
Effect of A2 gene on infectivity of the nonpathogenic parasite Leishmania tarentolae
10
Vaccine 28 (2010) 53–62 Contents lists available at ScienceDirect Vaccine journal homepage: www.elsevier.com/locate/vaccine Recombinant Leishmania tarentolae expressing the A2 virulence gene as a novel candidate vaccine against visceral leishmaniasis Amir Mizbani a,b , Tahereh Taheri a , Farnaz Zahedifard a , Yasaman Taslimi a , Hiva Azizi a , Kayhan Azadmanesh c , Barbara Papadopoulou d , Sima Rafati a,∗ a Molecular Immunology and Vaccine Research Laboratory, Pasteur Institute of Iran, Tehran, Iran b Department of Biotechnology, University College of Science, University of Tehran, Tehran, Iran c Hepatitis and AIDS Department, Pasteur Institute of Iran, Tehran, Iran d Research Centre in Infectious Diseases, CHUL Research Centre and Department of Microbiology and Immunology, Faculty of Medicine, Laval University, 2705 Laurier Blvd., Quebec (QC), Canada G1V 4G2 article info Article history: Received 22 August 2009 Received in revised form 21 September 2009 Accepted 25 September 2009 Available online 8 October 2009 Keywords: Visceral leishmaniasis Recombinant Leishmania tarentolae Live vaccine A2 gene Immune response abstract Visceral leishmaniasis is the most severe form of leishmaniasis. To date, there is no effective vaccine against this disease. Many antigens have been examined so far as protein- or DNA-based vaccines, but none of them conferred complete long-term protection. The use of live attenuated vaccines has recently emerged as a promising vaccination strategy. In this study, we stably expressed the Leishmania donovani A2 antigen in Leishmania tarentolae, a non-pathogenic member of the genus Leishmania, and evaluated its protective efficacy as a live vaccine against L. infantum challenge. Our results show that a single intraperi- toneal administration of the A2-recombinant L. tarentolae strain protects BALB/c mice against L. infantum challenge and that protective immunity is associated with high levels of IFN- production prior and after challenge. This is accompanied by reduced levels of IL-5 production after challenge, leading to a potent Th1 immune response. In contrast, intravenous injection elicited a Th2 type response, character- ized by higher levels of IL-5 and high humoral immune response, resulting in a less efficient protection. All together, these results indicate the promise of A2-expressing L. tarentolae as a safe live vaccine against visceral leishmaniasis. © 2009 Elsevier Ltd. All rights reserved. 1. Introduction Protozoan parasites of the genus Leishmania are causative agents of a broad spectrum of diseases, collectively known as leishmania- sis. The clinical symptoms of the disease range from asymptomatic self-healing cutaneous lesions (CL, caused by Leishmania major, L. tropica, and L. mexicana species) to mucocutaneous leishmaniasis (caused by L. braziliensis) and severe visceral (VL) infections (caused by L. infantum, L. donovani, and L. chagasi) [1,2]. Currently, over 12 million people in 88 countries are infected with this parasite, 350 million are at the risk of infection worldwide and 1.5–2 million new cases are reported every year [3–7]. VL is the most severe sys- temic disease among the three main categories of leishmaniasis and affects 500,000 people every year [6]. Moreover, VL has emerged as an opportunistic infection in HIV-1 infected patients in many parts of the world [8–10]. Currently, there is no effective vaccine against leishmaniasis and control of the disease is almost confined to chemotherapy. There are ∗ Corresponding author. Tel.: +98 21 66953311x2112; fax: +98 21 88742314. E-mail address: s [email protected] (S. Rafati). only a limited number of drugs available and each has its own dis- advantages. Conventional drugs require long-term administration periods and often induce serious side-effects due to their toxicity [11,12]. In addition increasing incidence of drug-resistant strains has hampered the control of the disease by chemotherapy [13–17]. Therefore, much attention has been given to developing effective vaccines. Although induction of lifelong protection against reinfection in recovered people demonstrates that a protective vaccine can be achieved, an effective vaccine against human leishmaniasis has yet to be discovered [18]. However, several vaccination strategies have been tried against experimental leishmaniasis, with partic- ular emphasis on their efficacy against CL rather than VL [19,20]. First-generation anti-leishmanial vaccines, composed of killed par- asites, were examined as an alternative but none has provided full protection thus far [18]. In addition, several antigens such as GP63 [21,22], GP46 [23], p36/LACK [24,25], CPB and CPA [26], LD1 [27], PSA-2 [28], TSA/LmSTI1 [29,30], PFR2 [31], HASPB1 [32], LeIF [33], LCR1 [34], and M2 [35] have been tested as recombinant proteins or DNA vaccines, but they have given only partial protection, and/or need some clinically unacceptable adjuvants to induce proper Th1 response in humans [36–38]. Also, it seems unlikely that a single 0264-410X/$ – see front matter © 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.vaccine.2009.09.114
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Effect of A2 gene on infectivity of the nonpathogenic parasite Leishmania tarentolae

Rc
AKa
b
c
d
L
a
ARR2AA
KVRLAI
1
osst(bmmntaao
c
0d
Vaccine 28 (2010) 53–62
Contents lists available at ScienceDirect
Vaccine
journa l homepage: www.e lsev ier .com/ locate /vacc ine
ecombinant Leishmania tarentolae expressing the A2 virulence gene as a novelandidate vaccine against visceral leishmaniasis
mir Mizbania,b, Tahereh Taheria, Farnaz Zahedifarda, Yasaman Taslimia, Hiva Azizi a,ayhan Azadmaneshc, Barbara Papadopouloud, Sima Rafati a,∗
Molecular Immunology and Vaccine Research Laboratory, Pasteur Institute of Iran, Tehran, IranDepartment of Biotechnology, University College of Science, University of Tehran, Tehran, IranHepatitis and AIDS Department, Pasteur Institute of Iran, Tehran, IranResearch Centre in Infectious Diseases, CHUL Research Centre and Department of Microbiology and Immunology, Faculty of Medicine,aval University, 2705 Laurier Blvd., Quebec (QC), Canada G1V 4G2
r t i c l e i n f o
rticle history:eceived 22 August 2009eceived in revised form1 September 2009ccepted 25 September 2009vailable online 8 October 2009
a b s t r a c t
Visceral leishmaniasis is the most severe form of leishmaniasis. To date, there is no effective vaccineagainst this disease. Many antigens have been examined so far as protein- or DNA-based vaccines, butnone of them conferred complete long-term protection. The use of live attenuated vaccines has recentlyemerged as a promising vaccination strategy. In this study, we stably expressed the Leishmania donovaniA2 antigen in Leishmania tarentolae, a non-pathogenic member of the genus Leishmania, and evaluated itsprotective efficacy as a live vaccine against L. infantum challenge. Our results show that a single intraperi-
eywords:isceral leishmaniasisecombinant Leishmania tarentolaeive vaccine2 gene
mmune response
toneal administration of the A2-recombinant L. tarentolae strain protects BALB/c mice against L. infantumchallenge and that protective immunity is associated with high levels of IFN-� production prior andafter challenge. This is accompanied by reduced levels of IL-5 production after challenge, leading to apotent Th1 immune response. In contrast, intravenous injection elicited a Th2 type response, character-ized by higher levels of IL-5 and high humoral immune response, resulting in a less efficient protection.All together, these results indicate the promise of A2-expressing L. tarentolae as a safe live vaccine against
visceral leishmaniasis.. Introduction
Protozoan parasites of the genus Leishmania are causative agentsf a broad spectrum of diseases, collectively known as leishmania-is. The clinical symptoms of the disease range from asymptomaticelf-healing cutaneous lesions (CL, caused by Leishmania major, L.ropica, and L. mexicana species) to mucocutaneous leishmaniasiscaused by L. braziliensis) and severe visceral (VL) infections (causedy L. infantum, L. donovani, and L. chagasi) [1,2]. Currently, over 12illion people in 88 countries are infected with this parasite, 350illion are at the risk of infection worldwide and 1.5–2 million
ew cases are reported every year [3–7]. VL is the most severe sys-emic disease among the three main categories of leishmaniasis andffects 500,000 people every year [6]. Moreover, VL has emerged as
n opportunistic infection in HIV-1 infected patients in many partsf the world [8–10].Currently, there is no effective vaccine against leishmaniasis andontrol of the disease is almost confined to chemotherapy. There are
∗ Corresponding author. Tel.: +98 21 66953311x2112; fax: +98 21 88742314.E-mail address: s [email protected] (S. Rafati).
