es bei den Rehen durch die Störversuche zu keinen längerfristigen Veränderungen der
Ruheherzfrequenz. Beim Rotwild kam es hingegen an Tagen mit Störversuchen zu
einer Absenkung der Ruheherzfrequenz" Möglicherweise kornpensieren die Tiere den
erhöhten Anteil von Aktivität an Tagen, an denen sie oft gestört wurden, durch an-
schließende tiefere Ruhephasen. Eine andere Erklärung läge in der erhöhten Aufmerk-

83
samkeit an unruhigen Tagen, die mit Braclykardie einhergeht.
Einmal wurde beobachtet, wie der Hirschspießer bei einern Gruppenrundgang zuerst
flüchtete, sich dann aber in einer Ecke des Geheges niederlegte und den Kopf flach am
Boden hielt" Erst bei weiterer Näherung der Personengruppe setzte er seine Flucht
fort. Auch Müt lrn-UsING u. Scut oBru (1967) beschreiben ein Sich-Drücken bei
adulten Cerviden, wenn sie nicht im Rudelverband eingeschlossen sind. WalrunR(1969) beobachtete in zwei Fällen, daß eine von Hunden gehetzte adulte Thomsonga-
zelle (Gazella tltomsomi) sich plötzlich hinlegte und von den Hunden überholt wurde,
ohne bernerkt zu werden" Er fügt aber hinzu, daß dieses Verhalten nur ausnahmsweise
auftritt" Auch Kunr (1991) erwähnt, daß ältere Rehe sich bei ganz hohen Feindreizen
drücken. Nach SIöHR (1982) ist eine bradykarde Reaktion verbunden mit passivem
Verhalten ein Zeichen von Unschlüssigkeit gegenüber einem übermächtigen Reiz.
Die Verhaltens- und Herzfrequenzreaktionen zeigten nach Störeinflüssen nicht immer
gleichgerichtete Veränderungen. So löste z.B. Feuer beim Rotwild starke und lang-
fristige Herzfrequenzreaktionen aus, jedoch gleich nach Beendigung des Störversuches
unterschied sich das Verhalten dieser Tiere nicht rnehr von demjenigen vor der Setzung
des Störreizes. In den meisten Fällen wiesen jedoch Verhaltens- und Herzfrequenzre-
aktionen ähnliche Tendenzen auf.
Die Nachteile einer alleinigen visuellen Beobachtung, wie z.B. fehlender Sichtkontakt
in deckungsreichern Gelände und zeitliche Begrenzung der Beobachtung, können durch
biotelemetrische Meßverfahren mit kontinuierlichen Aufzeichnungen aufgehoben wer-
den (ONTERSCHEKA et al. 1982).
Die Aktivierbarkeit und die Dauer der Aktivierung sind aus den Offline-Dateien rela-
tiv einfach und genau zu bestimmen. Dabei scheint die Aktivierbarkeit ein besseres
Maß für den Einfluß von anthropogenen Faktoren auf das Wild zu sein als die Dauer
der Aktivierung. Beim Rehwild war es für die Dauer der nachfolgenden Aktivphase
unbedeutend, durch welche Art von Störreiz das Tier aktiviert wurde.
Die Herzfrequenz gibt beim Tier einen guten Hinweis auf Beunruhigungen und ist vor
allem wegen ihres engen Zusammenhangs mit dem Energiestoffwechsel und ihrer rela-
tiv einfachen Meßbarkeit ein gern verwendeter Parameter zur Streßuntersuchung von
Tieren. Dabei erweist sich die Anzahl der zusätzlichen Herzschläge a1s besseres Maß
für das Ausmaß einer Beunruhigung als der maximale oder durchschnittliche Anstieg
der Herzfrequenz. Trotzdem ist die Herzfrequenz, allein verwendet, nicht sehr aussa-
gekräftig, da sie stark abhängig ist von der jeweiligen Aktivität des Tieres. In manchen

84 4 DISKUSSIOI\I
Situationen sinkt sie bei Störungen sogar ab, z.B. beim Sichern. Daher ist es sinnvoll,
die Herzfrequerzmessung mit einer Verhaltensanalyse zu kombinieren.
Die Ergebnisse dieser Untersuchung können nicht unmittelbar auf Tiere in der freien
Wildbahn übertragen werden. Folgende Kritikpunkte müssen berücksichtigt werden:
Die Tiere stammten aus Gehegehaltungen und waren Menschen z.T. gewöhnt,
obwohl sie nicht handzahm waren. VöUig neue Reize für die Tiere waren die
Pferderitte und Drachenflüge.
Das Gehege begrenzte die Fluchtmöglichkeit der Tiere" Da sie sich dem Störreiz
durch Flucht nicht entziehen konnten. war die Reaktion auf den gesetzten Störreiz
womöglich verlängert.
Der Flugdrachen glitt inuner entlang der gleichen Strecke. Nach einigen Versu-
chen fehlte der iJberraschungseffekt eines vom Himmel plötzlich auftauchenden
Drachengleiters (,,raumspezifische Habituation" ).
Der Schuß blieb ohne negative Folgen für die Tiere. Es bleibt die Frage offen, ob
Tiere (besonders Rotwild) im Rudelverband den Schuß mit einem erlegten Tier
in Zusammenhang bringen und diese Erfahrung sogar tradieren, Hinweise dafür
bringt Bürzlnn (1986).
Es muß noch erwähnt werden, daß der Traktor außerhalb des Geheges fuhr und
die Personen- und Pferderundgänge innerhalb des Geheges stattfanden. Der tren-
nende Zaun kann mögiicherweise auch einen Einfluß auf das Fluchtverhalten des
Tieres haben.
Die Tiere wurden ad libitum gefüttert. Dies kann möglicherweise einen Einfluß
auf die Ernährungssituation und das daraus folgende Verhalten iraben, z.B^ haben
Rentiere, die in physisch schlechter Kondition waren) anders auf Jäger reagiert
als gut genälirte Tiere (Sxocr,axo u. GnovAN 1988). Ozoca u. VnRIr,tn (1970)
konnten bei Weißwedelhirschen in Gehegehaltung nachweisen, daß untergewich-
tige und gutgenährte Tiere ähnliche jahres- und tageszeitliche Aktivitätsrnuster
zeigten, doch ließen die rnangelernährten Tiere in ihrem Verhalten eine stärkere
F luchtreaktion erkennen.
Beim Roiwild waren nur zwei junge Tiere vorhanden. Es fehlte ihnen der Sozial-
verband, der für das Rotwild von entscheidender Bedeutung ist. Nach Pornlx

