
The challenge of choosing environmental indicators of anthropogenic impacts in estuaries
Upload
southernmissCategory
view
1download
0
Effects of Sediment Contaminants and Environmental Gradients on Macrobenthic Community Trophic Structure in Gulf of Mexico Estuaries
Steven S. Brown; Gary R. Gaston; Chet F. Rakocinski; Richard W. Heard
Estuaries, Vol. 23, No.3. (Jun., 2000), pp. 411-424.
STOR
Stable URL: http://links.jstor.org/sici?sici=0160-8347%28200006%2923%3A3%3C411%3AEOSCAE%3E2.0.CO%3B2-S
Estuaries is currently published by Estuarine Research Federation.
Your use of the JSTOR archive indicates your acceptance of JSTOR' s Terms and Conditions of Use, available at http://www.jstor.org/aboutiterms.html. JSTOR's Terms and Conditions of Use provides, in part, that unless you
have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content in the JSTOR archive only for your personal, non-commercial use.
Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at http://www.jstor.org/journals/estuarine.html.
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed page of such transmission.
JSTOR is an independent not-for-profit organization dedicated to creating and preserving a digital archive of scholarly journals. For more information regarding JSTOR, please contact [email protected].
http://www .j stor.org/ Tue Jul 18 12:52:022006
®

Estuaries Vol. 23, No.3, p. 411-424 June 2000
Effects of Sediment Contaminants and Environmental
Gradients on Macrobenthic Community Trophic Structure
in Gulf of Mexico Estuaries
STEVEN s. BROWN l
GARY R. GASTON
Biology Department University of Mississippi University, Mississippi 38677
CHET F. RAKOCINSKI
RICHARD W. HEARD
University of Southern Mississippi Institute of Marine Sciences Gulf Coast Research Laboratory Ocean springs, Mississippi 39564
ABSTRACT: Macrobenthic communities from estuaries throughout the northern Gulf of Mexico were studied to assess the influence of sediment contaminants and natural environmental factors on macrobenthic community trophic structure. Community trophic data were also used to evaluate whether results from laboratory sediment toxicity tests were effective indicators of site-specific differences in benthic trophic structure. A multiple regression model consisting of five composite factors (principal components) was used to distinguish the effects of sediment contaminants and environmental variables on benthic community trophic structure. This model explained 33.5% of the variation in macrobenthic trophic diversity (p < 0.001), a variable derived from the distribution of taxa among nine original trophic categories. A significant negative relationship was found between principal components reflecting concentrations of sediment contaminants and macrobenthic trophic diversity.
Detritivores including surface deposit-feeders (SDF), subsurface deposit-feeders (SSDF), and filter feeders (FF) ~ere numerically dominant at 201 random sites, each group accounting for 25-30% of total macrobenthic abundance. The relative abundance of SDFs was considerably lower (12.1 ± 2.9% to 17.1 ± 4.4%) at sites where sediment contaminant concentrations exceeded minimum biological effects tbresholds (ER-L values from Long and Morgan 1990) tban at sites sampled at random (29.3 ± 5.7%). SSDFs were proportionally more abundant at contaminated sites (42.0 ± 7.7% to 63.6 ± 10.3%) versus random sites (27.5 ± 5.7%), and the relative abundance of SSDFs was positively correlated with concentrations of particular contaminants. Benthic trophic structure was also found to be a function of salinity, where the proportion of SSDFs was negatively correlated with salinity (p = 0.035, r = -0.223, n = 326). Silt-day content loaded fairly strongly on the first principal component, but trophic structure parameters were not significantly correlated with sediment grain size or dissolved oxygen (perhaps due, in part, to covariation). Results from laboratory sediment toxicity tests with mysids were predictive of differences in macrobenthic trophic structure in situ (i.e., mysid survival was negatively correlated with %SSDF; p < 0.001, r = -0.292, n = 326). Results from laboratory sediment toxicity tests with ampeliscid amphipods were not indicative of site-specific differences in benthic trophic structure.
Results from tbis study demonstrated that sediment contaminants cali be quite important in structuring macrobenthic communities in soft-bottom estuarine habitats. The fact that macro benthic trophic diversity decreased significantly with increasing sediment contamination indicates that important general differences in benthic community function may exist between contaminated and random sites. These data suggest that benthic trophic structure analysis may be an effective tool for assessing integrated community responses to chronic subletbal exposure and may be useful for assessing toxicological responses at ecologically relevant levels of organization.
Introduction Macrobenthic invertebrates play an important
functional role in estuaries and other aquatic eco-
I Corresponding author: present address: Rolun and Haas Company, Toxicology Department, 727 Norristown Road, P. O. Box 904, Spring House, Pennsylvania 19477-0904; tele: 215/619-5323.
© 2000 Estuarine Research Federation 411
systems. They alter geochemical conditions at the sediment-water interface, promote decomposition and nutrient cycling, and transfer energy to other food web components (Rhoads 1974; Boesch et al. 1976; Aller 1982; Tenore et al. 1984; Schaffner et al. 1987). Dominance by different macrobenthic trophic groups suggests differences in food-re-

412 s. S. Brown et al.
source availability and food web interactions, and may be indicative of differences in ecosystem-level processes (Commito and Ambrose 1985; Cohen and Briand 1990; Diaz and Schaffner 1990). Benthic community trophic structure is a potentially valuable criterion for integrating and assessing ecological responses along estuarine gradients.
One closely-held tenet of estuarine ecology is that macrobenthic community trophic structure varies along selected environmental gradients, most notably sediment type, in which greater proportions of deposit feeders and fewer suspension feeders are anticipated with increasing silt-clay content (Rhoads and Young 1970; Levinton 1972; Day et al. 1989; Peterson 1991). This relationship appears to be strongest for high-mesohaline (12-18%0) to polyhaline (18-25%0) environments. Community composition is influenced by an array of anthropogenic and non-anthropogenic factors, and direct causal relationships are typically not evident (Long and Chapman 1985; Diaz and Schaffner 1990; Gaston and Edds 1994). Evaluating the relative influence and interaction of different factors, including the effects of sediment-associated contaminants, poses a major challenge (Boesch and Rosenberg 1981; Ferraro et al. 1991; Diaz 1992).
Many contaminants partition to sediments, creating a major sink and potential source for organism exposure (Baker et al. 1980; Dickson et al. 1984; Jenne and Zachara 1984; Adams et al. 1985; Karickhoff and Morris 1985; Medine and McCutcheon 1989; Baudo et al. 1990; Burton 1992). Macrobenthos may be exposed to sediment-associated contaminants via several routes including dermal contact (sorption through interstitial and overlying water), sediment ingestion, and trophic transfer (Anderson et al. 1984; Rodgers et al. 1984; Landrum and Robbins 1990; Chapman et al. 1991; Di Toro et al. 1991). Feeding strategy may, therefore, be an important determinant of contaminant exposure (Matisoff et al. 1985; Landrum and Robbins 1990), and trophic structure may be an appropriate and sensitive measure of community response.
The purpose of this study was to evaluate the influence of sediment-associated contaminants and selected natural environmental gradients on macrobenthic community trophic structure. We were interested in determining whether macrobenthic trophic structure varied predictably in response to differences in sediment contaminant concentrations, salinity, sediment type, and dissolved oxygen. A second objective was to assess whether differences in macrobenthic community trophic structure were indicated or predicted by results from laboratory-based sediment toxicity tests.
Materials and Methods
SAMPUNG AND ANALYSIS
Physical, chemical, and biological data were collected during the summers of 1991 and 1992 from 333 stations in estuaries throughout the northern Gulf of Mexico (Anclote Anchorage, Florida to the Rio Grande River, Texas). A surface area-based, probabilistic sampling design was used to ensure that different estuarine environments were equitably represented, including large systems (>260 km2), small systems (>2.6 km2 to <260 km2), and large tidal rivers (>260 km2 with aspect ratio >20) (Summers et al. 1991, 1993; Summers and Engle 1993). Data were collected from 201 random base (i.e., reference) stations with sediments which did not contain contaminant concentrations exceeding published biological-effects thresholds (ER-Ls from Long and Morgan 1990). Data from these sites were used to characterize habitat conditions and benthic trophic structure for estuaries throughout the northern Gulf of Mexico. Data were also collected from 132 stations located in areas of known or suspected environmental degradation. Since a large proportion of total estuarine habitat in the northern Gulf of Mexico is distributed along the Louisiana coastline, a majority of sampling stations were located in this area.
Sampling stations were located using Loran-C. Surface water quality variables (e.g., temperature, salinity, dissolved oxygen, pH) were measured at all stations using a Hydrolab system interfaced with an on-board computer. Water quality variables were also measured 1 m above the bottom at 15-min intervals by overnight deployment of a Datasonde 3 (Hydrolab Corp.). Three replicate benthic grab samples were collected from each station with a modified van Veen sampler (413 cm2), samples were washed on a 500-J..Lm screen, and organisms were transferred to labeled jars containing 10% buffered formalin and Rose Bengal. Additional sediments were collected with the grab to provide material for sediment toxicity tests, while other samples were collected and the top 3 cm was subsampled for the analysis of contaminants, sediment grain-size distribution, total organic carbon, and acid-volatile sulfides (Heitmuller and Valente 1991; Summers et al. 1991).
Sediment grain-size distribution was analyzed using methods of Plumb (1981), including the determination of percent silt-clay, percent sand, and proportions of various sand and silt-clay fractions. Percent total organic carbon content was determined by combustion at 2,500°C of an acidified, dried, and ground subsample using a Leco CR-12 carbon analyzer with an infrared detector. Acid-volatile sulfide content of sediments was determined

using a modification of Boothman et al. (1990) with detection by ion-specific electrode. Surface sediments were analyzed for organic and inorganic compounds including 27 alkanes, 43 polynuclear aromatic hydrocarbons, 20 polychlorinated biphenyls, 24 pesticides, and 15 metals using methods detailed elsewhere (MacLeod et al. 1985; Krahn et al. 1988; Heitmuller and Valente 1991). Sediment characterizations and chemical analyses were subject to rigorous quality assurance procedures, including the analysis of sample duplicates, laboratory blanks, reagent blanks, and standard reference materials (Taylor 1987; Heitmuller and Valente 1991).
Sediment toxicity tests were conducted in the laboratory under static conditions using the infaunal tube-dwelling amphipod Ampelisca abdita (lO-d survival) and the epibenthic mysid shrimp Mysidopsis bahia (4-d survival). Methods for these assays followed descriptions provided in Swartz et al. (1985) and Graves et al. (1990).
