
Edgecombe, G.D., García-Bellido, D.C. & Paterson, J.R., 2011. A new leanchoiliid megacheiran...
16
A new leanchoiliid megacheiran arthropod from the lower Cambrian Emu Bay Shale, South Australia GREGORY D. EDGECOMBE, DIEGO C. GARCÍA−BELLIDO, and JOHN R. PATERSON Edgecombe, G.D., García−Bellido, D.C., and Paterson, J.R. 2011. A new leanchoiliid megacheiran arthropod from the lower Cambrian Emu Bay Shale, South Australia. Acta Palaeontologica Polonica 56 (2): 385–400. The Leanchoiliidae is well−known from abundant material of Leanchoilia, from the Burgess Shale and Chengjiang Konservat−Lagerstätten. The first Australian member of the group is Oestokerkus megacholix gen. et sp. nov., described from the Emu Bay Shale (Cambrian Series 2, Stage 4), at Buck Quarry, Kangaroo Island, South Australia, and is interme− diate in age between the well known leanchoiliid species from the Burgess Shale and Chengjiang. Phylogenetic analysis of “short great appendage” arthropods (Megacheira) in the context of the chelicerate stem group resolves the Australian species as sister to Burgess Shale, Utah, and Chengjiang Leanchoilia species, but most readily distinguished from Leanchoilia and Alalcomenaeus by a different telson shape, interpreted as being forked, widening distally, and with a few dorsally curved spines at the posterior angle. Leanchoiliid interrelationships are stable to alternative character weights, and Megacheira corresponds to a clade in most analyses. Key words: Arthropoda, Megacheira, Leanchoiliidae, Oestokerkus, Leanchoilia, Alalcomenaeus, midgut glands, phylog− eny, Cambrian, South Australia. Gregory D. Edgecombe [[email protected]], Department of Palaeontology, The Natural History Museum, Cromwell Road, London SW7 5BD, UK; Diego C. García−Bellido [[email protected]], Departamento de Paleontología, Instituto de Geología Econó− mica/Instituto de Geociencias (CSIC−UCM), José Antonio Novais 2, 28040−Madrid, Spain; John R. Paterson [[email protected]], Division of Earth Sciences, School of Environmental and Rural Science, Uni− versity of New England, Armidale, NSW 2351, Australia. Received 16 July 2010, accepted 7 February 2011, available online 9 February 2011. Introduction “Great appendage arthropods” have figured prominently in considerations of Cambrian arthropod systematics and the assignment of early arthropod lineages to the stem groups of major extant clades. An assemblage of taxa formalised as Megacheira Hou and Bergström, 1997, has variably been re− garded as belonging to the stem group of Chelicerata (Wills et al. 1998; Chen et al. 2004; Cotton and Braddy 2004) or to the stem group of all extant arthropods (Hou and Bergström 1997; Budd 2002; Daley et al. 2009). Megacheira, composed at least of the Cambrian taxa Leanchoiliida, Yohoiida, and Fortiforcipida, has been depicted as a clade (Hou and Berg− ström 1997: fig. 87), as a polyphyletic group in different parts of the euarthropod stem group (Bergström and Hou 2005), or as an unresolved polytomy in the chelicerate stem group (Chen et al. 2004: fig. 6). When conceived as a clade, the group is united by the “great appendage” having a few elongate spinose projections, the exopods being fringed with setae, and a large telson (Hou and Bergström 1997). The ex− treme ends of the spectrum on the phylogenetic position of Megacheira involve whether the so−called “short great ap− pendage” (Chen et al. 2004) is a precursor to the chelicera (Bousfeld 1995; Chen et al. 2004; Cotton and Braddy 2004; Bitsch and Bitsch 2007) or is the appendage of a segment that is anatomically anterior to the chelicera or antenna (Budd 2002). The anatomically best understood megacheirans belong to the Order Leanchoiliida Størmer, 1944, of which the gen− era Leanchoilia Walcott, 1912, and Alalcomenaeus Simo− netta, 1970, provide most available information. Alalcome− naeus cambricus Simonetta, 1970, has been revised based on Burgess Shale collections made through the 1980s and 1990s (Briggs and Collins 1999). Alalcomenaeus is apparently also represented in the Chengjiang fauna, based on A. elegans (Luo and Hu in Luo et al., 1999) (see Liu et al. 2007). The more common Leanchoilia has received even more study. The anatomy and taxonomy of Leanchoilia from both the Burgess Shale (García−Bellido and Collins 2007) and Chengjiang Lagerstätten (Liu et al. 2007) have recently been reviewed, the former updating descriptions by Bruton and Whittington (1983). Two species in the Burgess Shale (Lean− choilia superlata Walcott, 1912 and Leanchoilia persephone Simonetta, 1970) and a single species from Chengjiang (Leanchoilia illecebrosa [Hou, 1987]) have been redescribed based on extensive new collections. Leanchoilia illecebrosa subsumes several generic and specific names introduced by Luo et al. (1997) for Chengjiang material (see Liu et al. 2007 doi:10.4202/app.2010.0080 Acta Palaeontol. Pol. 56 (2): 385–400, 2011
Transcript of Edgecombe, G.D., García-Bellido, D.C. & Paterson, J.R., 2011. A new leanchoiliid megacheiran...

A new leanchoiliid megacheiran arthropod fromthe lower Cambrian Emu Bay Shale South Australia
GREGORY D EDGECOMBE DIEGO C GARCIacuteAminusBELLIDO and JOHN R PATERSON
Edgecombe GD GarciacuteaminusBellido DC and Paterson JR 2011 A new leanchoiliid megacheiran arthropod from thelower Cambrian Emu Bay Shale South Australia Acta Palaeontologica Polonica 56 (2) 385ndash400
The Leanchoiliidae is wellminusknown from abundant material of Leanchoilia from the Burgess Shale and ChengjiangKonservatminusLagerstaumltten The first Australian member of the group is Oestokerkus megacholix gen et sp nov describedfrom the Emu Bay Shale (Cambrian Series 2 Stage 4) at Buck Quarry Kangaroo Island South Australia and is intermeminusdiate in age between the well known leanchoiliid species from the Burgess Shale and Chengjiang Phylogenetic analysisof ldquoshort great appendagerdquo arthropods (Megacheira) in the context of the chelicerate stem group resolves the Australianspecies as sister to Burgess Shale Utah and Chengjiang Leanchoilia species but most readily distinguished fromLeanchoilia and Alalcomenaeus by a different telson shape interpreted as being forked widening distally and with a fewdorsally curved spines at the posterior angle Leanchoiliid interrelationships are stable to alternative character weightsand Megacheira corresponds to a clade in most analyses
Key words Arthropoda Megacheira Leanchoiliidae Oestokerkus Leanchoilia Alalcomenaeus midgut glands phylogminuseny Cambrian South Australia
Gregory D Edgecombe [gedgecombenhmacuk] Department of Palaeontology The Natural History MuseumCromwell Road London SW7 5BD UKDiego C GarciacuteaminusBellido [DiegoGBCgeoucmes] Departamento de Paleontologiacutea Instituto de Geologiacutea EconoacuteminusmicaInstituto de Geociencias (CSICminusUCM) Joseacute Antonio Novais 2 28040minusMadrid SpainJohn R Paterson [jpater20uneeduau] Division of Earth Sciences School of Environmental and Rural Science Uniminusversity of New England Armidale NSW 2351 Australia
Received 16 July 2010 accepted 7 February 2011 available online 9 February 2011
IntroductionldquoGreat appendage arthropodsrdquo have figured prominently inconsiderations of Cambrian arthropod systematics and theassignment of early arthropod lineages to the stem groups ofmajor extant clades An assemblage of taxa formalised asMegacheira Hou and Bergstroumlm 1997 has variably been reminusgarded as belonging to the stem group of Chelicerata (Willset al 1998 Chen et al 2004 Cotton and Braddy 2004) or tothe stem group of all extant arthropods (Hou and Bergstroumlm1997 Budd 2002 Daley et al 2009) Megacheira composedat least of the Cambrian taxa Leanchoiliida Yohoiida andFortiforcipida has been depicted as a clade (Hou and Bergminusstroumlm 1997 fig 87) as a polyphyletic group in differentparts of the euarthropod stem group (Bergstroumlm and Hou2005) or as an unresolved polytomy in the chelicerate stemgroup (Chen et al 2004 fig 6) When conceived as a cladethe group is united by the ldquogreat appendagerdquo having a fewelongate spinose projections the exopods being fringed withsetae and a large telson (Hou and Bergstroumlm 1997) The exminustreme ends of the spectrum on the phylogenetic position ofMegacheira involve whether the sominuscalled ldquoshort great apminuspendagerdquo (Chen et al 2004) is a precursor to the chelicera(Bousfeld 1995 Chen et al 2004 Cotton and Braddy 2004
Bitsch and Bitsch 2007) or is the appendage of a segment thatis anatomically anterior to the chelicera or antenna (Budd2002)
The anatomically best understood megacheirans belongto the Order Leanchoiliida Stoslashrmer 1944 of which the genminusera Leanchoilia Walcott 1912 and Alalcomenaeus Simominusnetta 1970 provide most available information Alalcomeminusnaeus cambricus Simonetta 1970 has been revised based onBurgess Shale collections made through the 1980s and 1990s(Briggs and Collins 1999) Alalcomenaeus is apparently alsorepresented in the Chengjiang fauna based on A elegans(Luo and Hu in Luo et al 1999) (see Liu et al 2007) Themore common Leanchoilia has received even more studyThe anatomy and taxonomy of Leanchoilia from both theBurgess Shale (GarciacuteaminusBellido and Collins 2007) andChengjiang Lagerstaumltten (Liu et al 2007) have recently beenreviewed the former updating descriptions by Bruton andWhittington (1983) Two species in the Burgess Shale (Leanminuschoilia superlata Walcott 1912 and Leanchoilia persephoneSimonetta 1970) and a single species from Chengjiang(Leanchoilia illecebrosa [Hou 1987]) have been redescribedbased on extensive new collections Leanchoilia illecebrosasubsumes several generic and specific names introduced byLuo et al (1997) for Chengjiang material (see Liu et al 2007
doi104202app20100080Acta Palaeontol Pol 56 (2) 385ndash400 2011
for synonymy) Leanchoilia protogonia Simonetta 1970from the Burgess Shale has been reillustrated and accepted asa valid species (Briggs et al 2008) In addition to its BurgessShale and Chengjiang species Leanchoilia has also been reminusported from the Spence Shale of Utah (Leanchoilia superminuslata [Briggs et al 2008]) Leanchoilia hanceyi Briggs andRobison 1984] and the Kaili Formation Guizhou China(Leanchoilia sp of Zhao et al 2005)
Herein we add new diversity and biogeographic data forLeanchoiliidae by documenting a new genus and speciesfrom the Emu Bay Shale Kangaroo Island South AustraliaEarlier collections made from the Emu Bay Shalemdashformingthe basis of previous studies (see Paterson and Jago 2006Paterson et al 2008 for an overview)mdashcame from the cliffand waveminuscut platform exposures outcropping on the shoreminusline immediately east of the mouth of Big Gully on the northminuseast coast of Kangaroo Island (Fig 1) The new collectionsdescribed herein come from a new inland site Buck Quarrythe excavation of which began in 2007 The locality and eleminusments of the arthropod fauna are documented by Paterson etal (2008) GarciacuteaminusBellido et al (2009) and Paterson et al(2010) The Emu Bay Shale correlates with the lower Camminusbrian (Series 2) Pararaia janeae Zone of the South Austraminuslian biostratigraphic scheme (Jago et al 2006) equivalent tothe earlyminusmid Canglangpuan Stage of China and the midndashlateBotoman of Siberia (Paterson and Brock 2007 Paterson et al2008)
Institutional abbreviationmdashSAM P South Australian Muminusseum Adelaide Australia palaeontological collections
Other abbreviationsmdashcn1ndash2 cephalic endopods cs ceminusphalic shield cx1ndash2 cephalic exopods eo excretory organga great appendage gaf great appendage flagellum gappeduncle of great appendage L prefix for left mg midgutglands ol overlap between adjacent trunk tergites R prefixfor right t1ndasht11 tergites of trunk segments 1ndash11 tn trunkendopods tr reinforcing ridges on telson ts telson marginalspines tx trunk exopods txm trunk exopod lobe marginsvm ventral margin of tergites
Material and methodsSpecimen photography used a Canon EOS 5D digital camerawith a Canon MPminusE 65 mm 1ndash5times macro lens and low angle fiminusbre optic directional lighting from the top left (unless otherminuswise stated see Figure captions) Original colour images wereconverted to 8minusbit grey scale Camera lucida drawings weremade using a Leica MZ6 microscope at times16 magnification
In the phylogenetic analysis data were analysed using theparsimony program TNT (Goloboff et al 2008) with an exminusact search (implicit enumeration) Character optimisationwas explored with WINCLADA (Nixon 2002) Multistatecharacters were treated as unordered Characters that havepartial uncertainty (of the form ldquoeither 0 or 1 but not 2rdquo) weretreated as either of the two possible alternatives (as impleminus
mented by NONA and PeeWee) Jackknife resampling (Farminusris et al 1996) and Bremer support (Bremer 1994) were usedas measures of nodal support Jackknifing used 1000 repliminuscates with 36 deletion each replicate a heuristic search inminusvolving 1000 random stepwise addition sequences and TBRbranch swapping Bremer support was calculated from colminuslections of suboptimal trees obtained by exact searches Imminusplied weighting (Goloboff 1993) with TNT used the defaultconcavity constant of k = 3 followed by tree search with imminusplicit enumeration
Systematic palaeontology
Phylum Arthropoda Siebold and Stannius 1845Class Megacheira Hou and Bergstroumlm 1997Order Leanchoiliida Stoslashrmer 1944Family Leanchoiliidae Raymond 1935Genus Oestokerkus novType species Oestokerkus megacholix sp nov see below
Etymology From Greek oiumlstos arrow and kerkus tail with reference tothe forked shape of the telson
DiagnosismdashLeanchoiliid with long cephalic shield morethan oneminusthird length of trunk excluding telson lateral eyerelatively large paratergal folds long maximum height ofthorax nearly 40 of its length midgut glands large twopairs in head as well developed as those in trunk exopodsfringed by long setae telson forked widening distally with afew dorsally curved spines at posterior angle
386 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Fig 1 Geology of the area near Buck Quarry north east coast of KangarooIsland South Australia (modified from GarciacuteaminusBellido et al 2009)
DiscussionmdashMembership of Oestokerkus in Leanchoiliidaesensu Liu et al (2007) a group that includes the formerly reminuscognised families Alalcomenaeidae Simonetta and DelleCave 1975 and Actaeidae Simonetta and Delle Cave 1975 isclearly demonstrated by the morphology of the frontal apminuspendage which has greatly elongated spinose projections deminusveloped on its distal two articles and the movable finger eachprojection terminating at the same length and bearing a longflagellum presence of 11 trunk segments anteroventrallyminuspositioned eyes pendant appendages that have a similar biraminusmous structure in the cephalon and trunk and spines fringingthe margin of a long telson
The distinction between Alalcomenaeus and Leanchoiliahas most recently been reviewed by Liu et al (2007) whoemphasised the shape of the telson as a diagnostic characterthat of Leanchoilia being pointed and that of Alalcomenaeusrounded The telson of Oestokerkus megacholix described beminuslow differs substantially from the other species of both genminusera The telson widens distally in the holotype of O megaminuscholix (Figs 2 3A2 B) and is inferred to be forked Marginalspines are confined to the posterior edge rather than being deminusveloped along the lateral margins of a lanceolate telson as inall species of Leanchoilia or a paddleminusshaped posteriorlyrounded telson in Alalcomenaeus To date specimens of Omegacholix are preserved only in lateral aspect most likelydue to gravitational settling of the body on its sidemdasha comminusmon burial position of other leanchoiliids (Zhang and Hou2007)mdashbecause of the deep dorsominusventral versus transverselynarrow proportions of the body thus our interpretation of thetelson (see description below) has not yet been confirmed indorsal aspect Phylogenetic analysis (see below) resolves Omegacholix as sister to the widely accepted grouping ofLeanchoilia so we have elected to erect a new genus for itrather than to widen the scope of Leanchoilia and be forced torediagnose that genus using characters other than telson shapeThis taxonomic framework is consistent with the generic conminuscepts of Liu et al (2007)
In addition to telson form Oestokerkus is further distinminusguished from other leanchoiliid genera by relative proporminustions of the cephalic shield and trunk pleurotergites The ceminusphalic shield of O megacholix occupies a greater proporminustion of body length than in Leanchoilia or Alalcomenaeusthe head being ca 36 the length of the trunk (excluding thetelson) compared with 28 in Alalcomenaeus cambricus25 in Leanchoilia illecebrosa 23 in Leanchoilia perseminusphone and 26 in Leanchoilia superlata The paratergalfold in O megacholix is also considerably deeper than inother leanchoiliids The height to length ratio of ca 39 inO megacholix (Fig 2) contrasts with ca 19ndash22 in thethree wellminusknown Burgess Shale and Chengjiang species ofLeanchoilia (eg Liu et al 2007 fig 2D E) as well as Acambricus From what is observed in other fossils from thislocalitymdashand is also the case in the Burgess Shale Leanminuschoiliamdashdespite the fossils being compressed flat there islittle distortion Therefore the dimensions discussed aboveare not considered to deviate in any significant way from
those the animals would have had in life (Fig 7) The largemidgut glands of O megacholix are approached in size bysome species of Leanchoilia notably L superlata (GarciacuteaminusBellido and Collins 2007 pl 2 5) in which their structureof radiating grooves is the same The position and size ofthe lateral eye in O megacholix resemble those of Acambricus in which laterally preserved specimens showthe eye to be bulging at the anterolateral margin of the ceminusphalic shield (Briggs and Collins 1999 pl 2 4 pl 6 3) butpreservation in dorsoventral aspect shows that the lateralpair of eyes to be adaxial to the cephalic margin (Briggs andCollins 1999 pl 1 1) No evidence is available to indicatewhether median eyes (either two as in Leanchoilia or threeas in Alalcomenaeus) are present in Oestokerkus becausethis requires a dorsoventral aspect though the resolution ofOestokerkus within Leanchoiliidae predicts the presence ofthis feature
We have not coded the number of cephalic appendages inO megacholix in the cladistic analysis but the only specimenthat preserves the endopods (see species description) is mostconsistent with two biramous cephalic appendages as hasbeen documented in A cambricus (Briggs and Collins 1999)and in both L superlata and L persephone (GarciacuteaminusBellidoand Collins 2007) and contrasting with three pairs of biraminusmous cephalic appendages in L illecebrosa (Chen et al 2004Liu et al 2007) We reject the opinion of Chen et al (2004)that Briggs and Collins (1999) probably miscounted forAlalcomenaeus because more recent observations on BurgessShale Leanchoilia show that two pairs are indeed present insome members of Leanchoiliidae (GarciacuteaminusBellido and Collins2007) The variability in this feature in a narrowlyminuscircumminusscribed monophyletic group (ie Leanchoilia) indicates that itis of fairly minor taxonomic weight within this family
Oestokerkus megacholix sp novFigs 2ndash7
Etymology From Greek mega large and cholix gut in reference to thevoluminous midgut glands
Type material Holotype SAM P43631 a b part and counterpart preminusserved in lateral aspect (Figs 2 3) from the Emu Bay Shale BuckQuarry preserving cephalon including spinose projections and flagellaof great appendages all 11 trunk segments and most of telson includminusing a few marginal spines midgut glands exposed in head and anteriortwominusthirds of trunk some exopod setae of cephalic appendages as wellas lobes and setae of those associated with entire trunk Paratypes Allpreserved in lateral aspect SAM P45167 (Fig 4) cephalon exposingeye and peduncle of great appendage most of trunk with exopod setaeand a few endopods SAM P43630ab (Fig 5B) part and counterpartpreserving cephalic shield spinose projections of great appendage andanterior eight trunk segments two midgut glands and exopod setae ofcephalic and trunk appendages exposed SAM P15531 (Fig 5A) partonly preserving cephalon including spinose projections of great apminuspendage and most of trunk with midgut glands SAM P15438 (unfigminusured) incomplete cephalic shield with parts of great appendage comminusplete trunk and apparently proximal part of telson SAM P45168 (Fig6) part only cephalon with partial great appendages most of trunkwith midgut glands and exopod setae SAM P45169a b (unfigured)cephalon with partial great appendages trunk with midgut glands SAMP45177a b SAM P45178a b (unfigured) two weathered specimens
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 387
388 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
55 mmmm
5 mm
anus
telsontr
tr
t11
t5
mg
txm
t1
cs
gap
vm
ga
gaf
txts
cx
Fig 2 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov Holotype SAM P43631a A Photograph B Interpretative cameralucida drawing Arrows indicate tips of trunk exopod setae Abbreviations cs cephalic shield cx cephalic exopods ga great appendage gaf great appendminusage flagellum gap peduncle of great appendage mg midgut glands t1 tergite of trunk segment 1 tr reinforcing ridges on telson ts telson marginalspines tx trunk exopods txm trunk exopod lobe margins vm ventral margin of tergites Boundaries between trunk tergites indicated by dashed linesabove dorsal margin
Fig 3 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov Holotype SAM P43631 A P43631a A1 Midgut glands and tergiteboundaries in dorsal part of trunk Arrows indicate furrows that may represent position of dorsal transverse tendons A2 Telson light from NE A3 Spinoseprojections and flagella of distal articles of frontal appendage and exopod setae of cephalic and anterior trunk appendages and posterior margins of lobes ofthe latter B P43631b telson light from NE Arrows indicate tips of exopod setae Abbreviations gaf great appendage flagellum mg midgut glands oloverlap between adjacent trunk tergites tr reinforcing ridges on telson ts telson marginal spines txm trunk exopod lobe margins vm ventral margin oftergites
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 389
SAM P45904a b (unfigured) poorlyminuspreserved cephalic shield trunkwith wellminuspreserved midgut glands proximal part of telson SAMP45905ab (unfigured) cephalic shield lacking great appendages butexposing an eye trunk with incomplete telson SAM P45906a b (unfigminusured) great appendages with spinose projections oriented anteriorlytrunk preserved to end of midgut glands SAM P45908a b (unfigured)cephalic shield great appendages with flagella most of trunk (telsonmissing) showing midgut glands and exopod setae SAM P45909 (unminusfigured) small specimen with great appendages and series of mineralminusised midgut glands SAM P45910a b (unfigured) incomplete great apminuspendages oriented anteriorly trunk with complete series of midgutglands but indistinct appendages
Type locality Buck Quarry Big Gully (Fig 1)
Type horizon Emu Bay Shale (Cambrian Series 2 Stage 4)
DiagnosismdashAs for genus
DescriptionmdashCephalic shield 36 length of the trunk sagitminustally dorsal margin of cephalic shield gently convex in lateralview anteromedian margin inferred to be evenly rounded(based on comparison with lateral versus dorsal preservation ofBurgess Shale Leanchoilia and Alalcomenaeus) lacking an upminusturned median process Eye visible in SAM P54167 (Fig 4A)and P45905 relatively large (20 sagittal length of cephalon)exposed under anterolateral part of cephalic shield (Fig 4A2)
390 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Fig 4 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov SAM P45167 A P45167a A1 cephalic shield and most of trunkA2 detail of cephalic shield B P45167b A2 and B light from NE Overlap between adjacent tergites offset on left (Lol) and right (Rol) sides of bodyBoundaries between cephalic shield and anterior trunk tergites delimited in B by dashed lines Arrows mark positions of inferred dorsal transverse tendonsAbbreviations cn1ndash2 cephalic endopods cs cephalic shield gap peduncle of great appendage ol overlap between adjacent trunk tergites t1ndasht3 tergitesof trunk segments 1ndash3 tn trunk endopods
Fig 5 Leanchoiliid megacheiran arthropod Oestokerkus megacholix genet sp nov A SAM P 15531 B SAM P43630b Light from NE Abbreviaminustions cs cephalic shield ga great appendage gap peduncle of great apminuspendage mg midgut glands ol overlap between adjacent trunk tergites txtrunk exopods
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 391
Great appendage robust proportions of peduncle and disminustal spineminusbearing articles as in other members of Leanchoiliminusidae more commonly preserved with posterior orientation
but anteriorly oriented in SAM P45906 and SAM P45910Each of the three long spinose projections terminating at thesame length the projections evenly tapering distally with
392 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Fig 6 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov SAM P45168 A head and anterior part of trunk B Detail of spinoseprojections of great appendages and excretory organs (sensu Bruton and Whittington 1983) Abbreviations cs cephalic shield eo excretory organ gagreat appendage mg midgut glands tx trunk exopods
margins straight for much of their length or slightly curvedterminally distal spinose projection distinctly stouter at itsbase than the two more proximal projections and more markminusedly tapering along its length Dark irregularminusshaped low reminuslief lump on basal spineminusbearing article of great appendagesof SAM P45168 (eo in Fig 6) and SAM P45908 correminussponding to excretory organ of Bruton and Whittington(1983) Long flagella extending from each of the threespinose projections seen only in SAM P43631 (Figs 23A3) of similar narrow width along its preserved extent anminusnulations indistinguishable
Trunk of 11 segments of even length Maximum height oftrunk relative to length (excluding telson) ca 39 bodydeepest at tergite 4 Trunk tergite boundaries marked as narminusrow ridges in several specimens corresponding to narrowband of overlap between adjacent tergites (ol in Figs 2 3A14A1 B) Some specimens show tergites to be bisected by anarrow but sharp furrow of uncertain identity (eg possiblyimpression of dorsal transverse tendon cf Stein 2010 fig12B C E) (arrows in Figs 3A1 4B)
Exopods with fringe of long setae on cephalic and trunkappendages (Fig 2) setal fringe gradually shortening fromabout the fourth trunk segment Setae project ventrally bymore than 30 height of tergites in deepest part of trunk(Fig 2B) setae flattened narrow and parallelminussided alongmost of their length tapering near their distal ends to termiminusnate as a pointed tip (Figs 2B 3A3 arrows) Endopods exminusposed only in SAM P45167 (Fig 4) two pairs associatedwith cephalon (Fig 4B) those of anterior segments of trunkof similar morphology to those in cephalon all slender lackminusing obvious endites podomere boundaries unrecognisable
Midgut glands occupy a large extent of the dorsoventralheight of the trunk two midgut glands in cephalon of similarsize to those in each of the first seven segments of the trunk(Figs 3A1 5A 6A) all developed as radiating grooves emaminusnating from a globular structure at their ventral edge as in
Leanchoilia superlata (GarciacuteaminusBellido and Collins 2007 pl2 6) Foregut trace in SAM P43631 (Fig 2) indicates that gutcurves ventrally in anterior part of cephalon mouth regionnot preserved Terminal part of hindgut (in trunk segment 11)with closely spaced longitudinal lineations (Fig 2) that apminusparently correspond to folds in the gut wall hindgut termiminusnating at base of telson
Telson most completely preserved in holotype (SAMP43631 Figs 2B 3A2 B) In this specimen the telson likerest of body compressed in lateral aspect Telson shows meminusdian and marginal reinforcing ridges connecting a flat trianminusgular surface this surface appears to be partially broken inSAM P43631 showing underlying surface of other (left) halfof telson Lateral preservation of specimen implies that topcorresponds to an axial structure and bottom to a marginalone 30ndash35 angle in posterior edge of vane connecting thesetwo structures requires that in dorsal aspect telson would apminuspear forked and widening distally Five stout pointed dorminussally curved marginal spines attached to outer posterior edgeon each side of telson
Phylogenetic analysis ofLeanchoiliidaeDetermining the systematic position of Oestokerkus demandsan appraisal of the relationships of Leanchoiliidae as a wholeCladistic analysis including the bestminusknown members ofMegacheira including leanchoiliids has been conducted byCotton and Braddy (2004) and Vannier et al (2009) The taxominusnomic sampling used by Cotton and Braddy (2004) includedseveral megacheiran genera representative chelicerates anumber of taxa representing groups that have been proposedas possible chelicerate relatives (eg Sidneyia AglaspisCheloniellon) in other analyses (Dunlop and Selden 1997Hou and Bergstroumlm 1997) and members of the major lineagesof the Artiopoda sensu Hou and Bergstroumlm (1997) ie trilominusbiteminusallied lamellipedians Because artiopodan interminusrelationminusships have remained quite stable through a series of phylogenminusetic analyses (Edgecombe and Ramskoumlld 1999 Cotton andBraddy 2004 Hendricks and Lieberman 2008 Paterson et al2010) and artiopodans are now often regarded as less closelyrelated to Megacheira than is Chelicerata (Chen et al 2004Cotton and Braddy 2004 Dunlop 2006) we have reduced thesampling of Artiopoda in the present analysis To augment thediversity of ldquoshort great appendagerdquo arthropods Haikoucarisis added based on the description of H ercaiensis by Chen etal (2004) Offacolus (Orr et al 2000 Sutton et al 2002) hasbeen allied to Chelicerata and is included as a terminal Beminuscause the present analysis is principally concerned with relaminustionships of and within Leanchoiliidae sensu Liu et al (2007)we follow Vannier et al (2009) in sampling Alalcomenaeusand Leanchoilia at the species level Within Chelicerata Cotminuston and Braddy (2004) and Hendricks and Lieberman (2008)sampled ancestral states in Xiphosura by coding the Devonian
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 393
Fig 7 Reconstruction of Oestokerkus megacholix gen et sp nov in lateralview Total length and annulations on the flagella and positions of podominusmere boundaries of the frontal appendage are based on Leanchoilia superminuslata (Bruton and Whittington 1983)
synziphosurine Weinbergina Our coding for Xiphosura alsoemploys information from extant xiphosurans because theysupply more reliable information on the homology of the partsof the trunk appendages (Suzuki and Bergstroumlm 2008) and eyeanatomy (Harzsch and Hafner 2006) among other charactersystems than do fossils alone
Our character set extracts characters from the matrix ofCotton and Braddy (2004) that vary in the revised taxonomicsample reformulates some characters to accommodate reminusvised homology concepts and adds new characters especiallyfor leanchoiliids In total 32 characters are included (Table 1)In this analysis the primary homology of the ldquoshort great apminuspendagerdquo and chelicera is coded (as described by character 4)and the taxon sampling broadly constrains megacheirans aschelicerate relatives (as defended by Chen et al 2004 Cottonand Braddy 2004) rather than testing the alternative that theycould be stem group Euarthropoda (Budd 2002 Bergstroumlmand Hou 2005) A test of the two alternatives involves samminuspling of the entire diversity of euarthropods and a range ofnonminuseuarthropod outgroups an exercise beyond the scope ofthis study
Characters1 Mineralised cuticle (0) absent (1) present For definiminus
tion and discussion of this character see Cotton andBraddy (2004 character 30) and Paterson et al (2010character 1)
2 Curvature of anterior margin of cephalic shield (0) rounminusded or straight (1) pointed medially (2) with multiplespinose projections This character codes for variability inthe shape of the cephalic shield in Leanchoiliidae (state 0and 1 sensu GarciacuteaminusBellido and Collins [2007 textminusfigs10 and 9 respectively]) State 2 is added to accommodateOffacolus the median spine of a series that fringes the ceminusphalic margin is not convincingly homologised with themedian projection in leanchoiliids
