
L'ethnographie en trois dimensions. Un entretien avec Jack Katz
Upload
khangminh22Category
view
25download
0
REPUBLIQUE ALGERIENNE DEMOCRATIQUE ET POPULAIRE
MINISTERE DE L'ENSEIGNEMENT SUPERIEUR ET DE LA RECHERCHE SCIENTIFIQUE
UNIVERSITIE ABOU BEKR BELKAID TLEMCEN
FACULTE DES SCIENCES DE LA NATURE ET DE LA VIE
DE LA TERRE ET DE L’UNIVERS
DEPARTEMENT DE BIOLOGIE
Laboratoire de Microbiologie Appliquée à l’Agroalimentaire, au Biomédical
et à l’Environnement « LAMAABE »
THESE
Présentée par
Mme ZENATI Fatima
Ep Lakehal
En vue de l’obtention du diplôme de Doctorat en biologie
Option : Microbiologie
Soutenue le …/…/2016
Devant le jury
Président Abdelouahid D.E. Professeur U. de Tlemcen
Examinateur Timinouni M. Professeur Institut Pasteur Maroc
Examinatrice Hassaine H. Professeur U. de Tlemcen
Examinateur Abbouni B. Professeur U. de Sidi Bel abbes
Directeur de thèse Bendahou M. Professeur U. de Tlemcen
Année Universitaire : 2015-2016
Effet inhibiteur des huiles essentielles de trois plantes aromatiques
sur Escherichia coli (BLSE) responsables d’infections urinaires
d’origine hospitalière


REPUBLIQUE ALGERIENNE DEMOCRATIQUE ET POPULAIRE
MINISTERE DE L'ENSEIGNEMENT SUPERIEUR ET DE LA RECHERCHE SCIENTIFIQUE
UNIVERSITIE ABOU BEKR BELKAID TLEMCEN
FACULTE DES SCIENCES DE LA NATURE ET DE LA VIE
DE LA TERRE ET DE L’UNIVERS
DEPARTEMENT DE BIOLOGIE
Laboratoire de Microbiologie Appliquée à l’Agroalimentaire, au Biomédical
et à l’Environnement « LAMAABE »
THESE
Présentée par
Mme ZENATI Fatima
Ep Lakehal
En vue de l’obtention du diplôme de Doctorat en biologie
Option : Microbiologie
Soutenue le …/…/2016
Devant le jury
Président Abdelouahid D.E. Professeur U. de Tlemcen
Examinateur Timinouni M. Professeur Institut Pasteur Maroc
Examinatrice Hassaine H. Professeur U. de Tlemcen
Examinateur Abbouni B. Professeur U. de Sidi Bel abbes
Directeur de thèse Bendahou M. Professeur U. de Tlemcen
Année Universitaire : 2015-2016
Effet inhibiteur des huiles essentielles de trois plantes aromatiques
sur Escherichia coli (BLSE) responsables d’infections urinaires
d’origine hospitalière

Dédicaces
Je dédie humblement ce travail :
A mes très chers parents
A mes sœurs Amina et Meriem et sa fille Nouran.
A mon époux. Kamel.
A mes très chère amies : Raniya, Naima, Rema, Zahra, Wassila, Nadia, Fatima, Nadjet,
Chaficka, Sara, Nassima, Khawla, Khadidja, Karima, Wafae, Zahira Et Samya. En
souvenir des moments inoubliables que nous avons passés ensemble. Je vous souhaite, à
travers ce travail et en témoignage de ma profonde affection, tout le bonheur et la réussite
que vous méritez.
A tous les membres de ma famille petits et grands. Veuillez trouver dans ce modeste travail
l’expression de mon affection la plus sincère.
F. Zenati

Remerciements
Avant toutes choses, je remercie Dieu, le tout puissant, pour m’avoir donné la force
et la patience pour réaliser ce travail.
Ce travail a été réalisé dans le laboratoire de microbiologie appliquée à
l’agroalimentaire, au biomédicale et a l’environnement (LAMAABE), université Aboubekr
Belkaïd (ABB) de Tlemcen. La caractérisation moléculaire des souches bactériennes a été
faite à l’Institut Pasteur de Casablanca au Maroc.
J’adresse mes vifs remerciements à mon Directeur de Thèse, monsieur Bendahou
Mourad, professeur à l’université ABB, pour sa grande disponibilité, son écoute, ses conseils
judicieux et son suivi tout au long de ce travail. Je le remercie également de m’avoir fait
confiance pour la réalisation de ce travail.
Je remercie le professeur Moussa-Boudjemaa Boumediène directeur du laboratoire
LAMAABE de m’avoir permis de réaliser ce travail au sein du laboratoire.
J’adresse mes remerciements et mes respects à monsieur Abdelouahid Djamel Eddine,
professeur à l’université ABB de Tlemcen, pour ces conseils précieux et pour avoir accepté de
présider le jury de cette thèse.
Je suis très reconnaissante à Monsieur Timinouni Mohammed, professeur et chef du
département de microbiologie responsable du laboratoire de Bactériologie Moléculaire à
l’Institut Pasteur à Casablanca (Maroc), de m’avoir accueilli au sein du Laboratoire. Je lui
témoigne toute ma reconnaissance pour sa bonne humeur, son sourire, son agréable assistance
durant mon stage et d’avoir accepté d’examiner ce travail.
Je remercie profondément Mme Hassaine Hafida, professeur à l’université ABB de
Tlemcen et responsable de la formation doctorale «Maitrise du développement microbien»
pour avoir accepté d’examiner ce travail. Ses encouragements, ces conseils précieux et sa
confiance m’ont permis d’avancer dans ce travail.
Un grand merci à Mr Abbouni Bouziane professeur à l’université de Sidi Belabess
pour avoir accepté d’examiner cette thèse.
Je remercie l’équipe de chimie des substances naturelles dirigée par le professeur Jean
Costa de l’université de Corse (France) pour l’analyse des huiles essentielles.

Tous mes remerciements vont aussi à l’équipe du laboratoire de bactériologie
moléculaire de l’Institut Pasteur de Casa (Maroc), particulièrement à Monsieur Barguigua
Abouddihaj et madame Kawter pour leur collaboration durant mon séjour scientifique au
laboratoire.
Je remercie tous les enseignants et les doctorants du laboratoire LAMAABE, en
particulier Mme Bendimred Nahida, qui m’ont soutenu et encourager tout au long de ma
thèse.
Merci aux membres de l’équipe des substances naturelles antimicrobiennes (SNA) de
LAMAABE, Dr Khadir Abdelmounaim, Dr Barka Mohamed Salih, Dr Borsali Mohamed
Nabil, Mr Messaoudi Omar, Dr Benbelaid Fethi et Mme Bellahcène Chafika, qui m’ont
aidé et soutenu dans mes travaux au laboratoire.
Enfin, merci à tous ceux qui, de loin ou de près ont contribué à la réalisation de ce
travail.

يسخشفى الانسؤونت ع االنخهاباث انبىنت و انعسونت ي E. coli BLSE عهى زىث عطرتثالديفعىل : العنوان
الملخص
د انقضاء عهى هذ انشكهت حى حجربت وف صذ.اإلساإ يقاويت انبكخرا نهضاداث انحىت يشكهت حققت حهذد صحت
Ecoli . ضذ (Cinnamomum cassia, Coriandrum sativum Ziziphora hispanica)ثالثت زىث عطرت
BLSEعرضت أكثر انساء هى أ انخائج بج إ . انسؤونت ع االنخهاباث انبىنت و انعسونت ي يسخشفى حهسا
E. coli BLSE يع انعهى ا سبت ℅63حثم انسالنت انرئست نهذا االنخهاب بسبت E. coliنالنخهاباث انبىنت وا
℅96 بسبت TEM1وBla CTX-M نهذ انبكخرا اها حهك سبت كبرة ي جاث PCR واظهر ححهم 32,5℅
حأثرا األكثر ه Cinnamomum cassia أ زىث عطرت بج ثالد نخأثر انعرضت E. coli BLSE إ.℅100و
يم وحركس /غ و1.97 اد انثبط يى يع انعهى ا قت حركس 33.23 انثبط انحانخ انهائت وانبىفهت يع قطر كهخاف
يم /غ و4,53 انبىفهى هىإلزانت األدىيم و انخركس /غ و 4,37 نبىفهىلااألدى انثبطيم وحركس /غ و2.5 األدىانقاحم
واظهر يسج انسىث انعطرت يع .نها يفعىل اقمZiziphora hispanica و Coriandrum sativum أيع انعهى
انخال ا انبكخرت اضهرث أ حركت قخم أ كا إضاف يفعىال E. coli BLSE 09ضذ céfotaxime انضاد انحىي
120 ف غضى Coriandrum sativum دققت 30 انبكخرا ف غضى حقخم يCinnamomum cassia زج
.دققت
انفعانت ضذ انبكخرا, زىث عطرت,انقاويت , ,PCR , E. coli BLSE االنخهاباث انبىنت:انكهاث انفخاحت

Titre : Effet inhibiteur des huiles essentielles de trois plantes aromatiques sur Escherichia coli
(BLSE) responsables d’infections urinaires d’origine hospitalière.
Résumé
La résistance des bactéries aux antibiotiques pose un vrai problème de santé publique.
Dans le but de trouver des alternatifs de lutte contre cette résistance, l’étude proposé est
d’évaluer l’effet inhibiteur de trois huiles essentielles (Cinnamomum cassia, Coriandrum
sativum et Ziziphora hispanica) sur des souches E. coli BLSE responsables d’infections
urinaires isolées au CHU de Tlemcen. Les résultats ont montré que l’infection urinaire
nosocomiale est importante chez les femmes et elle est due principalement à E. coli (63℅).
L’incidence d’Escherichia coli BLSE a été de 32,5%. L’analyse par PCR de E. coli BLSE ces
dernières, a révélé la présence des gènes Bla CTX-M et TEM1 avec un pourcentage élevé
(100℅ et 96℅). Les souches E. coli BLSE soumises à l’effet des trois huiles essentielles, ont
montré une grande sensibilité à l’huile essentielle de Cinnamomum cassia à l’état
planctonique et biofilm. Elle a donné en moyenne respectivement, une zone d’inhibition
> 33,25mm, une CMI=1,97mg/ml, CMB=2,5mg/ml et une CMIB=4,37 mg/ml,
CMEB=4,53mg/ml. Par contre Coriandrum sativum et Ziziphora hispanica sont moins actif.
En combinant les huiles à l’antibiotique céfotaxime contre E.coli BLSE 09, les huiles
essentielles de Cinnamomum cassia et Coriandrum sativum ont présenté un effet d’addition.
La cinétique de destruction des cellules bactériennes par l’huile essentielle de Cinnamomum
cassia a été obtenue au bout de 30 min, par contre il a fallu 240 min pour les détruire par
l’huile de Coriandrum sativum.
Mots clés : Infection urinaire ; E. coli BLSE ; PCR ; Résistance ; Huiles essentielles ; Activité
antibactérienne.

Title: Inhibitory effect of essential oils of three aromatic plants on Escherichia coli (ESBL)
responsible for urinary tract infections Hospital
Abstract
Bacterial resistance to antibiotics is a real public health problem. In order to find
alternative fight against this resistance, the proposed study is to evaluate the inhibitory effect
of three essential oils (Cinnamomum cassia, Coriandrum sativum and Ziziphora hispanica)
on ESBL E. coli strains responsible for urinary tract infections isolated at the University
Hospital of Tlemcen. The results showed that nosocomial urinary tract infection is high
among women and it is mainly due to E. coli (63℅). The effect of E. coli ESBL was 32,5%.
PCR analysis of ESBL E. coli, revealed the presence of genes Bla CTX-M and TEM1 with
high percentage (100℅ and 96℅). ESBL E. coli strains subject to the effect of three essential
oils, have shown great sensitivity to essential oil of Cinnamomum cassia at state planctonic
and biofilm. She gave an inhibition zone > 33,25mm, MIC=1,97mg/ml, MBC = 2,5mg/ml and
a MBIC=4,37mg/mL, MBEC=4,53mg/ml. While, Coriandrum sativum and Ziziphora
hispanica oils are less active. Essential oils combined with the antibiotic cefotaxime against
ESBL E. coli 09, showed an additive effect of Cinnamomum cassia and Coriandrum sativum
essential oils. The kinetics of destruction of bacterial cells with essential oil of Cinnamomum
cassia was obtained after 30 min, while Coriandrum sativum oil destroys bacterial cells after
240 min.
Keywords : Urinary tract infection ; ESBL E. coli ; PCR ; Résistance ; Essential oils
Antibactérial activity.

Sommaire
Introduction………………………………………………………………………………………1
Partie I : Synthèse bibliographique…………………………...4
Chapitre I : les infections urinaires………………………………………………………..........4
I.1. Généralités sur les infections urinaires………………………………………………..........4
I.1.1. Infection urinaire et colonisation………………………………………………………........4
I.1.2. Infection urinaire simple et compliquée………………………………………………..........5
I.1.3. Infection urinaire récidivante………………………………………………………..............6
I.1.4. Infection urinaire communautaire et nosocomiale………………………………………….6
I.1.5. Epidémiologie et facteurs de risque……………………………………………………..7
I.2. Mode de pénétration des bactéries dans les voies urinaires………………………….........7
I.2. La voie ascendante…………………………………………………………………………….7
I.2.2. La voie hématogène…………………………………………………………………............8
I.2.3. Extension directe à partir d’un organe de voisinage (voie lymphatique)……………………8
I.3. Les infections à E. coli……………………………………………………………………….8
I.3.1. Généralités…………………………………………………………………………………..8
I.3.2. Caractères morphologiques et culturaux…………………………………………………….9
I.3.3. Caractères biochimiques……………………………………………………………….........9
I.3.4. Caractères antigéniques……………………………………………………………….........10
I.3.5. Pouvoir pathogène………………………………………………………………………….10
I.3.6. Les substances élaborées...………………………………………………………………….11
I.3.7. Physiopathologie de l’infection urinaire à Escherichia coli…………………………..........11
I.4. Antibiotiques et facteurs de risque de résistance aux antibiotiques……………………..14
I.4.1. Antibiotiques des infections urinaires ………………………………………………..........20
I.4.2. Schéma de traitement des infections urinaires ……………………………………….........22
I.4.3. Resistance d’E.coli aux antibiotiques chez l'adulte …………………………………..........23
I.4.4. Les mécanismes de résistances bactériennes aux antibiotiques …………………………...29
I.4.5. Facteurs de risque connus d’infections à EP-blse …………………………………………30
I.4.6. Les moyens de lutte contre E. coli BLSE ………………………………………….........31
I.4.7. Préventions…………………………………………………………………………………30

Chapitre II : Produits naturels et infections urinaires ………………………………….........31
II.1. Les plantes aromatiques et médicinales……………………………………………..............31
II.2. Généralités sur les huiles essentielles ...……………………………………………….........32
II.2.1. Définition des huiles essentielles ...………………………………………………….........32
II.2.2. Répartition…………………………………………………………………………………32
II.3. Composition des huiles essentielles…………………………………………………………32
II.4. Méthodes utilisées d’extractions des huiles essentielles……………………………………33
II.5. Activité antimicrobienne……………………………………………………………….........33
II.5.1. Mécanismes d’action des huiles essentielles………………………………………………34
II.5.2. Place des plantes médicinales dans la lutte contre les résistances aux
antibiotiques…................................................................................................................................35
II.5.3. Méthodes d’évaluation de l’activité antimicrobienne………………………………...........36
II.5.4. Méthodes de détermination de la Concentration Minimale Inhibitrice.……………...........37
II.6. Domaines d’utilisation des huiles essentielles………………………………………….........38
II.7. Les principales voies d’utilisation des huiles essentielles…………………………………....39
II.8. Monographie des plantes sélectionnées………………………………………………...........40
Partie II : Matériel et méthodes………………………………44
II.1. Matériel microbien………………………………………………………………................44
II.1.1. Souches de références ……………………………………………………………..............44
II.1.2. Souches d’origine clinique ………………………………………………………………..44
II.2. Méthode……………………………………………………………………………………..44
II.2.1.Prélèvement………………………………………………………………………………...44
II.2.2. Bandelette urinaire ………………………………………………………………………...44
II.2.3. Examen bactériologique des urines ……………………………………………….............44
II.2.4. Antibiogramme ……………………………………………………………………………45
II.2.5. Détection des E .coli BLSE (Test de synergie) …………………………………………...45
II.2.6. Détermination de la CMI sur microplaque………………………………………………...46
II.2.7. Typage moléculaire d’E. coli BLSE……………………………………………….............47
II.2.7.1. Extraction d’ADN bactérien …………………………………………………………….47
II.2.7.2. PCR standard ……………………………………………………………………………47
II.2.7.3. Electrophorèse sur gel d’agarose………………………………………………………...55
II.3. Matériel végétal……………………………………………………………………….........56
II.3.1. Extraction des huiles essentielles…………………………………………………..............57
III.3.2. Analyse chimique des huiles essentielles………………………………………………....57

II.3.3. Evaluation de l’activité antibactérienne …………………………………………………...58
II.3.4. Étude de la combinaison des huiles essentielles avec les antibiotiques ……………….......61
II.3.5. La cinétique de destruction d’E. coli exposé aux huiles essentielles ………………….......62
Partie III : Résultats et discussions …………………………..63
III.1. Examens macroscopiques des urines ……………………………………………………….63
III.2. Résultats de la bandelette urinaire …………………………………………………………63
III.3. Résultats bactériologiques ………………………………………………………………....64
III.4. Fréquences des urines infectées en fonction du sexe……………………………………….65
III.5. Fréquences des urines infectées et non infectées en fonction de l’âge ……………………..66
III.6. Répartition des urines infectées et non infectées en fonction du lieu de prélèvement..........66
III.7. Étude de la résistance des entérobactéries isolées aux ntibiotiques………………………...67
III.8. Étude de la résistance d’E. coli isolée aux antibiotiques …………………………………..69
III.9. Phénotypes de résistance d’E. coli aux β-lactamines ………………………………………70
III.10. Biotypage des souches d’E. coli BLSE selon les profils numériques en API 20E
………..72
III.11.Répartition des profils numérique………………………………………………………….72
III.12. Répartition des souches d’E.coli BLSE en fonction des services………………………...73
III.13. Répartition des souches d’E.coli BLSE en fonction sexe ………………………………...73
III.14. Répartition des souches d’E. coli BLSE en fonction d’âge ……………………………..74
III.15. Répartition des patients infectés par E.coli BLSE en fonction des antécédents
médicaux…………………………………………………………………………………….........75
III.16. Fréquence d’E.coli BLSE durant les trois années d’étude ……………………………….75
III.17. Concentrations minimales inhibitrices de céfotaxime contre E. coli BLSE ……………...76
III.18. Profil moléculaire de la résistance d’E.coli BLSE aux β-lactamines...……………...........77
III.19. Recherche moléculaire de la résistance croisée……………………………………………79
III.20. Effet des HEs des plantes aromatiques retenues sur les souches d’E. coli BLSE ………...90
III.21. Étude de la combinaison des huiles essentielles avec la céfotaxime………………...........101
III.22. Cinétique de destruction d’E. coli avec les huiles essentielles……………………………102
Conclusion générale……………………………………………………………………………..105
Référence Bibliographique……………………………………………………………………..107
Annexes
Communications et Publication

Liste des tableaux
Tableau 1: Caractères biochimiques d’Escherichia coli ………………………………………………9
Tableau2 : Le traitement de la cystite aigüe...........................................................................………..20
Tableau 3: Le traitement des pyélonéphrites, prostatites et de la cystite compliquée de l’homme
………………………………………………………………………………………………………….21
Tableau 4: Le traitement de la pyélonéphrite simple …………………………………………............22
Tableau 5: Répartition des résistances d’E. coli aux ATB les plus fréquemment utilisés
dans le traitement des cystites simples ………………………………………………………………....23
Tableau6 : Phénotypes de résistance des entérobactéries aux aux β-lactamines ……………………...29
Tableau 7: Nomenclature des terpènes ………………………………………………………………..33
Tableau 8 : Composition chimique de l’huile essentielle de Coriandrum sativum ……………...........41
Tableau 9 : Composition chimique de l’HE essentielle de Ziziphora hispanica ……………….............42
Tableau 10 : Constituants chimiques de l’HE de C. cassia …………………………………………...43
Tableau 11: Amorces utilisées pour l’amplification des gènes de résistance aux β-lactamines
………………………………………………………………………………………………………….48
Tableau 12 : Les conditions d’amplification des gènes codants pour des bêta-lactamases…………...49
Tableau 13 : Amorces utilisées pour l’amplification des gènes plasmidiques de type AmpC………..49
Tableau 14 : Les conditions d’amplification des gènes codants pour AmpC…………………............50
Tableau15 : Amorces utilisées pour l’amplification des gènes de résistance aux quinolones ……......50
Tableau 16 : Les conditions d’amplification des gènes codants pour les quinolones………………....51
Tableau17 : Amorces utilisées pour l’amplification de gènes de résistance aac(6’)-Ib ………………51
Tableau 18 : Les conditions d’amplification de gènes de résistance aac(6’)-Ib ………………………51
Tableau19 : Amorces utilisées pour l’amplification des gènes de résistance aux aminosides………...52
Tableau 20: Les conditions d’amplification des gènes codants pour les aminosides………………….52
Tableau 21: Amorces utilisées pour l’amplification des gènes de virulence…………………………..53
Tableau 22 : Les conditions d’amplification des gènes codants pour les gènes de virulence53
Tableau 23 : Les amorces d’ERIC-PCR utilisées…………………………………………………......54
Tableau 24 : Les amorces utilisées pour groupement phylogénétique……………………………......54
Tableau25 : Les conditions d’amplification des gènes codants pour le groupement
phylogénétique………………………………………………………………………………………….55

Tableau 26 : Données sur les espèces végétales retenues et leurs utilisations………………………..56
Tableau 27: Répartition selon l’aspect macroscopique des urines d’origine hospitalière…………….63
Tableau 28: Résultat de la chimie des urines analysées d’origine hospitalière……………………….63
Tableau 29: Fréquences des urines infectées d’origine hospitalière………………………………….64
Tableau 30: Fréquences des urines infectées et non infectées en fonction du sexe…………………...65
Tableau 31 : Fréquences des urines infectées en fonction des tranches d’âge………………………..66
Tableau 32: Répartition des urines infectées et non infectées en fonction
du lieu de prélèvement………………………………………………………………………………….67
Tableau 33 : Comparaison des résistances bactériennes d’E. coli…………………………………….70
Tableau 34 : Biotypage des souches d’E. coli BLSE selon les profils numériques en API
20E………………………………………………………………………………………………..........72
Tableau 35 : Concentrations minimales inhibitrices (µg/mL) de céfotaxime contre E.coli
BLSE…………………………………………………………………………………………………..76
Tableau 36 : Profil moléculaire des 27 souches E. coli BLSE………………………………………78
Tableau 37: Les gènes qui codent pour les enzymes modificatrices des aminosides…………...........79
Tableau 38 : Les 16S ARNr methyltransphérases…………………………………………………….80
Tableau 39: Profil moléculaire de la résistance associée des souches d’E.coli BLSE aux
quinolones……………………………………………………………………………………………..81
Tableau 40 : Phénotype des facteurs de virulence……………………………………………............87
Tableau 41: Relation épidémiologique entre les 27 souches d’E.coli BLSE étudiées par ERIC/
PCR et groupement phylogénétique ………………………………………………………………….89
Tableau 42: Rendements en HE et caractéristiques organoleptiques…………………………...........90
Tableau 43 : Rendements en HE rapportés dans la littérature………………………………………..91
Tableau 44: Composition chimique (%) des huiles essentielles des plantes retenues…………….....92
Tableau 45 : Composés majoritaires des trois HEs rapportés dans la littérature………………….....93
Tableau 46 : Diamètres d’inhibitions (en mm) des huiles essentielles et de céfotaxime……………94
Tableau 47 : Concentrations minimales inhibitrices des huiles essentielles (mg/ml) et
de céfotaxime (μg/ml)………………………………………………………………………………...96
Tableau 48 : Evaluation de la formation du biofilm in vitro ……………………………………......98

Tableau 49. Effet des HEs étudiées vis–à–vis des souches d'E. coli en état du biofilm,
exprimées par les concentrations minimales inhibitrices (CMIB) et éradicatrices (CMEB)
du biofilm en mg/ml………………………………………………………………………………..99
Tableau 50 : CFI des huiles essentielles de Cinnamomum cassia et Coriandrum sativum…......102

Liste des figures
Figure 1 : Microphotographie d’E. Coli obtenue par microscopie électronique à balayage………..8
Figure 2 : Les différentes étapes d'infection urinaire à Escherichia coli..………………………... 12
Figure 3 : Physiopathologie de l’infection urinaire………………………………………………...13
Figure 4 : Structure de l'unité isoprénique………………………………………………………….32
Figure 5 : Cellules d’E. coli avant et après le traitement avec huile essentielle d’Origan,
sous microscope électronique a balayage ……………………………………………………….…35
Figure 6 : Photo de Coriandrum sativum ……………………………………………………….....41
Figure 7 : Photo de Ziziphora hispanica…………………………………………………………...42
Figure 8 : Photo de Cinnamomum cassia ………………………………………………………….43
Figure 9: Réalisation d’un test de synergie………………………………………………………...46
Figure 10 : Photo de l’appareil utilisé pour l’extraction des huiles essentielles par
hydrodistillation……………………………………………………………………………………..57
Figure 11 : Principe de la méthode de diffusion par disque …………………………………….....59
Figure 12 : Répartition des bactéries responsables d'infections urinaires d’origine hospitalière en
fonction de l'espèce………………………………………………………………………………….64
Figure 13: Pourcentage de résistance aux antibiotiques des entérobactéries isolées d’urines au
CHU de Tlemcen……………………………………………………………………………………68
Figure 14 : Pourcentage de résistance d’E. coli aux antibiotiques……………………………......69
Figure 15 : Fréquences des phénotypes de résistance aux β-lactamines des souches d’E .coli
isolées d’urines infectées au CHU de Tlemcen…………………………………………………….70
Figure 16 : Répartition des profils numériques d’E. coli BLSE par service (CHU
Tlemcen)……………………………………………………………………………………............72
Figure17 : Répartition des souches d’E.coli BLSE en fonction des services au CHU de
Tlemcen……………………………………………………………………………………………..73
Figure 18 : Répartition des souches d’E. coli BLSE en fonction de sexe…………………….......74
Figure 19 : Répartition des souches d’E.coli BLSE en fonction d’âge……………………...........74
Figure 20 : Répartition des patients infectés par E.coli BLSE en fonction des antécédents
médicaux……………………………………………………………………………………...........75
Figure 21: Fréquence d’E.coli BLSE responsables d’IU durant les trois années d’étude (CHU de
Tlemcen)………………………………………………………………………………………........76
Figure 22a : détection des gènes bla CTXM15 ……………………………………………...........77
Figure 22b : détection des gènes bla TEM1………………………………………………………..77

Figure 23 : Les enzymes modificatrices des aminosides…………………………………………..79
Figure 24 : Effet de l’huile essentielle de Cinnamomum cassia sur Pseudomonas aueruginosa
………………………………………………………………………………………………………………...95
Figure 25 : Cinétique de destruction de la souche Ecoli BLSE 09 exposée à l’huile essentielle de
Cinnamomum cassia pendant 15, 30, 45 et 75 mn à la concentration de CMB= 5 mg/ml………..102
Figure 26 : Cinétique de destruction de la souche Ecoli BLSE 09 exposée à l’huile essentielle de
Coriandrum sativum pendant 15, 30, 45, 75,90,120,150,180 et 240 mn aux concentrations CMB=
10, 20 et 40 mg/ml…………………………………………………………………………………103

Liste des abréviations
A.F.S.S.A.P.S : Agence Française de Sécurité Sanitaire et des Produits de Santé
ACC : Ambler class C
ADN : acide désoxyribonucléique.
AMC : Amoxiciline+ acide Clavulamique
AME: Aminoglycosides modifying enzymes
AmpC : céphalosporinase
API 20E: Analytical profile index 20E (E= Entérobactéries)
Arm: Aminoglycoside resistance methylase
ATB : Antibiotique
ATM : Aztreonam
ATP: Adénosine Triphosphate
BES : Brazilian extended-spectrum
BHIB : Bouillon Cœur cervelle
BLSE : béta-lactamases à spectre étendu.
C3G : céphalosporine de 1ere, 2eme, 3eme et 4eme génération
CASE: céphalosporinase
CC : complexe clonal
CFI : Concentration fractionnelle inhibitrice
CG/SM : chromatographie en phase gazeuse couplée à la spectrométrie de masse
CHU : Centre hospitalier universitaire
CIP : Ciprofloxacine
CIT :Gène codant pour une céphalosporinase
CLIN: Comité de lute contre les infections nosocomiales
CLSI : Clinical and Laboratory Standard Institute
CMB: Concentration(s) Minimale(s) Bactericide
CMEB : Concentration minimale éradicatrice du biofilm
CMI : Concentration minimale inhibitrice.
CMIB : Concentration minimale inhibitrice du biofilm
CMY : Cephamycinase (gène codant pour une céphalosporinase plasmidique
CNF1: cytotoxic necrotizing factor 1
CTIN : Comité Technique national des Infections Nosocomiales

CTX-M : cefotaximase-Munich
DHA : Dharhan hospital
DMSO : Diméthyl sulfoxyde
dNTP : désoxyribonucléotide triphosphate
DO : Densité optique
E. coli : Escherichia coli
EARS-Net : European Antimicrobial Resistance Surveillance Network
EBLSE: Entérobactéries productrices de β-lactamases à spectre étendu.
ECBU : examen cytobactériologique urinaire
ECDC :European Center for Disease prevention and Control
EDTA : acide éthylène diamine tétraacétique
ERIC-PCR : Enterobacterial Repetitive Intergenic Consensus- Polymerase chain reaction
FOS : Fosfomicine
FOX : Gène codant pour une céphalosporinase
FQ : Fluoroquinolones.
H.E : Huiles essentielles
hly :hémolysine
IU : Infection urinaire
IUN : Infection urinaire nosocomiale
LCR: Liquide céphalorachidien
LPS: Lipopolysaccharide
M P: Marqueur de poid
Mgcl2 :magnesium chloride
MH: Muller Hinton
MOX : Gène codant pour une céphalosporinase
ORL : Oto-rhino-laryngologie.
OXA : oxacillinase
PASE: pénicillinase
PB : Paires de bases
PCR : Polymérase Chain réaction.
PER : Pseudomonas extended resistance
PLP: Protéines de liaison aux pénicillines
PNA : La pyélonéphrite aiguë
qepA : quinoloneef-ffux pump

qnr : Quinolone resistance gene
QRDR : Quinolone Resistance Determining Region
Rmt: ARNr 16S méthyltransférase
rpm : rotation par minute
sfa:S-family adhesions
SFO : Serratia fonticola
SHV : sulfydryl variable
SPILF : Société de Pathologie Infectieuse de Langue Française
SXT : Triméthoprime + Sulfaméthoxazole.
TBE : TRIS Acide borique EDTA
TEM : TEMoneira- nom du patient
Ufc : unité formant colonie
VEB : Vietnam extended-spectrum
VIM : Verona integron–encoded metallo-β-lactamase

Introduction

Introduction
1
Introduction
Les infections en générale posent un véritable problème de santé publique du fait de leur
fréquence, leur gravité et leur coût socioéconomique. Leurs traitements par les antibiotiques
restent le moyen de choix, mais l’émergence de bactéries résistantes pose un problème
d’inefficacité de ces molécules anti-infectieuses. Dans les établissements de soins, les
infections dites nosocomiales sont préoccupantes du fait de la libre circulation des bactéries.
Les infections urinaires représentent un exemple élucidant des infections
nosocomiales contractées à l’hôpital. Elles sont plus fréquentes dans les services de moyen
séjour tels que le service d’urologie, où ces infections, sont souvent la résultante d’un certain
nombre de facteurs tels que les pratiques de soins, la flore liée au patient ou encore
l’environnement.
Les micro-organismes les plus incriminés dans les infections urinaires sont les bacilles
à gram négatif, hôtes naturels de l’intestin et de l’environnement, dont Escherichia coli
est la plus incriminée. Le moyen de lutte le plus fréquemment utilisé est l’antibiothérapie,
mais la prescription massive et probabiliste de ces molécules a été rapidement suivie par
l’émergence de bactéries résistantes. Même l’utilisation de nouvelles molécules de quinolones
s’est avérée inefficaces suite aux mécanismes de résistances développées par les bactéries
(Lazrak, 2014).
Les mécanismes sont (i) soit de natures enzymatiques telles que les β-lactamases qui
inactivent les β-lactamines par modification chimique ou (ii) soit non enzymatiques causés
par des mutations ou par des gènes d’origine inconnue qui s’intègre au niveau du génome
bactérien (Gutmann, 1987).
A ce jour, plus de 600 variantes de BLSE ont été décrites à travers le monde. On
distingue 11 familles différentes : TEM, SHV, CTX-M, PER, VEB, GES, TLA, BES, SFO,
FEC et OXA. Les BLSE les plus anciennes dérivent de TEM- 1,2 et de SHV-1 (Geser et al.,
2012).
Les souches d’E. coli productrices de beta-lactamases à spectre élargi (BLSE) sont
apparues dans les années 1980 (Knothe et al., 1983 ; Bradford, 2001). Elles sont dominées
actuellement par la production de BLSE de type CTX-M. Le problème d’échec thérapeutique
découle aussi des résistances associées, puisque 80% d’E. Coli CTX-M sont résistantes aux
fluoroquinolones de nature plasmidique attribuer aux gènes qnr ou encore à l’enzyme AAC6’-
1b-cr (López-pujol et al., 2011).

Introduction
2
Les infections causées par E. coli (BLSE) sont associées à une morbidité et une
mortalité élevées, à une prolongation de la durée de l’hospitalisation et à une augmentation
des coûts d’hospitalisation (Patterson, 2001 ; Masterton et al., 2003).
La fréquence élevée des bactéries résistantes aux antibiotiques complique la conduite
thérapeutique de cette pathologie, et justifie d’une part (i) d’une évaluation de l’efficacité de
ces médicaments et (ii) d’autre part la recherche de nouvelles molécules antimicrobiennes.
Parmi les sources prometteuses de molécules bioactives, les plantes aromatiques et
médicinales.
Jadis, les plantes étaient le seul moyen pour traiter les infections mêmes avant de
découvrir les microbes. Aujourd’hui, 80% des populations à travers le monde utilisent les
plantes médicinales pour se soigner, par manque d’accès aux médicaments prescrits par la
médecine moderne mais aussi parce que ces plantes ont souvent une réelle efficacité (Dibong
et al., 2011). Ils restent aux scientifiques de préciser les propriétés et de valider les usages des
plantes aromatiques et médicinales qui constituent un réservoir naturel de molécules
bioactives (Pelt, 2001).
Des travaux antérieurs ont montré l’effet antimicrobien des extraits des plantes sur les
microorganismes. L’exemple de la canneberge (Vaccinum macrocarpon) est aujourd’hui
reconnu dans le traitement des IU. D’ailleurs l’Agence française de sécurité sanitaire des
produits de santé (A.F.S.S.A.P.S) lui reconnaît depuis 2004 les propriétés suivantes :
« Contribue à diminuer la fixation de certaines bactéries Escherichia coli sur les parois des
voies urinaires ». L’importance de cette plante est liée aux pro-anthocyanidines et à la
présence des substances anti-oxydantes (Peyramaure, 2008). Cependant, peu de travaux ont
été réalisés sur un exemple clinique bien déterminé tel qu’E. coli (BLSE).
Dans le même axe de recherche de molécules bioactives issues des plantes aromatiques
et médicinales de la région de Tlemcen, nous nous sommes intéressé à l’étude de l’effet
inhibiteur de trois huiles essentielles Coriandrum sativum (kasbour) ; Ziziphora hispanica
(Fliou) et Cinnamomum cassia (qirfa) sur E. coli (BLSE) responsables d’infections urinaires
d’origine hospitalière. Les objectifs de ce travail sont la mise en évidence de la prévalence de
résistance d’E. coli aux antibiotiques, le typage moléculaire des souches isolées et surtout
l’évaluation de l’effet inhibiteur des huiles essentielles retenues sur E. coli (BLSE).
Le travail est structuré de la façon suivante :
La première partie est une synthèse bibliographique sur les infections urinaires et les
huiles essentielles.

Introduction
3
La deuxième partie consiste à la présentation du matériel et les méthodes utilisés dans
ce travail.
Et enfin, la troisième partie, sera consacrée à la présentation des résultats et à leurs
discussions.

Synthèse
bibliographique

Synthèse bibliographique
4
Partie I : Synthèse bibliographique
Chapitre I : Les infections urinaires
I.1. Généralités sur les infections urinaires
I.1.1. Infection urinaire et colonisation
A l’état normal, l’urine est stérile. Les infections urinaires (IU) se traduisent par la présence de
germes pathogènes dans l’urine à l’intérieur des voies excrétrices. Elles se définissent à l’aide de
critères biologiques: une bactériurie supérieure à 105/ml, une leucocyturie supérieure à 10/mm
3 ou
104/ml avec la présence d’au moins 10
5 colonies par ml, qu’il y ait ou non des signes cliniques
d’accompagnement (Gonthier et Heritier, 1988).
D'après la définition établie lors de la conférence de consensus organisée par la société de
pathologie infectieuse de langue française et l’association française d'urologie (SPILF et l’AFU,
2002 ; Clere, 2012), on parle d'infection urinaire si l'on est en présence d'au moins un des signes
cliniques suivants :
fièvre (> 38°C),
impériosité mictionnelle (envie pressante),
pollakiurie (envie fréquente) : la fréquence des mictions est plus élevée (> 8 mictions par 24
heures et/ou > 2 mictions nocturnes) mais le volume d'urine journalier n'est pas augmenté,
brulures mictionnelles ou douleurs sous-pubiennes (pesanteur pelvienne),
douleurs lombaires.
de dysurie (difficulté a la miction : évacuation lente et difficile, voire douloureuse des
urines, avec diminution du débit urinaire et sensation que la vessie ne se vide pas),
d'hématurie (sang dans les urines) en fin de miction dans 30% des cas,
d'urines troubles (pyurie ou leucocyturie) et malodorantes.
Selon Lobel et Soussy, (2007), l'infection urinaire (IU) correspond a l'agression d'un tissu par
un (ou plusieurs) microorganisme (s), générant une réponse inflammatoire et des symptômes de
nature et d'intensité variable selon le terrain. L’IU regroupe les infections des différents constituants
de l'appareil urinaire ou de certaines annexes comme :
l'urétrite : inflammation de l'urètre, considérée comme une maladie sexuellement
transmissible,
la cystite : inflammation de la vessie,
la pyélonéphrite : inflammation des reins,
la prostatite : inflammation de la prostate,
l'orchi-épididymite : inflammation de l’épididyme et des testicules.

Synthèse bibliographique
5
En général, la fièvre et les douleurs lombaires (souvent unilatérales) sont le signe d'une
atteinte parenchymateuse, c'est à dire d'une pyélonéphrite ou d'une prostatite (Leroy et Tattevin,
2012 ). A ces signes peuvent être associés des frissons, des sueurs et une altération de l'état général
( Rostoker et al., 1991).
La colonisation est à différencier de l'IU, elle correspond à la présence d’un (ou de plusieurs)
micro-organisme dans l’arbre urinaire sans qu’il ne génère par lui-même de manifestations
cliniques. Le concept de bactériurie asymptomatique est indissociable de celui de colonisation et
correspond à la même entité sans le rattacher à une notion de seuil (ufc/ml). Le terme de
colonisation est préférable à celui de bactériurie asymptomatique (Vildé et al., 2002).
1.1.2. Infection urinaire simple et compliquée
Depuis la parution des dernières recommandations de l’Agence française de sécurité sanitaire
des produits de santé en la matière (2008), les termes d’infections urinaires hautes ou basses, ont été
abandonnées. On emploie plus volontiers aujourd’hui les notions d’infections urinaires simples ou
compliquées. Ces recommandations font apparaitre de nouvelles stratégies thérapeutiques qui
tiennent compte de l’évolution des résistances des microorganismes les plus fréquemment retrouvés
dans les infections urinaires.
a. Les infections urinaires simples
Ce sont des infections urinaires survenant chez des patients ne présentant pas de facteur de
risque de complication ou de morbidité. L’âge (au-delà de 65 ans) n’est plus considéré comme un
facteur de risque de complication en l’absence de terrain problématique. Les IU dites simples
comprennent la cystite aigue simple et la pyélonéphrite simple (AFSSAPS, 2010).
b. Les infections urinaires compliquées
Elles surviennent chez des patients présentant au moins un facteur de risque de complication,
parmi les suivants :
Anomalie organique ou fonctionnelle de l’arbre urinaire (lithiase, tumeur, reflux…)
Certaines pathologies comme le diabète, l’insuffisance rénale, l’immunodépression
Un terrain particulier (grossesse, sexe masculin).
Les IU dites compliquées comprennent la cystite compliquée et la pyélonéphrite compliquée
(AFSSAPS, 2010).
Toute infection urinaire survenant chez l’homme est automatiquement considérée comme
compliquée et gérée comme une prostatite aigue (inflammation de la glande prostatique d’origine
bactérienne). Les infections urinaires survenant chez la femme enceinte sont également considérées
comme des IU compliquées (cystite aigue gravidique et pyélonéphrite aigue gravidique)
(AFSSAPS, 2010).

Synthèse bibliographique
6
I.1.3. Infection urinaire récidivante
Les IU sont dites récidivantes lorsqu'il y a au moins 4 épisodes par an ou un épisode datant de
moins de 3 mois (AFSSAPS, 2008). Elles concernent en général les femmes jeunes, en bonne santé,
sans anomalies fonctionnelles ou anatomiques du tractus urinaire. Les IU récidivantes sont malgré
tout à considérer comme des IU compliquées (Barrier Letertre, 2013).
Parmi ces IU, on distingue les rechutes (IU symptomatique causée par la même bactérie après
un traitement adéquat, ayant lieu moins d'un mois après la première IU), des réinfections (IU
symptomatiques causées par une autre bactérie, ou par la même bactérie après un traitement adapté
et un ECBU de contrôle négatif, survenant plus d'un mois après la première IU) ( Kodner et
Gupton, 2010). Ce dernier cas, réinfection par la même bactérie, est le plus fréquent. Le caractère
récurent d'une IU dépend surtout de la bactérie et moins des facteurs de risques (Foxman, 1990).
I.1.4. Infection urinaire communautaire et nosocomiale
Les IU communautaires surviennent en dehors d'une structure de soins (Marrhich, 2008).
Par contre les IU nosocomiales sont acquises dans une structure de soins, ou bien reliée à la prise en
charge du patient, et causée par des microorganismes dont l'origine est hospitalière (SPLIF et AFU,
2002 ; Eilenberg, 2005). L’IU nosocomiale peut concerner les personnes séjournant, visitant ou
travaillant à l'hôpital (Eilenberg, 2005). La fréquence est variable selon les résidents avec sonde et
sans sonde. De manière générale, les IU nosocomiales touchent tous les services hospitaliers. Les
principaux services touchés dans les hôpitaux sont : le service de chirurgie urologique, réanimation
médicale, pédiatrie, rééducations fonctionnelle de l’adulte et de la personne âgée, maternité et le
service de gynécologie (Vildé et al., 2002).
Les germes les plus fréquemment rencontrés à l’hôpital sont les bacilles Gram négatif avec
Escherichia coli en tête, Proteus, Klebsiella, Enterobacter, Serratia et Pseudomonas. Les germes
sont souvent multi-résistants du fait de nombreux traitements antibiotiques antérieurs.
La contamination se fait par la flore rectale ou lors de manipulations exogènes (Faucher et
Cudennec, 2002 ; SPILF et AFU, 2002).
Il existe peu d’études sur la morbidité des infections urinaires car elle est difficile à évaluer
lors d’un séjour hospitalier en raison des poly pathologies et autres infections qui peuvent entraîner
le décès. Le rôle de la bactériurie asymptomatique dans la morbidité génito-urinaire chez les sujets
âgés en institution est faible en comparaison de sa prévalence élevée. Il n’y a pas d’argument pour
un lien causal entre bactériurie asymptomatique et mortalité dans la population âgée dans les
dernières études. Les IU sont davantage un marqueur de maladie qu’une cause d’augmentation de la
mortalité. La mortalité de la pyélonéphrite est faible si la prise en charge adaptée est précoce. C’est
cependant la cause la plus fréquente de septicémie et de choc infectieux chez le sujet âgé. Les décès

Synthèse bibliographique
7
par septicémie à point de départ urinaire correspondent à moins de 10 % des décès totaux (Faucher
et Cudennec, 2002).
I.1.5. Epidémiologie et facteurs de risque
L’infection de l’appareil urinaire est fréquente chez la femme. 6 à 10 % des fillettes et femmes
jeunes ont une bactériurie. Ce taux augmente avec l’âge pour atteindre 20 % chez la femme adulte
et 25 à 50 % chez la femme de plus de quatre-vingts ans. Ainsi une femme sur trois aura une
infection urinaire avant l’âge de vingt-quatre ans, 40 à 50 % auront une infection au cours de leur
vie et 20 à 30 % d’entre elles ayant eu un premier épisode, récidiveront dans les trois à quatre mois
chez le sujet âgé. D’autres facteurs de risque sont responsables de l’IU comme la grossesse, le
diabète et l’existence d’anomalies urologiques (Sourander, 1966 ; Raz, 2001 ; Foxman, 2002).
I.2. Mode de pénétration des bactéries dans les voies urinaires
L’urine vésicale normale est stérile. Cependant on rencontre des bactéries de façon
permanente surtout chez la femme. La bactériurie dépend de 3 phénomènes :
- la vitesse de pénétration des bactéries dans la vessie,
- la vitesse de croissance de ces bactéries,
- la vitesse d’élimination ou de la destruction des bactéries (kodio, 1979 ; Fauchere, 1997 ; Epok,
1999).
I.2.1. La voie ascendante (Chartier, 2001)
La voie ascendante est la plus fréquente. A partir du méat (physiologiquement porteur de
Staphylocoques et de Streptocoques, mais pas de bacilles Gram -, les germes peuvent remonter vers
la vessie. Chez la femme, la brièveté de l’urètre, la proximité de l’anus et la tendance des bactéries
du rectum (bacilles Gram -) à coloniser le périnée, prédisposent à cette migration. L’anatomie du
bas appareil urinaire de la femme avec un urètre court et la proximité de la cavité vaginale explique
la possibilité de transmission des germes des régions rectales et vaginales à la vessie. La miction
ayant un rôle de « chasse » des germes remontants vers la vessie, toute perturbation de celle-ci
favorisera aussi cette ascension. Chez l’homme, l’urètre situé à distance de l’anus et les sécrétions
prostatiques antibactériennes (riches en zinc) rendent cette migration peu fréquente.
Au cours des infections urinaires basses, une fois la vessie contaminée, la persistance des
microorganismes est liée à leur capacité d’adhésion à l’épithélium vésical. Les bactéries présentes
dans la vessie engendrent une réponse inflammatoire qui se manifeste par la présence de
polynucléaires dans la vessie. Chez la femme comme chez l’homme, les infections urinaires sont
favorisées par :
les sondes à demeure,
tout obstacle à l’écoulement de l’urine (lithiase),

Synthèse bibliographique
8
l’état grabataire (pour les malades qui ne quittent pas le lit).
I.2.2. La voie hématogène :
Moins fréquente, elle survient lors d’une septicémie ou lors d’une bactériémie, surtout chez
l’immunodéprimé et le diabétique. La porte d’entrée infectieuse, inconstamment retrouvée peut être
variable : cutanée, O.R.L., dentaire (Chartier, 2001).
I.2.3. Extension directe à partir d’un organe de voisinage (voie lymphatique) :
Exemple de la maladie inflammatoire de l’intestin, suppuration pelvienne aiguë chez la
femme, abcès para vésical (Chartier, 2001).
I.3. Les infections à E. coli
I.3.1. Généralités
L’espèce E. coli, également appelé colibacille, est un bacille Gram-négatif (figure 1) de la
famille des Enterobacteriacae. Découvert en 1885 par Théodore Escherich dans les selles des
nourrissons, c’est un commensal commun de la flore intestinale de l’homme et des animaux à sang
chaud. Son établissement dans le tractus digestif s’effectue durant les premières heures suivant la
naissance (Kaper et al., 2004).
Figure 1: Microphotographie d’E. coli obtenue par microscopie électronique à balayage
(Clément, 2012).
Certaines souches peuvent devenir pathogènes par acquisition de gènes de virulence et elles
sont capables de causer des infections intestinales et extra intestinales à l’aide des facteurs de
virulence qui perturbent les processus cellulaires normaux (Kaper et al., 2004). Les facteurs de
virulence qui distinguent les différents pathotypes d’E.coli sont codés par des plasmides,
transposons, phages et les génomes des autres bactéries par transfert horizontal ou vertical des
gènes. L’îlot de pathogénicité peut-être identifié dans les souches pathogènes car il possède une
composition différente du reste du génome bactérien ce qui indique leur provenance d’autres
espèces (Whittan, 1996). E. coli est l’organisme le plus étudié dans les laboratoires de biologie

Synthèse bibliographique
9
moléculaire et constitue un modèle pour les études biochimiques et génétiques grâce à sa
découverte précoce et sa culture aisée (Donnenberg et Whittam, 2001).
I.3.2. Caractères morphologiques et culturaux
E.coli est une bactérie asporulée mesurant 2 à 4 μm de long sur 0,4 à 0,6 μm de large. C’est
une bactérie fine et allongée à extrémités arrondies, mobile grâce à une ciliature péritriche. Ce
germe non exigeant, sur gélose ordinaire donne des colonies lisses, brillantes et homogènes (Lobril,
1998).
I.3.3. Caractères biochimiques
E. coli possède une catalase mais est dépourvu d’oxydase. L’étude de l’activité enzymatique
et de la fermentation des sucres est réalisée à l’aide de micro-méthodes validées disponibles dans le
commerce sous forme de galeries (Gueye, 2007 ; Sibery et al., 2001). E. coli fermente le glucose et
le lactose avec une production de gaz, il est dépourvu d’une uréase, produit de l’indole, n’utilise pas
le citrate de Simmons comme source de carbone et ne produit pas d’hydrogène sulfuré. Ces
différents tests sont regroupés dans le tableau 1 (Gueye, 2007).
Tableau 1: Caractères biochimiques d’Escherichia coli (Gueye, 2007)
tests ADH B
Gal
CC CS Gel IND MAL PDA LDC ODC TDA URE NIT GLU LAC VP ESC H2S
résultats +∕- + + - - + - +∕- + + + +∕- - + + - - -
Légende : (+) Caractère positif ; (-): Caractère négatif ; (+/-): caractère inconstant.
ADH : Arginine dihydrolase ; β Gal: Bêta galacto-D pyranoside ; CC : Citrate de Christensen, CS :
citrate de Simmons ; Gel : Gélatinase ; H2S : Hydrogène sulfuré ; IND : Indole ; MAL : Maltose ;
PDA : Phényle alanine désaminase ; LDC : Lysine décarboxylase ; ODC : Ornithine
décarboxylase ; URE : Uréase ; NIT : Nitrate réductase ; VP : Réaction de Voges Proskauer ; TDA
: Tryptophane désaminase, GLU : Glucose, LAC : Lactose
Souches atypiques
Se sont des souches mutantes qui ont perdu ou acquis un caractère biochimique non habituel
chez l’espèce E. coli. Exemples :
Des variant indole - : pour ces souches, l’indole est le seule caractère qui a muté. (Le minor,
1993). A la différence des souches appartenant à l’espèce Escherichia fergusonii qui sont : indole -,
LDC -, ODC - (Pelmont, 1995).
Des variant H2S+, Uréase+, ces caractères sont codés par un plasmide qui peut déterminer une
résistance à un antibiotique tel que la tétracycline (Le minor, 1993 ; Pelmont, 1995).

Synthèse bibliographique
10
Alkalescens-dispar (A.D) : Auparavant les variants immobiles et agazogènes d’E. coli
portaient le nom de Alkalescens-dispar et étaient classés avec les Shigella. Aujourd’hui, on leurs
reconnaît le même pouvoir, la même écologie que ceux des autres E. coli, d’où leur intégration dans
ce genre. (Le minor, 1989 ; 1993). Les souches E.coli sont isolées des selles d’individus en bonne
santé. Ils sont immobiles, agazogènes, fermentent tardivement le lactose, ou lactose négative ;
ONPG négative.
I.3.4. Caractères antigéniques
Au sein de chaque genre, on individualise les espèces par l’étude des caractères
biochimiques et antigéniques. Les entérobactéries possèdent toutes, des antigènes de paroi
(somatiques) ou antigènes O. Les espèces mobiles quand à elles renferment en plus des antigènes O,
des antigènes H ou flagellaires : C’est le cas d’E. coli (Lobril, 1998 ; Edler, 2001 ; Flaudrois,
2004).
I.3.5. Pouvoir pathogène
Les entérobactéries sont des taxons isolées dans de nombreux aliments (céréales), et dans
divers produits pathologiques (urine, LCR, selles……). E. coli est souvent responsable de gastro-
entérites graves pouvant être mortelles dans certains cas à l’absence de traitement. Il est classé dans
le groupe des entérobactéries pathogènes spécifiques avec les Shigelles et les Salmonelles qui sont
responsables de dysenteries et fièvres typhoïdes graves (Collignon, 2002).
Il existe divers d’E coli à mécanismes d’actions différentes :
- ETEC : Enterotoxinogen E. coli, responsable de la « diarrhée des voyageurs » ou « turista » et des
syndromes épidémiques dans les pays du Tiers-monde ;
- EIEC : Enteroinvasive E. coli, encore appelé E. coli Shigella-like, responsable de syndromes
dysentériques avec invasion de la muqueuse intestinale ;
- EHEC : Enterohaemorragic E. coli, responsable de diarrhées sanglantes liées à la production de
toxines ;
- EPEC : Enteropathogen E. coli, responsable de gastro-entérites infantiles. (Perrière, 1992 ;
Collignon, 2002).
Infections urinaires
E. coli représente à lui seul l'agent responsable de la très grande majorité de cas d'infection
urinaire spontanée ou après instrumentation. L'infection urinaire basse à E .coli est vulgairement
appelée colibacillose. En fait, l'origine de l'infection est intestinale (infection par voie ascendante),
favorisée chez la femme par l'anatomie du bas appareil urinaire (urètre court) et par la fréquence des
rapports sexuels. E. coli est aussi dominant dans le rectum et la sphère génito-urinaire (Flandrois,
1997).

Synthèse bibliographique
11
Septicémies et méningites
Les E. coli sont isolés dans 20 % des septicémies et représentent 45% des septicémies dues
aux bacilles à Gram négatif. Les méningites sont rares, elles surviennent surtout chez le nourrisson
mais sont souvent graves. 80 % des E. coli isolés de méningites possèdent l'antigène K1,
polysaccharide acide dont la composition chimique et la spécificité immunologique sont identiques
à celle de l'antigène B de Neissseria meningitidis (Flandrois, 1997).
Suppuration diverses
Les E. coli de la flore fécale peuvent être en cause dans des péritonites, des cholécystites, des
salpingites et des suppurations postopératoires jouant le rôle de bactéries pyogènes (Flandrois,
1997).
I.3.6. Les substances élaborées :
E. coli peut produire des entérotoxines, des hémolysines, des enzymes capables de détruire
certains antibiotiques et des bactériocines, substances à action antibiotique qui détruisent
spécifiquement d'autres bactéries qui sont aussi dénommées colicines (Ferron, 1994)
1.3.7. Physiopathologie de l’infection urinaire à Escherichia coli:
Escherichia coli uropathogène s’attache à l’uroépithélium par des pili de type 1, qui se lient
aux récepteurs Uroplakin Ia et IIIa (figure 2). Cette liaison stimule des voies de signalisation
inconnues qui interviennent dans l'invasion et de l'apoptose. La liaison du pili de type 1 à l’α3β1
intégrines est également le médiateur de l’internalisation des bactéries dans les cellules
superficielles pour former des communautés bactériennes intracellulaires. Des concentrations
sublytiques de la toxine porogène hémolysine A (HlyA) peuvent inhiber l'activation de protéines
Akt et conduire à l'apoptose de l'hôte cellulaire et l’exfoliation. L’exfoliation de l’uroépithélium
expose les cellules de transition sous-jacentes pour une autre invasion d’E. coli uropathogène, et les
bactéries peuvent résider dans ces cellules en tant que des réservoirs intracellulaires qui peuvent être
impliqués dans des infections récurrentes (Matthew et al., 2010).

Synthèse bibliographique
12
Figure 2 : Les différentes étapes d'infection urinaire à Escherichia coli (Matthew et al., 2010).
L’infection urinaire débute par la colonisation du tube digestif par une souche uropathogène
qui, grâce à la présence de facteurs de virulence, colonise l’aire périurétrale et migre le long de
l’urètre vers la vessie, puis le long de l’uretère vers le rein (figure 3). La migration d’E. coli en
dépit du flux urinaire requiert son attachement à la surface des cellules épithéliales par
l’intermédiaire des différentes adhésines. Les flagelles ne semblent pas jouer un rôle déterminant
dans la remontée des voies urinaires par E. coli. Parmi les nombreuses adhésines, deux jouent un
rôle majeur : les pili de type 1 pour la colonisation de la vessie et les pili de type P pour l’invasion
du parenchyme rénal. L’interaction de l’adhésine FimH des pili de type 1 avec les récepteurs mono-
mannose à la surface des cellules vésicales facilite l’internalisation de la bactérie et conduit à la
formation de communautés bactériennes intracellulaires dans une matrice de biofilm. Elles sont
ensuite reléguées dans la lumière vésicale ou restent quiescentes faisant le lit de la récidive de
l’infection urinaire. Cette internalisation et cette persistance d’E. coli dans les voies urinaires basses
seraient renforcées en présence de la toxine CNF (facteur cytotoxique nécrosant1). Face à
l’infection de la vessie, seule l’immunité innée peut entrer en action. Durant les deux premières
heures, elle est essentiellement liée à l’action des défensines. Par la suite, l’adhésion via les pili de
type 1 et les pili de type P va être à l’origine de la réponse inflammatoire. En l’absence de CD14
libre dans les urines, cette adhésion sert de cofacteur lors de la reconnaissance du complexe LPS
(lipoplysaccharide binding protein) par les TLR4 (Toll-like receptor 4) des cellules vésicales.
L’activation des TLR4 induit la sécrétion de cytokines chimiotactiques aboutissant à un afflux de
polynucléaires neutrophiles. Les bactériuries asymptomatiques sont reliées soit à une déficience de
l’hôte en TLR4, soit à un phénomène de dépiliation des E. coli intra vésicaux. L’expression

Synthèse bibliographique
13
continue de fimbriae de type 1 confine l’infection à la vessie. Les souches d’E. coli responsables de
pyélonéphrites aiguës bloquent leur synthèse de fimbriae de type 1 en invalidant le promoteur en
position « off », et peuvent ainsi migrer le long des uretères en dépit du flux urinaire. La migration
est favorisée par l’expression des fimbriae de type P. Le passage de la bactérie dans la circulation
sanguine serait facilité par la déstabilisation de l’épithélium rénal par des cytotoxines comme
l’hémolysine α ou la toxine Sat. Une fois dans la circulation sanguine, la bactérie se protège du
système immunitaire par les différentes structures (capsule, LPS) et par les protéines de surfaces
permettant notamment d’échapper à l’activité bactéricide du complément et à la phagocytose. Tout
au long de sa progression, aussi bien dans les urines que dans le sang, E. coli va devoir trouver les
éléments nécessaires à sa croissance et notamment le fer en utilisant les nombreux systèmes de
capture. (Bonacorsi et al., 2014 ; Bidet et al., 2012)
Figure 3 : Physiopathologie de l’infection urinaire (Bidet et al., 2012)
I.4. Antibiotiques et facteurs de risque de résistance aux antibiotiques
Généralement, le traitement des infections urinaires par les ATB vise trois objectifs :
(i) l’infection selon une antibiothérapie adaptée (généralement par les fluoroquinolones, les
céphalosporines de 3ème génération et les aminosides).
(ii) la douleur en utilisant les antalgiques, antispasmodiques et anti-inflammatoires non
stéroïdiens.
(iii) la récidive en respectant les mesures hygiéniques et diététiques.