Macrobenthic samples were rewashed in the laboratory on a 500-j.Lm screen, sorted into major taxonomic groups, and placed into labeled vials containing 70% ethanol. Macrobe.nthos were identified to the lowest practical taxon (usually species) and enumerated, and voucher specimens were designated. Ten percent of all sorted samples were resorted, and identifications and counts were verified by senior taxonomists (Heitmuller and Valente 1991; Summers et al. 1991).
Macrobenthic taxa were assigned to trophic groups based on feeding morphology, feeding behavior, and food preferences documented in the peer-reviewed literature (e.g., Fauchald andjumars 1979; Diaz 1980; Dauer et al. 1981; Levin 1981; Maurer and Leathem 1981; Dauer 1984; Weinberg 1984; Commito and Ambrose 1985; Gaston 1987; Schaffner 1990; Rakocinski et al. 1997). Published data were unavailable for selected Crustacea and a small number « 10%) of other taxa, and trophic classifications for these organisms were based upon unpublished observations of the authors. Trophic guild selection followed Gaston and Nasci (1988), and included surface-deposit feeders (SDF), subsurface-deposit feeders (SSDF), suspension or filter feeders (FF), carnivores (CARN), omnivores (OMNI), and others (XXX). Approximately 10% of the taxa exhibited facultative (split) feeding behaviors and were assigned trophic designations indicating two feeding strategies (e.g., Strebwspio benedicti = SDF IFF). Abundances for these taxa were divided equally among the applicable guilds.
DATA ANALYSIS
Variation in macrobenthic trophic structure was examined across a range of sediment contaminant
Sediment Contaminants and Trophic Structure 413
concentrations and natural environmental gradients. Initial efforts involved simple qualitative comparisons of trophic structure between stations characterized by differences in contaminant levels and natural environmental variables. Macrobenthic trophic structure was compared between stations widely separated by differences in salinity (high> 25%0, low < 10%0), sediment type (sandy < 20% silt-clay, muddy> 80% silt-clay) and near-bottom dissolved oxygen (high > 5 ppm, low < 2 ppm). Similar comparisons of trophic structure were made between the 201 random sites and groups of stations characterized by concentrations of sediment contaminants (e.g., metals, polynuclear aromatic hydrocarbons (PAHs), PCBs, and DDTs) exceeding effects-range low (ER-L) thresholds posed by Long and Morgan (1990) and Long et al. (1995). These ER-L values represent concentrations below which adverse biological effects were observed in 10% of the cases examined by the authors. This qualitative approach was used for screening potential relationships between species distributions, trophic structure, and habitat variables which could be explored by multivariate analyses. Contingency tables (G-tests) were used to compare trophic distributions between sampling years and among different regions within the Louisianan Province.
Numerical abundance was used to calculate proportional (percent) representation by different macrobenthic trophic groups, and percentage data were arcsine transformed (Zar 1984). Relationships between trophic group percentages and different environmental and contaminant variables were evaluated using Pearson's pair-wise correlation analysis, and significant bivariate relationships were determined using Bonferroni-adjusted probabilities (0: = 0.05), which allowed for multiple comparisons. Correlation was followed by principal components analysis (PCA) in order to identifY groups of highly interrelated environmental variables. A Varimax rotated PCA was performed in order to reduce 21 natural environmental and contaminant variables to a subset of non-correlated composite variables (Rakocinski et al. 1997).
Two benthic trophic response variables, arcsinesquare root SSDF and trophic diversity, were subsequently utilized as dependent variables in separate multiple regression analyses with the composite PCA factors as independent variables. The Trophic Diversity Index was calculated analogously to the Shannon-Wiener Index (Pie lou 1975), i.e., H' = -I log (Pi) A, where P = proportion and i = trophic category (natural log scale; range = 0-1.9; maximum possible = 2.2). Calculations of trophic diversity were based on the distribution of taxa among the nine original trophic categories (Le.,

414 s. S. Brown et at.
TABLE 1. Macrobenthic community trophic structure by year for estuaries in the northern Gulf of Mexico. Data collected at 201 random stations (603 samples).
Mean Abundance m- 2 and Trophic Group Proportions (%) Trophic Group 1991 and 1992 1991 1992
SDF 833.2 (29.3) 724.8 (29.7) 943.3 (28.9) SSDF 782.5 (27.5) 532.5 (21.9) 1,035.0 (31.7) FF 712.7 (25.0) 737.3 (30.3) 687.9 (21.1) CARN 349.9 (12.3) 278.3 (11.4) 422.8 (13.0) OMNI 63.2 (2.2) 77.4 (3.2) 48.8 (1.5) XXX 104.9 (3.7) 86.2 (3.5) 123.8 (3.8) Total 2,846.4 (100) 2,436.5 (100) 3,261.6 (100)
CARN, FF, OMNI, SDF, SDF/CARN, SDF/FF, SDF /SSDF, SSDF, XXX). The XXX category was included in the calculations so that the sum of proportions for each sample would still equal 1.0. Since there were nine possible categories, the Trophic Diversity Index could range between 0 and 2.197 (i.e., even distribution with Pi = 0.111). Observed trophic diversity ranged between 0.007 and 1.882, and averaged 1.126 ::t 0.053 (mean ::t 2 SE; n = 319). Arcsine-square root SSDF and trophic diversity were regressed against various combinations of PCA factors, including their squared terms to examine possible simple non-linear relationships, to generate multiple regression models best explaining the observed variation in macrobenthic trophic structure. Final regression models were also compared with stepwise selection models to ensure the best fit.
Results
MACROBENTHIC TROPHIC STRUCTURE AND DOMINANT TAXA AT RANDOM SITES
Macrobenthic communities sampled at random sites in northern Gulf of Mexico estuaries were composed of equal proportions (25-30%) of the three detritivore feeding guilds (SDF, SSDF, and FF), which together accounted for 85% of total macrobenthic abundance (Table 1). Of the remainder, CARN accounted for 12% of total macrobenthic abundance, while OMNI and XXX represented <4% each. Trophic distributions were not different between years (G = 4.1; df = 5, P > 0.5), nor among large estuaries from Texas, Louisiana, Mississippi-Alabama, and Florida (G = 9.4; df = 15, P > 0.75). Approximately 100,000 organisms representing 840 macrobenthic taxa were collected during the two-year study. The six most abundant taxa collected from the random (or base) sites represented 35% of mean total macrobenthic density (2,846 m- 2). These taxa included Mediomastus californiensis (SSDF polychaete, 386 m- 2), Corophium cf., lacustre (SDF amphipod, 178 m-2), Mulinia latemlis (FF bivalve, 129 m-2), unidentified tubificids
TABLE 2. Numerically dominant macrobenthic taxa collected from 201 random stations (603 samples) in estuaries in the northern Gulf of Mexico (1991-1992).
Mean Taxa Trophic Group Abundance m- 2
Mediomastus californiensis SSDF 386 Corophium cf. lacustre SDF 178 Mulinia lateralis FF 129 Unidentified Tubificidae SSDF 110 ProUythinella louisianae SDF 109 Streblospio benedicti SDFjFF 85 Texadina sphinctostoma SDF 79 Paraprionospio pinnata SDFjFF 58 Spiochaetopterus costarum FF 54 Caecum johnsoni OMNI 47 Myriochele oculata SSDF 41 Hobsonia florida SDF 40 Unidentified Nemertea CARN 36 CrassineUa lunulata FF 35 Nemerteasp.B CARN 35 Rangia cuneata FF 35 Nemertea sp. A CARN 34 Tubificoides heterochaetus SSDF 33 Parandalia sp. A CARN 32 Ampelisca abdita FF 27 Notomastus latericeus SSDF 26 Magelona sp. H SDF 26 Acteocina canaliculata CARN 24 Balanus sp. FF 24 Prionospio pygmaea SDFjFF 24 Unidentified Maldanidae SSDF 23 Prionospio perkinsi SDFjFF 23 Phoronis muelleri FF 23 Petricola pholadiformis FF 22 Unidentified Ophiuroidea XXX 20
(SSDF oligochaete, 110 m-2), Probythinella louisianae (SDF gastropod, 109 m- 2), Strebwspio benedicti (SDF /FF polychaete, 85 m-2), and Texadina sphinctostoma (SDF gastropod, 79 m-2) (Table 2).
MACRO BENTHIC TROPHIC STRUCTURE VERSUS NATURAL ENVIRONMENTAL VARIABLES
Among 333 stations sampled, 92 were characterized as high salinity sites (>25%0) and 131 were classified as low salinity stations « 1 0%0). Species richness and mean total density were much higher at high salinity stations (675 total taxa, 7.3 taxa per station, 3,248 m- 2) than at low salinity sites (260 total taxa, 2.0 taxa per station, 2,260 m-2). Benthic communities at high salinity sites were composed of roughly equal proportions (20-30%) of SDF, SSDF, and FF, while low salinity stations supported higher proportions of SSDF (39.7 ::t 4.5% versus 29.2 ::t 5.2%) and SDF (35.8 ::t 4.9% versus 22.8 ::t 3.2%), and lower proportions of FF (16.9 ± 5.3% versus 21.2 ::t 3.5%) (Tables 3 and 4). The proportion of SSDF was negatively correlated with salinity (p = 0.035 (Bonferroni adjusted), r = -0.223, n = 326), while CARN, especially nemer-

TABLE 3. Macrobenthic community trophic structure at sta-tions in northern Gulf of Mexico estuaries with high (>25%0) versus low «10%0) salinity, sandy «20% silt-clay) versus mud-dy (>80% silt-clay) sediments, and high (>5.0 ppm) versus low «2.0 ppm) dissolved oxygen.
Percent of Total Macrobenthic Abundance (Mean ± (SD»
Salinity Sediment Dissolved Oxygen Trophic Group High Low Sandy Muddy High Low
(n) (92) (131) (34) (158) (238) (20) SDF 22.8 35.8 20.0 30.1 29.3 17.1
(3.2) (4.9) (5.2) (6.8) (6.3) (3.5) SSDF 29.2 39.7 29.3 32.8 31.3 36.7
(5.2) (4.5) (3.9) (4.2) (6.3) (6.8) FF 21.2 16.9 26.7 25.7 23.0 23.3
(3.5) (5.3) (6.2) (4.7) (4.2) (6.1) CARN 16.3 6.0 11.2 9.2 10.5 18.2
(2.1) (1.3) (3.6) (2.2) (3.1) (4.2) OMNI 3.8 0.3 5.6 0.5 2.1 0.1
(0.5) (0.1) (2.7) (0.3) (1.0) (0.1) xxx 6.7 1.3 7.2 1.7 3.8 4.6
(1.3) (0.3) (2.5) (1.3) (1.4) (2.5)
Sediment Contaminants and Trophic Structure 415
teans, were relatively more abundant at high salinity sites (16.3 ± 2.1 % versus 6.0 ± 1.3%).