3 Number of cephalic somitesappendages (0) antenna orfrontal appendage only (1) antenna or frontal appendageand two limb pairs (2) antenna or frontal appendage andthree limb pairs (3) antenna or frontal appendage and fourlimb pairs (4) antenna or frontal appendage and five limbpairs Coding for lamellipedians and Aglaspis is as discussedby Paterson et al (2010 character 2) Coding for Alalcomeminusnaeus cambricus (state 1) follows Briggs and Collins (1999)Leanchoilia illecebrosa (state 2) follows Liu et al (2007)and Leanchoilia persephone and Leanchoilia superlata (state1) follows GarciacuteaminusBellido and Collins (2007)
4 Form of first cephalic appendage (0) antenna (1) anteminusriorly directed raptorial appendage with reduced numberof podomeres each of the terminal podomeres bearing aspinose process on its distal margins a geniculation beminustween spinose podomeres and peduncle This characteris formulated to identify a homology between the frontalappendages of megacheirans and the chelicera of Cheliminus
cerata and state 1 corresponds to the putative apomorminusphic detail defended by Chen et al (2004) and Cottonand Braddy (2004) Vannier et al (2009 character 8)specify another shared property of ldquoshort great appendminusagesrdquo of the megacheiran (ss) type in their characterstate ldquobent at an anglerdquo referring to the geniculation beminustween articles of the peduncle or between the peduncleand distal spiniferous articles This property is sharedwith Chelicerata
5 Number of spineminusbearing articles on frontal appendage (0)three + movable finger (1) two + movable finger (2) one+ movable finger The number of spineminusbearing articles onthe frontal appendage (exclusive of the terminal movablefinger) varies from three in Yohoia (Whittington 1974 pl6 1ndash3 pl 7 3ndash6 pl 9 4ndash6) and Fortiforceps (Hou andBergstroumlm 1997 figs 31C D 32A 33B C E) to two inHaikoucaris (Chen et al 2004 fig 2A B) and Leanminuschoiliidae or only one the last being a chelicera
6 Long spinose projections on distal part of terminal threepodomeres of frontal appendage bearing a flagellum (0)absent (1) present The morphology of the great appendminusage in Leanchoiliidae is coded here We have not codedthe length of the spinose projections and the presence ofa flagellum as two independent characters Cotton andBraddyrsquos (2004) character 6 state 2 (ldquospinesrdquo longer thanthe entire podomere series) and character 8 state 1 (presminusence of an annulated flagellum) are thus grouped as a sinminusgle trait here
394 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Table 1 Characters coded for phylogenetic analysis Polymorphismindicated by ldquo+rdquo partial uncertainty by states separated by ldquordquo Abbreviminusations A Alalcomenaeus L Leanchoilia O Oestokerkus
Squamacula 0000minus minus minus2minus0 0110 00000 10minus minus0 1000 0
Cindarella 0030minus minus minus000 0010 00000 10minus minus0 11121 00
Kuamaia 000minus minus minus001 0011 10000 10minus minus0 11121 00
Kwanyinaspis 00 001 00112 1000 1000 1121 00
Olenoides 1020minus minus minus100 0011 00000 10minus minus0 01121 00
Retifacies 0020minus minus minus000 0010 00000 11000 1020 00
Aglaspis 10(23)0minus minus minus100 00101 00010 01001 0 00
Sidneyia 0000minus minus minus000 01102 00110 01101 00020 00
Emeraldella 0040minus minus minus2minus 0 00103 00110 01001 001(1+2)1 00
A cambricus 00111 10010 00001 00000 01110 01110 10
L illecebrosa 01211 11010 00001 01001 01210 11110 10
L persephone 00111 10010 00001 00000 01210 11110 10
L superlata 01111 11010 00001 01001 01210 11110 10
L hanceyi 00 00 0001 0000 01210
O megacholix 0011 100 00001 00000 01110 1110 0
Jianfengia 0021 0minus000 00005 00000 01000 010(12)0 0
Fortiforceps 00210 0minus00 00004 00000 01100 01230 20
Yohoia 00210 0minus000 00003 00011 01110 00010 (01)1
Haikoucaris 00211 0minus00 00003 00000 01(12)0 1010 0
Cheloniellon 0040minus minus minus100 00100 10010 01300 0030 00
Offacolus 02412 0minus200 0000 00010 01000 13minus minus 01
Xiphosura 00412 0minus100 10000 0010 01000 000minus minus 01
Eurypterida 00412 0minus100 10002 00010 01000 00minus minus minus 01
7 Length of flagellate great appendage (0) flagella termiminusnating at midlength of trunk (1) flagella extending to endof body Coding is applicable to Leanchoiliidae onlydistinguishing some Leanchoilia species that resembleAlalcomenaeus in having relative short great appendages(state 0) versus the longer appendages in L superlata andL illecebrosa
8 Position of lateral facetted eyes (0) ventral (1) dorsalsessile (2) absent Variation in Lamellipedia and severalother taxa coded herein follows Paterson et al (2010character 3)
9 Four or five ventral eyes arranged in a subtransverseband across head shield (0) absent (1) present A bandof ventral eyes is shared by the best known species ofLeanchoiliidae The distinction between three medianeyes in Alalcomenaeus cambricus (Briggs and Collins1999) and two in Leanchoilia illecebrosa (Liu et al2007) and both Leanchoilia superlata and L persephone(GarciacuteaminusBellido and Collins 2007) is uninformative forthe taxon sample here
10 Bulge in exoskeleton accommodating dropminusshaped venminustral eyes (0) absent (1) present This character followsEdgecombe and Ramskoumlld (1999 character 6) and Patminuserson et al (2010 character 5)
11 Dorsal median eyes (0) absent (1) present Coding for anabsence of dorsal median eyes in fossil taxa considers theabsence of either paired ocelli or a median eye tubercleand the character is scored as a chelicerate autapomorphy(see Cotton and Braddy 2004 character 24)
12 Width of doublure on cephalic shield (0) narrow or modminuserately wide (1) entire ventral side of head covered byexpansion of doublure The interpretation of state 1 inSidneyia and Squamacula is discussed by Cotton andBraddy (2004) and Paterson et al (2010 characters 12and 29)
13 Mouth covered by hypostome (0) absent (1) present Noldquoshort great appendagerdquo arthropods have been shown topossess a hypostome and a reconstruction of a smalllabrum in Alalcomenaeus (Briggs and Collins 1999) wasadmittedly conjectural The absence of a conspicuoushypostome contrasts with its ubiquitous presence in laminusmellipedians and several putative chelicerateminusallied taxathat have antennae (such as aglaspidids and Emeralminusdella) Chelicerates have a small epistomeminuslabrum (Bitschand Bitsch 2007) but like Cotton and Braddy (2004character 27 state 4) we code a hypostomal sclerite aslacking Kwanyinaspis has a large hypostome similar tothat found in aglaspidids (photograph of the holotype ofKwanyinaspis maotianshanensis shown to us by JavierOrtegaminusHernaacutendez [personal communication 2010])
14 Tergite articulations (0) extensive overlap of tergites (1)edge to edge articulations This character follows Edgeminuscombe and Ramskoumlld (1999 character 18) and Patersonet al (2010 character 20)
15 Number of trunk segments (0) nine (1) 11 (2) 12 (3)13 (4) 20 (5) 21 or 22 Coding is confined to taxa that do
not possess a pygidium because the precise number ofsomites associated with pygidia can sometimes not bedetermined with confidence Codings for Xiphosura andEurypterida use opisthosomal somite numbers followingShultz (2007 character 95)
16 Exoskeletal boundaries of anterior trunk segments (0)boundaries transverse or reflexed posterolaterally (1) reminusflexed anterolaterally This character follows Edgecombeand Ramskoumlld (1999 character 19) and Paterson et al(2010 character 21)
17 Trunk tergites with serrate lateral margins (0) absent (1)present Liu et al (2007 fig 2C) documented a row ofmarginal spines on the tergites of Leanchoilia illeceminusbrosa a featured shared by Leanchoilia superlata (egGarciacuteaminusBellido and Collins 2007 pl 1 7) but not otherterminals coded here
18 Differentiation in width of trunk tergites (0) trunk withgradational narrowing posteriorly (1) abrupt narrowingat boundary between prothorax and opisthothorax Anabrupt contraction in width between the boundary ofthe prominus and opisthothorax is observed in Emeraldellaand Sidneyia Yohoia has been reconstructed as having apronounced break in the shape of the trunk tergites beminushind segment 10 (Whittington 1974 textminusfig 2) thepoint at which the paratergal folds lose their spinose tipsand appendages are lacking Specimens preserved indorsal aspect (Whittington 1974 pl 10) suggest that thewidth of the opisthothorax is rather less abruptly narminusrowed and we code Yohoia as state 0
19 Appendages of posterior trunk segments (0) all trunktergites bearing appendages (1) one or more posteriortrunk segments lacking appendages Coding correspondsto Cotton and Braddyrsquos (2004) character 43 ldquopostabdominusmen of segments lacking appendagesrdquo
20 Paired tergal carinae (0) absent (1) present Liu et al(2007 265 fig 2F) recorded the presence of tergalcarinae in Leanchoilia illecebrosa shared with L superminuslata (eg Bruton and Whittington 1983 pl 17 106GarciacuteaminusBellido and Collins 2007 pl 1 5)
21 Pygidium (0) absent (1) present Coding of the pygiminusdium in many terminals herein was discussed by Paterminusson et al (2010 character 22)
22 Telson (0) absent (1) present Following Hendricks andLieberman (2008) a telson is a postminussegmental terminalstructure that lacks appendages Dunlop and Selden (1997)considered a telson to be absent in Cheloniellon but weidentify the conical projection posterior to the ultimate legpair and cerci (Stuumlrmer and Bergstroumlm 1978 figs 2 3 1011) as being most consistent with an identity as a telson
23 Telson shape (0) styliform (1) flapminuslike (2) lanceolate(3) short conical The outline of the telson in Haikouminuscaris ercaiensis is incompletely known but the mostcomplete specimen (Chen 2004 fig 472) constrains it aseither state 1 or state 2 A partial uncertainty coding isthus employed A unique state for Cheloniellon (state 3)is based on the interpretation in character 22
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 395
24 Telson fringed with spines (0) spines absent (1) spinespresent Presence of telson spines was introduced as a charminusacter by Hendricks and Lieberman (2008 character 34)
25 Paired flaps lateral to telson (0) absent (1) present Thischaracter describes the postminusventral plates of several taxa(see Edgecombe and Ramskoumlld 1999 character 25) Cotminuston and Braddy (2004 character 54) raised the possibilitythat the cerci of Cheloniellon could be homologous withpostminusventral plates (see also Hou and Bergstroumlm 1997 93)because they are similarly paired structures derived fromthe segment immediately anterior to the telson We havenot coded for this homology because the cerci of Chelominusniellon originate dorsally rather than ventrally
26 Midgut glands with submillimetric lamellae (0) absent(1) present Midgut glands in some stem group arthrominuspods and early crown group arthropods are metamericreniform organs with a characteristic lamellar structure(Butterfield 2002) Their morphology is identical inLeanchoilia and Oestokerkus notably including groovesthat radiate from a ventral median node (Figs 3A1 6A)The gut of Alalcomenaeus cambricus is more tubular(Briggs and Collins 1999 textminusfigs 10 11) and thoughan anterior six segments may display phosphatised latminuseral projections that likely represent gut diverticula (Wilminusson 2006) they have not been shown to have the detailedmorphology of Leanchoiliaminustype midgut glands Butterminusfield (2002 164) cited a figure from Bruton (1981 fig96) in support of ldquolamellarrdquo midgut glands in the cephaminuslon of Sidneyia Like Bruton (1981) in his description ofthe specimen we consider it likely that the structures inquestion are indeed gut diverticula but there is no basisfor identifying them more precisely as the reniformlamellar structures coded under this character
27 Exopod on first postminusantennal first postminusfrontal appendminusage segment (0) absent (1) present This character reminuslates to Cotton and Braddyrsquos (2004) character 3
28 Division of exopod (0) exopod an undivided flap (1)exopod differentiated into proximal and distal lobes (2)small flap with radiating lamellae (3) pediform exopoddivided into ca 6 articles Identification of the exopod inXiphosura follows Suzuki and Bergstroumlm (2008) Thischaracter groups conditions of the exopod shaft coded asindependent characters by Cotton and Braddy (2004)eg state 0 corresponds to their character 16 and state 1to their character 13 Our coding permits transformationbetween any two states without an extra implicit step
29 Exopod setae (0) short spiniform setae (1) long flatminustened tapering distally with slight separation (2)lamellar setae (flattened imbricating) (3) short finesetae A distinction is made between lamellipedian setaewhich are imbricated (state 2) and the pointed more disminustinctly separated and variably spiniform exopod setae ofleanchoiliids (state 1) This distinction is consistent withthe identification of megacheirans as nonminuslamellipedians(eg Hou and Bergstroumlm 1997 Bergstroumlm and Hou2005) Although Cotton and Braddy (2004 character 18)
regarded the form of the setae in Yohoia to be uncertainwe regard specimens figured by Whittington (1974 pl3 3) as conforming to the leanchoiliid state Squamaculadiffers from other taxa in its shorter spiniform setae(state 0) (Zhang et al 2004 textminusfig 1A B) Offacolushas longer spiniform projections on the distal articles ofthe exopod that have been variably identified as eitherspines or setae (Sutton et al 2002 fig 4b) but theirhomology with the setae that splay regularly along themargin of a flapminuslike exopod (eg states 0ndash3) is uncerminustain This and the following character for exopod setaeare left as ldquordquo for Offacolus A homology between thegill lamellae of Xiphosura and exopod setae of lamellipeminusdians has a long history (see Cotton and Braddy 2004character 18) We follow Suzuki and Bergstroumlm (2008)in identifying the xiphosuran gills as outgrowths of thebasis rather than of the exopod this perspective negates ahomology with the exopod setae of Lamellipedia Thepolymorphic coding for Emeraldella reflects the twoseparate states (1+2) displayed by the exopods in thetrunk (see Bruton and Whittington 1983 pl 8 45 47Edgecombe and Ramskoumlld 1999 fig 11)
30 Differentiation of setae along exopod (0) uniform orgradationally variable setae along length of exopod (1)lamellar setae confined to proximal lobe with distal lobebearing slender setae
31 Endopod segmentation (distal to coxa) (0) six or sevenpodomeres and terminal claw or telotarsal spurs (1)eight or more podomeres and terminal claw (2) ca 15podomeres State 1 is formulated to describe the segmenminustation of the leanchoiliid leg (GarciacuteaminusBellido and Collins2007 textminusfig 6 Liu et al 2007 fig 5) in contrast to thereduced number of podomeres in other euarthropods(Boxshall 2004) State 2 characterises Fortiforceps Apartial uncertainty coding is employed for Yohoia inwhich the precise number of podomeres is undeterminedand could be either state 0 or 1 (Whittington 1974 pl 74) but the spacing of the preserved podomere boundariesnegates the possibility of state 2
32 Endopods on anterior postminuscephalic segments (0) welldeveloped (1) rudimentary or lacking Following Cottonand Braddy (2004 character 11) the absence of trunkendopods in Yohoia (Whittington 1974) is coded as a puminustative homologue with the suppression of postminuscephalicendopods in Chelicerata
ResultsAs a convention for depicting the trees networks are rootedon Squamacula which separates artiopodan Lamellipedia(sensu Hou and Bergstroumlm 1997) from a group that includesMegacheira Chelicerata and Xenopoda all members ofwhich possess a telson
Under equal weights 14 shortest cladograms have a lengthof 77 steps with a Consistency Index of 064 and Retention Inminus
396 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
dex of 077 The strict consensus of these trees with supportvalues is shown in Fig 8 Leanchoiliidae is unambiguouslysupported in all 14 cladograms by four apomorphic characterstwo limbminusbearing cephalic segments posterior to the frontalappendage (character 3 state 1) long spinose projections onthe distal part of the terminal three podomeres of the frontalappendage that bear flagella (character 6 state 1) 11 trunksegments (character 15 state 1) and division of the exopodsinto two lobes (character 28 state 1) The arrangement of fouror five eyes in a subtransverse band across the cephalic shieldis unique to Leanchoiliidae (character 9 state 1) but is not anunambiguous autapomorphy of the group because the state isuncertain in close relatives notably Haikoucaris It is apominusmorphic for Leanchoiliidae only under particular optimisaminustions (Delayed Transformation)
The internal relationships of Leanchoiliidae are stableacross all 14 shortest cladograms Leanchoilia is united by thelanceolate telson (character 23 state 2) a unique unambiguminusous autapomorphy Within Leanchoilia a sister group relaminustionship between L superlata and L illecebrosa is well supminusported receiving a Bremer support of 3 and jackknife freminusquency of 86 These two species share four synapomorminusphies a medially pointed anterior cephalic margin (character2 state 1) flagella of the great appendage extending the lengthof the body (character 7 state 1) serrate lateral margins on thetrunk tergites (character 17 state 1) and paired tergal carinae(character 20 state 1) As such a species from Chengjiang (Lillecebrosa) and the Burgess Shale (L superlata) are moreclosely related to each other than are the two Burgess Shalecongeners a relationship that weakens the possibility that theoften cominusoccurring L superlata and L persephone could besexual dimorphs (GarciacuteaminusBellido and Collins 2007) ratherthan separate species The union of Leanchoilia and Oestominuskerkus is unambiguously supported by the shared midgutgland structure of both genera (character 26 state 1)
All shortest cladograms resolve Haikoucaris Yohoiaand Fortiforceps in that order as progressively less closelyrelated to Leanchoiliidae The union of these three taxa withLeanchoiliidae yields a clade that corresponds to Megacheirasensu Hou and Bergstroumlm (1997) apart from the ambiguousplacement of Jianfengia In half of the 14 shortest cladominusgrams Jianfengia is sister to the other megacheirans yieldminusing a monophyletic Megacheira ss In the remaining cladominusgrams Jianfengia is sister to Xenopoda sensu Hou andBergstroumlm (1997) Aglaspis and Chelicerata Haikoucarisand Leanchoiliidae are united by a frontal appendage comminusposed of two spineminusbearing articles in addition to the movminusable finger (character 5 state 1) Yohoia groups with thesetaxa based on spines fringing the telson (character 24 state1) and Fortiforceps is united with them at a broader levelbased on the flapminuslike telson shape (character 23 state 1)When Jianfengia is placed as sister to the other megacheiminusrans the entire clade is supported by the raptorial frontal apminuspendage (character 4 state 1) and the lack of a large hypominusstomal shield (character 13 state 0) In this set of cladominusgrams these features are nonminushomologous between Megaminus
cheira and Chelicerata whereas in some cladograms with thealternative resolution of Jianfengia as closer to cheliceratescharacter 4 is optimised as homologous in Megacheira andChelicerata
Reanalysis of the data with implied weights finds two optiminusmal trees (Fig 9) Relationships within Leanchoiliidae underequal weights are completely stable to reweighting as is theresolution of Haikoucaris as sister to Leanchoiliidae As suchthe character optimisations described above for the leanchoiminusliids are depicted the same in Fig 9 The main difference beminustween equal and implied weights involves the placement ofChelicerata Under equal weights (Fig 8) chelicerates are alminuslied to Xenopoda (here Emeraldella + Sidneyia + Chelominusniellon sensu clade L in Hou and Bergstroumlm 1997 fig 88)and Aglaspidida A clade composed of Aglaspidida Xenominuspoda and Chelicerata was similarly identified by Hou andBergstroumlm (1997 fig 88 their clade K) In our analysis thisgroup consistently finds support from one or more posteriortrunk segments lacking appendages (character 19 state 1) anddepending on alternative internal resolutions it is variably supminusported by dorsallyminuspositioned sessile eyes (character 8 state 1)and the absence of exopods on the first postminusantennalpostminuscheliceral limb (character 27 state 1) Under implied weightshowever chelicerates are more closely allied to Megacheirathan to aglaspidids or xenopodans (Fig 9)
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 397
2
582
76
Chelicera-morpha
Megacheira
62
2
53
50Leanchoiliidae
3
86
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
EurypteridaChelicerata
Fortiforceps
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia hanceyi
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
Fig 8 Strict consensus of 14 shortest cladograms (77 steps) based on charminusacter data in Table 1 Numbers above nodes are Bremer support those beminuslow nodes are jackknife frequencies gt50 Cheliceramorpha correspondsto a clade with the same composition in Cotton and Braddy (2004)
The major groupings in Fig 9 correspond closely to thoseidentified by Cotton and Braddy (2004) Notably the cladeuniting Aglaspis Xenopoda Offacolus Chelicerata Jianminusfengia and other Megacheira corresponds to their ldquoClade 3rdquo(Cotton and Braddy 2004 fig 8) for which they proposedthe name Cheliceramorpha The clade uniting CheloniellonAglaspis Sidneyia and Emeraldella in Fig 9 corresponds totheir ldquoClade 5rdquo and the clade that groups Megacheira andChelicerata is equivalent to their ldquoClade 4rdquo Their analysisdid not identify megacheirans as a monophyletic groupChelicerata was nested within the group and among taxaconsidered here Yohoia was resolved closer to cheliceratesthan to Leanchoiliidae In contrast Yohoia was regarded byBergstroumlm and Hou (2005) as more closely allied to Leanminuschoiliidae than to chelicerates a megacheiran grouping morecompatible with our cladograms The analysis of ldquogreat apminuspendage arthropodsrdquo by Vannier et al (2009) identified a
monophyletic Leanchoiliidae but a polyphyletic Megacheira(to the exclusion of Haikoucaris and Jianfengia) Howevertheir resolution of Fortiforceps and even more so Yohoia asthe closest relatives of Leanchoiliidae is as in our trees (apartfrom the different position of Haikoucaris)
In summary a major controversy in the systematics of theldquoshort great appendagerdquo arthropods remains whether thechelicerates are nested within the group or are instead moreclosely allied to taxa conventionally identified as lamelliminuspedians specifically Xenopoda and Aglaspidida We findthat both of these results are compatible with the availabledata depending on character weighting protocols Noneminustheless the broader megacheiranminuschelicerate issue does notcloud the status of leanchoiliids as a clade or the identificaminustion of their closest relatives (ie Haikoucaris and Yohoia)and the internal relationships of Leanchoiliidae are stableand supported by compelling characters
398 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
Eurypterida
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
2 7 17 20
3
2
1 1 1
26
1
281563
1 1 1 1
5
1
24
1
23 19 20 27 32
28Clade 4
1 0
4 13 11 27 0
154 1 1
3 19 32Clade 3
2 3
2 28
0
3 1 121
1
22
Clade 5
1 0
19 27
1
25
15 28 301 1
2312
1 150 1 3 3
15 16 2923
18
1 1
10 16
1 1
1 8
1
14
3
3
28 30
1 0
5
15
1 1
1
1
22
Fortiforceps
23
1 1 0 1
1 1
Leanchoilia hanceyi
2
2
1
1
Fig 9 Characters optimised on one of two shortest cladograms with imminusplied weights using a concavity constant k = 3 Black squares are nonminushomoplastic changes white squares homoplastic changes all changes areunambiguous Branches without strict support collapsed after optimisingcharacters Numbers for characters (above the branches) and states (belowthe branches) correspond to Table 1 ldquoClades 3ndash5rdquo correspond to groups sonumbered by Cotton and Braddy (2004 fig 8)
AcknowledgementsAccess to the field area has been most kindly allowed by the Buck famminusily We thank our collaborators in Emu Bay Shale research Jim Jago(University of South Australia Mawson Lakes) Jim Gehling andMike Lee (South Australian Museum Adelaide) for tremendous supminusport and advice as well as Mike Gemmell Dennis Rice Allison DaleyAaron Camens John Laurie Glenn Brock Pierre Kruse and Trevorand Jenny Worthy (all South Australian Museum) for their assistancein the field and Natalie Schroeder MaryminusAnne Binnie and RondaAtkinson (all South Australian Museum) for organisational support inthe lab and in the field Javier OrtegaminusHernaacutendez (University of Camminusbridge Cambridge UK) advised us on Kwanyinaspis and the journalrsquosreferees provided useful feedback TNT is available courtesy of theWilli Hennig Society Funding has been provided by an AustralianResearch Council Linkage grant (LP0774959) to the University ofAdelaide with generous financial assistance from Beach Energy Ltdand the South Australian Museum SeaLink provided logistical supminusport DGBrsquos travel to Australia has been supported by grants from theSpanish Research Council (CSIC PA 1001105) the Complutense Uniminusversity of Madrid and the Spanish Ministry of Education and Science(CGL2006minus12245BTE CGL2009minus07073 and RYC2007minus00090)
ReferencesBergstroumlm J and Hou XminusG 2005 Early Palaeozoic lamellipedian arthrominus
pods In S Koenemann and RA Jenner (eds) Crustacea and Arthrominuspod Relationships Crustacean Issues 16 73ndash93
Bitsch J and Bitsch C 2007 The segmental organization of the head reminusgion in Chelicerata a critical review of recent studies and hypothesesActa Zoologica 88 317ndash335
Bousfeld EL 1995 A contribution to the natural classification of Lower andMiddle Cambrian arthropods food gathering and feeding mechanismsAmphipacifica 2 3ndash34
Boxshall GA 2004 The evolution of arthropod limbs Biological Reviews79 253ndash300
Bremer K 1994 Branch support and tree stability Cladistics 10 295ndash304Briggs DEG and Collins D 1999 The arthropod Alalcomenaeus cambricus
Simonetta from the Middle Cambrian Burgess Shale of British ColumbiaPalaeontology 42 953ndash977
Briggs DEG and Robison RA 1984 Exceptionally preserved nonminustrilobite arthropods and anomalocaridids from the Middle Cambrian ofUtah University of Kansas Paleontological Contributions 111 1ndash23
Briggs DEG Lieberman BS Hendricks JR Halgedahl SL andJerrard RD 2008 Middle Cambrian arthropods from Utah Journal ofPaleontology 82 238ndash254
Bruton DL 1981 The arthropod Sidneyia inexpectans Middle CambrianBurgess Shale British Columbia Philosophical Transactions of theRoyal Society of London Series B 295 619ndash656
Bruton DL and Whittington HB 1983 Emeraldella and Leanchoiliatwo arthropods from the Burgess Shale Middle Cambrian British Cominuslumbia Philosophical Transactions of the Royal Society of London Seminusries B 300 553ndash582
Budd GE 2002 A palaeontological solution to the arthropod head probminuslem Nature 417 271ndash275
Butterfield NJ 2002 Leanchoilia guts and the interpretation of threeminusdiminusmensional structures in Burgess Shaleminustype fossils Paleobiology 28155ndash171
Chen JminusY 2004 The Dawn of Animal World [in Chinese] 366 pp JiangScience and Technology Press Nanjing
Chen JminusY Waloszek D and Maas A 2004 A new ldquogreatminusappendagerdquo arminusthropod from the Lower Cambrian of China and homology of cheliceratechelicerae and raptorial anterominusventral appendages Lethaia 37 3ndash20
Cotton TJ and Braddy SJ 2004 The phylogeny of arachnomorph arthrominuspods and the origin of the Chelicerata Transactions of the Royal Societyof Edinburgh Earth Sciences 94 169ndash193
Daley AC Budd GE Caron JminusB Edgecombe GD and Collins D2009 The Burgess Shale anomalocaridid Hurdia and its significancefor early euarthropod evolution Science 323 1597ndash1600
Dunlop JA 2006 New ideas about the euchelicerate stemminuslineage In CDeltshev and P Stoev (eds) European Arachnology 2005 Acta Zoominuslogica Bulgarica (Supplement 1) 9ndash27
Dunlop JA and Selden PA 1997 The early history and phylogeny of thechelicerates In RA Fortey and RH Thomas (eds) Arthropod Relaminustionships Systematics Association Special Volume 55 221ndash235
Edgecombe GD and Ramskoumlld L 1999 Relationships of CambrianArachnata and the systematic position of Trilobita Journal of Paleonminustology 73 263ndash287
Farris JS Albert VA Kaumlllersjouml M Lipcomb D and Kluge AG1996 Parsimony jackknifing outperforms neighborminusjoining Cladistics12 99ndash124
GarciacuteaminusBellido DC and Collins D 2007 Reassessment of the genusLeanchoilia (Arthropoda Arachnomorpha) from the Middle CambrianBurgess Shale British Columbia Canada Palaeontology 50 693ndash709
GarciacuteaminusBellido DC Paterson JR Edgecombe GD Jago JB GehlingJG and Lee MSY 2009 The bivalved arthropods Isoxys and Tuzoiawith softminuspart preservation from the lower Cambrian Emu Bay ShaleLagerstaumltte (Kangaroo Island Australia) Palaeontology 52 1221ndash1241
Goloboff PA 1993 Estimating character weights during tree searchCladistics 9 83ndash91
Goloboff PA Farris JS and Nixon KC 2008 TNT a free program forphylogenetic analysis Cladistics 24 774ndash786
Harzsch S and Hafner G 2006 Evolution of eye development in arthrominuspods phylogenetic aspects Arthropod Structure amp Development 35319ndash340
Hendricks JR and Lieberman BS 2008 New phylogenetic insights intothe Cambrian radiation of arachnomorph arthropods Journal of Paleminusontology 82 585ndash594
Hou XminusG and Bergstroumlm J 1997 Arthropods of the Lower CambrianChengjiang fauna southwest China Fossils and Strata 45 1ndash116
Jago JB Zang W Sun X Brock GA Paterson JR and SkovstedCB 2006 A review of the Cambrian biostratigraphy of South Austraminuslia Palaeoworld 15 406ndash423
Liu Y Hou XminusG and Bergstroumlm J 2007 Chengjiang arthropod Leanminuschoilia illecebrosa (Hou 1987) reconsidered GFF 129 263ndash272
Luo H Hu S Chen L Zhang S and Tao Y 1999 Early CambrianChengjiang fauna from Kunming region China [in Chinese] 129 ppYunnan Science and Technology Press Kunming
Luo H Hu S Zhang S and Tao Y 1997 New occurrence of the EarlyCambrian Chengjiang fauna in Haikou Kunming Yunnan Provinceand study on Trilobitoidea Acta Geologica Sinica 71 122ndash132
Nixon KC 2002 WINCLADA Version 19908 Cornell University IthacaNY
Orr PJ Siveter DJ Briggs DEG Siveter DJ and Sutton MD 2000A new arthropod from the Silurian KonservatminusLagerstaumltte of Hereminusfordshire Proceedings of the Royal Society Series B Biological Sciminusences 267 1497ndash1504
Paterson JR and Brock GA 2007 Early Cambrian trilobites fromAngorichina Flinders Ranges South Australia with a new assemblagefrom the Pararaia bunyerooensis Zone Journal of Paleontology 81116ndash142
Paterson JR and Jago JB 2006 New trilobites from the Lower CambrianEmu Bay Shale Lagerstaumltte at Big Gully Kangaroo Island South Ausminustralia Memoirs of the Association of Australasian Palaeontologists 3243ndash57
Paterson JR Edgecombe GD GarciacuteaminusBellido DC Jago JB andGehling JG 2010 Nektaspid arthropods from the lower CambrianEmu Bay Shale Lagerstaumltte South Australia with a reassessment oflamellipedian relationships Palaeontology 53 377ndash402
Paterson JR Jago JB Gehling JG GarciacuteaminusBellido DC Edgecombe
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 399
GD and Lee MSY 2008 Early Cambrian arthropods from the EmuBay Shale Lagerstaumltte South Australia 319ndash325 In I Raacutebano RGozalo and D GarciacuteaminusBellido (eds) Advances in Trilobite ResearchCuadernos del Museo Geominero 9 319ndash325
Shultz JW 2007 A phylogenetic analysis of the arachnid orders based onmorphological characters Zoological Journal of the Linnean Society150 221ndash265
Stein M 2010 A new arthropod from the Early Cambrian of North Greenminusland with a ldquogreat appendagerdquominuslike antennula Zoological Journal ofthe Linnean Society 158 477ndash500
Stuumlrmer W and Bergstroumlm J 1978 The arthropod Cheloniellon from theDevonian Hunsruumlck Shale Palaumlontologische Zeitschrift 52 57ndash81
Sutton MD Briggs DEG Siveter DJ Siveter DJ and Orr PD2002 The arthropod Offacolus kingi (Chelicerata) from the Silurian ofHerefordshire England computer based morphological reconstrucminustions and phylogenetic affinity Proceedings of the Royal Society B Biminusological Sciences 269 1195ndash1203
