
Effects of the fuel oil spilled by the Prestige tanker on reproduction parameters of wild mussel...
11
Effects of the fuel oil spilled by the Prestige tanker on reproduction parameters of wild mussel populations Maren Ortiz-Zarragoitia, Larraitz Garmendia, Mar ıa Carmen Barbero, Teresa Serrano, Ionan Marig omez and Miren P. Cajaraville * Received 23rd April 2010, Accepted 2nd August 2010 DOI: 10.1039/c0em00102c The aim of this work was to assess possible effects of the Prestige oil spill on reproduction parameters of mussels along the Galician and Bay of Biscay coast. Studied endpoints included sex ratio, gonad histology and vitellogenin-like proteins using the alkali-labile phosphate (ALP) method. A high prevalence of haemocytic infiltration of follicles and severe oocyte atresia was found in most localities in April 2003. Spawning gonads were observed in most impacted populations in the same sampling. In April 2004 mature small sized follicles were observed. No histopathological changes were observed in April 2005 and 2006, except a high prevalence of necrotic gametes in 6 out of 22 localities in April 2006. Female ALP levels showed high interindividual variability in April 2004, which was reduced in April 2005 and 2006. No xenoestrogenic effects were observed in male mussels. Overall, gamete alterations were detected during 2003–2004 and a recovery trend was observed afterwards. Introduction On November 19th, 2002, the oil tanker Prestige broke in half and sunk in international waters 133 miles from the north western coast of the Iberian Peninsula. The Prestige transported 77 000 tones of heavy fuel oil no. 2 according to the French classification (UK classification type no. 6; Russian classification M-100). More than 60000 tones of fuel oil were spilled affecting mainly the coast of Galicia on the north west of the Iberian Peninsula. During the following months spilled oil also affected the whole northern coast of the Iberian Peninsula reaching up to Brittany. 1,2 The Prestige oil spill (POS) has been considered the most important ecological disaster ever to occur in the affected area. 1,3 Severe loss of intertidal invertebrate communities has been measured on POS affected beaches. 4,5 The area impacted by the POS is rich in bivalve production, Galicia being the most important European producer of mussels. Mussel production surpasses 200 000 Tn per year and employs more than 8000 full-time workers in Galicia. 6 Mussels have been used worldwide as sentinel species of environmental pollution in coastal areas. 7,8 Mussels are excellent indicators of contamination levels in the marine environment as they are sessile and filter feed from surrounding water. Furthermore, they show a low ability to metab- olize xenobiotic compounds like polycyclic aromatic hydrocarbons (PAHs) and accumulate them at high levels. 9,10 Mussels are sensitive species to accidental oil spills as effects on different biomarkers have been observed after Exxon Valdez in Alaska, 11 Aegean Sea in Gali- cia, 12 Sea Empress in Wales, 13 Coral Bulker in North Portugal, 14 Erika in Brittany 15 and the case of Prestige. 16,17 Studied biological endpoints in mussels affected by an oil spill usually show a rapid recovery in 1 to 2 years after the oil spill. 12,13,18 However, long lasting effects could not be discounted as shown in the case of wildlife Biologia Zelularra eta Histologia Lab., Zoologia eta Biologia Zelularra Saila, Zientzia eta Teknologia Fakultatea, University of the Basque Country UPV/EHU, Sarriena z/g, 48940 Leioa, Basque Country (Spain). E-mail: [email protected]; Fax: +34 946013500; Tel: +34 946012697 Environmental impact Several works have used mussels as sentinel organism for chemical biomonitoring studies after oil spill events, but no studies have addressed the impact of oil spills on reproduction-parameters in wild mussel populations. Here we present for the first time a long- term (2003–2006) biomonitoring survey studying reproduction-related parameters, such as sex ratio, gonad histology and vitello- genin-like proteins, in 22 mussel populations from Galicia and the Bay of Biscay after the Prestige oil spill. Our results showed alterations in gamete development in mussels from the whole studied area in 2003 and 2004, but recovered later on (2005–2006). The applied reproduction-related biomarkers have been demonstrated to be a useful tool for pollution assessment and could be included in ongoing worldwide biomonitoring programmes studying mussels. 84 | J. Environ. Monit., 2011, 13, 84–94 This journal is ª The Royal Society of Chemistry 2011 Dynamic Article Links C < Journal of Environmental Monitoring Cite this: J. Environ. Monit., 2011, 13, 84 www.rsc.org/jem PAPER Published on 29 October 2010. Downloaded on 13/01/2014 15:25:06. View Article Online / Journal Homepage / Table of Contents for this issue
Transcript of Effects of the fuel oil spilled by the Prestige tanker on reproduction parameters of wild mussel...

Dynamic Article LinksC<Journal ofEnvironmentalMonitoringCite this: J. Environ. Monit., 2011, 13, 84
www.rsc.org/jem PAPER
Publ
ishe
d on
29
Oct
ober
201
0. D
ownl
oade
d on
13/
01/2
014
15:2
5:06
. View Article Online / Journal Homepage / Table of Contents for this issue
Effects of the fuel oil spilled by the Prestige tanker on reproductionparameters of wild mussel populations
Maren Ortiz-Zarragoitia, Larraitz Garmendia, Mar�ıa Carmen Barbero, Teresa Serrano, Ionan Marig�omezand Miren P. Cajaraville*
Received 23rd April 2010, Accepted 2nd August 2010
DOI: 10.1039/c0em00102c
The aim of this work was to assess possible effects of the Prestige oil spill on reproduction parameters of
mussels along the Galician and Bay of Biscay coast. Studied endpoints included sex ratio, gonad
histology and vitellogenin-like proteins using the alkali-labile phosphate (ALP) method. A high
prevalence of haemocytic infiltration of follicles and severe oocyte atresia was found in most localities
in April 2003. Spawning gonads were observed in most impacted populations in the same sampling. In
April 2004 mature small sized follicles were observed. No histopathological changes were observed in
April 2005 and 2006, except a high prevalence of necrotic gametes in 6 out of 22 localities in April 2006.
Female ALP levels showed high interindividual variability in April 2004, which was reduced in April
2005 and 2006. No xenoestrogenic effects were observed in male mussels. Overall, gamete alterations
were detected during 2003–2004 and a recovery trend was observed afterwards.
Introduction
On November 19th, 2002, the oil tanker Prestige broke in half
and sunk in international waters 133 miles from the north
western coast of the Iberian Peninsula. The Prestige transported
77 000 tones of heavy fuel oil no. 2 according to the French
classification (UK classification type no. 6; Russian classification
M-100). More than 60000 tones of fuel oil were spilled affecting
mainly the coast of Galicia on the north west of the Iberian
Peninsula. During the following months spilled oil also affected
the whole northern coast of the Iberian Peninsula reaching up to
Brittany.1,2 The Prestige oil spill (POS) has been considered the
most important ecological disaster ever to occur in the affected
Biologia Zelularra eta Histologia Lab., Zoologia eta Biologia ZelularraSaila, Zientzia eta Teknologia Fakultatea, University of the BasqueCountry UPV/EHU, Sarriena z/g, 48940 Leioa, Basque Country(Spain). E-mail: [email protected]; Fax: +34 946013500; Tel:+34 946012697
Environmental impact
Several works have used mussels as sentinel organism for chemical
addressed the impact of oil spills on reproduction-parameters in wil
term (2003–2006) biomonitoring survey studying reproduction-rela
genin-like proteins, in 22 mussel populations from Galicia and the
alterations in gamete development in mussels from the whole studied
applied reproduction-related biomarkers have been demonstrated to
in ongoing worldwide biomonitoring programmes studying mussel
84 | J. Environ. Monit., 2011, 13, 84–94
area.1,3 Severe loss of intertidal invertebrate communities has
been measured on POS affected beaches.4,5
The area impacted by the POS is rich in bivalve production,
Galicia being the most important European producer of mussels.
Mussel production surpasses 200 000 Tn per year and employs more
than 8000 full-time workers in Galicia.6 Mussels have been used
worldwide as sentinel species of environmental pollution in coastal
areas.7,8 Mussels are excellent indicators of contamination levels in
the marine environment as they are sessile and filter feed from
surrounding water. Furthermore, they show a low ability to metab-
olize xenobiotic compounds like polycyclic aromatic hydrocarbons
(PAHs) and accumulate them at high levels.9,10 Mussels are sensitive
species to accidental oil spills as effects on different biomarkers have
been observed after Exxon Valdez in Alaska,11 Aegean Sea in Gali-
cia,12 Sea Empress in Wales,13 Coral Bulker in North Portugal,14
Erika in Brittany15 and the case of Prestige.16,17 Studied biological
endpoints in mussels affected by an oil spill usually show a rapid
recovery in 1 to 2 years after the oil spill.12,13,18 However, long lasting
effects could not be discounted as shown in the case of wildlife
biomonitoring studies after oil spill events, but no studies have
d mussel populations. Here we present for the first time a long-
ted parameters, such as sex ratio, gonad histology and vitello-
Bay of Biscay after the Prestige oil spill. Our results showed
area in 2003 and 2004, but recovered later on (2005–2006). The
be a useful tool for pollution assessment and could be included
s.
This journal is ª The Royal Society of Chemistry 2011

Publ
ishe
d on
29
Oct
ober
201
0. D
ownl
oade
d on
13/
01/2
014
15:2
5:06
.