Thirty-four stations were characterized as sandy sites «20% silt-clay) and 158 stations were classified as muddy sites (>80% silt<lay). Higher species richness and mean total density were observed at sandy versus muddy sites (541 total taxa, 15.9 taxa per station, and 4,747 m-2 versus 411 total taxa, 2.6 taxa per station, and 2,219 m-2). Muddy stations supported slightly higher proportions of SDF (30.1 ± 6.8%) than did sandy sites (20.0 ± 5.2%) (Tables 3 and 4), and the SDF amphipod Corophium cf., lacustre was particularly abundant at fine-sediment stations (212 m-2 versus 15 m-2). Statistically significant relationships between sediment grain size and proportions of SDF and SSDF were not evident (e.g., r = 0.064 for SSDF, r = 0.008 for SDF; n = 326), perhaps due to covariation, despite the fact that subsequent regression analyses showed that silHlay content loaded fairly strongly on the first principal component (see below).
Two-hundred thirty-eight stations exhibited
TABLE 4. Mean density of the ten most abundant macrobenthic taxa at stations in estuaries of the northern Gulf of Mexico (1991-1992) characterized by high (>25%0) versus low «10%0) salinity, sandy «20% silt-clay) versus muddy (>80% silt-clay) sediments, and high (>5.0 ppm) versus low «2.0 ppm) near-bottom dissolved oxygen concentrations. - indicates taxa not among the most abundant for this habitat condition.
Salinity
Taxa (Trophic Group) High Low
(n) (92) (131) Mediomastus califumiensis (SSDF) 331 322 Acanthohaustorius sp. A (SSDF) 135 Notomastus latericeus (SSDF) 131 Paraprionospio pinnata (SDF IFF) 104 Caecumjohnsoni (OMNI) 99 Myriochele oculata (SSDF) 76 Crassinella lunulata (FF) 75 Unidentified Nemertea (CARN) 66 Nemertea sp. B (CARN) 63 Prionospio pygmo,ea (SDF IFF) 56 Unidentified Tubificidae (SSDF) 322 Corophium cf. lacustre (SDF) 271 Probythinella louisianae (SDF) 181 Texadina sphinctostoma (SDF) 136 Rangia cuneata (FF) 112 Streblospio benedicti (SDF IFF) 100 Tumficoides heterochaetus (SSDF) 79 Hobsonia florida (SDF) 66 Halmyrapseudes bahamensis (SSDF) 52 Cerapus benthophilus (SDF IFF) Ampelisca abdita (FF) Donax sp. (FF) Ampelisca holmesi (FF) Polygordius sp. (CARN) Mulinia lateralis (FF) Ischadium recurvum (FF) Unidentified Chaoboridae (CARN) Magelona sp. H (SDF) Unidentified Podocopida (XXX) Asychis elongatus (SSDF)
Mean Density (Abundance m-')
Sediment
Sandy Muddy
(34) (158) 343 320 367
96 232
203
180 212 121
71 59 75 61
199 211 173 107 105 92
153
Dissolved Oxygen
High
(238) 317
53
49
149 149 99 75 56 75
110
Low
(20) 413
225 269
66
182
242 154 121 58 54
416 s. S. Brown et al.
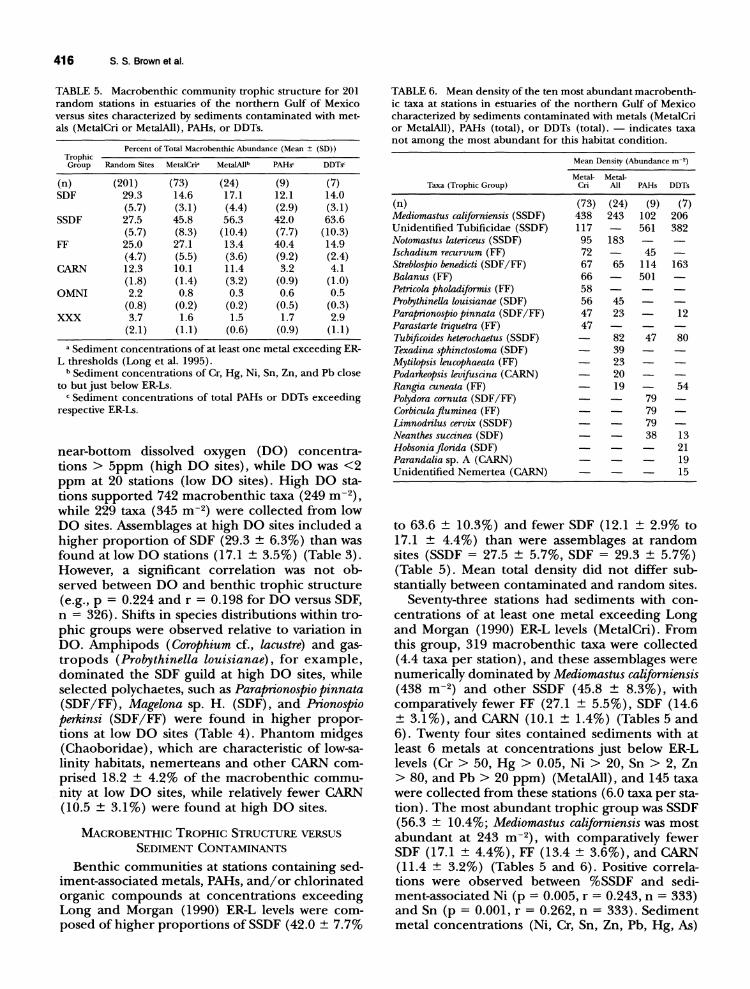
TABLE 5. Macrobenthic community trophic structure for 201 random stations in estuaries of the northern Gulf of Mexico versus sites characterized by sediments contaminated with met-als (MetalCri or MetaWl), PARs, or DDTs.
Percent of Total Macrobenthic Abundance (Mean ± (SO» Trophic Group Random Sites MetalCri" MelaWlb PAH," DDT,"
(n) (201) (73) (24) (9) (7) SDF 29.3 14.6 17.1 12.1 14.0
(5.7) (3.1) (4.4) (2.9) (3.1) SSDF 27.5 45.8 56.3 42.0 63.6
(5.7) (8.3) (10.4) (7.7) (10.3) FF 25.0 27.1 13.4 40.4 14.9
(4.7) (5.5) (3.6) (9.2) (2.4) CARN 12.3 10.1 11.4 3.2 4.1
(1.8) (1.4) (3.2) (0.9) (1.0) OMNI 2.2 0.8 0.3 0.6 0.5
(0.8) (0.2) (0.2) (0.5) (0.3) XXX 3.7 1.6 1.5 1.7 2.9
(2.1) (1.1) (0.6) (0.9) (1.1)
a Sediment concentrations of at least one metal exceeding ERL thresholds (Long et al. 1995).
b Sediment concentrations of Cr, Hg, Ni, Sn, Zn, and Pb close to but just below ER-Ls.
C Sediment concentrations of total PARs or DDTs exceeding respective ER-Ls.
near-bottom dissolved oxygen (DO) concentrations > 5ppm (high DO sites), while DO was <2 ppm at 20 stations (low DO sites). High DO stations supported 742 macrobenthic taxa (249 m- 2),
while 229 taxa (345 m- 2) were collected from low DO sites. Assemblages at high DO sites included a higher proportion of SDF (29.3 ± 6.3%) than was found at low DO stations (17.1 ± 3.5%) (Table 3). However, a significant correlation was not observed between DO and benthic trophic structure (e.g., p = 0.224 and r = 0.198 for DO versus SDF, n = 326). Shifts in species distributions within trophic groups were observed relative to variation in DO. Amphipods (Corophium cf., lacustre) and gastropods (Probythinella louisianae), for example, dominated the SDF guild at high DO sites, while selected polychaetes, such as Parapnonospio pinnata (SDFIFF) , Magelona sp. H. (SDF), and Prionospio perkinsi (SDF IFF) were found in higher proportions at low DO sites (Table 4). Phantom midges (Chaoboridae), which are characteristic of low-salinity habitats, nemerteans and other CARN comprised 18.2 ± 4.2% of the macrobenthic community at low DO sites, while relatively fewer CARN (10.5 ± 3.1 %) were found at high DO sites.
MACRO BENTHIC TROPHIC STRUCTURE VERSUS SEDIMENT CONTAMINANTS
Benthic communities at stations containing sediment-associated metals, PAHs, andlor chlorinated organic compounds at concentrations exceeding Long and Morgan (1990) ER-L levels were composed of higher proportions ofSSDF (42.0 ± 7.7%
TABLE 6. Mean density of the ten most abundant macrobenthic taxa at stations in estuaries of the northern Gulf of Mexico characterized by sediments contaminated with metals (MetalCri or MetaWl) , PARs (total), or DDTs (total). - indicates taxa not among the most abundant for this habitat condition.
Mean Density (Abundance m-2 )
Metal- Metal-Taxa (Trophic Group) eri All PAH, DDT,
(n) (73) (24) (9) (7) Mediomastus californiensis (SSDF) 438 243 102 206 Unidentified Tubificidae (SSDF) 117 561 382 Notomastus latericeus (SSDF) 95 183 Ischadium recurvum (FF) 72 45 Streblospio benedicti (SDF IFF) 67 65 114 163 Balanus (FF) 66 501 Petri cola Pholadiformis (FF) 58 Probythinella louisianae (SDF) 56 45 Parapnonospio pinnata (SDF IFF) 47 23 12 Parastarte triquetra (FF) 47 Tubificoides heterochaetus (SSDF) 82 47 80 Texadina sphinctostoma (SDF) 39 Mytilopsis leucophaeata (FF) 23 Podarkeopsis levifuscina (CARN) 20 Rangia cuneata (FF) 19 54 Polydora cornuta (SDF IFF) 79 Corbicula fluminea (FF) 79 Limnodrilus cervix (SSDF) 79 Neanthes succinea (SDF) 38 13 Hobsonia florida (SDF) 21 Parandalia sp. A (CARN) 19 Unidentified Nemertea (CARN) 15
to 63.6 ± 10.3%) and fewer SDF (12.1 ± 2.9% to 17.1 ± 4.4%) than were assemblages at random sites (SSDF = 27.5 ± 5.7%, SDF = 29.3 ± 5.7%) (Table 5). Mean total density did not differ substantially between contaminated and random sites.