Synthèse bibliographique
14
I.4.1. Antibiotiques des infections urinaires
I.4.1.1. Les β-Lactamines
a. Définition
Les -lactamines constituent la famille d'antibiotiques la plus importante, aussi bien par le
nombre et la diversité des molécules utilisables que par leurs indications en thérapeutique et en
prophylaxie des infections bactériennes. Cette famille qui regroupe les pénicillines, les
céphalosporines, les carbapénèmes et les monobactames, est caractérisée par la présence constante
du cycle β-lactame associé à des cycles et des chaînes latérales variables qui expliquent les
propriétés pharmacocinétiques et le spectre d'activité des différents produits. La grande variété de
leurs modes d'administration, leur large spectre d'activité antibactérien associé à une action
bactéricide, une bonne diffusion tissulaire, une bonne tolérance et un faible nombre d'interactions
médicamenteuses expliquent leur popularité et l'importance de leurs utilisations seules ou en
associations (Cavallo et al., 2004).
b. Classification des β-Lactamines
Les β--lactamines sont classées dans quatre familles : pénicillines, céphalosporines,
monobactames et carbapénèmes (Bryskier, 1999).
Les pénicillines :
- La benzylpénicilline a été la première pénicilline utilisée. Cette β-lactamine et ses dérivés
(pénicilline du groupe G) possèdent un spectre antibactérien qui couvre les cocci à Gram positif et
négatif et les bacilles à Gram positif. L’apparition de souches de Staphylococcus aureus
productrices de pénicillinases a nécessité la synthèse de molécules stables à l’hydrolyse par cette
enzyme.
- Les isoxazolyl-pénicillines (pénicilline du groupe M) possèdent des modifications structurales qui
permettent une augmentation de la stabilité à l’hydrolyse par les pénicillinases mais qui entraînent
souvent une diminution de l’activité antibactérienne.
- Les amino-pénicillines (pénicilline du groupe A) possèdent une activité sur les bacilles à Gram
négatif. L’ampicilline et l’amoxicilline appartiennent à ce groupe. L’amoxicilline ne diffère de
l’ampicilline que par un groupement hydroxyle qui lui confère une meilleure biodisponibilité par
voie orale que l’ampicilline. La pivampicilline fait partie d’un groupe d’ester de l’ampicilline qui
libère l’ampicilline suite à l’action des estérases digestives extra- et intracellulaires. Ces prodrogues
ont été développées pour améliorer la biodisponibilité de l’ampicilline par voie orale
(Ferres, 1983).

Synthèse bibliographique
15
- Les uréido-pénicillines et les carboxy-pénicillines possèdent un spectre élargi à certains bacilles
à Gram-négatif. Elles sont actives sur Pseudomonas aeruginosa et sur certaines souches
productrices de céphalosporinases. Elles sont réservées à la médecine humaine (Bibbal, 2008).
Les céphalosporines :
Les céphalosporines diffèrent chimiquement des pénicillines par le remplacement du cycle
thiazolidine par un cycle dihydrothiazine. Elles possèdent en effet pour structure commune l'acide
7-amino céphalosporanique. Par rapport à l'acide 6-aminopénicillanique, ce noyau possède un
carbone supplémentaire mais la distance séparant la fonction carboxylique de l'amide cyclique reste
conserver. Un élément important est la possibilité de substitution en C3 par des groupements
électrocapteurs. Ceux-ci permettront une meilleure délocalisation des électrons au niveau du cycle
bêta-lactame, rendant en principe les céphalosporines plus actives vis-à-vis des trans-peptidases en
comparaison des pénicillines. Ceci ne se traduit cependant pas toujours par un avantage clinique.
Grand nombre de céphèmes orales sont administrées sous forme de prodrogues afin d’augmenter
leur résorption. Les céphalosporines sont habituellement classées en fonction de leur spectre
d'activité, leur résistance aux β-lactamases, ainsi que de leur date de commercialisation. Et on
distingue les céphalosporines de première, deuxième, troisième et quatrième génération :
- Les céphalosporines de première génération : Elles présentent un spectre antibactérien commun
avec celui des pénicillines M et A et sont surtout utilisées contre les cocci Gram positif, à
l’exception des entérocoques et de certains staphylocoques résistants. Citons notamment le céfaclor,
la céfadroxil, la céfazoline, ou la céfatrizine qui constituent un premier choix dans la prophylaxie
chirurgicale.
- Les céphalosporines de deuxième génération : comprenant la céfuroxime, le céfamandole et la
céfoxitine qui sont caractérisées par une meilleure résistance aux -lactamases et un spectre d'action
plus large (Haemophilus influenzae et certaines entérobactéries), une activité à faible concentration
ainsi qu’une bonne diffusion tissulaire. Elles sont utilisées dans un grand nombre d'infections,
notamment respiratoires, urinaires et ostéo-articulaires.
- Les céphalosporines de troisième génération : telles que le céfotaxime, la ceftriaxone, la
ceftazidime. Elles sont moins efficaces que celles de première génération sur les germes Gram
positif mais beaucoup plus que celles de deuxième génération sur les entérobactéries. Certaines de
ces molécules sont particulièrement actives contre le bacille pyocyanique (Pseudomonas
aeruginosa). Leur prescription est généralement réservée aux infections sévères dues à des bacilles
Gram négatif. De même, elles sont utilisées avec un certain succès dans le cas des infections
nosocomiales. Elles sont actives à plus faible concentration et résistent mieux aux β-lactamases.

Synthèse bibliographique
16
- Les céphalosporines de quatrième génération : avec la cefpirome et la céfépime qui pourraient
remplacer les céphalosporines de 3e génération pour le traitement des infections à germes résistants.
Elles possèdent un spectre large, leur activité est améliorée sur les germes Gram positifs et sont plus
stables face aux -lactamases. De nombreuses céphalosporines ne sont pas résorbées par voie orale
et doivent donc être administrées par voie parentérale. Comme pour les pénicillines, les
céphalosporines présentent une courte demi-vie (à l'exception de la ceftriaxone) et doivent être
administrées de façon régulière (Wise, 1990). Ceci est d'autant plus important que le principal
paramètre d'efficacité soit le temps au delà duquel la concentration sérique est maintenue supérieure
à la concentration minimale inhibitrice.
Les monobactames :
À la fin des années 1970, toutes les nouvelles bêta-lactamines identifiées étaient produites par
les Actinomycètes. Sykes et al., (1981) entreprennent alors la recherche de nouveaux antibiotiques
produits par d’autres organismes dont les monobactames. Ces derniers sont des bêta-lactamines
monocycliques inactives sur les bactéries à Gram positif et les anaérobies. Ces antibiotiques sont en
revanche très actifs sur les entérobactéries et P. aeruginosa. L’activité anti Gram-négatif de
l’aztréonam, chef de file de cette classe est globalement comparable à celle des céphalosporines de
3ème génération comme la ceftazidime. L’aztréonam présente une bonne stabilité vis-à-vis des
bêta-lactamases de spectre restreint. De plus, les monobactames constituent les seules beta-
lactamines non hydrolysées par les métallo--lactamases sécrétrices (Figueiredo, 2012).
Les carbapénèmes :
Les carbapénémases sont des β-lactamases ayants une activité hydrolytique vis à vis des
carbapénèmes. Ces enzymes appartiennent à trois classes selon la classification d’Ambler
(Nordmann et Carrer, 2010) :
- la classe A correspond principalement aux enzymes de type KPC, IMI et GES. Elles ont la
particularité de voir leur activité in vitro totalement ou partiellement inhibée par l’acide boronique
et l’acide clavulanique. Elles hydrolysent toutes les β-lactamines.
- la classe B correspond aux métallo-β-lactamases de type VIM, IMP et NDM. Ces enzymes
hydrolysent très fortement toutes les β-lactamines à l’exception de l’aztréonam. Leur activité in
vitro n’est pas affectée par les inhibiteurs suicides de β-lactamases (acide clavulanique et
tazobactam). Ce sont des métallo-enzymes qui contiennent un ion zinc dans leur site actif
expliquant l’inhibition de leur activité par l’EDTA (chélateur des cations divalents) ou l’acide
dipicolinique.

Synthèse bibliographique
17
- la classe C correspond essentiellement aux enzymes de type oxacillinases (OXA-48, OXA-163,
OXA-181). Ces enzymes hydrolysent fortement les carbapénèmes mais pas ou peu les
céphalosporines de 3ème génération. Elles sont résistantes aux inhibiteurs suicides de β-lactamases
(acide clavulanique et tazobactam). Toutefois, leur présence est souvent couplée à la présence d’une
β-lactamase à spectre étendu (BLSE), ce qui conduit à une multirésistance des souches (Boutet-
dubois et al., 2012)
c. Inhibiteurs des -lactamases :
Les inhibiteurs des β-lactamases possèdent une faible activité antibactérienne intrinsèque. En
se liant à la β-lactamase, ils permettent l’activité de la β-lactamine à laquelle ils sont associés. Il en
résulte une action synergique et une augmentation de l’activité de la β-lactamine. Actuellement,
sont disponibles l’association :
- Amoxicilline-acide clavulanique (Augmentin) (Andre et al., 1998).
- Pipéracilline-tazobactam (Tazocillin)
- Sulbactam : En plus de son effet inhibiteur irréversible sur les β-lactamases, le sulbactam a une
activité antibiotique intrinsèque sur quelques germes, mais il est toujours utilisé en association avec
les antibiotiques détruisent par les β-lactamases.
- Sulbactam+ampicilline estérifiée : Unacim (Allain, 2008).
d. Mécanisme d’action des β-lactamines
Les bêta-lactamines agissent en inhibant la synthèse de la paroi bactérienne par le biais d’une
liaison à des cibles moléculaires spécifiques appelées les protéines liants les pénicillines (PLP). Le
peptidoglycane est un polymère réticulé, fait de chaines polysaccharidiques reliées par des peptides.
Ses précurseurs sont synthétisés dans le cytoplasme et assemblés à l’extérieur de la membrane
cytoplasmique. Lorsque les bactéries sont en phase de croissance, il existe simultanément des
phénomènes de synthèse et de destruction du peptidoglycane. L’équilibre entre ces deux
phénomènes est rompu par les antibiotiques inhibant la synthèse du peptidoglycane (Nauciel,
2001).
Le blocage de la phase finale de polymérisation représente le mode d’action des β-lactamines.
Chez les bactéries à Gram négatif, les β-lactamines doivent traverser la membrane externe pour
atteindre leurs cibles. Cette membrane agit comme une barrière hydrophobe et les β-lactamines, qui
sont le plus souvent des molécules hydrophiles, vont traverser cette barrière essentiellement par la
voie des porines (Cavallo et al., 2004) Après avoir traversées la membrane externe des bactéries à
Gram négatif, les β-lactamines diffusent facilement à travers le peptidoglycane, se trouvant ensuite
dans l’espace périplasmique.

Synthèse bibliographique
18
Le β-lactamines présente une analogie structurale entre leur noyau β-lactame et le dipeptide
terminal D-alanine-D-alanine du pentapeptide constitutif du peptidoglycane. Leur reconnaissance
par les trans-peptidases et les carboxypeptidases (PLP) aboutit à la fixation du cycle β-lactame sur
le site actif de ces enzymes. Cette fixation entraîne la formation d’un complexe pénicilloyl-enzyme
covalent provoquant l’inactivation de l’enzyme, l’inhibition de la synthèse du peptidoglycane et
enfin l’arrêt de la croissance bactérienne (Livermore, 1995).
I.4.1.2. Les quinolones
a. Définition
Les quinolones sont des molécules obtenues par synthèse chimique, qui dérivent d’acides
carboxyliques hétérocycliques diversement substitués. Toutes les quinolones actuelles présentent
une structure bicyclique, avec un azote en position 1, un carboxylate en position 3 et un carbonyle
en position 4. Les fluoroquinolones, ainsi appelées car contenants un atome de fluor en position 6,
dérivent de la quinoléine (Thomas, 2001).
b. Classification des quinolones
- Quinolones de première génération (A. nalidixique)
- Quinolones de deuxième génération (ofloxacine, ciprofloxacine, lévofloxacine, …)
- Quinolones de troisième génération (trovafloxacine, gémifloxacine, moxifloxacine)
- Quinolones de quatrième génération : des fluoroquinolones (garénoxacine) (Lafaurie, 2008).
c. Mécanisme d’action des quinolones
Les quinolones sont des antibiotiques bactéricides qui bloquent la réplication de l’ADN.
Avant d’atteindre leur cible, elles traversent les diverses structures membranaires de manière
passive. Elles se concentrent dans le cytoplasme où elles se lient à une topo-isomérase. Les
topoisomérases sont des enzymes qui modifient la topologie de l’ADN bactérien. Au cours du cycle
de réplication de l’ADN, le chromosome bactérien est soit surenroulé, soit relâché. Les
topoisomérases sont les enzymes qui permettent le passage d’un état à l’autre, en coupant l’ADN et
en le recollant. Les quinolones vont se fixer sur le complexe ADN-topo-isomérase en inhibant son
fonctionnement. La bactéricide s’expliquerait par la stabilisation des coupures d’ADN qui
déclencherait des phénomènes d’autolyse. Les deux enzymes cibles principales sont l’ADN-gyrase
et la topo-isomérase IV (Jehl et al., 2003 ; Gaudy et Buxeraud, 2005 ).
I.4.1.3. Les aminosides
a. Définition :
Le premier aminoside a été découvert en 1944 par Waksman (le premier à avoir défini les
antibiotiques), a été nommé streptomycine et a été utilisé contre la tuberculose. Cette molécule est
isolée de souches de Streptomyces griseus. Les aminosides sont principalement produits par

Synthèse bibliographique
19
Streptomyces (molécule se terminant par -myxine) ou Micromonospora (molécule se terminant par
-micine) qui sont tous les deux des actinomycètes (bactéries). Il faut préciser que quelques
aminosides, découverts dans les années 70, sont hémi-synthétiques (amikacine et netilmicine). Ils
possèdent un spectre d’activité antibactérien large justifiant que certaines spécialités soient
réservées à l’hôpital pour combattre les infections bactériennes sévères. Les principaux
inconvénients de cette classe d’antibiotiques sont leur ototoxicité et leur néphrotoxicité (Dorosz
et al., 2011).
b. Classification des aminosides:
Ils sont divisés en trois classes :
1 - Les déoxystreptamines bisusbstituées 4-5 qui comprennent: Néomycine B ou C, Paromomycine,
Lividomycine A ou B, Ribostamycine, Framycétine.
2 - Les déoxystreptamines bisubstituées 4-6 qui comprennent: Kanamycines A, B, C et dérivés,
Amikacine, Tobramycine, Dibékacine, Gentamicine, Sisomycine, Nétilmicine.
3 - Les autres : Streptomycine, Streptidine, Spectinomycine (Ezaitouni et al., 1999).
c. Mécanisme d’action des aminosides
L’activité anti-bactérienne des aminosides se décompose en plusieurs phases : une
accumulation intracellulaire (en trois phases) précède la fixation sur la cible (Casin et Collatz,
1997).
La première étape de la pénétration intracellulaire se fait par diffusion passive à travers du
peptidoglycane. Elle est rapide et non spécifique.
Les deux étapes suivantes (Phase énergie dépendante « EDP I et II» sont des transports actifs à
travers la membrane cytoplasmique. Ils nécessitent un apport d’énergie, et donc la présence d’ATP
et d’une chaîne respiratoire. EDP I permet une lente accumulation dans le cytoplasme. La fixation
progressive de la molécule sur sa cible entraîne une accélération du transfert de l’antibiotique au
travers de la membrane cytoplasmique, EDP II est donc une phase d’accumulation rapide. Ces deux
phases de transports actifs reposent sur l’existence d’un gradient électrique transmembranaire, qui
est modifié par certains paramètres biochimiques de l’environnement bactérien, tel le pH, la
présence d’O2…
La phase d’accumulation rapide permet la saturation des récepteurs ribosomaux de ces
molécules. Elles se fixent sur l’ARN 16S de la sous-unité 30S du ribosome bactérien, interférant
dans la synthèse protéique au niveau du site A (site accepteur de l’ARNt). Il y a donc inhibition de
la synthèse au stade d’initiation et d’élongation, avec synthèse de protéines non fonctionnelles,
voire létales.

Synthèse bibliographique
20
I.4.1.4. Les phénicoles (Coumbes, 1995 ; Drame, 1999).
a. Définition :
Ce sont des antibiotiques bactériostatiques à large spectre qui couvrent une grande variété de
germes à Gram positif et à Gram négatif.
b. Classification :
Nous distinguons dans ce groupe le chloramphénicol et le thiamphénicol.
c. Mécanisme d’action des phénicoles
Les phénicoles agissent en inhibant la synthèse protéique en se fixant au niveau de la sous
unité 50 S des ribosomes et empêchant la transpeptidation de l’ARN de transfert.
I.4.1.5. Les antibactériens de synthèse
Parmi les molécules synthétisées, nous citerons principalement les oxyquinoleines et la
notroxoline. L’association sulfamétoxazole-trimétoprime possède un large spectre permettant
d’exercer un effet bactéricide. Ainsi, le cotrimoxazole inhibe la synthèse des acides nucléiques en
agissant sur les deux enzymes principales de la voie de synthèse des bases puriques, le
sulfaméthoxazole inhibe le dihydrofolate synthétase (DHS) et le triméthoprime inhibe la
dihydrofolate réductase (DHR).
I.4.2. Schéma de traitement des infections urinaires
Les différents traitements des infections urinaires sont présentés dans les tableaux 2 et 3.
Tableau2 : Le traitement de la cystite aigüe (Vorkaufer, 2011)
1er
intention 2eme
intention
Cystie aigüe simple/
Cystite récidivante
Fosfomycine–
Trometamol
Dose Unique
Nitrofurantoine : 100mg (3/j pendant 5j)
Fluoroquinolones dose unique ou 3/j
Cystite compliquée Nitrofurantoine
100mg (3/j Pendant 7j)
Cefixime ou fluoroquinolones pendant 5j

Synthèse bibliographique
21
Tableau 3 : Le traitement des pyélonéphrites, prostatites et de la cystite compliquée de l’homme
(Vorkaufer, 2011).
Probabiliste Relais oral
(après antibiogramme)
Pyélonéphrite aigüe
Simple ou compliquée
Ceftriaxone
ou cefotaxime,
par voie injectable,
ou Fluoroquinolone
per os ou IV
(ciprofloxacine ou
levofloxacine ou
ofloxacine)
amoxicilline,
ou amoxicilline- acide
clavulanique,
ou cefixime,
ou fluoroquinolone
(ciprofloxacine, Levofloxacine,
ofloxacine),
ou sulfamethoxazole –
trimethoprime
Prostatite et Cystite
compliquée de l’homme
fluoroquinolone (ciprofloxacine
ou levofloxacine ou ofloxacine),
ou sulfamethoxazole trimethoprime.
I.4.2.1. Infection urinaire de la femme enceinte.
a. Bactériurie asymptomatique
Le traitement est à prescrire en fonction des résultats de l’antibiogramme.
b. Cystite aigüe gravidique
Traitement probabiliste : Céfixime, ou Nitrofurantoine, à réadapté aux résultats de
l’antibiogramme. Durée du traitement 5j (7j pour la nitrofurantoine).
c. Pyélonéphrite aigüe gravidique
Traitement probabiliste : céftriaxone ou céfotaxime, par voie injectable. Relais par voie orale
avec les résultats de l’antibiogramme. Durée totale de traitement : au moins 14 jours. (Vorkaufer,
2011)
I.4.2.2. Traitement d'une infection urinaire à EBLSE :
Les différents traitements sont présentés dans le tableau 4.

Synthèse bibliographique
22
Tableau 4: Le traitement de la pyélonéphrite simple (SPILF, 2014)
1er
choix 2ème
choix
3ème
choix(en l’absence
d’alternative)
Pyélonéphrite
simple
Fluoroquinolones-S
Fluoroquinolone
(ciprofloxacine,
lévofloxacine, ofloxacine)
Fluoroquinolones-S et TMP-
SMX-S TMP-SMX
Fluoroquinolones-R et
TMP-SMX-R
Amoxicilline+acide
clavulanique Si CMI < 8 mg/l
Pipéracilline+tazobactam Si
CMI < 8 mg/l
Céfotaxime Si CMI < 1 mg/l
Ceftriaxone Si CMI < 1 mg/l
Ceftazidime Si CMI < 1 mg/l
Céfépime Si CMI < 1 mg/l
Céfoxitine si
souche sensible, et
IU à
E. coli
Aminoside
(amikacine,
gentamicine,
tobramycine)
Carbapénème
1er
traitement :
Imipénème,
méropénème
Traitement de relais :
Ertapénème
I.4.3. Résistance d’E. coli aux antibiotiques chez l'adulte
E. coli est naturellement sensible à l'ensemble des pénicillines (exceptées les pénicillines G et
M), des céphalosporines, des carbapénèmes, des quinolones, des aminosides, à la fosfomycine, à la
nitrofurantoïne et au TMP-SMX (SPILF, 2014).
Il existe différents facteurs de risque de résistance aux antibiotiques dont le principal est
l'exposition antérieure aux antibiotiques. Les taux de résistance observés sont étroitement liés a la
quantité d’antibiotiques utilisée (Austin, 1999 ; Sotto, 2001 ; Hillier, 2007). L'exposition à une
famille d’antibiotiques peut sélectionner des bactéries résistantes à d'autres familles d’antibiotiques
par le biais de mécanismes croisés. Les résistances bactériennes se développent tout
particulièrement dans la flore digestive compte tenu de son abondance (109 bactéries /g de selles).
Comme les IU sont d'origine ascendante (espèces bactériennes de la flore périnéale, elle-même

Synthèse bibliographique
23
reflet de la flore digestive), cette pression de sélection a un retentissement clinique certain (Comes,
2011).
Ceci a été montré dans différents travaux rapportant une prévalence de résistance plus élevée
pour les souches isolées de patients préalablement exposés à des antibiotiques. Ainsi, De Mouy et
al., (1999) ont montré qu’une exposition aux -lactamines ou aux quinolones dans les 6 mois
précédant, une IU augmente le risque de sélection d’une souche résistante à ces antibiotiques. Pour
l'antibiothérapie probabiliste des cystites simples, pathologie bénigne dont le risque d'évolution vers
une PNA est très faible, les antibiotiques utilisables sont ceux dont le taux de résistance est inférieur
à 20% dans la population cible. Pour l'antibiothérapie probabiliste des autres IU (cystite à risque de
complication, cystite gravidique, PNA, IU masculine), un taux de résistance supérieur à 10% n'est
pas acceptable (SPILF, 2014). La fréquence des résistances acquises aux antibiotiques d’E. coli est
présentée dans le tableau 5.
Tableau 5: Répartition des résistances d’E. coli aux ATB les plus fréquemment utilisés dans
le traitement des cystites simples (Afssaps, 2009 ; Soussy et al ., 2000).
Antibiotiques Resistance (%)
Amoxicilline
Amoxicilline - acide clavulanique
Céphalosporines de 3eme
génération
Pivmecillinam
Sulfamethoxazole-trimethoprime
Quinolones de 1ere
generation
Fluoroquinolones
Fosfomycine trometamol
Nitrofurantoine
40 à 50
25 à 30
4
25
20
14
10
< 3
< 5
I.4.4. Les mécanismes de résistances bactériennes aux antibiotiques
Outre les résistances naturelles des bactéries, il existe un nombre important de mécanismes de
résistance développé par les bactéries en réponse à la pression de sélection des ATB. Ces
résistances relèvent d’un mécanisme biochimique dont le support est génétique et elles peuvent être
transmissibles à d’autres bactéries (Andremont et Pression, 2002). Les modes de résistance
connus actuellement résultants de la pression de sélection exercée par les ATB sont au nombre de
quatre :
L’inactivation enzymatique par la sécrétion d’une enzyme
L’efflux actif

Synthèse bibliographique
24
La modification de la cible
La diminution de la perméabilité (porines) à l’antibiotique
Une même bactérie peut présenter plusieurs de ces mécanismes de résistance (Bevilacqua, 2011).
I.4.4.1. Mécanismes de résistance d’E. coli
a. La résistance aux ß-lactamines
Dans le cas d’E. coli, la résistance aux β-lactamines est due à une inactivation de
l’antibiotique par l’acquisition d’enzymes. Nous avons identifié 6 phénotypes qui sont :
phénotype sensibles (P.S.) : Les souches de ce phénotype sont sensibles aux -lactamines. Il
s’agit des entérobactéries du groupe 1.
phénotype pénicillinase de bas niveau (P.B.N.) : Les souches sont résistantes à
l’amoxicilline et sensibles à l’association amoxicilline + acide clavulanique, à la céfalotine et à la
ceftriaxone.
phénotype pénicillinase haut niveau (P.H.N.) : Les souches sont résistantes à l’amoxicilline,
à l’association amoxicilline + acide clavulanique, à la céfalotine mais sont sensibles à la
ceftriaxone.
phénotype pénicillinase résistante aux inhibiteurs des bêta-lactamases pour TEM résistantes
aux inhibiteurs: Les souches sont résistantes à l’amoxicilline, l’association amoxicilline + acide
clavulanique mais sont sensibles à la céfalotine et à la ceftriaxone.
phénotype céphalosporinase inductible (C.Ind.) : Les souches de ce phénotype sont
résistantes à l’amoxicilline, à l’association amoxicilline + acide clavulanique, à la céfalotine mais
sont sensibles à la ceftriaxone et à la ticarcilline.
phénotype céphalosporinase déréprimée (C.D.) : Les souches sont résistantes à
l’amoxicilline, à l’association amoxicilline + acide clavulanique, à la ticarcilline, à la céfalotine et à
la ceftriaxone mais elles sont sensibles au mécillinam (Keita, 1999).
phénotype -lactamase à spectre élargi (B.L.S.E.) : Les β-lactamases à spectre élargi (BLSE)
constituent un vaste groupe de β-lactamases véhiculées habituellement par des plasmides. Ce type
de β-lactamases confère une résistance à l’ensemble des β-lactamines, à l’exception des
céphamycines (céfoxitine, céfotétan, latamoxef) et des carbapénèmes (imipénème, doripénème,
ertapénème, méropénème). Elles ont la propriété d’être inhibées par les inhibiteurs de bêta-
lactamases dont l’acide clavulanique, à la base même du principe de détection des BLSE. Plus de
200 BLSE ont été décrites, classées en 11 familles en fonction de leur séquence protéique. Les 4
familles majeures sont TEM, SHV, OXA et CTX-M, et sont le plus souvent d’origine plasmidique
(Géraldine, 2010).

Synthèse bibliographique
25
Caractéristiques moléculaires des BLSE:
Leur apparition s'explique par la mutation des gènes des plasmides de TEM et SHV (Bush et
al., 1995). En effet, une seule mutation sur le plasmide de TEM ou SHV peut provoquer une
modification du site actif de l'enzyme et donc une modulation de l'activité pouvant s'exprimer de
façon variable vis-à-vis des C3G (Paterson et Bonomo, 2005). Cette substitution en acide aminé
ne concerne qu'un nombre restreint de position mais certaines sont particulièrement importantes. Il
a été montré que, pour TEM, les acides aminés les plus mutés sont en position 104, 164, 238 et 240
(Perilli et al., 1997). Les acides substitués sont la lysine, la sérine, la leucine et l'histidine
(Philippon et Arlet, 2005). Cela a une importance car la mutation du gène entraînant une variation
de la structure protéique de l'enzyme synthétisé, l'affinité de l'enzyme pour la -lactamine peut en
être modifiée. Les mutations ne concernent jamais la sérine 70 du site actif (Paterson et Bonomo,
2005). Parfois, la mutation de l'acide aminé sur TEM peut entraîner une baisse de l'affinité de la
- lactamase pour l'ensemble des -lactamines et en particulier pour les inhibiteurs. Ces enzymes de
classe A ont une faible activité en entraînant une faible augmentation des CMI (Philippon et Arlet,
2005).
Par rapport aux -lactamases de type TEM, il existe relativement moins de mutants SHV avec
un nombre de mutations ponctuelles également réduit (Jacoby et Sutton, 1991). La position la plus
substituée est la 238 avec par exemple la SHV-2 sur laquelle la glycine est substituée par une serine.
Ainsi, en fonction de l'acide aminé substitué, la –lactamase développera une meilleure affinité
pour l'une des C3G (Huletsky et al., 1993).
Nouvelles BLSE:
Les BLSE ont une capacité de diffusion et d'évolution importante. Elles sont à l'origine de
nouvelles enzymes plasmidiques et sont en pleine expansion ces dernières années. Ces enzymes
sont appelés non-TEM non SHV avec un phénotype BLSE (Vahaboglu et al., 1996 ). Tout d'abord,
les CTX-M ou « céfotaximases » qui possèdent une capacité importante d'hydrolyser le céfotaxime
et l'aztréonam (Sabaté et al., 2000 ; Tzouvelekis et al., 2000). Par contre, la sensibilité à la
ceftazidime est conservée. Elles sont inhibées de façon plus importante par le tazobactam que par
l'acide clavulanique et elles ont une distribution mondiale (Bradford et al., 1999; Gniadkowski et
al., 1998). Depuis 1989, plus de 20 enzymes ont été mis en évidence chez des entérobactéries
comme Salmonella typhimurium, Escherichia coli et Kiebsiella pneumoniae (Gazouli et al., 1998 ;
Bonnet et al., 2000 ; Sabaté et al., 2000). Il a été montré une homologie à 98% entre les CTX-M
du groupe 1 (notamment CTX-M-2) et la -lactamase chromosomique de Kluyvera ascorbata (gène

Synthèse bibliographique
26
klu), bactérie de l'environnement, alors qu'il n'existe que 40% d'homologie avec les TEM et SHV
(Bonnet et al., 2000).
De point de vu classification, les premières CTX-M sont classées, selon leur séquence en
acides aminés, en quatre groupes (Philippon et Arlet, 2005):
- Groupe I : CTX-M-2, 4, 5, 6, 7, 20
- Groupe 2: CTX-M- 1, 3, 10, 11, 12, 15
- Groupe 3: CTX-M-9, 13, 14, 16, 17, 19, 21
- Groupe 4: CTX-M-8
Ces -lactamases possèdent une capacité de diffusion et d'évolution importante. En effet,
certaines continuent même de s'élargir pour devenirs actifs sur la ceftazidime, comme les CTX-M-
15 et 16 (Bonnet et al., 2000). D'autre part, les enzymes ayant un haut niveau de résistance à la
ceftazidime et parfois à l'aztréonam sont décrits chez Pseudomonas aeruginosa.. Il est possible de
les retrouver aussi chez Acinetobacter baumannii (Barrial et Scotet, 2006).
b. La résistance aux fluoroquinolones
Comme pour beaucoup d’antibiotiques, la résistance passe par deux mécanismes principaux :
réduction de la concentration intrabactérienne de l’antibiotique et altération de ses enzymes cibles.
Pour ce dernier mécanisme, il s’agit en fait de mutations dans les gènes codants pour l’ADN gyrase
et la topoisomérase IV. Ce type de mutation est stable et diffusé facilement dans le milieu bactérien.
La résistance clinique apparaît selon un mécanisme en marche d’escalier par accumulation de
plusieurs mutations génétiques. Cela peut induire une résistance de classes à l’ensemble des
quinolones, ou à seulement certaines molécules de la famille. Là encore la présence d’un groupe
méthoxy en position 8 comme pour la moxifloxacine et la gatifloxacine, est importante puisqu’elle
réduit la probabilité de sélection de résistants mutants (Dong et al., 1998 ; Zhao et al., 1997 ;
Dalhoff, 2001 ;Van bambeke, 2005 ). Quand à la réduction de la concentration d’antibiotiques
dans la bactérie, elle peut impliquer plusieurs mécanismes, dont des changements dans la membrane
bactérienne externe, comme la modification de structures protéiques appelées porines, qui assurent
une partie des échanges entre l’extérieur et l’intérieur de la bactérie. Ces systèmes d’échange
n’étant pas spécifiques des molécules transportées car l’acquisition de ce type de résistance peut
aussi entraîner une réduction de la sensibilité à d’autres classes d’antibiotiques, notamment les
carbapénèmes (Van bambeke, 2005). La réduction de la concentration de quinolones dans la
bactérie peut également être la conséquence de l’augmentation de sa sortie, par un système dit
d’efflux. Ce mécanisme peut soit rendre en lui-même la bactérie résistante à l’antibiotique s’il est
très puissant, soit permettre à la bactérie de survivre dans une concentration suboptimale
d’antibiotiques ; ce qui à terme favorise l’apparition de mutations génétiques du fait d’une pression

Synthèse bibliographique
27
majeure de sélection antibiotique jouant alors à son maximum d’efficacité. Il s’agit donc dans tous
les cas d’un mécanisme favorisant la résistance aux quinolones et sa faible spécificité tend à
permettre l’émergence de multirésistance. Enfin, les résistances aux quinolones peuvent aussi être
transmises par des plasmides. Toutefois ce phénomène bien décrit chez Klebsiella sp et E. coli
semble rare pour les autres familles bactériennes, excepté dans certaines régions géographiques
comme l’Asie orientale (Hopper, 2003).
La résistance d’E. coli aux fluoroquinolones a nettement augmenté au cours des 10 dernières
années mais elle est très variable selon le terrain: 3% à 25% aujourd'hui en France, selon la
présentation clinique et le terrain. Chez la femme entre 15 et 65 ans ; la résistance reste proche de
5%.Un traitement par quinolones dans les 6 mois précédents expose au risque de sélection de
souches moins sensibles. Il faut donc éviter les prescriptions répétées de fluoroquinolones chez un
même patient et ne pas les utiliser en traitement probabiliste chez un patient déjà traité par
quinolones dans les 6 mois précédents (quelle qu'en ait été l'indication). L'impact écologique
important des fluoroquinolones sur le microbiote intestinal rend obligatoire une stratégie d'épargne
et limite leur usage à des indications spécifiques (SPLIF, 2014).
c. Résistance aux aminosides
Résistance non enzymatique
Les mécanismes non enzymatiques peuvent être regroupés en trois grands types de
mécanisme : diminution de la perméabilité, efflux et modification de la cible des antibiotiques
(Euzéby, 2008). Les microorganismes producteurs d'aminosides ont développé un mécanisme
supplémentaire de résistance aux aminosides, leurs évitant le suicide: la méthylation post-
transcriptionnelle de l'ARNr (Galimand et al., 2003). La diminution de la perméabilité résulte
souvent d'une mutation affectant la structure des porines ou diminuant la synthèse des porines. Dans
le cas des aminosides, l'imperméabilité résulte d'un mécanisme différent. Elle est due à des
mutations modifiant le système de transport actif de ces molécules et provoquant une diminution
d'activité de tous les aminosides. C'est alors une modification de la membrane cytoplasmique
(Galimand et al., 2003).
Le système d'efflux actif est efficace grâce aux protéines transmembranaires ancrées dans la
membrane plasmique mais également dans la membrane externe des bactéries (Walsh, 2003). Ces
protéines sont spécifiques d'une classe d'antibiotique ou au contraire responsables de la multi-
résistance (Cattoir, 2004). Chez E. coli, le transporteur (Acri) appartenant à la famille "résistance-
nodulation-division"(RND) impliquée dans le transport des aminoglycosides fonctionne seule
(Rosenberg et al., 2000).

Synthèse bibliographique
28
Les modifications de la cible ribosomale relèvent de trois mécanismes : mutation de l'ARN
16S, mutation des protéines de la sous-unité 30S et méthylation de l'ARN 16S sur certaines bases
impliquées dans la fixation des aminosides au niveau du site A (Jean et Lambert, 2012). Ces
mutations de I'ARN 16S peuvent être responsables de la résistance à la streptomycine ou à la
spectinomycine, tandis que la mutation A1408G rapportée chez les mycobactéries confère la
résistance de type AGNT (Prammananan et al., 1998).
Les bactéries peuvent acquérir une résistance aux aminosides par l’altération de la cible
ribosomale. L‘existence chez la plupart des bactéries de plusieurs copies de l'opéron ARN
ribosomal constitue donc un facteur limitant à l'apparition de la résistance qui nécessite plusieurs
mutations (Jean et Lambert, 2012). Les mutations de certaines protéines de la sous-unité 30S du
ribosome S12 et S16 ont été observées comme responsables respectivement de la résistance à la
streptomycine et à la spectinomycine (Funatsu et Wittmann, 1972), mais cette résistance est
relativement rare (Carter et al., 2000).
La méthylation de l'ARN 16S est un mécanisme émergent qui confère une résistance de haut
niveau à tous les aminosides disponibles utilisés pour la thérapie systémique, à l'exception de
streptomycine. ArmA est la première méthylase responsable de ce type de résistance, porté par un
transposon situé sur un plasmide conjugatif (Galimand et al., 2003) et coder pour une enzyme qui
méthyle la position N7 de la guanine 1405 de l'ARN 16S au niveau du site A (Liou et aI., 2006).
Résistance enzymatique
L'inactivation enzymatique des aminosides est le mécanisme de résistance le plus souvent
observé. Il permet d'expliquer la résistance de plus de 95 % des souches d'entérobactéries résistantes
aux aminosides. Le déterminisme génétique est souvent plasmidique (Perichon et al., 2007). Il
existe trois classes d'enzymes différentes classées en fonction du radical qu'elles ajoutent à la
molécule d'aminoside : les N-acétyltransférases (AAC) qui neutralisent les fonctions N112, les O-
nucléotidyltransférases (ANT), et les O- phosphotransférases (APH) qui neutralisent les fonctions
OH (Doublet et al., 2004).
Ces enzymes sont codées sur des éléments génétiques mobiles, les plasmides, ce qui permet
leur échange entre les bactéries ( Schiessinger, 1988). La famille des phosphotransférases est une
large famille d'isozymes qui est responsable de l'O-phosphorylation. Il y a sept classes d'isozymes
qui catalysent la phosphorylation d'aminoglycosides: l'APH (3'), APH (2"), APH (3"), APH (4),
APH (7"), APH (6) et l'APH (9) (Poole, 2005). L'enzyme la plus étudiée et la mieux comprise de
cette famille est I'APH (3'). Cette enzyme à elle seule confère à la bactérie qui la produit une
résistance à la kanamycine, la néomycine, la paromomycine et l'amikacine (Wright et Thompson,

Synthèse bibliographique
29
1999). La famille des aminoglycosides acétyltransférases est constituée de 4 isozymes : soit les
AAC (1), AAC (3), AAC (2') et AAC (6) (Vakulenko et Mobashery, 2003).
Ces enzymes catalysent la N-acétylation. Les informations génétiques de ces enzymes sont
codées sur des transposons ou des intégrons. Elle peut aussi acétyler les aminoglycosides
possédants un hydroxyle en 6' (Ishino et al., 2004).La dernière famille d'enzymes ; les
aminoglycosides adényltransférases (ANT) catalyse le transfert catalytique d'un motif adényle entre
le complexe Mg-ATP et l'aminoglycoside .Il y a quatre isozymes ANT (6), ANT (4'), ANT (3"), et
ANT (2 "). Le gène codant pour l'ANT (2") est retrouvé dans la majorité des bactéries à Gram
négatif. Les informations génétiques pour ces enzymes sont localisées sur des transposons et des
intégrons (Vakulenko et Mobashery, 2003).
Phénotypes de résistance des entérobactéries aux -lactamines
Les phénotypes de résistance des entérobactéries aux -lactamines sont colligés dans le tableau 6.
Tableau6 : Phénotypes de résistance des entérobactéries aux β-lactamines (Jehl et al., 2003 ; Hedi,
2007)
a
phénotype conféré par une carbapénémase chez une souche de pseudomonas aeruginosa
I.4.5. Facteurs de risque connus d’infections à entérobactéries productrices BLSE
Devant l’augmentation de l’incidence des infections aux entérobactéries productrices BLSE
nosocomiales et dans la population générale, il est primordial de pouvoir identifier les patients à
risque afin de mieux cibler ceux qui nécessitent un traitement empirique de première intention par
carbapénèmes.

Synthèse bibliographique
30
1.4.5.1. Facteurs de risque d’infections nosocomiales
De nombreuses études ont été menées depuis ces 15 dernières années afin d’identifier les
facteurs de risque d’infections nosocomiales aux entérobactéries productrices BLSE.
Les plus fréquemment identifiés sont les suivants :
les traitements antibiotiques par céphalosporines et fluoroquinolones ;
les procédures invasives et la présence de matériel étranger, en particulier le sondage vésical
ou la ventilation mécanique et la présence de cathéters centraux ;
les antécédents d’hospitalisation, notamment en unités de soins intensifs ainsi que la durée
de séjour prolongé.
les facteurs liés au statut et comorbidités du patient avec l’âge avancé, la présence des
maladies chroniques (diabète, insuffisance rénale chronique, cirrhose hépatique, pathologie
pulmonaire chronique obstructive). (Rodríguez-Baño et al., 2006 ; Pena et al., 2006 ;
Lavigne et al., 2007 ; Trecarichi et al., 2012).
I.4.6. Les moyens de lutte contre E. coli BLSE
Les moyens de lutte contre E. coli BLSE recommandés par le haut conseil de santé publique
sont (Mondain, 2010) :
Surveillance épidémiologique : étudier la fréquence du portage de E. coli BLSE dans
différents types de population, évaluer l’évolution du nombre des infections, dépister les
entérobactéries BLSE en établissement de soins en cas de situation épidémique ou en cas de
facteurs de risque connus de BLSE : patients d’ Établissements d’hébergement pour personnes
âgées dépendantes, maisons de retraite ou l’unité de soins longue durée, patients ayants reçu une
antibiothérapie, antécédents de traitement par -lactamines ou fluoroquinolones, antécédents
d’hospitalisation, âge élevé, sexe féminin, existence de comorbidités, diabète, infections urinaires
récidivantes, sondage urinaire et chirurgie gynécologique.
Réduction de l’usage des antibiotiques : réduire les volumes utilisés, promouvoir le
recours à des antibiotiques autres que les C3G et les fluoroquinolones, révision des
recommandations concernant les traitements de 1ère intention.
Hygiène : à domicile et dans les collectivités autres que les établissements de santé et les
EHPAD, l’accent sera mis sur l’hygiène des mains, l’hygiène générale autour de la toilette et de
l’alimentation.
Formation :
• Sensibiliser les microbiologistes, recherche systématique de BLSE face à une entérobactérie
résistante, notification dans le compte-rendu du laboratoire.

Synthèse bibliographique
31
• Faire prendre conscience à la population de l’émergence d’un péril sanitaire.
• A l’hôpital : outre les mesures précitées, mettre en place des mesures de nature à prévenir la
transmission croisée et diminuer la dissémination d’E. coli BLSE dans l’environnement en
contrôlant les effluents.
I.4.7. Préventions
Des mesures simples de prévention peuvent être réalisées au quotidien afin de diminuer le
risque d'IU. Un traitement préventif est par ailleurs envisagé en cas d'IU récidivantes.
a. Mesures préventives non médicamenteuses
Certaines mesures non médicamenteuses sont recommandées, d'autres n'ont pas fait leurs
preuves mais sont classiquement admises :
- boire suffisamment d’eau (> 1,5 l /j),
- éviter de retenir un besoin d'uriner : avoir des mictions régulières et complètes,
- avoir une miction post-coïtale (efficacité non confirmée mais recommandée (AFSSAPS, 2008),
- réguler le transit intestinal : lutter contre la diarrhée ou la constipation,
- avoir une bonne hygiène intime quotidienne avec un savon adapté,
- éviter les douches vaginales,
- éviter les produits parfumes d'hygiène intime,
- s'essuyer de l'avant vers l’arrière,
- préférer des sous-vêtements en coton, pas trop serrés,
- éviter les spermicides et l'utilisation d'un diaphragme en cas d'IU récidivante (Bruyere et al.,
2008)
Chapitre II : Produits naturels et infections urinaires
Les infections urinaires représentent la seconde cause de consultation et de prescription
d’antibiotiques, juste derrière les infections respiratoires. L’apparition depuis quelques années de
résistances aux antibiotiques, augmentant la fréquence des rechutes et diminuant leur efficacité, a
rendu plus urgente la nécessité de trouver des alternatives naturelles. Le D-mannose, la bromélaïne,
les extraits de canneberge, d’hibiscus, de pissenlit et d’orthosiphon agissent de façon synergique
contre les micro-organismes responsables des infections urinaires (Ofek et al., 1982).
II.1. Les plantes aromatiques-médicinales
Les plantes aromatiques-médicinales (PAM) sont des drogues végétales dont au moins une
partie possède des propriétés médicamenteuses (Farnsworth et al., 1986). Environ 35 000 espèces
de plantes sont employées par le monde à des fins médicinales, ce qui constitue le plus large
éventail de biodiversité utilisé par les êtres humains. Les plantes médicinales continuent de

Synthèse bibliographique
32
répondre à un besoin important malgré l'influence croissante du système sanitaire moderne (Elqaj
et al., 2007)
II.2. Généralités sur les huiles essentielles
II.2.1. Définition des huiles essentielles
Les huiles essentielles (HE) sont des mélanges complexes de substances organiques
aromatiques liquides qu'on trouve naturellement dans diverses parties des végétaux. Elles sont très
concentrées, volatiles, non huileuses et sensibles à la décomposition sous l'effet de la chaleur.
Actuellement, leur utilisation en parfumerie et en alimentation est considérable ; c'est pour quoi
certains organismes de normalisation AFNOR NF et ISO ont donné une définition beaucoup plus
précise des huiles essentielles : l'huile essentielle est un produit obtenu par hydrodistillation et
séparée de la phase aqueuse par des procédés physiques. Cette définition est restrictive : elle exclut
d'une part les produits odorants d'origine animale, et d'autre part les essences obtenues selon d'autre
procédés d'extraction (Hammoudi, 2008).
II.2.2. Répartition
Les huiles essentielles peuvent être stockées dans divers organes, fleurs (origan), feuilles
(citronnelle), écorces (cannelier), bois (bios de rose), rhizomes (acore), fruits (badiane), ou grains
(carvi) (Degryse et al., 2008).
II.3. Composition chimique des huiles essentielles
Les huiles essentielles sont constituées principalement de deux groupes de composés odorants
distincts selon la voie métabolique empruntée. Il s’agit des terpènes, prépondérants dans la plupart
des essences et des dérivés du phénylpropane, retrouvées en tant que composé majoritaire dans
quelques unes, telles que les essences d’anis, de cannelle, de girofle, etc.… Divers autres
constituants minoritaires leurs sont associés. Les composés terpéniques sont issus d’une voie
métabolique secondaire de l’acide mévalonique. Suivant le nombre entier d'unités pentacarbonés
(C5) ramifiées, dérivées du 2-méthylbutadiène (isoprène) (Ganou, 1993) (figure4).
Figure 4 : Structure de l'unité isoprénique (Crete, 1965 ; Robles et al., 1995)

Synthèse bibliographique
33
On distingue plusieurs familles (tableau 7)
Tableau 7: Nomenclature des terpènes (Marriott et al., 2001)
Nombre d’atomes Unité d'isoprène Nomenclature
10 2 Monoterpènes (C10)
15 3 Sesquiterpènes (C15)
20 4 Diterpènes (C20)
25 5 Sesterpènes (C25)
30 6 Triterpènes (C30)
40 8 Tétraterpènes (C40)
II.4. Méthodes utilisées pour l’extraction des huiles essentielles
Le procédé le plus ancien consiste à briser les cellules productrices des huiles avec des pierres
ou instruments en bois à la température ambiante pour libérer leurs contenus (Baster et
Buchbauer, 2009). Les méthodes utilisées actuellement sont l’hydrodistillation, la distillation par
entraînement à la vapeur, l’extraction par les solvants volatils, l’extraction au CO2 supercritique et
l’'extraction par micro-onde.
II.4.1. Hydrodistillation
L’hydrodistillation est la technique de référence dans l’extraction des composés volatiles
d’une plante dans le domaine de la recherche. Cependant une verrerie adaptée a été mise en place
permettant à la fois la circulation en circuit quasi-fermé de l’eau sous forme aqueuse et gazeuse et la
cohobation de l’huile essentielle. Ces phénomènes ont étés rendus possibles à l’échelle du
laboratoire grâce à l’utilisation d’un appareillage de type Clevenger (Reverchon et De marco,
2006).
II.5. Activité antimicrobienne
Les huiles essentielles possèdent de nombreuses activités biologiques. En phytothérapie, elles
sont utilisées pour leurs propriétés antiseptiques contre les maladies infectieuses d'origine
bactérienne, par exemple contre les bactéries endocanalaires (Benblaid, 2015), la microflore
vaginale et les dermatophytes (Hammoudi, 2008).
L’utilisation des antibiotiques conduit dans la très grande majorité des cas à la sélection de
populations microbiennes résistantes. Cette résistance est due à des mutations chromosomiques ou à
l’acquisition de gènes de résistance portés par des éléments génétiques mobiles (plasmides, phages,
transposons, intégrons). Ces résistances ont conduit à chercher de nouveaux agents antimicrobiens
possédants une efficacité plus importante que les drogues synthétiques d’une part et bien accepté

Synthèse bibliographique
34
par l’organisme d’autre part (sans exercer des effets délétères sur la santé humaine) (Kempf et
Zeitouni, 2009).
Beaucoup de groupes de recherches ont étudié l’activité antimicrobienne des extraits de
plantes médicinales telles que fennel (Foeniculum vulgare), peppermint (Mentha piperita), thyme
(Thymus vulgaris). Ils ont trouvé que ces extraits sont actifs non seulement contre les bactéries mais
aussi contre les champignons, les levures et les virus (Jürgen et al., 2009). D’autres groupes de
chercheurs ont franchi une étape plus loin, ils ont isolé et identifié les métabolites responsables de
l’activité antimicrobienne des extraits de plantes, cette étape constitue une plateforme pour
plusieurs implications incluant l’industrie pharmaceutique, la médecine alternative et la thérapie
naturelle (Huang et al., 2008).
II.5.1. Mécanismes de l’action antimicrobienne des huiles essentielles
Les mécanismes par lesquels les huiles essentielles exercent leur activité antibactérienne sont
incomplètement compris, mais il y a un certain nombre de mécanismes proposés (Holley et Patel,
2005). L’action des huiles essentielles sur le développement des micro-organismes peut être
expliquée par l’altération de la perméabilité membranaire des germes en perturbant les systèmes de
transport ionique, le transport des électrons et la production d’énergie (Sikkema et al., 1995).
Smith-Palmer et al., (2001) ont montré que les bactéries à Gram positif sont plus sensibles à l’effet
des huiles essentielles que les bactéries à Gram négatif qui se caractérisent par une membrane
externe imperméable. Selon Cristiani, cette imperméabilité est due à la richesse de cette membrane
en lipo-polysaccharides la rendant plus hydrophile, ce qui empêche les terpènes hydrophobes d’y
adhérer (Cristiani et al., 2007).
D’autres études ont été effectuées sur la relation entre la présence de citral (mélange des
isomères néral et géraniale) dans le zeste des fruits des agrumes et l’inhibition de Penicillium
digitatum, Penicillium italicum et Geotrichum candidum qui sont les principales moisissures
responsables de la contamination des Citrus (Wuryatmo et al., 2003). Cette inhibition est due à la
présence d’un groupement carbonyle adjacent aux carbones α et β dans les aldéhydes insaturés α et
β ; néral et géraniale ; ceci polarise positivement le carbone β et l’aldéhyde peut agir en tant
qu’agent d’alkylation direct capable de lier les groupes nucléophiles cellulaires (Cosentino et al.,
1999).
Le mode d’action de certaines molécules antibactériennes a été décrit dans la littérature. Par
exemple : l’action antibactérienne de l’huile essentielle d’Origan est due à une augmentation de la
perméabilité de la membrane plasmique d’E. coli suivie d’une rupture de celle-ci, entraînant une
fuite du contenu cytoplasmique, (apparition de fuite d’ions potassium K+ des cellules
microbiennes), et donc la mort de la bactérie (figure 5).
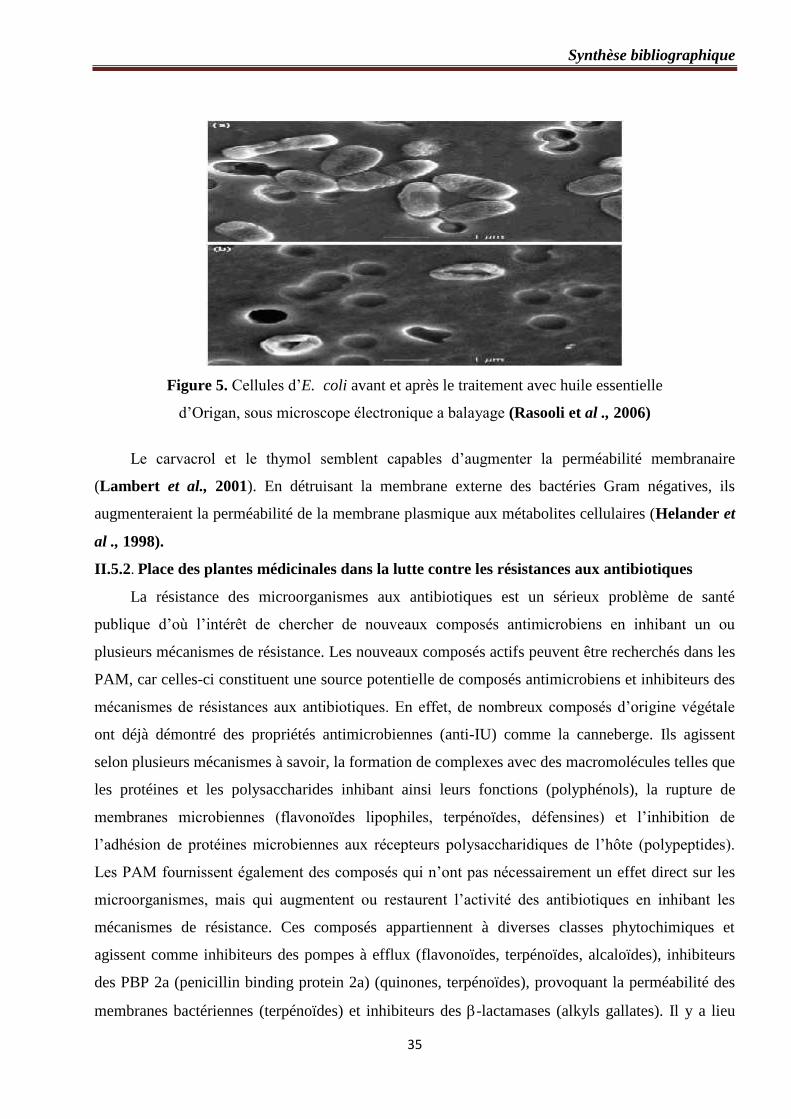
Synthèse bibliographique
35
Figure 5. Cellules d’E. coli avant et après le traitement avec huile essentielle
d’Origan, sous microscope électronique a balayage (Rasooli et al ., 2006)
Le carvacrol et le thymol semblent capables d’augmenter la perméabilité membranaire
(Lambert et al., 2001). En détruisant la membrane externe des bactéries Gram négatives, ils
augmenteraient la perméabilité de la membrane plasmique aux métabolites cellulaires (Helander et
al ., 1998).
II.5.2. Place des plantes médicinales dans la lutte contre les résistances aux antibiotiques
La résistance des microorganismes aux antibiotiques est un sérieux problème de santé
publique d’où l’intérêt de chercher de nouveaux composés antimicrobiens en inhibant un ou
plusieurs mécanismes de résistance. Les nouveaux composés actifs peuvent être recherchés dans les
PAM, car celles-ci constituent une source potentielle de composés antimicrobiens et inhibiteurs des
mécanismes de résistances aux antibiotiques. En effet, de nombreux composés d’origine végétale
ont déjà démontré des propriétés antimicrobiennes (anti-IU) comme la canneberge. Ils agissent
selon plusieurs mécanismes à savoir, la formation de complexes avec des macromolécules telles que
les protéines et les polysaccharides inhibant ainsi leurs fonctions (polyphénols), la rupture de
membranes microbiennes (flavonoïdes lipophiles, terpénoïdes, défensines) et l’inhibition de
l’adhésion de protéines microbiennes aux récepteurs polysaccharidiques de l’hôte (polypeptides).
Les PAM fournissent également des composés qui n’ont pas nécessairement un effet direct sur les
microorganismes, mais qui augmentent ou restaurent l’activité des antibiotiques en inhibant les
mécanismes de résistance. Ces composés appartiennent à diverses classes phytochimiques et
agissent comme inhibiteurs des pompes à efflux (flavonoïdes, terpénoïdes, alcaloïdes), inhibiteurs
des PBP 2a (penicillin binding protein 2a) (quinones, terpénoïdes), provoquant la perméabilité des
membranes bactériennes (terpénoïdes) et inhibiteurs des -lactamases (alkyls gallates). Il y a lieu

Synthèse bibliographique
36
d’ajouter quelques résultats importants publiés récemment comme la formation de biofilm par les
agents infectieux devenus inaccessibles aux antibiotiques, leur conférant ainsi une résistance. Les
métabolites secondaires de PAM peuvent inhibés les biofilms. Ainsi, les huiles essentielles de
Cymbopogon citratus et Syzygium aromaticum inhibent la formation des biofilms chez Candida
albicans (Khan et Ahmad, 2012). L’extrait aqueux d’un mélange de cinq drogues, présente une
double action, d’une part il inhibe la formation des biofilms chez Pseudomonas aeruginosa et
d’autre part il tue les microorganismes se trouvant dans la matrice (Wang et al., 2011). Une
macrolactone contenant le D-xylose, le L-rhamnose, la pescapréine, a montré un effet inhibiteur sur
les pompes à efflux des MRSA, augmentant ainsi l’action de la norfloxacine (Escobedo Martinez
et al., 2010). La liaison de deux triterpènes, l’acide oléanolique et l’acide ursolique, augmente
l’activité de -lactames vis-à-vis des souches de Staphylococcus aureus, Streptococcus epidermidis
et Listeria monocytogenes (Kurek et al., 2012).
II.5.3. Méthodes d’évaluation de l’activité antimicrobienne
Les méthodes d’évaluation de l’activité antimicrobienne des HE les plus couramment utilisées
sont la méthode de diffusion dans l’agar et la méthode de dilution d’agar et de bouillon (Malecky,
2007). Ces méthodes sont relativement rapides, peu coûteuses et n'exigent pas l'équipement de
laboratoire sophistiqué. Cependant, elles ne sont pas sans inconvénients (Wilkinson, 2006) du fait
de la faible solubilité des huiles essentielles dans l’eau, d’où la nécessité d’ajouter des solvants
(dimethylsulfoxide et l’éthanol) ou des détergents (tween 20) au milieu de culture ( Hammer et al.,
1999), et de la volatilité des huiles essentielles pendant l’incubation.
a. Méthode de diffusion dans l’agar ou aromatogramme
L’aromatogramme est basée sur une technique utilisée en bactériologie médicale, appelée
antibiogramme ou méthode par diffusion en milieu gélosé ou encore méthode des disques (Jacob et
al., 1979 ; Abdesselam, 2006 ; Razakarivony et al., 2009). Cette méthode a l’avantage d’être
d’une grande souplesse dans le choix des antibiotiques testés, de s’appliquer à un très grand nombre
d’espèces bactériennes et d’avoir été largement évaluée durant 50 ans d’utilisation mondiale
(Fauchère et Avril, 2002). Elle permet également de déterminer la sensibilité des différentes
espèces bactériennes vis-à-vis des huiles essentielles et autres agents antimicrobiens (Wilkinson,
2006). La technique consiste à utiliser des disques de papier imprégnés des différentes substances à
tester, puis déposés à la surface d’une gélose uniformément ensemencée avec une suspension de la
bactérie à étudier. Après incubation, les colonies se développent à la surface de la gélose laissant
des zones claires autour des disques appelées zones d’inhibition. Plus le diamètre de la zone
d’inhibition est grand, plus la souche est sensible à la substance testée (Conner et Beuchat, 1984).

