
Effects of mass fish removal on coregonid larval abundance in a large mesotrophic lake
14
Effects of mass fish removal on coregonid larval abundance in a large mesotrophic lake Olli Urpanen 1, *, Tapio Keskinen 1 , Timo J. Marjomäki 1 , Ville Sakomaa 1 , Hannu Salo 3 , Jukka Syrjänen 1 , Markku Viljanen 2 , Juha Karjalainen 1 with 6 figures and 2 tables Abstract: It has been proposed that perch populations can regulate the size of vendace (Coregonus albula (L.)) stocks due to predation and prevent the recovery of stocks by prolonging the low stock phase. Thus, the impacts of a mass fish removal on the densities of vendace and whitefish (C. lavaretus L. s.l.) larvae were examined in a mesotrophic Finnish lake (area 4 600 ha) with a Beyond BACI design with four control lakes. The goal of the mass removal was to improve coregonid stocks. Total biomass of fish removed was about 428 tons (33 kg ha -1 ) during four intensive fishing years (2001–2004). Major proportions of fish removed were roach (Rutilus rutilus) (46%) and perch (Perca fluviatilis) (24%). Newly hatched coregonid larvae densities were estimated for three years before and six years after the beginning of fishing. Regardless of slight increases in larval vendace densities, no significant differences in larval densities between study periods were found. This study confirmed previous results, but did not produce any new support for the perch predation hypothesis. The fish removal may not have been sufficiently high or the fishing period may have not been long enough to strengthen the very low spawning stock and affect the density of larval coregonids. Keywords: coregonid, larval, fish removal. Introduction Predation is one of the main factors affecting stock density for many fishes: it is usually size-dependent and larval and juvenile fishes are the most vulnerable. It is proposed that the populations of piscivorous fishes, especially perch, may regulate the size of vendace (Coregonus albula (L.)) stocks due to predation and prevent the recovery of stocks by pro- Authors’ addresses: 1 Department of Biological and Environmental Science, Aquatic Sciences, University of Jyväskylä, P.O. Box 35, 40014 Jyväskylä, Finland. 2 Ecological Research Institute, Faculty of Science and Forestry, University of Eastern Finland, P.O. Box 111, 80101 Joensuu, Finland. 3 Institute for Environmental Research, University of Jyväskylä, P.O. Box 35, 40014 Jyväskylä, Finland. * Corresponding author, e-mail: olli.urpanen@metsa.fi 1612-166X/12/0063-0383 $ 3.50 © 2012 E. Schweizerbart’sche Verlagsbuchhandlung, 70176 Stuttgart, Germany Advanc. Limnol. 63, p. 383–395 Biology and Management of Coregonid Fishes – 2008
Transcript of Effects of mass fish removal on coregonid larval abundance in a large mesotrophic lake

Effects of mass fi sh removal on coregonid larval abundance in a large mesotrophic lake
Olli Urpanen1,*, Tapio Keskinen1, Timo J. Marjomäki1, Ville Sakomaa1, Hannu Salo3, Jukka Syrjänen1, Markku Viljanen2, Juha Karjalainen1
with 6 fi gures and 2 tables
Abstract: It has been proposed that perch populations can regulate the size of vendace (Coregonus albula (L.)) stocks due to predation and prevent the recovery of stocks by prolonging the low stock phase. Thus, the impacts of a mass fi sh removal on the densities of vendace and whitefi sh (C. lavaretus L. s.l.) larvae were examined in a mesotrophic Finnish lake (area 4 600 ha) with a Beyond BACI design with four control lakes. The goal of the mass removal was to improve coregonid stocks. Total biomass of fi sh removed was about 428 tons (33 kg ha-1) during four intensive fi shing years (2001–2004). Major proportions of fi sh removed were roach (Rutilus rutilus) (46%) and perch (Perca fl uviatilis) (24%). Newly hatched coregonid larvae densities were estimated for three years before and six years after the beginning of fi shing. Regardless of slight increases in larval vendace densities, no signifi cant differences in larval densities between study periods were found. This study confi rmed previous results, but did not produce any new support for the perch predation hypothesis. The fi sh removal may not have been suffi ciently high or the fi shing period may have not been long enough to strengthen the very low spawning stock and affect the density of larval coregonids.
Keywords: coregonid, larval, fi sh removal.
Introduction
Predation is one of the main factors affecting stock density for many fi shes: it is usually size-dependent and larval and juvenile fi shes are the most vulnerable. It is proposed that the populations of piscivorous fi shes, especially perch, may regulate the size of vendace (Coregonus albula (L.)) stocks due to predation and prevent the recovery of stocks by pro-
Authors’ addresses:1 Department of Biological and Environmental Science, Aquatic Sciences, University of Jyväskylä,
P.O. Box 35, 40014 Jyväskylä, Finland.2 Ecological Research Institute, Faculty of Science and Forestry, University of Eastern Finland, P.O.
