
Effects of Low pH acute exposure on survival and gill morphology inTriturus italicus larvae
12
Effects of Low pH Acute Exposure on Survival and Gill Morphology in Triturus italicus Larvae ELVIRA BRUNELLI AND SANDRO TRIPEPI Department of Ecology, University of Calabria, Via P. Bucci, I-87036 Rende (Cosenza), Italy ABSTRACT We examined, from a morphological and ultrastructural point of view, the gill epithelium of Triturus italicus, both in basal conditions and after acute exposure to low pH. Our analysis of gill morphology began with the aim of determining lethal pH levels; we found that the pH value at which 50% of mortality occurs (LC50) is 4.0. We then investigated the effects of the larvae’s exposure to a critical value of pH (pH 4.5). No change was observed in the gill during the first 24 hr. After 48 hr, a cellular response was evident and the gills appeared covered with a dense mucous layer. Observations that were carried out by light microscopy (LM) and electron microscopy, both scanning (SEM) and transmission (TEM), showed considerable histological and ultrastructural changes. As regards the principal filament, the alterations resulted in the presence of an external keratinized layer. More changes affected the mitochondria-rich cells (MRCs) in both a qualitative and quantitative way; this cell type shows a wide surface and seems to protrude more than the near pavement cells that, in normal conditions, partially cover the MRCs, thus reducing their external surface. The microvilli were extremely lengthened and often anatomized each other. Changes in the secondary filament affected the thickness of the epithelium, which appeared considerably smaller in the gills of the newts exposed to acid stress. The ciliated cells appeared to be more numerous than in the control specimens and the MRCs showed a widening of the apical surface. J. Exp. Zool. 303A:946–957, 2005. r 2005 Wiley-Liss, Inc. The decline in amphibian populations over the last few years (Barinaga, ’90; Blaustein, ’90; Rowe et al., ’92; Blaustein et al., 2001) has led to an increasing interest in this complex phenomenon. At present, many investigators believe that the decrease in amphibian populations is a conse- quence of human activities, apart from (in addi- tion to) the regular fluctuation in animal populations (Pechman et al., ’91). Many amphibian species reproduce in small ponds; due to their small dimensions and their dependence on rainfall, these environments are particularly sensitive to any changes in abiotic parameters. The acidification of aquatic ecosystems is thought to be one of the possible causes of the decline of amphibians , particularly in Europe and North America (Battarbee and Charles, ’86; Bradford et al., ’92, ’94; Corn and Vertucci, ’92; Wissinger and Whiteman, ’92), and this is a result of acid rainfall (Lenhart and Steinberg, ’84; Battarbee and Charles, ’86; Davis and Stokes, ’86; Bo¨hmher and Rahmann, ’90); this phenom- enon occurs in temporary backwater ponds as well as in larger ponds if the substratum is not rich enough in carbonates (minerals with tamponing characteristics). The known effects of acid pH have been summarized by Pierce (’85) and Freda (’86). Several studies have been carried out concerning the effects of acidity both on embryos (Gosner and Black, ’57; Pough, ’76; Leuven et al., ’86; Andre ´n et al., ’88; Beattie et al., ’92; Grant and Licht, ’93) and on amphibian larvae (Pierce et al., ’84; Gascon and Bider, ’85; Cummins, ’86; Rosenberg and Pierce, ’95); particularly the effects on embryonic and larval development, the survival capacity of larvae after exposure to low pH (Cummins, ’86; Pierce and Wooten, ’92; Rowe et al., ’92; Published online in Wiley InterScience (www.interscience.wiley. com). DOI: 10.1002/jez.a.221. Received 19 April 2005; Accepted 25 July 2005 Abbreviations used: bl, basement lamella; C, capillary; CC, ciliated cell; LC, Leydig cell; m, mitochondria; MRC, mitochondria-rich cell; mv, microvilli; n, nucleus; PC, pavement cell; PF, primary filament; sg, sub-apical granules; SL, secondary lamellae. Correspondence to: Elvira Brunelli, Department of Ecology, University of Calabria, Via P. Bucci, 87036 Rende (Cosenza), Italy. E-mail: [email protected] r 2005 WILEY-LISS, INC. JOURNAL OF EXPERIMENTAL ZOOLOGY 303A:946–957 (2005)
Transcript of Effects of Low pH acute exposure on survival and gill morphology inTriturus italicus larvae

Effects of Low pH Acute Exposure on Survivaland Gill Morphology in Triturus italicus Larvae
ELVIRA BRUNELLI� AND SANDRO TRIPEPIDepartment of Ecology, University of Calabria, Via P. Bucci, I-87036 Rende(Cosenza), Italy
ABSTRACT We examined, from a morphological and ultrastructural point of view, the gillepithelium of Triturus italicus, both in basal conditions and after acute exposure to low pH. Ouranalysis of gill morphology began with the aim of determining lethal pH levels; we found that the pHvalue at which 50% of mortality occurs (LC50) is 4.0. We then investigated the effects of the larvae’sexposure to a critical value of pH (pH 4.5).
No change was observed in the gill during the first 24 hr. After 48 hr, a cellular response wasevident and the gills appeared covered with a dense mucous layer. Observations that were carriedout by light microscopy (LM) and electron microscopy, both scanning (SEM) and transmission(TEM), showed considerable histological and ultrastructural changes.
As regards the principal filament, the alterations resulted in the presence of an externalkeratinized layer. More changes affected the mitochondria-rich cells (MRCs) in both a qualitativeand quantitative way; this cell type shows a wide surface and seems to protrude more than the nearpavement cells that, in normal conditions, partially cover the MRCs, thus reducing their externalsurface. The microvilli were extremely lengthened and often anatomized each other. Changes in thesecondary filament affected the thickness of the epithelium, which appeared considerably smaller inthe gills of the newts exposed to acid stress. The ciliated cells appeared to be more numerous than inthe control specimens and the MRCs showed a widening of the apical surface. J. Exp. Zool.303A:946–957, 2005. r 2005 Wiley-Liss, Inc.
The decline in amphibian populations over thelast few years (Barinaga, ’90; Blaustein, ’90; Roweet al., ’92; Blaustein et al., 2001) has led to anincreasing interest in this complex phenomenon.At present, many investigators believe that thedecrease in amphibian populations is a conse-quence of human activities, apart from (in addi-tion to) the regular fluctuation in animalpopulations (Pechman et al., ’91).
Many amphibian species reproduce in smallponds; due to their small dimensions and theirdependence on rainfall, these environments areparticularly sensitive to any changes in abioticparameters.
The acidification of aquatic ecosystems isthought to be one of the possible causes of thedecline of amphibians , particularly in Europe andNorth America (Battarbee and Charles, ’86;Bradford et al., ’92, ’94; Corn and Vertucci, ’92;Wissinger and Whiteman, ’92), and this is a resultof acid rainfall (Lenhart and Steinberg, ’84;Battarbee and Charles, ’86; Davis and Stokes,’86; Bohmher and Rahmann, ’90); this phenom-enon occurs in temporary backwater ponds as well
as in larger ponds if the substratum is not richenough in carbonates (minerals with tamponingcharacteristics).