View Article Online
populations affected by the Exxon Valdez oil spill.11 Bivalve pop-
ulations from the Prince William Sound area were not completely
recovered 17 years after the oil spill.19
Studies dealing with reproduction effects after oil spill events are
scarce and most of them focus only on population abundances. It is
well known that complex oil mixtures can provoke deleterious
effects on wildlife reproduction.20 Fish populations inhabiting oil
and PAH polluted environments show endocrine disruption
effects, such as reduced plasma hormonal levels and reduced
reproduction success.21 After the Exxon Valdez oil spill, reduced
estradiol levels in plasma from marine fish species such as dolly
varden, yellowfin solea and pollock were measured. In dolly varden
low levels of gonadotropin-I, a neurohormone responsible for
steroid biosynthesis and gamete development, were also observed
together with delayed gamete development.22 Reduction in repro-
ductive success was observed in pink salmon and Pacific herring
from the Exxon Valdez oil spill area.23,24 Embryos obtained from
both species showed low survival ratios and severe morphological
deformities in the following 4 years after the oil spill.23,24
Most of the few works dealing with reproduction alterations
after oil spills have been carried out in vertebrates, few of them
focusing on invertebrates such as bivalve mollusks. Therefore, it
is important to study the possible effects of the POS on repro-
duction parameters in local mussel populations. The endocrine
system of bivalve mollusks is not well understood, but effects on
reproduction parameters in mollusks have been described after
exposure to environmental contaminants, both in the laboratory
and in the field (see review by Porte et al.).25 Mussels have been
shown to be sensitive to possible endocrine disruption effects
provoked by PAHs.25–27 Female mussels exposed in the labora-
tory during gametogenesis to North Sea oil showed low levels of
vitellogenin-like proteins in comparison to control females and
delayed gamete development accompanied by high percentage of
atretic oocytes, indicating possible anti-estrogenic effects.27
Similarly, high prevalences of atretic germ cells were observed in
mussels exposed to diesel oil and PAH derivatives.28,29 Anti-
estrogenic effects were also observed in field studies using soft
shell clams. Female clams inhabiting a PAH contaminated
harbor showed low vitellogenin-like protein levels and retarded
gamete development.30,31 Although several works have demon-
strated the anti-estrogenic activity of oil and PAHs,21,32 some
authors have observed estrogenic effects provoked by PAHs.33,34
The water soluble fraction of the Prestige fuel oil reduced clam
embryo survival35 but did not alter reproduction ability using
Daphnia magna bioassays.36 Polychaetes exposed to fuel oil no. 2
during the whole life cycle showed accelerated oocyte maturation
and suppression of fecundity.37 Furthermore, mussel seed
collected from POS affected areas showed reduced growth in
comparison to mussel seed from non-affected populations.38
Affected mussel seed populations showed higher levels of
triglycerides and carbohydrates.39,40
In the present work we aimed to investigate possible reproduction
effects after POS in local mussel populations, which could compro-
mise their reproduction ability and survival. For this purpose we
studied vitellogenin-like protein levels as a biomarker of endocrine
disruption. In fish, vitellogenin is a bulky phospholipoglycoprotein
synthesized by the liver of female fish under the regulation of estra-
diol. After being synthesized it is secreted to the blood and trans-
ported to the ovary, where it is endocytosed by growing oocytes.
This journal is ª The Royal Society of Chemistry 2011
Vitellogenin is the precursor protein of vitellins present in the oocytes
to feed the embryos.41 In fish, vitellogenin levels are used as a marker
of xenoestrogenicity in adult males and juvenile organisms.41–44 In
bivalve mollusks, vitellogenin synthesis occurs in the follicular cells
of the female gonad.45,46 Regulation of vitellogenin synthesis is not
well understood and hormonal control by estrogens is under
discussion.47–49 Nevertheless, bivalve mollusks are known to
synthesize steroid hormones50 and a relationship between estradiol
levels and vitellogenesis has been suggested.48 Due to the lack of
specific antibodies, measurement of vitellogenin or vitellogenin-like
protein levels in bivalves has been based on indirect methods, such as
the alkali-labile phosphate (ALP) method.25,51 Thus, we applied
measurement of ALP levels in combination with gonad histology to
study possible reproduction alterations in 22 mussel populations
differently affected by the POS in Galicia and the Bay of Biscay, for
a period of three years (2003–2006).
Materials and methods
Samplings and sample processing
For the purpose of the present work a biomonitoring study was
performed along the North Atlantic coast of the Iberian Penin-
sula. Mussels (Mytilus galloprovincialis) were collected from 22
different locations (Fig. 1) selected on the basis of the severity of
the impact of the oil spill and on the oil arrival conditions to the
coast (fresh spilled oil versus weathered oil). Based on the first
reports after the oil spill2 the impact was most severe in the coast
of Galicia, where we studied 8 locations comprised of highly
impacted sites (Agui~no, Caldebarcos, Camelle and Sega~no) and
less impacted sites (Estaca de Bares, Ons, C�ıes and Oia), all
affected by fresh spilled oil. We studied a further 13 localities in
the Bay of Biscay affected later on by weathered oil. We also
included one sampling point in the Natural Park of North
Littoral of Esposende (Sao Bartolomeu do Mar, Portugal) as
a possible non-impacted site. Samplings were performed in April,
July and September 2003 (17 localities), February 2004
(19 localities), April, July and October 2004 and 2005, and April
2006 (22 localities). At each sampling time 30–60 adult mussels,
3.5–4.5 cm long, were collected from each locality and directly
processed in the field. For histological studies 10–30 mussels
from each site were carefully opened by sectioning the adductor
muscle and directly fixed in 10% neutral buffered formalin. For
the study of vitellogenin-like protein levels, mantle samples
collected during April 2004, 2005 and 2006 were used since this is
the period when mussels at this latitude show the most developed
gonads.52–55 Mussel gonads (mantle tissue) from 20 animals were
dissected out, chilled on ice and a small piece of tissue was
removed to do a smear on a microscope slide. The sex of each
animal was determined after visualization of the smear under the
microscope. Afterwards the rest of the tissue was frozen indi-
vidually in liquid nitrogen. Once in the laboratory they were
stored at �80 �C until subsequent analysis.
Sex ratio, gamete development and histopathology of the gonad
The gonad (mantle tissue) from each mussel was dissected out
after fixation, placed in histological cassettes in 70� ethanol and
routinely processed for paraffin embedding in a Leica ASP300
Automatic tissue processor (Nussloch, Germany). 7 mm thick
J. Environ. Monit., 2011, 13, 84–94 | 85

Fig. 1 Map showing the study area, the location where the Prestige sunk and selected sampling points. In the adjacent table geographical coordinates
for the location of each sampling point are shown.
Publ
ishe
d on
29
Oct
ober
201
0. D
ownl
oade
d on
13/
01/2
014
15:2
5:06
.
View Article Online
sections were cut in a Leitz 1512 microtome (Vienna, Austria)
and stained with hematoxylin and eosin.56 The sex of each animal
was recorded and the total number of female and male mussels
was calculated for each sampling time and location, including the
sex data collected from mussels used for ALP measurements.
With these data, the sex ratio was calculated for each sampling
time and location.
Gamete developmental stages were distinguished in mussel
gonads following the classification of Seed.57 Then, a gonad
index (GI) value was assigned to each developmental stage
adapted from the description of Kim et al.:58 1 resting stage
(inactive or undifferentiated); 1.5 early gametogenic stage
(gametogenesis has begun but no ripe gametes visible);
3.5 advanced gametogenic stage (gametogenesis still progressing
and ripe gametes and developing gametes have about equal
proportions); 5 mature stage (gonad fully mature, follicles full of
ova or sperm); 3.5 spawning stage (active emission of gametes,
some follicles appear empty); 1.5 post-spawning stage (empty
follicles and only residual gametes remain). Afterwards, a mean
GI value for each location and sampling time was calculated.
In samples corresponding to April 2003, 2004, 2005 and 2006
a detailed histopathological study was carried out. Histopatho-
logical alterations such as haemocytic infiltration of follicles
containing mature gametes, severe gamete atresia, necrosis and
abnormal follicle development were scored for each animal and
reported as a percentage of total animals in each group, for males
and females separately.
Terminal transferase dUTP nick end labeling (TUNEL) assay
With the aim of identifying apoptotic gametes in the gonads of
studied mussel populations, we used a commercial TUNEL assay
system (DeadEndTM Colorimetric TUNEL system, Promega
86 | J. Environ. Monit., 2011, 13, 84–94
Corporation, Madison, WI, USA). Apoptotic cells were detected
in non-stained gonad histology sections. The fragmented DNA
of apoptotic cells was detected by incorporating a biotinylated
nucleotide at the 30–OH ends of the DNA fragments using the
enzyme terminal deoxynucleotidyl transferase and then staining
with diaminobenzidine (DAB) in a reaction catalyzed by the
enzyme streptavidin horseradish peroxidase.
Briefly, paraffin-embedded sections (7 mm thick) were depar-
affinized and rehydrated in graded ethanol steps. Then, slides were
washed in a 0.85% NaCl solution for 5 min and subsequently
immersed in phosphate buffered saline (PBS) solution for 5 min.
Sections were fixed in 10% neutral buffered formalin for 15 min
and then washed in PBS for 2 � 5 min. Following the manu-
facturer’s instructions, sections were covered with a 20 mg ml�1
proteinase K solution and incubated for 10–30 min at room
temperature in a wet chamber. Afterwards, sections were washed
in PBS for 5 min, refixed in 10% neutral buffered formalin for
5 min and rinsed in PBS for 2 � 5 min. The excess of liquid was
removed by tapping the slides and sections were covered for 5 min
with the equilibration buffer supplied in the commercial kit. After
carefully removing the excess of liquid, sections were covered with
recombinant terminal deoxynucleotidyl transferase (rTdT) reac-
tion mix and incubated at 37 �C for 60 min. Then, slides were
immersed in SSC buffer for 15 min. After rinsing the slides with
PBS for 2� 5 min, endogenous peroxidase was blocked with 0.3%
hydrogen peroxide solution for 5 min. Slides were washed in PBS
for 2� 5 min and incubated in a solution (1 : 500) of streptavidin
horseradish peroxidase for 30 min. Before reaction visualization,
slides were rinsed in PBS for 2 � 5 min and subsequently incu-
bated in the DAB solution supplied in the kit for 10 min in the
dark. Finally, slides were rinsed in deionized water for 2 � 5 min
and mounted in Kaiser’s glycerol gelatine. Apoptotic nuclei
appeared stained dark brown. Negative control sections were run
This journal is ª The Royal Society of Chemistry 2011

Publ
ishe
d on
29
Oct
ober
201
0. D
ownl
oade
d on
13/
01/2
014
15:2
5:06
.