Box 111, 80101 Joensuu, Finland.3 Institute for Environmental Research, University of Jyväskylä, P.O. Box 35, 40014 Jyväskylä,
Finland.* Corresponding author, e-mail: [email protected]
1612-166X/12/0063-0383 $ 3.50© 2012 E. Schweizerbart’sche Verlagsbuchhandlung, 70176 Stuttgart, Germany
Advanc. Limnol. 63, p. 383–395Biology and Management of Coregonid Fishes – 2008

384 O. Urpanen et al.
longing low stock phases (HELMINEN & SARVALA 1994, HEIKINHEIMO 2001, VALKEAJÄRVI & MARJOMÄKI 2004). Based on dynamic modelling, HEIKINHEIMO (2001) suggested that perch has more effect on vendace stocks than brown trout (Salmo trutta m. lacustris) due to more dense stocks and fl exible behaviours.
Mass fi sh removals have been used in biomanipulation projects in attempts to change the structure of fi sh and plankton communities and to improve water quality (e.g., SHAPIRO & WRIGHT 1984). The targets for biomanipulation have usually been small lakes and eutrophic lakes (JEPPESEN et al. 2007). The abundances of self-sustaining populations of coregonids in these lakes have been low, and the fi sh communities have been dominated by cyprinids. Only a few biomanipulated lakes have been large (area >10 000 ha), one of them being Lake Vesijärvi, a eutrophied lake in southern Finland where about 400 kg ha-1 of fi sh were removed during 1989–1993 (KAIRESALO et al. 1999). Besides an improvement in water quality, consid-erable changes in fi sh community structure occurred. Instead of cyprinid dominance, natural stocks of pike (Esox lucius), pikeperch (Sander lucioperca), whitefi sh (Coregonus lavaretus) and vendace increased their proportions in fi sh catches (KAIRESALO et al. 1999, KETO & TALL-BERG 2000). However, substantial fi shing effort was needed to maintain the new structure and prevent the return of a dense roach stock (HORPPILA & PELTONEN 1994, KAIRESALO et al. 1999).
In this study, we analyzed whether the mass removal of perch (Perca fl uviatilis) and cyprinids, potential predators and competitors of coregonids, can improve the recruitment success of vendace and whitefi sh in northern Lake Päijänne, Central Finland. Larval densities of coregonids were monitored before (1999–2001) and after (2002–2007) the beginning of mass removals in Lake Päijänne and four control lakes, and were analyzed as a Before-After-Control-Impact (BACI) design. The aim of the restoration project, which was funded by the local fi sheries administration, was to increase the economic values of the fi sh community and to improve opportunities for the professional fi shery.
Material and methods
Study and control lakes
Mass removals of percids and cyprinids were performed in northern Lake Päijänne (Fig. 1) located near the city of Jyväskylä, Finland. The lake is mesotrophic with a total phosphorus concentration of 15 mg m-3 and a water colour of 40 mg Pt l-1 (Table 1). The lake area is 15 700 ha and the mass fi sh removals were performed in a 4 600 ha area in the far northern part of the study area (Fig. 1). Coregonid larval
Table 1. Characteristics of study and control lakes.
Lakes Area(ha)
Max. depth(m)
Mean depth(m)
Total P(mg m-3)
Colour(Pt mg
l-1)
Secchi depth(m)
N. Päijänne 15 700 94 16 15 40 3.7S. Konnevesi 13 000 56 13 8 25 4.2Puulavesi 19 000 60 12 4 15 5.3Onkamo 3 227 13 4 20 15 3.9SW. Pyhäjärvi 15 400 26 5 31 15 3.3

Effects of mass fi sh removal on coregonid larval abundance 385
Fig. 1. Locations of lakes and study area in Northern Päijänne.
sampling covered the entire study area. The control lakes were Lakes southern Konnevesi, Onkamo, Puulavesi and southwestern Pyhäjärvi (Fig. 1, Table 1).
Based on gillnet fi shing, about 40% of the fi sh community in northern Päijänne consisted of perch and roach in the fi rst year (2001) of the fi sh removal project (ALAJA 2008). Most of the perch population consisted of small individuals (<15 cm in total length). The proportion of coregonids was low (<1 %) and pikeperch was the most common predatory fi sh. In northern Päijänne, the populations of coregon-ids were scarce, mainly because the spawning stocks were low.

386 O. Urpanen et al.
Fish removal and monitoring
Intensive mass fi sh removals were performed in 2001–2004 (annually from ice-off to the end of June) by professional fi shers. The majority of catches were caught by trap nets. The effects of mass removals on the fi sh community were monitored by gillnet fi shing in Lake Päijänne. Nets included 9 mesh sizes (from knot to knot) 10, 12, 15, 20, 25, 30, 35, 45 and 55 mm. Net locations were based on a stratifi ed random sampling design. The fi shing period of each net was approximately 24 h. Gillnet fi shing was done in July–September in 2001–2007. The annual fi shing effort was 72–103 net days. The catch-per-unit-effort (CPUE, g or individual net day-1) was used as an index of fi sh stock abundance.
