
Effect of elevated CO2 levels on leaf starch, nitrogen and photosynthesis of plants growing at three...
10
ORIGINAL ARTICLE Yusuke Onoda Tadaki Hirose Kouki Hikosaka Effect of elevated CO 2 levels on leaf starch, nitrogen and photosynthesis of plants growing at three natural CO 2 springs in Japan Received: 14 February 2006 / Accepted: 19 July 2006 / Published online: 13 September 2006 ȑ The Ecological Society of Japan 2006 Abstract Plant communities around natural CO 2 springs have been exposed to elevated CO 2 levels over many generations and give us a unique opportunity to inves- tigate the effects of long-term elevated CO 2 levels on wild plants. We searched for natural CO 2 springs in cool temperate climate regions in Japan and found three springs that were suitable for studying long-term re- sponses of plants to elevated levels of CO 2 : Ryuzin- numa, Yuno-kawa and Nyuu. At these CO 2 springs, the surrounding air was at high CO 2 concentration with no toxic gas emissions throughout the growth season, and there was natural vegetation around the springs. At each site, high-CO 2 (HC) and low-CO 2 (LC) plots were established, and three dominant species at the shrub layers were used for physiological analyses. Although the microenvironments were different among the springs, dicotyledonous species growing at the HC plots tended to have more starch and less nitrogen per unit dry mass in the leaves than those growing at the LC plots. In contrast, monocotyledonous species growing in the HC and LC plots had similar starch and nitrogen concentrations. Photosynthetic rates at the mean growth CO 2 concentration were higher in HC plants than LC plants, but photosynthetic rates at a common CO 2 concentration were lower in HC plants. Efficiency of water and nitrogen use of leaves at growth CO 2 con- centration was greatly increased in HC plants. These results suggest that natural plants growing in elevated CO 2 levels under cool temperate climate conditions have down-regulated their photosynthetic capacity but that they increased photosynthetic rates and resource use efficiencies due to the direct effect of elevated CO 2 con- centration. Keywords Down-regulation of photosynthesis Long-term response to elevated CO 2 Photosynthetic nitrogen use efficiency Water use efficiency Wild plants Introduction Over 20 years a number of studies have investigated the response of plants to elevated levels of CO 2 . Open-top chambers (OTC) and free-air CO 2 enrichment (FACE) have been used to obtain a more realistic picture of plant responses to elevated CO 2 concentrations in natural conditions. However, most results came from experi- ments that had run a maximum of 10 years (Ainsworth et al. 2003). Because many ecological processes (plant– plant interaction, microbe activity, decomposition and soil condition) progress much slower and because these changes could also modulate plant responses (e.g., Luo and Reynolds 1999; Kohut 2003), there are still many uncertainties as to how plants respond to long-term (>10 years) elevated levels of CO 2 . The vegetation around natural CO 2 springs is valuable, because it is the only vegetation that has been exposed to high CO 2 concentration for a very long time (Koch 1994; Miglietta et al. 1994; Kohut 2003). Detailed field studies of natural CO 2 springs were started in Italy in the early 1990s. Miglietta and Raschi (1993) and Miglietta et al. (1993a) described several natural CO 2 springs where CO 2 had been emitted from vents in the Mediterranean climate. Studies on plant responses to elevated CO 2 levels have been carried out at CO 2 springs with respect to photosynthesis (Jones et al. 1995), photoinhibition (Stylinski et al. 2000), water Y. Onoda T. Hirose K. Hikosaka Graduate School of Life Sciences, Tohoku University, Aoba, Sendai, Japan Y. Onoda (&) Section of Plant Ecology and Biodiversity, Institute of Environmental Biology, Utrecht University, P. O. Box 800.84, 3508 TB Utrecht, The Netherlands E-mail: [email protected] Tel.: +31-30-2516876 Fax: +31-30-2536700 T. Hirose Department of International Agriculture Development, Tokyo University of Agriculture, Tokyo, Japan Ecol Res (2007) 22: 475–484 DOI 10.1007/s11284-006-0030-z
Transcript of Effect of elevated CO2 levels on leaf starch, nitrogen and photosynthesis of plants growing at three...

ORIGINAL ARTICLE
Yusuke Onoda Æ Tadaki Hirose Æ Kouki Hikosaka
Effect of elevated CO2 levels on leaf starch, nitrogen and photosynthesisof plants growing at three natural CO2 springs in Japan
Received: 14 February 2006 / Accepted: 19 July 2006 / Published online: 13 September 2006� The Ecological Society of Japan 2006
Abstract Plant communities around natural CO2 springshave been exposed to elevated CO2 levels over manygenerations and give us a unique opportunity to inves-tigate the effects of long-term elevated CO2 levels onwild plants. We searched for natural CO2 springs in cooltemperate climate regions in Japan and found threesprings that were suitable for studying long-term re-sponses of plants to elevated levels of CO2: Ryuzin-numa, Yuno-kawa and Nyuu. At these CO2 springs, thesurrounding air was at high CO2 concentration with notoxic gas emissions throughout the growth season, andthere was natural vegetation around the springs. At eachsite, high-CO2 (HC) and low-CO2 (LC) plots wereestablished, and three dominant species at the shrublayers were used for physiological analyses. Althoughthe microenvironments were different among thesprings, dicotyledonous species growing at the HC plotstended to have more starch and less nitrogen per unitdry mass in the leaves than those growing at the LCplots. In contrast, monocotyledonous species growing inthe HC and LC plots had similar starch and nitrogenconcentrations. Photosynthetic rates at the mean growthCO2 concentration were higher in HC plants than LCplants, but photosynthetic rates at a common CO2
concentration were lower in HC plants. Efficiency ofwater and nitrogen use of leaves at growth CO2 con-centration was greatly increased in HC plants. These
results suggest that natural plants growing in elevatedCO2 levels under cool temperate climate conditions havedown-regulated their photosynthetic capacity but thatthey increased photosynthetic rates and resource useefficiencies due to the direct effect of elevated CO2 con-centration.
Keywords Down-regulation of photosynthesis ÆLong-term response to elevated CO2 Æ Photosyntheticnitrogen use efficiency Æ Water use efficiency ÆWild plants
Introduction
Over 20 years a number of studies have investigated theresponse of plants to elevated levels of CO2. Open-topchambers (OTC) and free-air CO2 enrichment (FACE)have been used to obtain a more realistic picture of plantresponses to elevated CO2 concentrations in naturalconditions. However, most results came from experi-ments that had run a maximum of 10 years (Ainsworthet al. 2003). Because many ecological processes (plant–plant interaction, microbe activity, decomposition andsoil condition) progress much slower and because thesechanges could also modulate plant responses (e.g., Luoand Reynolds 1999; Kohut 2003), there are still manyuncertainties as to how plants respond to long-term(>10 years) elevated levels of CO2. The vegetationaround natural CO2 springs is valuable, because it is theonly vegetation that has been exposed to high CO2
concentration for a very long time (Koch 1994; Migliettaet al. 1994; Kohut 2003).
