
The Intersection of Well-Being and Advice - Fidelity Clearing ...

REGULAR ARTICLE
Effect of woody vegetation clearing on nutrient and carbonrelations of semi-arid dystrophic savanna
Susanne Schmidt & Rebecca E. Lamble &
Rod J. Fensham & Ilyas Siddique
Received: 2 June 2009 /Accepted: 11 November 2009 /Published online: 25 November 2009# Springer Science + Business Media B.V. 2009
Abstract Throughout the savanna biome, woodyvegetation is cleared to increase productivity ofherbaceous pasture. While clearing can result inincreased pasture production of semi-arid dystrophicsavannas in the short term, it is uncertain whetherproduction is sustained in the long term. There isinsufficient knowledge of how clearing affects soilnutrient and organic carbon (SOC) stocks. Usingcleared-uncleared site pairs, we evaluated techniquesfor time-integrated assessment of nutrient and carbonrelations in Australian savanna. Short-term in situresin incubation showed that soil at cleared sites had ahigher time-integrated availability of ammonium andnitrate, indicating that nitrogen (N) may turn overfaster and/or is taken up slower at cleared sites thanuncleared savanna. Nitrate and ammonium availabil-ity was approximately 2-fold higher in spring than insummer, likely due to greater uptake and/or loss ofnitrate during summer rains. Nitrate was a prominent
N source for evergreen trees, especially beforesummer rain, pointing to a role of trees as permanentN sinks. Stable isotope signatures of soil andvegetation indicate that N input occurs via N2 fixingmicrobiotic crusts and Acacia species. 30 years afterclearing, SOC contained more C4 grass-derivedcarbon than uncleared savanna, but this shift in Csource was not associated with the net C gain oftenobserved in grasslands. Interactions between alterednutrient and C relations and composition of theunderstorey should be assessed in context ofintroduced buffelgrass (Cenchrus ciliaris) whichhad higher macronutrient concentrations than nativegrasses. Heterogeneity of the studied soils highlightsthe need for replication at several spatial scales toinfer long-term dynamics with space-for-time chro-nosequences. We conclude that the techniquespresented here are useful for gaining knowledge ofthe biogeochemical processes governing savannasand the systems that result from clearing.
Keywords 15-N natural abundance . 13-C naturalabundance .Eucalyptus savanna .Macronutrients .
Soil organic carbon . Soil nitrogen . Tree clearing
Introduction
Tropical savannas cover ~15% of the Earth’s landsurface and are complex landscapes where grasses,herbs, shrubs and trees co-exist in dynamic equilib-
Plant Soil (2010) 331:79–90DOI 10.1007/s11104-009-0233-5
Responsible Editor: Wim van der Putten.
S. Schmidt (*) : R. E. Lamble :R. J. Fensham : I. SiddiqueSchool of Biological Sciences,The University of Queensland,Brisbane, QLD 4071, Australiae-mail: [email protected]
R. J. FenshamQueensland Herbarium,Environmental Protection Agency,Mt Coot-tha Road,Toowong, QLD 4066, Australia

rium. Savannas occupy 20% of the Australiancontinent and extensive clearing of savannas hasresulted from widespread pastoralism (Whitehead etal. 2000). Worldwide the integrity of tropical savan-nas is threatened by fragmentation and transformationto agriculture and forestry, and clearing rates inAustralia have been ~400,000 ha per year (Wilson etal. 2002). Most woody vegetation clearing (“clearing”is used hereafter) occurs in semi-arid dystrophicsavanna (Fairfax and Fensham 2000; Wilson et al.2002). There is evidence that removal of trees fromthese savannas increases pasture production in theshort-term (<10 years) (Scanlan 2002), but long-termconsequences of clearing are largely unknown(Schmidt and Lamble 2002). In semi-arid Eucalyptussavanna and Acacia woodland, pasture productiondecreased significantly in the long term (~30 years)(Kaur et al. 2005).
Interactions between woody and herbaceous sa-vanna taxa are complex and evidence exists forfacilitation and competition (Scholes and Archer1997). One possible outcome is that sustainablepasture production results from clearing if pasturespecies maintain ecosystem function. Conversely,unsustainable land use and degradation will resultfrom clearing if woody vegetation is integral forecosystem function. We used the existing configura-tion of the landscape to examine processes associatedwith clearing. Four cleared and adjacent unclearedsites were studied within a single land type in theDesert Upland Bioregion, one of 15 biodiversityhotspots in Australia (Commonwealth of Australia(2008) http://www.environment.gov.au/biodiversity/hotspots/national-hotspots.html#1). Clearing of Euca-lyptus dominated savannas in the Desert UplandsBioregion is not recommenced since inherently lowsoil fertility and low water availability, combined withcertain management practices, hinder successfulmaintenance of pastures (Turner 1978; Thompson etal. 2003).
Nutrient relations can be quantified by determiningpool sizes and rates of transformation. We contrastwidely used measures of total soil C, N and soil-extractable macronutrients with field incubation ofion exchange resin. In situ resin incubations allowtime-integrated quantification of N turnover in soil todetect processes involved in nutrient loss versus on-site retention (Siddique et al. 2008). Soil N relationswere examined because N is likely to (co)-limit plant
productivity in the dystrophic soils of the DesertUpland, and because N is linked to soil C storage(Neff et al. 2002). We measured macronutrientcontent of grasses to determine if soil nutrientavailability is mirrored by vegetation, since availabil-ity of soil macronutrients increased in the first decadeafter clearing but decreased in the longer term(Sangha et al. 2005). Nitrogen composition of xylemsap was analysed to discern the role of trees fornutrient capture. Nitrate is transported in xylem sapfrom roots to shoots only and is therefore a marker forN uptake from soil (Schmidt and Stewart 1998,2003).
Tropical and subtropical ecosystems contain ~30%of the global SOC pool (Dalal and Carter 2000), anddeforestation is associated with declining SOC (Powerset al. 2004). Reasons for loss of SOC include reducedC input into soil and increased decomposition rates(Guo and Gifford 2002). Stable isotope ratios wereevaluated as integrators of ecosystem N and C cycles.δ15N signals are linked to plant N relations (Robinson2001), and an open N cycle commonly results in 15N-enrichment of soil and vegetation (Högberg 1997).Different pathways of CO2-fixation have distinct δ13Cvalues, and indicate the contribution of C3-trees andC4-grasses to SOC (Baldesdent and Mariotti 1996).
The suite of techniques evaluated here includes time-integrated processes over short (resin incubation) andlonger (stable isotope ratios) time scales and ecophys-iological processes (nutrient contents of vegetation) togain insights into biogeochemical processes underlyingsavanna and the systems that result from clearing. Thestudied sites were cleared 1.5, 2, 10 and 30 years agoand each cleared site had an adjacent, uncleared site forcomparison. Apart from clearing, sites had comparablemanagement including a long grazing history.We aimedto evaluate time-integrated techniques in combinationwith ecophysiological markers for assessing savannafunction in context of existing knowledge of the effectsof clearing.