264-410X/$ – see front matter © 2009 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2009.09.114
© 2009 Elsevier Ltd. All rights reserved.
only a limited number of drugs available and each has its own dis-advantages. Conventional drugs require long-term administrationperiods and often induce serious side-effects due to their toxicity[11,12]. In addition increasing incidence of drug-resistant strainshas hampered the control of the disease by chemotherapy [13–17].Therefore, much attention has been given to developing effectivevaccines.
Although induction of lifelong protection against reinfection inrecovered people demonstrates that a protective vaccine can beachieved, an effective vaccine against human leishmaniasis hasyet to be discovered [18]. However, several vaccination strategieshave been tried against experimental leishmaniasis, with partic-ular emphasis on their efficacy against CL rather than VL [19,20].First-generation anti-leishmanial vaccines, composed of killed par-asites, were examined as an alternative but none has provided fullprotection thus far [18]. In addition, several antigens such as GP63[21,22], GP46 [23], p36/LACK [24,25], CPB and CPA [26], LD1 [27],
PSA-2 [28], TSA/LmSTI1 [29,30], PFR2 [31], HASPB1 [32], LeIF [33],LCR1 [34], and M2 [35] have been tested as recombinant proteins orDNA vaccines, but they have given only partial protection, and/orneed some clinically unacceptable adjuvants to induce proper Th1response in humans [36–38]. Also, it seems unlikely that a single
5 accine
apintvpttat
oneogbfdpLrpt
acehvcTtHn[v
fioridtiabv
iatifi
2
2
pl
4 A. Mizbani et al. / V
ntigen can elicit a fully protective immunity against a complexarasite like Leishmania [36]. On the other hand, there is increas-
ng evidence that presence of a small number of live parasites isecessary to maintain durable immunity [39,40], and the only wayo meet this requirement is by using attenuated live vaccines. Liveaccines can simulate the natural course of infection more closely,resent a rich repertoire of antigens to the host immune system inhe way that the wild type parasite does and persist for a longerime, leading to a more potent and durable immunity. Using livevirulent organisms as a vaccine has proven to be more efficacioushan traditional subunit vaccines [41].
Attenuated strains using either long-term in vitro cultures [42]r selection for temperature sensitivity [43] or chemical mutage-esis [44], and parasite culture under drug pressure [45] are notasily applicable to human use, because there is always a riskf reversion of the organism to its virulent state. The advent ofene transfection technologies for Leishmania has made it possi-le to obtain attenuated organisms by targeting genes importantor virulence or intracellular survival of the parasite, such as dihy-rofolate reductase thymidylate synthase (DHFR-TS) [46], cysteineroteinase genes [47], HSP100 heat shock protein [48], SIR2 [49],PG2 [50], biopterin transporter BT1 [51], and the trypanothioneeductase [52]. Using intrinsically non-pathogenic species is also aromising approach, exemplified by BCG vaccine against Mycobac-erium tuberculosis infection.
Among different species of Leishmania, Leishmania tarentolae islizard parasite which is non-pathogenic to humans. The parasite
an differentiate into amastigotes, but is not able to persist longnough within mammalian macrophages [53,54]. Recent studiesave shown that it can be used as a live vaccine against L. dono-ani due to its capability to efficiently target antigen-presentingells, induce maturation of dendritic cells, and elicit a protectiveh1 immune response [53]. Interestingly, a recombinant L. taren-olae expressing HIV-1 Gag protein has been used as a candidateIV-1 vaccine and shown to induce strong cell-mediated immu-ity in mice and to decrease HIV-1 replication in an ex-vivo system55], suggesting that this species can be used as a promising live-ectored vaccine against intracellular pathogens.
We have recently shown that among some important virulenceactors, A2 is not present in L. tarentolae [56]. A2 was first identifiedn L. donovani as a gene family that is expressed only in the amastig-te stage [57]. A2 proteins are comprised mainly of a 10-amino acidepeated sequence and range in size from 45 to 110 kDa, depend-ng on the number of repeats [58]. A2-specific antibodies have beenetected in 90% of sera samples from VL patients, which confirmshat A2 is expressed in the human host [59]. In addition, a signif-cant protection against L. donovani and L. amazonensis infectionsssociated with both humoral and cellular immune responses haseen obtained when using A2 as a recombinant protein or a DNAaccine in BALB/c mice, and more recently in dogs [60–65].
In this study, we engineered a recombinant L. tarentolae express-ng the L. donovani-specific A2 protein and used it as a live vaccinegainst L. infantum infection in BALB/c mice. We provide evidencehat intravenous (i.v.) and to a larger extent intraperitoneal (i.p.)mmunization with the recombinant L. tarentolae-A2 strain elicitedavorable immune responses and significant levels of protectivemmunity against L. infantum infectious challenge.
. Materials and methods
.1. DNA constructs
The A2 gene (with 3′UTR sequences) was amplified fromKSNEOA2-1 vector [58] (kindly provided by Dr. Greg Mat-
ashewski, Microbiology and Immunology Department, McGill
28 (2010) 53–62
University, Montreal, Quebec, Canada) by hot-start PCR (Hot-Star kit, Qiagen) using the following primers: (forward,5′-TTGAAGCTTACCGAGCACAATGAAGATCC-3′); and (reverse,5′-AACAAGCTTAGCAGAGGAAGTCAGCAAGG-3′) including HindIIIrestriction sites indicated as underlined. For amplification, DNAwas denatured at 95 ◦C for 5 min, 30 cycles of 94 ◦C for 30 s, 61 ◦C for30 s and 72 ◦C for 90 s, and a cycle of 72 ◦C for 10 min. The amplifiedfragment was cloned into the pDrive cloning vector (Qiagen). Aftersequence confirmation, the A2 fragment was subcloned into theHindIII site of vector pNEO-GFP [66] upstream of the GFP geneto generate pNEO-GFP-A2, and the correct insert orientation wasconfirmed by restriction analysis.
2.2. Parasite growth and transfections
The L. tarentolae Tar II (ATCC 30267) parasites were grownat pH 7.2 and 26 ◦C in M199 medium (Sigma) supplementedwith 10% heat-inactivated fetal calf serum (FCS, Gibco), 40 mMHEPES, 0.1 mM adenosine, 5 �g/ml hemin and 50 �g/ml gentam-icin. For transfection, 4 × 107 log-phase parasites were washed andre-suspended in 300 �l of electroporation buffer (21 mM HEPES,137 mM NaCl, 5 mM KCl, 0.7 mM Na2HPO4, 6 mM glucose; pH 7.5)containing 15 �g of plasmid DNA, stored on ice for 10 min, and elec-troporated using Bio-Rad Gene Pulser Ecell at 450 V and 500 �F asdescribed previously [67]. Transfectants were selected with G418(Sigma).
2.3. RNA extraction and reverse-transcription PCR
For L. tarentolae, RNA samples were extracted from both pro-mastigote and amastigote-like forms of the parasite. L. tarentolaeamastigote-like forms were prepared by culturing the parasitesat pH 4.5 and 37◦C for 10 h. In the case of L. infantum, amastig-otes were obtained by infecting mouse peritoneal macrophageswith stationary-phase parasites. Briefly, peritoneal exudate cellsfrom naïve BALB/c mice were used as a source of residentmacrophages (MQ). The peritoneal cavity was washed with 5 mlcold RPMI 1640 (Sigma) media, and then the media containingperitoneal macrophages was recovered. Cells were cultured inRPMI-1640 supplemented with 10% FCS, 2 mM glutamine, 50 nM2-mercaptoethanol, 10 mM HEPES and 40 �g/ml gentamicin ata density of 106 cells/well, and incubated at 37 ◦C in 5% CO2for 24 h. In order to infect MQ, the stationary-phase L. infantumpromastigotes were added at the ratio of 10:1 parasites ver-sus macrophages. The cultures were washed after 4 h to excludefree parasites and then incubated for 5 h. Infected attached MQswere collected using cold PBS. RNA was extracted using RNeasykit (Qiagen) and treated with RNase-free DNase for 30 min at37 ◦C to eliminate any remaining DNA. cDNA synthesis was per-formed using the Qiagen Omniscript RT Kit from 1 �g of RNA.To detect the A2 cDNA, two primers were designed to amplifya 154-bp region within the A2 3′-UTR starting at the proximityof A2 stop codon (AUF: 5′-GCAAGAGCAACAGCAGAGC-3′; AUR: 5′-CTTCCAACCTTGCGACTTTC-3′), and a hot-start PCR (Qiagen HotStarKit) was performed at 95 ◦C for 5 min, 40 cycles of 95 ◦C for 30 s,55 ◦C for 30 s and 72 ◦C for 45 s, and a cycle of 72 ◦C for 10 min.