View Article Online
in parallel, where autoclaved deionized water was used instead of
rTdT in the reaction mix. Positive control slides of human pro-
myelocytic leukemia cells (HL60) previously treated with acti-
nomycin to induce apoptosis, were also included.
Alkali-labile phosphate (ALP) method
Levels of vitellogenin-like proteins in mussel gonads were
measured by the ALP method following the protocol described in
Gagn�e et al..59 Briefly, the gonad from each animal was homog-
enized in 25 mM Hepes-NaOH buffer, pH 7.4, containing 125 mM
NaCl, 1 mM dithiothreitol and 1mM EDTA using a glass-teflon
pestle in an ice containing bath with a Braun Potter S homogenizer
(Melsungen, Germany). The obtained homogenate was centri-
fuged at 12 000 g for 30 min at 2 �C in a Beckman Coulter Optima
L-90 K ultracentrifuge (Palo Alto, CA, USA). The obtained
supernatant was carefully removed and an aliquot of 200 ml was
mixed with acetone (35% final acetone concentration) and
centrifuged at 10 000 g for 5 min for the precipitation of proteins.
Another aliquot (75 ml) of the original supernatant was stored at
�80 �C for protein concentration determination. The obtained
pellet was dissolved in 200 ml of 1 M NaOH solution for 30 min at
60 �C in a water-shaking bath. Levels of inorganic free phosphates
were determined using the phosphomolybdenum assay adapted
from Stanton.60 A subsample of 75 ml was mixed with 125 ml tri-
chloroacetic acid, 630 ml ultrapure water, 170 ml molybdenum
reagent (0.02 M ammonium molybdate tetrahydrated and 5.25 M
H2SO4 solution), and 50 ml Fiske-Subbarow reducer (Sigma, St.
Louis, MO, USA). After incubating for 10 min, the absorbance
was measured at 660 nm using a Multiskan Spektrum spectro-
photometer (Thermo Labsystems, Chantilly, VA, USA). For
inorganic phosphate standard curve, a series of KH2PO4
concentrations was used. ALP levels in gonads are given as mg
phosphates/mg protein. Protein concentration was measured
using the DC Protein Assay of Bio-Rad (Hercules, CA, USA)
based on the Lowry method adapted to microtiter plates.61
Statistics
Sex ratio bias was studied using the G test of association,
comparing total number of female and male mussels and
normalizing for theoretical gender bias (1 : 1). Other statistical
analyses were performed using the SPSS 10.0 software (SPSS,
Inc., Chicago, IL, USA). After testing normality (Kolmogorov–
Smirnov’s test) and homogeneity (Levene’s test) of the data, the
effect of factors sampling time and location, as well as their
interaction on ALP levels was studied by two-way analysis of
variance (ANOVA). Statistically significant differences in ALP
levels among groups were studied using one-way ANOVA fol-
lowed by the Duncan’s post-hoc test. In case of GI values, non-
parametric ANOVA (Kruskall–Wallis) was applied followed by
Mann–Whitney’s U test. The interactive effect of both sampling
time and location on GI values was analyzed by Friedman’s test.
Correlation studies between GI and ALP levels were performed
using the Spearman’s correlation index. Differences in percent-
ages of histopathological alterations among groups were studied
by Chi-squared test. Statistical significance was established at
p < 0.05 for all studied tests. Finally, classification studies based
on hierarchical cluster analysis were performed with GI and ALP
This journal is ª The Royal Society of Chemistry 2011
values and a principal component analysis was run to study the
association between studied endpoints and to explain the vari-
ability among sampled populations.
Results
Sex ratio
Sex ratio values for each sampling location at each sampling time
are shown in Table 1. No statistically significant bias in the sex
ratio of the whole studied population (G value ¼ 0.59, p ¼ 0.44)
was detected in comparison with the theoretical sex ratio in
mussel populations (1 : 1). The observed sex ratio was 1 : 1.02
(female : male) for the total of three years of study. When the sex
ratio was analyzed separately for each sampling time, a statisti-
cally significant bias to more males was observed in September
2003 (G value ¼ 27.98; p < 0.01), July 2004 (G value ¼ 4.14,
p < 0.05) and October 2004 (G value ¼ 6.86, p < 0.05). On the
other hand, in April 2004 significantly more females were
observed (G value ¼ 7.09, p < 0.05). Among studied locations
only C�ıes (G value ¼ 7.77, p < 0.05) showed a significant bias to
females (1.7 : 1). Finally, we found two hermaphrodite animals,
one in Mutriku (April 2005) and the other in Ons (July 2005).
They both showed separate male and female follicles in the
mantle tissue (see Fig. 2c below).
Gamete development and gonad index
Microscopical observation of the gonad showed differences in
gamete development patterns among stations (Fig. 3). Further-
more, Friedman’s test showed that both sampling time and
location had significant effects on gamete development (c2 ¼2495.48, p < 0.05). At the beginning of the present study in April
2003, most mussel populations showed mature gonads, with the
exceptions of mussels from Oia, Estaca de Bares and San Vicente
that showed a high number of animals in advanced gametogen-
esis. It is interesting to note that mussels from Agui~no and Cal-
debarcos, located close to the most impacted area, showed
gonads mainly at spawning stage in April 2003. In July 2003,
mussels from all locations were at more advanced developmental
stages than in April 2003, except for mussels from Llanes, which
remained in the mature stage. In most locations, some mussels
were at the post-spawning stage and in Camelle and Suances the
beginning of a second gametogenesis cycle could be observed. In
September 2003, the gonad of most mussels was at a post-
spawning stage and mussels from most locations had started the
next reproductive cycle.
In February 2004 most mussel populations showed gonads in
advanced gametogenesis or in mature phases, with the exception
of mussels from Suances where all mussels showed early game-
togenic gonads (Fig. 3). In April 2004, mussels from most loca-
tions were spawning and some had reached the post-spawning
stage. However, most mussels from Caldebarcos, Estaca de
Bares, Pedre~na and all in Muskiz showed mature gonads (Fig. 3).
In July 2004, spawning but also mature gonads were predomi-
nant in studied mussel populations, indicating the possibility of
a second gametogenic cycle in some mussels of 16 out of 22
locations. Only mussels from Ons, C�ıes, Caldebarcos, Camelle
and Muskiz did not show this process. In October 2004, most
J. Environ. Monit., 2011, 13, 84–94 | 87
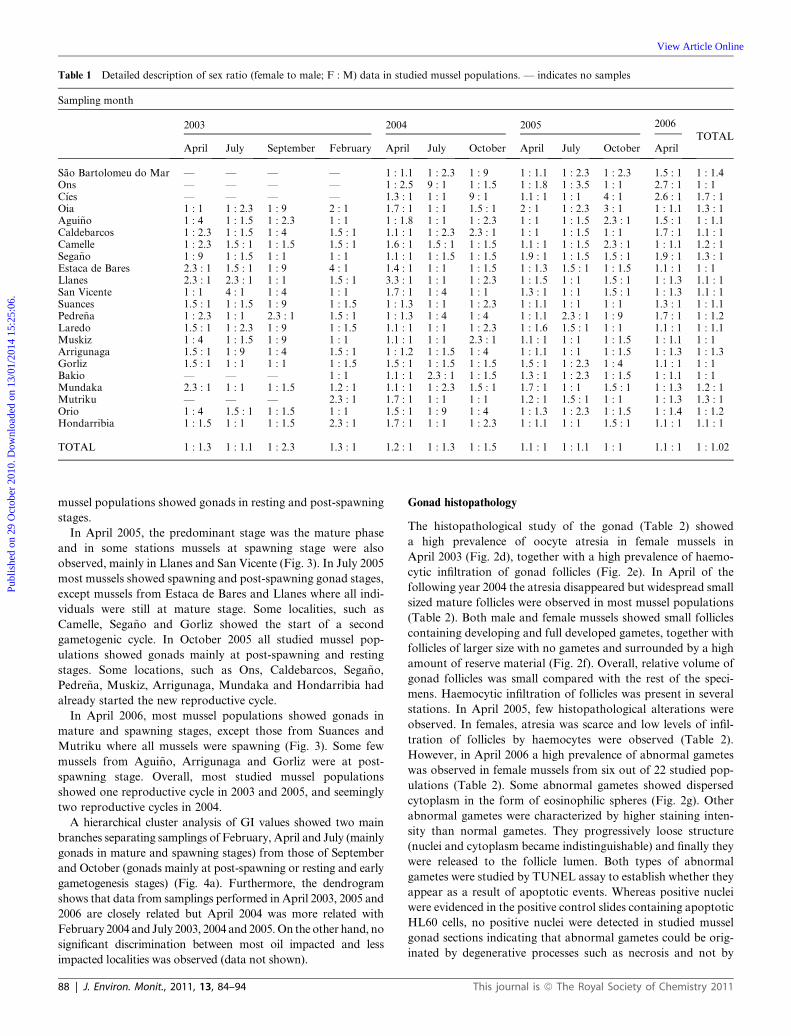
Table 1 Detailed description of sex ratio (female to male; F : M) data in studied mussel populations. — indicates no samples
Sampling month
2003 2004 2005 2006
TOTALApril July September February April July October April July October April
