
Effects of contact shape on the scaling of biological attachments
16
doi: 10.1098/rspa.2004.1326 , 305-319 461 2005 Proc. R. Soc. A Ralph Spolenak, Stanislav Gorb, Huajian Gao and Eduard Arzt attachments Effects of contact shape on the scaling of biological References http://rspa.royalsocietypublishing.org/content/461/2054/305#related-urls Article cited in: Email alerting service here right-hand corner of the article or click Receive free email alerts when new articles cite this article - sign up in the box at the top http://rspa.royalsocietypublishing.org/subscriptions go to: Proc. R. Soc. A To subscribe to This journal is © 2005 The Royal Society on May 18, 2011 rspa.royalsocietypublishing.org Downloaded from
Transcript of Effects of contact shape on the scaling of biological attachments

doi: 10.1098/rspa.2004.1326, 305-319461 2005 Proc. R. Soc. A
Ralph Spolenak, Stanislav Gorb, Huajian Gao and Eduard Arzt attachmentsEffects of contact shape on the scaling of biological
Referenceshttp://rspa.royalsocietypublishing.org/content/461/2054/305#related-urls
Article cited in:
Email alerting service hereright-hand corner of the article or click Receive free email alerts when new articles cite this article - sign up in the box at the top
http://rspa.royalsocietypublishing.org/subscriptions go to: Proc. R. Soc. ATo subscribe to
This journal is © 2005 The Royal Society
on May 18, 2011rspa.royalsocietypublishing.orgDownloaded from

Proc. R. Soc. A (2005) 461, 305–319doi:10.1098/rspa.2004.1326
Published online 2 November 2004
Effects of contact shape on the scalingof biological attachments
By Ralph Spolenak, Stanislav Gorb,
Huajian Gao and Eduard Arzt
Max Planck Institute for Metals Research,Heisenbergstraße 3, 70569 Stuttgart, Germany
([email protected]; [email protected]; [email protected]; [email protected])
Adhesion of biological systems has recently received much research attention: thesurvival of organisms ranging from single cells and mussels to insects, spiders andgeckos relies crucially on their mechanical interaction with their environments. Forspiders, lizards and possible other ‘dry’ adhesive systems, explanations for adhesionare based on van der Waals interaction, and the adhesion of single-contact elementshas been described by the classical Johnson–Kendall–Roberts (JKR) model derivedfor spherical contacts. However, real biological contacts display a variety of shapesand only rarely resemble a hemisphere. Here, we theoretically assess the influence ofvarious contact shapes on the pull-off force for single contacts as well as their scalingpotential in contact arrays. It is concluded that other shapes, such as a toroidalcontact geometry, should lead to better attachment; such geometries are observed inour microscopic investigations of hair-tip shapes in beetles and flies.
Keywords: adhesion; insects; Johnson–Kendall–Roberts (JKR) theory;Derjaguin–Muller–Toporov (DMT) theory; contact mechanics; surface patterning
1. Introduction
The mechanical interaction of animals with their substratum plays an importantrole in their locomotion. In particular, animals that can cling to vertical walls andeven ceilings have been at the centre of research interest for a long time. Amongthem are insects, such as flies (Bauchhenss 1979), beetles (Stork 1980) and spiders(Homann 1957). The largest animal with pronounced clinging ability is the gecko,whose adhesion has been studied extensively in recent investigations (Hiller 1968;Autumn et al . 2000, 2002; Autumn & Peattie 2002; Russell 2002).
The micromechanisms of adhesion in these animals are still under debate. Whereasadhesion is definitely enhanced by capillary forces in animals producing secretions(flies, beetles), ‘dry’ attachment systems (e.g. in spiders and geckos) are believedto rely entirely on van der Waals forces between the finely structured feet and asubstratum. Based on the action of such surface forces, theories of contact mechanicscan be applied to these biological attachment devices. The most common model foradhesion is the Johnson–Kendall–Roberts (JKR) theory (Johnson et al . 1971), whichbalances the elastic distortion of the contact pair against the reduction in surface
Received 31 October 2003Revised 11 March 2004Accepted 23 March 2004
305c© 2004 The Royal Society
on May 18, 2011rspa.royalsocietypublishing.orgDownloaded from

