
Pharmacokinetics of new 625 mg nelfinavir formulation during pregnancy and postpartum
Upload
independentCategory
view
4download
0
Animal Reproduction Science 114 (2009) 1–13
Contents lists available at ScienceDirect
Animal Reproduction Science
journal homepage: www.elsevier.com/locate/anireprosci
Effect of prepartum diet on postpartum ovarian activity inHolstein cows in a pasture-based dairy system
D. Cavestanya,b,∗, C. Vinolesa, M.A. Crowec, A. La Mannaa, A. Mendozab
a National Agricultural Research Institute, INIA La Estanzuela, C. Correos 39173, 70000 Colonia, Uruguayb Faculty of Veterinary, University of Uruguay, Lasplaces 1620, 11600 Montevideo, Uruguayc School of Agriculture, Food Science and Veterinary Medicine and Conway Institute,University College Dublin, Belfield, Dublin 4, Ireland
a r t i c l e i n f o
Article history:Received 4 April 2008Received in revised form 11 July 2008Accepted 4 August 2008Available online 7 August 2008
Keywords:Cattle-prepartum supplementationOvarian follicular dynamicsOvarian activityIGF-INEFA
a b s t r a c t
The hypothesis was that supplementation during the late prepar-tum period will differentially affect reproductive and productivevariables according to parity. Primiparous (n = 22) and multiparous(n = 22) pregnant autumn calving Holstein cows were stratified intwo groups according to parity (primiparous or multiparous) andwithin each group were randomly assigned to two treatments: (a)low supplemented (LS) or (b) high supplemented (HS) prepartumdiet. The LS group was offered 5.2 kg/cow/day (DM basis) of wheatsilage, and the HS group 4.7 kg cow/day (DM basis)/of corn silageand 3.6 kg (DM basis) of wheat bran + 12 g of urea. Both groupsgrazed on natural pastures. After calving, all cows received the samediet. The experimental period was from 3 weeks before calving to7 weeks postpartum (PP); body condition score (BCS) and bloodsamples for hormonal analyses were obtained weekly and ovarianultrasonography was conducted three times per week. The loss inBCS around calving was less pronounced in HS cows, but only mul-tiparous supplemented cows maintained BCS throughout the study.Non-esterified fatty acids (NEFA) increased during the prepartumperiod in the LS but not in the HS cows, with peak values occurringon day 14 PP in all groups. During the remainder of the experi-ment NEFA was greater in LS than in HS cows. Prepartum treatmentdid not affect the proportion of cows that had ovulations from thefirst dominant follicle postpartum, but decreased the interval tofirst ovulation in multiparous cows (22.9 compared with 38.2 days;P < 0.05). This was associated with greater plasma IGF-I concentra-tions at the time the dominant follicle of the first follicular wave
∗ Corresponding author at: Faculty of Veterinary, University of Uruguay, Lasplaces 1620, 11600 Montevideo, Uruguay.Tel.: +598 2 6232455; fax: +598 2 6232455.
E-mail address: [email protected] (D. Cavestany).
0378-4320/$ – see front matter © 2008 Elsevier B.V. All rights reserved.doi:10.1016/j.anireprosci.2008.08.007

2 D. Cavestany et al. / Animal Reproduction Science 114 (2009) 1–13
reached its maximum diameter (8.0 compared with 3.6 nmol/L;P < 0.05). However, prepartum treatment had no effect on onsetof ovarian activity in primiparous cows. Supplementation had noeffect on milk production or milk protein percentage but increasedmilk fat percentage. We conclude that feeding a high-supplementedprepartum diet to multiparous cows allowed them to maintain BCSaround calving, and this was associated with greater concentrationsof IGF-I and an earlier onset of estrous cycles after calving.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
It is well established that during the last month of pregnancy and the early postpartum period, highproducing cows undergo a period of negative energy balance (NEB) as the final growth of the fetusand milk production requirements exceed the energy obtained through feeding (Grummer, 1995;Butler, 2003). The severity of the NEB is more pronounced in primiparous cows, as they have energyrequirements for their own growth in addition to the energy demands for foetal growth, mammarydevelopment and their first lactation (Bell, 1995).
The interval to first postpartum (PP) ovulation is related to the period of NEB, as metabolites andmetabolic hormones convey information from the cow’s metabolic status to her central nervous system(Butler, 2003). Cows in NEB have greater concentrations of growth hormone (GH) and non-esterifiedfatty acids (NEFA), and lesser concentrations of insulin-like growth factor-I (IGF-I), insulin and glu-cose (Canfield and Butler, 1991). Moreover, cows having ovulations within the first 35 days PP havegreater concentrations of IGF-I, as well as glucose and insulin, and lesser concentrations of NEFA and�-hydroxybutyrate (BHB; Huszenicza et al., 2001).
Re-initiation of postpartum ovarian activity is closely related to the feeding and management duringthe transition period and the metabolic and physiological changes that it implies (Bell, 1995). A high-energy diet prepartum may favour adaptation of ruminal microorganisms to PP diets (Ingvartsen andAndersen, 2000) and increase absorption of volatile fatty acids in the rumen, leading to an increasein insulin and a decrease of fatty acid mobilization from the adipose tissue (Holcomb et al., 2001;Doepel et al., 2002). However, the effect of different prepartum diets on reproductive performanceis controversial, with studies showing a positive (Chagas et al., 2006), negative (Keady et al., 2001)or no response on it (Pushpakumara et al., 2003). Moreover, the impact of different transition dietson metabolic variables and fertility has not been evaluated for primiparous and multiparous cowsseparately under grazing conditions.
Our hypothesis was that feeding a high-supplemented diet during the prepartum period will reducethe magnitude of PP NEB, diminishing the mobilization of adipose tissue, thus, leading to lesser concen-trations of NEFA and BHB, and greater concentrations of IGF-I, that may promote enhanced folliculargrowth, and induce an earlier onset of ovulation PP. We speculated that the benefit might be more pro-nounced in primiparous than in multiparous cows. The aim of this study was to evaluate the effects ofamount of supplementation during the late prepartum period in primiparous and multiparous Hol-stein cows under grazing conditions, on milk production, metabolic profiles, ovarian follicular growthand the interval to first PP ovulation.
