
Efficient gene transfer into human primary blood lymphocytes by surface-engineered lentiviral...
10
doi:10.1182/blood.V99.7.2342 2002 99: 2342-2350 Marielle Maurice, Els Verhoeyen, Patrick Salmon, Didier Trono, Stephen J. Russell and François-Loi?c Cosset polypeptide activating - surface-engineered lentiviral vectors that display a T cell Efficient gene transfer into human primary blood lymphocytes by http://bloodjournal.hematologylibrary.org/content/99/7/2342.full.html Updated information and services can be found at: (523 articles) Gene Therapy Articles on similar topics can be found in the following Blood collections http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requests Information about reproducing this article in parts or in its entirety may be found online at: http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprints Information about ordering reprints may be found online at: http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtml Information about subscriptions and ASH membership may be found online at: Copyright 2011 by The American Society of Hematology; all rights reserved. American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036. Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the For personal use only. by guest on May 30, 2013. bloodjournal.hematologylibrary.org From
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Efficient gene transfer into human primary blood lymphocytes by surface-engineered lentiviral...

doi:10.1182/blood.V99.7.23422002 99: 2342-2350
Marielle Maurice, Els Verhoeyen, Patrick Salmon, Didier Trono, Stephen J. Russell and François-Loi?c Cosset polypeptide
activating−surface-engineered lentiviral vectors that display a T cellEfficient gene transfer into human primary blood lymphocytes by
http://bloodjournal.hematologylibrary.org/content/99/7/2342.full.htmlUpdated information and services can be found at:
(523 articles)Gene Therapy �Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the
For personal use only. by guest on May 30, 2013. bloodjournal.hematologylibrary.orgFrom

GENE THERAPY
Efficient gene transfer into human primary blood lymphocytes bysurface-engineered lentiviral vectors that display a T cell–activating polypeptideMarielle Maurice, Els Verhoeyen, Patrick Salmon, Didier Trono, Stephen J. Russell, and Francois-Loıc Cosset
In contrast to oncoretroviruses, lentivi-ruses such as human immunodeficiencyvirus 1 (HIV-1) are able to integrate theirgenetic material into the genome of non-proliferating cells that are metabolicallyactive. Likewise, vectors derived fromHIV-1 can transduce many types of non-proliferating cells, with the exception ofsome particular quiescent cell types suchas resting T cells. Completion of reversetranscription, nuclear import, and subse-quent integration of the lentivirus ge-nome do not occur in these cells unlessthey are activated via the T-cell receptor(TCR) or by cytokines or both. However,to preserve the functional properties of
these important gene therapy target cells,only minimal activation with cytokines orTCR-specific antibodies should be per-formed during gene transfer. Here wereport the characterization of HIV-1–de-rived lentiviral vectors whose virion sur-face was genetically engineered to dis-play a T cell-activating single-chainantibody polypeptide derived from theanti-CD3 OKT3 monoclonal antibody. In-teraction of OKT3 IgGs with the TCR canactivate resting peripheral blood lympho-cytes (PBLs) by promoting the transitionfrom G 0 to G1 phases of the cell cycle.Compared to unmodified HIV-1–basedvectors, OKT3-displaying lentiviral vec-
tors strongly increased gene delivery infreshly isolated PBLs by up to 100-fold.Up to 48% transduction could be ob-tained without addition of PBL activationstimuli during infection. Taken together,these results show that surface-engi-neered lentiviral vectors significantly im-prove transduction of primary lympho-cytes by activating the target cells.Moreover these results provide a proof ofconcept for an approach that may haveutility in various gene transfer applica-tions, including in vivo gene delivery.(Blood. 2002;99:2342-2350)
© 2002 by The American Society of Hematology
Introduction
Efficient gene transfer into T lymphocytes may allow the treatmentof several genetic dysfunctions of the hematopoietic system, suchas severe combined immunodeficiency,1,2 and the development ofnovel therapeutic strategies for diseases such as cancers andacquired immunodeficiency syndrome.3 To reach this goal, it isessential to preserve the functional properties of the transducedcells, that is, their capacity to appropriately react on stimulation ofthe immune system. Therefore, only minimal ex vivo manipulationof the cells should be performed during gene transfer becausegrowth factor/cytokine combinations, used to force cell prolifera-tion, often lead to skewing of cell populations and may alter theirability to respond to novel antigens.4,5 Ideally, gene delivery shouldbe best achieved in vivo to minimize contacts of target cells withnonphysiologic cell culture reagents that may induce differentia-tion or proliferation.
Vectors derived from retroviruses are probably among the mostsuitable tools to achieve a long-term gene transfer because they allowstable integration of a transgene and its propagation in daughter cells. Todate, vectors derived from oncoretroviruses such as murine leukemiaviruses (MLVs) have been widely used for gene transfer into human Tcells,6 essentially because of the simplicity of their manipulation.Perhaps one of the most important drawbacks associated with the use of
such vectors is their inability to transduce nonproliferating target cells.Indeed, following internalization of the vector into the target cellcytoplasm and reverse transcription, transport of the preintegrationcomplex to the nucleus requires the breakdown of the nuclear mem-branes during mitosis.7,8This provides a formidable barrier to the use ofMLV-based vectors in the many gene therapy protocols for which targetcells are quiescent or for which induction of cell proliferation is to beavoided. Thus, the recent emergence of lentiviral vectors may provide avaluable alternative to overcome this problem owing to the lentivirusmechanism that allows mitosis-independent nuclear import of thepreintegration complex and infection of nonproliferating cells.9-11
Several studies have now established the capacity of these vectorsderived from human immunodeficiency virus 1 (HIV-1) to transducevarious types of nonproliferating cells both in vitro and in vivo.12
However, some cell types that are important gene therapy targets arerefractory to gene transfer with lentiviral vectors, despite the most recentimprovements brought into their structures.13-16This includes, in particu-lar, early progenitor hematopoietic stem cells in G0,17 monocytes,18,19
and resting T lymphocytes.14 That the parental virus, HIV-1, can enterinto restingT lymphocytes but does not replicate,20-24has been attributedto multiple post-entry blocks. This includes, in particular, (1) defects ininitiation and completion of the reverse-transcription process,20,23-25(2)
From the Laboratoire Vectorologie Retrovirale et Therapie Genique, Unite deVirologie Humaine, U412 INSERM, IFR 74 and ENS de Lyon, France;Department of Genetics and Microbiology, Faculty of Medicine, University ofGeneva, Geneva, Switzerland; and the Molecular Medicine Program, MayoClinic, Rochester, Minnesota.
Submitted July 25, 2001; accepted November 9, 2001.
Supported by Agence Nationale pour la Recherche contre le SIDA (ANRS), theEuropean Community (QLK3-1999-00859), AFIRST, Association Francaisecontre les Myopathies (AFM), Association pour la Recherche contre le Cancer(ARC), Centre National de la Recherche Scientifique (CNRS), and Institut
National de la Sante Et de la Recherche Medicale (INSERM). M.M. and E.V.contributed equally to this work.
Reprints: Francois-Loıc Cosset, Laboratoire de Vectorologie Retrovirale etTherapie Genique, INSERM U412, Ecole Normale Superieure de Lyon, 46Allee d’Italie, 69364 Lyon Cedex 07, France; e-mail: [email protected].
The publication costs of this article were defrayed in part by page chargepayment. Therefore, and solely to indicate this fact, this article is herebymarked ‘‘advertisement’’ in accordance with 18 U.S.C. section 1734.
© 2002 by The American Society of Hematology
2342 BLOOD, 1 APRIL 2002 z VOLUME 99, NUMBER 7
For personal use only. by guest on May 30, 2013. bloodjournal.hematologylibrary.orgFrom

lack of adenosine triphosphate-dependent nuclear import,9,26 and (3)lack of integration of the proviral genome.27Low levels of nucleotides inthe resting cells do not entirely explain the restricted HIV-1 replicationbecause artificially raising intracellular nucleotide pools increasedreverse-transcription products but not the level of productive infection.28
However, it was recently reported that inducing the resting T cells toenter into the G1b phase of the cell cycle by stimulation through theT-cell receptor (TCR) and CD28 costimulation receptor, using anti-CD3plus anti-CD28 antibodies, was sufficient to render the cells susceptibleto HIV-1 infection and replication.25 Moreover, exposing T cells tocytokines that do not trigger cell division could render thempermissive to transduction with HIV-1 vectors.29 These findingssuggest that partial activation of resting T cells is sufficient for genetransfer by HIV-1–derived vectors and that DNA synthesis ormitosis of these cells is not necessary.
Here we sought to refine the structure of lentiviral vectorparticles to overcome their inability to transduce nonactivated Tcells. Because extensive activation of resting T cells with combina-tions of stimulating factors should be avoided ex vivo and may notbe possible to achieve in vivo, we have designed novel lentiviralvector particles that transiently provide a minimal stimulus to thetarget cells during gene delivery. As a proof of concept, weestablish here that lentiviral vectors whose virion surface wasengineered to display a TCR-activating polypeptide could effi-ciently transduce primary resting T lymphocytes without additionof soluble T-cell activation stimuli in the cell culture medium.HIV-1–derived vectors were copseudotyped with an MLV-derivedchimeric envelope glycoprotein fused to an anti-CD3 single-chainantibody variable fragment (scFv) and the vesicular stomatitis virusG (VSV-G) glycoprotein. This anti-CD3 scFv was derived from theOKT3 monoclonal antibody, which activates the TCRs. Impor-tantly, the surface-modified lentiviral vectors specifically recog-nized CD3 and triggered activation of freshly isolated peripheralblood T cells. This minimal stimulation was sufficient to allowgene transfer in up to 48% of the lymphocytes, that is, 100-foldmore than the performance of unmodified lentiviral vectors innonactivated T cells.
