
Effects of copper on Enteromorpha flexuosa (Chlorophyta) in vitro
9
Ecotoxicology and Environmental Safety 58 (2004) 117–125 Effects of copper on Enteromorpha flexuosa (Chlorophyta) in vitro Leonardo R. Andrade, a Marcos Farina, a and Gilberto M. Amado Filho b, a Laborato ´rio de Biomineraliza@a˜o, Instituto de Cieˆncias Biome´dicas/CCS, Universidade Federal do Rio de Janeiro, Cidade Universita´ria, 21941-590 Rio de Janeiro, RJ, Brazil b Programa Zona Costeira, Instituto de Pesquisas Jardim Botaˆnico/MMA, Rua Pacheco Lea˜o 915, 22460-030, Rio de Janeiro, RJ, Brazil Received 11 October 2002; received in revised form 17 June 2003; accepted 20 June 2003 Abstract Enteromorpha species are widely used as biomonitors of copper (Cu) contamination in coastal waters, but the effects of Cu at the subcellular level and possible mechanisms of metal resistance are scarcely known. To contribute to the understanding of the Cu accumulation process in macroalgae species, we exposed adult individuals of Enteromorpha flexuosa to 50, 250, and 500 mg Cu/L in seawater for 5 days under controlled conditions. Thereafter, gross photosynthesis rate (GPR) and metal accumulation were measured. Conventional transmission electron microscopy (CTEM) and energy-dispersive X-ray analysis (EDXA) were used to study the metal accumulation process at the cellular level. Treatments with 250 and 500 mg Cu/L were observed to cause an inhibition of the GPR in the algae. Cu accumulation in samples exposed to 500 mg Cu/L was 52847561 mg Cu/g (dry wt), whereas in control samples (no Cu addition), accumulation was 971 mg/g. In cells of plants undergoing the 50 mg Cu/L treatment, the cytoplasm was clearly vacuolated. Thickening of cell walls and increase of relative number of starch granules and of lipid bodies were the main cellular changes observed in plants exposed to 250 and 500 mg Cu/L. EDXA of algae cells after 250 and 500 mg Cu/L exposure detected Cu mainly in vacuole precipitates. Cu was also detected in chloroplasts and in some epiphytic bacteria. It was concluded that E. flexuosa did not avoid penetration of Cu into the cytoplasm and consequently its toxic effects in concentrations of 250 and 500 mg Cu/L. Precipitates containing Cu in vacuoles should be related to a metal immobilization mechanism, minimizing the Cu toxicity for cells. The epiphytic bacteria can act as a biofilter diminishing the availability of free Cu +2 for algae accumulation. r 2003 Elsevier Inc. All rights reserved. Keywords: Algae; Copper; Enteromorpha; Epiphytic bacteria; Ultrastructure; X-ray microanalysis 1. Introduction Various sources of Cu, including industrial and domestic wastes, agricultural practices, copper mine drainage, copper-based pesticides, and antifouling paints, have contributed to a progressive increase in Cu concentrations in aquatic environments (Reed and Moffat, 1983; Ho, 1987; Nor, 1987; Correa et al., 1996; Callow and Callow, 2002). The free-Cu +2 or hydrated forms (Cu +2 H 2 O) are considered the most toxic Cu species for marine organisms because of their high availability for accu- mulation (Lobban et al., 1985; Sunda and Guillard, 1996). Physicochemical characteristics of metallic ions in seawater are influenced by environmental features such as pH, temperature, salinity, redox potential, products from biological activities, and dissolved organic matter (Karez et al., 1994; Suzuki et al., 1995; Amado Filho et al., 1997a, b; Gledhill et al., 1997). The free-Cu +2 species exist at very low concentrations in coastal waters as a result of their considerable capacity to form complexes with organic matter (Coale and Bruland, 1990). Copper (Z ¼ 29) is an essential microelement for algae, participating in important biological reactions as an enzymatic cofactor (amine oxidase and cytochrome c oxidase) and electron carrier in the photosynthetic process (plastocyanin and Cu-free cytochrome c 6 ) (Lobban et al., 1985; Linder, 1991). As a result of several characteristics, Enteromorpha species (Ulvophyceae) are widely used as biomonitors to assess the dissolved Cu fraction in aquatic environ- ments. These include its sedentary and cosmo- politan nature, its abundant populations, and its capacity to accumulate high Cu concentrations ARTICLE IN PRESS Corresponding author. Fax: +55-21-22942295. E-mail address: gfi[email protected] (G.M. Amado Filho). 0147-6513/$ - see front matter r 2003 Elsevier Inc. All rights reserved. doi:10.1016/S0147-6513(03)00106-4
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Effects of copper on Enteromorpha flexuosa (Chlorophyta) in vitro

Ecotoxicology and Environmental Safety 58 (2004) 117–125
ARTICLE IN PRESS
�Corresp
E-mail a
0147-6513/$
doi:10.1016
Effects of copper on Enteromorpha flexuosa (Chlorophyta) in vitro
Leonardo R. Andrade,a Marcos Farina,a and Gilberto M. Amado Filhob,�
aLaboratorio de Biomineraliza@ao, Instituto de Ciencias Biomedicas/CCS, Universidade Federal do Rio de Janeiro, Cidade Universitaria,
21941-590 Rio de Janeiro, RJ, BrazilbPrograma Zona Costeira, Instituto de Pesquisas Jardim Botanico/MMA, Rua Pacheco Leao 915, 22460-030, Rio de Janeiro, RJ, Brazil
Received 11 October 2002; received in revised form 17 June 2003; accepted 20 June 2003
Abstract
Enteromorpha species are widely used as biomonitors of copper (Cu) contamination in coastal waters, but the effects of Cu at the
subcellular level and possible mechanisms of metal resistance are scarcely known. To contribute to the understanding of the Cu
accumulation process in macroalgae species, we exposed adult individuals of Enteromorpha flexuosa to 50, 250, and 500 mgCu/L in
seawater for 5 days under controlled conditions. Thereafter, gross photosynthesis rate (GPR) and metal accumulation were
measured. Conventional transmission electron microscopy (CTEM) and energy-dispersive X-ray analysis (EDXA) were used to
study the metal accumulation process at the cellular level. Treatments with 250 and 500 mgCu/L were observed to cause an
inhibition of the GPR in the algae. Cu accumulation in samples exposed to 500mgCu/L was 52847561 mgCu/g (dry wt), whereas incontrol samples (no Cu addition), accumulation was 971mg/g. In cells of plants undergoing the 50 mgCu/L treatment, the
cytoplasm was clearly vacuolated. Thickening of cell walls and increase of relative number of starch granules and of lipid bodies
were the main cellular changes observed in plants exposed to 250 and 500mgCu/L. EDXA of algae cells after 250 and 500 mgCu/Lexposure detected Cu mainly in vacuole precipitates. Cu was also detected in chloroplasts and in some epiphytic bacteria. It was
concluded that E. flexuosa did not avoid penetration of Cu into the cytoplasm and consequently its toxic effects in concentrations of
250 and 500mgCu/L. Precipitates containing Cu in vacuoles should be related to a metal immobilization mechanism, minimizing theCu toxicity for cells. The epiphytic bacteria can act as a biofilter diminishing the availability of free Cu+2 for algae accumulation.