Detailed field studies of natural CO2 springs werestarted in Italy in the early 1990s. Miglietta and Raschi(1993) and Miglietta et al. (1993a) described severalnatural CO2 springs where CO2 had been emitted fromvents in the Mediterranean climate. Studies on plantresponses to elevated CO2 levels have been carried out atCO2 springs with respect to photosynthesis (Jones et al.1995), photoinhibition (Stylinski et al. 2000), water
Y. Onoda Æ T. Hirose Æ K. HikosakaGraduate School of Life Sciences, Tohoku University,Aoba, Sendai, Japan
Y. Onoda (&)Section of Plant Ecology and Biodiversity,Institute of Environmental Biology,Utrecht University, P. O. Box 800.84,3508 TB Utrecht, The NetherlandsE-mail: [email protected].: +31-30-2516876Fax: +31-30-2536700
T. HiroseDepartment of International Agriculture Development,Tokyo University of Agriculture, Tokyo, Japan
Ecol Res (2007) 22: 475–484DOI 10.1007/s11284-006-0030-z

relations (Chaves et al. 1995; Jones et al. 1995; Tognettiet al. 1996, 1998, 1999), non-structural carbohydrate(Korner and Miglietta 1994), isoprene emission (Rap-parini et al. 2004; Scholefield et al. 2004), growth (Mi-glietta et al. 1993b), tree-ring expansion (Hattenschwileret al. 1997; Tognetti et al. 2000) and litter quality (Ga-hrooee 1998; Cotrufo et al. 1999). Although a number ofstudies has been done in Italy, only a few comparablestudies have been conducted in different climatic regions(e.g., New Zealand, Newton et al. 1996; Iceland, Cooket al. 1998; Venezuela, Fernandez et al. 1998; Slovenia,Vodnik et al. 2002). Differences in precipitation andtemperature may influence the effect of elevated CO2
levels on plants (Long 1991; Idso and Idso 1994; Drakeet al. 1997). Furthermore, interspecific variations in CO2
responses may be evident across plant communities thatwere established at different CO2 springs (Poorter 1993).Therefore, studies on natural CO2 springs in differentclimates are crucial to generalize plant response to long-term elevated concentrations of CO2.
The Japan Archipelago is situated on the Pacificvolcanic belt and has at least 82 CO2 springs (Kimbara1992; Cook et al. 2000). The islands are influenced by theAsian monsoon climate, with relatively high rainfall andhumidity throughout the year, which is very differentfrom the Mediterranean climate. In this study we aimedto identify natural CO2 springs in Japan as a place tostudy plant responses to CO2 enrichment. We firstsearched CO2 springs that were suitable for biologicalresearch and described the CO2 environment to establishstudy sites. In contrast to the previous studies, whichused one or two springs, we used three CO2 springs,which overcame the general drawbacks of CO2 springstudy such as lack of real control and replication.
In this study we also studied leaf traits of plantsgrowing around natural CO2 springs. As CO2 is a sub-strate for photosynthesis, elevated CO2 concentrationenhances photosynthesis (e.g., Mott 1990). However,when plants grow under prolonged exposure to elevatedCO2 concentrations, this increase is often offset bydown-regulation of photosynthetic capacity (e.g., Longet al. 2004). Accumulation of non-structural carbohy-drates cause down-regulation by decreasing the amountand/or activity of photosynthetic proteins (Sheen 1990,Stitt and Krapp 1999). The extent of down-regulation is
sensitive to growing conditions (e.g., Arp 1991). As thegrowing conditions at CO2 springs would have beeninfluenced by ecosystem level feedbacks that occurredover a very long time (Luo and Reynolds 1999; Hiko-saka 2003), plants growing around CO2 springs can beused to test whether their CO2 responses are differentfrom those obtained in short-term growth experiments.We investigated how much starch was accumulated andto what extent photosynthesis down-regulated in plantsgrowing around three CO2 springs.
Materials and methods
A search for CO2 springs followed former geologicalinvestigations (Kimbara 1992; Tsurumi et al. 1999;Cook et al. 2000). We visited many springs and assessedwhether they could be used for biological research withrespect to the following aspects: (1) the CO2 concen-trations around the springs are continuously at highlevels, (2) toxic gasses for plants (e.g., H2S, SO2) are notemitted or are at levels too low to affect plant growth, (3)the springs are cold, (4) natural vegetation is abundantaround the CO2 spring. From preliminary surveys(1999–2002), three sites (Ryuzin-numa, Yuno-kawa andNyuu) had been found to be suitable for plant scienceresearch and are described in this paper.
Ryuzin-numa and Yuno-kawa are located at 570–575 m above sea level (a.s.l.), at the foot of MountHakkoda in Aomori Prefecture (Table 1; Fig. 1a). Thedistance between the two CO2 springs is 1.0 km,implying that they are likely to share the same source ofCO2. Nyuu is located at 1,000 m a.s.l., at the foot ofMount Gassan in Yamagata Prefecture, about 300 kmsouth of Ryuzin-numa and Yuno-kawa (Fig. 1a). TheseCO2 springs are situated in areas having a cool–tem-perate climate (Table 1), receiving high rainfall fromspring to autumn and a large amount of snow duringwinter. They are located in the National Parks, andnatural vegetation around the springs was generally wellpreserved (Fig. 1).
A CO2 concentration monitoring system wasconstructed with a portable infrared gas analyzer(LI-2820, Li-Cor, Lincoln, Nb., USA, full-scale20,000 lmol mol�1), a six-channel air sampling system
Table 1 Geographical co-ordinates, air pressure, mean air temperature, monthly mean rainfall from June to August in 2003, and theapproximate area where mean CO2 concentration was above 500 lmol mol�1 at three natural CO2 springs
CO2
springsLatitude–longitude Altitude
(m)Airpressure(kPa)
Meantemperaturea
(�C)
Rainfalla
(mm)HighCO2 area(m2)
Ryuzin-numa 40�41¢N–140�55¢E 575 95 17.1 186 >300Yuno-kawa 40�41¢N–140�55¢E 570 95 17.1 186 >400Nyuu 38�32¢N–139�59¢E 1,000 90 15.8 212 >600
aMeteorological data for Ryuzin-numa and Yuno-kawa are from the Sukayu meteorological station (a.s.l. 890 m) 7–8 km away and thosefor Nyuu are from the Hakkosawa meteorological station (a.s.l. 700 m) 3.5 km away. Temperature was adjusted for difference in altitudewith a lapse rate of 0.006�C m�1
476

(Meiwa Co. Ltd., Tokyo, Japan), a data logger (Li-1400,Li-Cor) and 12 V car batteries (38B19R, Nihon-Denki,Kyoto, Japan). Six sampling tubes were fixed 1 m abovethe ground at appropriate positions around the CO2
springs (see Fig. 1). CO2 concentrations were measuredevery second and logged as mean values every 30 s. Themonitoring was done for 2–4 days every month duringthe growing season (June–October). Hand-held CO2
monitors (Telaire 7001, TELAIRE, Goleta, Calif., USA,full-scale 10%) with data loggers (H08–007-02, Onset,Bourne, Mass., USA) were used for point-to-pointmeasurements and also for measurements at low CO2
areas. These CO2 analyzers were calibrated with a pureN2 and a CO2 standard gas (Tanuma-Sanso, Sendai,Japan) every month.
The concentrations of H2S and SO2 were measuredby H2S- and SO2 detecting tubes (no. 4LT and no. 5Lbfor H2S and SO2, respectively, Gastec, Ayase, Japan) atpoints where maximum concentrations were expected.
Relative light intensities around the CO2 springs weremeasured with light-sensitive films that fade and changetheir transmittance in proportion to the amount ofaccumulated light absorption (Opt leaf R-2D, Taisei-Kako, Tokyo, Japan) (Hikosaka et al. 2001). Films wereattached to plastic plates and placed horizontally at var-ious distances from the springs 1 m above the ground for3 days in July in 2002 (Ryuzin-numa) and in 2003 (Yuno-kawa and Nyuu). Films that were exposed to full lightintensity in open areas were used as a reference. The ex-tent of discoloration was measured at 560 nm with aspectrophotometer (UV-160A, Shimadzu, Kyoto, Japan).