306 R. Spolenak, S. Gorb, H. Gao and E. Arzt
spherical flat
conical toroidal
filament-like band-like suction cup
(a) (b)
(c) (d)
(e) ( f ) (g)
Figure 1. Shapes of attachment devices in nature and their hypothetical evolution paths (shownas arrows). (a) Bug Pyrrhocoris apterus, smooth pulvillus. (b) Grasshopper Tettigonia viridis-sima, surface of the attachment pad. (c) Fly Myathropa florea, unspecialized hairs on the leg.(d) Fly Calliphora vicina, seta of the pulvilli. (e) Beetle Harmonia axyridis, seta of the secondtarsal segment. (f) Beetle Chrysolina fastuosa, seta of the second tarsal segment. (g) Male beetleDytiscus marginatus, suction cups on the vertical side of the foreleg tarsi.
energy due to the formation of the adhesive contact. Application of this model toliving systems has, despite simplifying assumptions, been surprisingly successful:calculations give adhesion forces of similar magnitude as those found in experiments(Autumn et al . 2002; Sitti & Fearing 2002). As an important consequence of theJKR model, the principle of ‘contact splitting’ has been identified (Arzt et al . 2002;Autumn et al . 2002; Arzt et al . 2003): for dimensional reasons inherent in the JKRanalysis, the adhesion force for a given apparent contact area increases as the totalcontact is split up into ever-finer contact elements. In this way, an experimentallyobserved trend could be explained quantitatively: to be able to hold their own weightwhen adhering to the ceiling, heavier animals must develop finer contact elements,which range in diameter from a few micrometres in small beetles down to 200 nm ingeckos.
In all previous contact-mechanics analyses of biological systems it was assumedthat the terminal elements can be modelled as having hemispherical shape. Micro-scopic studies, however, show that nature provides a variety of contact shapes, mostof which differ substantially from spherical (figure 1). For example, we can see infigure 1d that the contact region of the seta of a fly looks more like a horseshoe thana hemisphere. The objective of the present paper is to analyse the effects of differentshapes on the adhesive force in the framework of contact mechanics. This will bedone with a view to single contacts as well as to the potential of contact splittingfor optimizing adhesion. It will be shown that shape effects are far from negligible:shapes other than spherical can, in principle, purely for geometric reasons, lead toan increase of adhesion forces by more than an order of magnitude. These results
Proc. R. Soc. A (2005)
on May 18, 2011rspa.royalsocietypublishing.orgDownloaded from

Effects of contact shape on the scaling of biological attachments 307
not only improve the understanding of biological attachment systems, they can alsoguide the design of artificial contact structures.
The paper is organized as follows. First, in § 2, fundamental solutions of contactmechanics are compiled and reviewed. Then, in § 3, we discuss how these differentgeometries lend themselves to the principle of contact splitting and interpret thefindings in the light of biological devices.
2. Contact-mechanics fundamentals
Two alternative models dominate the world of contact mechanics: the model ofJohnson–Kendall–Roberts for elastic solids; and the model of Derjaguin–Muller–Toporov (DMT)/Bradley (Derjaguin et al . 1975; Bradley 1932) for stiff solids. Thetransition between the two models was analysed by Tabor (1977) and Maugis (1992)and later by Greenwood (1997). Here we will use Tabor’s parameter µ (Tabor 1977;Greenwood 1997; Pashley 1984), which is given by
µ =R1/3γ2/3
(E∗)2/3ε, (2.1)
where R is the radius of the contacting sphere, γ is the work of adhesion (γ =γ1 + γ2 − γ12, where γ1, γ2 and γ12 are the surface energies of the two solids andtheir interfacial energy, respectively), E∗ is the reduced modulus of the contactingpartners (E∗ = [(1 − ν2
1)/E1 + (1 − ν22)/E2)]−1, Ei and νi are Young’s modulus and
Poisson’s ratio, respectively, of materials 1 and 2), and ε is the interatomic distanceat the closest contact point. Following Tabor, the JKR theory is applicable for µ > 3.This means that the JKR model will be favoured by the comparatively low modulusvalues of biological systems, which may, however, be offset by their small size.
Figure 2 shows the dependence of Tabor’s parameter on contact radius and reducedmodulus. Two regions delineate the range of parameters accessible in nature andthrough technology. Although the boundaries of the parameter space come close tothe transition region between the two theories, in general the JKR theory prevailsfor biological systems. The application of the JKR theory in the subsequent analysisseems, therefore, to be justified. The JKR theory has the additional advantage thatits framework is consistent with linear-elastic fracture mechanics, which is usefulin determining the pull-off forces for certain geometries. It should be noted, how-ever, that for very small or relatively stiff systems the DMT theory may becomeapplicable.
Let us now analyse different contact shapes as shown in figure 3. The JKR theoryassumes that short-range contact forces act only inside the contact area. The contactgeometry differs from the original Hertzian shape without adhesive forces. At thepoint of pull-off the incremental decrease in elastic energy Uelastic is counterbalancedby the incremental increase in surface energy Usurface, such that
dUelastic
dA+
dUsurface
dA= 0, (2.2)
where A is the true contact area of a single contact. When the elastic energy gainedthrough detachment by dA equals the work of adhesion, the contact will becomeunstable and will detach. The geometry before this point is reached has significantconsequences, as will be shown below.
Proc. R. Soc. A (2005)
on May 18, 2011rspa.royalsocietypublishing.orgDownloaded from

308 R. Spolenak, S. Gorb, H. Gao and E. Arzt
µ = 105
µ = 104
µ = 10−1
µ = 103
µ = 102
µ = 10 µ = 1
JKR
DMT
bio-attachment
artificial systems10−2
1
102
104
106
E∗
(MPa
)
10−2 1 102 104 106
R (µm)
Figure 2. The transition between the DMT model and the JKR model: Tabor’s parameter µ asa function of contact radius R and effective modulus E∗ (γ = 0.05 J m−2, ε = 2 A). The solidline indicates the transition between the DMT and the JKR theory. The two ellipses delineateroughly the range of parameters in natural attachment devices and the range accessible byartificial attachment systems.
Alternatively, one can apply the principles of linear-elastic fracture mechanics. Forplane strain, the stress-intensity factor KI is related to the energy release rate G inthe following way:
G =1
2E∗ K2I . (2.3)
Unstable crack growth, which corresponds to detachment of the contact, occurs whenthe energy release rate G exceeds a critical value Gc. In the absence of rate effects,Gc can be equated to the work of adhesion γ.
In the following compilation of mathematical solutions, one of the two equivalentapproaches will be taken, depending on the geometry in question.
(a) Sphere
In the JKR model the contact of a spherical element of radius R with a semi-infinite half-space is easily treated by the linear-elastic fracture-mechanics approach.According to the Hertz theory, the load P1 is given as a function of the contactradius a by
P1 =4πE∗a3
3R. (2.4)
With the addition of adhesive forces, the stress singularity at the edge of the contactcorresponds to a stress-intensity factor KI:
KI =Pa
2a√
πa, (2.5)
Proc. R. Soc. A (2005)
on May 18, 2011rspa.royalsocietypublishing.orgDownloaded from