Synthèse bibliographique
37
b. Méthode de diffusion en puits
Méthode proposée par Cooper et Woodman en 1946 et reprise par Shroeder et Messing en
1949. Elle assure une diffusion radiale de l’huile essentielle a partir d’un puits en donnant une zone
d’inhibition claire facilement mesurable. La méthode consiste à découper un trou circulaire dans la
gélose et y verser une solution de l’huile essentielle de concentration connue. L’huile essentielle
diffuse radialement en donnant une zone d’inhibition circulaire a la surface de la gélose
préalablement ensemencée avec la suspension bactérienne (Eymard, 2003)
c. Méthode de dilution
Les huiles essentielles à tester peuvent également être directement mélangées en concentration
connue au milieu de culture, qu’il soit solide ou liquide (exige la dispersion homogène par un
émulsifiant). Le milieu est ensuite inoculé à un taux déterminé de microorganismes, après
incubation, on note la présence ou l’absence de culture. La lecture peut-être visuelle ou a l’aide d’un
spectrophotomètre, le degré d’inhibition est en rapport avec la turbidité du milieu (Robert-Demuet,
1995).
d. Méthode de micro-atmosphère
Cette technique consiste à cultiver les microorganismes à tester dans les boites de Pétri sur
milieu de culture approprié. La différence réside principalement dans la position du disque
imprégné d’huile essentielle qui est déposé au centre du couvercle de la boite de Pétri, renversée
après fixation de l’huile essentielle sur le disque. Celui-ci n’est donc pas en contact avec le milieu
gélosé. L’huile s’évapore dans l’atmosphère de la boite, elle peut exercer son effet inhibiteur sur les
microorganismes testés (Pibiri, 2005).
II.5. 4. Méthodes de détermination de la Concentration Minimale Inhibitrice
Le but des méthodes de dilution en bouillon et en gélose est de déterminer la concentration la
plus faible de l’antimicrobien testé qui inhibe la croissance de la bactérie testée (la CMI,
habituellement exprimée en mg/ml ou mg/L). Cependant, la CMI ne représente pas toujours une
valeur absolue. La « véritable » CMI est un point entre la plus basse concentration qui empêche la
croissance de la bactérie et la concentration inférieure immédiate. Concernant les huiles essentielles,
les techniques de détermination de la concentration minimale inhibitrice CMI sont décrites par
plusieurs études (Bendahou et al., 2008 ; Chaker, 2010 ).
a. La dilution en bouillon
La dilution en bouillon est une technique dans laquelle une suspension bactérienne (à une
concentration optimale ou appropriée prédéterminée) est testée contre des concentrations variables
d’un agent antimicrobien dans un milieu liquide. La méthode de dilution en bouillon peut être
effectuée dans des tubes contenants un volume minimum de 2 ml (macrodilution) ou dans de plus

Synthèse bibliographique
38
petits volumes à l’aide de plaques de microtitration (microdilution). L’utilisation de ces plaques
avec un protocole documenté, y compris les précisions sur les micro-organismes de référence
appropriée peut faciliter la comparaison des résultats entre analyses.
b. La dilution en gélose
La dilution en gélose implique l’incorporation d’un agent antimicrobien dans un milieu gélosé
à des concentrations variables, en général une dilution en série de 2 en 2, suivie de
l’ensemencement d’un inoculum bactérien défini à la surface de la gélose de la boîte.
II.6. Domaines d’utilisation des HE
II.6.1. Industrie alimentaire
Les HE sont utilisés dans la conservation des denrées alimentaires, elles y sont rajoutées pour
rehausser le goût et pour empêcher le développement des contaminants alimentaires (Lachowicz et
al., 1998; Cosentino et al., 1999 ; Nielson et Rios, 2000 ; Skandamis et Nychas, 2001). Plusieurs
travaux ont montré que les HE de thym, de cannelle, d’origan et d’autres plantes aromatiques ont un
effet inhibiteur sur la croissance et la toxinogenèse de plusieurs bactéries et champignons
responsables d’infections alimentaires (Montes-belmont et Carvajal, 1998 ; Nielsen et Rios,
2000)
II.6.2. Parfumerie et cosmétologie
Un grand nombre d’HE (400 à 500) est utilisé dans l’élaboration de la majorité des parfums et
produits de toilette. Ces essences servent à préserver ces cosmétiques grâce à leur activité
antiseptique tout en leur assurant une odeur agréable (Roulier, 1992). De même, certains
constituants chimiques isolés à partir d’HE peuvent faire l’objet de transformations chimiques
donnant naissance à de nouvelles odeurs; ainsi, à partir de l’eugénol tiré de l’essence de girofle, on
aboutira à l’isogénol qui a une odeur d’œillet (Vigne, 1987).
II.6.3. Désinfection des locaux
Des essences naturelles (de citron et de lilas) à activité bactéricide, acaricide et fongistatique
entrent dans la composition d’un produit, le «paragerm», solution volatile qui s’est révélé sans
toxicité pour l’homme aux doses utilisées (Mallea et al., 1979).
Les HE étant volatiles, on peut envisager leur utilisation en tant qu’agents de préservation
pour le contrôle de l’hygiène de l’air des systèmes de climatisation, notamment dans le milieu
hospitalier, entraînant un effet bénéfique au niveau de la qualité de l’air (De billerbeck et al.,
2002). Les composés majoritaires de certaines HE (thymol et carvacrol) sont utilisés dans certains
pays comme additifs aux déchets animaux afin d’empêcher leur dégradation, qui pourrait générer de
mauvaises odeurs, réduire les agents pathogènes et conserver les nutriments des déchets jusqu’à leur
recyclage comme agents fertilisants (Varel, 2002).

Synthèse bibliographique
39
II.6.4. Aromathérapie
L’aromathérapie est une branche de la phytothérapie qui utilise les HE pour traiter un certain
nombre de maladies. Beaucoup d’ouvrages décrivent des préparations à base d’HE diverses
prescrites pour le traitement de plusieurs maladies. Cependant, ces prescriptions ne possèdent pas
de bases scientifiques rigoureuses car elles sont souvent tirées de pratiques empiriques (Valnet,
1990). Actuellement, plusieurs publications scientifiques fondées utilisent les HE ou leur composés
majoritaires pour tester leurs effets in vitro (Lima et al., 1993; Viollon et al., 1993; Viollon et
Chaumont, 1994). Ces effets sont également testés in vivo sur des modèles animaux afin de traiter
certaines infections expérimentales d’origine bactérienne provoquées par Helicobacter pylori chez
la souris par exemple (Bergonzelli et al., 2003; Ohno et al., 2003), ou de champignons
opportunistes (Surech et al., 1997; Manohar et al., 2001).
II.6. 5. Médecine dentaire
En médecine dentaire, l’exemple le plus couramment utilisé est la Listerine: solution
constituée d’HE de thymol et d’eucalyptol utilisée pour le lavage de la cavité orale et des dents et
qui possède une activité bactéricide sur les microorganismes de la salive et de la plaque dentaire
(Kato et al., 1990). Plusieurs HE ont donné des résultats cliniques très satisfaisants dans la
désinfection de la pulpe dentaire et dans le traitement et la prévention des caries (Pellecuer et al.,
1980; Sourai, 1989; Schwartz et al., 1992). Plusieurs auteurs ont rapporté les propriétés
antimicrobiennes d’un certain nombre d’HE et de leurs composés majoritaires sur les bactéries de la
cavité orale (Shapiro et al., 1994; Didry et al., 1994 ; Hammer et al., 2003). Une autre équipe de
chercheurs a décrit les propriétés antifongiques de l’HE de l’arbre à thé (Melaleuca alternifolia) sur
les infections oropharyngées chez les patients ayants présentés une résistance aux antifongiques
classiques (Vasquez et al., 2000).
II.7 Les principales voies d’utilisation des huiles essentielles (Baudoux et al., 2006 ;Willem,
2002)
II.7.1. La diffusion atmosphérique
Lors de la diffusion dans l’atmosphère, il faut prendre soin de choisir des huiles essentielles
labélisées biologiques, pures, et adaptées afin d’éviter les allergies et les contre-indications.
Certaines huiles essentielles peuvent être irritantes pour les muqueuses respiratoires. Il faut éviter de
diffuser en continu dans une pièce close et toute la nuit en présence d’une personne qui dort, mais
plutôt une quinzaine de minutes, une à trois fois par jour. Le diffuseur doit être placé de façon à ne
pas projeter directement vers le visage ou les yeux. Il faut utiliser un diffuseur qui ne chauffe pas les
huiles essentielles afin qu’elles ne s’oxydent pas. Cette voie d’administration est préférée dans

Synthèse bibliographique
40
certaines indications comme pour les huiles essentielles utilisées pour une indication respiratoire
comme l’Eucalyptus globulus, le Pin.
II.7.2. La voie interne
La voie interne peut être utilisée avec beaucoup de précaution.
La voie orale :
L’ingestion ne doit jamais se faire pure : il faut toujours les diluer avec de l’huile végétale ou
par exemple dans du miel car celles-ci ne sont pas solubles dans l’eau et laisser fondre sous la
langue. Il existe des capsules à avaler déjà prêtes avec une base d’huile végétale. Il est préférable de
ne jamais ingérer plus de trois gouttes d’une même huile essentielle plus de trois fois par jour.
La voie rectale :
La voie rectale, avec l’emploi de suppositoires est le mode d’utilisation préconisé dans les
infections broncho-pulmonaires. Cette voie permet une absorption rapide et efficace des principes
actifs des huiles essentielles en évitant le circuit digestif.
La voie gynécologique :
Elle permet une action rapide localement avec l’emploi d’ovules vaginaux fabriqués sur le
même modèle que les suppositoires en aromathérapie.
II.7.3. La voie externe
La voie cutanée :
La voie cutanée peut être utilisée dès trois ans en effleurage. Elle est beaucoup utilisée en
aromathérapie. L’huile essentielle est appliquée pure ou en mélange avec une huile végétale
préférentiellement au niveau des poignets ou du plexus solaire.
Le bain :
On peut également mettre quelques gouttes d’huile essentielle dans un bain. Là encore, la
dilution avec une huile végétale hydrosoluble est recommandée pour éviter tout risque de réaction
cutanée du fait de leur insolubilité et ainsi de leur contact avec la peau en trop grande concentration.
Les huiles essentielles sont toujours insolubles dans l’eau, pour cette raison, il faut utiliser un
dispersant en quantité quatre fois supérieure à celle de l’huile essentielle pour disperser le tout dans
le bain
II.8. Monographie des plantes sélectionnées
II.8.1. Coriandrum sativum
Coriandrum sativum est une plante odorante appartenant à la famille des Apiaceae.
Noms vernaculaires: En Français (coriandre) et en Arabe: kasbour
Nom scientifique: Coriandrum sativum L. (figure 6).

Synthèse bibliographique
41
Figure 6: Photo de Coriandrum sativum
Composition chimique: la composition chimique de l’HE de C. sativum est donnée dans
le tableau 8.
Tableau 8 : Composition chimique de l’huile essentielle de Coriandrum sativum (Craker et al.,
2004)
Constituants %
Heptanal
α-pinene
Camphene
Sabinene
β-pinene
Myrcene
ρ-cymene
Limonene
γ-terpinene
Linalool
Camphor
Borneol
Terpine-4-ol
Decanal
Geraniol
2-decenal
Geranyl-acetate
Tetradecane
2.06
3.97
0.33
0.26
0.38
0.77
2.16
1.28
4.64
77.48
2.60
0.18
0.17
0.46
0.46
0.16
1.06
0.05
Utilisations traditionnelles : Les grains de coriandre sont traditionnellement utilisés dans
le traitement symptomatique des troubles digestifs tels que : ballonnement épigastrique, lenteur de
la digestion, éructations et flatulences, la toux, le dysfonctionnement vésiculaire, la fièvre,
(Bruneton, 2009) et contre les infections urinaires (Verbeke, 2006 ; Ghourri et al., 2014).

Synthèse bibliographique
42
Usage et propriétés pharmacologie
Utilisations principales : eupeptique, spasmolytique, antimicrobien, carminatif. Le coriandre aurait
aussi des propriétés hypolipémiantes et hypoglycémiantes.
Utilisations secondaires : En usage externe ; elle entre dans la composition de pommades contre
les rhumatismes et douleurs articulaires.
2.8.1. Ziziphora hispanica:
Ziziphora hispanica est une plante odorante appartenant à la famille des lamiacées.
Noms vernaculaires: en Français: Menthe pouliot (Boullard, 2001) et en Arabe: Fliou
Nom scientifique: Ziziphora hispanica L. (Quezel et Santa, 1963) (figure 7).
Figure 7: Photo de Ziziphora hispanica
Composition chimique: La composition chimique est donnée dans le tableau 9.
Tableau 9: Composition chimique de l’HE essentielle de Ziziphora hispanica (Bekhchi et al.,
2007)
Constituants % Constituants %
α-pinene 0.52 (3Z,5E)-1,3,5-undecatriene 0.41
Cyclohexanone-3-methyl 0.24 1-Dodecene 0.36
Sabinene 0.11 α-terpineol 0.71
β-pinene 0.5 Pulegone 78.6
β-myrcene 0.3 Piperitenone 2.9
Isolimonene-trans 0.1 8-hydroxy-p-menthan-3-one 2.24
Limonene 1.4 1,3-Dimethyl pyrogallate 0.98
Iso menthone 0.11 α-copaene 0.2
1,8-Cineole 0.1 β-bourbonene 0.1
(-)-L-Isopulegol 0.1 β-caryophyllene 0.4
Camphor 0.06 γ-cadinene 0.1
Trans-isopulegone 1.09 Mint furanone-2 0.59
Menthofuran DB5-785 1.26 Caryophyllene oxide 0.11
neo-Menthol
0.05 2-Pentenoic acid, methyl ester,
(E)
0.18

Synthèse bibliographique
43
Utilisations traditionnelles
Le décocté, le macérât et la poudre végétale de la partie aérienne de cette plante, sont
préconisés pour les douleurs d'estomac. Cependant le décocté est utilisé pour la céphalée, les
douleurs abdominales, la toux, l'ictère et l’infection urinaire. Aussi, la poudre végétale de la partie
aérienne est utilisée sous forme de cataplasme pour soulager les douleurs de la céphalée (Ozturk et
Ercisli, 2007; Zargari, 1995)
Cinnamomum cassia
Cinnamomum cassia est une plante odorante appartenant à la famille des Lauraceae.
Noms vernaculaires: en Français: cannelle de Ceylan, cannellier de Ceylan et en Arabe: quirfa,
Nom scientifique: Cinnamomum cassia (Wichtl et Anton, 2003) (figure8).
Figure 8 : Photo de Cinnamomum cassia
Constituants chimiques : La composition chimique est cosignée dans le tableau 10.
Tableau 10 : Constituants chimiques de l’HE de C. cassia (Geng et al., 2011)
Les composés principaux %
L'aldéhyde trans-cinnamique
L'acétate de cinnamyle
L'eugénol
La coumarine
Le trans-2méthoxycinnamaldéhyde
70 - 90
1 - 6
≤ 0.5
1.5 - 4.0
3.0 - 15
Utilisations traditionnelles
La drogue est surtout utilisée comme aromatisant, correcteur de gout et comme épice. L’huile
essentielle est utilisée dans les dysménorrhées et comme hemostyptique (Bruneton, 2009) et contre
les infections urinaires (Verbeke, 2006 ; Ghourri et al., 2014), La drogue est également utilisée
dans les bronchites chroniques , comme stimulants de l’appétit (Bruneton, 2009 ), troubles
digestifs tels que ballonnements épigastriques (Bruneton, 2009).

Matériels et
méthodes

Matériels et méthodes
44
Partie II : Matériels et méthodes
II.1. Matériel microbien
II.1.1. Souches de références
Dans ce travail nous avons utilisé des souches de référence ATCC (American Type Culture
Collection) conservées à LAMAABE : Pseudomonas aeruginosa ATCC27853, Escherichia coli
ATCC25922, Proteus mirabilis ATCC 35659 et Klebsiella pneumoniae ATCC 70603.
II.1.2. Souches d’origine clinique
Des prélèvements cliniques (urines) ont été réalisés de mai 2011 à juin 2013 sur des patients
hospitalisés plus de 48h dans trois services à risque du Centre Hospitalo- Universitaire (CHU) de
Tlemcen à savoir les services d’urologie, de médecine interne et de maternité.
II.2. Méthodes
II.2.1. Prélèvements
Les prélèvements et l’acheminement des échantillons ont été effectués selon les
recommandations du CTIN et du CLIN (1998). Après une désinfection soigneuse du méat urétral au
savon, les urines sont recueillies dans des pots stériles après élimination du premier jet. Les
échantillons sont transportés dans une glacière et traités dans les plus brefs délais au laboratoire.
Chaque prélèvement est accompagné d’une fiche de renseignement comportant le sexe, l’âge, le
service d’hospitalisation et la durée du séjour (voir annexe 1 : Enquête sur les infections urinaires).
II.2.2. Bandelette urinaire
L’intérêt essentiel du diagnostic par les bandelettes urinaires (leucocytes et nitrites) réside dans
sa facilité de réalisation et dans sa valeur prédictive négative. Elle permet de confirmer une
infection urinaire par une activité de leucocytes et de nitrites. Pour cela, des bandelettes (Aragen)
sont trempées dans des urines fraîchement émises et dans des conditions aseptiques. La lecture doit
se faire 1 ou 2 minutes après le trempage par comparaison avec une gamme étalon de couleur
accompagnant la boite contenant les bandelettes (Champetier et al., 1998).
II.2.3. Examen bactériologique des urines
Le but de l’examen bactériologique des urines et de rechercher les entérobactéries à Gram
négatif.
a. Examens direct et macroscopique : Cet examen permet d’apprécier la leucocyturie,
l’aspect de l’urine (couleur, odeur, consistance……) et les éléments figurés de l’urine (hématies,
sédiments………) (Darbas et al., 2007).
b. Examen bactériologique : La culture quantitative des urines contribue à définir l’infection
urinaire. Dans des tubes, on a fait des dilutions de l’urine avec de l’eau physiologique stérile à 9%

Matériels et méthodes
45
pour obtenir des dilutions de 10-1
à 10-4
. Puis 1 ml de chaque dilution est ensemencé sur gélose
nutritive. La boite de Pétrie est laissée sécher pendant 30min à la température ambiante puis
incuber pendant 24h à 37 °C. Après incubation, les boites considérées sont celles contenants entre
30 et 300 colonies. Le nombre ainsi déterminé est multiplié par la dilution donnant le nombre de
bactéries par millilitre d’urine (Veron et Kazmierczak, 1971).. Urine est infecté si le nombre de
bactéries significatif d’une infection urinaire est >10 5 UFC/ml (Corroyer-simovicet Faure, 2011).
c. Isolement, purification et identification : L’isolement des entérobactéries Gram négatif a
été réalisé par repiquage successif sur bouillon nutritif et sur le milieu Mac conkey (Fluka) puis
incuber 18 à 24 h à 37°C. Les souches purifiées par repiquage successif sur GN sont ensemencées
sur de la GN inclinée puis conservée à 4°C. L’identification des souches est contrôlée après
vérification de leur pureté par :
l’étude des caractères macroscopiques ;
l’étude des caractères microscopiques (forme des colonies, mobilité, coloration de Gram)
l’étude des caractères biochimiques (API 20E Biomerieux®, France).
II.2.4. Antibiogramme
Un antibiogramme a été réalisé pour chaque souche isolée par la méthode de diffusion à partir
de disques chargés sur gélose de Muller Hinton selon les recommandations du Clinical Laboratory
Standards Institute (CLSI, 2008). Les géloses ont été ensemencées à l'aide d'un écouvillon à partir
d'une suspension bactérienne à 0.5 Mac Farland (une densité optique égale à 620 nm). Des disques
d'antibiotiques ont été déposés à la surface des géloses.
Les antibiotiques utilisés (oxoid) sont :Amoxicilline (25 µg), l'amoxicilline / acide clavulanique (30
µg), ticarcilline (75 µg), acide ticarcilline / acide clavulanique (85 µg), pipéracilline (75µg),
pipéracilline/tazobactam(85µg), céphalotine (30µg),céfotaxime (30 µg), la ceftazidime (30 µg),
céfépime (30 µg), l'imipénème (10 µg), l'aztréonam (30 µg), la céfoxitine (30 µg), la gentamicine
(15 µg), l'amikacine (30 µg) l'acide nalidixique (30µg), la ciprofloxacine (5 µg).
II.2.5. Détection des E.coli BLSE (Test de synergie)
Le test consiste à rechercher une image de synergie entre un disque d’antibiotique contenant un
inhibiteur de ß-lactamases et un disque de céphalosporine de 3ème
génération (CTX, CAZ). Cette
image est dite « bouchon de champagne » (Philippon et Arlet, 2005).
La recherche de ß-lactamases à spectre étendu se fait dans les conditions standards de
l’antibiogramme, en disposant 2 disques : amoxicilline + acide clavulanique (AMC) et une
céphalosporine de 3ème
génération (CTX) à une distance de 20 a 30mm sur gélose Mulleur Hinton.
L’incubation se fait à 37°C pendant 18h (figure 9).

Matériels et méthodes
46
Figure 9: Réalisation d’un test de synergie (El Brahmi, 2013)
La production de BLSE se traduit par l’apparition d’une image de synergie ou bouchon de
champagne entre le disque AMC et C3G.
II.2.6. Détermination de la CMI sur microplaque (CLSI, 2008)
Des microplaques à fond en U (plaque à microtitration) ont été utilisées pour la détermination
des CMI. Une plaque à 96 puits permet la détermination des CMI des souches E.coli BLSE vis-à-
vis de la céfotaxime.
II.2.6.1. Technique :
Dissoudre 2,048 mg de poudre titrée d’antibiotique dans le volume adéquat du solvant
correspondant (Léau distillé), pour obtenir une solution mère à 2048 μg/ml.
Introduire 25 μl de solution de céfotaxime de façon à avoir des concentrations intermédiaires
allant de 512 μg/ml a 0,063 μg/ml
Ajouter la suspension bactérienne standardisée : A partir d’une culture pure de bactéries en
phase exponentielle, une colonie est introduite dans 5 à 10 ml d’eau physiologique stérile
(ou de bouillon MH), de telle sorte que l’inoculum sera ajusté à une turbidité standard de 0,5
McFarland correspond à une densité optique de 0,08‒0,1 à une longueur d’onde de 625 nm.
Ceci correspond approximativement à 108
CFU/ml. Diluer la suspension d’opacité 0,5 MF
au 1/10ème. Inoculer les cupules de la microplaque avec 5 µl de suspension bactérienne
afin d’obtenir une concentration finale de 5.105 CFU/ml. Pour chaque série, réaliser un
témoin sans antibiotique.
Ajouter dans chaque cupule 5 μl de la suspension bactérienne ajustée à 108 bactérie/ml
(Larif et al., 2013)
Répartir dans les cupules 70μl de bouillon MH, la concentration d’antibiotique obtenue va
ainsi de 512 μg/ml à 0.063 μg/ml.

Matériels et méthodes
47
Recouvrir la plaque d’un couvercle et incuber à 37º C pendant 24h
Après incubation, voir l’opacité ou le dépôt au fond de la cupule.
la CMI de chaque antibiotique correspond à la 1ère cupule ‘claire’ (pas de culture par
rapport au témoin sans antibiotique).
II.2.7. Typage moléculaire d’E. coli BLSE
La caractérisation moléculaire des souches d’E. Coli BLSE responsables d’IU a été réalisé au
laboratoire de Bactériologie Moléculaire de l’Institut Pasteur à Casablanca (Maroc). Les gènes de
résistance aux β-lactamines (bla CTX-M1, bla CTX-M2, bla CTX-M9 , bla CTX-M14 , bla CTX-M15 , bla CTX 28,
blaTEM-1 et bla SHV-12), aux aminosides (armA,rmtB,rmtA et rmtC ), aux quinolones (aac6’-Ib-cr,
qnrA, qnrB, qnrS), les AmpC (MOX ,FOX ,CMY,EBC,DHA et CIT), les gènes de virulence (pap,
hly,sfa et Cnf) et le groupement phylogénétique ont été identifiés par réaction d’amplification
génique de type PCR simplex ou multiplex (Guessennd et al., 2008 ; Perichon et al., 2007 ;. Doi
et al., 2004 ; Nordmann et al., 2010, Versalovic et al., 1991 ; Farshad et Emamghorashi ,
2009) cette dernière nécessite tout d’abord une étape d’extraction d’ADN qui sera suivi d’une
caractérisation et quantification des différents types des gènes.
II.2.7.1. Extraction d’ADN bactérien :
L’extraction de l’ADN bactérien a été effectuée par la technique de choc thermique. A partir
d’une culture jeune de 18 à 24 heures à 37°C sur gélose ordinaire, quelques colonies sont mises en
suspension dans 500 µL d’eau ultra-pure (eau de biologie moléculaire, dépourvue d’ADNase et
ARNase) et portées à ébullition pendant 10 min puis transférées directement dans la glace (0°C)
pendant 5 min. Après une centrifugation de 14500 trs/min pendant 10 min, 300µL du surnageant
riche en ADN sont récupérés et conservé à -20 °C .
II.2.7.2. PCR standard :
Les réactions d'amplification des gènes codants pour les β-lactamases ont été réalisées avec des
couples d'amorces spécifiques. A chaque réaction de PCR standard nous avons utilisé un ADN
d’une souche produisant le gène à tester comme témoin positif.
La réaction PCR permet d'amplifier in vitro une région spécifique d'un acide nucléique donné
afin d'obtenir une quantité suffisante pour le détecter et l’étudier. Pour se faire une série de réactions
permettant la réplication d’une matrice d’ADN double brin est répétée en boucle. Ainsi, au cours de
la réaction PCR, les produits obtenus à la fin de chaque cycle servent de matrice pour le cycle
suivant. L’amplification est donc exponentielle.
Pour avoir une réplication d’un ADN double brin, il faut agir en trois étapes. (i) Il faut
dénaturer l’ADN pour obtenir des matrices simples brin ; (ii) borner et amorcer la réplication de la

Matériels et méthodes
48
séquence à amplifier à l’aide d’oligonucléotides amorces spécifiques ; (iii) réaliser la réaction de
polymérisation du brin complémentaire. A la fin de chaque cycle, les produits sont sous forme
d'ADN double brin.
Les gènes de résistance aux β-lactamines : Les β-lactamases de classe A d’Ambler ; bla
TEM 1, bla SHV12, bla CTX-M1, bla CTX-M2, bla CTX-M9 , bla CTX-M14 , bla CTX-M15 , bla CTX 28 ont été
identifiés par La PCR standard en utilisant le protocole décrit par Guessennd et al., (2008). Les
gènes bla TEM, bla SHV, et bla CTX-M ont été amplifiées en utilisant des amorces spécifiques (Tableau
11) à partir de l’ADN total.
Tableau 11: Amorces utilisées pour l’amplification des gènes de résistance aux β-lactamines
Gènes séquences séquences (5’--3’) Taille des fragments
(pb)
blaCTX-M 1
blaCTX-M 2
blaCTX-M 9
blaCTX-M15
blaCTX-M14
blaCTX-M28
blaTEM
blaSHV
CTX-M1(+)
CTX-M1(-)
CTX-M2(+)
CTX-M2(-)
CTX-M9(+)
CTX-M9(-)
CTX-M15(+)
CTX-M15(-)
CTX-M14(+)
CTX-M14(-)
CTX-M28(+)
CTX-M28(-)
a-216
a-217
Os-5
Os-6
GGTTAAAAAATCACTGCGTC
TTGGTGACGATTTTAGCCGC
ATGATGACTCAGAGCATTCG
TGGGTTACGATTTTCGCCGC
ATGGTGACAAAGAGAGTGCA
CCCTTCGGCGATGATTCTC
GAC GAT GTC ACT GGC TGA GC
AGC CG C CGA CGC TAA TAC A
GCT GGA GAA AAG CAG CGG AG
GTA AGC TGA CGC AAC GTC TG
GAC GAT GTC ACT GGC TGA GC
AGC CG C CGA CGC TAA TAC A
ATAAAATTCTTGAAGACGAAA
GACAGTTACCAATGCTTAATCA
CGCCGGGTTATTCTTATTTGTCGC
CGCCGGGTTATTCTTATTTGTCGC
863
865
869
499
477
499
1079
795
(+) : amorce sens, (-) : amorce antisens
Le milieu réactionnel, commun pour toute les PCR effectuées, comprenait dans un volume final de
25 μl: 0.25 μl de dNTP (dATP,dGTP,dCTP,dGTP)(10 mM ); 2,5μl tampon de réaction10 X, 0,10 μl
de Taq polymérase (1U), 0,5 μl de chaque amorce (20 pmol/ μl), 2μl d'ADN génomique , 1,25 μl
de Mgcl2 (50Mm pour blaCTX et blaTEM et 25 Mm pour blaSHV); et 17,9 μl de H2O stérile ; suivant
le programme figurant dans le Tableau 12.

Matériels et méthodes
49
Tableau 12 : Les conditions d’amplification des gènes codants pour des bêta-lactamases
Gènes Etape
initiale
Dénaturation/hybridation/élongation Nombre
de cycles
Etape finale
CTX-M 94 ºC 5min 94 ºC 1min/60 ºC1min/72 ºC 1min 30 72 ºC 7 min
SHV 94 ºC 5min 94 ºC 1min/60 ºC1min/72 ºC 1min 30 72 ºC 7 min
TEM 94 ºC 5min 94 ºC 1min/50C1min/72 ºC 1min 30 72 ºC 7 min
Les souches de référence utilisées sont : E. coli U2A1790 (CTX-M-1), E. coli U2A1799
(CTX-M-9), Salmonella sp. U2A2145 (CTX-M-2), Salmonella sp. U2A1446 (TEM-1 and SHV-12)
sont utilisées comme contrôles positifs (Guessennd et al., 2008).
Dépistage des gènes codants pour de céphalosporinase plasmidique de type Amp C :
Les gènes Ampc plasmidiques dérivent de gène chromosomique présent chez certaines
entérobactéries. La recherche des gènes AmpC (MOX, CIT, FOX, DHA, CMY et DHA) a été
effectuée par PCR multiplex en utilisant le protocole décrit par Perichon et al., (2007) Pour
amplifier ces gènes, nous avons utilisé les amorces décrites par Nordmann et al., (2010 ) qui
permettent d’amplifier les différents variant de chaque famille de gènes blaAmpC plasmidiques
(tableau 13).
Tableau 13 : Amorces utilisées pour l’amplification des gènes plasmidiques de type AmpC.
Gènes séquences séquences (5’--3’) Taille des
fragments (pb)
CIT
DHA
EBC
FOX
MOX
CMY
CIT(+)
CIT(-)
DHA(+)
DHA(-)
EBC(+)
EBC(-)
FOX(+)
FOX(-)
MOX(+)
MOX(-)
CMY(+)
CMY(-)
CGCCGGGTTATTCTTATTTGTCGC
CGCCGGGTTATTCTTATTTGTCGC
TGGCCAGAACTGACAGGCAAA
TTTCTCCTGAACGTGGCTGGC
AAC TTT CAC AGG TGT GCT GGG T
CCG TAC GCA TAC TGG CTT TGC
TCGGTAAAGCCGATGTTGCGG
CTTCCACTGCGGCTGCCAGTT
AACATGGGGTATCAGGGAGATG
CAAAGCGCGTAACCGGATTGG
GACAGCCTCTTTCTCCACA
TGGAACGAAGGCTACGTA
795
462
405
302
190
1000
(+) : amorce sens, (-) : amorce antisens

Matériels et méthodes
50
Le milieu réactionnel pour le PCR effectuée
comprenait dans un volume final de 25 μl: 0.25 μl de dNTP (dATP,dGTP,dCTP,dGTP)(10 mM ),
2,5μl tampon de réaction 10 X, 0,10 μl de Taq polymérase (5U/ μl), 0,25 μl de chaque amorce (20
pmol/ μl), 2μl d'ADN génomique 1,25 μl de Mgcl2 (25Mm); et 15,9 μl H2O stérile; suivant le
programmes figurant dans le Tableau 14.
Tableau 14 : Les conditions d’amplification des gènes codants pour AmpC
Gène Etape
initiale
Dénaturation/hybridation/élongation Nombre
de cycles
Etape
finale
AmpC 94ºC3 min 94ºC 30s, 64ºC 30s, 72ºC 1min 25 72ºC 7 min
La souche Klebsiella pneumoniae U2A 2240 (gène dha ) ,utilisé comme contrôle positif.
Les gènes de résistance aux quinolones qnrA, qnrB et qnrS : Le dépistage du gène qnr
est réalisé par la technique de PCR Multiplex sur Ecoli BLSE détectées, en utilisant les amorces
spécifiques pour les gènes qnrA, qnrB et qnrS (Tableau 15) selon les protocoles décrit par
Guessennd et al., (2008).
Tableau15 : Amorces utilisées pour l’amplification des gènes de résistance aux quinolones
Gènes Séquences Séquences (5’--3’) Taille des fragments
(pb)
qnrA
qnrB
qnrS
qnrA(+)
qnrA(-)
qnrB(+)
qnrB(-)
qnrS (+)
qnrS (-)
TGGCGAAAAAAATT(GA)ACAGAA
GAGCAACGA(TC)GCCTGGTAG
GACGTGCTAACTTGCGTGAT
AACACCTCGACTTAAGTCTGA
ATGACTGAGCATGACCTTG
AACCATGTACACGGCTGG
594
388
476
(+) : amorce sens, (-) :amorce antisens
Les souches d’Escherichia coli, E. coli U2A2118 (qnrA), E. coli U2A2119 (qnrB), E. coli
U2A2120 (qnrS), sont utilisées comme contrôles positifs. Le milieu réactionnel pour le PCR
effectuée comprenait dans un volume final de 25 μl ; 0.5 μl de dNTP (dATP,dGTP,dCTP,dGTP)(5
mM ); 2,5μl tampon de réaction 10 X, 0.1μl de Taq polymérase (5U/ μl), 0,5 μl de chaque amorce
(20 pmol/ μl), 2μl d'ADN génomique,1,25 μl de Mgcl2 (25Mm) et 15.65 μl H2O stérile. Pour
l’amplification des échantillons, 25 μl de ce mélange sont mis dans chaque puits de la plaque. La

Matériels et méthodes
51
plaque contenant le mélange réactionnel est introduite dans le thermocycleur. L’ADN a été amplifié
selon le programme suivant (Tableau 16).
Tableau 16 : Les conditions d’amplification des gènes codants pour les quinolones
Gènes Etape
initiale
Dénaturation/hybridation/élongation Nombre
de
cycles
Etape finale
qnr(A,B et S) 94 ºC 5min 94°C 1min,60°C 45s, 72°C 1 min 30 72ºC 10 min
Recherche de gène aac (6’)-Ib : Le dépistage du gène aac (6’)-Ib est réalisé par la
technique de PCR simplex sur Ecoli BLSE détectées, selon le protocole décrit par Perichon et al.,
(2007 en utilisant les amorces spécifiques pour le gènes aac (6’)-Ib (Tableau 17).
Tableau17: Amorces utilisées pour l’amplification de gène aac(6’)-Ib
Gènes séquences séquences (5’--3’) Taille des fragments
(pb)
aac(6’)-Ib
aac(6’)-Ib(+)
aac(6’)-Ib(-)
AAGTAAGTGACTGGGGTGAGC
GATGTAAGCTCCTGGGGATTCAC
variable
(+) : amorce sens, (-) : amorce antisens
La souche d’E. coli U2A1528 (aac (6’)-Ib), est utilisée comme contrôles positifs. Le milieu
réactionnel pour le PCR effectuée comprenait dans un volume final de 25 μl ; 0.25 μl de dNTP
(dATP,dGTP,dCTP,dGTP) (5 mM ); 2.5μl tampon de réaction 10 X, 0,1μl de Taq polymérase
(5U/ μl), 0,5 μl de chaque amorce (20 pmol/ μl), 2 μl d'ADN génomique , 1,25 μl de Mgcl2 (25mM)
et 17.9 μl H2O stérile. L’ADN a été amplifié selon le programme suivant (Tableau 18).
Tableau 18 : Les conditions d’amplification des gènes codants pour le gène aac(6`)-Ib
Type de
-lactamase
Etape
initiale
Dénaturation/hybridation/élongation Nombre
de
cycles
Etape finale
aac(6`)-Ib 94 ºC 5min 94°C 1min, 55°C 1min, 72°C 1 min 30 72 ºC 7 min
Les gènes de résistance aux aminosides : La recherche des gènes de résistance aux
aminosides (armA, rmtB, rmtA, rmtC), a été effectuée par PCR multiplex selon le protocole décrit
par Doi et al., (2004) en utilisant les amorces spécifiques pour les aminosides (tableau 19).

Matériels et méthodes
52
Tableau19 : Amorces utilisées pour l’amplification des gènes de résistance aux aminosides
Gènes séquences séquences (5’--3’) Taille des fragments
(pb)
armA
rmtA
rmtB
rmtC
armA(+)
armA(-)
rmtA(+)
rmtA(-)
rmtB(+)
rmtB(-)
rmtC(+)
rmtC(-)
TATGGGGGTCTTACTATTCTGCCTAT
TCTTCCATTCCCTTCTCCTTT
CTAGCGTCCATCCTTTCCTC
TTTGCTTCCATGCCCTTGCC
TCAACGATGCCCTCACCTC
GCAGGGCAAAGGTAAAATCC
GCCAAAGTACTCACAAGTGG
CTCAGATCTGACCCAACAAG
774
315
556
846
(+) : amorce sens, (-) :amorce antisens
Le milieu réactionnel pour le PCR effectuée comprenait dans un volume final de 25 μl: 0.23
μl de dNTP(dATP,dGTP,dCTP,dGTP)(2.5mM) ; 2.3μl tampon de réaction l0 X, 1.16 μl de
Mgcl2(25Mm); ,0.09 μl Taq polymérase (2U/ μl), 0,93 μl de chaque amorce(20 pmol/ μl),, 2 μl
d'ADN génomique et 11,78 μl de H2O stérile . L’ADN a été amplifié selon le programme suivant
(Tableau 20).
Tableau 20: Les conditions d’amplification des gènes codants pour les aminosides
Gènes Etape
initiale
Dénaturation/hybridation/élongation Nombre
de cycles
Etape finale
rmt A 94 ºC 5min 94°C 30 S, 55°C 1min, 72°C 1 min 30 72 ºC 7 min
rmt B 94 ºC 5min 94°C 30 S, 55°C 1min, 72°C 1 min 30 72 ºC 7 min
rmt C 94 ºC 5min 94°C 30 S, 55°C 1min, 72°C 1 min 30 72 ºC 7 min
armA 94 ºC 5min 94°C 1min,55°C 45s, 72°C 1 min 30 72 ºC 7 min
Les gènes de virulence : La recherche des gènes pili (pap), S-family adhesions (sfa),
hémolysine (hly), et cytotoxic necrotizing factor 1 (cnf1) ont été effectuées par PCR multiplex en
utilisant le protocole décrit par Farshad et Emamghorashi , (2009) (Tableau 21).

Matériels et méthodes
53
Tableau 21: Amorces utilisées pour l’amplification des gènes de virulence
Gènes séquences séquences (5’--3’) Taille des
fragments (pb)
Cnf
pap
hly
sfa
cnf (+)
cnf(-)
pap (+)
pap (-)
hly(+)
hly (-)
sfa (+)
sfa(-)
TTATATAGTCGTCAAGATCACTAA
GCTTTACAATATTGAC
GGCGTTTGCTTCCATGCCCTTGCC
ATA TCC TTT CTG CAG GGA TGC AATA
AACAAGGATAAGCACTGTTCTGGCT
ACCATATAAGCGGTCATTCCCGTCA
CGGAGGAGTAATTACAAACCTGGCA
GAGAACTGCCCGGGTGCATACTCT
634
328
1177
410
(+) : amorce sens, (-) :amorce antisens
Le milieu réactionnel pour le PCR effectuée comprenait dans un volume final de 50 μl: 1.25
μl de dNTP(dATP,dGTP,dCTP,dGTP) ; 5 μl tampon de réaction l0 X, 225 μl de Mgcl2 , 0.3 μl
Taq polymérase, 1 μl de chaque amorce, 4μl d'ADN génomique et 26.95 μl H2O.. L’ADN a été
amplifié selon le programme figurant dans le Tableau 22.
Tableau 22 : Les conditions d’amplification des gènes codants pour les gènes de virulence
Type de - -
lactamase
Etape
initiale
Dénaturation/hybridation/élongation Nombre
de
cycles
Etape
finale
Hly 94 ºC 5min 94°C 1min, 63°C 30 s, 72°C 3 min 30 72 ºC 7 min
Sfa 94 ºC 5min 94°C 1min, 63°C 30 s, 72°C 3 min 30 72 ºC 7 min
Cnf 94 ºC 5min 94°C 1min, 63°C 30 s, 72°C 3 min 30 72 ºC 7 min
Pap 94 ºC 5min 94°C 1min, 63°C 30 s, 72°C 3 min 30 72 ºC 7 min
ERIC-PCR (Enterobacterial Repetitive Intergenic Consensus)
ERIC-PCR (Enterobacterial Repetitive Intergenic Consensus) est une technique génotypique de
typage épidémiologique pour établir le degré de parenté entre les souches cliniques, si les bandes
sont identiques ou différentes. Elle utilise des séquences ERIC (tableau 23) pour l’amplification par
PCR ( Versalovic et al., 1991).

Matériels et méthodes
54
Tableau 23 : Les amorces d’ERIC-PCR utilisées
Gènes séquences séquences (5’--3’) Taille des
fragments (pb)
ERIC-PCR ERIC-2
ERIC-1
AAGTAAGTGACTGGGGTGAGCG
ATGTAAGCTCCTGGGGATTCAC Variable
(+) : amorce sens, (-) : amorce antisens
L’ADN total de chaque souche a été testé par ERIC-PCR. Pour cette réaction, l’ADN a été
amplifié à l’aide de l’amorce ERIC avec le programme suivant: 7 min à 94C° et 1 min à 94C° ;
puis 36 cycles de 1min à 52C°, 6 min à 65C°, 8 min à 35C° et à la fin 10 min à 72C°. Les
conditions de préparation de mixte pour ERIC-PCR ont été réalisées comme suivantes: eau distillée
stérile: 11, 17μl, 1 μl dNTP (50 μM )(Eurogentec) ,1.5μl de MgCl2(2.5 mM) , 2.5μl de Tampon
1X , 0.5 μl de chaque amorce ERIC (20 pmol/ μl) , 0.4 μl de Taq ADN polymérase (Promega) (1.5
U), 0.43 μl de BSA (1,7 mg /ml) 2.5 μl de DMSO (10%) et 5 μl d’extrait d’ADN brut. L’ADN a
été amplifié dans un volume final de 25 μl, dans un thermocycleur puis les produits de la PCR ont
été migrés sur un gel d’agarose à 1.5% dans une cuve pour électrophorèse pendant 1 heure. Après la
migration, le gel a été soumis sous U.V pour la révélation.
Phylogénie d’E. coli : La détermination des groupements phylogénétiques d’E. coli (A1,
A2, B1, B2, et D), a été réalisée par réaction d’amplification génique de type PCR multiplex en
utilisant la combinaison de trois ADN marqueurs chuA, yjaA et le fragment ADN d’E. coli
(TspE4.C2) (Clermont et al., 2009) (tableau24).
Tableau 24 : Les amorces utilisées pour groupement phylogénétique
Gènes séquences séquences (5’--3’) Taille des
fragments (pb)
chuA
yjaA
TspE4.C2
ChuA 1
chuA 2
yjaA 1
yjaA2
TspE4.C2.1
TspE4.C2.2
GACGAACCAACGGTCAGGAT
TGCCGCCAGTACCAAAGACA
TGAAGTGTCAGGAGACGCTG
ATGGAGAATGCGTTCCTCAAC
GAGTAATGTCGGGGCATTCA
CGCGCCAACAAAGTATTACG
288
211
152
Le milieu réactionnel pour le PCR effectuée comprenait dans un volume final de 25 μl ; 0.5
μl de dNTP (dATP,dGTP,dCTP,dGTP)(5 mM ); 2,5μl tampon de réaction 10 X, 0,075μl de Taq
polymerase (5U/ μl), 0,5 μl de chaque amorce (20 pmol/ μl), 0.5μl d'ADN génomique et 15,65 μl

Matériels et méthodes
55
H2O stérile,1,25 μl de Mgcl2 (25Mm); suivant les programmes figurants sur le. Pour l’amplification
des échantillons, 25 μl de ce mélange sont mis dans chaque puits de la plaque. La plaque contenant
le mélange réactionnel est introduite dans le thermocycleur (Tableau 25). L’ADN a été amplifié
selon le programme suivant.
Tableau 25 : Les conditions d’amplification des gènes codants pour le groupement phylogénétique
Type de -
lactamase
Etape
initiale
Dénaturation/hybridation/élongation Nombre
de cycles
Etape finale
chuA 94 ºC 5min 94°C 30S, 59°C 30S, 72°C 30S 30 72 ºC 7 min
yjaA 94 ºC 5min 94°C 30 S, 59°C 30S, 72°C 30S 30 72 ºC 7 min
TspE4.C2 94 ºC 5min 94°C 30 S, 59°C 30S, 72°C 30S 30 72 ºC 7 min
II.2.7.3. Electrophorèse sur gel d’agarose (Sambrook et Russell, 2001)
Principe : En milieu basique, les fragments d’ADN sont chargés négativement. Placés
dans un champ électrique, ils vont donc se déplacer vers l’anode, mais leurs charges respectives
étant à peu près équivalentes, c’est leur masse moléculaire qui va régler leur vitesse de déplacement
à travers les mailles du gel dans lequel ils ont été placés. Plus les fragments sont petits, plus ils vont
migrer rapidement et donc loin de leur point de départ.
Préparation du gel d’agarose :
Réaliser le gel d’agarose par dissolution à chaud de 1 g de poudre d’agarose dans 150 ml
tampon TBE 0.5X et refroidir jusqu’à une température voisine de 50°C.
Couler le gel dans un moule dont les 2 extrémités ont été préalablement fermées par du
ruban adhésif
Disposer le peigne nécessaire à la réalisation des puits dans le gel
Laisser solidifier
Enlever délicatement le peigne et le ruban adhésif
Placer le moule avec le gel dans la cuve d’électrophorèse : les puits sont placés du côté de la
cathode et la cuve est remplie de tampon TBE jusqu’au niveau supérieur du gel.
Ensemencement :
Répartir dans les tubes Eppendorfs 8 μl de chaque solution d’ADN
Ajouter 2 μl de tampon de charge
Mélanger à l’aide d’une micropipette puis transférer les 10 μl des mélanges dans les puits
du gel.

Matériels et méthodes
56
Migration : Brancher le générateur après avoir raccordé la cuve. Appliquer un voltage de
90 volts pendant 4 heures.
Révélation : Colorer le gel en l’immergeant dans une solution de TBE 0.5 X contenant du
BET à une concentration de 0,5 μg/ml pendant 30 à 45 minutes à la température ambiante.
II.3. Matériel végétal
Suite à une enquête préliminaire chez les herboristes de la ville de Tlemcen et de quelques
vendeurs ambulants (voir questionnaire sur l’utilisation des plantes anti-Infections Urinaires en
annexe 1) sur les plantes utilisées contre les infections urinaires, nous avons sélectionnés trois
espèces qui sont : Cinnamomum cassia (Kirfa), Coriandrum sativum (Kasbour) et Ziziphora
hispanica (Fliou). Ces trois espèces ont été choisies pour les raisons suivantes :
Les trois plantes sont utilisées par la population de Tlemcen pour traiter les infections
urinaires et comme complément alimentaire (alicaments)
La disponibilité du matériel végétal au marché
Les HEs sont reconnues comme substances antimicrobiennes (Furletti et al., 2011 ;
Bekhchi, 2007 ; El hadri et al., 2014).
Les trois plantes ont été achetées à l’herboristerie du marché de la ville de Tlemcen. Les
données sur les trois plantes sont résumées dans le tableau 26.
Tableau 26 : Données sur les espèces végétales retenues et leurs utilisations
Plantes Appellation
locale
Famille Utilisations Origine Parties
utilisées
Cinnamomum
cassia
qirfa Lauraceae stimulants de l’appétit,
trouble digestifs et
infection urinaire
Marché
de
Tlemcen
Ecorce
Coriandrum
sativum
kasbour Apiaceae troubles digestifs, toux,
dysfonctionnement
vésiculaire, fièvre et
infection urinaire
Grains
Ziziphora
hispanica
Fliou Lamiacées douleurs d'estomac,
céphalée, douleurs
abdominales, toux, ictère
et infection urinaire
Plante
entière

Matériels et méthodes
57
II.3.1. Extraction des huiles essentielles
L’extraction a été effectuée par hydrodistillation a l’aide d’un appareil de type Clevenger selon
la technique recommandée par la pharmacopée européenne (European-Pharmacopoeia, 2005). La
matière végétale fraiche (200‒400 g) est ajoutée a l’eau dans un ballon de 2 litre placé au dessus
d’une source de chaleur (figure 10). Le ballon est lié à une colonne qui communique avec un
réfrigérant, permettant la condensation des vapeurs d’eau chargée de gouttelettes d’HE. Après
l’extraction du maximum de l’huile essentielle, la partie huileuse flottante est récupérée puis
déshydratée sur du sulfate de magnésium (MgSO4) et conservée à 4 °C.
Figure 10 : Photo de l’appareil utilisé pour l’extraction des huiles essentielles par hydrodistillation
II.3.2. Analyse chimique des huiles essentielles
Les analyses GC et GC/SM ont été effectuées au laboratoire des Produits Naturels de Corse
(France) en utilisant un appareil Perkin Elmer GC Autosystem équipé d'un injecteur unique et de
deux détecteurs à ionisation de flamme (FID). L'appareil est doté de deux colonnes de silice fondue
capillaires (60 m x 0,22 mm de diamètre, épaisseur de film 0,25 um) avec différentes phases
stationnaires : Rtx-1 (polydiméthylsiloxane) et Rtx- Wax (polyéthylène glycol). Programme de
température : 60-230 °C à 2 °C / min puis maintenus isotherme à 230 °C (30 min). Gaz vecteur :

Matériels et méthodes
58
hélium (1 ml/min). Températures d'injecteur et du détecteur ont eu lieu à 280 °C. Injection a été
réalisée avec un rapport de division de 1:80. Volume injecté : 0,1 μl.
Les analyses CG/SM ont été effectuées à l'aide d'un détecteur Perkin Elmer TurboMass,
directement couplé à un XL Perkin Elmer Autosystem équipé de colonnes de silice fondue
capillaires (60 m×0,22 mm de diamètre, épaisseur du film 0,25 um), Rtx-1 (polydiméthylsiloxane)
et Rtx-Wax (polyéthylène glycol). Autres conditions CG étaient les mêmes que décrit ci-dessus.
CG/SM (IE) les conditions : température de la source d'ions : 150 °C ; énergie d'ionisation : 70 eV ;
spectres électroniques de masse à ionisation ont été acquises sur la gamme de masse de 35 à 350
Da. Temps de numérisation : 1 s. L’injection a été réalisée avec un rapport de division de 1:80.
L’identification des composants est fondée : (i) la comparaison de leurs indices de rétention
(RI GC) sur des colonnes apolaires et polaires, déterminée par rapport au temps de rétention d'une
série de n-alcanes avec une interpolation linéaire, avec les composées authentiques ou des données
de la littérature, et (ii) sur l'ordinateur correspondant à des bibliothèques spectrales de masses
commerciales (Adams, 1995; Köning et al., 2001; Mc lafferty et Stauffer, 1994; NIST, 1999) et
avec la comparaison des spectres avec ceux de notre bibliothèque personnelle. La quantité relative
de chaque composant a été réalisée sur la base de leurs surfaces de pics sur les deux colonnes GC
capillaires Rtx-1 et Rtx-Wax, sans correction du facteur de réponse du FID.
II.3.3. Evaluation de l’activité antibactérienne
II.3.3.1. Technique en milieu solide: La méthode de Vincent (Aromatogramme)
La méthode de diffusion par disques sur gélose (méthode de Kirby-Bauer) a été utilisée pour
Évaluer l’activité antibactérienne des huiles essentielles par la formation de zones d’inhibition
(Carson et Riley, 1995).Des disques en papier filtre à 6 mm de diamètre sont imprègnes de 5 μl
d’huile essentielle et déposés sur la surface gélosée pré-ensemencée par écouvillonnage avec de la
suspension microbienne standardisée. Les souches bactériennes sont ensemencées sur gélosé Muller
Hinton (Fluka, Inde) et incubées à 37 °C pendant 24 h (figure11).Les résultats sont lus par la
mesure des diamètres des zones d’inhibition en millimètres (mm) selon la fourchette proposée par
Ponce et al., (2003) comme suit :
6 mm ≤ Ø ≤ 8 mm : non sensible 9mm ≤ Ø ≤ 14 mm : sensible
15 mm ≤ Ø ≤ 19 mm : très sensible Ø ≥20 mm : extrêmement sensible

Matériels et méthodes
59
Figure 11 : Principe de la méthode de diffusion par disque (Ganou, 1993)
II.3.3.2. Détermination des CMI de la croissance
Les concentrations minimales inhibitrices (CMI) ont été déterminées à l’aide des plaques de
microtitration à 96 puits (Wiegand et al., 2008). Les inocula standardisés à 108 ufc/ml sont dilués
au 1/1000 par le même milieu de culture (bouillon Cœur-Cervelle) pour avoir la concentration de
105
UFC/ml. Les puits de la première rangée verticale sont remplis par 200 μl de la suspension
microbienne standardisée a 105 ufc/ml comme premier témoin positif. Les puits de la deuxième
rangée sont remplis par 200 μl d’un mélange contenant du Tween 80 dilué à 0,1 % (v/v) dans la
suspension microbienne standardisée à 105 ufc/ml afin de vérifier l’effet du Tween 80 sur les
souches testées.
Le reste des dix rangées est rempli par les différentes concentrations de chaque huile essentielle.
Pour cela, une série de dilutions au 1/2 a été préparée à partir d’une solution mère contenant 400
mg/ml d’huile essentielle et 1 % de Tween 80 dilués ensemble dans le bouillon Cœur- Cervelle
(BHIB). Les autres solutions filles contenaient déjà un mélange de Tween 80 dilue à 1 % dans le
BHIB (dans la même concentration que la solution mère), dans un volume total égale à la moitié du
volume total de la solution mère, pour garder la concentration de Tween 80 constante à 1 %. Les dix
puits dans chaque rangée sont remplis par 180 μl de suspension puis 20 μl de la concentration
correspondante allant de la plus grande à la plus faible concentration dans une gamme finale de 4 %
jusqu’à 0,0039 %. La concentration finale du Tween 80 dans les puits était 0,1 %.