The known effects of acid pH have beensummarized by Pierce (’85) and Freda (’86).Several studies have been carried out concerningthe effects of acidity both on embryos (Gosner andBlack, ’57; Pough, ’76; Leuven et al., ’86; Andrenet al., ’88; Beattie et al., ’92; Grant and Licht, ’93)and on amphibian larvae (Pierce et al., ’84; Gasconand Bider, ’85; Cummins, ’86; Rosenberg andPierce, ’95); particularly the effects on embryonicand larval development, the survival capacityof larvae after exposure to low pH (Cummins,’86; Pierce and Wooten, ’92; Rowe et al., ’92;
Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/jez.a.221.
Received 19 April 2005; Accepted 25 July 2005
Abbreviations used: bl, basement lamella; C, capillary; CC,ciliated cell; LC, Leydig cell; m, mitochondria; MRC, mitochondria-richcell; mv, microvilli; n, nucleus; PC, pavement cell; PF, primaryfilament; sg, sub-apical granules; SL, secondary lamellae.�Correspondence to: Elvira Brunelli, Department of Ecology,
University of Calabria, Via P. Bucci, 87036 Rende (Cosenza), Italy.E-mail: [email protected]
r 2005 WILEY-LISS, INC.
JOURNAL OF EXPERIMENTAL ZOOLOGY 303A:946–957 (2005)

Grant and Licht, ’93; Griffiths and De Wijer, ’94;Whiteman et al., ’95), the effects of acidity onswimming capability (Kutka, ’94), on the preda-tory skill (Griffiths, ’93), and on the choice ofreproductive sites (Horne and Dunson, ’94) havebeen studied in detail.
There may be different mechanisms throughwhich the toxicity of an acid pH can act onamphibian larvae; one of the main consequencesis an ionic equilibrium alteration (Freda andDunson, ’84; McDonald et al., ’84; Rowe et al., ’92).
In amphibian larvae, the gills perform animportant role in osmoregulation and in themaintenance of acid–base equilibrium. Histo-chemical and cytochemical studies have shownthe presence of Na1-K1-ATPase in the basolateralmembrane of the pavement cells, whereas carbo-nic anhydrase has been found in the mitochon-dria-rich cell (MRC) (Lewinson et al., ’82; Katoand Kurihara, ’88; Warburg et al., ’94).
Many studies have been published on the effectsof acidification on amphibians but only a few haveconsidered gill morphology and ultrastructure(Linnenbach et al., ’87; Bohmher and Rahmann,’90), and no previous study has examined theeffects of low pH on the MRC and on thekeratinization pattern of the epithelium.
The aim of this study was to give a structuraland ultrastructural description of the gill appara-tus of Triturus italicus larvae (about which wehave previously described only the structure andthe ultrastructure of the main filament (Tripepiand Brunelli, 2000)) and to evaluate the effects ofexposure to a nonlethal value of acid pH on thisorgan that is actively involved in acid–baseregulation. Furthermore, the experiments carriedout are intended to contribute to the discussionregarding the survival of Amphibia by evaluatingtolerance to low pH in T. italicus larvae, endemicspecies to peninsular Italy.
MATERIALS AND METHODS
Maintenance of larvae and survival study
All the larvae used in the study were collectedfrom a pond in the vicinity of Cosenza (Calabria,Southern Italy), which had a pH of 7.2–7.5 duringthe breeding season. The larvae were caught bymeans of a close-mesh net, transported to thelaboratory, and kept in aquaria of 50 l filled withwater from the original pond (Table 1) for a 5-dayacclimatization period.
The developmental stage was determined by thepresence of certain distinctive morphological
features and by using appropriate developmentalchronological tables (Gallien and Bidaud, ’59).Forty larvae at the stage 53–55a of Gallien andBidaud and with a mean mass of 1.570.2 g wereselected for the experiment. They were kept undera natural light/dark cycle and fed with Artemiasalina (after thorough washing in fresh water) onalternate days for the entire duration of theexperiment. All the aquaria were kept at roomtemperature.
After the acclimatization period, groups of eightlarvae each were moved to specially preparedaquaria containing either natural pond water orpond water acidified with the addition of HCl indrops, respectively. The pH level was measuredwith a DeltaOHM HD8705 pH meter with auto-matic temperature compensation, and measure-ments were taken every 4 hr during the day andevery 8 hr during the night. The pH value wasregulated by adding water or HCl as required,thereby excluding any pH buffering due to thewater ion composition.
The first step in the experiment was to deter-mine the lethal level of pH by exposing the larvaeto decreasing pH values.
LC50 (the condition at which 50% of the speci-mens are expected to die) was determined bygraphic analysis of survival rates after 96 hr usingthe method of Della Croce et al. (2001).
All the control experiments were carried out onanimals of the same origin and at the samedevelopmental stage, bred simultaneously in non-treated water. All the aquaria were subjected tocontinuous monitoring during the day and mon-
TABLE 1. Analysis of test water taken from a natural pondin the vicinity of Cosenza (Calabria, Southern Italy)—analysis
was performed according to APAT IRSA-CNR (2003)
Parameter Values1
Conductivity 679 mspH 7.44Cl� 11.60 mg/lSO4
2� 121.22 mg/lCa21 132.21 mg/lMg21 12.36 mg/lNa1 16.72 mg/lK1 2.52 mg/lAl31 44 mg/lFe21 20 mg/lCu11 7 mg/lAlkalinity 230 mg/l CaCO3
1At 18.11C.
LOW pH EFFECTS ON TRITURUS ITALICUS GILLS 947

itoring every 4 hr during the night; in this way,feeding frequency and any anomalous swimmingbehavior were annotated.
To test swimming activity in both the controland the pH 4-exposed group, four 60-min observa-tion periods took place. During each of theseperiods the aquaria were monitored using aPanasonic NV-GX7 videocamera; any kind ofmotion every 60 sec was considered and scored asmoving or not moving.
Concerning feeding behavior, we evaluated thefrequency of food intake only by counting thenumber of attempts that the larvae made to feedduring the feeding interval; during this period,four 1-hr observations were made in which feedingbehaviors were estimated and reported for bothtreatment (pH 4.5) and control groups.
To compare variations in behavioral observa-tions, the two groups (control sample and pH 4.5sample) were evaluated using the statisticalprogram InStat (GraphPad) and by applying theMann–Whitney U-test for unpaired data, withPo0.05 as the fiducial limit. The data arepresented as mean7s.e.m.
Morphological analysis of the gillapparatus
The removal of the gill occurred after 24 and48 hr exposure. The animals were anesthetizedwith tricaine methane sulfonate—MS 222 (San-doz, Sigma, St. Louis, MO) and decapitated.Animals were handled according to the recom-mendations of the Ethical Committee and underthe supervision of authorized investigators.
Light and electron microscopy
The removal and dissection phases were per-formed while the specimens were kept constantlycovered with a fixative solution of glutaraldehydein phosphate buffer. After removal, specimensundergoing light microscopy (LM) and transmis-sion electron microscopy (TEM) were subjected toaldehyde fixation (glutaraldehyde at 3% in phos-phate buffer, 0.05 M, pH 7.5) and post-fixation inosmium tetroxide (1% in the same buffer); thenthey were dehydrated in graded ethanol soaked inpropylene-oxide and embedded in Epon-Araldite.