Seventy-three stations had sediments with concentrations of at least one metal exceeding Long and Morgan (1990) ER-L levels (MetaICri). From this group, 319 macrobenthic taxa were collected (4.4 taxa per station), and these assemblages were numerically dominated by Mediomastus californiensis (438 m-2) and other SSDF (45.8 ± 8.3%), with comparatively fewer FF (27.1 ± 5.5%), SDF (14.6 ± 3.1 %), and CARN (10.1 ± 1.4%) (Tables 5 and 6). Twenty four sites contained sediments with at least 6 metals at concentrations just below ER-L levels (Cr > 50, Hg > 0.05, Ni > 20, Sn > 2, Zn > 80, and Pb > 20 ppm) (MetalAlI), and 145 taxa were collected from these stations (6.0 taxa per station). The most abundant trophic group was SSDF (56.3 ± 10.4%; Mediomastus californiensis was most abundant at 243 m- 2), with comparatively fewer SDF (17.1 ± 4.4%), FF (13.4 ± 3.6%), and CARN (11.4 ± 3.2%) (Tables 5 and 6). Positive correlations were observed between %SSDF and sediment-associated Ni (p = 0.005, r = 0.243, n = 333) and Sn (p = 0.001, r = 0.262, n = 333). Sediment metal concentrations (Ni, Cr, Sn, Zn, Pb, Hg, As)
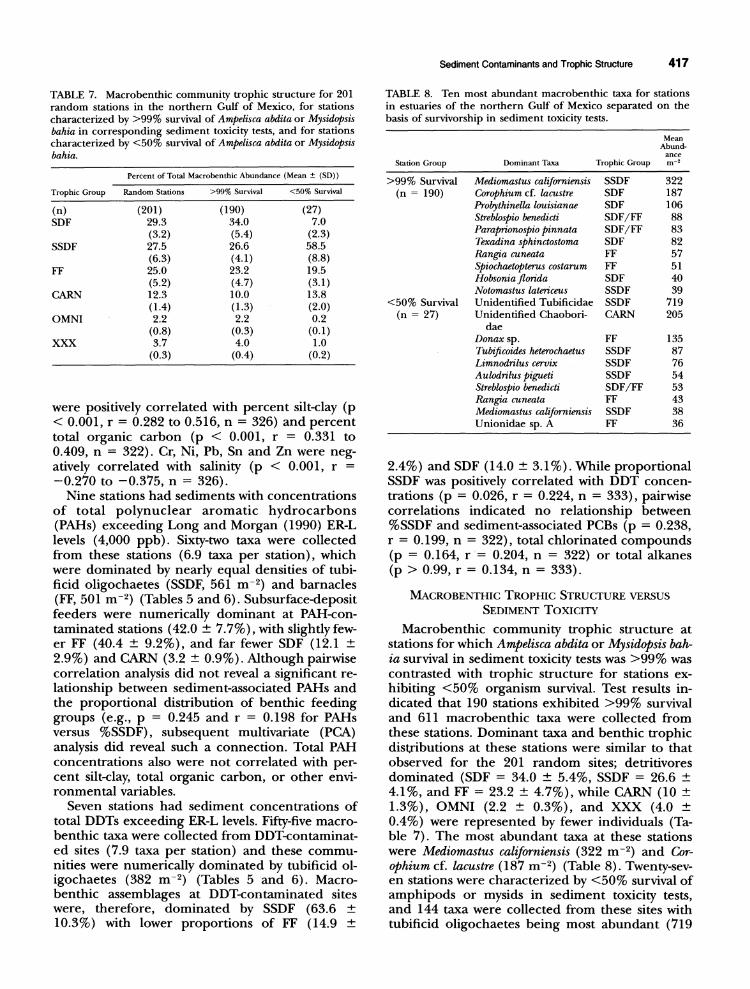
TABLE 7. Macrobenthic community trophic structure for 201 random stations in the northern Gulf of Mexico, for stations characterized by >99% survival of Ampelisca abdita or Mysidopsis bahia in corresponding sediment toxicity tests, and for stations characterized by <50% survival of Ampelisca abdita or Mysidopsis bahia.
Percent of Total Macrobenthic Abundance (Mean :!: (SD»
Trophic Group Random Stations >99% Survival <50% Survival
(n) (201) (190) (27) SDF 29.3 34.0 7.0
(3.2) (5.4) (2.3) SSDF 27.5 26.6 58.5
(6.3) (4.1) (8.8) FF 25.0 23.2 19.5
(5.2) (4.7) (3.1) CARN 12.3 10.0 13.8
(1.4) (1.3) (2.0) OMNI 2.2 2.2 0.2
(0.8) (0.3) (0.1) XXX 3.7 4.0 1.0
(0.3) (0.4) (0.2)
were positively correlated with percent silt-clay (p < 0.001, r = 0.282 to 0.516, n = 326) and percent total organic carbon (p < 0.001, r = 0.331 to 0.409, n = 322). Cr, Ni, Pb, Sn and Zn were negatively correlated with salinity (p < 0.001, r = -0.270 to -0.375, n = 326).
Nine stations had sediments with concentrations of total polynuclear aromatic hydrocarbons (PAHs) exceeding Long and Morgan (1990) ER-L levels (4,000 ppb). Sixty-two taxa were collected from these stations (6.9 taxa per station), which were dominated by nearly equal densities of tubificid oligochaetes (SSDF, 561 m-2) and barnacles (FF, 501 m-2) (Tables 5 and 6). Subsurface-deposit feeders were numerically dominant at PAH-contaminated stations (42.0 ± 7.7%), with slightly fewer FF (40.4 ± 9.2%), and far fewer SDF (12.1 ± 2.9%) and CARN (3.2 ± 0.9%). Although pairwise correlation analysis did not reveal a significant relationship between sediment-associated PAHs and the proportional distribution of benthic feeding groups (e.g., p = 0.245 and r = 0.198 for PAHs versus %SSDF), subsequent multivariate (PCA) analysis did reveal such a connection. Total PAH concentrations also were not correlated with percent silt-clay, total organic carbon, or other environmental variables.
Seven stations had sediment concentrations of total DDTs exceeding ER-L levels. Fifty-five macrobenthic taxa were collected from DDT-contaminated sites (7.9 taxa per station) and these communities were numerically dominated by tubificid oligochaetes (382 m-2) (Tables 5 and 6). Macrobenthic assemblages at DDT-contaminated sites were, therefore, dominated by SSDF (63.6 ± 10.3%) with lower proportions of FF (14.9 ±
Sediment Contaminants and Trophic Structure 417
TABLE 8. Ten most abundant macrobenthic taxa for stations in estuaries of the northern Gulf of Mexico separated on the basis of survivorship in sediment toxicity tests.
Station Group
>99% Survival (n = 190)
<50% Survival (n = 27)
Dominant Taxa
Mediomastus californiensis Corophium cf. lacustre Probythinella louisianae Streblospio benedicti Paraprionospio pinnata Texadina sphinctostoma Ro.ngia cuneata Spiochaetopterus costarum Hobsonia florida Notomastus latericeus Unidentified Tubificidae Unidentified Chaobori-
dae Donaxsp. Thbificoides heterochaetus Limnodrilus cervix A ulodrilus pigueti Streblospio benedicti Ro.ngia cuneata Mediomastus californiensis Unionidae sp. A
Mean Abund-
ance Trophic Group m-'
SSDF 322 SDF 187 SDF 106 SDF/FF 88 SDF/FF 83 SDF 82 FF 57 FF 51 SDF 40 SSDF 39 SSDF 719 CARN 205
FF 135 SSDF 87 SSDF 76 SSDF 54 SDF/FF 53 FF 43 SSDF 38 FF 36
2.4%) and SDF (14.0 ± 3.1 %). While proportional SSDF was positively correlated with DDT concentrations (p = 0.026, r = 0.224, n = 333), pairwise correlations indicated no relationship between %SSDF and sediment-associated PCBs (p = 0.238, r = 0.199, n = 322), total chlorinated compounds (p = 0.164, r= 0.204, n = 322) or total alkanes (p> 0.99, r = 0.134, n = 333).
MACROBENTHIC TROPHIC STRUcrURE VERSUS SEDIMENT TOXICIlY
Macrobenthic community trophic structure at stations for which Ampelisca abdita or Mysidopsis bahia survival in sediment toxicity tests was >99% was contrasted with trophic structure for stations exhibiting <50% organism survival. Test results indicated that 190 stations exhibited >99% survival and 611 macrobenthic taxa were collected from these stations. Dominant taxa and benthic trophic distributions at these stations were similar to that observed for the 201 random sites; detritivores dominated (SDF = 34.0 ± 5.4%, SSDF = 26.6 ± 4.1%, and FF = 23.2 ± 4.7%), while CARN (10 ± 1.3%), OMNI (2.2 ± 0.3%), and XXX (4.0 ± 0.4%) were represented by fewer individuals (Table 7). The most abundant taxa at these stations were Mediomastus californiensis (322 m-2) and Corophium cf. lacustre (187 m-2) (Table 8). Twenty-seven stations were characterized by <50% survival of amphipods or mysids in sediment toxicity tests, and 144 taxa were collected from these sites with tubificid oligochaetes being most abundant (719