2. Materials and methods
2.1. Animals and treatments
The trial was conducted in the experimental dairy herd of the research station of the NationalAgricultural Research Institute (INIA) in La Estanzuela (Colonia, Uruguay), using 22 primiparous and 22multiparous pregnant Holstein cows. At the start of the experiment, primiparous cows had an averageBW of 501 ± 57 (mean ± S.E.M.) kg and BCS of 2.85 ± 0.07, and multiparous cows had an average BW

D. Cavestany et al. / Animal Reproduction Science 114 (2009) 1–13 3
of 563 ± 65 kg and BCS of 2.53 ± 0.06 (scale 1–5, Edmonson et al., 1989). The experimental period wasfrom 21 days before the expected calving date to 49 days PP. The cows were expected to calve at thebeginning of the austral autumn (March), and 21 days before the individual expected calving datethey were stratified in two groups on the basis of lactation number (primiparous or multiparous) andwithin each group were randomly assigned to the following prepartum treatments: (1) low (LS) and (2)high (HS) amounts of supplementation. Each treatment had 11 primiparous cows and 11 multiparouscows. Time of parturitions was controlled, and at 15 days postpartum a gynaecologycal examination(vaginoscopy) was done to all animals.
During the prepartum, multiparous and primiparous cows from both groups grazed in separatepaddocks of natural pastures (10.4% CP, 1.34 Mcal NEL/kg dry matter (DM)). Cows from the LS groupreceived a supplement, which consisted of 5.2 kg (DM basis) of whole-plant wheat silage (9.0% CP,1.29 Mcal NEL/kg DM) per cow and day, and received no concentrate. Cows from the HS group receiveddaily (at 08:00 h) a concentrate consisting in 3.6 kg (DM) of wheat bran (16.4% CP, 1.69 Mcal NEL/kgDM) + 0.012 kg of urea, and 4.7 kg (DM) of corn silage (6.0% CP, 1.46 Mcal NEL/kg DM), to complete asupplement with 10.9% CP and 1.56 Mcal NEL/kg DM. In both treatments, all feeds were administeredin individual feeders. After parturition, both groups received a common diet, that consisted in 7.0 kg(DM) of a commercial concentrate (19.3% CP, 1.70 Mcal NEL/kg DM) administered twice a day duringeach milking (07:30 and 14:30 h), and 4.7 kg (DM)/day of corn silage (6.3% CP, 1.46 Mcal NEL/kg DM) inindividual feeders, and had access to a daily strip of pasture containing alfalfa (Medicago sativa), whiteclover (Trifolium repens), and tall fescue (Festuca arundinacea) (21.1% CP, 1.45 Mcal NEL/kg DM). Thesame operator determined the BCS weekly, from the beginning of the experiment until day 49 PP. Milkproduction was recorded daily and averaged for each week. A composite sample of four consecutivemilkings during each week was taken during the first 5 weeks postpartum to determine fat and proteinpercentage with a Bentley 2000 (Bentley Instruments Inc., Chaska, MN 55318, USA).
2.2. Determination of dry matter intake
Total dry matter intake (TDMI) was estimated during weeks −2, −1 (prepartum), 1 and 2 (postpar-tum) relative to parturition. Silage (SDMI) and concentrate (CDMI) dry matter intake were determinedby difference between the amounts offered minus the amounts refused. Pasture dry matter intake(PDMI) was estimated using chromium oxide (Cr2O3) as an external marker. From day 21 beforeexpected calving to day 16 PP, each cow was orally dosed with 14 g of chromium oxide in a gelatinecapsule (Torpac, Fairfield, NJ, USA) once daily after AM concentrate feeding (both during the pre- andpostpartum) and fecal samples were taken from days −15 to −12, −8 to −5, 6 to 9 and 13 to 16, whichwere considered as weeks −2, −1, 1 and 2, respectively. Fecal samples were collected at 15:00 h duringthe prepartum period and before PM milking during the PP, and were dried, ground through a 1-mmscreen, and analyzed to determine chromium (Cr) content by atomic absorption spectrophotometer(PerkinElmer 3300, PerkinElmer, Wellesley, MA, USA). Total organic matter fecal output (OMFO) wascalculated as follows: total OMFO = [(g of Cr dosed per day)/(g of Cr/g of fecal DM)] × % organic matter(OM) in feces. The OMFO from other feeds than pastures was calculated as: OMFO of feed X = OM intakeof X feed × (1 − in vitro OM digestibility (IVOMD) of feed X). The OMFO from pasture was estimated bydifference between total OMFO and OMFO from the other feeds, and then used to determine PDMI asfollows: PDMI = [kg OMFO from pasture/(1 − pasture IVOMD)]/% pasture OM. To estimate the IVOMDof the pasture consumed by the cows in the pre- and postpartum periods, 15 quadrates (0.2 m × 0.5 m)were cut at a height of 5 cm during each fecal extraction period, dried and ground through a 1-mmscreen, and then analyzed following the procedure of Tilley and Terry (1963). Samples of every feedwere taken during each fecal extraction period and together with fecal samples were analyzed for ashto estimate fecal OM content.
2.3. Determination of postpartum ovarian activity
Follicular development was monitored by three times per week ovarian ultrasonography using a7.5 MHz linear probe (Aloka 500, Aloka, Tokyo, Japan) from day 8 to 50 postpartum, and a blood sample

4 D. Cavestany et al. / Animal Reproduction Science 114 (2009) 1–13
was taken each time for IGF-I determination. All follicles visible on the screen were measured andovarian maps drawn for their subsequent analysis. Follicles were classified according to their diameterin size classes (Class I: 2–5 mm; Class II: 6–9 mm; Class III: 10–14 mm; Largest: >15 mm). In animals thatdid not have ovulations from the first PP dominant follicle the evaluation of follicle growth continueduntil ovulation or 49 days PP (whatever occurred first). After ovulation, ultrasonography was carriedout weekly, and a blood sample was collected to determine progesterone concentrations. Ovulationwas determined by the disappearance of the largest follicle followed by the formation of a corpusluteum on the same location, which was confirmed by increased plasma progesterone concentrations.The re-initiation of ovarian cycles was defined as the day when progesterone increased from basalconcentrations in two consecutive samples of >1.6 nmol/L or one sample of >3.2 nmol/L (Meikle et al.,2004). The interval from calving to ovulation for cows not ovulating at the end of the ultrasonographicperiod was arbitrarily assigned as 49 days. The reproductive variable evaluated was days from calvingto first ovulation.