Semithin sections with a thickness of 1–2 mmwere stained using the technique described byHumphrey and Pittman (methylene blue, azure II,and basic fuchsine), observed, and photographedby an LM LEITZ Dialux EB 20.
Ultrathin sections (600–900 A) were treated withuranyl acetate and lead citrate and coated inEdwards EM 400. The observations were carriedout by a TEM ZEISS EM 900.
Specimens undergoing scanning electron micro-scopy (SEM) were dehydrated and dried withhexamethyldisilazane, according to the methodsreported by Nation (’83), put on the appropriatestubs, covered with gold, and observed by SEMCambridge Stereoscan 360.
Immunohistochemistry
After fixation in Bouin liquid for 24 hr, thespecimens were dehydrated and placed in paraffin.Paraffined sections of 10 mm were used for theimmunolabeling, which was performed follow-ing the indirect immunofluorescence technique(Coons et al., ’55).
The sections were treated for 20 min with 2%H2O2 in 0.1 phosphate buffer solution (PBS) toinhibit the endogenous peroxidases and thenincubated for 30 min in a moist chamber with20% normal goat serum to block nonspecific sites.Unwashed sections were then incubated overnightat 41C with a mouse monoclonal antibody toCytokeratin Type II at working dilutions of 1:100(Clone Determination AE3—Class IgG 1 IgGa5;Progen-Biotechnik Heidelberg). The AE3 antibodyrecognizes all members of the basic cytokeratinsubfamily and reacts with cells of epithelial origin,including simple and stratified epithelia andepidermis.
After washings in PBS, fluorescein-isothiocyanate-conjugated g-globulin goat anti-mouse (diluted1:200 Sigma) was used as the second antiserum ata dilution of 1:200 for 1 hr at room temperature.Immunolabeling specificity was verified by sub-stituting the primary antiserum with nonimmunegoat serum or with PBS. The observations werecarried out on a Confocal Laser Scanning Micro-scope Leica TCS SP2.
RESULTS
Survival
We calculated that the pH value at which 50%of mortality occurs in 96 hr (LC50) is 4.0 forT. italicus larvae. No mortality occurred in thecontrol group or in the sublethal group. Within afew hours after being put in water with a pH valueof 4.0, the larvae showed behavior anomalies:reduced swimming activity, low reactivity to
E. BRUNELLI AND S. TRIPEPI948
stimuli, and decrease in feeding (results notshown).
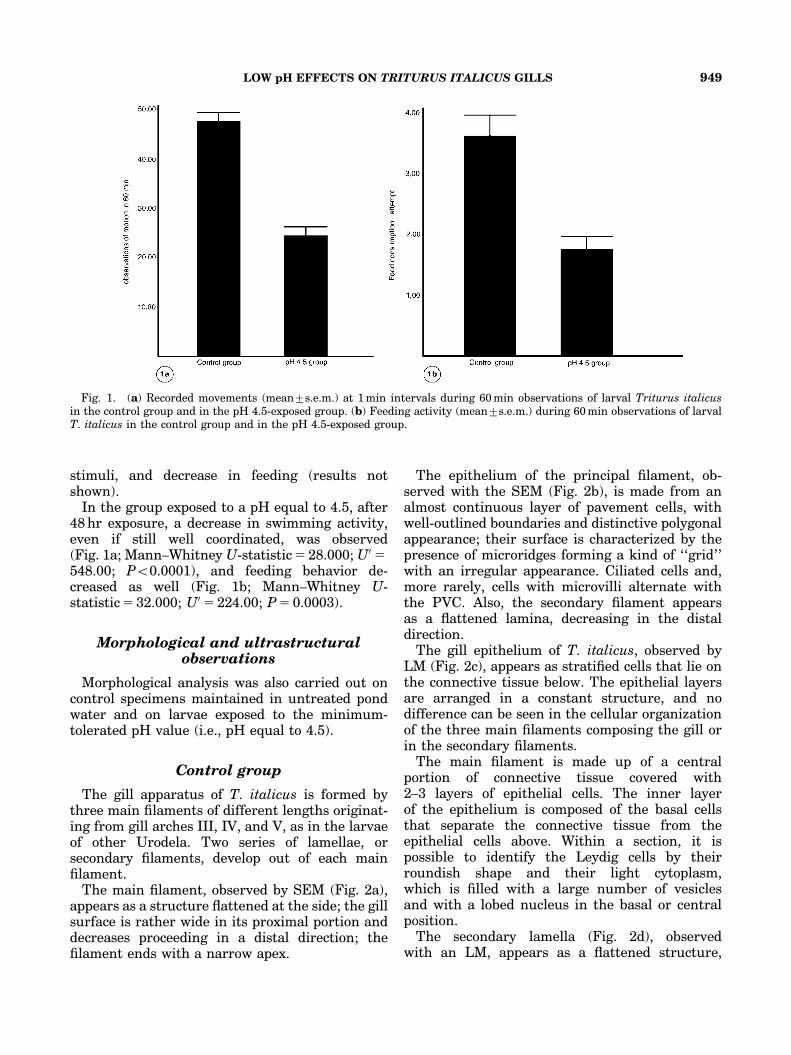
In the group exposed to a pH equal to 4.5, after48 hr exposure, a decrease in swimming activity,even if still well coordinated, was observed(Fig. 1a; Mann–Whitney U-statistic 5 28.000; U05548.00; Po0.0001), and feeding behavior de-creased as well (Fig. 1b; Mann–Whitney U-statistic 5 32.000; U05 224.00; P 5 0.0003).
Morphological and ultrastructuralobservations
Morphological analysis was also carried out oncontrol specimens maintained in untreated pondwater and on larvae exposed to the minimum-tolerated pH value (i.e., pH equal to 4.5).
Control group
The gill apparatus of T. italicus is formed bythree main filaments of different lengths originat-ing from gill arches III, IV, and V, as in the larvaeof other Urodela. Two series of lamellae, orsecondary filaments, develop out of each mainfilament.
The main filament, observed by SEM (Fig. 2a),appears as a structure flattened at the side; the gillsurface is rather wide in its proximal portion anddecreases proceeding in a distal direction; thefilament ends with a narrow apex.
The epithelium of the principal filament, ob-served with the SEM (Fig. 2b), is made from analmost continuous layer of pavement cells, withwell-outlined boundaries and distinctive polygonalappearance; their surface is characterized by thepresence of microridges forming a kind of ‘‘grid’’with an irregular appearance. Ciliated cells and,more rarely, cells with microvilli alternate withthe PVC. Also, the secondary filament appearsas a flattened lamina, decreasing in the distaldirection.
The gill epithelium of T. italicus, observed byLM (Fig. 2c), appears as stratified cells that lie onthe connective tissue below. The epithelial layersare arranged in a constant structure, and nodifference can be seen in the cellular organizationof the three main filaments composing the gill orin the secondary filaments.