2.4. Northern blot hybridization
Northern blot analysis was performed with the DIG System(Roche Applied Science) according to instructions of the manu-
facturer with minor modifications. For each sample, 3 �g of totalRNA was denatured, blotted onto Hybond + membrane (Roche), andUV cross-linked to the membrane with the intensity of 0.12 J/cm2.Using the “PCR DIG Probe Synthesis Kit” (Roche Applied Science), aDIG-labeled probe was prepared by PCR amplification of a 1.7 kb
accine
saNDte(ack
2
e3sTtiCs4asppw24w
2
abp(2bTatww(aamsaut
2
oti5aoi3
5 pmol of each forward and reverse primers, 12.5 �l Qiagen Quan-
A. Mizbani et al. / V
equence containing A2 ORF and part of its 3′-UTR using ADFnd ADR primers. After incubating in prehybridization buffer (10%-lauroylsarcosine, 0.02% SDS (w/v), 10% Blocking reagent, 0.02%enhardt solution, 2.5% Herring sperm in SSPE 5×) at 65 ◦C for 5 h,
he probe was added and incubation continued for 16 h. After sev-ral washing steps using 2× SSPE, anti Dig AP conjugated antibody1:5000 in blocking buffer) were used and incubated for 30 mint room temperature. Hybridized probes were detected usingolor substrate solution according to manual of Roche detectionit.
.5. Fluorescence microscopy and flow cytometry analysis
Both promastigote and amastigote forms of L. tarentolae werexamined for GFP expression. Promastigotes were centrifuged in000 rpm for 15 min and after washing once with PBS, cells were re-uspended in Hayman solution and mounted on microscope slides.o obtain amastigotes, mouse peritoneal macrophages were cul-ured on glass coverslips for 24 h to attach to the surface, and thennfected with stationary-phase L. tarentolae as described above.overslips were examined for GFP-expressing intracellular para-ites 24 h post-infection. Green fluorescence was excited with a88 nm argon-ion laser and was captured using a combination of510–530 nm bandpass emission filter. For flow cytometric mea-
urement, both non-infected and infected macrophages with eitherNEO-GFP or pNEO-GFP-A2-transfected L. tarentolae were pre-ared as described above. After cell detachment on ice and washingith PBS, the infection rate was monitored using at least a total of
0,000 events counted by PAS flow cytometer (Partec, Germany)8 h post-infection. The results were analyzed using Flowmax soft-are (Partec, Germany).
.6. Western blot analysis
Promastigote, amastigote, and amastigote-like forms of the par-sites were prepared as described above. Cells were then harvestedy centrifugation at 3000 rpm for 15 min and washed in PBS. Theellets were immediately lysed in 2× SDS-PAGE sample buffer4.5 mM Tris–HCl, pH 6.8, 10%, v/v glycerol, 2%, w/v SDS, 5%, v/v-mercaptoethanol, 0.05%, w/v bromophenol blue) on ice and thenoiled for 5 min. Samples were then loaded on a 15% SDS-PAGE.he gels were transferred onto a nitrocellulose membrane usingBio-Rad wet blotting system and incubated with blocking solu-
ion (PBS with 0.15% Tween 20 and 2.5% BSA) for 1 h. Washingas performed 3 times with 0.15% Tween 20 in PBS, and blotsere incubated overnight with anti-A2 monoclonal antibody C9
kindly provided by Dr. Greg Matlashewski) as the first antibodyt a 1:100 dilution. The membranes were washed three timesnd incubated for 90 min with peroxidase-conjugated goat anti-ouse IgG (1:1000, Sigma) as secondary antibodies. Unbound
econdary antibodies were removed by washing as describedbove. Diaminobenzidine tetrahydrochloride (DAB, Sigma) weresed as the substrate to detect the desired bands of the pro-eins.
.7. Immunization and challenge infection
Six groups of 8–10-week-old female BALB/c mice (n = 8) werebtained from the breeding stock maintained at the Pasteur Insti-ute of Iran. Two different routes of immunization, including i.p. and.v. were chosen. In the case of i.p. immunization, total number of
× 106 stationary-phase promastigotes were used. In the case of i.v.dministration, 2 × 108 stationary-phase L. tarentolae promastig-tes were injected. Groups 1 (i.v. route) and 2 (i.p. route) weremmunized with L. tarentolae expressing the GFP only and groups(i.v. route) and 4 (i.p. route) with recombinant L. tarentolae-A2.
28 (2010) 53–62 55
The fifth group received PBS only and the sixth group correspondsto naïve animals. Six weeks later, animals were challenged with107 virulent stationary-phase L. infantum promastigotes throughthe lateral tail vein. The parasite burden in the spleens and liverswas quantitatively determined 4 weeks after challenge by real timePCR.
2.8. Determination of antibody responses
Sera were taken from all groups before and 4 weeks after chal-lenge. Total IgG, IgG1 and IgG2a were measured using SolubleLeishmania Antigen (SLA) as coating antigen and performed byELISA. Briefly, 96-well plates (Greiner) were coated with 10 �g/mlSLA in PBS and incubated at 4 ◦C overnight. Then plates wereblocked with 100 �l of 1% BSA in PBS for 2 h at 37 ◦C. 100 �l ofserum samples (1:500 dilutions) was then added and incubated for2 h at 37 ◦C. After three washes, horseradish peroxidase-conjugatedgoat anti-mouse IgG (1:5000, Sigma) or biotinylated rabbit anti-mouse IgG1 (1:10,000) or IgG2a (1:1000, Zymed) were added andincubated for 2 h at 37 ◦C, and then plates were incubated for1 h at 37 ◦C with streptavidine-conjugated horseradish peroxidase(BRL, Gaithersburg, MD). After four washes, plates were devel-oped with O-phenylenediamine (OPD) as substrate. Reactions werestopped with 4N sulfuric acid and the absorbance was measuredat 492 nm.
2.9. Cytokine assay
Four mice from each group were sacrificed before and also 4weeks after challenge and spleens were homogenized. After lysisusing ACK lysis buffer (0.15 M NH4Cl, 10 mM KHCO3 and 0.1 mMNa2EDTA), splenocytes were washed and re-suspended in com-plete RPMI medium (RPMI-1640 supplemented with 5% FCS, 1%l-glutamine, 1% HEPES, 0.1% 2ME, 0.1% gentamicin). Cells werethen seeded at a density of 2 × 105 cells/well in the presence of SLA(10 �g/ml) or medium alone. Concanavalin A (Con A; 5 �g/ml) wasalso used in all experiments as the positive control. Plates wereincubated for 5 days at 37 ◦C in 5% CO2 humidified atmosphere.The IFN-� and IL-5 production in supernatants of splenocyte cul-tures was measured by sandwich ELISA kits (R&D, Minneapolis, MN,USA), according to the manufacturer’s instructions. All experimentswere run in duplicates.
2.10. Real time PCR
Real time PCR was used to quantify parasite burden inspleens and livers 4 weeks after challenge. Four mice from eachgroup were sacrificed and genomic DNA was extracted from10 mg of spleen and 30 mg of liver tissues using DNeasy Blood& Tissue kit (Qiagen). Two set of primers were designed byusing Primer Express software version 1.3.1 (Applied Biosys-tems), targeting a region of kinetoplastid minicircle DNA of L.infantum (forward: 5′-GCGCATACTGCAGTGAATTGAA-3′; reverse:5′-CTACCCGGAGGACCAGAAAAGT-3′). Absolute copy number of thetarget sequence was measured using Applied Biosystem 7500 realtime PCR system. L. infantum genomic DNA was used in 10-folddilutions corresponding to 2 × 105 parasites and used in real timePCR to draw the standard curve. For quantification of parasites intissues, 600 ng of DNA was subjected to the reaction containing
tiFast SYBR Green Master Mix in total volume of 25 �l. All reactionswere performed in duplicate. Conditions for PCR amplification wereas follows: 95 ◦C for 5 min; 40 cycles consisting of 95 ◦C for 15 s, 60◦
C for 20 s, and 72 ◦C for 40 s. Specific amplification of the targetregion was confirmed by gel electrophoresis of the PCR products.