2.4. Blood biochemistry and hormone determination
Blood samples were obtained weekly in the morning, before feeding concentrates, by jugu-lar venipuncture in heparinized vacutainers, from day −21 to 49 (parturition: day 0), centrifuged,plasma separated, and stored at −20 ◦C until analysis. Metabolites were analyzed on a Vitalab Spectra2 autoanalyzer (Vital Scientific NV, Dieren, The Netherlands) according to the following method-ologies: �-hydroxybutyrate (BHB) was determined by a D-3-Hydroxybutyrate kit (Ranbut, RandoxLab, Ardmore, UK), and NEFA by ACS-ACOD (acyl-CoA synthetase and acyl-CoA-oxydase) (WakoChemicals, Richmond, VA 23237, USA) in the Veterinary Diagnostic Laboratory “Miguel C. Rubino”(DILAVE, Uruguay). The intra- and inter-assay CV was ≤3.7% and ≤9.6% for all variables, respec-tively.
The plasma sample corresponding to the day when the dominant follicle of the first follicular waveattained its maximum diameter was analyzed for IGF-I at the School of Agriculture, Food Scienceand Veterinary Medicine, University College Dublin, Ireland, by a previously validated RIA (Spicer etal., 1988). Mean inter-assay (n = 6) and intra-assay (n = 4) CV for a pooled plasma sample containing11.3 nmol/L IGF-I were 12.9% and 13.3 %, respectively and for a pooled sample containing 38.9 nmol/LIGF-I were 9.6% and 9.7%. Progesterone was determined at the Laboratory of Nuclear Techniques ofthe Faculty of Veterinary, University of Uruguay, with a commercial kit (Coat-a-Count, DPC, DiagnosticProducts Co., Los Angeles, CA, 90045, USA). The assay sensitivity was 0.1 nmol/L and intra- and inter-assay CV were 6% and 11%, respectively.
2.5. Statistical analysis
Milk production, body condition score, metabolites, hormone concentrations, and follicle growthwere analyzed by the PROC MIXED procedure (SAS v. 9.1, Statistical Analysis System, SAS Institute,Cary NC, USA, 2007) and the model included as fixed effects: treatment, parity, weekly observa-tions, and their interactions. The covariance structure was autoregressive order 1 and cow withintreatment was set as a random effect. Intake was analyzed with the same model but separatelyfor the prepartum and postpartum period. Concentrations of IGF-I and reproductive variables wereanalyzed by the PROC GLM (SAS) and the effects included in each model were treatment, par-ity and their interaction. Previous results are presented as least square mean ± S.E.M., and werecompared with LSD test. The proportion of cows having ovulations during the first wave of ovar-ian follicular development postpartum was analyzed with a generalized linear model (GENMODprocedure, binomial distribution), which included the effects of treatment, parity and its inter-action. Results are presented as the probability for a cow to have ovulations during the firstwave of ovarian follicular development, and the 95% confidence interval for that probability. Pear-son’s correlation coefficient was calculated to study the relationship between IGF-I concentrationsand diameter of dominant follicle of the first follicular wave. Significance was always declared atP < 0.05.
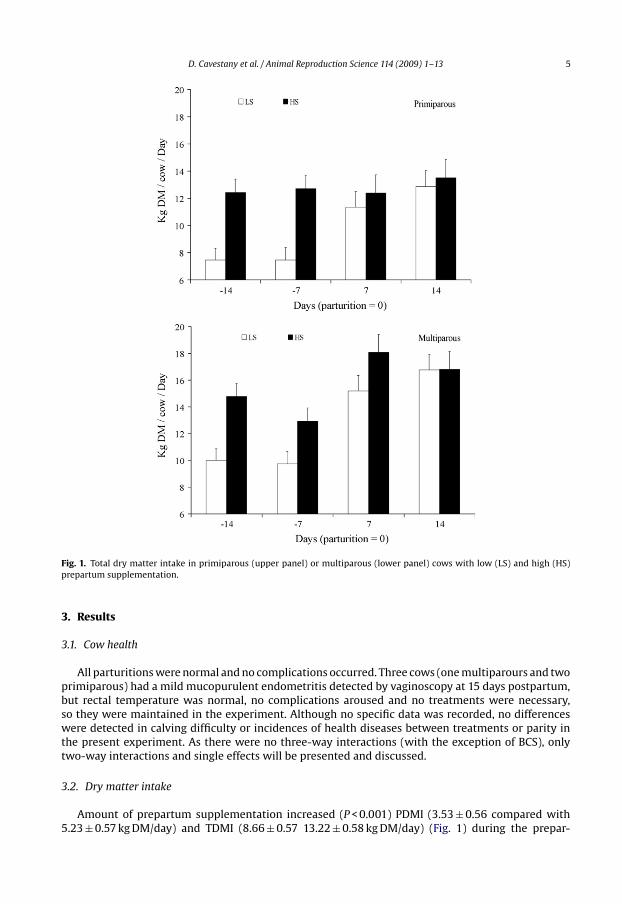
D. Cavestany et al. / Animal Reproduction Science 114 (2009) 1–13 5
Fig. 1. Total dry matter intake in primiparous (upper panel) or multiparous (lower panel) cows with low (LS) and high (HS)prepartum supplementation.
3. Results
3.1. Cow health
All parturitions were normal and no complications occurred. Three cows (one multiparours and twoprimiparous) had a mild mucopurulent endometritis detected by vaginoscopy at 15 days postpartum,but rectal temperature was normal, no complications aroused and no treatments were necessary,so they were maintained in the experiment. Although no specific data was recorded, no differenceswere detected in calving difficulty or incidences of health diseases between treatments or parity inthe present experiment. As there were no three-way interactions (with the exception of BCS), onlytwo-way interactions and single effects will be presented and discussed.