Materials and methods
Study area
The Desert Uplands biogeographical region covers~75,000 km2 (Fig. 1, Thackway and Creswell 1995)and is characterised by low altitude Tertiary plateau
80 Plant Soil (2010) 331:79–90

with well-developed sand-sheets. Climate is semi-aridwith highly variable seasonal rainfall mostly fromNovember to March. Mean annual rainfall at Jericho inthe southern Desert Uplands is 516 mm with 71 %falling in summer between October and March (Clewettet al. 1994). Summer and winter daily maximumtemperatures are 30–35°C and 20–25°C, respectively(Fairfax and Fensham 2000). The area is prone tooccasional severe drought (Fensham and Holman1999). Each of four cleared-uncleared site pairs waslocated in Eucalyptus melanophloia open woodlandwith a sparse canopy on undulating sand plains(Queensland regional ecosystem 10.5.5a; EPA 2007;http://www.epa.qld.gov.au/media/nature_conservation/biodiversity/desert_uplands/Factsheets/100505.htm).This regional ecosystem (hereafter ‘E. melanophloiasavanna’) dominates large areas of the Desert Uplandsbioregion and harbours relatively high plant speciesdiversity and high to very high animal species diversity(Thompson et al. 2003). Besides dominant Eucalyptusmelanophloia, common trees were Eucalyptus popul-nea, Acacia species and Proteaceae. Common nativegrasses were sampled including Aristida jerichoensis,Bothriochloa ewartiana, Heteropogon contortus, The-meda triandra and Triodia pungens.
Eucalyptus dominated savannas have been exten-sively cleared with the intention to enhance pasturegrowth. Typically savanna is cleared by chain pullingthe woody vegetation and burning 1 to 2 years afterclearing. Fertilisers are not used. Clearing is usuallyfollowed by sowing the exotic pasture grass Cenchrusciliaris (buffelgrass), but establishment success isvariable. The study sites were selected to represent achronosequence of 1.5-, 2-, 10- and 30-year oldcleared sites of E. melanophloia savanna. The 1.5-year cleared (23°50’S, 146°08’E) and 10-year clearedsites (23°50’S, 146°05’E) were situated on grazingproperty ‘Summerdell’, and the 2-year (23°24’S, 146°30’E) and 30-year cleared sites (23°26’S, 146°30’E)were situated on grazing property ‘Monklands’(Fig. 1). Buffelgrass was present at cleared sites butwas not the dominant grass species. Cleared sites hadsome resprouts of E. melanophloia which were multi-stemmed and up to 3 m tall. Sites were selected inadjacent uncleared savanna after field inspectionsuggested compatible soil types. Uncleared sites forcomparison were grazed savanna remnants between2.2 and 10 ha in size and at more than 0.5 km distanceto cattle watering points.
Fig. 1 Location of the study sites, grazing properties“Monklands” and “Summerdell”, in the Desert Uplandsbiogeographical region in Queensland, Australia. Images ofcleared (with some remnant trees shown in the centre) anduncleared savanna (Regional Ecosystem 10.5.5) study site
Plant Soil (2010) 331:79–90 81

All study sites were located on upper slopepositions of moderately well drained, broad and flatsand plateaus, which have high saturated hydraulicconductivity in the topsoil and moderately high tohigh in the subsoil (McKenzie et al. 2007), and areeasily compacted (Thompson et al. 2003). Sites werelocated on Kandosol soils, which lack abrupt texturecontrast between the fine sandy loam or coarsertopsoils and the structure less subsoils (Jacquier etal. 2001; McKenzie et al. 2007), without signs of ashallow hardpan (<0.6 m depth). Sites had similarover- and under-storey vegetation components, soiltype, land management practices, and cattle stockingrates typical for the region.
Sampling protocol
In order to distinguish nutrient relations with respectto the highly variable rainfall in the region (Fenshamand Holman 1999), we sampled immediately beforeand after a period with major summer rainfall.Following the 2001 dry season (winter) which wasdrier than long-term averages (total rainfall <15 mmat Summerdell and <60 mm at Monklands duringJuly-Sept), we sampled vegetation and soil in spring(Oct 2001). Immediately after the major rainfallperiod in summer 2001–2002 (480 and 207 mmNov-Jan at Summerdell and Monklands, respective-ly), we re-sampled the sites in Jan-Feb 2002. Inuncleared savanna, leaves and xylem sap weresampled from three trees per species: the dominantover-storey species Eucalyptus melanophloia (Myrta-ceae), Acacia coriacea and A. excelsa (Mimosaceae),and an unidentified Hakea species (Proteaceae). Atcleared sites, grasses and E. melanophloia resproutswere sampled. Leaf samples were taken from 3–5individuals of the main native grass species occurringat the study sites, with seven to nine grass speciessampled at each site (pair sampling regime). Sampledgrass species were native Aristida holathera, Bothrio-chloa ewartiana, Chrysopogon fallax, Cymbopogonrefractus, Digitaria ammophila, Eriachne sp., Triodiapungens, and Themeda triandra and introducedCenchrus ciliaris.
Soil was sampled in January 2002 at 0–5 cm and at30–40 cm depth using a pneumatic soil auger (15 cmdiameter). Nine soil samples were taken at random ateach study site within an area representative of theoverall site; we avoided sampling in zones of visible
soil erosion or dense patches of vegetation. The ninesoil samples from were taken from an area ofapproximately 300 m2 and were pooled randomlyinto three soil samples per site. In uncleared savanna,the same sampling protocol was applied. At each site,we took nine soil samples were taken from a typicalarea containing trees and grasses, but avoiding thedirect vicinity of trees. Rather, soil samples weretaken at >2 m distance from tree trunks to avoidshallow roots in the canopy zone. Samples werepooled to produce three final soil samples foranalysis. An image of typical cleared and unclearedsavanna of this study is shown in Fig. 1.
Plant tissue and soil samples were air-dried in thefield and oven-dried for several days at 60°C. Severalintact pieces of microbiotic crust (10–20 cm2) weretaken from the soil surface with a hand trowel, takingcare to minimise contamination with mineral soil.Samples were subsequently ground to powder with aball-mill, except those for extraction of nitrate andammonium.
Plant and soil analyses
Elements and stable isotopes
Ground leaves (0.25 g dry weight) were digested in15 ml nitric perchloric acid for element analysis anddiluted with distilled water to 25 ml (Rayment andHigginson 1992). Soil and microbiotic crust (3 g dryweight) were extracted with 30 ml of 1 M NH4Cl(pH 7.5). Samples were analysed via ICPAES(Rayment and Higginson 1992) for P, K, Ca andMg. Prior to soil C analysis, inorganic C was removedby adding dilute sulfuric acid and subsequentlyneutralising with KOH. Leaves, soil and microbioticcrust were analysed for N and C content (%)and natural abundance of 15N and 13C (δ15N, δ13C)with an isotope ratio mass spectrometer (Roboprep-Tracermass, Europa PDZ, UK).
Soil ammonium and nitrate
Samples were hand-sifted to 2 mm and 3 g of soilwas mixed with 30 ml 2 M potassium chloride(KCl), shaken for 1 h on an orbital shaker andfiltered. Ammonium and nitrate were determinedcolourimetrically (Schmidt et al. 1998). For a time-integrated measure of plant-available soil nitrate and
82 Plant Soil (2010) 331:79–90
ammonium, five nylon mesh bags containing mixed-bed ion exchange resin were inserted per site and soildepth (5 and 40 cm) in October 2001 and February2002. Resin samplers remained in situ for 3 to 4 days.Disruption of the soil profile occurred as a result ofaugering to 40 cm depth, and the soil was subsequentlyfilled back into the hole. Resin bags were storedrefrigerated in sealed plastic bags until analysis. Resinwas extracted as detailed in Schmidt et al. (1998).