Effects of contact shape on the scaling of biological attachments 309
2R
2a
2R 2R
2R
2R
h α
F
2r
(a) (b) (c)
(d) (e)
Figure 3. Side and plan view of contact shapes studied: (a) sphere, (b) flat punch/verticalcylinder, (c) torus/horizontal cylinder, (d) suction cup, (e) elastic band (in each case, the shadedarea denotes contact).
where Pa is the load introduced by the adhesive forces. By equating the energyrelease rate G with the work of adhesion γ and using equation (2.3), the net load Pis obtained as
P = P1 − Pa =4πE∗a3
3R−
√8πE∗γa3. (2.6)
The pull-off force Pc is derived from the condition dP/da = 0, giving
Pc = −32πγR. (2.7)
As is common in contact mechanics, the minus sign is used to denote tensile forces.Note that the pull-off force, for this case only, is independent of the elastic propertiesof the two contacting surfaces.
(b) Horizontal cylinder and torus
The case of a cylinder of radius R in line contact with a semi-infinite half-spacecan be treated as follows (Chaudhury et al . 1996). According to the Hertz theorythe load P1l per unit length is
P1l =πE∗a2
4R. (2.8)
In the presence of adhesive forces the stress singularity at the edge of the contactcorresponds to a stress-intensity factor KI:
KI =Pal√πa
, (2.9)
where Pal is the load per unit length introduced by adhesive forces. By equating theenergy release rate G with the work of adhesion γ and using equation (2.3), the net
Proc. R. Soc. A (2005)
on May 18, 2011rspa.royalsocietypublishing.orgDownloaded from

310 R. Spolenak, S. Gorb, H. Gao and E. Arzt
load Pl per unit length is obtained as
Pl = P1l − Pal =πE∗a2
4R−
√2πE∗γa. (2.10)
Differentiation leads to a pull-off force Pcl per unit length of
Pcl = −32(πγ2E∗R/2)1/3. (2.11)
From this solution, the pull-off force for a slender torus can be approximated. Assum-ing r � R, we can model this case by a cylinder of radius r and length 2Rπ. Thenthe pull-off force Pc is
Pc = −3(πE∗γ2r/2)1/3πR. (2.12)
For scaling purposes one can assume self-similarity with, for example, r = R/10,which leads to
Pc = −1.1(πE∗γ2)1/3πR4/3. (2.13)
It is seen that, unlike for the sphere, the results for cylindrical geometries do dependon the elastic properties of the materials involved.
(c) Vertical cylinder (flat punch)
For comparison, we include in our analysis the solution for a flat punch, i.e. acylinder standing upright against a semi-infinite half-space. We follow the fracture-mechanics approach of Kendall (1971) and Maugis (1999). If the modulus of thecylinder is higher than that of the half-space, the geometry resembles that for anotched cylindrical bar. Using Paris & Sih’s (1965) stress-intensity factor,
KI =P
2a√
πa, (2.14)
and applying the same procedure as before and assuming that a = R results in apull-off force of
Pc = −√
8πE∗R3γ. (2.15)
The assumption that the contact radius a is equal to the punch radius R is onlyapplicable when the punch has a higher modulus than the half-space. For an elasticpunch on a stiff half-space the contact radius needs to be smaller than the punchradius, introducing a flaw size (R − a). Following Bueckner (1973), equation (2.14)is then modified by a parameter a < 1:
KI =P
2a√
πaβ. (2.16)
Thus the difference (R − a) corresponds to the initial crack length for the fracture-mechanics problem. If one assumes a = 0.8R, the pull-off force Pc is reducedby ca. 10% and equation (2.15) can still be used as a first-order approximation.
Proc. R. Soc. A (2005)
on May 18, 2011rspa.royalsocietypublishing.orgDownloaded from