3.2. Dry matter intake
Amount of prepartum supplementation increased (P < 0.001) PDMI (3.53 ± 0.56 compared with5.23 ± 0.57 kg DM/day) and TDMI (8.66 ± 0.57 13.22 ± 0.58 kg DM/day) (Fig. 1) during the prepar-
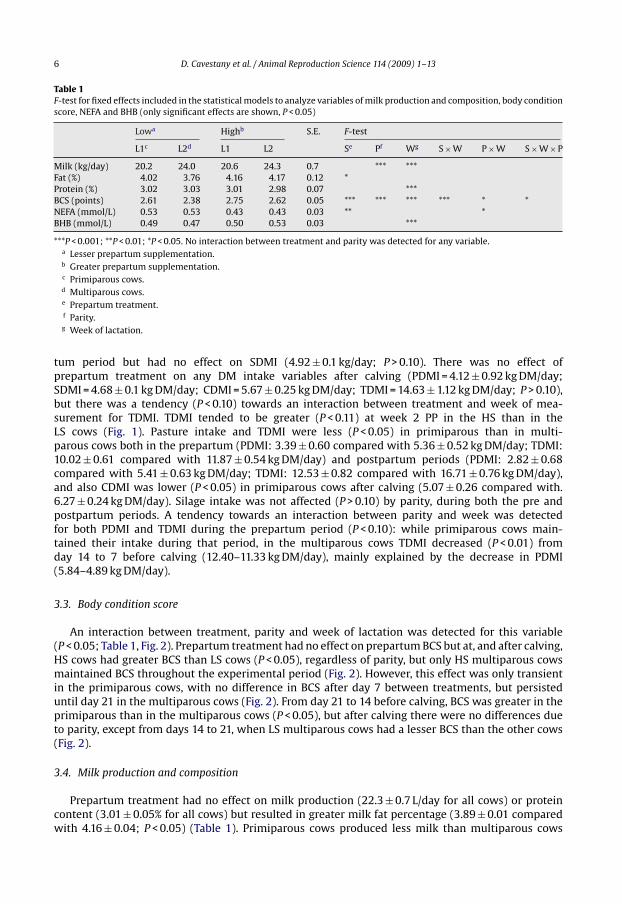
6 D. Cavestany et al. / Animal Reproduction Science 114 (2009) 1–13
Table 1F-test for fixed effects included in the statistical models to analyze variables of milk production and composition, body conditionscore, NEFA and BHB (only significant effects are shown, P < 0.05)
Lowa Highb S.E. F-test
L1c L2d L1 L2 Se Pf Wg S × W P × W S × W × P
Milk (kg/day) 20.2 24.0 20.6 24.3 0.7 *** ***Fat (%) 4.02 3.76 4.16 4.17 0.12 *Protein (%) 3.02 3.03 3.01 2.98 0.07 ***BCS (points) 2.61 2.38 2.75 2.62 0.05 *** *** *** *** * *NEFA (mmol/L) 0.53 0.53 0.43 0.43 0.03 ** *BHB (mmol/L) 0.49 0.47 0.50 0.53 0.03 ***
***P < 0.001; **P < 0.01; *P < 0.05. No interaction between treatment and parity was detected for any variable.a Lesser prepartum supplementation.b Greater prepartum supplementation.c Primiparous cows.d Multiparous cows.e Prepartum treatment.f Parity.g Week of lactation.
tum period but had no effect on SDMI (4.92 ± 0.1 kg/day; P > 0.10). There was no effect ofprepartum treatment on any DM intake variables after calving (PDMI = 4.12 ± 0.92 kg DM/day;SDMI = 4.68 ± 0.1 kg DM/day; CDMI = 5.67 ± 0.25 kg DM/day; TDMI = 14.63 ± 1.12 kg DM/day; P > 0.10),but there was a tendency (P < 0.10) towards an interaction between treatment and week of mea-surement for TDMI. TDMI tended to be greater (P < 0.11) at week 2 PP in the HS than in theLS cows (Fig. 1). Pasture intake and TDMI were less (P < 0.05) in primiparous than in multi-parous cows both in the prepartum (PDMI: 3.39 ± 0.60 compared with 5.36 ± 0.52 kg DM/day; TDMI:10.02 ± 0.61 compared with 11.87 ± 0.54 kg DM/day) and postpartum periods (PDMI: 2.82 ± 0.68compared with 5.41 ± 0.63 kg DM/day; TDMI: 12.53 ± 0.82 compared with 16.71 ± 0.76 kg DM/day),and also CDMI was lower (P < 0.05) in primiparous cows after calving (5.07 ± 0.26 compared with.6.27 ± 0.24 kg DM/day). Silage intake was not affected (P > 0.10) by parity, during both the pre andpostpartum periods. A tendency towards an interaction between parity and week was detectedfor both PDMI and TDMI during the prepartum period (P < 0.10): while primiparous cows main-tained their intake during that period, in the multiparous cows TDMI decreased (P < 0.01) fromday 14 to 7 before calving (12.40–11.33 kg DM/day), mainly explained by the decrease in PDMI(5.84–4.89 kg DM/day).
3.3. Body condition score
An interaction between treatment, parity and week of lactation was detected for this variable(P < 0.05; Table 1, Fig. 2). Prepartum treatment had no effect on prepartum BCS but at, and after calving,HS cows had greater BCS than LS cows (P < 0.05), regardless of parity, but only HS multiparous cowsmaintained BCS throughout the experimental period (Fig. 2). However, this effect was only transientin the primiparous cows, with no difference in BCS after day 7 between treatments, but persisteduntil day 21 in the multiparous cows (Fig. 2). From day 21 to 14 before calving, BCS was greater in theprimiparous than in the multiparous cows (P < 0.05), but after calving there were no differences dueto parity, except from days 14 to 21, when LS multiparous cows had a lesser BCS than the other cows(Fig. 2).
3.4. Milk production and composition
Prepartum treatment had no effect on milk production (22.3 ± 0.7 L/day for all cows) or proteincontent (3.01 ± 0.05% for all cows) but resulted in greater milk fat percentage (3.89 ± 0.01 comparedwith 4.16 ± 0.04; P < 0.05) (Table 1). Primiparous cows produced less milk than multiparous cows
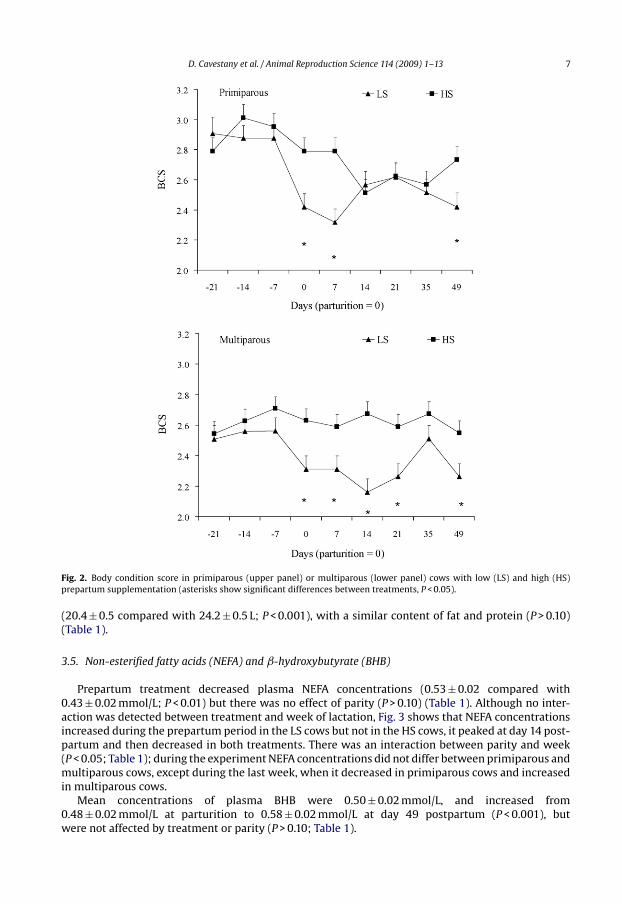
D. Cavestany et al. / Animal Reproduction Science 114 (2009) 1–13 7
Fig. 2. Body condition score in primiparous (upper panel) or multiparous (lower panel) cows with low (LS) and high (HS)prepartum supplementation (asterisks show significant differences between treatments, P < 0.05).