The main filament is made up of a centralportion of connective tissue covered with2–3 layers of epithelial cells. The inner layerof the epithelium is composed of the basal cellsthat separate the connective tissue from theepithelial cells above. Within a section, it ispossible to identify the Leydig cells by theirroundish shape and their light cytoplasm,which is filled with a large number of vesiclesand with a lobed nucleus in the basal or centralposition.
The secondary lamella (Fig. 2d), observedwith an LM, appears as a flattened structure,
Fig. 1. (a) Recorded movements (mean7s.e.m.) at 1 min intervals during 60 min observations of larval Triturus italicusin the control group and in the pH 4.5-exposed group. (b) Feeding activity (mean7s.e.m.) during 60 min observations of larvalT. italicus in the control group and in the pH 4.5-exposed group.
LOW pH EFFECTS ON TRITURUS ITALICUS GILLS 949
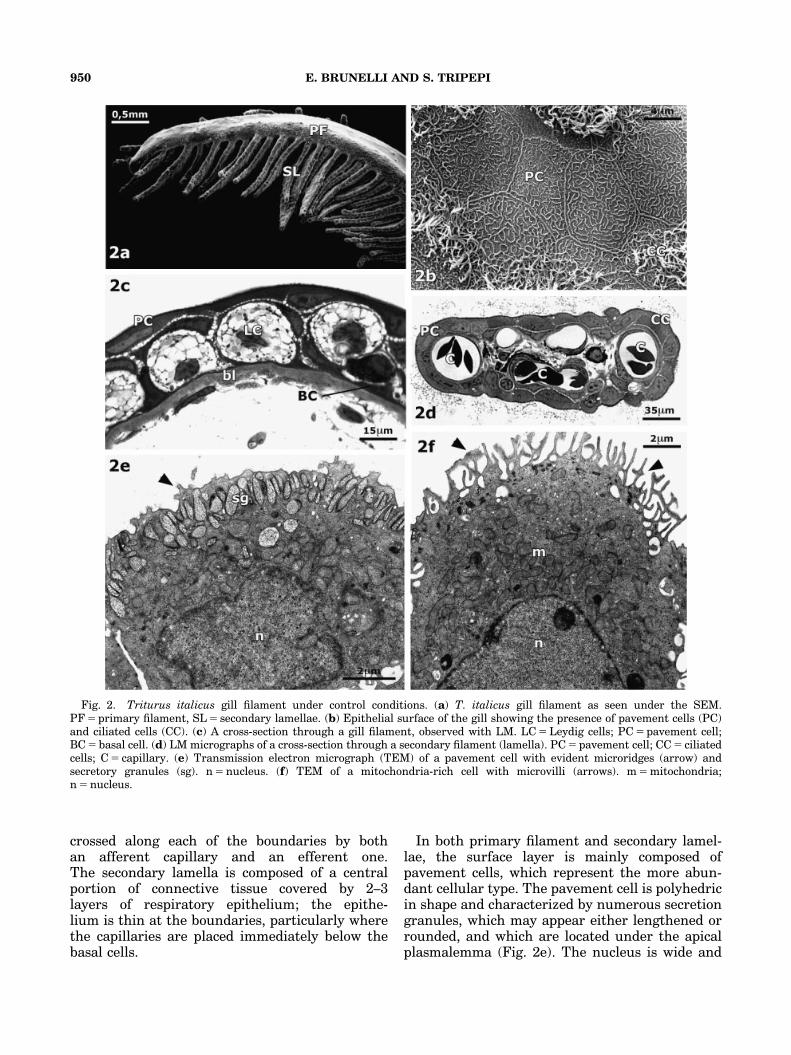
crossed along each of the boundaries by bothan afferent capillary and an efferent one.The secondary lamella is composed of a centralportion of connective tissue covered by 2–3layers of respiratory epithelium; the epithe-lium is thin at the boundaries, particularly wherethe capillaries are placed immediately below thebasal cells.
In both primary filament and secondary lamel-lae, the surface layer is mainly composed ofpavement cells, which represent the more abun-dant cellular type. The pavement cell is polyhedricin shape and characterized by numerous secretiongranules, which may appear either lengthened orrounded, and which are located under the apicalplasmalemma (Fig. 2e). The nucleus is wide and
Fig. 2. Triturus italicus gill filament under control conditions. (a) T. italicus gill filament as seen under the SEM.PF 5 primary filament, SL 5 secondary lamellae. (b) Epithelial surface of the gill showing the presence of pavement cells (PC)and ciliated cells (CC). (c) A cross-section through a gill filament, observed with LM. LC 5 Leydig cells; PC 5 pavement cell;BC 5 basal cell. (d) LM micrographs of a cross-section through a secondary filament (lamella). PC 5 pavement cell; CC 5 ciliatedcells; C 5 capillary. (e) Transmission electron micrograph (TEM) of a pavement cell with evident microridges (arrow) andsecretory granules (sg). n 5 nucleus. (f) TEM of a mitochondria-rich cell with microvilli (arrows). m 5 mitochondria;n 5 nucleus.
E. BRUNELLI AND S. TRIPEPI950

sometimes lobed. The cytoplasm contains a well-developed Golgi apparatus, some mitochondria,and often an abundant rough endoplasmic reticu-lum; observed by TEM, the microridges appear asa short protrusion of the apical membrane and areoften covered with a thin mucous layer.
When observed by TEM, the MRC (Fig. 2f)shows a typical pear shape, and there arenumerous apical microvilli on its surface. Themost notable feature of these cells is that theycontain a large number of mitochondria that fillthe whole cytoplasm. The nucleus is wide, round-ish, and usually in the basal position. Connectionsamong adjacent cells are performed by tightjunctions and desmosomes.
pH-exposed group
After 24 hr exposure, our analysis did not showany modification in the gill apparatus, and nodifference, on either the structural or ultrastruc-tural level, resulted between control specimensand treated ones. On the contrary, macroscopicmodifications occurred after 48 hr exposure; whenobserved by a stereoscopic microscope, the gillsappeared covered with a dense mucous layer.