85
(1988) rea€lieren Rudel mit älteren und erfahrenen Leittieren bei Störungen oft
nur mit Sichern, während Rudei mit jungen Leittieren in vergleichbaren Situatio-
nen flüchten. Ebenso reagieren kleine Gruppen eher mit Flucht als ein größeres
R.udel.
Die Versuche unter Gehegebedingungen bringen aber auch große Vorteile. Die gute Be-
obachtbarkeit der Tiere erleichtert die Interpretation der biotelemetrisch übertragenen
Meßwerte, da immer wieder mit Videoaufnahmen verglichen werden kann. In freier
Wildbahn kommt es viel häufiger zu unkontrollierten Störungen, ctie eine Deutung der
Akiivität und cler Herzfrequenz der Tiere erschweren. Weiterführende tlntersuchun-
gen in einem Großgehege und anschließend in freier Wildbalin stellen eine sinnvolle
Ergänzung zu den grundlegenden Erkenntnissen aus dem Kleingehege dar.

86 5 ZUSAMMEATTASSUAIG
5 Zusarnmenfassung
An drei Rehen (Capreolus capreolu.s) und zwei Rothirschen (Ceruus elaphus) wurden
gezielte Störversuc.he in einern Kleingehege durchgeführt. Die Tiere waren mit im-
plantierten Sendern ausgerüstet, die die Herz{requenz und die Stellung des Kopfes
(gehoben/gesenkt) übermittelten. Die Untersuchung erstreckte sich auf den Vergleich
der Herzfrequenz und des Verhaltens vor, während uncl nach einer Störreizsetzung und
auf die Erfassung des gesarnten Aktivitätsmusters.
Das Rehwild zeigte Mitte Oktober einen markanten Einschnitt in seinem Aktivitäts-rhythmus. Es wechselte von überwiegend inaktivern Verhalten während des Lichttages
(15-29% aktiv) auf vermehrte Tagesaktivität (50-61%). Das Rotwild war während des
Lichttages die ganze Untersuchungsperiode über (Dezember bis März) zu etwa 50-60%
aktiv. Beim Rehwild wurde durch die Störversuche die Dauer der einzelnen Liege-
phasen verkürzt. Gleichzeitig erhöhte sich jedoch deren Anzahl, sodaß das gesamte
Aktivitätsbudget. gleichgültig ob nur auf den Lichttag oder auf 24 Stunden bezogen,
durch Störversuche unbeeinflußt blieb. Das Rotwild neigte an Tagen mit Störversuchen
zu vermehrter Aktivität arn Lichttag bei gleichbleibender nächtlicher Aktivität.
Sowohl Reh- als auch Rotwild waren durch optische Störreize (Personen, Hund, Pferde
und Flugdrachen) viel leichter aktivierbar und zeigten deutlichere Herzfrequenzreak-
tionen als durch akustische Störreize (Motorsägelärm, Radio, Hundegebell). Eine Aus-
nahme bildet cler Schuß, der als akustischer Reiz ebenfalls hohe Reaktionen auslöste.
Die Aktivierbarkeit war abhängig von der Tages- und Jahreszeit. Am frühen Vormittag
und am späten I\achmittag wurden die Tiere eher aktiviert als um die Mittagszeit. Die
Rehe waren irn Spätherbst signifikant leichter zu beunruhigen als im Frühherbst"
Nach Personenrundgängen und Schüssen zeigte das Rotwild im Vergleich mit un-
gestörten Tieren rnindestens 10 Minuten lang ein verändertes Verhalten, während die-
selben Tiere nach Pferdelitteri, Drachenflügen, Traktorfahrten und den meisten aku-
stischen Störreizen schon in der ersten Minute nach Beendigung des Störreizes wieder
ein ungestörtes Verhalten zeigten. Beim Reh wurde irn frühen Herbst das Verhalten
durch Störreize nur geringfügig beeinflußt, im späten Herbst dauerte es dagegen nach
optischen Störreizen etwa 7 Minuten, bis die Tiere wieder ein ungestörtes Verhalten
zeigten.
Optische Störrbize verursachten bei Rot- und Rehwild generell kurzfristig hohe Herzfre-
quenzreaktionen. Flugdrachen lösten jedoch beim Rehwild im Vergleich zum Rotwild

87
bedeutend stärkere Reaktionen aus. Akustische Störreize hatten nur geringe Herzfre-
quenzreaktionen zur Folge, abgesehen von Schüssen, die sowohl beim Rotwild als auch
ganz besonders beim Rehwild starke Herzfrequenzerhöhungen auslösten. Eine Habi-
tuation fand beim Rotwild nur gegenüber Pferden statt.
Langfristige Veränderungen der Ruheherzfrequenz konnten beim Rehwild nicht festge-
stellt werden, während es beim Rotwild an Tagen mit Störversuchen zu einer Senkung
der Ruheherzfrequenz kam.
Insgesamt zeigte Rehwild eher kurze und heftige Reaktionen auf Störreize, während
Rotwild zLt zwar länger anhaltenden, jedoch weniger ausgeprägten Reaktionen ten-
dierte.
Summary
Three roe deer (Capreolus capreolus) and two red deer (Ceruus elaphus) were investiga-
ted during planned disturbance trials. By means of implanted telemetry transmitters
heart rates and positions of the head (up/down) were recorded. The study included
comparing heart rates and behavior before, while and after confronting the animals
with various stimuli and recording their complete activity patterns"
The observed roe deer altered their activity pattern in the third week of October.
Being mostly inactive during daytime in the first half of October (15-29% active), they
sudclenly changed to more daytirne activity (50-61%) afterwards. Red deer were active
50 60% of the daytirne throughout the investigation period from December to March.
By roe deer tlie length of resting bouts were shortened on days including disturbance
trials. At the same time the number of resting bouts per day increased, so that the total
activity budgets for daytime as well as for 24 hours were not influenced by disturbance
trials. Red deer tended to be more active during daytime when disturbed without
changing their nighttime activity.
Both roe deer and red deer exhibited stronger behavioral reactions ("activation") to-
wards optical stirnuli (humans, dogs, horses and hanggliders) than to acoustic stimuli
(cirain saw, radio music, dog barking) with exception of gunshots, which also elicited
high reactions" Activation was associated with daytime and season. Both roe and

88 5 ZUSAMMENFASSUAIG
red deer were rnore often activated in the morning hours and iate afternoon than at
noontime and roe deer were rnore easily activated in late fall than in early autumn.
The behavior of recl deer was disturbed for at least 10 minutes after gunshots ancl
persons passing by, whiie riders, hanggliclers, tractors and most acoustic stimuli had
only mild reactions as consequence. In early autumn roe deer were hardly disturbed,
whereas in late fall disturbed behavior after optical trials persisted about 7 minutes.
Optical stimuli generally caused liigh shorttirne heart rate reactions in deer. Hanggli-
ders elicited higher heart rate reactions in roe deer than in red deer" Acoustic stimuli
evoked only mild heart rate reactions, except for gunshots, which caused strong heart
rate reactions both in red deer and especially in roe deer. Red deer habituated only
towards horses"
Roe deer dicl not show any longtirne heart rate changes throughout the whole investi-
gation period, whereas reci deer tended to have lower resting heart rates on days on
which disturbance trials were carried out.
Altogether the reactions of roe cleer towarcls stimuli were short and strong, whereas
red deer tended to show less obvious but persisting reactions.