418 S. S. Brown et al.
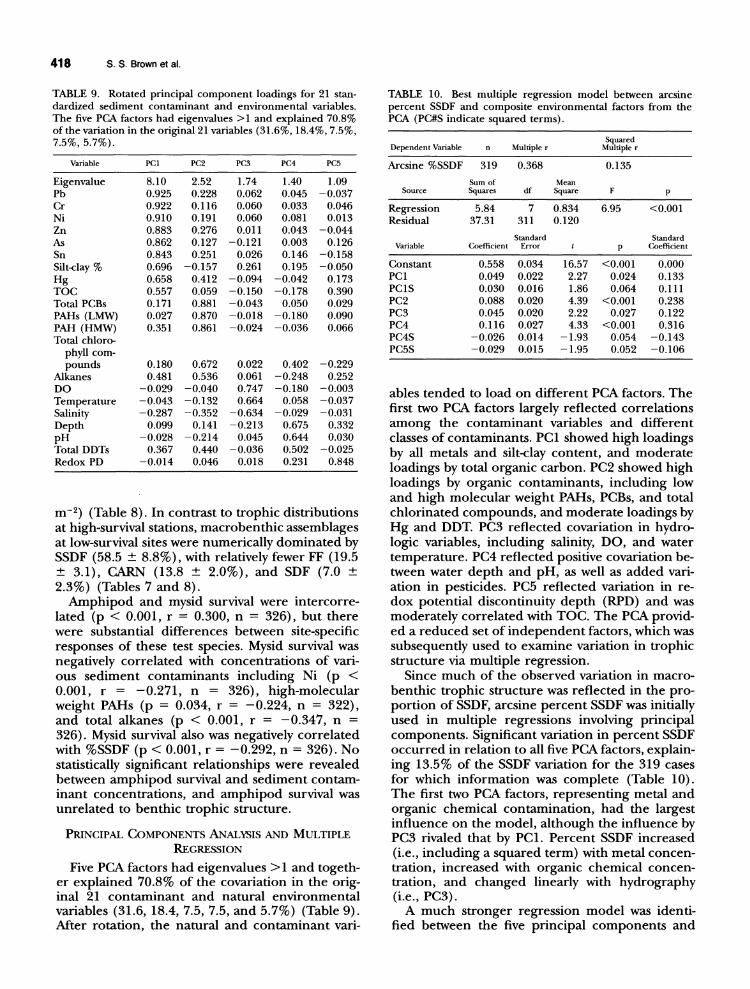
TABLE 9. Rotated principal component loadings for 21 stan-dardized sediment contaminant and environmental variables. The five PCA factors had eigenvalues> I and explained 70.8% of the variation in the original 21 variables (31.6%, 18.4%,7.5%, 7.5%, 5.7%).
Variable PCl PC2 PC3 PC4 PC5
Eigenvalue 8.10 2.52 1.74 1.40 1.09 Pb 0.925 0.228 0.062 0.045 -0.037 Cr 0.922 0.1l6 0.060 0.033 0.046 Ni 0.910 0.191 0.060 0.081 0.013 Zn 0.883 0.276 0.011 0.043 -0.044 As 0.862 0.127 -0.121 0.003 0.126 Sn 0.843 0.251 0.026 0.146 -0.158 Silt·day % 0.696 -0.157 0.261 0.195 -0.050 Hg 0.658 0.412 -0.094 -0.042 0.173 TOC 0.557 0.059 -0.150 -0.178 0.390 Total PCBs 0.171 0.881 -0.043 0.050 0.029 PAHs (LMW) 0.027 0.870 -0.018 -0.180 0.090 PAH (HMW) 0.351 0.861 -0.024 -0.036 0.066 Total chloro-
phyll com-pounds 0.180 0.672 0.022 0.402 -0.229
Alkanes 0.481 0.536 0.061 -0.248 0.252 DO -0.029 -0.040 0.747 -0.180 -0.003 Temperature -0.043 -0.132 0.664 0.058 -0.037 Salinity -0.287 -0.352 -0.634 -0.029 -0.031 Depth 0.099 0.141 -0.213 0.675 0.332 pH -0.028 -0.214 0.045 0.644 0.030 Total DDTs 0.367 0.440 -0.036 0.502 -0.025 Redox PD -0.014 0.046 0.018 0.231 0.848
m- 2) (Table 8). In contrast to trophic distributions at high-survival stations, macrobenthic assemblages at low-survival sites were numerically dominated by SSDF (58.5 ± 8.8%), with relatively fewer FF (19.5 ± 3.1), CARN (13.8 ± 2.0%), and SDF (7.0 ± 2.3%) (Tables 7 and 8).
Amphipod and mysid survival were intercorrelated (p < 0.001, r = 0.300, n = 326), but there were substantial differences between site-specific responses of these test species. Mysid survival was negatively correlated with concentrations of various sediment contaminants including Ni (p < 0.001, r = -0.271, n = 326), high-molecular weight PAHs (p = 0.034, r = -0.224, n = 322), and total alkanes (p < 0.001, r = -0.347, n = 326). Mysid survival also was negatively correlated with %SSDF (p < 0.001, r = -0.292, n = 326). No statistically significant relationships were revealed between amphipod survival and sediment contaminant concentrations, and amphipod survival was unrelated to benthic trophic structure.
PRINCIPAL COMPONENTS ANALYSIS AND MULTIPLE REGRESSION
Five PCA factors had eigenvalues> 1 and together explained 70.8% of the covariation in the original 21 contaminant and natural environmental variables (31.6, 18.4,7.5,7.5, and 5.7%) (Table 9). Mter rotation, the natural and contaminant vari-
TABLE 10. Best multiple regression model between arcsine percent SSDF and composite environmental factors from the PCA (PC#S indicate squared terms).
Dependent Variable n Multiple r Squared
Multiple r
Arcsine %SSDF 319 0.368 0.135
Sum of Mean Source Squares df Square F p
Regression 5.84 7 0.834 6.95 <0.001 Residual 37.31 311 0.120
Standard Standard Variable Coefficient Error p Coefficient
Constant 0.558 0.034 16.57 <0.001 0.000 PCl 0.049 0.022 2.27 0.024 0.133 PC1S 0.030 0.016 1.86 0.064 0.1l1 PC2 0.088 0.020 4.39 <0.001 0.238 PC3 0.045 0.020 2.22 0.027 0.122 PC4 0.1l6 0.027 4.33 <0.001 0.316 PC4S -0.026 0.014 -1.93 0.054 -0.143 PC5S -0.029 0.015 -1.95 0.052 -0.106
abies tended to load on different PCA factors. The first two PCA factors largely reflected correlations among the contaminant variables and different classes of contaminants. PC1 showed high loadings by all metals and silt-clay content, and moderate loadings by total organic carbon. PC2 showed high loadings by organic contaminants, including low and high molecular weight PAHs, PCBs, and total chlorinated compounds, and moderate loadings by Hg and DDT. PC3 reflected covariation in hydrologic variables, including salinity, DO, and water temperature. PC4 reflected positive covariation between water depth and pH, as well as added variation in pesticides. PC5 reflected variation in redox potential discontinuity depth (RPD) and was moderately correlated with TOC. The PCA provided a reduced set of independent factors, which was subsequently used to examine variation in trophic structure via multiple regression.
Since much of the observed variation in macrobenthic trophic structure was reflected in the proportion of SSDF, arcsine percent SSDF was initially used in multiple regressions involving principal components. Significant variation in percent SSDF occurred in relation to all five PCA factors, explaining 13.5% of the SSDF variation for the 319 cases for which information was complete (Table 10). The first two PCA factors, representing metal and organic chemical contamination, had the largest influence on the model, although the influence by PC3 rivaled that by PCl. Percent SSDF increased (i.e., including a squared term) with metal concentration, increased with organic chemical concentration, and changed linearly with hydrography (i.e., PC3).
A much stronger regression model was identified between the five principal components and

TABLE 11. Best multiple regression model between trophic diversity and composite environmental factors from the PCA (PC#S- indicate squared terms).
n Multiple r
Trophic Diversity 319 0.578
Sum of Mean Source Squares df Square
Regression 23.80 5 4.76 Residual 47.32 313 0.151
Standard Variable Coefficient Error
Constant 1.182 0.025 48.12 PCl -0.144 0.022 -6.57 PC2 -0.144 0.022 -6.52 PC3 -0.141 0.022 -6.38 PC4S -0.039 0.011 -3.50 PC5 -0.055 0.022 -2.51
Squared Multiple r
0.335
F p
31.48 <0.001
Standard p Coefficient
<0.001 0.000 <0.001 -0.303 <0.001 -0.303 <0.001 -0.298
0.001 -0.165 0.013 -0.116
trophic diversity, which was used as an index of benthic trophic structure (Table 11). This model accounted for 33.5% of the variation in trophic diversity. The first three PCA factors had equally strong influences on the model, and trophic diversity declined significandy with all five PCA factors, especially with respect to organic contaminants and metals.
Discussion RELATIONSHIPS BETWEEN SEDIMENT CONTAMINANTS,
ENVIRONMENTAL GRADIENTS, AND MACROBENTHIC
COMMUNI1Y TROPHIC STRUCTURE
Macrobenthic community trophic structure was clearly related to sediment contaminant concentra·tions as well as natural environmental variables in northern Gulf of Mexico estuaries. A multiple regression model consisting of five composite factors (principal components) representing contaminant and environmental variables (metals, PAHs, DO/ temperature/salinity, chlorinated compounds, water depth/redox potential discontinuity) explained 33.5% of the variation in trophic diversity (p < 0.001). A significant negative correlation was found between sediment contaminant concentration and benthic trophic diversity. The relative abundance of SDF was substantially lower at sites where sediment contaminant levels exceeded Long and Morgan (1990) ER-L biological effects thresholds (12.1 ± 2.9% to 17.1 ± 4.4%) than at stations sampled at random (29.3 ± 5.7%). SSDFwere proportionally more abundant at contaminated sites (42.0 ± 7.7% to 63.6 ± 10.3%) versus random sites (27.5 ± 5.7%), and the relative abundance of SSDF was positively correlated with concentrations of selected contaminants. While results from this study indicate that benthic trophic structure was determined by both natural environmental gradi-
Sediment Contaminants and Trophic Structure 419
ents and sediment contamination, diagnostic interpretations about these relationships would benefit from further analyses (i.e., canonical correspondence analysis) (Rakocinski et al. 1997).
The fact that trophic diversity decreased with increasing sediment contamination suggests important differences between contaminated and random sites in terms of benthic community function. Reduced trophic diversity suggests a shift in benthic community function that may have important ramifications for material cycling, decomposition, and other ecosystem processes (Diaz and Schaffner 1990). Reduced taxonomic richness and trophic diversity could also result in the loss of functionally redundant species at contaminated sites, which could in turn have negative effects on ecosystem functions and services (Cairns and Pratt 1986).