Materials and methods
Cells
The 293T human embryo kidney cell line was grown in Dulbecco modifiedEagle medium (DMEM; Life Technologies, Cergy Pontoise, France)supplemented with 10% fetal calf serum (FCS). The Jurkat human Tleukemia cell line was grown in RPMI 1640 (Life Technologies) supple-mented with 10% FCS. Human peripheral blood mononuclear cells(PBMCs) were separated from fresh blood of healthy donors using aFicoll-Hypaque/Percoll gradient (Amersham Pharmacia Biotech, Orsay,France). Peripheral blood lymphocytes (PBLs) were enriched from thePBMC fraction by an overnight plastic adherence at 37°C to removeadherent monocytes. The nonadherent cells were resuspended in RPMI1640 medium supplemented with 10% FCS. PBLs were checked for CD3marker expression (75%-85% were CD31) and the percentage of cells inthe different phases of the cell cycle was determined by flow cytometry($ 99% were in G0/G1).
Packaging and vectors constructs
The pCMVD8.230 and pCMVD8.9131 HIV-1 packaging plasmids have beendescribed elsewhere. The HPPT-EF1a-GFP vector contains the EF1ainternal promoter driving theGFP (green fluorescent protein) gene and thecPPT/CTS (central poly-purine track and central termination sequence)cis-acting signal, which stimulates nuclear import.15,16 The pCMVD8.2
packaging-deficient construct contains all HIV-1 genes exceptenv, andpCMVD8.91 lacks the HIV-1 accessory genesvif, vpr, vpu, andnef. TheCMV1intron plasmid, containing the MLV packaging sequences (S.Chapel-Fernandes and F-L. Cosset, unpublished material, 1996), and theMFG-GFP plasmid, encoding an MLV-based vector containing theGFPmarker gene, were used to generate MLV-derived vectors. phCMV-G32
encodes the VSV-G glycoprotein under control of the human cytomegalo-virus (hCMV) promoter and rabbitb-globin intron II and polyadenyla-tion sequences.
Envelope glycoprotein constructs
ThescFvgene encoding a polypeptide that binds the human CD3 complexof TCRs was derived from the OKT3 monoclonal antibody33 and wasdescribed elsewhere.34 The OKT3 scFv complementary DNA (cDNA) wasfused in the 4070A(amphotropic) MLVenvgene,35at a position correspond-ing to the first amino acid of the SU (surface) subunit of the amphotropicMLV envelope glycoprotein, as described previously.34 To reduce sterichindrance between the scFv polypeptide and the amphotropic envelopeglycoprotein, a peptide linker, 7 amino acids long, was inserted betweenthese 2 domains.34,36 The resulting chimeric glycoprotein was namedOKT3SU (Figure 1). The SUx mutation,37 which inhibits furin-mediatedcleavage of the MLV glycoprotein in the producer cells, was inserted in theOKT3SU construct, thus resulting in a second chimera, named OKT3SUx.This was achieved by replacing the Lys-Tyr-Lys-Arg furin cleavage site bythe IPe-Glu-Gly-Arg peptide.37 Both the OKT3SU and OKT3SUx chimeraswere expressed using the phCMV-G expression vector backbone.32
Production of retroviral vectors
Pseudotyped HIV-1–derived vectors were generated as previously de-scribed30 by transient transfection of 293T cells. Then 8.6mg of theHPPT-EF1a-GFP vector construct, 8.6mg of either pCMVD8.2 orpCMVD8.91 packaging constructs, and 2.7mg of the envconstruct, forexample, the VSV-G–expressing construct (phCMV-G) were used tocotransfect 293T cells. When HIV-1–derived vectors were copseudotypedwith VSV-G and OKT3SU or OKT3SUx chimeric glycoproteins, anequimolar quantity of envelope-expressing plasmids was used. A similarprotocol was used to produce MLV-derived vectors with the CMV1intronand the MFG-GFP expressing constructs. Plasmid DNAs were transfectedinto 2.63 106 293T cells seeded the day before in 10-cm diameter platesusing the calcium-phosphate transfection system according to manufactur-er’s recommendations (Life Technologies). The medium (12 mL/plate) wasreplaced 16 hours after transfection, and supernatant was harvested 24hours later, low-speed centrifuged (2000 rpm for 5 minutes at 4°C), filteredthrough 0.45-mm pore-sized membranes, and directly used in thedifferent assays.
Figure 1. Schematic representation of chimeric retroviral glycoproteins display-ing an anti-CD3 scFv. A cDNA encoding an scFv derived from the OKT3 monoclonalantibody raised against the human CD3 was fused to the amino-terminal end of the4070A-MLV env gene, at a position corresponding to the first codon of the SUenvelope subunit. A spacer, 7 amino acids long, was inserted between the scFvpolypeptide and the envelope glycoprotein to minimize steric hindrances. The SU/TMcleavage site was inactivated by replacing the Lys-Tyr-Lys-Arg wild-type amino acidsequence by the IPe-Glu-Gly-Arg sequence. L indicates leader-signal peptide;ligand, anti-CD3 scFv polypeptide; SU, surface envelope subunit; TM, transmem-brane envelope subunit.
T-CELL TRANSDUCTION BY SURFACE-MODIFIED HIV VECTORS 2343BLOOD, 1 APRIL 2002 z VOLUME 99, NUMBER 7
For personal use only. by guest on May 30, 2013. bloodjournal.hematologylibrary.orgFrom

Infection assays
To determine transduction efficiency and infectious titers of HIV-1–derivedvectors, 293T target cells were seeded at a density of 23 105/well in 6-wellplates 1 day before transduction. Serial dilutions of vector preparationswere added to the 293T cells. The percentage of GFP1 cells was determinedby FACS analysis, following homogenization of transduced cells in trypsinand resuspension in phosphate-buffered saline (PBS). The titer was derivedfrom the percentage of GFP1 cells after transduction of 33 105 target cellswith 1 mL viral supernatant. The infectious titers are expressed as 293Ttransducing units (TU/mL).
To infect human primary lymphocytes, 1 mL viral supernatant contain-ing 53 105 to 53 106 TU, as determined on 293T target cells, was addedto 1 to 33 105 lymphocytes suspended in 0.5 mL RPMI supplemented with10% FCS in 24-well plates. Multiplicities of infection (MOIs) weredetermined on proliferating 293T cells and are indicated in all lymphocytetransduction experiments. Human primary lymphocytes were infected inthe same conditions as 293T control cells and infection efficiency wasdetermined by flow cytometry at day 6. For T-cell stimulation and controlinfection assays performed with vectors pseudotyped with VSV-G only,anti-CD3 (HIT3a, Pharmingen) and anti-CD28 (CD28.2, Pharmingen)antibodies were used at a final concentration of 1mg/mL/point. Activationwith soluble anti-CD3/anti-CD28 antibodies was preferred to activationwith immobilized (plastic-coated) antibodies, to provide a true comparisonwith the OKT3SU-displaying virions. This resulted in approximately 30%lower transduction efficiencies than obtained in transduction protocols thatused immobilized OKT3 antibodies (data not shown).4,14,25,28No differ-ences in transduction efficiency were found when comparing OKT3 versusHIT3a anti-CD3 monoclonal antibodies for T-cell activation and infection(data not shown).
For the short exposure of PBLs to virus supernatants, the cells werewashed with RPMI medium 24 hours after infection, resuspended in RPMIcontaining 10% FCS supplemented with or without recombinant interleukin(IL) 2 (rIL-2; 1 ng/mL; R & D Systems, Abingdon, United Kingdom) andmaintained up to day 6 after infection for FACS analysis.
Antibodies, immunoblots, cell surface staining, andbinding assays
Anti-SU (Viromed Biosafety Labs, Camden, NJ) was a goat antiserumraised against the Rausher leukemia virus gp70, used diluted to 1:2000 forWestern blots. Anti-MLV-CA (Viromed Biosafety Labs) was a goatantiserum raised against the Rausher leukemia virus p30 capsid protein(CA), used diluted to 1:10 000 for Western blots. Anti-VSV-G P5D4monoclonal antibody (1mg/mL; Sigma, St Louis, MO) was diluted to1:1000 for Western blots. Anti-HIV-1-CA (p24 capsid) monoclonal anti-body (8mg/mL) was diluted to 1:8000 in immunoblots. For Western blotanalysis, lysates of vector producer cells and virion samples were preparedas previously described.38
Anti-CD3 was used at a 1:125 dilution for cell surface staining. Stainingfor CD25, CD69, and CD71 T-cell activation markers was performedwith phycoerythrin-conjugated anti-CD25, anti-CD69, and anti-CD71antibodies (BD-Pharmingen, Pont de Claix, France) at a 1:25 dilution.Target cells (53 105 cells/point) were incubated with the appropriateantibodies for 45 minutes at 4°C. Fluorescence of living cells wasanalyzed with a fluorescent-activated cell sorter (FACSCalibur, BectonDickinson, Pont de Claix, France).