Suzuki Y and Bergstroumlm J 2008 Respiration in trilobites a reevaluationGFF 130 211ndash229
Vannier J GarciacuteaminusBellido DC Hu XminusX and Chen AminusL 2009 Arthrominuspod visual predators in the early pelagic ecosystem evidence from the
Burgess Shale and Chengjiang biotas Proceedings of the Royal SocietyB Biological Sciences 276 2567ndash2574
Whittington HB 1974 Yohoia Walcott and Plenocaris n gen arthropodsfrom the Burgess Shale Middle Cambrian British Columbia Geologiminuscal Survey of Canada Bulletin 231 1ndash21
Wills MA Briggs DEG Fortey RA Wilkinson M and SneathPHA 1998 An arthropod phylogeny based on fossil and recent taxaIn GD Edgecombe (ed) Arthropod Fossils and Phylogeny 33ndash105Columbia University Press New York
Wilson LA 2006 Food for thought a morphological and taphonomic studyof fossilised digestive systems from Early to Middle Cambrian taxa275 pp Unpublished PhD thesis University of Cambridge Cambridge
Zhang XminusG and Hou X 2007 Gravitational constraints on the burial ofChengjiang fossils Palaios 22 448ndash453
Zhang XminusL Han J Zhang Z Liu H and Shu D 2004 Redescription ofthe Chengjiang arthropod Squamacula clypeata Hou and Bergstroumlm fromthe Lower Cambrian Southminuswest China Palaeontology 47 605ndash617
Zhao Y Zhu M Babcock LE Yuan L Parsley RL Peng J YangX and Wang Y 2005 Kaili Biota a taphonomic window on diversifiminuscation of metazoans from the basal Middle Cambrian Guizhou ChinaActa Geologica Sinica 79 751ndash765
400 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011

for synonymy) Leanchoilia protogonia Simonetta 1970from the Burgess Shale has been reillustrated and accepted asa valid species (Briggs et al 2008) In addition to its BurgessShale and Chengjiang species Leanchoilia has also been reminusported from the Spence Shale of Utah (Leanchoilia superminuslata [Briggs et al 2008]) Leanchoilia hanceyi Briggs andRobison 1984] and the Kaili Formation Guizhou China(Leanchoilia sp of Zhao et al 2005)
Herein we add new diversity and biogeographic data forLeanchoiliidae by documenting a new genus and speciesfrom the Emu Bay Shale Kangaroo Island South AustraliaEarlier collections made from the Emu Bay Shalemdashformingthe basis of previous studies (see Paterson and Jago 2006Paterson et al 2008 for an overview)mdashcame from the cliffand waveminuscut platform exposures outcropping on the shoreminusline immediately east of the mouth of Big Gully on the northminuseast coast of Kangaroo Island (Fig 1) The new collectionsdescribed herein come from a new inland site Buck Quarrythe excavation of which began in 2007 The locality and eleminusments of the arthropod fauna are documented by Paterson etal (2008) GarciacuteaminusBellido et al (2009) and Paterson et al(2010) The Emu Bay Shale correlates with the lower Camminusbrian (Series 2) Pararaia janeae Zone of the South Austraminuslian biostratigraphic scheme (Jago et al 2006) equivalent tothe earlyminusmid Canglangpuan Stage of China and the midndashlateBotoman of Siberia (Paterson and Brock 2007 Paterson et al2008)
Institutional abbreviationmdashSAM P South Australian Muminusseum Adelaide Australia palaeontological collections
Other abbreviationsmdashcn1ndash2 cephalic endopods cs ceminusphalic shield cx1ndash2 cephalic exopods eo excretory organga great appendage gaf great appendage flagellum gappeduncle of great appendage L prefix for left mg midgutglands ol overlap between adjacent trunk tergites R prefixfor right t1ndasht11 tergites of trunk segments 1ndash11 tn trunkendopods tr reinforcing ridges on telson ts telson marginalspines tx trunk exopods txm trunk exopod lobe marginsvm ventral margin of tergites
Material and methodsSpecimen photography used a Canon EOS 5D digital camerawith a Canon MPminusE 65 mm 1ndash5times macro lens and low angle fiminusbre optic directional lighting from the top left (unless otherminuswise stated see Figure captions) Original colour images wereconverted to 8minusbit grey scale Camera lucida drawings weremade using a Leica MZ6 microscope at times16 magnification
In the phylogenetic analysis data were analysed using theparsimony program TNT (Goloboff et al 2008) with an exminusact search (implicit enumeration) Character optimisationwas explored with WINCLADA (Nixon 2002) Multistatecharacters were treated as unordered Characters that havepartial uncertainty (of the form ldquoeither 0 or 1 but not 2rdquo) weretreated as either of the two possible alternatives (as impleminus
mented by NONA and PeeWee) Jackknife resampling (Farminusris et al 1996) and Bremer support (Bremer 1994) were usedas measures of nodal support Jackknifing used 1000 repliminuscates with 36 deletion each replicate a heuristic search inminusvolving 1000 random stepwise addition sequences and TBRbranch swapping Bremer support was calculated from colminuslections of suboptimal trees obtained by exact searches Imminusplied weighting (Goloboff 1993) with TNT used the defaultconcavity constant of k = 3 followed by tree search with imminusplicit enumeration
Systematic palaeontology
Phylum Arthropoda Siebold and Stannius 1845Class Megacheira Hou and Bergstroumlm 1997Order Leanchoiliida Stoslashrmer 1944Family Leanchoiliidae Raymond 1935Genus Oestokerkus novType species Oestokerkus megacholix sp nov see below
Etymology From Greek oiumlstos arrow and kerkus tail with reference tothe forked shape of the telson
DiagnosismdashLeanchoiliid with long cephalic shield morethan oneminusthird length of trunk excluding telson lateral eyerelatively large paratergal folds long maximum height ofthorax nearly 40 of its length midgut glands large twopairs in head as well developed as those in trunk exopodsfringed by long setae telson forked widening distally with afew dorsally curved spines at posterior angle
386 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Fig 1 Geology of the area near Buck Quarry north east coast of KangarooIsland South Australia (modified from GarciacuteaminusBellido et al 2009)
DiscussionmdashMembership of Oestokerkus in Leanchoiliidaesensu Liu et al (2007) a group that includes the formerly reminuscognised families Alalcomenaeidae Simonetta and DelleCave 1975 and Actaeidae Simonetta and Delle Cave 1975 isclearly demonstrated by the morphology of the frontal apminuspendage which has greatly elongated spinose projections deminusveloped on its distal two articles and the movable finger eachprojection terminating at the same length and bearing a longflagellum presence of 11 trunk segments anteroventrallyminuspositioned eyes pendant appendages that have a similar biraminusmous structure in the cephalon and trunk and spines fringingthe margin of a long telson
The distinction between Alalcomenaeus and Leanchoiliahas most recently been reviewed by Liu et al (2007) whoemphasised the shape of the telson as a diagnostic characterthat of Leanchoilia being pointed and that of Alalcomenaeusrounded The telson of Oestokerkus megacholix described beminuslow differs substantially from the other species of both genminusera The telson widens distally in the holotype of O megaminuscholix (Figs 2 3A2 B) and is inferred to be forked Marginalspines are confined to the posterior edge rather than being deminusveloped along the lateral margins of a lanceolate telson as inall species of Leanchoilia or a paddleminusshaped posteriorlyrounded telson in Alalcomenaeus To date specimens of Omegacholix are preserved only in lateral aspect most likelydue to gravitational settling of the body on its sidemdasha comminusmon burial position of other leanchoiliids (Zhang and Hou2007)mdashbecause of the deep dorsominusventral versus transverselynarrow proportions of the body thus our interpretation of thetelson (see description below) has not yet been confirmed indorsal aspect Phylogenetic analysis (see below) resolves Omegacholix as sister to the widely accepted grouping ofLeanchoilia so we have elected to erect a new genus for itrather than to widen the scope of Leanchoilia and be forced torediagnose that genus using characters other than telson shapeThis taxonomic framework is consistent with the generic conminuscepts of Liu et al (2007)
In addition to telson form Oestokerkus is further distinminusguished from other leanchoiliid genera by relative proporminustions of the cephalic shield and trunk pleurotergites The ceminusphalic shield of O megacholix occupies a greater proporminustion of body length than in Leanchoilia or Alalcomenaeusthe head being ca 36 the length of the trunk (excluding thetelson) compared with 28 in Alalcomenaeus cambricus25 in Leanchoilia illecebrosa 23 in Leanchoilia perseminusphone and 26 in Leanchoilia superlata The paratergalfold in O megacholix is also considerably deeper than inother leanchoiliids The height to length ratio of ca 39 inO megacholix (Fig 2) contrasts with ca 19ndash22 in thethree wellminusknown Burgess Shale and Chengjiang species ofLeanchoilia (eg Liu et al 2007 fig 2D E) as well as Acambricus From what is observed in other fossils from thislocalitymdashand is also the case in the Burgess Shale Leanminuschoiliamdashdespite the fossils being compressed flat there islittle distortion Therefore the dimensions discussed aboveare not considered to deviate in any significant way from
those the animals would have had in life (Fig 7) The largemidgut glands of O megacholix are approached in size bysome species of Leanchoilia notably L superlata (GarciacuteaminusBellido and Collins 2007 pl 2 5) in which their structureof radiating grooves is the same The position and size ofthe lateral eye in O megacholix resemble those of Acambricus in which laterally preserved specimens showthe eye to be bulging at the anterolateral margin of the ceminusphalic shield (Briggs and Collins 1999 pl 2 4 pl 6 3) butpreservation in dorsoventral aspect shows that the lateralpair of eyes to be adaxial to the cephalic margin (Briggs andCollins 1999 pl 1 1) No evidence is available to indicatewhether median eyes (either two as in Leanchoilia or threeas in Alalcomenaeus) are present in Oestokerkus becausethis requires a dorsoventral aspect though the resolution ofOestokerkus within Leanchoiliidae predicts the presence ofthis feature
We have not coded the number of cephalic appendages inO megacholix in the cladistic analysis but the only specimenthat preserves the endopods (see species description) is mostconsistent with two biramous cephalic appendages as hasbeen documented in A cambricus (Briggs and Collins 1999)and in both L superlata and L persephone (GarciacuteaminusBellidoand Collins 2007) and contrasting with three pairs of biraminusmous cephalic appendages in L illecebrosa (Chen et al 2004Liu et al 2007) We reject the opinion of Chen et al (2004)that Briggs and Collins (1999) probably miscounted forAlalcomenaeus because more recent observations on BurgessShale Leanchoilia show that two pairs are indeed present insome members of Leanchoiliidae (GarciacuteaminusBellido and Collins2007) The variability in this feature in a narrowlyminuscircumminusscribed monophyletic group (ie Leanchoilia) indicates that itis of fairly minor taxonomic weight within this family
Oestokerkus megacholix sp novFigs 2ndash7
Etymology From Greek mega large and cholix gut in reference to thevoluminous midgut glands
Type material Holotype SAM P43631 a b part and counterpart preminusserved in lateral aspect (Figs 2 3) from the Emu Bay Shale BuckQuarry preserving cephalon including spinose projections and flagellaof great appendages all 11 trunk segments and most of telson includminusing a few marginal spines midgut glands exposed in head and anteriortwominusthirds of trunk some exopod setae of cephalic appendages as wellas lobes and setae of those associated with entire trunk Paratypes Allpreserved in lateral aspect SAM P45167 (Fig 4) cephalon exposingeye and peduncle of great appendage most of trunk with exopod setaeand a few endopods SAM P43630ab (Fig 5B) part and counterpartpreserving cephalic shield spinose projections of great appendage andanterior eight trunk segments two midgut glands and exopod setae ofcephalic and trunk appendages exposed SAM P15531 (Fig 5A) partonly preserving cephalon including spinose projections of great apminuspendage and most of trunk with midgut glands SAM P15438 (unfigminusured) incomplete cephalic shield with parts of great appendage comminusplete trunk and apparently proximal part of telson SAM P45168 (Fig6) part only cephalon with partial great appendages most of trunkwith midgut glands and exopod setae SAM P45169a b (unfigured)cephalon with partial great appendages trunk with midgut glands SAMP45177a b SAM P45178a b (unfigured) two weathered specimens
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 387
388 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
55 mmmm
5 mm
anus
telsontr
tr
t11
t5
mg
txm
t1
cs
gap
vm
ga
gaf
txts
cx
Fig 2 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov Holotype SAM P43631a A Photograph B Interpretative cameralucida drawing Arrows indicate tips of trunk exopod setae Abbreviations cs cephalic shield cx cephalic exopods ga great appendage gaf great appendminusage flagellum gap peduncle of great appendage mg midgut glands t1 tergite of trunk segment 1 tr reinforcing ridges on telson ts telson marginalspines tx trunk exopods txm trunk exopod lobe margins vm ventral margin of tergites Boundaries between trunk tergites indicated by dashed linesabove dorsal margin
Fig 3 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov Holotype SAM P43631 A P43631a A1 Midgut glands and tergiteboundaries in dorsal part of trunk Arrows indicate furrows that may represent position of dorsal transverse tendons A2 Telson light from NE A3 Spinoseprojections and flagella of distal articles of frontal appendage and exopod setae of cephalic and anterior trunk appendages and posterior margins of lobes ofthe latter B P43631b telson light from NE Arrows indicate tips of exopod setae Abbreviations gaf great appendage flagellum mg midgut glands oloverlap between adjacent trunk tergites tr reinforcing ridges on telson ts telson marginal spines txm trunk exopod lobe margins vm ventral margin oftergites
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 389
SAM P45904a b (unfigured) poorlyminuspreserved cephalic shield trunkwith wellminuspreserved midgut glands proximal part of telson SAMP45905ab (unfigured) cephalic shield lacking great appendages butexposing an eye trunk with incomplete telson SAM P45906a b (unfigminusured) great appendages with spinose projections oriented anteriorlytrunk preserved to end of midgut glands SAM P45908a b (unfigured)cephalic shield great appendages with flagella most of trunk (telsonmissing) showing midgut glands and exopod setae SAM P45909 (unminusfigured) small specimen with great appendages and series of mineralminusised midgut glands SAM P45910a b (unfigured) incomplete great apminuspendages oriented anteriorly trunk with complete series of midgutglands but indistinct appendages
Type locality Buck Quarry Big Gully (Fig 1)
Type horizon Emu Bay Shale (Cambrian Series 2 Stage 4)
DiagnosismdashAs for genus
DescriptionmdashCephalic shield 36 length of the trunk sagitminustally dorsal margin of cephalic shield gently convex in lateralview anteromedian margin inferred to be evenly rounded(based on comparison with lateral versus dorsal preservation ofBurgess Shale Leanchoilia and Alalcomenaeus) lacking an upminusturned median process Eye visible in SAM P54167 (Fig 4A)and P45905 relatively large (20 sagittal length of cephalon)exposed under anterolateral part of cephalic shield (Fig 4A2)
390 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Fig 4 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov SAM P45167 A P45167a A1 cephalic shield and most of trunkA2 detail of cephalic shield B P45167b A2 and B light from NE Overlap between adjacent tergites offset on left (Lol) and right (Rol) sides of bodyBoundaries between cephalic shield and anterior trunk tergites delimited in B by dashed lines Arrows mark positions of inferred dorsal transverse tendonsAbbreviations cn1ndash2 cephalic endopods cs cephalic shield gap peduncle of great appendage ol overlap between adjacent trunk tergites t1ndasht3 tergitesof trunk segments 1ndash3 tn trunk endopods
Fig 5 Leanchoiliid megacheiran arthropod Oestokerkus megacholix genet sp nov A SAM P 15531 B SAM P43630b Light from NE Abbreviaminustions cs cephalic shield ga great appendage gap peduncle of great apminuspendage mg midgut glands ol overlap between adjacent trunk tergites txtrunk exopods
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 391
Great appendage robust proportions of peduncle and disminustal spineminusbearing articles as in other members of Leanchoiliminusidae more commonly preserved with posterior orientation
but anteriorly oriented in SAM P45906 and SAM P45910Each of the three long spinose projections terminating at thesame length the projections evenly tapering distally with
392 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Fig 6 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov SAM P45168 A head and anterior part of trunk B Detail of spinoseprojections of great appendages and excretory organs (sensu Bruton and Whittington 1983) Abbreviations cs cephalic shield eo excretory organ gagreat appendage mg midgut glands tx trunk exopods
margins straight for much of their length or slightly curvedterminally distal spinose projection distinctly stouter at itsbase than the two more proximal projections and more markminusedly tapering along its length Dark irregularminusshaped low reminuslief lump on basal spineminusbearing article of great appendagesof SAM P45168 (eo in Fig 6) and SAM P45908 correminussponding to excretory organ of Bruton and Whittington(1983) Long flagella extending from each of the threespinose projections seen only in SAM P43631 (Figs 23A3) of similar narrow width along its preserved extent anminusnulations indistinguishable
Trunk of 11 segments of even length Maximum height oftrunk relative to length (excluding telson) ca 39 bodydeepest at tergite 4 Trunk tergite boundaries marked as narminusrow ridges in several specimens corresponding to narrowband of overlap between adjacent tergites (ol in Figs 2 3A14A1 B) Some specimens show tergites to be bisected by anarrow but sharp furrow of uncertain identity (eg possiblyimpression of dorsal transverse tendon cf Stein 2010 fig12B C E) (arrows in Figs 3A1 4B)
Exopods with fringe of long setae on cephalic and trunkappendages (Fig 2) setal fringe gradually shortening fromabout the fourth trunk segment Setae project ventrally bymore than 30 height of tergites in deepest part of trunk(Fig 2B) setae flattened narrow and parallelminussided alongmost of their length tapering near their distal ends to termiminusnate as a pointed tip (Figs 2B 3A3 arrows) Endopods exminusposed only in SAM P45167 (Fig 4) two pairs associatedwith cephalon (Fig 4B) those of anterior segments of trunkof similar morphology to those in cephalon all slender lackminusing obvious endites podomere boundaries unrecognisable
Midgut glands occupy a large extent of the dorsoventralheight of the trunk two midgut glands in cephalon of similarsize to those in each of the first seven segments of the trunk(Figs 3A1 5A 6A) all developed as radiating grooves emaminusnating from a globular structure at their ventral edge as in
Leanchoilia superlata (GarciacuteaminusBellido and Collins 2007 pl2 6) Foregut trace in SAM P43631 (Fig 2) indicates that gutcurves ventrally in anterior part of cephalon mouth regionnot preserved Terminal part of hindgut (in trunk segment 11)with closely spaced longitudinal lineations (Fig 2) that apminusparently correspond to folds in the gut wall hindgut termiminusnating at base of telson
Telson most completely preserved in holotype (SAMP43631 Figs 2B 3A2 B) In this specimen the telson likerest of body compressed in lateral aspect Telson shows meminusdian and marginal reinforcing ridges connecting a flat trianminusgular surface this surface appears to be partially broken inSAM P43631 showing underlying surface of other (left) halfof telson Lateral preservation of specimen implies that topcorresponds to an axial structure and bottom to a marginalone 30ndash35 angle in posterior edge of vane connecting thesetwo structures requires that in dorsal aspect telson would apminuspear forked and widening distally Five stout pointed dorminussally curved marginal spines attached to outer posterior edgeon each side of telson
Phylogenetic analysis ofLeanchoiliidaeDetermining the systematic position of Oestokerkus demandsan appraisal of the relationships of Leanchoiliidae as a wholeCladistic analysis including the bestminusknown members ofMegacheira including leanchoiliids has been conducted byCotton and Braddy (2004) and Vannier et al (2009) The taxominusnomic sampling used by Cotton and Braddy (2004) includedseveral megacheiran genera representative chelicerates anumber of taxa representing groups that have been proposedas possible chelicerate relatives (eg Sidneyia AglaspisCheloniellon) in other analyses (Dunlop and Selden 1997Hou and Bergstroumlm 1997) and members of the major lineagesof the Artiopoda sensu Hou and Bergstroumlm (1997) ie trilominusbiteminusallied lamellipedians Because artiopodan interminusrelationminusships have remained quite stable through a series of phylogenminusetic analyses (Edgecombe and Ramskoumlld 1999 Cotton andBraddy 2004 Hendricks and Lieberman 2008 Paterson et al2010) and artiopodans are now often regarded as less closelyrelated to Megacheira than is Chelicerata (Chen et al 2004Cotton and Braddy 2004 Dunlop 2006) we have reduced thesampling of Artiopoda in the present analysis To augment thediversity of ldquoshort great appendagerdquo arthropods Haikoucarisis added based on the description of H ercaiensis by Chen etal (2004) Offacolus (Orr et al 2000 Sutton et al 2002) hasbeen allied to Chelicerata and is included as a terminal Beminuscause the present analysis is principally concerned with relaminustionships of and within Leanchoiliidae sensu Liu et al (2007)we follow Vannier et al (2009) in sampling Alalcomenaeusand Leanchoilia at the species level Within Chelicerata Cotminuston and Braddy (2004) and Hendricks and Lieberman (2008)sampled ancestral states in Xiphosura by coding the Devonian
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 393
Fig 7 Reconstruction of Oestokerkus megacholix gen et sp nov in lateralview Total length and annulations on the flagella and positions of podominusmere boundaries of the frontal appendage are based on Leanchoilia superminuslata (Bruton and Whittington 1983)
synziphosurine Weinbergina Our coding for Xiphosura alsoemploys information from extant xiphosurans because theysupply more reliable information on the homology of the partsof the trunk appendages (Suzuki and Bergstroumlm 2008) and eyeanatomy (Harzsch and Hafner 2006) among other charactersystems than do fossils alone
Our character set extracts characters from the matrix ofCotton and Braddy (2004) that vary in the revised taxonomicsample reformulates some characters to accommodate reminusvised homology concepts and adds new characters especiallyfor leanchoiliids In total 32 characters are included (Table 1)In this analysis the primary homology of the ldquoshort great apminuspendagerdquo and chelicera is coded (as described by character 4)and the taxon sampling broadly constrains megacheirans aschelicerate relatives (as defended by Chen et al 2004 Cottonand Braddy 2004) rather than testing the alternative that theycould be stem group Euarthropoda (Budd 2002 Bergstroumlmand Hou 2005) A test of the two alternatives involves samminuspling of the entire diversity of euarthropods and a range ofnonminuseuarthropod outgroups an exercise beyond the scope ofthis study
Characters1 Mineralised cuticle (0) absent (1) present For definiminus
tion and discussion of this character see Cotton andBraddy (2004 character 30) and Paterson et al (2010character 1)
2 Curvature of anterior margin of cephalic shield (0) rounminusded or straight (1) pointed medially (2) with multiplespinose projections This character codes for variability inthe shape of the cephalic shield in Leanchoiliidae (state 0and 1 sensu GarciacuteaminusBellido and Collins [2007 textminusfigs10 and 9 respectively]) State 2 is added to accommodateOffacolus the median spine of a series that fringes the ceminusphalic margin is not convincingly homologised with themedian projection in leanchoiliids
3 Number of cephalic somitesappendages (0) antenna orfrontal appendage only (1) antenna or frontal appendageand two limb pairs (2) antenna or frontal appendage andthree limb pairs (3) antenna or frontal appendage and fourlimb pairs (4) antenna or frontal appendage and five limbpairs Coding for lamellipedians and Aglaspis is as discussedby Paterson et al (2010 character 2) Coding for Alalcomeminusnaeus cambricus (state 1) follows Briggs and Collins (1999)Leanchoilia illecebrosa (state 2) follows Liu et al (2007)and Leanchoilia persephone and Leanchoilia superlata (state1) follows GarciacuteaminusBellido and Collins (2007)
4 Form of first cephalic appendage (0) antenna (1) anteminusriorly directed raptorial appendage with reduced numberof podomeres each of the terminal podomeres bearing aspinose process on its distal margins a geniculation beminustween spinose podomeres and peduncle This characteris formulated to identify a homology between the frontalappendages of megacheirans and the chelicera of Cheliminus
cerata and state 1 corresponds to the putative apomorminusphic detail defended by Chen et al (2004) and Cottonand Braddy (2004) Vannier et al (2009 character 8)specify another shared property of ldquoshort great appendminusagesrdquo of the megacheiran (ss) type in their characterstate ldquobent at an anglerdquo referring to the geniculation beminustween articles of the peduncle or between the peduncleand distal spiniferous articles This property is sharedwith Chelicerata
5 Number of spineminusbearing articles on frontal appendage (0)three + movable finger (1) two + movable finger (2) one+ movable finger The number of spineminusbearing articles onthe frontal appendage (exclusive of the terminal movablefinger) varies from three in Yohoia (Whittington 1974 pl6 1ndash3 pl 7 3ndash6 pl 9 4ndash6) and Fortiforceps (Hou andBergstroumlm 1997 figs 31C D 32A 33B C E) to two inHaikoucaris (Chen et al 2004 fig 2A B) and Leanminuschoiliidae or only one the last being a chelicera
6 Long spinose projections on distal part of terminal threepodomeres of frontal appendage bearing a flagellum (0)absent (1) present The morphology of the great appendminusage in Leanchoiliidae is coded here We have not codedthe length of the spinose projections and the presence ofa flagellum as two independent characters Cotton andBraddyrsquos (2004) character 6 state 2 (ldquospinesrdquo longer thanthe entire podomere series) and character 8 state 1 (presminusence of an annulated flagellum) are thus grouped as a sinminusgle trait here
394 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Table 1 Characters coded for phylogenetic analysis Polymorphismindicated by ldquo+rdquo partial uncertainty by states separated by ldquordquo Abbreviminusations A Alalcomenaeus L Leanchoilia O Oestokerkus
Squamacula 0000minus minus minus2minus0 0110 00000 10minus minus0 1000 0
Cindarella 0030minus minus minus000 0010 00000 10minus minus0 11121 00
Kuamaia 000minus minus minus001 0011 10000 10minus minus0 11121 00
Kwanyinaspis 00 001 00112 1000 1000 1121 00
Olenoides 1020minus minus minus100 0011 00000 10minus minus0 01121 00
Retifacies 0020minus minus minus000 0010 00000 11000 1020 00
Aglaspis 10(23)0minus minus minus100 00101 00010 01001 0 00
Sidneyia 0000minus minus minus000 01102 00110 01101 00020 00
Emeraldella 0040minus minus minus2minus 0 00103 00110 01001 001(1+2)1 00
A cambricus 00111 10010 00001 00000 01110 01110 10
L illecebrosa 01211 11010 00001 01001 01210 11110 10
L persephone 00111 10010 00001 00000 01210 11110 10
L superlata 01111 11010 00001 01001 01210 11110 10
L hanceyi 00 00 0001 0000 01210
O megacholix 0011 100 00001 00000 01110 1110 0
Jianfengia 0021 0minus000 00005 00000 01000 010(12)0 0
Fortiforceps 00210 0minus00 00004 00000 01100 01230 20
Yohoia 00210 0minus000 00003 00011 01110 00010 (01)1
Haikoucaris 00211 0minus00 00003 00000 01(12)0 1010 0
Cheloniellon 0040minus minus minus100 00100 10010 01300 0030 00
Offacolus 02412 0minus200 0000 00010 01000 13minus minus 01
Xiphosura 00412 0minus100 10000 0010 01000 000minus minus 01
Eurypterida 00412 0minus100 10002 00010 01000 00minus minus minus 01
7 Length of flagellate great appendage (0) flagella termiminusnating at midlength of trunk (1) flagella extending to endof body Coding is applicable to Leanchoiliidae onlydistinguishing some Leanchoilia species that resembleAlalcomenaeus in having relative short great appendages(state 0) versus the longer appendages in L superlata andL illecebrosa
8 Position of lateral facetted eyes (0) ventral (1) dorsalsessile (2) absent Variation in Lamellipedia and severalother taxa coded herein follows Paterson et al (2010character 3)
9 Four or five ventral eyes arranged in a subtransverseband across head shield (0) absent (1) present A bandof ventral eyes is shared by the best known species ofLeanchoiliidae The distinction between three medianeyes in Alalcomenaeus cambricus (Briggs and Collins1999) and two in Leanchoilia illecebrosa (Liu et al2007) and both Leanchoilia superlata and L persephone(GarciacuteaminusBellido and Collins 2007) is uninformative forthe taxon sample here
10 Bulge in exoskeleton accommodating dropminusshaped venminustral eyes (0) absent (1) present This character followsEdgecombe and Ramskoumlld (1999 character 6) and Patminuserson et al (2010 character 5)
11 Dorsal median eyes (0) absent (1) present Coding for anabsence of dorsal median eyes in fossil taxa considers theabsence of either paired ocelli or a median eye tubercleand the character is scored as a chelicerate autapomorphy(see Cotton and Braddy 2004 character 24)
12 Width of doublure on cephalic shield (0) narrow or modminuserately wide (1) entire ventral side of head covered byexpansion of doublure The interpretation of state 1 inSidneyia and Squamacula is discussed by Cotton andBraddy (2004) and Paterson et al (2010 characters 12and 29)
13 Mouth covered by hypostome (0) absent (1) present Noldquoshort great appendagerdquo arthropods have been shown topossess a hypostome and a reconstruction of a smalllabrum in Alalcomenaeus (Briggs and Collins 1999) wasadmittedly conjectural The absence of a conspicuoushypostome contrasts with its ubiquitous presence in laminusmellipedians and several putative chelicerateminusallied taxathat have antennae (such as aglaspidids and Emeralminusdella) Chelicerates have a small epistomeminuslabrum (Bitschand Bitsch 2007) but like Cotton and Braddy (2004character 27 state 4) we code a hypostomal sclerite aslacking Kwanyinaspis has a large hypostome similar tothat found in aglaspidids (photograph of the holotype ofKwanyinaspis maotianshanensis shown to us by JavierOrtegaminusHernaacutendez [personal communication 2010])
14 Tergite articulations (0) extensive overlap of tergites (1)edge to edge articulations This character follows Edgeminuscombe and Ramskoumlld (1999 character 18) and Patersonet al (2010 character 20)
15 Number of trunk segments (0) nine (1) 11 (2) 12 (3)13 (4) 20 (5) 21 or 22 Coding is confined to taxa that do
not possess a pygidium because the precise number ofsomites associated with pygidia can sometimes not bedetermined with confidence Codings for Xiphosura andEurypterida use opisthosomal somite numbers followingShultz (2007 character 95)
16 Exoskeletal boundaries of anterior trunk segments (0)boundaries transverse or reflexed posterolaterally (1) reminusflexed anterolaterally This character follows Edgecombeand Ramskoumlld (1999 character 19) and Paterson et al(2010 character 21)
17 Trunk tergites with serrate lateral margins (0) absent (1)present Liu et al (2007 fig 2C) documented a row ofmarginal spines on the tergites of Leanchoilia illeceminusbrosa a featured shared by Leanchoilia superlata (egGarciacuteaminusBellido and Collins 2007 pl 1 7) but not otherterminals coded here