Sao Bartolomeu do Mar — — — — 1 : 1.1 1 : 2.3 1 : 9 1 : 1.1 1 : 2.3 1 : 2.3 1.5 : 1 1 : 1.4Ons — — — — 1 : 2.5 9 : 1 1 : 1.5 1 : 1.8 1 : 3.5 1 : 1 2.7 : 1 1 : 1C�ıes — — — — 1.3 : 1 1 : 1 9 : 1 1.1 : 1 1 : 1 4 : 1 2.6 : 1 1.7 : 1Oia 1 : 1 1 : 2.3 1 : 9 2 : 1 1.7 : 1 1 : 1 1.5 : 1 2 : 1 1 : 2.3 3 : 1 1 : 1.1 1.3 : 1Agui~no 1 : 4 1 : 1.5 1 : 2.3 1 : 1 1 : 1.8 1 : 1 1 : 2.3 1 : 1 1 : 1.5 2.3 : 1 1.5 : 1 1 : 1.1Caldebarcos 1 : 2.3 1 : 1.5 1 : 4 1.5 : 1 1.1 : 1 1 : 2.3 2.3 : 1 1 : 1 1 : 1.5 1 : 1 1.7 : 1 1.1 : 1Camelle 1 : 2.3 1.5 : 1 1 : 1.5 1.5 : 1 1.6 : 1 1.5 : 1 1 : 1.5 1.1 : 1 1 : 1.5 2.3 : 1 1 : 1.1 1.2 : 1Sega~no 1 : 9 1 : 1.5 1 : 1 1 : 1 1.1 : 1 1 : 1.5 1 : 1.5 1.9 : 1 1 : 1.5 1.5 : 1 1.9 : 1 1.3 : 1Estaca de Bares 2.3 : 1 1.5 : 1 1 : 9 4 : 1 1.4 : 1 1 : 1 1 : 1.5 1 : 1.3 1.5 : 1 1 : 1.5 1.1 : 1 1 : 1Llanes 2.3 : 1 2.3 : 1 1 : 1 1.5 : 1 3.3 : 1 1 : 1 1 : 2.3 1 : 1.5 1 : 1 1.5 : 1 1 : 1.3 1.1 : 1San Vicente 1 : 1 4 : 1 1 : 4 1 : 1 1.7 : 1 1 : 4 1 : 1 1.3 : 1 1 : 1 1.5 : 1 1 : 1.3 1.1 : 1Suances 1.5 : 1 1 : 1.5 1 : 9 1 : 1.5 1 : 1.3 1 : 1 1 : 2.3 1 : 1.1 1 : 1 1 : 1 1.3 : 1 1 : 1.1Pedre~na 1 : 2.3 1 : 1 2.3 : 1 1.5 : 1 1 : 1.3 1 : 4 1 : 4 1 : 1.1 2.3 : 1 1 : 9 1.7 : 1 1 : 1.2Laredo 1.5 : 1 1 : 2.3 1 : 9 1 : 1.5 1.1 : 1 1 : 1 1 : 2.3 1 : 1.6 1.5 : 1 1 : 1 1.1 : 1 1 : 1.1Muskiz 1 : 4 1 : 1.5 1 : 9 1 : 1 1.1 : 1 1 : 1 2.3 : 1 1.1 : 1 1 : 1 1 : 1.5 1 : 1.1 1 : 1Arrigunaga 1.5 : 1 1 : 9 1 : 4 1.5 : 1 1 : 1.2 1 : 1.5 1 : 4 1 : 1.1 1 : 1 1 : 1.5 1 : 1.3 1 : 1.3Gorliz 1.5 : 1 1 : 1 1 : 1 1 : 1.5 1.5 : 1 1 : 1.5 1 : 1.5 1.5 : 1 1 : 2.3 1 : 4 1.1 : 1 1 : 1Bakio — — — 1 : 1 1.1 : 1 2.3 : 1 1 : 1.5 1.3 : 1 1 : 2.3 1 : 1.5 1 : 1.1 1 : 1Mundaka 2.3 : 1 1 : 1 1 : 1.5 1.2 : 1 1.1 : 1 1 : 2.3 1.5 : 1 1.7 : 1 1 : 1 1.5 : 1 1 : 1.3 1.2 : 1Mutriku — — — 2.3 : 1 1.7 : 1 1 : 1 1 : 1 1.2 : 1 1.5 : 1 1 : 1 1 : 1.3 1.3 : 1Orio 1 : 4 1.5 : 1 1 : 1.5 1 : 1 1.5 : 1 1 : 9 1 : 4 1 : 1.3 1 : 2.3 1 : 1.5 1 : 1.4 1 : 1.2Hondarribia 1 : 1.5 1 : 1 1 : 1.5 2.3 : 1 1.7 : 1 1 : 1 1 : 2.3 1 : 1.1 1 : 1 1.5 : 1 1.1 : 1 1.1 : 1
TOTAL 1 : 1.3 1 : 1.1 1 : 2.3 1.3 : 1 1.2 : 1 1 : 1.3 1 : 1.5 1.1 : 1 1 : 1.1 1 : 1 1.1 : 1 1 : 1.02
Publ
ishe
d on
29
Oct
ober
201
0. D
ownl
oade
d on
13/
01/2
014
15:2
5:06
.
View Article Online
mussel populations showed gonads in resting and post-spawning
stages.
In April 2005, the predominant stage was the mature phase
and in some stations mussels at spawning stage were also
observed, mainly in Llanes and San Vicente (Fig. 3). In July 2005
most mussels showed spawning and post-spawning gonad stages,
except mussels from Estaca de Bares and Llanes where all indi-
viduals were still at mature stage. Some localities, such as
Camelle, Sega~no and Gorliz showed the start of a second
gametogenic cycle. In October 2005 all studied mussel pop-
ulations showed gonads mainly at post-spawning and resting
stages. Some locations, such as Ons, Caldebarcos, Sega~no,
Pedre~na, Muskiz, Arrigunaga, Mundaka and Hondarribia had
already started the new reproductive cycle.
In April 2006, most mussel populations showed gonads in
mature and spawning stages, except those from Suances and
Mutriku where all mussels were spawning (Fig. 3). Some few
mussels from Agui~no, Arrigunaga and Gorliz were at post-
spawning stage. Overall, most studied mussel populations
showed one reproductive cycle in 2003 and 2005, and seemingly
two reproductive cycles in 2004.
A hierarchical cluster analysis of GI values showed two main
branches separating samplings of February, April and July (mainly
gonads in mature and spawning stages) from those of September
and October (gonads mainly at post-spawning or resting and early
gametogenesis stages) (Fig. 4a). Furthermore, the dendrogram
shows that data from samplings performed in April 2003, 2005 and
2006 are closely related but April 2004 was more related with
February 2004 and July 2003, 2004 and 2005. On the other hand, no
significant discrimination between most oil impacted and less
impacted localities was observed (data not shown).
88 | J. Environ. Monit., 2011, 13, 84–94
Gonad histopathology
The histopathological study of the gonad (Table 2) showed
a high prevalence of oocyte atresia in female mussels in
April 2003 (Fig. 2d), together with a high prevalence of haemo-
cytic infiltration of gonad follicles (Fig. 2e). In April of the
following year 2004 the atresia disappeared but widespread small
sized mature follicles were observed in most mussel populations
(Table 2). Both male and female mussels showed small follicles
containing developing and full developed gametes, together with
follicles of larger size with no gametes and surrounded by a high
amount of reserve material (Fig. 2f). Overall, relative volume of
gonad follicles was small compared with the rest of the speci-
mens. Haemocytic infiltration of follicles was present in several
stations. In April 2005, few histopathological alterations were
observed. In females, atresia was scarce and low levels of infil-
tration of follicles by haemocytes were observed (Table 2).
However, in April 2006 a high prevalence of abnormal gametes
was observed in female mussels from six out of 22 studied pop-
ulations (Table 2). Some abnormal gametes showed dispersed
cytoplasm in the form of eosinophilic spheres (Fig. 2g). Other
abnormal gametes were characterized by higher staining inten-
sity than normal gametes. They progressively loose structure
(nuclei and cytoplasm became indistinguishable) and finally they
were released to the follicle lumen. Both types of abnormal
gametes were studied by TUNEL assay to establish whether they
appear as a result of apoptotic events. Whereas positive nuclei
were evidenced in the positive control slides containing apoptotic
HL60 cells, no positive nuclei were detected in studied mussel
gonad sections indicating that abnormal gametes could be orig-
inated by degenerative processes such as necrosis and not by
This journal is ª The Royal Society of Chemistry 2011
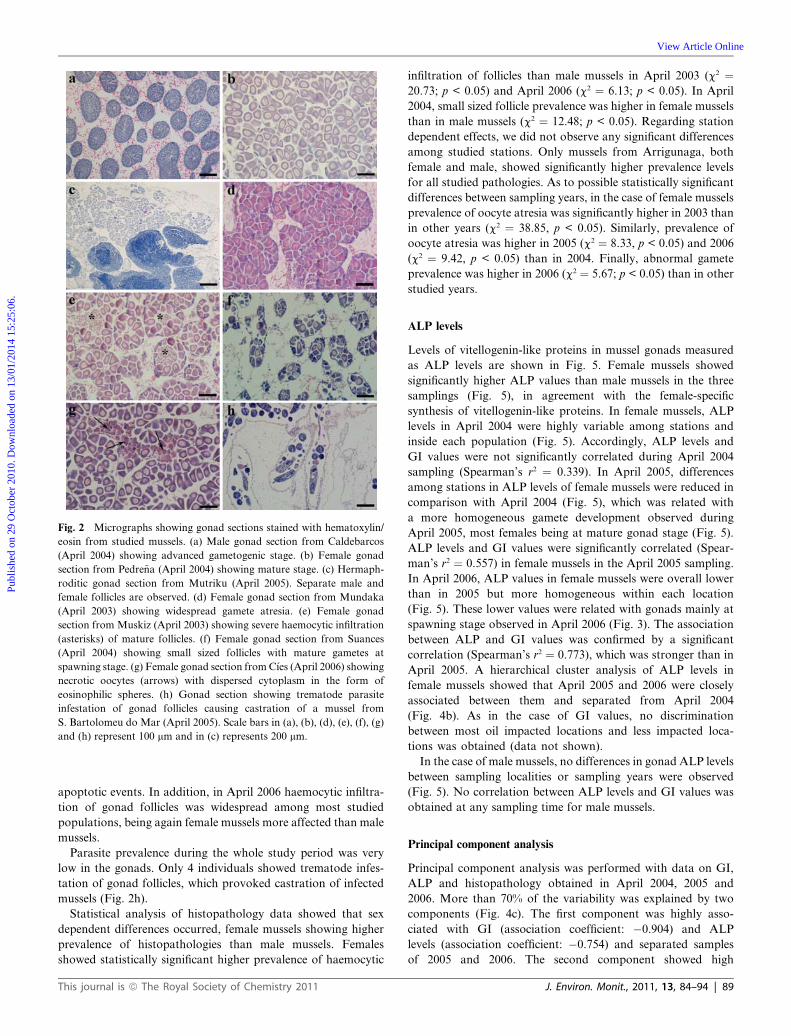
Fig. 2 Micrographs showing gonad sections stained with hematoxylin/
eosin from studied mussels. (a) Male gonad section from Caldebarcos
(April 2004) showing advanced gametogenic stage. (b) Female gonad
section from Pedre~na (April 2004) showing mature stage. (c) Hermaph-
roditic gonad section from Mutriku (April 2005). Separate male and
female follicles are observed. (d) Female gonad section from Mundaka
(April 2003) showing widespread gamete atresia. (e) Female gonad
section from Muskiz (April 2003) showing severe haemocytic infiltration
(asterisks) of mature follicles. (f) Female gonad section from Suances
(April 2004) showing small sized follicles with mature gametes at
spawning stage. (g) Female gonad section from C�ıes (April 2006) showing
necrotic oocytes (arrows) with dispersed cytoplasm in the form of
eosinophilic spheres. (h) Gonad section showing trematode parasite
infestation of gonad follicles causing castration of a mussel from
S. Bartolomeu do Mar (April 2005). Scale bars in (a), (b), (d), (e), (f), (g)
and (h) represent 100 mm and in (c) represents 200 mm.