At the three study sites, high CO2 (HC) and low CO2
(LC, close to ambient CO2) plots were established. Weselected LC plots by the following four criteria: (1) CO2
concentration was close to ambient at 370 lmol mol�1;(2) not far from the HC plot (to eliminate effects of anyunexpected environmental difference); (3) similar lightenvironment to HC plots; (4) some overlap of specieswith HC plot.2
To measure soil nutrients we sampled the surface soil(0–5 cm depth) of each plot in June 2003 (n=5). The soilwas sieved (2 mm wire mesh), and 10 g of the wet soilwas mixed with 50 ml of 2 M KCl. Nitrate and ammo-nium in the soil solution extract were determinedphotometrically with the Cataldo method and theindophenol blue method, respectively (Keeney andNelson 1982). The same soil solution was used for thedetermination of soil pH. The soil analysis was con-ducted again for Ryuzin-numa in August 2004 becauseof a possible error in the extraction procedure.
Physiological analysis
Three dominant species (see Table 3) at the shrub layerat each CO2 spring were used for analysis of leaf starch,nitrogen and carbon. Eight leaves were sampled fromabout 1 m above the ground surface (except for Plan-tago asiatica, 10 cm) at about noon from differentindividuals at HC and LC plots. The sampling was doneon 4 August 2002, 6 August 2003 and 28 August 2003for Ryuzin-numa, Yuno-kawa and Nyuu, respectively.
Ryuzin-numaYuno-kawaNyuu
(a)
Forest Herb & Shrub Reed Marsh Water Spring
10 m
LC
HC
HC
10 mLC
Track
1 2 345 6
10 m
HC
LCRoad
654321
Path
123456
(b) Ryuzin-numa
(c) Yuno-kawa (d) Nyuu
Fig. 1 Location of CO2 springsstudied (a) and vegetation atRyuzin-numa (b), Yuno-kawa(c) and Nyuu (d). HC (highCO2) and LC (low CO2,ambient CO2) plots wereestablished in the area withsimilar light climates. Numbers(1–6) in the figures indicate theplace at which CO2
concentrations were monitored
477

Leaves were brought to the laboratory in a cooledpolystyrene box (<5�C). Within 10 h of being sampled,eight disks of 1 cm diameter were punched out fromeach leaf. Four were weighed after being dried in anoven at 75�C for more than 3 days, and their totalnitrogen and carbon were determined with an NC ana-lyzer (NC-80, Shimadzu, Kyoto, Japan). The other fourleaf disks were frozen in liquid nitrogen and stored at�80�C until required for starch analysis.
The amount of starch was determined according toRufty and Huber (1983). Two disks from each leaf werehomogenized in liquid nitrogen and suspended in 1.5 mlof 80% ethanol. The suspension was heated at 80�C for30 min and centrifuged at 10,000 g for 10 min, and thesupernatant was removed. This extraction procedurewas repeated three times. The pellet was dried in an ovenfor 1 day, and then suspended in 1.0 ml of 0.2 M KOHand heated at 90�C for 30 min. After cooling, 0.2 ml of1.0 M acetic acid and 1.0 ml of amyloglucosidase(35 units ml�1 in 50 mM Na acetate buffer, pH 4.5,Rhizopus mold, Sigma, St Louis, USA) were added, andthe mixture was incubated at 55�C for 30 min. Thesuspension was centrifuged at 10,000 g for 10 min, andthe supernatant was used for determination of glucosewith the glucose B test Wako (Wako, Osaka, Japan).
CO2 response curves of photosynthesis were deter-mined on one dominant species from each spring(Polygonum sachalinense in Ryuzin-numa and Yuno-kawa, and Plantago asiatica in Nyuu) on 2–3 July, 4–6August and 26–27 August 2003 for Ryuzin-numa,Yuno-kawa and Nyuu, respectively (n=8 at each plot).At Ryuzin-numa and Yuno-kawa, photosynthesis wasmeasured on attached, fully expanded young leaves withan open gas-exchange system (LI-6400, Li-Cor). AtNyuu, fully expanded young leaves were cut at the pet-iole and used for the measurement in the laboratory.During measurements, leaf temperature was kept at20�C, and the vapor pressure deficit in the chamber at<1.0 kPa. Photosynthetic photon flux density (PPFD)was set at a saturating light intensity for plants at eachspring (1,200 lmol m�2 s�1, 800 lmol m�2 s�1,1,200 lmol m�2 s�1 for plants at Ryuzin-numa, Yuno-kawa and Nyuu, respectively).
We analyzed the CO2 response curve of photosyn-thesis in accordance with the model of Farquhar et al.(1980) to determine the maximum RuBP carboxylationrate (Vcmax) and the maximum electron transport ratedriving RuBP regeneration (Jmax). Michaelis–Mentenconstants of Rubisco (ribulose-1,5-bisphosphate car-boxylase/oxygenase) and the CO2 compensation point inthe absence of day respiration followed Bernacchi et al.(2001). Day respiration was assumed to be 0.02 of Vcmax
(von Caemmerer 2000).Water use efficiency (WUE) was defined as the pho-
tosynthetic rate at the mean growth CO2 concentration(Agrowth) divided by the transpiration rate. Similarly,photosynthetic N use efficiency (PNUE) was defined asAgrowth divided by leaf nitrogen concentration per unitarea (Narea).
Statistical tests were performed with STATVIEWv. 5.0 (SAS Institute, Cary, Ind., USA). The effects ofCO2 (HC and LC) and sites (Ryuzin-numa, Yuno-kawaand Nyuu) on leaf characteristics were evaluated byanalysis of variance (ANOVA). The t -test was used totest the difference between HC and LC.
In the present study the genus name is used when aplant name is abbreviated, because some species havethe same initial for their genus name, which would makea confusion of the species if the genus names wereabbreviated.
Results
Site characteristics
Ryuzin-numa, which is characterized by the presence ofa pond (Fig. 1b), was a semi-open space surrounded bya birch–oak forest with more than 20 species of shrubsand herbs. A gradient of CO2 concentrations was ob-served along a distance from the vent. Mean daytimeCO2 was about 680 lmol mol�1 (at 95 kPa) at thenearest monitoring point (Fig. 2) and 375 lmol mol�1
at 50 m from the vent. There was no clear seasonal trendin CO2 concentrations during the growth season,(Fig. 2). Diurnally, CO2 concentration was, at most,twice as high in the night as in the day. CO2 concen-tration was generally higher near the surface and grad-ually decreased with height, but it was still higher thanambient CO2 at 5 m above the ground (data not shown).The area with [CO2] above 500 lmol mol�1 wasapproximately 300 m2. Near the CO2 springs, Polygo-num sachalinense, Sasa kurilensis, Hydrangea paniculataand Calamagrostis matsumurae were abundant. Lyco-podium obscurum and Lycopodium clavatum (lycophytes)that are rare at the foot of Mount Hakkoda were alsoobserved.
Yuno-kawa is located in a gentle valley area withmature beech–oak forests. The CO2 concentration herewas relatively stable and stayed at around530 lmol mol�1 (at 95 kPa) (Fig. 2). Low wind speedsdue to the dense forest and depressed topography mighthave reduced CO2 fluctuations. The vertical gradient of[CO2] was fairly small from 0.1 m to 5 m above ground,and no diurnal change in CO2 concentration was ob-served (data not shown). CO2 concentration was highalong the stream and decreased gradually with distancefrom the springs but was still higher than the ambientCO2 300 m downstream from the spring. The approxi-mate area where mean [CO2] was above 500 lmol mol�1
was greater than 400 m2. About 20 shade-tolerant spe-cies grew around the springs, of which Tiarella po-lyphylla, Sanicula chinensism, Sasa and Polygonum wererelatively abundant.