18 Differentiation in width of trunk tergites (0) trunk withgradational narrowing posteriorly (1) abrupt narrowingat boundary between prothorax and opisthothorax Anabrupt contraction in width between the boundary ofthe prominus and opisthothorax is observed in Emeraldellaand Sidneyia Yohoia has been reconstructed as having apronounced break in the shape of the trunk tergites beminushind segment 10 (Whittington 1974 textminusfig 2) thepoint at which the paratergal folds lose their spinose tipsand appendages are lacking Specimens preserved indorsal aspect (Whittington 1974 pl 10) suggest that thewidth of the opisthothorax is rather less abruptly narminusrowed and we code Yohoia as state 0
19 Appendages of posterior trunk segments (0) all trunktergites bearing appendages (1) one or more posteriortrunk segments lacking appendages Coding correspondsto Cotton and Braddyrsquos (2004) character 43 ldquopostabdominusmen of segments lacking appendagesrdquo
20 Paired tergal carinae (0) absent (1) present Liu et al(2007 265 fig 2F) recorded the presence of tergalcarinae in Leanchoilia illecebrosa shared with L superminuslata (eg Bruton and Whittington 1983 pl 17 106GarciacuteaminusBellido and Collins 2007 pl 1 5)
21 Pygidium (0) absent (1) present Coding of the pygiminusdium in many terminals herein was discussed by Paterminusson et al (2010 character 22)
22 Telson (0) absent (1) present Following Hendricks andLieberman (2008) a telson is a postminussegmental terminalstructure that lacks appendages Dunlop and Selden (1997)considered a telson to be absent in Cheloniellon but weidentify the conical projection posterior to the ultimate legpair and cerci (Stuumlrmer and Bergstroumlm 1978 figs 2 3 1011) as being most consistent with an identity as a telson
23 Telson shape (0) styliform (1) flapminuslike (2) lanceolate(3) short conical The outline of the telson in Haikouminuscaris ercaiensis is incompletely known but the mostcomplete specimen (Chen 2004 fig 472) constrains it aseither state 1 or state 2 A partial uncertainty coding isthus employed A unique state for Cheloniellon (state 3)is based on the interpretation in character 22
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 395
24 Telson fringed with spines (0) spines absent (1) spinespresent Presence of telson spines was introduced as a charminusacter by Hendricks and Lieberman (2008 character 34)
25 Paired flaps lateral to telson (0) absent (1) present Thischaracter describes the postminusventral plates of several taxa(see Edgecombe and Ramskoumlld 1999 character 25) Cotminuston and Braddy (2004 character 54) raised the possibilitythat the cerci of Cheloniellon could be homologous withpostminusventral plates (see also Hou and Bergstroumlm 1997 93)because they are similarly paired structures derived fromthe segment immediately anterior to the telson We havenot coded for this homology because the cerci of Chelominusniellon originate dorsally rather than ventrally
26 Midgut glands with submillimetric lamellae (0) absent(1) present Midgut glands in some stem group arthrominuspods and early crown group arthropods are metamericreniform organs with a characteristic lamellar structure(Butterfield 2002) Their morphology is identical inLeanchoilia and Oestokerkus notably including groovesthat radiate from a ventral median node (Figs 3A1 6A)The gut of Alalcomenaeus cambricus is more tubular(Briggs and Collins 1999 textminusfigs 10 11) and thoughan anterior six segments may display phosphatised latminuseral projections that likely represent gut diverticula (Wilminusson 2006) they have not been shown to have the detailedmorphology of Leanchoiliaminustype midgut glands Butterminusfield (2002 164) cited a figure from Bruton (1981 fig96) in support of ldquolamellarrdquo midgut glands in the cephaminuslon of Sidneyia Like Bruton (1981) in his description ofthe specimen we consider it likely that the structures inquestion are indeed gut diverticula but there is no basisfor identifying them more precisely as the reniformlamellar structures coded under this character
27 Exopod on first postminusantennal first postminusfrontal appendminusage segment (0) absent (1) present This character reminuslates to Cotton and Braddyrsquos (2004) character 3
28 Division of exopod (0) exopod an undivided flap (1)exopod differentiated into proximal and distal lobes (2)small flap with radiating lamellae (3) pediform exopoddivided into ca 6 articles Identification of the exopod inXiphosura follows Suzuki and Bergstroumlm (2008) Thischaracter groups conditions of the exopod shaft coded asindependent characters by Cotton and Braddy (2004)eg state 0 corresponds to their character 16 and state 1to their character 13 Our coding permits transformationbetween any two states without an extra implicit step
29 Exopod setae (0) short spiniform setae (1) long flatminustened tapering distally with slight separation (2)lamellar setae (flattened imbricating) (3) short finesetae A distinction is made between lamellipedian setaewhich are imbricated (state 2) and the pointed more disminustinctly separated and variably spiniform exopod setae ofleanchoiliids (state 1) This distinction is consistent withthe identification of megacheirans as nonminuslamellipedians(eg Hou and Bergstroumlm 1997 Bergstroumlm and Hou2005) Although Cotton and Braddy (2004 character 18)
regarded the form of the setae in Yohoia to be uncertainwe regard specimens figured by Whittington (1974 pl3 3) as conforming to the leanchoiliid state Squamaculadiffers from other taxa in its shorter spiniform setae(state 0) (Zhang et al 2004 textminusfig 1A B) Offacolushas longer spiniform projections on the distal articles ofthe exopod that have been variably identified as eitherspines or setae (Sutton et al 2002 fig 4b) but theirhomology with the setae that splay regularly along themargin of a flapminuslike exopod (eg states 0ndash3) is uncerminustain This and the following character for exopod setaeare left as ldquordquo for Offacolus A homology between thegill lamellae of Xiphosura and exopod setae of lamellipeminusdians has a long history (see Cotton and Braddy 2004character 18) We follow Suzuki and Bergstroumlm (2008)in identifying the xiphosuran gills as outgrowths of thebasis rather than of the exopod this perspective negates ahomology with the exopod setae of Lamellipedia Thepolymorphic coding for Emeraldella reflects the twoseparate states (1+2) displayed by the exopods in thetrunk (see Bruton and Whittington 1983 pl 8 45 47Edgecombe and Ramskoumlld 1999 fig 11)
30 Differentiation of setae along exopod (0) uniform orgradationally variable setae along length of exopod (1)lamellar setae confined to proximal lobe with distal lobebearing slender setae
31 Endopod segmentation (distal to coxa) (0) six or sevenpodomeres and terminal claw or telotarsal spurs (1)eight or more podomeres and terminal claw (2) ca 15podomeres State 1 is formulated to describe the segmenminustation of the leanchoiliid leg (GarciacuteaminusBellido and Collins2007 textminusfig 6 Liu et al 2007 fig 5) in contrast to thereduced number of podomeres in other euarthropods(Boxshall 2004) State 2 characterises Fortiforceps Apartial uncertainty coding is employed for Yohoia inwhich the precise number of podomeres is undeterminedand could be either state 0 or 1 (Whittington 1974 pl 74) but the spacing of the preserved podomere boundariesnegates the possibility of state 2
32 Endopods on anterior postminuscephalic segments (0) welldeveloped (1) rudimentary or lacking Following Cottonand Braddy (2004 character 11) the absence of trunkendopods in Yohoia (Whittington 1974) is coded as a puminustative homologue with the suppression of postminuscephalicendopods in Chelicerata
ResultsAs a convention for depicting the trees networks are rootedon Squamacula which separates artiopodan Lamellipedia(sensu Hou and Bergstroumlm 1997) from a group that includesMegacheira Chelicerata and Xenopoda all members ofwhich possess a telson
Under equal weights 14 shortest cladograms have a lengthof 77 steps with a Consistency Index of 064 and Retention Inminus
396 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
dex of 077 The strict consensus of these trees with supportvalues is shown in Fig 8 Leanchoiliidae is unambiguouslysupported in all 14 cladograms by four apomorphic characterstwo limbminusbearing cephalic segments posterior to the frontalappendage (character 3 state 1) long spinose projections onthe distal part of the terminal three podomeres of the frontalappendage that bear flagella (character 6 state 1) 11 trunksegments (character 15 state 1) and division of the exopodsinto two lobes (character 28 state 1) The arrangement of fouror five eyes in a subtransverse band across the cephalic shieldis unique to Leanchoiliidae (character 9 state 1) but is not anunambiguous autapomorphy of the group because the state isuncertain in close relatives notably Haikoucaris It is apominusmorphic for Leanchoiliidae only under particular optimisaminustions (Delayed Transformation)
The internal relationships of Leanchoiliidae are stableacross all 14 shortest cladograms Leanchoilia is united by thelanceolate telson (character 23 state 2) a unique unambiguminusous autapomorphy Within Leanchoilia a sister group relaminustionship between L superlata and L illecebrosa is well supminusported receiving a Bremer support of 3 and jackknife freminusquency of 86 These two species share four synapomorminusphies a medially pointed anterior cephalic margin (character2 state 1) flagella of the great appendage extending the lengthof the body (character 7 state 1) serrate lateral margins on thetrunk tergites (character 17 state 1) and paired tergal carinae(character 20 state 1) As such a species from Chengjiang (Lillecebrosa) and the Burgess Shale (L superlata) are moreclosely related to each other than are the two Burgess Shalecongeners a relationship that weakens the possibility that theoften cominusoccurring L superlata and L persephone could besexual dimorphs (GarciacuteaminusBellido and Collins 2007) ratherthan separate species The union of Leanchoilia and Oestominuskerkus is unambiguously supported by the shared midgutgland structure of both genera (character 26 state 1)
All shortest cladograms resolve Haikoucaris Yohoiaand Fortiforceps in that order as progressively less closelyrelated to Leanchoiliidae The union of these three taxa withLeanchoiliidae yields a clade that corresponds to Megacheirasensu Hou and Bergstroumlm (1997) apart from the ambiguousplacement of Jianfengia In half of the 14 shortest cladominusgrams Jianfengia is sister to the other megacheirans yieldminusing a monophyletic Megacheira ss In the remaining cladominusgrams Jianfengia is sister to Xenopoda sensu Hou andBergstroumlm (1997) Aglaspis and Chelicerata Haikoucarisand Leanchoiliidae are united by a frontal appendage comminusposed of two spineminusbearing articles in addition to the movminusable finger (character 5 state 1) Yohoia groups with thesetaxa based on spines fringing the telson (character 24 state1) and Fortiforceps is united with them at a broader levelbased on the flapminuslike telson shape (character 23 state 1)When Jianfengia is placed as sister to the other megacheiminusrans the entire clade is supported by the raptorial frontal apminuspendage (character 4 state 1) and the lack of a large hypominusstomal shield (character 13 state 0) In this set of cladominusgrams these features are nonminushomologous between Megaminus
cheira and Chelicerata whereas in some cladograms with thealternative resolution of Jianfengia as closer to cheliceratescharacter 4 is optimised as homologous in Megacheira andChelicerata
Reanalysis of the data with implied weights finds two optiminusmal trees (Fig 9) Relationships within Leanchoiliidae underequal weights are completely stable to reweighting as is theresolution of Haikoucaris as sister to Leanchoiliidae As suchthe character optimisations described above for the leanchoiminusliids are depicted the same in Fig 9 The main difference beminustween equal and implied weights involves the placement ofChelicerata Under equal weights (Fig 8) chelicerates are alminuslied to Xenopoda (here Emeraldella + Sidneyia + Chelominusniellon sensu clade L in Hou and Bergstroumlm 1997 fig 88)and Aglaspidida A clade composed of Aglaspidida Xenominuspoda and Chelicerata was similarly identified by Hou andBergstroumlm (1997 fig 88 their clade K) In our analysis thisgroup consistently finds support from one or more posteriortrunk segments lacking appendages (character 19 state 1) anddepending on alternative internal resolutions it is variably supminusported by dorsallyminuspositioned sessile eyes (character 8 state 1)and the absence of exopods on the first postminusantennalpostminuscheliceral limb (character 27 state 1) Under implied weightshowever chelicerates are more closely allied to Megacheirathan to aglaspidids or xenopodans (Fig 9)
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 397
2
582
76
Chelicera-morpha
Megacheira
62
2
53
50Leanchoiliidae
3
86
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
EurypteridaChelicerata
Fortiforceps
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia hanceyi
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
Fig 8 Strict consensus of 14 shortest cladograms (77 steps) based on charminusacter data in Table 1 Numbers above nodes are Bremer support those beminuslow nodes are jackknife frequencies gt50 Cheliceramorpha correspondsto a clade with the same composition in Cotton and Braddy (2004)
The major groupings in Fig 9 correspond closely to thoseidentified by Cotton and Braddy (2004) Notably the cladeuniting Aglaspis Xenopoda Offacolus Chelicerata Jianminusfengia and other Megacheira corresponds to their ldquoClade 3rdquo(Cotton and Braddy 2004 fig 8) for which they proposedthe name Cheliceramorpha The clade uniting CheloniellonAglaspis Sidneyia and Emeraldella in Fig 9 corresponds totheir ldquoClade 5rdquo and the clade that groups Megacheira andChelicerata is equivalent to their ldquoClade 4rdquo Their analysisdid not identify megacheirans as a monophyletic groupChelicerata was nested within the group and among taxaconsidered here Yohoia was resolved closer to cheliceratesthan to Leanchoiliidae In contrast Yohoia was regarded byBergstroumlm and Hou (2005) as more closely allied to Leanminuschoiliidae than to chelicerates a megacheiran grouping morecompatible with our cladograms The analysis of ldquogreat apminuspendage arthropodsrdquo by Vannier et al (2009) identified a
monophyletic Leanchoiliidae but a polyphyletic Megacheira(to the exclusion of Haikoucaris and Jianfengia) Howevertheir resolution of Fortiforceps and even more so Yohoia asthe closest relatives of Leanchoiliidae is as in our trees (apartfrom the different position of Haikoucaris)
In summary a major controversy in the systematics of theldquoshort great appendagerdquo arthropods remains whether thechelicerates are nested within the group or are instead moreclosely allied to taxa conventionally identified as lamelliminuspedians specifically Xenopoda and Aglaspidida We findthat both of these results are compatible with the availabledata depending on character weighting protocols Noneminustheless the broader megacheiranminuschelicerate issue does notcloud the status of leanchoiliids as a clade or the identificaminustion of their closest relatives (ie Haikoucaris and Yohoia)and the internal relationships of Leanchoiliidae are stableand supported by compelling characters
398 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
Eurypterida
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
2 7 17 20
3
2
1 1 1
26
1
281563
1 1 1 1
5
1
24
1
23 19 20 27 32
28Clade 4
1 0
4 13 11 27 0
154 1 1
3 19 32Clade 3
2 3
2 28
0
3 1 121
1
22
Clade 5
1 0
19 27
1
25
15 28 301 1
2312
1 150 1 3 3
15 16 2923
18
1 1
10 16
1 1
1 8
1
14
3
3
28 30
1 0
5
15
1 1
1
1
22
Fortiforceps
23
1 1 0 1
1 1
Leanchoilia hanceyi
2
2
1
1
Fig 9 Characters optimised on one of two shortest cladograms with imminusplied weights using a concavity constant k = 3 Black squares are nonminushomoplastic changes white squares homoplastic changes all changes areunambiguous Branches without strict support collapsed after optimisingcharacters Numbers for characters (above the branches) and states (belowthe branches) correspond to Table 1 ldquoClades 3ndash5rdquo correspond to groups sonumbered by Cotton and Braddy (2004 fig 8)
AcknowledgementsAccess to the field area has been most kindly allowed by the Buck famminusily We thank our collaborators in Emu Bay Shale research Jim Jago(University of South Australia Mawson Lakes) Jim Gehling andMike Lee (South Australian Museum Adelaide) for tremendous supminusport and advice as well as Mike Gemmell Dennis Rice Allison DaleyAaron Camens John Laurie Glenn Brock Pierre Kruse and Trevorand Jenny Worthy (all South Australian Museum) for their assistancein the field and Natalie Schroeder MaryminusAnne Binnie and RondaAtkinson (all South Australian Museum) for organisational support inthe lab and in the field Javier OrtegaminusHernaacutendez (University of Camminusbridge Cambridge UK) advised us on Kwanyinaspis and the journalrsquosreferees provided useful feedback TNT is available courtesy of theWilli Hennig Society Funding has been provided by an AustralianResearch Council Linkage grant (LP0774959) to the University ofAdelaide with generous financial assistance from Beach Energy Ltdand the South Australian Museum SeaLink provided logistical supminusport DGBrsquos travel to Australia has been supported by grants from theSpanish Research Council (CSIC PA 1001105) the Complutense Uniminusversity of Madrid and the Spanish Ministry of Education and Science(CGL2006minus12245BTE CGL2009minus07073 and RYC2007minus00090)
ReferencesBergstroumlm J and Hou XminusG 2005 Early Palaeozoic lamellipedian arthrominus
pods In S Koenemann and RA Jenner (eds) Crustacea and Arthrominuspod Relationships Crustacean Issues 16 73ndash93
Bitsch J and Bitsch C 2007 The segmental organization of the head reminusgion in Chelicerata a critical review of recent studies and hypothesesActa Zoologica 88 317ndash335
Bousfeld EL 1995 A contribution to the natural classification of Lower andMiddle Cambrian arthropods food gathering and feeding mechanismsAmphipacifica 2 3ndash34
Boxshall GA 2004 The evolution of arthropod limbs Biological Reviews79 253ndash300
Bremer K 1994 Branch support and tree stability Cladistics 10 295ndash304Briggs DEG and Collins D 1999 The arthropod Alalcomenaeus cambricus
Simonetta from the Middle Cambrian Burgess Shale of British ColumbiaPalaeontology 42 953ndash977
Briggs DEG and Robison RA 1984 Exceptionally preserved nonminustrilobite arthropods and anomalocaridids from the Middle Cambrian ofUtah University of Kansas Paleontological Contributions 111 1ndash23
Briggs DEG Lieberman BS Hendricks JR Halgedahl SL andJerrard RD 2008 Middle Cambrian arthropods from Utah Journal ofPaleontology 82 238ndash254
Bruton DL 1981 The arthropod Sidneyia inexpectans Middle CambrianBurgess Shale British Columbia Philosophical Transactions of theRoyal Society of London Series B 295 619ndash656
Bruton DL and Whittington HB 1983 Emeraldella and Leanchoiliatwo arthropods from the Burgess Shale Middle Cambrian British Cominuslumbia Philosophical Transactions of the Royal Society of London Seminusries B 300 553ndash582
Budd GE 2002 A palaeontological solution to the arthropod head probminuslem Nature 417 271ndash275
Butterfield NJ 2002 Leanchoilia guts and the interpretation of threeminusdiminusmensional structures in Burgess Shaleminustype fossils Paleobiology 28155ndash171
Chen JminusY 2004 The Dawn of Animal World [in Chinese] 366 pp JiangScience and Technology Press Nanjing
Chen JminusY Waloszek D and Maas A 2004 A new ldquogreatminusappendagerdquo arminusthropod from the Lower Cambrian of China and homology of cheliceratechelicerae and raptorial anterominusventral appendages Lethaia 37 3ndash20
Cotton TJ and Braddy SJ 2004 The phylogeny of arachnomorph arthrominuspods and the origin of the Chelicerata Transactions of the Royal Societyof Edinburgh Earth Sciences 94 169ndash193
Daley AC Budd GE Caron JminusB Edgecombe GD and Collins D2009 The Burgess Shale anomalocaridid Hurdia and its significancefor early euarthropod evolution Science 323 1597ndash1600
Dunlop JA 2006 New ideas about the euchelicerate stemminuslineage In CDeltshev and P Stoev (eds) European Arachnology 2005 Acta Zoominuslogica Bulgarica (Supplement 1) 9ndash27
Dunlop JA and Selden PA 1997 The early history and phylogeny of thechelicerates In RA Fortey and RH Thomas (eds) Arthropod Relaminustionships Systematics Association Special Volume 55 221ndash235
Edgecombe GD and Ramskoumlld L 1999 Relationships of CambrianArachnata and the systematic position of Trilobita Journal of Paleonminustology 73 263ndash287
Farris JS Albert VA Kaumlllersjouml M Lipcomb D and Kluge AG1996 Parsimony jackknifing outperforms neighborminusjoining Cladistics12 99ndash124
GarciacuteaminusBellido DC and Collins D 2007 Reassessment of the genusLeanchoilia (Arthropoda Arachnomorpha) from the Middle CambrianBurgess Shale British Columbia Canada Palaeontology 50 693ndash709
GarciacuteaminusBellido DC Paterson JR Edgecombe GD Jago JB GehlingJG and Lee MSY 2009 The bivalved arthropods Isoxys and Tuzoiawith softminuspart preservation from the lower Cambrian Emu Bay ShaleLagerstaumltte (Kangaroo Island Australia) Palaeontology 52 1221ndash1241
Goloboff PA 1993 Estimating character weights during tree searchCladistics 9 83ndash91
Goloboff PA Farris JS and Nixon KC 2008 TNT a free program forphylogenetic analysis Cladistics 24 774ndash786
Harzsch S and Hafner G 2006 Evolution of eye development in arthrominuspods phylogenetic aspects Arthropod Structure amp Development 35319ndash340
Hendricks JR and Lieberman BS 2008 New phylogenetic insights intothe Cambrian radiation of arachnomorph arthropods Journal of Paleminusontology 82 585ndash594
Hou XminusG and Bergstroumlm J 1997 Arthropods of the Lower CambrianChengjiang fauna southwest China Fossils and Strata 45 1ndash116
Jago JB Zang W Sun X Brock GA Paterson JR and SkovstedCB 2006 A review of the Cambrian biostratigraphy of South Austraminuslia Palaeoworld 15 406ndash423
Liu Y Hou XminusG and Bergstroumlm J 2007 Chengjiang arthropod Leanminuschoilia illecebrosa (Hou 1987) reconsidered GFF 129 263ndash272
Luo H Hu S Chen L Zhang S and Tao Y 1999 Early CambrianChengjiang fauna from Kunming region China [in Chinese] 129 ppYunnan Science and Technology Press Kunming
Luo H Hu S Zhang S and Tao Y 1997 New occurrence of the EarlyCambrian Chengjiang fauna in Haikou Kunming Yunnan Provinceand study on Trilobitoidea Acta Geologica Sinica 71 122ndash132
Nixon KC 2002 WINCLADA Version 19908 Cornell University IthacaNY
Orr PJ Siveter DJ Briggs DEG Siveter DJ and Sutton MD 2000A new arthropod from the Silurian KonservatminusLagerstaumltte of Hereminusfordshire Proceedings of the Royal Society Series B Biological Sciminusences 267 1497ndash1504
Paterson JR and Brock GA 2007 Early Cambrian trilobites fromAngorichina Flinders Ranges South Australia with a new assemblagefrom the Pararaia bunyerooensis Zone Journal of Paleontology 81116ndash142
Paterson JR and Jago JB 2006 New trilobites from the Lower CambrianEmu Bay Shale Lagerstaumltte at Big Gully Kangaroo Island South Ausminustralia Memoirs of the Association of Australasian Palaeontologists 3243ndash57
Paterson JR Edgecombe GD GarciacuteaminusBellido DC Jago JB andGehling JG 2010 Nektaspid arthropods from the lower CambrianEmu Bay Shale Lagerstaumltte South Australia with a reassessment oflamellipedian relationships Palaeontology 53 377ndash402
Paterson JR Jago JB Gehling JG GarciacuteaminusBellido DC Edgecombe
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 399
GD and Lee MSY 2008 Early Cambrian arthropods from the EmuBay Shale Lagerstaumltte South Australia 319ndash325 In I Raacutebano RGozalo and D GarciacuteaminusBellido (eds) Advances in Trilobite ResearchCuadernos del Museo Geominero 9 319ndash325
Shultz JW 2007 A phylogenetic analysis of the arachnid orders based onmorphological characters Zoological Journal of the Linnean Society150 221ndash265
Stein M 2010 A new arthropod from the Early Cambrian of North Greenminusland with a ldquogreat appendagerdquominuslike antennula Zoological Journal ofthe Linnean Society 158 477ndash500
Stuumlrmer W and Bergstroumlm J 1978 The arthropod Cheloniellon from theDevonian Hunsruumlck Shale Palaumlontologische Zeitschrift 52 57ndash81
Sutton MD Briggs DEG Siveter DJ Siveter DJ and Orr PD2002 The arthropod Offacolus kingi (Chelicerata) from the Silurian ofHerefordshire England computer based morphological reconstrucminustions and phylogenetic affinity Proceedings of the Royal Society B Biminusological Sciences 269 1195ndash1203
Suzuki Y and Bergstroumlm J 2008 Respiration in trilobites a reevaluationGFF 130 211ndash229
Vannier J GarciacuteaminusBellido DC Hu XminusX and Chen AminusL 2009 Arthrominuspod visual predators in the early pelagic ecosystem evidence from the
Burgess Shale and Chengjiang biotas Proceedings of the Royal SocietyB Biological Sciences 276 2567ndash2574
Whittington HB 1974 Yohoia Walcott and Plenocaris n gen arthropodsfrom the Burgess Shale Middle Cambrian British Columbia Geologiminuscal Survey of Canada Bulletin 231 1ndash21
Wills MA Briggs DEG Fortey RA Wilkinson M and SneathPHA 1998 An arthropod phylogeny based on fossil and recent taxaIn GD Edgecombe (ed) Arthropod Fossils and Phylogeny 33ndash105Columbia University Press New York
Wilson LA 2006 Food for thought a morphological and taphonomic studyof fossilised digestive systems from Early to Middle Cambrian taxa275 pp Unpublished PhD thesis University of Cambridge Cambridge
Zhang XminusG and Hou X 2007 Gravitational constraints on the burial ofChengjiang fossils Palaios 22 448ndash453
Zhang XminusL Han J Zhang Z Liu H and Shu D 2004 Redescription ofthe Chengjiang arthropod Squamacula clypeata Hou and Bergstroumlm fromthe Lower Cambrian Southminuswest China Palaeontology 47 605ndash617
Zhao Y Zhu M Babcock LE Yuan L Parsley RL Peng J YangX and Wang Y 2005 Kaili Biota a taphonomic window on diversifiminuscation of metazoans from the basal Middle Cambrian Guizhou ChinaActa Geologica Sinica 79 751ndash765
400 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011

DiscussionmdashMembership of Oestokerkus in Leanchoiliidaesensu Liu et al (2007) a group that includes the formerly reminuscognised families Alalcomenaeidae Simonetta and DelleCave 1975 and Actaeidae Simonetta and Delle Cave 1975 isclearly demonstrated by the morphology of the frontal apminuspendage which has greatly elongated spinose projections deminusveloped on its distal two articles and the movable finger eachprojection terminating at the same length and bearing a longflagellum presence of 11 trunk segments anteroventrallyminuspositioned eyes pendant appendages that have a similar biraminusmous structure in the cephalon and trunk and spines fringingthe margin of a long telson
The distinction between Alalcomenaeus and Leanchoiliahas most recently been reviewed by Liu et al (2007) whoemphasised the shape of the telson as a diagnostic characterthat of Leanchoilia being pointed and that of Alalcomenaeusrounded The telson of Oestokerkus megacholix described beminuslow differs substantially from the other species of both genminusera The telson widens distally in the holotype of O megaminuscholix (Figs 2 3A2 B) and is inferred to be forked Marginalspines are confined to the posterior edge rather than being deminusveloped along the lateral margins of a lanceolate telson as inall species of Leanchoilia or a paddleminusshaped posteriorlyrounded telson in Alalcomenaeus To date specimens of Omegacholix are preserved only in lateral aspect most likelydue to gravitational settling of the body on its sidemdasha comminusmon burial position of other leanchoiliids (Zhang and Hou2007)mdashbecause of the deep dorsominusventral versus transverselynarrow proportions of the body thus our interpretation of thetelson (see description below) has not yet been confirmed indorsal aspect Phylogenetic analysis (see below) resolves Omegacholix as sister to the widely accepted grouping ofLeanchoilia so we have elected to erect a new genus for itrather than to widen the scope of Leanchoilia and be forced torediagnose that genus using characters other than telson shapeThis taxonomic framework is consistent with the generic conminuscepts of Liu et al (2007)
In addition to telson form Oestokerkus is further distinminusguished from other leanchoiliid genera by relative proporminustions of the cephalic shield and trunk pleurotergites The ceminusphalic shield of O megacholix occupies a greater proporminustion of body length than in Leanchoilia or Alalcomenaeusthe head being ca 36 the length of the trunk (excluding thetelson) compared with 28 in Alalcomenaeus cambricus25 in Leanchoilia illecebrosa 23 in Leanchoilia perseminusphone and 26 in Leanchoilia superlata The paratergalfold in O megacholix is also considerably deeper than inother leanchoiliids The height to length ratio of ca 39 inO megacholix (Fig 2) contrasts with ca 19ndash22 in thethree wellminusknown Burgess Shale and Chengjiang species ofLeanchoilia (eg Liu et al 2007 fig 2D E) as well as Acambricus From what is observed in other fossils from thislocalitymdashand is also the case in the Burgess Shale Leanminuschoiliamdashdespite the fossils being compressed flat there islittle distortion Therefore the dimensions discussed aboveare not considered to deviate in any significant way from
those the animals would have had in life (Fig 7) The largemidgut glands of O megacholix are approached in size bysome species of Leanchoilia notably L superlata (GarciacuteaminusBellido and Collins 2007 pl 2 5) in which their structureof radiating grooves is the same The position and size ofthe lateral eye in O megacholix resemble those of Acambricus in which laterally preserved specimens showthe eye to be bulging at the anterolateral margin of the ceminusphalic shield (Briggs and Collins 1999 pl 2 4 pl 6 3) butpreservation in dorsoventral aspect shows that the lateralpair of eyes to be adaxial to the cephalic margin (Briggs andCollins 1999 pl 1 1) No evidence is available to indicatewhether median eyes (either two as in Leanchoilia or threeas in Alalcomenaeus) are present in Oestokerkus becausethis requires a dorsoventral aspect though the resolution ofOestokerkus within Leanchoiliidae predicts the presence ofthis feature
We have not coded the number of cephalic appendages inO megacholix in the cladistic analysis but the only specimenthat preserves the endopods (see species description) is mostconsistent with two biramous cephalic appendages as hasbeen documented in A cambricus (Briggs and Collins 1999)and in both L superlata and L persephone (GarciacuteaminusBellidoand Collins 2007) and contrasting with three pairs of biraminusmous cephalic appendages in L illecebrosa (Chen et al 2004Liu et al 2007) We reject the opinion of Chen et al (2004)that Briggs and Collins (1999) probably miscounted forAlalcomenaeus because more recent observations on BurgessShale Leanchoilia show that two pairs are indeed present insome members of Leanchoiliidae (GarciacuteaminusBellido and Collins2007) The variability in this feature in a narrowlyminuscircumminusscribed monophyletic group (ie Leanchoilia) indicates that itis of fairly minor taxonomic weight within this family
Oestokerkus megacholix sp novFigs 2ndash7
Etymology From Greek mega large and cholix gut in reference to thevoluminous midgut glands
Type material Holotype SAM P43631 a b part and counterpart preminusserved in lateral aspect (Figs 2 3) from the Emu Bay Shale BuckQuarry preserving cephalon including spinose projections and flagellaof great appendages all 11 trunk segments and most of telson includminusing a few marginal spines midgut glands exposed in head and anteriortwominusthirds of trunk some exopod setae of cephalic appendages as wellas lobes and setae of those associated with entire trunk Paratypes Allpreserved in lateral aspect SAM P45167 (Fig 4) cephalon exposingeye and peduncle of great appendage most of trunk with exopod setaeand a few endopods SAM P43630ab (Fig 5B) part and counterpartpreserving cephalic shield spinose projections of great appendage andanterior eight trunk segments two midgut glands and exopod setae ofcephalic and trunk appendages exposed SAM P15531 (Fig 5A) partonly preserving cephalon including spinose projections of great apminuspendage and most of trunk with midgut glands SAM P15438 (unfigminusured) incomplete cephalic shield with parts of great appendage comminusplete trunk and apparently proximal part of telson SAM P45168 (Fig6) part only cephalon with partial great appendages most of trunkwith midgut glands and exopod setae SAM P45169a b (unfigured)cephalon with partial great appendages trunk with midgut glands SAMP45177a b SAM P45178a b (unfigured) two weathered specimens