r 2003 Elsevier Inc. All rights reserved.
Keywords: Algae; Copper; Enteromorpha; Epiphytic bacteria; Ultrastructure; X-ray microanalysis
1. Introduction
Various sources of Cu, including industrial anddomestic wastes, agricultural practices, copper minedrainage, copper-based pesticides, and antifoulingpaints, have contributed to a progressive increase inCu concentrations in aquatic environments (Reed andMoffat, 1983; Ho, 1987; Nor, 1987; Correa et al., 1996;Callow and Callow, 2002).The free-Cu+2 or hydrated forms (Cu+2 �H2O) are
considered the most toxic Cu species for marineorganisms because of their high availability for accu-mulation (Lobban et al., 1985; Sunda and Guillard,1996). Physicochemical characteristics of metallic ions inseawater are influenced by environmental features suchas pH, temperature, salinity, redox potential, products
onding author. Fax: +55-21-22942295.
ddress: [email protected] (G.M. Amado Filho).
- see front matter r 2003 Elsevier Inc. All rights reserved.
/S0147-6513(03)00106-4
from biological activities, and dissolved organic matter(Karez et al., 1994; Suzuki et al., 1995; Amado Filhoet al., 1997a, b; Gledhill et al., 1997). The free-Cu+2
species exist at very low concentrations in coastal watersas a result of their considerable capacity to formcomplexes with organic matter (Coale and Bruland,1990).Copper (Z ¼ 29) is an essential microelement for
algae, participating in important biological reactions asan enzymatic cofactor (amine oxidase and cytochromec oxidase) and electron carrier in the photosyntheticprocess (plastocyanin and Cu-free cytochrome c6)(Lobban et al., 1985; Linder, 1991).As a result of several characteristics, Enteromorpha
species (Ulvophyceae) are widely used as biomonitorsto assess the dissolved Cu fraction in aquatic environ-ments. These include its sedentary and cosmo-politan nature, its abundant populations, and itscapacity to accumulate high Cu concentrations

ARTICLE IN PRESSL.R. Andrade et al. / Ecotoxicology and Environmental Safety 58 (2004) 117–125118
(Riquelme et al., 1997; Correa et al., 1999; Haritonidisand Malea, 1999).Biofouling on ship hulls results in an increase in
roughness and leads to an increase in hydrodynamicdrag as the vessel moves through water, resulting inadditional fuel consumption and necessitating periodichull cleaning, antifouling paint removal, and repainting.The genus Enteromorpha has been considered the majormacrofouling alga commonly found adhering to boathulls protected with antifouling paints (Callow andCallow, 2002). Copper-based antifouling paints arewidely used and have a profound toxic effect on theenvironment, principally on initial-stage growth inseveral organisms.Electron microscopy (EM) analytical techniques, such
as energy-dispersive X-ray analysis (EDXA), have beensystematically employed in biological sciences to identifychemical elements in cells. X-ray microanalysis has beenapplied to localize metals within the cells of differentalgae species, contributing to our comprehension ofmechanisms of tolerance and resistance (Silverberg,1975; Lignell et al., 1982; Ballan-Dufran@ais et al.,1991; Pellegrini et al., 1991, 1993; Tretyn et al., 1996;Webster and Gadd, 1996; Amado Filho et al., 1999;Correa et al., 1999; Stengel and Dring, 2000; Andradeet al., 2002; Talarico, 2002).To contribute to our understanding of the process by
which marine green algae accumulate Cu, we exposedadult individuals of E. flexuosa to Cu concentrations inseawater under controlled conditions. Thereafter, grossphotosynthesis rate (GPR) and metal concentrationswere determined. Samples were also studied by conven-tional transmission electron microscopy (CTEM) andEDXA.
2. Materials and methods
2.1. Experimental conditions
Adult individuals (8–10 cm long) of E. flexuosa werecollected at the intertidal zone of Barra da Tijuca beach(city of Rio de Janeiro, Brazil), an area considered notto be contaminated by metals, where the Cu concentra-tion in seawater was in a range of 0.6–1.6 mg/L (Reeset al., 1998). In the laboratory, the algae samples werewashed in seawater and transferred to a glass aquariumwith 3000mL of seawater (35 PSU) filtered in Millipore0.45 mm pore size. The algae were maintained for 3 daysin the laboratory for acclimatization before the begin-ning of the experiment. The laboratory conditions wereas follow: temperature 2172�C; photon flux density100 mmol/m2/s; light/dark cycle 12 h light/12 h dark.Four treatments were carried out: control (no Cuaddition), 50, 250, and 500 mgCu/L, added as Cu2SO4 �H2O (Sigma) to the filtered seawater. Approximately
10 g (wet wt) of E. flexuosa were cultured in a volume of500mL, in triplicates for each treatment, during 5 days.The seawater medium was changed on day 3. After theexperimental period, algae samples were taken fordissolved-oxygen measurements, Cu quantification,and TEM analysis.