Supernatant of the 83A25 hybridoma,39 secreting a rat monoclonalantibody against MLV SU, was used undiluted for binding assays. Bindingassays on Jurkat T cells (5-83 105 cells/point) were performed aspreviously described.37 For competition binding assays, Jurkat cells werepreincubated for 45 minutes at 4°C with an anti-CD3 antibody (HIT3a, 1mg/mL, Pharmingen) diluted to 1:100 to saturate TCRs, then exposed tovirus. Cells were then processed as previously described38 except that 10ng/mL mouse IgGs (Sigma) was added with the secondary antibody toinhibit nonspecific antibody recognition.
Cell cycle analysis
Freshly isolated human PBLs (1-33 105 cells) were maintained in RPMIsupplemented with 10% FCS. PBLs were stimulated for different timeperiods after cultivation with 1 mL filtered viral supernatants or withstimulating agents (soluble anti-CD3 or soluble anti-CD28 antibodies).Cells were then centrifuged and fixed in 70% ethanol-30% PBS. The cellswere stored at 4°C until FACS analysis. Just before FACS analysis, thefixed cells were pelleted and resuspended in 1 mL PBS containing 50mgRNase and 10mg/mL propidium iodide. The cells were incubated for 30minutes at 37°C and the cell cycle was then analyzed with a fluorescence-activated cell sorter (FACSCalibur, Becton Dickinson).
Results
Engineering of the viral surface glycoprotein with anOKT3-derived scFv
Previous data from our laboratory have shown that the MLVenvelope glycoprotein can be engineered to display various typesof polypeptides at its amino terminus.40 Because the wild-typeMLV glycoprotein is efficiently incorporated on HIV-1–derivedlentivirus vectors,30 we sought to display a T cell-activatingpolypeptide on HIV-1 vector particles (Figure 1). Thus, an scFvderived from the OKT3 monoclonal antibody, which recognizesand activates the TCR (TCR/CD3/zcomplex, named TCR herein),was fused to the amino terminus of the SU subunit of the MLVenvelope glycoprotein (Figure 1). The position of insertion of theOKT3-derived scFv was chosen to allow its functional display onvirions.37,38 The resulting chimera was named OKT3SU. Becausethe SU and TM subunits of the MLV envelope glycoprotein are notcovalently held together,41 partial dissociation of the chimeric SUfrom the envelope complex, and thus from the viral surface, mayoccur.37 Therefore, a second TCR-activating chimeric glycopro-tein, derived from OKT3SU, was designed to prevent the potentialloss of T cell-activating polypeptides from the viral particles. In theresulting chimera, named OKT3SUx, the cleavage site between theSU and TM envelope subunits was inactivated by substitution witha noncleavable linker peptide37 (Figure 1).
Characterization of surface-engineered lentiviralvector particles
The HIV-1–derived vectors were generated by transient transfec-tions of 293T cells with plasmids encoding the viral core proteins(pCMVD8.2 or pCMVD8.91), the gene transfer vector (HPPT-EF1a-GFP), the VSV-G glycoprotein (phCMV-G), and either ofthe 2 TCR-activating glycoproteins (OKT3SU or OKT3SUx).Coexpression of VSV-G protein with either OKT3SU or OKT3SUxwas necessary to render the viral particles fully infectious (data notshown), consistent with previous results.37 Lentiviral particles,harvested in supernatants of producer cells 2 days after transfec-tion, were concentrated by ultracentrifugation. Incorporation of thechimeric glycoproteins on the virions was assessed by immunoblot-ting. Standardization of the amount of viral particles loaded on gelswas determined using anticapsid antibodies (Figure 2). Immunode-tection of the viral pellets using antibodies against the VSV-G orMLV glycoproteins indicated that the OKT3SU and OKT3SUxchimeras were coincorporated with the VSV-G glycoprotein on thevirions. The OKT3SUx chimeric glycoprotein was less abundant inthe viral pellet than the OKT3SU chimera, indicating a reducedlevel of viral incorporation of the former glycoprotein. Differencesin the electrophoretic mobilities of the OKT3SU and OKT3SUx
2344 MAURICE et al BLOOD, 1 APRIL 2002 z VOLUME 99, NUMBER 7
For personal use only. by guest on May 30, 2013. bloodjournal.hematologylibrary.orgFrom
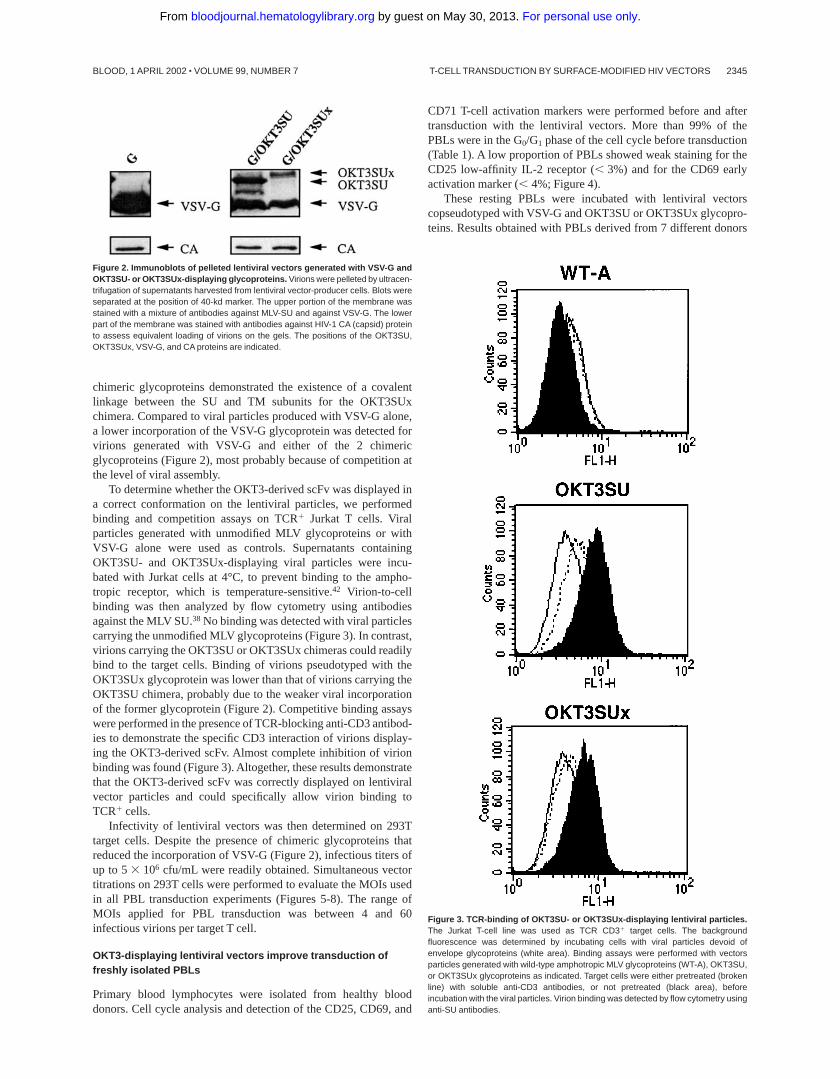
chimeric glycoproteins demonstrated the existence of a covalentlinkage between the SU and TM subunits for the OKT3SUxchimera. Compared to viral particles produced with VSV-G alone,a lower incorporation of the VSV-G glycoprotein was detected forvirions generated with VSV-G and either of the 2 chimericglycoproteins (Figure 2), most probably because of competition atthe level of viral assembly.