Matériels et méthodes
60
II.3.3.3. Concentration minimale bactéricide (CMB)
La Concentration Minimale Bactéricide (CMB) correspond à la plus faible concentration en
huile essentielle capable de tuer plus de 99,9 % de l’inoculum bactérien initial (soit moins de 0,01
% de survivants). Elle définit l’effet bactéricide d’une huile essentielle.
La gélose nutritive coulée dans des boîtes de Pétri est ensemencée en stries par 100 μl des
contenus des puits ayant la concentration minimale inhibitrice et des puis ayant une concentration
supérieure à la CMI. La CMB est déterminée après une incubation de 24 heures à 37°C ( Ganiere et
al ., 2004 ; Andrews, 2001)
II.3.3.4. Evaluation de la formation de biofilm in vitro
Test d’adhésion sur des microplaques de titration à 96 puits : Le test d‘adhésion pour
les souches était réalisé selon Extremina et al., (2010) avec quelques modifications. Pour cela, les
microplaques de titration à 96 puits ont été utilisées.
Ensemencement des souches à partir de GN inclinée dans 5mL de bouillon Cœur-Cervelle
(BHIB). L‘incubation est effectuée à 37°C pendant 24heures
Remplir les 12 puits de la microplaque avec 100μL de BHIB stérile qui vont servir de
témoin
Puis remplir les autres puits avec 100μL d'inoculum à 108
UFC/ml.
Incuber les microplaques à 37°C pendant 24 et 48 heures
Rincer les plaques trois fois avec de l‘eau physiologique stérile (pH 7,2) Laisser sécher
pendant 15 minutes
Colorer les puits au cristal violet à 0,2% pendant 15 minutes à température ambiante puis
verser le surplus du cristal violet et rincer trois fois à (EPS)
Laisser sécher pour une nuit à l‘air libre
Solubiliser le cristal violet en ajoutant 150μL d‘acide acétique à 33% dans chaque puits et
laisser pendant 15 minutes avant de lire les Densité Optique à 630nm en utilisant un lecteur
de microplaques (Awareness Technologies Stat Fax 3200).
Lecture des résultats :
La classification des résultats obtenus présente sur la base du DO témoin. Les souches ont été
classées comme suit : DO ≤ DOt(Témoin) : non formatrice du biofilm, DOt × 2 ≤ DO ≤ DOt × 4
: Modérée, DOt × 4 ≤ DO : Fortement formatrice du biofilm (Christensen et al., 1985).

Matériels et méthodes
61
Détermination des concentrations minimales inhibitrices de biofilm (CMIB) :
Obtention de biofilm : Les concentrations inhibitrices de biofilm (CMIB) des
HEs ont été déterminées comme décrit par Nostro et al., (2007). Premièrement, la microplaque à
96 puits a été remplie en distribuant 100 µL d'inoculum à 108
UFC/ml dans chaque puits. Après
24 h d'incubation à 37°C, l’inoculum flottant a été doucement enlevé et tous les puits ont été lavés
trois fois avec de l’eau physiologique stérile. Dix concentrations de chaque extrait déjà solubilisé
dans le TW80 ont été préparées par la série de dilution 1/2 dans le bouillon Mueller-Hinton stérile.
Ensuite tous les puits ont été remplis par 70 µL de bouillon Mueller-Hinton stérile et 30 µL de
chaque concentration d’huile essentielle pour obtenir des concentrations finales de 0,23 mg/ml à
120 mg /ml pour l’huile essentielle.
Lecture des résultats : La concentration inhibitrice de biofilm (CMIB) a été
déterminée après 24 h d’incubation à 37°C comme la concentration la plus basse sans culture dans
le puit visuellement déterminée.
Détermination des concentrations minimales éradicatrices de biofilm (CMEB) : Les
concentrations bactéricides de biofilm des huiles essentielles ont été déterminées comme décrit par
Nostro et al., (2007). Premièrement, la microplaque à 96 puits a été remplie par 70 µL de
bouillon Mueller-Hinton stérile et 30 µL de chaque concentration dans chaque puits. Après 24 h
d'incubation à 37°C, l’inoculum a été doucement enlevé et tous les puits ont été lavés trois fois
avec de l’eau physiologique stérile. Ensuite tous les puits ont été remplis par 100 µL de bouillon
Mueller-Hinton stérile.
Lecture des résultats : La concentration bactéricide de biofilm a été
déterminée après 24 h d’incubation à 37°C comme la concentration la plus basse sans culture dans
le puits visuellement déterminée
II.3.4. Étude de la combinaison des huiles essentielles avec les antibiotiques
Le test de combinaison a été effectué sur une souche de référence (E. coli ATCC 25922) en
combinant les huiles essentielles avec un antibiotique qui est la céfotaxime selon la méthode de
Vitale et al., (2005) et Mahboubi et Bidgoli, (2010) ; Sur microplaques de 96 puits, on introduit
verticalement 10 μl des concentrations de l’antibiotique a des concentrations décroissantes qui
débutent de 256 μg/ml et horizontalement 10 μl des concentrations de l’huile essentielle avec des
concentrations décroissantes qui débutent de 400 μg/ml. En fin on ajoute 180 μl de l’inoculum à

Matériels et méthodes
62
105 UFC/ml dans tous les puits. L’interprétation des résultats se fait en calculant la concentration
fractionnelle inhibitrice (CFI).
CFI = CFI l’huile + CFI de la céfotaxime
CFI de l’huile =CMI de l’huile combiné avec la cefotaxime
CMI de l’huile seul
CFI de la cefotaxime = CMI de la cefotaxime combiné avec l′huile
CMI de la cefotaxime seul
La CFI est interprété comme montrant l'effet synergique si elle est ≤ 0.5, l’effet indifférent quand
elle est > 0,5-2 et comme antagonistes quand elle est >2 (Mahboubi et Ghazian Bidgoli, 2010).
II.3.5. La cinétique de destruction d’E. coli exposé aux huiles essentielles
Pour l’étude de la cinétique de destruction d’E. coli BLSE par les huiles essentielles, nous
avons choisi la souche E. coli 09 BLSE parce qu’elle possède les gènes de résistance de quinolones,
des aminosides et les gènes de virulence. Pour cela, nous avons suivi le protocole expérimental de
Tangjitjaroenkun et al., (2012) . La préparation de l’inoculum a été faite en cultivant la souche a
testé dans 5 ml de cœur cervelle pendants 18 h d’incubation, on remplit les tube par la suspension
microbienne standardisée a 105 puis on ajoute l’huile essentielle pour avoir les concentrations
suivantes : CMB, 2 CMB et 4 CMB et de Tween 80 (1 %). Après une incubation sous agitation en
continu de 15, 30, 45, 60,90, 120,150 et 180 min à température 37C °, ensuite la culture est
centrifugé pendant 10 min a 250 rpm a 25 C°. Après, le culot est récupéré ensuite il est suspendu
dans 9 ml d’eau physiologique stérile pour avoir une série de dilution 10-1
a 10-4
, 100 μL de chaque
dilution ont été étalés sur le milieu gélose nutritif pour énumérer les colonies.
N.B. Chaque essai est réalisé en triple

Résultats et
discussion

Résultats et discussion
63
Partie III : Résultats et discussion
III.1. Examens macroscopiques des urines
Les résultats de l’examen macroscopique des urines prélevées au CHU de Tlemcen sont
présentés dans le tableau 27.
Tableau 27: Répartition selon l’aspect macroscopique des urines prélevées à partir des patients
hospitalisés
Nous remarquons que sur les 441 échantillons d’urines prélevées à partir des patients hospitalisés
; 121 échantillons étaient d’aspects troubles et 320 échantillons étaient d’aspects limpides.
Nous anticipons ici les résultats bactériologiques (voir ci-dessous), les urines troubles étaient toutes
infectées alors que 11/320 urines limpides étaient aussi infectées. Au mali, Ya bi (2006) a trouvé
sur un total de 131 urines infectés d’origine hospitalière, 70,23 % étaient d’aspects limpides et
29,77 % étaient d’aspects troubles. Ce qui nous amène à dire que l’aspect macroscopique des urines
ne présume pas de l’infection.
III.2. Résultats de la bandelette urinaire
Les résultats chimiques des urines sont enregistrés dans le tableau 28.
Tableau 28: Résultat de la chimie des urines analysées d’origine hospitalière
Aspects Présence Absence Total
Leucocytes > 104/ml 132 320 441
Nitrites 132 320 441
D’après le tableau 28 ; on constate la présence de leucocytes et de nitrites dans 132 urines
analysées dont 121 troubles et 11 limpides. Legras en 1993, a trouvé que tous les urines contenants
des leucocytes ≥104/ml et des nitrites étaient infectées. Ces résultats permettent de suspecter la
présence des microorganismes responsables d’une infection urinaire. C’est pourquoi la leucocyturie
garde une place de choix dans le diagnostic de l’infection urinaire.
Aspect Effectif Pourcentage (%)
Trouble 121 27.43
Limpide 320 72.56
Total 441 100
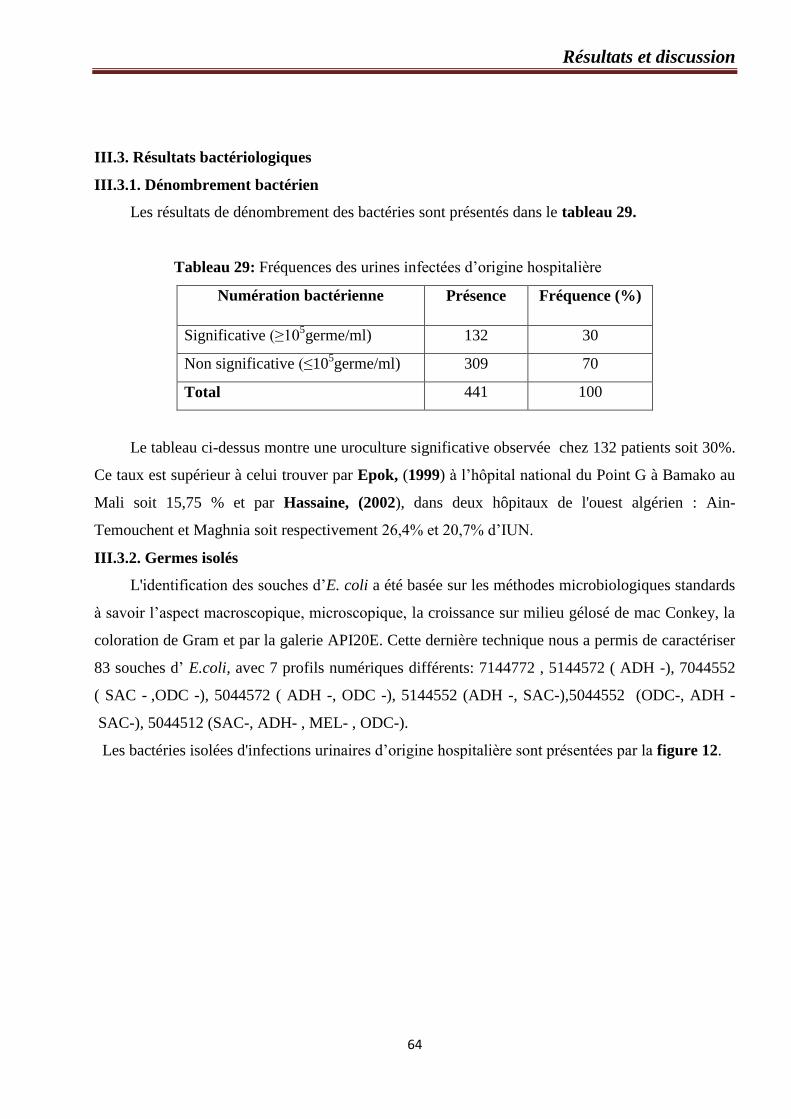
Résultats et discussion
64
III.3. Résultats bactériologiques
III.3.1. Dénombrement bactérien
Les résultats de dénombrement des bactéries sont présentés dans le tableau 29.
Tableau 29: Fréquences des urines infectées d’origine hospitalière
Numération bactérienne Présence Fréquence (%)
Significative (≥105germe/ml) 132 30
Non significative (≤105germe/ml) 309 70
Total 441 100
Le tableau ci-dessus montre une uroculture significative observée chez 132 patients soit 30%.
Ce taux est supérieur à celui trouver par Epok, (1999) à l’hôpital national du Point G à Bamako au
Mali soit 15,75 % et par Hassaine, (2002), dans deux hôpitaux de l'ouest algérien : Ain-
Temouchent et Maghnia soit respectivement 26,4% et 20,7% d’IUN.
III.3.2. Germes isolés
L'identification des souches d’E. coli a été basée sur les méthodes microbiologiques standards
à savoir l’aspect macroscopique, microscopique, la croissance sur milieu gélosé de mac Conkey, la
coloration de Gram et par la galerie API20E. Cette dernière technique nous a permis de caractériser
83 souches d’ E.coli, avec 7 profils numériques différents: 7144772 , 5144572 ( ADH -), 7044552
( SAC - ,ODC -), 5044572 ( ADH -, ODC -), 5144552 (ADH -, SAC-),5044552 (ODC-, ADH -
SAC-), 5044512 (SAC-, ADH- , MEL- , ODC-).
Les bactéries isolées d'infections urinaires d’origine hospitalière sont présentées par la figure 12.

Résultats et discussion
65
Figure 12 : Répartition des bactéries responsables d'infections urinaires d’origine hospitalière en
fonction de l'espèce
D’après la figure 12, on remarque qu’E. coli est l'espèce la plus fréquemment isolée (63℅),
suivie de Klebsiella pneumoniae (13℅) et de Proteus mirabilis (6℅). Les autres espèces sont
présentées en faible proportion comme Pseudomonas aueroginosa (3℅), Acinetobacter baumani
(3℅), Enterobacter sakazakic (3℅), Enterobacter cloacae (2℅), Klebsiella oxytoca (2℅),
Aeromonas hydrophila (1℅), Serratia marecesens (1℅), Citrobacter freundii (1℅), Providencia
stuartii (1℅), Pseudomona putida (1℅).
La fréquence d’isolement de l’espèce E.coli à l’hôpital de Tlemcen est presque équivalente à
celle de Zahlane et al., (2009) qui ont trouvé 65℅ d’E coli à l’hôpital de Marrakech (Maroc). Par
contre, la fréquence d’isolement est inferieure à celle de Valeri et ces collaborateurs qui ont trouvé
à l’hôpital de Paris 80 % d’E.coli (1998). Thabet et al., (2008) ont isolé 71℅ d’E.coli en Tunisie.
Par contre Barka et al., (2014) ont trouvé 19.16℅ d’E.coli à l’hôpital de Mahnia.
L’isolement fréquent d’E.coli à partir des prélèvements urinaires peut être expliqué par la
physiopathologie de ce type d’infection qui est en général ascendante. Il existe une forte
colonisation du périnée par les entérobactéries d'origine digestive et en particulier E.coli (Larabi et
al., 2003).
III.4. Fréquences des urines infectées en fonction du sexe :
Les résultats des urines infectées et non infectées en fonction du sexe sont données dans le
tableau 30.
Tableau 30: Fréquences des urines infectées et non infectées en fonction du sexe.
Sexe Infectées Non infectées Fréquence (%)
Femmes (F) 90 176 33.83
Hommes (H) 42 133 24

Résultats et discussion
66
D’après les résultats ci-dessus, on remarque que les urines infectées provenant du sexe
féminin (33,83℅) sont supérieurs aux urines infectées provenant du sexe masculin (24℅). Le
rapport Ratio F/H est égal à 1,4. La fréquence élevée des infections urinaires chez la femme par
rapport à celle de l’homme est en concordance avec celle rapportée par la littérature (Alexander,
1998 ; Saint et Lipsky, 1999 ; Masrar et al., 2000 ; Wagenlehner et Naber, 2001). Plus de la
moitié des femmes ont au moins un épisode de cystite bactérienne au cours de leur vie (Brumfitt et
al., 1991). L’infection urinaire chez la femme peut être interprétée par plusieurs facteurs ; liés à la
nature anatomique et physiologique de son appareil urinaire. A titre d’exemple ; la longueur de
l’urètre très réduite (4cm), les changements hormonaux et physiologiques qui parviennent au cours
des périodes de grossesse, d’accouchement et de ménopause favorisants ainsi ces incidences
d’infections (Brumfitt et al., 1991).L'homme est relativement plus protégé vue la structure
anatomique de son appareil urinaire et la distance qui sépare l'anus de son méat urinaire permet de
réduire les contaminations fécales (Lepelletier, 1997).
III.5. Fréquences des urines infectées et non infectées en fonction de l’âge
Les résultats des urines infectées et non infectées en fonction d’âge sont montrés dans le
tableau 31
Tableau 31 : Fréquences des urines infectées en fonction des tranches d’âge.
âges 0-19 20-39 40-59 ≥ 60
Infectées 2 60 20 50
Non infectées 8 117 74 110
Fréquence (%) 20 33.89 21.27 31.25
Le tableau ci-dessus montre que les patients qui ont un âge compris entre 20 et 39 ans
montrent une incidence d'infection urinaire importante (33.89%). Cette tranche d’âge coïncide avec
la phase sexuellement la plus active chez l’homme, ce qui est en concordance avec les résultats de
la littérature (Larabi et al., 2003). Aussi, l’infection est présente chez les sujets âgées plus de ≥ 60
ans (31,25℅). L’infection urinaire chez les patients âgés est aggravée par la fréquence des formes
sévères et compliquées, du fait de l’état d’immunodépression due à l’âge. Cette prévalence élevée
peut être expliquée par de nombreux facteurs plus ou moins intriqués, anatomiques, fonctionnels ou
immunologiques (Lobel et Soussy, 2007). Chez le sujet âgé la diminution du débit urinaire du fait
d’une baisse des apports hydriques, la réduction du tonus musculaire des parois des voies urinaires,
notamment celles de la vessie, entraînent après chaque miction une stase vésicale responsable de la
prolifération des germes par réduction de l’effet chasse due chez la femme à un prolapsus
(descente) de la vessie et du vagin, et chez l'homme à une affection de la prostate (adénome). Chez

Résultats et discussion
67
la femme ménopausée, la carence hormonale modifie la flore vaginale et provoque la disparition
des lactobacilles et une alcanisation du pH favorisant ainsi la colonisation des urines par les souches
uropathogènes (Boscia et al., 1986).
III.6. Répartition des urines infectées et non infectées en fonction du lieu de prélèvement
Les résultats des urines infectées et non infectées en fonction du lieu de prélèvement sont
donnés dans le tableau 32.
Tableau 32: Répartition des urines infectées et non infectées en fonction du lieu de prélèvement.
Service
Urine
infectée
Urine non
infectée
Pourcentage
(%)
Médecine interne 47 93 33.57
Maternité 44 111 28.38
Urologie 41 105 28.08
D’après ce tableau, on constate que les urines les plus infectées provenaient du service de la
médecine interne (33℅) suivi des services de maternité et d’urologie (28℅ par service). Ce résultat,
peut être expliqué par une nette dominance des patients diabétiques admis au service.
En effet, l'infection urinaire est une pathologie fréquente chez les sujets diabétiques pouvant
avoir des séquelles sur des organes vitaux comme les reins (Haraj et al., 2012).
Cette fréquence globale est variable d'une étude à l'autre en fonction des cadres d'étude et de
la population étudiée. Ainsi, Alebiosu et al., (2003) ont trouvé une fréquence de 26,6%, chez les
diabétiques de type 2 au CHU Olabisi Onabanjo, à Ibadan ; Kayima et al., (1996) ont rapporté un
taux de 11,1 % de bactériurie asymptomatique chez les diabétiques au CHU de Nairobi au Kenya
alors que Traoré et al., (1994) ont estimé cette prévalence à 19,5% à l'hôpital point "G", à Bamako.
Certains facteurs sont à l'origine de ces fréquences élevées de l'lU chez le diabétique. Ce sont
notamment la glycosurie, l'altération des fonctions des polynucléaires neutrophiles, l'anomalie
immunitaire, l'augmentation de l'adhésion de certaines souches d'E.coli aux cellules de l'urothélium
et une anomalie fonctionnelle et (ou) anatomique des voies urinaires liées à la neuropathie
autonome.
III.7. Étude de la résistance des entérobactéries isolées aux antibiotiques
Les résultats d’étude de la résistance des entérobactéries aux antibiotiques sont présentés par
la figure 13 (voir annexe 2, annexe3 et annexe 7).

Résultats et discussion
68
Figure 13: Pourcentage de résistance aux antibiotiques des entérobactéries isolées d’urines au
CHU de Tlemcen
D’après la figure 13, on note que la résistance des souches aux aminopénicillines, était élevée
(90,20%), alors qu’en présence d'acide clavulanique, cette résistance baissait pour atteindre le taux
de 62,08%. Kayima et al., (1996) avaient obtenu des résultats inferieurs avec une résistance de
66,7% pour l’amoxicilline.
Les taux de résistance des souches d’entérobactéries aux β-lactamines sont élevés. Nos
résultats montrent que l’imipenème est la seule β-lactamine active pouvant être utilisée comme un
traitement alternatif pour ces souches résistantes. Selon les anciennes études, l’imipenème est
considéré, pour longtemps, comme un antibiotique de choix pour les cliniciens (Mansour et al.,
2008). Plusieurs études ont rapporté l’efficacité de cette molécule sur les souches d’entérobactéries
productrices de BLSE. Une sensibilité de 100 % a été rapportée par Goossens et Grabein, (2005).
Dans notre étude, le taux de résistance des entérobactéries aux C3G observé était de 45,41%.
Ce taux est inferieur par apport à celui rapporté par Ben romdhane et al., (2007) soit 71% et par
Thabet et al., (2008) avec 70,8% en Tunisie. Le même constat est observé en Algérie à l’hôpital de
Dr Dorban (Annaba) 42,85% (Ramoul, 2013) et 43,56% à l’hôpital de Sidi Bel Abbes (Souna,
2010).
Pour les autres résistances associées au β-lactamines, on retrouve la ciprofloxacine qui a
montré une activité importante sur les entérobactéries (28,54%). Touati et al., (2006), ont rapporté
un taux de 0% de résistance à la ciprofloxacine. Par contre d’autres auteurs ont rapporté des taux de
résistance variables à cette molécule, soit respectivement 55% et 80% de souches BLSE sont
résistantes à la ciprofloxacine (Eisner et al., 2006 ; Goossens et Grabein, 2005).
CTX:Céfotaxine ; FOX:Cefotaxime ; CF:Céfalotine ; FEP:Péfloxacine ; PRL:Pipéraciline,
TZP:Pipéraciline+Tazobactam ; CAZ:Ceftazidine ; ATM:Aztreonam ;
TIC:Ticarciline ;TIM:Ticaciline+Acide Clavulanique ; AMX:Amoxicilline ;
AMC:Amoxicilline+Acide Clavulanique ; IMP:Imipinème ; GN;Gentamicine ;
AK:Amikacine ; CIP:Ciprofloxacine NA : Acide Nalidixique.

Résultats et discussion
69
La résistance des souches aux aminosides observée dans notre étude est faible 16,52% pour la
gentamycine et 14,14% pour l’amikacine. Des taux de 76% de résistance aux aminosides sont
rapportés par Touati et al., (2006). L’amikacine reste la molécule la plus efficace avec 85,86% de
souches sensibles comme cela a été rapporté dans la bibliographie (Gangoue-Pieboji, 2007;
Wu et al., 2007).
III.8. Étude de la résistance d’ E. coli isolée aux antibiotiques
Les résultats d’étude de la résistance d’E.coli aux antibiotiques sont présentés par la figure
14 (voir annexe 2, annexe3 et annexe 4).
Figure 14 : Pourcentage de résistance d’E. coli aux antibiotiques
D’après la figure 14, on constate que les taux de résistance les plus élevés sont observés avec
les pénicillines (amoxicilline 83,13℅, ticarciline 75,9℅, pipéraciline 61,44℅). L’association des
pénicillines avec les inhibiteurs de -lactamase a permis de récupérer l’activité de ces molécules.
En effet, on note des taux de 62,65℅ pour l’amoxiciline +acide clavulanique, 24,09℅ pour
pipéraciline+tazobactam et 39,75℅ pour ticarciline +acide clavulanique. Nous avons cependant
noté l'activité supérieure de l'association pipéracilline + tazobatam que l'association amoxiciline +
acide clavulanique. Ceci pouvant s'expliquer par le fait que l'association pipéracilline + tazobactam
est peu utilisée que l'association amoxiciline + acide clavulanique à l'Hôpital de Tlemcen. Pour les
céphalosporines de troisième génération (céfotaxime et ceftazidime) on note une résistance
importante 56,62℅.
CTX:Céfotaxine ; FOX:Cefotaxime ; CF:Céfalotine ; FEP:Péfloxacine ; PRL:Pipéraciline,
TZP:Pipéraciline+Tazobactam ; CAZ:Ceftazidine ; ATM:Aztreonam ;
TIC:Ticarciline ;TIM:Ticaciline+Acide Clavulanique ; AMX:Amoxicilline ;
AMC:Amoxicilline+Acide Clavulanique ; IMP:Imipinème ; GN;Gentamicine ;
AK:Amikacine ; CIP:Ciprofloxacine NA : Acide Nalidixique.

Résultats et discussion
70
Les niveaux des résistances bactériennes varient d’un pays à l’autre et d’une année à l’autre.
Aussi, la connaissance de la situation locale et de son évolution sont nécessaires pour le choix de
l’antibiothérapie de première intention (El bakkouri et al., 2009). Pour les autres classes
d’antibiotiques, les aminosides et les quinolones ont montré une bonne activité sur E. coli
(18℅ à 39℅).
En Tunisie, Thabet et al., (2008), ont constaté une résistance de cette espèce aux pénicillines
et aux fluoroquinolones. A El Jadida (Maroc) les résistances les plus élevées ont été observées pour
l’amoxicilline (61,2%), l’association triméthoprime sulfaméthoxazole (33,7%), l’acide nalidixique
(26,52%) et la ciprofloxacine (42%) (Tassouiket, 2014). Le tableau 33 donne une comparaison de
résistance d’E. coli aux antibiotiques testés.
Tableau 33 : Comparaison des résistances bactériennes d’E. coli
AMX AMC C3G AK GN FQ ATB (%)
Références
79,0
97,4
85,3
-
83,13
67,0
-
39,2
50,0
62,65
6,0-6,5
37,2-36,5
-
11,0
56,62
9,0
-
37,0
6,0
18,07
10,5
50,7
-
21,0
19,27
34,5-37
31,9
72,0
25,0
30,11
Tassouiket, 2014
Kashef, 2010
Kothari, 2008
Moukrad, 2012
Nos résultats
D’après le tableau 33, on remarque que la résistance aux céphalosporines de 3eme
génération des souches isolées dans notre étude est beaucoup plus élevée et que la résistance à
l’amoxiciline et aux fluroquinolones est identique par rapport aux résultats des travaux antérieurs.
III.9. Phénotypes de résistance d’E. coli aux β-lactamines
Les résultats de phénotypes de résistance aux β-lactamines sont illustrés par la figure 15
(voir annexe 4).
Figure15: Fréquences des phénotypes de résistance aux β-lactamines des souches
d’E .coli isolées d’urines infectées au CHU de Tlemcen.
Case: Céphalosporinases
Pase : Pénicillinases
S : Sensible

Résultats et discussion
71
L'analyse des phénotypes de résistance aux -lactamines d’E. coli isolée au CHU de Tlemcen
a montré une nette dominance des souches productrices de -lactamases à spectre étendu BLSE
(32.5%). Cette fréquence est très importante par rapport à celle observée à l’hôpital Mostafa Pacha
à Alger soit 7,9% (Ramdani-Bouguessa et al., 2006). En Tunisie, Thabet et al., (2008) ont isolé
1,8% d’E.coli BLSE. Selon le réseau de surveillance de la résistance des antibiotiques en Europe
2009 (EARSNet), l'incidence des E. coli productrices de BLSE varie entre 85 à 100% dans plus de
la moitié des pays déclarants y compris l'Allemagne, la France, la Belgique, le Danemark, la
Norvège, la Finlande, l'Irlande, l'Espagne, le Royaume-Uni, l'Italie, le Portugal, la Grèce et
l'Autriche. Cependant les plus faibles proportions ont été signalées par l'Islande (1,8%), en Estonie
(2,2%) et la Norvège (2,3%) (Gagliotti et al., 2011).
Cette différence de fréquence peut être expliquée par la variation géographique, la variabilité
des facteurs épidémiologiques, des politiques d’utilisation des antibiotiques et des mesures
d’hygiène hospitalière entre les différentes institutions (Mkouar et al., 2008).
Cette émergence de bactéries BLSE est liée à l'utilisation accrue des antibiotiques de type
céphalosporines de 3ème génération quelques années avant l'apparition des premières BLSE. De
nombreuses études ont été effectuées ces dernières années sur le sujet et ont souvent montré le rôle
prépondérant des antibiotiques lors de l'émergence des BLSE (Dutour et al., 2002). En effet, les
antibiotiques agissent à plusieurs niveaux : ils peuvent par exemple : transformer la flore habituelle
des patients, favoriser la colonisation par des bactéries résistantes ou encore sélectionner des
souches résistantes et faciliter leurs disséminations. En d'autres termes, les antibiotiques exercent
une pression de sélection non-négligeable (Ferron, 1989 ; Flandrois, 1997 ; Eveillard et al.,
2000 ; Gonullu et al., 2008). Cette pression de sélection est d'autant plus marquée que le nombre de
patients traités est important et que la durée de l'antibiothérapie est longue (Issa maigardie, 2006).
De plus, on peut constater que la restriction de l'utilisation des antibiotiques a permis la diminution
du nombre de BLSE (Kounta, 1999 ; Karim et al., 2001).
En générale l’acquisition des bactéries productrices de BLSE concerne des patients
immunodéprimés, suite à une hospitalisation prolongée et après exposition à des manœuvres
invasives tels que les prélèvements protégés distaux. D’autres facteurs de risque sont la
malnutrition, l’admission en réanimation, l’antécédent de prise d’antibiotique tel que l’exposition
préalable aux C3G (aussi aux fluoroquinolones et aminosides), le nombre d’antibiotiques
administrés et la durée du traitement (Ruppé, 2010). Ainsi le milieu hospitalier joue un rôle de
réservoir des souches multi-résistantes dont la dissémination est aggravée par la difficulté d’y
appliquer des mesures d’hygiène (port de gants et lavage des mains). Depuis plus de 20 ans, la
résistance des entérobactéries aux céphalosporines de 3eme génération (C3G) ne cesse de se

Résultats et discussion
72
renforcer notamment par l’acquisition de -lactamases à spectre élargi (BLSE) (Belmonte et al.,
2010).
III.10. Biotypage des souches d’E. coli BLSE selon les profils numériques en API 20E
Les résultats de biotypage des souches d’E coli BLSE selon les profils numériques en API
20E sont présentés dans le tableau 34.
Tableau 34 : Biotypage des souches d’E. coli BLSE selon les profils numériques en API 20E
Profil 5044572 5144572 5144552 5044552 7044572 7144572 5044512
Fréquence(%) 25.92 37.03 7.4 11.11 7.4 7.4 3.7
D’après les résultats de tableau 27, on note que sur les 27 souches d’E .coli BLSE isolées des
différents services hospitaliers de CHU de Tlemcen, 7 profils numériques différents ont été
enregistrés. Le profil 5144572 est dominant avec une fréquence de (37.03%), suivi du profil
numérique 5044572 (25.92%), puis du profil numérique 5044552 (11.11%). Davies et al., (1977)
ont trouvé chez les patients infectés par E coli les mêmes profils des biotypes 5144552, 7144572 et
7044572
III.11. Répartition des profils numériques d’E. coli par service d’hospitalisation
Les résultats de répartition des profils numériques par service d’hospitalisation sont présentés
dans la figure 16.
Figure 16 : Répartition des profils numériques d’E. coli BLSE par service (CHU Tlemcen)
La relation profil numérique et origine de la souche nous informe encore une fois que le
phénotype 5144572 est dominant dans les trois services et que sa fréquence varie entre 11% et
14%. Ainsi le service de médecine enregistre le taux le plus élevé 14%, suivis du service de
maternité et d’urologie avec 11% pour chacun.

Résultats et discussion
73
III.12. Répartition des souches d’E. coli BLSE en fonction des services
Les résultats de la répartition des 27 souches d’E. coli BLSE en fonction des services sont
illustrés par la figure 17 (voir annexe5).
Figure17 : Répartition des souches d’E. coli BLSE en fonction des services au CHU de Tlemcen
Les résultats ci-dessus montrent que la fréquence d’E. coli BLSE varie en fonction des
services. Le service d’urologie reste de loin le pourvoyeur essentiel de ces souches (37,1%), suivi
du service de maternité avec une fréquence de 33,3%. Le service de médecine interne marque la
plus basse fréquence 29,6%.
Le taux élevé d’E. coli BLSE enregistré en urologie peut être expliqué par le fait que les
patients admis dans le service d’urologie sont particulièrement vulnérables aux infections à des
bactéries résistantes à cause de leurs défenses immunitaires amoindries et leur exposition fréquente
aux antibiotiques (Ebertin, 1997).
D’après bertrand et al., (2003), la prévalence des phénotypes de résistance observés constitue
le plus souvent un reflet fidèle des habitudes de prescription.
Donc on peut dire que le principal facteur de risque incriminé dans cette augmentation de
résistance à l’hôpital de Tlemcen est la conséquence de la consommation croissante de C3G.
Il est nécessaire de préciser que les surfaces ainsi que les objets se trouvant dans
l’environnement du patient pouvaient être une source de contamination par les bactérie
productrices de BLSE (Tietz et al., 2004).
III.13. Répartition des souches d’E. coli BLSE en fonction sexe
Les résultats de la répartition des 27 souches d’E. coli BLSE en fonction du sexe sont notés
dans la figure 18(voir annexe5).

Résultats et discussion
74
Figure 18 : Répartition des souches d’E. coli BLSE en fonction de sexe
D’après la figure 18, on note une dominance féminine au sein de la population des personnes
infectées. La littérature nous indique qu'une femme a plus de risque de présenter une infection
urinaire que l'homme (Johanson et al., 1972 ; Garibaldi et al., 1974) à cause de la position de son
urètre. Les mêmes auteurs ont montré que le taux de l'acquisition d'une bactériémie était en rapport
avec le sexe. (Vittori et al., 2012 ; Park et al., 2012).
III.14. Répartition des souches d’E. coli BLSE en fonction d’âge
Les résultats de la répartition des 27 souches d’E. coli BLSE en fonction d’âge sont présentés
dans la figure 19(voir annexe5).
Figure 19 : Répartition des souches d’E. coli BLSE en fonction d’âge
La figure ci-dessus montre que l’infection urinaire est élevée chez les patients qui ont une
tranche d’âge comprise entre 20 et 39 ans, suivie par des patients qui ont un âge ≥ 60 ans.
L'incidence des infections urinaires nosocomiales par E. coli BLSE croît avec l’âge. Khoury
et al., (1985), ont constaté une proportion importante des infections urinaires chez les sujets âgés.

Résultats et discussion
75
Butreau et Botto, (1997), ont montré que le risque d'infection nosocomiale est multiplié par 5 chez
les sujets de plus de 65 ans. En effet, les patients âgés présentent le plus souvent des facteurs de
risque à l'infection urinaire notamment la présence de maladies métaboliques ou débilitantes
(Ben-Ami et al., 2009) .
III.15. Répartition des patients infectés par E. coli BLSE en fonction des antécédents
médicaux.
Les résultats de la répartition des patients infectés par les 27 souches d’E. coli BLSE en
fonction des antécédents médicaux sont illustrés dans la figure 20 (voir annexe 5).
Figure 20: Répartition des patients infectés par E. coli BLSE en fonction des antécédents médicaux
D’après la figure 20, on note que le pourcentage d’infection urinaire est élevé chez les
diabétiques et les femmes enceintes.
En effet, un diabète déséquilibré est associé à un risque plus élevé de survenue d'IU. Le
glucose présent dans les urines en cas de diabète favorise la croissance bactérienne. Il en est de
même pour un diabète compliqué avec une neuropathie vésicale, qui est à l'origine de reflux vésico-
rénaux et donc facteur de risque de pyélonéphrite (Rostoker et al., 1991). Les femmes enceintes
sont particulièrement à risque en raison de la pression exercée par le bébé sur le système urinaire et
à des changements hormonaux inhérents à la grossesse.
III.16. Fréquence d’E. coli BLSE durant les trois années d’étude
Les résultats de la fréquence des 27 souches d’E.coli BLSE durant les trois années d’étude
sont illustrés dans la figure 21.

Résultats et discussion
76
Figure 21: Fréquence d’E. coli BLSE responsables d’IU durant les trois années d’étude
(CHU de Tlemcen)
D’après la figure 21, on note qu’il ya une augmentation importante de souches d’E. coli
BLSE durant l’année 2013 avec une fréquence de 55,5℅. Nous avons montré que dans un intervalle
de trois ans, la progression de la résistance des germes vis-à-vis des différentes familles
d’antibiotiques s'est multipliée.
L’acquisition par les bactéries de moyens de lutte contre l’effet létal des antibiotiques ou les
transformations génotypiques qui en résultent, progressent dans le même sens que cette résistance
rendant la bactérie de plus en plus virulente (Ecdc, 2011).
Il est actuellement prouvé que l'utilisation des antibiotiques, notamment les céphalosporines de
3éme génération dans un but thérapeutique est le facteur de risque le plus important dans le
développement de résistances bactériennes qui est devenue un problème majeur de santé public
(Rubin et Samore, 2002).
III.17. Concentrations minimales inhibitrices de céfotaxime contre E. coli BLSE
Les résultats des concentrations minimales inhibitrices de céfotaxime contre E. coli BLSE
sont illustrés dans le tableau 35.
Tableau 35 : Concentrations minimales inhibitrices (µg/mL) de céfotaxime contre E. coli BLSE
E.coli 01 02 03 04 05 06 08 09 10 11 12 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29
CMI 128 32 4 4 64 8 4 4 8 64 16 4 6 4 64 64 4 4 64 16 4 64 4 8 4 4 4
D’après le tableau 35, on note que les valeurs absolues des CMI de céfotaxime des 27
souches d’E. coli résistantes oscillent dans un intervalle allant de 128µg/ml à 4 µg/ml. Ce sont des

Résultats et discussion
77
valeurs très élevées comparées aux valeurs trouvées par Lerma et al., (2008) soit de 0,007 a 0,125
µg/ml.
III.18. Profil moléculaire de la résistance d’E. coli BLSE aux β-lactamines
III.18.1. PCR Standard
L’amplification par PCR standard en utilisant les amorces spécifiques qui codent pour les
BLSE et la révélation des gènes par l’électrophorèse sur gel d’agarose ont montré que les 27
souches d’E. coli BLSE isolées dans cette étude se sont avérées productrices de moins de deux
gènes qui codent pour le BLSE (figures 22a, 22b et tableau 36).
Figure 22a : détection des gènes bla CTXM15 Figure 22b : détection des gènes bla TEM1
T1+: bla CTXM15 - T2+: bla CTXM15
D’après le tableau 36, on remarque que les 27 souches d’E.coli BLSE (100%) possèdent le
gène bla CTX-M «CTX-M 14 (7,40℅), CTX-M15 (92,59℅), CTX-M28 (3,7℅), CTX-M1 (3,7℅)»,
1 seule souche (3,7%) possède le gène bla SHV et 26 souches (96,29%) possèdent le gène bla TEM.
100 pb 200pb 500 pb 1000 pb
100 pb 200pb 500 pb 800 pb
T1+ T2+ 17 4 3 2 1 MP
19 14 21 18
MP 19 20 17 16 15 14 13 T1+ T2+

Résultats et discussion
78
Tableau 36 : Profil moléculaire des 27 souches E. coli BLSE
Référence de la
souche
Gènes codant pour les BLSE
TEM1 SHV12 CTXM-14,1, 28 CTXM-15
Ec01 TEM1 - - CTXM-15
Ec02 TEM1 - CTXM-15
Ec03 TEM1 - CTXM-14 CTXM-15
Ec04 TEM1 - CTXM-15
Ec05 TEM1 - CTXM-14 CTXM-15
Ec06 TEM1 - - CTXM-15
Ec08 TEM1 - - CTXM-15
Ec09 TEM1 - - CTXM-15
Ec10 TEM1 - - CTXM-15
Ec11 TEM1 - - CTXM-15
Ec12 TEM1 - - CTXM-15
Ec14 TEM1 - - CTXM-15
Ec15 TEM1 - - CTXM-15
Ec16 TEM1 - - CTXM-15
Ec17 - - - CTXM-15
Ec18 TEM1 - - CTXM-15
Ec19 TEM1 - - CTXM-15
Ec20 TEM1 - CTXM-1 -
Ec21 TEM1 - CTXM-28 -
Ec22 TEM1 - - CTXM-15
Ec23 TEM1 - - CTXM-15
Ec24 TEM1 - - CTXM-15
Ec25 TEM1 - - CTXM-15
Ec26 TEM1 - - CTXM-15
Ec27 TEM1 - - CTXM-15
Ec28 TEM1 - - CTXM-15
Ec29 TEM1 SHV12 - CTXM-15

Résultats et discussion
79
III.19. Recherche moléculaire de la résistance croisée
III.19.1-Résistance aux aminosides
A- Les enzymes modificatrices des aminosides
Les résultats des gènes qui codent pour les enzymes modificatrices des aminosides sont
présentés dans le tableau 37 et a par la figure 23.
Tableau 37: Les gènes qui codent pour les enzymes modificatrices des aminosides
Référence de la souche Gènes codant pour les
BLSE
Référence de la souche Gènes codant pour les
BLSE
Ec 01 aac(6')-Ib-cr Ec16 aac(6')-Ib-cr
Ec 02 aac(6')-Ib-cr Ec17 -
Ec 03 - Ec18 -
Ec 04 aac(6')-Ib-cr Ec19 aac(6')-Ib-cr
Ec 05 aac(6')-Ib-cr Ec21 -
Ec 06 - Ec22 -
Ec 08 aac(6')-Ib-cr Ec23 -
Ec 09 aac(6')-Ib-cr Ec24 -
Ec 10 - Ec25 -
Ec 11 aac(6')-Ib-cr Ec26 -
Ec 12 - Ec27 -
Ec14 aac(6')-Ib-cr Ec 28 aac(6')-Ib-cr
Ec15 aac(6')-Ib-cr
Ec 29 aac(6')-Ib-cr
T1+: aac(6’)-Ib- T2+: aac(6’)-Ib
Figure 23 : Les enzymes modificatrices des aminosides
MP T1+ T2+ 22 17 19 21 18
1000pb 600pb 200pb

Résultats et discussion
80
D’après le tableau 37 et la figure 23, on note que la PCR standard des gènes qui codent pour
les enzymes modificatrices des aminosides ont fait apparaitre la présence de 13 souches d’E. coli
BLSE (48,14%) qui portent le gène aac(6)-lb.
B- Les 16S ARNr methyltransphérases
Les résultats des gènes qui codent pour le 16s ARNr methyltransférase des souches qui
résistent aux aminosides sont présentés dans le tableau 38.
Tableau 38 : Les 16S ARNr methyltransphérases
Référence de la
souche
Gènes codant pour les BLSE
armA rmt B
rmt A
rmtC
Ec 03 - - - -
Ec 04 - - - -
Ec 05 - - - -
Ec 06 - - -
Ec 08 armA rmt B - -
Ec 09 armA rmt B - -
Ec 12 - - - -
Ec 14 - - - -
Ec 16 - - - -
Ec 17 armA - - -
Ec 21 - - - -
Ec 22 - - - -
D’après le tableau 38, on note que la PCR a montré que 3/12 souches d’E. coli BLSE
(25%) possèdent le(s) gène(s) armA et/ou rmt B qui code(nt) pour le 16s ARNr methyltransférase
responsable(s) de la résistance élevée à tout les aminosides. En revanche, aucune souche d’E. coli
BLSE ne possède les gènes rmt A et rmt C.
III.19.2. Résistance aux quinolones
Les résultats des gènes qui codent pour les quinolones sont présentés dans le tableau 39.

Résultats et discussion
81
Tableau 39: Profil moléculaire de la résistance associée des souches d’E. coli BLSE aux
quinolones.
Référence de la
souche
Gènes codant pour les BLSE
qnrB qnrA qnrS
Ec 01 - - -
Ec 02 - qnrA1 -
Ec 03 - - -
Ec04 qnrB1 - -
Ec 05 - -
Ec 06 - - -
Ec 08 - - -
Ec 09 - - -
Ec 10 - - -
Ec 11 - - -
Ec 12 - - -
Ec 14 - - -
Ec 15 - - -
Ec 16 - - -
Ec 17 - - -
Ec 18 qnrB1 - -
Ec 19 - - -
Ec 20 - - -
Ec 21 - - -
Ec 22 - - -
Ec 23 - - -
Ec 24 - - -
Ec 25 - - -
Ec 26 - - -
Ec 27 qnrB1 - -
Ec 28 qnrB1 - -
Ec 29 _ - -
D’après le tableau 39 ; on note que l’amplification par PCR standard a montré que 5
souches E. coli BLSE (18.51%) isolées dans cette étude possèdent le gène qnrb ou qnra qui

Résultats et discussion
82
codent pour la résistance aux fluoroquinolones. Le gène qnr a toujours coexisté avec le gène
aac(6’)-Ib-cr, ce dernier code pour la résistance simultanée aux aminosides et à la ciprofloxacine.
Le support génétique des enzymes BLSE produits par les 27 souches d’E.coli isolées dans
notre étude était varié. Le type CTX-M a été détecté chez les 27 souches (100%), le type SHV chez
une seule souche (3.7%) et 26 souches (96,44%) hébergeaient le gène TEM.
Cette étude souligne, d’une part l’augmentation de la fréquence de la production de BLSE par
les souches d’E.coli isolées en Algérie, et d’autre part confirme la propagation mondiale du gène
CTX-M-15 (Carrer et Nordmann, 2011). Dans la présente étude, la diversité génétique des gènes
BLSE peut être attribuée à des gènes déjà détectés (TEM-1, SHV-12, CTXM-14, 15, 28,1) (Baba
Ahmed-Kazi, 2013 ; Messai et al., 2008 ; Nedjai et al., 2012 ; Ramdani-Bouguessa et al, 2003 ;
Touati et al., 2006)
La souche BLSE SHV-12 a été isolée pour la première fois en 1997 en Suisse chez une
souche de K. pneumoniae isolée d’un patient venant de l’Afrique du Nord (Nüesch-Inderbinen et
al., 1997). Depuis lors, cette BLSE a une répartition mondiale : Australie (Howard et al., 2002),
Canada (Mulvey et al., 2004), Chine (Chanawong et al., 2002), Croatie (Bedenic et al., 2001),
France (Weill et al., 2004), Grèce (Neonakis et al., 2003), Inde (Dhawan et al., 2003), Italie
(Perilli et al., 2002), Japon (Yagi et al., 2000), Corée (Kim et al., 1998), Espagne (Briñas et al.,
2003), Taiwan (Yan et al, 2000), Thaillande (Chanawong et al., 2001), Etats- Unis (Wong-
Beringer et al., 2002), Afrique du Sud (Kruger et al., 2004), Tanzanie (Blomberg et al., 2005),
Cameroun (Gangoué Piéboji, 2000), RCA (Frank et al., 2006), Tunisie (Ben-Hamouda et al.,
2004), Sénégal (Cardinale et al., 2000), Mali (Weill et al., 2004 et Algérie (Iabadene et al., 2008 ;
Iabadene et al., 2009 ; Berrazeg et al., 2013).
Les souches BLSE de type SHV, TEM et CTX-M-1 ont été rapportées dans différentes études
algériennes. Touati et al., (2008a) ont révélé la présence de CTX-M-15 dans des souches
environnementales de K. pneumoniae et E. cloacae, sachant que CTX-M-15 fait partie du groupe
de CTX-M-1 (Gangoué Piéboji, 2007). Messai et al., (2008) ont également rapporté des souches
de K. pneumoniae portant les gènes bla TEM et bla CTX-M-1. Dans une autre étude, Touati et al.,
(2010) ont rapporté en plus de la présence de CTX-M-15, la détection de SHV-12 chez des souches
de l’environnement. Au Bénin, Ahoyo et al., (2007) ont également rapporté la présence du gène bla
SHV chez des souches environnementales d' E. coli. La BLSE du type CTX-M a été rapportée pour
la première fois en Europe Occidentale en 1989 et c’était la CTX-M-1. Cette enzyme a été détectée
en Allemagne, en Italie et en France chez des espèces variées d’Entérobactéries. La CTX-M-15 a
été identifiée pour la première fois en Inde et au Japon et a été par la suite mise en évidence dans
plusieurs autres pays (Canada, Russie, Bulgarie, Pologne, France, Royaume uni, Turquie et

Résultats et discussion
83
Taiwan). Trois autres types de CTX-M ont été rapportés en Afrique, la CTX-M-2 (Afrique du Sud),
la CTX-M-3 (RCA) et la CTX-M-12 (Kenya) (Gangoué Piéboji, 2007).
La résistance aux β-lactamines chez les entérobactéries est dominée par la production de
BLSE de type CTX-M-3 et CTX-M-15 (Baba Ahmed-Kazi Tani et Arlet, 2014). Plusieurs études
ont rapporté ces BLSE dans différents régions en Algérie : à Annaba (Bouzidi et al., 2011; Nejai et
al., 2012), à Constantine (Naas et al., 2005; Naas et al., 2011), à Bejaia (Touati et al., 2006;
Touati et al., 2008 b, Gharout-Sait et al., 2012), à Alger (Ramdani-Bouguessa et al., 2006;
Iabadene et al., 2008; Iabadene et al., 2009; Ramdani-Bouguessa et al., 2011; Touati et al.,
2012; Kermas et al., 2012) et à Tlemcen (Baba Ahmed et al., 2012; Baba Ahmed-Kazi Tani et
al., 2013).
Il faut noter que les différents types de BLSE (CTX-M-1, TEM et SHV) trouvés dans notre
étude font partie des BLSE dites transférables, ce qui explique la facilité du transfert entre les
entérobactéries (Philippon, 2013). La présence des souches environnementales productrices de
BLSE de différents types peut agir comme un réservoir de gènes de résistance qui peuvent être
transmis horizontalement à des souches cliniques ou environnementales. En plus, elle constitue un
facteur de risque des épidémies nosocomiales (Touati et al., 2008a). Par conséquent,
l’environnement inanimé des patients (surfaces et objets), contaminé par des entérobactéries multi
résistantes peut servir comme un réservoir secondaire pour une transmission croisée. En effet,
plusieurs auteurs, ont rapporté des souches environnementales qui ont été identifiées comme
sources d’épidémies (Touati et al., 2010). Par ailleurs, la coexistence des différentes -lactamases
dans une même bactérie peut poser un problème de diagnostic et de thérapeutique (Roh et al.,
2008).
La résistance aux quinolones est codée par différents types de gènes dont les gènes qnr qui
ont été identifiés dans différentes espèces d’entérobactéries essentiellement K. pneumoniae,
Enterobacter spp., E. coli et Salmonella enterica, aussi bien en milieu hospitalier que
communautaire (Rodriguez-Martinez et al., 2011). Elle est souvent associée à la production de
bêta-lactamases à spectre élargi Guessennd et al., 2008).
Dans notre étude la répartition des gènes de résistance plasmidique aux quinolones montre
que les gènes qnr sont présents chez 5 souche (qnrB1 chez 4 souches et qnrA1 chez une
souche).qnrA1 a été identifié en 1998 sur un large plasmide conjuguait isolé à partir d’une souche
de K. pneumoniae résistante à la ciprofloxacine (Martinez-Martinez et al., 1998). Les gènes qnr B
et qnr S ont été décrits dans peu de pays (Guessennd et al., 2008).

Résultats et discussion
84
En Algérie, les gènes de résistance aux fluoroquinolones sont d’identification plus récente
dont les plus répandus sont les déterminants qnr (Baba Ahmed-Kazi Tani et Arlet, 2014). Aussi,
les gènes qnr (A, B, S) ont été détectés chez des souches hospitalières d’entérobactéries produisants
des BLSE (CTX-M, SHV-12 et VEB-1) (Iabadene et al., 2008; Meradi et al., 2011 ; Touati et al.,
2008 b). Plus récemment les gènes qnr ont été isolés chez des souches d’E.coli produisantes des
BLSE de type CTX-M et responsables d’infections urinaires en milieu communautaire à Bejaia
(Gharout-Sait et al., 2012).
D’autres études basées sur des approches moléculaires ont montré la relation entre les
déterminants de la résistance aux quinolones et la production de BLSE (Rodriguez-Martinez et al.,
2011; Poirel et al., 2006; Ambrozic Avgustin et al., 2007; Fihman et al., 2008; Lavilla et al.,
2008). Le aac(6’)-Ib-cr a été découvert pour la première fois en 2006 chez une souche d’E.coli
qnrA-positive en Chine (Robicsek et al., 2006). Le gène aac (6')-lb-cr confère une résistance
simultanée à la ciprofloxacine et aux aminosides (Bouchakour et al., 2010). En Algérie, il a été
détecté pour la première fois en 2009 chez une souche d’E. cloacae (Meradi et al., 2011) et dans
des travaux antérieures (Baba Ahmed-Kazi, 2013).
L’étude des associations des gènes de résistance qnr et aac(6’)-Ib-cr entre eux d’une part, et
avec le phénotype BLSE d’autre part montre que les souches EBLSE contenant le gene aac(6’)-Ib-
cr sont les plus nombreuses (48%), suivies des E coli co-exprimant les gènes qnr et aac(6’)-Ib-cr
(Guessennd et al., 2008). Le gène aac(6’)-Ib-cr est souvent associé à d’autres gènes de résistance
aux quinolones tels les différents variant qnr, les BLSEs (CTX-M-1, CTX-M-14, CTX-M-15, CTX-
M-24), les céphalosporinases plasmidiques (DHA-1) et les carbapénémases (KPC-2) (Strahilevitz
et al., 2009). Il a été essentiellement identifié chez des souches cliniques d’E.coli et de K.
pneumoniae (Poirel et al., 2012).
En outre, le problème lié aux BLSE est surtout la présence fréquente de corésistances rendant
les souches multirésistantes (Leotard et Negrin, 2010). En effet, les BLSE sont généralement
portées par de grands plasmides qui portent aussi des -lactamines, tels que les gènes de résistance
aux classes d’antibiotiques non aminoglycosides, les quinolones et le
triméthoprime/sulfaméthoxazole. Aussi, l’utilisation de ces antibiotiques contribue à la sélection de
souches productrice de BLSE (Paterson et Bonomo, 2005).
L’incidence d’apparition des résistances aux aminosides a augmenté ces dernières années et
particulièrement chez les souches productrices de BLSE (Spanu et al., 2002).
Parmi les autres AMEs, 3 souches d’E.coli BLSE hébergent le gène armA et/ou rmtB. Toutes
les souches qui possèdent le gène armA et rmtB ont exprimé de hauts niveaux de résistance à
l’amikacine et à la gentamicine. Cependant, ce gène a été trouvé chez des souches de

Résultats et discussion
85
K. pneumoniae isolées chez quatre patients d’origine Algérienne. Ce qui a été rapporté par une
enquête dans les hôpitaux Belges entre 2000 et 2005 (Bogaerts et al., 2007). En Algérie, le gène
armA a également été trouvé chez Salmonella enterica productrice de BLSE à Constantine (Naas et
al., 2009) et Salmonella non-typhi productrice de BLSE de type CTX-M-15 à Annaba en 2009
(Bouzidi et al., 2011). Récemment, le gène armA a été identifié en combinaison avec le gène
NDM-1 chez K. pneumoniae au Népal. Notre taux d’isolement du gène armA est plus élevé que
celui constaté chez les entérobactéries productrices de BLSE en Grèce (0,2%) en 2012 et en France
(1,3%) en 2007(Bercot et al., 2008). Par ailleurs, la prévalence du gène ArmA reste inférieure à
celle trouvée chez les souches de K. pneumoniae « BMR » en Chine (54,5%) en 2009 (Yang et al.,
2011). Le gène rmtB a été détecté principalement en Asie et en Europe chez les Entérobactéries
(Bogaerts et al., 2007). L’équipe japonaise de Yamane et al., (2005) ont constaté que le gène
rmtB est trouvé à la fois dans Klebsiella spp. et E. coli avec un taux de prévalence de 0,008% et
0,02% respectivement.
La méthylase ARNr 16S semble donc être plus répandue en Asie qu'en Europe et en
Amérique. La résistance transférable aux C3G et aux aminosides par production de BLSE et le 16s
ARNr methyltranspherase entraîne l’émergence de bactéries multi-résistantes BMR, limitant ainsi
les possibilités thérapeutiques. La dissémination de ces BMR peut impliquer l’association de
plusieurs mécanismes : propagation de souches épidémiques ou gènes de résistance. Les
investigations concernant cette dissémination nécessitent une compréhension des mécanismes et les
risques potentiels qui s’y rapportent.
Ainsi, un traitement efficace doit être sélectionné en fonction de l'antibiogramme afin de
limiter l'utilisation des carbapénèmes lorsque cela est possible. En effet, la classe des carbapénèmes
comme l’imipenème est le traitement de premier choix pour traiter les bactéries productrices des
enzymes BLSE parce que ces enzymes ne l’hydrolysent pas (Nordmann et Carrer, 2010).
Actuellement, les carbapénèmes sont de plus en plus utilisés dans le monde entier car c'est souvent
le médicament de dernier recours pour traiter les infections graves causées par les entérobactéries
productrices de BLSE. Néanmoins, Il a été démontré que, lors de ce traitement, une fréquence
élevée de la résistance à l'imipénème peut être sélectionnée (Belbel et al., 2014).
III.19.3. Caractérisation des céphalosporinases plasmidiques
Parmi les 6 souches isolées qui résistent a la cefoxitine, deux souches d’E.coli (Ec 19 et Ec
26) ont été détectées positives pour le gène blamox et blaCMY-2, soit une prévalence de 33%.
L’amplification a permis de préciser la dissémination des céphalosporinases plasmidiques AmpC
de type MOX, CMY-2 chez les deux souches isolées en 2013 et 2012 à partir de deux services :

Résultats et discussion
86
médecine interne et urologie. En revanche, aucune souche d’E. coli BLSE ne possède les gènes bla
dha et bla FOX. Des études réalisées en Egypte, Pakistan et Tunisie ont montré les mêmes données
où il y’a la diffusion des céphalosporinases plasmidiques MOX associés avec les mécanismes de
production de β-lactamases à spectre étendue BLSE chez les souches d’E. coli (Wassef et al., 2014
; Cherif et al., 2009). Aux États-Unis Kaye et al., (2004), ont montré qu’il y’a une diffusion des
céphalosporinases plasmidiques.
Ces enzymes ont été plus récemment individualisées chez des entérobactéries comme E. coli,
K. pneumoniae, Klebsiella oxytoca, P. mirabilis et Salmonella spp. Ces céphalosporinases
plasmidiques sont très proches génétiquement des céphalosporinases AmpC chromosomiques et le
transfert des gènes codants les céphalosporinases sur les plasmides implique des éléments
génétiques mobiles (Bush et al., 1995 ; Philippon et al., 2002). Généralement, les gènes codants
les enzymes AmpC sont localisés sur des plasmides dont la taille varie de 7 à 180 kb (Philippon et
al., 2002).
La dissémination de la multirésistance et la diffusion des gènes de résistance sont liées à
l’existence d’éléments génétiques mobiles entre bactéries d’une même espèce ou d’espèces
différentes, ainsi qu’à l’existence de structures génétiques permettant de cumuler de nombreux
gènes de résistance au sein d’une même souche (Skurnik et et al., 2006).
III.19.4. Étude du phénotype des facteurs de virulence
Les résultats des gènes de virulence sont consignés dans le tableau 40. On note que
l’amplification par PCR standard montre que 12 souches d’E. coli sur 27 possèdent l’opéron
d’addition pap et/ou l’opéron sfa et n’expriment pas le gène hly. Par contre, une souche d’E. coli
possède le gène cnf1.
Les travaux d’Andreu et al., (1989) ont identifié le gène de virulence de type fimbriae I
(FimH) chez E. coli isolées chez des patients qui ont une pyélonéphrite, cystite, et UTI avec des
pourcentages 97%, 97% et 90%, respectivement. En outre, la fréquence de papC a été de 73%, 0%
et 20%, respectivement. Blanco en 1997 a indiqué que les taux de sfa , papC et afa d’ E. coli
isolées chez des patients UTI était de 53%, 54% et 2%, respectivement. Dans une autre étude,
Santo en 2007 a trouvé des gènes de virulence chez E. coli de sfa (19%), aérobactine (76%), papC
(11%), afa (32%) et hémolysine (96%). Alors que Tiba et al., (2008) ont trouvé des gènes différents
chez E. coli provenant de patients atteints de cystite de virulence FimH (97,5%), papC (32,7%),
afa (27,8%), IUCC (25,9%), hly (25,3%), et afa (6,2%).