(20.4 ± 0.5 compared with 24.2 ± 0.5 L; P < 0.001), with a similar content of fat and protein (P > 0.10)(Table 1).
3.5. Non-esterified fatty acids (NEFA) and ˇ-hydroxybutyrate (BHB)
Prepartum treatment decreased plasma NEFA concentrations (0.53 ± 0.02 compared with0.43 ± 0.02 mmol/L; P < 0.01) but there was no effect of parity (P > 0.10) (Table 1). Although no inter-action was detected between treatment and week of lactation, Fig. 3 shows that NEFA concentrationsincreased during the prepartum period in the LS cows but not in the HS cows, it peaked at day 14 post-partum and then decreased in both treatments. There was an interaction between parity and week(P < 0.05; Table 1); during the experiment NEFA concentrations did not differ between primiparous andmultiparous cows, except during the last week, when it decreased in primiparous cows and increasedin multiparous cows.
Mean concentrations of plasma BHB were 0.50 ± 0.02 mmol/L, and increased from0.48 ± 0.02 mmol/L at parturition to 0.58 ± 0.02 mmol/L at day 49 postpartum (P < 0.001), butwere not affected by treatment or parity (P > 0.10; Table 1).

8 D. Cavestany et al. / Animal Reproduction Science 114 (2009) 1–13
Fig. 3. Milk fat percentage (upper panel) and plasma NEFA concentrations (lower panel) in cows with low (LS) and high (HS)prepartum supplementation (primiparous and multiparous cows pooled).
3.6. Re-initiation of ovarian activity
There were no effects of treatment or parity, or interaction between the two factors, in the numberof follicles in Class I (12.3 ± 0.4 mm), Class II (2.6 ± 0.2), Class III (0.6 ± 0.1) or >15 mm (0.10 ± 0.02)(P > 0.10). From day 8 to 16 PP, total number of follicles of Class I increased from 11.3 ± 0.8 to 12.7 ± 0.8;Class II from 2.5 ± 0.4 to 3.0 ± 0.4; Class III from 0.5 ± 0.2 to 0.7 ± 0.1; and the total number of follicles(>2 mm) during this period increased from 14.3 ± 0.9 to 16.4 ± 0.9 (P < 0.05). Most variables describ-ing the characteristics of re-initiation of postpartum ovarian activity were not affected (P > 0.10) bytreatment, parity or their interaction, as shown in Table 2, but an interaction between treatment andparity was detected for the interval from calving to first ovulation (P < 0.05); a greater prepartum sup-plementation reduced the length of the interval from calving to first ovulation in the multiparous(P < 0.05) but not in the primiparous cows (P > 0.10). However, the proportion of animals ovulatingthe dominant follicle of the first wave PP was not affected by treatment, parity or their interaction(P > 0.10), although it was numerically less in the multiparous cows of the LS group than in the othercows (Table 2).

D. Cavestany et al. / Animal Reproduction Science 114 (2009) 1–13 9
Table 2F-test for fixed effects included in the statistical models to analyze characteristics of the postpartum ovarian follicular dynamics(only significant effects are shown, P < 0.05)
Lowc Highd S.E. F-test
L1e L2f L1 L2 Sg Ph S × P
Diameter of the dominant follicle of wave 1 atday 8 PP (mm)
9.0 7.3 8.1 7.7 1.0
Maximum diameter of the first wave dominantfollicle (mm)
14.5 14.1 13.0 14.5 0.8
Day of maximum diameter of the first wavedominant follicle
14.7 14.1 15.2 14.5 0.8
Day of end of the first follicular wave(ovulation or regression of the follicle)
21.1 21.1 23.8 18.5 1.9
Number of waves for cows ovulating beforeday 50 PP
1.4 1.4 1.1 1.2 0.2
Number of waves for cows not havingovulations (by day 49 PP)
3.7 3.7 3.3 4.5 0.5
Proportion of cows ovulating from thedominant follicle of the first follicular wave
0.55 0.27 0.50 0.72 –i
Days from calving to first ovulation 33.6a 38.2a 37.8a 22.9b 4.6 *Plasma IGF-I concentrations by day of
maximum diameter of the first wavedominant follicle
7.1a 3.6b 6.4a 8.0a 0.1 0.06 **
**P < 0.01; *P < 0.05.a,bMeans with different superscripted letters within a row differ, P < 0.05.
c Lesser prepartum supplementation.d Greater prepartum supplementation.e Primiparous cows.f Multiparous cows.g Prepartum treatment.h Parity.i 95% confidence intervals for low-L1, low-L2, high-L1 and high-L2 were [0.23–0.83], [0.07–0.64], [0.21–0.79] and [0.36–0.93],
respectively.
3.7. IGF-I concentrations
An interaction was detected between treatment and parity for plasma IGF-I concentrations at thetime when the dominant follicle of the first follicular wave postpartum achieved its maximum diameter(P < 0.01; Table 2). Plasma concentrations were less in multiparous cows in the LS group than in cowsfrom other treatment × parity groups, which did no differ among them (P > 0.10).
3.8. Characteristics of cows ovulating during the experiment
When data from all cows were combined, the interval from parturition to first ovulation was33.1 ± 4.6 days. Only 66% of the cows (29/44) had ovulations before day 49 postpartum, and thesehad a greater BCS (2.70 ± 0.07 compared with 2.40 ± 0.07; P < 0.01) at calving and a larger follicle(9.5 ± 0.5 mm compared with 6.9 ± 0.7 mm; P < 0.01) on day 7 to 9 postpartum than those that did nothave ovulations during this period. In cows that had ovulations, the dominant follicle of the first follic-ular wave was larger (15.7 ± 0.4 mm compared with 14.1 ± 0.6 mm; P < 0.05), and IGF-I concentrationswere greater than in those not having ovulations (8.0 ± 0.6 nmol/L compared with 4.2 ± 0.1 nmol/L;P < 0.05). NEFA concentrations on days 35 and 49 postpartum were less in cows that had ovulations(day 35: 0.8 ± 0.2 nmol/L and day 49: 0.9 ± 0.2 nmol/L), compared with those that did not have ovu-lations (day 35: 2.1 ± 0.3 nmol/L and day 49: 1.8 ± 0.3 nmol/L; P < 0.05). As shown in Fig. 4 there wasa significant positive correlation between the size of the dominant follicle and IGF-I concentrations(r = 0.55, P < 0.001, n = 44).