Xylem sap
Xylem sap was collected from trees using a hand-heldvacuum pump (Schmidt and Stewart 1998) from 1-2 cmthick branches between 6.00 and 9.00 am, kept on iceand stored at −80°C. Amino acids and amides wereanalysed with a post-column, ninhydrin derivatisationreverse-phase HPLC (Vasanits and Molnar-Perl 1999)using 50 μl of sample derivatised with PITC (Model6300, Beckman Instruments, Palo Alto). Nitrate inxylem sap was analysed colourimetrically (Schmidtand Stewart 1998). The proportion of nitrate wascalculated as a percentage of the total concentration oflow-molecular weight N compounds in xylem sap.
Statistics
T-tests were performed between soils of cleared andcorresponding uncleared sites (GraphPad Prism soft-ware). One-way ANOVAwas used to compare δ15N andN concentrations of tree genera. Two-way ANOVAwith interactions was used to test the effects of clearingand season on soil macronutrients (Fig. 2), and effectsof clearing and grass species (or native versus intro-duced grasses) on shoot nutrient concentrations, usingR 2.6.0 (R Core Team 2007). Log-transformationshomogenised heteroskedasticity and normalised skewedresiduals of grass nutrient concentrations.
Results
Soil nitrate and ammonium
KCl-extraction and in situ ion exchange resin incu-bation were compared to discern the most sensitivetechnique for assessing soil ammonium and nitratelevels in the studied soil. Levels of extractable nitrate(NO3
-) and ammonium (NH4+) were 0.35-0.62 mg
NO3− kg−1 dry soil and 4.7–7.4 mg NH4
+ kg−1 drysoil, but no consistent differences between clearedand uncleared sites were observed. NH4
+/NO3− ratios
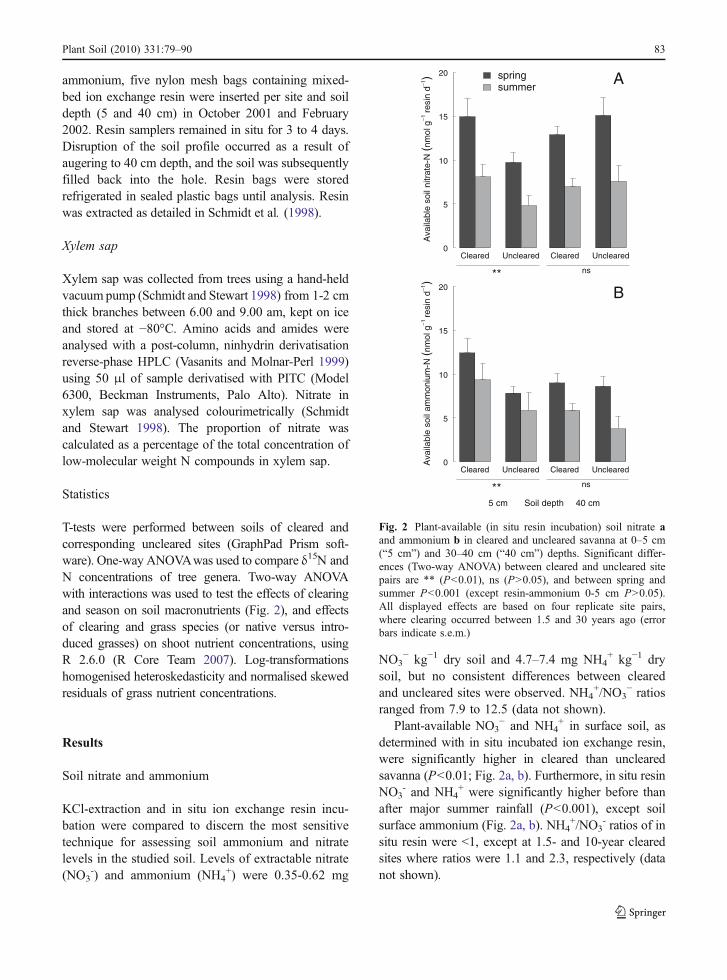
ranged from 7.9 to 12.5 (data not shown).Plant-available NO3
− and NH4+ in surface soil, as
determined with in situ incubated ion exchange resin,were significantly higher in cleared than unclearedsavanna (P<0.01; Fig. 2a, b). Furthermore, in situ resinNO3
- and NH4+ were significantly higher before than
after major summer rainfall (P<0.001), except soilsurface ammonium (Fig. 2a, b). NH4
+/NO3- ratios of in
situ resin were <1, except at 1.5- and 10-year clearedsites where ratios were 1.1 and 2.3, respectively (datanot shown).
springsummer
Cleared Uncleared
**
0
5
10
15
20
Ava
ilabl
e so
il ni
trat
e-N
(nm
ol g
−1 re
sin
d−1)
Cleared Uncleared
ns
A
Cleared Uncleared
**5 cm Soil depth
0
5
10
15
20
Ava
ilabl
e so
il am
mon
ium
-N (n
mol
g−1
resi
n d−1
)
Cleared Uncleared
ns
40 cm
B
Fig. 2 Plant-available (in situ resin incubation) soil nitrate aand ammonium b in cleared and uncleared savanna at 0–5 cm(“5 cm”) and 30–40 cm (“40 cm”) depths. Significant differ-ences (Two-way ANOVA) between cleared and uncleared sitepairs are ** (P<0.01), ns (P>0.05), and between spring andsummer P<0.001 (except resin-ammonium 0-5 cm P>0.05).All displayed effects are based on four replicate site pairs,where clearing occurred between 1.5 and 30 years ago (errorbars indicate s.e.m.)