Larval coregonid sampling
Newly hatched coregonid larval densities were estimated in northern Päijänne and in the four control lakes during three years (1999–2001) before the beginning of the intensive fi shing and six years (2002–2007) after the start of the fi shing period. Hatching of coregonid larvae in Finnish lakes occurs in early spring (KARJALAINEN et al. 2000, URPANEN et al. 2005) and larval sampling was conducted annually in May, immediately after ice-off in each lake. Thus, coregonid larval densities in 2001 were not affected by the mass fi sh removal in the same year, and the data of larval densities in that year were suitable for Beyond BACI-design as a before sampling period (see later in methods).
Estimates of the total number and density of newly hatched larvae were based on a stratifi ed random sampling design. The consistent survey procedure outlined by KARJALAINEN et al. (1998) was used in all years and lakes. Generally, 20 littoral and 10 pelagic sampling sites (1 x 1 km) were sampled annually in each lake. At each littoral site, larvae were sampled from fi ve different strata: stratum I, from shore-line to 0.5 m of bottom depth; stratum II, 0.5–1 m; stratum III, 1–2 m; stratum IV, generally 2–4 m; and stratum V, pelagic. Larvae from the shallowest littoral areas (stratum I) were collected with a manually pushed tube-net. Four tube-net replicates were taken and pooled from stratum I in each sampling site. Samples from strata II, III, IV and V were gathered by rectangular bongo nets with rounded corners (30 x 60 cm, mouth area 0.24 m2) that were pushed in front of a boat equipped with a jet engine. Net length was 2.5 m with a mesh size of 0.50 mm. Generally, two bongo nets were used in each sampling tow. In stratum II, sampling was done at only one water depth layer (0–30 cm) and consisted of two pooled bongo net samples. In strata III, IV and V, one net was used to collect a sample from the water depth layer of 0–30 cm and another net was used for the 30–60 cm layer. The time spent on each bongo-net haul at a speed of 4–6 km h-1 was 4 minutes in the littoral areas and 10 minutes in the pelagic areas. The volume of each sample was measured by a fl ow meter. All larval samples were preserved in a 1:1 mixture of 70% ethanol and 1% formalin. Species identifi cation of larvae was done in the laboratory according to KARJALAINEN et al. (1992).
Calculations
Population size of newly hatched larvae for a whole lake was estimated based on the samples taken from the different sampling sites, strata and depth layers. First, the whole lake average density Dstratum,layer (individuals 100 m-3) for each stratum and depth layer were estimated from the stratum- and layer-specifi c observations, dstratum,layer,site, in the different sampling sites. Second, the average density in each stratum, Dstratum, (individuals 100 m-3) was estimated based on observed densities at the depth layers of 0–30 cm and 30–60 cm and the predicted densities at the depth layers of >60 cm (described in more detail in URPANEN et al. (2009). Thirdly, the larval population size (million individuals) for the whole lake was calculated by weighting the stratum-specifi c average densities by the volumes of the different strata: in strata I, II and III, the volumes included depth layers from surface to bottom and in strata IV and V from surface to depth of 2 m.

Effects of mass fi sh removal on coregonid larval abundance 387
Larval density estimates between the northern and southern parts of northern Päijänne were com-pared using 2-way ANOVA with study area and sampling period (years before (3 years) and after (6 years) intensive fi shing) as explanatory variables. Data was log (x+1) -transformed before analysis.
The effect of mass fi sh removal on coregonid larval densities in northern Päijänne was analyzed using asymmetric analysis of variance, i.e., a Beyond BACI (Before-After-Control-Impact) design, following the procedure described by UNDERWOOD (1992: pages 156–159).
Results
Fish removal
In 2001–2004, 428 tonnes of fi sh (93 kg ha-1) were removed from the northern part of north-ern Päijänne (Fig. 1), mainly by trap net. Ninety-six percent of the total catch by weight consisted of roach (Rutilus rutilus), bream (Abramis brama), perch and bleak (Alburnus alburnus) (Fig. 2). The proportion of pikeperch, pike and brown trout in the catch biomass was only 0.1%. The proportions of whitefi sh and vendace in the total catch biomass were only 0.35% and <0.01%, respectively. Thus, intensive mass fi sh removal was not supposed to affect the spawning stock of either species or their larval densities in successive years. The mass removal did not signifi cantly affect the gillnet CPUE of perch or roach (Fig. 3), although a decrease of ca. 22% in perch CPUE was observed from 2001 to 2004.
Larval densities
Overall mean densities of newly hatched vendace and whitefi sh larvae in northern Päijänne were very low during the study period compared to the other lakes (Fig. 4). Mean (± S.E.) ven-dace density in northern Päijänne was 386 ± 96 ind. ha-1 in the whole study period. In contrast, in Lake Puulavesi, the mean (± S.E.) density of vendace larvae was 6665 ± 2042 ind. ha-1.