Publ
ishe
d on
29
Oct
ober
201
0. D
ownl
oade
d on
13/
01/2
014
15:2
5:06
.
View Article Online
apoptotic events. In addition, in April 2006 haemocytic infiltra-
tion of gonad follicles was widespread among most studied
populations, being again female mussels more affected than male
mussels.
Parasite prevalence during the whole study period was very
low in the gonads. Only 4 individuals showed trematode infes-
tation of gonad follicles, which provoked castration of infected
mussels (Fig. 2h).
Statistical analysis of histopathology data showed that sex
dependent differences occurred, female mussels showing higher
prevalence of histopathologies than male mussels. Females
showed statistically significant higher prevalence of haemocytic
This journal is ª The Royal Society of Chemistry 2011
infiltration of follicles than male mussels in April 2003 (c2 ¼20.73; p < 0.05) and April 2006 (c2 ¼ 6.13; p < 0.05). In April
2004, small sized follicle prevalence was higher in female mussels
than in male mussels (c2 ¼ 12.48; p < 0.05). Regarding station
dependent effects, we did not observe any significant differences
among studied stations. Only mussels from Arrigunaga, both
female and male, showed significantly higher prevalence levels
for all studied pathologies. As to possible statistically significant
differences between sampling years, in the case of female mussels
prevalence of oocyte atresia was significantly higher in 2003 than
in other years (c2 ¼ 38.85, p < 0.05). Similarly, prevalence of
oocyte atresia was higher in 2005 (c2 ¼ 8.33, p < 0.05) and 2006
(c2 ¼ 9.42, p < 0.05) than in 2004. Finally, abnormal gamete
prevalence was higher in 2006 (c2 ¼ 5.67; p < 0.05) than in other
studied years.
ALP levels
Levels of vitellogenin-like proteins in mussel gonads measured
as ALP levels are shown in Fig. 5. Female mussels showed
significantly higher ALP values than male mussels in the three
samplings (Fig. 5), in agreement with the female-specific
synthesis of vitellogenin-like proteins. In female mussels, ALP
levels in April 2004 were highly variable among stations and
inside each population (Fig. 5). Accordingly, ALP levels and
GI values were not significantly correlated during April 2004
sampling (Spearman’s r2 ¼ 0.339). In April 2005, differences
among stations in ALP levels of female mussels were reduced in
comparison with April 2004 (Fig. 5), which was related with
a more homogeneous gamete development observed during
April 2005, most females being at mature gonad stage (Fig. 5).
ALP levels and GI values were significantly correlated (Spear-
man’s r2 ¼ 0.557) in female mussels in the April 2005 sampling.
In April 2006, ALP values in female mussels were overall lower
than in 2005 but more homogeneous within each location
(Fig. 5). These lower values were related with gonads mainly at
spawning stage observed in April 2006 (Fig. 3). The association
between ALP and GI values was confirmed by a significant
correlation (Spearman’s r2 ¼ 0.773), which was stronger than in
April 2005. A hierarchical cluster analysis of ALP levels in
female mussels showed that April 2005 and 2006 were closely
associated between them and separated from April 2004
(Fig. 4b). As in the case of GI values, no discrimination
between most oil impacted locations and less impacted loca-
tions was obtained (data not shown).
In the case of male mussels, no differences in gonad ALP levels
between sampling localities or sampling years were observed
(Fig. 5). No correlation between ALP levels and GI values was
obtained at any sampling time for male mussels.
Principal component analysis
Principal component analysis was performed with data on GI,
ALP and histopathology obtained in April 2004, 2005 and
2006. More than 70% of the variability was explained by two
components (Fig. 4c). The first component was highly asso-
ciated with GI (association coefficient: �0.904) and ALP
levels (association coefficient: �0.754) and separated samples
of 2005 and 2006. The second component showed high
J. Environ. Monit., 2011, 13, 84–94 | 89

Fig. 3 Graphs showing gamete development and gonad index from each station at each sampling time. Stacking-bar graphs show the percentage of
individuals at each gametogenic stage at each sampling time. Superimposed continuous lines represent gonad index values for each sampling point at
each sampling month. F: February; A: April; J: July; S: September; O: October.
Publ
ishe
d on
29
Oct
ober
201
0. D
ownl
oade
d on
13/
01/2
014
15:2
5:06
.
View Article Online
association with small sized follicle prevalence (association
coefficient: �0.696) and haemocytic infiltration of gonad
follicles (association coefficient: 0.654) and separated samples
of 2004 from those of 2005 and 2006. Therefore, variability in
90 | J. Environ. Monit., 2011, 13, 84–94
the spatial distribution of studied samples was based on
gamete development (GI and ALP levels) and observed
histopathologies such as small sized follicles and haemocytic
infiltration.
This journal is ª The Royal Society of Chemistry 2011
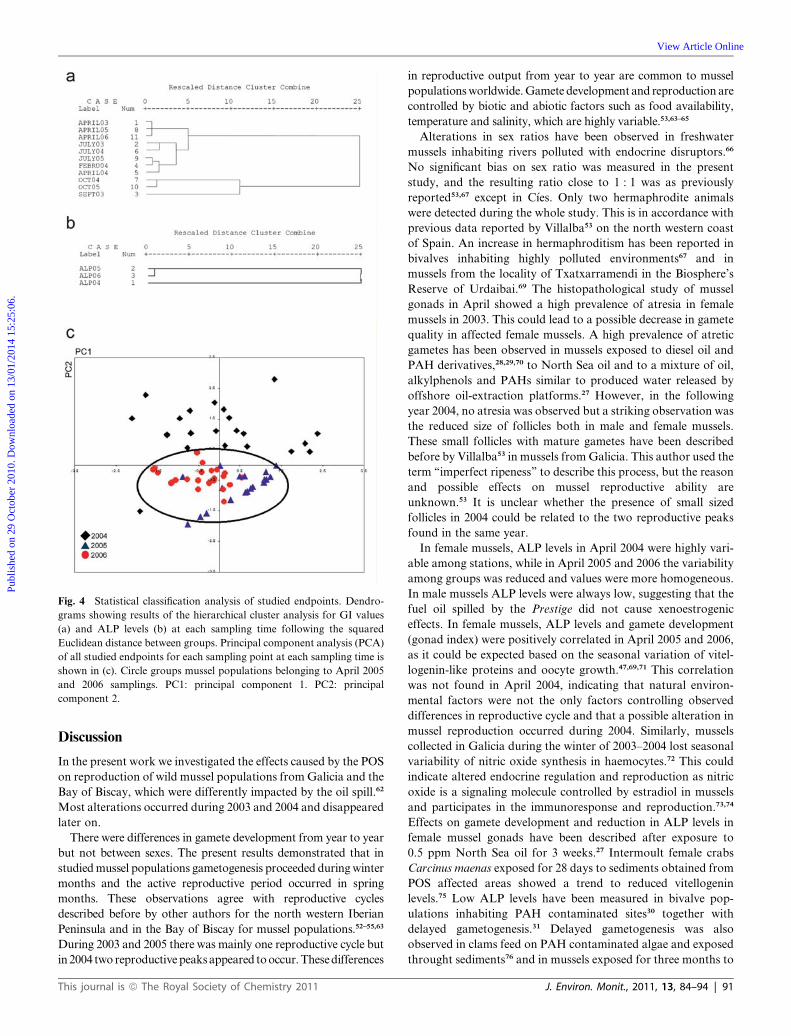
Fig. 4 Statistical classification analysis of studied endpoints. Dendro-
grams showing results of the hierarchical cluster analysis for GI values
(a) and ALP levels (b) at each sampling time following the squared
Euclidean distance between groups. Principal component analysis (PCA)
of all studied endpoints for each sampling point at each sampling time is
shown in (c). Circle groups mussel populations belonging to April 2005
and 2006 samplings. PC1: principal component 1. PC2: principal
component 2.
Publ
ishe
d on
29
Oct
ober
201
0. D
ownl
oade
d on
13/
01/2
014
15:2
5:06
.
View Article Online
Discussion
In the present work we investigated the effects caused by the POS
on reproduction of wild mussel populations from Galicia and the
Bay of Biscay, which were differently impacted by the oil spill.62
Most alterations occurred during 2003 and 2004 and disappeared
later on.