Plant Soil (2010) 331:79–90 83
Soil total N and C content and stable isotopecomposition
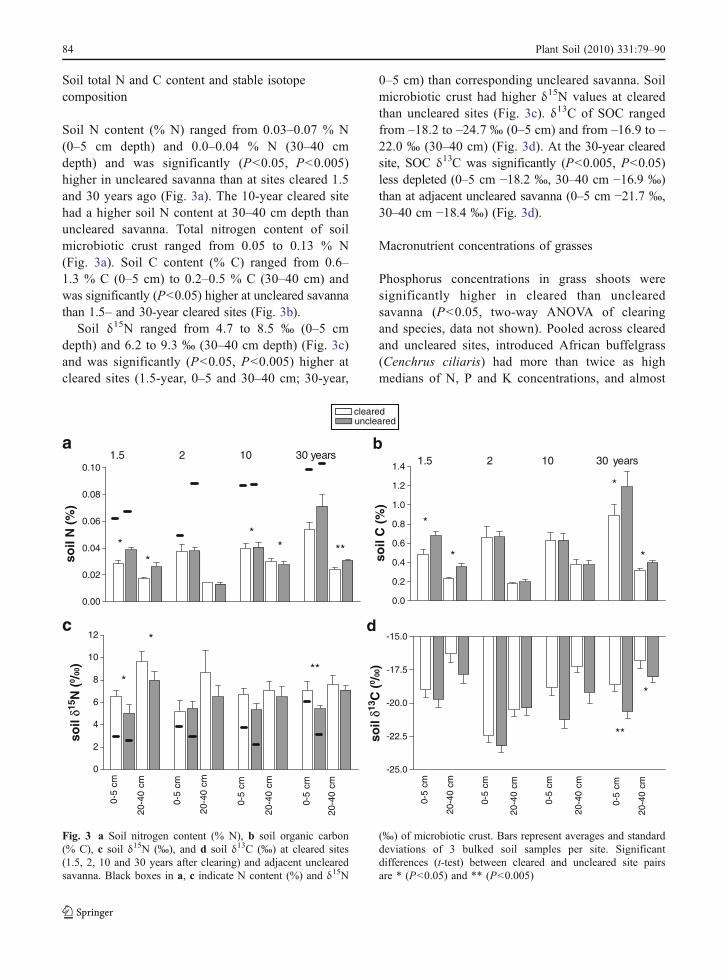
Soil N content (% N) ranged from 0.03–0.07 % N(0–5 cm depth) and 0.0–0.04 % N (30–40 cmdepth) and was significantly (P<0.05, P<0.005)higher in uncleared savanna than at sites cleared 1.5and 30 years ago (Fig. 3a). The 10-year cleared sitehad a higher soil N content at 30–40 cm depth thanuncleared savanna. Total nitrogen content of soilmicrobiotic crust ranged from 0.05 to 0.13 % N(Fig. 3a). Soil C content (% C) ranged from 0.6–1.3 % C (0–5 cm) to 0.2–0.5 % C (30–40 cm) andwas significantly (P<0.05) higher at uncleared savannathan 1.5– and 30-year cleared sites (Fig. 3b).
Soil δ15N ranged from 4.7 to 8.5 ‰ (0–5 cmdepth) and 6.2 to 9.3 ‰ (30–40 cm depth) (Fig. 3c)and was significantly (P<0.05, P<0.005) higher atcleared sites (1.5-year, 0–5 and 30–40 cm; 30-year,
0–5 cm) than corresponding uncleared savanna. Soilmicrobiotic crust had higher δ15N values at clearedthan uncleared sites (Fig. 3c). δ13C of SOC rangedfrom –18.2 to –24.7 ‰ (0–5 cm) and from –16.9 to –22.0 ‰ (30–40 cm) (Fig. 3d). At the 30-year clearedsite, SOC δ13C was significantly (P<0.005, P<0.05)less depleted (0–5 cm −18.2 ‰, 30–40 cm −16.9 ‰)than at adjacent uncleared savanna (0–5 cm −21.7 ‰,30–40 cm −18.4 ‰) (Fig. 3d).
Macronutrient concentrations of grasses
Phosphorus concentrations in grass shoots weresignificantly higher in cleared than unclearedsavanna (P<0.05, two-way ANOVA of clearingand species, data not shown). Pooled across clearedand uncleared sites, introduced African buffelgrass(Cenchrus ciliaris) had more than twice as highmedians of N, P and K concentrations, and almost
0.00
0.02
0.04
0.06
0.08
0.10
cleareduncleared
**
**
1.5 2 10 30 years
**
soil
N (
%)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4 1.5 2 10 30 years
*
*
*
*soil
C (
%)
0
2
4
6
8
10
12
0-5
cm
20-4
0 cm
0-5
cm
20-4
0 cm
0-5
cm
20-4
0 cm
0-5
cm
20-4
0 cm
*
soil
δ15N
(0 /0
0)
*
**
-25.0
-22.5
-20.0
-17.5
-15.0
0-5
cm
20-4
0 cm
0-5
cm
20-4
0 cm
0-5
cm
20-4
0 cm
0-5
cm
20-4
0 cm
*
**soil
δ13C
(0 /0
0)
a b
c d
Fig. 3 a Soil nitrogen content (% N), b soil organic carbon(% C), c soil δ15N (‰), and d soil δ13C (‰) at cleared sites(1.5, 2, 10 and 30 years after clearing) and adjacent unclearedsavanna. Black boxes in a, c indicate N content (%) and δ15N
(‰) of microbiotic crust. Bars represent averages and standarddeviations of 3 bulked soil samples per site. Significantdifferences (t-test) between cleared and uncleared site pairsare * (P<0.05) and ** (P<0.005)
84 Plant Soil (2010) 331:79–90
50% higher S concentrations in shoot biomass thaneight native grass species sampled (P<0.001, two-way ANOVA of clearing and species, data notshown), despite high variability in C. ciliaris.
δ 15N of vegetation and microbiotic crust
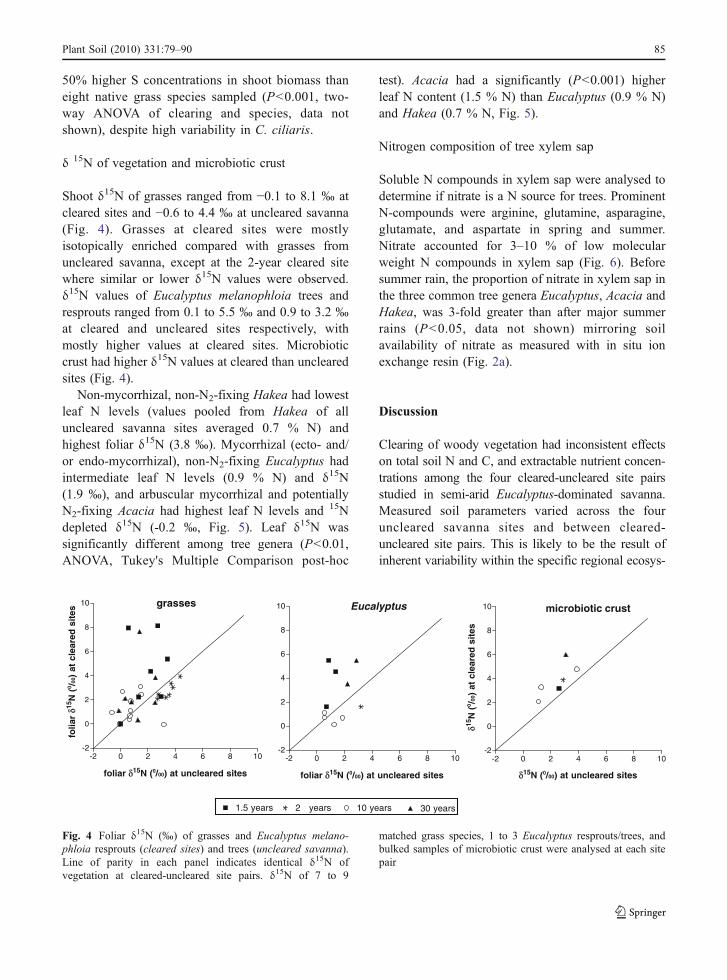
Shoot δ15N of grasses ranged from −0.1 to 8.1 ‰ atcleared sites and −0.6 to 4.4 ‰ at uncleared savanna(Fig. 4). Grasses at cleared sites were mostlyisotopically enriched compared with grasses fromuncleared savanna, except at the 2-year cleared sitewhere similar or lower δ15N values were observed.δ15N values of Eucalyptus melanophloia trees andresprouts ranged from 0.1 to 5.5 ‰ and 0.9 to 3.2 ‰at cleared and uncleared sites respectively, withmostly higher values at cleared sites. Microbioticcrust had higher δ15N values at cleared than unclearedsites (Fig. 4).