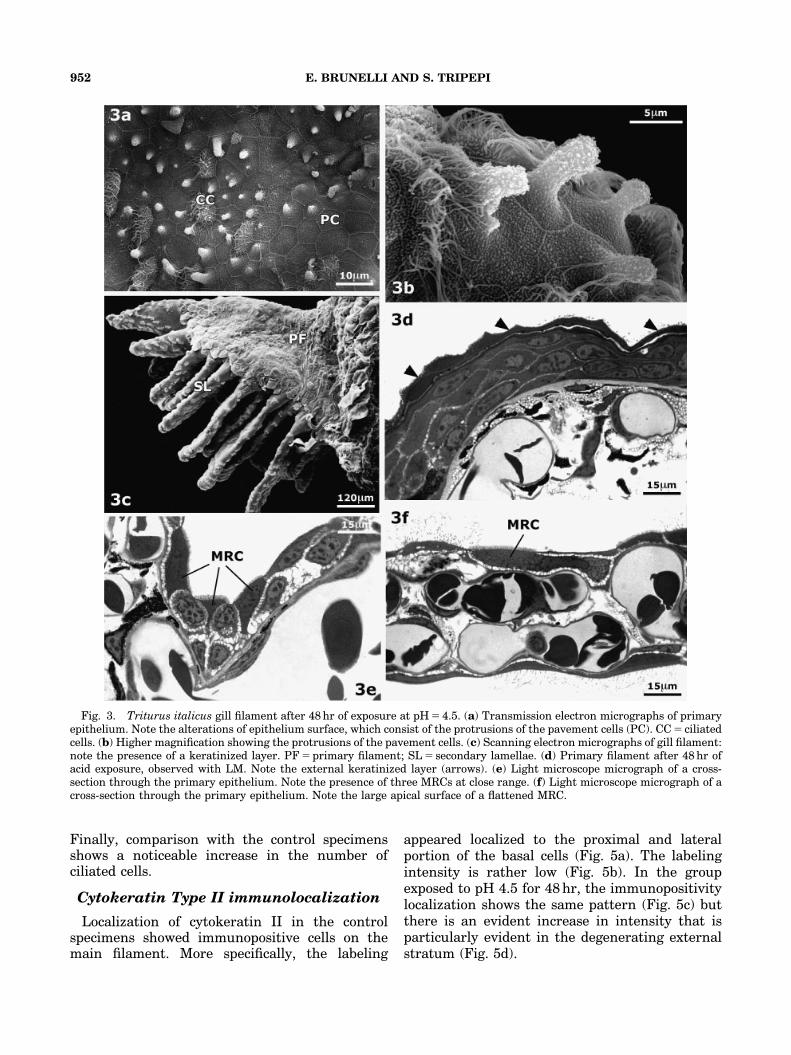
Observed by SEM, after removal of the mucus,the gill shows frequent superficial irregularitiesoriginating from the pavement cells (Fig. 3a).These cells keep their polygonal shape and areinterposed with numerous ciliated cells. Irregula-rities are very frequent and affect both the threemain filaments and, to a minor extent, thesecondary filaments.
At high magnification (Fig. 3b), the irregula-rities appear as electron-clear digitiform protru-sions departing from the central region of thePVC; the peripheral surface keeps its own organi-zation and junctional margins and microridges canbe seen. Observing the cellular protrusions, andparticularly their terminal portion, it can be seenthat the microridges start losing their typicalorganization to assume a flocculent appearance.
In other cases (Fig. 3c), one can observe akeratinized and irregular cellular layer uniformlycovering the cells below.
This structure appears at the level of the mainfilament and particularly on the first of the three gillfilaments, and never reaches the respiratory lamel-lae. Observations with an LM (Fig. 3d) show thisclearly because this layer, strongly stained, coversnearly the whole surface of the main filament. Inmore than one area it is possible to see thesekeratinized cells constituting this layer and the
presence of a flocculent substance on its surface; theepithelium below appears thinned and flattenedcompared with that of the control specimens.
Further ultrastructural alterations involve thecellular types that reach the surface but are notdirectly involved in keratinization. In particular,noticeable qualitative and quantitative modifica-tions can be seen in the MRC. These are muchmore numerous than under normal conditions;two or more of them often appear close to eachother (Fig. 3e).
Moreover, these cells show a wide surface areaand appear to protrude in relation to the adjacentpavement cells, which under normal conditionspartially cover the MRC, thus reducing theirexternal surface.
No alteration has been found regarding theLeydig cells and club-shaped cells.
In the secondary filament, keratinization phe-nomena are less frequent and are limited to theproximal portion; therefore, we cannot see acontinuous keratinized layer similar to that foundin the main filament. The decrease in epitheliumthickness is particularly evident in the secondaryfilaments (Fig. 3f). We also observed an increase inthe surface of the vessels, which almost entirelyfilled the central portion of the connective tissue.
The ciliated cells appear to be more numerousthan usual, and the MRCs show a widening of theapical surface, as occurs in the main filament.
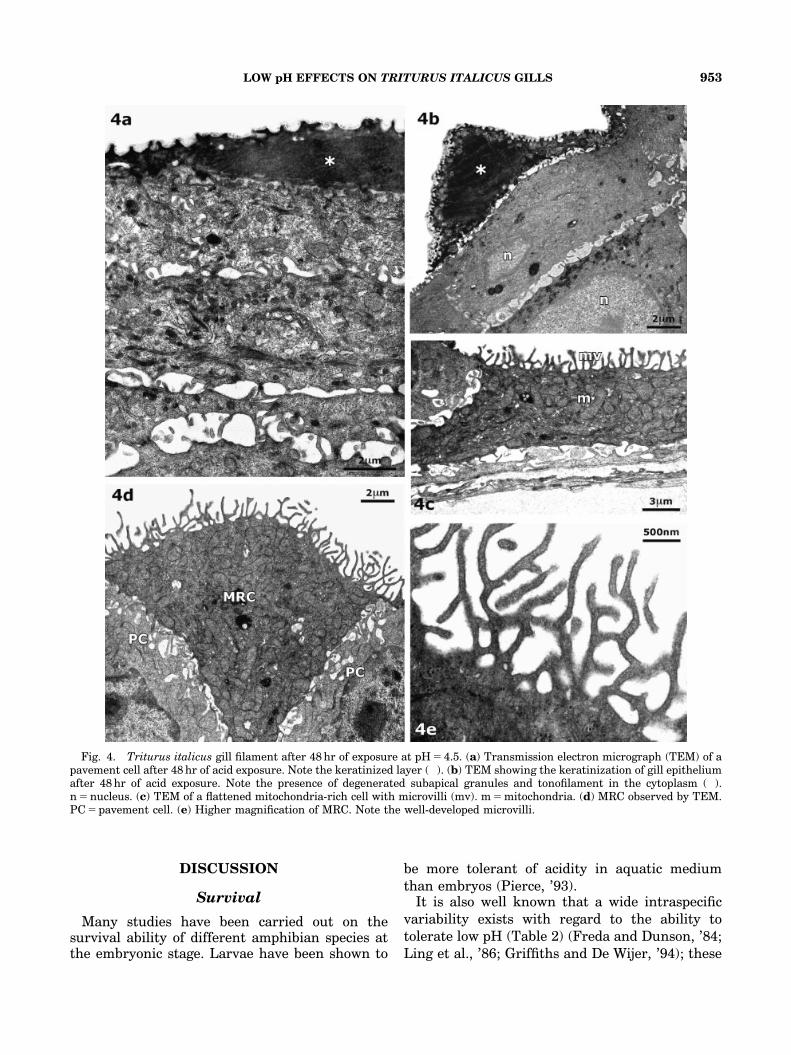
To confirm the cellular nature of the keratinizedlayer, some sections were taken from the mainfilament (Figs. 4a and b) and observed by TEM;it was possible to recognize subapical secretiongranules, strongly degenerated, and the profile ofthe microridges which were covered with mucoussubstance. The cytoplasm appeared completelyfilled with electron-dense material.