Résultats et discussion
87
Tableau 40 : phénotype des facteurs de virulence
E.coli 01 02 03 04 05 06 08 09 10 11 12 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29
sfa - - - - Sfa - Sfa Sfa - - - Sfa - - Sfa - - - - - - - - - - sfa sfa
pap - - - Pap - - - - - Pap - - - Pap - - - - Pap - - - pap - - pap -
cnf1 - - - - -
- -
-
- -
- -
- - - - - - - - - - - - - - cnf1

Résultats et discussion
88
Il semble que certains facteurs d’uropathogénicité comme les systèmes d’adhésion pap et sfa
jouer un rôle majeur dans la survenue d’infection urinaire symptomatique. En outre, l'incidence des
gènes de virulence chez E. coli isolées à partir adultes roumain avec UTI était FimH (86%), sfa / foc
(23%), papC (36%), et afa (14%) (Usein et al., 2001).
Les facteurs d’uropathogénicité des E. coli et leurs déterminants génétiques sont bien connus.
Ils sont soumis à une régulation complexe qui dépend essentiellement des conditions de
l’environnement dans lequel la bactérie doit se développer. Selon de nombreuses études
épidémiologiques, dans les IUS, les facteurs d’uropathogénicité des E. coli peuvent être corrélés
avec la symptomatologie des cystites et des pyélonéphrites (Michel, 2014).
En fait, il n’existe pas de bactérie uropathogènes type et on peut considérer que plus une
bactérie possède de facteurs d’uropathogénicité, plus elle est pathogène pour son hôte. Le nombre
de facteurs présents est variable mais certains comme les systèmes d’adhésion Pap ou les toxines
comme l’hémolysine α ou le Cnf semblent plus spécifiques de l’uropathogénicité. En outre, certains
de ces facteurs sont capables de se transmettre par transfert horizontal lorsque les opérons qui les
codent se trouvent regroupés sur des loci du chromosome dénommés par Hacker et al., (2003).
Ilots de pathogénicité. Au total, les bactéries retrouvées dans les infections urinaires possédant ces
facteurs doivent être soumises à un traitement antibiotique bien codifié selon le type d’infection.
II.19.5. Détermination de la relation épidémiologique
L’analyse génotypique par ERIC-PCR des souches d’E. coli BLSE a montré une grande
diversité de souches : 13 différents génotypes parmi les 27 souches étudiées. La détermination des
groupes phylogénétiques a montré que 9 clones appartiennent au groupe phylogénétique A0 et A1,
3 clones au groupe phylogénétique B1, 4 clones au groupe phylogénétique B2 et 1 clone au groupe
phylogénétique D1 (tableau41).

Résultats et discussion
89
Tableau 41: Relation épidémiologique entre les 27 souches d’E. coli BLSE étudiées par ERIC/-
PCR et groupement phylogénétique
Code service groupement
Phylogenetique
ERIC
profile
Ec 06 Maternité A0 c
Ec 12 Urologie A0 f
Ec 22 Médecine interne A0 j
Ec 24 maternité A0 e
Ec 26 Urologie A0 p
Ec 27 Maternité A0 o
Ec 01 Maternité A1 a
Ec 02 Maternité A1 q
Ec 04 Maternité A1 c
Ec 05 Maternité A1 c
Ec 08 Maternité A1 e
Ec 09 Urologie A1 f
Ec 14 Urologie A1 g
Ec 18 Médecine interne A1 c
Ec 23 Urologie A1 k
Ec 03 Maternité B1 b
Ec 10 Urologie B1 f
Ec 16 Médecine interne B1 h
Ec 19 Médecine interne B1 h
Ec 25 Médecine interne B1 i
Ec 20 Médecine interne B22 h
Ec 11 Urologie B23 f
Ec 15 Urologie B23 h
Ec 17 Médecine interne B23 h
Ec 21 Médecine interne B23 i
Ec 29 Urologie B23 n
Ec 28 Urologie D1 e
La phylogénie moléculaire a pour but de reconstruire les relations de parenté entre les
séquences de nucléotides ou d’acides aminés. On peut ainsi étudier les relations de parenté entre les
espèces qui les portent mais, aussi, l’évolution du génome.
D’après le tableau 41, on remarque que les souches isolées sont du groupe phylogénétique B1
et A1. Alors que les souches du groupement phylogénétique B2 et D1 sont moins fréquents.
Selon la littérature, les souches du groupe phylogénétique B1 et A1 sont moins virulentes et
dite commensale. Il existe une corrélation entre la résistance aux antibiotiques, les facteurs de
virulence et le groupement phylogénétique des souches. Les souches de groupe B1 et A sont
fréquemment résistantes a de nombreux antibiotique alors que les souches B2 demeurent
relativement sensible (Moran et al., 2006 ; Corvec et al., 2007 ).

Résultats et discussion
90
Les souches de groupe B1, A et D sont plus fréquemment associes a une rareté des facteur de
virulence, a une résistance aux antibiotiques, a une origine nosocomiale de l’infection, a une porte
d’entré urinaire ou non et au statut immunodéprimé (Moran et al., 2006 ; Corvec et al., 2007 ).
Conclusion :
Les infections urinaires posent un problème majeur de la santé publique, du fait de leur
fréquence très élevée, leur coût culminant de traitement et les multiples échecs de l’antibiothérapie
à cause des bactéries multi résistantes incriminées dans ces infections.
Les résultats de cette étude ont montré que sur 441 échantillons urinaires provenant des
patients hospitalisés 132 répondaient aux critères d'infection urinaire. Le rapport Ratio F/H est égal
à 1,4. L’infection urinaire est plus importante chez les tranches d’âge comprises entre 20 et 39 ans.
Les agents infectieux impliqués dans ces infections étaient surtout E. coli, l'espèce la plus
fréquemment isolée (63℅). La résistance des souches d'E. coli isolées aux ATB est très importante:
83,13% sont résistantes à l’amoxiciline, 62,65% à l’amoxicilline plus l’acide clavulanique et 56,62
% aux céphalosporines de 3eme génération.
L'analyse des phénotypes de résistance aux -lactamines d’E .coli isolées au CHU de Tlemcen,
a montrée une nette dominance des souches productrices de -lactamases à spectre étendu BLSE
(32,5%) appartenant au génotype CTX-M 15.
III.20. Effet des HEs des plantes aromatiques retenues sur les souches d’E. coli BLSE
III.20.1. Résultats des extractions
Les rendements en HE des trois plantes retenues et leurs caractères organoleptiques sont
résumés dans le tableau 42.
Tableau 42 : Rendements en HE et caractéristiques organoleptiques
Plante Partie
utilisée
Propriétés organoleptiques Teneur
(%)
Durée
d’extraction
(h) Aspect Couleur Odeur Saveur
Cinnamomum
cassia
Ziziphora
hispanica
Coriandrum
sativum
Ecorces
Feuilles
et fleurs
Graines
Liquide
Liquide
Liquide
Jaune foncé
Jaune pale
Jaune clair
légèrement
sucrée
Fraîche
légèrement
sucrée et
citronnée
Aromatique
Suave
Aromatique
1.7
1.5
1.2
8
3
5

Résultats et discussion
91
D’après le tableau ci–dessus, on remarque que les rendements en HEs sont variables selon les
espèces. En général, nous avons obtenu de bons rendements en HE comparés à ceux obtenus par
d’autres chercheurs (tableau 43).
Tableau 43 : Rendements en HE rapportés dans la littérature.
Espèces Teneur (%)
Cinnamomum cassia
Ziziphora hispanica
Coriandrum sativum
Nos résultats Autres travaux Références
1.7
1.5
1.2
0,5 - 2
0.5
0,7
Richard, (2008)
Bekhchi et al., (2007)
Skiredj et al., (2012)
L’espèce Cinnamomum cassia a donné un rendement en HE (1,7%) très proche de celui
obtenu par Richard, 2008 mais les rendements en HE obtenus pour Ziziphora hispanica (1,5%) et
Coriandrum sativum (1,2%) sont supérieurs à ceux obtenus par bekhchi et al., (2007) et Skiredj et
al., (2012). Cette différence de rendements en HEs dépend de plusieurs facteurs biotiques et
abiotiques comme la période de récolte, le type d’organes récoltés, le sol et la pluviométrie
(Schmidt, 2010).
III.20.2. Analyse chimique des huiles essentielles
Les résultats des analyses chimiques par CG/SM des huiles essentielles sont présentés dans le
tableau 44 et illustrés par les chromatogrammes (voir annexe 8).
Les résultats de l’analyse chimique montrent que les HEs de C. sativum et de Z. hispanica
sont riches en monoterpènes oxygénés, respectivement 95,71% et 93,66%, alors que l’HE de
C. cassia est riche en phénylpropanoides (80,12%).

Résultats et discussion
92
Tableau 44: Composition chimique (%) des huiles essentielles des plantes retenues
Composés Ira Irp C. sativum Z. hispanica C. cassia
α–Pinène 931 1022 0.79 0.49 1.69 Camphène 943 1066 - - 0.67
–Pinène 970 1110 - 0.55 0.50
p–Cymène 1011 1268 1.36 - -
Limonène 1020 1199 0.68 3.36 - 1,8-Cinéole 1022 1209 - - 2.52
(Z)–beta–Ocimène 1034 1247 0.24 - - Linalol 1081 1544 85.57 - 0.22
(E) oxyde de Linalol 1044 1469 0.50 - -
(Z) oxide de Linalol 1073 1469 0.44 - - Camphre 1123 1517 4.78 - 1.37 p–menth–3–en–8–ol 1135 1595 - 2.46 - Bornéol 1148 1698 - - 0.44 (E)-Isopulégone 1149 1572 - 0.88 - Terpinène- 4-ol 1161 1600 - - 0.77
α–Terpinéol 1179 1700 0.48 - 1.17 (Z)-Cinnamaldéhyde 1182 1867 - - 0.96 Pulégone 1216 1645 - 88.33 -
Géraniol 1232 1844 2.76 - - Pipéritone 1232 1730 - 0.34 - (E)-Cinnamaldéhyde 1233 2044 - - 75.85 (E)–Pipéritone oxide 1235 1724 - 0.46 - Thymol 1266 2189 - 0.38 - Acétate de Bornyl 1269 1515 - - 0.78 Carvacrol 1278 2219 - 0.20 - Pipériténone 1315 1909 - 0.61 -
Acétate de néryl 1342 1725 1.18 - - α –Copaène 1379 1488 - - 0.11 Coumarin 1391 1488 - - 0.46 Acétate de Cinnamyl 1413 2152 - - 3.31
Total
98.78 98.06 90.82
Monoterpènes
hydrocarbonés 3.07
4.40
2.86
Monoterpènes
oxygénés 95.71
93.66
7.27
Phénylpropanoides - - 80.12
Sesquiterpènes
hydrocarbonés -
-
0.11
Sesquiterpènes
oxygénés -
-
0.46 Ira : Indice de rétention sur colonne apolaire, Irp : Indice de rétention sur colonne polaire
- : Absence ; ID : identification ; SM : spectrométrie de masse.
En effet, le linalol est le composé majoritaire de l’HE de C. sativum (85,57%) alors que la
pulégone est le composé majoritaire de l’HE de Z. hispanica (88,33%). Cependant, le
cinnamaldéhyde est le composé majoritaire de l’HE de C. cassia (75,85%).

Résultats et discussion
93
En comparant nos résultats avec ceux de la littérature, on remarque que les mêmes composés
majoritaires des HEs de Ziziphora hispanica et de Coriandrum sativum sont retrouvés mais avec
une variabilité quantitative (tableau 45).
Tableau 45 : Composés majoritaires des trois HEs rapportés dans la littérature.
Espèces Nos résultats Autres travaux Références
Ziziphora hispanica
Coriandrum sativum
Cinnamomum cassia
Pulégone (88.33℅)
Linalol (85.57℅)
Cinnamaldehyde
(75.85℅)
Pulégone (79,5℅)
Linalol (57,57℅)
Cinnamaldehyde
(42,37℅)
Bekhchi et al., (2007)
Khani et Rahdari, (2012)
Chang et al., (2013)
D’après le tableau on remarque que l’espèce Ziziphora hispanica a donné un pourcentage de
Pulégone (88,33%) très proche de celui obtenu par bekhchi et al., (2007), mais les pourcentages
de linalol (85,57%) et du cinnamaldéhyde (75,85℅) sont supérieurs à ceux obtenus par Khani et
Rahdari, (2012) et Chang et al., (2013) respectivement.
III.20.3. Etude du pouvoir antibactérien des huiles essentielles
Pour rappel, l’étude a été faite sur les souches E.coli BLSE responsables d’infections
urinaires. Pour comparaison, nous avons élargi les tests sur d’autres souche responsables d’infection
urinaire (Klebsiella pneumonia, Proteus mirabilis et Pseudomonas aeruginosa), sur des souches
E.coli sensibles et sur des souches de référence.
III.20.3.1. Méthode des disques
Les résultats du pouvoir antimicrobien des trois HEs et de céfotaxime par la méthode des
disques sont consignés dans le tableau 46 et la figure 24.
On remarque que l’HE de C. cassia est la plus active sur les souches testées en particulier sur
les souches E. coli BLSE. Par contre, les huiles de C. sativum et Z. hispanica ont montré une
moyenne voir une faible activité.
En effet, l’HE de Cinnamum à donné des zones d’inhibitions de 27 a 39 mm pour toutes les
souches sauf pour P. aeruginosa (12 à 19 mm) malgré le faible volume utilisé d’huile (5μl).
Selon les moyennes des zones d’inhibitions on peut classer l’efficacité des HEs par ordre
décroissant comme suit : C. cassia (27.35 mm), C. sativum et Z. hispanica ( 11 mm).
Selon la sensibilité des souches, E. coli, Proteus et Klebsiella ont montré la plus grande
sensibilité à l’HE de C. cassia par rapport aux autres HEs. Les souches E. coli BLSE ont enregistré
un diamètre moyen 33 mm, alors qu’on note une résistance de P. aeruginosa vis-à-vis des huiles
de C. sativum et Z. hispanica.

Résultats et discussion
94
Tableau 46 : Diamètres d’inhibitions (en mm) des huiles essentielles et de céfotaxime
HE et ATB
Souches
Z. hispanica C. cassia C. sativum Céfotaxime
Escherichia coli ATCC 25922 14±1 30±1 13±1 29
Escherichia coli 72 S 17±1 35±0 31±1 27±1
Escherichia coli 73 S 17±1 32±1 29±1 30±0
Escherichia coli 74 S 16±0 39±1 16±1 31±0
Escherichia coli 75 S 13±1 38±1 11± 1 28±1
Moyenne E.coli S 15.75 36 21.75 29
Escherichia coli 21 BLSE 08±1 32±1 9±1 21±1
Escherichia coli 17 BLSE 09±1 36±1 10±0 19±1
Escherichia coli 05 BLSE 09±0 34 ±1 10±1 19±1
Escherichia coli 02 BLSE 12±1 35±1 13±1 15±1
Escherichia coli 09 BLSE 10±0 34±1 11±0 11±1
Escherichia coli 16 BLSE 08±1 32±1 09±1 10±1
Escherichia coli 15 BLSE 08±1 34±1 08±1 11±1
Escherichia coli 04 BLSE 07±1 29±1 07±1 15±1
Escherichia coli 06 BLSE 10±1 36±1 11±1 15±1
Escherichia coli 29 BLSE 09±1 32±0 10±0 11±0
Escherichia coli 20 BLSE 12±0 33±1 13±1 10±0
Escherichia coli 19 BLSE 10±1 32±1 10±1 11±1
Moyenne E.coli BLSE 09.33 33.25 10.08 14
P. aueruginosa ATCC 27853 06±1 15±1 06±0 08±1
Pseudomonas aueruginosa S 06±0 19±1 06±0 10±0
Pseudomonas aueruginosa R 06±0 12±0 06±0 13±1
Proteus mirabilis ATCC 35659 09±0 34±1 12±1 19±0
Proteus mirabilis S 10±1 39±0 13±1 19±1
Proteus mirabilis R 08±1 30 ±1 09±1 20±0
K. pneumoniae ATCC 70603 12±1 21±1 14±1 19±1
Klebsiella pneumoniae S 13±1 32±1 14±1 20±1
Klebsiella pneumoniae R 07±1 27±1 10±1 21±1
Moyenne générale 9.67 27.35 11.23 18.41
S : sensible ; R : résistante
En comparant l’effet des HEs au céfotaxime, on remarque que l’effet de C. cassia est plus
élevé que l’antibiotique soit respectivement 27,35 mm et 18,4.

Résultats et discussion
95
Figure 24 : Effet de l’huile essentielle de Cinnamomum cassia sur Pseudomonas aueruginosa
Nos résultats obtenus concordent avec ceux de Vilas et Kamble (2013). Ces auteurs ont
obtenu des zones d’inhibition de 12 à 15 mm vis–à–vis d’E. coli avec l’HE de C. sativum.
Par contre, Ali Khan et al., (2013) n’ont pas trouvé d’activité sur B. cereus, S. aureus,
P. aeruginosa et E. coli, alors que Shahid et Mohammad, (2013) ont obtenu des zones d’inhibition
entre 13 et 20 mm.
Pour Z. hispanica, Bekhchi et al., (2007) ont obtenu des zones d’inhibition de 10,33 mm vis–
à–vis d’E. coli. Par contre pour C. cassia, Al-saghir, (2009) ont obtenu environ 26 mm vis–à–vis
d’E.coli.
Historiquement, les souches de Pseudomonas aeruginosa ont toujours présenté une résistance
vis-à-vis de des huiles essentielles (Sivropoulou et al., 1995; Mimica-Dukié et al., 2003 ; Salehi et
al., 2005). Mais dans notre étude, P. aeruginosa s’est montré sensible à l’HE de cassia.
D’après Kalemba et Kunicka, (2003), la méthode de diffusion sur gélose n’est pas une
technique idéale pour l’évaluation du potentiel antimicrobien des HEs vis–à–vis des
microorganismes à l’état planctonique. Son utilisation reste uniquement pour des criblages rapides.
Par contre, la détermination des CMIs est une technique plus fiable, car cette méthode permet la
quantification de l’activité antimicrobienne de l’agent testé.
II.20.3.2. Résultats des concentrations minimales inhibitrices et bactéricides
Les résultats des concentrations minimales inhibitrices (CMI) et bactéricides (CMB) des HEs
et de céfotaxime vis-à-vis des souches responsables d’infection urinaire et des souches de
références sont présentés dans le tableau 47.

Résultats et discussion
96
Tableau 47 : Concentrations minimales inhibitrices des huiles essentielles (mg/ml) et de
céfotaxime (μg/ml).
HE
Souches
Z. hispanica C. cassia C. sativum
Céfotaxime
CMI CMB CMI CMB CMI CMB CMI
E. coli ATCC 25922 2.5 5.0 0.63 1.25 2.5 2.5 0.125
Escherichia coli 72 S 2.5 2.5
0.63 1.25 1.25 1.25 1.0
Escherichia coli 73 S 2.5 2.5 0.31 0.63 1.25 1.25 1.0
Escherichia coli 74 S 2.5 2.5 0.63 0.63 2.5 5.0 1.0
Escherichia coli 75 S 2.5 2.5 0.63 0.63 2.5 5.0 1.0
Moyenne E. coli S 2.5 2.5 0.55 0.78 1.87 3.12 1.0
Escherichia coli 21 BLSE 5.0 10.0 2.5 2.5 5.0 10.0 64.0
Escherichia coli 17BLSE 5.0 5.0 1.25 2.5 5.0 10.0 64.0
Escherichia coli 05BLSE 5.0 10.0 1.25 2.5 5.0 10.0 64.0
Escherichia coli 02BLSE 5.0 10.0 1.25 2.5 5.0 10.0 32.0
Escherichia coli 09BLSE 10.0 20.0 5.0 5.0 5.0 10.0 4.0
Escherichia coli 16 BLSE 10.0 20.0 2.5 5.0 10.0 20.0 4.0
Escherichia coli 15BLSE 10.0 20.0 1.25 1.25 20.0 40.0 64.0
Escherichia coli 04BLSE 20.0 40.0 2.5 2.5 10.0 20.0 4.0
Escherichia coli 06 BLSE 20.0 40.0 2.5 2.5 10.0 20.0 8.0
Escherichia coli 29BLSE 20.0 40.0 1.25 1.25 10.0 20.0 4.0
Escherichia coli 20 BLSE 20.0 40.0 1.25 1.25 10.0 20.0 4.0
Escherichia coli 19 BLSE 20.0 40.0 1.25 1.25 10.0 20.0 4.0
Moyenne E.coli BLSE 12.5 24.58 1.97 2.5 8.75 17.5 26.66
P. aueruginosa ATCC 27853 2.5 2.5 - - 16.0
Pseudomonas aueruginosa S - - 2.5 2.5 - - 8.0
Pseudomonas aueruginosa R - - 5.0 5.0 - - 64
P. mirabilis ATCC 35659 5.0 5.0 0.31 0.31 5.0 5.0 2.0
Proteus mirabilis S 5.0 5.0 0.31 0.31 2.5 2.5 0.5
Proteus mirabilis R 10 10 1.25 1.25 5.0 10 4.0
K. pneumoniae ATCC 70603 2.5 2.5 0.16 0.16 2.5 2.5 2.0
Klebsiella pneumoniae S 5 5.0 0.31 0.31 5.0 5.0 0.5
Klebsiella pneumoniae R 2.5 5.0 0.16 0.16 2.5 2.5 4.0
Moyenne générale 5.27 7.17 1.30 1.41 3.95 5.62 10.73
S : sensible aux ATB ; R : résiste aux ATB

Résultats et discussion
97
D’après le tableau 41, on note que les plus faibles CMI et CMB enregistrées sont celles de
l’HE de C. cassia. En effet, celle-ci a montré l’activité la plus forte avec une CMI moyenne ≤ 1,30
mg/ml et une CMB moyenne ≤ 1,41 mg/ml. Mais comparé à l’ATB céfotaxime, ce dernier est actif
à une concentration moyenne basse (10,73μg/ml) que les trois HEs.
De point de vue sensibilité des souches, toutes les souches testées étaient sensibles à l’HE de
C. cassia avec des CMI et des CMB beaucoup plus faibles que celles des HEs de C. sativum et
Z. hispanica. Par contre, Pseudomonas aueruginosa s’est révélée plus résistante aux HEs de
coriandre et de fliou. Les souches E. coli BLSE ont enregistrés des CMI presque dix fois plus
petites que celles de C. sativum et Z. hispanica. Ce résultat confirme le diamètre moyen de la zone
d’inhibition obtenu (33,25 mm).
Les mêmes résultats sont obtenus pour les souches de K. pneumoniae et Proteus mirabilis qui
ont enregistrées des concentrations très faibles (CMI 1.25 mg/ml et CMB 1.25 mg/ml) d’HE de
cassia alors que par la méthode des disques, elles ont donnés des zones d’inhibitions moyennes de
34 et 27 mm respectivement.
Duman et al., (2010), ont montré que l'activité de l'huile essentielle de coriandre était
supérieure à l'activité de son constituant principal, le linalool. Le même résultat a été obtenu par
Inouye et al., (2011) qui ont trouvé que le linalol possède une activité modéré sur plusieurs
bactéries. Silva et al., (2011) ont trouvé que toutes les souches testées ont été inhibées par l'huile de
coriandre avec différents degrés d’inhibition mais P. aeruginosa était la souche la plus résistante
avec CMI (1,6% v/v). Pour E. coli ATCC 25922, ils ont enregistré une CMI= CMB= 0.2%.
Pour l’HE de C. cassia, les travaux de Regiane et al., (2014) ont enregistré une CMI de 88
mg/ml vis–à–vis d’E. coli. Canillac et Mourey, (2001), Friedman et al., (2002), Bruneton,
(2009), Pozzatti et al., (2010) et Unlu et al., (2010) ont démontré que C. cassia, a une activité
bactéricide contre E.coli . Dans une étude réalisée par Hofer et al., (2009) les huiles essentielles
extraites à partir des feuilles de la cannelle de Ceylan (riche en eugénol) ont une activité inhibitrice
et bactéricide sur S. aureus, E. coli et S. enterica. Par contre, Pseudomonas aeruginosa étai
résistante. Ce résultat contraste avec le notre, puisque P. aeruginosa s’est montré légèrement
sensible à l’HE (ϕmoy = 15 mm, CMI et CMB=2,5 à 5 mg/ml). Gill et Holley, (2004 et 2006) ont
montré que l’huile essentielle de C. cassia riche en cinnamaldéhyde (composé majoritaire de l’HE
de cassia) possède une activité antimicrobienne. En effet, ce composé inhibe les ATP synthétases
bactériennes et provoque une diminution de la production d’ATP intracellulaire. Chez E. aerogenes,
le cinnamaldéhyde pourrait interférer avec les mécanismes du quorum-sensing (Niu et al., 2006).
Chez B. cereus, le cinnamaldéhyde provoque un changement de la morphologie cellulaire (Kwon et

Résultats et discussion
98
al., 2003) et agit sur la croissance, l'ultra-structure des hyphes et les facteurs de virulence d'A.
fumigatus, A. niger et T. rubrum. Les membranes cellulaires et certaines structures endo-
membranaires de la cellule fongique sont des sites d'action possibles (Khan et Ahmad, 2011).
Concernant l’HE de Z. hispanica, les études antérieures réalisés par Sezik et al., (1991) ;
Meral et al., (2002 ) ont montré que l’activité antibactérienne est due à des composant majeure et
mineur tel que pulégone, pipéritenone et 1,8-cinéole, donc on peut dire que l’activité
antibactérienne des HEs est due un une synergie entre les composant chimique. Bekhchi et al., en
(2007) n’ont pas montré d’activité antibactérienne intéressante. Cette faible efficacité est due
probablement à la faible activité de la pulégone (composé majoritaire de l’HE) (Arras et Usai,
2001).
Bien que l’HE de cassia est la plus efficace sur les souches testées, du essentiellement à son
composé majoritaire le cinnamaldéhyde (reconnu pour son activité antimicrobienne), il en demeure
que l’effet des composés mineurs n’est pas négligeable pour exhiber une activité antibactérienne
impliqué dans les phénomène de synergie entre les différents constituant qui peuvent être a l’origine
d’une activité antibacterienne beaucoup plus prononcée que celle prévisible par les compose
majoritaire ( Hammer et al., 2003; Burt, 2004).
III.20.3.3. Evaluation de la formation du biofilm in vitro par la microplaque
Les résultats de la formation du biofilm in vitro sont présentés dans le Tableau 48.
Tableau 48 : Evaluation de la formation du biofilm in vitro.
Technique Nombre de souches d’E. coli BLSE
Absence Faible Modéré Fort
Microplaque 0 Ec(6,22 et 24) Ec(1,2,3,5,8,9,11,12,14,15,
16, 18, 21, 23, 25, 26, et 27)
Ec(4,10,19,20,17,28 et 29)
Ec : Ecoli
D’après le tableau ci-dessus, on remarque que la capacité d‘adhésion variait largement d‘une
souche à une autre. Parmi les 27 souches testées, 3 (11.11%) étaient estimées comme faiblement
formatrices de biofilm, 17 (62.96%) comme intermédiaires et 7 (25.92%) sont considérés comme
fortement formatrices de biofilm
On remarque que les souches qui possèdent le gène pap sont plus formatrice de biofilm par
contre les souches qui possèdent le gène sfa sont moyenne ou faiblement formatrice de biofilm

Résultats et discussion
99
Sargol et al., (2015) ont montré que 94% des souches formatrice de biofilm contient le
gène pap. Escherichia coli est l'un des principaux agents responsables des infections des voies
urinaires (UTI) à travers le monde. La capacité de cette bactérie pour former des biofilms sur des
dispositifs médicaux tels que des cathéters joue un rôle important dans le développement des
infections urinaires pour cela on a testé nos huile essentielle sur ces souches
III.20.3.4. Résultats des concentrations minimales inhibitrices et éradicatrices de biofilm
Les résultats de CMIB et CMEB des HEs envers les souches d’infection urinaire à l’état
biofilm sont présentés dans le Tableau 49.
Tableau 49. Effet des HEs étudiées vis–à–vis des souches d'E. coli en état du biofilm, exprimée par
les concentrations minimales inhibitrices (CMIB) et éradicatrices (CMEB) du biofilm en mg/ml.
HE
Souches
Z. hispanica
C. cassia C. sativum
CMIB CMEB CMIB CMEB CMIB CMEB
Escherichia coli ATCC 25922 7.5 15 0.93 0.93 3.75 7.5
Escherichia coli 72 S 3.75 7.5 1.875 3.75 7.5 7.5
Escherichia coli 73 S 7.5 7.5 0.93 0.93 3.75 7.5
Escherichia coli 74 S 3.75 7.5 1.875 3.75 3.75 7.5
Escherichia coli 75 S 7.5 15 1.875 3.75 3.75 7.5
Moyenne E. coli S 5.62 9.37 1.63 3.045 4.68 7.5
Escherichia coli 21 BLSE 60 60 7.5 7.5 30 60
Escherichia coli 17BLSE 30 30 3.75 3.75 15 30
Escherichia coli 05 BLSE 30 30 3.75 3.75 15 15
Escherichia coli 02 BLSE 15 30 1.875 3.75 7.5 7.5
Escherichia coli 09 BLSE 30 30 3.75 3.75 15 30
Escherichia coli 16BLSE 30 30 3.75 3.75 15 30
Escherichia coli 15 BLSE 15 15 3.75 3.75 15 15
Escherichia coli 04 BLSE 30 30 3.75 3.75 15 15
Escherichia coli 06BLSE 15 15 1.875 1.875 7.5 15
Escherichia coli 29 BLSE 30 60 3.75 3.75 15 15
Escherichia coli 20 BLSE 60 60 7.5 7.5 30 30
Escherichia coli 19 BLSE 60 60 7.5 7.5 30 30
Moyenne E.coli BLSE 33.75 37.5 4.37 4.53 17.5 24.37

Résultats et discussion
100
P. aueruginosa ATCC 27853 - - - - - -
Pseudomona aueruginosa S - - - - - -
Pseudomona aueruginosa R - - - - - -
P. mirabilis ATCC 35659 7.5 15 0.46 0.46 7.5 7.5
Proteus mirabilis S 7.5 15 0.46 0.46 3.75 7.5
Proteus mirabilis R 15 15 1.875 1.875 7.5 7.5
K. pneumoniae ATCC 70603 3.75 7.5 0.93 0.93 3.75 7.5
Klebsiella pneumoniae S 7.5 7.5 1.875 1.875 7.5 7.5
Klebsiella pneumoniae R 3.75 7.5 0.93 0.93 3.75 3.75
Moyenne générale 10.20 14.37 1.49 1.67 6.63 8.95
S : sensible aux ATB ; R : résiste aux ATB
D’après les résultats ci–dessous, on remarque bien que les CMIBs (concentration minimale
inhibitrice du biofilm) sont plus ou moins grandes par rapport à celles des CMIs. Comme à l’état
planctonique, l’huile de C. cassia est la plus active vis–à–vis des souches responsables d’infection
urinaire en biofilm. Cette HE peut inhiber la croissance de ces microorganismes sous forme de
biofilms avec des concentrations intéressantes, qui varient entre 0,46 à 7,5 mg/ml. Pour
C. sativum et Z. hispanica, leurs HE sont également actives mais avec des taux supérieurs. Aussi,
on remarque que les souches qui possèdent le gène pap sont plus résistantes que les souches qui
possèdent le gène sfa et que les CMEBs sont légèrement supérieures à celles des CMIBs.
Selon les souches, on constate qu’E.coli BLSE sous forme de biofilms sont plus ou moins
résistantes vis–à–vis des HEs que les souches de référence (sensible aux antibiotiques). Cette légère
résistance des souches cliniques est expliquée par le faite que le phénotype de formation des
biofilms est beaucoup plus répondu chez les souches multi–résistantes que chez les souches
sensibles (Sanchez et al., 2013). D’autre part, il est connu que la matrice extracellulaire du biofilm
entrave l'efficacité de nombreux agents antimicrobiens (Davies, 2003). Par conséquent, on peut dire
que les souches cliniques d’E. coli BLSE en biofilm ont été légèrement résistantes vis–à–vis des
HEs probablement en raison de leur grande capacité de formation des biofilms. Cependant, même si
on a noté une légère résistance des souches cliniques d’E. coli en biofilm envers les HEs étudiées,
ces agents antimicrobiens naturels restent très actifs et intéressants, surtout l’huile de C. cassia.
En comparant nos résultats de l’effet d'inhibiteur des HEs étudiées vis–à–vis des souches en
état de biofilm avec des travaux antérieurs, Andersen , (2001) ont trouvé que le cinnamaldehyde et
l’eugénol (principaux composants de l'HE de la cannelle) sont capables d’inhibés la formation de
biofilm par Escherichia coli ATCC 33456.
Des expériences récentes ont montré que l'effet des deux composés sur la réduction de la
signalisation cellulaire et la formation de biofilm est additif, non synergique. Dans l'ensemble, la

Résultats et discussion
101
présence d'eugénol et de cinnamaldéhyde dans Cinnamomum spp. pourrait faire partie d'une réponse
évolutive pour réduire l'ampleur de la colonisation bactérienne de la phyllosphère ( Niu et Gilbert,
2004).
D’après Bazargani et Rohloff, (2016), l’huile essentielle de coriandre avait une activité
antibiofilm élevée contre un biofilm formé par S. aureus et E. coli avec une CMIB de 0,8 ul/ml et
1,6 ul/ml, respectivement. Alors, que nous avons trouvé avec la même huile une CMIB de 17,5
mg/ml contre E.coli BLSE.
Les biofilms sont responsables d’infections chroniques et posent de nombreux problèmes dans
le domaine médical. Les infections liées à des biofilms touchent majoritairement les personnes
légèrement ou fortement immunodéprimés et impliquent souvent des bactéries commensales
comme Staphylococcus epidermidis, Staphylococcus aureus et Pseudomonas aeruginosa
(Costerton et al., 1999). Elles sont caractérisées par des symptômes qui surgissent de façon
récurrente et contribuent de manière très importante aux infections nosocomiales. En effet, les
biofilms ont la capacité de se développer sur divers instruments médicaux : sondes urinaires,
cathéters veineux, tubes de ventilation artificielle, prothèses orthopédiques. Dans la majorité des
cas, la seule solution efficace est le retrait de l’instrument infecté. D’autre part, la contamination des
systèmes de climatisation, de ventilation et de distribution d’eau par des biofilms abritant des
microorganismes pathogènes, contribue à la propagation des infections en milieux hospitaliers ou
non hospitaliers, mais également dans les environnement agroalimentaires où les biofilms sont une
source importante de nuisance (donlan et costerton, 2002).
Nombreuses études actuelles se focalisent sur les huiles essentielles du fait de leurs effets
bénéfiques dans la prévention des infections urinaires. L’adhérence des bactéries à l’uroépithélium
est la première étape dans la pathogénicité des infections urinaires suivie par la multiplication
bactérienne et la colonisation du tractus urinaire (Lowe et Fagelman, 2001). L’adhérence va
permettre notamment aux Escherichia coli de remonter via l’urètre dans la vessie en évitant leur
élimination par le flux urinaire. Pour permettre cette adhérence, les bactéries ont des prolongements
appelés fimbriae ou adhésines. De nature protéique, ces adhésines sont spécifiques, se liant aux
récepteurs hydrocarbonés correspondants à la surface des cellules uroépithéliales (Beachey, 1981).
III.21. Étude de la combinaison des huiles essentielles avec la céfotaxime
L’étude de la combinaison de l’antibiotique céfotaxime a été réalisée avec deux huiles
essentielles qui sont : Cinnamomum cassia et Coriandrum sativum. Les résultats de la combinaison
sont présentés dans le tableau 50.

Résultats et discussion
102
Tableau 50: CFI des huiles essentielles de Cinnamomum cassia et Coriandrum sativum
Espèces CMI HE
combinée à
l’ATB /CMI de
HE seul
CFI de l’HE CMI ATB
combinée à HE
/CMI d’AB
seul
CFI ATB CFI Interprétation
Coriandrum sativum.
Cinnamomum cassia
1.25/2.5
0.31/0.63
0.5
0.5
0.0625/0.125
0.0625/0.125
0.5
0.5
1
1
Effet d’addition
Effet d’addition
Selon la formule CFI = CFI l’huile + CFI de la cefotaxime, toutes les HEs testées
présentent juste l’effet d’addition, avec des CFI égales à 1. Tandis qu’aucun effet de synergie ou
d’antagonisme envers E. coli n’a été observé.
En comparant nos résultats de la combinaison avec des travaux antérieurs, Yap et al., (2013)
ont montré que Cinnamomum verum présente un effet synergétique avec pipéracilline et
Ampicilline.
Les travaux sur les combinaisons des agents antimicrobiens en clinique ont montré leurs
efficacités en augmentant l’effet antimicrobien et en diminuant la concentration des antibiotiques.
Ces derniers gardent toujours leurs effets inhibiteurs surtout lorsqu’ils possèdent une forte synergie
entre eux (Denes et Hidri, 2009). Plusieurs travaux ont montré la possibilité de l’association des
antibiotiques avec les huiles essentielles dans le but d’augmenter leurs effets a de faibles doses et
ainsi pour diminuer la toxicité de plusieurs antibiotiques (Shin et Kang, 2003 ; Rosato et al.,
2008).
III.22. Cinétique de destruction d’E. coli avec les huiles essentielles
Afin d’évaluer la rapidité de l’activité antibactérienne pour se rapprocher d’un contexte
d’utilisation clinique, nous avons mesuré le taux de réduction de la population bactérienne par les
deux huiles essentielle les plus actifs (Cinnamomum cassia et Coriandrum sativum) contre E. coli
BLSE 09 à la concentration de CMB (figures 25 et 26).
Figure 25 : Cinétique de destruction de la souche Ecoli BLSE 09 exposée à l’huile essentielle de
Cinnamomum cassia pendant 15, 30, 45 et 75 mn à la concentration de CMB= 5 mg/ml.

Résultats et discussion
103
Figure 26 : Cinétique de destruction de la souche Ecoli BLSE 09 exposée à l’huile essentielle de
Coriandrum sativum pendant 15, 30, 45, 75,90,120,150,180 et 240 mn aux concentrations CMB=
10, 20 et 40 mg/ml.
Les résultats des figures 25 et 26 montrent que l’HE de Cinnamomum cassia a détruit toutes
les cellules bactériennes au bout de 75 min, alors qu’il a fallu 240 min pour détruire toutes les
cellules par Coriandrum sativum.
Senhaji et al., (2007), en étudiant la cinétique de destruction de la souche E.coli O157: H7 par
l’huile essentielle de Cinnamomum zeylanicum, ont trouvé que la souche est détruite au bout de 60
min à la concentration de 0.025℅.
Vu les résultats ci-dessus, on peut dire que l’HE de Cinnamomum cassia était la plus active
sur les souches E. coli BLSE avec une CMI= 1,97mg/ml et une CMB= 2,5mg/ml. Alors les HEs de
C. sativum et Z. hispanica étaient moins actives. Cette variabilité d’effet des HEs est due à
plusieurs paramètres dont la composition chimique. En effet, le cinnamaldéhyde, composant
majoritaire de Cinnamomum cassia, est très actif sur les souches alors que la pulégone et linalool
sont reconnu moins actif (Khani et Rahdari, 2012 et Chang et al., 2013).
Nombreux antibiotiques sont toxiques de manière intrinsèque, citons notamment la
néphrotoxicité des aminosides, les troubles de l'hémostase causés par les β-lactamines et les
troubles de la minéralisation dentaire et osseuse provoqués par les tétracyclines. Jamais de tels
effets indésirables n'ont été observés avec l'emploi des HEs non caustiques (Jouault, 2012. C’est
vrai, certaines sont toxiques, et c'est seulement par ignorance des précautions d'emploi les plus
basiques que des accidents surviennent. Mais respectant des conditions d'utilisation bien établies,
comme on le fait déjà pour les antibiotiques, le risque de survenue d'accidents iatrogènes serait
proche de zéro.
Il ne faut jamais utiliser les HEs à l’état pur. Bien que l’HE de cannelle à cinnamaldéhyde est
irritante pour les muqueuses et la peau à faible dose, son utilisation reste. La concentration

Résultats et discussion
104
maximale est de 0.5% et ne doit être utilisée qu'en application externe. Elle est contre indiquée en
cas de gestation (Bruneton, 2009). La toxicité aiguë de l'huile essentielle est très faible (DL50 =
4.16 g/kg chez le rat par voie orale) (Bruneton, 2009). La consommation de cannelle ne présente
pas de risques à la dose journalière acceptable pour l'humain a été fixée à 1.25 mg/kg de
cinnamaldéhyde (Bruneton, 2009).
Ajoutons que de nombreux patients peuvent supporter les effets indésirables constants de la
plupart des antibiotiques : effets digestifs (nausées, maux de ventre, diarrhées), apparition de
mycoses (candidose buccale, intestinale, vaginale). Pour tenter de prévenir ces effets secondaires, il
est fréquent de voir prescrits sur la même ordonnance l'antibiotique associé à un probiotique comme
un extrait de cannelle (HE) car cette association n’entraine pas ces désagréments. En effet les HEs
n'entraînent pas ce genre de désagréments puisque la microflore intestinale de l'être humain est
accoutumée depuis des milliers d'années à être en contact avec ces molécules naturelles, que l'on
trouve pour la plupart dans les épices et les herbes condimentaires. En effet, il a été montré que les
bactéries commensales, celles faisant partie du pool habituellement retrouvé dans l'organisme
humain, exprimaient une résistance aux HE de 8 à 32 fois supérieure par rapport aux bactéries
pathogènes (Hammer et al., 1999).

Conclusion générale

Conclusion générale
105
Conclusion générale
La résistance aux antibiotiques est devenue un problème majeur de santé publique en
Algérie et à travers le monde. En effet, au cours de ces dix dernières années, nous avons
constaté une importante augmentation de la résistance aux antibiotiques en particulier chez les
bacilles à Gram négatif.
La multirésistance des souches d’E.coli BLSE, représente une vraie menace pour la santé
publique, puisqu’elle touche plusieurs molécules allant des β-lactamines, aux aminosides et
aux quinolones. Le risque étant la difficulté de traitement des infections à E. coli produisant
les gènes de résistance en milieu hospitalier.
L’objectif assigné à cette étude était d’évaluer l’effet inhibiteur de trois plantes
aromatiques : Cinnamomum cassia, Coriandrum sativum et Ziziphora hispanica sur
Escherichia coli (BLSE) responsables d’infections urinaires d’origine hospitalière.
D'après les résultats de l'étude rétrospective, 132 malades ayant eu des infections
urinaires (ECBU) ont été recensés durant la période allant de 2011 à 2013. La fréquence de
ces infections urinaires était élevée chez les femmes (33 ,83%) par rapport à celle des
hommes (24 %). Les patients qui ont un âge compris entre 20 et 40 ans ont montré une
incidence d'infection urinaire importante (33,89%). La fréquence des infections urinaires était
élevée dans le service de la médecine interne (33℅) suivi des services de maternité et
d’urologie (28℅ par service).
Parmi les germes isolés, Escherichia coli a été le plus fréquent (63%). On a constaté que
les taux de résistance les plus élevés sont observés avec les pénicillines (amoxicilline
83,13℅, ticarciline 75,9℅ et pipéraciline 61,44℅). L’association des pénicillines avec les
inhibiteurs de -lactamase a permis de récupérer l’activité de ces molécules. En effet, on note
des taux de 62,65℅ pour l’amoxiciline+acide clavulanique, 24,09℅ pour
pipéraciline+tazobactam et 39,75℅ pour ticarciline+acide clavulanique. Pour les
céphalosporines de troisième génération (céfotaxime et ceftazidime) on a noté une résistance
importante (56,62℅). Pour les autres classes d’antibiotiques, les aminosides et les quinolones
ont montré une bonne activité sur E. coli (18℅ à 39℅).
L’analyse des extraits d’ADN d’E. coli BLSE par amplification a révélé la présence des
gène de résistance aux -lactamine bla CTX-15 et TEM-1 avec un pourcentage élevée, des
gènes de résistance aux quinolones et aux aminoside à savoir les gènes aac(6)-lb, qnrB, qnrA,
armA et rmt B, et les gène de virulence l’opéron d’adition pap, et/ou l’opéron sfa et le gène
toxine d’hémolyse cnf1. L’étude ERIC/PCR à montré qu’il y a un polymorphisme de souches

Conclusion générale
106
E. coli BLSE, ce qui pourrait annuler l’hypothèse d’une épidémie ayant pour origine la même
souche clonale.
L’analyse chimique des trois huiles essentielles par CG/SM a révélé que ces huiles sont
chimiquement variées. L’huile de Cinnamomum cassia est riche en phénylpropanoides dont le
cinnamaldéhyde est le composé majoritaire (75,85%), alors que les huiles de Coriandrum
sativum et Ziziphora hispanica sont riches respectivement en linalol (85.57℅) et en pulégone
(88.33℅).
L’activité antimicrobienne des trois huiles essentielles envers E. coli BLSE, a montré
que l’huile de Cinnamomum cassia est la plus active contre ces souches testées à l’état
planctonique et biofilm. A l’état sessile, l’huile a donné un diamètre moyen d’inhibition de
33,25 mm, une CMImoy = 1,97mg/ml et une CMBmoy = 2,5 mg/ml. A l’état de biofilm, elle
a donné une CMIBmoy = 4,37 mg/ml et une CMEBmoy = 4,53 mg/ml. Par contre les huiles
essentielles de Coriandrum sativum et Ziziphora hispanica sont moins actif. Le même
résultat a été obtenu vis-à-vis des souches Klebsiella pneumoniae et Proteus mirabilis où
l’huile de Cinnamomum c’est révélée la plus active.
La combinaison des huiles avec l’antibiotique céfotaxime a montré que l’huile de
Cinnamomum cassia et Coriandrum sativum présente un effet d’addition, alors, que le suivi
de la cinétique de destruction des cellules en fonction du temps, a montré que l’huile
essentielle de Cinnamomum cassia a détruit toutes les cellules bactériennes au bout de 30
min, mais il a fallu 240 min pour détruire toutes les cellules par Coriandrum sativum.
Les résultats obtenus affirment bien le pouvoir antimicrobien des huiles essentielles,
notamment l'HE de cannelle, qui a montré une forte activité in vitro contre les souches E. coli
BLSE d’origine hospitalière à une faible concentration.
L’utilisation de la cannelle comme alicament (utilisation alimentaire et thérapeutique)
constitue un alternatif aux antibiotiques pour lutter contre la flore pathogène (uropathogene)
tout en préservant la flore résidente. Cependant, il serait intéressant de valoriser la cannelle et
autres plantes aromatiques in vivo, d’élargir l’étude sur d’autres souches résistantes à d’autres
antibiotiques, d’étudier l’effet de ces huiles sur les types de β-lactamases et pourquoi pas
élaborer des produits pharmaceutiques à base d’huiles essentielles contre les diverses
infections.

Références

Références Bibliographiques
107
Références bibliographiques :
Abdesselam, Z. (2006). Les huiles essentielles, un pouvoir antimicrobien avéré. Nutra News.,
pp. 6-16.
Adams, R.P.(1995).Identification of Essential Oils Components by Gas
Chromatography/Mass.
AFSSAPS (Agence Française de Sécurité Sanitaire et des Produits de Santé). (2009).
Recommandations De Bonne Pratique. Prévention et Traitement De La Maladie
Thromboembolique Veineuse En Médecine.
AFSSAPS (Agence Française de Sécurité Sanitaire et des Produits de Santé). (2008).
Recommandations de bonne pratique. Diagnostic et antibiothérapie des infections urinaires
bactériennes communautaires chez l‘adulte. [En ligne]. [Consulté en Mars 2010].
http://www.afssaps.fr/Infos-de-securite/Recommandations-de-bonne-pratique.
AFSSAPS (Agence Française de Sécurité Sanitaire et des Produits de Santé). (2010).
Annales du contrôle national de qualité des analyses de biologie médicales.
Ahoyo, A. T., Baba-Moussa, L., Anago, A. E., Avogbe, P., Missihoun, T. D., Loko, F.,
Dramane, K. (2007). Incidence d'infections liées à Escherichia coli producteur de bêta
lactamase à spectre élargi au Centre hospitalier départemental du Zou et Collines au
Bénin. Médecine et maladies infectieuses,37(11), 746-752.
Alebiosu, C. O., Osinupebi, O. A., et Olajubu, F. A. (2003). Significant asymptomatic
bacteriuria among Nigerian type 2 diabetics. Journal of the National Médical
Association, 95(5), 344.
Alexander, T. (1998). Infections urinaires et génitales. Médecine générale. ESTEM; 96:56–
65.collection Medicine General, livre-Fnac.
Ali Khan, D., Hassan, F., Ullah, H., Karim, S., Baseer, A.,Ali, A. M ., Ubaid, M .,Ali
Khan, S., Ghulam, M .(2013).Antibacterial Activity of Phyllantus Emblica, Coriandrum
Sativum, Culinaris Medic, Lawsonia Alba and Cucumis Sativus. Acta Polonaise
Pharmaceutique.
Allain, P. (2008). Bêta-lactamines, pénicillines et céphalosporines.Les medicaments. 3 me
edition.
AL-Saghir, M. G. (2009). Antibacterial assay of Cinnamomum cassia (Nees and Th. Nees)
Nees ex blume bark and Thymus vulgaris L. leaf extracts against five pathogens.Journal of
Biological Sciences, 9(3), 280-2.

Références Bibliographiques
108
Ambrozic, A. J., Keber, R., Zerjavic, K., Orazem, T., Grabnar, M. (2007). Emergence of
the quinolone resistance-mediating gene aac (6')-Ib-cr in extended-spectrum-beta-lactamase-
producing Klebsiella isolates collected in Slovenia between 2000 and 2005.Antimicrobial
agents and chemotherapy, 51(11), 4171-4173.
Andre, M. H., Lortholary, O., Bryskier, A. (1998). Classification des antibiotiques :
relation structure-activité. Encycl Méd Chir (Elsevier, Paris), AKOS Encyclopédie Pratique
de Médecine. 5-0015. P6
Anderson, M. (2001). "Risk assessment and epidemiology of dental caries: review of the
literature." Pediatr Dent 24(5): 377–385.4.
Andremont, A., Pression. (2002). De sélection antibiotiques, flores commensales et
évolutions de la résistance. J Pédiatr Puericulture ; 15(3) : 160-65.
Andreu, A., Xercavins, M., Fernandez, F.(1989).Type 1 fimbriae, P fimbriae and X
adhesins in Escherichia strains causing pyelonephritis, cystitis and recurrent urinary
infections]. Med Clin (Barc). ;92(11):409–12.[PubMed].
Andrews, J.M. (2001). The development of the BSAC standardization method of disc
diffusion testing. JAC. Ed: British society for antimicrobial chemotherapy. (48) S129-42.
Arras, G ., Usai, M. (2001). Fungitoxic activity oftwelve essential oils against four
postharvest Citrus pathogens: Chemical analysis of Thymus capitatus (L.) Hoffing. Oil a dits
effect in subatmospheric pressure conditions. J ofFood Protection, 64: 1025 – 1029.
Austin, D.J., Kristinsson, K.G. et Anderson, R.M.(1999). The relation ship between the
volume of antimicrobial consumption in human communities and the frequency of resistance.
Proc Natl Acad Sci USA.96: 1152-6.
Baba Ahmed Z. B., Arlet ,G. (2014) . News of antibiotic resistance among Gram-negative
bacilli in Algeria. PubMed.
Baba Ahmed Z. B.. Decré, D., Genel, N., Boucherit-Otmani, Z., Arlet, G., Drissi, M.
(2013).Molecular and epidemiological characterization of enterobacterial multi-drug-resistant
strains in Tlemcen Hospital (Algeria) (2008-2010). Microb Drug Resist ; 19:185-90.
Baba Ahmed, Z. B., Ayad, A., Mesli, E., Messai, Y., Bakour, R., & Drissi, M. (2012).
CTX-M-15 extended-spectrum â-lactamases in Enterobacteriaceae in the intensive care unit
of Tlemcen Hospital, Algeria. Eastern Mediterranean health journal, 18(4), 382.
Barrial, K., Scotet, J. (2006). Classification raisonnée des β-lactamases chez les bacilles
Gram négatif. Perspective d'évolution. Tigaud de bactériologie. 3-10.

Références Bibliographiques
109
Barrier, L. C.(2013). Infections Urinaires Chez La Personne Âgée : Difficultés Du
Diagnostic Microbiologique Et Impact De La Prescription Des Ecbu Pour La Prise En Charge
Des Personnes Agées Au Chu D'angers .these de doctorat .universitie angers .
Barka M.S., Benyoub N., Zaoui, H .(2014). Prevalence of urogenital tract infections during
pregnancy in Maghnia hospital (Algeria).
Baster H. C., Buchbauer, G. (2009) Handbook of essential oils: Science, Technologie, and
application. Edit. CRC. Press, 975p.
Baudoux, D., Blanchard, J-M., Malotaux, A.F.(2006). Les cahiers pratiques
d‘aromathérapie selon l‘école française, Soins palliatifs Edition Inspir.
Bazargani, M. M., Rohloff, J. (2016). Antibiofilm activity of essential oils and plant extracts
against Staphylococcus aureus and Escherichia coli biofilms.Food Control, 61, 156-164.
Beachey, E.H.(1981). Bacterial adherence: adhesin-receptor interactions mediating the
attachment of bacteria to mucosal surfaces. J Infect Dis ; 143:325–45.
Bedenić, B., Randegger, C., Stobberingh, E., Hächler, H. (2001). Molecular epidemiology
of extended-spectrum β-lactamases from Klebsiella pneumoniae strains isolated in Zagreb,
Croatia. European Journal of Clinical Microbiology and Infectious Diseases, 20(7), 505-508.
Bekhechi, C., Atik Bekkara, F., Abdelouahid, D. E ., Liu, Kai ., Casanova, J ., Tomi, F
.(2007). Composition and Antibacterial Activity of the Essential Oil of Ziziphora
hispanica (L.) from Algeria . Journal of Essential Oil Bearing Plants.pages 318-323.
Belbel, Z., H. Chettibi, M. Dekhil, A. Ladjama, S. Nedjai, J. M. Rolain. (2014). Outbreak
of an armA Methyltransferase-Producing ST39 Klebsiella pneumoniae Clone in a Pediatric
Algerian Hospital. Microb.Drug Resist.
Belmonte, O., Drouet, D., Alba, J., Moiton, M. P., Kuli, B., Lugagne-Delpon, N., Jaffar-
Bandjee, M. C. (2010). Évolution de la résistance des entérobactéries aux antibiotiques sur
l‘île de la Réunion: émergence des bêta-lactamases à spectre élargi. Pathologie
Biologie, 58(1), 18-24.
Ben Romdhane, F., Bougherra, C.H., Sahnoun O., Loussaief, C.H., Kacem B., Mastouri,
M., Tabka-Stambouli, R., Chakroun, M., Bouzouaia N. (2007). Les bactéries
multirésistantes isoles chez les malades hospitalises dans unservice de maladies infectieuses.
Revue Tunisienne Infectiology. 1:12-15.

Références Bibliographiques
110
Ben-Ami, R., Rodríguez-Baño, J., Arslan, H., Pitout, J. D., Quentin, C., Calbo, E. S.,
Garau, J. (2009). A multinational survey of risk factors for infection with extended-spectrum
β-lactamase-producing Enterobacteriaceae in nonhospitalized patients. Clinical Infectious
Diseases, 49(5), 682-690.
Benbelaid, F. (2015). Effets des huiles essentielles de quelques plantes aromatiques sur
Enterococcus faecalis responsable d‘infections d‘origine dentaire (Doctoral
dissertation).Universitie de Tlemcen.
Bendahou, M., Muselli, A., Grignon-Dubois, M., Benyoucef, M., Desjobert, J. M.,
Bernardini, A. F., Costa, J. (2008). Antimicrobial activity and chemical composition of
Origanum glandulosum Desf. essential oil and extract obtained by microwave extraction:
Comparison with hydrodistillation. Food Chemistry,106(1), 132-139.
Ben-Hamouda, T., Foulon, T., & Ben-Mahrez, K. (2004). Involvement of SHV-12 and
SHV-2a encoding plasmids in outbreaks of extended-spectrum β-lactamase-producing
Klebsiella pneumoniae in a Tunisian neonatal ward.Microbial Drug Resistance, 10(2), 132-
138.
Berçot, B., Poirel, L., Nordmann, P. (2008). Plasmid-mediated 16S rRNA methylases
among extended-spectrum β-lactamase-producing Enterobacteriaceae isolates. Antimicrobial
agents and chemotherapy, 52(12), 4526-4527.
Bergonzelli, G.E., Donnicola, D., Porta, N.,Corthesy-Theulaz, I.E. (2003). Essential oils as
components of a diet-based approach to management of Helicobacter infection. Antimicrob.
Agents Chemother. 47: 3240-46.
Berrazeg, M., Drissi, M., Medjahed, L., & Rolain, J. M. (2013). Hierarchical clustering as
a rapid tool for surveillance of emerging antibiotic-resistance phenotypes in Klebsiella
pneumoniae strains. Journal of medical microbiology,62(6), 864-874.
Bertrand, X., Blasco, G., Belle, E., Boillot, A., Capellier, G.,Talon, D. (2003). Importance
de la transmission croisée dans l‘épidémiologie de Pseudomonas aeruginosa en service de
soins intensifs. In Annales francaises d'anesthesie et de reanimation (Vol. 22, No. 6, pp. 505-
509). Elsevier Masson.
.
Bevilacqua, S.(2011). Évaluation de l‘impact d‘une équipe opérationnelle en infectiologie sur
la consommation et le coût des antibiotiques au CHU de Nancy. (Essai d‘intervention
contrôlé).these de doctorat .universitie de nancy.
Bibbal, D.(2008). Impact des beta_lactamine sur l‘emergence d‘enterobacterie resistantes
dans la flore dégestive chez les porcs : caracterisation et strategie de prévention. thèse de
doctorat .universitie de Toulouse.