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 387
388 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
55 mmmm
5 mm
anus
telsontr
tr
t11
t5
mg
txm
t1
cs
gap
vm
ga
gaf
txts
cx
Fig 2 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov Holotype SAM P43631a A Photograph B Interpretative cameralucida drawing Arrows indicate tips of trunk exopod setae Abbreviations cs cephalic shield cx cephalic exopods ga great appendage gaf great appendminusage flagellum gap peduncle of great appendage mg midgut glands t1 tergite of trunk segment 1 tr reinforcing ridges on telson ts telson marginalspines tx trunk exopods txm trunk exopod lobe margins vm ventral margin of tergites Boundaries between trunk tergites indicated by dashed linesabove dorsal margin
Fig 3 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov Holotype SAM P43631 A P43631a A1 Midgut glands and tergiteboundaries in dorsal part of trunk Arrows indicate furrows that may represent position of dorsal transverse tendons A2 Telson light from NE A3 Spinoseprojections and flagella of distal articles of frontal appendage and exopod setae of cephalic and anterior trunk appendages and posterior margins of lobes ofthe latter B P43631b telson light from NE Arrows indicate tips of exopod setae Abbreviations gaf great appendage flagellum mg midgut glands oloverlap between adjacent trunk tergites tr reinforcing ridges on telson ts telson marginal spines txm trunk exopod lobe margins vm ventral margin oftergites
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 389
SAM P45904a b (unfigured) poorlyminuspreserved cephalic shield trunkwith wellminuspreserved midgut glands proximal part of telson SAMP45905ab (unfigured) cephalic shield lacking great appendages butexposing an eye trunk with incomplete telson SAM P45906a b (unfigminusured) great appendages with spinose projections oriented anteriorlytrunk preserved to end of midgut glands SAM P45908a b (unfigured)cephalic shield great appendages with flagella most of trunk (telsonmissing) showing midgut glands and exopod setae SAM P45909 (unminusfigured) small specimen with great appendages and series of mineralminusised midgut glands SAM P45910a b (unfigured) incomplete great apminuspendages oriented anteriorly trunk with complete series of midgutglands but indistinct appendages
Type locality Buck Quarry Big Gully (Fig 1)
Type horizon Emu Bay Shale (Cambrian Series 2 Stage 4)
DiagnosismdashAs for genus
DescriptionmdashCephalic shield 36 length of the trunk sagitminustally dorsal margin of cephalic shield gently convex in lateralview anteromedian margin inferred to be evenly rounded(based on comparison with lateral versus dorsal preservation ofBurgess Shale Leanchoilia and Alalcomenaeus) lacking an upminusturned median process Eye visible in SAM P54167 (Fig 4A)and P45905 relatively large (20 sagittal length of cephalon)exposed under anterolateral part of cephalic shield (Fig 4A2)
390 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Fig 4 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov SAM P45167 A P45167a A1 cephalic shield and most of trunkA2 detail of cephalic shield B P45167b A2 and B light from NE Overlap between adjacent tergites offset on left (Lol) and right (Rol) sides of bodyBoundaries between cephalic shield and anterior trunk tergites delimited in B by dashed lines Arrows mark positions of inferred dorsal transverse tendonsAbbreviations cn1ndash2 cephalic endopods cs cephalic shield gap peduncle of great appendage ol overlap between adjacent trunk tergites t1ndasht3 tergitesof trunk segments 1ndash3 tn trunk endopods
Fig 5 Leanchoiliid megacheiran arthropod Oestokerkus megacholix genet sp nov A SAM P 15531 B SAM P43630b Light from NE Abbreviaminustions cs cephalic shield ga great appendage gap peduncle of great apminuspendage mg midgut glands ol overlap between adjacent trunk tergites txtrunk exopods
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 391
Great appendage robust proportions of peduncle and disminustal spineminusbearing articles as in other members of Leanchoiliminusidae more commonly preserved with posterior orientation
but anteriorly oriented in SAM P45906 and SAM P45910Each of the three long spinose projections terminating at thesame length the projections evenly tapering distally with
392 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Fig 6 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov SAM P45168 A head and anterior part of trunk B Detail of spinoseprojections of great appendages and excretory organs (sensu Bruton and Whittington 1983) Abbreviations cs cephalic shield eo excretory organ gagreat appendage mg midgut glands tx trunk exopods
margins straight for much of their length or slightly curvedterminally distal spinose projection distinctly stouter at itsbase than the two more proximal projections and more markminusedly tapering along its length Dark irregularminusshaped low reminuslief lump on basal spineminusbearing article of great appendagesof SAM P45168 (eo in Fig 6) and SAM P45908 correminussponding to excretory organ of Bruton and Whittington(1983) Long flagella extending from each of the threespinose projections seen only in SAM P43631 (Figs 23A3) of similar narrow width along its preserved extent anminusnulations indistinguishable
Trunk of 11 segments of even length Maximum height oftrunk relative to length (excluding telson) ca 39 bodydeepest at tergite 4 Trunk tergite boundaries marked as narminusrow ridges in several specimens corresponding to narrowband of overlap between adjacent tergites (ol in Figs 2 3A14A1 B) Some specimens show tergites to be bisected by anarrow but sharp furrow of uncertain identity (eg possiblyimpression of dorsal transverse tendon cf Stein 2010 fig12B C E) (arrows in Figs 3A1 4B)
Exopods with fringe of long setae on cephalic and trunkappendages (Fig 2) setal fringe gradually shortening fromabout the fourth trunk segment Setae project ventrally bymore than 30 height of tergites in deepest part of trunk(Fig 2B) setae flattened narrow and parallelminussided alongmost of their length tapering near their distal ends to termiminusnate as a pointed tip (Figs 2B 3A3 arrows) Endopods exminusposed only in SAM P45167 (Fig 4) two pairs associatedwith cephalon (Fig 4B) those of anterior segments of trunkof similar morphology to those in cephalon all slender lackminusing obvious endites podomere boundaries unrecognisable
Midgut glands occupy a large extent of the dorsoventralheight of the trunk two midgut glands in cephalon of similarsize to those in each of the first seven segments of the trunk(Figs 3A1 5A 6A) all developed as radiating grooves emaminusnating from a globular structure at their ventral edge as in
Leanchoilia superlata (GarciacuteaminusBellido and Collins 2007 pl2 6) Foregut trace in SAM P43631 (Fig 2) indicates that gutcurves ventrally in anterior part of cephalon mouth regionnot preserved Terminal part of hindgut (in trunk segment 11)with closely spaced longitudinal lineations (Fig 2) that apminusparently correspond to folds in the gut wall hindgut termiminusnating at base of telson
Telson most completely preserved in holotype (SAMP43631 Figs 2B 3A2 B) In this specimen the telson likerest of body compressed in lateral aspect Telson shows meminusdian and marginal reinforcing ridges connecting a flat trianminusgular surface this surface appears to be partially broken inSAM P43631 showing underlying surface of other (left) halfof telson Lateral preservation of specimen implies that topcorresponds to an axial structure and bottom to a marginalone 30ndash35 angle in posterior edge of vane connecting thesetwo structures requires that in dorsal aspect telson would apminuspear forked and widening distally Five stout pointed dorminussally curved marginal spines attached to outer posterior edgeon each side of telson
Phylogenetic analysis ofLeanchoiliidaeDetermining the systematic position of Oestokerkus demandsan appraisal of the relationships of Leanchoiliidae as a wholeCladistic analysis including the bestminusknown members ofMegacheira including leanchoiliids has been conducted byCotton and Braddy (2004) and Vannier et al (2009) The taxominusnomic sampling used by Cotton and Braddy (2004) includedseveral megacheiran genera representative chelicerates anumber of taxa representing groups that have been proposedas possible chelicerate relatives (eg Sidneyia AglaspisCheloniellon) in other analyses (Dunlop and Selden 1997Hou and Bergstroumlm 1997) and members of the major lineagesof the Artiopoda sensu Hou and Bergstroumlm (1997) ie trilominusbiteminusallied lamellipedians Because artiopodan interminusrelationminusships have remained quite stable through a series of phylogenminusetic analyses (Edgecombe and Ramskoumlld 1999 Cotton andBraddy 2004 Hendricks and Lieberman 2008 Paterson et al2010) and artiopodans are now often regarded as less closelyrelated to Megacheira than is Chelicerata (Chen et al 2004Cotton and Braddy 2004 Dunlop 2006) we have reduced thesampling of Artiopoda in the present analysis To augment thediversity of ldquoshort great appendagerdquo arthropods Haikoucarisis added based on the description of H ercaiensis by Chen etal (2004) Offacolus (Orr et al 2000 Sutton et al 2002) hasbeen allied to Chelicerata and is included as a terminal Beminuscause the present analysis is principally concerned with relaminustionships of and within Leanchoiliidae sensu Liu et al (2007)we follow Vannier et al (2009) in sampling Alalcomenaeusand Leanchoilia at the species level Within Chelicerata Cotminuston and Braddy (2004) and Hendricks and Lieberman (2008)sampled ancestral states in Xiphosura by coding the Devonian
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 393
Fig 7 Reconstruction of Oestokerkus megacholix gen et sp nov in lateralview Total length and annulations on the flagella and positions of podominusmere boundaries of the frontal appendage are based on Leanchoilia superminuslata (Bruton and Whittington 1983)
synziphosurine Weinbergina Our coding for Xiphosura alsoemploys information from extant xiphosurans because theysupply more reliable information on the homology of the partsof the trunk appendages (Suzuki and Bergstroumlm 2008) and eyeanatomy (Harzsch and Hafner 2006) among other charactersystems than do fossils alone
Our character set extracts characters from the matrix ofCotton and Braddy (2004) that vary in the revised taxonomicsample reformulates some characters to accommodate reminusvised homology concepts and adds new characters especiallyfor leanchoiliids In total 32 characters are included (Table 1)In this analysis the primary homology of the ldquoshort great apminuspendagerdquo and chelicera is coded (as described by character 4)and the taxon sampling broadly constrains megacheirans aschelicerate relatives (as defended by Chen et al 2004 Cottonand Braddy 2004) rather than testing the alternative that theycould be stem group Euarthropoda (Budd 2002 Bergstroumlmand Hou 2005) A test of the two alternatives involves samminuspling of the entire diversity of euarthropods and a range ofnonminuseuarthropod outgroups an exercise beyond the scope ofthis study
Characters1 Mineralised cuticle (0) absent (1) present For definiminus
tion and discussion of this character see Cotton andBraddy (2004 character 30) and Paterson et al (2010character 1)
2 Curvature of anterior margin of cephalic shield (0) rounminusded or straight (1) pointed medially (2) with multiplespinose projections This character codes for variability inthe shape of the cephalic shield in Leanchoiliidae (state 0and 1 sensu GarciacuteaminusBellido and Collins [2007 textminusfigs10 and 9 respectively]) State 2 is added to accommodateOffacolus the median spine of a series that fringes the ceminusphalic margin is not convincingly homologised with themedian projection in leanchoiliids
3 Number of cephalic somitesappendages (0) antenna orfrontal appendage only (1) antenna or frontal appendageand two limb pairs (2) antenna or frontal appendage andthree limb pairs (3) antenna or frontal appendage and fourlimb pairs (4) antenna or frontal appendage and five limbpairs Coding for lamellipedians and Aglaspis is as discussedby Paterson et al (2010 character 2) Coding for Alalcomeminusnaeus cambricus (state 1) follows Briggs and Collins (1999)Leanchoilia illecebrosa (state 2) follows Liu et al (2007)and Leanchoilia persephone and Leanchoilia superlata (state1) follows GarciacuteaminusBellido and Collins (2007)
4 Form of first cephalic appendage (0) antenna (1) anteminusriorly directed raptorial appendage with reduced numberof podomeres each of the terminal podomeres bearing aspinose process on its distal margins a geniculation beminustween spinose podomeres and peduncle This characteris formulated to identify a homology between the frontalappendages of megacheirans and the chelicera of Cheliminus
cerata and state 1 corresponds to the putative apomorminusphic detail defended by Chen et al (2004) and Cottonand Braddy (2004) Vannier et al (2009 character 8)specify another shared property of ldquoshort great appendminusagesrdquo of the megacheiran (ss) type in their characterstate ldquobent at an anglerdquo referring to the geniculation beminustween articles of the peduncle or between the peduncleand distal spiniferous articles This property is sharedwith Chelicerata
5 Number of spineminusbearing articles on frontal appendage (0)three + movable finger (1) two + movable finger (2) one+ movable finger The number of spineminusbearing articles onthe frontal appendage (exclusive of the terminal movablefinger) varies from three in Yohoia (Whittington 1974 pl6 1ndash3 pl 7 3ndash6 pl 9 4ndash6) and Fortiforceps (Hou andBergstroumlm 1997 figs 31C D 32A 33B C E) to two inHaikoucaris (Chen et al 2004 fig 2A B) and Leanminuschoiliidae or only one the last being a chelicera
6 Long spinose projections on distal part of terminal threepodomeres of frontal appendage bearing a flagellum (0)absent (1) present The morphology of the great appendminusage in Leanchoiliidae is coded here We have not codedthe length of the spinose projections and the presence ofa flagellum as two independent characters Cotton andBraddyrsquos (2004) character 6 state 2 (ldquospinesrdquo longer thanthe entire podomere series) and character 8 state 1 (presminusence of an annulated flagellum) are thus grouped as a sinminusgle trait here
394 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Table 1 Characters coded for phylogenetic analysis Polymorphismindicated by ldquo+rdquo partial uncertainty by states separated by ldquordquo Abbreviminusations A Alalcomenaeus L Leanchoilia O Oestokerkus
Squamacula 0000minus minus minus2minus0 0110 00000 10minus minus0 1000 0
Cindarella 0030minus minus minus000 0010 00000 10minus minus0 11121 00
Kuamaia 000minus minus minus001 0011 10000 10minus minus0 11121 00
Kwanyinaspis 00 001 00112 1000 1000 1121 00
Olenoides 1020minus minus minus100 0011 00000 10minus minus0 01121 00
Retifacies 0020minus minus minus000 0010 00000 11000 1020 00
Aglaspis 10(23)0minus minus minus100 00101 00010 01001 0 00
Sidneyia 0000minus minus minus000 01102 00110 01101 00020 00
Emeraldella 0040minus minus minus2minus 0 00103 00110 01001 001(1+2)1 00
A cambricus 00111 10010 00001 00000 01110 01110 10
L illecebrosa 01211 11010 00001 01001 01210 11110 10
L persephone 00111 10010 00001 00000 01210 11110 10
L superlata 01111 11010 00001 01001 01210 11110 10
L hanceyi 00 00 0001 0000 01210
O megacholix 0011 100 00001 00000 01110 1110 0
Jianfengia 0021 0minus000 00005 00000 01000 010(12)0 0
Fortiforceps 00210 0minus00 00004 00000 01100 01230 20
Yohoia 00210 0minus000 00003 00011 01110 00010 (01)1
Haikoucaris 00211 0minus00 00003 00000 01(12)0 1010 0
Cheloniellon 0040minus minus minus100 00100 10010 01300 0030 00
Offacolus 02412 0minus200 0000 00010 01000 13minus minus 01
Xiphosura 00412 0minus100 10000 0010 01000 000minus minus 01
Eurypterida 00412 0minus100 10002 00010 01000 00minus minus minus 01
7 Length of flagellate great appendage (0) flagella termiminusnating at midlength of trunk (1) flagella extending to endof body Coding is applicable to Leanchoiliidae onlydistinguishing some Leanchoilia species that resembleAlalcomenaeus in having relative short great appendages(state 0) versus the longer appendages in L superlata andL illecebrosa
8 Position of lateral facetted eyes (0) ventral (1) dorsalsessile (2) absent Variation in Lamellipedia and severalother taxa coded herein follows Paterson et al (2010character 3)
9 Four or five ventral eyes arranged in a subtransverseband across head shield (0) absent (1) present A bandof ventral eyes is shared by the best known species ofLeanchoiliidae The distinction between three medianeyes in Alalcomenaeus cambricus (Briggs and Collins1999) and two in Leanchoilia illecebrosa (Liu et al2007) and both Leanchoilia superlata and L persephone(GarciacuteaminusBellido and Collins 2007) is uninformative forthe taxon sample here
10 Bulge in exoskeleton accommodating dropminusshaped venminustral eyes (0) absent (1) present This character followsEdgecombe and Ramskoumlld (1999 character 6) and Patminuserson et al (2010 character 5)
11 Dorsal median eyes (0) absent (1) present Coding for anabsence of dorsal median eyes in fossil taxa considers theabsence of either paired ocelli or a median eye tubercleand the character is scored as a chelicerate autapomorphy(see Cotton and Braddy 2004 character 24)
12 Width of doublure on cephalic shield (0) narrow or modminuserately wide (1) entire ventral side of head covered byexpansion of doublure The interpretation of state 1 inSidneyia and Squamacula is discussed by Cotton andBraddy (2004) and Paterson et al (2010 characters 12and 29)
13 Mouth covered by hypostome (0) absent (1) present Noldquoshort great appendagerdquo arthropods have been shown topossess a hypostome and a reconstruction of a smalllabrum in Alalcomenaeus (Briggs and Collins 1999) wasadmittedly conjectural The absence of a conspicuoushypostome contrasts with its ubiquitous presence in laminusmellipedians and several putative chelicerateminusallied taxathat have antennae (such as aglaspidids and Emeralminusdella) Chelicerates have a small epistomeminuslabrum (Bitschand Bitsch 2007) but like Cotton and Braddy (2004character 27 state 4) we code a hypostomal sclerite aslacking Kwanyinaspis has a large hypostome similar tothat found in aglaspidids (photograph of the holotype ofKwanyinaspis maotianshanensis shown to us by JavierOrtegaminusHernaacutendez [personal communication 2010])
14 Tergite articulations (0) extensive overlap of tergites (1)edge to edge articulations This character follows Edgeminuscombe and Ramskoumlld (1999 character 18) and Patersonet al (2010 character 20)
15 Number of trunk segments (0) nine (1) 11 (2) 12 (3)13 (4) 20 (5) 21 or 22 Coding is confined to taxa that do
not possess a pygidium because the precise number ofsomites associated with pygidia can sometimes not bedetermined with confidence Codings for Xiphosura andEurypterida use opisthosomal somite numbers followingShultz (2007 character 95)
16 Exoskeletal boundaries of anterior trunk segments (0)boundaries transverse or reflexed posterolaterally (1) reminusflexed anterolaterally This character follows Edgecombeand Ramskoumlld (1999 character 19) and Paterson et al(2010 character 21)
17 Trunk tergites with serrate lateral margins (0) absent (1)present Liu et al (2007 fig 2C) documented a row ofmarginal spines on the tergites of Leanchoilia illeceminusbrosa a featured shared by Leanchoilia superlata (egGarciacuteaminusBellido and Collins 2007 pl 1 7) but not otherterminals coded here
18 Differentiation in width of trunk tergites (0) trunk withgradational narrowing posteriorly (1) abrupt narrowingat boundary between prothorax and opisthothorax Anabrupt contraction in width between the boundary ofthe prominus and opisthothorax is observed in Emeraldellaand Sidneyia Yohoia has been reconstructed as having apronounced break in the shape of the trunk tergites beminushind segment 10 (Whittington 1974 textminusfig 2) thepoint at which the paratergal folds lose their spinose tipsand appendages are lacking Specimens preserved indorsal aspect (Whittington 1974 pl 10) suggest that thewidth of the opisthothorax is rather less abruptly narminusrowed and we code Yohoia as state 0
19 Appendages of posterior trunk segments (0) all trunktergites bearing appendages (1) one or more posteriortrunk segments lacking appendages Coding correspondsto Cotton and Braddyrsquos (2004) character 43 ldquopostabdominusmen of segments lacking appendagesrdquo
20 Paired tergal carinae (0) absent (1) present Liu et al(2007 265 fig 2F) recorded the presence of tergalcarinae in Leanchoilia illecebrosa shared with L superminuslata (eg Bruton and Whittington 1983 pl 17 106GarciacuteaminusBellido and Collins 2007 pl 1 5)
21 Pygidium (0) absent (1) present Coding of the pygiminusdium in many terminals herein was discussed by Paterminusson et al (2010 character 22)
22 Telson (0) absent (1) present Following Hendricks andLieberman (2008) a telson is a postminussegmental terminalstructure that lacks appendages Dunlop and Selden (1997)considered a telson to be absent in Cheloniellon but weidentify the conical projection posterior to the ultimate legpair and cerci (Stuumlrmer and Bergstroumlm 1978 figs 2 3 1011) as being most consistent with an identity as a telson
23 Telson shape (0) styliform (1) flapminuslike (2) lanceolate(3) short conical The outline of the telson in Haikouminuscaris ercaiensis is incompletely known but the mostcomplete specimen (Chen 2004 fig 472) constrains it aseither state 1 or state 2 A partial uncertainty coding isthus employed A unique state for Cheloniellon (state 3)is based on the interpretation in character 22
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 395
24 Telson fringed with spines (0) spines absent (1) spinespresent Presence of telson spines was introduced as a charminusacter by Hendricks and Lieberman (2008 character 34)
25 Paired flaps lateral to telson (0) absent (1) present Thischaracter describes the postminusventral plates of several taxa(see Edgecombe and Ramskoumlld 1999 character 25) Cotminuston and Braddy (2004 character 54) raised the possibilitythat the cerci of Cheloniellon could be homologous withpostminusventral plates (see also Hou and Bergstroumlm 1997 93)because they are similarly paired structures derived fromthe segment immediately anterior to the telson We havenot coded for this homology because the cerci of Chelominusniellon originate dorsally rather than ventrally
26 Midgut glands with submillimetric lamellae (0) absent(1) present Midgut glands in some stem group arthrominuspods and early crown group arthropods are metamericreniform organs with a characteristic lamellar structure(Butterfield 2002) Their morphology is identical inLeanchoilia and Oestokerkus notably including groovesthat radiate from a ventral median node (Figs 3A1 6A)The gut of Alalcomenaeus cambricus is more tubular(Briggs and Collins 1999 textminusfigs 10 11) and thoughan anterior six segments may display phosphatised latminuseral projections that likely represent gut diverticula (Wilminusson 2006) they have not been shown to have the detailedmorphology of Leanchoiliaminustype midgut glands Butterminusfield (2002 164) cited a figure from Bruton (1981 fig96) in support of ldquolamellarrdquo midgut glands in the cephaminuslon of Sidneyia Like Bruton (1981) in his description ofthe specimen we consider it likely that the structures inquestion are indeed gut diverticula but there is no basisfor identifying them more precisely as the reniformlamellar structures coded under this character
27 Exopod on first postminusantennal first postminusfrontal appendminusage segment (0) absent (1) present This character reminuslates to Cotton and Braddyrsquos (2004) character 3
28 Division of exopod (0) exopod an undivided flap (1)exopod differentiated into proximal and distal lobes (2)small flap with radiating lamellae (3) pediform exopoddivided into ca 6 articles Identification of the exopod inXiphosura follows Suzuki and Bergstroumlm (2008) Thischaracter groups conditions of the exopod shaft coded asindependent characters by Cotton and Braddy (2004)eg state 0 corresponds to their character 16 and state 1to their character 13 Our coding permits transformationbetween any two states without an extra implicit step
29 Exopod setae (0) short spiniform setae (1) long flatminustened tapering distally with slight separation (2)lamellar setae (flattened imbricating) (3) short finesetae A distinction is made between lamellipedian setaewhich are imbricated (state 2) and the pointed more disminustinctly separated and variably spiniform exopod setae ofleanchoiliids (state 1) This distinction is consistent withthe identification of megacheirans as nonminuslamellipedians(eg Hou and Bergstroumlm 1997 Bergstroumlm and Hou2005) Although Cotton and Braddy (2004 character 18)
regarded the form of the setae in Yohoia to be uncertainwe regard specimens figured by Whittington (1974 pl3 3) as conforming to the leanchoiliid state Squamaculadiffers from other taxa in its shorter spiniform setae(state 0) (Zhang et al 2004 textminusfig 1A B) Offacolushas longer spiniform projections on the distal articles ofthe exopod that have been variably identified as eitherspines or setae (Sutton et al 2002 fig 4b) but theirhomology with the setae that splay regularly along themargin of a flapminuslike exopod (eg states 0ndash3) is uncerminustain This and the following character for exopod setaeare left as ldquordquo for Offacolus A homology between thegill lamellae of Xiphosura and exopod setae of lamellipeminusdians has a long history (see Cotton and Braddy 2004character 18) We follow Suzuki and Bergstroumlm (2008)in identifying the xiphosuran gills as outgrowths of thebasis rather than of the exopod this perspective negates ahomology with the exopod setae of Lamellipedia Thepolymorphic coding for Emeraldella reflects the twoseparate states (1+2) displayed by the exopods in thetrunk (see Bruton and Whittington 1983 pl 8 45 47Edgecombe and Ramskoumlld 1999 fig 11)
30 Differentiation of setae along exopod (0) uniform orgradationally variable setae along length of exopod (1)lamellar setae confined to proximal lobe with distal lobebearing slender setae
31 Endopod segmentation (distal to coxa) (0) six or sevenpodomeres and terminal claw or telotarsal spurs (1)eight or more podomeres and terminal claw (2) ca 15podomeres State 1 is formulated to describe the segmenminustation of the leanchoiliid leg (GarciacuteaminusBellido and Collins2007 textminusfig 6 Liu et al 2007 fig 5) in contrast to thereduced number of podomeres in other euarthropods(Boxshall 2004) State 2 characterises Fortiforceps Apartial uncertainty coding is employed for Yohoia inwhich the precise number of podomeres is undeterminedand could be either state 0 or 1 (Whittington 1974 pl 74) but the spacing of the preserved podomere boundariesnegates the possibility of state 2
32 Endopods on anterior postminuscephalic segments (0) welldeveloped (1) rudimentary or lacking Following Cottonand Braddy (2004 character 11) the absence of trunkendopods in Yohoia (Whittington 1974) is coded as a puminustative homologue with the suppression of postminuscephalicendopods in Chelicerata
ResultsAs a convention for depicting the trees networks are rootedon Squamacula which separates artiopodan Lamellipedia(sensu Hou and Bergstroumlm 1997) from a group that includesMegacheira Chelicerata and Xenopoda all members ofwhich possess a telson
Under equal weights 14 shortest cladograms have a lengthof 77 steps with a Consistency Index of 064 and Retention Inminus
396 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
dex of 077 The strict consensus of these trees with supportvalues is shown in Fig 8 Leanchoiliidae is unambiguouslysupported in all 14 cladograms by four apomorphic characterstwo limbminusbearing cephalic segments posterior to the frontalappendage (character 3 state 1) long spinose projections onthe distal part of the terminal three podomeres of the frontalappendage that bear flagella (character 6 state 1) 11 trunksegments (character 15 state 1) and division of the exopodsinto two lobes (character 28 state 1) The arrangement of fouror five eyes in a subtransverse band across the cephalic shieldis unique to Leanchoiliidae (character 9 state 1) but is not anunambiguous autapomorphy of the group because the state isuncertain in close relatives notably Haikoucaris It is apominusmorphic for Leanchoiliidae only under particular optimisaminustions (Delayed Transformation)
The internal relationships of Leanchoiliidae are stableacross all 14 shortest cladograms Leanchoilia is united by thelanceolate telson (character 23 state 2) a unique unambiguminusous autapomorphy Within Leanchoilia a sister group relaminustionship between L superlata and L illecebrosa is well supminusported receiving a Bremer support of 3 and jackknife freminusquency of 86 These two species share four synapomorminusphies a medially pointed anterior cephalic margin (character2 state 1) flagella of the great appendage extending the lengthof the body (character 7 state 1) serrate lateral margins on thetrunk tergites (character 17 state 1) and paired tergal carinae(character 20 state 1) As such a species from Chengjiang (Lillecebrosa) and the Burgess Shale (L superlata) are moreclosely related to each other than are the two Burgess Shalecongeners a relationship that weakens the possibility that theoften cominusoccurring L superlata and L persephone could besexual dimorphs (GarciacuteaminusBellido and Collins 2007) ratherthan separate species The union of Leanchoilia and Oestominuskerkus is unambiguously supported by the shared midgutgland structure of both genera (character 26 state 1)
All shortest cladograms resolve Haikoucaris Yohoiaand Fortiforceps in that order as progressively less closelyrelated to Leanchoiliidae The union of these three taxa withLeanchoiliidae yields a clade that corresponds to Megacheirasensu Hou and Bergstroumlm (1997) apart from the ambiguousplacement of Jianfengia In half of the 14 shortest cladominusgrams Jianfengia is sister to the other megacheirans yieldminusing a monophyletic Megacheira ss In the remaining cladominusgrams Jianfengia is sister to Xenopoda sensu Hou andBergstroumlm (1997) Aglaspis and Chelicerata Haikoucarisand Leanchoiliidae are united by a frontal appendage comminusposed of two spineminusbearing articles in addition to the movminusable finger (character 5 state 1) Yohoia groups with thesetaxa based on spines fringing the telson (character 24 state1) and Fortiforceps is united with them at a broader levelbased on the flapminuslike telson shape (character 23 state 1)When Jianfengia is placed as sister to the other megacheiminusrans the entire clade is supported by the raptorial frontal apminuspendage (character 4 state 1) and the lack of a large hypominusstomal shield (character 13 state 0) In this set of cladominusgrams these features are nonminushomologous between Megaminus
cheira and Chelicerata whereas in some cladograms with thealternative resolution of Jianfengia as closer to cheliceratescharacter 4 is optimised as homologous in Megacheira andChelicerata
Reanalysis of the data with implied weights finds two optiminusmal trees (Fig 9) Relationships within Leanchoiliidae underequal weights are completely stable to reweighting as is theresolution of Haikoucaris as sister to Leanchoiliidae As suchthe character optimisations described above for the leanchoiminusliids are depicted the same in Fig 9 The main difference beminustween equal and implied weights involves the placement ofChelicerata Under equal weights (Fig 8) chelicerates are alminuslied to Xenopoda (here Emeraldella + Sidneyia + Chelominusniellon sensu clade L in Hou and Bergstroumlm 1997 fig 88)and Aglaspidida A clade composed of Aglaspidida Xenominuspoda and Chelicerata was similarly identified by Hou andBergstroumlm (1997 fig 88 their clade K) In our analysis thisgroup consistently finds support from one or more posteriortrunk segments lacking appendages (character 19 state 1) anddepending on alternative internal resolutions it is variably supminusported by dorsallyminuspositioned sessile eyes (character 8 state 1)and the absence of exopods on the first postminusantennalpostminuscheliceral limb (character 27 state 1) Under implied weightshowever chelicerates are more closely allied to Megacheirathan to aglaspidids or xenopodans (Fig 9)