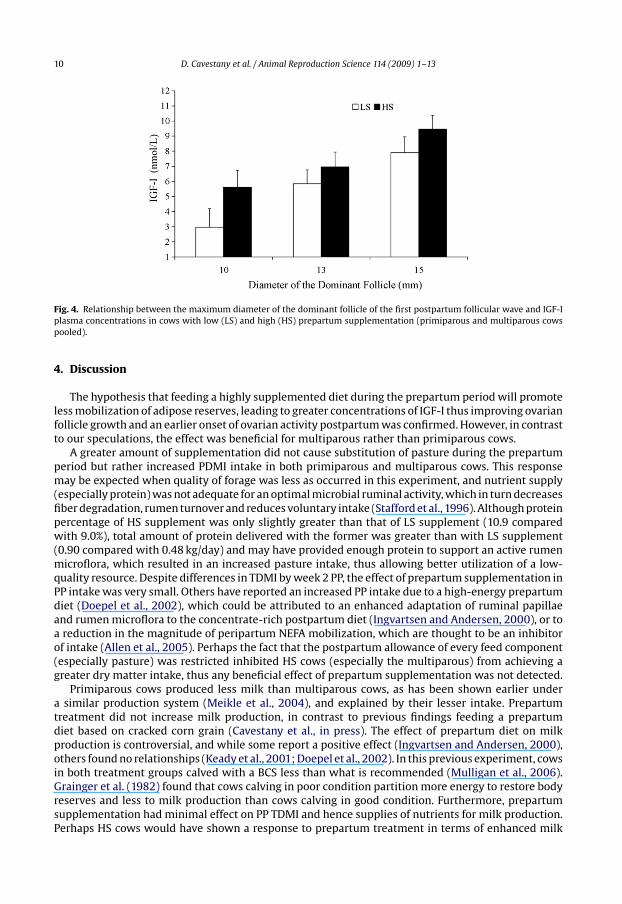
10 D. Cavestany et al. / Animal Reproduction Science 114 (2009) 1–13
Fig. 4. Relationship between the maximum diameter of the dominant follicle of the first postpartum follicular wave and IGF-Iplasma concentrations in cows with low (LS) and high (HS) prepartum supplementation (primiparous and multiparous cowspooled).
4. Discussion
The hypothesis that feeding a highly supplemented diet during the prepartum period will promoteless mobilization of adipose reserves, leading to greater concentrations of IGF-I thus improving ovarianfollicle growth and an earlier onset of ovarian activity postpartum was confirmed. However, in contrastto our speculations, the effect was beneficial for multiparous rather than primiparous cows.
A greater amount of supplementation did not cause substitution of pasture during the prepartumperiod but rather increased PDMI intake in both primiparous and multiparous cows. This responsemay be expected when quality of forage was less as occurred in this experiment, and nutrient supply(especially protein) was not adequate for an optimal microbial ruminal activity, which in turn decreasesfiber degradation, rumen turnover and reduces voluntary intake (Stafford et al., 1996). Although proteinpercentage of HS supplement was only slightly greater than that of LS supplement (10.9 comparedwith 9.0%), total amount of protein delivered with the former was greater than with LS supplement(0.90 compared with 0.48 kg/day) and may have provided enough protein to support an active rumenmicroflora, which resulted in an increased pasture intake, thus allowing better utilization of a low-quality resource. Despite differences in TDMI by week 2 PP, the effect of prepartum supplementation inPP intake was very small. Others have reported an increased PP intake due to a high-energy prepartumdiet (Doepel et al., 2002), which could be attributed to an enhanced adaptation of ruminal papillaeand rumen microflora to the concentrate-rich postpartum diet (Ingvartsen and Andersen, 2000), or toa reduction in the magnitude of peripartum NEFA mobilization, which are thought to be an inhibitorof intake (Allen et al., 2005). Perhaps the fact that the postpartum allowance of every feed component(especially pasture) was restricted inhibited HS cows (especially the multiparous) from achieving agreater dry matter intake, thus any beneficial effect of prepartum supplementation was not detected.
Primiparous cows produced less milk than multiparous cows, as has been shown earlier undera similar production system (Meikle et al., 2004), and explained by their lesser intake. Prepartumtreatment did not increase milk production, in contrast to previous findings feeding a prepartumdiet based on cracked corn grain (Cavestany et al., in press). The effect of prepartum diet on milkproduction is controversial, and while some report a positive effect (Ingvartsen and Andersen, 2000),others found no relationships (Keady et al., 2001; Doepel et al., 2002). In this previous experiment, cowsin both treatment groups calved with a BCS less than what is recommended (Mulligan et al., 2006).Grainger et al. (1982) found that cows calving in poor condition partition more energy to restore bodyreserves and less to milk production than cows calving in good condition. Furthermore, prepartumsupplementation had minimal effect on PP TDMI and hence supplies of nutrients for milk production.Perhaps HS cows would have shown a response to prepartum treatment in terms of enhanced milk

D. Cavestany et al. / Animal Reproduction Science 114 (2009) 1–13 11
production if total feed allowance had not been restricted. The increased milk fat percentage in HScows could be explained by the increased BCS at calving due to prepartum supplementation, whichhas been positively related to milk fat content in the subsequent lactation (Grainger et al., 1982) or toa more efficient use of available NEFA from circulation.
The reduced prepartum intake in LS compared with HS cows probably resulted in a poorer energybalance in the former that led to early mobilization of their reserves, and was reflected in a lesser BCSand increased NEFA concentrations before calving. This situation persisted after calving, but the lesserBCS in LS multiparous cows than in the other animals suggests that NEB was more pronounced in theformer. Total dry matter intake was reduced during the last week prepartum in the multiparous but notin the primiparous cows. This effect may have had a more adverse effect in LS than in HS multiparouscows, because the former could not compensate the decrease in TDMI with a more nutrient-densediet, as did the HS cows. Body condition score did not recover by the end of the experiment in anytreatment × parity group, which suggests that they had not yet recovered from NEB.
The concentrations of BHB, which reflects the conversion of NEFA to ketone bodies induced by theenergetic demands of parturition and onset of lactation (Ingvartsen and Andersen, 2000), remainedgreater for a longer period after calving than reported by Moorby et al. (2000) and Cavestany et al.(2005), probably due to differences in diet composition or dry matter intake. The long-term increasein BHB reflects the slow recovery in BCS in all cows and further suggests that postpartum feed allowancerestricted excessively for these cows.