Nyuu is located on a gentle hill-slope facing west andis covered by a species-rich beech-oak forest. CO2
emissions at this site were the largest among the threeCO2 springs, and a large area with high CO2 levels was
478
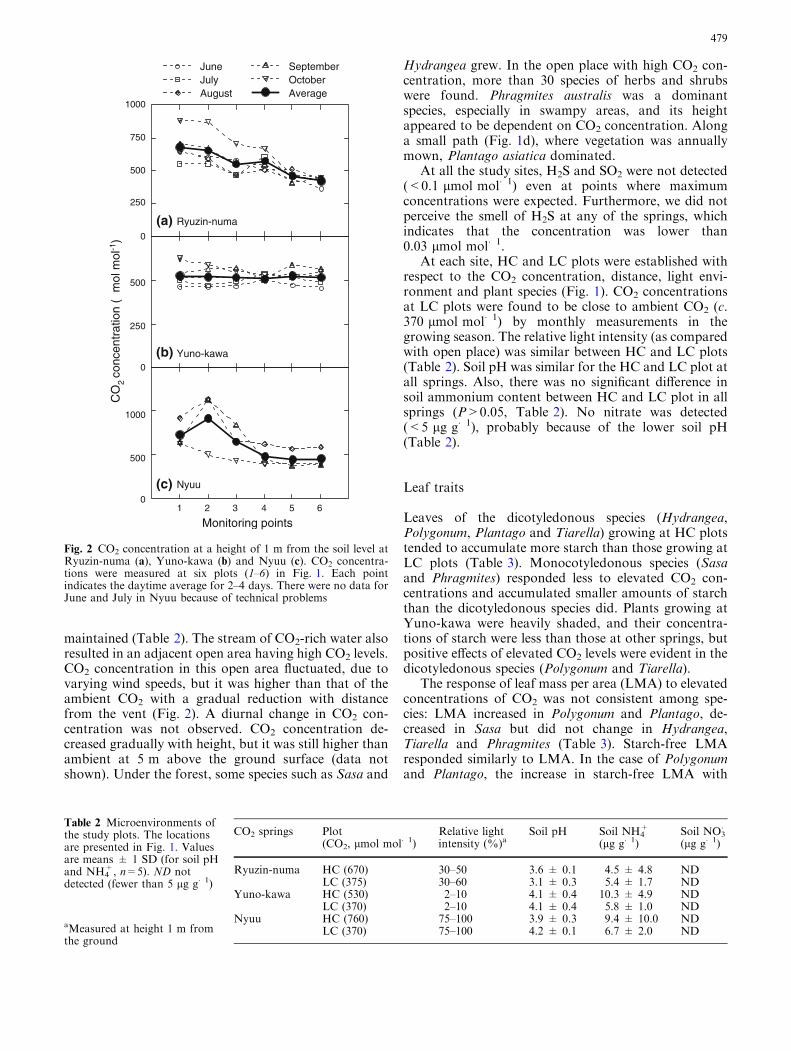
maintained (Table 2). The stream of CO2-rich water alsoresulted in an adjacent open area having high CO2 levels.CO2 concentration in this open area fluctuated, due tovarying wind speeds, but it was higher than that of theambient CO2 with a gradual reduction with distancefrom the vent (Fig. 2). A diurnal change in CO2 con-centration was not observed. CO2 concentration de-creased gradually with height, but it was still higher thanambient at 5 m above the ground surface (data notshown). Under the forest, some species such as Sasa and
Hydrangea grew. In the open place with high CO2 con-centration, more than 30 species of herbs and shrubswere found. Phragmites australis was a dominantspecies, especially in swampy areas, and its heightappeared to be dependent on CO2 concentration. Alonga small path (Fig. 1d), where vegetation was annuallymown, Plantago asiatica dominated.
At all the study sites, H2S and SO2 were not detected(<0.1 lmol mol�1) even at points where maximumconcentrations were expected. Furthermore, we did notperceive the smell of H2S at any of the springs, whichindicates that the concentration was lower than0.03 lmol mol�1.
At each site, HC and LC plots were established withrespect to the CO2 concentration, distance, light envi-ronment and plant species (Fig. 1). CO2 concentrationsat LC plots were found to be close to ambient CO2 (c.370 lmol mol�1) by monthly measurements in thegrowing season. The relative light intensity (as comparedwith open place) was similar between HC and LC plots(Table 2). Soil pH was similar for the HC and LC plot atall springs. Also, there was no significant difference insoil ammonium content between HC and LC plot in allsprings (P>0.05, Table 2). No nitrate was detected(<5 lg g�1), probably because of the lower soil pH(Table 2).
Leaf traits
Leaves of the dicotyledonous species (Hydrangea,Polygonum, Plantago and Tiarella) growing at HC plotstended to accumulate more starch than those growing atLC plots (Table 3). Monocotyledonous species (Sasaand Phragmites) responded less to elevated CO2 con-centrations and accumulated smaller amounts of starchthan the dicotyledonous species did. Plants growing atYuno-kawa were heavily shaded, and their concentra-tions of starch were less than those at other springs, butpositive effects of elevated CO2 levels were evident in thedicotyledonous species (Polygonum and Tiarella).
The response of leaf mass per area (LMA) to elevatedconcentrations of CO2 was not consistent among spe-cies: LMA increased in Polygonum and Plantago, de-creased in Sasa but did not change in Hydrangea,Tiarella and Phragmites (Table 3). Starch-free LMAresponded similarly to LMA. In the case of Polygonumand Plantago, the increase in starch-free LMA with
JuneJulyAugust
1 2 3 4 5 6
(a) Ryuzin-numa
(b)
(c)
Yuno-kawa0
250
500
Nyuu0
500
1000
0
250
500
750
1000
SeptemberOctoberAverage
Monitoring points
CO
2 co
ncen
trat
ion
(µm
ol m
ol-1
)
Fig. 2 CO2 concentration at a height of 1 m from the soil level atRyuzin-numa (a), Yuno-kawa (b) and Nyuu (c). CO2 concentra-tions were measured at six plots (1–6) in Fig. 1. Each pointindicates the daytime average for 2–4 days. There were no data forJune and July in Nyuu because of technical problems
Table 2 Microenvironments ofthe study plots. The locationsare presented in Fig. 1. Valuesare means ± 1 SD (for soil pHand NH4
+, n=5). ND notdetected (fewer than 5 lg g�1)
aMeasured at height 1 m fromthe ground
CO2 springs Plot(CO2, lmol mol�1)
Relative lightintensity (%)a
Soil pH Soil NH4+
(lg g�1)Soil NO3
�
(lg g�1)
Ryuzin-numa HC (670) 30–50 3.6 ± 0.1 4.5 ± 4.8 NDLC (375) 30–60 3.1 ± 0.3 5.4 ± 1.7 ND
Yuno-kawa HC (530) 2–10 4.1 ± 0.4 10.3 ± 4.9 NDLC (370) 2–10 4.1 ± 0.4 5.8 ± 1.0 ND
Nyuu HC (760) 75–100 3.9 ± 0.3 9.4 ± 10.0 NDLC (370) 75–100 4.2 ± 0.1 6.7 ± 2.0 ND
479

elevated CO2 was smaller than the increase in LMA dueto substantial starch accumulation (Table 3).