5 accine
2
wD
3
3g
i
Flappaalm
6 A. Mizbani et al. / V
.11. Statistical analysis
All data comparisons were examined for significance by one-ay ANOVA and Student’s t-test using ‘SigmaPlot’ software.ifferences were considered statistically significant when P < 0.05.
. Results
.1. Generation of recombinant L. tarentolae expressing the A2ene
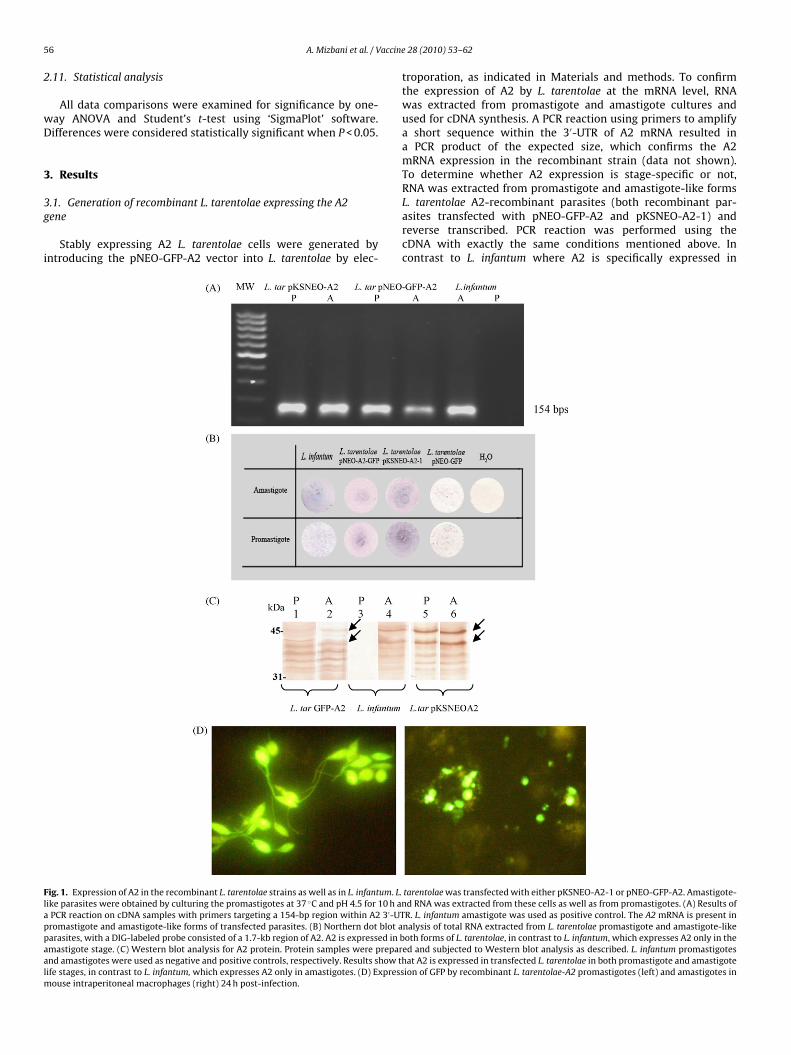
Stably expressing A2 L. tarentolae cells were generated byntroducing the pNEO-GFP-A2 vector into L. tarentolae by elec-
ig. 1. Expression of A2 in the recombinant L. tarentolae strains as well as in L. infantum. L.ike parasites were obtained by culturing the promastigotes at 37 ◦C and pH 4.5 for 10 h anPCR reaction on cDNA samples with primers targeting a 154-bp region within A2 3′-UTromastigote and amastigote-like forms of transfected parasites. (B) Northern dot blot aarasites, with a DIG-labeled probe consisted of a 1.7-kb region of A2. A2 is expressed in bmastigote stage. (C) Western blot analysis for A2 protein. Protein samples were preparnd amastigotes were used as negative and positive controls, respectively. Results show thife stages, in contrast to L. infantum, which expresses A2 only in amastigotes. (D) Express
ouse intraperitoneal macrophages (right) 24 h post-infection.
28 (2010) 53–62
troporation, as indicated in Materials and methods. To confirmthe expression of A2 by L. tarentolae at the mRNA level, RNAwas extracted from promastigote and amastigote cultures andused for cDNA synthesis. A PCR reaction using primers to amplifya short sequence within the 3′-UTR of A2 mRNA resulted ina PCR product of the expected size, which confirms the A2mRNA expression in the recombinant strain (data not shown).To determine whether A2 expression is stage-specific or not,RNA was extracted from promastigote and amastigote-like forms
L. tarentolae A2-recombinant parasites (both recombinant par-asites transfected with pNEO-GFP-A2 and pKSNEO-A2-1) andreverse transcribed. PCR reaction was performed using thecDNA with exactly the same conditions mentioned above. Incontrast to L. infantum where A2 is specifically expressed intarentolae was transfected with either pKSNEO-A2-1 or pNEO-GFP-A2. Amastigote-d RNA was extracted from these cells as well as from promastigotes. (A) Results ofR. L. infantum amastigote was used as positive control. The A2 mRNA is present innalysis of total RNA extracted from L. tarentolae promastigote and amastigote-likeoth forms of L. tarentolae, in contrast to L. infantum, which expresses A2 only in the
ed and subjected to Western blot analysis as described. L. infantum promastigotesat A2 is expressed in transfected L. tarentolae in both promastigote and amastigote
ion of GFP by recombinant L. tarentolae-A2 promastigotes (left) and amastigotes in

A. Mizbani et al. / Vaccine 28 (2010) 53–62 57
F entola4 fectivL ts wer
af(
sftFaTaAfNtciLdfm
a(wtti2
ig. 2. Percentage of mouse peritoneal macrophages infected with transfected L. tar8 h post-infection. Uninfected macrophages were used as a negative control (A). In. tarentolae pNEO-GFP. The experiment was repeated three times and similar resul
mastigotes, in L. tarentolae pNEO-GFP-A2 and pKSNEO-A2 trans-ectants A2 is expressed similarly in both developmental stagesFig. 1A).
Consistently, Northern blot analysis using an A2-specific probehows that A2 is expressed at approximately equal level in bothorms of the A2-transfected L. tarentolae, in contrast to L. infan-um which expresses A2 only in amastigotes, as demonstrated inig. 1B. Both promastigote and amastigote-like forms of L. tarentolaes well as L. infantum were also examined for A2 protein expression.otal protein was obtained and subjected to Western blot analysiss described in Section 2. Results presented in Fig. 1C show that2 protein is expressed in both promastigote and amastigote-like
orms of transfected L. tarentolae. Overall, the results of RT-PCR,orthern blot hybridization and Western blot analysis are consis-
ent with the exogenously provided A2 gene in L. tarentolae beingonstitutively expressed. Interestingly, the 3′UTR of the A2 mRNAs not able to promote amastigote-specific regulation, as it does in. infantum. Although further investigation is required to draw anyefinitive conclusions, we hypothesize that the mechanism or theactors responsible for stage-specific regulation of the A2 gene are
issing or not functional in L. tarentolae.Expression of GFP in transfected parasites in both promastigote
nd amastigote stages was confirmed by fluorescence microscopyFig. 1D). Mouse intraperitoneal macrophages were infected
ith GFP-expressing L. tarentolae and the infectivity rates ofhe recombinant L. tarentolae-A2 parasites were compared to L.arentolae-GFP by measuring fluorescence levels by FACS. As shownn Fig. 2, recombinant L. tarentolae-GFP-A2 (48.60%) were close to-fold more infective than L. tarentolae-GFP (27.44%).
e with either pNEO-GFP (B) or pNEO-GFP-A2 (C), as determined by flow cytometryity rates are almost doubled in the case of A2-expressing L. tarentolae compared toe obtained.