Most characteristics of the dominant follicle and the first follicular wave were not affected by theprepartum treatment. Although it did not reach statistical significance, probably due to the smallnumber of cows used in the experiment, the proportion of multiparous cows that had ovulations fromthe first dominant follicle was numerically greater in the HS than in the LS cows, and this was reflectedin a shorter interval to first ovulation postpartum in HS cows. This was in agreement with previousreports for dairy cows in confined production systems (Shrestha et al., 2004). Moreover, the greaterconcentrations of IGF-I in HS than in LS multiparous cows reinforces the key role that IGF-I plays onfollicle development (Spicer et al., 2002; Webb et al., 2004). However, despite having a longer intervalfrom calving to first ovulation, IGF-I concentrations in primiparous cows did not differ from that ofHS multiparous cows, indicating that the association between IGF-I and the length of the intervalfrom calving to first ovulation is not found in all situations (Chagas et al., 2006), and that other bloodtraits should also be considered to predict the moment of onset of postpartum ovarian activity, atleast in primiparous cows. The delayed resumption of ovarian activity in the LS multiparous cowsstrengthens the idea that these animals were under more severe NEB than the other cows, which wasalso reflected in their lesser plasma IGF-I concentrations. Situations of NEB are associated with greaterGH concentrations but a decrease in its hepatic receptors; thus causing a decrease in IGF-I production(Butler, 2003). When cows start to recover from NEB, the associated increase in insulin concentrationsinduces the GH hepatic receptor and stimulates IGF-I production (Butler, 2003). Although insulin wasnot measured in this study, our results suggest that HS but not LS multiparous cows recovered fromNEB very soon after parturition, situation thus promoting an enhanced endocrine environment forfollicle maturation and ovulation.
In grazing conditions, primiparous cows calving with a BCS < 3 have a late resumption of ovariancycles in comparison to primiparous cows that calve with a BCS > 3 (Meikle et al., 2004). In this experi-ment, although a high-supplemented prepartum diet resulted in a less acute loss of BCS around calving,both HS and LS primiparous cows calved with a BCS < 3, suggesting that treatment was not sufficientto maintain adequate body reserves and thus to promote a fast recovery from NEB after calving, andexplains the lack of treatment effect on the interval from calving to first postpartum ovulation betweenprimiparous cows. Because energy demands are greater in primiparous than in multiparous cows, itmay be necessary to combine a high-supplemented prepartum energy diet with a high-supplementedpostpartum diet, to stimulate a faster recovery from NEB and a fast resumption of postpartum ovarianactivity.
Early resumption of ovarian activity is important to allow for more luteal phases before the firstservice which has been shown to increase fertility of cows in both confined (Townson et al., 2002)and grazing (Cavestany et al., 2001) dairy production systems. In the present study, only 66% of thecows had ovulations before day 49 postpartum and the interval from parturition to first ovulation was

12 D. Cavestany et al. / Animal Reproduction Science 114 (2009) 1–13
longer than previous reports (Savio et al., 1990; Rajamahendran and Taylor, 1990). When data fromall cows were combined, those cows that had ovulations before day 50 postpartum had an enhancedBCS at calving and less mobilization of fat reserves postpartum associated with greater IGF-I concen-trations that stimulated the development of a larger dominant follicle before ovulation. In the presentstudy, there was a positive correlation between IGF-I and the size of the dominant follicle, which isin agreement with reports by Beam and Butler (1999). IGF-I has direct effects at the ovary, regulatingthe size, estradiol production and health of dominant follicles (Austin et al., 2001; Canty et al., 2006).These results are consistent with the findings in multiparous supplemented cows, reinforcing theconcept that less severe NEB and fast recovery from NEB are associated with enhanced reproductiveperformance (Beam and Butler, 1999).
5. Conclusions
Greater supplementation during the prepartum period in multiparous cows promoted themaintenance of body condition score postpartum, which was associated with greater circulating con-centrations of IGF-I and an earlier onset of estrous cycles in the present study. The high-supplementedprepartum diet increased milk fat content but did not alter milk production or protein content.
Acknowledgments
The authors are thankful to Dr. Ana Meikle from the Laboratory of Nuclear Techniques of the Facultyof Veterinary of Montevideo, Uruguay, for the progesterone analysis, Dr. Gonzalo Uriarte of DILAVE,Miguel Rubino, for the analysis of the metabolites, and Niamh Hynes (University College Dublin) forassistance with IGF-I assays. The help of Federica Albanell, Sebastián Belassi, Federico Olariaga, MaríaNoel Pérez, Anita Silva and Ignacio Torres for their participation in the fieldwork is very much appre-ciated. The trial was financed partially by INIA, and by a grant to D. Cavestany of the CSIC, Universityof Uruguay.
References
Allen, M.S., Bradford, B.J., Harvatine, K.J., 2005. The cow as a model to study food intake regulation. Ann. Rev. Nutr. 25, 523–547.Austin, E.J., Mihm, M., Evans, A.C.O., Knight, P.G., Ireland, J.L.H., Ireland, J.J., Roche, J.F., 2001. Alterations in intrafollicular regulatory
factors and apoptosis during selection of follicles in the first follicular wave of the bovine oestrous cycle. Biol. Reprod. 64,839–848.
Beam, S.W., Butler, W.R., 1999. Effects of energy balance on follicular development and first ovulation in postpartum dairy cows.J. Reprod. Fertil. (Suppl.) 54, 411–424.
Bell, A., 1995. Regulation of organic nutrient metabolism during transition from late pregnancy to early lactation. J. Anim. Sci.73, 2804–2819.
Butler, W.R., 2003. Energy balance relationships with follicular development, ovulation and fertility in postpartum dairy cows.Livest. Prod. Sci. 83, 211–218.
Canfield, R.W., Butler, W.R., 1991. Energy balance, first ovulation and the effects of naloxone on LH secretion in early postpartumdairy cows. J. Anim. Sci. 69, 740–746.
Canty, M.J., Boland, M.P., Evans, A.C.O., Crowe, M.A., 2006. Alterations in follicular IGFBP-2, -3 and -4 mRNA expression andintrafollicular IGFBP concentrations during the first follicle wave in beef heifers. Anim. Reprod. Sci. 93, 199–217.