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 397
2
582
76
Chelicera-morpha
Megacheira
62
2
53
50Leanchoiliidae
3
86
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
EurypteridaChelicerata
Fortiforceps
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia hanceyi
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
Fig 8 Strict consensus of 14 shortest cladograms (77 steps) based on charminusacter data in Table 1 Numbers above nodes are Bremer support those beminuslow nodes are jackknife frequencies gt50 Cheliceramorpha correspondsto a clade with the same composition in Cotton and Braddy (2004)
The major groupings in Fig 9 correspond closely to thoseidentified by Cotton and Braddy (2004) Notably the cladeuniting Aglaspis Xenopoda Offacolus Chelicerata Jianminusfengia and other Megacheira corresponds to their ldquoClade 3rdquo(Cotton and Braddy 2004 fig 8) for which they proposedthe name Cheliceramorpha The clade uniting CheloniellonAglaspis Sidneyia and Emeraldella in Fig 9 corresponds totheir ldquoClade 5rdquo and the clade that groups Megacheira andChelicerata is equivalent to their ldquoClade 4rdquo Their analysisdid not identify megacheirans as a monophyletic groupChelicerata was nested within the group and among taxaconsidered here Yohoia was resolved closer to cheliceratesthan to Leanchoiliidae In contrast Yohoia was regarded byBergstroumlm and Hou (2005) as more closely allied to Leanminuschoiliidae than to chelicerates a megacheiran grouping morecompatible with our cladograms The analysis of ldquogreat apminuspendage arthropodsrdquo by Vannier et al (2009) identified a
monophyletic Leanchoiliidae but a polyphyletic Megacheira(to the exclusion of Haikoucaris and Jianfengia) Howevertheir resolution of Fortiforceps and even more so Yohoia asthe closest relatives of Leanchoiliidae is as in our trees (apartfrom the different position of Haikoucaris)
In summary a major controversy in the systematics of theldquoshort great appendagerdquo arthropods remains whether thechelicerates are nested within the group or are instead moreclosely allied to taxa conventionally identified as lamelliminuspedians specifically Xenopoda and Aglaspidida We findthat both of these results are compatible with the availabledata depending on character weighting protocols Noneminustheless the broader megacheiranminuschelicerate issue does notcloud the status of leanchoiliids as a clade or the identificaminustion of their closest relatives (ie Haikoucaris and Yohoia)and the internal relationships of Leanchoiliidae are stableand supported by compelling characters
398 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
Eurypterida
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
2 7 17 20
3
2
1 1 1
26
1
281563
1 1 1 1
5
1
24
1
23 19 20 27 32
28Clade 4
1 0
4 13 11 27 0
154 1 1
3 19 32Clade 3
2 3
2 28
0
3 1 121
1
22
Clade 5
1 0
19 27
1
25
15 28 301 1
2312
1 150 1 3 3
15 16 2923
18
1 1
10 16
1 1
1 8
1
14
3
3
28 30
1 0
5
15
1 1
1
1
22
Fortiforceps
23
1 1 0 1
1 1
Leanchoilia hanceyi
2
2
1
1
Fig 9 Characters optimised on one of two shortest cladograms with imminusplied weights using a concavity constant k = 3 Black squares are nonminushomoplastic changes white squares homoplastic changes all changes areunambiguous Branches without strict support collapsed after optimisingcharacters Numbers for characters (above the branches) and states (belowthe branches) correspond to Table 1 ldquoClades 3ndash5rdquo correspond to groups sonumbered by Cotton and Braddy (2004 fig 8)
AcknowledgementsAccess to the field area has been most kindly allowed by the Buck famminusily We thank our collaborators in Emu Bay Shale research Jim Jago(University of South Australia Mawson Lakes) Jim Gehling andMike Lee (South Australian Museum Adelaide) for tremendous supminusport and advice as well as Mike Gemmell Dennis Rice Allison DaleyAaron Camens John Laurie Glenn Brock Pierre Kruse and Trevorand Jenny Worthy (all South Australian Museum) for their assistancein the field and Natalie Schroeder MaryminusAnne Binnie and RondaAtkinson (all South Australian Museum) for organisational support inthe lab and in the field Javier OrtegaminusHernaacutendez (University of Camminusbridge Cambridge UK) advised us on Kwanyinaspis and the journalrsquosreferees provided useful feedback TNT is available courtesy of theWilli Hennig Society Funding has been provided by an AustralianResearch Council Linkage grant (LP0774959) to the University ofAdelaide with generous financial assistance from Beach Energy Ltdand the South Australian Museum SeaLink provided logistical supminusport DGBrsquos travel to Australia has been supported by grants from theSpanish Research Council (CSIC PA 1001105) the Complutense Uniminusversity of Madrid and the Spanish Ministry of Education and Science(CGL2006minus12245BTE CGL2009minus07073 and RYC2007minus00090)
ReferencesBergstroumlm J and Hou XminusG 2005 Early Palaeozoic lamellipedian arthrominus
pods In S Koenemann and RA Jenner (eds) Crustacea and Arthrominuspod Relationships Crustacean Issues 16 73ndash93
Bitsch J and Bitsch C 2007 The segmental organization of the head reminusgion in Chelicerata a critical review of recent studies and hypothesesActa Zoologica 88 317ndash335
Bousfeld EL 1995 A contribution to the natural classification of Lower andMiddle Cambrian arthropods food gathering and feeding mechanismsAmphipacifica 2 3ndash34
Boxshall GA 2004 The evolution of arthropod limbs Biological Reviews79 253ndash300
Bremer K 1994 Branch support and tree stability Cladistics 10 295ndash304Briggs DEG and Collins D 1999 The arthropod Alalcomenaeus cambricus
Simonetta from the Middle Cambrian Burgess Shale of British ColumbiaPalaeontology 42 953ndash977
Briggs DEG and Robison RA 1984 Exceptionally preserved nonminustrilobite arthropods and anomalocaridids from the Middle Cambrian ofUtah University of Kansas Paleontological Contributions 111 1ndash23
Briggs DEG Lieberman BS Hendricks JR Halgedahl SL andJerrard RD 2008 Middle Cambrian arthropods from Utah Journal ofPaleontology 82 238ndash254
Bruton DL 1981 The arthropod Sidneyia inexpectans Middle CambrianBurgess Shale British Columbia Philosophical Transactions of theRoyal Society of London Series B 295 619ndash656
Bruton DL and Whittington HB 1983 Emeraldella and Leanchoiliatwo arthropods from the Burgess Shale Middle Cambrian British Cominuslumbia Philosophical Transactions of the Royal Society of London Seminusries B 300 553ndash582
Budd GE 2002 A palaeontological solution to the arthropod head probminuslem Nature 417 271ndash275
Butterfield NJ 2002 Leanchoilia guts and the interpretation of threeminusdiminusmensional structures in Burgess Shaleminustype fossils Paleobiology 28155ndash171
Chen JminusY 2004 The Dawn of Animal World [in Chinese] 366 pp JiangScience and Technology Press Nanjing
Chen JminusY Waloszek D and Maas A 2004 A new ldquogreatminusappendagerdquo arminusthropod from the Lower Cambrian of China and homology of cheliceratechelicerae and raptorial anterominusventral appendages Lethaia 37 3ndash20
Cotton TJ and Braddy SJ 2004 The phylogeny of arachnomorph arthrominuspods and the origin of the Chelicerata Transactions of the Royal Societyof Edinburgh Earth Sciences 94 169ndash193
Daley AC Budd GE Caron JminusB Edgecombe GD and Collins D2009 The Burgess Shale anomalocaridid Hurdia and its significancefor early euarthropod evolution Science 323 1597ndash1600
Dunlop JA 2006 New ideas about the euchelicerate stemminuslineage In CDeltshev and P Stoev (eds) European Arachnology 2005 Acta Zoominuslogica Bulgarica (Supplement 1) 9ndash27
Dunlop JA and Selden PA 1997 The early history and phylogeny of thechelicerates In RA Fortey and RH Thomas (eds) Arthropod Relaminustionships Systematics Association Special Volume 55 221ndash235
Edgecombe GD and Ramskoumlld L 1999 Relationships of CambrianArachnata and the systematic position of Trilobita Journal of Paleonminustology 73 263ndash287
Farris JS Albert VA Kaumlllersjouml M Lipcomb D and Kluge AG1996 Parsimony jackknifing outperforms neighborminusjoining Cladistics12 99ndash124
GarciacuteaminusBellido DC and Collins D 2007 Reassessment of the genusLeanchoilia (Arthropoda Arachnomorpha) from the Middle CambrianBurgess Shale British Columbia Canada Palaeontology 50 693ndash709
GarciacuteaminusBellido DC Paterson JR Edgecombe GD Jago JB GehlingJG and Lee MSY 2009 The bivalved arthropods Isoxys and Tuzoiawith softminuspart preservation from the lower Cambrian Emu Bay ShaleLagerstaumltte (Kangaroo Island Australia) Palaeontology 52 1221ndash1241
Goloboff PA 1993 Estimating character weights during tree searchCladistics 9 83ndash91
Goloboff PA Farris JS and Nixon KC 2008 TNT a free program forphylogenetic analysis Cladistics 24 774ndash786
Harzsch S and Hafner G 2006 Evolution of eye development in arthrominuspods phylogenetic aspects Arthropod Structure amp Development 35319ndash340
Hendricks JR and Lieberman BS 2008 New phylogenetic insights intothe Cambrian radiation of arachnomorph arthropods Journal of Paleminusontology 82 585ndash594
Hou XminusG and Bergstroumlm J 1997 Arthropods of the Lower CambrianChengjiang fauna southwest China Fossils and Strata 45 1ndash116
Jago JB Zang W Sun X Brock GA Paterson JR and SkovstedCB 2006 A review of the Cambrian biostratigraphy of South Austraminuslia Palaeoworld 15 406ndash423
Liu Y Hou XminusG and Bergstroumlm J 2007 Chengjiang arthropod Leanminuschoilia illecebrosa (Hou 1987) reconsidered GFF 129 263ndash272
Luo H Hu S Chen L Zhang S and Tao Y 1999 Early CambrianChengjiang fauna from Kunming region China [in Chinese] 129 ppYunnan Science and Technology Press Kunming
Luo H Hu S Zhang S and Tao Y 1997 New occurrence of the EarlyCambrian Chengjiang fauna in Haikou Kunming Yunnan Provinceand study on Trilobitoidea Acta Geologica Sinica 71 122ndash132
Nixon KC 2002 WINCLADA Version 19908 Cornell University IthacaNY
Orr PJ Siveter DJ Briggs DEG Siveter DJ and Sutton MD 2000A new arthropod from the Silurian KonservatminusLagerstaumltte of Hereminusfordshire Proceedings of the Royal Society Series B Biological Sciminusences 267 1497ndash1504
Paterson JR and Brock GA 2007 Early Cambrian trilobites fromAngorichina Flinders Ranges South Australia with a new assemblagefrom the Pararaia bunyerooensis Zone Journal of Paleontology 81116ndash142
Paterson JR and Jago JB 2006 New trilobites from the Lower CambrianEmu Bay Shale Lagerstaumltte at Big Gully Kangaroo Island South Ausminustralia Memoirs of the Association of Australasian Palaeontologists 3243ndash57
Paterson JR Edgecombe GD GarciacuteaminusBellido DC Jago JB andGehling JG 2010 Nektaspid arthropods from the lower CambrianEmu Bay Shale Lagerstaumltte South Australia with a reassessment oflamellipedian relationships Palaeontology 53 377ndash402
Paterson JR Jago JB Gehling JG GarciacuteaminusBellido DC Edgecombe
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 399
GD and Lee MSY 2008 Early Cambrian arthropods from the EmuBay Shale Lagerstaumltte South Australia 319ndash325 In I Raacutebano RGozalo and D GarciacuteaminusBellido (eds) Advances in Trilobite ResearchCuadernos del Museo Geominero 9 319ndash325
Shultz JW 2007 A phylogenetic analysis of the arachnid orders based onmorphological characters Zoological Journal of the Linnean Society150 221ndash265
Stein M 2010 A new arthropod from the Early Cambrian of North Greenminusland with a ldquogreat appendagerdquominuslike antennula Zoological Journal ofthe Linnean Society 158 477ndash500
Stuumlrmer W and Bergstroumlm J 1978 The arthropod Cheloniellon from theDevonian Hunsruumlck Shale Palaumlontologische Zeitschrift 52 57ndash81
Sutton MD Briggs DEG Siveter DJ Siveter DJ and Orr PD2002 The arthropod Offacolus kingi (Chelicerata) from the Silurian ofHerefordshire England computer based morphological reconstrucminustions and phylogenetic affinity Proceedings of the Royal Society B Biminusological Sciences 269 1195ndash1203
Suzuki Y and Bergstroumlm J 2008 Respiration in trilobites a reevaluationGFF 130 211ndash229
Vannier J GarciacuteaminusBellido DC Hu XminusX and Chen AminusL 2009 Arthrominuspod visual predators in the early pelagic ecosystem evidence from the
Burgess Shale and Chengjiang biotas Proceedings of the Royal SocietyB Biological Sciences 276 2567ndash2574
Whittington HB 1974 Yohoia Walcott and Plenocaris n gen arthropodsfrom the Burgess Shale Middle Cambrian British Columbia Geologiminuscal Survey of Canada Bulletin 231 1ndash21
Wills MA Briggs DEG Fortey RA Wilkinson M and SneathPHA 1998 An arthropod phylogeny based on fossil and recent taxaIn GD Edgecombe (ed) Arthropod Fossils and Phylogeny 33ndash105Columbia University Press New York
Wilson LA 2006 Food for thought a morphological and taphonomic studyof fossilised digestive systems from Early to Middle Cambrian taxa275 pp Unpublished PhD thesis University of Cambridge Cambridge
Zhang XminusG and Hou X 2007 Gravitational constraints on the burial ofChengjiang fossils Palaios 22 448ndash453
Zhang XminusL Han J Zhang Z Liu H and Shu D 2004 Redescription ofthe Chengjiang arthropod Squamacula clypeata Hou and Bergstroumlm fromthe Lower Cambrian Southminuswest China Palaeontology 47 605ndash617
Zhao Y Zhu M Babcock LE Yuan L Parsley RL Peng J YangX and Wang Y 2005 Kaili Biota a taphonomic window on diversifiminuscation of metazoans from the basal Middle Cambrian Guizhou ChinaActa Geologica Sinica 79 751ndash765
400 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
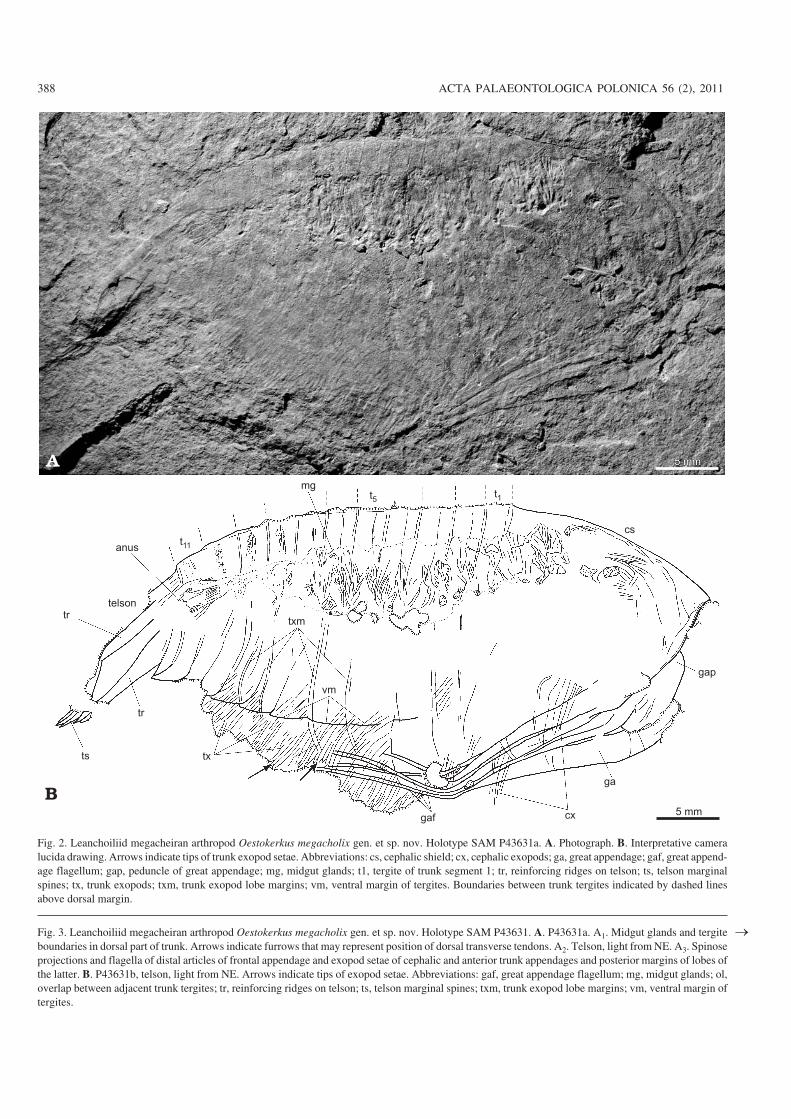
388 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
55 mmmm
5 mm
anus
telsontr
tr
t11
t5
mg
txm
t1
cs
gap
vm
ga
gaf
txts
cx
Fig 2 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov Holotype SAM P43631a A Photograph B Interpretative cameralucida drawing Arrows indicate tips of trunk exopod setae Abbreviations cs cephalic shield cx cephalic exopods ga great appendage gaf great appendminusage flagellum gap peduncle of great appendage mg midgut glands t1 tergite of trunk segment 1 tr reinforcing ridges on telson ts telson marginalspines tx trunk exopods txm trunk exopod lobe margins vm ventral margin of tergites Boundaries between trunk tergites indicated by dashed linesabove dorsal margin
Fig 3 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov Holotype SAM P43631 A P43631a A1 Midgut glands and tergiteboundaries in dorsal part of trunk Arrows indicate furrows that may represent position of dorsal transverse tendons A2 Telson light from NE A3 Spinoseprojections and flagella of distal articles of frontal appendage and exopod setae of cephalic and anterior trunk appendages and posterior margins of lobes ofthe latter B P43631b telson light from NE Arrows indicate tips of exopod setae Abbreviations gaf great appendage flagellum mg midgut glands oloverlap between adjacent trunk tergites tr reinforcing ridges on telson ts telson marginal spines txm trunk exopod lobe margins vm ventral margin oftergites
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 389
SAM P45904a b (unfigured) poorlyminuspreserved cephalic shield trunkwith wellminuspreserved midgut glands proximal part of telson SAMP45905ab (unfigured) cephalic shield lacking great appendages butexposing an eye trunk with incomplete telson SAM P45906a b (unfigminusured) great appendages with spinose projections oriented anteriorlytrunk preserved to end of midgut glands SAM P45908a b (unfigured)cephalic shield great appendages with flagella most of trunk (telsonmissing) showing midgut glands and exopod setae SAM P45909 (unminusfigured) small specimen with great appendages and series of mineralminusised midgut glands SAM P45910a b (unfigured) incomplete great apminuspendages oriented anteriorly trunk with complete series of midgutglands but indistinct appendages
Type locality Buck Quarry Big Gully (Fig 1)
Type horizon Emu Bay Shale (Cambrian Series 2 Stage 4)
DiagnosismdashAs for genus
DescriptionmdashCephalic shield 36 length of the trunk sagitminustally dorsal margin of cephalic shield gently convex in lateralview anteromedian margin inferred to be evenly rounded(based on comparison with lateral versus dorsal preservation ofBurgess Shale Leanchoilia and Alalcomenaeus) lacking an upminusturned median process Eye visible in SAM P54167 (Fig 4A)and P45905 relatively large (20 sagittal length of cephalon)exposed under anterolateral part of cephalic shield (Fig 4A2)
390 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Fig 4 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov SAM P45167 A P45167a A1 cephalic shield and most of trunkA2 detail of cephalic shield B P45167b A2 and B light from NE Overlap between adjacent tergites offset on left (Lol) and right (Rol) sides of bodyBoundaries between cephalic shield and anterior trunk tergites delimited in B by dashed lines Arrows mark positions of inferred dorsal transverse tendonsAbbreviations cn1ndash2 cephalic endopods cs cephalic shield gap peduncle of great appendage ol overlap between adjacent trunk tergites t1ndasht3 tergitesof trunk segments 1ndash3 tn trunk endopods
Fig 5 Leanchoiliid megacheiran arthropod Oestokerkus megacholix genet sp nov A SAM P 15531 B SAM P43630b Light from NE Abbreviaminustions cs cephalic shield ga great appendage gap peduncle of great apminuspendage mg midgut glands ol overlap between adjacent trunk tergites txtrunk exopods
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 391
Great appendage robust proportions of peduncle and disminustal spineminusbearing articles as in other members of Leanchoiliminusidae more commonly preserved with posterior orientation
but anteriorly oriented in SAM P45906 and SAM P45910Each of the three long spinose projections terminating at thesame length the projections evenly tapering distally with
392 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Fig 6 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov SAM P45168 A head and anterior part of trunk B Detail of spinoseprojections of great appendages and excretory organs (sensu Bruton and Whittington 1983) Abbreviations cs cephalic shield eo excretory organ gagreat appendage mg midgut glands tx trunk exopods
margins straight for much of their length or slightly curvedterminally distal spinose projection distinctly stouter at itsbase than the two more proximal projections and more markminusedly tapering along its length Dark irregularminusshaped low reminuslief lump on basal spineminusbearing article of great appendagesof SAM P45168 (eo in Fig 6) and SAM P45908 correminussponding to excretory organ of Bruton and Whittington(1983) Long flagella extending from each of the threespinose projections seen only in SAM P43631 (Figs 23A3) of similar narrow width along its preserved extent anminusnulations indistinguishable
Trunk of 11 segments of even length Maximum height oftrunk relative to length (excluding telson) ca 39 bodydeepest at tergite 4 Trunk tergite boundaries marked as narminusrow ridges in several specimens corresponding to narrowband of overlap between adjacent tergites (ol in Figs 2 3A14A1 B) Some specimens show tergites to be bisected by anarrow but sharp furrow of uncertain identity (eg possiblyimpression of dorsal transverse tendon cf Stein 2010 fig12B C E) (arrows in Figs 3A1 4B)
Exopods with fringe of long setae on cephalic and trunkappendages (Fig 2) setal fringe gradually shortening fromabout the fourth trunk segment Setae project ventrally bymore than 30 height of tergites in deepest part of trunk(Fig 2B) setae flattened narrow and parallelminussided alongmost of their length tapering near their distal ends to termiminusnate as a pointed tip (Figs 2B 3A3 arrows) Endopods exminusposed only in SAM P45167 (Fig 4) two pairs associatedwith cephalon (Fig 4B) those of anterior segments of trunkof similar morphology to those in cephalon all slender lackminusing obvious endites podomere boundaries unrecognisable
Midgut glands occupy a large extent of the dorsoventralheight of the trunk two midgut glands in cephalon of similarsize to those in each of the first seven segments of the trunk(Figs 3A1 5A 6A) all developed as radiating grooves emaminusnating from a globular structure at their ventral edge as in
Leanchoilia superlata (GarciacuteaminusBellido and Collins 2007 pl2 6) Foregut trace in SAM P43631 (Fig 2) indicates that gutcurves ventrally in anterior part of cephalon mouth regionnot preserved Terminal part of hindgut (in trunk segment 11)with closely spaced longitudinal lineations (Fig 2) that apminusparently correspond to folds in the gut wall hindgut termiminusnating at base of telson
Telson most completely preserved in holotype (SAMP43631 Figs 2B 3A2 B) In this specimen the telson likerest of body compressed in lateral aspect Telson shows meminusdian and marginal reinforcing ridges connecting a flat trianminusgular surface this surface appears to be partially broken inSAM P43631 showing underlying surface of other (left) halfof telson Lateral preservation of specimen implies that topcorresponds to an axial structure and bottom to a marginalone 30ndash35 angle in posterior edge of vane connecting thesetwo structures requires that in dorsal aspect telson would apminuspear forked and widening distally Five stout pointed dorminussally curved marginal spines attached to outer posterior edgeon each side of telson
Phylogenetic analysis ofLeanchoiliidaeDetermining the systematic position of Oestokerkus demandsan appraisal of the relationships of Leanchoiliidae as a wholeCladistic analysis including the bestminusknown members ofMegacheira including leanchoiliids has been conducted byCotton and Braddy (2004) and Vannier et al (2009) The taxominusnomic sampling used by Cotton and Braddy (2004) includedseveral megacheiran genera representative chelicerates anumber of taxa representing groups that have been proposedas possible chelicerate relatives (eg Sidneyia AglaspisCheloniellon) in other analyses (Dunlop and Selden 1997Hou and Bergstroumlm 1997) and members of the major lineagesof the Artiopoda sensu Hou and Bergstroumlm (1997) ie trilominusbiteminusallied lamellipedians Because artiopodan interminusrelationminusships have remained quite stable through a series of phylogenminusetic analyses (Edgecombe and Ramskoumlld 1999 Cotton andBraddy 2004 Hendricks and Lieberman 2008 Paterson et al2010) and artiopodans are now often regarded as less closelyrelated to Megacheira than is Chelicerata (Chen et al 2004Cotton and Braddy 2004 Dunlop 2006) we have reduced thesampling of Artiopoda in the present analysis To augment thediversity of ldquoshort great appendagerdquo arthropods Haikoucarisis added based on the description of H ercaiensis by Chen etal (2004) Offacolus (Orr et al 2000 Sutton et al 2002) hasbeen allied to Chelicerata and is included as a terminal Beminuscause the present analysis is principally concerned with relaminustionships of and within Leanchoiliidae sensu Liu et al (2007)we follow Vannier et al (2009) in sampling Alalcomenaeusand Leanchoilia at the species level Within Chelicerata Cotminuston and Braddy (2004) and Hendricks and Lieberman (2008)sampled ancestral states in Xiphosura by coding the Devonian
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 393
Fig 7 Reconstruction of Oestokerkus megacholix gen et sp nov in lateralview Total length and annulations on the flagella and positions of podominusmere boundaries of the frontal appendage are based on Leanchoilia superminuslata (Bruton and Whittington 1983)
synziphosurine Weinbergina Our coding for Xiphosura alsoemploys information from extant xiphosurans because theysupply more reliable information on the homology of the partsof the trunk appendages (Suzuki and Bergstroumlm 2008) and eyeanatomy (Harzsch and Hafner 2006) among other charactersystems than do fossils alone
Our character set extracts characters from the matrix ofCotton and Braddy (2004) that vary in the revised taxonomicsample reformulates some characters to accommodate reminusvised homology concepts and adds new characters especiallyfor leanchoiliids In total 32 characters are included (Table 1)In this analysis the primary homology of the ldquoshort great apminuspendagerdquo and chelicera is coded (as described by character 4)and the taxon sampling broadly constrains megacheirans aschelicerate relatives (as defended by Chen et al 2004 Cottonand Braddy 2004) rather than testing the alternative that theycould be stem group Euarthropoda (Budd 2002 Bergstroumlmand Hou 2005) A test of the two alternatives involves samminuspling of the entire diversity of euarthropods and a range ofnonminuseuarthropod outgroups an exercise beyond the scope ofthis study
Characters1 Mineralised cuticle (0) absent (1) present For definiminus
tion and discussion of this character see Cotton andBraddy (2004 character 30) and Paterson et al (2010character 1)
2 Curvature of anterior margin of cephalic shield (0) rounminusded or straight (1) pointed medially (2) with multiplespinose projections This character codes for variability inthe shape of the cephalic shield in Leanchoiliidae (state 0and 1 sensu GarciacuteaminusBellido and Collins [2007 textminusfigs10 and 9 respectively]) State 2 is added to accommodateOffacolus the median spine of a series that fringes the ceminusphalic margin is not convincingly homologised with themedian projection in leanchoiliids
3 Number of cephalic somitesappendages (0) antenna orfrontal appendage only (1) antenna or frontal appendageand two limb pairs (2) antenna or frontal appendage andthree limb pairs (3) antenna or frontal appendage and fourlimb pairs (4) antenna or frontal appendage and five limbpairs Coding for lamellipedians and Aglaspis is as discussedby Paterson et al (2010 character 2) Coding for Alalcomeminusnaeus cambricus (state 1) follows Briggs and Collins (1999)Leanchoilia illecebrosa (state 2) follows Liu et al (2007)and Leanchoilia persephone and Leanchoilia superlata (state1) follows GarciacuteaminusBellido and Collins (2007)
4 Form of first cephalic appendage (0) antenna (1) anteminusriorly directed raptorial appendage with reduced numberof podomeres each of the terminal podomeres bearing aspinose process on its distal margins a geniculation beminustween spinose podomeres and peduncle This characteris formulated to identify a homology between the frontalappendages of megacheirans and the chelicera of Cheliminus
cerata and state 1 corresponds to the putative apomorminusphic detail defended by Chen et al (2004) and Cottonand Braddy (2004) Vannier et al (2009 character 8)specify another shared property of ldquoshort great appendminusagesrdquo of the megacheiran (ss) type in their characterstate ldquobent at an anglerdquo referring to the geniculation beminustween articles of the peduncle or between the peduncleand distal spiniferous articles This property is sharedwith Chelicerata
5 Number of spineminusbearing articles on frontal appendage (0)three + movable finger (1) two + movable finger (2) one+ movable finger The number of spineminusbearing articles onthe frontal appendage (exclusive of the terminal movablefinger) varies from three in Yohoia (Whittington 1974 pl6 1ndash3 pl 7 3ndash6 pl 9 4ndash6) and Fortiforceps (Hou andBergstroumlm 1997 figs 31C D 32A 33B C E) to two inHaikoucaris (Chen et al 2004 fig 2A B) and Leanminuschoiliidae or only one the last being a chelicera
6 Long spinose projections on distal part of terminal threepodomeres of frontal appendage bearing a flagellum (0)absent (1) present The morphology of the great appendminusage in Leanchoiliidae is coded here We have not codedthe length of the spinose projections and the presence ofa flagellum as two independent characters Cotton andBraddyrsquos (2004) character 6 state 2 (ldquospinesrdquo longer thanthe entire podomere series) and character 8 state 1 (presminusence of an annulated flagellum) are thus grouped as a sinminusgle trait here
394 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Table 1 Characters coded for phylogenetic analysis Polymorphismindicated by ldquo+rdquo partial uncertainty by states separated by ldquordquo Abbreviminusations A Alalcomenaeus L Leanchoilia O Oestokerkus
Squamacula 0000minus minus minus2minus0 0110 00000 10minus minus0 1000 0
Cindarella 0030minus minus minus000 0010 00000 10minus minus0 11121 00
Kuamaia 000minus minus minus001 0011 10000 10minus minus0 11121 00
Kwanyinaspis 00 001 00112 1000 1000 1121 00
Olenoides 1020minus minus minus100 0011 00000 10minus minus0 01121 00
Retifacies 0020minus minus minus000 0010 00000 11000 1020 00
Aglaspis 10(23)0minus minus minus100 00101 00010 01001 0 00
Sidneyia 0000minus minus minus000 01102 00110 01101 00020 00
Emeraldella 0040minus minus minus2minus 0 00103 00110 01001 001(1+2)1 00
A cambricus 00111 10010 00001 00000 01110 01110 10
L illecebrosa 01211 11010 00001 01001 01210 11110 10
L persephone 00111 10010 00001 00000 01210 11110 10
L superlata 01111 11010 00001 01001 01210 11110 10
L hanceyi 00 00 0001 0000 01210
O megacholix 0011 100 00001 00000 01110 1110 0
Jianfengia 0021 0minus000 00005 00000 01000 010(12)0 0
Fortiforceps 00210 0minus00 00004 00000 01100 01230 20
Yohoia 00210 0minus000 00003 00011 01110 00010 (01)1
Haikoucaris 00211 0minus00 00003 00000 01(12)0 1010 0
Cheloniellon 0040minus minus minus100 00100 10010 01300 0030 00
Offacolus 02412 0minus200 0000 00010 01000 13minus minus 01
Xiphosura 00412 0minus100 10000 0010 01000 000minus minus 01
Eurypterida 00412 0minus100 10002 00010 01000 00minus minus minus 01
7 Length of flagellate great appendage (0) flagella termiminusnating at midlength of trunk (1) flagella extending to endof body Coding is applicable to Leanchoiliidae onlydistinguishing some Leanchoilia species that resembleAlalcomenaeus in having relative short great appendages(state 0) versus the longer appendages in L superlata andL illecebrosa
8 Position of lateral facetted eyes (0) ventral (1) dorsalsessile (2) absent Variation in Lamellipedia and severalother taxa coded herein follows Paterson et al (2010character 3)
9 Four or five ventral eyes arranged in a subtransverseband across head shield (0) absent (1) present A bandof ventral eyes is shared by the best known species ofLeanchoiliidae The distinction between three medianeyes in Alalcomenaeus cambricus (Briggs and Collins1999) and two in Leanchoilia illecebrosa (Liu et al2007) and both Leanchoilia superlata and L persephone(GarciacuteaminusBellido and Collins 2007) is uninformative forthe taxon sample here
10 Bulge in exoskeleton accommodating dropminusshaped venminustral eyes (0) absent (1) present This character followsEdgecombe and Ramskoumlld (1999 character 6) and Patminuserson et al (2010 character 5)
11 Dorsal median eyes (0) absent (1) present Coding for anabsence of dorsal median eyes in fossil taxa considers theabsence of either paired ocelli or a median eye tubercleand the character is scored as a chelicerate autapomorphy(see Cotton and Braddy 2004 character 24)
12 Width of doublure on cephalic shield (0) narrow or modminuserately wide (1) entire ventral side of head covered byexpansion of doublure The interpretation of state 1 inSidneyia and Squamacula is discussed by Cotton andBraddy (2004) and Paterson et al (2010 characters 12and 29)
13 Mouth covered by hypostome (0) absent (1) present Noldquoshort great appendagerdquo arthropods have been shown topossess a hypostome and a reconstruction of a smalllabrum in Alalcomenaeus (Briggs and Collins 1999) wasadmittedly conjectural The absence of a conspicuoushypostome contrasts with its ubiquitous presence in laminusmellipedians and several putative chelicerateminusallied taxathat have antennae (such as aglaspidids and Emeralminusdella) Chelicerates have a small epistomeminuslabrum (Bitschand Bitsch 2007) but like Cotton and Braddy (2004character 27 state 4) we code a hypostomal sclerite aslacking Kwanyinaspis has a large hypostome similar tothat found in aglaspidids (photograph of the holotype ofKwanyinaspis maotianshanensis shown to us by JavierOrtegaminusHernaacutendez [personal communication 2010])
14 Tergite articulations (0) extensive overlap of tergites (1)edge to edge articulations This character follows Edgeminuscombe and Ramskoumlld (1999 character 18) and Patersonet al (2010 character 20)
15 Number of trunk segments (0) nine (1) 11 (2) 12 (3)13 (4) 20 (5) 21 or 22 Coding is confined to taxa that do
not possess a pygidium because the precise number ofsomites associated with pygidia can sometimes not bedetermined with confidence Codings for Xiphosura andEurypterida use opisthosomal somite numbers followingShultz (2007 character 95)