2.2. Gross photosynthesis rate (GPR)
GPR was determined by measurements of oxygenlevels produced by algae with a dissolved-oxygenanalyzer (Clark-O2 probe YSI 5000) according toThomas (1988). Algae samples (1 g wet wt) from theCu treatments (50, 250, and 500 mgCu/L) and thecontrol were transferred to 300-mL bottles (in tripli-cate), with or without illumination. Illuminated bottleswere called light bottle (LB) and those withoutillumination, dark bottle (DB). The seawater used tofill the LB and DB was the same for each treatment. Thealgae were maintained for 5 h at 2172�C with photonflux density of 100 mmol/m2/s in LB. Oxygen levels weremeasured in both LB and DB. The final results wereobtained by using the following formula: GPR=x/algaebiomass (mg dry wt), where x ¼ ½ðLB � DBÞ=T :GPR isthe GPR (mg O2/mg/h); LB is the LB oxygenconcentration (mg/L); DB is the DB oxygen concentra-tion (mg/L); T is the incubation time (h).
2.3. Accumulation of Cu
After 5 days of Cu exposition, algae subsamples werewashed in distilled water and dried at 50�C. Samples of250mg (triplicates) were digested in 10mL HNO3
(Merck PA 65%) in closed Teflon flasks in a microwaveoven (CEM-MDS-2000) for 30min. The samples weredried and 5mL of HCl (0.1M) was added. Concentra-tions of Cu and Mg (a constitutive element of thechlorophyll molecule) were measured by flame atomicabsorption spectrophotometry (AAS-Varian AA-1475).The results were expressed in mg/g dry wt. Analyticalprocedures were tested by comparative analysis ofInternational Atomic Energy Agency (IAEA)-certifiedreference material IAEA-140 (sea plant homogenate,Fucus). The results obtained were within the confidencelimit (significance level a ¼ 0:05).The existence of significant differences among Cu
accumulation results and among GPR results in thetested copper treatments were determined by one-wayanalysis of variance (ANOVA) and by post hoc‘‘Tukey’’ multiple comparisons test.
2.4. Conventional TEM
Pieces of B9mm2 from the median region of thethallus (5 cm above the base) were cut with razor bladesunder a stereomicroscope and immediately fixed in 2.5%

ARTICLE IN PRESS
Table 1
GPR of E. flexuosa exposed to different Cu concentrations
Treatments Gross photosynthetic ratea
Control 27.6777.62
50 mgCu/L 23.6874.95
250mgCu/L �0.9970.25
500mgCu/L �0.84570.29
aResults shown as mg O2/mg/h (mean7SD).
Table 2
Copper and magnesium accumulation in E. flexuosa exposed to
different copper concentrations for 5 days
Treatments Metal accumulationa
Copper Magnesium
Control 9.5071.61 19,938.117396.45
50 mg/L 860.55710.10 16,282.017316.67
250mg/L 4160.757331.43 13,018.087217.21
500mg/L 5284.917561.22 13,662.537427.24
aResults are shown in mg/g dry wt (mean7SD).
L.R. Andrade et al. / Ecotoxicology and Environmental Safety 58 (2004) 117–125 119
glutaraldehyde in 0.1M cacodylate buffer (pH 7.2) infiltered seawater in a microwave oven (Pelco LMP 3450)for 10 s (2 ) at 40�C. The samples were postfixed in 1%buffered OsO4 under microwaves for 10 s (2 ),dehydrated through an acetone series, and embeddedin Spurr low-viscosity resin. Ultrathin sections (70 nm)were obtained using a Reichert–Jung ultramicrotome,placed in copper grids (300 mesh), and stained with 5%uranyl acetate for 30min and 2% lead citrate for 5min.Observations were done in a Zeiss 900 EM, operated at80 kV. To compare cell ultrastructural changes, numberof vesicles, lipid bodies, and starch granules weredetermined in at least 15 cells from each treatment.These quantifications were done by using the softwareSigma Scan Pro (Version 3.0), and the results arepresented as mean7SD (see Results section).
2.5. Analytical TEM
For analytical purposes, the samples were obtainedand fixed as already described, without the steps ofpostfixation and dehydration. Thereafter, they wereembedded in Nanoplast resin, a special water-solublemelamine resin. The samples were kept at 40�C for 2days (infiltration) and at 60�C for 1 day (polymeriza-tion).For EDXA, a LEO 912 EM equipped with an
Oxford-Link analytical system was used. The standardspecimen holder was replaced by a beryllium specimenholder to avoid undesirable copper peaks due to electronscattering on the holder and the EM column. Non-stained ultrathin sections were placed on nylon grids.Typical acquisition data were as follow: sample tiltangle=15�; livetime=1000 s; deadtime=20%; acceler-ating voltage=80 kV; beam spot sizeD63 nm (dia-meter). The control spectra were obtained fromelectron-lucent cytoplasm regions, resin, and a gridregion without section, to evaluate a possible Cu signalfrom the microscope column.
3. Results
3.1. Gross photosynthesis rate
At the end of the experimental period, we observedplant general color to vary from dark green in controlsamples to light green in samples that were treated with500 mgCu/L. The GPR results showed no significantdifference (ANOVA, P ¼ 0:51; F ¼ 0:52; df=5) be-tween the control (27.6777.62mg O2/mg/h) andthe 50 mgCu/L treatment (23.6874.95mg O2/mg/h)(Table 1). The two other Cu treatments, 250 and500 mgCu/L, showed similar GPR data and significantinhibition in relation to the control (ANOVA, control250 mgCu/L, P ¼ 0:002; F ¼ 48:78; df=5; control
500 mgCu/L, P ¼ 0:002; F ¼ 47:41; df=5). The GPRwas negative in these two treatments (Table 1).
3.2. Copper accumulation
A significant increase in Cu accumulation (ANOVA,Po0:001; F ¼ 166; df=11) was observed in samplesfrom the three Cu-added treatments when comparedwith the control samples (Table 2). An increase of twoorders of magnitude was seen in individual plantsreceiving 50 mgCu/L and of three orders of magnitudein individual plants receiving 250 and 500 mgCu/Ltreatments (Table 2). In contrast, we found a significantreduction (ANOVA, P ¼ 0:03; F ¼ 5:11; df=11) in Mgconcentration between control and Mg-added treat-ments (Table 2).