Références Bibliographiques
111
Bidet, P., Bonarcorsi, S., Bingen, E.(2012). Facteurs de pathogénicité et physiopathologie
des Escherichia coli extra- intestinaux. Elsevier Masson ; Archives de pédiatrie .19 : S80-S92.
Blanco, M., Blanco, J. E., Rodrıguez, E., Abalia, I., Alonso, M. P., Blanco, J. (1997).
Detection of virulence genes in uropathogenic Escherichia coli by polymerase chain reaction
(PCR): comparison with results obtained using phenotypic methods. Journal of
microbiological methods, 31(1), 37-43.
Blomberg, B., Jureen, R., Manji, K. P., Tamim, B. S., Mwakagile, D. S., Urassa, W. K.,
Langeland, N. (2005). High rate of fatal cases of pediatric septicemia caused by gram-
negative bacteria with extended-spectrum beta-lactamases in Dar es Salaam,
Tanzania. Journal of clinical microbiology, 43(2), 745-749.
Bogaerts, P., Galimand, M., Bauraing, C., Deplano, A., Vanhoof, R., De, M. R.,
Rodriguez-Villalobos, H., Struelens, M., Glupczynski, Y. (2007). Emergence of ArmA and
RmtB aminoglycoside resistance 16S rRNA methylases in Belgium. J. Antimicrob.
Chemother. 59, 459-464.
Bonacorsi, S., Mariani-Kurkdjian, P., Desmarest M., Doit, C.(2014) .Les entérobactéries
productrices de bêtalactamase à spectre étendu en pédiatrie : Elsevier Masson SAS. Archives
de Pédiatrie.21:181-182.
Bonnet, R., Sampaio, J. L. M., Labia, R., De Champs, C., Sirot, D., Chanal, C., & Sirot,
J. (2000).A Novel CTX-M β-Lactamase (CTX-M-8) in Cefotaxime
ResistantEnterobacteriaceae Isolated in Brazil. Antimicrobial agents and
chemotherapy, 44(7), 1936-1942.
Boscia, J. A., Kobasa, W. D., Knight, R. A., Abrutyn, E., Levison, M. E., & Kaye, D.
(1986). Epidemiology of bacteriuria in an elderly ambulatory population. The American
journal of medicine, 80(2), 208-214.
Bouchakour, M., Zerouali, K., Claude, J. D. P. G., Amarouch, H., El Mdaghri, N.,
Courvalin, P., Timinouni, M. (2010). Plasmid-mediated quinolone resistance in expanded
spectrum beta lactamase producing enterobacteriaceae in Morocco. The Journal of Infection
in Developing Countries, 4(12), 779-803.
Boullard, B. (2001). Plantes médicinales du monde: croyances et réalités. De Boeck
Secundair.
Boutet-Dubois, A., Pantel, A., Sotto, A., Philippe Lavigne, J. (2012). Les entérobactéries
productrices de carbapénémases. Lettre d'information du CClin Sud-Est destinée aux Acteurs
de la Lutte contre les Infections Nosocomiales & Associées aux Soins.
Bouzidi, N., Aoun, L., Dekhil, M., Granier, S. A., Poirel, L., Brisabois, A., Millemann,
Y. (2011). Co-occurrence of aminoglycoside resistance gene armA in non-Typhi Salmonella

Références Bibliographiques
112
isolates producing CTX-M-15 in Algeria. Journal of antimicrobial chemotherapy, 66(9),
2180-2181.
Bradford, P. A. (2001). Extended-spectrum β-lactamases in the 21st century:
characterization, epidemiology, and detection of this important resistance threat. Clinical
microbiology reviews, 14(4), 933-951
Bradford, P. A. (1999). Automated thermal cycling is superior to traditional methods for
nucleotide sequencing of blasHv genes. Antimicrob. Agents Chemother. 43: 2960-2963.
Briñas, L., Moreno, M. A., Teshager, T., Zarazaga, M., Sáenz, Y., Porrero, C., ... &
Torres, C. (2003). β-lactamase characterization in Escherichia coli isolates with diminished
susceptibility or resistance to extended-spectrum cephalosporins recovered from sick animals
in Spain. Microbial Drug Resistance, 9(2), 201-209.
Brumfitt, W., Gillespie, W.A.(1991). The mysterious ―urethral syndrome‖. A rapid and
accurate test for bacteriuria would improve its management. BMJ 1991;303:1-2.
Bruneton, J.( 2009). Pharmacognosie, phytochimie, plantes médicinales, 4e édition, Tec &
Doc, Paris, 1269 pp.
Bruyère, F., Cariou, G., Boiteux, J.P., Hoznek, A., Mignard, J.P., Escaravage
,L.,Bernard, L., Sotto, A., Soussy, C.J., Coloby, P., CIAFU.( 2008). Généralités. Progrès
en Urologie, 18 Suppl. 1, S14-S18.
Bryskier, A. (1999). Antibiotiques agents antibactériens et antifongiques. Ed Ellipses.
Burt, S. A. (2004). Essential oils: their antibacterial properties and potentiel applications in
foods-a rewiew. International J offoodMicrobiology, 94: 223 - 253.
Bush, K., Jacoby, G.A, Medeiros, A.A .(1995). A functional classification scheme for
betalactamases and its correlation with molecular structure. Antimicrob. Agents. Chemother.
39:1211–33.
Butreau-Lemaire, M., Botto, H. (1997). Infections urinaires nosocomiales.Progrès en
urologie, 7(4), 674-682.
Canillac, N., Mourey, A. (2001). Antimicrobial activity of the essential oil of Picea
excelsa on Listeria, Staphylococcus aureus and coliform bacteria. Food Microbiol 18:261-
268.
Cardinale, E.,Colbachini,P., Perrier-Gros-Claude,J.D.,Gassama, A.,Aidara-
Kane , A.(2000). Dual emergence in food and humans of a novel multiresistant serotype

Références Bibliographiques
113
of Salmonella in Senegal: Salmonella enterica subsp. enterica serotype 35:c:1,2. J. Clin.
Microbiol. 39:2373-2374. [PMC free article] [PubMed]
Carrer, A., Nordmann, P. (2011). CTX-M-15-producing Klebsiella pneumoniae: a change
in the epidemiology of ESBL. Pathol.Biol.(Paris) 59:e133-e135.
Carter, A. P., Clemons, W. M., Brodersen, D. E., Morgan-Warren, R. J., Wimberly, B.
T., Ramakrishnan, V. (2000). Functional insights from the structure of the 30S ribosomal
subunit and its interactions with antibiotics.Nature, 407(6802), 340-348.
Carson C.F., Riley T.V. (1995). "Antimicrobial activity of the major components of the
essential oil of Melaleuca alternifolia." J. Appl. Bacteriol. 78(3): 264–269.
Casin, I., Collatz, E.(1997). Mécanisme de résistance aux aminosides. Med Thérap. 3 (1) :
86-96.
Cattoir, V. (2004). Efflux-mediated antibiotics resistance in bacteria. Pathol. Biol. 52, 607-
616.
Cavallo, J.D., Fabre, R., Jehl F., Rapp, C., Garrabé, E.(2004). Bêtalactamines. EMC-
Maladies infectieuses. 1 : 129-202.
Chaker, E.K. (2010). Caractérisations chimiques et biologiques d‘extraits de plantes
aromatiques oubliées de Midi-Pyrénées, Thèse de Doctora université de Toulouse,p.70-71.
Champetier de Ribes, G., Ranaivoson, G., Ravaonindrina, N., Rakotonjanabelo, A. L.,
Rasolofonirina, N., Roux, J. F., Yasumoto, T. (1998). Un problème de santé publique
réémergent à Madagascar: les intoxications collectives par consommation d'animaux marins.
Aspects épidémiologiques, cliniques et toxicologiques des épisodes notifiés de janvier 1993 à
janvier 1998. Archives de l'Institut Pasteur de Madagascar, 64(1-2), 71-76.
Chanawong, A., M'Zali, F. H., Heritage, J., Lulitanond, A., Hawkey, P. M. (2001). SHV-
12, SHV-5, SHV-2a and VEB-1 extended-spectrum β-lactamases in Gram-negative bacteria
isolated in a university hospital in Thailand. Journal of Antimicrobial Chemotherapy, 48(6),
839-852.
Chanawong, A., M'Zali, F. H., Heritage, J., Xiong, J. H., Hawkey, P. M. (2002). Three
Cefotaximases, CTX-M-9, CTX-M-13, and CTX-M-14, among Enterobacteriaceae in the
People's Republic of China. Antimicrobial agents and chemotherapy, 46(3), 630-637.
Chang, C. T., Chang, W. L., Hsu, J. C., Shih, Y., Chou, S. T. (2013). Chemical
composition and tyrosinase inhibitory activity of Cinnamomum cassia essential oil. Botanical
Studies, 54(10), 1-7

Références Bibliographiques
114
Chartier, E.(2001). Infections urinaires (Généralités) ; Urologie ; Med-Line ; 2ème édition ;
31-36
Chérif, T., Mabrouka, S ., Decré, D., Boutiba-Ben Boubaker, I.,Arlet, G.( 2009).
Cooccurrence of Multiple AmpC β-Lactamases in Escherichia coli, Klebsiella pneumoniae,
and Proteus mirabilis in Tunisia.antimicrobial agent and chemotherapy 60:44–51.
doi:10.1128/AAC.00828-15.
Christensen, G.D., Simpson, W.A., Younger, J.A., Baddour, L.M., Barrett, F.F., Melton,
D.M. (1985). Adherence of coagulase negative Staphylococci to plastic tissue cultures: a
quantitative model for the adherence of staphylococci to medical devices. Journal of Clinical
Microbiology. 22:996–1006
Clément, N. M. (2012). L‘entérotoxine STb d‘Escherichia coli affecte les jonctions serrées
des cellules intestinales épithéliales. Mémoire présenté à la Faculté de médecine vétérinaire en
vue de l‘obtention du grade de maître ès sciences (M. Sc.) en sciences vétérinaires option
microbiologie. Université de Montréal.
Clere, N. (2012). Comment venir à bout des infections urinaires. Actualités
Pharmaceutiques, 51(516), 33-34.
Clermont, O., Dhanji, H., Upton, M., Gibreel, T., Fox, A., Boyd, D.,Frank, T. (2009).
Rapid detection of the O25b-ST131 clone of Escherichia coli encompassing the CTX-M-15-
producing strains. Journal of Antimicrobial Chemotherapy, 64(2), 274-277.
CLIN (Comité de Lutte contre les Infections Nosocomiales.(1998). Hygiène dans les
établissements de Rééducation Fonctionnelle (Soins de Suite et de Réadaptation).
CLSI (Clinical and Laboratory Standards Institute. Performance Standards for
Antimicrobial Susceptibility Testing; Eighteenth Informational Supplement. (2008).
document M100-S18 (ISBN 1- 56238-653-0). Clinical and Laboratory Standards Institute,
940 West Valley Road, Suite 1400, Wayne, Pennsylvania 19087-1898 USA, 2008.
Collignon, A. (2002). Nutrition, Croissance bactérienne . Cours de bactériologie fichier pdf. :
2, 3, 8.
Comes, Jean-François.(2011). Épidémiologie bactérienne des infections cystites non
compliquées en Lorraine.- 93 p. Th : Méd. : Nancy .
Conner, D.,Beuchat, L. R. (1984). Effects of essential oils from plants on growth of food
spoilage yeasts. Journal of food science, 49(2), 429-434.

Références Bibliographiques
115
Cooper, K. E., Woodman, D. (1946). The diffusion of antiseptics through agar gels, with
special reference to the agar cup assay method of estimating the activity of penicillin. The
Journal of pathology and bacteriology, 58(1), 75-84
Corroyer-Simovic B.,Faure, K. (2011). Prévention et prise en charge des infections urinaires
en EHPAD. Risque infectieux en EPHAD
Corvec, S., Poirel, L., Naas, T., Drugeon, H., Nordmann, P. (2007). Genetics and
expression of the carbapenem-hydrolyzing oxacillinase gene blaOXA-23 in Acinetobacter
baumannii. Antimicrobial agents and chemotherapy, 51(4), 1530-1533.
Cosentino, S., Tuberoso, C. I. G., Pisano, B., Satta, M. L., Mascia, V., Arzedi, E.,
Palmas, F. (1999). In‐vitro antimicrobial activity and chemical composition of Sardinian
thymus essential oils. Letters in applied microbiology,29(2), 130-135.
Costerton ,J.W., Stewart, P.S., Greenberg, E.P. (1999) .Bacterial biofilms: a common
cause of persistent infections.Science. PubMed.;284:1318–22.
Coumbes, A.(1995). Antibiotiques en Pharmacologie 1. Paris. Edition concours médical
,153-156 18.
Craker, L.E., Simon, J.E., A, Jatisatienr., Lewinsohn, E .(2004). Composition of
Coriander Essential Oil from Brazil. Future for Medicinal and Aromatic Plants.
Crété, P. (1965). Precis de botanique. v. 2: Systematique des angiospermes.
Cristani, M., D'Arrigo, M., Mandalari, G., Castelli, F., Sarpietro, M. G., Micieli, D.,
Trombetta, D. (2007). Interaction of four monoterpenes contained in essential oils with
model membranes: implications for their antibacterial activity. Journal of Agricultural and
Food Chemistry, 55(15), 6300-6308.
CTIN, S. (1998). Isolement septique. Recommandations pour les établissements de soins.
Ministère de l’Emploi et de la Solidarité, 12-15.
Dalhoff, A.(2001). Comparative in vitro and in vivo activity of the C-8 methoxy quinolone
moxifloxacin and the C-8 chlorine quinolone BAY y 3118. Clin Infect Dis,; 32(Suppl 1):S16-
22.
Darbas, H., Marchandin, H., Bourgeois, N., Michaux- Charachon, S., (2007). Diagnostic
et suivi des infections urinaires: le bon usage de l'examen cytobactériologique des urines.
Faculté de Médecine Montpelier- Nîmes. France.

Références Bibliographiques
116
Davies, D. (2003). Understanding biofilm resistance to antibacterial agents.Nat. Rev. Drug.
Discov. 2(2): 114–122.
Davies, B. I. (1977). Biochemical typing of urinary Escherichia coli strains by means of the
API20E Enterobacteriaceae system. J. Med. Microbiol. 10:293-298.
De Billerbeck, V. G., Roques, C., Vanière, P., Marquier, P. (2002). Activité
antibactérienne et antifongique de produits à base d'huiles essentielles.Hygiènes, 10(3), 248-
251.
De Mouy, D., Cavallo, J.D., Armengaud, M., Arzouni, J.P., Berges, J.L., Bouilloux, J.P.,
Charbit, N., Cirioni, N., Fabre, R., Garrabe, E., Galinier, J., Gayon, A., Grobost, F.,
Larribet, G., Lepargneur, J.P.(1999). Urinary tract infection in an urban population:
etiology and antibiotic sensitivity as a function of patient history. Presse Med; 28: 1624-8.
Degryse, A., Delplala, I.,Voinier, M.(2008).Risque Et Bénéfices Possibles Huiles
Essentielles. École Des Hautes Etudes En Santé Publique, P. 2-44.
Denes, É., Hidri, N. (2009). Synergie et antagonisme en antibiothérapie. Antibiotiques. 11,
106-115.
Dhawan, B., Bonnet, R., Shukla, N. K., Mathur, P., Das, B. K., Kapil, A. (2003). Infection
with an extended-spectrum β-lactamase-producing strain of Serratia marcescens following
tongue reconstruction. Journal of clinical microbiology, 41(5), 2233-2234.
Dibong, S. D., Mpondo, E. M., Ngoye, A., Kwin, M. F. (2011). Plantes médicinales utilisées
par les populations bassa de la région de Douala au Cameroun. International Journal of
Biological and Chemical Sciences, 5(3).
Didry, N., Dubreuil, L., Pinkas, M. (1994). Activity of thymol, carvacrol, cinnamaldehyde
and eugenol on oral bacteria. Pharmaceutical Acta Helvetiae, 69(1), 25-28.
Doi, Y., Yokoyama, K., Yamane, K., Wachino, J., Shibata, N., Yagi, T., Shibayama, K.,
Kato, H., Arakawa, Y .(2004).Plasmid-mediated 16S rRNA methylase in Serratia
marcescens conferring high-level resistance to aminoglycosides. Antimicrob Agents
Chemother 48: 491–496 [PMC free article] [PubMed]
Dong, Y., Xu, C., Zhao, X., Domagala, J., Drlica, K.(1998). Fluoroquinolone action
against mycobacteria: effects of C-8 substituents on growth, survival, and resistance.
Antimicrob Agents Chemother, 1998;42:2978-84.
Donlan, R. M., Costerton, J. W. (2002). Biofilms: survival mechanisms of clinically
relevant microorganisms. Clinical microbiology reviews, 15(2), 167-193.

Références Bibliographiques
117
Donnenberg, M. S., Whittam, T. S. (2001). Pathogenesis and evolution of virulence in
enteropathogenic and enterohemorrhagic Escherichia coli. J. Clin. Inv 107:539-548.
Dorosz, P.H., Vital Durand D., Le Jeunne C.(2011). Guide pratique des médicaments.
30eme éd. ;Maloine, Paris; p1892.
Doublet, B., Butaye, P., Imberechts, H., Chaslus-Dancla, E., Cloeckaert, A. (2004).
Salmonella Agona harboring genomic Island 1-A. Emerg. Infect. Dis. 10 (4): 756-8
Drame, B.G.(1999). Phénotypes de résistance des souches bactériennes isolées au CHU A.
LE DANTEC Thèse pharm., Dakar, n°5.
Duman, A. D., Telci, I., Dayisoylu, K. S., Digrak, M., Demirtas, I., Alma, M. H. (2010).
Evaluation of bioactivity of linalool-rich essential oils from Ocimum basilucum and
Coriandrum sativum varieties. Nat Prod Commun 5, 969–974.
Ebertin, T. (1997). Les infections microbiennes. eds Nathan Université. Coll. Sciences 1997,
128.
ECDC. Transatlantic Taskforce on Antimicrobial Resistance. Recommendations for future
collaboration between the U.S. and the EU. 2011. Disponible à :
http://www.ecdc.europa.eu/en/activities/diseaseprogrammes/TATFAR/Documents/210911_
TATFAR_Report.pdf [Consulté le 20/07/2013]
Edler, L. (2001).Biometry - The Role of the Biostatistician. Introduction to Clinical Drug
Research. Vienna School of Clinical Drug Research. 22 – 26.
Edqm .(2004). European Pharmacopoeia 5th Ed. Main Volume 5.0, 2005 with
Su... by Council of Europe.
Eilenberg, E.(2005). Analyse terminologiques des définitions données à l'infection
nosocomiale et proposition d'une définition. Rev Méd Inter , 26 ,572-577 .
Eisner, A., Fagan, E. J., Feierl, G., Kessler, H. H., Marth, E., Livermore, D.
M.,Woodford N. (2006). Emergence of Enterobacteriaceae isolates producing CTX-M
extended spectrum beta-lactamase in Austria. Antimicrob Agents Chemother.50:785-7.
El Bakkouri, J., Belabbes, H., Zerouali, K., Belaiche, A., Messaouidi, D., Gros, Claude,
JD., El Mdaghri, N. (2009).Résistance aux Antibiotiques d‘Escherichia coli Uropathogène
communautaire et consommation d‘Antibiotiques à Casablanca (Maroc). European Journal of
Scientific Research , 36,1 : 49-55.

Références Bibliographiques
118
EL Brahmi, R.(2013). Profil Epidemiologique Et De Resistance Des Bacteries Multi
Resistantes Au Chu Hassan Ii De Fès Universite Sidi Mohammed Ben Abdellah Pour
L'obtention Du Doctorat En Medecine
El Hadri, S., Chahbi, Z., Baizri, H. (2014). Faut-il toujours croire ce qu‘on voit à l‘IRM
hypothalamo-hypophysaire?. In Annales d'Endocrinologie(Vol. 75, No. 5, pp. 448-449).
Elsevier Masson.
Elqaj, M., Ahami, A., Belghyti, D. (2007). La phytothérapie comme alternative à la
résistance des parasites intestinaux aux antiparasitaires. Journée scientifique "ressources
naturelles et antibiotiques". Maroc.science and education.
Epok, J .E. C.(1999). L‘infection urinaire à Bamako, aspect épidémiologique et étiologique
Thèse, Pharm, Bamako,these de doctorat. Faculté de Médecine de Pharmacie et
d‘Odontostomatologie mali.
Escherich, T. (1885). Die Darmbacterien des Neugeborenen und Saglings. Fortschr. Med.
3:515-522. 20.
Escobedo-Martínez, C., Cruz-Morales, S., Fragoso-Serrano, M., Rahman, M. M.,
Gibbons, S.,Pereda-Miranda, R. (2010). Characterization of a xylose containing
oligosaccharide, an inhibitor of multidrug resistance in Staphylococcus aureus, from Ipomoea
pes-caprae. Phytochemistry, 71(14), 1796-1801.
Euzéby, J.P. (2008). Résistance bactérienne aux antibiotiques, Abrégé de bactériologie
générale et médicale. (pt9) :2615-22.
Eveillard, M., Biendo, M., Canarelli, B., Daoudi, F., Laurans, G., Rousseau, F. Al.(2000).
Diffusion des entérobactéries productrices de bêta-lactamase à spectre élargi et évolution de
leur incidence sur une période de 16 mois dans un centre hospitalier universitaire. Pathol
Biol.49 : 515-21.
Extremina, C. I., Da Fonseca, A. F., Granja, P. L., Fonseca, A. P. (2010). Anti-adhesion
and antiproliferative cellulose triacetate membrane for prevention of biomaterial-centred
infections associated with Staphylococcus epidermidis.International journal of antimicrobial
agents, 35(2), 164-168.
Eymard, S. (2003). Mise en évidence et suivi de l'oxydation des lipides au cours de la
conservation et de la transformation du chinchard (Trachurus trachurus): choix des
procédés (Doctoral dissertation, Université de Nantes).
Ezaitouni, F., Rhou, H., Benamar, L., Ouzeddoun, N., Bayahya, R., Balafrej L.( 1999).
Rein et aminosides. Médecine du Maghreb.77 :1.

Références Bibliographiques
119
Farnsworth, N. R., Akerele, O., Bingel, A. S., Soejarto, D. D. Guo, Z.( 1986). Places des
plantes médicinales dans la thérapeutique. Bulletin de l'organisation mondiale de la
santé.64(2) : 159-164.
Faucher, N., Cudennec, T.(2002). Les infections urinaires bactériennes. Hôpital Sainte
Périne, Paris.
Fauchere, J.L. (1997). Bactério fiches - Techniques en bactériologie clinique ; Ellipses –
Paris.
Fauchère, J. L., Avril, J. L. (2002). Bactériologie générale et médicale. Ellipses.
Ferres, H., Nunn, B. (1983). Penicillin metabolites and platelet function. Lancet 2, 226– 227.
Ferron, A.(1989). Bactériologie médicale à l‘usage des étudiants en médecine. La Madeleine
: Crouans et Roques, 375p.
Ferron, A.(1994). Bactériologie médicale. La Madeleine : Crouan et Roque ; 377p.
Figueiredo, S.(2012). Acinetobacter spp. et reservoir de genes de carbapenemases. Medecine
humaine et pathologie. Universit´e Paris Sud – Paris.
Fihman, V., Lartigue MF., Jacquier H., Meunier F., Schnepf N., Raskine L., Riahi
J., Sanson-le Pors MJ., Berçot, B. (2008).Appearance of aac (6')-Ib-cr gene among
extended-spectrum β-lactamase-producing Enterobacteriaceae in a French hospital. J Infect .
56: 454-9.
Farshad, S., Emamghorashi, F. (2009). The prevalence of virulence genes of E. coli strains
isolated from children with urinary tract infection. Saudi Journal of Kidney Diseases and
Transplantation, 20(4), 613.
Flaudrois, J. P.(2004). Bactério Géné/ croissance bactérienne Cours de Bactériologie
Médicale DCEM1 UFR Médecine Lyon Sud- Laboratoire de Biométrie, Biologie Evolutive
UMR 5558. : 1, 3, 10.
Flandrois, J.P.(1997). Bactériologie médicale, collection Azay, Presses universitaires de
Lyon, 309 pages
Foxman, B. (2002). Epidemiology of urinary tract infections: incidence, morbidity, and
economic costs. The American journal of medicine, 113(1), 5-13.
Frank, T., Arlet, G., Gautier, V., Talarmin, A., Bercion, R. (2006). Extended-spectrum
beta-lactamase-producing Enterobacteriaceae, Central African Republic. Emerg Infect
Dis, 12(5), 863-865.. Ben-Hamouda, T., Foulon, T., & Ben-Mahrez, K. (2004). Involvement
of SHV-12 and SHV-2a encoding plasmids in outbreaks of extended-spectrum β-lactamase-

Références Bibliographiques
120
producing Klebsiella pneumoniae in a Tunisian neonatal ward.Microbial Drug
Resistance, 10(2), 132-138.
Friedman, M., Henika, P., Mandrell, R., Robert, E .(2002).Bactericidal Activities of Plant
Essential Oils and Some of Their Isolated Constituents against Campylobacter
jejuni, Escherichia coli, Listeria monocytogenes, and Salmonella enterica Journal of Food
Protection®, Number 10. pp. 1527-1673, pp. 1545-1560(16).
Funatsu, G., Wittmann, H. G. (1972). Ribosomal proteins: XXXIII. Location of amino-acid
replacements in protein S12 isolated from Escherichia coli mutants resistant to
streptomycin. Journal of molecular biology, 68(3), 547-550.
Furletti, V. F., Teixeira, I. P., Obando-Pereda, G., Mardegan, R. C., Sartoratto, A.,
Figueira, G. M.,Höfling, J. F. (2011). Action of Coriandrum sativum L. essential oil upon
oral Candida albicans biofilm formation. Evidence-Based Complementary and Alternative
Medicine.
Gagliotti, C., Balode, A., Baquero, F., Degener, J., Grundmann, H., Gür, D., Rossolini,
G. M. (2011). EARS-Net Participants (Disease Specific Contact Points for AMR).
Escherichia coli and Staphylococcus aureus: bad news and good news from the European
Antimicrobial Resistance Surveillance Network (EARS-Net, formerly EARSS), 2002 to
2009. Euro Surveill, 16(11), 19819.
Galimand, M. , Courvalin, P.,Lambert, T.(2003). Plasmid-Mediated High-Level
Resistance to Aminoglycosides in Enterobacteriaceae Due to 16S rRNA Methylation.
Antimicrob. Agents Chemother.
Gangoue Pieboji, J. (2007). Caractérisation des -lactamases et leur inhibition par les
extraits des plantes médicinales. Centre d'ingénierie des protéines. Université de Liége. Thèse
de doctorat. p 1-27.
Gangoué-Piéboji, J. (2000). Résistance des bacilles a gram-négatif aux antibiotiques:
prévalence et caractérisation des âlactamases a spectre élargi a l’hopital Central de
Yaoundé (Doctoral dissertation, Thèse de Doctorat de 3e cycle, Université de Yaoundé I,
2000: P 210).
Ganiere, J.P., Mangion C., Peridy M.(2004). Détermination des Concentrations Minimales
Inhibitrices et Bactéricides de la cefquinome, la marbofloxacine, la tylosine et la spiramycine
en solution dans du lait vis-à-vis de bactéries isolées de mammites bovines. Revue Médecine
vétirinaire.155 (8): 411-416.
Ganou, L. (1993). Contribution a l‘étude des mécanismes fondamentaux de
l‘hydrodistillation des huiles essentielle. Thèse de l‘INP Toulouse, France.
Garibaldi, R.A., Burke J.P. , Dickman, M.L., Smith, C.B.(1974). Factors predisposing to
bacteriura during indwelling urethral catheterization . N Engl. J . Med.:215-219

Références Bibliographiques
121
Gaudy, C., Buxeraud, J.(2005). Antibiotiques: pharmacologie et thérapeutique, Paris,
Elsevier.
Gazouli, M., Tzelepi, E., Sidorenko, S. V., Tzouvelekis L. S. (1998). Sequence of the gene
encoding a plasmid-mediated cefotaxime-hydrolyzing class A B-lactamase (CTX-M-4):
involvement of serine 237 in cephalosporin hydrolysis. Antimicrob Agents Chemother. 42:
1259-1262.
Géraldine, J.(2010)..Emergence des entérobactéries sécrétrices de β- lactamases à spectre
élargi. Communication de G. Arlet, lors du 38e Colloque national des biologistes des hôpitaux
.Muller M.P. Résistance des bacilles Gram négatif aux β-lactamases.Cattoir V. Les nouvelles
BLSE.
Geng, S., Cui, Z., Huang, X., Chen, Y., Xu, D., Xiong, P. (2011). Variations in essential oil
yield and composition during Cinnamomum cassia bark growth.Industrial Crops and
Products, 33(1), 248-252.
Geser, N., Stephan, R., Hächler, H. (2012). Occurrence and characteristics of extended-
spectrum β-lactamase (ESBL) producing Enterobacteriaceae in food producing animals,
minced meat and raw milk. BMC veterinary research,8(1), 1.
Gharout-Sait, A., Touati A., Benallaoua, S., Guillard, T., Brasme, L., Madoux, J., de
Champs, C. (2012). CTX-M from community-acquired urinary tract infections in Algeria.
African Journal of Microbiology Research 6, 5306-5313.
Gniadkowski, M., Schneider, I., Palucha, A., Jungwirth, R., Mikiewicz, B., Bauernfeind,
A.( 1998). Cefotaxime-resistant Enterobacteriaceae isolates from a hospital in Warsaw,
Poland: Identification of a new CTX-M-3 cefotaxime-hydrolyzing B-lactamase that is closely
related to the CTX-M- 1/MEN-1 enzyme. Antimicrob Agents Chemother. 42: 827-832.
Gonthier, R., Heritier, P.h. (1988). Néphro-Urologie. Editions Collection Prép‘internat,
Lyon, 285 p.
Ghourri, M., Zidane, L., Douira, A. (2014). La phytothérapie et les infections urinaires (La
pyélonéphrite et la cystite) au Sahara Marocain (Tan-Tan). Journal of Animal &Plant
Sciences, 20(3), 3171-3193.
Gill, AO ., Holley, R.A .(2004) .Mechanisms of bactericidal action of cinnamaldehyde
against Listeria monocytogenes and of eugenol against L. monocytogenes and Lactobacillus
sakei. Appl. Environ. Microbiol. 70: 5750-5755 .

Références Bibliographiques
122
Gill, A.O ., Holley, R.A. (2006). Inhibition of membrane bound ATPases of Escherichia coli
and Listeria monocytogenes by plant oil aromatics. Int. J. Food Microbiol. 111: 170-174.
Gonullu, N., Aktas, Z., Kayacan, C. B., Salcioglu, M., Carattoli, A., Yong, D. E., Walsh,
T. R. (2008). Dissemination of CTX-M-15 β-lactamase genes carried on Inc FI and FII
plasmids among clinical isolates of Escherichia coli in a university hospital in Istanbul,
Turkey. Journal of clinical microbiology, 46(3), 1110-1112.
Goossens, H., Grabein,B. (2005).Prevalence and antimicrobial susceptibility data for
extended-spectrum beta-lactamase- and AmpC-producing Enterobacteriaceae from the
MYSTIC Program in Europe and the United States (1997-2004). Diagn Microbiol Infect Dis.
53:257-64
Guessennd, N., Bremont, S., Gbonon, V., Kacou-NDouba, A., Ekaza, E., Lambert, T.,
Dosso M., Courvalin, P. (2008). Résistance aux quinolones de type qnr chez les
entérobactéries productrices de bêta-lactamases à spectre élargi à Abidjan en Côte
d‘Ivoire. Pathologie Biologie, 56(7), 439-446.
Gueye, O.(2007). Utilisation des méthodes biométriques dans l‘identificationde quelques
bacilles Gram négative Thèse pharm. Dakar N° 36 .
Gutmann, A., Ben‐Porath, S. (1987). Democratic education. John Wiley & Sons, Ltd.
Häcker, U., Nystedt, S., Barmchi, M. P., Horn, C., Wimmer, E. A. (2003). piggyBac-
based insertional mutagenesis in the presence of stably integrated P elements in
Drosophila. Proceedings of the National Academy of Sciences,100(13), 7720-7725.
Hammer, K .A., Dry, L., Johnson M., Michalak E.M.,Carson, C.F., Riley, T.V. (2003)
Susceptibility of oral bacteria to Melaleuca alternifolia (tea tree) oil in vitro. Oral. Microbiol.
Immunol. 18: 389-392.
Hammer, K. A., Carson, C. F., Riley, T. V. (1999). Antimicrobial activity of essential oils
and other plant extracts. Journal of applied microbiology, 86(6), 985-990.
Hamoudi, R. (2008).Contribution à la mis en évidence de principes actifs de
plantes Teuriumpoliumgeryriiprovonant de la région Tamanrasset.Magister universite Kasdi
MerbahOuargla, P. 15-43.
Haraj, N., Elaziz, S., Chadli.(2012).prevalance del‘infections urinaire chey la femme CHU
de sasblonc.
Hassaine, H.,Larbi, M., Maamar, H.(2002). Essai de dépistage des infections urinaires
d'origine hospitalières dans deux centres hospitaliers en Algérie: Hôpital d'Ain-Témouchent et
Maghnia.

Références Bibliographiques
123
Hedi, M. (2007). Mecanisme De Resistance Aux Antibiotiques, MCU-PH, Service
DeBactériologie-Hygiène, CHU Amiens
Helander, I. M., Alakomi, H. L., Latva-Kala, K., Mattila-Sandholm, T., Pol, I., Smid, E.
J.,VON Wright, A. (1998). Characterization of the action of selected essential oil
components on Gram-negative bacteria. Journal of agricultural and food chemistry, 46(9),
3590-3595.
Hillier, S., Roberts, Z., Dunstan, F., Butler, C., Howard, A., Palmer, S.(2007). Prior
antibiotics and risk of antibiotic-resistant community-acquired urinary tract infection: a case-
control study. J Antimicrob Chemother; 60: 92-9.
Hofer, S. B., Mrsic-Flogel, T. D., Bonhoeffer, T., Hubener, M. (2009). Experience leaves a
lasting structural trace in cortical circuits. Nature .PubMed.457 313–317
10.1038/nature07487.
Holley, R. A., Patel D. (2005). Improvement in shelf-life and safety of perishable compounds
from leaf essential oil and smoke antimicrobials. Food Microbiol. 22 ; 273-292.
Hopper DC.(2003). Mechanisms the resistance of of quinolone resistance. . Quinolone
antimicrobial agents 3 ed. ASM Press, Washington DC,; p. 41-67.
Howard, S. C., Budovskaya, Y. V., Chang, Y. W., & Herman, P. K. (2002). The C-
terminal domain of the largest subunit of RNA polymerase II is required for stationary phase
entry and functionally interacts with the Ras/PKA signaling pathway. Journal of Biological
Chemistry, 277(22), 19488-19497.
Huang, G., Jiang, J., Dai, D. (2008). Antioxidative and antibacterial activity of the methanol
extract of Artemisia anomala S. Moore. African Journal of Biotechnol.7 (9): 1335-1338.
Huletsky, A., Knox, J. R., Levesque, R. C.( 1993). Role of Ser-238 and Lys-240 in the
hydrolysis of 3rd
generation cephalosporin's by SI-IV-type B-lactamases probed by site-
directed mutagenesis and 3-dimensional modeling. J. Biol. Chem. 268: 3690-3697.
Iabadene, H., Messai, Y., Ammari, H., Alouache, S., Verdet, C., Bakour, R., Arlet, G.
(2009). Prevalence of plasmid-mediated AmpC β-lactamases among Enterobacteriaceae in
Algiers hospitals. International journal of antimicrobial agents, 34(4), 340-342.
Iabadene, H., Messai, Y., Ammari, H., Ramdani-Bouguessa, N., Lounes, S., Bakour, R.,
and Arlet, G.(2008). Dissemination of ESBL and Qnr determinants in Enterobacter cloacae
in Algeria. J. Antimicrob. Chemother. 62, 133-136.

Références Bibliographiques
124
Inouye, S., Yamaguchi H., Takizawa, T. (2011). Screening of the antibacterial effects of a
variety of essential oils on respiratory tract pathogens, using a modified dilution assay
method. J Infect Chemother. Pubmed. Intermediates, 1-13.
Ishino, K., Ishikawa, J., Ikeda, Y., Hotta, K. (2004). Characterization of a bifunctional
aminoglycoside-modifying enzyme with novel substrate specificity and its gene from a
clinical isolate of methicillin-resistant Staphylococcus aureus with high arbekacin
resistance. The Journal of antibiotics, 57(10), 679-686.
Issa Maigardie, B. (2006). Prévalence des infections nosocomiales au centre hospitalier
universitaire du Point G. Thèse de doctorat en Pharm, Bamako.
Jacob, M., Pellecuer, J.,Tomei, R. (1979). Centre régional d‘étude et de développement des
plantes à usage pharmaceutique. Rivista Italiana EPPOS,11, 26-30.
Jacoby, G. A., Sutton, L. (1991). Properties of plasmids responsible for production of
extendedspectrum B-lactamases. Antimicrob. Agents Chemother. 35: 164-169.
Jean, C. N. Lambert T. (2012). Interprétation phénotypique de l'antibiogramme vis-à- vis
des aminosides. Rev francophone des laboratoires Vol. 42 - N° 445. p : 75-77.
Jehl, F., Chomarat, M., Gerard, A.(2003). De l'antibiogramme à la prescription, 2ème
édition, Marcy-L‘étoile, Biomérieux.
Johanson, W. G., PIERCE, A. K., Sanford, J. P., Thomas, G. D. (1972). Nosocomial
respiratory infections with gram-negative bacilli: the significance of colonization of the
respiratory tract. Annals of Internal Medicine, 77(5), 701-706.
Jouault, S. (2012). La qualité des huiles essentielles et son influence sur leur efficacité et sur
leur toxicité. Université de Lorraine, Nancy.
Jürgen, R., Paul, S., Ulrike S.,Reinhard S. (2009). Essential Oils of Aromatic Plants with
Antibacterial, Antifungal, Antiviral, and Cytotoxic PropertiesŔ an Overview: Forsch
Komplementmed.16: 79Ŕ90.
Kalemba, D., Kunicka, A. (2003). Antibacterial and antifungal properties of essential
oils. Current medicinal chemistry, 10(10), 813-829.
Kaper, J. B., Nataro, J. P., Mobley, H. L. T.( 2004). Pathogenic Escherichia coli. Nat. Rev.
Microbiol. 2: 123-140.
Karim, A., Poirel, L., Nagarajan, S., Nordman, P.(2001). Plasmid-mediated extended-
spectrum beta-lactamase (CTX-M-3 like) from India and gene association with insertion
sequence ISEcp1. FEMS Microbiol Lett ; 201 : 237-41.

Références Bibliographiques
125
Kashef N., Esmaeeli Djavid G., Shahbazi S.(2010). Antimicrobial susceptibility patterns of
community-acquired uropathogens in Tehran, Iran. J Infect Dev Ctries 2010; 2010; 4(4):202-
206.
Kato, T., Lijima, H., Ishihara, K., Kaneto,T., Hirai, K., Naito, Y.Okuda, K. (1990).
Antibacterial effect listerine on oral bacteria. Bull. Tokyo. Dent. coll. 31(4): 301-307.
Kaye, K. S., Gold, H. S., Schwaber, M. J., Venkataraman, L., Qi, Y., De Girolami, P. C.,
Tenover, F. C. (2004). Variety of β-lactamases produced by amoxicillin-clavulanate-resistant
Escherichia coli isolated in the northeastern United States. Antimicrobial agents and
chemotherapy, 48(5), 1520-1525.
Kayima, J. K., Otieno, L. S., Twahir, A., Njenga, E. (1996). Asymptomatic bacteriuria
among diabetics attending Kenyatta National Hospital. East African medical journal, 73(8),
524-526.
Keita, A.(1999). Résistance aux antibiotiques des bactéries isolées chez les malades en
consultation externe au service de bactériologie à l‘I.N.R.S.P. (Institut national de recherche
en Santé publique).
Kempf I, Zeitouni S.(2009) .Cout biologique de la résistance aux antibiotiques : Analyse et
conséquences. PATBIO. Oct;2852:6.
Kermas, R., Touati, A., Brasme, L.(2012). Le Magrex-Debar E, Mehrane S, Weill FX, et al.
Characterization of extended-spectrum beta-lactamase-producing Salmonella enterica
serotype Brunei and Heidelberg at the Hussein Dey hospital in Algiers (Algeria). Foodborne
Pathog Dis 2012; 9: 803-8.
Khan, M. S. A., Ahmad, I. (2012). Biofilm inhibition by Cymbopogon citratus and
Syzygium aromaticum essential oils in the strains of Candida albicans.Journal of
Ethnopharmacology, 140(2), 416-423.
Khan, M. S., Ahmad, I. (2011). In vitro antifungal, anti-elastase and anti-keratinase activity
of essential oils of Cinnamomum-, Syzygium- and Cymbopogon-species against Aspergillus
fumigatus and Trichophyton rubrum. Phytomedicine19, 48-55.
Khani, A., Rahdari, T. (2012). Chemical composition and insecticidal activity of essential
oil from Coriandrum sativum seeds against Tribolium confusum and Callosobruchus
maculatus. ISRN pharmaceutics, 2012.

Références Bibliographiques
126
Khoury, S., Chatelain, C., Pechere, J.C., Bocon-Gibod L., Richand, C.(1985). Urologie:
Pathologie infectieuse et parasitaire. Paris: Masson, 1985 : 400-448.
Kim, J., Kwon, Y., Pai, H., Kim, J.W., Cho, DT. (1998).Survey of Klebsiella pneumoniae
strains producing extended-spectrum beta-lactamases: prevalence of SHV-12 and SHV-2a in
Korea. J clin Microbiol 1998; 36: 1446-1449.
Knothe, H., Shah, P. D. P., Krcmery, V., Antal, M., Mitsuhashi, S. (1983). Transferable
resistance to cefotaxime, cefoxitin, cefamandole and cefuroxime in clinical isolates of
Klebsiella pneumoniae and Serratia marcescens.Infection, 11(6), 315-317.
Kodio, A. (1979). Etude des infections urinaires au laboratoire de l‘hôpital national du Point
G (à propos de 200 examens bactériologiques) Thèse Pharm, Bamako.
Kodner, C.M., Thomas Gupton, E.K.(2010). Recurrent Urinary Tract Infections in Women
: Diagnosis and Management , Am Fam Physician, 2010, 82, (6), 638-643.
Köning, W.A., Hochmuth, D.H., Joulain, D. (2001). Terpenoids and related constituents of
essential oils. University of Hambourg, Institute of Organic Chemistry, Hambourg, Germany.
Kothari, A., Saga, V.(2008). Antibiotic resistance in pathogens causing community-acquired
urinary tract infections in India: a multicenter study. J Infect Developing Countries ; 2(5):354-
358.
Kounta, L.A. (1999).Sensibilité et évolution de la résistance des entérobactéries aux
antibiotiques à Bamako. Thèse de doctorat en Pharm, Bamako, 1999 ;123p.
Kruger, T., Szabo, D., Keddy, K. H., Deeley, K., Marsh, J. W., Hujer, A. M., Paterson,
D. L. (2004). Infections with nontyphoidal Salmonella species producing TEM-63 or a novel
TEM enzyme, TEM-131, in South Africa.Antimicrobial agents and chemotherapy, 48(11),
4263-4270.
Kurek, A., Nadkowska, P., Pliszka, S., Wolska, K. I. (2012). Modulation of antibiotic
resistance in bacterial pathogens by oleanolic acid and ursolic acid.Phytomedicine, 19(6),
515-519.
Kunikazu, Y., Jun-ichi, W., Yohei, D., Hiroshi, K., Yoshichika, A.(2005). Global Spread
of Multiple Aminoglycoside Resistance Genes. Emerging Infectious Diseases, pp951-953
Kwon, J.A., Yu, C.B., Park, H.D .(2003). Bacteriocidal effects and inhibition of cell
separation of cinnamic aldehyde on Bacillus cereus. Lett. Appl. Microbiol. 37: 61-65.

Références Bibliographiques
127
Lachowicz, K.J., Jones, G.P., Briggs D.R., Bienvenu, F.E., Wan, J., Wilcok,A.,
Coventry, M.J. (1998). The synergetic preservative effects of the essential oils of sweet basil
(Ocimum basilicum) against acid tolerant food microflora. Lett. Appl. Micribiol. 26: 209-214.
Lafaurie, M.(2008).aminoside et fluroquinolone .DU antibiotiques et antibiotherape l‘hopital
de saint louis .
Lambert, R.J, Skandamis, P.N., Coote, P.J, Nychas, G.J .(2001). A study of the minimum
inhibitory concentration and mode of action of oregano essential oil, thymol and carvacrol. J.
Appl. Microbiol. 91: 453-462.
Larabi, K., Masmoudi, A., Fendri, C. (2003). Etude bactériologique et phynotypes de
résistance des germes responsables d‘infections urinaires dans un CHU de Tunis : à propos de
1930 cas. Revue, Médecine et maladies infectieuses
Larif, M., Ouhssine, M., Soulaymani, A., Elmidaoui, A. (2013). Potential effluent oil mills
and antibacterial activity polyphenols against some pathogenic strains. Research on Chemical
Intermediates, 1-13.
Lavigne, J. P., Marchandin, H., Delmas, J., Moreau, J., Bouziges, N., Lecaillon, E.,
Sotto, A. (2007). CTX-M β-lactamase-producing Escherichia coli in French hospitals:
prevalence, molecular epidemiology, and risk factors.Journal of clinical microbiology, 45(2),
620-626
Lavilla, S., Gonzalez-Lopez, J. J., Sabate, M., Garcia-Fernandez, A., Larrosa, M. N.,
Bartolome, R. M., Prats, G. (2008). Prevalence of qnr genes among extended-spectrum β-
lactamase-producing enterobacterial isolates in Barcelona, Spain. Journal of antimicrobial
chemotherapy, 61(2), 291-295.
Lazrak, A., Fraisse, G., Leconte, A., Papillon, P.,Souyri, B. (2014). Vers une méthodologie
générique de modélisation par réseaux de neurones des systèmes énergétiques dans le
bâtiment.
Le Minor, L., Richard, C. (1993). Méthodes de laboratoire pour l'identification des
entérobactéries. Institut Pasteur.
LE Minor,L . (1989a).famille des Entérobactéacéae .
Legras, A., Cattier, B., Perrotin, D. (1993). Dépistage des infections urinaires dans un
service de réanimation: intérêt des bandelettes réactives. Médecine et maladies
infectieuses, 23(1), 34-3.

Références Bibliographiques
128
Lepelletier, D. (1997). Enquete Epidemiologique Sur Les Infections A Escherichia Coli Au
Chu De Nantes (Doctoral Dissertation).
Lerma, M., Cebrián, L., Giménez, M. J., Coronel, P., Gimeno, M., Alfaro, L. A., de
Lomas Barrionuevo, J. G. (2008). B-lactam susceptibility of Escherichia coli isolates from
urinary tract infections exhibiting different resistance phenotypes. Revista Española de
Quimioterapia, 21(3), 149-152.
Leroy, H.,Tattevin P.(2012). Infections urinaires », EMC - Traité Médecine AKO. 7, (2), 1-
6
Leotard, S., Negrin, N. (2010). [Epidemiology of Enterobacteriaceae producing extended-
spectrum beta-lactamase in Grasse Hospital (2005-2008)].Pathologie-biologie, 58(1), 35-38.
Lima, E.O., Gompertz O.F., Giesbrecht, A.M.Paulo, M.O. (1993). In vitro antifungal
activity of essential oils obtained from plants against dermatophytes. Mycoses. 36: 333-336.
Liou, G. F., Yoshizawa, S., Courvalin, P., Galimand, M. (2006). Aminoglycoside
resistance by ArmA-mediated ribosomal 16S methylation in human bacterial
pathogens. Journal of molecular biology, 359(2), 358-364.
Livermore, D.M.(1995). Beta-lactamases in laboratory and clinical resistance. Clinical
Microbiology Reviews;8(4):557-584.
Lobel, B ., Soussy, C . (2007). Les infections urinaires. Paris : Springer-Verlag .p.75
Lobril ,J. R.(1998). Réévaluation du modèle de croissance de Monod : effets des
antibiotiques sur l‘énergie de maintenance. Thèse Université de Lyon I, France. 42, 77.
López-Pujol, J., Zhang, F. M., Sun, H. Q., Ying, T. S., Ge, S. (2011). Mountains of
southern China as ―plant museums‖ and ―plant cradles‖: Evolutionary and conservation
insights. Mountain Research and Development,31(3), 261-269.
Lowe, F. C., Fagelman, E. (2001). Cranberry juice and urinary tract infections: what is the
evidence?. Urology, 57(3), 407-413.
Mahboubi, M., Bidgoli, F. G. (2010). Antistaphylococcal activity of Zataria multiflora
essential oil and its synergy with vancomycin. Phytomedicine, 17(7), 548-550.
Malecky, M. (2007). Métabolisme des terpénoïdes chez les caprins. Thèse de doctorat :
Institut des Sciences et Industries du Vivant et de l‘Environnement, AgroParisTech (France).
Mallea, M., Soler, M., Anfosso, F., Charpin, J. (1979). Activité antifongique d‘essences
aromatiques. Pathol. Biol. 27: 597-60.

Références Bibliographiques
129
Manohar, V., Ingram, C., Gray, J.D., Talpur, N.A., Echard, B.W., Bagchi, D., Preuss
,H.G. (2001). Antifungal activities of origanum oil against Candida albicans. Mol. Cel.
Biochem. 228: 111-117.
Mansour, W., Bouallegue, O., Dahmen, S., Boujaafar, N. (2008) .Caractérisation des
mécanismes enzymatiques de résistance aux b-lactamines chez des souches de Acinetobacter
baumannii isolées à l‘hôpital universitaire Sahloul, Sousse en Tunisie (2005). Pathologie
Biologie. 56 :116–120.
Marrhich, B.(2008). Les Antibiotiques Utilises Dans Les Infections Urinaires.
Marriott, P. J., Shellie, R., Cornwell, C. (2001). Gas chromatographic technologies for the
analysis of essential oils. Journal of Chromatography A,936(1), 1-22.
Martínez-Martínez, L., Pascual, A., Jacoby, G. A. (1998). Quinolone resistance from a
transferable plasmid. The Lancet, 351(9105), 797-799.
Masrar, A., Ouali, M., Bebouda, A., Zouhdi, M., Alaoui, M. A. (2000). L'infection
urinaire nosocomiale en milieu urologique: enquête prospective.Revue marocaine de biologie
infectiologie, 6(2), 45-66.
Masterton, R., Drusano, G., Paterson, D.L., Park, G. 2003. Appropriate antimicrobial
treatment in nosocomial infections-the clinical challenges. J Hosp Infect. 55: 1–12.
Matthew, A., Croxen, B.,Brett, F.(2010). Molecular mechanisms of Escherichia coli
pathogenicity. Nature Reviews Microbiology 8, 26-38.
Mc Lafferty, F.W., Stauffer, D.B. (1994). Wiley Registry of Mass Spectra Data, 6th Ed.
Mass spectrometry Library Search System Bench-Top/PBM, Version 3.10d.
Palisade:Newfield.
Meradi, L., Djahoudi, A., Abdi, A., Bouchakour, M., Perrier Gros Claude, J. D.,
Timinouni, M. (2011). Qnr and aac (6')-Ib-cr types quinolone resistance among
Enterobacteriaceae isolated in Annaba, Algeria. Pathol. Biol. (Paris) 59, e73-e78.
Meral, G. E., Konyalioglu, S., Ozturk, B. (2002). Essential oil composition and antioxidant
activity of endemic Ziziphora taurica subsp. cleonioides.Fitoterapia, 73(7), 716-718.
Messai, Y., Iabadene H., Benhassine T., Alouache S., Tazir T., Gautier T., Arlet G.,
Bakour R. (2008). Prevalence and characterization of extended-spectrum beta- lactamases in
Klebsiella pneumoniae in Algiers hospitals (Algeria). Pathol.Biol.(Paris)56:319-325.
Michel, C. (2014). Infections urinaires gravidiques: analyse de leur prise en charge dans le
service d'hospitalisation des grossesses du CHU Estaing à Clermont-Ferrand.

Références Bibliographiques
130
Microbiol. 56(4): 519–523.
Mimica-Dukić, N., Bozin, B., Soković, M., Mihajlović, B., & Matavulj, M. (2003).
Antimicrobial and antioxidant activities of three Mentha species essential oils. Planta
medica, 69(5), 413-419.
Mkaouar, D., Mahjoubi, F., Mezghani, S., Znazen, A., Ktari, S., Hammami, A. (2008).
Étude de la résistance des entérobactéries aux céphalosporines de troisième génération dans
les hôpitaux de Sfax, Tunisie (1999–2005).Médecine et maladies infectieuses, 38(6), 293-298.
Mondain, V.(2010).E.coli BLSE : une menace grandissante
Montes-Belmont, R.,Carvajal, M. (1998) .Control of Aspergillus flavus in maiz with plant
essential oils and their components. J. food. Prot. 61: 616-619.
Moran, G. J., Krishnadasan, A., Gorwitz, R. J., Fosheim, G. E., McDougal, L. K., Carey,
R. B., Talan, D. A. (2006). Methicillin-resistant S. aureus infections among patients in the
emergency department. New England Journal of Medicine, 355(7), 666-674.
Moukrad, N., Rhazi, F., Makoudi, Y. (2012). Prévalence de la multi-résistance bactérienne
aux antibiotiques des infections urinaires dans la ville de Meknes (Maroc) et son évolution
dans le temps. J Sci Lib, 4, 1-17.
Mulvey, M. R., Bryce, E., Boyd, D., Ofner-Agostini, M., Christianson, S., Simor, A. E.,
Paton, S. (2004). The Canadian Hospital Epidemiology Committee of the Canadian
Nosocomial Infection Surveillance Program, Health Canada. 2004. Ambler class A extended-
spectrum beta-lactamase-producing Escherichia coli and Klebsiella spp. Canadian hospitals.
Antimicrob. Agents Chemother, 48, 1204-1214.
Naas, T., Bentchouala, C., Cuzon, G., Yaou, S., Lezzar, A., Smati, F., Nordmann, P.
(2011). Outbreak of Salmonella enterica serotype Infantis producing ArmA 16S RNA
methylase and CTX-M-15 extended-spectrum β-lactamase in a neonatology ward in
Constantine, Algeria. International journal of antimicrobial agents, 38(2), 135-139.
Naas, T., Bentchouala, C., Lima, S., Lezzar, A., Smati, F., Scheftel, J. M., & Nordmann,
P. (2009). Plasmid-mediated 16S rRNA methylases among extended-spectrum-β-lactamase-
producing Salmonella enterica Senftenberg isolates from Algeria. Journal of antimicrobial
chemotherapy, dkp312.
Naas, T., Lezzar, A., Bentchouala, C., Smati, F., Scheftel, J. M., Monteil, H., Nordmann,
P. (2005). Multidrug-resistant Salmonella enterica serotype Senftenberg isolates producing
CTX-M β-lactamases from Constantine, Algeria. Journal of Antimicrobial
Chemotherapy, 56(2), 439-440.

Références Bibliographiques
131
Nauciel, P. (2001). Bactériologie médicale. Ed Masson.
Nedjai, S., Barguigua, A., Djahmi, N., Jamali, L., Zerouali, K., Dekhil, M., Timinouni
,M. (2012). Prevalence and characterization of extended spectrum beta-lactamases in
Klebsiella-Enterobacter-Serratia group bacteria, in Algeria. Med.Mal Infect. 42:20-29.
Neonakis, I. K., Scoulica, E. V., Dimitriou, S. K., Gikas, A. I., Tselentis, Y. J. (2003).
Molecular epidemiology of extended-spectrum β-lactamases produced by clinical isolates in a
university hospital in Greece: detection of SHV-5 in Pseudomonas aeruginosa and prevalence
of SHV-12. Microbial Drug Resistance, 9(2), 161-165.
Nielsen, P. V., Rios, R. (2000). Inhibition of fungal growth on bread by volatile components
from spices and herbs, and the possible application in active packaging, with special emphasis
on mustard essential oil. International Journal of Food Microbiology, 60(2), 219-229.
NIST .(1999). National Institute of Standards and Technology, PC version 1.7 of the
NIST/EPA/NIH
Niu, C., Gilbert, E. S. (2004). Colorimetric method for identifying plant essential oil
components that affect biofilm formation and structure. Applied and Environmental
Microbiology, 70(12), 6951-6956.
.
Niu, C., Afre, S., Gilbert, E. S. (2006). Subinhibitory concentrations of cinnamaldehyde
interfere with quorum sensing. Letters in applied microbiology,43(5), 489-494.
Nordmann, P.,Carrer, A.( 2010). Carbapenemases in enterobacteriaceae. Arch. Pediatr. 17
Suppl 4, S154-S162.
Nordmann, M., Cabrera, M., Perz, A., Bröcker, C., Ostrowicz, C., Engelbrecht-Vandré,
S., Ungermann, C. (2010). The Mon1-Ccz1 complex is the GEF of the late endosomal Rab7
homolog Ypt7. Current Biology, 20(18), 1654-1659.
Nostro, A., Sudano Roccaro, A., Bisignano, G., Marino, A., Cannatelli, M.A., Pizzimenti
,F.C., Cioni, P.L., Procopio, F., Blanco, A.R. (2007). "Effects of oregano, carvacrol and
thymol on Staphylococcus aureus and Staphylococcus epidermidis biofilms." J. Med.
Microbiol. 56(4): 519–523.
Nüesch-Inderbinen, M. T., Kayser, F. H., & Hächler, H. E. R. B. E. R. T. (1997). Survey
and molecular genetics of SHV beta-lactamases in Enterobacteriaceae in Switzerland: two
novel enzymes, SHV-11 and SHV-12. Antimicrobial agents and chemotherapy, 41(5), 943-
949.
Ofek, I., Edwin H.B.(1982).The importance of mannose specific adhesins (lectins) in
infections caused by Escherichia coli. Scand. J. Infect. Dis. Suppl. 33 : 61-7.