The central portion is mainly composed oftonofilaments joined to form an amorphous mass.The keratinized structure tends to grow awayfrom the epithelial strata below which are com-posed of pavement cells; these cells always appearrather lengthened, and it is possible to detect onlya few short microridges and small secretiongranules. Mitochondria and bundles of tonofila-ments are present in the cytoplasm (Fig. 4a). TheMRCs (Figs. 4c and d) always show a wide surfaceand often assume a lengthened and flattenedshape (Fig. 4e); it is always possible to detect thenumerous mitochondria that almost entirely fillthe cytoplasm.
The microvilli (Fig. 4d) appear extremely length-ened and often anatomized each other (Fig. 4f).
LOW pH EFFECTS ON TRITURUS ITALICUS GILLS 951
Finally, comparison with the control specimensshows a noticeable increase in the number ofciliated cells.
Cytokeratin Type II immunolocalization
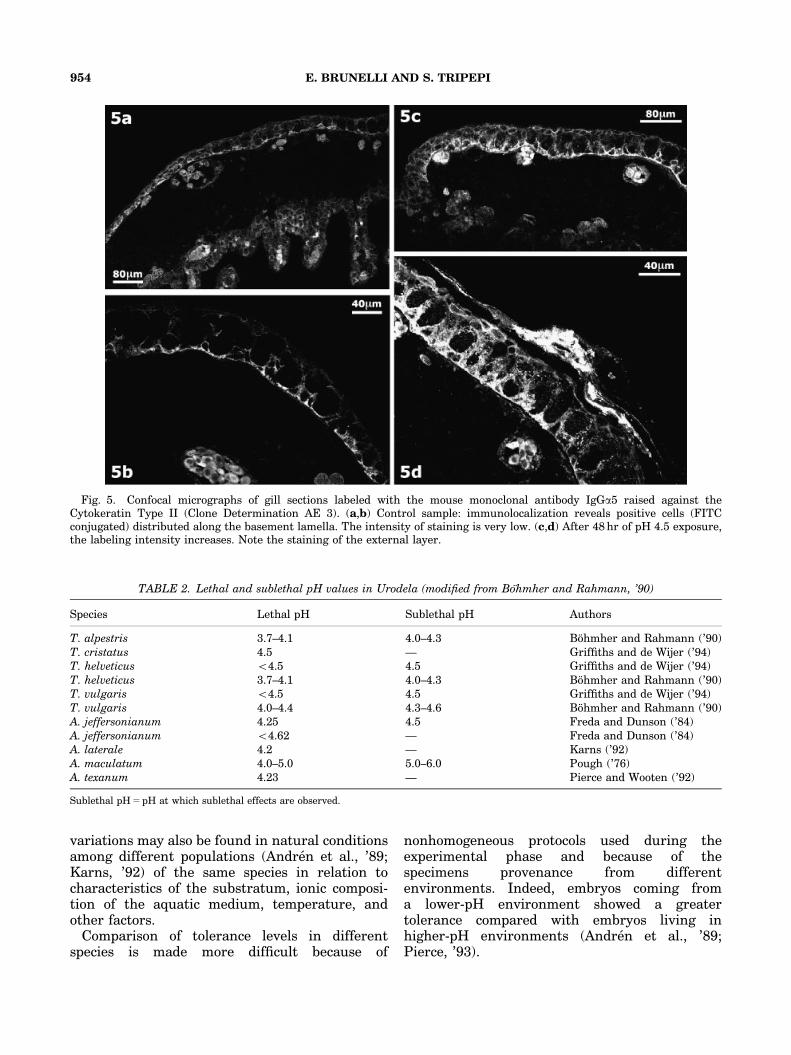
Localization of cytokeratin II in the controlspecimens showed immunopositive cells on themain filament. More specifically, the labeling
appeared localized to the proximal and lateralportion of the basal cells (Fig. 5a). The labelingintensity is rather low (Fig. 5b). In the groupexposed to pH 4.5 for 48 hr, the immunopositivitylocalization shows the same pattern (Fig. 5c) butthere is an evident increase in intensity that isparticularly evident in the degenerating externalstratum (Fig. 5d).
Fig. 3. Triturus italicus gill filament after 48 hr of exposure at pH 5 4.5. (a) Transmission electron micrographs of primaryepithelium. Note the alterations of epithelium surface, which consist of the protrusions of the pavement cells (PC). CC 5 ciliatedcells. (b) Higher magnification showing the protrusions of the pavement cells. (c) Scanning electron micrographs of gill filament:note the presence of a keratinized layer. PF 5 primary filament; SL 5 secondary lamellae. (d) Primary filament after 48 hr ofacid exposure, observed with LM. Note the external keratinized layer (arrows). (e) Light microscope micrograph of a cross-section through the primary epithelium. Note the presence of three MRCs at close range. (f) Light microscope micrograph of across-section through the primary epithelium. Note the large apical surface of a flattened MRC.
E. BRUNELLI AND S. TRIPEPI952
DISCUSSION
Survival
Many studies have been carried out on thesurvival ability of different amphibian species atthe embryonic stage. Larvae have been shown to
be more tolerant of acidity in aquatic mediumthan embryos (Pierce, ’93).
It is also well known that a wide intraspecificvariability exists with regard to the ability totolerate low pH (Table 2) (Freda and Dunson, ’84;Ling et al., ’86; Griffiths and De Wijer, ’94); these
Fig. 4. Triturus italicus gill filament after 48 hr of exposure at pH 5 4.5. (a) Transmission electron micrograph (TEM) of apavement cell after 48 hr of acid exposure. Note the keratinized layer (�). (b) TEM showing the keratinization of gill epitheliumafter 48 hr of acid exposure. Note the presence of degenerated subapical granules and tonofilament in the cytoplasm (�).n 5 nucleus. (c) TEM of a flattened mitochondria-rich cell with microvilli (mv). m 5 mitochondria. (d) MRC observed by TEM.PC 5 pavement cell. (e) Higher magnification of MRC. Note the well-developed microvilli.
LOW pH EFFECTS ON TRITURUS ITALICUS GILLS 953
variations may also be found in natural conditionsamong different populations (Andren et al., ’89;Karns, ’92) of the same species in relation tocharacteristics of the substratum, ionic composi-tion of the aquatic medium, temperature, andother factors.
Comparison of tolerance levels in differentspecies is made more difficult because of
nonhomogeneous protocols used during theexperimental phase and because of thespecimens provenance from differentenvironments. Indeed, embryos coming froma lower-pH environment showed a greatertolerance compared with embryos living inhigher-pH environments (Andren et al., ’89;Pierce, ’93).
Fig. 5. Confocal micrographs of gill sections labeled with the mouse monoclonal antibody IgGa5 raised against theCytokeratin Type II (Clone Determination AE 3). (a,b) Control sample: immunolocalization reveals positive cells (FITCconjugated) distributed along the basement lamella. The intensity of staining is very low. (c,d) After 48 hr of pH 4.5 exposure,the labeling intensity increases. Note the staining of the external layer.