Gaston and Young (1992) found that benthic trophic diversity also was lower in contaminated bayous of the Upper Calcasieu Estuary (Louisiana), and these communities were dominated by shallow-burrowing oligochaetes (Tubificidae, SSDF). Observations of increased dominance by SSDF at contaminated sites seem to conflict with data for the lower Chesapeake Bay (Diaz and Schaffner 1990) and Puget Sound (Weston 1990), where macrobenthic assemblages were characterized by reductions in the relative abundance of deep-burrowing species. Benthic communities of northern Gulf of Mexico estuaries are characterized by burrowers (SSDF) that live near the sediment-water interface and few deep-burrowing species (Gaston et al. 1998). For instance, most Mediomastus californiensis and small tubificids (SSDF) inhabit the top 2 cm of the sediment. Such differences may be explained by differences among species within the subsurface-deposit feeding guild, leading to broad-scale dissimilarities be"tween benthic communities in the northern Gulf of Mexico and lower Chesapeake Bay /Puget Sound. Benthic assemblages in lower Chesapeake Bay include several large-bodied, deep-burrowing species which promote bioturbation (Schaffner et al. 1987; Diaz and Schaffner 1990; Schaffner et al. 1987). Benthic communities in estuaries of the northern Gulf of Mexico, and northern Chesapeake Bay, are characterized by a limited subsurface component typically consisting of a few relatively small species (e.g., M. californiensis, tubificids). These characteristic differences in community structure may be translated into differences in community and ecosystem-level functional parameters, including benthic-pelagic coupling and solute/chemical dynamics (exchange) at the sediment-water interface.
Although species assemblages present at contaminated and random sites have been described

420 s. S. Brown at al.
herein, the purpose of the present study was not to identify macrobenthic species which might serve as indicators of contaminant-induced stress. Rakocinski et al. (1997) utilized canonical correspondence analysis to examine this data set with respect to this question, and, indeed, contaminant-sensitive and tolerant taxa were identified.
The depth distribution of macrobenthos in the northern Gulf of Mexico, as in other estuaries, is a reflection of physical and chemical variables and stresses including hypoxia. In the present study, macrobenthic species richness and density were considerably higher at well-oxygenated stations than at sites influenced by low DO, and SDF were relatively more abundant as well. A significant correlation between macrobenthic trophic structure (i.e., %SSDF) and DO was not revealed in the present study, perhaps due to low statistical power (as indicated in the Bonferroni adjusted levels of rejection). Gaston and Edds (1994) similarly found lower species diversity and density in response to oxygen stress, while reporting no effect of DO on trophic structure. As in the present study, Gaston and Edds (1994) found that deposit-feeding polychaetes, such as Magelona cr., phyllisae and Paraprionospio pinnata, dominated assemblages at sites with low DO. Rabalais and Harper (1992) also described lower macrobenthic species richness and density as a result of seasonal hypoxia near the Mississippi River Delta, although effects on trophic structure were not explored.
Many investigators have described shifts in macrobenthic trophic structure as a function of sediment grain-size distribution (sensu Zobell and Feltham 1938; Sanders 1956,1958; Rhoads and Young 1970; Day et al. 1989; Peterson 1991). In this study, trophic structure was shown to be more closely related to sediment contaminant concentrations than sediment grain size, as indicated in the correlation and multiple regression analyses. Deposit feeders dominated both sandy (approximately 59%) and muddy (approximately 63%) stations in northern Gulf of Mexico estuaries, indicating the importance of sedimentary particulate detritus in these systems. The number of different depositfeeding species suggested a wide variety of food resources (sensu Whitlatch 1981). Evidence from the present study combined with gut-contents analysis indicated that benthic species in Gulf of Mexico estuaries focused on near-bottom seston and detritus (Gaston et al. 1997).
Salinity had a significant influence on benthic trophic structure. Trophic diversity increased with increasing salinity, and %SSDF was negatively correlated with salinity. Deposit feeders (SDF and SSDF) were numerically dominant at low-salinity stations, while FF, CARN, and other groups were
poorly represented. While there is a temptation to attribute differences in benthic trophic structure solely to salinity or other environmental variables, the multiple regression analyses showed that contamination factors were at least as strongly related to trophic structure as natural environmental factors including salinity.
In oligohaline-mesohaline portions of Chesapeake Bay, suspension-feeders (FF) and deposit feeders were dominant and FF bivalves dominated faunal biomass (Diaz and Schaffner 1990). Deposit and suspension-feeding species also were dominant off the Louisiana coast, and trophic distributions did not appear to be affected by discharged brine (Gaston et al. 1985; Gaston and Edds 1994). Results from the present study indicating dominance by deposit feeders at low-salinity stations further indicates the importance of sediment-associated detritus as a source of energy for macrobenthos in these estuaries.
ARE RESULTS FROM SEDIMENT TOXICfIY TESTS
PREDICTIVE OF DIFFERENCES IN MACROBENTHIC
COMMUNI1Y TROPHIC STRUCTURE?
Macrobenthic trophic structure was strongly correlated with results from laboratory sediment toxicity tests conducted with Mysidopsis (now Americamysis) bahia. A significant negative relationship was found between mysid survival and %SSDF, and mysid survival was negatively correlated with sediment-associated Ni, PARs, and total alkanes. Survival of amphipods (Ampelisca abdita) , on the other hand, was not correlated with contaminant variables or trophic structure indicators. Stations exhibiting <50% survival of either mysids or amphipods were characterized by macrobenthic communities dominated by SSDF. Stations exhibiting >99% survival of either mysids or amphipods were characterized by communities which were evenly distributed among the major functional feedinggroups (SDF, SSDF, FF), similar to that observed for random stations.
Spies (1989) suggested that results from sediment bioassays may be misleading because toxicity covaries with natural sediment characteristics (e.g., sediment grain size, organic carbon) and other factors. Others have argued that toxicity test organisms are sensitive to sediment contaminants and that such tests are robust in the face of variation in grain size and other natural environmental variables (e.g., Swartz et al. 1985; Long and Buchman 1989; Chapman et al. 1991). In this study, results from sediment toxicity tests with mysids, but not amphipods, were good indicators of the potential adverse effects of contaminants on structural and functional characteristics of macrobenthic communities. Differences in the toxicological respons-

es of these test species may have been due to differences in life history characteristics which influence exposure conditions, or physiological attributes which affect relative sensitivity.
Spies (1989) also expressed reservations about the correlative use of sediment chemistry data, toxicity test results, and benthic community responses embodied in the sediment quality triad approach, suggesting that this approach overlooks biotic interactions which may confound interpretations. While there is certainly a need to better understand the role of biotic interactions in studies of community and ecosystem-level effects of contaminants, the triad approach does not overlook these considerations. The benthic community component of the triad integrates such interactions into a weight-of-evidence assessment.
WHY INCREASED DOMINANCE BY SUBSURFACE DEPOSIT-FEEDERS AT CONTAMINATED SITES?
Our finding that the proportion of subsurface deposit-feeders increased with increasing levels of sediment contamination is at odds with conventional thinking in benthic ecology regarding the effects of organic enrichment on community structure (Pearson and Rosenberg 1978; Rhoads et al. 1978; Diaz and Schaffner 1990; Weston 1990). This can probably be attributed to the fact that smallbodied, shallow-burrowing subsurface depositfeeding organisms (e.g., Mediomastus californiensis) are very abundant in the Gulf of Mexico. Dominance by small SSDF organisms may be indicative of frequent disturbance in shallow Gulf of Mexico estuaries (Gaston et al. 1998). While the abundance of M. californiensis did not increase significandy at stations characterized by sediments contaminated with PAHs and DDTs, the numbers of very small, shallow-burrowing, subsurface depositfeeding oligochaetes did (Tables 2 and 6). As the abundance of SSDF increased, the proportion of SDF decreased. In another study using these data to examine Species-Abundance-Biomass relationships in response to sediment contamination, we noted a marked decrease in the B/ A ratio with increasing contamination, suggesting that large-bodied (i.e., equilibrium) taxa declined with increasing contamination (Rakocinski et al. In press).
Benthic macroinvertebrates are major components of aquatic food webs, and their life histories make them especially susceptible to sediment contaminant exposure effects. The individual and collective responses of macrobenthic organisms are, therefore, useful for assessing risks from contaminants to aquatic ecosystems (Cairns 1985; Long' and Chapman 1985; Chapman 1989; U.S. Environmental Protection Agency 1989; Chapman et al. 1991, 1992; Bartell et al. 1992; Suter 1993). Results
Sediment Contaminants and Trophic Structure 421
from this study demonstrated that sediment contaminants are quite important in structuring macrobenthic communities in soft-bottom estuarine habitats. It follows that differences in community structure may result in shifts in community / ecosystem function. These results help demonstrate that trophic structure may be a useful and ecologically meaningful response variable in studies of estuarine ecotoxicology, as Diaz (1992) suggests. The potential benefits of developing and using community trophic response indicators are analogous to those derived from studies of aquatic insect functional feeding-groups in stream ecology (sensu Vannote et al. 1980; Barnes and Minshall 1983; Cummins et al. 1983; Cushing et al. 1983; Minshall et al. 1983; Power et al. 1988; Plaf'kin et al. 1989; Barbour et al. 1992). The observations made in this study suggest that trophic response variables may provide an effective means for assessing integrated community responses to chronic sublethal exposure. These kinds of data provide a link between community information and the effects of contaminants on the functional qualities of aquatic ecosystems.
ACKNOWLEDGMENTS
This study was conducted under the auspices of the U.S. Environmental Protection Agency's (USEPA) Environmental Monitoring and Assessment Program. We are grateful to the personnel at the USEPA Environmental Research Laboratory in Gulf Breeze, Florida for their support, and we especially thank K. Summers, J. Macauley, V. Engle, and T. Heitmuller. We thank A. McAllister, D. Hard, R. Woods, and S. LeCroy for laboratory and taxonomic assistance, and W. Walker for his support and encouragement. Finally, we are grateful for the thorough review of the manuscript provided by an anonymous reviewer, whose insightful and constructive comments strengthened the manuscript.
LITERATURE CITED
ADAMS, W. J., R. A. KiMERLE, AND R. G. MOSHER. 1985. Aquatic safety assessment of chemicals sorbed to sediments, p. 429-453. In R. D. Cardwell, R. Purdy, and R. C. Bahner (eds.), Aquatic Toxicology and Hazard Assessment: Seventh Symposium. American Society for Testing and Materials, Philadel-phia, Pennsylvania. .
ALLER, R. C. 1982. The effects of macrobenthos on chemical properties of marine sediment and overlying water, p. 53-102. In P. L. McCall and M. J. S. Tevesz (eds.), Animal-Sediment Relations. Plenum Publishing Corporation, New York.