16 Exoskeletal boundaries of anterior trunk segments (0)boundaries transverse or reflexed posterolaterally (1) reminusflexed anterolaterally This character follows Edgecombeand Ramskoumlld (1999 character 19) and Paterson et al(2010 character 21)
17 Trunk tergites with serrate lateral margins (0) absent (1)present Liu et al (2007 fig 2C) documented a row ofmarginal spines on the tergites of Leanchoilia illeceminusbrosa a featured shared by Leanchoilia superlata (egGarciacuteaminusBellido and Collins 2007 pl 1 7) but not otherterminals coded here
18 Differentiation in width of trunk tergites (0) trunk withgradational narrowing posteriorly (1) abrupt narrowingat boundary between prothorax and opisthothorax Anabrupt contraction in width between the boundary ofthe prominus and opisthothorax is observed in Emeraldellaand Sidneyia Yohoia has been reconstructed as having apronounced break in the shape of the trunk tergites beminushind segment 10 (Whittington 1974 textminusfig 2) thepoint at which the paratergal folds lose their spinose tipsand appendages are lacking Specimens preserved indorsal aspect (Whittington 1974 pl 10) suggest that thewidth of the opisthothorax is rather less abruptly narminusrowed and we code Yohoia as state 0
19 Appendages of posterior trunk segments (0) all trunktergites bearing appendages (1) one or more posteriortrunk segments lacking appendages Coding correspondsto Cotton and Braddyrsquos (2004) character 43 ldquopostabdominusmen of segments lacking appendagesrdquo
20 Paired tergal carinae (0) absent (1) present Liu et al(2007 265 fig 2F) recorded the presence of tergalcarinae in Leanchoilia illecebrosa shared with L superminuslata (eg Bruton and Whittington 1983 pl 17 106GarciacuteaminusBellido and Collins 2007 pl 1 5)
21 Pygidium (0) absent (1) present Coding of the pygiminusdium in many terminals herein was discussed by Paterminusson et al (2010 character 22)
22 Telson (0) absent (1) present Following Hendricks andLieberman (2008) a telson is a postminussegmental terminalstructure that lacks appendages Dunlop and Selden (1997)considered a telson to be absent in Cheloniellon but weidentify the conical projection posterior to the ultimate legpair and cerci (Stuumlrmer and Bergstroumlm 1978 figs 2 3 1011) as being most consistent with an identity as a telson
23 Telson shape (0) styliform (1) flapminuslike (2) lanceolate(3) short conical The outline of the telson in Haikouminuscaris ercaiensis is incompletely known but the mostcomplete specimen (Chen 2004 fig 472) constrains it aseither state 1 or state 2 A partial uncertainty coding isthus employed A unique state for Cheloniellon (state 3)is based on the interpretation in character 22
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 395
24 Telson fringed with spines (0) spines absent (1) spinespresent Presence of telson spines was introduced as a charminusacter by Hendricks and Lieberman (2008 character 34)
25 Paired flaps lateral to telson (0) absent (1) present Thischaracter describes the postminusventral plates of several taxa(see Edgecombe and Ramskoumlld 1999 character 25) Cotminuston and Braddy (2004 character 54) raised the possibilitythat the cerci of Cheloniellon could be homologous withpostminusventral plates (see also Hou and Bergstroumlm 1997 93)because they are similarly paired structures derived fromthe segment immediately anterior to the telson We havenot coded for this homology because the cerci of Chelominusniellon originate dorsally rather than ventrally
26 Midgut glands with submillimetric lamellae (0) absent(1) present Midgut glands in some stem group arthrominuspods and early crown group arthropods are metamericreniform organs with a characteristic lamellar structure(Butterfield 2002) Their morphology is identical inLeanchoilia and Oestokerkus notably including groovesthat radiate from a ventral median node (Figs 3A1 6A)The gut of Alalcomenaeus cambricus is more tubular(Briggs and Collins 1999 textminusfigs 10 11) and thoughan anterior six segments may display phosphatised latminuseral projections that likely represent gut diverticula (Wilminusson 2006) they have not been shown to have the detailedmorphology of Leanchoiliaminustype midgut glands Butterminusfield (2002 164) cited a figure from Bruton (1981 fig96) in support of ldquolamellarrdquo midgut glands in the cephaminuslon of Sidneyia Like Bruton (1981) in his description ofthe specimen we consider it likely that the structures inquestion are indeed gut diverticula but there is no basisfor identifying them more precisely as the reniformlamellar structures coded under this character
27 Exopod on first postminusantennal first postminusfrontal appendminusage segment (0) absent (1) present This character reminuslates to Cotton and Braddyrsquos (2004) character 3
28 Division of exopod (0) exopod an undivided flap (1)exopod differentiated into proximal and distal lobes (2)small flap with radiating lamellae (3) pediform exopoddivided into ca 6 articles Identification of the exopod inXiphosura follows Suzuki and Bergstroumlm (2008) Thischaracter groups conditions of the exopod shaft coded asindependent characters by Cotton and Braddy (2004)eg state 0 corresponds to their character 16 and state 1to their character 13 Our coding permits transformationbetween any two states without an extra implicit step
29 Exopod setae (0) short spiniform setae (1) long flatminustened tapering distally with slight separation (2)lamellar setae (flattened imbricating) (3) short finesetae A distinction is made between lamellipedian setaewhich are imbricated (state 2) and the pointed more disminustinctly separated and variably spiniform exopod setae ofleanchoiliids (state 1) This distinction is consistent withthe identification of megacheirans as nonminuslamellipedians(eg Hou and Bergstroumlm 1997 Bergstroumlm and Hou2005) Although Cotton and Braddy (2004 character 18)
regarded the form of the setae in Yohoia to be uncertainwe regard specimens figured by Whittington (1974 pl3 3) as conforming to the leanchoiliid state Squamaculadiffers from other taxa in its shorter spiniform setae(state 0) (Zhang et al 2004 textminusfig 1A B) Offacolushas longer spiniform projections on the distal articles ofthe exopod that have been variably identified as eitherspines or setae (Sutton et al 2002 fig 4b) but theirhomology with the setae that splay regularly along themargin of a flapminuslike exopod (eg states 0ndash3) is uncerminustain This and the following character for exopod setaeare left as ldquordquo for Offacolus A homology between thegill lamellae of Xiphosura and exopod setae of lamellipeminusdians has a long history (see Cotton and Braddy 2004character 18) We follow Suzuki and Bergstroumlm (2008)in identifying the xiphosuran gills as outgrowths of thebasis rather than of the exopod this perspective negates ahomology with the exopod setae of Lamellipedia Thepolymorphic coding for Emeraldella reflects the twoseparate states (1+2) displayed by the exopods in thetrunk (see Bruton and Whittington 1983 pl 8 45 47Edgecombe and Ramskoumlld 1999 fig 11)
30 Differentiation of setae along exopod (0) uniform orgradationally variable setae along length of exopod (1)lamellar setae confined to proximal lobe with distal lobebearing slender setae
31 Endopod segmentation (distal to coxa) (0) six or sevenpodomeres and terminal claw or telotarsal spurs (1)eight or more podomeres and terminal claw (2) ca 15podomeres State 1 is formulated to describe the segmenminustation of the leanchoiliid leg (GarciacuteaminusBellido and Collins2007 textminusfig 6 Liu et al 2007 fig 5) in contrast to thereduced number of podomeres in other euarthropods(Boxshall 2004) State 2 characterises Fortiforceps Apartial uncertainty coding is employed for Yohoia inwhich the precise number of podomeres is undeterminedand could be either state 0 or 1 (Whittington 1974 pl 74) but the spacing of the preserved podomere boundariesnegates the possibility of state 2
32 Endopods on anterior postminuscephalic segments (0) welldeveloped (1) rudimentary or lacking Following Cottonand Braddy (2004 character 11) the absence of trunkendopods in Yohoia (Whittington 1974) is coded as a puminustative homologue with the suppression of postminuscephalicendopods in Chelicerata
ResultsAs a convention for depicting the trees networks are rootedon Squamacula which separates artiopodan Lamellipedia(sensu Hou and Bergstroumlm 1997) from a group that includesMegacheira Chelicerata and Xenopoda all members ofwhich possess a telson
Under equal weights 14 shortest cladograms have a lengthof 77 steps with a Consistency Index of 064 and Retention Inminus
396 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
dex of 077 The strict consensus of these trees with supportvalues is shown in Fig 8 Leanchoiliidae is unambiguouslysupported in all 14 cladograms by four apomorphic characterstwo limbminusbearing cephalic segments posterior to the frontalappendage (character 3 state 1) long spinose projections onthe distal part of the terminal three podomeres of the frontalappendage that bear flagella (character 6 state 1) 11 trunksegments (character 15 state 1) and division of the exopodsinto two lobes (character 28 state 1) The arrangement of fouror five eyes in a subtransverse band across the cephalic shieldis unique to Leanchoiliidae (character 9 state 1) but is not anunambiguous autapomorphy of the group because the state isuncertain in close relatives notably Haikoucaris It is apominusmorphic for Leanchoiliidae only under particular optimisaminustions (Delayed Transformation)
The internal relationships of Leanchoiliidae are stableacross all 14 shortest cladograms Leanchoilia is united by thelanceolate telson (character 23 state 2) a unique unambiguminusous autapomorphy Within Leanchoilia a sister group relaminustionship between L superlata and L illecebrosa is well supminusported receiving a Bremer support of 3 and jackknife freminusquency of 86 These two species share four synapomorminusphies a medially pointed anterior cephalic margin (character2 state 1) flagella of the great appendage extending the lengthof the body (character 7 state 1) serrate lateral margins on thetrunk tergites (character 17 state 1) and paired tergal carinae(character 20 state 1) As such a species from Chengjiang (Lillecebrosa) and the Burgess Shale (L superlata) are moreclosely related to each other than are the two Burgess Shalecongeners a relationship that weakens the possibility that theoften cominusoccurring L superlata and L persephone could besexual dimorphs (GarciacuteaminusBellido and Collins 2007) ratherthan separate species The union of Leanchoilia and Oestominuskerkus is unambiguously supported by the shared midgutgland structure of both genera (character 26 state 1)
All shortest cladograms resolve Haikoucaris Yohoiaand Fortiforceps in that order as progressively less closelyrelated to Leanchoiliidae The union of these three taxa withLeanchoiliidae yields a clade that corresponds to Megacheirasensu Hou and Bergstroumlm (1997) apart from the ambiguousplacement of Jianfengia In half of the 14 shortest cladominusgrams Jianfengia is sister to the other megacheirans yieldminusing a monophyletic Megacheira ss In the remaining cladominusgrams Jianfengia is sister to Xenopoda sensu Hou andBergstroumlm (1997) Aglaspis and Chelicerata Haikoucarisand Leanchoiliidae are united by a frontal appendage comminusposed of two spineminusbearing articles in addition to the movminusable finger (character 5 state 1) Yohoia groups with thesetaxa based on spines fringing the telson (character 24 state1) and Fortiforceps is united with them at a broader levelbased on the flapminuslike telson shape (character 23 state 1)When Jianfengia is placed as sister to the other megacheiminusrans the entire clade is supported by the raptorial frontal apminuspendage (character 4 state 1) and the lack of a large hypominusstomal shield (character 13 state 0) In this set of cladominusgrams these features are nonminushomologous between Megaminus
cheira and Chelicerata whereas in some cladograms with thealternative resolution of Jianfengia as closer to cheliceratescharacter 4 is optimised as homologous in Megacheira andChelicerata
Reanalysis of the data with implied weights finds two optiminusmal trees (Fig 9) Relationships within Leanchoiliidae underequal weights are completely stable to reweighting as is theresolution of Haikoucaris as sister to Leanchoiliidae As suchthe character optimisations described above for the leanchoiminusliids are depicted the same in Fig 9 The main difference beminustween equal and implied weights involves the placement ofChelicerata Under equal weights (Fig 8) chelicerates are alminuslied to Xenopoda (here Emeraldella + Sidneyia + Chelominusniellon sensu clade L in Hou and Bergstroumlm 1997 fig 88)and Aglaspidida A clade composed of Aglaspidida Xenominuspoda and Chelicerata was similarly identified by Hou andBergstroumlm (1997 fig 88 their clade K) In our analysis thisgroup consistently finds support from one or more posteriortrunk segments lacking appendages (character 19 state 1) anddepending on alternative internal resolutions it is variably supminusported by dorsallyminuspositioned sessile eyes (character 8 state 1)and the absence of exopods on the first postminusantennalpostminuscheliceral limb (character 27 state 1) Under implied weightshowever chelicerates are more closely allied to Megacheirathan to aglaspidids or xenopodans (Fig 9)
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 397
2
582
76
Chelicera-morpha
Megacheira
62
2
53
50Leanchoiliidae
3
86
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
EurypteridaChelicerata
Fortiforceps
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia hanceyi
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
Fig 8 Strict consensus of 14 shortest cladograms (77 steps) based on charminusacter data in Table 1 Numbers above nodes are Bremer support those beminuslow nodes are jackknife frequencies gt50 Cheliceramorpha correspondsto a clade with the same composition in Cotton and Braddy (2004)
The major groupings in Fig 9 correspond closely to thoseidentified by Cotton and Braddy (2004) Notably the cladeuniting Aglaspis Xenopoda Offacolus Chelicerata Jianminusfengia and other Megacheira corresponds to their ldquoClade 3rdquo(Cotton and Braddy 2004 fig 8) for which they proposedthe name Cheliceramorpha The clade uniting CheloniellonAglaspis Sidneyia and Emeraldella in Fig 9 corresponds totheir ldquoClade 5rdquo and the clade that groups Megacheira andChelicerata is equivalent to their ldquoClade 4rdquo Their analysisdid not identify megacheirans as a monophyletic groupChelicerata was nested within the group and among taxaconsidered here Yohoia was resolved closer to cheliceratesthan to Leanchoiliidae In contrast Yohoia was regarded byBergstroumlm and Hou (2005) as more closely allied to Leanminuschoiliidae than to chelicerates a megacheiran grouping morecompatible with our cladograms The analysis of ldquogreat apminuspendage arthropodsrdquo by Vannier et al (2009) identified a
monophyletic Leanchoiliidae but a polyphyletic Megacheira(to the exclusion of Haikoucaris and Jianfengia) Howevertheir resolution of Fortiforceps and even more so Yohoia asthe closest relatives of Leanchoiliidae is as in our trees (apartfrom the different position of Haikoucaris)
In summary a major controversy in the systematics of theldquoshort great appendagerdquo arthropods remains whether thechelicerates are nested within the group or are instead moreclosely allied to taxa conventionally identified as lamelliminuspedians specifically Xenopoda and Aglaspidida We findthat both of these results are compatible with the availabledata depending on character weighting protocols Noneminustheless the broader megacheiranminuschelicerate issue does notcloud the status of leanchoiliids as a clade or the identificaminustion of their closest relatives (ie Haikoucaris and Yohoia)and the internal relationships of Leanchoiliidae are stableand supported by compelling characters
398 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
Eurypterida
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
2 7 17 20
3
2
1 1 1
26
1
281563
1 1 1 1
5
1
24
1
23 19 20 27 32
28Clade 4
1 0
4 13 11 27 0
154 1 1
3 19 32Clade 3
2 3
2 28
0
3 1 121
1
22
Clade 5
1 0
19 27
1
25
15 28 301 1
2312
1 150 1 3 3
15 16 2923
18
1 1
10 16
1 1
1 8
1
14
3
3
28 30
1 0
5
15
1 1
1
1
22
Fortiforceps
23
1 1 0 1
1 1
Leanchoilia hanceyi
2
2
1
1
Fig 9 Characters optimised on one of two shortest cladograms with imminusplied weights using a concavity constant k = 3 Black squares are nonminushomoplastic changes white squares homoplastic changes all changes areunambiguous Branches without strict support collapsed after optimisingcharacters Numbers for characters (above the branches) and states (belowthe branches) correspond to Table 1 ldquoClades 3ndash5rdquo correspond to groups sonumbered by Cotton and Braddy (2004 fig 8)
AcknowledgementsAccess to the field area has been most kindly allowed by the Buck famminusily We thank our collaborators in Emu Bay Shale research Jim Jago(University of South Australia Mawson Lakes) Jim Gehling andMike Lee (South Australian Museum Adelaide) for tremendous supminusport and advice as well as Mike Gemmell Dennis Rice Allison DaleyAaron Camens John Laurie Glenn Brock Pierre Kruse and Trevorand Jenny Worthy (all South Australian Museum) for their assistancein the field and Natalie Schroeder MaryminusAnne Binnie and RondaAtkinson (all South Australian Museum) for organisational support inthe lab and in the field Javier OrtegaminusHernaacutendez (University of Camminusbridge Cambridge UK) advised us on Kwanyinaspis and the journalrsquosreferees provided useful feedback TNT is available courtesy of theWilli Hennig Society Funding has been provided by an AustralianResearch Council Linkage grant (LP0774959) to the University ofAdelaide with generous financial assistance from Beach Energy Ltdand the South Australian Museum SeaLink provided logistical supminusport DGBrsquos travel to Australia has been supported by grants from theSpanish Research Council (CSIC PA 1001105) the Complutense Uniminusversity of Madrid and the Spanish Ministry of Education and Science(CGL2006minus12245BTE CGL2009minus07073 and RYC2007minus00090)
ReferencesBergstroumlm J and Hou XminusG 2005 Early Palaeozoic lamellipedian arthrominus
pods In S Koenemann and RA Jenner (eds) Crustacea and Arthrominuspod Relationships Crustacean Issues 16 73ndash93
Bitsch J and Bitsch C 2007 The segmental organization of the head reminusgion in Chelicerata a critical review of recent studies and hypothesesActa Zoologica 88 317ndash335
Bousfeld EL 1995 A contribution to the natural classification of Lower andMiddle Cambrian arthropods food gathering and feeding mechanismsAmphipacifica 2 3ndash34
Boxshall GA 2004 The evolution of arthropod limbs Biological Reviews79 253ndash300
Bremer K 1994 Branch support and tree stability Cladistics 10 295ndash304Briggs DEG and Collins D 1999 The arthropod Alalcomenaeus cambricus
Simonetta from the Middle Cambrian Burgess Shale of British ColumbiaPalaeontology 42 953ndash977
Briggs DEG and Robison RA 1984 Exceptionally preserved nonminustrilobite arthropods and anomalocaridids from the Middle Cambrian ofUtah University of Kansas Paleontological Contributions 111 1ndash23
Briggs DEG Lieberman BS Hendricks JR Halgedahl SL andJerrard RD 2008 Middle Cambrian arthropods from Utah Journal ofPaleontology 82 238ndash254
Bruton DL 1981 The arthropod Sidneyia inexpectans Middle CambrianBurgess Shale British Columbia Philosophical Transactions of theRoyal Society of London Series B 295 619ndash656
Bruton DL and Whittington HB 1983 Emeraldella and Leanchoiliatwo arthropods from the Burgess Shale Middle Cambrian British Cominuslumbia Philosophical Transactions of the Royal Society of London Seminusries B 300 553ndash582
Budd GE 2002 A palaeontological solution to the arthropod head probminuslem Nature 417 271ndash275
Butterfield NJ 2002 Leanchoilia guts and the interpretation of threeminusdiminusmensional structures in Burgess Shaleminustype fossils Paleobiology 28155ndash171
Chen JminusY 2004 The Dawn of Animal World [in Chinese] 366 pp JiangScience and Technology Press Nanjing
Chen JminusY Waloszek D and Maas A 2004 A new ldquogreatminusappendagerdquo arminusthropod from the Lower Cambrian of China and homology of cheliceratechelicerae and raptorial anterominusventral appendages Lethaia 37 3ndash20
Cotton TJ and Braddy SJ 2004 The phylogeny of arachnomorph arthrominuspods and the origin of the Chelicerata Transactions of the Royal Societyof Edinburgh Earth Sciences 94 169ndash193
Daley AC Budd GE Caron JminusB Edgecombe GD and Collins D2009 The Burgess Shale anomalocaridid Hurdia and its significancefor early euarthropod evolution Science 323 1597ndash1600
Dunlop JA 2006 New ideas about the euchelicerate stemminuslineage In CDeltshev and P Stoev (eds) European Arachnology 2005 Acta Zoominuslogica Bulgarica (Supplement 1) 9ndash27
Dunlop JA and Selden PA 1997 The early history and phylogeny of thechelicerates In RA Fortey and RH Thomas (eds) Arthropod Relaminustionships Systematics Association Special Volume 55 221ndash235
Edgecombe GD and Ramskoumlld L 1999 Relationships of CambrianArachnata and the systematic position of Trilobita Journal of Paleonminustology 73 263ndash287
Farris JS Albert VA Kaumlllersjouml M Lipcomb D and Kluge AG1996 Parsimony jackknifing outperforms neighborminusjoining Cladistics12 99ndash124
GarciacuteaminusBellido DC and Collins D 2007 Reassessment of the genusLeanchoilia (Arthropoda Arachnomorpha) from the Middle CambrianBurgess Shale British Columbia Canada Palaeontology 50 693ndash709
GarciacuteaminusBellido DC Paterson JR Edgecombe GD Jago JB GehlingJG and Lee MSY 2009 The bivalved arthropods Isoxys and Tuzoiawith softminuspart preservation from the lower Cambrian Emu Bay ShaleLagerstaumltte (Kangaroo Island Australia) Palaeontology 52 1221ndash1241
Goloboff PA 1993 Estimating character weights during tree searchCladistics 9 83ndash91
Goloboff PA Farris JS and Nixon KC 2008 TNT a free program forphylogenetic analysis Cladistics 24 774ndash786
Harzsch S and Hafner G 2006 Evolution of eye development in arthrominuspods phylogenetic aspects Arthropod Structure amp Development 35319ndash340
Hendricks JR and Lieberman BS 2008 New phylogenetic insights intothe Cambrian radiation of arachnomorph arthropods Journal of Paleminusontology 82 585ndash594
Hou XminusG and Bergstroumlm J 1997 Arthropods of the Lower CambrianChengjiang fauna southwest China Fossils and Strata 45 1ndash116
Jago JB Zang W Sun X Brock GA Paterson JR and SkovstedCB 2006 A review of the Cambrian biostratigraphy of South Austraminuslia Palaeoworld 15 406ndash423
Liu Y Hou XminusG and Bergstroumlm J 2007 Chengjiang arthropod Leanminuschoilia illecebrosa (Hou 1987) reconsidered GFF 129 263ndash272
Luo H Hu S Chen L Zhang S and Tao Y 1999 Early CambrianChengjiang fauna from Kunming region China [in Chinese] 129 ppYunnan Science and Technology Press Kunming
Luo H Hu S Zhang S and Tao Y 1997 New occurrence of the EarlyCambrian Chengjiang fauna in Haikou Kunming Yunnan Provinceand study on Trilobitoidea Acta Geologica Sinica 71 122ndash132
Nixon KC 2002 WINCLADA Version 19908 Cornell University IthacaNY
Orr PJ Siveter DJ Briggs DEG Siveter DJ and Sutton MD 2000A new arthropod from the Silurian KonservatminusLagerstaumltte of Hereminusfordshire Proceedings of the Royal Society Series B Biological Sciminusences 267 1497ndash1504
Paterson JR and Brock GA 2007 Early Cambrian trilobites fromAngorichina Flinders Ranges South Australia with a new assemblagefrom the Pararaia bunyerooensis Zone Journal of Paleontology 81116ndash142
Paterson JR and Jago JB 2006 New trilobites from the Lower CambrianEmu Bay Shale Lagerstaumltte at Big Gully Kangaroo Island South Ausminustralia Memoirs of the Association of Australasian Palaeontologists 3243ndash57
Paterson JR Edgecombe GD GarciacuteaminusBellido DC Jago JB andGehling JG 2010 Nektaspid arthropods from the lower CambrianEmu Bay Shale Lagerstaumltte South Australia with a reassessment oflamellipedian relationships Palaeontology 53 377ndash402
Paterson JR Jago JB Gehling JG GarciacuteaminusBellido DC Edgecombe
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 399
GD and Lee MSY 2008 Early Cambrian arthropods from the EmuBay Shale Lagerstaumltte South Australia 319ndash325 In I Raacutebano RGozalo and D GarciacuteaminusBellido (eds) Advances in Trilobite ResearchCuadernos del Museo Geominero 9 319ndash325
Shultz JW 2007 A phylogenetic analysis of the arachnid orders based onmorphological characters Zoological Journal of the Linnean Society150 221ndash265
Stein M 2010 A new arthropod from the Early Cambrian of North Greenminusland with a ldquogreat appendagerdquominuslike antennula Zoological Journal ofthe Linnean Society 158 477ndash500
Stuumlrmer W and Bergstroumlm J 1978 The arthropod Cheloniellon from theDevonian Hunsruumlck Shale Palaumlontologische Zeitschrift 52 57ndash81
Sutton MD Briggs DEG Siveter DJ Siveter DJ and Orr PD2002 The arthropod Offacolus kingi (Chelicerata) from the Silurian ofHerefordshire England computer based morphological reconstrucminustions and phylogenetic affinity Proceedings of the Royal Society B Biminusological Sciences 269 1195ndash1203
Suzuki Y and Bergstroumlm J 2008 Respiration in trilobites a reevaluationGFF 130 211ndash229
Vannier J GarciacuteaminusBellido DC Hu XminusX and Chen AminusL 2009 Arthrominuspod visual predators in the early pelagic ecosystem evidence from the
Burgess Shale and Chengjiang biotas Proceedings of the Royal SocietyB Biological Sciences 276 2567ndash2574
Whittington HB 1974 Yohoia Walcott and Plenocaris n gen arthropodsfrom the Burgess Shale Middle Cambrian British Columbia Geologiminuscal Survey of Canada Bulletin 231 1ndash21
Wills MA Briggs DEG Fortey RA Wilkinson M and SneathPHA 1998 An arthropod phylogeny based on fossil and recent taxaIn GD Edgecombe (ed) Arthropod Fossils and Phylogeny 33ndash105Columbia University Press New York
Wilson LA 2006 Food for thought a morphological and taphonomic studyof fossilised digestive systems from Early to Middle Cambrian taxa275 pp Unpublished PhD thesis University of Cambridge Cambridge
Zhang XminusG and Hou X 2007 Gravitational constraints on the burial ofChengjiang fossils Palaios 22 448ndash453
Zhang XminusL Han J Zhang Z Liu H and Shu D 2004 Redescription ofthe Chengjiang arthropod Squamacula clypeata Hou and Bergstroumlm fromthe Lower Cambrian Southminuswest China Palaeontology 47 605ndash617
Zhao Y Zhu M Babcock LE Yuan L Parsley RL Peng J YangX and Wang Y 2005 Kaili Biota a taphonomic window on diversifiminuscation of metazoans from the basal Middle Cambrian Guizhou ChinaActa Geologica Sinica 79 751ndash765
400 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011

doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 389
SAM P45904a b (unfigured) poorlyminuspreserved cephalic shield trunkwith wellminuspreserved midgut glands proximal part of telson SAMP45905ab (unfigured) cephalic shield lacking great appendages butexposing an eye trunk with incomplete telson SAM P45906a b (unfigminusured) great appendages with spinose projections oriented anteriorlytrunk preserved to end of midgut glands SAM P45908a b (unfigured)cephalic shield great appendages with flagella most of trunk (telsonmissing) showing midgut glands and exopod setae SAM P45909 (unminusfigured) small specimen with great appendages and series of mineralminusised midgut glands SAM P45910a b (unfigured) incomplete great apminuspendages oriented anteriorly trunk with complete series of midgutglands but indistinct appendages
Type locality Buck Quarry Big Gully (Fig 1)
Type horizon Emu Bay Shale (Cambrian Series 2 Stage 4)
DiagnosismdashAs for genus
DescriptionmdashCephalic shield 36 length of the trunk sagitminustally dorsal margin of cephalic shield gently convex in lateralview anteromedian margin inferred to be evenly rounded(based on comparison with lateral versus dorsal preservation ofBurgess Shale Leanchoilia and Alalcomenaeus) lacking an upminusturned median process Eye visible in SAM P54167 (Fig 4A)and P45905 relatively large (20 sagittal length of cephalon)exposed under anterolateral part of cephalic shield (Fig 4A2)
390 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Fig 4 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov SAM P45167 A P45167a A1 cephalic shield and most of trunkA2 detail of cephalic shield B P45167b A2 and B light from NE Overlap between adjacent tergites offset on left (Lol) and right (Rol) sides of bodyBoundaries between cephalic shield and anterior trunk tergites delimited in B by dashed lines Arrows mark positions of inferred dorsal transverse tendonsAbbreviations cn1ndash2 cephalic endopods cs cephalic shield gap peduncle of great appendage ol overlap between adjacent trunk tergites t1ndasht3 tergitesof trunk segments 1ndash3 tn trunk endopods
Fig 5 Leanchoiliid megacheiran arthropod Oestokerkus megacholix genet sp nov A SAM P 15531 B SAM P43630b Light from NE Abbreviaminustions cs cephalic shield ga great appendage gap peduncle of great apminuspendage mg midgut glands ol overlap between adjacent trunk tergites txtrunk exopods
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 391
Great appendage robust proportions of peduncle and disminustal spineminusbearing articles as in other members of Leanchoiliminusidae more commonly preserved with posterior orientation
but anteriorly oriented in SAM P45906 and SAM P45910Each of the three long spinose projections terminating at thesame length the projections evenly tapering distally with
392 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Fig 6 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov SAM P45168 A head and anterior part of trunk B Detail of spinoseprojections of great appendages and excretory organs (sensu Bruton and Whittington 1983) Abbreviations cs cephalic shield eo excretory organ gagreat appendage mg midgut glands tx trunk exopods
margins straight for much of their length or slightly curvedterminally distal spinose projection distinctly stouter at itsbase than the two more proximal projections and more markminusedly tapering along its length Dark irregularminusshaped low reminuslief lump on basal spineminusbearing article of great appendagesof SAM P45168 (eo in Fig 6) and SAM P45908 correminussponding to excretory organ of Bruton and Whittington(1983) Long flagella extending from each of the threespinose projections seen only in SAM P43631 (Figs 23A3) of similar narrow width along its preserved extent anminusnulations indistinguishable
Trunk of 11 segments of even length Maximum height oftrunk relative to length (excluding telson) ca 39 bodydeepest at tergite 4 Trunk tergite boundaries marked as narminusrow ridges in several specimens corresponding to narrowband of overlap between adjacent tergites (ol in Figs 2 3A14A1 B) Some specimens show tergites to be bisected by anarrow but sharp furrow of uncertain identity (eg possiblyimpression of dorsal transverse tendon cf Stein 2010 fig12B C E) (arrows in Figs 3A1 4B)
Exopods with fringe of long setae on cephalic and trunkappendages (Fig 2) setal fringe gradually shortening fromabout the fourth trunk segment Setae project ventrally bymore than 30 height of tergites in deepest part of trunk(Fig 2B) setae flattened narrow and parallelminussided alongmost of their length tapering near their distal ends to termiminusnate as a pointed tip (Figs 2B 3A3 arrows) Endopods exminusposed only in SAM P45167 (Fig 4) two pairs associatedwith cephalon (Fig 4B) those of anterior segments of trunkof similar morphology to those in cephalon all slender lackminusing obvious endites podomere boundaries unrecognisable
Midgut glands occupy a large extent of the dorsoventralheight of the trunk two midgut glands in cephalon of similarsize to those in each of the first seven segments of the trunk(Figs 3A1 5A 6A) all developed as radiating grooves emaminusnating from a globular structure at their ventral edge as in
Leanchoilia superlata (GarciacuteaminusBellido and Collins 2007 pl2 6) Foregut trace in SAM P43631 (Fig 2) indicates that gutcurves ventrally in anterior part of cephalon mouth regionnot preserved Terminal part of hindgut (in trunk segment 11)with closely spaced longitudinal lineations (Fig 2) that apminusparently correspond to folds in the gut wall hindgut termiminusnating at base of telson