Non-mycorrhizal, non-N2-fixing Hakea had lowestleaf N levels (values pooled from Hakea of alluncleared savanna sites averaged 0.7 % N) andhighest foliar δ15N (3.8 ‰). Mycorrhizal (ecto- and/or endo-mycorrhizal), non-N2-fixing Eucalyptus hadintermediate leaf N levels (0.9 % N) and δ15N(1.9 ‰), and arbuscular mycorrhizal and potentiallyN2-fixing Acacia had highest leaf N levels and 15Ndepleted δ15N (-0.2 ‰, Fig. 5). Leaf δ15N wassignificantly different among tree genera (P<0.01,ANOVA, Tukey's Multiple Comparison post-hoc
test). Acacia had a significantly (P<0.001) higherleaf N content (1.5 % N) than Eucalyptus (0.9 % N)and Hakea (0.7 % N, Fig. 5).
Nitrogen composition of tree xylem sap
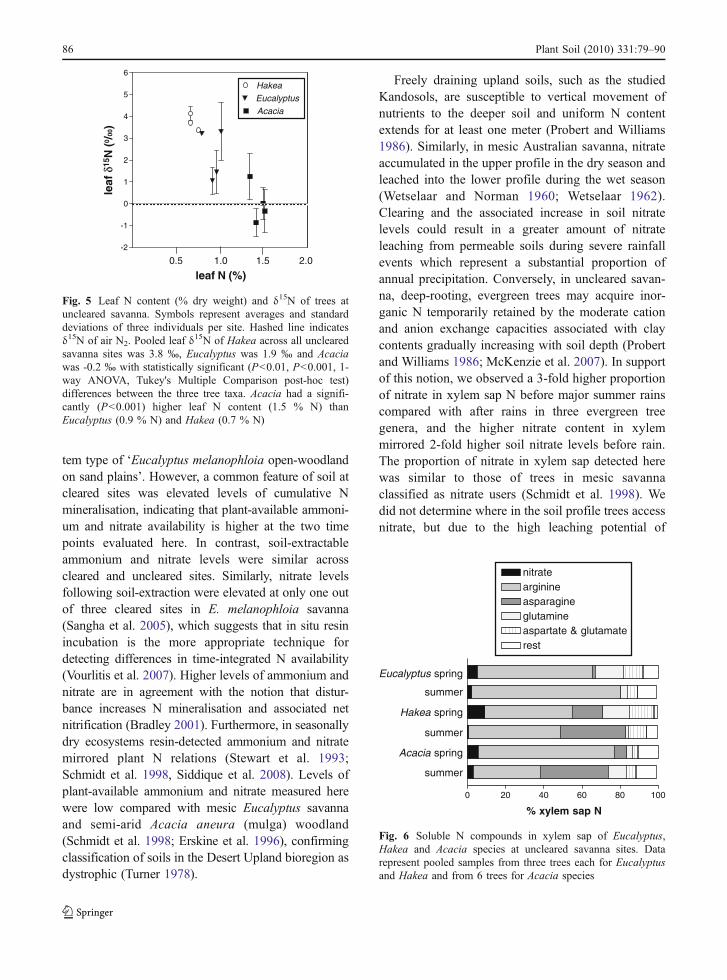
Soluble N compounds in xylem sap were analysed todetermine if nitrate is a N source for trees. ProminentN-compounds were arginine, glutamine, asparagine,glutamate, and aspartate in spring and summer.Nitrate accounted for 3–10 % of low molecularweight N compounds in xylem sap (Fig. 6). Beforesummer rain, the proportion of nitrate in xylem sap inthe three common tree genera Eucalyptus, Acacia andHakea, was 3-fold greater than after major summerrains (P<0.05, data not shown) mirroring soilavailability of nitrate as measured with in situ ionexchange resin (Fig. 2a).
Discussion
Clearing of woody vegetation had inconsistent effectson total soil N and C, and extractable nutrient concen-trations among the four cleared-uncleared site pairsstudied in semi-arid Eucalyptus-dominated savanna.Measured soil parameters varied across the fouruncleared savanna sites and between cleared-uncleared site pairs. This is likely to be the result ofinherent variability within the specific regional ecosys-
-2 0 2 4 6 8 10-2
0
2
4
6
8
10 grasses
foliar δ15N (0/00) at uncleared sites
folia
r δ15
N (
0 /00)
at
clea
red
sit
es
-2 0 2 4 6 8 10-2
0
2
4
6
8
10 Eucalyptus
foliar δ15N (0/00) at uncleared sites
-2 0 2 4 6 8 10-2
0
2
4
6
8
10
1.5 years 10 years 30 years2 years
microbiotic crust
δ15N
(0 /0
0) a
t cl
eare
d s
ites
δ15N (0/00) at uncleared sites
Fig. 4 Foliar δ15N (‰) of grasses and Eucalyptus melano-phloia resprouts (cleared sites) and trees (uncleared savanna).Line of parity in each panel indicates identical δ15N ofvegetation at cleared-uncleared site pairs. δ15N of 7 to 9
matched grass species, 1 to 3 Eucalyptus resprouts/trees, andbulked samples of microbiotic crust were analysed at each sitepair
Plant Soil (2010) 331:79–90 85
tem type of ‘Eucalyptus melanophloia open-woodlandon sand plains’. However, a common feature of soil atcleared sites was elevated levels of cumulative Nmineralisation, indicating that plant-available ammoni-um and nitrate availability is higher at the two timepoints evaluated here. In contrast, soil-extractableammonium and nitrate levels were similar acrosscleared and uncleared sites. Similarly, nitrate levelsfollowing soil-extraction were elevated at only one outof three cleared sites in E. melanophloia savanna(Sangha et al. 2005), which suggests that in situ resinincubation is the more appropriate technique fordetecting differences in time-integrated N availability(Vourlitis et al. 2007). Higher levels of ammonium andnitrate are in agreement with the notion that distur-bance increases N mineralisation and associated netnitrification (Bradley 2001). Furthermore, in seasonallydry ecosystems resin-detected ammonium and nitratemirrored plant N relations (Stewart et al. 1993;Schmidt et al. 1998, Siddique et al. 2008). Levels ofplant-available ammonium and nitrate measured herewere low compared with mesic Eucalyptus savannaand semi-arid Acacia aneura (mulga) woodland(Schmidt et al. 1998; Erskine et al. 1996), confirmingclassification of soils in the Desert Upland bioregion asdystrophic (Turner 1978).