ANDERSON,J., W. BIRGE,J. GENTlLE,J. LAKE,J. RODGERS,JR., AND R. SWARTZ. 1984. Biological effects, bioaccumulation, and ecotoxicology of sediment-associated chemicals, p. 267-296. In K. L. Dickson, A. W. Maki, and W. A. Brungs (eds.), Fate and Effects of Sediment-Bound Chemicals in Aquatic Systems. Pergamon Press, Elmsford, New York.
BAKER, R. A. (ED.). 1980. Contaminants and Sediments, Volume 1. Ann Arbor Science Publishers, Ann Arbor, Michigan.
BARBOUR, M. T., J. L. PLAFKlN, B. P. BRADLEY, C. G. GRAVES, AND R. W. WISSEMAN. 1992. Evaluation of EPA's rapid bioassessment benthic metrics: Metric redundancy and variability among reference stream sites. Environmental Toxicology and Chemistry 11:437-450.

422 s. S. Brown et al.
BARNES, J. R AND G. W. MINSHALL. 1983. Stream Ecology: Application and Testing of General Ecological Theory. Plenum Press, New York.
BARTELL, S. M., R H. GARDNER, AND R V. O'NEILL. 1992. Ecological Risk Estimation. Lewis Publishers, Inc., Boca Raton, Florida.
BAUDO, R,J. P. GIESY, AND H. MUNTAU. 1990. Sediments: Chemistry and Toxicity of In-Place Pollutants. Lewis Publishers, Inc., Chelsea, Michigan.
BOESCH, D: F. AND R ROSENBERG. 1981. Response to stress in marine benthic communities, p. 179-200. In G. W. Barrett and R Rosenberg (eds.), Stress Effects on Natural Ecosystems. Wiley and Sons, Inc., New York.
BOESCH, D. F., M. L. WASS, AND R W. VIRNSTEIN. 1976. The dynamics of estuarine benthic communities, p. 245-256. In M. Wiley (ed.), Estuarine Processes. Academic Press, New York.
BOOTHMAN, W., D. DI TORO, AND J. MAHONY. 1990. Determination of acid volatile sulfides in sediments, p. 94-102. In Laboratory Methods Manual, U.S. Environmental Protection Agency Environmental Monitoring Systems Laboratory, Cincinnati, Ohio.
BURTON, JR., G. A. 1992. Sediment Toxicity Assessment. Lewis Publishers, Inc., Boca Raton, Florida.
CAIRNS, JR., J. (ED.). 1985. Multispecies Toxicity Testing. Pergamon Press, New York.
CAIRNS, JR., J. AND J. R PRATT. 1986. On the relation between structural and functional analyses of ecosystems. Environmental Toxicology and Chemistry 5:785-786.
CHAPMAN, P. M. 1989. Current approaches to developing sediment quality criteria. Environmental Toxicology and Chemistry 8: 589-599.
CHAPMAN, P. M., E. R LONG, R C. SWARTZ, T. H. DEWITT, AND R PASTOROK. 1991. Sediment toxicity tests, sediment chemistry, and benthic ecology do provide new insights into the significance and management of contaminated sediments-a reply to Robert Spies. Environmental Toxicology and Chemistry 10:1-4.
CHAPMAN, P. M., E. A. POWER, AND G. A. BURTON, JR. 1992. Integrative assessments in aquatic ecosystems, p. 313-340. In G. A. Burton,Jr. (ed.), Sediment Toxicity Assessment. Lewis Publishers, Inc., Boca Raton, Florida.
COHEN, J. E. AND F. BRIAND. 1990. Trophic links in community food webs, p. 31-41. In J. E. Cohen, F. Briand, and C. M. Newman (eds.), Community Food Webs. Springer-Verlag, New York.
COMMITO,J. A. AND W. G. AMBROSE. 1985. Multiple trophic levels in soft-bottom communities. Marine Ecology Progress Series 26: 289-293.
CUMMINS, K. W., J. R SEDELL, F. J. SWANSON, G. W. MINSHALL, S. G. FISHER, C. E. CUSHING, R C. PETERSON, AND R L. VANNOTE. 1983. Organic matter budgets for stream ecosystems: Problems in their evaluation, p. 299-353. In J. R Barnes and G. W. Minshall (eds.), Stream Ecology: Application and Testing of General Ecological Theory. Plenum Press, New York.
CUSHING, C. E., C. D. McINTIRE, K. W. CUMMINS, G. W. MINSHALL, R C. PETERSON,J. R SEDELL, AND R L. VANNOTE. 1983. Relationships among chemical, physical, and biological indices along river continua based on multivariate analysis. Archives of Hydrobiologia 98:317-326.
DAUER, D. M. 1984. The use of polychaete feeding guilds as biological variables. Marine Pollution Bulletin 15:301-305.
DAUER, D. M., C. A. MAYBURY, AND R M. EWING. 1981. Feeding behavior and general ecology of several spionid polychaetes from the Chesapeake Bay. Journal of Experimental Marine Biology and Ecology 54:21-38.
DAY, JR., J. w., C. A. S. HALL, W. M. KEMP, AND A. YANEZ-ARANCIBIA. 1989. Estuarine Ecology. John Wiley and Sons, Inc., New York.
DIAZ, R J. 1980. Ecology of tidal freshwater and estuarine tubificidae (Oligochaeta), p. 319-330. In R O. Brinkhurst and D. G. Cook (eds.), Aquatic Oligochaete Biology. Plenum Press, New York.
DIAZ, R J. 1992. Ecosystem assessment using estuarine and marine benthic community structure, p. 67-85. In G. A. Burton (ed.), Sediment Toxicity Testing. Lewis Publishers, Chelsea, Michigan.
DIAZ, R J. AND L. C. SCHAFFNER. 1990. The functional role of estuarine benthos, p. 25-56. In M. Haire and E. C. Krome (eds.), Perspectives on the Chesapeake Bay: Advances in Estuarine Science. Report No. CBP/TRS41/90. Chesapeake Research Consortium, Gloucester, Virginia.
DICKSON, K. L., A. W. MAKI, AND W. A. BRUNGS. 1984. Fate and Effects of Sediment-Bound Chemicals in Aquatic Systems. Pergamon Press, Elmsford, New York.
DI TORO, D. M., C. S. ZAlmA, D. J. HANSEN, W. J. BERRY, R C. SWARTZ, C. E. COWEN, S. P. PAVLOU, H. E. ALLEN, N. A. THOMAS, AND P. R PAQUIN. 1991. Technical basis for establishing sediment quality criteria for nonionic organic chemicals using equilibrium partitioning. Environmental Toxicology and Chemistry 10:1541-1583.
FAUCHALD, K. AND P. A. JUMARS. 1979. The diet of worms: A study of polychaete feeding guilds. Oceanography and Marine Biology Annual Review 17:193-284.
FERRARO, S. P., R C. SWARTZ, F. A. COLE, AND D. W. SCHULTS. 1991. Temporal changes in the benthos along a pollution gradient: Discriminating the effects of natural phenomena from sewage-industrial wastewater effects. Estuarine Coastal and Shelf Science 33:383-407.
GASTON, G. R 1987. Benthic Polychaeta of the Middle Atlantic Bight: Feeding and distribution. Marine Ecology Progress Series 36:251-262.
GASTON, G. R, BROWN, S. S., C. F. RAKOCINSKI, S. S. BROWN, AND C. M. CLEVELAND. 1998. Trophic function in estuaries: Response of macrobenthos to natural and contaminant gradients. Marine and Freshwater Research 49:833-846.
GASTON, G. R, C. M. CLEVELAND, S. S. BROWN, AND C. F. RAKOCINSKI. 1997. Benthic-pelagic coupling in northern Gulf of Mexico estuaries: Do benthos feed directly on phytoplankton? Gulf Research Reports 9:231-237.
GASTON, G. RAND K. A. EDDS. 1994. Long-term study of benthic communities on the continental shelf off Cameron, Louisiana: A review of brine effects and hypoxia. Gulf Research Reports 9:57-64.
GASTON, G. R AND J. C. NASCI. 1988. Trophic structure of macrobenthic communities in the Calcasieu Estuary, Louisiana. Estuaries 11:201-211.
GASTON, G. R, P. A. RUTLEDGE, AND M. L. WALTHER. 1985. The effects of hypoxia and brine on recolonization by macrobenthos off Cameron, Louisiana (USA). Contributions to Marine Science 28:79-83.
GASTON, G. R AND J. C. YOUNG. 1992. Effects of contaminants on macrobenthic communities in the upper Calcasieu Estuary, Louisiana. Bulletin of Environmental Contamination and Toxicology 49:922-928.
GRAVES, R L.;J. LAZORCHAK, R VALENTI, D. McMULLEN, AND K. WINKS. 1990. Environmental monitoring and assessment program laboratory methods manual near coastal demonstration project. U.S. Environmental Protection Agency Environmental Monitoring Systems Laboratory, Cincinnati, Ohio.
HEITMULLER, T. AND R VALENTE. 1991. Near coastal Louisianian Province monitoring quality assurance project plan. EPA/ 600/X-91/XXX. U.s. Environmental Protection Agency Office of Research and Development. Gulf Breeze, Florida.
JENNE, E. A. AND J. M. ZACHARA. 1984. Factors influencing the sorption of metals, p. 83-98. In K. L. Dickson, A. W. Maki, and W. A. Brungs (eds.), Fate and Effects of Sediment-Bound

Chemicals in Aquatic Systems. Pergamon Press, Elmsford, New York.
KARIcKHOFF, S. W. AND K. R MORRIs. 1985. Sorption dynamics of hydrophobic pollutants in sediment suspensions. Environmental Toxicology and Chemistry 4:467-479.
KRAHN, M. M., C. A WIGREN, R W. PEARCE, L. K. MOORE, R. G. BOGAR, W. D. MAcLEOD, S. L. CHAN, AND D. W. BROWN. 1988. Standard Analytical Procedures of the NOAA National Analytical Facility, 1988. New HPLC Cleanup and Revised Extraction Procedures for Organic Contaminants. National Oceanic and Atmospheric Administration Technical Memorandum, National Marine Fisheries Service F/NWC-153. U.S. Department of Commerce, National Oceanic and Atmospheric Administration, National Marine Fisheries Service; Seattle, Washington.
LANDRUM, P. F. AND]. A. ROBBINS. 1990. Bioavailability of sediment-associated contaminants to benthic invertebrates, p. 237-264. In R Baudo,]. Giesy, and H. MUnlau (eds.), Chemistry and Toxicity of In-Place Pollutants. Lewis Publishers, Boca Raton, Florida.