In northern Päijänne, the density of larval vendace did not differ either between the north-ern and southern parts of the study area or between before and after time periods (P > 0.05). However, the area × time period interaction was signifi cant (F1,159 = 8.831, P < 0.01). Mean (± SE) density of vendace larvae in the northern part of the study area was 147 ± 39 ind. ha-1 before fi sh removal and 345 ± 67 ind ha-1 after the start of intensive fi shing (Fig. 5). Densities of whitefi sh larvae were 11 ± 7 ind. ha-1 and 5 ± 4 ind. ha-1, respectively, and no differences between areas or time periods were found (P > 0.05).
Overall vendace larval density in northern Päijänne was 935 (95% c.l. 198–2069) ind. ha-1 in 2007 (Fig. 5), i.e., the highest density observed during the study period. In the same year, larval density of vendace also increased in lakes southern Konnevesi and Puulavesi (Fig. 6). In 2007, the larval density of whitefi sh was also higher compared to previous years in all lakes except in southwestern Lake Pyhäjärvi.
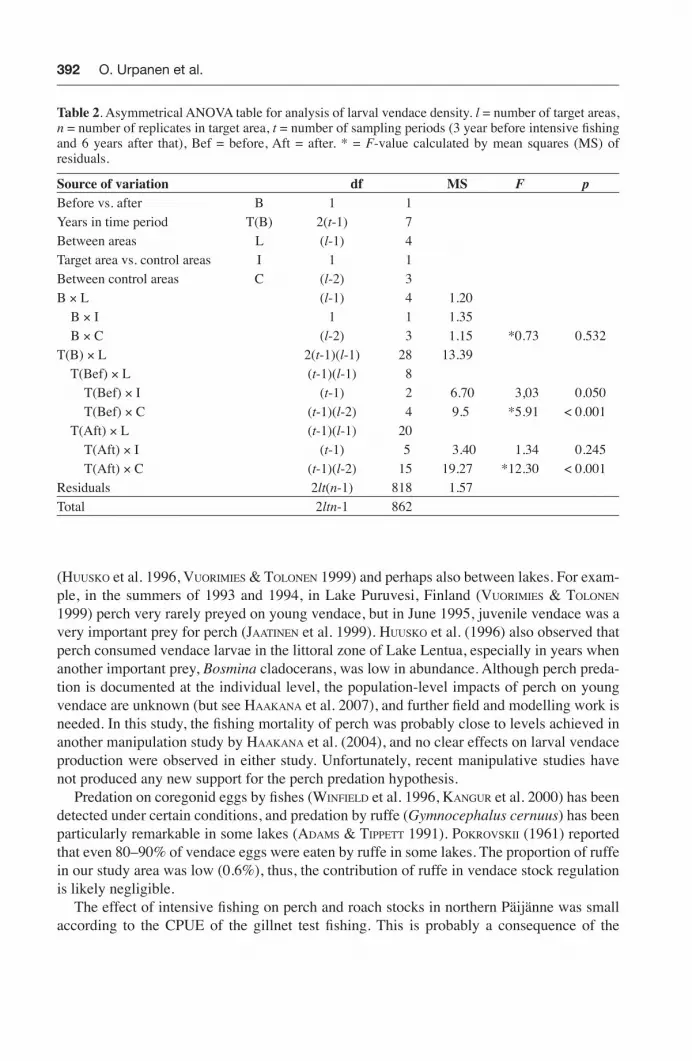
In the BACI analysis for vendace, there was a temporal interaction among control lakes during the after-period (T(Aft) x C, F15, 818 = 12.30, P < 0.001) (Table 2). Because of this, we conducted a comparison for the mean squares of two interactions: years x impact vs. control lakes and years x control lakes. As this comparison was not signifi cant (MS T(Aft) × I / MS T(Aft) x C, F5,15 = 0.17, P = 0.97), no impact could be detected. For whitefi sh, the statistical

388 O. Urpanen et al.
Fig. 2. Catches of different fi sh species during intensive fi shing in 2001–2004.
Fig. 3. Gillnet CPUE (± S.E.) (g net day-1) of perch and roach in northern Päijänne.
procedure was exactly the same (T(Aft) x C, F10, 658 = 4.01, P < 0.001), and again, no impact was found (MS T(Aft) × I / MS T(Aft) x C, F5, 10 = 0.21, P = 0.95). Thus, the mass removal of cyprinids and perch from northern Päijänne had no effect on the densities of larval vendace or whitefi sh.