Freely draining upland soils, such as the studiedKandosols, are susceptible to vertical movement ofnutrients to the deeper soil and uniform N contentextends for at least one meter (Probert and Williams1986). Similarly, in mesic Australian savanna, nitrateaccumulated in the upper profile in the dry season andleached into the lower profile during the wet season(Wetselaar and Norman 1960; Wetselaar 1962).Clearing and the associated increase in soil nitratelevels could result in a greater amount of nitrateleaching from permeable soils during severe rainfallevents which represent a substantial proportion ofannual precipitation. Conversely, in uncleared savan-na, deep-rooting, evergreen trees may acquire inor-ganic N temporarily retained by the moderate cationand anion exchange capacities associated with claycontents gradually increasing with soil depth (Probertand Williams 1986; McKenzie et al. 2007). In supportof this notion, we observed a 3-fold higher proportionof nitrate in xylem sap N before major summer rainscompared with after rains in three evergreen treegenera, and the higher nitrate content in xylemmirrored 2-fold higher soil nitrate levels before rain.The proportion of nitrate in xylem sap detected herewas similar to those of trees in mesic savannaclassified as nitrate users (Schmidt et al. 1998). Wedid not determine where in the soil profile trees accessnitrate, but due to the high leaching potential of
0 20 40 60 80 100
nitratearginineasparagineglutamineaspartate & glutamaterest
Acacia spring
summer
summer
Eucalyptus spring
Hakea spring
summer
% xylem sap N
Fig. 6 Soluble N compounds in xylem sap of Eucalyptus,Hakea and Acacia species at uncleared savanna sites. Datarepresent pooled samples from three trees each for Eucalyptusand Hakea and from 6 trees for Acacia species
-2
-1
0
1
2
3
4
5
6
Acacia
Eucalyptus
Hakea
0.5 1.0 1.5 2.0
leaf N (%)
leaf
δ15
N (
0 /00)
Fig. 5 Leaf N content (% dry weight) and δ15N of trees atuncleared savanna. Symbols represent averages and standarddeviations of three individuals per site. Hashed line indicatesδ15N of air N2. Pooled leaf δ15N of Hakea across all unclearedsavanna sites was 3.8 ‰, Eucalyptus was 1.9 ‰ and Acaciawas -0.2 ‰ with statistically significant (P<0.01, P<0.001, 1-way ANOVA, Tukey's Multiple Comparison post-hoc test)differences between the three tree taxa. Acacia had a signifi-cantly (P<0.001) higher leaf N content (1.5 % N) thanEucalyptus (0.9 % N) and Hakea (0.7 % N)
86 Plant Soil (2010) 331:79–90

Kandosols a considerable proportion of soil nitratemay be accessed in the deeper soil profile. Nitrateincreased in the xylem sap following long dry periodswhich indicates that trees acquire nitrate from the soilat times when roots of herbaceous species are inactive(Cui and Caldwell 1997). In support of this notion,Australian savanna trees are associated with ‘islandsof fertility’ characterised by a higher N status thaninter-canopy soil (Jackson and Ash 1998, 2001;Ludwig et al. 2004). It has to be established whethersavanna trees are a continuous sink for nitratethroughout the year, effectively preventing accumula-tion of nitrate in the soil and thereby preventingleaching losses which could affect post-clearingpastures.
In this context, uncertainty exists whether intro-duced buffelgrass (Cenchrus ciliaris) uses subsoil Nand how buffelgrass affects overall nutrient cycling incleared savanna. We observed markedly higher con-centrations of N, P, K and S in shoot biomass ofbuffelgrass compared to all common native grasses,but the effects of such nutrient accumulation oninterspecific competition, redistribution and losses ofnutrients from the ecosystem remain to be examined.For instance, buffelgrass can result in intensified firesand associated N (Jackson 2005) and potentially Sloss. 100 years after clearing mulga woodland, SOCin Kandosol under buffelgrass pastures had declinedby 31% in the top 5 cm (Dalal et al. 2005), andchanges of soil C and N stocks were stronglycorrelated (Harms et al. 2005). In Central Queenslandat 8 out of 31 sites, buffelgrass on solodic soils ofcleared brigalow (Acacia harpophylla) was associatedwith 16-34% reductions of SOC and N in the top10 cm (Graham et al. 1981), but similar to Harms etal. (2005), change in SOC or soil N was notassociated with time since clearing. Thus, it appearsthat other factors impact on the fate of SOC and thuson soil N, which may include initial composition ofsoil, vegetation and climate characteristics and land-scape position. Similarly, conversion of native Mex-ican grassland to buffelgrass pasture resulted in a 25and 40% decline of soil N and C stocks, respectively,at some sites, but no change or elevated soil N and Cstocks at other sites (Ibarra-Flores et al. 1999).Further, we observed that buffelgrass had highershoot macronutrient contents than native grasses,irrespective of clearing history of the site. Buffelgrass,a popular pasture species, is abundant or dominant on
30-50 Mha in Queensland alone (Woinarski 2001)and covers seven of the ten ‘Grazing Land Manage-ment Zones’ in Australia (Martin et al. 2006). Exoticgrasses can alter soil biogeochemistry (D’Antonio andVitousek 1992; Rossiter-Rachor et al. 2008, 2009),and the sources of variability and interactions of theeffects of clearing and presence of buffelgrass have tobe established.
The less depleted δ13C values of SOC at the 30-year cleared site points to a greater contribution of C4-grasses compared with uncleared savanna but this wasnot associated with the net C gain often observed ingrasslands. In buffelgrass pasture, Dalal et al. (2005)detected a greater proportion of C4-grass derived SOCand a decline in SOC quality compared to that of theoriginal mulga (Acacia aneura) ecosystem. It wasproposed that pasture soils dominated by Africangrasses gain C because exotic grasses sequester Cdeeper into the soil than native grasses (Fisher et al.1994), but a meta-analysis of the effects of forest-to-pasture conversion showed that soil C stocks mayeither increase or decrease in regions with <1,000 mmmean annual rainfall (Guo and Gifford 2002). There isevidence from studies of sandy soils that the activepool of C4-derived SOC decomposes faster thanequivalent C3-derived SOC in mixed C3/C4 soils(Wynn and Bird 2007), and this could contribute tothe observed net loss of SOC here and in previousstudies.
Soil of the 1.5- and 30-year cleared sites whichshowed a net reduction in soil N stocks also hadgreater 15N enrichment than adjacent unclearedsavanna. 15N enrichment is indicative of N loss fromecosystems (Högberg 1997), and isotopic enrichmentof soil was mirrored by grasses, microbiotic crust andEucalyptus resprouts at cleared sites, confirming thenotion that δ15N of bulk soil is broadly reflected byvegetation (Handley et al. 1999). The 2–3‰ lowerδ15N of microbiotic crust relative to soil indicatesactive N2 fixation, which is supported by acetylenereduction capacity of hydrated microbiotic crust(unpublished results). In arid ecosystems, microbioticcrusts fix considerable amounts of N, and clearing aswell as grazing change composition and presenceof microbiotic crusts (Evans and Johansen 1999).Similarly, Acacia coriacea and A. excelsa had the15N-depleted signatures that can be indicative of N2-fixation. It is likely that the studied Acacia species fixN2 because there are no known non-nodulating
Plant Soil (2010) 331:79–90 87

Acacia species in sub-genus Phyllodineae (Sprent2001). Nitrogen input from biological N2 fixation isdifficult to ascertain in natural ecosystems, butestimates for tropical savannas range from 16 to44 kg N ha−1 y−1, with symbiotic and asymbiotic N2
fixation contributing 0.7–35 kg and 9.5 kg N ha−1
y−1, respectively (Cleveland et al. 1999). The large-scale invasion by N2-fixing exotic shrubs and trees inAustralian rangelands (Martin et al. 2006) suggests arole for woody N2-fixers in grazed rangelands. Aninventory across ecosystem N pools could assign N2
fixing function to ecosystem components, includingas woody taxa, forbs, graminoids/non-symbiotic N2
fixers and microbiotic crusts, which would improveunderstanding of N fluxes in savanna (Bustamante etal 2006).