There were differences in gamete development from year to year
but not between sexes. The present results demonstrated that in
studied mussel populations gametogenesis proceeded during winter
months and the active reproductive period occurred in spring
months. These observations agree with reproductive cycles
described before by other authors for the north western Iberian
Peninsula and in the Bay of Biscay for mussel populations.52–55,63
During 2003 and 2005 there was mainly one reproductive cycle but
in 2004 two reproductive peaks appeared to occur. These differences
This journal is ª The Royal Society of Chemistry 2011
in reproductive output from year to year are common to mussel
populations worldwide. Gamete development and reproduction are
controlled by biotic and abiotic factors such as food availability,
temperature and salinity, which are highly variable.53,63–65
Alterations in sex ratios have been observed in freshwater
mussels inhabiting rivers polluted with endocrine disruptors.66
No significant bias on sex ratio was measured in the present
study, and the resulting ratio close to 1 : 1 was as previously
reported53,67 except in C�ıes. Only two hermaphrodite animals
were detected during the whole study. This is in accordance with
previous data reported by Villalba53 on the north western coast
of Spain. An increase in hermaphroditism has been reported in
bivalves inhabiting highly polluted environments67 and in
mussels from the locality of Txatxarramendi in the Biosphere’s
Reserve of Urdaibai.69 The histopathological study of mussel
gonads in April showed a high prevalence of atresia in female
mussels in 2003. This could lead to a possible decrease in gamete
quality in affected female mussels. A high prevalence of atretic
gametes has been observed in mussels exposed to diesel oil and
PAH derivatives,28,29,70 to North Sea oil and to a mixture of oil,
alkylphenols and PAHs similar to produced water released by
offshore oil-extraction platforms.27 However, in the following
year 2004, no atresia was observed but a striking observation was
the reduced size of follicles both in male and female mussels.
These small follicles with mature gametes have been described
before by Villalba53 in mussels from Galicia. This author used the
term ‘‘imperfect ripeness’’ to describe this process, but the reason
and possible effects on mussel reproductive ability are
unknown.53 It is unclear whether the presence of small sized
follicles in 2004 could be related to the two reproductive peaks
found in the same year.
In female mussels, ALP levels in April 2004 were highly vari-
able among stations, while in April 2005 and 2006 the variability
among groups was reduced and values were more homogeneous.
In male mussels ALP levels were always low, suggesting that the
fuel oil spilled by the Prestige did not cause xenoestrogenic
effects. In female mussels, ALP levels and gamete development
(gonad index) were positively correlated in April 2005 and 2006,
as it could be expected based on the seasonal variation of vitel-
logenin-like proteins and oocyte growth.47,69,71 This correlation
was not found in April 2004, indicating that natural environ-
mental factors were not the only factors controlling observed
differences in reproductive cycle and that a possible alteration in
mussel reproduction occurred during 2004. Similarly, mussels
collected in Galicia during the winter of 2003–2004 lost seasonal
variability of nitric oxide synthesis in haemocytes.72 This could
indicate altered endocrine regulation and reproduction as nitric
oxide is a signaling molecule controlled by estradiol in mussels
and participates in the immunoresponse and reproduction.73,74
Effects on gamete development and reduction in ALP levels in
female mussel gonads have been described after exposure to
0.5 ppm North Sea oil for 3 weeks.27 Intermoult female crabs
Carcinus maenas exposed for 28 days to sediments obtained from
POS affected areas showed a trend to reduced vitellogenin
levels.75 Low ALP levels have been measured in bivalve pop-
ulations inhabiting PAH contaminated sites30 together with
delayed gametogenesis.31 Delayed gametogenesis was also
observed in clams feed on PAH contaminated algae and exposed
throught sediments76 and in mussels exposed for three months to
J. Environ. Monit., 2011, 13, 84–94 | 91
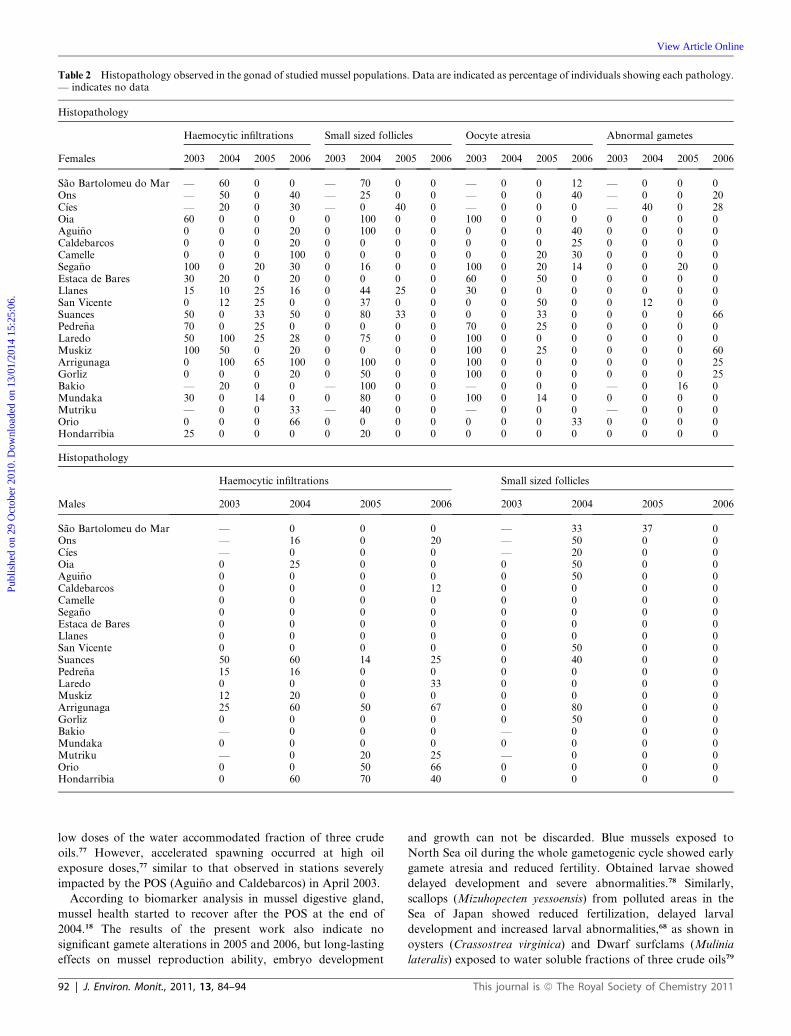
Table 2 Histopathology observed in the gonad of studied mussel populations. Data are indicated as percentage of individuals showing each pathology.— indicates no data
Histopathology
Haemocytic infiltrations Small sized follicles Oocyte atresia Abnormal gametes
Females 2003 2004 2005 2006 2003 2004 2005 2006 2003 2004 2005 2006 2003 2004 2005 2006
Sao Bartolomeu do Mar — 60 0 0 — 70 0 0 — 0 0 12 — 0 0 0Ons — 50 0 40 — 25 0 0 — 0 0 40 — 0 0 20C�ıes — 20 0 30 — 0 40 0 — 0 0 0 — 40 0 28Oia 60 0 0 0 0 100 0 0 100 0 0 0 0 0 0 0Agui~no 0 0 0 20 0 100 0 0 0 0 0 40 0 0 0 0Caldebarcos 0 0 0 20 0 0 0 0 0 0 0 25 0 0 0 0Camelle 0 0 0 100 0 0 0 0 0 0 20 30 0 0 0 0Sega~no 100 0 20 30 0 16 0 0 100 0 20 14 0 0 20 0Estaca de Bares 30 20 0 20 0 0 0 0 60 0 50 0 0 0 0 0Llanes 15 10 25 16 0 44 25 0 30 0 0 0 0 0 0 0San Vicente 0 12 25 0 0 37 0 0 0 0 50 0 0 12 0 0Suances 50 0 33 50 0 80 33 0 0 0 33 0 0 0 0 66Pedre~na 70 0 25 0 0 0 0 0 70 0 25 0 0 0 0 0Laredo 50 100 25 28 0 75 0 0 100 0 0 0 0 0 0 0Muskiz 100 50 0 20 0 0 0 0 100 0 25 0 0 0 0 60Arrigunaga 0 100 65 100 0 100 0 0 100 0 0 0 0 0 0 25Gorliz 0 0 0 20 0 50 0 0 100 0 0 0 0 0 0 25Bakio — 20 0 0 — 100 0 0 — 0 0 0 — 0 16 0Mundaka 30 0 14 0 0 80 0 0 100 0 14 0 0 0 0 0Mutriku — 0 0 33 — 40 0 0 — 0 0 0 — 0 0 0Orio 0 0 0 66 0 0 0 0 0 0 0 33 0 0 0 0Hondarribia 25 0 0 0 0 20 0 0 0 0 0 0 0 0 0 0
Histopathology
Haemocytic infiltrations Small sized follicles
Males 2003 2004 2005 2006 2003 2004 2005 2006
Sao Bartolomeu do Mar — 0 0 0 — 33 37 0Ons — 16 0 20 — 50 0 0C�ıes — 0 0 0 — 20 0 0Oia 0 25 0 0 0 50 0 0Agui~no 0 0 0 0 0 50 0 0Caldebarcos 0 0 0 12 0 0 0 0Camelle 0 0 0 0 0 0 0 0Sega~no 0 0 0 0 0 0 0 0Estaca de Bares 0 0 0 0 0 0 0 0Llanes 0 0 0 0 0 0 0 0San Vicente 0 0 0 0 0 50 0 0Suances 50 60 14 25 0 40 0 0Pedre~na 15 16 0 0 0 0 0 0Laredo 0 0 0 33 0 0 0 0Muskiz 12 20 0 0 0 0 0 0Arrigunaga 25 60 50 67 0 80 0 0Gorliz 0 0 0 0 0 50 0 0Bakio — 0 0 0 — 0 0 0Mundaka 0 0 0 0 0 0 0 0Mutriku — 0 20 25 — 0 0 0Orio 0 0 50 66 0 0 0 0Hondarribia 0 60 70 40 0 0 0 0
Publ
ishe
d on
29
Oct
ober
201
0. D
ownl
oade
d on
13/
01/2
014
15:2
5:06
.
View Article Online
low doses of the water accommodated fraction of three crude
oils.77 However, accelerated spawning occurred at high oil
exposure doses,77 similar to that observed in stations severely
impacted by the POS (Agui~no and Caldebarcos) in April 2003.