3.2. Immunization with live recombinant L. tarentolae A2protects mice against L. infantum challenge
We first examined whether immunization with the A2-recombinant L. tarentolae was protective against L. infantuminfectious challenge. Different groups of mice were immunizedeither with recombinant L. tarentolae parasites expressing GFP onlyor with L. tarentolae-GFP-A2 via the i.p. or i.v. routes and challengedwith virulent stationary-phase L. infantum 6 weeks later. The degreeof protection against infection was determined by quantifying para-site burden in the spleen and the liver by real time PCR 4 weeks afterchallenge. As shown in Fig. 3, immunization with A2-recombinant L.tarentolae via the i.p. route drastically (p < 0.05) reduced the infec-tion levels in both the liver (Fig. 3A) and the spleen (Fig. 3B) incontrast to the naïve control mice (group 5) (p = 0.008 and 0.01,respectively). Indeed, almost no parasites were detected in thespleens of mice vaccinated with the A2-recombinant L. tarentolae(Fig. 3B). Interestingly, this decrease in parasite burden was morepronounced in the case of A2-recombinant L. tarentolae-immunizedmice than in mice vaccinated with L. tarentolae expressing theGFP only. However, in contrast to liver, no significant differenceswere observed between the spleen parasite burdens in these twogroups when the vaccine regimens were administered by the i.v.route (p = 0.13). Results of immunization with A2-recombinant
L. tarentolae indicate that there is a highly significant differencebetween the levels of protection elicited depending on the routeof immunization, i.p. vs. i.v. In this case, the i.p. immunizationseems to result in higher levels of protection as compared to thei.v. immunization. These data emphasize the importance of the
58 A. Mizbani et al. / Vaccine
Fig. 3. Protection levels of immunized BALB/c mice challenged with L. infantum.Groups of BALB/c mice (n = 8) were immunized with GFP recombinant or A2-recombinant L. tarentolae via i.p. or i.v. routes. Groups 1 (i.v. route) and 2 (i.p. route)were immunized with L. tarentolae-GFP and groups 3 (i.v. route) and 4 (i.p. route)with recombinant L. tarentolae expressing A2. Group 5 received PBS and group 6corresponds to naïve animals. Six weeks after immunization, mice were challengedi 6
4we
ir
3
rpctbLmofSbitli((
.v. with 5 × 10 L. infantum promastigotes. Four mice were killed from each groupweeks after challenge and parasite burden in the liver (A) and the spleen (B)as quantified by real time PCR. The data are represented as the mean ± S.E. and
xperiment was performed two times.
mmunization route in stimulating the right branch of the immuneesponse.
.3. Protective immunity is associated with a potent Th1 response
The above experiments demonstrate that immunization withecombinant L. tarentolae expressing A2 confers a significantrotection against L. infantum infection in mice. We thereforeharacterized the type of immune response contributing to protec-ive immunity. We first focused on the cellular immune responseecause it is well established that high levels of protection againsteishmania infections require the presence of a strong Th1 cell-ediated immunity [68–70]. We therefore determined the level
f IFN-� and IL-5 production in cultures of splenocytes isolatedrom immunized and control mice in response to L. infantumLA as a recall antigen. Levels of IFN-� and IL-5 were comparedetween different groups after the infectious challenge as shown
n Fig. 4. Splenocytes from mice immunized via the i.p. route with
he recombinant L. tarentolae A2 produced significantly higherevels (∼1.7-fold) of IFN-� (p = 0.001) after challenge in compar-son to mice immunized with L. tarentolae expressing GFP onlyFig. 4A), while similar levels of IL-5 were detected in both groupsFig. 4B). Furthermore, significantly higher levels of IL-5 were gen-28 (2010) 53–62
erally observed after i.v. administration as compared to the i.p.immunization (p = 0.02), which along with IFN-� data confirm thati.p. injection leads to a stronger Th1 response. Immunization withboth GFP recombinant and A2-recombinant L. tarentolae by the i.p.route resulted in a higher IFN-� production levels in comparisonto the i.v. immunization, further stressing the substantial role ofthe route of immunization in the type of the immune response(Fig. 4A and B). Moreover, the post-challenge IFN-� production wasincreased significantly (p = 0.002) in comparison to the IFN-� lev-els before challenge in the group immunized with A2-expressing L.tarentolae via the i.p. route (Fig. 4A).
3.4. Immunization via the i.v. route induces higher level ofantibody production against L. infantum SLA
The level of total IgG, IgG1 and IgG2a antibodies against SLAfrom both L. infantum and L. tarentolae was determined by ELISAbefore and 4 weeks after challenge in all groups. As shown in Fig. 5,although IgG2a levels are similar in all groups immunized with L.tarentolae prior or after challenge with L. infantum, immunizationthrough the i.v. route leads to considerably higher levels (2–4-fold)of IgG1 antibodies in comparison to the i.p. route. Interestingly, incontrast to groups 1 and 2 (immunized with L. tarentolae-GFP, i.v.and i.p. respectively), the level of IgG1 did not increase in groups3 and 4 (immunized with L. tarentolae-GFP-A2, i.v. and i.p. respec-tively) both before and after challenge. In other words, only i.p.immunization with L. tarentolae has kept the IgG1 production atlow levels after challenge (although more pronounced in group 4in comparison to group 3). This is consistent with cytokine produc-tion which shows that IFN-� levels increase also in the case of i.p.immunization with recombinant L. tarentolae A2.
4. Discussion
Despite several efforts towards a vaccine development againstvisceral leishmaniasis (VL), no effective vaccine currently exists forhuman use. It is well established that infection with Leishmaniaresults in a natural protective immunity against subsequent chal-lenges. Based on this fact, leishmanization has been used for manyyears to generate protection, however, at the expense of unpleasantclinical episodes in many cases. Therefore, the induction of protec-tive immunity using either inactivated or attenuated live vaccineswould be a significant step in the control of the disease [71], andpreclinical data suggest that this approach holds the most promisefor an anti-leishmanial vaccine [19]. Furthermore, different experi-mental studies both in human and mice support this strategy. Nylénet al. [72] showed that live parasites were more effective in inducingthe production of IFN-� from human PBMCs than killed parasites.It is also demonstrated that differences in early cytokine responsein mice inoculated with live and killed parasites are primarily dueto loss of infectivity [73]. Infection with live parasites (virulent andavirulent) leads to substantial activation and IFN-� production fromboth CD4+ and CD8+ T cells whereas inoculation of killed parasitesonly leads to activation of CD4+ T cells [73]. Another report also indi-cates that live parasites are important for maintaining Leishmaniaspecific effector cells that mediate immediate protection follow-ing secondary challenge in contrast to killed parasites, which aredefective in their ability to maintain effector as well as memorycells [74].
In this study, we generated a recombinant L. tarentolae strain
expressing the L. donovani A2 gene shown to play a role in viscer-alization [75] and evaluated its potential as a live vaccine againstVL. Interestingly, the A2-expressing recombinant parasites showedhigher macrophage infectivity in comparison to GFP-expressing L.tarentolae used as a control. Immunization (i.v. and i.p.) of BALB/c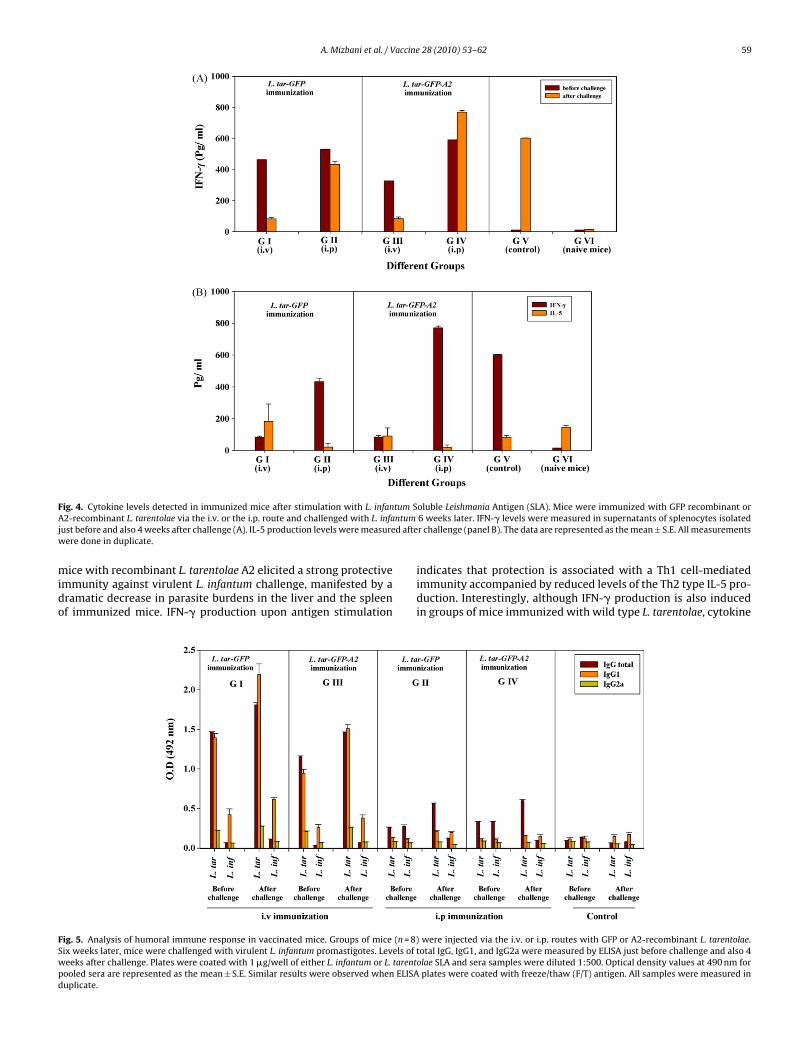
A. Mizbani et al. / Vaccine 28 (2010) 53–62 59
F tum SA ntumj d aftew
mido
FSwpd
ig. 4. Cytokine levels detected in immunized mice after stimulation with L. infan2-recombinant L. tarentolae via the i.v. or the i.p. route and challenged with L. infa
ust before and also 4 weeks after challenge (A). IL-5 production levels were measureere done in duplicate.