In addition to altered N and C relations, we foundthat P contents of grasses were elevated at clearedsites, which is a potential cause for increased pastureproduction at least in the short to medium term afterclearing (Scanlan 2002; Kaur et al. 2005). Causes forimproved P nutrition of grasses at cleared sitesinclude reduced competition from woody species,but how well native and exotic grasses acquire Pdepends on species’ abilities as well as soil properties.Old soils, such as the Kandosols studied here, supportvegetation characterised by the ability to efficientlyaccess and use soil nutrients, and among woodyspecies, Proteaceae which are ubiquitous in Australianecosystems, are prominent P mobilisers (Lambers etal. 2008). In E. melanophloia savanna, extractable Plevels in the top 30 cm of the soil were elevated10 years after clearing but declined to below pre-clearing levels 30 years after clearing (Sangha et al.2005). A more detailed investigation of longer term(>30 years) cleared sites has to determine thegenerality of these findings.
In summary, spatial heterogeneity in total andextractable soil nutrient concentrations may obscurethe effects of clearing semi-arid savanna trees onplant-available nutrient dynamics and potentialimplications for nutrient loss from the ecosystem.Assessment of savanna clearing needs to account fordifferences between slow nutrient cycling during dryperiods and pulses of rapid transformations duringheavy rainfall events, and their interactions withspecies and functional community composition, aswell as ecological variability. Understanding theserelationships will be essential to inform strategies to
efficiently limit the impact of climate change andexotic plant invasion while developing sustainableuse of semi-arid rangelands (Henry et al. 2005). Wepropose that approaches which combine measure-ment of ecophysiology and soil processes are mostsuitable.
Acknowledgements This study was supported by a grantfrom the CRC for Tropical Savannas. We thank the owners of“Summerdell” and “Monklands” for access to their property.
References
Baldesdent J, Mariotti A (1996) Measurement of soil organicmatter turnover using 13C natural abundance. In: BouttonTW, Yamasaki S (eds) Mass spectrometry of soil. Dekker,New York, pp 83–111
Bradley RL (2001) An alternative explanation for the post-disturbance NO3
- flush in some forest ecosystems. EcolLetters 4:412–416
Bustamante MMC, Medina E, Asner GP, Nardoto GB, Garcia-Montiel DC (2006) Nitrogen cycling in tropical andtemperate savannas. Biogeochem 79:209–237
Cleveland CC, Townsend AR, Schimel DS, Fisher H, HowarthRW, Hedin LO, Perakis SS, Latty ER, von Fischer JC,Elseroad A, Wasson MF (1999) Global patterns ofterrestrial biological nitrogen (N2) fixation in naturalecosystems. Global Biogeochem Cycles 13:623–645
Clewett JF, Clarkson NM, Owens DT, Arbrecht DG (1994)Australian RAINMAN, Version 2.2. Department of Pri-mary Industries, Brisbane
Commonwealth of Australia, Department of the Environment,Water, Heritage and the Arts (2008) http://www.environment.gov.au/biodiversity/hotspots/national-hotspots.html#1Accessed 31 Dec 2008
Cui M, Caldwell MM (1997) A large ephemeral release ofnitrogen upon wetting of dry soil and correspondingresponses in the field. Plant Soil 191:291–299
Dalal RC, Carter JO (2000) Soil organic matter dynamicsand carbon sequestration in Australian tropical soils.In: Lal R, Kimble JM, Stewart BA (eds) Globalclimate change and tropical ecosystems. CRC, BocaRaton, pp 285–314
Dalal RC, Harms BP, Krull E, Wang WJ (2005) Total soilorganic matter and its labile pool following mulga (Acaciaaneura) clearing for pasture development and cropping 1.Total and labile carbon. Aust J Soil Res 43:13–20
D’Antonio CM, Vitousek PM (1992) Biological invasions byexotic grasses, the grass/fire cycle, and global change. AnnRev Ecol Syst 23:63–87
EPA (2007). Regional ecosystem description database (REDD).Version 5.2, Updated Nov. 2007. Queensland Herbarium,Environmental Protection Agency, Brisbane.
Erskine PD, Stewart GR, Schmidt S, Turnbull MH, Unkovich M,Pate JS (1996)Water availability—a physiological constrainton nitrate utilisation in plants of Australian semi-arid mulgawoodlands. Plant Cell Environ 19:1149–1159
88 Plant Soil (2010) 331:79–90

Evans RD, Johansen JR (1999) Microbiotic crusts andecosystem processes. Crit Rev Plant Sci 18:183–225
Fairfax RJ, Fensham RJ (2000) The effect of exotic pasturedevelopment on floristic diversity in central Queensland,Australia. Biol Cons 94:11–21
Fensham RJ, Holman JE (1999) Temporal and special patternsin drought-related tree dieback in Australian savanna. JAppl Ecol 36:1035–55
Fisher MJ, Rao IM, Ayarza MA, Lascano CE, Sanz JI, ThomasRJ, Vera RR (1994) Carbon storage by introduced deep-rooted grasses in the South American Savannas. Nature371:236–238
Graham TWG, Webb AA, Waring SA (1981) Soil nitrogenstatus and pasture productivity after clearing of brigalow(Acacia harpophylla). Aust J Exp Agr Animal Husb21:109–118
Guo LB, Gifford RM (2002) Soil carbon stocks and land usechange: a meta analysis. Global Change Biol 8:345–360
Harms BP, Dalal RC, Cramp AP (2005) Changes in soil carbonand soil nitrogen after tree clearing in the semi-aridrangelands of Queensland. Aust J Bot 53:639–650
Handley LL, Austin AT, Robinson D, Scrimgeour CM, RavenJA, Heaton THE, Schmidt S, Stewart GR (1999) The 15Nnatural abundance (δ15N) of ecosystems samples reflectsmeasures of water availability. Aust J Plant Phys 26:185–199
Högberg P (1997) Tansley review no. 95. 15N naturalabundance in soil-plant systems. New Phytol 137:179–203
Henry B, Mitchell C, Cowie A, Woldring O, Carter J (2005) Aregional interpretation of rules and good practise forgreenhouse accounting: Northern Australian savannasystems. Aust J Bot 53:589–605
Ibarra-Flores F, Cox JR, Martin-Rivera M, Crowl TA, NortonBE (1999) Soil physicochemical changes followingbuffelgrass establishment in Mexico. Arid Soil Res Rehab13:39–52
Jackson J (2005) Is there a relationship between herbaceousspecies richness and buffel grass (Cenchrus ciliaris)?Austral Ecol 30:505–517
Jackson J, Ash AJ (1998) Tree-grass relationships in openeucalypt woodlands of north-eastern Australia: influenceof trees on pasture productivity, forage quality and speciesdistribution. Agrofor Syst 40:159–176
Jackson J, Ash AJ (2001) The role of trees in enhancing soilnutrient availability for native perennial grasses in openeucalypt woodlands of north-east Queensland. Aust J AgrRes 52:377–386