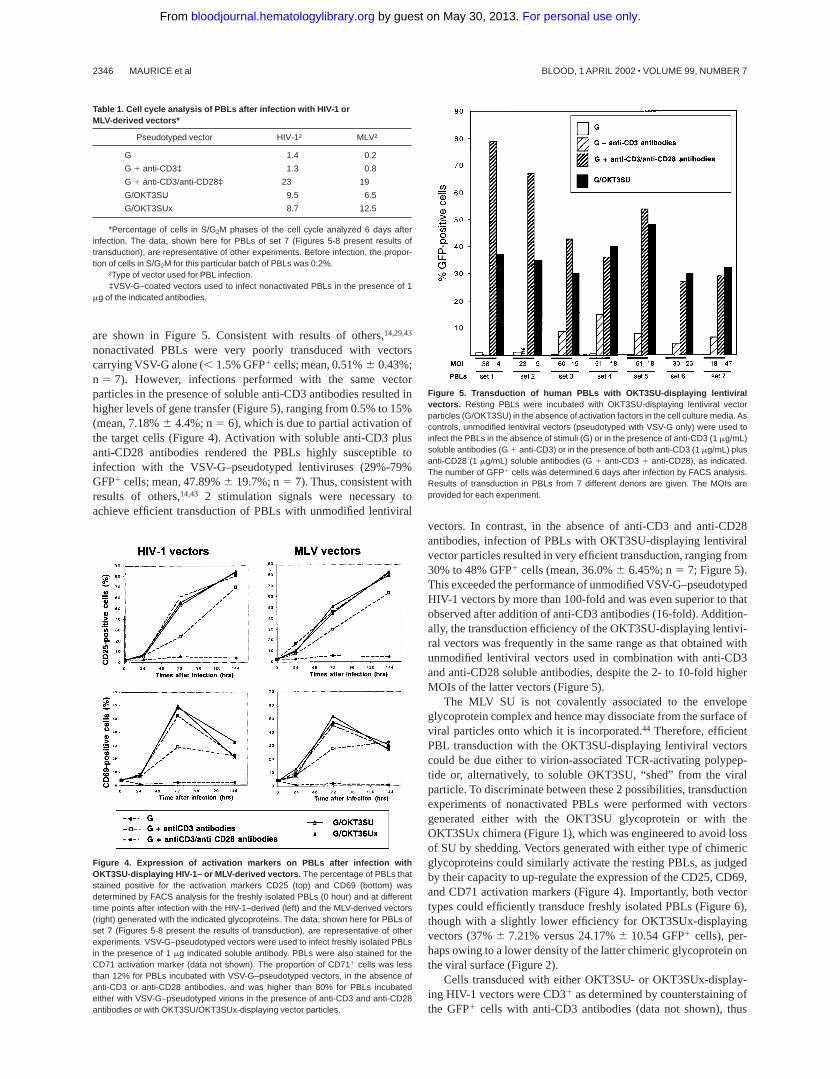
To determine whether the OKT3-derived scFv was displayed ina correct conformation on the lentiviral particles, we performedbinding and competition assays on TCR1 Jurkat T cells. Viralparticles generated with unmodified MLV glycoproteins or withVSV-G alone were used as controls. Supernatants containingOKT3SU- and OKT3SUx-displaying viral particles were incu-bated with Jurkat cells at 4°C, to prevent binding to the ampho-tropic receptor, which is temperature-sensitive.42 Virion-to-cellbinding was then analyzed by flow cytometry using antibodiesagainst the MLV SU.38 No binding was detected with viral particlescarrying the unmodified MLV glycoproteins (Figure 3). In contrast,virions carrying the OKT3SU or OKT3SUx chimeras could readilybind to the target cells. Binding of virions pseudotyped with theOKT3SUx glycoprotein was lower than that of virions carrying theOKT3SU chimera, probably due to the weaker viral incorporationof the former glycoprotein (Figure 2). Competitive binding assayswere performed in the presence of TCR-blocking anti-CD3 antibod-ies to demonstrate the specific CD3 interaction of virions display-ing the OKT3-derived scFv. Almost complete inhibition of virionbinding was found (Figure 3). Altogether, these results demonstratethat the OKT3-derived scFv was correctly displayed on lentiviralvector particles and could specifically allow virion binding toTCR1 cells.
Infectivity of lentiviral vectors was then determined on 293Ttarget cells. Despite the presence of chimeric glycoproteins thatreduced the incorporation of VSV-G (Figure 2), infectious titers ofup to 53 106 cfu/mL were readily obtained. Simultaneous vectortitrations on 293T cells were performed to evaluate the MOIs usedin all PBL transduction experiments (Figures 5-8). The range ofMOIs applied for PBL transduction was between 4 and 60infectious virions per target T cell.
OKT3-displaying lentiviral vectors improve transduction offreshly isolated PBLs
Primary blood lymphocytes were isolated from healthy blooddonors. Cell cycle analysis and detection of the CD25, CD69, and
CD71 T-cell activation markers were performed before and aftertransduction with the lentiviral vectors. More than 99% of thePBLs were in the G0/G1 phase of the cell cycle before transduction(Table 1). A low proportion of PBLs showed weak staining for theCD25 low-affinity IL-2 receptor (, 3%) and for the CD69 earlyactivation marker (,4%; Figure 4).
These resting PBLs were incubated with lentiviral vectorscopseudotyped with VSV-G and OKT3SU or OKT3SUx glycopro-teins. Results obtained with PBLs derived from 7 different donors
Figure 3. TCR-binding of OKT3SU- or OKT3SUx-displaying lentiviral particles.The Jurkat T-cell line was used as TCR CD31 target cells. The backgroundfluorescence was determined by incubating cells with viral particles devoid ofenvelope glycoproteins (white area). Binding assays were performed with vectorsparticles generated with wild-type amphotropic MLV glycoproteins (WT-A), OKT3SU,or OKT3SUx glycoproteins as indicated. Target cells were either pretreated (brokenline) with soluble anti-CD3 antibodies, or not pretreated (black area), beforeincubation with the viral particles. Virion binding was detected by flow cytometry usinganti-SU antibodies.
Figure 2. Immunoblots of pelleted lentiviral vectors generated with VSV-G andOKT3SU- or OKT3SUx-displaying glycoproteins. Virions were pelleted by ultracen-trifugation of supernatants harvested from lentiviral vector-producer cells. Blots wereseparated at the position of 40-kd marker. The upper portion of the membrane wasstained with a mixture of antibodies against MLV-SU and against VSV-G. The lowerpart of the membrane was stained with antibodies against HIV-1 CA (capsid) proteinto assess equivalent loading of virions on the gels. The positions of the OKT3SU,OKT3SUx, VSV-G, and CA proteins are indicated.
T-CELL TRANSDUCTION BY SURFACE-MODIFIED HIV VECTORS 2345BLOOD, 1 APRIL 2002 z VOLUME 99, NUMBER 7
For personal use only. by guest on May 30, 2013. bloodjournal.hematologylibrary.orgFrom
are shown in Figure 5. Consistent with results of others,14,29,43
nonactivated PBLs were very poorly transduced with vectorscarrying VSV-G alone (,1.5% GFP1 cells; mean, 0.51%6 0.43%;n 5 7). However, infections performed with the same vectorparticles in the presence of soluble anti-CD3 antibodies resulted inhigher levels of gene transfer (Figure 5), ranging from 0.5% to 15%(mean, 7.18%6 4.4%; n5 6), which is due to partial activation ofthe target cells (Figure 4). Activation with soluble anti-CD3 plusanti-CD28 antibodies rendered the PBLs highly susceptible toinfection with the VSV-G–pseudotyped lentiviruses (29%-79%GFP1 cells; mean, 47.89%6 19.7%; n5 7). Thus, consistent withresults of others,14,43 2 stimulation signals were necessary toachieve efficient transduction of PBLs with unmodified lentiviral
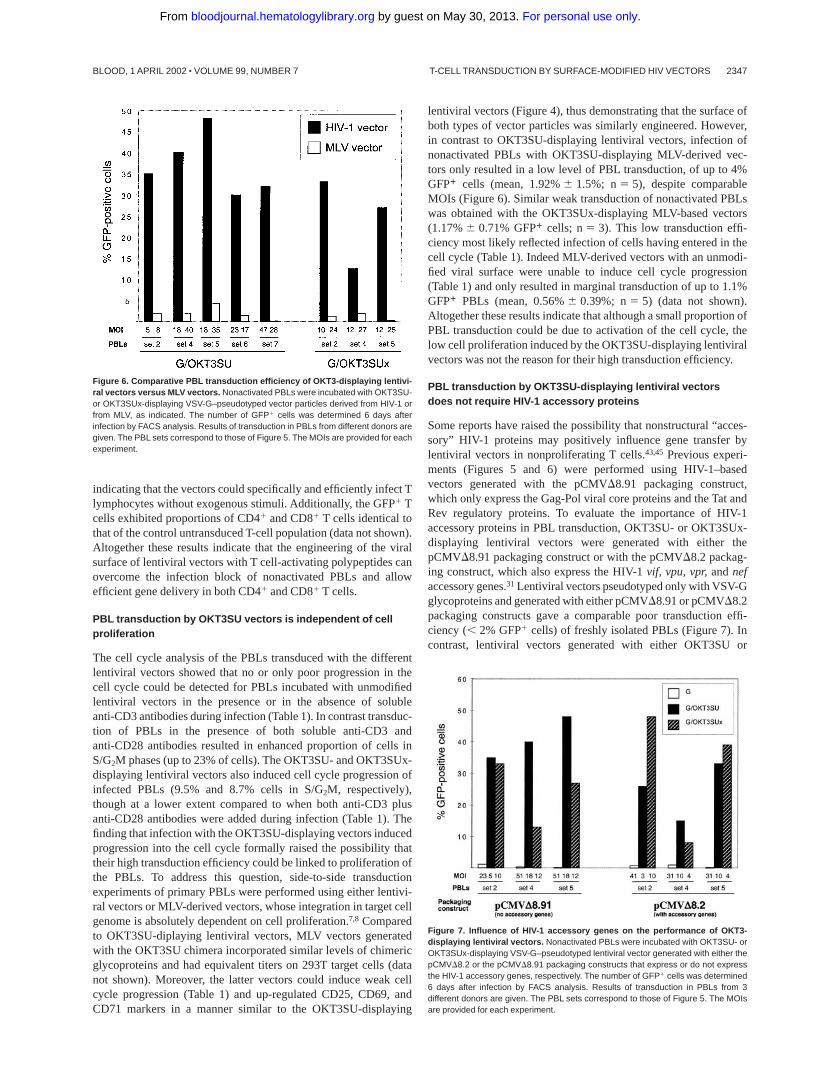
vectors. In contrast, in the absence of anti-CD3 and anti-CD28antibodies, infection of PBLs with OKT3SU-displaying lentiviralvector particles resulted in very efficient transduction, ranging from30% to 48% GFP1 cells (mean, 36.0%6 6.45%; n5 7; Figure 5).This exceeded the performance of unmodified VSV-G–pseudotypedHIV-1 vectors by more than 100-fold and was even superior to thatobserved after addition of anti-CD3 antibodies (16-fold). Addition-ally, the transduction efficiency of the OKT3SU-displaying lentivi-ral vectors was frequently in the same range as that obtained withunmodified lentiviral vectors used in combination with anti-CD3and anti-CD28 soluble antibodies, despite the 2- to 10-fold higherMOIs of the latter vectors (Figure 5).