Effects of contact shape on the scaling of biological attachments 311
(d) General axisymmetric profiles
Yao & Gao (2004) have recently derived the general JKR solution for contactbetween a half-space and an axisymmetric object described by the polynomial:
h(r) =rq
Rq−1 , (2.17)
where r is the horizontal distance to the centre of the shape along the flat surface, his the distance between the object and the surface of the half-space, and q is a shapeparameter. This expression contains several special cases: a sharp cone (q = 1), aparabolic function approximating the shape of a sphere (q = 2), and a flat punch(q → ∞). For the pull-off force Pc the following proportionalities are predicted:
Pc ∝ R3(q−1)/(2q−1)(E∗)(q−2)/(2q−1)γ(q+1)/(2q−1). (2.18)
We note that the dependence of Pc on R is always positive (for q > 1), whereas thedependence on E∗ is negative for 1 � q < 2. For q → ∞, the pull-off force scales as
Pc ∝ R3/2(E∗)1/2γ1/2. (2.19)
which shows exactly the same scaling as equation (2.15). For q = 2 we get thewell-known JKR expression:
Pc ∝ Rγ. (2.20)
It is readily seen from equation (2.18) that only in this case is the pull-off forceindependent of the modulus.
(e) Film peeling: contact of an elastic tape
In this case we address the problem from an energetic point of view followingKendall (1975). The work done in peeling an elastic tape of modulus E, width 2Rand thickness h over a distance δx is
Gδx2R = f(1 − cos α)δx + FδxF
4ERh, (2.21)
where F is the force acting in the direction of the tape and α is the peel angle. Thesecond term on the right-hand side of equation (2.21) is the work done by the forceF minus the elastic energy stored in the peeled-off segment of the tape. Equating Gto the work of adhesion γ yields a quadratic equation in Fc:
F 2c + 4Fc(1 − cos α)ERh − 8ER2hγ = 0. (2.22)
Solving equation (2.22) and taking the component of Fc normal to the surface, i.e. thepull-off force Pc = −Fc sin α, gives the following relation:
Pc = −2γRg(α, λ). (2.23)
The function g is defined as
g(α, λ) =2 sin α
√(1 − cos α)2 + 2λ + (1 − cos α)
, (2.24)
where α is the peel-off angle and λ is defined as γ/Eh. Note that the peeling tapeleads to a pull-off force that, like the JKR solution for the sphere, depends linearlyon the size parameter R. A more detailed analysis including nonlinear elasticity isin preparation.
Proc. R. Soc. A (2005)
on May 18, 2011rspa.royalsocietypublishing.orgDownloaded from
312 R. Spolenak, S. Gorb, H. Gao and E. Arzt
50 µm
50 nm 1.5 µm
10 µm
flat punch torus suction sphere
1.5 µm
104
102
1
10−2
10−2 1 10210−1 10 103 10−2 1 10210−1 10 103
radius (µm)
pull
off
forc
e (µ
N)
radius (µm)
(a) (b)
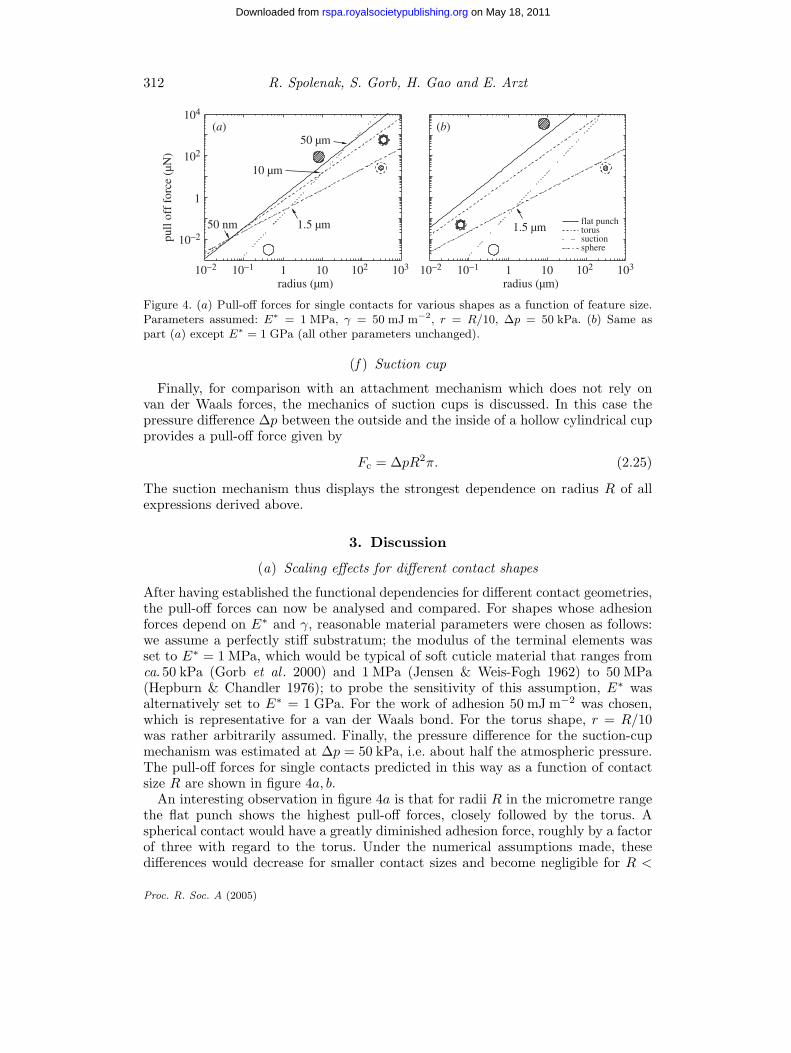
Figure 4. (a) Pull-off forces for single contacts for various shapes as a function of feature size.Parameters assumed: E∗ = 1 MPa, γ = 50 mJ m−2, r = R/10, ∆p = 50 kPa. (b) Same aspart (a) except E∗ = 1 GPa (all other parameters unchanged).
(f ) Suction cup
Finally, for comparison with an attachment mechanism which does not rely onvan der Waals forces, the mechanics of suction cups is discussed. In this case thepressure difference ∆p between the outside and the inside of a hollow cylindrical cupprovides a pull-off force given by
Fc = ∆pR2π. (2.25)
The suction mechanism thus displays the strongest dependence on radius R of allexpressions derived above.
3. Discussion
(a) Scaling effects for different contact shapes
After having established the functional dependencies for different contact geometries,the pull-off forces can now be analysed and compared. For shapes whose adhesionforces depend on E∗ and γ, reasonable material parameters were chosen as follows:we assume a perfectly stiff substratum; the modulus of the terminal elements wasset to E∗ = 1 MPa, which would be typical of soft cuticle material that ranges fromca. 50 kPa (Gorb et al . 2000) and 1 MPa (Jensen & Weis-Fogh 1962) to 50 MPa(Hepburn & Chandler 1976); to probe the sensitivity of this assumption, E∗ wasalternatively set to E∗ = 1 GPa. For the work of adhesion 50 mJ m−2 was chosen,which is representative for a van der Waals bond. For the torus shape, r = R/10was rather arbitrarily assumed. Finally, the pressure difference for the suction-cupmechanism was estimated at ∆p = 50 kPa, i.e. about half the atmospheric pressure.The pull-off forces for single contacts predicted in this way as a function of contactsize R are shown in figure 4a, b.
An interesting observation in figure 4a is that for radii R in the micrometre rangethe flat punch shows the highest pull-off forces, closely followed by the torus. Aspherical contact would have a greatly diminished adhesion force, roughly by a factorof three with regard to the torus. Under the numerical assumptions made, thesedifferences would decrease for smaller contact sizes and become negligible for R <
Proc. R. Soc. A (2005)
on May 18, 2011rspa.royalsocietypublishing.orgDownloaded from