Jacquier, D. W., Mckenzie, N. J., Brown, K. L., Isbell, R. F.,Paine, T. A. (2001). The Australian soil classification—aninteractive key. Version 1.0
Kaur K, Jalota RK, Midmore DJ, Rolfe J (2005) Pastureproduction in cleared and uncleared grazing systems ofCentral Queensland, Australia. Rangeland J 27:143–147
Lambers H, Raven JA, Shaver GR, Smith SE (2008) Plantnutrient-acquisition strategies change with soil age. TrendsEcol & Evol 23:95–103
Ludwig F, De Kroon H, Berendse F, Prins HHT (2004) Theinfluence of savanna trees on nutrient, water and lightavailability and the understorey vegetation. Plant Ecol170:93–105
Martin TG, Campbell S, Grounds S (2006) Weeds of Australianrangelands. Rangeland J 28:3–26
Mckenzie, N., Jacquier, D., Gregory, L., Pitman, C., Swan, G.,McKenzie, A. (2007). Australian soil resource informationsystem ASRIS (Last updated: Nov. 9, 2007). CSIRO Landand Water, Canberra. http://www.asris.csiro.au/. Accessed25 August 2008
Neff JC, Townsend AR, Gleixner G, Lehman SJ, Turnbull J,Bowman WD (2002) Variable effects of nitrogen additionsof the stability and turnover of soil carbon. Nature419:915–917
Powers JS, Read JM, Denslow JS, Guzman SM (2004)Estimating soil carbon fluxes following land-coverchange: a test of some critical assumptions for a regionin Costa Rica. Global Change Biol 10:170–181
Probert ME, Williams J (1986) The nitrogen status of redand yellow earths in the semi-arid tropics as influencedby Caribbean stylo (Stylosanthes hamata) grown atvarious rates of applied phosphorus. Aust J Soil Res24:405–421
R Core Team. (2007). R: A language and environment forstatistical computing. R foundation for statistical comput-ing, Vienna, Austria. http://www.R-project.org. Accessed28 August 2008
Rayment GE, Higginson FR (1992) Australian laboratoryhandbook of soil and water chemicals. Inkata, Melbourne
Robinson D (2001) δ15N as an integrator of the nitrogen cycle.Trends Ecol Evol 16:153–162
Rossiter-Rachor N, Setterfield SA, Douglas MM, Hutley LB,Cook GD (2008) Andropogon gayanus (gamba grass)invasion increases fire-mediated nitrogen losses in thetropical savannas of Northern Australia. Ecosystems11:77–88
Rossiter-Rachor NA, Setterfield SA, Douglas MM, Hutley LB,Cook GD, Schmidt S (2009) Invasive Andropogongayanus (gamba grass) is an ecosystem transformer ofnitrogen relations in Australia’s tropical savanna. EcolAppl 19:1546–1560
Sangha KK, Jalota RK, Midmore D (2005) Impact of treeclearing on soil pH and nutrient availability in grazingsystems of central Queensland, Australia. Aust J Soil Res43:51–60
Scanlan JC (2002) Some aspects of tree-grass dynamics inQueensland’s grazing lands. Rangeland J 24:56–82
Scholes RJ, Archer SR (1997) Tree-grass interactions insavannas. Ann Rev Ecol Syst 28:517–544
Schmidt S, Stewart GR (1998) Transport, storage and mobi-lisation of nitrogen by trees and shrubs in the wet/drytropics of northern Australia. Tree Phys 18:403–410
Schmidt S, Lamble RE (2002) Nutrient dynamics in Queens-land savannas: implications for the sustainability of landclearing for pasture production. Rangeland J 24:96–111
Schmidt S, Stewart GR (2003) δ15N values of tropical savannaand monsoon forest species reflect root specialisations andsoil nitrogen status. Oecologia 134:569–577
Schmidt S, Stewart GR, Turnbull MH, Erskine PD, Ashwath N(1998) Nitrogen relations in natural and disturbed plantcommunities in tropical Australia. Oecologia 117:95–105
Siddique I, Engel VL, Parrotta JA, Lamb D, Nardoto GB,Ometto JPHB, Martinelli LA, Schmidt S (2008) Domi-nance of legume trees alters nutrient relations in mixed
Plant Soil (2010) 331:79–90 89

species forest restoration plantings within seven years.Biogeochem 88:89–101
Sprent JI (2001) Nodulation in legumes. Royal BotanicGardens, Kew
Stewart GR, Pate JS, Unkovich M (1993) Characteristics ofinorganic nitrogen assimilation of plants in fire-proneMediterranean-type vegetation. Plant Cell Environ16:351–363
Thackway R, Creswell ID (1995) An interim biogeographicregionalisation for australia: A framework for settingpriorities in the national reserves system cooperativeprogram, Version 4’. Australian Nature ConservationAgency, Canberra
Thompson, E. J., Sharp, D., Kutt, A., & Morgan, G. (2003).10.5.5 Silver-leaved ironbark open-woodland on sandplains. In: Regional ecosystems of the desert uplandsfactsheets. Queensland Herbarium, environmental pro-tection agency, Brisbane. http://www.epa.qld.gov.au/media/nature_conservation/biodiversity/desert_uplands/Factsheets/100505.htm. Accessed 25 August 2008
Turner, E. J. (1978). Land systems. In: Western arid region landuse study. Part IV. Technical bulletin No. 23, Departmentof Primary Industries, Brisbane
Vasanits A, Molnar-Perl I (1999) Temperature, eluent flow-rateand column effects on the retention and quantificationproperties of phenylthiocarbamyl derivatives of aminoacids in reverse-phase high-performance liquid chroma-tography. J Chromat 832:109–122
Vourlitis GL, Pasquini S, Zorba G (2007) Plant and soil Nresponse of southern Californian semi-arid shrublandsafter 1 year of experimental N deposition. Ecosyst10:263–279
Wetselaar R, Norman MJT (1960) Recovery of available soilnitrogen by annual fodder crops at Katherine, NorthernTerritory. Aust J Agr Res 11:693–704
Wetselaar R (1962) Nitrate distribution in tropical soils. III.Downward movement and accumulation of nitrate in thesubsoil. Plant Soil 16:19–31
Whitehead, P. J., Woinarski, J., Jacklyn, P., Fell, D., Williams,D. (2000). Defining and measuring the health of savannalandscapes: A north Australian perspective. Tropicalsavannas cooperative research centre, Charles DarwinUniversity
Wilson BA, Nelder VJ, Accad A (2002) The extent and statusof remnant vegetation in Queensland and its implicationfor statewide vegetation management and legislation.Rangeland J 24:6–35
Woinarski J (2001) A review of changes in status andthreatening processes. In: Whitehead P, Woinarski J,Fisher A, Fensham JR, Beggs K (eds) Developing ananalytical framework for monitoring biodiversity inAustralia’s rangelands. Report for the national land andwater resources audit. Tropical Savanna CRC, Darwin
Wynn JG, Bird MI (2007) C4-derived soil organic carbondecomposes faster than its C3 counterpart in mixed C3/C4soils. Global Change Biol 13:2206–2217
90 Plant Soil (2010) 331:79–90
Copyright © 2022 FDOKUMEN




