TABLE 2. Lethal and sublethal pH values in Urodela (modified from Bohmher and Rahmann, ’90)
Species Lethal pH Sublethal pH Authors
T. alpestris 3.7–4.1 4.0–4.3 Bohmher and Rahmann (’90)T. cristatus 4.5 — Griffiths and de Wijer (’94)T. helveticus o4.5 4.5 Griffiths and de Wijer (’94)T. helveticus 3.7–4.1 4.0–4.3 Bohmher and Rahmann (’90)T. vulgaris o4.5 4.5 Griffiths and de Wijer (’94)T. vulgaris 4.0–4.4 4.3–4.6 Bohmher and Rahmann (’90)A. jeffersonianum 4.25 4.5 Freda and Dunson (’84)A. jeffersonianum o4.62 — Freda and Dunson (’84)A. laterale 4.2 — Karns (’92)A. maculatum 4.0–5.0 5.0–6.0 Pough (’76)A. texanum 4.23 — Pierce and Wooten (’92)
Sublethal pH 5 pH at which sublethal effects are observed.
E. BRUNELLI AND S. TRIPEPI954

Nevertheless, the lethal pH value found for thelarvae of T. italicus has been observed to becomparable with that found for embryos of otherspecies belonging to the same genus (Table 2).
According to Griffths (’93), we think that acidityeffects are both type-specific and stage-specific.Moreover, Griffiths (’93) pointed out in an inhi-bition of feeding activity T. vulgaris andT. helveticus after exposure to a pH value equalto 4.5. The same author supposes that thefailure in feeding could be due, as noted in fishes,to alterations in the chemosensory system. Wehave found, at the same pH, anomalies in thefeeding behavior of T. italicus; further studieson the chemosensory system could confirmthese hypothesis.
The hypoactivity found in the larvae ofT. italicus can be seen in other urodeles afterintense exposure to acid pH (Freda and Taylor,’92; Kutka, ’94), and this can be correlated withthe decrease in food search and capture activities.
Morphological changes in the gillapparatus
Examination of T. italicus larvae after a 48 hrperiod of exposure to a pH value of 4.5 showedremarkable histological changes in both the mainfilament and the secondary filament. The presenceof a mucous secretion covering the whole gillsurface has already been observed, regardingfishes, in the gills of Salmo trutta exposed to acidstress (Freda and Taylor, ’92; Kutka, ’94).
This secretion is also released from the vacuolesof the pavement cells under normal conditions(Kato and Kurihara, ’88), and its function isprobably to create a mucous coating to facilitatethe exchange of gases.
Alterations affecting the principal filament con-sist mainly in the presence of a keratinizedexternal layer. This phenomenon, confirmed byimmunolocalization of the Cytokeratin Type II,can be seen in the epidermis, as well. Therefore,similar to what happens in the epidermis duringmetamorphosis (Fox, ’86), it appears that the cellsof the external layer degenerate, leaving a residualstructure. The histological and ultrastructuralchanges observed can interfere with respiratoryand osmoregulation processes.
The thinning of the epithelium and the in-creased dimensions of the vasal component seemto indicate the need to facilitate these processes,which are probably complicated by the presence ofthe external keratinized layer. These phenomena
are also evident in the secondary filaments, whichact as the main sites of respiratory exchanges.
The numerical increase of the MRCs, the greaterextension of their apical surface, and the presenceof extremely lengthened microvilli could be anadaptive response of the gill epithelium to osmoticand ionic stress. At low pH, in fact, the perme-ability of the ‘‘tight junctions’’ increases, inducingan ionic loss through the so-called paracellularpathways; this is probably due to the fact that theprotons remove the calcium from negativelycharged glycoproteins that seal the ‘‘tight junc-tions’’ (Ferreira and Hill, ’82).
In fishes and amphibians, a high sensitivity to anacid pH is associated with a lower survival timeand a greater loss of whole-body ions (Bohmherand Rahmann, ’90). Freda and McDonald (’88)indicate Na efflux as the most important factor.Furthermore, other factors should be considered:in particular, aluminum, zinc, and cadmiumconcentrations present in the sediments (Linnen-bach et al., ’87). In fact, elevated but still lowconcentrations of cations (H1, Al31, Na1) increasethe permeability of Na channel, whereas higherAl levels induce a decrease in the uptake of Na(Wood and McDonald, ’87).
The role performed by MRCs in acid–baseregulation could explain the need for a greaterexchange surface with the purpose of increasingthe ionic transport rate through the apicalmembrane.
Changes in the osmoregulation process relatedto the morphological modifications shown in thegill epithelium could represent an importantfactor in tracking the decline of amphibianpopulations in acidified regions.
ACKNOWLEDGMENTS
This work was supported by MemoBioamarMURST Project. We thank E. Perrotta andF. Cofone for technical assistance.
LITERATURE CITED
Andren C, Henrikson L, Olsson M, Nilson G. 1988. Effect ofpH and aluminium on embryonic and early larval stages ofthe Swedish brown frogs Rana arvalis, Rana temporariaand Rana dalmatina. Holarctic Ecol 11:127–135.
Andren C, Marden M, Nilson G. 1989. Tolerance of low pH in apopulation of moor frogs, Rana arvalis, from an acid andneutral environment: a possible case of rapid evolutionaryresponse to acidification. Oikos 56:215–223.
APAT IRSA-CNR. 2003. Metodi analitici per le acque,Vol. 29/2003, IGER Ed., Roma.
Barinaga M. 1990. Where have all the froggies gone?. Science247:1033–1034.
LOW pH EFFECTS ON TRITURUS ITALICUS GILLS 955

Battarbee RW, Charles DF. 1986. Diatom-based pH recon-struction studies of acidic lakes in Europe and NorthAmerica: a synthesis. Water Air Soil Pollut 18:289–309.
Beattie RC, Tyler-Jones R, Baxter MJ. 1992. The effects ofpH, aluminum concentration and temperature on theembryonic develoment of the European common frog, Ranatemporaria. J Zool Lond 228:557–570.
Blaustein AR. 1990. Declining amphibian populations a globalphenomenon? Bull Ecol Soc Am 71:127–128.
Blaustein AR, Wildy EL, Belden LK, Hatch A. 2001. Influenceof abiotic and biotic factors on amphibians in ephemeralponds with special reference to long-toed salamanders(Ambystoma macrodactylum). Israel J Zool 47:333–345.
Bohmer J, Rahmann H. 1990. Influence of water acidificationon amphibians. Fortschr Zool 38:287–309.
Bradford DF, Swanson C, Gordon MS. 1992. Effects of low pHand aluminium on two declining species of amphibians inthe Sierra Nevada, California. J Herpetol 26:369–377.
Bradford DF, Gordon MS, Johnson DF, Andrews RD,Jennings WB. 1994. Acidic deposition as an unlikely causefor amphibian population declines in the Sierra Nevada,California. Biol Conserv 69:155–161.
Coons AH, Leduc EH, Connolly JM. 1955. Studies onantibody. I. A method for the histochemical demonstrationof specific antibody and its application to a study of thehyperimmune rabbit. J Exp Med 102:49–59.
Corn PS, Vertucci FA. 1992. Descriptive risk assessment ofthe effects of acid deposition on Rocky Mountain amphi-bians. J Herpetol 26:361–369.