Effects of contact shape on the scaling of biological attachments 313
Table 1. Functional dependencies for various contact shapes
(The exponents s, m, k, r and p (as defined in the first column) are listed for different contactshapes and mechanisms. P ′
c is the total pull-off force after splitting one contact into n singlecontacts; NA is the areal density of contacts; and m is the mass of an object adhering to theceiling.)
elastic flat suctionhemisphere torus tape punch cup general
Pc ∝ R5 1 43 1 3
2 23(q − 1)2q − 1
Pc ∝ Em 0 13 0 1
2 0q − 22q − 1
Pc ∝ γk 1 23 1 1
2 0q + 12q − 1
P ′c = nrPc
12 , 1a 1
3 , 12a 1
214 0
(q + 1)2(2q − 1)
NA ∝ mp 23 , 1
3a 1, 2
3a 2
343 ∞ 2(2q − 1)
3(q + 1)
aCurvature invariance (see analysis by Arzt et al . (2003)).
100 nm. These differences are somewhat magnified when using the second value ofE∗ = 1 GPa (figure 4b).
Because of the different size dependencies, the order of the three contact shapes isreversed below a critical radius R0, which is defined by the intersection of the linesin figure 4. Equating the pull-off force for the sphere (equation (2.7)) with that ofthe torus (equation (2.13)) results in
R0 � 0.80γ
E∗ . (3.1)
The same result is obtained for the crossover between the flat-punch solution (equa-tion (2.15)) and that of the sphere, except for a numerical pre-factor of 0.88. Thismeans that for very soft cuticle material, for instance with E∗ = 50 kPa, the cross-over radius R0 will be of the order of 1 µm.
Further details of the scaling behaviour are displayed in table 1. The value s, whichreflects the slope in figure 4, determines the potential for improving the pull-off forceby scaling the contact size. A larger s value favours a particular geometry at largesize; this is clearly visible for the suction-cup mechanism (s = 2), which rapidlyloses its effectiveness below ca. 10 µm compared with other mechanisms (figure 4a).In terms of functional relationship, the elastic tape scales like the sphere; the pull-offforce, however, depends on the peeling angle α.
Table 1 also summarizes the dependencies on E∗ and γ. Whereas the sphericalcontact, the elastic tape (assuming that the modulus is not too low, λ � 1 inequation (2.24)) and the suction cup are independent of elastic properties, moderatedependencies are present for the torus and the flat punch. The work of adhesion enterslinearly into the pull-off-force expressions for the sphere and the tape, whereas thedependencies for the other shapes are weaker.
After considering the scaling of single contacts, we now turn to the question ofhow contact splitting will affect the pull-off forces (see the last line in table 1 and
Proc. R. Soc. A (2005)
on May 18, 2011rspa.royalsocietypublishing.orgDownloaded from