89
6 Literaturverzeichnis
ALsRecHr, L.(1988): Die Gewöhnung von Gernsen an den Menschen. Wildtiere 3,
1-4"
Ar.runNN, Margaret(1958): The flight distance in flee-ranging big garne. J. Wildi.
Manage. 22,207-209.
AscHoFF, J.(1966): Circadian activity pattern with two peaks. Ecology 47" 657-662"
BRtL, l{.J., AulaNnR. Jn.(1979): Changing heart rates of herring gulls when ap-
proached by humans. In: Atr,tt.INER, C"J. und MncDoxar,o, D"W.(Hrsg.): A
handbook on biotelemetry and radio-tracking. Pergamon Press. Oxford" 589-
594.
Ba.nru. W.-E.(1984): Menschliche Störungen in der Natur Ii. Wild und Hund 10,
37-42.
BEIER, P., IvIcCuLLoucII, D.R.(1990): Factors influencing white-tailed deer acti-
vity pattelns and habitat use" Wildl. N{onogr. 109, 5i S.
BsRcsn.uo, A.T., JAKTMCHUI{. R.D., CaRRutr'rERS, D.R.(1984): The buffalo of
tlre nortli: Caribou (Rart.oi.fer tarandus) and hurnan developrnents. Artic. 37,
7-22.
Blacxsuar,v, J.I(.(1986): Objective rleasures of welfare in farming environments.
Australian Veterinary Journal 63, 361-364.
Boyr,n. S.A., SausoN, F.B.(1985): Effects of nonconsumptive recreation on wild-
life: A review" Wildl. Soc. Bull. 13, 110-116"
BuRCH, W.(1988): Human ecology and environrnental management. In: Acnp, J.K.
und DRRRyLL. R.J.(Hrsg.): Ecosystem lnanagernent for parks and wilderness,
Llniversity of Wasliington Pless, Seattle and London. 145-159.
Bümupn, K.(1980): tlntersuchungerl zurn Einfluß von Größe und Zusarnrnensetzung
des Wintelsprunges beim Rehwild auf die Zeitdauer verschiedener Verhaltenswei-
sen. Z. Ja,gdwiss. 26, 181-i94.
BürzlnR, W.(197a): I(zlmpf- und Paarungsverhalten, sozia,le Rangordnung und Ak-
tivitätsperioclili beinr Rotirirsch (Cervus elaphus L.). Parey. Berlin.

90 6 LITERATTIRVERZEICHNIS
BÜrzr,nR, W.(1986): Rotwild: Biologie-Verhalten-Llniwelt-Hege" BLV Jagdbuch, 3.
Aufl., BLV Verlagsgesellschaft, Mänchen.
C,tNNoN, W.B.(1929): Bodily changes in pain, hunger, fear and rage: An account of
recerrt researches into the function of emotional excitement. New York. Appleton.
Cnonnr,uxD, G.(1981): Daily and seasonal activity pattern of roe deer in a boreal
habitat. Viltrevy 11, 315-353"
CBoBnun, A., LovARr, S.(1985): The impact of tourism on chamois feeding activi-
ties in an area of the Abruzzo National Park,, Italy. The biology and management
of mountain ungulates" 216-225.
CuntsreNSEN, N"L.(1988): Succession and natural disturbance: Paradigms, pro-
blems, and preservation of natural ecosystems. In: AcBE, J.K. und DaRRvtt,R.J.(Hrsg.): Ecosystem management for parks and wilderness, University of Wa-
shington Press. Seattle and London. 62-86.
CottINs, W.ts., URl,,Bss, P.J., AustIlt, D.D.(1978): Elk diets and activities on
different lodgepole pine habitat segments. J. Wildl. Manage. 42,799-810.
Cznc:u, B.(1991): trlk behavior in response to human disturbance at the Mount
St. Helens National Volcanic Monument" Applied Animal Behaviour Science 29,
269-277.
Drntoxo, S., LAZARUS, J.(197a): The problem of vigilance in animal life. Brain,
Behav., Evol. 9. 60-79"
Er,r,nNsBRG, H.(1978): Zul Populationsökologie des Rehes (Capreolus capreoius L.,
Cervidae) in Mitteleuropa. Spixiana - Zeitschrift für Zoologie" Suppl. 2.
EspuaRx, Y., LlucvATN, R.(1979): Cardiac responses in alarmed red deer calves.
Beiravioral Processes 4. 179-186.
Esptrnnx, Y., LANGVATN, R.(1985): Developement and habituation of cardiac and
beliavioral responses in young red deer calves (Ceruus elaphus) exposed to alarm
stinruli. J. Mamm. 66,702-711.
FlNcv, S., WuIrn, R.(1985): Predicting enelgy expenditures for activities from
heart rates of caribou. Proceedings of the 1985 Artic Science Conf. 52.