Effects of mass fi sh removal on coregonid larval abundance 389
Discussion
Regardless of a slight increase in the density of larval vendace in northern Päijänne after the fi sh removal treatment, no signifi cant differences in larval densities between before- and after-periods were found. In 2007, the density of coregonid larvae was high in most lakes and the increases in densities might have resulted from abiotic factors (e.g., weather conditions) that occurred at large scales, since vendace stocks have been shown to fl uctuate synchronous-ly at the geographical scale of this study (MARJOMÄKI et al. 2004). Thus, we did not observe clear effects of the manipulative treatment on the reproductive output of coregonids, possibly because 1) predation and competition by perch or cyprinids did not regulate recruitment of
Fig. 4. Mean (± S.E.) larval densities of vendace (a) and whitefi sh (b) in northern Lake Päijänne and control lakes summed over 1999–2007.

390 O. Urpanen et al.
Fig. 5. Mean (± 95 % c.l.) larval densities of vendace (a) and whitefi sh (b) in northern Lake Päijänne during 1999–2007. Start of intensive fi shing indicated by arrows.
the coregonids in northern Päijänne, or 2) fi sh removal (only a 22% decrease in perch CPUE during the test fi shing) was not high enough to decrease predation and competition by perch or cyprinids, which affect the recruitment of coregonids.
The potential positive effects of the mass removal of perch and cyprinids on coregonids are reductions in predation and competition. Perch is a potential predator of vendace larvae (see below) but competition with cyprinids is not as obvious. Roach is an omnivorous cyprinid with the ability to use planktonic and benthic food resources, but piscivorous feeding has not been reported (EIE & BORGSTRØM 1981, HORPPILA 1994, KAHL & RADKE 2006). Thus, preda-tion by roach on vendace is unlikely. In pelagic habitats, vendace is expected to be a better competitor for zooplankton compared to roach and perch due to having a more effective

Effects of mass fi sh removal on coregonid larval abundance 391
Fig. 6. Mean ln(larval density (ind ha-1) +1) of vendace (a) and whitefi sh (b) in northern Lake Päijänne and control lakes during 1999–2007. Start of intensive fi shing indicated by arrows.
lower jaw and a higher number of gill rakers (SVÄRDSÖN 1976, HAMRIN 1986). Vendace can also forage at low temperatures and low light levels in the hypolimnion (HAMRIN & PERSSON 1986, PERSSON et al. 1991). Moreover, BEIER (2001) has shown that roach and vendace favour different lake habitats. On the other hand, piscivorous perch favour the presence of vendace (APPELBERG et al. 1990, PERSSON et al. 1991), but this may be the result of interactions among perch, roach and vendace, instead of direct perch predation on vendace (BEIER 2001).
Some correlative fi eld results (VALKEAJÄRVI & MARJOMÄKI 2004) and modelling stud-ies (HELMINEN & SARVALA 1994, HEIKINHEIMO 2001) indicated that vendace recruitment and reproductive success may be strongly affected by perch predation. Further, direct evidence of noticeable perch predation on larval (HUUSKO & SUTELA 1992, HUUSKO et al. 1996, HAAKANA et al. 2007) and juvenile coregonids (JAATINEN et al. 1999, TOLONEN 2000) is available. The contribution of coregonids in the diet of perch, however, varies remarkably between years
392 O. Urpanen et al.
(HUUSKO et al. 1996, VUORIMIES & TOLONEN 1999) and perhaps also between lakes. For exam-ple, in the summers of 1993 and 1994, in Lake Puruvesi, Finland (VUORIMIES & TOLONEN 1999) perch very rarely preyed on young vendace, but in June 1995, juvenile vendace was a very important prey for perch (JAATINEN et al. 1999). HUUSKO et al. (1996) also observed that perch consumed vendace larvae in the littoral zone of Lake Lentua, especially in years when another important prey, Bosmina cladocerans, was low in abundance. Although perch preda-tion is documented at the individual level, the population-level impacts of perch on young vendace are unknown (but see HAAKANA et al. 2007), and further fi eld and modelling work is needed. In this study, the fi shing mortality of perch was probably close to levels achieved in another manipulation study by HAAKANA et al. (2004), and no clear effects on larval vendace production were observed in either study. Unfortunately, recent manipulative studies have not produced any new support for the perch predation hypothesis.
Predation on coregonid eggs by fi shes (WINFIELD et al. 1996, KANGUR et al. 2000) has been detected under certain conditions, and predation by ruffe (Gymnocephalus cernuus) has been particularly remarkable in some lakes (ADAMS & TIPPETT 1991). POKROVSKII (1961) reported that even 80–90% of vendace eggs were eaten by ruffe in some lakes. The proportion of ruffe in our study area was low (0.6%), thus, the contribution of ruffe in vendace stock regulation is likely negligible.
The effect of intensive fi shing on perch and roach stocks in northern Päijänne was small according to the CPUE of the gillnet test fi shing. This is probably a consequence of the
Table 2. Asymmetrical ANOVA table for analysis of larval vendace density. l = number of target areas, n = number of replicates in target area, t = number of sampling periods (3 year before intensive fi shing and 6 years after that), Bef = before, Aft = after. * = F-value calculated by mean squares (MS) of residuals.