Mass-based leaf N concentration (Nmass) tended todecrease with elevated CO2 in plants at Ryuzin-numaand Nyuu (Table 3). Plants growing at Yuno-kawa wereheavily shaded and did not reduce leaf N concentration,and Tiarella had even higher Nmass at the HC plot. TheCO2 response of Nmass in the monocotyledonous specieswas small compared with that of the dicotyledonousspecies. Nitrogen concentration per unit structural mass(starch-free Nmass) was still lower in HC plants than inLC plants for most of the species showing low leaf Nconcentrations. This suggests that the reduction in leafN concentration with elevated CO2 level could be as-cribed not only to dilution by starch accumulationbut also to different allocation patterns of N in the plant.The response of leaf N concentration per unit area(Narea) to elevated CO2 levels varied among species,due to the various CO2 responses of LMA (Narea=Nmass·LMA). Differences in leaf carbon concentrationbetween the HC and LC were fairly small and were notsignificant in most species except for Polygonum inYuno-kawa, which increased by 2.4% under elevatedCO2 concentrations.
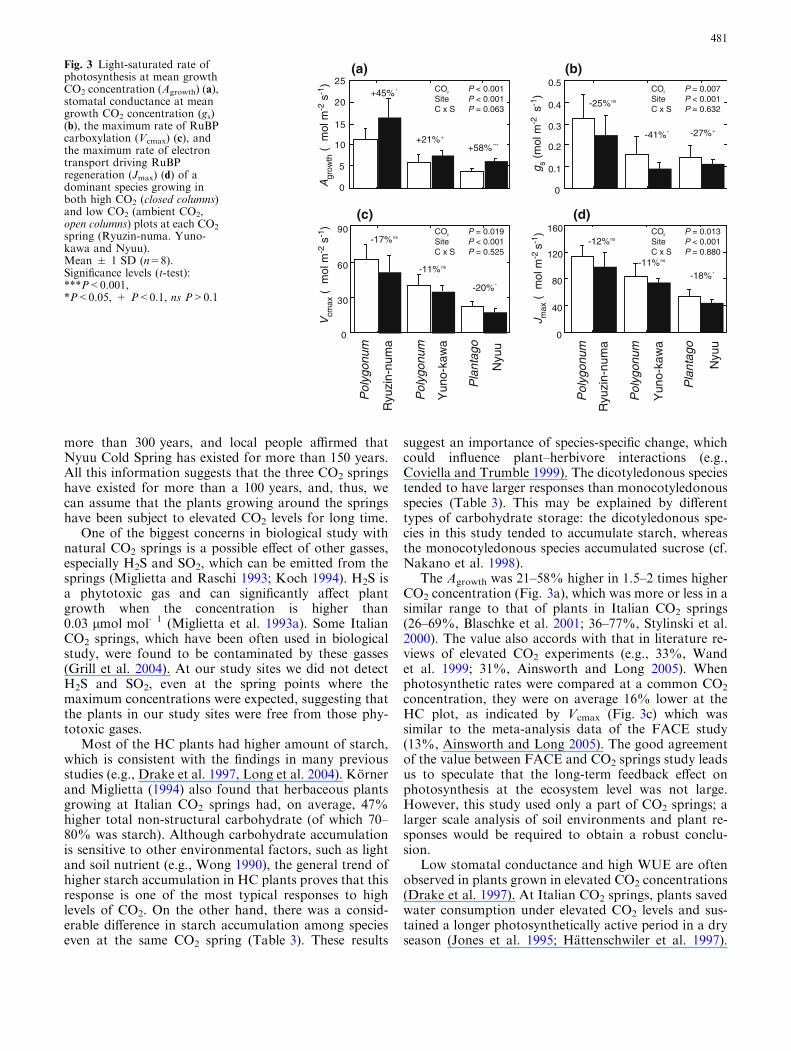
Photosynthetic rates at the mean growth CO2 con-centration (Agrowth; 670 lmol mol�1, 530 lmol mol�1
and 760 lmol mol�1 for plants at HC plots of Ryuzin-numa, Yuno-kawa and Nyuu, respectively, and370 lmol mol�1 for plants at all LC plots) were con-sistently higher in plants growing in HC than in LC plots(Fig. 3a). Stomatal conductance at the mean growthCO2 was generally lower in plants at HC plots (Fig. 3b).The maximum RuBP carboxylation rate (Vcmax) andmaximum electron transport rate (Jmax) were consis-tently lower in plants at HC plots, although the differ-ences were not always significant (Fig. 3c, d).
The higher Agrowth and lower stomatal conductancein plants growing at the HC plot led to significantlyhigher instantaneous WUE (Fig. 4a). Similarly, PNUEwas significantly higher in plants growing at HC plots(Fig. 4b). Vcmax/Narea, which indicates fraction of leafnitrogen in Rubisco, was not significantly different be-tween the two plots (Fig. 4c).
Discussion
The present study described three CO2 springs in Ja-pan. Although no scientific study on the history ofthese CO2 springs was available, examination of oldpublic documents suggested a long history of CO2
emissions at these springs. Both Ryuzin-numa andYuno-kawa were described in a document produced byAomori City (Anonymous 1953), in which these springswere presumed to have existed for more than a hundredyears. Information on Nyuu is rather limited, but theadjacent hot spring (500 m from Nyuu) has been re-garded as a sacred place by the Yudonosan Shrine forT
able
3Leafcharacteristics
ofplantsgrowingattheHCandLCplotsofthreenaturalCO
2springs.M
monocotyledonous,D
dicotyledonous,Starchconcentration,LMAleafmass
per
area,st-freeLMAstarch-freeLMA,N
massN
concentration,st-freeN
starch-freemass
base
Nconcentration,N
areaN
concentrationper
unitareaandcarbonconcentrationare
shown.