According to biomarker analysis in mussel digestive gland,
mussel health started to recover after the POS at the end of
2004.18 The results of the present work also indicate no
significant gamete alterations in 2005 and 2006, but long-lasting
effects on mussel reproduction ability, embryo development
92 | J. Environ. Monit., 2011, 13, 84–94
and growth can not be discarded. Blue mussels exposed to
North Sea oil during the whole gametogenic cycle showed early
gamete atresia and reduced fertility. Obtained larvae showed
delayed development and severe abnormalities.78 Similarly,
scallops (Mizuhopecten yessoensis) from polluted areas in the
Sea of Japan showed reduced fertilization, delayed larval
development and increased larval abnormalities,68 as shown in
oysters (Crassostrea virginica) and Dwarf surfclams (Mulinia
lateralis) exposed to water soluble fractions of three crude oils79
This journal is ª The Royal Society of Chemistry 2011
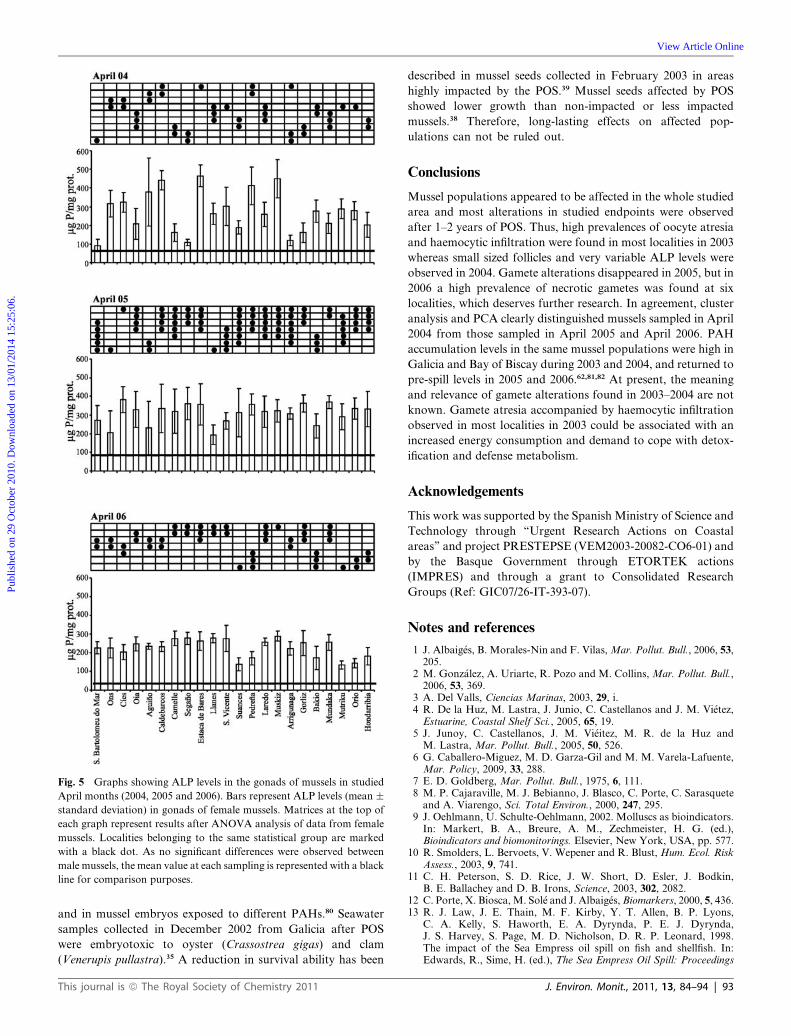
Fig. 5 Graphs showing ALP levels in the gonads of mussels in studied
April months (2004, 2005 and 2006). Bars represent ALP levels (mean �standard deviation) in gonads of female mussels. Matrices at the top of
each graph represent results after ANOVA analysis of data from female
mussels. Localities belonging to the same statistical group are marked
with a black dot. As no significant differences were observed between
male mussels, the mean value at each sampling is represented with a black
line for comparison purposes.
Publ
ishe
d on
29
Oct
ober
201
0. D
ownl
oade
d on
13/
01/2
014
15:2
5:06
.
View Article Online
and in mussel embryos exposed to different PAHs.80 Seawater
samples collected in December 2002 from Galicia after POS
were embryotoxic to oyster (Crassostrea gigas) and clam
(Venerupis pullastra).35 A reduction in survival ability has been
This journal is ª The Royal Society of Chemistry 2011
described in mussel seeds collected in February 2003 in areas
highly impacted by the POS.39 Mussel seeds affected by POS
showed lower growth than non-impacted or less impacted
mussels.38 Therefore, long-lasting effects on affected pop-
ulations can not be ruled out.
Conclusions
Mussel populations appeared to be affected in the whole studied
area and most alterations in studied endpoints were observed
after 1–2 years of POS. Thus, high prevalences of oocyte atresia
and haemocytic infiltration were found in most localities in 2003
whereas small sized follicles and very variable ALP levels were
observed in 2004. Gamete alterations disappeared in 2005, but in
2006 a high prevalence of necrotic gametes was found at six
localities, which deserves further research. In agreement, cluster
analysis and PCA clearly distinguished mussels sampled in April
2004 from those sampled in April 2005 and April 2006. PAH
accumulation levels in the same mussel populations were high in
Galicia and Bay of Biscay during 2003 and 2004, and returned to
pre-spill levels in 2005 and 2006.62,81,82 At present, the meaning
and relevance of gamete alterations found in 2003–2004 are not
known. Gamete atresia accompanied by haemocytic infiltration
observed in most localities in 2003 could be associated with an
increased energy consumption and demand to cope with detox-
ification and defense metabolism.
Acknowledgements
This work was supported by the Spanish Ministry of Science and
Technology through ‘‘Urgent Research Actions on Coastal
areas’’ and project PRESTEPSE (VEM2003-20082-CO6-01) and
by the Basque Government through ETORTEK actions
(IMPRES) and through a grant to Consolidated Research
Groups (Ref: GIC07/26-IT-393-07).
Notes and references
1 J. Albaig�es, B. Morales-Nin and F. Vilas, Mar. Pollut. Bull., 2006, 53,205.
2 M. Gonz�alez, A. Uriarte, R. Pozo and M. Collins, Mar. Pollut. Bull.,2006, 53, 369.
3 A. Del Valls, Ciencias Marinas, 2003, 29, i.4 R. De la Huz, M. Lastra, J. Junio, C. Castellanos and J. M. Vi�etez,
Estuarine, Coastal Shelf Sci., 2005, 65, 19.5 J. Junoy, C. Castellanos, J. M. Vi�eitez, M. R. de la Huz and
M. Lastra, Mar. Pollut. Bull., 2005, 50, 526.6 G. Caballero-Miguez, M. D. Garza-Gil and M. M. Varela-Lafuente,
Mar. Policy, 2009, 33, 288.7 E. D. Goldberg, Mar. Pollut. Bull., 1975, 6, 111.8 M. P. Cajaraville, M. J. Bebianno, J. Blasco, C. Porte, C. Sarasquete
and A. Viarengo, Sci. Total Environ., 2000, 247, 295.9 J. Oehlmann, U. Schulte-Oehlmann, 2002. Molluscs as bioindicators.
In: Markert, B. A., Breure, A. M., Zechmeister, H. G. (ed.),Bioindicators and biomonitorings. Elsevier, New York, USA, pp. 577.
10 R. Smolders, L. Bervoets, V. Wepener and R. Blust, Hum. Ecol. RiskAssess., 2003, 9, 741.
11 C. H. Peterson, S. D. Rice, J. W. Short, D. Esler, J. Bodkin,B. E. Ballachey and D. B. Irons, Science, 2003, 302, 2082.
12 C. Porte, X. Biosca, M. Sol�e and J. Albaig�es, Biomarkers, 2000, 5, 436.13 R. J. Law, J. E. Thain, M. F. Kirby, Y. T. Allen, B. P. Lyons,
C. A. Kelly, S. Haworth, E. A. Dyrynda, P. E. J. Dyrynda,J. S. Harvey, S. Page, M. D. Nicholson, D. R. P. Leonard, 1998.The impact of the Sea Empress oil spill on fish and shellfish. In:Edwards, R., Sime, H. (ed.), The Sea Empress Oil Spill: Proceedings
J. Environ. Monit., 2011, 13, 84–94 | 93

Publ
ishe
d on
29
Oct
ober
201
0. D
ownl
oade
d on
13/
01/2
014
15:2
5:06
.
View Article Online
of the International Conference held in Cardiff, 11–13 February 1998,Chartered Institute of Water and Environmental Management,London, UK, pp. 109–136.
14 S. M. Moreira, M. Moreira-Santos, R. Ribeiro and L. Guilhermino,Ecotoxicology, 2004, 13, 619.
15 G. Boquen�e, S. Chantereau, C. Cl�erendeau, E. Beausir, D. M�enard,B. Raffin, C. Minier, T. Burgeot, A. P. Leszkowicz andJ. F. Narbonne, Aquat. Living Resour., 2004, 17, 309.
16 I. Marig�omez, M. Soto, C. Cancio, A. Orbea, L. Garmendia andM. P. Cajaraville, Mar. Pollut. Bull., 2006, 53, 287.
17 A. Orbea, L. Garmendia, I. Marig�omez and M. P. Cajaraville, Mar.Ecol.: Prog. Ser., 2006, 306, 177.
18 M. P. Cajaraville, L. Garmendia, A. Orbea, R. Werding, A. G�omez-Mendikute, U. Izagirre, M. Soto and I. Marig�omez, Mar. Environ.Res., 2006, 62, S337.
19 Exxon Valdez Oil Spill Trustee Council, 2006. Exxon Valdez Oil SpillRestoration Plan. Update on injured resources and services, 2006,Anchorage, AK, USA. pp. 42.
20 WHO/IPCS (World Health Organization/International Progamme onChemical Safety), Global assessment of the state-of-the-science ofendocrine disruptors, 2002, WHO/IPCS/EDC/02.2, Geneva,Switzerland.