314 R. Spolenak, S. Gorb, H. Gao and E. Arzt
tota
l for
ce (
N)
human attachment human attachment
punchtorussphere
10−2
1
102
104
106 10103105107
1 102 105 106 108 1010 1012 1 102 105 106 108 1010 1012
10103105107
(a) (b)
number of contacts number of contacts
contact size (nm) contact size (nm)
Figure 5. (a) Comparison of pull-off forces for different shapes in terms of contact-splittingefficiency. The total apparent contact area is 100 cm2. Thin dashed lines: contact-invariantscaling (Arzt et al . 2003). Note that contact-invariant scaling must never cross the flat-punchcurve. The horizontal dashed black line indicates the force necessary for human attachment.Parameters: E∗ = 1 MPa, γ = 0.05 J m−2. (b) Same as part (a) except E∗ = 100 MPa (all otherparameters unchanged).
figure 5). Using the JKR formalism, it was recently shown (Arzt et al . 2003) thatsplitting a macroscopic contact into n contact elements, the total apparent contactarea remaining unchanged, increases the pull-off force to
P ′c = nrPc, (3.2)
where Pc is the pull-off force of the macroscopic contact without splitting. Note thatr is a measure for the efficiency of contact splitting: larger values result in highergains in pull-off force. Table 1 lists the corresponding r values for the different shapesand mechanisms.
It is readily seen that the spherical contact, especially under curvature-invariantconditions, offers the best splitting efficiency (r = 1), followed by the elastic tape (r =12) and the torus (r = 1
3 or 12). The flat punch does not respond favourably (r = 1
4).The adhesion of suction cups cannot be improved by contact splitting (r = 0), exceptfor better defect tolerance in the presence of dust particles or similar flaws.
In our earlier analysis, we also considered the areal density NA of spherical terminalelements required to support an animal of mass m hanging from the ceiling. Undercertain simplifying assumptions (e.g. spherical shape, self-similar scaling of weightand foot area) the following relationship was obtained:
NA = fmp, (3.3)
where f is approximately constant and p = 23 for self-similar scaling and p = 1
3 forcurvature invariance of the terminal elements. The p values for the other shapes andmechanisms are also included in table 1. Note that p is an inverse measure of thesplitting efficiency, with smaller values giving larger effects.
As expected, the most efficient shape for contact splitting is again the curvature-invariant spherical contact (p = 1
3), followed by the curvature-invariant torus or the
Proc. R. Soc. A (2005)
on May 18, 2011rspa.royalsocietypublishing.orgDownloaded from

Effects of contact shape on the scaling of biological attachments 315
self-similar spherical contact (p = 23). The scaling of contact-invariant shapes stops
when the contact radii R for the sphere and r for the torus become comparable withthe feature size. The self-similar torus exhibits a scaling law with p = 1, implyinga reduced efficiency for contact splitting; however, the toroidal single contact has arelatively high pull-off force to start with. The flat punch with a 4
3 scaling law, onthe other hand, in contrast to its high single-contact pull-off force, would be veryinefficient in contact splitting.
These relationships are illustrated in figure 5. There the total pull-off force isplotted as a function of the number of contact elements in an apparent contact areaof 100 cm2. In figure 5a, the values for E∗ and γ are the same as in figure 4a. Shapeswith lower pull-off forces as single contacts show higher contact-splitting efficiencyso that the forces converge at contact sizes of ca. 100 nm. If one wanted to providethe force necessary for attachment of a weight comparable with a human being(e.g. 10 000 N, for safety), the flat punch or the torus would still be the preferredshapes. Note that over a small variation in contact size the splitting efficiency canbe increased by using the criterion of curvature invariance. This changes the slopein figure 4 from 1
2 to 1 for the spherical contact and from 13 to 1
2 for the toroidalcase. However, the limit of this strategy is reached when the contacts have beenpressed flat, i.e. at the line for the flat punch. Figure 5b shows the same plot forE∗ = 100 MPa. Note that as the flat punch and the torus depend on the modulustheir pull-off forces are increased significantly.
The apparent advantage of the flat-punch geometry over all other shapes requiresan important qualification: more than the other shapes, the flat punch requires per-fect matching between the punch and the surface of the substratum. In nature or intechnical applications surfaces are far from perfect and roughness or dirt particleswill significantly reduce the theoretical pull-off forces. The flat punch is thereforenot a ‘robust’ contact geometry. Note that the reduction in pull-off force dependson the actual contact area and the effective modulus. Larger contact area and largereffective modulus will result in bigger reductions. This implies that a flat-punch con-tact would be feasible only if very finely structured and made of very-low-modulusmaterial.
(b) Application to biological systems
Cuticle outgrowths of various shapes in insects have evolved to fulfil several func-tions including attachment (Gorb 2001). Terminal elements of attachment devicesrange from pointed tips to spatulae, horseshoe-like structures and suction cups (fig-ure 1). Hemispheres and flat punches can also be found as contact shapes, but thesestructures are not homologous to protuberances mentioned above, i.e. they did notevolve out of the same structures and materials, but appeared independently in dif-ferent animal groups.
It is remarkable that some of the theoretical conclusions on contact shape areindeed reflected in the biological attachment systems. The flat punch, for example, ishardly found in nature. Only the contact elements of the grasshopper do bear someresemblance to it; they consist of material with a very low modulus of 30–70 kPa(Gorb et al . 2000), in keeping with the theoretical requirement stated above. Echi-noids have tube feet reminiscent of a flat punch, but their adhesion is ensured by atwo-component chemical adhesive (Flammang 1996).
Proc. R. Soc. A (2005)
on May 18, 2011rspa.royalsocietypublishing.orgDownloaded from