Values
are
means±
1SD
(n=
7–8).Significance
(t-test):nsnotsignificant,+
P<
0.1,*P<
0.05,**P<
0.01,***P<
0.001
Name
Species(m
onocotyledonous
ordicotyledonous)
Site
Starch(%
)LMA
(gm�2)
st-freeLMA
(gm�2)
Nmass(%
)st-freeN
(%)
Narea(g
m�2)
C(%
)
Ryuzin-numa
Hydrangea
paniculata
(D)
LC
3.50±
1.05
44.5
±9.4
42.9
±8.6
2.90±
0.31
3.01±
0.32
1.28±
0.24
48.4
±1.2
HC
6.18±
0.83*
43.8
±6.5
ns
41.2
±6.5
ns
2.20±
0.40**
2.35±
0.41**
0.95±
0.11**
47.4
±0.7
ns
Polygonum
sachalinense
(D)
LC
4.36±
1.03
35.3
±4.8
33.8
±4.6
3.80±
0.80
4.00±
0.80
1.31±
0.18
50.8
±1.5
HC
12.0
±4.17***
44.1
±7.1
*39.2
±2.1
+2.92±
1.03+
3.28±
1.08ns
1.23±
0.44ns
49.7
±1.2
ns
Sasa
kurilensis(M
)LC
1.07±
0.28
60.6
±3.5
60.0
±3.5
2.67±
0.26
2.70±
0.26
1.61±
0.14
45.8
±0.9
HC
2.02±
1.10*
54.8
±5.3
*53.7
±5.3
*2.42±
0.20+
2.47±
0.20+
1.33±
0.37**
46.5
±1.3
ns
Yuno-kawa
Polygonum
sachalinense
(D)
LC
0.97±
0.48
23.9
±2.3
23.6
±2.3
4.56±
0.33
4.61±
0.32
1.09±
0.10
49.2
±0.7
HC
1.97±
0.85*
27.6
±4.1
*27.0
±3.8
+4.53±
0.35ns
4.63±
0.35ns
1.24±
0.15*
51.6
±1.0
***
Sasa
kurilensis(M
)LC
0.43±
0.21
51.5
±4.7
51.2
±4.8
2.51±
0.24
2.52±
0.24
1.29±
0.11
47.9
±0.6
HC
0.39±
0.04ns
45.1
±2.5
**
45.0
±2.5
**
2.58±
0.24ns
2.58±
0.24ns
1.16±
0.10*
48.0
±0.6
ns
Tiarellapolyphylla(M
)LC
0.29±
0.20
20.5
±1.6
20.5
±1.6
3.15±
0.34
3.16±
0.34
0.64±
0.04
46.3
±1.0
HC
0.79±
0.35**
21.4
±4.4
ns
21.3
±4.3
ns
4.80±
0.53***
4.84±
0.52***
1.01±
0.12***
45.3
±1.4
ns
Nyuu
Hydrangea
paniculata
(D)
LC
4.64±
2.56
45.5
±10.1
43.6
±10.9
2.99±
0.60
3.14±
0.68
1.33±
0.26
47.8
±1.9
HC
6.19±
7.27ns
48.7
±8.9
ns
45.2
±5.7
ns
2.95±
0.50ns
3.14±
0.42ns
1.41±
0.24ns
47.5
±1.2
ns
Phragmites
australis(M
)LC
1.41±
0.40
55.0
±4.6
55.1
±4.3
3.13±
0.25
3.18±
0.26
1.75±
0.20
47.5
±0.5
HC
1.28±
0.20ns
55.0
±4.5
ns
54.3
±4.5
ns
2.87±
0.29ns
2.91±
0.30ns
1.57±
0.11+
47.8
±0.5
ns
Plantagoasiatica
(D)
LC
1.11±
0.48
26.1
±3.2
25.8
±3.2
2.78±
0.48
2.81±
0.48
0.72±
0.08
43.3
±1.4
HC
6.96±
3.54***
30.1
±5.4
+27.9
±4.0
ns
1.97±
0.38**
2.11±
0.35**
0.58±
0.05***
42.1
±0.8
+
480
more than 300 years, and local people affirmed thatNyuu Cold Spring has existed for more than 150 years.All this information suggests that the three CO2 springshave existed for more than a 100 years, and, thus, wecan assume that the plants growing around the springshave been subject to elevated CO2 levels for long time.
One of the biggest concerns in biological study withnatural CO2 springs is a possible effect of other gasses,especially H2S and SO2, which can be emitted from thesprings (Miglietta and Raschi 1993; Koch 1994). H2S isa phytotoxic gas and can significantly affect plantgrowth when the concentration is higher than0.03 lmol mol�1 (Miglietta et al. 1993a). Some ItalianCO2 springs, which have been often used in biologicalstudy, were found to be contaminated by these gasses(Grill et al. 2004). At our study sites we did not detectH2S and SO2, even at the spring points where themaximum concentrations were expected, suggesting thatthe plants in our study sites were free from those phy-totoxic gases.
Most of the HC plants had higher amount of starch,which is consistent with the findings in many previousstudies (e.g., Drake et al. 1997, Long et al. 2004). Kornerand Miglietta (1994) also found that herbaceous plantsgrowing at Italian CO2 springs had, on average, 47%higher total non-structural carbohydrate (of which 70–80% was starch). Although carbohydrate accumulationis sensitive to other environmental factors, such as lightand soil nutrient (e.g., Wong 1990), the general trend ofhigher starch accumulation in HC plants proves that thisresponse is one of the most typical responses to highlevels of CO2. On the other hand, there was a consid-erable difference in starch accumulation among specieseven at the same CO2 spring (Table 3). These results
suggest an importance of species-specific change, whichcould influence plant–herbivore interactions (e.g.,Coviella and Trumble 1999). The dicotyledonous speciestended to have larger responses than monocotyledonousspecies (Table 3). This may be explained by differenttypes of carbohydrate storage: the dicotyledonous spe-cies in this study tended to accumulate starch, whereasthe monocotyledonous species accumulated sucrose (cf.Nakano et al. 1998).
The Agrowth was 21–58% higher in 1.5–2 times higherCO2 concentration (Fig. 3a), which was more or less in asimilar range to that of plants in Italian CO2 springs(26–69%, Blaschke et al. 2001; 36–77%, Stylinski et al.2000). The value also accords with that in literature re-views of elevated CO2 experiments (e.g., 33%, Wandet al. 1999; 31%, Ainsworth and Long 2005). Whenphotosynthetic rates were compared at a common CO2
concentration, they were on average 16% lower at theHC plot, as indicated by Vcmax (Fig. 3c) which wassimilar to the meta-analysis data of the FACE study(13%, Ainsworth and Long 2005). The good agreementof the value between FACE and CO2 springs study leadsus to speculate that the long-term feedback effect onphotosynthesis at the ecosystem level was not large.However, this study used only a part of CO2 springs; alarger scale analysis of soil environments and plant re-sponses would be required to obtain a robust conclu-sion.
Low stomatal conductance and high WUE are oftenobserved in plants grown in elevated CO2 concentrations(Drake et al. 1997). At Italian CO2 springs, plants savedwater consumption under elevated CO2 levels and sus-tained a longer photosynthetically active period in a dryseason (Jones et al. 1995; Hattenschwiler et al. 1997).
Pol
ygon
um
Yun
o-ka
wa
Pol
ygon
um
Ryu
zin-
num
a
Pla
ntag
o
Nyu
u
Pol
ygon
um
Yun
o-ka
wa
Pol
ygon
um
Ryu
zin-
num
a
(c)
30
60
90
(d)
00
40
80
120
160
(a)
0
5
10
15
20
25(b)
0
0.1
0.2
0.3
0.4
0.5+45% *
+21% +
+58% ***
-25%ns
-41%* -27%+
-17%ns
-11%ns
-20%*
-12%ns
-11%ns
-18%*
CO2 P < 0.001Site P < 0.001C x S P = 0.063
CO2 P = 0.007Site P < 0.001C x S P = 0.632
CO2 P = 0.013Site P < 0.001C x S P = 0.880
CO2 P = 0.019Site P < 0.001C x S P = 0.525
J max
(µm
ol m
-2 s
-1)
g s (m
ol m
-2 s
-1)
Agr
owth
(µm
ol m
-2 s
-1)
Vcm
ax (
µmol
m-2
s-1
)
Pla
ntag
o
Nyu
u
Fig. 3 Light-saturated rate ofphotosynthesis at mean growthCO2 concentration (Agrowth) (a),stomatal conductance at meangrowth CO2 concentration (gs)(b), the maximum rate of RuBPcarboxylation (Vcmax) (c), andthe maximum rate of electrontransport driving RuBPregeneration (Jmax) (d) of adominant species growing inboth high CO2 (closed columns)and low CO2 (ambient CO2,open columns) plots at each CO2
spring (Ryuzin-numa. Yuno-kawa and Nyuu).Mean ± 1 SD (n=8).Significance levels (t-test):***P<0.001,*P<0.05, + P<0.1, ns P>0.1
481

Reduction in stomatal conductance with elevated CO2
concentration was also observed in humid climate re-gions like Japan (Fig. 3b). This may be explained by themechanism of the plant to keep a constant ratio ofintercellular CO2 to air CO2 concentration (Wong et al.1979; Drake et al. 1997). Furthermore, as stomatalconductance is associated with leaf temperature throughheat dissipation (Kimball et al. 1995), the lower con-ductance may ameliorate the low temperature stress inplants growing in cool climates.