91
FpnNs, P. N., MacAlerN p-Lnxy, I. H., Goss-CusrARD, J.D. ( 1 979): Telemetry of
heart rate as a possible method of estimating energy expenditure in the redshank.
In: AltlarvpR UND MACDouar,n (Hrsg.): A handbook on biotelemetry and
radio tracking. Pergamon Press. Oxford. 595-602.
FisctttrR, A., Gossow, H.(1985): Zw winterlichen Einstandswahl des Rotwildes in
Abhängigkeit von Sicherheits- und Klimaschutzansprüchen" XVIIth Congress of
the lnternational Union of Game Biologists" Brussels, Sept. 17-21, 1985. 275-222.
FnRson, D", RtcurE, J"S.D., FRasER, A.F.(1975): The terrn "stress" in a veteri-
nary context" Br. Vet. J. 131, 653-662.
FRnoov, D.J"(1979): Measuring heart rates of mule deer using a repeater-type tele-
metry system. Proc" 2"d Int" Conf. Wildl. Biotelern. 744-155.
FRoo»v, D.J.(19Ea): Heart rate for activities of mule deer at pasture^ J" Wildl.
Manage. 48, 962-969.
FRooov, D.J., BRoNaucH, W.M., FowtoR, 1\4.C.(1986): Responses of mule deer
to disturbance by persons afoot and snowmobiles. Wildl. Soc" Bull. 14, 63-68"
GatssmrpR, R.(1988): Telemeirisch errnittelte Reaktionen des Reirwildes auf simu-
lierte Störungen im Nationalpark Bayerischer Wald. Fachhochschule Weihenste-
phan" Diplornarbeit"
GtrIst, V.(1971): Is big game harassment harmful? Oilweek 22, 12-13.
GEoRctt. B.(1976): tlntersuchungen über das Nebennierengewicht des Rehes" Z.
Jagdwiss. 22, 85-97.
GnoRctt, B"(1980): Einflüsse menschlicher Störungen auf Standortwahl und Akti-vitätsmuster weiblicher Rothirsche. Verhandlungen der Gesellschaft für Ökologie,
Freising-Weihenstephan 1979. Band VIII, i63-168.
GaoRctt, B.(1981): Activity patterns of fernale red deer (Ceraus elaphus L.) in the
Alps. Oecologia 49, 127 -736.
GooRctt, B.(198a): Activity patterns of free-ranging red deer Ceraus elalthus L. Acta
Zool. Fennica 172, 81-83.
GuoRctt, 8., ScunöDER, W.(1978): Radiotelemetrisch gemessene Aktivität weib-
lichen Rotwildes (Cervus elaphus L.). Z.Jagdwiss" 24, g-23.

6 LITERATURVERZEICI{IüIS
GnoncIt, 8.. Scunöoen, W.(1981): Raurnnutzungs- und Aktivitätsmuster von
Rothirschen unter besonderer Beräcksichtigung menschlicher Störungen. Wild-biol. Gesellschaft, München 1977 e.V.
GRorm, R.A., BnnR, G.D.(1990): Seasonal cycles and daily activity patterns of
rocky rnountain elk. J. Wildl. Manage. 54,272-279.
GnztuBx, B.(1972): Grzimeks Tierleben. Enzyklopädie des Tierreiches. Kindler
Verlag. Zürich.
Guruönr,, V.(1987): Radiotelemetrische Untersuchungen zum Raum-Zeit-System
des Rehwildes, Capreolus c. capreolus (Linn6, 1758). Institut für Biographie der
Universität des Saarlandes. Diplomarbeit.
Hann. J"(1988): Disturbance behaviour of chamois in an alpine tourist area of Aus-
tria" Mountain Research and Developement. 8, 65,73.
HuonuaNm, G.(1973): Zur Biologie des Damwildes" In: Marnmalia depicta 9.
Parey. Hamburg.
HBRnoLD, H.(1990): Reaktion von Rehwild auf Störungen durch Menschen" Trans"
1gth IUGB Congress. Trondheim 1989. ,114-420.
HornltEtsrER, H.(1979): Untersuchungen über die Reaktionen von Reh- und Dam-
wild auf verschiedene Umwelteinflüße unter Einsatz der telemetrischen Erfassung
von Herzfrequenz und EKG. Hannover. Tierärztliche Hochschule. Dissertation.
HoltnR, J.B., URnnn, W.E., HAyES, H.H., SrlvoR, H.(1976): Predicting me-
tabolic rate frorn telemetered heart rate in white-tailed deer. J. Wildl. Manage^
10,626-629.
Hooo, R.E., Ilcr,ts, J.M.Q97a): Behavioral responses of white-tailed deer to in-
tensive ranching operations. J. Wildl" Manage. 38, 488-498.
Honolst, B.(1976): Sonre thoughts and observations on harassurent and bighorn
sheep. Biennial Meeting of the lrlorthern Wilcl Sheep Council, Jackson, Wyoming,
Febr. 10-20. 1976. 1-14.
Itutl,tpt,ir,tauN, I(.(1982): Wörterbuch der Verhaltensforschung. Parey. Berlin.
JncossBN, Nadine K.(1979a): Alarrn bradykardie in white-tailed deer fawns (Odo-
coileus airginianus). ,J. Mamm. 60, 343-349.
92

93
JlcosspN, Na,dine K.(1979b): Changes in heart rate with growth and activity of
white-tailed deer fawns. Comp. Biochem. Physiol. 624, 885-888"
JnwxtNs, W.L., KRucBR, J.M.(1973): Modern concepts of the animal's physiolo-
gical response to stress. In: Youlc, E.(Hrsg.): The capture and care of wild
animals. Ralpli Curtis books. Hollywood. 172-182.
JnppusoN, J"L.(1984): Human disturbance of roe deer and red deer: Preliminary
results" Commun. Inst. For. Fenn. 120, 113-118.
JppppsnN, J.L.(1987): hnpact of human disturbance on home range, movements and
activity of red cleer in a Danish environment" Danish Review of Game Biology
13, 1-38.
Jnppospl. J.L.(1989): Activity patterns of free-ranging roe deer ( Capreolus capreo-
lus) at KalO. Danish Review of Game Biology 13,,1-23.
JoNps, A.R., PRICE, S.(1990): Can stress in deer be rneasured? Deer 8,25-27"
KaulrnRuEyER, K.E., MancutruroN, R.L.(1977): Seasonal change in circadian
activity of radio-monitored deer" J. Wildl. Manage. 41, 315-317.
KnnacnR, T.J.(1990): Physiological monitoring of gray wolves (Canis lupus) by
radiotelemetry. J" Mamm. 7I, 258-26I.
KuRT, F.(1991): Das Reh in der Kulturlandschaft: Sozialverhalten und Ökologie
eines Anpassers. Parey. Hamburg.
LovtNn, S.(1985): A definition of stress? In: MoapRG, G.P.(Hrsg.): Animal stress.
Waverly Press Inc" Baltimore, Maryland" 51-69.
Ltee, J.W.(1981): Activity of free-ranging elk as determined by a motion-sensing
biotelemetry system. Proc. 3'd Int. Conf. on Wildl" Biot., 1-12.
Ltne, J"W., Los Mancuu, C.(1979): Biotelernetric monitoring of heart rate and
activity in elk" Proc. 2"d Int. Conf. on Wildl. Biot., 2l-32"
MacAnruuR, R..A., JouNstoN, R"H., GEIST, V.(1979): Factors influencing heart
rate in free-ranging bighorn sheep: A physiological appraoch to the study of
wilcllife harassrnent. Can. J. Zool. 57 , 2010-2021.