ice with recombinant L. tarentolae A2 elicited a strong protectivemmunity against virulent L. infantum challenge, manifested by aramatic decrease in parasite burdens in the liver and the spleenf immunized mice. IFN-� production upon antigen stimulation
ig. 5. Analysis of humoral immune response in vaccinated mice. Groups of mice (n = 8)ix weeks later, mice were challenged with virulent L. infantum promastigotes. Levels ofeeks after challenge. Plates were coated with 1 �g/well of either L. infantum or L. tarentooled sera are represented as the mean ± S.E. Similar results were observed when ELISAuplicate.
oluble Leishmania Antigen (SLA). Mice were immunized with GFP recombinant or6 weeks later. IFN-� levels were measured in supernatants of splenocytes isolatedr challenge (panel B). The data are represented as the mean ± S.E. All measurements
indicates that protection is associated with a Th1 cell-mediatedimmunity accompanied by reduced levels of the Th2 type IL-5 pro-duction. Interestingly, although IFN-� production is also inducedin groups of mice immunized with wild type L. tarentolae, cytokine
were injected via the i.v. or i.p. routes with GFP or A2-recombinant L. tarentolae.total IgG, IgG1, and IgG2a were measured by ELISA just before challenge and also 4olae SLA and sera samples were diluted 1:500. Optical density values at 490 nm for
plates were coated with freeze/thaw (F/T) antigen. All samples were measured in

6 accine
lnaatsnrOu[thiwatalamhnv
ccblcIlLtbirair
eniosarllcoitrtEbpfm
Aer[
[
[
[
[
[
[
[
[
[
[
[
[
0 A. Mizbani et al. / V
evels are increased in the group immunized with the recombi-ant L. tarentolae A2 and especially when the vaccine regimen isdministered via the i.p. route. This highlights the effect of A2ntigen in eliciting a potent Th-1 type response. This is consis-ent with epitope mapping studies on A2, which have shown thateveral A2 epitopes located in the repetitive sequences are recog-ized by CD8+ T cells and also an epitope in its N-terminal part isecognized by IFN-�-producing Th1-associated CD4+ T cells [64].ur data are also in agreement with previously published reportssing A2 in a vaccine regimen as part of a recombinant protein61,63] or a DNA vaccine [65] or a live recombinant viral vec-or [64] demonstrating the potential of A2 antigen to induce bothumoral and cellular immune responses and protective immunity
n mice and more recently in vaccinated dogs [60]. Furthermore,ild type L. tarentolae has been previously shown to be protective
gainst L. donovani infection [53]. Recently, it has been reportedhat qualitative differences in early immune response with live par-sites contribute to their ability to induce protective immunity. Theongevity of effectors cells and differences in their ability to inducend maintain different subsets of memory cells are among theost important factors [73]. In fact, we propose that these factors
ave possibly played a role in potentiating the protective immu-ity elicited by A2 in the presence of the recombinant L. tarentolaeector.
Characterization of the antibody response also corroborated theytokine assay findings. It has been shown previously that Th2ellular response is associated with increased level of IgG1 anti-ody subclass [76], whereas Th1 response, characterized by high
evels of IFN-� production, promotes the generation of IgG2a sub-lass [77]. Therefore, it can be concluded that maintaining thegG1 level at a constant level after challenge and before chal-enge, as observed in the group immunized with A2-recombinant. tarentolae through the i.p. route, is probably due to the induc-ion of a more pronounced Th1 response. In addition, it haseen well established that the increased level of IgG1 antibod-
es titers is indicative of the L. infantum load [78]. The antibodyesponse shown here indicates that the least IgG1 level is associ-ted with the least parasite burden, both observed in the groupmmunized with the recombinant L. tarentolae A2 via the i.p.oute.
Studies on the effect of immunization route on the type oflicited immune response (Th1 or Th2) have shown that immu-ization via the i.p. rather than the i.v., s.c. and i.m. routes results
n higher levels of IFN-� and IgG2a antibody, and in lower levelsf Th2-associated cytokines (IL-4 and IL-10) and IgG1 antibodyubclass [79]. Here, we show that intravenous immunization,s opposed to intraperitoneal immunization, elicited a Th2-likeesponse, characterized by significantly lower level of IFN-�, higherevels of IL-5, and high humoral immune response, resulting iness efficient protection. Recently, it has been reported that spleenells of BALB/c mice immunized with L. donovani promastig-te membrane antigens (LAg) through different routes (e.g. s.c.,.m., and i.v. in comparison to i.p.) have an increased capacityo produce pre-challenge TGF-� during an in vitro antigen-recallesponse [80]. Interestingly, i.p. immunization induced produc-ion of pre-challenge IFN-�, IL-12 and IL-4 with a Th1 bias.xposure to antigen-stimulated splenocyte supernatants of i.p.ut not s.c. immunized mice activated macrophages for in vitroarasite killing. Therefore, the route of injection could induce dif-erent immunological responses in animal model such as BALB/c
ice.
In conclusion, the results presented here demonstrate that2-expressing recombinant L. tarentolae shows a promise as anffective live vaccine against L. infantum infection. L. tarentolaeepresents a completely safe live vector, as previously reported53]. There is no sign of any pathology with L. tarentolae even
[
28 (2010) 53–62
in immunocompromised SCID mice. Although non-pathogenic, L.tarentolae can efficiently target antigen-presenting cells and differ-entiate inside macrophages [53]. It is clearly incapable of promotingdisease, however, our PCR analysis data indicated that this parasitecan be found in the spleen and the liver of mice infected with theA2-recombinant L. tarentolae, albeit at low levels, 4 weeks afterimmunization (data not shown). Therefore, it appears that L. tar-entolae is able to persist at very low levels in mice. In addition, A2per se, has proven to be a promising vaccine candidate, capableof inducing potent Th1 immune response against L. infantum infec-tious challenge. A2 has also been associated with the visceralizationprocess [75]. It is therefore possible that L. tarentolae expressing A2may be better armed to survive within the host than the wild typeL. tarentolae strain. We are currently pursuing this hypothesis inour laboratory.
Acknowledgments
We are thankful to Prof. Greg Matlashewski (Departmentof Microbiology and Immunology, McGill University, Montreal,Canada) for providing us with vector pKSNEOA2-1. We would alsolike to thank Shima Safayeian for genomic DNA preparation.
References
[1] Badaro R, Jones TC, Carvalho EM. New perspectives on a subclinical form ofvisceral leishmaniasis. J Infect Dis 1986;154:1003–11.
[2] Pearson RD, De Queiroz, Sousa A. Clinical spectrum of leishmaniasis. Clin InfectDis 1996;22:1–13.
[3] Desjeux P. Leishmania and HIV in gridlock. Publication nos.WHO/CTD/LEISH/98.9, UNAIDS/98.23. Geneva: World Health Organization;1998.
[4] Desjeux P. Leishmaniasis: current situation and new perspectives. CompImmunol Microbiol Infect Dis 2004;27:305–18.
[5] Graham PCC. Introduction. In: Peters W, Killick-Kendrick R, editors. The leish-maniasis in biology and medicine. New York: Academic Press; 1987.
[6] Herwaldt BL. Leishmaniasis. Lancet 1999;354:1191–9.[7] World Health Organization. Leishmania/HIV co-infection in South-
Western Europe 1990–1998: retrospective analysis of 965 cases, Geneva.WHO/LEISH/2000 2000;42:1–12.