21 P. Matthiessen, Pure Appl. Chem., 2003, 75, 2197.22 S. Y. Sol, L. L. Johnson, B. H. Horness and T. K. Collier, Mar. Pollut.
Bull., 2000, 40, 1139.23 S. D. Rice, R. E. Thomas, R. A. Heintz, A. C. Wertheimer,
M. L. Murphy, M. G. Carls, J. W. Short, A. Moles, Synthesis oflong-term impacts to pink salmon following the Exxon Valdez oilspill: persistence, toxicity, sensitivity, and controversy. Final Reportproject 99329, 2001,pp. 77.
24 M. G. Carls, G. D. Marty and J. E. Hose, Can. J. Fish. Aquat. Sci.,2002, 59, 153.
25 C. Porte, G. Janer, L. C. Lorusso, M. Ortiz-Zarragoitia,M. P. Cajaraville, C. Fossi and L. Canesi, Comp. Biochem. Physiol.Part C, 2006, 143, 303.
26 J. Lintelmann, A. Katayama, N. Kurihara, L. Shore and A. Wenzel,Pure Appl. Chem., 2003, 75, 631.
27 M. Ortiz-Zarragoitia and M. P. Cajaraville, Arch. Environ. Contam.Toxicol., 2006, 50, 361.
28 D. M. Lowe, Mar. Ecol. Res., 1988, 22, 243.29 D. Lowe and R. K. Pipe, Mar. Environ. Res., 1987, 22, 243.30 F. Gagn�e, C. Blaise, J. Pellerin and S. Gauthier-Clerc, Mar. Environ.
Res., 2002, 53, 295.31 S. Gauthier-Clerc, J. Pellerin, C. Blaise and F. Gagn�e, Comp.
Biochem. Physiol., Part C, 2002, 131, 457.32 J. M. Navas and H. Segner, Aquat. Toxicol., 2000, 51, 79.33 J. M. Nicolas, Aquat. Toxicol., 1999, 45, 77.34 M. M. H. Van Lipzig, N. P. E. Vermeulen, R. Gusinu, J. Legler,
H. Frank, A. Seidel and J. H. N. Meerman, Environ. Toxicol.Pharmacol., 2005, 19, 41.
35 R. Beiras and L. Saco-Alvarez, Water, Air, Soil Pollut., 2006, 177,457.
36 J. M. Navas, M. Bab�ın, S. Casado, C. Fern�andez and J. V. Tarazona,Mar. Environ. Res., 2006, 62, S352.
37 S. S. Rossi and J. W. Anderson, Water, Air, Soil Pollut., 1978, 9, 155.38 L. G. Peteiro, J. M. F. Babarro, U. Labarta and M. J. Fern�andez-
Reiriz, ICES J. Mar. Sci., 2006, 63, 1005.39 U. Labarta, M. J. Fern�andez-Reiriz, J. L. Garrido,
J. M. F. Babarro, J. M. Bayona and J. Albaig�es, Mar. Ecol.:Prog. Ser., 2005, 302, 135.
40 L. G. Peteiro, U. Labarta and M. J. Fern�andez-Reiriz, Comp.Biochem. Physiol. Part C, 2007, 145, 588.
41 A. Arukwe and A. Goksøyr, Comp. Hepatol., 2003, 2, 4.42 M. G. Marin and V. Matozzo, Mar. Pollut. Bull., 2004, 48, 835.43 A. Goksøyr, J. Toxicol. Environ. Health, Part A, 2006, 69, 175.44 T. H. Hutchinson, G. T. Ankley, H. Segner and C. R. Tyler, Environ.
Health Persp., 2006, 114(Suppl. 1), 106.45 T. Matsumoto, A. M. Nakamura, K. Mori and T. Kayano, Zool. Sci.,
2003, 20, 37.46 M. Osada, M. Harata, M. Kishida and A. Kijama, Mol. Reprod. Dev.,
2004, 67, 273.47 C. Blaise, F. Gagn�e, J. Pellerin and P. D. Hansen, Environ. Toxicol.,
1999, 14, 455.48 M. Osada, T. Takamura, H. Sato and K. Mori, J. Exp. Zool., 2003,
299a, 172.
94 | J. Environ. Monit., 2011, 13, 84–94
49 A. M. Puinean, P. Labadie, E. M. Hill, M. Osada, M. Kishida,R. Nakao, A. Novillo, I. P. Callard and J. M. Rotchell, Aquat.Toxicol., 2006, 79, 376.
50 G. Janer and C. Porte, Ecotoxicology, 2007, 16, 145.51 V. Matozzo, F. Gagn�e, M. G. Marin, F. Ricciardi and C. Blaise,
Environ. Int., 2008, 34, 531.52 S. Ogueta, B. J. G�omez, T. Serrano, M. Soto, Proceedings of the 5th
National Congress on Aquaculture (Sant Carles de la R�apita, Spain).Barcelona University Press, Barcelona, Spain, 1995, 169.
53 A. Villalba, Aquaculture, 1995, 130, 269.54 A. T. Mikhailov, M. Torrado, J. M�endez and M. J. L�opez, Mar. Biol.,
1996, 126, 77.55 M. P. Su�arez, C. Alvarez, P. Molist and F. San Juan, J. Shellfish Res.,
2005, 24, 531.56 M. Gamble, I. Wilson, The hematoxylin and eosin. In: Bancroft,
J. D., Gamble, M. (ed.), Theory and practice of histologicaltechniques.Churchill Livingstone-Elsevier Science, London, UK,2002, pp. 125–138.
57 R. Seed, Oecologia, 1969, 3, 277.58 Y. Kim, K. A. Ashton-Alcox, E. N. Powell, Gonadal analysis. NOAA
Histological techniques for marine bivalve mollusks: Update. NOAATechnical Memories NOS NCCOS 27,Silver Spring, USA, 2006, pp.1–18.
59 G. Gagn�e, C. Blaise, J. Pellerin, E. Pelletier, M. Douville, S. Gauthier-Clerc and L. Viglino, Comp. Biochem. Physiol., Part C, 2003, 134, 189.
60 M. G. Stanton, Anal. Biochem., 1968, 22, 27.61 H. J. Fryer, G. E. Davis, M. Manthorpe and S. Varon, Anal.
Biochem., 1986, 153, 262.62 L. Bartolom�e, M. Deusto, N. Etxebarria, P. Navarro, A. Usobiaga
and O. Zuloaga, J. Chromatogr., A, 2007, 1157, 369.63 J. C�aceres-Mart�ınez and A. Figueras, Aquaculture, 1998, 162, 141.64 R. I. E. Newell, Species profiles: life histories and environmental
requirements of coastal fishes and invertebrates (North and Mid-Atlantic) –blue mussel. US Fish and Wildlife Service of Biology andReproduction 82 (11.102). US Army Corps of Engineers, TREL-82-4, 1989, 25.
65 F. G. Figueiras, U. Labarta and M. J. Fern�andez-Reiriz,Hydrobiologia, 2002, 484, 121.
66 C. Blaise, F. Gagn�e, M. Salazar, S. Salazar, S. Trottier andP. D. Hansen, Fresenius Environ. Bull., 2003, 12, 865.
67 E. Kenchington, B. MacDonald, L. Cao, D. Tsagkarakis andE. Zouros, Genetics, 2002, 161, 1579.
68 M. A. Vaschenko, I. G. Syasina, P. M. Zhadan and L. A. Medvedeva,Hydrobiologia, 1997, 352, 231.
69 M. Ortiz-Zarragoitia and M. P. Cajaraville, Ecotoxicol. Environ. Saf.,2010, 73, 693.
70 A. Goksøyr, A. Arukwe, J. Larsson, M. P. Cajaraville, L. Hauser,B. D. Nilsen, D. Lowe, P. Matthiessen, Molecular/cellular processesand the impact on reproduction. In: Lawrence, A., Hemingway, K.(ed.), Effects of pollution on fish. Blackwell Publishing, Oxford, UK,2003, pp. 179–220.
71 C. Blaise, F. Gagn�e, J. Pellerin, P. D. Hansen and S. Trottier, Environ.Toxicol., 2002, 17, 170.
72 A. Novas, R. Barcia and J. I. Ramos-Mart�ınez, Aquat. Toxicol., 2007,85, 285.
73 L. Barbin, I. Boarini, P. G. Borasio, P. Barion, S. Fiorini, R. Rossiand C. Bioindi, Gen. Comp. Endocrinol., 2003, 130, 215.
74 G. B. Stefano, P. Cadet, K. Mantione, J. J. Cho, D. Jones andW. Zhu, Endocrinology, 2003, 144, 1234.
75 C. Morales-Caselles, M. L. Mart�ın D�ıaz, I. Riba and T. A. DelValls,Fresenius Environ. Bull., 2009, 18, 140.
76 H. Frouin, J. Pellerin, M. Fournier, E. Pelletier, P. Richard,N. Pichaud, C. Rouleau and F. Garnerot, Aquat. Toxicol., 2007, 82,120.
77 M. P. Cajaraville, I. Marig�omez and E. Angulo, Comp. Biochem.Physiol., Part C: Pharmacol., Toxicol. Endocrinol., 1992, 102, 103.
78 T. Baussant, M. Ortiz-Zarragoitia, M. P. Cajaraville,R. K. Bechmann, I. C. Taban, S. Sanni. Mar. Pollut. Bull., submitted.
79 A. Renzoni, Mar. Pollut. Bull., 1975, 6, 125.80 J. Bellas, L. Saco- �Alvarez, O. Nieto and R. Beiras, Mar. Pollut. Bull.,
2008, 57, 493.81 J. A. Soriano, L. Vi~nas, M. A. Franco, J. J. Gonz�alez, M. H. Nguyen,
J. M. Bayona and J. Albaig�es, J. Environ. Monit., 2007, 9, 1018.82 B. Fern�andez, M. Albentosa, L. Vi~nas, A. Franco, J. J. Gonz�alez and
J. A. Campillo, Ecotoxicology, 2010, 19, 735.
This journal is ª The Royal Society of Chemistry 2011




