Cavestany, D., Blanc, J.E., Kulcsár, M., Uriarte, G., Chilibroste, P., Meikle, A., Febel, H., Ferraris, A., Krall, E., 2005. Studies of thetransition cow under a pasture-based milk production system: Metabolic profiles. J. Vet. Med. Ser. A 52, 1–7.
Cavestany, D., Galina, C.S., Vinoles, C., 2001. Efecto de las características del reinicio de la actividad ovárica posparto en laeficiencia reproductiva de vacas Holstein en pastoreo. Arch. Med. Vet. (Chile) 33, 217–226.
Cavestany, D., Kulcsár, M., Crespi, D., Chilliard, Y., La Manna, A., Balogh, O., Keresztes, M., Delavaud, C., Huszenicza, G., Meikle, A.,in press. Effect of prepartum energetic supplementation on productive and reproductive characteristics, and metabolic andhormonal profiles in dairy cows under grazing conditions. Reprod. Domest. Anim. doi:10.1111/j.1439-0531.2007.01044.x.
Chagas, L.M., Rhodes, F.M., Blache, D., Gore, P.J.S., Macdonald, K.A., Verkerk, G.A., 2006. Precalving effects on metabolic responsesand postpartum anestrus in grazing primiparous dairy cows. J. Dairy Sci. 89, 1981–1989.
Doepel, L., Lapierre, H., Kennelly, J.J., 2002. Peripartum performance and metabolism of dairy cows in response to prepartumenergy and protein intake. J. Dairy Sci. 85, 2315–2334.
Edmonson, A.J., Lean, I.J., Weaver, L.D., Farver, T., Webster, G., 1989. A body scoring chart for Holstein dairy cows. J. Dairy Sci. 72,68–78.
Grainger, C., Wilhelms, G.D., McGowan, A.A., 1982. Effect of body condition at calving and level of feeding in early lactation onmilk production of dairy cows. Aust. J. Exp. Agric. Anim. Husb. 22, 9–17.
Grummer, R.R., 1995. Impact of changes in organic nutrient metabolism on feeding the transition cow. J. Anim. Sci. 73, 2820–2833.

D. Cavestany et al. / Animal Reproduction Science 114 (2009) 1–13 13
Holcomb, C.S., Van Horn, H.H., Head, H.H., Hall, M.B., Wilcox, C.J., 2001. Effects of prepartum dry matter intake and foragepercentage on postpartum performance of lactating dairy cows. J. Dairy Sci. 84, 2051–2058.
Huszenicza, G., Kulcsár, M., Nikolic, J.A., Schmidt, J., Korodi, P., Katai, L., Dieleman, S., Ribiczei-Szabo, P., Rudas, P., 2001. Plasmaleptin concentration and its interrelation with some blood metabolites, metabolic hormones and the resumption of cyclicovarian function in postpartum dairy cows supplemented with monensin or inert fat in feed. In: Diskin, M.G. (Ed.), In Fertilityin the High-producing Dairy Cow. British Society of Animal Science, Edinburgh, UK, pp. 405–409.
Ingvartsen, K.L., Andersen, J.B., 2000. Integration of metabolism and intake regulation: a review focusing on periparturientanimals. J. Dairy Sci. 83, 1573–1597.
Keady, T.W.J., Mayne, C.S., Fitzpatrick, D.A., McCoy, M.A., 2001. Effect of concentrate feed level in late gestation on subsequentmilk yield, milk composition, and fertility of dairy cows. J. Dairy Sci. 84, 1468–1479.
Meikle, A., Kulcsar, M., Chilliard, Y., Febel, H., Delavaud, C., Cavestany, D., Chilibroste, P., 2004. Effects of parity and body conditionat parturition on endocrine and reproductive parameters of the cow. Reproduction 127, 727–737.
Moorby, J.M., Dewhurst, R.J., Tweed, J.K.S., Dhanoa, M.S., Beck, N.F.G., 2000. Effects of altering the energy and protein supply ondairy cows during the dry period. 2. Metabolic and hormonal responses. J. Dairy Sci. 83, 1795–1805.
Mulligan, F.J., O’Grady, L., Rice, D.A., Doherty, M.L., 2006. A herd health approach to dairy cow nutrition and production diseasesof the transition cow. Anim. Reprod. Sci. 96, 331–353.
Pushpakumara, P.G., Gardner, N.H., Reynolds, C.K., Beever, D.E., Wathes, D.C., 2003. Relationships between transition period diet,metabolic parameters and fertility in lactating dairy cows. Theriogenology 60, 1165–1185.
Rajamahendran, R., Taylor, C., 1990. Characterization of ovarian activity in postpartum dairy cows using ultrasound imaging andprogesterone profiles. Anim. Reprod. Sci. 22, 171–180.
Savio, J.D., Boland, M.P., Hynes, N., Roche, J.F., 1990. Resumption of follicular activity in the early post-partum period in dairycows. J. Reprod. Fertil. 88, 569–579.
Shrestha, H.K., Nakao, T., Higaki, T., Suzuki, T., Akita, M., 2004. Resumption of postpartum ovarian cyclicity in high-producingHolstein cows. Theriogenology 61, 637–649.
Spicer, L.J., Chase Jr., C.C., Rutter, L.M., 2002. Relationship between serum insulin-like growth factor-I and genotype during thepostpartum interval in beef cows. J. Anim. Sci. 80, 716–722.
Spicer, L.J., Echternkamp, S.E., Canning, S.F., Hammond, J.M., 1988. Relationship between concentrations of immunoreactiveinsulin-like growth factor-I in follicular fluid and various biochemical markers of differentiation in bovine antral follicles.Biol. Reprod. 39, 573–580.
Stafford, S.D., Cochran, R.C., Vanzant, E.S., Fritz, J.O., 1996. Evaluation of the potential of supplements to substitute for low-quality,tallgrass-prairie forage. J. Anim. Sci. 74, 639–647.
Tilley, J.M.A., Terry, R.A., 1963. A two-step technique for the in vitro digestion of forage crops. J. Br. Grass. Soc. 18, 104–111.Townson, D.H., Tsang, P.C., Butler, W.R., Frajblat, M., Griel Jr., L.C., Johnson, C.J., Milvae, R.A., Niksic, G.M., Pate, J.L., 2002. Rela-
tionship of fertility to ovarian follicular waves before breeding in dairy cows. J. Anim. Sci. 80, 1053–1058.Webb, R., Garnsworthy, P.C., Gong, J.G., Armstrong, D.G., 2004. Control of follicular growth: Local interactions and nutritional
influences. J. Anim. Sci. 82 (Electronic Supplement), E63–E74.
Copyright © 2022 FDOKUMEN




