3.3. Ultrastructural observations
Conventional TEM of E. flexuosa control cellsshowed their typical ultrastructural features, such as:parietal lobed chloroplast associated to rounded pyr-enoid surrounded by starch granules, lipid bodies,central nucleus, and large vacuole (Figs. 1 and 2).In cells of plants from 50 mgCu/L treatment, the
cytoplasm was more vacuolated and several exocyticvesicles of different types were seen (Fig. 3). A highnumber of small cytoplasmic vesicles (1273 vesicles percell) in relation to the control cells (371 vesicles per cell)were found associated with Golgi apparatus (GA)(Fig. 4), and large vesicles containing minor vesicles intheir interior were abundant at the plasmalemma–cellwall interface (Fig. 5). The stacks of GA in 50 mgCu/Ltreatment cells were irregularly arranged (Fig. 4) in
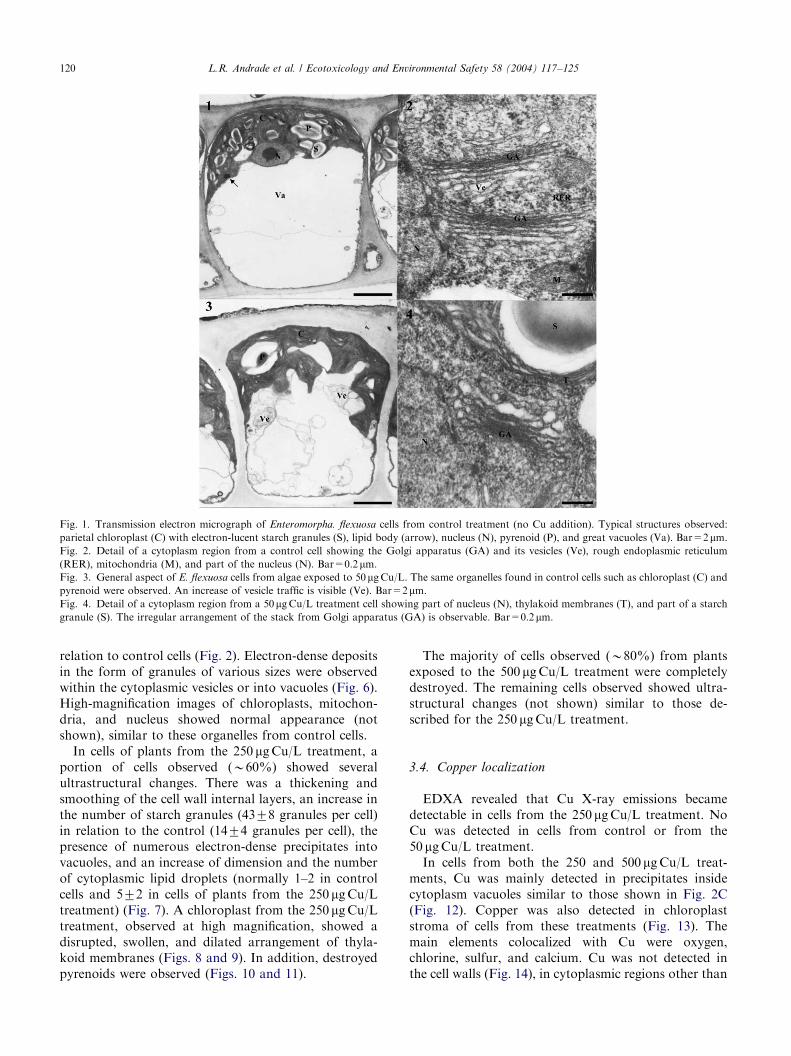
ARTICLE IN PRESS
Fig. 1. Transmission electron micrograph of Enteromorpha. flexuosa cells from control treatment (no Cu addition). Typical structures observed:
parietal chloroplast (C) with electron-lucent starch granules (S), lipid body (arrow), nucleus (N), pyrenoid (P), and great vacuoles (Va). Bar=2mm.Fig. 2. Detail of a cytoplasm region from a control cell showing the Golgi apparatus (GA) and its vesicles (Ve), rough endoplasmic reticulum
(RER), mitochondria (M), and part of the nucleus (N). Bar=0.2mm.Fig. 3. General aspect of E. flexuosa cells from algae exposed to 50 mgCu/L. The same organelles found in control cells such as chloroplast (C) and
pyrenoid were observed. An increase of vesicle traffic is visible (Ve). Bar=2mm.Fig. 4. Detail of a cytoplasm region from a 50mgCu/L treatment cell showing part of nucleus (N), thylakoid membranes (T), and part of a starch
granule (S). The irregular arrangement of the stack from Golgi apparatus (GA) is observable. Bar=0.2mm.
L.R. Andrade et al. / Ecotoxicology and Environmental Safety 58 (2004) 117–125120
relation to control cells (Fig. 2). Electron-dense depositsin the form of granules of various sizes were observedwithin the cytoplasmic vesicles or into vacuoles (Fig. 6).High-magnification images of chloroplasts, mitochon-dria, and nucleus showed normal appearance (notshown), similar to these organelles from control cells.In cells of plants from the 250 mgCu/L treatment, a
portion of cells observed (B60%) showed severalultrastructural changes. There was a thickening andsmoothing of the cell wall internal layers, an increase inthe number of starch granules (4378 granules per cell)in relation to the control (1474 granules per cell), thepresence of numerous electron-dense precipitates intovacuoles, and an increase of dimension and the numberof cytoplasmic lipid droplets (normally 1–2 in controlcells and 572 in cells of plants from the 250 mgCu/Ltreatment) (Fig. 7). A chloroplast from the 250 mgCu/Ltreatment, observed at high magnification, showed adisrupted, swollen, and dilated arrangement of thyla-koid membranes (Figs. 8 and 9). In addition, destroyedpyrenoids were observed (Figs. 10 and 11).
The majority of cells observed (B80%) from plantsexposed to the 500 mgCu/L treatment were completelydestroyed. The remaining cells observed showed ultra-structural changes (not shown) similar to those de-scribed for the 250 mgCu/L treatment.