LEVIN, L. A. 1981. Dispersion, feeding behavior and competition in two spionid polychaetes. Journal of Marine Research 39:99-117.
LEVINTON,]. S. 1972. Stability and trophic structure in depositfeeding and suspension-feeding communities. American Naturalist 106:472-486.
LoNG, E. R AND M. F. BUCHMAN. 1989. An evaluation of candidate measures of biological effects for the National Status and Trends Program. National Oceanic and Atmospheric Administration Technical Memorandum NOS OMA 45. U.S. Department of Commerce, Seattle, Washington.
LoNG, E. RAND P. M. CHAPMAN. 1985. A sediment quality triad: Measures of sediment contamination, toxicity and infaunal community composition in Puget Sound. Marine Pollution Bulletin 16:405-415.
LONG, E. R, D. D. MAcDONALD, S. L. SMITH, AND F. D. CALDER. 1995. Incidence of adverse biological effects within ranges of chemical concentrations in marine and estuarine sediments. Environmental Management 19:81-97.
LONG, E. R AND L. G. MORGAN. 1990. The potential for biological effects of sediment-sorbed contaminants tested in the National Status and Trends Program. National Oceanic and Atmospheric Administration Technical Memorandum NOS OMA 52. National Oceanic and Atmospheric Administration National Ocean Service. Seattle, Washington.
MACLEOD, W. D., D. W. BROWN, A]. FRIEDMAN, D. G. BURROWS, O. MAYNES, R. W. PEARCE, C. A WIGREN, AND R G. BOGAR. 1985. Standard Analytical Procedures of the NOAA National Analytical Facility, 1985-1986, Extractable Toxic Organic Compounds, 2nd edition. National Oceanic and Atmospheric Administration Technical Memorandum NMFS F /NWC-92. U.S. Department of Commerce, National Oceanic and Atmospheric Administration, National Marine Fisheries Service, Seattle, Washington.
MATISOFF, G.,]. B. FISHER, AND S. MATIS. 1985. Effects of benthic macroinvertebrates on the exchange of solutes between sediments and freshwater. Hydrobiologia 122:19-33.
MAURER, D. AND W. LEATHEM. 1981. Polychaete feeding guilds from Georges Bank, USA Marine Biology 62:161-171.
MEDINE, A]. AND S. C. MCCUTCHEON. 1989. Fate and transport of sediment-associated contaminants, p. 225-292. In]. Saxena (ed.), Hazard Assessment of Chemicals, Volume 6. Hemisphere Publishing Company, New York.
MINSHALL, G. W., R C. PETERSON, K. W. CUMMINS, T. L. BOTT, ]. R. SEDELL, C. E. CUSHING, AND R L. VANNOTE. 1983. Interbiome comparison of stream ecosystem dynamics. Ecological Monographs 53:1-25.
PEARSON, T. H. AND R ROSENBERG. 1978. Macrobenthic succession in relation to organic enrichment and pollution of the
Sediment Contaminants and Trophic Structure 423
marine environment. Oceanography and Marine Biology Annual Review 16:229-311.
PETERSON, C. H. 1991. Intertidal zonation of marine invertebrates in sand and mud. American Scientist 79:236-249.
PIELOU, E. C. 1975. Ecological Diversity. John Wiley and Sons, Inc., New York.
PLAFKIN,]. F., M. T. BARBOUR, K. D. PORTER, S. K. GROSS, AND R M. HUGHES. 1989. Rapid Bioassessment Protocols for Use in Streams and Rivers: Benthic Invertebrates and Fish. EPA 444/4-89-001. U.S. Environmental Protection Agency, Washington, D.C.
PWMB,JR., R H. 1981. Procedures for handling and chemical analysis of sediment and water samples. U.S. Environmental Protection Agency/U.S. Army Corps of Engineers Technical Committee on criteria for dredged and fill material. Vicksburg, Mississippi.
POWER, M. E., R]. STOUT, C. E. CUSHING, P. P. HARPER, F. R HAUER, W.]. MATTHEWS, P. B. MoYLE, B. STATZNER, AND I. R WAiS DE BADGEN. 1988. Biotic and abiotic controls in river and stream communities. Journal of the North American Benthological Society 7:456-479.
RABALAIS, N. N. AND D. E. HARPER. 1992. Studies of benthic biota in areas affected by moderate and severe hypoxia. Proceedings of National Oceanic and Atmospheric Administration workshop on nutrient enhanced coastal ocean productivity. October 1991. Texas A&M University Sea Grant Program, College Station, Texas.
RAKOCINSKI, C. F., S. S. BROWN, G. R GASTON, R. W. HEARD, W. W. WALKER, AND]. K. SUMMERS. 1997. Macrobenthicresponses to natural and contaminant-related gradients in northern Gulf of Mexico estuaries. Ecological Applications 7:1278--1298.
RAKOCINSKI, C. F., S. S. BROWN, G. R GASTON, R. W. HEARD, W. W. WALKER, AND]. K. SUMMERS. in press. Species-AbundanceBiomass responses by estuarine macrobenthos to sediment chemical contamination. Journal of Aquatic Ecosystem Stress and RecUIJery.
RHOADS, D. C. 1974. Organism-sediment relations on the muddy sea floor. Oceanography and Marine Biology Annual Review 12: 263-300.
RHOADS, D. C., P. L. McCALL, AND]. L. YiNGST. 1978. Disturbance and production on the estuarine seafloor. American Scientist 66:577-586.
RHOADS, D. C. AND D. K. YOUNG. 1970. The influence of depositfeeding organisms on sediment stability and community trophic structure. Journal of Marine Research 28:150-178.
RODGERS, JR.,]. H., K. L. DICKSON, F. Y SALEH, AND C. A STAPLES. 1984. Bioavailability of sediment-bound chemicals to aquatic organisms-Some theory, evidence, and research needs, p. 245-266. In K. L. Dickson, A. W. Maki, and W. A Brungs (eds.), Fate and Effects of Sediment-Bound Chemicals in Aquatic Systems. Pergamon Press, Elmsford, New York.
SANDERS, H. L. 1956. Oceanography of Long Island Sound, 1952-1954. X. Biology of marine bottom communities. Bulletin of the Bingham Oceanographic Collection 15:345-414.
SANDERS, H. L. 1958. Benthic studies in Buzzards Bay. I. Animalsediment relationships. Limnology and Oceanography 3:245-258.
SCHAFFNER, L. C. 1990. Small-scale organism distributions and patterns of species diversity: Evidence for positive interactions in an estuarine benthic community. Marine Ecology Progress Series 61:107-117.
SCHAFFNER, L. C., R]. DIAZ, C. R OLSEN, AND I. L. LARsEN. 1987. Faunal characteristics and sediment accumulation processes in the James River estuary, Virginia. Estuarine and Coastal Shelf Science 25:211-226.
SPIES, R B. 1989. Sediment bioassays, chemical contaminants and benthic ecology: New insights or just muddy water? Marine Environmental Research 27:73-75.
SUMMERS,]. K. AND V. D. ENGLE. 1993. Evaluation of sampling strategies to characterize dissolved oxygen conditions in

424 s. S. Brown et at.
northern Gulf of Mexico estuaries. Environmental Monitoring and Assessment 24:219-229.
SUMMERS,]. K.,]. M. MACAULEY, AND P. T. HEITMULLER. 1991. Implementation plan for monitoring the estuarine waters of the Louisianian Province-1991. EPAj600jR-91j228. U.S. Environmental Protection Agency Environmental Research Laboratory, Gulf Breeze, Florida.
SUMMERS,]. K.,]. M. MACAULEY, P. T. HEITMULLER, V. D. ENGLE, A. M. ADAMS, AND G. T. BROOKS. 1993. Statistical summary: EMAP-Estuaries Louisianian Province-1991. EPAj 620 jR-93 j 007, January. U.S. Environmental Protection Agency Environmental Research Laboratory, Gulf Breeze, Florida.
SUTER, JR., G. W. 1993. Ecological Risk Assessment. Lewis Publishers, Boca Raton, Florida.
SWARTZ, R C., W. A. DEBEN,]. K. JONES,]. O. LAMBERSON, AND F. A. COLE. 1985. Phoxocephalid amphipod bioassay for marine sediment toxicity, p. 284-307. In RD. Cardwell, R Purdy, and R C. Bahner (eds.), Aquatic Toxicology and Hazard Assessment: Seventh Symposium. American Society of Testing and Materials, Philadelphia, Pennsylvania.
TAYLOR, ]. K. 1987. Quality assurance of chemical measurements. Lewis Publishers, Inc., Chelsea, Michigan.
TENORE, K. R, R B. HANSON,]. MCCLAIN, A. E. MACCUBBIN, AND R E. HODSON. 1984. Changes in composition and nutritive value to a benthic deposit feeder of decomposing detritus pools. Bulletin of Marine Science 35:299-311.
U.S. ENVIRONMENTAL PROTECTION AGENCY. 1989. Risk Assessment Guidance Document for Superfund, Volume 2, Environmental Evaluation Manual, Interim Final. EPAj540jl-89j 001, March. U.S. Environmental Protection Agency Office of Emergency and Remedial Response, Washington, D.C.
VANNOTE, R L., G. W. MINSHALL, K. W. CUMMINS,]. R SEDELL, AND C. E. CUSHING. 1980. The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences 37: 130-137.
WEINBERG,]. R 1984. Interactions between functional groups in soft substrata: Do species differences matter? Journal of Experimental Marine Biology and Ecology 80: 11-28.
WESTON, D. P. 1990. Quantitative examination of macro benthic community changes along an organic enrichment gradient. Marine Ecology Progress Series 61:233-244.
WHITLATCH, R B. 1981. Animal-sediment relationships in intertidal marine benthic habitats: Some determinants of depositfeeding species diversity. Journal of Experimental Marine Biology and Ecology 53:31-45.
ZAR,]. H. 1984. Biostatistical Analysis, 2nd edition. PrenticeHall, Inc., Englewood Cliffs, New Jersey.
ZOBELL, C. E. AND C. B. FELTHAM. 1938. Bacteria as food for certain marine invertebrates. Journal of Marine Research 1 :312-327.
Received for consideration, April 6, 1998 Accepted for publication, February 11, 2000
Copyright © 2022 FDOKUMEN




