The MLV SU is not covalently associated to the envelopeglycoprotein complex and hence may dissociate from the surface ofviral particles onto which it is incorporated.44 Therefore, efficientPBL transduction with the OKT3SU-displaying lentiviral vectorscould be due either to virion-associated TCR-activating polypep-tide or, alternatively, to soluble OKT3SU, “shed” from the viralparticle. To discriminate between these 2 possibilities, transductionexperiments of nonactivated PBLs were performed with vectorsgenerated either with the OKT3SU glycoprotein or with theOKT3SUx chimera (Figure 1), which was engineered to avoid lossof SU by shedding. Vectors generated with either type of chimericglycoproteins could similarly activate the resting PBLs, as judgedby their capacity to up-regulate the expression of the CD25, CD69,and CD71 activation markers (Figure 4). Importantly, both vectortypes could efficiently transduce freshly isolated PBLs (Figure 6),though with a slightly lower efficiency for OKT3SUx-displayingvectors (37%6 7.21% versus 24.17%6 10.54 GFP1 cells), per-haps owing to a lower density of the latter chimeric glycoprotein onthe viral surface (Figure 2).
Cells transduced with either OKT3SU- or OKT3SUx-display-ing HIV-1 vectors were CD31 as determined by counterstaining ofthe GFP1 cells with anti-CD3 antibodies (data not shown), thus
Table 1. Cell cycle analysis of PBLs after infection with HIV-1 orMLV-derived vectors*
Pseudotyped vector HIV-1† MLV†
G 1.4 0.2
G 1 anti-CD3‡ 1.3 0.8
G 1 anti-CD3/anti-CD28‡ 23 19
G/OKT3SU 9.5 6.5
G/OKT3SUx 8.7 12.5
*Percentage of cells in S/G2M phases of the cell cycle analyzed 6 days afterinfection. The data, shown here for PBLs of set 7 (Figures 5-8 present results oftransduction), are representative of other experiments. Before infection, the propor-tion of cells in S/G2M for this particular batch of PBLs was 0.2%.
†Type of vector used for PBL infection.‡VSV-G–coated vectors used to infect nonactivated PBLs in the presence of 1
mg of the indicated antibodies.
Figure 4. Expression of activation markers on PBLs after infection withOKT3SU-displaying HIV-1– or MLV-derived vectors. The percentage of PBLs thatstained positive for the activation markers CD25 (top) and CD69 (bottom) wasdetermined by FACS analysis for the freshly isolated PBLs (0 hour) and at differenttime points after infection with the HIV-1–derived (left) and the MLV-derived vectors(right) generated with the indicated glycoproteins. The data, shown here for PBLs ofset 7 (Figures 5-8 present the results of transduction), are representative of otherexperiments. VSV-G–pseudotyped vectors were used to infect freshly isolated PBLsin the presence of 1 mg indicated soluble antibody. PBLs were also stained for theCD71 activation marker (data not shown). The proportion of CD711 cells was lessthan 12% for PBLs incubated with VSV-G–pseudotyped vectors, in the absence ofanti-CD3 or anti-CD28 antibodies, and was higher than 80% for PBLs incubatedeither with VSV-G–pseudotyped virions in the presence of anti-CD3 and anti-CD28antibodies or with OKT3SU/OKT3SUx-displaying vector particles.
Figure 5. Transduction of human PBLs with OKT3SU-displaying lentiviralvectors. Resting PBLs were incubated with OKT3SU-displaying lentiviral vectorparticles (G/OKT3SU) in the absence of activation factors in the cell culture media. Ascontrols, unmodified lentiviral vectors (pseudotyped with VSV-G only) were used toinfect the PBLs in the absence of stimuli (G) or in the presence of anti-CD3 (1 mg/mL)soluble antibodies (G 1 anti-CD3) or in the presence of both anti-CD3 (1 mg/mL) plusanti-CD28 (1 mg/mL) soluble antibodies (G 1 anti-CD3 1 anti-CD28), as indicated.The number of GFP1 cells was determined 6 days after infection by FACS analysis.Results of transduction in PBLs from 7 different donors are given. The MOIs areprovided for each experiment.
2346 MAURICE et al BLOOD, 1 APRIL 2002 z VOLUME 99, NUMBER 7
For personal use only. by guest on May 30, 2013. bloodjournal.hematologylibrary.orgFrom
indicating that the vectors could specifically and efficiently infect Tlymphocytes without exogenous stimuli. Additionally, the GFP1 Tcells exhibited proportions of CD41 and CD81 T cells identical tothat of the control untransduced T-cell population (data not shown).Altogether these results indicate that the engineering of the viralsurface of lentiviral vectors with T cell-activating polypeptides canovercome the infection block of nonactivated PBLs and allowefficient gene delivery in both CD41 and CD81 T cells.
PBL transduction by OKT3SU vectors is independent of cellproliferation
The cell cycle analysis of the PBLs transduced with the differentlentiviral vectors showed that no or only poor progression in thecell cycle could be detected for PBLs incubated with unmodifiedlentiviral vectors in the presence or in the absence of solubleanti-CD3 antibodies during infection (Table 1). In contrast transduc-tion of PBLs in the presence of both soluble anti-CD3 andanti-CD28 antibodies resulted in enhanced proportion of cells inS/G2M phases (up to 23% of cells). The OKT3SU- and OKT3SUx-displaying lentiviral vectors also induced cell cycle progression ofinfected PBLs (9.5% and 8.7% cells in S/G2M, respectively),though at a lower extent compared to when both anti-CD3 plusanti-CD28 antibodies were added during infection (Table 1). Thefinding that infection with the OKT3SU-displaying vectors inducedprogression into the cell cycle formally raised the possibility thattheir high transduction efficiency could be linked to proliferation ofthe PBLs. To address this question, side-to-side transductionexperiments of primary PBLs were performed using either lentivi-ral vectors or MLV-derived vectors, whose integration in target cellgenome is absolutely dependent on cell proliferation.7,8 Comparedto OKT3SU-diplaying lentiviral vectors, MLV vectors generatedwith the OKT3SU chimera incorporated similar levels of chimericglycoproteins and had equivalent titers on 293T target cells (datanot shown). Moreover, the latter vectors could induce weak cellcycle progression (Table 1) and up-regulated CD25, CD69, andCD71 markers in a manner similar to the OKT3SU-displaying
lentiviral vectors (Figure 4), thus demonstrating that the surface ofboth types of vector particles was similarly engineered. However,in contrast to OKT3SU-displaying lentiviral vectors, infection ofnonactivated PBLs with OKT3SU-displaying MLV-derived vec-tors only resulted in a low level of PBL transduction, of up to 4%GFP1 cells (mean, 1.92%6 1.5%; n5 5), despite comparableMOIs (Figure 6). Similar weak transduction of nonactivated PBLswas obtained with the OKT3SUx-displaying MLV-based vectors(1.17%6 0.71% GFP1 cells; n5 3). This low transduction effi-ciency most likely reflected infection of cells having entered in thecell cycle (Table 1). Indeed MLV-derived vectors with an unmodi-fied viral surface were unable to induce cell cycle progression(Table 1) and only resulted in marginal transduction of up to 1.1%GFP1 PBLs (mean, 0.56%6 0.39%; n5 5) (data not shown).Altogether these results indicate that although a small proportion ofPBL transduction could be due to activation of the cell cycle, thelow cell proliferation induced by the OKT3SU-displaying lentiviralvectors was not the reason for their high transduction efficiency.
PBL transduction by OKT3SU-displaying lentiviral vectorsdoes not require HIV-1 accessory proteins
Some reports have raised the possibility that nonstructural “acces-sory” HIV-1 proteins may positively influence gene transfer bylentiviral vectors in nonproliferating T cells.43,45 Previous experi-ments (Figures 5 and 6) were performed using HIV-1–basedvectors generated with the pCMVD8.91 packaging construct,which only express the Gag-Pol viral core proteins and the Tat andRev regulatory proteins. To evaluate the importance of HIV-1accessory proteins in PBL transduction, OKT3SU- or OKT3SUx-displaying lentiviral vectors were generated with either thepCMVD8.91 packaging construct or with the pCMVD8.2 packag-ing construct, which also express the HIV-1vif, vpu, vpr,andnefaccessory genes.31 Lentiviral vectors pseudotyped only with VSV-Gglycoproteins and generated with either pCMVD8.91 or pCMVD8.2packaging constructs gave a comparable poor transduction effi-ciency (,2% GFP1 cells) of freshly isolated PBLs (Figure 7). Incontrast, lentiviral vectors generated with either OKT3SU or
Figure 6. Comparative PBL transduction efficiency of OKT3-displaying lentivi-ral vectors versus MLV vectors. Nonactivated PBLs were incubated with OKT3SU-or OKT3SUx-displaying VSV-G–pseudotyped vector particles derived from HIV-1 orfrom MLV, as indicated. The number of GFP1 cells was determined 6 days afterinfection by FACS analysis. Results of transduction in PBLs from different donors aregiven. The PBL sets correspond to those of Figure 5. The MOIs are provided for eachexperiment.