3.4. Copper localization
EDXA revealed that Cu X-ray emissions becamedetectable in cells from the 250 mgCu/L treatment. NoCu was detected in cells from control or from the50 mgCu/L treatment.In cells from both the 250 and 500 mgCu/L treat-
ments, Cu was mainly detected in precipitates insidecytoplasm vacuoles similar to those shown in Fig. 2C(Fig. 12). Copper was also detected in chloroplaststroma of cells from these treatments (Fig. 13). Themain elements colocalized with Cu were oxygen,chlorine, sulfur, and calcium. Cu was not detected inthe cell walls (Fig. 14), in cytoplasmic regions other than
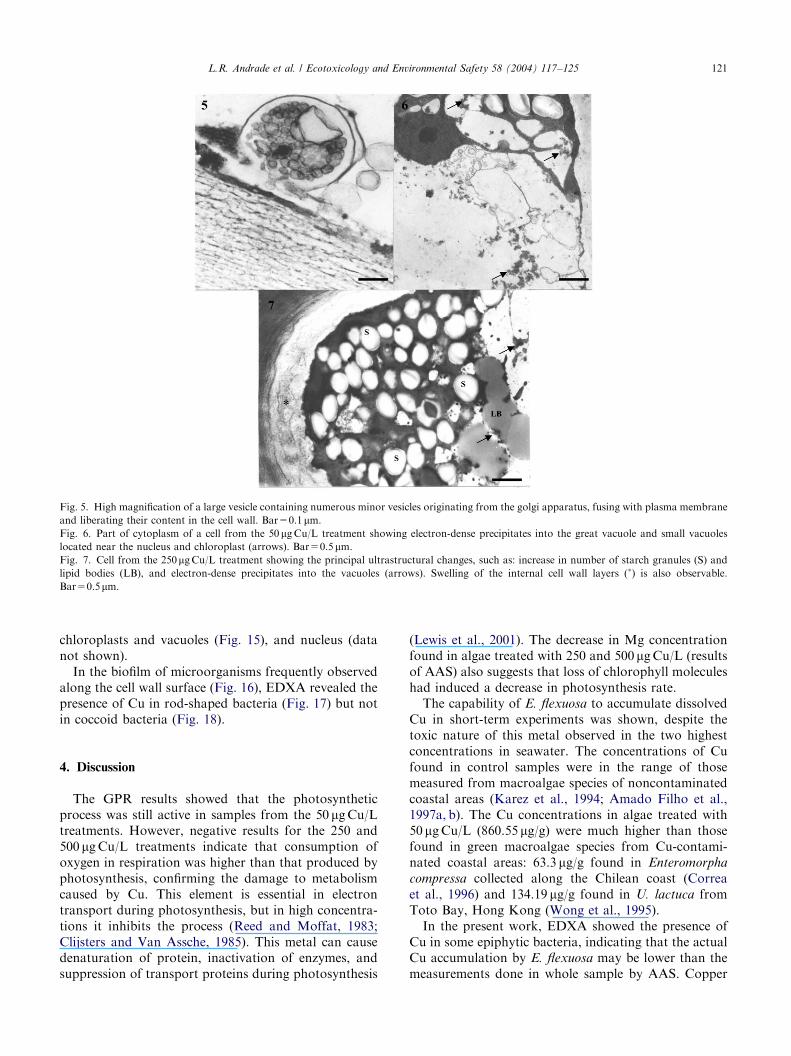
ARTICLE IN PRESS
Fig. 5. High magnification of a large vesicle containing numerous minor vesicles originating from the golgi apparatus, fusing with plasma membrane
and liberating their content in the cell wall. Bar=0.1mm.Fig. 6. Part of cytoplasm of a cell from the 50 mgCu/L treatment showing electron-dense precipitates into the great vacuole and small vacuoles
located near the nucleus and chloroplast (arrows). Bar=0.5mm.Fig. 7. Cell from the 250mgCu/L treatment showing the principal ultrastructural changes, such as: increase in number of starch granules (S) and
lipid bodies (LB), and electron-dense precipitates into the vacuoles (arrows). Swelling of the internal cell wall layers (�) is also observable.
Bar=0.5mm.
L.R. Andrade et al. / Ecotoxicology and Environmental Safety 58 (2004) 117–125 121
chloroplasts and vacuoles (Fig. 15), and nucleus (datanot shown).In the biofilm of microorganisms frequently observed
along the cell wall surface (Fig. 16), EDXA revealed thepresence of Cu in rod-shaped bacteria (Fig. 17) but notin coccoid bacteria (Fig. 18).
4. Discussion
The GPR results showed that the photosyntheticprocess was still active in samples from the 50 mgCu/Ltreatments. However, negative results for the 250 and500 mgCu/L treatments indicate that consumption ofoxygen in respiration was higher than that produced byphotosynthesis, confirming the damage to metabolismcaused by Cu. This element is essential in electrontransport during photosynthesis, but in high concentra-tions it inhibits the process (Reed and Moffat, 1983;Clijsters and Van Assche, 1985). This metal can causedenaturation of protein, inactivation of enzymes, andsuppression of transport proteins during photosynthesis
(Lewis et al., 2001). The decrease in Mg concentrationfound in algae treated with 250 and 500 mgCu/L (resultsof AAS) also suggests that loss of chlorophyll moleculeshad induced a decrease in photosynthesis rate.The capability of E. flexuosa to accumulate dissolved
Cu in short-term experiments was shown, despite thetoxic nature of this metal observed in the two highestconcentrations in seawater. The concentrations of Cufound in control samples were in the range of thosemeasured from macroalgae species of noncontaminatedcoastal areas (Karez et al., 1994; Amado Filho et al.,1997a, b). The Cu concentrations in algae treated with50 mgCu/L (860.55 mg/g) were much higher than thosefound in green macroalgae species from Cu-contami-nated coastal areas: 63.3 mg/g found in Enteromorpha
compressa collected along the Chilean coast (Correaet al., 1996) and 134.19 mg/g found in U. lactuca fromToto Bay, Hong Kong (Wong et al., 1995).In the present work, EDXA showed the presence of
Cu in some epiphytic bacteria, indicating that the actualCu accumulation by E. flexuosa may be lower than themeasurements done in whole sample by AAS. Copper

ARTICLE IN PRESS
Fig. 8. Detail of a chloroplast region of a cell exposed to 250mgCu/L, showing disrupted, swollen, and dilated arrangement of thylakoid membranes.Bar=0.2mm.Fig. 9. Detail of a chloroplast region of a cell from the control treatment showing the typical arrangement of thylakoid membranes. Bar=0.5 mm.Fig. 10. View of a pyrenoid of a cell from the 250mgCu/L treatment, completely destroyed. Some starch granules are also observed. Bar=0.2 mm.Fig. 11. Pyrenoid of a control cell forming a starch granule (�). Some plastoglobules (arrows) and thylakoid membranes are visible. Bar=0.2 mm.