94 6 LITERATURVERZEICHNIS
MacAnruuR, R.A., Gnts.r. V., JottxstoN, R.H.(1982): Cardiac and behavioral
responses of mountain sheep to human disturbance. J. Wildl. Manage. 46, 351-
358.
Maurz, W., FAIR, J.(1980): Energy expenditure and heart rate for activities of
white-tailed deer" J. Wildl. Manage. 44,333-342.
MoRTBNs, Claudia(198a): Zur Verhaltensynchronisation bei gruppenweise gehalte-
nen Rehen (Capreolus capreolus). Behavior 89,220-240"
Mtllpn, F.L., GUNN, Anne(198a): Muskox defense formations in response to heli-
copters in the Canadian high artic. Biol. Pap" Univ. Alaska Spec. Rep. 4, 123-
126.
MILLER, NT.E.(1982): Motivation and psychological stress. In: Prarl , D.W.(Hrsg.):
The physiological mechanisms of nrotivation. Springer Verlag, N.Y. 409-432.
MoBERG, G.P.(1985a): Biological response to stress: Key to assessment of animal
well-being? In: MonoRc, G.P.(Hrsg.): Animal Stress. Waverly Press Inc.,
Baltimore, Maryland. 215-267.
MoBERG, G.P.(1985b): Influence of stress on reproduction: A measure of weli-being.
In: MosERG, G.P"(Hrsg.): Animal Stress. Waverly Press Inc., Baltimore, Ma-
ryland" 245-267.
MopN, 4., Cupvr,rBn, Susan(1977): Analysis of telemetered ECG signals from
white-tailed deer" Proc. 1"t Int. Conf. on Wilcll. Biotel. 118-125"
Motit't, 4., DnILA FERA, M.A., HILLER, A.L., BuxroN, Bonnie(1977): Heart
rates of white-tailed deer fawns in response to recorded wolf howls" Can" J. Zoo7.
56,1207-1210.
MooN, A.N., Wut'r.tnltoRE,, Susan, Buxtoll, Bonnie (1982): Effects of disturbance
by snowmobiles on ireart rate of captive wirite-tailed deer. New York Fish and
Game Journal 29, 176-1E3.
Mnltx. W.(1990): Disturbance of the roe deer (Capreolus capreolus) in agrocoenoses
of southern Moravia. Folia, Zoologica 39, 25-35"

95
MÜlrrn-UsING, D.. Scutor:rri. R.(1967): Das Verhalten der Hirsche (Cervidae),
10. In: Hplucxp, J.-G., LeNcnRr(EN, H.v., SrRRcx, D., WERMUTH, H.
(Hrsg.): Handbuch der Zoologie, 8. Band, Walter de Gruyter & Co. Berlin. 1-60"
OnoBnscunxa, K.(1979): Untersuchungen der Wechselwirkung zwischen Rotwild-
population und Umwelt unter besonderer Berücksichtigung der Ursachen der
Wildschäden. Österreichs Weiclwerk 9, 480-485.
ONnpRsctloxa, K.(1985): Auswirkungen der tlmweltveränderungen auf das Rot-,
Reh- und Gamswild. Beiträge zur Umweltgestaltung- Alpine lJmweltprobleme
,,Der Wald als Lebensraum", Facharbeitstagung Achenkirch 1985, 31-35.
ON»onscuEI(A, K.(1986): Ist die Fütterung des Rotwildes in der Kulturlandschaft
des alpinen Raumes eine biologische Absurdität oder ein Beitrag zur Erhaltung
der Funktion des Ökosystems? Tagungsbericht ClC-Rotwildtagung Graz 1986,
386-395.
ONopnscsEI(A, K., ScuosnR, F.. Soltlrnn, F.(1982): Vergleich zwischen opti-
schen und biotelemetrischen Methoden bei der Beobachtung des Rehwildes. Cbl.
ges. Porstwesen 99, 129-139.
Ozocl, J.J., VrnME, L.J.(i970): Winter feeding patterns of penned white-tailed
deer. J. Wildl" Manage. 34, 431-439.
PBrnRsoN. R., MouNTFoRT, G., HorloM, P.A.D.(1976): Die Vögel Europas.
11. erw. Aufl. Parey, Hamburg.
Pprnax, N,{.(1988): Skilangiauf und Rothirsch (Ceruus elaphus Linn6, 175S) in der
Eifel. Z. Jagdwiss. 34, 105-114.
Prarn, D.W (1982): Motivational concepts: Definitions and distinctions. In: Pr.trr'(Hrsg.): The physiological mechanisms of motivation. Springer Verlag, N.Y" 3-24"
PnBtrEpR, J., HARTFIEL, W.(1984): Beziehungen zwischen der Winterfütterung
und dem Schälverhalten des Rotrvildes in der Eifel. Z. Jagdwiss. 30,243-255.
Ptnl-owsxt, Z.(1983): Das Feldreh - Eine Bereicherung der Wildtierfauna unserer
Feldreviere. In: HorltaxN, R.R.(Hrsg.): Wildbiol. Inform. für den Jäger VLStuttgart. 29-35.