[8] Desjeux P, Alvar J. Leishmania/HIV co-infections: epidemiology in Europe. AnnTrop Med Parasitol 2003;97(Suppl. 1):S17–28.
[9] Sinha PK, Pandey K, Bhattacharya SK. Diagnosis & management of Leishma-nia/HIV co-infection. Indian J Med Res 2005;121:407–14.
10] World Health Organization. Advances in the battle against leishmaniasis. TDRNews 1998;57:2.
11] Croft SL. Recent development in the chemotherapy of leishmaniasis. TrendsPharmacol Sci 1988;9:376–81.
12] Murray HW, Berman JD, Wright SD. Immunochemotherapy for intracellularLeishmania donovani infection: �-interferon plus pentavalent antimony. J InfectDis 1988;157:973–8.
13] Carter KC, Sundar S, Spickett C, Pereira OC, Mullen AB. The in vivo susceptibilityof Leishmania donovani to sodium stibogluconate is drug specific and can bereversed by inhibiting glutathione biosynthesis. Antimicrob Agents Chemother2003;47:1529–35.
14] Pérez-Victoria FJ, Castanys S, Gamarro F. Leishmania donovani resistance tomiltefosine involves a defective inward translocation of the drug. AntimicrobAgents Chemother 2003;47:2397–403.
15] Ponte-Sucre A. Physiological consequences of drug resistance in Leishmania andtheir relevance for chemotherapy. Kinetoplastid Biol Dis 2003;2:14.
16] Sundar S. Drug resistance in Indian visceral leishmaniasis. Trop Med Int Health2001;6:849–54.
17] Sundar S, More DK, Singh MK, Singh VP, Sharma S, Makharia A, et al. Failure ofpentavalent antimony in visceral leishmaniasis in India: report from the centerof the Indian epidemic. Clin Infect Dis 2000;31:1104–7.
18] Modabber F. Vaccines against leishmaniasis. Ann Trop Med Parasitol1995;89(Suppl. 1):83–8.
19] Handman E. Leishmaniasis: current status of vaccine development. Clin Micro-biol Rev 2001;14:229–43.
20] Sukumaran B, Madhubala R. Leishmaniasis: current status of vaccine develop-ment. Curr Mol Med 2004;4:667–79.
21] Connell ND, Medina-Acosta E, McMaster WR, Bloom BR, Russell DG. Effec-
tive immunization against cutaneous leishmaniasis with recombinant bacilleCalmette-Guerin expressing the Leishmania surface proteinase gp63. Proc NatlAcad Sci USA 1993;90:11473–7.22] Mendonca SCF, Russell DG, Coutinho SG. Analysis of the human T cell respon-siveness to purified antigens of Leishmania: lipophosphoglycan (LPG) andglycoprotein 63 (gp 63). Clin Exp Immunol 1991;83:472–8.

accine
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
A. Mizbani et al. / V
23] McMahon-Pratt D, Rodriguez D, Rodriguez J-R, Zhang Y, Manson K, BergmanC, et al. Recombinant vaccinia viruses expressing GP46/M-2 protect againstLeishmania infection. Infect Immun 1993;61:3351–9.
24] Gurunathan S, Sacks DL, Brown DR, Reiner SL, Charest H, Glaichenhaus N, et al.Vaccination with DNA encoding the immunodominant LACK parasite antigenconfers protective immunity to mice infected with Leishmania major. J Exp Med1997;186:1137–47.
25] Mougneau E, Altare F, Wakil AE, Zheng S, Coppola T, Wang Z-E, et al. Expressioncloning of a protective Leishmania antigen. Science 1995;268:563–6.
26] Rafati S, Baba AA, Bakhshayesh M, Vafa M. Vaccination of BALB/c mice withLeishmania major amastigote-specific cysteine proteinase. Clin Exp Immunol2000;120:134–8.
27] Dole VS, Raj VS, Ghosh A, Madhubala R, Myler PJ, Stuart KD. Immunizationwith recombinant LD1 antigens protects against experimental leishmaniasis.Vaccine 2000;19:423–30.
28] Handman E, Symons FM, Baldwin TM, Curtis JM, Scheerlinck J-PY. Protectivevaccination with promastigote surface antigen 2 from Leishmania major ismediated by a TH1 type of immune response. Infect Immun 1995;63:4261–7.
29] Skeiky YAW, Coler RN, Brannon M, Stromberg E, Greeson K, Thomas CraneR, et al. Protective efficacy of a tandemly linked, multi-subunit recombi-nant leishmanial vaccine (Leish-111f) formulated in MPL adjuvant. Vaccine2002;20:3292–303.
30] Webb JR, Campos-Neto A, Ovendale PJ, Martin TI, Stromberg EJ, Badaro R, et al.Human and murine immune responses to a novel Leishmania major recombi-nant protein encoded by members of a multicopy gene family. Infect Immun1998;66:3279–89.
31] Saravia NG, Hazbón MH, Osorio Y, Valderrama L, Walker J, Santrich C, et al.Protective immunogenicity of the paraflagellar rod protein 2 of Leishmaniamexicana. Vaccine 2005;23:984–95.
32] Stager S, Smith DF, Kaye PM. Immunization with a recombinant stage-regulatedsurface protein from Leishmania donovani induces protection against visceralleishmaniasis. J Immunol 2000;165:7064–71.
33] Skeiky YAW, Kennedy M, Kaufman D, Borges MM, Guderian JA, Scholler JK, etal. LeIF: a recombinant Leishmania protein that induces an IL-12-mediated Th1cytokine profile. J Immunol 1998;161:6171–9.
34] Wilson ME, Young BM, Andersen KP, Weinstock JV, Metwali A, Ali KM, et al.A recombinant Leishmania chagasi antigen that stimulates cellular immuneresponses in infected mice. Infect Immun 1995;63:2062–9.
35] Champsi J, McMahon-Pratt D. Membrane glycoprotein M-2 protects againstLeishmania amazonensis infection. Infect Immun 1988;56:3272–9.
36] Coler RN, Reed SG. Second-generation vaccines against leishmaniasis. TrendsParasitol 2005;21:244–9.
37] Murray HW, Berman JD, Davies CR, Saravia NG. Advances in leishmaniasis.Lancet 2005;366:1561–77.
38] Piedrafita D, Xu D, Hunter D, Harrison RA, Liew FY. Protective immuneresponses induced by vaccination with an expression genomic library of Leish-mania major. J Immunol 1999;163:1467–72.
39] Belkaid Y, Piccirillo CA, Mendez S, Shevach EM, Sacks DL. CD4+CD25+ reg-ulatory T cells control Leishmania major persistence and immunity. Nature2002;420:502–7.
40] Uzonna JE, Wei G, Yurkowski D, Bretscher P. Immune elimination of Leishmaniamajor in mice: implications for immune memory, vaccination, and reactivationdisease. J Immunol 2001;167:6967–74.
41] Rivier D, Bovay P, Shah R, Didisheim S, Mauel J. Vaccination against Leishmaniamajor in a CBA mouse model of infection: role of adjuvants and mechanism ofprotection. Parasite Immunol 1999;21:461–73.
42] Mitchell GF, Handman E, Spithill TW. Vaccination against cutaneous leishma-niasis in mice using nonpathogenic cloned promastigotes of Leishmania majorand importance of route of injection. Aust J Exp Biol Med Sci 1984;62:145–53.
43] Gorczynski RM. Immunization of susceptible BALB/c mice against Leishmaniabraziliensis. II. Use of temperature-sensitive avirulent clones of parasite forvaccination purposes. Cell Immunol 1985;94:11–20.
44] Kimsey PB, Theodos CM, Mitchen TK, Turco SJ, Titus RG. An avirulentlipophosphoglycan-deficient Leishmania major clone induces CD4+ T cellswhich protect susceptible BALB/c mice against infection with virulent L. major.Infect Immun 1993;61:5205–13.
45] Daneshvar H, Coombs GH, Hagan P, Phillips RS. Leishmania mexicana and Leish-mania major: attenuation of wild-type parasites and vaccination with theattenuated lines. J Infect Dis 2003;187:1662–8.
46] Titus RG, Gueiros-Filho FJ, De Freitas LAR, Beverley SM. Development of asafe live Leishmania vaccine line by gene replacement. Proc Natl Acad Sci USA1995;92:10267–71.
47] Alexander J, Coombs GH, Mottram JC. Leishmania mexicana cysteine proteinase-deficient mutants have attenuated virulence for mice and potentiate a Th1response. J Immunol 1998;161:6794–801.