Figure 7. Influence of HIV-1 accessory genes on the performance of OKT3-displaying lentiviral vectors. Nonactivated PBLs were incubated with OKT3SU- orOKT3SUx-displaying VSV-G–pseudotyped lentiviral vector generated with either thepCMVD8.2 or the pCMVD8.91 packaging constructs that express or do not expressthe HIV-1 accessory genes, respectively. The number of GFP1 cells was determined6 days after infection by FACS analysis. Results of transduction in PBLs from 3different donors are given. The PBL sets correspond to those of Figure 5. The MOIsare provided for each experiment.
T-CELL TRANSDUCTION BY SURFACE-MODIFIED HIV VECTORS 2347BLOOD, 1 APRIL 2002 z VOLUME 99, NUMBER 7
For personal use only. by guest on May 30, 2013. bloodjournal.hematologylibrary.orgFrom
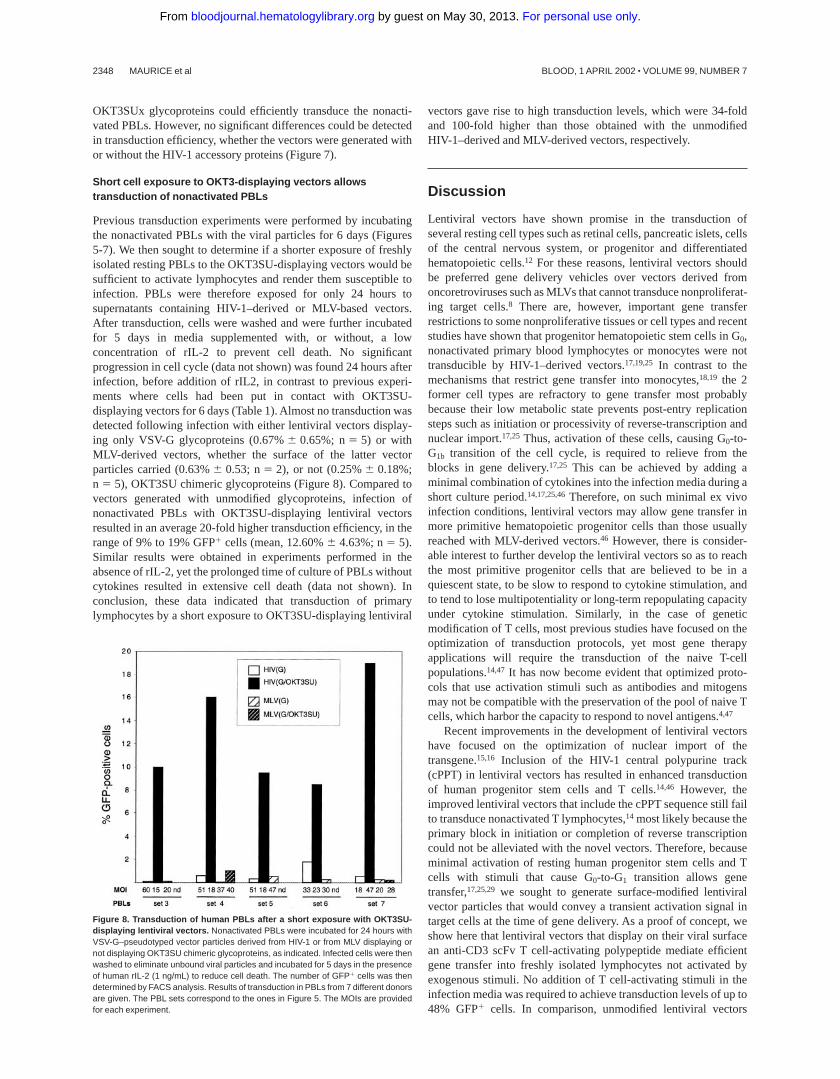
OKT3SUx glycoproteins could efficiently transduce the nonacti-vated PBLs. However, no significant differences could be detectedin transduction efficiency, whether the vectors were generated withor without the HIV-1 accessory proteins (Figure 7).
Short cell exposure to OKT3-displaying vectors allowstransduction of nonactivated PBLs
Previous transduction experiments were performed by incubatingthe nonactivated PBLs with the viral particles for 6 days (Figures5-7). We then sought to determine if a shorter exposure of freshlyisolated resting PBLs to the OKT3SU-displaying vectors would besufficient to activate lymphocytes and render them susceptible toinfection. PBLs were therefore exposed for only 24 hours tosupernatants containing HIV-1–derived or MLV-based vectors.After transduction, cells were washed and were further incubatedfor 5 days in media supplemented with, or without, a lowconcentration of rIL-2 to prevent cell death. No significantprogression in cell cycle (data not shown) was found 24 hours afterinfection, before addition of rIL2, in contrast to previous experi-ments where cells had been put in contact with OKT3SU-displaying vectors for 6 days (Table 1). Almost no transduction wasdetected following infection with either lentiviral vectors display-ing only VSV-G glycoproteins (0.67%6 0.65%; n5 5) or withMLV-derived vectors, whether the surface of the latter vectorparticles carried (0.63%6 0.53; n5 2), or not (0.25%6 0.18%;n 5 5), OKT3SU chimeric glycoproteins (Figure 8). Compared tovectors generated with unmodified glycoproteins, infection ofnonactivated PBLs with OKT3SU-displaying lentiviral vectorsresulted in an average 20-fold higher transduction efficiency, in therange of 9% to 19% GFP1 cells (mean, 12.60%6 4.63%; n5 5).Similar results were obtained in experiments performed in theabsence of rIL-2, yet the prolonged time of culture of PBLs withoutcytokines resulted in extensive cell death (data not shown). Inconclusion, these data indicated that transduction of primarylymphocytes by a short exposure to OKT3SU-displaying lentiviral
vectors gave rise to high transduction levels, which were 34-foldand 100-fold higher than those obtained with the unmodifiedHIV-1–derived and MLV-derived vectors, respectively.
Discussion
Lentiviral vectors have shown promise in the transduction ofseveral resting cell types such as retinal cells, pancreatic islets, cellsof the central nervous system, or progenitor and differentiatedhematopoietic cells.12 For these reasons, lentiviral vectors shouldbe preferred gene delivery vehicles over vectors derived fromoncoretroviruses such as MLVs that cannot transduce nonproliferat-ing target cells.8 There are, however, important gene transferrestrictions to some nonproliferative tissues or cell types and recentstudies have shown that progenitor hematopoietic stem cells in G0,nonactivated primary blood lymphocytes or monocytes were nottransducible by HIV-1–derived vectors.17,19,25 In contrast to themechanisms that restrict gene transfer into monocytes,18,19 the 2former cell types are refractory to gene transfer most probablybecause their low metabolic state prevents post-entry replicationsteps such as initiation or processivity of reverse-transcription andnuclear import.17,25 Thus, activation of these cells, causing G0-to-G1b transition of the cell cycle, is required to relieve from theblocks in gene delivery.17,25 This can be achieved by adding aminimal combination of cytokines into the infection media during ashort culture period.14,17,25,46Therefore, on such minimal ex vivoinfection conditions, lentiviral vectors may allow gene transfer inmore primitive hematopoietic progenitor cells than those usuallyreached with MLV-derived vectors.46 However, there is consider-able interest to further develop the lentiviral vectors so as to reachthe most primitive progenitor cells that are believed to be in aquiescent state, to be slow to respond to cytokine stimulation, andto tend to lose multipotentiality or long-term repopulating capacityunder cytokine stimulation. Similarly, in the case of geneticmodification of T cells, most previous studies have focused on theoptimization of transduction protocols, yet most gene therapyapplications will require the transduction of the naive T-cellpopulations.14,47 It has now become evident that optimized proto-cols that use activation stimuli such as antibodies and mitogensmay not be compatible with the preservation of the pool of naive Tcells, which harbor the capacity to respond to novel antigens.4,47
Recent improvements in the development of lentiviral vectorshave focused on the optimization of nuclear import of thetransgene.15,16 Inclusion of the HIV-1 central polypurine track(cPPT) in lentiviral vectors has resulted in enhanced transductionof human progenitor stem cells and T cells.14,46 However, theimproved lentiviral vectors that include the cPPT sequence still failto transduce nonactivated T lymphocytes,14 most likely because theprimary block in initiation or completion of reverse transcriptioncould not be alleviated with the novel vectors. Therefore, becauseminimal activation of resting human progenitor stem cells and Tcells with stimuli that cause G0-to-G1 transition allows genetransfer,17,25,29 we sought to generate surface-modified lentiviralvector particles that would convey a transient activation signal intarget cells at the time of gene delivery. As a proof of concept, weshow here that lentiviral vectors that display on their viral surfacean anti-CD3 scFv T cell-activating polypeptide mediate efficientgene transfer into freshly isolated lymphocytes not activated byexogenous stimuli. No addition of T cell-activating stimuli in theinfection media was required to achieve transduction levels of up to48% GFP1 cells. In comparison, unmodified lentiviral vectors
Figure 8. Transduction of human PBLs after a short exposure with OKT3SU-displaying lentiviral vectors. Nonactivated PBLs were incubated for 24 hours withVSV-G–pseudotyped vector particles derived from HIV-1 or from MLV displaying ornot displaying OKT3SU chimeric glycoproteins, as indicated. Infected cells were thenwashed to eliminate unbound viral particles and incubated for 5 days in the presenceof human rIL-2 (1 ng/mL) to reduce cell death. The number of GFP1 cells was thendetermined by FACS analysis. Results of transduction in PBLs from 7 different donorsare given. The PBL sets correspond to the ones in Figure 5. The MOIs are providedfor each experiment.