Source of variation df MS F pBefore vs. after B 1 1Years in time period T(B) 2(t-1) 7Between areas L (l-1) 4Target area vs. control areas I 1 1Between control areas C (l-2) 3B × L (l-1) 4 1.20 B × I 1 1 1.35 B × C (l-2) 3 1.15 *0.73 0.532T(B) × L 2(t-1)(l-1) 28 13.39 T(Bef) × L (t-1)(l-1) 8 T(Bef) × I (t-1) 2 6.70 3,03 0.050 T(Bef) × C (t-1)(l-2) 4 9.5 *5.91 < 0.001 T(Aft) × L (t-1)(l-1) 20 T(Aft) × I (t-1) 5 3.40 1.34 0.245 T(Aft) × C (t-1)(l-2) 15 19.27 *12.30 < 0.001Residuals 2lt(n-1) 818 1.57Total 2ltn-1 862

Effects of mass fi sh removal on coregonid larval abundance 393
fi shing method used and the large study area. Despite economic support, fi shers were not able to reach catch levels that could have collapsed the perch and roach stocks. Trap nets were only used in the littoral zone, thus the large pelagic area remained almost untouched. Although the monitoring methods used were not able to detect changes in the fi sh com-munity, a large number of fi sh was removed. It is worth noting that the intensive fi shing on the perch population may also cause compensatory effects leading to strong year-classes (ROMARE & BERGMAN 1999) and/or accelerated growth rates of individual fi sh. Highly abun-dant year-classes of perch may lead to increased predation pressure on coregonid larvae. On the other hand, higher perch growth rates may increase the time that coregonid larvae remain vulnerable to predation. In southwestern Lake Pyhäjärvi, the main predators of vendace lar-vae (10–15 mm TL) are thought to be young perch that are >10 cm (HELMINEN & SARVALA 1994) and juvenile vendace (20–100 mm TL), which are vulnerable to predation by large perch (20–30 cm) (HELMINEN et al. 1997).
Altogether, we conclude that the fi sh removal was insuffi cient to affect the densities of larval coregonids. Despite considerable fi nancial support, fi shermen could not fi sh more intensively in such a large area. Thus, short-term (2–4 years) intensive fi sh removals seem to be an ineffi cient way to restore vendace stocks in large lakes. On the other hand, in northern Päijänne, the spawning stocks of coregonids, especially vendace, have been low for a long period, which is typical for the prolonged recovery of vendace. Low spawning stocks can produce only a relatively low abundance of larvae, which leads to low year-class strength in autumn due to high natural mortality during the fi rst growing season. In some large Finnish lakes, low spawning stocks have been strengthened by transfers of mature adults, but with-out clear success; the vendace stock has not recovered (JURVELIUS et al. 1995, HUUSKONEN et al. 2004). In small Polish lakes (surface area mostly <1000 ha), stockings of newly hatched larvae are widely and successfully used to support vendace stocks that experienced unfavour-able environmental conditions during egg incubation (CJERNIEWSKI & WAWRZYNIAC 2006). In large lakes, larval or juvenile stockings of vendace are rarely applied since the required number of larvae is high (up to 20 000 ind. ha-1) and consequently costs are extensive. Pro-ductive coregonid stocks are necessary for most Finnish inland commercial fi sheries and long low-stock periods threaten the fi sheries. Unfortunately, recent methods to regenerate low vendace stocks have not been successful and local fi shermen have been required to adjust their fi shing behaviors (e.g., they must try to obtain licenses for new lakes and areas, MUJE et al. 2004) for variable coregonid resources.
Acknowledgements
We thank all fi shers who joined the fi sh removal project as well as all persons who helped in larval fi eld sampling over the years. Finally, we are grateful to the anonymous reviewers for valuable comments on the manuscript.

394 O. Urpanen et al.
References
ADAMS, C. E. & TIPPETT, R. (1991): Powan, Coregonus lavaretus (L.), ova predation by newly introduced ruffe, Gymnocephaluus cernuus (L.), in Loch Lomond, Scotland. – Aquacult. Fish. Manage. 22: 239–246.
ALAJA, H. (2008): Effects of fi sh mass removal on fi sh community and fi sh growth in lakes Northern Päijänne and Jyväsjärvi during 2001–2007. – In: SALO, H. (Ed.): Development of lakes Northern Päijänne and Jyväsjärvi – rehabilitation of fi shery. Institute for Environmental Research, University of Jyväskylä. pp. 49–87. [In Finnish].
APPELBERG, M., EKSTRÖM, C. & HÖRNSTRÖM, E. (1990): Lake Stora Härsjön – a case study for integrated monitoring of the effects of liming. – Information från Sötvattenslaboratoriet, Drottningholm 1. [In Swedish with English summary].