Références Bibliographiques
132
Ohno, T., Kita, M., Yamaoka, Y., Imamura, S., Yamamoto, T., Mitsufuji, S., Imanishi, J.
(2003). Antimicrobial activity of essential oils against Helicobacter pylori. Helicobacter, 8(3),
207-215.
Ozturk, S., & Ercisli, S. (2007). Antibacterial activity and chemical constitutions of
Ziziphora clinopodioides. Food control, 18(5), 535-540.
Park, Y. S., Adams-Haduch, J. M., Shutt, K. A., Yarabinec, D. M., Johnson, L. E.,
Hingwe, A.,Doi, Y. (2012). Clinical and Microbiologic Characteristics of Cephalosporin-
Resistant Escherichia coli in Three Centers in the United States. Antimicrobial agents and
chemotherapy, AAC-05650.
Paterson, D. Bonomo, R. (2005). Extended-spectrum 3-1actamases: a clinical update.
Clinical microbiology reviews. 657-686.
Patterson, J.E. (2001). Antibiotic utilization: is there an effect on antimicrobial resistance.
Chest. 119: 426–30.
Peyramaure, S .L. (2008). Troubles urinaires et prostatiques. Actualités pharmaceutiques,
N°480.
Perichon, B., Courvalin, P., Galimand, M. (2007). Transferable resistance to
aminoglycosides by methylation of G1405 in 16S rRNA ESBL genes in the Moroccan
community http://jmm.sgmjournals.org 1351 Downloaded from
www.microbiologyresearch.org by IP: 197.119.109.83 On: Fri, 06 May 2016 18:58:41 and to
hydrophilic fluoroquinolones by QepA-mediated efflux in Escherichia coli. Antimicrob
Agents Chemother 51, 2464–246.
Pellecuer, J., Jacob ,M., Simeon De Bouchberg, M. (1980) .Essais d‘utilisation d‘huiles
essentielles de plantes aromatiques méditerranéennes en odontologie conservatrice. Plant.
Med. Phytother. 14: 83-98.
Pelmont, J. (1995). Bactéries et environnement adaptation physiologique. Volume 1. Office
des publications universitaires, Ben-Aknoun (Alger)
Pelt, J.M. (2001) Les nouveaux actifs naturels. Marabout. Paris
Pena, C., Gudiol, C., Tubau, F., Saballs, M., Pujol, M., Dominguez, M.A., Calatayud,
L, Ariza, J., Gudiol, F.(2006). Risk-factors for acquisition of extended-spectrum beta-
lactamase-producing Escherichia coli among hospitalised patients. Clin Microbiol Infect
;12(3):279-284. 26.

Références Bibliographiques
133
Perilli, M., Dell'Amico, E., Segatore, B., de Massis, M. R., Bianchi, C., Luzzaro, F.,
Amicosante, G. (2002). Molecular characterization of extended-spectrum β-lactamases
produced by nosocomial isolates of Enterobacteriaceae from an Italian nationwide
survey. Journal of clinical microbiology, 40(2), 611-614.
Perilli, M., Felici, A., Franceschini, N., De Santis, A. N. N. A. L. I. S. A., Pagani, L.,
Luzzaro, F., Amicosante, G. (1997). Characterization of a new TEM-derived beta-lactamase
produced in a Serratia marcescens strain.Antimicrobial agents and chemotherapy, 41(11),
2374-2382.
Pibiri, M. C. (2005). Assainissement microbiologique de l" air et des systèmes de ventilation
au moyen d" huiles essentielles.
Perrière, G.(1992). Application d‘une présentation par objet des connaissances de
modélisation de certains aspects de l‘expression des gènes chez E.coli UCBL. Thèse
Université de Lyon I, France. 14, 77.
Philippon, A., Arlet, G. (2005). Les B-lactamases chez les bacilles à gram négatif: que de
nouveautés en 15 ans! Antibiotiques 2005; 7: 247-259.
Philippon, A. (2013). Les bêta-lactamases à spectre élargi ou étendu (BLSE).Immuno-
analyse & Biologie Spécialisée, 28(5), 287-296.
Philippon, A., Arlet, G., Jacoby, G.A. (2002). Plasmid-determined AmpC-type
Blactamases. Antimicrob. Agents Chemother. 46:1-11.
Poirel, L., Leviandier, C., Nordmann, P.(2006). Prevalene and genetic analysis of plasmid
mediated quinolone resistance determinants Qnr A and Qnr S in Enterobacteriaceae isolates
from a French university hospital. Antimicrob Agents Chemother .50: 3992- 7.
Poirel, L., Potron, A., Nordmann, P. (2012). OXA-48-like carbapenemases: the phantom
menace. Journal of Antimicrobial Chemotherapy, dks121.
Poole, K. (2005). Efflux-mediated antimicrobial resistance. J. Antimicrob. Chemother. 56:20–
51. 10.1093/jac/dki171.
Ponce, A. G., Fritz, R., Del Valle, C., Roura, S. I. (2003). Antimicrobial activity of essential
oils on the native microflora of organic Swiss chard. LWT-Food Science and
Technology, 36(7), 679-684.
Pozzatti, P., Loreto, É.S., Mario, D.A.N., Rossato, L.,Santuario, J.M., Alves, S.H.(2010).
Activities of essential oils in the inhibition of Candida albicans and Candida dubliniensis
germ tube formation. Journal de Mycologie Médicale, v. 20, p. 185-189.

Références Bibliographiques
134
Prammananan, T., Sander, P., Brown, B. A., Frischkorn, K., Onyi, G. O., Zhang, Y.,
Wallace, R. J. (1998). A single 16S ribosomal RNA substitution is responsible for resistance
to amikacin and other 2-deoxystreptamine aminoglycosides in Mycobacterium abscessus and
Mycobacterium chelonae.Journal of Infectious Diseases, 177(6), 1573-1581..
Quezel, P., & Santa, S. (1963). Nouvelle Flore de l‘Algerie et des Regions Desertiques et
Meridionales II, eds du Centre National de la Recherche Scientifique.
Ramdani-Bouguessa, N., Manageiro, V., Jones-Dias, D., Ferreira, E., Tazir, M. ,Canica
,M. (2011). Role of SHV beta-lactamase variants in resistance of clinical Klebsiella
pneumoniae strains to beta-lactams in an Algerian hospital. J.Med.Microbiol. 60:983-987
Ramdani-Bouguessa, N., Bes, M., Meugnier, H., Forey, F., Reverdy, M. E., Lina, G.,
Etienne, J. (2006). Detection of methicillin-resistant Staphylococcus aureus strains resistant
to multiple antibiotics and carrying the Panton-Valentine leukocidin genes in an Algiers
hospital. Antimicrobial agents and chemotherapy, 50(3), 1083-1085.
Ramdani-Bouguessa, N., Rahal, K. (2003). Serotype distribution and antimicrobial
resistance of Streptococcus pneumoniae isolated in Algiers, Algeria. Antimicrobial agents and
chemotherapy, 47(2), 824-826.
Ramoul, A., Hammami, S., Dekhil, M., Aimiri, S.,Slim, A., Boutiba-Ben Boubaker,
I.
(2013). Phenotypic and genotypic characterization of clinical multidrug resistant
Acinetohacter baumannii from Algerian intensive care units. African Journal of Microbiology
Research Volume 7(10), pp. 868- 874.
Rasooli, I., Rezaei, M. B., Allameh, A. (2006). Growth inhibition and morphological
alterations of Aspergillus niger by essential oils from Thymus eriocalyx and Thymus x-
porlock. Food Control, 17(5), 359-36
Razakarivony, A.A., Andriamihaja, B., Razanamahefa, B. (2009). Etude chimique de
l‘huile essentielle des feuilles de Callistemon rigidium. Actes du symposium biomad.
Université d‘Antananarivo. 28 p.
Regiane Ribeiro dos ,Santos. , Nathália, Ramos .,de Melo, Joyce .,Fagundes, Gomes ,
Motta , Erika, Fraga de Souza ., Reinaldo Francisco, Teófilo.(2014). minimale
concentration oftwo oils of different species of Cinnamon against microorganisms
alimentares.
Reverchon, E., De Marco, I. (2006). Supercritical fluid extraction and fractionation of
natural Matter. J. of Supercritical Fluids, 38 : 146-166.
Richard, H. (2008). Epices et herbes aromatiques.

Références Bibliographiques
135
Robicsek, A., Jacoby, G,A.,Hooper, D,C.(2006). The worldwide emergence of plasmid
mediated quinolone resistance. Lancet Infect Dis . 6: 629-40.
Robert-Demuet, S. (1995). Méthodes de dilutions. In Antibiotiques et antibiogrammes, 131-
137. Montréal-Canada
Robles, M., Aregullin, M., West, J., Rodriguez, E. (1995). Recent studies on the
zoopharmacognosy, pharmacology and neurotoxicology of sesquiterpene lactones. Planta
medica, 61(3), 199-203.
Rodríguez-Martínez, J. M., Cano, M. E., Velasco, C., Martínez-Martínez, L., Pascual, Á.
(2011). Plasmid-mediated quinolone resistance: an update. Journal of Infection and
Chemotherapy, 17(2), 149-182.
Rodríguez-Baño, J., Navarro, M.D., Romero, L., Muniain, M.A., Perea, E.J., Pérez-
Cano, R., Jose, R. H., Alvaro, P.(2006). Clinical and Molecular Epidemiology of Extended-
Spectrum β Lactamase—Producing Escherichia coli as a Cause of Nosocomial Infection or
Colonization: Implications for Control. Clin Infect Dis. 142(1):37-45.
Roh, K.H., Uh, Y., Kim, J.S, Kim HS., Shin, D.H., Song, W.(2008). First outbreak of
multidrugresistant Klebsiella pneumoniae producing both SHV-12-type extended-spectrum
betalactamase and DHA-1-Type AmpC beta-lactamase at a Korean Hospital Yonsei. Med;
49(1): 53-7.
Rosato, A., Vitali, C., Gallo, D., Balenzano, L., Mallamaci. R.(2008). The inhibition of
Candida species by selected essential oils and their synergism with amphotericin B.
Phytomedicine 15. 635– 638.
Rosenberg, E. Y., Ma, D., & Nikaido, H. (2000). AcrD of Escherichia coli is an
aminoglycoside efflux pump. Journal of Bacteriology, 182(6), 1754-1756.
Rostoker, G., Benmaadi, A., Lagrue G.(1991) . Infections urinaires hautes : pyélonéphrites
, Encycl. méd.-chir. Urol.
Roulier, G. (1992). Les huiles essentielles pour votre santé; Traité pratique d‘aromathérapie:
Propriétés et indications thérapeutiques des essences de plantes. Ed Dangles. France.
Rubin, M.A., Samore, M.H.(2002). Antimicrobial Use and Resistance. Curr Infect Dis Rep
; 4 : 491-7.
Ruppé, E. (2010). Epidemiology of expanded-spectrum beta-lactamases: the rise of CTX-M.
Antibiotiques.12:3-16.
Sabaté, M., Tarragó, R., Navarro, F., Miró, E., Vergés, C., Barbé, J., Prats, G.(2000)
Cloning and sequence of the gene encoding a novel cefotaxime-hydrolyzing β-lactamase

Références Bibliographiques
136
(CTX-M-9) from Escherichia coli in Spain.Antimicrob Agents Chemother. ;44:1970–
1973. [PMC free article] [PubMed]
Saint, S ., Lipsky, B.A. (1999). Preventing catheter-related bacteriuria. Arch Intern Med. Apr
26;159(8):800–8. [PubMed].
Salehi, P., Sefidkon, F., Bazzaz Tolami, L. (2005), Es- essential oil of Nepeta crispa Willd.
from Iran. Z. Nasential oil composition of Salvia xanthocheila from turforsch. 59c, 653Ð656.
Iran. J. Essent. Oil Res. 17, 442Ð443.
Sambrook, J., & Russell, D. W. (2001). Molecular Cloning: A Laboratory Manual .
Sanchez, C.J., Mende, K., Beckius, M.L., Akers, K.S., Romano, D.R., Wenke, J.C. ,
Murray, C.K. (2013). "Biofilm formation by clinical isolates and the implications in chronic
infections." BMC infectious diseases 13(1): 47.
Santo, E., Salvador, M.M., Marin, J.M.(2007). Multidrug-resistant urinary tract isolates of
escherichia coli from Ribeirao Preto, Sao Paulo, Brazil. Braz J Infect Dis. ;11(6):575–
8. [PubMed]
Sargol, F., Hossein, S., Kafil,M., Reza, N., Mohammad, A., Roghaya, N., Mohammad
,A.(2015). Relation ship of biofilm formation and defferent virulence gene .PMC.
Schlessinger, D. (1988). Failure of aminoglycoside antibiotics to kill anaerobic, low-pH, and
resistant cultures. Clin. Microbiol. Rev. 1:54-59. [PMC free article] [PubMed
Schroeder, M. P., & Messing, A. M. (1949). Methods for comparing the antibacterial
activity of essential oils and other aqueous insoluble compounds.Bull Nat Formulary
Comm, 17, 213-8.
Schmidt, E. (2010). Production of essential oils. Handbook of Essential Oils: Science,
Technology, and Applications. Can Başer K. H. etBuchbauer G. Florida, CRC Press: 83–119.
Schwartz, R., Davi, R., Hilton, T.J. (1992). Effect of temporary cements on the bond
strength of a resin cement. Am. J. Dent. 5: 147-150.
Senhaji, O., Faid, M., chraq K. (2007). Inactivation of Escherichia coli O157:H7 by
essential oil from Cinnamomum zeylanicum. Braz. J. Infect. Dis., 11(2): 234-236.
Sezik, E., Ttimen G., Baser, K. H. C. (1991). Ziziphora tenuior L., a new source of
pulegone. Flavour and Fragrance J., 6: 101 - 103.
Shahid, M., Mohammad, F. (2013). Recent advancements in natural dye applications: a
review. Journal of Cleaner Production, 53, 310-331.

Références Bibliographiques
137
Shapiro, S., Meier, A., Guggenheim, B. (1994). The antimicrobial activity of essential oils
and essential oil components towards oral bacteria. Oral. Microbiol. Immunol. 9: 202- 208.
Shin, S., Kang, C. (2003). Antifungal activity of the essential oil of Agastache rugosa Kuntze
and its
Sibery, G., Brahmadathan, K.n., Rajeswar, p., Lalitha, M.k., Mark C., Jacob, T.(2001).
Comparison of different culture media and storage temperature for the long-term preservation
of Streptococcus pneumonia Bulletin of the World Health Organization page 44.
Sikkema, J.A., de, Bont., Poolman, B. (1995). Mechanisms of membrane toxicity of
hydrocarbons, Journal of Applied Microbiology. Wiley Online Library. p 63-69.
Skirej, A.,Elattir, H ., Elfadl ,A.(2012).L‘art de produire les legumes et les fruits au maroc
Silva, F., Ferreira, S., Queiroz, J. A., Domingues, F. C. (2011). Coriander (Coriandrum
sativum L.) essential oil: its antibacterial activity and mode of action evaluated by flow
cytometry. Journal of medical microbiology, 60(10), 1479-1486.
Sivropoulou, A., Kokkini, S., Lanaras, T., & Arsenakis, M. (1995). Antimicrobial activity
of mint essential oils. Journal of Agricultural and Food Chemistry,43(9), 2384-2388.
Skandamis, P.N., Nychas, G.J.E. (2001). Effect of oregano essential oil on microbiological
and physicichemical attributes of minced meat stored in air and modified atmospheres. J.
Appl. Microbiol. 91: 1011-22.
Skurnik, D., Ruimy, R., Andremont, A., Amorin, C., Rouquet, P., Picard, B., Denamur,
E. (2006). Effect of human vicinity on antimicrobial resistance and integrons in animal faecal
Escherichia coli. Journal of Antimicrobial chemotherapy, 57(6), 1215-1219.
Smith-Palmer, A., Stewart, J., Fyfe, L. (2001). The potential application of plant essential
oils as natural food preservatives in soft cheese. Food Microbiology, 18(4), 463-470.
Société de Pathologie Infectieuse de Langue Française (SPILF) et l’Association
Française d’Urologie (AFU).2002. Conférence sur l‘ Infections urinaires nosocomiales de
l‘adulte
Sotto, A., Merle, De Boever, C., Fabro-Perray, P., Gouby, A., Sirot, D. ,Jourdan
J.(2001). Risk factors for antibiotic-resistant Escherichia coli isolated from hospitalised
patients with urinary tract infections: a prospective study. J Clin Microbiol; 39: 438-44.
Souna, D.(2010). Epidémiologie de la résistance aux antibiotiques des entérobactéries au
niveau du C.H.U de Sidi Bel Abbes mémoire de magister .universitie de Tlemcen .

Références Bibliographiques
138
Sourai, P.G. (1989). Antimicrobial action of dental materials used in operative dentistry: a
reiew. Odontostomatol. Proodos. 43(5): 399-408.
Sourander, L.B . (1966). Urinary tract infection in the aged an epidemiological study. Am
Med Intern Fenn 55: (45): 7-55.
Soussy, C. J., Cavallo, J. D., Courcol, R., Drugeon, H., Fahre, R., Jarlier, V., Sirot, J.
(2000). Sensibilité aux antibiotiques de souches d'Escherichia coli isolées en 1998 et 1999:
résultats d'une enquête multicentrique française.Médecine et maladies infectieuses, 30(10),
650-656.
Spanu, T., Luzzaro, F., Perilli, M., Amicosante, G., Toniolo, A., Fadda, G., Italian ESBL
Study Group. (2002). Occurrence of extended-spectrum β-lactamases in members of the
family Enterobacteriaceae in Italy: implications for resistance to β-lactams and other
antimicrobial drugs. Antimicrobial Agents and Chemotherapy, 46(1), 196-202.
SPILF .(2014). Diagnostic Et Antibiotherapie Des Infections Urinaires Bacteriennes
Communautaires De L‘adulte.
Strahilevitz, J., Jacoby, G. A., Hooper, D. C., Robicsek, A. (2009). Plasmid-mediated
quinolone resistance: a multifaceted threat. Clinical microbiology reviews, 22(4), 664-689.
Suresh, B., Sriram, S., Dhanaraj, S.A., Elango, K., chinnaswamay, K. (1997).
Anticandidosique activity of Santolina chamaecyparissus volatile oil. J. Ethnopharmacol. 55:
151-159.
Sykes, R.B.,Bonner D.P.,Bush K., Georgopapadakou N.H., Wells J.S. (1981).
Monobactams--monocyclic beta-lactam antibiotics produced by bacteria. J Antimicrob
Chemother 8 Suppl E:1-16.
Tangjitjaroenkun, J., Chavasiri, W., Thunyaharn, S., Yompakdee, C. (2012).
Bactericidal effects and time–kill studies of the essential oil from the fruits of Zanthoxylum
limonella on multi-drug resistant bacteria. Journal of Essential Oil Research, 24(4), 363-370.
Tassouiket, S. (2014). Sensibilité aux antibiotiques d‘Escherichia coli responsables
d‘infections urinaires communautaires à l‘Institut Pasteur de Casablanca (Doctoral
dissertation).
Thabet, L., Messadi, A., Mbarek, M., Turki, A., Meddeb, B.Ben Redjeb, S.
(2008).Surveillance Des Bactéries Multi Résistantes aux antibiotiques dans un hôpital
tunisien. Journale de la societie tunisiene des science medicinale. 86: 992-5.

Références Bibliographiques
139
Thomas, J. (2001). New quinolones and the impact on resistance. Drug Discovery Today.
6(10): 529-536.
Tiba, M.R, Yano, T., Leite, D.D.S.(2008). Genotypic characterization of virulence factors
in Escherichia coli strains from patients with cystitis. Rev Inst Med Trop. 2008;50(5):255–
260. [PubMed].
Tietz, A., Francioli, P., Widmer, A.F. (2004).Bêtalactamases à spectre étendu : implications
pour l ‘ hygiène hospitalière. Swiss noso (11, Numéro 4, article 2).
Touati, A., Medboua, C., Touati, D., Denine, R., Brasme, L., de Champs, C. (2012).
CTXM-15- producing Enterobacteriaceae isolates causing bloodstream infections at the
BeniMessous hospital in Algeirs (Algeria). Int Res J Microbiol. 3: 181-185.
Touati, A., Zenati, K., Brasme, L., Benallaoua, S., De Champs, C. (2010). Extended-
spectrum β-lactamase characterisation and heavy metal resistance of Enterobacteriaceae
strains isolated from hospital environmental surfaces.Journal of Hospital Infection, 75(1), 78-
79.
Touati, A., Brasme, L., Benallaoua, S., Madoux, J., Gharout, A., De Champs, C.
(2008a). Enterobacter cloacae and Klebsiella pneumoniae isolates producing CTX-M-15
recovered from hospital environmental surfaces from Algeria. Journal of Hospital
Infection, 68(2), 183-185
.
Touati, A., Brasme L., Benallaoua, S., Gharout, A., Madoux, J. de Champs, C. (2008b).
First report of qnrB-producing Enterobacter cloacae and qnrA-producing Acinetobacter
baumannii recovered from Algerian hospitals. Diagn. Microbiol. Infect. Dis. 60, 287-290.
Touati, A., Benallaoua S., Forte D, Madoux J., Brasme L. and C. C. De. (2006). First
report of CTX-M-15 and CTX-M-3 beta-lactamases among clinical isolates of
Enterobacteriaceae in Bejaia, Algeria. Int.J.Antimicrob.Agents 27:397-402.
Traore, H., Keita, B., Keita, S., Diallo, F., Dembele, M., Diallo, D., Pichard, E. (1994).
Etude clinique et bactériologique de l'infection urinaire dans le service de médecine interne à
l'hôpital national du point «G», Bamako, Mali. Médecine d'Afrique noire, 41(6), 336-342.
Trecarichi, E.M., Cauda, R., Tumbarello, M.(2012). Detecting risk and predicting patient
mortality in patients with extended-spectrum β-lactamase-producing Enterobacteriaceae
bloodstream infections. Future Microbiol.(10):1173-1189.
Tzouvelekis, L. S., Tzelepi, E., Tassios, P. T., Legakis, N. J. (2000). CTX-M-type B-
lactamases: an emerging group of extended-spectrum enzymes. Int. J. Antimicrob. Agents 14:
137-143.

Références Bibliographiques
140
Unlu, M., Ergene, E., Unlu, G.V, Zeytinoglu, H.S, Vural, N.(2010). Composition,
antimicrobial activity and in vitro cytotoxicity of essential oil from cinnamomum zeylanicum
blume (lauraceae). Food Chem Toxicol. PubMed. 48: 3274-3280. 10.1016/j.fct.2010.09.001.
Usein, C. R., Damian, M., Tatu‐Chitoiu, D., Capusa, C., Fagaras, R., Tudorache, D.,
Bouguénec, C. (2001). Prevalence of virulence genes in Escherichia coli strains isolated from
Romanian adult urinary tract infection cases. Journal of cellular and molecular
medicine, 5(3), 303-310..
Vahaboglu, H., Dodanli, S., Eroglu, C., Ôzttirk, R., Soyletir, G., Yildirim, 1., Avkan, V.
(1996). Characterization of multiple-antibiotic-resistant Salmonella typhimurium strains:
molecular epidemiology of PER-1 -producing isolates and evidence for nosocomial plasmid
exchange by a clone. J. Clin. Microbiol. 34: 2942-2946.
Vakulenko, S. B., Mobashery, S. (2003). Versatility of aminoglycosides and prospects for
their future. Clinical microbiology reviews, 16(3), 430-450.
Valeri, A., Joulin, V., Fournier, G. (1998). Prostatites. 1998 Enc 8 ycl Med Chir,
Néphrologie-Urologie.
Valnet, J. (1990). Aromathérapie: traitement des maladies par les essences des plantes.
11ème Maloine edition. Paris.
Van Bambeke, F., Michot, J.M., Van, Eldere, J., Tulkens P.M. (2005).Quinolones . an
update. Clin Microbiol Infect;11:256-80.
Varel, V.H. (2002). Live stock manure odor abatement with plant derived oils and nitrogen
conservation with urease inhibitors: A review 1. J. Anim. Sci. 80: E1-E7.
Vazquez J.A., Arganoza M.T., Boikov D., Vaishampayan J.K., Akins R.A. (2000). In
vitro susceptibilities of Candida and Aspergillus species to Melaleuca alternifolia (tea tree)
oil. Rev Iberoam. Micol. 17: 60-63.
Verbeke, N.(2006). L‗Aromatherapie Comme Alternative Credible A L‘antibiotherapie
Versalovic, J., Koeuth, T., Lupski, R. (1991). Distribution of repetitive DNA sequences in
eubacteria and application to finerpriting of bacterial enomes.Nucleic acids research, 19(24),
6823-6831.
Vigne, P. (1987). La France et ses productions aromatiques végétales actuelles. Parfums,
Cosmetiques, Arômes. 78: 97-103.
Vilas A., Kamble,L .(2013). Anti-bacterial studies of essential oils against mdr escherichia
coli isolated from different clinical specimens.an International peer reviewed journal Vol.4.

Références Bibliographiques
141
Vildé, J.L.,Byl, B.,Pothier, P.,Mangin, P.H .,Coloby., Davin, J.L., (2002).Infections
urinaires nosocomiales de l‘adulte.
Viollon, C., Chaumont ,J.P. (1994) Antifungal properties of essential oils and their main
components upon Cryptococcus neoformans. Mycopathologia. 128: 151-153.
Viollon, C., Leger, D., Chaumont, J.P. (1993). Activités antagonistes in vitro de certains
composés volatils naturels vis-à-vis de germes de la flore vaginale. Plant. Med. Phytother. 26:
17-22.
Vitale, R., Afeltra, J., Dannaoui, E., 2005. Antifungal Combinations, in: Ernst, E., Rogers,
P.D. (Eds.).
Vittori, M., D'Addessi A., Sasso, F.(2012) .Microbiological follow-up of nosocomial
infections in a single urological center. Urologia. Dec;79(19):147–51. [PubMed].
Vorkaufer, S.(2011). Les infections urinaires communautaires bactériennes de l‘adulte : prise
en charge diagnostique et thérapeutique. Résultats de deux tours d‘un audit clinique réalisé
par 66 médecins généralistes lorrains.- 104p. Th: Méd. : Nancy
Veron, M.,Kazmierczak, A. (1971). L'exploration cytobactériologique au cours des
infections urinaires. Médecine et Maladies Infectieuses, 1(5), 235-241.
Walsh,C.(2003) .naturel and produced immunity versus acquired resistance .in
antibiotics:action origins ,resistance pp1-106 ed ASMpres wasligh.
Wagenlehne, F.M.E.,Naber, K.G.(2001). Uncomplicated urinary tract infections in
women. Current Opin Urol.;11(1):49–53. [PubMed
Wang, Y., Wang, T., Hu, J., Ren, C., Lei, H., Hou, Y., Brantner, A. H. (2011). Anti-
biofilm activity of TanReQing, a Traditional Chinese Medicine used for the treatment of acute
pneumonia. Journal of ethnopharmacology, 134(1), 165-17.
Wassef, I., Behiry, M., Younan, M., El Guindy, N., Mostafa, S.,Abada
E.(2014).Genotypic identification of AmpC β-lactamases production in gram-
negative Bacilli isolates,‖ Jundishapur Journal of Microbiology, vol. 7, no. 1, Article ID
e8856.
Weill, F. X., Demartin, M., Tandé, D., Espié, E., Rakotoarivony, I., Grimont, P. A.
(2004). SHV-12-like extended-spectrum-β-lactamase-producing strains of Salmonella
enterica serotypes Babelsberg and Enteritidis isolated in France among infants adopted from
Mali. Journal of clinical microbiology, 42(6), 2432-2437.

Références Bibliographiques
142
Wiegand, I., Hilpert, K., Hancock, R.E.W., 2008. Agar and broth dilution methods to
determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nature
Protocols 3, 163-175.
Whittan, T. S. (1996). Génetic variation and evolutionary processes in natural populations of
Escherichia Coli. In: F.C.NERDHARDT (ed.) E. coli and salmonella: cellular and molecular
biology. Washington,DC,USA: ASM Press.
Wilkinson, J. M. (2006). Methods for testing the antimicrobial activity of extracts. Modern
Phytomedicine: Turning Medicinal Plants into Drugs, 157-171.
Wichtl, M., Anton, R. (2003). Plantes thérapeutiques, tradition, pratique officinale, science et
thérapeutique. 2 e édition. EMInter/Tec & Doc éditions, Paris, 587-589.
Willem, J.P. (2002). Les huiles essentielles médecine d'avenir. Dauphin, Paris. 318 P
Wiegand, I., Hilpert, K., Hancock, R.E.W. (2008). Agar and broth dilution methods to
determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nature
Protocols 3, 163-175.
Wise, R.(1990).The pharmacokinetics of the oral cephalosporins - A review. J Antimicrob
Chemother, 26(SupplE):13-20.
Wong-Beringer, A., Hindler, J., Loeloff, M., Queenan, A. M., Lee, N., Pegues, D. A.,
Bush, K. (2002). Molecular correlation for the treatment outcomes in bloodstream infections
caused by Escherichia coli and Klebsiella pneumoniae with reduced susceptibility to
ceftazidime. Clinical infectious diseases, 34(2), 135-146.
Wright, G. D., Thompson, P. R. (1999). Aminoglycoside phosphotransferases: proteins,
structure, and mechanism. Front Biosci, 4, D9-D21.
Wu, J.J., Ko, W.C., Tsai, S.H.,Yan, J.J. (2007). Prevalence of plasmid-mediated quinolones
resistance determinants QnrA, QnrB, and QnrS among clinical isolates of Enterobacter
cloacae in a Taiwanese hospital; 51(4): 1223-1227.
Wuryatmo, E., Klieber, A., Scott, E. S. (2003). Inhibition of citrus postharvest pathogens
by vapor of citral and related compounds in culture.Journal of agricultural and food
chemistry, 51(9), 2637-2640.
Ya Bi, F. A. R.(2006). Profil antibiotypique des bacteries responsables d‘infection urinaire
communautaire. These De Doctorat .Universite De Bamako.
Yagi, T., Kurokawa, H., Shibata, N., Shibayama, K., Arakawa, Y. (2000). A preliminary
survey of extended-spectrum β-lactamases (ESBLs) in clinical isolates of Klebsiella
pneumoniae and Escherichia coli in Japan. FEMS microbiology letters, 184(1), 53-56.

Références Bibliographiques
143
Yan, J. J., Wu, S. M., Tsai, S. H., Wu, J. J., Su, I. J. (2000). Prevalence of SHV-12 among
Clinical Isolates ofKlebsiella pneumoniae Producing Extended-Spectrum β-Lactamases and
Identification of a Novel AmpC Enzyme (CMY-8) in Southern Taiwan. Antimicrobial agents
and chemotherapy, 44(6), 1438-1442.
Yang, X., Strobel, M., Tian, L., Barennes, H., Buisson, Y. (2011). [Bacterial flora in acute
exacerbations of chronic obstructive pulmonary disease (COPD) in Kunming,
China]. Medecine et maladies infectieuses, 41(4), 186-191.
Yap, P.S., Lim, S.H., Hu, C.P.Yiap, B.C.( 2013). Combination of essential oils and
antibiotics reduce antibiotic resistance in plasmid-conferred multidrug resistant
bacteria. Phytomedicine. PubMed;20:710–713. doi: 10.1016/j.phymed.2013.02.013.
Zahlane.,Zougarhi, L .,Soraa, N.,Haouach, K., Admou, B., Arsalane, L.,Chabaa L(2009).
Épidémiologie et résistance aux antibiotiques des entérobactéries isolées d‘infections urinaires
à l‘hôpital universitaire Ibn Tofail de Marrakech.
Zargari, A .(1995). Iranian Medicinal plants. Tehran University press, Tehran, Iran, 4: 103-
104.
Zhao, X., Xu, C., Domagala, J.,Drlica, K. (1997).DNA topoisomerase targets of the
fluoroquinolones: a strategy for avoiding bacterial resistance. Proc Natl Acad Sci USA,
;94:13991-6.
.
.

Annexe

Annexe1
Fiche d’enquête sur les infections urinaires
ETAT CIVIL
Nom :...................................Prénom : ....................................
Sexe : M / F
Femmes : enceinte : O / N
Age …………. ou date de naissance : ….. / .…. / …..
Adresse du patient (nom du quartier) : .…………….……
CLINIQUE
Date du prélèvement : ….. / .…. / ….. h
Motif de la consultation : Douleur / Fièvre>38°C / Ctrl après traitement / ne sais pas /
Autres : ………..
Anomalie rénale ou vaginale : N / anomalie rénale :…………….
Anomalie vaginale : ……………..
Terrain diabétique : O / N
Hospitalisation de moins de 6 mois dans une clinique ou un hôpital : O / N
Si oui, nom de la clinique ou de l’hôpital : …………………….
Présence de sonde urinaire il y a moins de 7 jours : O / N
Antécédent infection urinaire : O / N

Annexe1
Questionnaire sur l’utilisation des plantes anti-Infections Urinaires
1- Information sur les plantes anti-IU
Connaissez- vous des plantes traditionnelles pour le traitement des IU?
Oui Non
Si oui, les quelles et comment ?………………………………………………………
2- Utilisez-vous les plantes traditionnelles pour traiter les IU ?
Oui Non
Si oui, les quelles ?…………………………………………………………………….
3- Sous quelle forme vous utilisez ?
…………………………………………………………………………………………
4- Quelle est la fréquence avec laquelle vous utilisez les plantes médicinales ?
………………………………………………………………………………………….
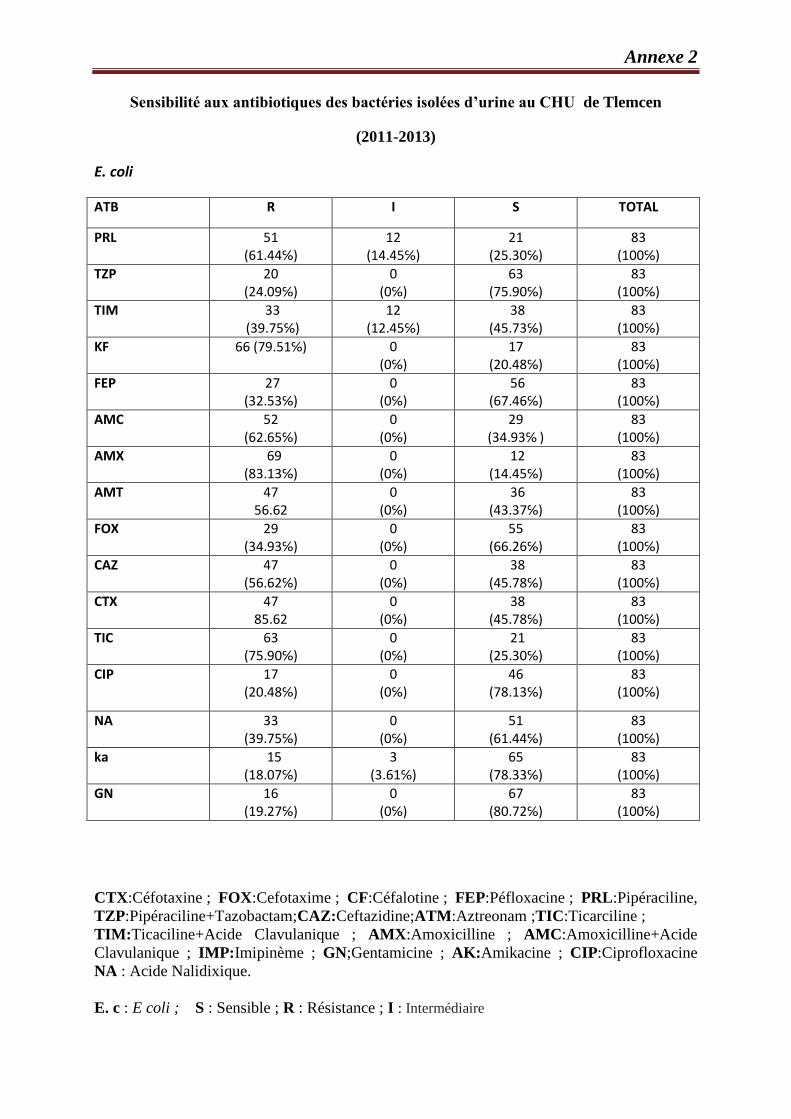
Annexe 2
Sensibilité aux antibiotiques des bactéries isolées d’urine au CHU de Tlemcen
(2011-2013)
E. coli
ATB R I S TOTAL
PRL 51 (61.44℅)
12 (14.45℅)
21 (25.30℅)
83 (100℅)
TZP 20 (24.09℅)
0 (0℅)
63 (75.90℅)
83 (100℅)
TIM 33 (39.75℅)
12 (12.45℅)
38 (45.73℅)
83 (100℅)
KF 66 (79.51℅) 0 (0℅)
17 (20.48℅)
83 (100℅)
FEP 27 (32.53℅)
0 (0℅)
56 (67.46℅)
83 (100℅)
AMC 52 (62.65℅)
0 (0℅)
29 (34.93℅ )
83 (100℅)
AMX 69 (83.13℅)
0 (0℅)
12 (14.45℅)
83 (100℅)
AMT 47 56.62
0 (0℅)
36 (43.37℅)
83 (100℅)
FOX 29 (34.93℅)
0 (0℅)
55 (66.26℅)
83 (100℅)
CAZ 47 (56.62℅)
0 (0℅)
38 (45.78℅)
83 (100℅)
CTX 47 85.62
0 (0℅)
38 (45.78℅)
83 (100℅)
TIC 63 (75.90℅)
0 (0℅)
21 (25.30℅)
83 (100℅)
CIP 17 (20.48℅)
0 (0℅)
46 (78.13℅)
83 (100℅)
NA 33 (39.75℅)
0 (0℅)
51 (61.44℅)
83 (100℅)
ka 15 (18.07℅)
3 (3.61℅)
65 (78.33℅)
83 (100℅)
GN 16 (19.27℅)
0 (0℅)
67 (80.72℅)
83 (100℅)
CTX:Céfotaxine ; FOX:Cefotaxime ; CF:Céfalotine ; FEP:Péfloxacine ; PRL:Pipéraciline,
TZP:Pipéraciline+Tazobactam;CAZ:Ceftazidine;ATM:Aztreonam ;TIC:Ticarciline ;
TIM:Ticaciline+Acide Clavulanique ; AMX:Amoxicilline ; AMC:Amoxicilline+Acide
Clavulanique ; IMP:Imipinème ; GN;Gentamicine ; AK:Amikacine ; CIP:Ciprofloxacine
NA : Acide Nalidixique.
E. c : E coli ; S : Sensible ; R : Résistance ; I : Intermédiaire

Annexe 2
Enterobacter sakazakic
ATB R I S TOTAL
PRL 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
TZP 0 (0℅)
1 (25℅)
3 (75℅)
4 (100℅)
TIM 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
KF 3 (75℅)
0 (0℅)
1 (25℅)
4 (100℅)
FEP 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
AMC 3 (75℅)
0 (0℅)
1 (25℅)
4 (100℅)
AMX 3 (75℅)
0 (0℅)
1 (25℅)
4 (100℅)
AMT 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
FOX 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
CAZ 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
CTX 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
TIC 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
CIP 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
NA 2 (50℅)
0 (0℅)
2 (50℅)
4 (100℅)
KA 0 (0℅)
0 (0℅)
4 (100℅)
4 (100℅)
IMP 0 (0℅)
0 (0℅)
4 (100℅)
4 (100℅)
GN 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
CTX:Céfotaxine ; FOX:Cefotaxime ; CF:Céfalotine ; FEP:Péfloxacine ; PRL:Pipéraciline,
TZP:Pipéraciline+Tazobactam;CAZ:Ceftazidine;ATM:Aztreonam ;TIC:Ticarciline ;
TIM:Ticaciline+Acide Clavulanique ; AMX:Amoxicilline ; AMC:Amoxicilline+Acide
Clavulanique ; IMP:Imipinème ; GN;Gentamicine ; AK:Amikacine ; CIP:Ciprofloxacine
NA : Acide Nalidixique.
E. c : E coli ; S : Sensible ; R : Résistance ; I : Intermédiaire

Annexe 2
Enterobacter cloacae
ATB R I S TOTAL
PRL 2 (66.66℅)
0 (0℅)
1 (33.33℅)
3 (100℅)
TZP 2 (66.66℅)
0 (0℅)
2 (66.66℅)
3 (100℅)
TIM 2 (66.66℅)
0 (0℅)
1 (33.33℅)
3 (100℅)
KF 2 (66.66℅)
0 (0℅)
1 (33.33℅)
3 (100℅)
FEP 0 (0℅)
0 (0℅)
3 (100℅)
3 (100℅)
AMC 2 (66.66℅)
0 (0℅)
1 (33.33℅)
3 (100℅)
AMX 2 (66.66℅)
0 (0℅)
1 (33.33℅)
3 (100℅)
AMT 2 (66.66℅)
0 (0℅)
1 (33.33℅)
3 (100℅)
FOX 2 (66.66℅)
0 (0℅)
1 (33.33℅)
3 (100℅)
CAZ 2 (66.66℅)
0 (0℅)
1(33.33℅) 3 (100℅)
CTX 2 (66.66℅)
0 (0℅)
1 (33.33℅)
3 (100℅)
TIC 2 (66.66℅)
0 (0℅)
1 (33.33℅)
3 (100℅)
CIP 1 (33.33℅)
0 (0℅)
2 (66.66℅)
3 (100℅)
NA 2 (66.66℅)
0(0℅) 1 (33.33℅)
3 (100℅)
KA 1 (33.33℅)
0(0℅) 2 (66.66℅)
3 (100℅)
IMP 0 (0℅)
0(0℅) 3 (100℅)
3 (100℅)
GN 1 (33.33℅)
0(0℅) 2 (66.66℅)
3 (100℅)
CTX:Céfotaxine ; FOX:Cefotaxime ; CF:Céfalotine ; FEP:Péfloxacine ; PRL:Pipéraciline,
TZP:Pipéraciline+Tazobactam;CAZ:Ceftazidine;ATM:Aztreonam ;TIC:Ticarciline ;
TIM:Ticaciline+Acide Clavulanique ; AMX:Amoxicilline ; AMC:Amoxicilline+Acide
Clavulanique ; IMP:Imipinème ; GN;Gentamicine ; AK:Amikacine ; CIP:Ciprofloxacine
NA : Acide Nalidixique.
E. c : E coli ; S : Sensible ; R : Résistance ; I : Intermédiaire

Annexe 2
Proteus mirabilis
ATB R I S TOTAL
TZP 0 (0℅)
2 (33.33℅)
2 (33.33℅)
6 (100℅)
PRL 3 (50℅)
1 (33.33℅)
2 (33.33℅)
6 (100℅)
TIM 2 (33.33℅)
1 (33.33℅)
3 (50℅)
6 (100℅)
KF 3 (50℅)
0 (0℅)
3 (50℅)
6 (100℅)
FEP 2 (33.33℅)
0 (0℅)
4 (66.66℅)
6 (100℅)
AMC 3 (50℅)
0 (0℅)
3 (50℅)
6 (100℅)
AMX 4 (66.66℅)
0 (0℅)
2 (33.33℅)
6 (100℅)
AMT 2 (33.33℅)
0 (0℅)
4 (66.66℅)
6 (100℅)
FOX 2 (33.33℅)
0 (0℅)
4 (66.66℅)
6 (100℅)
CAZ 2 (33.33℅)
0 (0℅)
4 (66.66℅)
6 (100℅)
CTX 2 (33.33℅)
0 (0℅)
4 (66.66℅)
6 (100℅)
TIC 4 (66.66℅)
0 (0℅)
2 (33.33℅)
6 (100℅)
CIP 3 (50℅)
1 (16.66℅)
2 (33.33℅)
6 (100℅)
NA 4 (66.66℅)
1 (16.66℅)
2 (33.33℅)
6 (100℅)
KA 2 (33.33℅)
0 (0℅)
4 (66.66℅)
6 (100℅)
GN 2 (33.33℅)
0 (0℅)
4 (66.66℅)
6 (100℅)
IMP 0 (0℅)
0 (0℅)
6 (100℅)
6 (100℅)
CTX:Céfotaxine ; FOX:Cefotaxime ; CF:Céfalotine ; FEP:Péfloxacine ; PRL:Pipéraciline,
TZP:Pipéraciline+Tazobactam;CAZ:Ceftazidine;ATM:Aztreonam ;TIC:Ticarciline ;
TIM:Ticaciline+Acide Clavulanique ; AMX:Amoxicilline ; AMC:Amoxicilline+Acide
Clavulanique ; IMP:Imipinème ; GN;Gentamicine ; AK:Amikacine ; CIP:Ciprofloxacine
NA : Acide Nalidixique.
E. c : E coli ; S : Sensible ; R : Résistance ; I : Intermédiaire

Annexe 2
Aeromonas hydrophila
ATB R I S TOTAL
PRL 0 (0℅)
0 (100℅)
1 (100℅)
1 (100℅)
TZP 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
TIM 0 (0℅)
1 (100℅)
0 (0℅)
1 (100℅)
KF 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
FEP 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
AMC 0 (0℅)
1 (100℅)
0 (0℅)
1 (100℅)
AMX 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
AMT 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
FOX 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
CAZ 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
CTX 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
TIC 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
CIP 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
NA 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
KA 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
GN 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
IMP 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
CTX:Céfotaxine ; FOX:Cefotaxime ; CF:Céfalotine ; FEP:Péfloxacine ; PRL:Pipéraciline,
TZP:Pipéraciline+Tazobactam;CAZ:Ceftazidine;ATM:Aztreonam ;TIC:Ticarciline ;
TIM:Ticaciline+Acide Clavulanique ; AMX:Amoxicilline ; AMC:Amoxicilline+Acide
Clavulanique ; IMP:Imipinème ; GN;Gentamicine ; AK:Amikacine ; CIP:Ciprofloxacine
NA : Acide Nalidixique.
E. c : E coli ; S : Sensible ; R : Résistance ; I : Intermédiaire

Annexe 2
Acinetobacter baumanii
ATB R I S TOTAL
PRL 2 (50℅)
0 (0℅)
3 (75℅)
4 (100℅)
TZP 0 (0℅)
1 (25℅)
3 (75℅)
4 (100℅)
TIM 1 (25℅)
1 (25℅)
2 (50℅)
4 (100℅)
KF 4 (100℅)
0 (0℅)
0 (0℅)
4 (100℅)
FEP 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
AMC 4 (100℅)
0 (0℅)
0 (0℅)
4 (100℅)
AMX 4 (100℅)
0 (0℅)
0 (0℅)
4 (100℅)
AMT 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
FOX 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
CAZ 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
CTX 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
TIC 2 (50℅)
0 (0℅)
2 (50℅)
4 (100℅)
CIP 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
NA 2 (50℅)
0 (0℅)
2 (50℅)
4 (100℅)
KA 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
GN 1 (25℅)
0 (0℅)
3 (75℅)
4 (100℅)
IMP 0 (0℅)
0 (0℅)
4 (100℅)
4 (100℅)
CTX:Céfotaxine ; FOX:Cefotaxime ; CF:Céfalotine ; FEP:Péfloxacine ; PRL:Pipéraciline,
TZP:Pipéraciline+Tazobactam;CAZ:Ceftazidine;ATM:Aztreonam ;TIC:Ticarciline ;
TIM:Ticaciline+Acide Clavulanique ; AMX:Amoxicilline ; AMC:Amoxicilline+Acide
Clavulanique ; IMP:Imipinème ; GN;Gentamicine ; AK:Amikacine ; CIP:Ciprofloxacine
NA : Acide Nalidixique.
E. c : E coli ; S : Sensible ; R : Résistance ; I : Intermédiaire

Annexe 2
Providencia stuartii
ATB R I S TOTAL
PRL 0 (0℅)
0 (100℅)
1 (100℅)
1 (100℅)
TZP 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
TIM 0 (0℅)
1 (100℅)
0 (0℅)
1 (100℅)
KF 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
FEP 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
AMC 0 (0℅)
1 (100℅)
0 (0℅)
1 (100℅)
AMX 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
AMT 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
FOX 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
CAZ 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
CTX 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
TIC 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
CIP 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
NA 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
KA 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
GN 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
IMP 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
CTX:Céfotaxine ; FOX:Cefotaxime ; CF:Céfalotine ; FEP:Péfloxacine ; PRL:Pipéraciline,
TZP:Pipéraciline+Tazobactam;CAZ:Ceftazidine;ATM:Aztreonam ;TIC:Ticarciline ;
TIM:Ticaciline+Acide Clavulanique ; AMX:Amoxicilline ; AMC:Amoxicilline+Acide
Clavulanique ; IMP:Imipinème ; GN;Gentamicine ; AK:Amikacine ; CIP:Ciprofloxacine
NA : Acide Nalidixique.
E. c : E coli ; S : Sensible ; R : Résistance ; I : Intermédiaire

Annexe 2
Citrobacter freundii
ATB R I S TOTAL
PRL 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
TZP 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
TIM 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
KF 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
FEP 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
AMC 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
AMX 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
AMT 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
FOX 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
CAZ 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
CTX 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
TIC 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
CIP 1 (0℅)
0 (0℅)
0 (0℅)
1 (100℅)
NA 1 (0℅)
0 (0℅)
0 (0℅)
1 (100℅)
KA 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
GN 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
IMP 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
CTX:Céfotaxine ; FOX:Cefotaxime ; CF:Céfalotine ; FEP:Péfloxacine ; PRL:Pipéraciline,
TZP:Pipéraciline+Tazobactam;CAZ:Ceftazidine;ATM:Aztreonam ;TIC:Ticarciline ;
TIM:Ticaciline+Acide Clavulanique ; AMX:Amoxicilline ; AMC:Amoxicilline+Acide
Clavulanique ; IMP:Imipinème ; GN;Gentamicine ; AK:Amikacine ; CIP:Ciprofloxacine
NA : Acide Nalidixique.
E. c : E coli ; S : Sensible ; R : Résistance ; I : Intermédiaire

Annexe 2
Serratia marecesens
ATB R I S TOTAL
PRL 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
TZP 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
TIM 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
KF 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
FEP 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
AMC 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
AMX 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
AMT 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
FOX 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
CAZ 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
CTX 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
TIC 1 (100℅)
0 (0℅)
0 (0℅)
1 (100℅)
CIP 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
NA 1 (0℅)
0 (0℅)
0 (0℅)
1 (100℅)
KA 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
GN 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
IMP 0 (0℅)
0 (0℅)
1 (100℅)
1 (100℅)
CTX:Céfotaxine ; FOX:Cefotaxime ; CF:Céfalotine ; FEP:Péfloxacine ; PRL:Pipéraciline,
TZP:Pipéraciline+Tazobactam;CAZ:Ceftazidine;ATM:Aztreonam ;TIC:Ticarciline ;
TIM:Ticaciline+Acide Clavulanique ; AMX:Amoxicilline ; AMC:Amoxicilline+Acide
Clavulanique ; IMP:Imipinème ; GN;Gentamicine ; AK:Amikacine ; CIP:Ciprofloxacine
NA : Acide Nalidixique.
E. c : E coli ; S : Sensible ; R : Résistance ; I : Intermédiaire

Annexe 2
Klebsiella oxytoca
ATB R I S TOTAL
TZP 0 (0℅)
1 (33.33℅)
2 (66.66℅)
3 (100℅)
PRL 2 (66.66℅)
0 (0℅)
1 (33.33℅)
3 (100℅)
TIM 0 (0℅)
1 (33.33℅)
2 (66.66℅)
3 (100℅)
KF 3 (100℅)
0 (0℅)
0 (0℅)
3 (100℅)
FEP 1 (33.33℅)
0 (0℅)
2 (66.66℅)
3 (100℅)
AMC 2 (66.66℅)
0 (0℅)
1 (33.33℅)
3 (100℅)
AMX 3 (100℅)
0 (0℅)
0 (0℅)
3 (100℅)
AMT 1 (33.33℅)
0 (0℅)
2 (66.66℅)
3 (100℅)
FOX 1 (33.33℅)
0 (0℅)
2 (66.66℅)
3 (100℅)
CAZ 1 (33.33℅)
0 (0℅)
2 (66.66℅)
3 (100℅)
CTX 1 (33.33℅)
0 (0℅)
2 (66.66℅)
3 (100℅)
TIC 2 (66.66℅)
0 (0℅)
1 (33.33℅)
3 (100℅)
CIP 1 (33.33℅)
1 (33.33℅)
1 (33.33℅)
3 (100℅)
NA 2 (66.66℅)
1 (33.33℅)
0 (0℅)
3 (100℅)
KA 1 (33.33℅)
1 (33.33℅)
1 (33.33℅)
3 (100℅)
GN 1 (33.33℅)
1 (33.33℅)
1 (33.33℅)
3 (100℅)
IMP 0 (0℅)
0 (0℅)
3 (100℅)
3 (100℅)
CTX:Céfotaxine ; FOX:Cefotaxime ; CF:Céfalotine ; FEP:Péfloxacine ; PRL:Pipéraciline,
TZP:Pipéraciline+Tazobactam;CAZ:Ceftazidine;ATM:Aztreonam ;TIC:Ticarciline ;
TIM:Ticaciline+Acide Clavulanique ; AMX:Amoxicilline ; AMC:Amoxicilline+Acide
Clavulanique ; IMP:Imipinème ; GN;Gentamicine ; AK:Amikacine ; CIP:Ciprofloxacine
NA : Acide Nalidixique.
E. c : E coli ; S : Sensible ; R : Résistance ; I : Intermédiaire

Annexe 2
Klebsiella pneumoniae
ATB R I S TOTAL
TZP 0 (0℅)
0 (0℅)
16 (100℅)
16 (100℅)
PRL 15 (93.75℅)
1 (6.25℅)
0 (0℅)
16 (100℅)
TIM 9 (56.5℅)
1 (6.25℅)
5 (31.25℅)
16 (100℅)
KF 13 (81.25℅)
0 (0℅)
3 (18.75℅)
16 (100℅)
FEP 12 (75℅)
0 (0℅)
3 (18.75℅)
16 (100℅)
AMC 10 (62.5℅)
0 (0℅)
6 (37.5℅)
16 (100℅)
AMX 15 (93.75℅)
0 (0℅)
1 (6.25℅)
16 (100℅)
AMT 12 (75℅)
0 (0℅)
4 (25℅)
16 (100℅)
FOX 5 (31.25℅)
0 (0℅)
11 (68.75℅)
16 (100℅)
CAZ 12 (75℅)
0 (0℅)
4 (25℅)
16 (100℅)
CTX 12 (75℅)
0 (0℅)
4 (25℅)
16 (100℅)
TIC 15 (93.75℅)
0 (0℅)
1 (6.25℅)
16 (100℅)
CIP 3 (18.75℅)
0 (0℅)
13 (81.25℅)
16 (100℅)
NA 5 (31.25℅)
0 (0℅)
11 (68.75)
16 (100℅)
KA 2 (12.5℅)
0 (0℅)
14 (87.5℅)
16 (100℅)
GN 2 (12.5℅)
0 (0℅)
14 (87.5℅)
16 (100℅)
IMP 0 (0℅)
0 (0℅)
16 (100℅)
16 (100℅)
CTX:Céfotaxine ; FOX:Cefotaxime ; CF:Céfalotine ; FEP:Péfloxacine ; PRL:Pipéraciline,
TZP:Pipéraciline+Tazobactam;CAZ:Ceftazidine;ATM:Aztreonam ;TIC:Ticarciline ;
TIM:Ticaciline+Acide Clavulanique ; AMX:Amoxicilline ; AMC:Amoxicilline+Acide
Clavulanique ; IMP:Imipinème ; GN;Gentamicine ; AK:Amikacine ; CIP:Ciprofloxacine
NA : Acide Nalidixique.
E. c : E coli ; S : Sensible ; R : Résistance ; I : Intermédiaire

Annexe 3
Pourcentage de résistance des entérobactéries aux antibiotiques
Espèce TZP PRL TIM KF FEP IMP AMC AMX ATM FOX CAZ TIC CTX CIP NA GN AK
E. coli 24,09 61.44 39,75 79.51 32.53 0 62.65
83.13 56.62 34.93 56.62 75.90 56.62 20.48 39.75
19.27
18,07
Enterobacter sakazakic 0 25 25 75 25 0 75 75 25 25 25 25 25 25 50 25 0
Enterobacter cloacae 66,66 66,66 66,66 66,66 0 0 66,66 66,66 66,66 66,66 66,66 66,66 66,66 33,33 66,66 33,33 33,33
Proteus mirabilis 0 50 33,33 50 33.33 0 50 66,66 33,33 33,33 33,33 66,66 33,33 50 66,66 33,33 33,33
Acinetobacter
baumanii
0 50 25 100 25 0 100 100 25 25 25 50 25 25 50 25 25
Providencia stuartii 0 100 0 0 0 0 0 100 0 0 0 100 0 0 0 0 0
Aeromonas hydrophila 0 100 0 0 0 0 0 100 0 0 0 100 0 0 0 0 0
Citrobacter freundii 0 100 100 100 0 0 100 100 100 0 100 100 100 100 100 0 0
Serratia marecesens 0 100 100 100 0 0 100 100 100 0 100 100 100 0 100 0 0
Klebsiella oxytoca 0 66.66 0 100 33,33 0 66,66 100 33,33 33,33 33,33 66,66 33,33 33,33 66,66 33,33 33,33
Klebsiella pneumoniae 0 93,75 56,25 81,25 75 0 62,5 93,75 75 31,25 75 93,75 68.75 18,75 31,25 12,5 12,5
CTX:Céfotaxine ; FOX:Cefotaxime ; CF:Céfalotine ; FEP:Péfloxacine ; PRL:Pipéraciline, TZP:Pipéraciline+Tazobactam ; CAZ:Ceftazidine ;
ATM:Aztreonam ; TIC:Ticarciline ;TIM:Ticaciline+Acide Clavulanique ; AMX:Amoxicilline ; AMC:Amoxicilline+Acide Clavulanique ;
IMP:Imipinème ; GN;Gentamicine ; AK:Amikacine ; CIP:Ciprofloxacine NA : Acide Nalidixique.