2348 MAURICE et al BLOOD, 1 APRIL 2002 z VOLUME 99, NUMBER 7
For personal use only. by guest on May 30, 2013. bloodjournal.hematologylibrary.orgFrom

whose viral surface expressed only the VSV-G glycoprotein wereunable to transduce freshly isolated T cells.
Optimal activation of resting T cells requires signaling via theTCR and additional stimulation provided through accessory mol-ecules present on the surface of antigen-presenting cells (APCs).For highly purified resting T cells, multivalent APC stimulation canbe mimicked ex vivo with immobilized anti-CD3 antibodies andcauses a G0-to-G1a transition. However, this does not allowcompletion of HIV-1 reverse transcription25 or efficient transduc-tion with HIV-1–based vectors.29 Costimulation with immobilizedanti-CD3 and anti-CD28 antibodies allows progression to G1b
48,49
and allows successful infection by wild-type HIV-125 or transduc-tion by lentiviral vectors.14 Prolonged cell exposure to high loads ofimmobilized anti-CD3 and anti-CD28 antibodies induces cellproliferation50 and allows transduction by vectors derived fromeither lentiviruses or oncoretroviruses.4,14 Our results confirm thenotion that induction of cell division is not a prerequisite forefficient gene transfer in PBLs with lentiviral vectors. No HIV-1accessory proteins, such as vpr or vif, were required to achievehigh-level transduction in the PBLs, in agreement with results ofothers.13,14 Interestingly, our data show that comparable transduc-tion levels can be achieved with either the surface-modified,OKT3SU-displaying, lentiviral vectors or with the unmodifiedvectors used in combination with both soluble anti-CD3 andanti-CD28 antibodies. This indicates that an exogenous costimula-tion signal may not be needed for transduction with the OKT3SU-displaying lentiviral vectors. These results, which are in contrast tothe poor transduction levels obtained with VSV-G–pseudotypedlentiviral vectors in the presence of anti-CD3 antibodies alone, arelikely to be explained by the fact that the multivalent presentationon viral particles of TCR-binding polypeptides may strongly favorTCR cross-linking. Additionally, local concentration of TCR-binding polypeptides could be much higher when presented onvirions compared to when provided to cells as soluble or immobi-lized anti-CD3 antibodies. In agreement with this hypothesis,concentrations of immobilized anti-CD3 antibodies 10-fold higherthan those classically used in T-cell activation and transductionexperiments, that is, around 1mg/well,14,25were found sufficient toallow infection by HIV-1 and could also induce cell proliferation.50
Alternatively, concomitant to partial activation induced by theOKT3SU-displaying virions, costimulation could be providedeither by residual APCs likely to be present in the preparations ofPBLs25,49 or by costimulatory molecules that may have beencoincorporated on the surface of the lentiviral particles.51
T-cell receptor cross-linking induced by surface-modified lenti-viral vectors may have adverse effects on the phenotype of thetransduced T cells, such as inducing anergy or modifying theirability to respond to novel antigens.52 An important component ofthe homeostasis of the immune system is the maintenance of the
peripheral naive T-cell repertoire. Human T cells can be dividedinto naive and memory subsets based, respectively, on expressionof RA and RO isoforms of the CD45 molecule. In contrast tomemory T cells, naive cells exhibit few effectors functions (eg,cytokine production), are less susceptible to activation-induced celldeath, and display robust proliferation in response to TCR-mediated activation signals. It is important that the compartment ofnaive CD45RA1 cells is maintained through life. Although TCRactivation induced on infection with OKT3SU-displaying viralparticles is short, the change in T-cell phenotype cannot be formallyexcluded. In fact, transduction of PBLs with OKT3SU-displayinglentiviral vectors was found to shift the T cells to the memoryphenotype (data not shown), in agreement with results of othersusing TCR stimulation by anti-CD3/anti-CD28 antibodies to allowinfection of resting PBLs by retroviral vectors.4 It should bepointed out that the approach reported here was designed to provideproof of concept of gene transfer strategies that may have utilityelsewhere. Here, we show for the first time that it is possible toefficiently transduce quiescent PBLs by a lentiviral vector thatdisplays a T cell-activating polypeptide. For the reasons discussedabove, it might be preferable to display cytokines like IL-2, IL-7,and IL-4, or combinations of these polypeptides, on the surface ofthe lentiviral vectors. These cytokines are known to promotelong-term survival of resting T cells while maintaining the pheno-type of the subset of naive cells.53,54 Previously, we described thegeneration of MLV-derived vectors that displayed IL-2 chimericenvelope glycoproteins (IL-2SU).37 These vectors allowed efficienttransduction of G0/G1-arrested cell lines expressing the IL-2receptor.37 However, lentiviral vectors pseudotyped with VSV-Gand chimeric IL-2SU envelope glycoproteins were not able totransduce freshly isolated PBLs, despite their capacity to weaklyup-regulate the CD25 and CD69 activation markers (data notshown). This result was most likely due to the low expression ofhigh-affinity IL-2 receptors on resting PBLs. Alternatively, IL-7 isa promising candidate because it is known that it activates PBLsand permits infection with lentiviral vectors into both memory andnaive T cells.29,47Moreover, IL-7 contributes to the maintenance ofthe adult CD45RA1 T-cell pool.55,56 Characterization of lentiviralvectors that display some of these polypeptides on the viral surfaceare currently underway in our laboratory and will be valuable toolsto selectively activate specific T-cell subsets, allowing numerousstudies of the physiology of lymphocytes.
Acknowledgments
We are grateful to Naomi Taylor and Jacqueline Marvel forstimulating discussions and for critical reading of the manuscript.
References
1. Blaese RM, Culver KW, Miller AD, et al. T-lym-phocyte directed gene therapy for ADA-SCID:initial trial results after 4 years. Science. 1995;270:275-280.
2. Bordignon C, Notaranglo LD, Nobili N, et al. Genetherapy in peripheral blood lymphocytes andbone marrow for ADA- immunodeficient patients.Science. 1995;270:470-475.
3. Buchschacher GL, Wong-Staal F. Approaches togene therapy for human immunodeficiency virusinfection. Hum Gene Ther. 2001;12:1013-1019.
4. Dardalhon V, Jaleco S, Rebouissou C, et al.Highly efficient gene transfer in naive human T
cells with a murine leukemia virus-based vector.Blood. 2000;96:885-893.
5. Ferrand C, Robinet E, Contassot E, et al. Retrovi-rus-mediated gene transfer in primary T lympho-cytes: influence of the transduction/selection pro-cess and of ex vivo expansion on the T cellreceptor b chain hypervariable region repertoire.Hum Gene Ther. 2000;11:1151-1164.
6. Anderson WF. Human gene therapy. Nature.1998;392:25-30.
7. Miller DG, Adam MA, Miller AD. Gene transfer byretrovirus vectors occurs only in cells that are ac-tively replicating at the time of infection. Mol CellBiol. 1990;10:4239-4242.
8. Roe T, Reynolds TC, Yu G, Brown PO. Integrationof murine leukemia virus DNA depends on mito-sis. EMBO J. 1993;12:2099-2108.
9. Bukrinsky MI, Sharova N, Dempsey MP, et al.Active nuclear import of human immunodefi-ciency virus type 1 preintegration complexes.Proc Natl Acad Sci U S A. 1992;89:6580-6584.
10. Lewis P, Hensel M, Emerman M. Human immu-nodeficiency virus infection of cells arrested in thecell cycle. EMBO J. 1992;11:3053-3058.
11. Weinberg JB, Matthews TJ, Cullen BR, MalimMH. Productive human immunodeficiency virustype 1 (HIV-1) infection of non proliferative humancells. J Exp Med. 1991;174:1477-1482.
T-CELL TRANSDUCTION BY SURFACE-MODIFIED HIV VECTORS 2349BLOOD, 1 APRIL 2002 z VOLUME 99, NUMBER 7
For personal use only. by guest on May 30, 2013. bloodjournal.hematologylibrary.orgFrom

12. Vigna E, Naldini L. Lentiviral vectors: excellenttools for experimental gene transfer and promis-ing candidates for gene therapy. J Gene Med.2000;2:308-316.
13. Salmon P, Kindler V, Ducrey O, Chapuis B, ZublerRH, Trono D. High-level transgene expression inhuman hematopoietic progenitors and differenti-ated blood lineages after transduction with im-proved lentiviral vectors. Blood. 2000;96:3392-3398.
14. Dardalhon V, Herpers B, Noraz N, et al. Lentivi-rus-mediated gene transfer in primary T cells isenhanced by a central DNA flap. Gene Ther.2001;8:190-198.
15. Zennou V, Petit C, Guetard D, Nerhbass U, Mon-tagnier L, Charneau P. HIV-1 genome nuclearimport is mediated by a central DNA flap. Cell.2000;101:173-185.
16. Follenzi A, Ailles L, Bakovic S, Geuna M, NaldiniL. Gene transfer by lentiviral vectors is limited bynuclear translocation and rescued by HIV-1 polsequences. Nat Genet. 2000;25:217-222.