48] Hübel A, Krobitsch S, Hörauf A, Clos J. Leishmania major Hsp100 is requiredchiefly in the mammalian stage of the parasite. Mol Cell Biol 1997;17:5987–95.
49] Silvestre R, Cordeiro-da-Silva A, Santarém N, Vergnes B, Sereno D, Ouaissi A.SIR2-deficient Leishmania infantum induces a defined IFN-�/IL-10 pattern thatcorrelates with protection. J Immunol 2007;179:3161–70.
50] Uzonna JE, Späth GF, Beverley SM, Scott P. Vaccination with phosphoglycan-deficient Leishmania major protects highly susceptible mice from virulent chal-lenge without inducing a strong Th1 response. J Immunol 2004;172:3793–7.
51] Papadopoulou B, Roy G, Breton M, Kündig C, Dumas C, Fillion I, et al. Reducedinfectivity of a Leishmania donovani biopterin transporter genetic mutant andits use as an attenuated strain for vaccination. Infect Immun 2002;70:62–8.
[
[
28 (2010) 53–62 61
52] Dumas C, Ouellette M, Tovar J, Cunningham ML, Fairlamb AH, Tamar S, et al.Disruption of the trypanothione reductase gene of Leishmania decreases itsability to survive oxidative stress in macrophages. EMBO J 1997;16:2590–8.
53] Breton M, Tremblay MJ, Ouellette M, Papadopoulou B. Live nonpathogenic par-asitic vector as a candidate vaccine against visceral leishmaniasis. Infect Immun2005;73:6372–82.
54] Wilson VCLC, Southgate BA. Lizard leishmania. In: Lumsden WH, Evans DA,editors. Biology of the Kinetoplastida. San Diego, CA: Academic Press; 1978. p.244–68.
55] Breton M, Zhao C, Ouellette M, Tremblay MJ, Papadopoulou B. A recombi-nant non-pathogenic Leishmania vaccine expressing human immunodeficiencyvirus 1 (HIV-1) Gag elicits cell-mediated immunity in mice and decreases HIV-1 replication in human tonsillar tissue following exposure to HIV-1 infection. JGen Virol 2007;88:217–25.
56] Azizi H, Hassani K, Taslimi Y, Shateri Najafabadi H, Papadopoulou B, RafatiS. Searching for virulence factors in the non-pathogenic parasite to humansLeishmania tarentolae. Parasitology 2009;136:723–35.
57] Charest H, Matlashewski G. Developmental gene expression in Leishmaniadonovani: differential cloning and analysis of an amastigotestage-specific gene.Mol Cell Biol 1994;14:2975–84.
58] Zhang WW, Charest H, Ghedin E, Matlashewski G. Identification and overex-pression of the A2 amastigote-specific protein in Leishmania donovani. MolBiochem Parasitol 1996;78:79–90.
59] Ghedin E, Zhang WW, Charest H, Sundar S, Kenney RT, Matlashewski G. Anti-body response against a Leishmania donovani amastigote-stage-specific proteinin patients with visceral leishmaniasis. Clin Diagn Lab Immunol 1997;4:530–5.
60] Fernandes AP, Costa MMS, Coelho EAF, Michalick MSM, de Freitas E, MeloMN, et al. Protective immunity against challenge with Leishmania (Leishma-nia) chagasi in beagle dogs vaccinated with recombinant A2 protein. Vaccine2008;26:5888–95.
61] Ferraz Coelho EA, Pereira Tavares CA, Amorim Carvalho FA, Chaves KF, Teix-eira KN, Rodrigues RC, et al. Immune responses induced by the Leishmania(Leishmania) donovani A2 antigen, but not by the LACK antigen, are protec-tive against experimental Leishmania (Leishmania) amazonensis infection. InfectImmun 2003;71:3988–94.
62] Ghosh A, Labrecque S, Matlashewski G. Protection against Leishmania donovaniinfection by DNA vaccination: increased DNA vaccination efficiency throughinhibiting the cellular p53 response. Vaccine 2001;19:3169–78.
63] Ghosh A, Zhang WW, Matlashewski G. Immunization with A2 protein resultsin amixed Th1/Th2 and a humoral response which protects mice against Leish-mania donovani infections. Vaccine 2001;20:59–66.
64] Resende DM, Caetano BC, Dutra MS, Penido MLO, Abrantes CF, Verly RM, etal. Epitope mapping and protective immunity elicited by adenovirus express-ing the Leishmania amastigote specific A2 antigen: Correlation with IFN-� andcytolytic activity by CD8+ T cells. Vaccine 2008;26:4585–93.
65] Zanin FHC, Coelho EAF, Tavares CAP, Marques-da-Silva EA, Silva Costa MM,Rezende SA, et al. Evaluation of immune responses and protection inducedby A2 and nucleoside hydrolase (NH) DNA vaccines against Leishmania cha-gasi and Leishmania amazonensis experimental infections. Microbes Infect2007;9:1070–7.
66] Rochette A, McNicoll F, Girard J, Breton M, Leblanc E, Bergeron MG, et al.Characterization and developmental gene regulation of a large gene familyencoding amastin surface proteins in Leishmania spp. Mol Biochem Parasitol2005;140:205–20.
67] Papadopoulou B, Roy G, Ouellette M. A novel antifolate resistance gene on theamplified H circle of Leishmania. EMBO J 1992;11:3601–8.
68] Liew FY, O’Donnell CA. Immunology of leishmaniasis. Adv Parasitol1993;32:161–259.
69] Miralles GD, Stoeckle MY, McDermott DF, Finkelman FD, Murray HW. Th1 andTh2 cell-associated cytokines in experimental visceral leishmaniasis. InfectImmun 1994;62:1058–63.
70] Tripathi P, Singh V, Naik S. Immune response to Leishmania: paradox ratherthan paradigm. FEMS Immunol Med Microbiol 2007;51:229–42.
71] Amaral VF, Teva A, Oliveira-Neto MP, Silva AJ, Pereira MS, Cupolillo E, et al. Studyof the safety, immunogenicity and efficacy of attenuated and killed Leishmania(Leishmania) major vaccines in a rhesus monkey (Macaca mulatta) model ofthe human disease. Mem Inst Oswaldo Cruz 2002;97:1041–8.
72] Nylén S, Mörtberg U, Kovalenko D, Satti I, Engström K, Bakhiet M, et al. Differen-tial induction of cellular responses by live and dead Leishmania promastigotesin healthy donors. Clin Exp Immunol 2001;124:43–53.
73] Okwor I, Liu D, Uzonna J. Qualitative differences in the early immune responseto live and killed Leishmania major: Implications for vaccination strategiesagainst Leishmaniasis. Vaccine 2009;27:2554–62.
74] Mendez S, Reckling SK, Piccirillo CA, Sacks D, Belkaid Y. Role for CD4+ CD25+
regulatory T cells in reactivation of persistent Leishmaniasis and control ofconcomitant immunity. J Exp Med 2004;200:201–10.
75] Zhang WW, Matlashewski G. Characterization of the A2-A2rel gene cluster inLeishmania donovani: involvement of A2 in visceralization during infection. MolMicrobiol 2001;39:935–48.
76] Warren HS, Vogel FR, Chedid LA. Current status of immunological adjuvants.
Annu Rev Immunol 1986;4:369–88.77] Snapper CM, Paul WE. Interferon-� and B cell stimulatory factor-1 reciprocallyregulate Ig isotype production. Science 1987;236:944–7.
78] Basu R, Bhaumik S, Basu JM, Naskar K, De T, Roy S. Kinetoplastid membraneprotein-11 DNA vaccination induces complete protection against both pen-tavalent antimonial-sensitive and -resistant strains of Leishmania donovani

6 accine
[
2 A. Mizbani et al. / V
that correlates with inducible nitric oxide synthase activity and IL-4 genera-tion: evidence for mixed Th1- and Th2-like responses in visceral leishmaniasis.Immunology 2005;174:7160–71.
79] Kaur S, Kaur T, Garg N, Mukherjee S, Raina P, Athokpam V. Effect of doseand route of inoculation on the generation of CD4+ Th1/Th2 type of immune
[
28 (2010) 53–62
response in murine visceral leishmaniasis. Parasitol Res 2008;103:1413–9.
80] Bhowmick S, Mazumdar T, Ali N. Vaccination route that induces transform-ing growth factor beta production fails to elicit protective immunity againstLeishmania donovani infection. Infect Immun 2009;77:1514–23.



