316 R. Spolenak, S. Gorb, H. Gao and E. Arzt
The suction-cup mechanism occurs in nature, for example, in water beetles (fig-ure 1g) and in mites (Gorb 2001). In all of these cases the size of the cups never liesbelow ca. 10 µm. This is in agreement with the theoretical finding (figure 4) that thesuction mechanism rapidly loses its effectivity for smaller sizes. It can, however, bea potent attachment mechanism in the ‘macroscopic’ world, as is demonstrated bytechnical and household items which rely on underpressure for their attachment.
This leaves the torus-like structures, which are found in attachment devices ofthe fly and the beetle (figure 1). In terms of contact mechanics the torus is a pre-ferred shape as it combines relatively high absolute forces with an appreciable scalingpotential. Our study has shown that for very small contact elements the influence ofthe shape diminishes. The hemisphere and the elastic tape (which have been foundto scale identically) then become efficient contact geometries. Cases in point are thesetulae in anthropods (e.g. spiders) and the spatulae in lizards—the smallest terminalcontact elements known.
Finally, it has to be borne in mind that in the biological world adhesion may notonly be the result of van der Waals-type surface forces as discussed here. Althoughthere is evidence (Autumn et al . 2002) that capillary forces are not important ingecko adhesion, their importance in this and other systems cannot be ruled outconclusively. In addition, animals such as the grasshopper, the fly and the beetleproduce secretions in their attachment structures, whereas spiders and lizards donot. The function of the secretion to promote adhesion is not entirely understood. Itis, however, reassuring that capillary forces in spherical contacts scale linearly withthe diameter (Israelachvili 1992), which is identical to the scaling behaviour predictedby the JKR model for van der Waals interaction. The influence of contact shape oncapillary adhesion requires further investigation. Another complication is the well-known fact that natural materials in attachment structures also exhibit viscoelasticproperties (Gorb et al . 2000). These properties are expected to have a significanteffect on the kinetics of adhesion and are the subject of further studies.
(c) Implications for artificial adhesive systems
Several preliminary conclusions may be drawn in connection with the design ofartificial contact elements. The present paper suggests that the shape of contactelements, which appears to have been largely disregarded in the past, can be animportant consideration. The toroidal contact is certainly worth investigating fur-ther as it exhibits high single-contact adhesion in conjunction with relatively highpotential for contact splitting. It is also important to note that in this case the pull-off force depends on the effective modulus, in contrast with the predictions of theJKR theory for a sphere. Therefore, elastic and maybe even viscoelastic properties ofcontacts will play an important role. In general, higher moduli would be beneficial aslong as the contact is still compliant enough not to be dominated by local roughness(Kendall 1971; Persson 2003). Contact splitting remains a primary strategy: it notonly increases the total pull-off forces but also improves the defect tolerance, i.e. thefailure of a single contact element does not affect the adhesion of the ensemble signifi-cantly. Furthermore, if single contacts are attached to very compliant pillars or hairs,the attachment device can adhere to arbitrary surfaces at the cost of little elasticenergy stored (Persson 2003; Sitti & Fearing 2002; Geim et al . 2003; Peressadko &Gorb 2004).
Proc. R. Soc. A (2005)
on May 18, 2011rspa.royalsocietypublishing.orgDownloaded from

Effects of contact shape on the scaling of biological attachments 317
Overall, the present study underscores the need to understand the several variablesinvolved in forming a dry adhesive contact. The optimum contact requires a balanceof contact size, shape and material, all of which are dependent on one another.The principles of this optimization procedure, as inspired by nature and refined bymechanical modelling, are the subject of current work in progress.
4. Conclusions
This paper has considered the effects of shape and of scaling on adhesion of dry sys-tems dominated by van der Waals-type surface forces. Hence the modelling is applica-ble to model systems and to natural systems that are known to rely on van der Waalsinteraction, i.e. spiders and lizards. The following conclusions can be drawn.
(i) Investigation of Tabor’s parameter shows that most natural attachment devicescan be treated in the contact-mechanics framework of the Johnson–Kendall–Roberts model. Only very small and stiff contact elements may require adescription by alternative models.
(ii) Besides contact size, contact shape can exert a strong influence on adhesion.Big differences arise for comparatively larger contact elements of relativelystiff material. At very small contact sizes (several tens of nanometres) and lowmodulus (below 1 MPa) the influence of shape becomes negligible.
(iii) The best shape for perfectly smooth surfaces is, unsurprisingly, the flat punch,followed by the torus and the sphere. However, the punch is sensitive to rough-ness and dirt effects and does not provide a ‘robust’ contact. The suction-cupmechanism, by comparison, can only compete at large contact sizes and isinefficient in the sub-micrometre regime.
(iv) The total pull-off force can be improved by splitting the contact into manyfiner contacts of identical shape. The benefit of this ‘contact-splitting’ strategydepends also on the shape. In general, lower absolute forces have a highersplitting potential.
(v) Natural attachment systems seem to reflect some of the theoretical findingsof this study. Very compliant contact elements (e.g. in the grasshopper) haveshapes resembling a flat punch, whereas stiffer contact elements exhibit torus-like shapes (e.g. in insects) or band-like structures (e.g. the gecko). The combi-nation of optimum size and shape of artificial contact elements should allow thegeneration of higher contact forces than encountered so far in natural systems.
References
Arzt, E., Enders, S. & Gorb, S. 2002 Towards a micromechanical understanding of biologicalsurface devices. Z. Metallk. 93, 345–353.
Arzt, E., Gorb, S. & Spolenak, R. 2003 From micro to nano contacts in biological attachmentdevices. Proc. Natl Acad. Sci. USA 100, 10 603–10 606.
Autumn, K. & Peattie, A. M. 2002 Mechanisms of adhesion in geckos. Integr. Compar. Biol.42(6), 1081–1090.
Autumn, K., Liang, Y. A., Hsieh, S. T., Zesch, W., Wai, P. C., Kenny, T. W., Fearing, R. &Full, R. J. 2000 Adhesive force of a single gecko foot-hair. Nature 405, 681–685.
Proc. R. Soc. A (2005)
on May 18, 2011rspa.royalsocietypublishing.orgDownloaded from