6 LITERATURVERZEICHNIS
Poncns, S.W.(1985): Spontaneous oscillations in heart rate: Potential index of
stress. In: MosnRG, G.P.(Hrsg.): Animal stress. Waverly Press, Inc" Baltimore,
Maryland. 97-111.
RRBsrBlo, F.v., Nouuaus, A.H., Scuarcu, W.(1985): Das Rehwild. Naturge-
schichte, Hege und Jagd. 9.Aufl. Parey. Hamburg.
RnBsrulo, F.v., RBulrcl(E, I(.(1988): Das Rotwild. Naturgeschichte, Hege und
Jagdausübung. 9.Aufl. Parey. Hamburg"
Raunn-GRoss, E}., Gossow, H., HAMR, J., Czd.aanr. H.(1988): Beiträge zur
Verhaltensökologie der Gemse (R. rupricapra r.) in touristisch stark belasteten
alpinen Gebieten" CIC: Gamswildsymposium. Ljubljana 25.126.10.1988. 127-
r43.
Rotcuuor,E, J.(1980): Jahreszeit- und fJiotopabhängigkeit der Rudelbildung beim
Rehwild. Spixiana 3, 193-208.
RBIcunor,F, J., Scunltnl, H.(1988): Segelsport und l{aturschutz - gehört das Se-
geln aus ökologischer Sicht zur ,,ruhigen Erholung"? Zeitschrift für angewandte
Umweltforschung. 343-354.
RntuosBn, F., MAyER, H., HolzrNcER. A., ZaNor, J.(1987): Einfluß von Som-
mer- und Wintertourismus auf Waldschäden durch Schalenwild im Angertal (Bad-
gastein). Cbl. ges. Forstwesen 104, 95-118.
RosHcuovsKII, M.P., I(oNovALov, N.1., BozmosrKov, V.S.(1976): Cardiac com-
ponent in emotional stress in elk Alces alces and reindeer Ran,gifer tarandus. J.
Evol. Bioch. Physiol. 12,347-349.
RusuBN, J.(i991): Problems associated with the interpretation of physiological data
in the assessment of animal welfare. Appliecl Animal Behaviour Science 28, 381-
386.
Rvxtnl, JR.,, Eor,vARD, J.(1985): Towards a definition of ecological disturbance.
Australian Jourirai of Ecology 10, 361-365.
ScuaNnRv, R.(1989): Lehrbuch der Psychophysiologie. Psychologie Verlags Union,
2. überarb. u. erw. Aufl., München-Weinheim.
96

97
ScunuNBR'r, A., TRAUTN{ANN, A.(1987): Lehrbuch der Veterinär-Physiologie. 7"
Aufl., Parey. Berlin.
Scur,Btor, W.M." SunlrpR, M.D.(1983): The effect of spatial context on habitua-
tion to a predator model. Z. Tierpsychol. 61, 67-70"
Scttl,tto:r, Karoline (1992): Über den Einfluß von Fütterung und Jagd auf das Raum-
Zeit-Verhalten von alpinen Rotwild. Z" Jagdwiss. 38, 88-100.
ScuoanR, F.(1986): A versatile receiving and data acquisition system for physiolo-
gical and ethological research. In: Arbeitskreis Telemetrie e.V.(Hrsg.): ETC 86
- Proceedings of the European Telernetry Conference, Garmisch-Partenkirchen
1986. Eigenverlag Arbeitskreis Telernetrie e.V. 351-476"
ScuoanR, F., PRstnRK. W., SoultnR, F.(1982): Impiantablelong-range, long-life,
heart late and activity transmitter for free-ranging animals. In: Stanford Univer-
sity: Proceedings of the 7tl' international symposium on biotelemetry, Stanford
i982" 108-1 11.
Scuoron, F., BucNAR, W., WacNnR, J"(1988): Software package for acquisition
and evaluation of biotelemetric data from domestic and wildlife animals. In:
AurRNoR, C.J.(Hrsg.): Biotelemetry X, Proceedings of tire 10th internat. sym-
posium on biotelemetry, Fayetteville. Arkansas, USA 1988" The tlniversity of
Arkansas Press, Fayetteville, London" 700-708.
Scnulrz, R..D., Bltltrv, J.A.(1978): Responses of national park elk to human
activity. J. Wildl. Manage. 42, 91-100.
Sulve, H.(1936): A syndrome produced by cliverse nocuous agents" Nature 138, 32.
Snlvn, H.(1952): The story of the adaptation syndrome. Acta Inc. Montreal"
Sual:roR, M.D.(1984): Predator-prey behavior and habituation. In: Prpxs, H.V.S.
und PnrRINovICH, L.(Hrsg.): Habituation, sensitizatiotl and behavior. Acade-
mic Press, Inc. Orlando. 349-383"
SttvBR. Helenette, Colovos, I§.F., HolrnR, J"8", Havns, H.H.(1969): Fasting
metabolism of white-tailed deer" J. Wildl" Manage. 33, 490-498.
Sxocllrvo. T., GRovAN, B.(1988): The effects of human disturbance on the activity
of wild reincieer in different physical condition. Rangifer 8, 11-20"

98 6 LITERATURVERZEICHNIS
Slttru, E.N., Wontu, D.J.(1979): Atropineeffect on fear bradykardie of the eastern
cottontail rabbit, Sylailagus fl"oridanus. In: AUIRNER und MacDoNnr,o(Hrsg.):
A handbook on biotelemetry and radio-tracking. Pergamon Press" Oxford" 549-
556"
SpaRRowE, R.D.. SpRrNcnn, P.F.(1970): Seasonal activitypatterns of white-tailed
deer in eastern South Dakota" J. Wildl. N4anage" 34,420-431.
SQutss, R.C", I(tttnntl, J.F.Jn., AuonRsoN, D.R.(1986): Birnodal distribution
of estimated conception dates in rocky mountain elk. J. Wildl. Manage. 50,
1 18-122.
StRoleR, R.(1985): Reaktionen des Rehwildes auf Störungen in baliungsnahen Wald-
gebieten am Beispiel des Nürnberger Reichwaldes. Fachhochschuie Weihenste-
phan. Fachbereich Forstwirtschaft. Diplomarbeit"
Sroup, R.E.(1983): tleart rate responses of bighorn sheep to environmental factors
and harassment. Masters Degree Project, Faculty of Environrnental Design, tlniv.of Calgary. Alberta, Canada^
SröHn, W.(1982): Telemet,rische Langzeituntersuchung der Herzfrequenz von Tupaia
belangeri: Basalwerte sowie phasische und tonische Reaktionen auf nichtsoziale
und soziale Belastungen. Universität Bayreuth. Fakultät Biologie, Cliemie und
Geowissensciraften. Dissertation.
Silolr:r, G.II.(1981): What is animal stress and how is it rneasured? J. Animal
Science 52, 150-153.
Slout, J.F., Scuwae, E.R.(1979): Telemetry of heart rate as a measure of the
effectiveness of clispersal inducing stimuli in seagulls. In: AIITANER und Mac-DoNalo (Hrsg.): A irandbook on biotelemetry and radio-tracking. Pergamon
Press. Oxford" 603-609.
Srusso. C., PasslRCiE, H.(1979): Rehwild. Verlag J" Neumann-Neudamm" Mel-
sungen.
SwpnNrv, J.R., MaRcutNToN. R.L.. SwppNny, J.M.(1971): Responses of radio-
rnonitored white-tailed cleer chased by hunting dogs. J. Wildl. Manage. 35, 707-
7i6.