L.R. Andrade et al. / Ecotoxicology and Environmental Safety 58 (2004) 117–125122
was also detected by EDXA in epiphytic bacteria inE. compressa collected in a Cu-contaminated area innorthern Chile (Correa et al., 1999). Riquelme et al.(1997) observed that Cu concentrations in E. compressa
samples without bacterial film (mechanically removed)decreased significantly in comparison to samples whosebiofilm was not removed before metal analysis. Stengeland Dring (2000) proposed that the variation of Cu andFe accumulated by Ascophyllum nodosum in three sitesin Ireland were influenced by the distinct degree ofepiphytism in this alga, principally because X-raymicroanalysis by SEM showed that most of the ironwas located in epiphytic pennate diatoms.The increased number of starch granules and the
disarrangement of thylakoid bands indicate that Cucaused changes in E. flexuosa cell metabolism. Theincrease in the number of storage product as starchgranules and lipid droplets in green algae are influencedby stress conditions. Among the physiological mechan-isms that have been suggested to explain tolerance orresistance and accumulation capability of heavy metalsby algae are metallic complex formation with metabo-lites produced and/or exuded by algae (Ragan and
Glombitza, 1986; Lombardi and Vieira, 1999, 2000);adsorption, immobilization, and precipitation of toxicmetals in extracellular and cellular compartments such ascell walls, phosphate-rich granules, lipid bodies, va-cuoles, nucleus, and physodes (Silverberg, 1975; Lignellet al., 1982; Pellegrini et al., 1991, 1993; Wong et al.,1995; Webster and Gadd, 1996; Tretyn et al., 1996,Amado Filho et al., 1999; Andrade et al., 2002); metalbinding in specific organic-metallic compounds such asmetalloproteins and phytochelatins (Robinson, 1989;Robinson et al., 1993, Lombardi and Vieira, 1999, 2000).The cell wall acts as a cation exchange system, binding
and selecting metal ions that will be absorbed within thecell (Mariani et al., 1990). However, in our experimentalconditions the increase of cell wall by newly formedpolysaccharides in E. flexuosa cell walls did not preventCu from reaching the cytoplasm. Silverberg (1975)observed that the vacuole was the major site for Pbdeposition and retention in the green alga Stigeoclonium
tenue. Similar findings were reported for differentseaweeds collected in copper mine areas along thenorthern coast of Chile. Correa et al. (1999) observedelectron-dense granules of diverse sizes scattered within
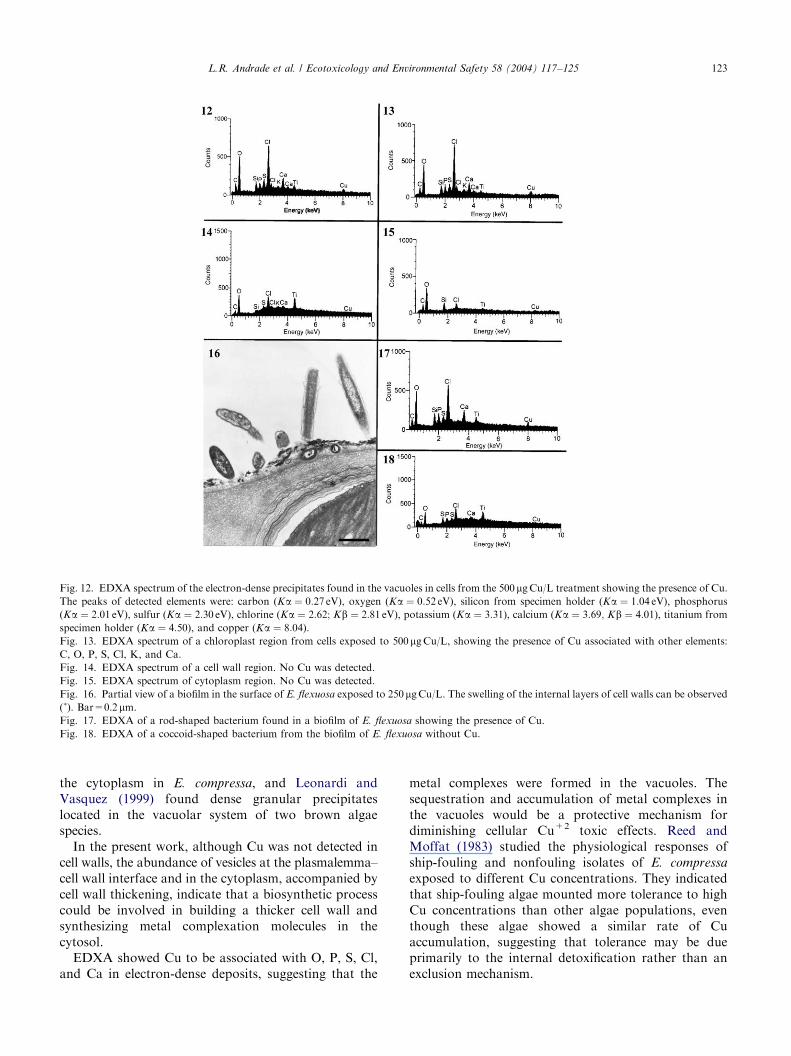
ARTICLE IN PRESS
Fig. 12. EDXA spectrum of the electron-dense precipitates found in the vacuoles in cells from the 500 mgCu/L treatment showing the presence of Cu.
The peaks of detected elements were: carbon (Ka ¼ 0:27 eV), oxygen (Ka ¼ 0:52 eV), silicon from specimen holder (Ka ¼ 1:04 eV), phosphorus
(Ka ¼ 2:01 eV), sulfur (Ka ¼ 2:30 eV), chlorine (Ka ¼ 2:62; Kb ¼ 2:81 eV), potassium (Ka ¼ 3:31), calcium (Ka ¼ 3:69; Kb ¼ 4:01), titanium from
specimen holder (Ka ¼ 4:50), and copper (Ka ¼ 8:04).