Cummins CP. 1986. Effects of aluminium and low pHon growth and development in Rana temporaria tadpoles.Oecologia 69:248–252.
Davis RB, Stockes PM. 1986. Overview of historical andpalaeoecological studies of acidic air pollution and its effects.Water Air Soil Pollut 30:311–318.
Della Croce N, Cattaneo, Vietti R, Danovaro R. 2001. Ecologiae protezione dell’ambiente marino costiero, UTET Ed.,Torino.
Ferreira KTG, Hill BS. 1982. The effect of low external pH onproperties of the paracellular pathway and junctionalstructure on isolated frog skin. J Physiol 332:59–67.
Fox H. 1986. Changes in amphibian skin during larvaldevelopment and metamorphosis. In: Balls M, Bownes M,editors. Metamorphosis. Oxford: Oxford University Press.p 59–81.
Freda J, Dunson WA. 1984. Sodium balance of amphibianlarvae exposed to low environmental pH. Physiol Zool 57:435–443.
Freda J, McDonald DG. 1988. Physiological correlates ofinterspecific variation in acid tolerance in fish. J Exp Biol136:243–258.
Freda J, Taylor DH. 1992. Behavioral response of amphibianlarvae to acid water. J Herpetol 26:429–433.
Freda J. 1986. The influence of acidic pond water onamphibians: a review. Water Air Soil Pollut 30:439–450.
Gallien L, Bidaud O. 1959. Table cronologique du developpe-ment chez Triturus helveticus Razoumowsky. Bull Soc ZoolFrance 22–32.
Gascon C, Bider JR. 1985. The effect of pH on bullfrog, Ranacatesbeiana, and green frog, Rana clamitans melanota,tadpoles. Can Field Nat 99:259–261.
Gosner KL, Black IH. 1957. The effects of acidity on thedevelopment and hatching of New Jersey frogs. Ecology38:256–262.
Grant KP, Licht LE. 1993. Acid tolerance of Anuran embryosand larvae from Central Ontario. J Herpetol 27:1–6.
Griffiths RA, De Wijer P. 1994. Differential effects of pH andtemperature on embryonic development in the British newt(Triturus). J Zool Lond 234:613–622.
Griffiths RA. 1993. The effect of pH on feeding behaviour innewt larvae (Triturus: Amphibia). J Zool Lond 231:285–290.
Horne MT, Dunson WA. 1994. Exclusion of the JeffersonSalamander, Ambystoma jeffersonianum, from some poten-tial breeding ponds in Pennsylvania: effects of pH, tem-perature and metals on embryonic development. ArchEnviron Contam Toxicol 27:323–330.
Karns DR. 1992. Effects of acidic bog habitats on amphibianreproduction in a northern Minnesota peatland. J Herpetol26:401–412.
Kato S, Kurihara K. 1988. Electron microscopic studies of theexternal gill epithelium of the salamander, Hynobius dunniTago, followed throughout larval life. Okajimas Folia AnatJapan 65:177–202.
Kutka FJ. 1994. Low pH effects on swimming activity ofAmbystoma Salamander larvae. Environ Toxicol Chem 13:1821–1824.
Lenhart B, Steinberg C. 1984. Limnochemische und limno-biologische Auswirkungen der Versauerung von kalkarmenOberflachengewassern. Informatinsberichte Bayer. Land-esamt f. Wasserwirtschaft. 210p.
Leuven RSEW, Den Hartog C, Christiaans MMC, HeijligersWHC. 1986. Effects of water acidification on the distributionpattern and the reproductive success of amphibians.Experientia 42:495–503.
Lewinson D, Rosenberg M, Warburg MR. 1982. Mitochondria-rich cells in Salamander larva epidermis; ultrastructuraldescription and carbonic anhydrase activity. Biol Cell 46:75–84.
Ling RW, VanAmberg JP, Werner JK. 1986. Pond acidity andits relationship to larval development of Ambystomamaculatum and Rana sylvatica in upper Michigan.J Herpetol 20:230–236.
Linnenbach M, Marthaler R, Gebhardt H. 1987. Effect ofacid water on gills and epidermis in brown trout (Salmotrutta L.) and in tadpoles of the common frog (Ranatemporaria L.). In: Witters H, Vanderborght O, editors.Ecophysiology of acid stress in aquatic organism. Ann Soc RZool Belg Arch Environ Contam Toxicol 117:365–374.
McDonald DG, Ozog JL, Simons BP. 1984. The influence oflow pH environments on ion regulation in the larval stagesof the anuran amphibian, Rana clamitans. Can J Zool 62:2171–2177.
Nation JL. 1983. A new method using hexamethyl-disilazanefor preparation of soft insect tissues for scanning electronmicroscopy. Stain Technol 58:347–351.
Pechman JHK, Scott DE, Semlitsh RD, Caldwell JP, Vitt LJ,Gibbons JW. 1991. Declining amphibian populations: theproblem of separating human impacts from natural fluctua-tions. Science 253:893–895.
Pierce BA, Hoskin JB, Epstein E. 1984. Acid tolerance inConnecticut wood frogs (Rana sylvatica). J Herpetol 18:159–167.
Pierce BA, Wooten DK. 1992. Acid tolerance of Ambystomatexanum from Central Texas. J Herpetol 26:230–232.
Pierce BA. 1985. Acid tolerance in amphibians. Bioscience 35:239–243.
Pierce BA. 1993. The effects of acid precipitation onamphibians. Ecotoxicology 2:65–77.
E. BRUNELLI AND S. TRIPEPI956

Pough FH. 1976. Acid precipitation and embryonic mortalityof spotted salamanders Ambystoma maculatum. Science.19:68–70.
Rosenberg EA, Pierce BA. 1995. Effect on initial masson growth and mortality at low pH in tadpoles ofPseudacris clarkii and Bufo valliceps. J Herpetol 29:181–185.
Rowe CL, Sadinsky WJ, Dunson WA. 1992. Effects of acuteand chronic acidification on three larval amphibians thatbreed in temporary ponds. Arch Environ Contam Toxicol23:339–350.
Tripepi S, Brunelli E. 2000. Ultrastructural observation onthe gill main filament of Triturus italicus larva. Mus Reg SciNat Torino 187–192.
Warburg MR, Lewinson D, Rosenberg M. 1994. Structure andfunction of Salamandra skin and gills. Mertensiella 4:423–452.
Whiteman HH, Howard RD, Whitten KA. 1995. Effects of pHon embryo tolerance and adult behavior in the tigersalamander: Ambystoma tigrinum tigrinum. Can J Zool 73:1529–1537.
Wissinger SA, Whiteman HH. 1992. Fluctuation in a RockyMountain population of salamanders: anthropogenic acidi-fication or natural variation? J Herpetol 26:377–391.
Wood CM, McDonald D. 1987. The physiology of acid/aluminium and toxic effects on Atlantic salmo. In: WittersH, Vanderborght O, editors. Ecophysiology of acid stress inaquatic organisms. Ann Soc R Zool Belg. 117:399–410.
LOW pH EFFECTS ON TRITURUS ITALICUS GILLS 957




