Annexe 4
Phénotype de résistance d’E. coli
Code TZP PRL TIM KF FEP IMP AMC AMX ATM FOX CAZ TIC CTX CIP NA GN KA Phénotype
E .c 01 S R R R R S R R
R
S R R R S S S I BLSE
E .c 02 S R R R R S R R R S R R R S R S S BLSE
E .c 03 S R S R R S S R R R R R R S S R R BLSE
E .c04 S R S R R S S R R S R R R S S S S BLSE
E .c 05 S R R R R S R R R S R R R R R R R BLSE
E .c 06 S R S R R S S R R R R R R S S R R BLSE
E .c 08 S R R R R S R R R S R R R S R R S BLSE
E .c 09 S R S R R S S R R S R R R S R S R BLSE
E .c 10 S R S R R S S R R S R R R R R S R BLSE
E .c 11 S R S R R S S R R S R R R S S S S BLSE
E .c 12 S R S R R S S R R S R R R S R S S BLSE
E .c 14 S R S R R S S R R S R R R S R S R BLSE
E .c 15 S R R R R S R R R S R R R R R R R BLSE
E .c 16 S R S R R S S R R S R R R R R S S BLSE
E .c 17 S R R R R S R R R S R R R R S S R BLSE

Annexe 4
E .c 18 S R R R R S R R R S R R R R S S R BLSE
E .c 19 S R R R R S R R R R R R R S R S S BLSE
E .c 20 S R S R R S S R R S R R R S S S S BLSE
E .c 21 S R S R R S S R R S R R R S S S S BLSE
E .c 22 S R S R R S S R R R R R R S R R S BLSE
E .c 23 S R R R R S R R R R R R R R S S S BLSE
E .c 24 S R R R R S R R R S R R R S R S I BLSE
E .c 25 S R R R R S R R R S R R R R S S S BLSE
E .c 26 S R R R R S R R R R R R R S S S I BLSE
E .c 27 S R R R R S R R R S R R R S R S S BLSE
E .c 28 S R S R R S S R R S R R R S R S S BLSE
E .c 29 S R S R R S S R R S R R R S R S R BLSE
E .c 30 R R R R S S R R R R R R R S R R R Case
E .c 31 R R R R S S R R R R R R R R R R R Case
E .c 32 S S S R S S R R S R S S S S S S S Case
E .c33 S S S R S S R R S R S S S S S S S Case
E .c 34 R R R R S S R R R R R R R S R S S Case
E .c 35 R R R R S S R R R R R R R S R S S Case

Annexe 4
E .c 36 R R R R S S R R R R R R R S R S S Case
E .c37 R R R R S S R R R R R R R S S S S Case
E .c 38 R R R R S S R R R R R R R R R R R Case
E .c 39 R R R R S S R R R R R R R R R S S Case
E .c 40 S S S R S S R R S R S S S S S S S Case
E .c 41 R R R R S S R R R R R R R S R S S Case
E .c 42 S S S R S S R R S R S S S S S S S Case
E .c 43 S S S R S S R R S R S S S S S S S Case
E .c 44 R R R R S S R R R R R R R R R S S Case
E .c 45 R R R R S S R R R R R R R R R R S Case
E .c 46 R R R R S S R R R R R R R S R R S Case
E .c 47 R R R R S S R R R R R R R S S S S Case
E .c 48 R R R R S S R R R S R R R S R R S Case
E .c 49 R R R R S S R R R S R R R R R R S Case
E .c 50 R R R R S S R R R S R R R R R R S Case
E .c 51 S S S R S S R R S R S S S S S S S Case
E .c 52 R R R R S S R R R R R R R S R R S Case
E .c 53 R R R R S S R R R R R R R R R S S Case

Annexe 4
E .c 54 R R R R S S R R R R R R R R R R R Case
E .c 55 R R R R S S R R R R R R R R R R R Case
E .c 56 S R S S S S S R S S S R S S S S S Pase
E .c 57 S R S S S S S R S S S R S S S S S Pase
E .c 58 S I I R S S R R S S S R S S S S S Pase
E .c59 S I I R S S R R S S S R S S S S S Pase
E .c 60 S I I R S S R R S S S R S S S S S Pase
E .c 61 S I I R S S R R S S S R S S S S S Pase
E .c 62 S I I R S S R R S S S R S S S S S Pase
E .c 63 S I I R S S R R S S S R S S S S S Pase
E .c 64 S I I R S S R R S S S R S S S S S Pase
E .c 65 S I I R S S R R S S S R S S S S S Pase
E .c 66 S R S S S S S R S S S R S S S S S Pase
E .c67 S I I R S S R R S S S R S S S S S Pase
E .c 68 S I I R S S R R S S S R S S S S S Pase
E .c 69 S I I R S S R R S S S R S S S S S Pase
E .c 70 S I I R S S R R S S S R S S S S S Pase
E.c 71 S R S S S S S R S S S R S S S S S Pase

Annexe 4
E .c 72 S S S S S S S S S S S S S S S S S Sensible
E .c 73 S S S S S S S S S S S S S S S S S Sensible
E .c 74 S S S S S S S S S S S S S S S S S Sensible
E .c 75 S S S S S S S S S S S S S S S S S Sensible
E .c 76 S S S S S S S S S S S S S S S S S Sensible
E .c 77 S S S S S S S S S S S S S S S S S Sensible
E .c 78 S S S S S S S S S S S S S S S S S Sensible
E .c 79 S S S S S S S S S S S S S S S S S Sensible
E .c 80 S S S S S S S S S S S S S S S S S Sensible
E .c 81 S S S S S S S S S S S S S S S S S Sensible
E .c 82 S S S S S S S S S S S S S S S S S Sensible
E .c 83 S S S S S S S S S S S S S S S S S Sensible
E .c 84 S S S S S S S S S S S S S S S S S Sensible
E. c 85 S S S S S S S S S S S S S S S S S Sensible
CTX:Céfotaxine ; FOX:Cefotaxime ; CF:Céfalotine ; FEP:Péfloxacine ; PRL:Pipéraciline, TZP:Pipéraciline+Tazobactam ; CAZ:Ceftazidine ;
ATM:Aztreonam ; TIC:Ticarciline ;TIM:Ticaciline+Acide Clavulanique ; AMX:Amoxicilline ; AMC:Amoxicilline+Acide Clavulanique ;
IMP:Imipinème ; GN;Gentamicine ; AK:Amikacine ; CIP:Ciprofloxacine NA : Acide Nalidixique.
E c : E coli ; S : sensible ; R :résistance ;BLSE: β-lactamase à spectre étendu ;Case :Céphalosporinase ;Pase :pénicillinase

Annexe 5
Données sur les patients infectés par E. coli
Code Service Date de
prélèvement
Sexe
Age Durée
d'hospitalisation
Antécédents
d'hospitalisation Motifs d'hospitalisation
Antécédents
d’infection
urinaire
Acte
adhésif
E .c 01 Maternité avr-2011 Femme 35 7 nom Anémie nom voie
veineuse
E .c 02 Maternité févr-2013 Femme 22 8 oui 2fois'maternité' HTA +enceinte'7 mois' oui nom
E .c 03 Maternité avr-2012 Femme 31 10 oui 3 fois 'maternité' MAP+enceinte'6mois' nom nom
E .c04 Maternité mars-2013 Femme 37 15 oui 2 fois 'maternité' HTA +enceinte'9 mois' oui nom
E .c 05 Maternité avr-2013 Femme 38 14 nom Problème
rénale+enceinte'8mois' nom nom
E .c 06 Maternité mai-2013 Femme 30 10 nom MAP nom nom
E .c 08 Maternité juin-2013 Femme 31 14 oui 2 fois 'maternité' MAP+enceinte'9mois' oui nom
E .c 09 Urologie nov-2011 Homme 91 2 oui Rétrécissement urétral nom nom
E .c 10 Urologie févr-2011 Homme 57 3 oui problème rénale
oui voie
veineuse
E .c 11 Urologie juin-2013 Homme 69 2 nom Tumeur vésicale nom voie
veineuse
E .c 12 Urologie Nov-2011 Homme 63 2 nom Rétrécissement urétral oui nom
E .c 14 Urologie nov-2012 Homme 90 2 oui 3fois'urologie Rétrécissement urétral nom nom
E .c 15 Urologie mars-2013 Homme 42 3 non Tumeur vésicale nom nom
E .c 16 Médecine Interne janv-2012 Femme 95 10 oui 2fois'urologie Diabète nom Nom
E .c 17 Médecine Interne juin-2013 Femme 56 15 nom Hépatite+diabète oui nom
E .c 18 Médecine Interne juin-2012 Femme 65 20 Oui (maternité et
médecine interne) Diabète nom nom
E .c 19 Médecine Interne déc-2012 Femme 40 15 Oui (maternité) Diabète oui nom
E .c 20 Médecine Interne févr-2012 Femme 37 11 Oui (maternité et
médecine interne) Diabète nom
voie
veineuse
E .c 21 Médecine Interne nov-2011 Femme 83 22 Oui (maternité et Gangrène oui voie
Données sur les patients infectés par E coli

Annexe 5
Données sur les patients infectés par E. coli
médecine interne) veineuse
E .c 22 Médecine Interne Févr2013 Homme 35 8 Ou i(maternité ) Diabète nom nom
E .c 23 Urologie mai-2013 Homme 30 13 Oui (maternité) Tumeur Vésical nom nom
E .c 24 Maternité mars-2013 Femme 39 4 nom HTA+ grossesse nom nom
E .c 25 Médecine Interne juin-2013 Femme 55 20 nom Gangrène nom nom
E .c 26 Urologie juin-2013 Homme 58 3 nom Abcès Rénale nom voie
veineuse
E .c 27 Maternité mars-2013 Femme 35 10 Oui (maternité) Anémie nom nom
E .c 28 Urologie nov-2012 Femme 62 3 nom Tumeur prostate' 2 Ans' oui nom
E .c 29 Urologie avr-2013 homme 79 2 non Tumeur de Vessie oui nom
E .c 30 Maternité
16/01/2012 Femme 35 20 nom Map oui voie
veineuse
E .c 31 Maternité 11/04/2011 Femme 23 15 nom HTA nom nom
E .c 32 Maternité 2011 Femme 30 11 nom Cancer utérus nom nom
E .c33 Maternité
04/02/2013 Femme 35 2j oui Kyste d'ovaire nom voie
veineuse
E .c 34 Maternité
05/02/2013 Femme 27 10 oui HTA nom voie
veineuse
E .c 35 Maternité 20/02/2013 Femme 28 15 nom Map nom nom
E .c 36 Maternité 2013 Femme 21 5 nom Map nom nom
E .c37 Maternité 16/01/2012 Femme 23 21 nom Abcès rénale nom nom
E .c 38 Urologie 16/10/2011 Femme 40 15 nom Adénome prostate 4 ans nom voie
veineuse
E .c 39 Urologie 09/11/2011 Homme 70 2 nom Tumeur de vessie oui nom
E .c 40 Urologie 2013 Homme 69 2 nom Tumeur de vessie oui nom
E .c 41 Urologie 2013 Homme 82 2 oui Tumeur de vessie oui nom
E .c 42 Urologie 2012 Homme 42 3 nom Insuffisante rénale oui nom

Annexe 5
Données sur les patients infectés par E. coli
E .c 43 Urologie 05/06/2012 Femme 65 15 oui Gangrène oui voie
veineuse
E .c 44 Urologie 2012 Homme 44 20 nom Plait infecte nom nom
E .c 45 Médecines Interne 16/11/2012 Homme 30 10 nom Plait infecte nom nom
E .c 46 Médecines Interne 25/05/2012 Femme 70 9 oui Plait infecte oui nom
E .c 47 Médecines Interne 15/02/2012 Femme 72 21 oui Diabète oui nom
E .c 48 Médecines Interne 2013 Homme 1941 15 oui Diabète oui nom
E .c 49 Médecines Interne 2013 Femme 24 17 nom Plait infecte non nom
E .c 50 Médecines Interne 2013 Femme 36 20 nom Plait infecte nom nom
E .c 51 Médecines Interne 30/05/2012 Homme 26 10 nom HTA nom nom
E .c 52 Maternité
Mars-11 Femme 34 5 non Map non voie
veineuse
E .c 53 Maternité
Juin-13 Femme 28 10 oui 2 fois 'maternité' HTA oui voie
veineuse
E .c 54 Maternité
Mars-13 Femme 34 7 non Map oui voie
veineuse
E .c 55 Maternité Juin-13 Femme 28 15 oui 2 fois 'maternité' Diabète oui nom
E .c 56 Maternité
29/08/2012 Femme 35 15 nom Diabète nom voie
veineuse
E .c 57 Maternité 10/06/2013 Femme 38 4 oui Kyste d'ovaire nom nom
E .c 58 Maternité 15/02/2012 Femme 45 16 oui Map nom nom
E .c59 Maternité 08/03/2013 Femme 35 8 nom Traumatisme rénale nom nom
E .c 60 Urologie 31/05/2011 Homme 37 3 nom Rétrécissement urétral
nom voie
veineuse
E .c 61 Urologie 31/05/2012 Homme 50 4 oui Abcès rein oui voie
veineuse
E .c 62 Urologie 05/05/2013 Femme 36 6 oui Tumeur de vessie oui voie
veineuse

Annexe 5
Données sur les patients infectés par E. coli
E .c 63 Urologie 07/04/2013 Homme 39 5 nom Lithiase rénale nom voie
veineuse
E .c 64 Urologie 27/03/2012 Femme 36 3 oui Hépatite oui voie
veineuse
E .c 65 Médecines Interne 15/06/2013 Femme 23 15 nom Diabète oui nom
E .c 66 Médecines Interne 04/01/2011 Femme 79 16 oui Thyroïde oui nom
E .c67 Médecines Interne 30/05/2012 Femme 26 10 non HTA bom nom
E .c 68 Médecines Interne
08/05/2012 Homme 67 15 nom Diabète oui voie
veineuse
E .c 69 Médecines Interne
22/08/2013 Homme 83 20 nom Diabète oui voie
veineuse
E .c 70 Médecines Interne 17/05/2013 Homme 77 17 nom Anémié oui nom
E.c 71 Maternité 10/04/2013 Femme 36 15 oui Diabète oui nom
E .c 72 Maternité
22/04/2011 Femme 33 11 oui HTA nom voie
veineuse
E .c 73 Maternité 12/05/2011 Femme 30 10 nom Diabète nom nom
E .c 74 Maternité 29/05/2011 Femme 35 6 oui HTA nom nom
E .c 75 Maternité 10/05/2011 Femme 25 12 oui Tumeur de vessie nom nom
E .c 76 Urologie 17/04/2013 Homme 61 2 nom Adénome prostate
nom voie
veineuse
E .c 77 Urologie 21/11/2011
Homme 90 2 nom Lithiase rénale nom voie
veineuse
E .c 78 Urologie 04/02/2012 Femme 45 4 oui Tumeur de vessie oui voie
veineuse
E .c 79 Urologie 07/03/2011 Homme 65 2 nom Lithiase rénale nom voie
veineuse
E .c 80 Urologie 05/08/2013 Femme 45 4 nom Diabète nom voie
veineuse
E .c 81 Médecines Interne 04/02/2011 Femme 54 10 oui Diabète oui voie

Annexe 5
Données sur les patients infectés par E. coli
MAP : accouchement prématuré
HTA : hypertension artérielle
Ec : E coli
veineuse
E .c 82 Médecines Interne
12/12/2013 Femme 31 7 nom Diabète nom voie
veineuse
E .c 83 Médecines Interne 08/02/2012 Femme 31 9 oui Diabète nom nom
E .c 84 Médecines Interne 16/03/2012 Homme 31 7 nom Diabète nom nom
E. c 85 Médecines Interne 15/05/2011 Homme 47 5 nom Map nom nom

Annexe 6
Caractéristiques d’E. coli BLSE
Code Gène de BLSE
Gène de
resistance aux
β-lactamine
Gène de resistance
aux quinolone
Gène de
resistance aux
aminosides
Genes de
virulence
groupement
phylogenetique ERIC
profile
E .c06 blaCTX-M-15 blaTEM-1 armA A0 c
E.c12 blaCTX-M-15 blaTEM-1 A0 f
E .c22 blaCTX-M-15 blaTEM-1 A0 j
E.c24 blaCTX-M-15 blaTEM-1 A0 e
E. c26 blaCTX-M-15 blaTEM-1, blaCMY-2 A0 p
E.c27 blaCTX-M-15 blaTEM-1 qnrB1 A0 o
E. c 01 blaCTX-M-15 blaTEM-1 aac(6')-Ib-cr A1 a
E.c02 blaCTX-M-15 blaTEM-1 qnrA1, aac(6')-Ib-cr A1 q
E .c04 blaCTX-M-15 blaTEM-1 qnrB1, aac(6')-Ib-cr pap A1 c
E.c05 blaCTX-M-15, blaCTX-M-14 blaTEM-1 aac(6')-Ib-cr sfa A1 c
E.c08 blaCTX-M 15 blaTEM-1 aac(6')-Ib-cr armA ,rmtB sfa A1 e
E.c09 blaCTX-M-15 blaTEM-1 aac(6')-Ib-cr armA,rmtB sfa A1 f
E.c14 blaCTX-M-15 blaTEM-1 aac(6')-Ib-cr sfa, A1 g
E.c18 blaCTX-M-15 blaTEM-1 qnrB1 A1 c
E.c23 blaCTX-M-15 blaTEM-1 aac(6')-Ib-cr A1 k
E.c03 blaCTX-M-15, blaCTX-M-14 blaTEM-1 B1 b
E.c10 blaCTX-M-15 blaTEM-1 pap B1 f
E.c16 blaCTX-M-15 blaTEM-1 aac(6')-Ib-cr B1 h
E.c19 blaCTX-M-15 blaTEM-1, blaMOX pap B1 h
E.c25 blaCTX-M-15 blaTEM-1 B1 i
E.c20 blaCTX-M-1 blaTEM-1 pap B22 h
E.c11 blaCTX-M-15 blaTEM-1 aac(6')-Ib-cr B23 f
E.c15 blaCTX-M-15 blaTEM-1 aac(6')-Ib-cr sfa B23 h
E.c17 blaCTX-M-15 armA pap B23 h
E.c21 blaCTX-M-28 blaTEM-1 sfa,cf1 B23 i
E.c29 blaCTX-M-15, blaSHV-12 blaTEM-1 aac(6')-Ib-cr sfa,pap B23 n
E.c28 blaCTX-M-15 blaTEM-1 qnrB1, aac(6')-Ib-cr D1 e

Annexe7
Phénotype de résistance des Entérobactéries
Espece TZP PRL TIM KF FEP IMP AMC AMX ATM FOX CAZ TIC CTX CIP NA GN KA Phénotype
Enterobacter sakazakic I R R R R S R R R R R R R R R R S BLSE
Enterobacter sakazakic S S S R S S R R S S S S S S R S S CASE
Enterobacter sakazakic S S S R S S R R S S S S S S S S S CASE
Enterobacter sakazakic S S S S S S S S S S S S S S S S S Sensible
Enterobacter cloacae R R R R S S R R R R R R R R R R R CASE
Enterobacter cloacae R R R R S S R R R R R R R S R S S CASE
Enterobacter cloacae S S S S S S S S S S S S S S S S S Sensible
Proteus mirabilis I R R R R S R R R R R R R R R R R BLSE
Proteus mirabilis I R R R R S R R R R R R R R R R R BLSE
Proteus mirabilis S I I R S S R R S S S R S R R S S PASE
Proteus mirabilis S R S S S S S R S S S R S S R S S PASE
Proteus mirabilis S S S S S S S S S S S S S S S S S Sensible
Proteus mirabilis S S S S S S S S S S S S S S S S S Sensible
Acinetobacter baumanii S R R R R S R R R R R R R R R R R BLSE
Acinetobacter baumanii S S S R S S R R S S S S S S R S S CASE
Acinetobacter baumanii S S S R S S R R S S S S S S S S S CASE
Acinetobacter baumanii I R I R S S R R S S S R S S S S S PASE
Providencia stuartii S R S S S S S R S S S R S S S S S PASE
Aeromonas hydrophila S R S S S S S R S S S R S S S S S PASE
Citrobacter freundii S R R R R S R R R S R R R R R S S BLSE
Serratia marecesens S R R R R S R R R S R R R S R S S BLSE
Klebsiella oxytoca S R S R R S S R R R R R R R R R R BLSE
Klebsiella oxytoca S S S R S S R R S S S S S S R S S CASE
Klebsiella oxytoca S R I R S S R R S S S R S I I I I PASE
Klebsiella pneumoniae S R R R R S R R R R R R R R R R R BLSE
Klebsiella pneumoniae S R R R R S R R R R R R R R R R R BLSE
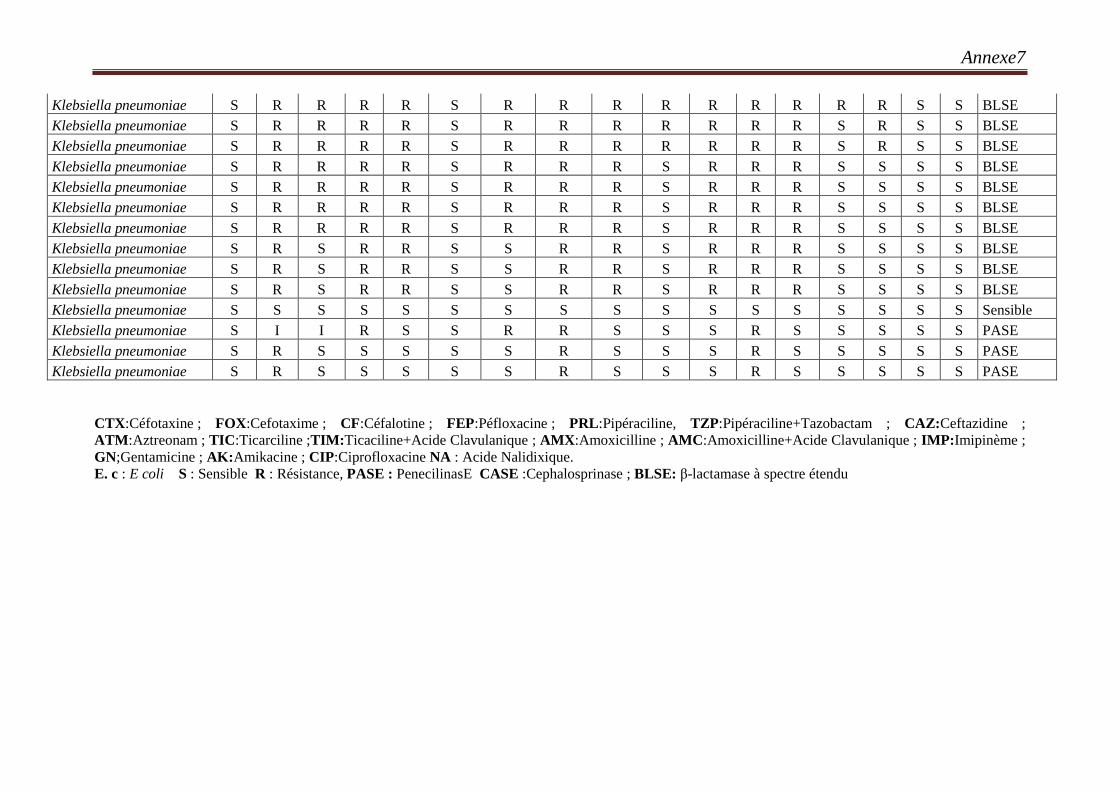
Annexe7
Klebsiella pneumoniae S R R R R S R R R R R R R R R S S BLSE
Klebsiella pneumoniae S R R R R S R R R R R R R S R S S BLSE
Klebsiella pneumoniae S R R R R S R R R R R R R S R S S BLSE
Klebsiella pneumoniae S R R R R S R R R S R R R S S S S BLSE
Klebsiella pneumoniae S R R R R S R R R S R R R S S S S BLSE
Klebsiella pneumoniae S R R R R S R R R S R R R S S S S BLSE
Klebsiella pneumoniae S R R R R S R R R S R R R S S S S BLSE
Klebsiella pneumoniae S R S R R S S R R S R R R S S S S BLSE
Klebsiella pneumoniae S R S R R S S R R S R R R S S S S BLSE
Klebsiella pneumoniae S R S R R S S R R S R R R S S S S BLSE
Klebsiella pneumoniae S S S S S S S S S S S S S S S S S Sensible
Klebsiella pneumoniae S I I R S S R R S S S R S S S S S PASE
Klebsiella pneumoniae S R S S S S S R S S S R S S S S S PASE
Klebsiella pneumoniae S R S S S S S R S S S R S S S S S PASE
CTX:Céfotaxine ; FOX:Cefotaxime ; CF:Céfalotine ; FEP:Péfloxacine ; PRL:Pipéraciline, TZP:Pipéraciline+Tazobactam ; CAZ:Ceftazidine ;
ATM:Aztreonam ; TIC:Ticarciline ;TIM:Ticaciline+Acide Clavulanique ; AMX:Amoxicilline ; AMC:Amoxicilline+Acide Clavulanique ; IMP:Imipinème ;
GN;Gentamicine ; AK:Amikacine ; CIP:Ciprofloxacine NA : Acide Nalidixique.
E. c : E coli S : Sensible R : Résistance, PASE : PenecilinasE CASE :Cephalosprinase ; BLSE: β-lactamase à spectre étendu

Annexe8
Chromatogramme CPG/SM de l'HE brute de Ziziphora hispanica
Chromatogramme CPG/SM de l'HE brute de Coriandrum sativum
Chromatogramme CPG/SM de l'HE brute de Cinnamomum cassia

Annexe8

Annexe 9
Tampon et milieux de culture
1- Les tampons
Tampon TBE 1 X
Tris base…………………………………………………………10,78g
Acide borique………………………………….. ………………..5,50 g
EDTA 0,5 M ……………………………………………………. 0,58 g
Eau distillée …………………………………………………….1 litre
Tampon de charge
Glycérol …………………………………………………………...3 ml
Bleu de bromophénol ……………………………………………75 mg
Eau distillée ……………………………………………………….7 ml
2-Les milieux de culture :
Gélose nutritif :
Pour 1 litre de milieu :
Tryptone...........................................................................................5,0 g
Extrait de viande ..............................................................................3,0 g
Agar agar bactériologique..............................................................12,0 g
pH du milieu prêt-à-l’emploi à 25°C : 7,0 ± 0,2.
Muller Hinton
Pour 1 litre de milieu :
Hydrolysât acide de caséine ..........................................................17,5 g
Infusion de viande............................................................................2,0 g
Amidon soluble ................................................................................1,5 g
Agar agar bactériologique..............................................................17,0 g
pH du milieu prêt-à-l’emploi à 25°C : 7,3 ± 0,2.
Bouillon cœur-cervelle
Pour 1 litre de milieu :

Annexe 9
Extrait cœur-cervelle......................................................................17,5 g
Peptone pancréatique de gélatine ...................................................10,0 g
Chlorure de sodium..........................................................................05,0 g
Phosphate disodique .........................................................................2,5 g
Glucose..............................................................................................2,0 g
pH du milieu prêt-à-l’emploi à 25°C : 7,4 ± 0,2.
2- Gélose de MacConkey
Pour 1 litre de milieu :
Peptone pancréatique de gélatine ...................................................17,0 g
Tryptone...........................................................................................1,5 g
Peptone pepsique de viande ...........................................................1,5 g
Lactose ..........................................................................................10,0 g
Sels biliaires.....................................................................................1,5 g
Chlorure de sodium..........................................................................5,0 g
Rouge neutre ..............................................................................30,0 mg
Cristal violet ..................................................................................1,0 mg
Agar agar bactériologique..............................................................13,5 g
pH du milieu prêt-à-l’emploi à 25°C : 7,1 ± 0,2.

Travaux
personnelle

Travaux personnelles
Travaux personnelles
Publications:
Fatima Zenati, Fethi Benbelaid, Abdelmounaim Khadir, Chafika Bellahsene, Mourad
Bendahou, 2014.Antimicrobial effects of three essential oils on multidrug resistant bacteria
responsible for urinary infections. Journal of Applied Pharmaceutical Science Vol. 4 (11), pp.
015-018
Fatima Zenati, Abouddihaj Barguigua, Fethi Benbelaïd, Abdelmounaïm Khadir, Chafika
Bellahsene, Mourad Bendahou, Hassaïne Hafida, Mohammed Timinouni. Characterization of
extended-spectrum β-lactamase-produced by Escherichia coli isolates from patients with
urinary tract infection hospitalized in western Algeria (en cours)
Communications:
Fatima Zenati, Mourad Bendahou, Chafika Bellahsene, Fethi Benbelaid, Abdelmounaim
Khadir, 2013.Antimicrobial effects of essential oils on resistant bacteria responsible for
urinary infections.4th
international workshop on industrial biotechnology .April 10-11 2013,
Tlemcen;Algeria
Fatima Zenati, Mourad Bendahou, Chafika Bellahsene, Fethi Benbelaid, Abdelmounaim
Khadir, 2013. Activité antimicrobienne des huiles essentielles sur germes responsable
d’infection urinaire d’origine hospitalière. Forum sur le développent des sciences de la vie et
de l’univers.14-15 mai, Université de Tlemcen.
Fatima Zenati, Mourad Bendahou, Chafika Bellahsene, Fethi Benbelaid, Abdelmounaim
Khadir, 2014. Activité antimicrobienne des huiles essentielles sur germes responsable
d’infection urinaire d’origine hospitalière. Les 20emes
journées nationales de microbiologie .12
et 13 novembre, Jijel, Algérie.
Fatima Zenati, Mourad Bendahou, Chafika Bellahsene, Fethi Benbelaid, Abdelmounaim
Khadir, 2014. Effet de certains extraits des plantes aromatiques et médicinales sur les
bactéries responsables des infections urinaire. La 1eme
journée scientifique des sciences de
l’agroalimentaire, environnement et santé. 3 juin , Université de Tlemcen.

Publications

© 2014 Fatima Zenati et al. This is an open access article distributed under the terms of the Creative Commons Attribution License -NonCommercial-ShareAlike Unported License (http://creativecommons.org/licenses/by-nc-sa/3.0/).
Journal of Applied Pharmaceutical Science Vol. 4 (11), pp. 015-018, November, 2014 Available online at http://www.japsonline.com DOI: 10.7324/JAPS.2014.4113 ISSN 2231-3354
Antimicrobial effects of three essential oils on multidrug resistant bacteria responsible for urinary infections Fatima Zenati, Fethi Benbelaid, Abdelmounaim Khadir, Chafika Bellahsene, Mourad Bendahou
Laboratory of Applied Microbiology in Food, Biomedical and Environment (LAMAABE), Aboubekr Belkaïd University, PO Box 119, 13000 Tlemcen, Algeria.
ARTICLE INFO
ABSTRACT
Article history: Received on: 30/06/2014 Revised on: 18/07/2014 Accepted on: 02/08/2014 Available online: 27/11/2014
Urinary tract infections (UTIs) are ones of the most common nosocomial infections worldwide. Lesions which are mainly caused by microorganisms that inhabiting in hospitals, known and characterized by their both resistance against antibiotics and high ability of biofilms formation. In this study, we have evaluated the effect of three essential oils, which are Cinnamomum cassia, Coriandrum sativum and Ziziphora hispanica, against bacterial species most responsible for UTIs. A total of 18 bacterial strains were tested, which varies between reference strains and clinical multidrug resistant. Cassia oil was the most antimicrobial active against all strains, with interesting MICs values which doesn’t exceed 5 mg/ml. The finding of this study indicate that essential oil appears as an excellent solution for treatment of nosocomial UTIs, especially against failure problems seen in care services, which are over common in the last years.
Key words: Urinary tract infections, Essential oils, Antimicrobial activity, Multidrug resistance.
INTRODUCTION
Urinary Tract Infections (UTIs) are among the most common bacterial infections accounting for morbidity and mortality in all the human populations (Hooton, 2000), especially in women (Todar, 2006). Most of UTIs are hospital-acquired infections in which they represent up to 40% and 34% of infections acquired in long-stay units (Michelet and Tattevin, 2003). Regarding pathogenic responsible, Enterobacteriaceae were the most common bacteria detected, in which they causes up to 84.3% of UTIs (Gales et al., 2000). Others pathogens are also identified, especially Enterococcus faecalis (Singh et al., 2007) and Staphylococcus spp. (Kuroda et al., 2005; Muder et al., 2006).
UTIs present a real problem currently, not only because these types of infections are in increasing in last years, notably in developed countries (Nicolle, 2012), but also the pathogens responsible are commonly multidrug resistant bacteria to antibiotics used in routine (Manikandan et al., 2011).
.
* Corresponding Author Mourad Bendahou, Laboratory of Applied Microbiology in Food, Biomedical and Environment (LAMAABE), Aboubekr Belkaïd University, PO Box 119, 13000 Tlemcen, Algeria. Email : [email protected]
The resistance against antibiotics was detected in several genera, including Escherichia, Enterobacter, Klebsiella, Proteus, Salmonella, Serratia and Pseudomona (Noor et al., 2004). There is an urgent need thus, for highlighted new antimicrobial agents which will be used against treatment failures seen in UTIs related to multidrug resistance. Plants preparations have long been used in treatment of all kinds of human infectious diseases (Lai and Roy, 2004), including UTIs. Presently, it’s has been clear that plants are a promising and wealthy source for safe and effective new antimicrobial agents (Benbelaïd et al., 2013).
Among which, essential oils (EOs) are a mixture of violates compound derived from secondary metabolism of aromatic plants, and are one of the most interesting biomolecules of natural origin (Edris, 2007). Through scientific researches, some EOs possess an antimicrobial potency against microorganisms (Bakkali et al., 2008), activity which can be valorized in medicine against nosocomial infections, such as UTIs. Therefore, EOs appears as a probable solution against treatment failures seen in UTIs. The purpose of this study was to investigate the antimicrobial potency of three EOs against multidrug resistant bacteria responsible for urinary infections. We have tested multidrug resistant clinical strains, which are obtained from patients with UTIs, compared with other reference strains sensitive to antibiotics.

016 Zenati et al. / Journal of Applied Pharmaceutical Science 4 (11); 2014: 015-018 MATERIALS AND METHODS
Plant material Three medicinal plants were selected for this study,
including Coriandrum sativum (C. sativum), Ziziphora hispanica (Z. hispanica), and Cinnamomum cassia (C. cassia). The species choice was based on literature survey and their use in traditional medicine by Algerian population. The plant materials used for EOs obtention were purchased from commercial sources.
EOs obtention
EOs were obtained from dried plant materials by hydrodistillation for 3 h, using a Clevenger-type apparatus. Recovered EOs were dried using magnesium sulfate then stored at +4°C until tested.
Bacterial strains
A total of 18 bacterial strains were tested in this study, including reference and clinical ones (Table 1). The species are nine Escherichia coli (E. coli) strains, three Pseudomonas aeruginosa (P. aeruginosa), three Klebsiella pneumoniae (K. pneumoniae) and three Proteus mirabilis (P. mirabilis).
Clinical strains were collected from patients with UTIs recruited at the University Hospital of Tlemcen, in the following services, internal medicine, urology, and maternity. UTIs in patients were confirmed by detection of significant bacteremia by urine culture and sensitivity method. Bacterial growth was carried out on Mac Conkey agar (Conda Pronadisa™, Spain) and Nutrient agar (Conda Pronadisa™, Spain), and the diagnosis of UTIs in urine samples was based on the presence of ≥ 105 CFU of microorganisms per ml in urine culture (Cardoso et al., 1998). Isolated bacterial strains were firstly identified by conventional biochemical methods, and the API 20E (BioMerieux® SA, Lyon, France) were used for final identification confirmation. Antibiogram
The multidrug resistance in clinical strains was determined according to the Clinical and Laboratory Standard Institute (CLSI, 2006). Antibiogram was carried out with the following antimicrobial agent-containing disks: Amoxicillin (25 μg), amoxicillin/clavulanic acid (30 μg), ticarcillin (75 μg), ticarcilline/clavulanic acid (85 μg), piperacillin (75 μg), piperacillin/tazobactam (85 μg), cephalotin (30 μg), cefuroxime (30 μg), cefixime (30 μg), cefotaxime (30 μg), ceftazidime (30 μg), cefepime (30 μg), imipenem (10 μg), aztreonam (30 μg), cefoxitin (30 μg), gentamicin (15 μg), tobramycin (10 μg), nalidixic acid (30 μg), ofloxacin (5 μg), ciprofloxacin (5 μg), and fosfomycin (50 μg) (Oxoid®, England). The resistance phenotype of studied strains is presented in Table 1.
EOs antibacterial activity Preparation and standardization of inocula
For inocula preparation, four to five isolated colonies of tested organisms were picked by sterile inoculating loop and
inoculated in tubes of BHIB (5 ml in each). The inoculated tubes were incubated at 37oC for 18 hours and standardized to 108CFU/ml (CLSI, 2006). Agar disc diffusion method
The agar well diffusion method was used for the antimicrobial evaluations. 18 ml of Mueller Hinton agar medium (Conda Pronadisa™, Spain) was casted in each Petri dish (9 cm) and then inoculated by swabbing with a suspension of 18h culture already standardized at 108 CFU/ml, as recommended by (CLSI, 2006). After that, discs of filter paper (6 mm diameter) thoroughly moistened with 5 μl of the oil were placed on surface of inoculated agar, then incubated at 37°C for 24 h. The result was determined by measuring diameters of the inhibition zones in millimeters. Determination of the minimum inhibitory concentration (MIC)
The minimum inhibitory concentration (MIC) of studied essential oils was determined by micro dilution method described by (Wiegand et al., 2008). The essential oils were dissolved using Tween 80, then diluted by ½ at ten concentrations in external tubes ranging from 400 to 0.7 mg/ml. After that, the 96-well plates were prepared by distributing 180 μl of 105 CFU/ml inoculum with 20μl of each concentration of essential oil. The final concentration of oil in plat was ranged between 40 to 0.07 mg/ml, and the final concentration of Tween 80 was 1% in each well. The MIC was defined as the lowest concentration of essential oil inhibiting visible growth. RESULTS AND DISCUSSION
The antibiogram of different clinical isolates showed several susceptible and resistant phenotypes. From the eight strains of E. coli, four were resistant, among these last two strains showed increased MDR. Regarding the species P. aeruginosa from two collected strains one was resistant. The same result was obtained by P. mirabilis and K. pneumoniae (see Table 1).
The antimicrobial activity results of EOs against strains collected from urinary infections show that the most important activity is recorded by C. cassia, since this oil showed significant activity against all Gram positive and negative strains including P. aeruginosa, which is known by its resistance to essential oils. In addition, the greater diameters were obtained by this EO with inhibition zone of 39 mm for some strains such as the case of E. coli. Then the essential oil of C. sativum also gave satisfactory results with inhibitions zones up to 31 mm but with no inhibition effect against P. aeruginosa. The EO of Z. hispanica gave similar results to C. sativum since it didn’t inhibit the species P. aeruginosa. However, its effect was somewhat weaker than C. sativum that the greatest diameter was 17 mm (see table 2). From the inhibitions zones obtained and according to Ponce et al. (2003) the strains were extremely susceptible to C. cassia and C. sativum but they are also very sensitive to Z. hispanica. The results of minimum inhibitory concentrations MICs correlate with those of

Zenati et al. / Journal of Applied Pharmaceutical Science 4 (11); 2014: 015-018 017
the zones of inhibition since the smaller MICs were obtained with the larger inhibitions zones and vice versa. UTIs are the second most common type of infections in human body, which are one of the most serious health problem affecting millions of people each year. UTIs involve infection in the kidneys, ureters, bladder or urethra (Anjum et al., 2004).
For minimized risks of mortality, morbidity, and any renal damage which can be caused by UTIs, clinicians should use the appropriate antibiotic in treatment. Choosing the specific antimicrobial agents appear so difficult, especially because of resistance to antimicrobial agents remarked in bacteria responsible for nosocomial UTIs.
In this study, results obtained are corresponding to literature, in which we have found that almost of bacteria responsible for UTIs are especially E. coli, and at less degrees K. pneumoniae, P. mirabilis and P. aeruginosa (Hooton, 2012; Barber et al., 2013). Also, it’s been clear that pathogens responsible for UTIs are commonly from hospital origin, species
which are multi-resistant to antibiotics (Manikandan et al., 2011) and possess a high ability to biofilm formation (Tenke et al., 2012). Therefore, the resolution of UTIs problem depend especially in hygiene in hospitals and most important in highlight of new antimicrobial agents in case of treatment failures. Among selected essential oils for this study, the oil of C. cassia has shown an interesting antimicrobial activity against all studied bacterial species. According to the literature, it’s has been clear that cassia oils are possess an strong antimicrobial activity (Ooi et al., 2006; Yang et al., 2012), with interesting MICs values which varies between 0.018 and 0.7 mg/mL against Gram positive and negative bacteria and fungi. Also, cassia oil was active against P. aeruginosa a specie which are commonly resistant not only to antibiotics (Zavascki et al., 2010), but also against almost essential oils (Mann et al., 2000; Bekhechi et al., 2008). Given that the use of EOs in antiseptic is better than use of terpenoids alone (Benbelaïd et al., 2014), cassia oil seems to be an ingesting alternative treatment in nosocomial UTIs.
Table . 1: Data about studied clinical strains.
Strains Resistance phenotype Escherichia coli 1 Sensitive Escherichia coli 2 Sensitive Escherichia coli 3 Sensitive Escherichia coli 4 Sensitive Escherichia coli 5 TCC,TIC, FEP, KF, PRL, CXM, AMX, CAZ, ATM, CTX,AMC Escherichia coli 6 TCC,TIC, FEP, KF, PRL, CXM, AMX, CAZ, ATM, CTX,AMC Escherichia coli 7 AMX ,TIC ,CXM Escherichia coli 8 AMX ,TIC ,CXM,TCC Pseudomonas aeruginosa 1 Sensitive Pseudomonas aeruginosa 2 TCC, TOB, FF, CN Proteus mirabilis 1 Sensitive Proteus mirabilis 2 TCC,TIC, FEP, KF, PRL, CXM, AMX, CAZ, ATM, CTX Klebsiella pneumoniae 1 Sensitive Klebsiella pneumoniae 2 TCC,TIC, FEP, KF, PRL, CXM, AMX, CAZ, ATM, CTX,AMC Table . 2: Antimicrobial activity of studied essential oils against bacteria responsible for nosocomial UTIs.
Strains Cinnamomum cassia Coriandrum sativum Ziziphora hispanica Iz MIC Iz MIC Iz MIC
Escherichia coli ATCC 25922 30±1 0.63 13±1 2.50 14±1 2.50 Escherichia coli 1 35±1 0.63 31±1 1.25 17±1 2.50 Escherichia coli 2 32±1 0.30 29±1 1.25 17±1 2.50 Escherichia coli 3 39±1 0.63 16±1 2.50 16±1 2.50 Escherichia coli 4 38±1 0.63 11±1 2.50 13±1 2.50 Escherichia coli 5 26±1 2.50 11±1 5.00 11±1 5.00 Escherichia coli 6 30±1 1.25 11±1 5.00 11±1 5.00 Escherichia coli 7 27±1 1.25 11±1 5.00 12±1 5.00 Escherichia coli 8 29±1 1.25 11±1 5.00 11±1 5.00 Pseudomonas aeruginosa ATCC 27853 15±1 2.50 - - - - Pseudomonas aeruginosa 1 19±1 2.50 - - - - Pseudomonas aeruginosa 2 12±1 5.00 - - - - Proteus mirabilis ATCC 35659 34±2 0.31 12±1 5.00 9±1 5.00 Proteus mirabilis 1 39±1 0.31 13±1 2.50 10±1 5.00 Proteus mirabilis 2 30±1 1.25 9±1 05.00 8±1 10.0 Klebsiella pneumoniae ATCC 70603 21±1 0.16 14±1 2.50 12±1 2.50 Klebsiella pneumoniae 1 32±1 0.31 14±1 5.00 13±1 5.00 Klebsiella pneumoniae 2 27±1 0.16 10±1 2.50 7±1 2.50 Iz: inhibition zones (mm). MIC: minimum inhibitory concentration (mg/ml).

018 Zenati et al. / Journal of Applied Pharmaceutical Science 4 (11); 2014: 015-018 CONCLUSION
In conclusion, UTIs in human are considered as the most serious health problems facing the world. The present study has revealed the importance of natural products to control antibiotic resistant in bacteria which are being a threat to human health. This scientific study can serve as an important platform for the development of inexpensive, safe and effective medicines. REFERENCES
Anjum F, Kadri S, Ahmad I , Ahmad S. A study of recurrent urinary tract infection among women attending out patient department in SMHS Hospital, Srinagar, Kashmir, India. JK Pract, 2004; 11: 272–273.
Bakkali F, Averbeck S, Averbeck D , Idaomar M. Biological effects of essential oils – A review. Food Chem Toxicol, 2008; 46: 446–475.
Barber AE, Norton JP, Spivak AM , Mulvey MA. Urinary tract infections–current and emerging management strategies. Clin Infect Dis, 2013; 57: 719–724.
Bekhechi C, Atik–Bekkara F , Abdelouahid D. Composition et activité antibactérienne des huiles essentielles d’Origanum glandulosum d’Algérie. Phytothérapie, 2008; 6: 153–159.
Benbelaïd F, Khadir A, Abdoune MA , Bendahou M. Phytochemical screening and in vitro antimicrobial activity of Thymus lanceolatus Desf. from Algeria. Asian Pac J Trop Dis, 2013; 3: 454–459.
Benbelaïd F, Khadir A, Abdoune MA, Bendahou M, Muselli A , Costa J. Antimicrobial activity of some essential oils against oral multidrug-resistant Enterococcus faecalis in both planktonic and biofilm state. Asian Pac J Trop Biomed, 2014; 4: 463–472.
Cardoso CL, Muraro CB, Siqueira VL , Guilhermetti M. Simplified technique for detection of significant bacteriuria by microscopic examination of urine. J Clin Microbiol, 1998; 36: 820–823.
CLSI. Performance Standards for Antimicrobial Disk Susceptibility Tests. Clinical and Laboratory Standards Institute, 2006; 26.
Edris AE. Pharmaceutical and therapeutic potentials of essential oils and their individual volatile constituents–a review. Phytother. Res., 2007; 21: 308–323.
Gales AC, Jones RN, Gordon KA, Sader HS, Wilke WW, Beach ML, Pfaller MA , Doern GV. Activity and spectrum of 22 antimicrobial agents tested against urinary tract infection pathogens in hospitalized patients in Latin America: report from the second year of the SENTRY antimicrobial surveillance program J Antimicrob Chemother, 2000; 45: 295–303.
Hooton TM. Pathogenesis of urinary tract infections–an update. J Antimicrob Chemother, 2000; 46: 1–7.
Hooton TM. Uncomplicated urinary tract infection. New Engl J Med, 2012; 366: 1028–1037.
Kuroda M, Yamashita A, Hirakawa H, Kumano M, Morikawa K, Higashide M, Maruyama A, Inose Y, Matoba K , Toh H. Whole genome sequence of Staphylococcus saprophyticus reveals the pathogenesis of uncomplicated urinary tract infection. Proc Natl Acad Sci U S A, 2005; 102: 13272–13277.
Lai P , Roy J. Antimicrobial and chemopreventive properties of herbs and spices. Curr. Med. Chem., 2004; 11: 1451–1460.
Manikandan S, Ganesapandian S, Singh M , Kumaraguru A. Antimicrobial susceptibility pattern of urinary tract infection causing human pathogenic bacteria. Asian J Med Sci, 2011; 3: 56–60.
Manikandan S, Ganesapandian S, Singh M , Kumaraguru A. Emerging of Multidrug Resistance Hum an Pathogens from Urinary Tract Infections. Curr Res Bacteriol, 2011; 4: 9–15.
Mann C, Cox S , Markham J. The outer membrane of Pseudomonas aeruginosa NCTC 6749 contributes to its tolerance to the essential oil of Melaleuca alternifolia (tea tree oil). Lett Appl Microbiol, 2000; 30: 294–297.
Michelet C , Tattevin P. Infections urinaires nosocomiales chez l’immunocompétent en milieu médical: qui traiter, quand traiter et comment traiter? Med Mal Infect, 2003; 33: 457–468.
Muder RR, Brennen C, Rihs JD, Wagener MM, Obman A, Stout JE , Victor LY. Isolation of Staphylococcus aureus from the urinary tract: association of isolation with symptomatic urinary tract infection and subsequent staphylococcal bacteremia. Clin Infect Dis, 2006; 42: 46–50.
Nicolle LE. 2012. Urinary tract infections.in: Vincent J-L , Hall JB, ed. Encyclopedia of Intensive Care Medicine. Berlin Springer-Verlag 2359–2364.
Noor N, Ajaz M, Rasool SA , Pirzada ZA. Urinary tract infections associated with multidrug resistant enteric bacilli–characterization and genetical studies. Pak J Pharm Sci, 2004; 17: 115–123.
Ooi LS, Li Y, Kam S-L, Wang H, Wong EY , Ooi VE. Antimicrobial activities of cinnamon oil and cinnamaldehyde from the Chinese medicinal herb Cinnamomum cassia Blume. Am J Chin Med, 2006; 34: 511–522.
Ponce AG, Fritz R, del Valle C , Roura SI. Antimicrobial activity of essential oils on the native microflora of organic Swiss chard. LWT -Food Sci. Technol., 2003; 36: 679–684.
Singh KV, Nallapareddy SR , Murray BE. Importance of the ebp (endocarditisand biofilm-associated pilus) locus in the pathogenesis of Enterococcus faecalis ascending urinary tract infection. J Infect Dis, 2007; 195: 1671–1677.
Tenke P, Köves B, Nagy K, Hultgren SJ, Mendling W, Wullt B, Grabe M, Wagenlehner FM, Cek M , Pickard R. Update on biofilm infections in the urinary tract. World J Urol, 2012; 30: 51–57.
Todar K. 2006. Todar's online textbook of bacteriology. Madison, USA: University of Wisconsin-Madison Department of Bacteriology.
Wiegand I, Hilpert K , Hancock REW. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat Protoc, 2008; 3: 163–175.
Yang CH, Yang CS, Hwang ML, Chang CC, Li RX , Chang LY. Antimicrobial activity of various parts of Cinnamomum cassia extracted with different extraction methods. J Food Biochem, 2012; 36: 690–698.
Zavascki AP, Carvalhaes CG, Picão RC , Gales AC. Multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii–resistance mechanisms and implications for therapy. Expert Rev Anti Infect Ther, 2010; 8: 71–93.
How to cite this article:
Fatima Zenati, Fethi Benbelaid, Abdelmounaim Khadir, Chafika Bellahsene, Mourad Bendahou. Antimicrobial effects of three essential oils on multidrug resistant bacteria responsible for urinary infections. J App Pharm Sci, 2014; 4 (11): 015-018.

Publication (en cours)
Title: Characterization of extended-spectrum β-lactamase-produced by Escherichia coli
isolates from patients with urinary tract infection hospitalized in western Algeria
Order of authors: Fatima Zenati1, Abouddihaj Barguigua
2,3, Fethi Benbelaïd
1,
Abdelmounaïm Khadir1, Chafika Bellahsene
1, Mourad Bendahou
1, Hassaïne Hafida
1,
Mohammed Timinouni2,*
1 Laboratory of Applied Microbiology in Food, Biomedical and Environment (LAMAABE),
AboubekrBelkaïd University, PO Box 119, 13000 Tlemcen, Algeria 2
Molecular bacteriology laboratory, Pasteur Institute of Morocco, 1Place Louis Pasteur,
20360, Casablanca, Morocco 3 Microbiology laboratory, Faculty of medicine and pharmacy, 1 Street Hospital, 20360,
Casablanca, Morocco
Corresponding author: Mohammed Timinouni
Corresponding Author Institution: Pasteur Institute of Morocco, Casablanca
Address of corresponding author: MolecularBacteriology Laboratory, Pasteur Institute of
Morocco.1, Place Louis Pasteur.Casablanca (20360) Morocco ; Tel : + (212) 05 22 43 44 50 ;
Fax: + (212) 05 22 26 09 57 ; E-mail:[email protected]
Running title: ESBL producing E. coli isolated from Algeria
Abstract
Background & objectives: Extended spectrum β-lactamases (ESBLs) produce bacteria have
emerged as a major threat worldwide with limited treatment options. The present study was
aimed to determine the occurrence of ESBLs in Escherichia coli, their molecular types and
associated risk factors.
Methods: A total of 83 clinical isolates of E. coli collected between 2011 and 2013 in
the Tlemcen university hospital Algeria were examined phenotypically for ESBL production.
ESBL strains were further typed for the blaTEM/SHV/CTX-M genes by PCR using specific primers.
Resistance to other antimicrobial agents was also studied. Various risk factors associated with
ESBL infections were analysed by logistic regressions.
Results: ESBLs were found in 32.5 percent E. coli isolates. The majority of typed isolates
harboured two or more ESBL genes. Overall bla CTX-M was the commonest genotype (100%)
followed by blaTEM (96%) and blaSHV (4%) either alone or in combination. The frequencies of
resistance to amoxicillin and cephalotin, which are antibiotics used in the treatment first-line
bladder infections, are relatively elevated in both groups.
Interpretation & conclusions: Our study showed high ESBL occurrence with CTX-M.
Keywords: Extended-spectrum β-lactamase; Urinary tract infections; Escherichia coli.

يسخشفى الانسؤونت ع االنخهاباث انبىنت و انعسونت ي E. coli BLSE عهى زىث عطرتثالديفعىل : العنوان
الملخص
,Cinnamomum cassia)د انقضاء عهى هذ انشكهت حى حجربت ثالثت زىث عطرت وف صذ.اإلساإ يقاويت انبكخرا نهضاداث انحىت يشكهت حققت حهذد صحت
Coriandrum sativum Ziziphora hispanica) ضذ . Ecoli BLSEأكثر انساء هى أ انخائج بج إ . انسؤونت ع االنخهاباث انبىنت و انعسونت ي يسخشفى حهسا
نهذ انبكخرا اها حهك PCR واظهر ححهم E. coli BLSE ℅32,5 يع انعهى ا سبت ℅63حثم انسالنت انرئست نهذا االنخهاب بسبت E. coliعرضت نالنخهاباث انبىنت وا
األكثر ه Cinnamomum cassia أ زىث عطرت بج ثالد نخأثر انعرضت E. coli BLSE إ.℅100 و℅96 بسبت TEM1وBla CTX-M سبت كبرة ي جاث
األدى انثبطيم وحركس /غ و2.5 األدىيم وحركس انقاحم /غ و1.97 اد انثبط يى يع انعهى ا قت حركس 33.23 انثبط انحانخ انهائت وانبىفهت يع قطر كهخا ف حأثرا
واظهر يسج انسىث .نها يفعىل اقمZiziphora hispanica و Coriandrum sativum أيم يع انعهى /غ و4,53 انبىفهى هىإلزانت األدىيم و انخركس /غ و 4,37 نبىفهىلا
حقخم Cinnamomum cassia انخال ا انبكخرت اضهرث أ زج حركت قخم أ كا إضاف يفعىال E. coli BLSE 09ضذ céfotaxime انعطرت يع انضاد انحىي
.دققت 120 ف غضى Coriandrum sativum و دققت 30 انبكخرا ف غضى
انفعانت ضذ انبكخرا, زىث عطرت,انقاويت , ,PCR , E. coli BLSE االنخهاباث انبىنت:انكهاث انفخاحت
Title: Inhibitory effect of essential oils of three aromatic plants on Escherichia coli (ESBL) responsible for urinary tract infections Hospita
Abstract
Bacterial resistance to antibiotics is a real public health problem. In order to find alternative fight against this resistance, the proposed study is to
evaluate the inhibitory effect of three essential oils (Cinnamomum cassia, Coriandrum sativum and Ziziphora hispanica) on ESBL E. coli strains
responsible for urinary tract infections isolated at the University Hospital of Tlemcen. The results showed that nosocomial urinary tract infection is high
among women and it is mainly due to E. coli (63℅). The effect of E. coli ESBL was 32,5%. PCR analysis of ESBL E. coli, revealed the presence of
genes Bla CTX-M and TEM1 with high percentage (100℅ and 96℅). ESBL E. coli strains subject to the effect of three essential oils, have shown great
sensitivity to essential oil of Cinnamomum cassia at state planctonic and biofilm. She gave an inhibition zone > 33,25mm, MIC=1,97mg/ml, MBC =
2,5mg/ml and a MBIC=4,37mg/mL, MBEC=4,53mg/ml. While, Coriandrum sativum and Ziziphora hispanica oils are less active. Essential oils
combined with the antibiotic cefotaxime against ESBL E. coli 09, showed an additive effect of Cinnamomum cassia and Coriandrum sativum essential
oils. The kinetics of destruction of bacterial cells with essential oil of Cinnamomum cassia was obtained after 30 min, while Coriandrum sativum oil
destroys bacterial cells after 240 min.
Keywords : Urinary tract infection ; ESBL E. coli ; PCR ; Résistance ; Essential oils Antibactérial activity.
Titre : Effet inhibiteur des huiles essentielles de trois plantes aromatiques sur Escherichia coli (BLSE) responsables d’infections
urinaires d’origine hospitalière.
Résumé
La résistance des bactéries aux antibiotiques pose un vrai problème de santé publique. Dans le but de trouver des alternatifs de
lutte contre cette résistance, l’étude proposé est d’évaluer l’effet inhibiteur de trois huiles essentielles (Cinnamomum cassia, Coriandrum
sativum et Ziziphora hispanica) sur des souches E. coli BLSE responsables d’infections urinaires isolées au CHU de Tlemcen. Les
résultats ont montré que l’infection urinaire nosocomiale est importante chez les femmes et elle est due principalement à E. coli (63℅).
L’incidence d’Escherichia coli BLSE a été de 32,5%. L’analyse par PCR de E. coli BLSE ces dernières, a révélé la présence des gènes
Bla CTX-M et TEM1 avec un pourcentage élevé (100℅ et 96℅). Les souches E. coli BLSE soumises à l’effet des trois huiles
essentielles, ont montré une grande sensibilité à l’huile essentielle de Cinnamomum cassia à l’état planctonique et biofilm. Elle a donné
en moyenne respectivement, une zone d’inhibition > 33,25mm, une CMI=1,97mg/ml, CMB=2,5mg/ml et une CMIB=4,37 mg/ml,
CMEB=4,53mg/ml. Par contre Coriandrum sativum et Ziziphora hispanica sont moins actif. En combinant les huiles à l’antibiotique
céfotaxime contre E.coli BLSE 09, les huiles essentielles de Cinnamomum cassia et Coriandrum sativum ont présenté un effet
d’addition. La cinétique de destruction des cellules bactériennes par l’huile essentielle de Cinnamomum cassia a été obtenue au bout de
30 min, par contre il a fallu 240 min pour les détruire par l’huile de Coriandrum sativum.
Mots clés : Infection urinaire ; E. coli BLSE ; PCR ; Résistance ; Huiles essentielles ; Activité antibactérienne.
Copyright © 2022 FDOKUMEN




