Conclusions
We described three CO2 springs, where wild plants havebeen exposed to CO2 enrichment for hundreds of yearswithout the effects of sulfur gases. The CO2 responsesin leaf traits observed in the present study were quali-
tatively not different from those in previous experi-mental studies. However, there was a large differencesin CO2 response in starch accumulation among species,even within the same CO2 spring, suggesting that theresponse was quantitatively species-specific. Althoughdown-regulation of photosynthesis was observed athigh CO2 plots, the photosynthetic rate at growth CO2
concentration was stimulated as a result of the directeffect of high CO2 levels. Similar results between FACEand CO2 spring studies suggest that the feedback effecton photosynthesis at an ecosystem level would not belarge.
Acknowledgments The landowners of CO2 springs (Hakkoda-On-sen Co., Tashiro Bokuya-Chikusan Kumiai and Yudonosanshrine) kindly gave us a permission to use their land for this study.We thank Makoto Tsurumi (Hirosaki Univ.) and Atsuki Uchida(Nippon Geophysical Prospecting Co.) for geological informationof Ryuzin-numa and Yuno-kawa, and Taku Noro (Ministry ofLand, Infrastructure and Transport) for climate data at Hakkos-awa meteorological station. We also thank Mark Lieffering (Ag-Research, NZ) for his comments on the draft manuscript, KoujiYonekura (Tohoku Univ.) for sorting plant species, and YukoYasumura, Hisashi Tsujisawa and Riichi Oguchi (Tohoku Univ.)for their technical assistance. Noriyuki Osada (Tohoku Univ.)kindly provided his data of soil nutrients and also gave us con-structive comments on the draft manuscript. This study was partlysupported by grants-in-aid from JSPS for Young Research Fellows(Y.O.), and from the Japan Ministry of Education, Culture, Sports,Science and Technology to K.H. and T.H.
References
Ainsworth EA, Long SP (2005) What have we learned from 15years of free-air CO2 enrichment (FACE)? A meta-analyticreview of the responses of photosynthesis, canopy propertiesand plant production to rising CO2. New Phytol 165:351–372
Ainsworth EA, Davey PA, Hymus GJ, Osborne CE, Rogers A,Blum H, Nosberger J, Long SP (2003) Is stimulation of leafphotosynthesis by elevated carbon dioxide concentrationmaintained in the long term? A test with Lolium perenne grownfor ten years at two nitrogen fertilization levels under free airCO2 enrichment (FACE). Plant Cell Environ 26:705–714
Anonymous (1953) A plan to change land utilization in Ara-kawaand Komagome-gawa areas. Official document of Aomori City,Aomori (in Japanese)
Arp WJ (1991) Effects of source-sink relations on photosyntheticacclimation to elevated carbon dioxide. Plant Cell Environ14:869–876
Bernacchi CJ, Singsaas EL, Pimentel C, Portis AR Jr., Long SP(2001) Improved temperature response functions for models ofRubisco-limited photosynthesis. Plant Cell Environ 24:253–259
Blaschke L, Schulte M, Raschi A, Slee N, Rennenberg H, Polle A(2001) Photosynthesis, soluble and structural carbon com-pounds in two Mediterranean oak species (Quercus pubescensand Q. ilex) after lifetime growth at naturally elevated CO2
concentrations. Plant Biol 3:288–297von Caemmerer S (2000) Biochemical models of leaf photosyn-
thesis. CSIRO publishing, CanberraChaves MM, Pereira JS, Cerasoli S, Clifton-Brown J, Miglietta F,
Raschi A (1995) Leaf metabolism during summer drought inQuercus ilex trees with lifetime exposure to elevated CO2. JBiogeogr 22:255–259
Cook AC, Tissue DT, Roberts SW, Oechel WC (1998) Effects oflong-term elevated [CO2] from natural CO2 springs on Nardusstricta: photosynthesis, biochemistry, growth and phenology.Plant Cell Environ 21:417–425
0
5
10
15
20 (b)
0
20
40
60(c)
(a)
0
5
10
15 CO2 P < 0.001Site P < 0.001C x S P = 0.262
CO2 P < 0.001Site P < 0.001C x S P = 0.024
CO2 P = 0.570Site P < 0.001C x S P = 0.440
+93%***
+90%***
+105%**
+59%***
+40%***
+96%***
-8%ns
+2%ns +1%ns
WU
E (
mm
ol m
ol-1
)P
NU
E (
µmol
gN
-1 s
-1)
Vcm
ax/N
area
(µm
ol g
N-1
s-1
)
Pol
ygon
um
Ryu
zin-
num
a
Pol
ygon
um
Yun
o-ka
wa
Pla
ntag
o
Nyu
u
Fig. 4 Water use efficiency (WUE, photosynthetic rate divided bytranspiration rate) (a), photosynthetic N use efficiency (PNUE,photosynthetic rate divided by leaf N) (b) and Vcmax divided by leafN (c) of a dominant species growing in both high CO2 (closedcolumns) and low CO2 (ambient CO2, open columns) plots at eachCO2 spring (Ryuzin-numa, Yuno-kawa and Nyuu). Mean ± 1 SD(n=8). Significance levels (t-test): ***P<0.001, ns P>0.05
482

Cook AC, Vourlitis GL, Harazono Y (2000) Evaluating the po-tential for long-term elevated CO2 exposure studies using CO2
springs in Japan. J Agric Meteorol 56:31–40Cotrufo MF, Raschi A, Lanini M, Ineson P (1999) Decomposition
and nutrient dynamics of Quercus pubescens leaf litter in anaturally enriched CO2 Mediterranean ecosystem. Func Ecol13:343–351
Coviella CE, Trumble JT (1999) Effects of elevated atmosphericcarbon dioxide on insect–plant Interactions. Conserv Biol13:700–712
Drake BG, Gonzalez-Meler MA, Long SP (1997) More efficientplants—a consequence of rising atmospheric CO2. Ann RevPlant Physiol Plant Mol Biol 48:609–639
Farquhar GD, von Caemmerer S, Berry JA (1980) A biochemicalmodel of photosynthetic CO2 assimilation in leaves of C3 spe-cies. Planta 149:78–90
FernandezMD, Pieters A, Donoso C, Tezara W, AzkueM, HerreraC, Rengifo E,HerreraA (1998) Effects of a natural source of veryhigh CO2 concentration on the leaf gas exchange, xylem waterpotential and stomatal characteristics of plants of Spatiphylumcannifolium and Bauhinia multinervia. New Phytol 138:689–697
Gahrooee FR (1998) Impacts of elevated atmospheric CO2 on litterquality, litter decomposability and nitrogen turnover rate oftwo oak species in a Mediterranean forest ecosystem. GlobalChange Biol 4:667–677
Grill D, Muller M, Tausz M, Strnad B, Wonisch A, Raschi A(2004) Effects of sulphurous gases in two CO2 springs on totalsulphur and thiols in acorns and oak seedlings. Atmos Environ38:3775–3780
Hattenschwiler S, Miglietta F, Raschi A, Korner C (1997) Thirtyyears of in situ tree growth under elevated CO2—a model forfuture forest responses. Global Change Biol 3:463–471
Hikosaka K (2003) A model of dynamics of leaves and nitrogen ina plant canopy: an integration of canopy photosynthesis, leaflife span, and nitrogen use efficiency. Am Nat 162:149–164