BEIER, U. (2001): Habitat distribution and size structure in freshwater fi sh communities: effects of vendace on interactions between perch and roach. – J. Fish Biol. 59: 1437–1454.
CJERNIEWSKI, P. & WAWRZYNIAC, W. (2006): Management of vendace (Coregonus albula (L.)) in the lakes of Northwest Poland in the late twentieth and early twenty-fi rst centuries. – Arch. Pol. Fish. 14: 105–121.
EIE, J. A. & BORGSTRØM, R. (1981): Distribution and food of roach (Rutilus rutilus L.) and perch (Perca fl uviatilis L.) in Lake Årungen, Norway. – Verhandlungen der Internationalen Vereinigung für Limnologie 21: 1257–1263.
HAAKANA, H., HUUSKONEN, H. & VILJANEN, M. (2004): AHTI – Fish Removal of perch in large lakes – processing of the catch and restoration of vendace stock. – Rep. of Karelian Inst., Univ. Joensuu 9/2004: 1–48 [In Finnish].
HAAKANA, H., HUUSKONEN, H. & KARJALAINEN, J. (2007): Predation of perch on vendace larvae: diet composition in an oligotrophic lake and digestion time of the larvae. – J. Fish Biol. 70: 1171–1184.
HAMRIN, S.F. (1986): Vertical distribution and habitat partitioning between different size classes of vendace, Coregonus albula, in thermally stratifi ed lakes. – Can. J. Fish. Aquat. Sci 43: 1617–1625.
HAMRIN, S.F. & PERSSON, L. (1986): Asymmetrical competition between age classes as a factor causing population oscillations in an obligate planktivorous fi sh species. – Oikos 47: 223–232.
HEIKINHEIMO, O. (2001): Effect of predation on the low-density dynamics of vendace: signifi cance of the functional response. – Can. J. Fish. Aquat. Sci. 58: 1909–1923.
HELMINEN, H. & SARVALA, J. (1994): Population regulation of vendace (Coregonus albula) in Lake Pyhäjärvi, southern Finland. – J. Fish Biol. 45: 387–400.
HELMINEN, H., SARVALA, J. & KARJALAINEN, J. (1997): Patterns in vendace recruitment in Lake Pyhäjärvi south-west Finland. – J. Fish Biol. 51: 303–316.
HORPPILA, J. (1994): The diet and growth of roach (Rutilus rutilus (L.)) in Lake Vesijärvi and possible changes in the course of biomanipulation. – Hydrobiologia 294: 35–41.
HORPPILA, J. PELTONEN, H. (1994): The fate of a roach Rutilus rutilus stock under an extremely strong fi shing pressure and its predicted development after the cessation of mass removal. – J. Fish Biol. 45: 777–786.
HUUSKO, A. & SUTELA, T. (1992): Fish predation on vendace (Coregonus albula L.) larvae in Lake Lentua, Northern Finland. – Pol. Arch. Hydrobiol. 39: 381–391.
HUUSKO, A., VUORIMIES, O. & SUTELA, T. (1996). Temperature- and light-mediated predation by perch on vendace larvae. – J. Fish Biol. 49: 441–457.
HUUSKONEN, H., HAAKANA, H. & AHO, T. (2004). Stock transfer in vendace: an evaluation using microsatellite markers. – Ann. Zool. Fennici 41: 69–74.
JAATINEN, R., VUORIMIES, O. & AUVINEN, H. (1999): Perch predation on larval and juvenile vendace in L. Puruvesi (Harvanselkä). Kalatutkimuksia Fiskundersökningar 162: 27–44 [In Finnish].
JEPPESEN, E., MEERHOFF, M., JACOBSEN, B. A., HANSEN, R. S., SØNDERGAARD, M., JENSEN, J. P., LAURIDSEN, T. L., MAZZEO, N. & BRANCO, C. W. C. (2007): Restoration of shallow lakes by nutrient control and biomanipulation—the successful strategy varies with lake size and climate. – Hydrobiologia 581: 269–285.

Effects of mass fi sh removal on coregonid larval abundance 395
JURVELIUS, J., AUVINEN, H., SIKANEN, A., AUVINEN, S. & HEIKKINEN, T. (1995): Stock transfers of vendace (Coregonus albula L.) into Lake Puruvesi, Finland. – In: LUCZYNSKI, M. et al. (Eds.) Biology and Management of Coregonid Fishes. – Arch. Hydrobiol. Spec. Issues Advanc. Limnol. 46: 413–420.
KAHL, U. & RADKE, R. J. (2006): Habitat and food resource use of perch and roach in a deep mesotrophic reservoir: enough space to avoid competition? – Ecol. Freshw. Fish 15: 48–56.
KAIRESALO, T., LAINE, S., LUOKKANEN, E., MALINEN, T. & KETO, J. (1999): Direct and indirect mechanisms behind successful biomanipulation. – Hydrobiologia 395/396: 99–106.