318 R. Spolenak, S. Gorb, H. Gao and E. Arzt
Autumn, K., Sitti, M., Liang, Y. C. A., Peattie, A. M., Hansen, W. R., Sponberg, S., Kenny,T. W., Fearing, R., Israelachvili, J. N. & Full, R. J. 2002 Evidence for van der Waals adhesionin gecko setae. Proc. Natl Acad. Sci. USA 99, 12 252–12 256.
Bauchhenss, E. 1979 Die Pulvillen von Calliphora erythrocephala Meig. (Diptera, Brachycera)als Adhasionsorgane. Zoomorphologie 93, 99–123.
Bradley, R. S. 1932 The cohesive force between solid surfaces and the surface energy of solids.Phil. Mag. 13, 853–862.
Bueckner, H. F. 1973 Field singularities and related integral representations (crack analysis). InMechanics of fracture, vol. 1. Methods of analysis and solutions of crack problems, pp. 239–314. Groningen: Noordhoff.
Chaudhury, M. K., Weaver, T., Hui, C. Y. & Kramer, E. J. 1996 Adhesive contact of a cylindricallens and a flat sheet. J. Appl. Phys. 80(1), 30–37.
Derjaguin, B. V., Muller, V. M. & Toporov, Yu. P. 1975 Effect of contact deformations on theadhesion of particles. J. Colloid Interface Sci. 53, 314–326.
Flammang, P. 1996 Adhesion in echinoderms. Echinoderm Stud. 5, 1–60.Geim, A. K., Dubonos, S. V., Grigorieva, K. S., Novoselov, K. S., Zhukov, A. A. & Shapoval,
S. Y. 2003 Microfabricated adhesive mimicking gecko foot-hair. Nat. Mater. 2(7), 461–463.Gorb, S. N. 2001 Attachment devices of insect cuticle, p. 18. Deventer: Kluwer.Gorb, S. N., Jiao, Y. & Scherge, M. 2000 Ultrastructural architecture and mechanical properties
of attachment pads in Tettigonia viridissima (Orthoptera Tettigoniidae). J. Compar. Physiol.A186, 821–831.
Greenwood, J. A. 1997 Adhesion of elastic spheres. Proc. R. Soc. Lond. A453, 1277–1297.Hepburn, H. R. & Chandler, H. D. 1976 Material properties of arthropod cuticles: the arthrodial
membranes. J. Compar. Physiol. B100, 177–198.Hiller, U. 1968 Untersuchungen zum Feinbau und zur Funktion der Haftborsten von Reptilien.
Z. Morphol. Tiere 62, 307–362.Homann, H. 1957 Haften Spinnen an einer Wasserhaut? Naturwissenschaften 44(11), 318–319.Israelachvili, J. N. 1992 Intermolecular and surface forces, 2nd edn, p. 332. Academic.Jensen, M. & Weis-Fogh, T. 1962 Biology and physics of locust flight. V. Strength and elasticity
of locust cuticle. Phil. Trans. R. Soc. Lond. B245(721), 137–169.Johnson, K. L., Kendall, K. & Roberts, A. D. 1971 Surface energy and contact of elastic solids.
Proc. R. Soc. Lond. A324, 301–320.Kendall, K. 1971 The adhesion and surface energy of elastic solids. J. Phys. D4, 1186–1195.Kendall, K. 1975 Thin film peeling—the elastic term. J. Phys. D4, 1450–1452.Maugis, D. 1992 Adhesion of spheres: the JKR–DMT transition using a Dugdale model. J.
Colloid Interface Sci. 150, 243–269.Maugis, D. 1999 Contact, adhesion and rupture of elastic solids. In Solid state science, p. 234.
Springer.Paris, P. C. & Sih, G. C. 1965 Stress analysis of cracks. In Fracture toughness and testing
applications. ASTM STM 381, 30–83.Pashley, M. D. 1984 Further consideration of the DMT model for elastic contact. Colloids Surf.
12, 69–77.Peressadko, A. & Gorb, S. 2004 When less is more: experimental evidences for adhesion enhance-
ment by division of contact area. J. Adhes. 80(4), 247–261.Persson, B. N. J. 2003 On the mechanism of adhesion in biological systems. J. Chem. Phys.
118(16), 7614–7621.Russell, A. P. 2002 Integrative functional morphology of the gekkotan adhesive system (Reptilia:
Gekkota). Integr. Compar. Biol. 42(6), 1154–1163.Sitti, M. & Fearing, R. S. 2002 Nanomolding based fabrication of synthetic gecko foot-hairs. In
Proc. 2nd IEEE Conf. on Nanotechnology, Piscataway, NJ, USA, 2002, pp. 137–140. Piscat-away, NJ: IEEE Press.
Proc. R. Soc. A (2005)
on May 18, 2011rspa.royalsocietypublishing.orgDownloaded from

Effects of contact shape on the scaling of biological attachments 319
Stork, N. E. 1980 Experimental analysis of adhesion of Chrysolina polita (Chrysomelidae,Coleoptera) on a variety of surfaces. J. Exp. Biol. 88, 91–107.
Tabor, D. 1977 Surface forces and surface interactions. J. Colloid Interface Sci. 58, 2–13.Yao, H. & Gao, H. 2004 Shape insensitive optimal adhesion of nanoscale fibrillar structures.
Proc. Natl Acad. Sci. USA 101(21), 7851–7856.
Proc. R. Soc. A (2005)
on May 18, 2011rspa.royalsocietypublishing.orgDownloaded from




