99
TuRNon, D.C.(1980): A multi-variate analysis of roe deer (Aapreolus capreolus)
population activity. Revue Suisse Zool. 87, 991-1002.
lJmorRwooD, R.(1982): Vigilance behaviour in grazing African antelopes. Behavior
79, 81-107.
W.tcNnR, J.(1991): EKG-Ableitungsorte bei Wildwiederkäuert7 ^)r Herzfrequenz-
übertragung mittels implantierbarer Telernetriesender" Wien. Veterinärmedizi-
nische Universität. Dissertation.
Wnlsu, Noreen E., FRNcv, S.G., McCanu, T.R., PRNt<, L.F.(1992): Habitat use
by the Porcupine Caribou Herd during predicted insect harassment," J. Wildl"
Manage. 56,465-473.
Wl.lruoR, F.R.(1969): Flight behaviour and the avoidance of predators in Thom-
son's gazelle (Gazella thomsortz Guenther, 1884). Behavior 34, 184-221.
Waao, A.L.(1964): The response of elk and rnule deer to firewood gathering on the
Medicine Bow Range in southcentral Wyoming. Proceedings of the 1984 western
states and provinces elk workshop. 28-40.
Wpn.Nnn J.-S.(1976): Telemetrische Messungen an Rehen zur Prüfung der Reaktion
auf simulierte Umweltreize. Hannover. Tierärztlicire Hochschule" Dissertation"
WurrrpN, K.R.. Ganxon, G.W., MArrER, F.J., H,q,nRrs, R.B.(1992): Produc-
tivitv ancl early calf survival in the Porcupine Caribou Herd. J" Wildl. Manage.
56: 201-212.
ZBHNTBn, H.-C.(1991): ,,Ab wann ist eine Störung eine Störung?" Diskussionsbei-
trag der Projektgruppe Etho-Ökologie der Ruhr-Universität Bochum,, gehalten
auf der 124. Jahresversamrnlung d. Deutschen Ornithologen-Gesellschaft in In-
terlaken/Schweiz, 18.-24. Sept. 1 991.
ZtuttvtnR, H., VossnL, G., FRöHtIcH. W.D.(1989): Antizipatorische und ereig-
nisbezogene Veränderungen cler Schlagfrecluenz des Herzens als Indikatoren der
Aufinerksamkeitsregulation. Arch" Psychol. 14I, 251-272.

100 7 ANHANG
der in dieser Arbeit untersuchten oder in Zitaten aufschei-
7 Anhang
Tabelle 222 Yerzeichnis
nenden Tierarten"
Damhirsch
Delawarenrnöwe
Dickhornschaf
trlch
Florida-Wal clkaninchen
Gemse
Grünfink
Hühner
Karil:u
Maultierhirsch
Moschusochse
Reh
Rentier
Rothirsch
Rotschenkel
Segelflosser
Silbennöwe
Thornsongazelle
Tupaja
Wapiti
Weißwedelhirsch
Wolf
Dama datna
Lat'us delautarensis
Oais cartadensis canadensis
Alces alces
S3llui,lagtrs fl o ridanus
Rupricapra rupricapra
Chlori,s ch,loris
Gallus spec.
R arr.g ifer tarandus cari.bou
Odocoilu.c hemionus
Oaibos moschatus
Capreolus cap'reolus
Rart,gif er tarand,us tarandus
Ceruus elaphu,s
Tri,rt,ga totanus
Pterophyllum eimicki
Larus argentatus
Gazella thomsomi
Tupaia belangeri
Ceruus elaphus
O docoileus rsirginianus
Canis lupus
fallow deer
ring-billed gull
bighorn sheep
moose (elk)
eastern cottontail rabbit
chamois
greenfinch
fowl
caribou
mule deer
muskox
roe deer
reincleer
red deer
redshank
angel fish
herring gull
Thomson's gazelle
tree shrew
elk (wapiti)
white-tailed deer
gray wolf
Die Nomenklatur dieser Tabelle hält sich bei den Vögeln an Ptrtnnsom et al. (1976),
bei den Säugetieren an GRzTMEK(1968).

'(pU^+oU rutaq rnu ua3
-urrclg pun ue{rog 'ua1qn5 'uauqrO 'rllor}uo}I 'ro11u;1o) PII'\t}oU Pun -qeg Ieq uesle^a
-suall"rlre1 aPuarutuol.Io^\ ]Iazqlns:r.\ 'laP PuerqPA\ uals8gnuq arg :67 EunpllqqY
ualqns
suam{a0
«--r(=EJ
ueflIa0
*[- §1 \i v \
\\\--\7t4
ulallnqrs
Kfl UH\ iJ '--:-) \r\ b/
$nT rep u-r" ?Irq'+g 'rluox'}{"JIo }r-ul"'
r\ {(§ ,ß-)» Ct\§) ä-"5
-2")<_- ) .J
flOII
){ö/\__.__.- \
Vüaqarz
r:\'.r'\i )\/ Y
-+'/\f
-.\-- \ t\)^)-i
t. 1
ii' +, r1\ llr/,''\ 'r"-- L' l\ , ,/'i/
,r----:h\_ -.1&
ueFqrs
\) \ I\ , )t)1,../ tJ\lt:./t.- -/. I
üasY lltu"'ua?urrd5
üe{3ogt .\\\
^ßv\1;'^-\--/
z1"sq"ül"H
\iNlj\ 2\ ,- //lq lL'I r-'
V
üerr€qJS
Eaqarz rm-"
§M>ilU
eflorluox 'roHeJIo
ursrills'n uaqels
I lai,' lVl
) '-\"\ .-_--___\
i'\ >l^I
uaBa1ut11
tlloraBura"'
-4-=\\ft\\YJ /\- ll
ry-/
auqeuJn€ra11nfuelun 1do11 1iul"'
,--=- ,' -.1,,
uaqrer;doY
sa8rlsuog
uazlnd
üa1[Bqra^
-+Jo;[Eox
uasY
It t,t + i/ ltll t'. t' tYl / ',
.) --)'- ,r) F!
tt/
uaBarl
.r+uox'+)tsflo
/'qnesBrrnflIep
uai{alsJnY
3rrn3a,rtaq1lo.4
fuaqalg
TOI
uaBarl
uazlrd ?r-m"
F<-u)\-J _§: i \s