KANGUR, K., KANGUR, A. & KANGUR, P. (2000): Diet composition and food consumption level of ruffe, Gymnocephalus cernuus (L.), in Lake Peipsi. – Proc. Estonian Acad. Sci. Biol. Ecol. 49: 121–135.
KARJALAINEN, J., AUVINEN, H., HELMINEN, H., MARJOMÄKI, T.J., NIVA, T., SARVALA, J. & VILJANEN, M. (2000): Unpredictability of fi sh recruitment: interannual variation in young-of-the-year abundance. – J. Fish Biol. 56: 837–857.
KARJALAINEN, J., HELMINEN, H., HIRVONEN, A., SARVALA, J. & VILJANEN, M. (1992): Identifi cation of vendace (Coregonus albula (L.)) and whitefi sh (Coregonus lavaretus) larvae by the counts of myomeres. – Arch. Hydrobiologia 125:167–173.
KARJALAINEN, J., OLLIKAINEN, S. & VILJANEN, M. (1998): Estimation of the year-class of newly hatched fi sh larvae in Finnish lakes – how sampling design can infl uence abundance estimations? – Arch. Hydrobiol. Spec. Issues Advanc. Limnol. 50: 73–80.
KETO, J. & TALLBERG, P. (2000): The recovery of Vesijärvi, a lake in southern Finland: water quality and phytoplankton interpretations. – Boreal. Env. Res. 5: 15–26.
MARJOMÄKI, T.J., AUVINEN, H., HELMINEN, H., HUUSKO. A., SARVALA. J., VALKEAJÄRVI. P., VILJANEN, M. & KARJALAINEN. J. (2004): Spatial synchrony in the inter-annual population variation of vendace (Coregonus albula (L.)) in Finnish lakes. – Ann. Zool. Fennici 41: 225–240.
MUJE, K., LINDROOS, M., MARJOMÄKI, T. & KARJALAINEN, J. (2004). Interlocked sustainable use of multiple fi sh stocks—modelling biological and socio-economic conditions in Finnish vendace (Coregonus albula (L.)) fi sheries.– Ann. Zool. Fennici 41: 375–390.
POKROVSKII, V. V. (1961): Basic environmental factors determining the abundance of whitefi sh. – Trudy Soveshchanii 13: 228–234.
PERSSON, L., DIEHL, S., JOHANSSON, L., ANDERSSON, G. & HAMRIN, S.F. (1991): Shifts in fi sh communities along the productivity gradient of temperate lakes—patterns and the importance of size–structured interactions. – J. Fish Biol. 38: 281–293.
ROMARE, P. & BERGMAN, E. (1999): Juvenile fi sh expansion following biomanipulation and its effect on zooplankton. – Hydrobiologia 404: 89–97.
SHAPIRO J. & WRIGHT, D.I. (1984). Lake restoration by biomanipulation: Round Lake, Minnesota, the fi rst two years. – Freshwater Biol. 14: 371–383.
SVÄRDSÖN, G. (1976): Interspecifi c population dominance in fi sh communities. – Rep. Inst. Freshw. Res., Drottningholm 55: 144–171.
TOLONEN, M. (2000): Diet and food consumption of perch in L. S Konnevesi. University of Jyväskylä. M.Sc. Thesis 1–56 [In Finnish with English abstract].
UNDERWOOD, A. J. (1992): Beyond BACI: the detection of environmental impacts on populations in the real, but variable, world. – J. Exp. Mar. Biol. Ecol. 161: 145–178.
URPANEN, O., HUUSKONEN, H., MARJOMÄKI, T. J. & KARJALAINEN, J. (2005): Growth and size-selective mortality of vendace (Coregonus albula (L.)) and whitefi sh (C. lavaretus L.) larvae. – Boreal. Env. Res. 10: 225–238.
URPANEN, O., MARJOMÄKI, T. J., VILJANEN, M., HUUSKONEN, H., & KARJALAINEN, J. (2009): Population size estimation of larval coregonids in large lakes: Stratifi ed sampling design with a simple prediction model for vertical distribution. – Fish. Res. 96: 109–117.
VALKEAJÄRVI, P. & MARJOMÄKI, T. (2004): Perch (Perca fl uviatilis) as a factor in recruitment variations of vendace (Coregonus albula) in lake Konnevesi, Finland. – Ann. Zool. Fennici 41: 329–338.
VUORIMIES, O. & TOLONEN, K. (1999): Diet of perch in different habitats of L. Puruvesi. Kalatutkimuksia Fiskundersökningar 162: 1–26 [In Finnish].
WINFIELD, I. J., ADAMS, C. E. & FLETCHER, J. M. (1996): Recent introductions of the ruffe (Gymnocephalus cernuus) to three United Kingdom lakes containing Coregonus species. – Ann. Zool. Fennici. 33: 459–466





