Fig. 13. EDXA spectrum of a chloroplast region from cells exposed to 500 mgCu/L, showing the presence of Cu associated with other elements:
C, O, P, S, Cl, K, and Ca.
Fig. 14. EDXA spectrum of a cell wall region. No Cu was detected.
Fig. 15. EDXA spectrum of cytoplasm region. No Cu was detected.
Fig. 16. Partial view of a biofilm in the surface of E. flexuosa exposed to 250mgCu/L. The swelling of the internal layers of cell walls can be observed(�). Bar=0.2 mm.Fig. 17. EDXA of a rod-shaped bacterium found in a biofilm of E. flexuosa showing the presence of Cu.
Fig. 18. EDXA of a coccoid-shaped bacterium from the biofilm of E. flexuosa without Cu.
L.R. Andrade et al. / Ecotoxicology and Environmental Safety 58 (2004) 117–125 123
the cytoplasm in E. compressa, and Leonardi andVasquez (1999) found dense granular precipitateslocated in the vacuolar system of two brown algaespecies.In the present work, although Cu was not detected in
cell walls, the abundance of vesicles at the plasmalemma–cell wall interface and in the cytoplasm, accompanied bycell wall thickening, indicate that a biosynthetic processcould be involved in building a thicker cell wall andsynthesizing metal complexation molecules in thecytosol.EDXA showed Cu to be associated with O, P, S, Cl,
and Ca in electron-dense deposits, suggesting that the
metal complexes were formed in the vacuoles. Thesequestration and accumulation of metal complexes inthe vacuoles would be a protective mechanism fordiminishing cellular Cu+2 toxic effects. Reed andMoffat (1983) studied the physiological responses ofship-fouling and nonfouling isolates of E. compressa
exposed to different Cu concentrations. They indicatedthat ship-fouling algae mounted more tolerance to highCu concentrations than other algae populations, eventhough these algae showed a similar rate of Cuaccumulation, suggesting that tolerance may be dueprimarily to the internal detoxification rather than anexclusion mechanism.

ARTICLE IN PRESSL.R. Andrade et al. / Ecotoxicology and Environmental Safety 58 (2004) 117–125124
In conclusion, the AAS results indicated that E.
flexuosa can accumulate high Cu concentrations inshort-term experimental conditions, and the reductionin Mg concentration can be associated with negativeGPR results. According to the ultrastructure studies,this species did not avoid Cu penetration into theprotoplasm and its toxic effects, as confirmed by thesubcellular changes observed. The Cu-rich precipitatesfound in the vacuoles can be related to a metalimmobilization process that minimizes the Cu toxicityfor cells. The epiphytic bacteria can act as a biofilterdiminishing the availability of free Cu+2 for algaeaccumulation.
Acknowledgments
The authors thank Flavio C. Miguens (UENF) for theuse of the Leo 912 electron microscope, Rita C. Mansofor the dissolved-oxygen analysis, and Mair Machadofor technical support. This study is part of a doctoralthesis developed by L.R. Andrade at UFRJ—Brasil,council by FAPERJ. Financial support came fromPRONEX/CNPq, and research grants from CNPq(521688/96-5) and FAPERJ (E-26/170.336/98) toG.M.A.-F.
References
Amado Filho, G.M., Karez, C.S., Andrade, L.R., Yoneshigue-
Valentin, Y., Pfeiffer, W.C., 1997a. Effects on growth and
accumulation of zinc in six seaweed species. Ecotoxicol. Environ.
Saf. 37, 223–228.
Amado Filho, G.M., Andrade, L.R., Reis, R.P., Bastos, W.R.,
Pfeiffer, W.C., 1997b. Heavy metal concentration in seaweed
species from the Abrolhos Reef region, Brazil. Proceedings
of the VIII International Coral Reef Symposium, Vol. 2,
pp. 1843–1846.
Amado Filho, G.M., Andrade, L.R., Karez, C.S., Farina, M., Pfeiffer,
W.C., 1999. Brown algae species as biomonitors of Zn and Cd
at Sepetiba Bay, Rio de Janeiro, Brazil. Mar. Environ. Res. 48,
213–224.
Andrade, L.R., Farina, M., Amado Filho, G.M., 2002. Role of cell
walls of Padina gymnospora (Phaeophyta) in Cd accumulation.
Phycologia 41 (1), 39–48.
Ballan-Dufran@ais, C., Marcaillou, C., Amiard-Triquet, C., 1991.
Response of the phytoplankton alga Tetraselmis suecica to copper
and silver exposure: vesicular metal bioaccumulation and lack of
starch bodies. Biol. Cell 72, 103–112.
Callow, M.E., Callow, J.A., 2002. Marine biofouling: a sticky
problem. Biologist 49 (1), 1–5.
Clijsters, H., Van Assche, F., 1985. Inhibition of photosynthesis by
heavy metals. Photosynth. Res. 7, 31–40.
Coale, K.H., Bruland, K.W., 1990. Copper complexation in the
Northeast Pacific. Limnol. Oceanogr. 33, 1084–1101.
Correa, J.A., Gonzales, P., Sanchez, P., Munoz, J., Orellana, M.C.,
1996. Copper–algae interactions: inheritance or adaptation?
Environ. Monit. Assess. 40, 41–54.
Correa, J.A., Castilla, J.C., Ramirez, M.A., Varas, M., Lagos, N.,
Vergara, S., Moenne, A., Roman, D., Brown, M.T., 1999. Copper,
copper mine tailings and their effects on marine algae in Northern
Chile. J. Appl. Phycol. 11, 57–67.
Gledhill, M., Nimmo, M., Hill, S.J., Brown, M.T., 1997. The toxicity
of copper (II) species to marine algae, with particular reference to
macroalgae. J. Phycol. 33 (1), 2–11.
Haritonidis, S., Malea, P., 1999. Bioaccumulation of metals by the
green alga Ulva rigida from Thermaikos Gulf, Greece. Environ.
Pollut. 104, 365–372.