102 7 ANHANG
PROT.-NR.:DATUI{:
NA\IE: T1€ O
Ä (Asen)Fü (Fürterua6 )
Hu ( KuDen /Tr (Trinken)
L (Lregenl i 1 mit \\'red"rläuenS (S!.beni t - ohne \\'rederl;uenSs (Stehen uud Sichern)OK iOlIakt. Xonlr-)
Z (Ziehen)T iTroll)Fl ( Fiuchl )
1 ! zur StörquelieI - *et vom 5lorreEL r inrrunciivtduell
Sch ( Schütteln)P (Putzen)KR i Xoplreiben )
Abbildung 50: Beispiel eines Verhaltensprotokolls für die Ausrnertung der Videoauf-
zeichnungen.
Verhaiaen tl hrzert(Besinn
lJauerIsec I
Bemerkungen
5J lo.r+lLt 5Y5 l2
\\ U<c I trr+ 3
r'I 5r) Jl 3r
T-125,2,)
l/J +l 3Y.+ z
li ?c l)IL
13<,JJ /Y
rl Sf)/L t7s5
)q.J 3
A(rr ),1LV
qJ 51i ..-,
r; S/t' ZC
(5 a/ l6A, JI t4(5 Sr ?
r, 5g lcs5 5 2e, +i r5 55r zc fSC:K 53': T<) c5 5(
! erhalten a arzelti Beginn
l) a uerlseci
Bemerkuugen
a
5f" 3b
5- 2,)L q2
5t 55r8 l7
.1- 15 Z}.J Qt lL
at_ 3?5 Y3 (. (,)
a 5;te ?
5 <7 Z
AK_l
-)) 37 5i t(<) >)-, )
+ IC tCK
l\+ l")z15- ?rJ) r3
OK u1- +
55 2
/ri .i'-i"n s'r /-ä
?r.*. o6er'Jä.lb-t
-
\ V.
53" .,1§
trr'-,< !tccsS I(tr)iL)LZt(!)
3
T- 36
55 3t 238cSss t)
Y5 zz35 ,3 ''
2+
lü"s
/<.)r

= ,:'
=
=
r)
o
05
]S
00I
06
0lo0r
00r
0
LT
L6g6
co
o9
,tr)
E
ia
Z
IIIIZ
II
Z
tö
IIs
9Ih1
OI
L
6
IIc
b
I8
v
I6
6
OI
SI
ZT
8
tIe
I9
1
I_
to
0
o
n
?
6
zt8
8
9
L
8
€
II9
IZ
IIL
L
6
OI
OJ
t€t6OZ
(x)I
88
tto
tte
J6
.16
C?
tto
LIt»o
[€ttL}L8
ۤ)
EL
LA
0a
D?
,c
tt
IL
tj
I
ti,-
i
:
i
:.
-
T
:
:
:
:
;ii.
Z
Z
s
,t,tII{I6
OI
8
8
8
8
?t9IttLILI6I6I6I6I9IstrIZT
?tAI17,
?t9ILIoz
9Z
90rLITL6ct
ct
t9z-t0,c-c
Z9LOIII TI6II6tr6€r6PI6?T6PI8€r€ort6t8966 7,t
68r9€r6IIOI 8I2Zlzt 9r8t 6r
oooo
LT
CC
0t)!e8
!et806
e8
t6
AJ,
t8
IItz
Z
IZ
IZ
ttZ
t
IZ
8I9I9tqt
6I9I8III8tIZ
LIVI
9
v
6
6
IIZ
L
8
o
9
6
TI
IIZI
ZI
1I
ZI
I TII 9I
0tSI
r9lZI
, II
.I€
'0t,62
'82.LZ
'97,
'92
'bz
'Ee
.ZZ
.IZ
'o('6r'8I.LI
'9I'gr'br'8I.ZI
.II'0I'6
'8
').
'9
'9
'v
.Z
.I
SgAArDuL u?ru crnleradual
STI.{AIDUI ulu {rnl"radrual
Saxou ulul q
Jn]?JaduraI
raqua^oNIaqol{oraquraldes
(lrqrN : :ua8a5 O) saErlsuoS:S
iluazo:4 ur 3un11o.tnafl:B la>pe1spur16-r11 l(runurrxetrl lurnurtutry lpa.ta.1a11rursa8
-eJ) D. ut .rnlu.radtua; 'pll^arlaE rnJ runErllazsqlnsra^ IIII uelePBrullll :82 allaqBl
t0T

104 7 ANHANG
im Versuchszeitraum für Rotwild. Temperatril in "Cl (Ta-
Maximum); \\I:\Vindstärke; B:Beu,ölkung in Prozent;
Nebel: + Schneefall)
Tabelle 24: Klimadaten
gesmittelwert; Minimum;
S:Sonstiges (O Regenl :
Januar Februar MärzTemperatur
c min mar w B S
Temperatur
i min mar w B S
Temperatur
i min rnar w B S
1.
2.
4.
5.
6.
7.
8.
9.
10.
11.
t').13.
74.
15.
16.
17.
18.
19.
20.
27.
22.
24.
25.
26.
27.
28.
29.
30.
31.
427306961361114764-1 I3166383062-t452774105472-74
-4 -8 -1-4 -8 -1-5 -8 -2-4 -10 4
-3-71-6 -9 -3-4 -6 -2-4 -7 -1-1-20
a-221-1 4
-2-51-1 -2 -l
1 -1 3
-1-420-33
-8 -12 -3
3
3
1
a
2
1
n
1
,1
2
2
1
2
1
1
1
1
1
1
1
3
1
2
2
,4
3
100
67
90
43
67
63
87
53
93
80
83
o7
97
37
0
3
3
0
0
100
83
97
100
67
30
63
100
E;93
)J
10
o
=oo
o
o
=
-10 -14 -6-8 -12 -3-7 -11 -2-7 -11 -3-8 -13 -4
-13 -1.7 -9-7 -9 -4-6 -10 -2-3 -5 -1-2-62
2-26-5 -8 -1-2 -4 -1-3-50-4-80-2-630-34
-2-94-3-71-2-62-1-52
1-474 -2 11
8 5 ll1-364092-1.60 -5 3
2
2
2
3
2
2
2
1
3
2
4
2
2
1
1
1
1
2
2
II2
2
2
o
67
50
43
IU
100
47
93
100
90
97
47
43
97
43
3
73
80
43
50
97
53
50
60
*=
*=
1
2
2
8
7
6
8
9
10
10
10
10
t215
72
11
13
11
5
3
5
2
,t)
-1 2
14
15131524? 1'
41039594 11
2731 16
I lt
, t-
4 ltl4165747 16
10 27
6 19
6 16
7197164715
03033 10
3
3
2
2
2
2
3
1
1
1
1
2
1
I
2
2
2
3
2
2
1
2
2
2
2
3
2
93
100
100
100
100
97
100
a7
97
83
100
93
3
7
'l
40
6L)
70
.J
33
90
6T
97
100
97
50
100
100
o
o
o
o




