17. Sutton RE, Reitsma MJ, Uchida N, Brown PO.Transduction of human progenitor hematopoieticstem cells by human immunodeficiency virus type1-based vectors is cell cycle dependent. J Virol.1999;73:3649-3660.
18. Kootstra NA, Zwart BM, Schuitemaker H. Dimin-ished human immunodeficiency virus type 1 re-verse transcription and nuclear transport in pri-mary macrophages arrested in early G(1) phaseof the cell cycle. J Virol. 2000;74:1712-1717.
19. Neil S, Martin F, Ikeda Y, Collins M. Postentry re-striction to human immunodeficiency virus-basedvector transduction in human monocytes. J Virol.2001;75:5448-5456.
20. Stevenson M, Stanwick TL, Dempsey MP, Lam-onica CA. HIV-1 replication is controlled at thelevel of T cell activation and proviral integration.EMBO J. 1990;9:1551-1560.
21. Sun Y, Pinchuk LM, Agy MB, Clark EA. Nuclearimport of HIV-1 DNA in resting CD41 T cells re-quires a cyclosporin A-sensitive pathway. J Im-munol. 1997;158:512-517.
22. Tang S, Patterson B, Levy JA. Highly purified qui-escent human peripheral blood CD41 T cells areinfectible by human immunodeficiency virus butdo not release virus after activation. J Virol. 1995;69:5659-5665.
23. Zack JA, Haislip AM, Krogstad P, Chen ISY. In-completely reverse-transcribed human immuno-deficiency virus type 1 genomes in quiescentcells can function as intermediates in the retrovi-ral life cycle. J Virol. 1992;68:1717-1725.
24. Zack JA. The role of the cell cycle in HIV-1 infec-tion. Adv Exp Med Biol. 1995;374:27-31.
25. Korin YD, Zack JA. Progression to the G1b phaseof the cell cycle is required for completion of hu-man immunodeficiency virus type 1 reverse tran-scription in T cells. J Virol. 1998;72:3161-3168.
26. Bukrinsky MI, Haggerty S, Dempsey MP, et al. Anuclear localization signal within HIV-1 matrixprotein that governs infection of non-dividingcells. Nature. 1993;365:666-669.
27. Spina CA, Guatelli JC, Richman DD. Establish-ment of a stable, inducible form of human immu-
nodeficiency virus type 1 DNA in quiescent CD4lymphocytes in vitro. J Virol. 1995;69:2977-2988.
28. Korin YD, Zack JA. Nonproductive human immu-nodeficiency virus type 1 infection in nucleoside-treated Go lymphocytes. J Virol. 1999;73:6526-6532.
29. Unutmaz D, KewalRamani VN, Marmon S, Litt-man DR. Cytokine signals are sufficient for HIV-1infection of resting human T lymphocytes. J ExpMed. 1999;189:1735-1746.
30. Naldini L, Blomer U, Gallay P, et al. In vivo genedelivery and stable transduction of nondividingcells by a lentiviral vector. Science. 1996;272:263-267.
31. Zufferey R, Nagy D, Mandel R, Naldini L, TronoD. Multiply attenuated lentiviral vector achievesefficient gene delivery in vivo. Nat Biotechnol.1997;15:871-875.
32. Yee JK, Friedmann T, Burns JC. Generation ofhigh-titer pseudotyped retroviral vectors with verybroad host range. Methods Cell Biol. 1994;43:99-112.
33. Kung PC, Goldstein G, Reinherz EL, SchlossmanSF. Monoclonal antibodies defining distinctivehuman T cell surface antigens. Science. 1979;206:347-349.
34. Ager S, Nilson BHK, Morling FJ, Peng KW, Cos-set F-L, Russell SJ. Retroviral display of antibodyfragments; interdomain spacing strongly influ-ences vector infectivity. Hum Gene Ther. 1996;7:2157-2164.
35. Ott D, Friedrich R, Rein A. Sequence analysis ofamphotropic and 10A1 murine leukemia virus:close relationship to mink cell focus forming vi-ruses. J Virol. 1990;64:757-766.
36. Valsesia-Wittmann S, Morling FJ, Nilson BHK,Takeuchi Y, Russell SJ, Cosset F-L. Improvementof retroviral retargeting by using amino acid spac-ers between an additional binding domain andthe N terminus of Moloney murine leukemia virusSU. J Virol. 1996;70:2059-2064.
37. Maurice M, Mazur S, Bullough FJ, et al. Efficientgene delivery to quiescent IL2-dependent cells bymurine leukemia virus-derived vectors harboringIL2 chimeric envelopes glycoproteins. Blood.1999;94:401-410.
38. Cosset F-L, Morling FJ, Takeuchi Y, Weiss RA,Collins MKL, Russell SJ. Retroviral retargeting byenvelopes expressing an N-terminal binding do-main. J Virol. 1995;69:6314-6322.
39. Evans LH, Morrison RP, Malik FG, Portis J, BrittWJ. A neutralizable epitope common to the enve-lope glycoproteins of ecotropic, polytropic, xeno-tropic and amphotropic murine leukemia viruses.J Virol. 1990;64:6176-6183.
40. Russell SJ, Cosset F-L. Modifying the host rangeproperties of retroviral vectors. J Gene Med.1999;1:300-311.
41. Pinter A, Kopelman R, Li Z, Kayman SC, SandersDA. Localization of the labile disulfide bond be-tween SU and TM of the murine leukemia virusenvelope protein complex to a highly conservedCWLC motif in SU that resembles the active-sitesequence of thiol-disulfide exchange enzymes.J Virol. 1997;71:8073-8077.
42. Battini JL, Rodrigues P, Muller R, Danos O,Heard J-M. Receptor-binding properties of a puri-
fied fragment of the 4070A amphotropic murineleukemia virus envelope glycoprotein. J. Virol.1996;70:4387-4393.
43. Chinnasamy D, Chinnasamy N, Enriquez MJ,Otsu M, Morgan RA, Candotti F. Lentiviral-medi-ated gene transfer into human lymphocytes: roleof HIV-1 accessory proteins. Gene Ther. 2000;96:1309-1316.
44. Gliniak BC, Kozak SL, Jones RT, Kabat D. Disul-fide bonding controls the processing of retroviralenvelope glycoproteins. J Biol. Chem. 1991;266:22991-22997.
45. Costello E, Munoz M, Buetti E, Meylan PRA,Diggelmann H, Thali M. Gene transfer into stimu-lated and unstimulated T lymphocytes by HIV-1-derived lentiviral vectors. Gene Ther. 2000;7:596-604.
46. Sirven A, Pflumio F, Zennou V, et al. The humanimmunodeficiency virus type-1 central DNA flap isa crucial determinant for lentiviral vector nuclearimport and gene transduction of human hemato-poietic stem cells. Blood. 2000;96:4103-4110.
47. Dardalhon V, Jaleco S, Kinet S, et al. Interleu-kin-7 differentially regulates cell cycle progressionand HIV-1 based vector infection in neonatal andadult CD41 T cells. Proc Natl Acad Sci U S A.2001;98:9277-9282.
48. Chambers CA, Allison JP. Co-stimulation in T cellresponses. Curr Opin Immunol. 1997;9:396-404.
49. Lanzavecchia A, Lezzi G, Viola A. From TCR en-gagement to T cell activation: a kinetic view of Tcell behavior. Cell. 1999;96:1-4.
50. Spina CA, Prince HE, Richman DD. Preferentialreplication of HIV-1 in the CD45RO memory cellsubset of primary CD4 lymphocytes in vitro. J ClinInvest. 1997;99:1774-1785.
51. Bounou S, Dumais N, Tremblay MJ. Attachmentof human immunodeficiency virus-1 (HIV-1) par-ticles bearing host-encoded B7–2 proteins leadsto nuclear factor-kappa B- and nuclear factor ofactivated T cells-dependent activation of HIV-1long terminal repeat transcription. J Biol Chem.2001;276:6359-6369.
52. Contassot E, Ferrand C, Angonin R, et al. Ganci-clovir-sensitive acute graft-versus-host disease inmice receiving herpes simplex virus-thymidinekinase-expressing donor T cells in a bone marrowtransplantation setting. Transplantation. 2000;69:503-508.
53. Boise L, Minn AJ, June CH, Lindsten T, Thomp-son CB. Growth factors can enhance lymphocytesurvival without committing the cell undergo celldivision. Proc Natl Acad Sci U S A. 1995;92:5491-5495.
54. Geiselhart LA, Humphries CA, Gregorio TA, MouS, Subleski J, Komschlies KL. IL-7 administrationalters the CD4:CD8 ratio, increases T cell num-bers, and increases T cell function in the absenceof activation. J Immunol. 2001;166:3019-3027.
55. Soares MVD, Borthwick NJ, Maini MK, JanossyG, Salmon M, Akbar AN. IL-7 dependent extrathy-mic expansion of CD45RA1 T cells enables pres-ervation of a naive repertoire. J Immunol. 1998;161:5909-5917.
56. Hassan J, Reen DJ. IL-7 promotes the survivaland maturation but not differentiation of humanpost-thymic CD41 T cells. Eur J Immunol. 1998;28:3057-3065.
2350 MAURICE et al BLOOD, 1 APRIL 2002 z VOLUME 99, NUMBER 7
For personal use only. by guest on May 30, 2013. bloodjournal.hematologylibrary.orgFrom




