Ho, Y., 1987. Metals in 19 intertidal macroalgae in Hong Kong waters.
Mar. Pollut. Bull. 18, 564–566.
Karez, C.S., Magalhaes, V.F., Pfeiffer, W.C., Amado Filho, G.M.,
1994. Trace metal accumulation by algae in Sepetiba Bay, Brazil.
Environ. Pollut. 83, 351–356.
Leonardi, P.I., Vasquez, J.A., 1999. Effects of copper pollution on
the ultrastructure of Lessonia spp. Hydrobiologia 398/399,
375–383.
Lewis, S., Donkin, M.E., Depledge, M.H., 2001. Hsp70 expression in
Enteromorpha intestinalis (Chlorophyta) exposed to environmental
stressors. Aquat. Toxicol. 51, 277–291.
Lignell, A., Roomans, G.M., Pedersen, M., 1982. Localization of
adsorbed cadmium in Fucus vesiculosus L. by X-ray microanalysis.
Z. Pflanzenphysiol. Bd. 105, 103–109.
Linder, M.C., 1991. Biochemistry of Copper. Plenum Press, New
York, pp. 34–78.
Lobban, C.S., Harrison, P.J., Duncan, M.J., 1985. The Physiological
Ecology of Seaweeds. Cambridge University Press, New York,
pp. 1–242.
Lombardi, A.T., Vieira, A.A.H., 1999. Lead- and copper-complexing
extracellular ligands released by K. aperta (Chlorococcales,
Chlorophyta). Phycologia 38 (4), 283–288.
Lombardi, A.T., Vieira, A.A.H., 2000. Copper complexation
by Cyanophyta and Chlorophyta exudates. Phycologia 39 (2),
118–125.
Mariani, P., Tolomio, C., Baldan, B., Braghetta, P., 1990. Cell wall
ultrastructure and cation localization in some benthic marine algae.
Phycologia 29, 253–262.
Nor, Y.M., 1987. Ecotoxicology of copper to aquatic biota: a review.
Environ. Res. 43, 274–282.
Pellegrini, L., Pellegrini, M., Delivopoulos, S., Berail, G., 1991. The
effects of cadmium on the fine structure of the brown alga
Cystoseira barbata forma repens Zinova et Kalugina. Br. Phycol.
J. 26, 1–8.
Pellegrini, M., Laugie, A., Sergent, M., Phan-Tan-Lun, R., Valls, R.,
Pellegrini, L., 1993. Interactions between the toxicity of heavy
metals cadmium, copper, zinc in combinations and detoxifying role
of calcium in the brown alga Cystoseira barbata. J. Appl. Phycol.
5, 351–361.
Ragan, M.A., Glombitza, K.W., 1986. Phlorotannins, brown algal
polyphenols. In: Round, F.E., Chapman, D.J. (Eds.), Progress
in Phycological Research. Biopress, Bristol, pp. 129–241.
Reed, R.H., Moffat, L., 1983. Copper toxicity and copper tolerance
in Enteromorpha compressa (L.) Grev. J. Exp. Mar. Biol. Ecol.
69, 85–103.
Rees, J.G., Breward, N., Maddock, J.E.L., Ferguson, A.J., Williams,
T.M., 1998. Land Ocean Contamination Study (LOCS): assess-
ment of contamination by metals and selected organic compounds
in coastal sediments and waters of Sepetiba Bay, Rio de Janeiro
State, Brazil. Technical Report WC/98/30, Overseas Geology
Series, British Geological Survey, Nottingham.
Riquelme, C., Rojas, A., Flores, V., Correa, J.A., 1997. Epiphytic
bacteria in a copper-enriched environment in Northern Chile. Mar.
Pollut. Bull. 34 (10), 816–820.
Robinson, N.J., 1989. Algal metallothioneins: secondary metabolites
and proteins. J. Appl. Phycol. 1, 5–18.
Robinson, N.J., Tommey, A.M., Kuske, C., Jackson, P.J., 1993. Plant
metallothioneins. Biochem. J. 295, 1–10.

ARTICLE IN PRESSL.R. Andrade et al. / Ecotoxicology and Environmental Safety 58 (2004) 117–125 125
Silverberg, B.A., 1975. Ultrastructural localization of lead in Stigeoclo-
nium tenue (Chlorophyceae, Ulotrichales) as demonstrated by
cytochemical and X-ray microanalysis. Phycologia 14, 265–274.
Stengel, D.B., Dring, M.J., 2000. Copper and iron concentrations in
Ascophyllum nodosum (Fucales, Phaeophyta) from different sites
in Ireland and after culture experiments in relation to thallus age
and epiphytism. J. Exp. Mar. Biol. Ecol. 246, 145–161.
Sunda, W.G., Guillard, R.R.L., 1996. The relationship between cupric ion
activity and the toxicity to phytoplankton. J. Mar. Res. 34, 511–529.
Suzuki, Y., Kuma, K., Matsunaga, K., 1995. Bioavailable iron species
in seawater measured by macroalga (Laminaria japonica) uptake.
Mar. Biol. 123, 173–178.
Talarico, L., 2002. Fine structure and X-ray microanalysis of a red
macrophyte cultured under cadmium stress. Environ. Pollut. 120
(3), 813–821.
Thomas, M.T.H., 1988. Photosynthesis and respiration of aquatic
macro-flora using the light and dark bottle oxygen method
and dissolved oxygen analyzer. In: Lobban, C.S., Chapman, D.J.,
Kremer, B.P. (Eds.), Experimental Phycology: A Labo-
ratory Manual. Cambridge University Press, Cambridge,
pp. 64–77.
Tretyn, A., Grolig, F., Magdowski, G., Wagner, G., 1996. Selective
binding of Ca, Zn, Cu and K by the physods of the green
alga Mougeotia scalaris. Folia Histochem. Cytobiol. 34 (2),
103–108.
Webster, E., Gadd, G.M., 1996. Cadmium replaces calcium in the cell
wall of Ulva lactuca. Biometals 9 (3), 241–244.
Wong, S.L., Wainwright, J.F., Pimenta, J., 1995. Quantification of
total and metal toxicity in wastewaters using algal bioassays.
Aquat. Toxicol. 31, 57–75.




