Hikosaka K, Nagashima H, Harada Y, Hirose T (2001) A simpleformulation of interaction between individuals competing forlight in a monospecific stand. Func Ecol 15:642–646
Idso KE, Idso SB (1994) Plant responses to atmospheric CO2
enrichment in the face of environmental constraints: a review ofthe past 10 years’ research. Agric Forest Meteorol 69:153–203
Jones MB, Clifton Brown J, Raschi A, Miglietta F (1995) Theeffects on Arbutus unedo L. of long-term exposure to elevatedCO2. Global Change Biol 1:295–302
Keeney DR, Nelson DW (1982) Nitrogen—Inorganic Forms. In:Page AL (Ed) Methods of soil analysis, part 2. AmericanSociety of Agronomy, Madison, pp 643–698
Kimball BA, Pinter PJ, Garcia RL, Lamorte RL, Wall GW,Hunsaker DJ, Wechsung G, Wechsung F, Kartschall T (1995)Productivity and water use of wheat under free-air CO2
enrichment. Global Change Biol 1:429–442Kimbara K (1992) Distribution map and catalogue of hot and
mineral springs in Japan. Geological Survey of Japan, TsukubaKoch GW (1994) The use of natural situation of CO2 enrichment in
studies of vegetation response to increasing atmospheric CO2.In: Schulze ED, Mooney HA (Eds) Design and execution ofexperiments on CO2 enrichment. Commission of the EuropeanCommunities, Luxembourg, pp 381–392
Kohut R (2003) The long-term effects of carbon dioxide on naturalsystems: issues and research needs. Environ Int 29:171–180
Korner C, Miglietta F (1994) Long term effects of naturally ele-vated CO2 on Mediterranean grassland and forest trees. Oec-ologia 99:343–351
Long SP (1991) Modification of the response of photosyntheticproductivity to rising temperature by atmospheric carbondioxide concentrations: has its importance been underesti-mated? Plant Cell Environ 14:729–740
Long SP, Ainsworth EA, Rogers A, Ort DR (2004) Rising atmo-spheric carbon dioxide: plants FACE the future. Annu RevPlant Biol 55:591–628
Luo Y, Reynolds J (1999) Validity of extrapolating field CO2
experiments to predict carbon sequestration in natural ecosys-tems. Ecology 80:1568–1583
Miglietta F, Raschi M (1993) Studying the effect of elevated CO2 inthe open in a naturally enriched environment in Central Italy.Vegetation 104/105:391–400
Miglietta F, Raschi A, Bettarini I, Resti R, Selvi F (1993a) NaturalCO2 springs in Italy: a resource for examining long-term re-sponse of vegetation to rising atmospheric CO2 concentrations.Plant Cell Environ 16:873–878
Miglietta F, Raschi A, Resti R, Resti R, Selvi F (1993b) Growthand onto-morphogenesis of soybean (Glycine max Merril) in anopen, naturally CO2-enriched environment. Plant Cell Environ16:909–918
Miglietta F, Raschi A, Bettarini I, Baldiani M, van Gardingen P(1994) Carbon dioxide springs and their use for experimenta-tion. In: Schulze ED, Mooney HA (Eds) Design and executionof experiments on CO2 enrichment. Commission of the Euro-pean communities, Luxembourg, pp 393–403
Mott KA (1990) Sensing of atmospheric CO2 by plants. Plant CellEnviron 13:731–737
Nakano H, Makino A, Mae T (1998) The responses of Rubiscoprotein to long-term exposure to elevated CO2 in rice and beanleaves. Photosynth Mech Eff 5:3391–3394
Newton PCD, Bell CC, Clark H (1996) Carbon dioxide emissionsfrom mineral springs in Northland and the potential of thesesites for studying the effects of elevated carbon dioxide onpastures. New Zeal J Agric Res 39:33–40
Poorter H (1993) Interspecific variation in the growth response ofplants to an elevated ambient CO2 concentration. Vegetation104/105:77–97
Rapparini F, Baraldi R, Miglietta F, Loreto F (2004) Isoprenoidemission in trees of Quercus pubescens and Quercus ilex withlifetime exposure to naturally high CO2 environment. Plant CellEnviron 27:381–391
Rufty TW, Huber SC (1983) Changes in starch formation andactivities of sucrose phosphate synthesis and cytoplasmic fruc-tose-1–6-bisphosphatase in response to source-sink alterations.Plant Physiol 72:474–480
Scholefield PA, Doick KJ, Herbert BMJ, Hewitt CNS, SchnitzlerJP, Pinelli P, Loreto F (2004) Impact of rising CO2 on emissionsof volatile organic compounds: isoprene emission from Phrag-mites australis growing at elevated CO2 in a natural carbondioxide spring. Plant Cell Environ 27:393–401
Sheen J. (1990) Metabolic repression of transcription in higherplants. Plant Cell 2:1027–1038
Stitt M, Krapp A (1999) The interaction between elevated carbondioxide and nitrogen nutrition: the physiological and molecularbackground. Plant Cell Environ 22:583–621
Stylinski CD, Oechel WC, Gamon JA, Tissue DT, Miglietta F,Raschi A (2000) Effects of lifelong [CO2] enrichment on car-boxylation and light utilization of Quercus pubescens Willd.examined with gas exchange, biochemistry and optical tech-niques. Plant Cell Environ 23:1353–1362
Tognetti R, Giovannelli A, Longobucco A, Miglietta F, Raschi A.(1996) Water relations of oak species growing in the naturalCO2 spring of Rapolano (central Italy). Ann Des Sci Forest53:475–485
Tognetti R, Longobucco A, Miglietta F, Raschi A (1998) Tran-spiration and stomatal behaviour of Quercus ilex plants duringthe summer in a Mediterranean carbon dioxide spring. PlantCell Environ 21:613–622
Tognetti R, Longobucco A, Miglietta F, Raschi A (1999) Waterrelations, stomatal response and transpiration of Quercus pu-bescens trees during summer in a Mediterranean carbon dioxidespring. Tree Physiol 19:261–270
Tognetti R, Cherubini P, Innes JL (2000) Comparative stem-growth rates of Mediterranean trees under background andnaturally enhanced ambient CO2 concentrations. New Phytol146:59–74
483

Tsurumi M, Hirabayashi J, Yoshimura K, Kondo H, Sasaki M,Ishida S (1999) Report on the volcanic gas in the HakkodaMountain. Official document of Aomori Prefecture, Aomori (inJapanese)
Vodnik D, Pfanz H, Wittmann C, Macek I, Kastelec D, Turk B,Batic F (2002) Photosynthetic acclimation in plants growingnear a carbon dioxide spring. Phyton 42:239–244
Wand SJE, Midgley GF, Jones MH, Curtis PS (1999) Response ofwild C4 and C3 grass (Poaceae) species to elevated atmospheric
CO2 concentration: a meta-analytic test of current theories andperceptions. Global Change Biol 5:723–741
Wong SC (1990) Elevated atmospheric partial pressure of carbondioxide and plant growth ii. non-structural carbohydrate con-tent in cotton plants and its effect on growth parameters.Photosynth Res 23:171–180
Wong SC, Cowan IR, Farquhar GD (1979) Stomatal conductancecorrelates with photosynthetic capacity. Nature 282:424–426
484




