Telson most completely preserved in holotype (SAMP43631 Figs 2B 3A2 B) In this specimen the telson likerest of body compressed in lateral aspect Telson shows meminusdian and marginal reinforcing ridges connecting a flat trianminusgular surface this surface appears to be partially broken inSAM P43631 showing underlying surface of other (left) halfof telson Lateral preservation of specimen implies that topcorresponds to an axial structure and bottom to a marginalone 30ndash35 angle in posterior edge of vane connecting thesetwo structures requires that in dorsal aspect telson would apminuspear forked and widening distally Five stout pointed dorminussally curved marginal spines attached to outer posterior edgeon each side of telson
Phylogenetic analysis ofLeanchoiliidaeDetermining the systematic position of Oestokerkus demandsan appraisal of the relationships of Leanchoiliidae as a wholeCladistic analysis including the bestminusknown members ofMegacheira including leanchoiliids has been conducted byCotton and Braddy (2004) and Vannier et al (2009) The taxominusnomic sampling used by Cotton and Braddy (2004) includedseveral megacheiran genera representative chelicerates anumber of taxa representing groups that have been proposedas possible chelicerate relatives (eg Sidneyia AglaspisCheloniellon) in other analyses (Dunlop and Selden 1997Hou and Bergstroumlm 1997) and members of the major lineagesof the Artiopoda sensu Hou and Bergstroumlm (1997) ie trilominusbiteminusallied lamellipedians Because artiopodan interminusrelationminusships have remained quite stable through a series of phylogenminusetic analyses (Edgecombe and Ramskoumlld 1999 Cotton andBraddy 2004 Hendricks and Lieberman 2008 Paterson et al2010) and artiopodans are now often regarded as less closelyrelated to Megacheira than is Chelicerata (Chen et al 2004Cotton and Braddy 2004 Dunlop 2006) we have reduced thesampling of Artiopoda in the present analysis To augment thediversity of ldquoshort great appendagerdquo arthropods Haikoucarisis added based on the description of H ercaiensis by Chen etal (2004) Offacolus (Orr et al 2000 Sutton et al 2002) hasbeen allied to Chelicerata and is included as a terminal Beminuscause the present analysis is principally concerned with relaminustionships of and within Leanchoiliidae sensu Liu et al (2007)we follow Vannier et al (2009) in sampling Alalcomenaeusand Leanchoilia at the species level Within Chelicerata Cotminuston and Braddy (2004) and Hendricks and Lieberman (2008)sampled ancestral states in Xiphosura by coding the Devonian
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 393
Fig 7 Reconstruction of Oestokerkus megacholix gen et sp nov in lateralview Total length and annulations on the flagella and positions of podominusmere boundaries of the frontal appendage are based on Leanchoilia superminuslata (Bruton and Whittington 1983)
synziphosurine Weinbergina Our coding for Xiphosura alsoemploys information from extant xiphosurans because theysupply more reliable information on the homology of the partsof the trunk appendages (Suzuki and Bergstroumlm 2008) and eyeanatomy (Harzsch and Hafner 2006) among other charactersystems than do fossils alone
Our character set extracts characters from the matrix ofCotton and Braddy (2004) that vary in the revised taxonomicsample reformulates some characters to accommodate reminusvised homology concepts and adds new characters especiallyfor leanchoiliids In total 32 characters are included (Table 1)In this analysis the primary homology of the ldquoshort great apminuspendagerdquo and chelicera is coded (as described by character 4)and the taxon sampling broadly constrains megacheirans aschelicerate relatives (as defended by Chen et al 2004 Cottonand Braddy 2004) rather than testing the alternative that theycould be stem group Euarthropoda (Budd 2002 Bergstroumlmand Hou 2005) A test of the two alternatives involves samminuspling of the entire diversity of euarthropods and a range ofnonminuseuarthropod outgroups an exercise beyond the scope ofthis study
Characters1 Mineralised cuticle (0) absent (1) present For definiminus
tion and discussion of this character see Cotton andBraddy (2004 character 30) and Paterson et al (2010character 1)
2 Curvature of anterior margin of cephalic shield (0) rounminusded or straight (1) pointed medially (2) with multiplespinose projections This character codes for variability inthe shape of the cephalic shield in Leanchoiliidae (state 0and 1 sensu GarciacuteaminusBellido and Collins [2007 textminusfigs10 and 9 respectively]) State 2 is added to accommodateOffacolus the median spine of a series that fringes the ceminusphalic margin is not convincingly homologised with themedian projection in leanchoiliids
3 Number of cephalic somitesappendages (0) antenna orfrontal appendage only (1) antenna or frontal appendageand two limb pairs (2) antenna or frontal appendage andthree limb pairs (3) antenna or frontal appendage and fourlimb pairs (4) antenna or frontal appendage and five limbpairs Coding for lamellipedians and Aglaspis is as discussedby Paterson et al (2010 character 2) Coding for Alalcomeminusnaeus cambricus (state 1) follows Briggs and Collins (1999)Leanchoilia illecebrosa (state 2) follows Liu et al (2007)and Leanchoilia persephone and Leanchoilia superlata (state1) follows GarciacuteaminusBellido and Collins (2007)
4 Form of first cephalic appendage (0) antenna (1) anteminusriorly directed raptorial appendage with reduced numberof podomeres each of the terminal podomeres bearing aspinose process on its distal margins a geniculation beminustween spinose podomeres and peduncle This characteris formulated to identify a homology between the frontalappendages of megacheirans and the chelicera of Cheliminus
cerata and state 1 corresponds to the putative apomorminusphic detail defended by Chen et al (2004) and Cottonand Braddy (2004) Vannier et al (2009 character 8)specify another shared property of ldquoshort great appendminusagesrdquo of the megacheiran (ss) type in their characterstate ldquobent at an anglerdquo referring to the geniculation beminustween articles of the peduncle or between the peduncleand distal spiniferous articles This property is sharedwith Chelicerata
5 Number of spineminusbearing articles on frontal appendage (0)three + movable finger (1) two + movable finger (2) one+ movable finger The number of spineminusbearing articles onthe frontal appendage (exclusive of the terminal movablefinger) varies from three in Yohoia (Whittington 1974 pl6 1ndash3 pl 7 3ndash6 pl 9 4ndash6) and Fortiforceps (Hou andBergstroumlm 1997 figs 31C D 32A 33B C E) to two inHaikoucaris (Chen et al 2004 fig 2A B) and Leanminuschoiliidae or only one the last being a chelicera
6 Long spinose projections on distal part of terminal threepodomeres of frontal appendage bearing a flagellum (0)absent (1) present The morphology of the great appendminusage in Leanchoiliidae is coded here We have not codedthe length of the spinose projections and the presence ofa flagellum as two independent characters Cotton andBraddyrsquos (2004) character 6 state 2 (ldquospinesrdquo longer thanthe entire podomere series) and character 8 state 1 (presminusence of an annulated flagellum) are thus grouped as a sinminusgle trait here
394 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Table 1 Characters coded for phylogenetic analysis Polymorphismindicated by ldquo+rdquo partial uncertainty by states separated by ldquordquo Abbreviminusations A Alalcomenaeus L Leanchoilia O Oestokerkus
Squamacula 0000minus minus minus2minus0 0110 00000 10minus minus0 1000 0
Cindarella 0030minus minus minus000 0010 00000 10minus minus0 11121 00
Kuamaia 000minus minus minus001 0011 10000 10minus minus0 11121 00
Kwanyinaspis 00 001 00112 1000 1000 1121 00
Olenoides 1020minus minus minus100 0011 00000 10minus minus0 01121 00
Retifacies 0020minus minus minus000 0010 00000 11000 1020 00
Aglaspis 10(23)0minus minus minus100 00101 00010 01001 0 00
Sidneyia 0000minus minus minus000 01102 00110 01101 00020 00
Emeraldella 0040minus minus minus2minus 0 00103 00110 01001 001(1+2)1 00
A cambricus 00111 10010 00001 00000 01110 01110 10
L illecebrosa 01211 11010 00001 01001 01210 11110 10
L persephone 00111 10010 00001 00000 01210 11110 10
L superlata 01111 11010 00001 01001 01210 11110 10
L hanceyi 00 00 0001 0000 01210
O megacholix 0011 100 00001 00000 01110 1110 0
Jianfengia 0021 0minus000 00005 00000 01000 010(12)0 0
Fortiforceps 00210 0minus00 00004 00000 01100 01230 20
Yohoia 00210 0minus000 00003 00011 01110 00010 (01)1
Haikoucaris 00211 0minus00 00003 00000 01(12)0 1010 0
Cheloniellon 0040minus minus minus100 00100 10010 01300 0030 00
Offacolus 02412 0minus200 0000 00010 01000 13minus minus 01
Xiphosura 00412 0minus100 10000 0010 01000 000minus minus 01
Eurypterida 00412 0minus100 10002 00010 01000 00minus minus minus 01
7 Length of flagellate great appendage (0) flagella termiminusnating at midlength of trunk (1) flagella extending to endof body Coding is applicable to Leanchoiliidae onlydistinguishing some Leanchoilia species that resembleAlalcomenaeus in having relative short great appendages(state 0) versus the longer appendages in L superlata andL illecebrosa
8 Position of lateral facetted eyes (0) ventral (1) dorsalsessile (2) absent Variation in Lamellipedia and severalother taxa coded herein follows Paterson et al (2010character 3)
9 Four or five ventral eyes arranged in a subtransverseband across head shield (0) absent (1) present A bandof ventral eyes is shared by the best known species ofLeanchoiliidae The distinction between three medianeyes in Alalcomenaeus cambricus (Briggs and Collins1999) and two in Leanchoilia illecebrosa (Liu et al2007) and both Leanchoilia superlata and L persephone(GarciacuteaminusBellido and Collins 2007) is uninformative forthe taxon sample here
10 Bulge in exoskeleton accommodating dropminusshaped venminustral eyes (0) absent (1) present This character followsEdgecombe and Ramskoumlld (1999 character 6) and Patminuserson et al (2010 character 5)
11 Dorsal median eyes (0) absent (1) present Coding for anabsence of dorsal median eyes in fossil taxa considers theabsence of either paired ocelli or a median eye tubercleand the character is scored as a chelicerate autapomorphy(see Cotton and Braddy 2004 character 24)
12 Width of doublure on cephalic shield (0) narrow or modminuserately wide (1) entire ventral side of head covered byexpansion of doublure The interpretation of state 1 inSidneyia and Squamacula is discussed by Cotton andBraddy (2004) and Paterson et al (2010 characters 12and 29)
13 Mouth covered by hypostome (0) absent (1) present Noldquoshort great appendagerdquo arthropods have been shown topossess a hypostome and a reconstruction of a smalllabrum in Alalcomenaeus (Briggs and Collins 1999) wasadmittedly conjectural The absence of a conspicuoushypostome contrasts with its ubiquitous presence in laminusmellipedians and several putative chelicerateminusallied taxathat have antennae (such as aglaspidids and Emeralminusdella) Chelicerates have a small epistomeminuslabrum (Bitschand Bitsch 2007) but like Cotton and Braddy (2004character 27 state 4) we code a hypostomal sclerite aslacking Kwanyinaspis has a large hypostome similar tothat found in aglaspidids (photograph of the holotype ofKwanyinaspis maotianshanensis shown to us by JavierOrtegaminusHernaacutendez [personal communication 2010])
14 Tergite articulations (0) extensive overlap of tergites (1)edge to edge articulations This character follows Edgeminuscombe and Ramskoumlld (1999 character 18) and Patersonet al (2010 character 20)
15 Number of trunk segments (0) nine (1) 11 (2) 12 (3)13 (4) 20 (5) 21 or 22 Coding is confined to taxa that do
not possess a pygidium because the precise number ofsomites associated with pygidia can sometimes not bedetermined with confidence Codings for Xiphosura andEurypterida use opisthosomal somite numbers followingShultz (2007 character 95)
16 Exoskeletal boundaries of anterior trunk segments (0)boundaries transverse or reflexed posterolaterally (1) reminusflexed anterolaterally This character follows Edgecombeand Ramskoumlld (1999 character 19) and Paterson et al(2010 character 21)
17 Trunk tergites with serrate lateral margins (0) absent (1)present Liu et al (2007 fig 2C) documented a row ofmarginal spines on the tergites of Leanchoilia illeceminusbrosa a featured shared by Leanchoilia superlata (egGarciacuteaminusBellido and Collins 2007 pl 1 7) but not otherterminals coded here
18 Differentiation in width of trunk tergites (0) trunk withgradational narrowing posteriorly (1) abrupt narrowingat boundary between prothorax and opisthothorax Anabrupt contraction in width between the boundary ofthe prominus and opisthothorax is observed in Emeraldellaand Sidneyia Yohoia has been reconstructed as having apronounced break in the shape of the trunk tergites beminushind segment 10 (Whittington 1974 textminusfig 2) thepoint at which the paratergal folds lose their spinose tipsand appendages are lacking Specimens preserved indorsal aspect (Whittington 1974 pl 10) suggest that thewidth of the opisthothorax is rather less abruptly narminusrowed and we code Yohoia as state 0
19 Appendages of posterior trunk segments (0) all trunktergites bearing appendages (1) one or more posteriortrunk segments lacking appendages Coding correspondsto Cotton and Braddyrsquos (2004) character 43 ldquopostabdominusmen of segments lacking appendagesrdquo
20 Paired tergal carinae (0) absent (1) present Liu et al(2007 265 fig 2F) recorded the presence of tergalcarinae in Leanchoilia illecebrosa shared with L superminuslata (eg Bruton and Whittington 1983 pl 17 106GarciacuteaminusBellido and Collins 2007 pl 1 5)
21 Pygidium (0) absent (1) present Coding of the pygiminusdium in many terminals herein was discussed by Paterminusson et al (2010 character 22)
22 Telson (0) absent (1) present Following Hendricks andLieberman (2008) a telson is a postminussegmental terminalstructure that lacks appendages Dunlop and Selden (1997)considered a telson to be absent in Cheloniellon but weidentify the conical projection posterior to the ultimate legpair and cerci (Stuumlrmer and Bergstroumlm 1978 figs 2 3 1011) as being most consistent with an identity as a telson
23 Telson shape (0) styliform (1) flapminuslike (2) lanceolate(3) short conical The outline of the telson in Haikouminuscaris ercaiensis is incompletely known but the mostcomplete specimen (Chen 2004 fig 472) constrains it aseither state 1 or state 2 A partial uncertainty coding isthus employed A unique state for Cheloniellon (state 3)is based on the interpretation in character 22
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 395
24 Telson fringed with spines (0) spines absent (1) spinespresent Presence of telson spines was introduced as a charminusacter by Hendricks and Lieberman (2008 character 34)
25 Paired flaps lateral to telson (0) absent (1) present Thischaracter describes the postminusventral plates of several taxa(see Edgecombe and Ramskoumlld 1999 character 25) Cotminuston and Braddy (2004 character 54) raised the possibilitythat the cerci of Cheloniellon could be homologous withpostminusventral plates (see also Hou and Bergstroumlm 1997 93)because they are similarly paired structures derived fromthe segment immediately anterior to the telson We havenot coded for this homology because the cerci of Chelominusniellon originate dorsally rather than ventrally
26 Midgut glands with submillimetric lamellae (0) absent(1) present Midgut glands in some stem group arthrominuspods and early crown group arthropods are metamericreniform organs with a characteristic lamellar structure(Butterfield 2002) Their morphology is identical inLeanchoilia and Oestokerkus notably including groovesthat radiate from a ventral median node (Figs 3A1 6A)The gut of Alalcomenaeus cambricus is more tubular(Briggs and Collins 1999 textminusfigs 10 11) and thoughan anterior six segments may display phosphatised latminuseral projections that likely represent gut diverticula (Wilminusson 2006) they have not been shown to have the detailedmorphology of Leanchoiliaminustype midgut glands Butterminusfield (2002 164) cited a figure from Bruton (1981 fig96) in support of ldquolamellarrdquo midgut glands in the cephaminuslon of Sidneyia Like Bruton (1981) in his description ofthe specimen we consider it likely that the structures inquestion are indeed gut diverticula but there is no basisfor identifying them more precisely as the reniformlamellar structures coded under this character
27 Exopod on first postminusantennal first postminusfrontal appendminusage segment (0) absent (1) present This character reminuslates to Cotton and Braddyrsquos (2004) character 3
28 Division of exopod (0) exopod an undivided flap (1)exopod differentiated into proximal and distal lobes (2)small flap with radiating lamellae (3) pediform exopoddivided into ca 6 articles Identification of the exopod inXiphosura follows Suzuki and Bergstroumlm (2008) Thischaracter groups conditions of the exopod shaft coded asindependent characters by Cotton and Braddy (2004)eg state 0 corresponds to their character 16 and state 1to their character 13 Our coding permits transformationbetween any two states without an extra implicit step
29 Exopod setae (0) short spiniform setae (1) long flatminustened tapering distally with slight separation (2)lamellar setae (flattened imbricating) (3) short finesetae A distinction is made between lamellipedian setaewhich are imbricated (state 2) and the pointed more disminustinctly separated and variably spiniform exopod setae ofleanchoiliids (state 1) This distinction is consistent withthe identification of megacheirans as nonminuslamellipedians(eg Hou and Bergstroumlm 1997 Bergstroumlm and Hou2005) Although Cotton and Braddy (2004 character 18)
regarded the form of the setae in Yohoia to be uncertainwe regard specimens figured by Whittington (1974 pl3 3) as conforming to the leanchoiliid state Squamaculadiffers from other taxa in its shorter spiniform setae(state 0) (Zhang et al 2004 textminusfig 1A B) Offacolushas longer spiniform projections on the distal articles ofthe exopod that have been variably identified as eitherspines or setae (Sutton et al 2002 fig 4b) but theirhomology with the setae that splay regularly along themargin of a flapminuslike exopod (eg states 0ndash3) is uncerminustain This and the following character for exopod setaeare left as ldquordquo for Offacolus A homology between thegill lamellae of Xiphosura and exopod setae of lamellipeminusdians has a long history (see Cotton and Braddy 2004character 18) We follow Suzuki and Bergstroumlm (2008)in identifying the xiphosuran gills as outgrowths of thebasis rather than of the exopod this perspective negates ahomology with the exopod setae of Lamellipedia Thepolymorphic coding for Emeraldella reflects the twoseparate states (1+2) displayed by the exopods in thetrunk (see Bruton and Whittington 1983 pl 8 45 47Edgecombe and Ramskoumlld 1999 fig 11)
30 Differentiation of setae along exopod (0) uniform orgradationally variable setae along length of exopod (1)lamellar setae confined to proximal lobe with distal lobebearing slender setae
31 Endopod segmentation (distal to coxa) (0) six or sevenpodomeres and terminal claw or telotarsal spurs (1)eight or more podomeres and terminal claw (2) ca 15podomeres State 1 is formulated to describe the segmenminustation of the leanchoiliid leg (GarciacuteaminusBellido and Collins2007 textminusfig 6 Liu et al 2007 fig 5) in contrast to thereduced number of podomeres in other euarthropods(Boxshall 2004) State 2 characterises Fortiforceps Apartial uncertainty coding is employed for Yohoia inwhich the precise number of podomeres is undeterminedand could be either state 0 or 1 (Whittington 1974 pl 74) but the spacing of the preserved podomere boundariesnegates the possibility of state 2
32 Endopods on anterior postminuscephalic segments (0) welldeveloped (1) rudimentary or lacking Following Cottonand Braddy (2004 character 11) the absence of trunkendopods in Yohoia (Whittington 1974) is coded as a puminustative homologue with the suppression of postminuscephalicendopods in Chelicerata
ResultsAs a convention for depicting the trees networks are rootedon Squamacula which separates artiopodan Lamellipedia(sensu Hou and Bergstroumlm 1997) from a group that includesMegacheira Chelicerata and Xenopoda all members ofwhich possess a telson
Under equal weights 14 shortest cladograms have a lengthof 77 steps with a Consistency Index of 064 and Retention Inminus
396 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
dex of 077 The strict consensus of these trees with supportvalues is shown in Fig 8 Leanchoiliidae is unambiguouslysupported in all 14 cladograms by four apomorphic characterstwo limbminusbearing cephalic segments posterior to the frontalappendage (character 3 state 1) long spinose projections onthe distal part of the terminal three podomeres of the frontalappendage that bear flagella (character 6 state 1) 11 trunksegments (character 15 state 1) and division of the exopodsinto two lobes (character 28 state 1) The arrangement of fouror five eyes in a subtransverse band across the cephalic shieldis unique to Leanchoiliidae (character 9 state 1) but is not anunambiguous autapomorphy of the group because the state isuncertain in close relatives notably Haikoucaris It is apominusmorphic for Leanchoiliidae only under particular optimisaminustions (Delayed Transformation)
The internal relationships of Leanchoiliidae are stableacross all 14 shortest cladograms Leanchoilia is united by thelanceolate telson (character 23 state 2) a unique unambiguminusous autapomorphy Within Leanchoilia a sister group relaminustionship between L superlata and L illecebrosa is well supminusported receiving a Bremer support of 3 and jackknife freminusquency of 86 These two species share four synapomorminusphies a medially pointed anterior cephalic margin (character2 state 1) flagella of the great appendage extending the lengthof the body (character 7 state 1) serrate lateral margins on thetrunk tergites (character 17 state 1) and paired tergal carinae(character 20 state 1) As such a species from Chengjiang (Lillecebrosa) and the Burgess Shale (L superlata) are moreclosely related to each other than are the two Burgess Shalecongeners a relationship that weakens the possibility that theoften cominusoccurring L superlata and L persephone could besexual dimorphs (GarciacuteaminusBellido and Collins 2007) ratherthan separate species The union of Leanchoilia and Oestominuskerkus is unambiguously supported by the shared midgutgland structure of both genera (character 26 state 1)
All shortest cladograms resolve Haikoucaris Yohoiaand Fortiforceps in that order as progressively less closelyrelated to Leanchoiliidae The union of these three taxa withLeanchoiliidae yields a clade that corresponds to Megacheirasensu Hou and Bergstroumlm (1997) apart from the ambiguousplacement of Jianfengia In half of the 14 shortest cladominusgrams Jianfengia is sister to the other megacheirans yieldminusing a monophyletic Megacheira ss In the remaining cladominusgrams Jianfengia is sister to Xenopoda sensu Hou andBergstroumlm (1997) Aglaspis and Chelicerata Haikoucarisand Leanchoiliidae are united by a frontal appendage comminusposed of two spineminusbearing articles in addition to the movminusable finger (character 5 state 1) Yohoia groups with thesetaxa based on spines fringing the telson (character 24 state1) and Fortiforceps is united with them at a broader levelbased on the flapminuslike telson shape (character 23 state 1)When Jianfengia is placed as sister to the other megacheiminusrans the entire clade is supported by the raptorial frontal apminuspendage (character 4 state 1) and the lack of a large hypominusstomal shield (character 13 state 0) In this set of cladominusgrams these features are nonminushomologous between Megaminus
cheira and Chelicerata whereas in some cladograms with thealternative resolution of Jianfengia as closer to cheliceratescharacter 4 is optimised as homologous in Megacheira andChelicerata
Reanalysis of the data with implied weights finds two optiminusmal trees (Fig 9) Relationships within Leanchoiliidae underequal weights are completely stable to reweighting as is theresolution of Haikoucaris as sister to Leanchoiliidae As suchthe character optimisations described above for the leanchoiminusliids are depicted the same in Fig 9 The main difference beminustween equal and implied weights involves the placement ofChelicerata Under equal weights (Fig 8) chelicerates are alminuslied to Xenopoda (here Emeraldella + Sidneyia + Chelominusniellon sensu clade L in Hou and Bergstroumlm 1997 fig 88)and Aglaspidida A clade composed of Aglaspidida Xenominuspoda and Chelicerata was similarly identified by Hou andBergstroumlm (1997 fig 88 their clade K) In our analysis thisgroup consistently finds support from one or more posteriortrunk segments lacking appendages (character 19 state 1) anddepending on alternative internal resolutions it is variably supminusported by dorsallyminuspositioned sessile eyes (character 8 state 1)and the absence of exopods on the first postminusantennalpostminuscheliceral limb (character 27 state 1) Under implied weightshowever chelicerates are more closely allied to Megacheirathan to aglaspidids or xenopodans (Fig 9)
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 397
2
582
76
Chelicera-morpha
Megacheira
62
2
53
50Leanchoiliidae
3
86
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
EurypteridaChelicerata
Fortiforceps
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia hanceyi
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
Fig 8 Strict consensus of 14 shortest cladograms (77 steps) based on charminusacter data in Table 1 Numbers above nodes are Bremer support those beminuslow nodes are jackknife frequencies gt50 Cheliceramorpha correspondsto a clade with the same composition in Cotton and Braddy (2004)
The major groupings in Fig 9 correspond closely to thoseidentified by Cotton and Braddy (2004) Notably the cladeuniting Aglaspis Xenopoda Offacolus Chelicerata Jianminusfengia and other Megacheira corresponds to their ldquoClade 3rdquo(Cotton and Braddy 2004 fig 8) for which they proposedthe name Cheliceramorpha The clade uniting CheloniellonAglaspis Sidneyia and Emeraldella in Fig 9 corresponds totheir ldquoClade 5rdquo and the clade that groups Megacheira andChelicerata is equivalent to their ldquoClade 4rdquo Their analysisdid not identify megacheirans as a monophyletic groupChelicerata was nested within the group and among taxaconsidered here Yohoia was resolved closer to cheliceratesthan to Leanchoiliidae In contrast Yohoia was regarded byBergstroumlm and Hou (2005) as more closely allied to Leanminuschoiliidae than to chelicerates a megacheiran grouping morecompatible with our cladograms The analysis of ldquogreat apminuspendage arthropodsrdquo by Vannier et al (2009) identified a
monophyletic Leanchoiliidae but a polyphyletic Megacheira(to the exclusion of Haikoucaris and Jianfengia) Howevertheir resolution of Fortiforceps and even more so Yohoia asthe closest relatives of Leanchoiliidae is as in our trees (apartfrom the different position of Haikoucaris)
In summary a major controversy in the systematics of theldquoshort great appendagerdquo arthropods remains whether thechelicerates are nested within the group or are instead moreclosely allied to taxa conventionally identified as lamelliminuspedians specifically Xenopoda and Aglaspidida We findthat both of these results are compatible with the availabledata depending on character weighting protocols Noneminustheless the broader megacheiranminuschelicerate issue does notcloud the status of leanchoiliids as a clade or the identificaminustion of their closest relatives (ie Haikoucaris and Yohoia)and the internal relationships of Leanchoiliidae are stableand supported by compelling characters
398 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
Eurypterida
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
2 7 17 20
3
2
1 1 1
26
1
281563
1 1 1 1
5
1
24
1
23 19 20 27 32
28Clade 4
1 0
4 13 11 27 0
154 1 1
3 19 32Clade 3
2 3
2 28
0
3 1 121
1
22
Clade 5
1 0
19 27
1
25
15 28 301 1
2312
1 150 1 3 3
15 16 2923
18
1 1
10 16
1 1
1 8
1
14
3
3
28 30
1 0
5
15
1 1
1
1
22
Fortiforceps
23
1 1 0 1
1 1
Leanchoilia hanceyi
2
2
1
1
Fig 9 Characters optimised on one of two shortest cladograms with imminusplied weights using a concavity constant k = 3 Black squares are nonminushomoplastic changes white squares homoplastic changes all changes areunambiguous Branches without strict support collapsed after optimisingcharacters Numbers for characters (above the branches) and states (belowthe branches) correspond to Table 1 ldquoClades 3ndash5rdquo correspond to groups sonumbered by Cotton and Braddy (2004 fig 8)
AcknowledgementsAccess to the field area has been most kindly allowed by the Buck famminusily We thank our collaborators in Emu Bay Shale research Jim Jago(University of South Australia Mawson Lakes) Jim Gehling andMike Lee (South Australian Museum Adelaide) for tremendous supminusport and advice as well as Mike Gemmell Dennis Rice Allison DaleyAaron Camens John Laurie Glenn Brock Pierre Kruse and Trevorand Jenny Worthy (all South Australian Museum) for their assistancein the field and Natalie Schroeder MaryminusAnne Binnie and RondaAtkinson (all South Australian Museum) for organisational support inthe lab and in the field Javier OrtegaminusHernaacutendez (University of Camminusbridge Cambridge UK) advised us on Kwanyinaspis and the journalrsquosreferees provided useful feedback TNT is available courtesy of theWilli Hennig Society Funding has been provided by an AustralianResearch Council Linkage grant (LP0774959) to the University ofAdelaide with generous financial assistance from Beach Energy Ltdand the South Australian Museum SeaLink provided logistical supminusport DGBrsquos travel to Australia has been supported by grants from theSpanish Research Council (CSIC PA 1001105) the Complutense Uniminusversity of Madrid and the Spanish Ministry of Education and Science(CGL2006minus12245BTE CGL2009minus07073 and RYC2007minus00090)
ReferencesBergstroumlm J and Hou XminusG 2005 Early Palaeozoic lamellipedian arthrominus
pods In S Koenemann and RA Jenner (eds) Crustacea and Arthrominuspod Relationships Crustacean Issues 16 73ndash93
Bitsch J and Bitsch C 2007 The segmental organization of the head reminusgion in Chelicerata a critical review of recent studies and hypothesesActa Zoologica 88 317ndash335
Bousfeld EL 1995 A contribution to the natural classification of Lower andMiddle Cambrian arthropods food gathering and feeding mechanismsAmphipacifica 2 3ndash34
Boxshall GA 2004 The evolution of arthropod limbs Biological Reviews79 253ndash300
Bremer K 1994 Branch support and tree stability Cladistics 10 295ndash304Briggs DEG and Collins D 1999 The arthropod Alalcomenaeus cambricus
Simonetta from the Middle Cambrian Burgess Shale of British ColumbiaPalaeontology 42 953ndash977
Briggs DEG and Robison RA 1984 Exceptionally preserved nonminustrilobite arthropods and anomalocaridids from the Middle Cambrian ofUtah University of Kansas Paleontological Contributions 111 1ndash23
Briggs DEG Lieberman BS Hendricks JR Halgedahl SL andJerrard RD 2008 Middle Cambrian arthropods from Utah Journal ofPaleontology 82 238ndash254
Bruton DL 1981 The arthropod Sidneyia inexpectans Middle CambrianBurgess Shale British Columbia Philosophical Transactions of theRoyal Society of London Series B 295 619ndash656
Bruton DL and Whittington HB 1983 Emeraldella and Leanchoiliatwo arthropods from the Burgess Shale Middle Cambrian British Cominuslumbia Philosophical Transactions of the Royal Society of London Seminusries B 300 553ndash582
Budd GE 2002 A palaeontological solution to the arthropod head probminuslem Nature 417 271ndash275
Butterfield NJ 2002 Leanchoilia guts and the interpretation of threeminusdiminusmensional structures in Burgess Shaleminustype fossils Paleobiology 28155ndash171
Chen JminusY 2004 The Dawn of Animal World [in Chinese] 366 pp JiangScience and Technology Press Nanjing
Chen JminusY Waloszek D and Maas A 2004 A new ldquogreatminusappendagerdquo arminusthropod from the Lower Cambrian of China and homology of cheliceratechelicerae and raptorial anterominusventral appendages Lethaia 37 3ndash20
Cotton TJ and Braddy SJ 2004 The phylogeny of arachnomorph arthrominuspods and the origin of the Chelicerata Transactions of the Royal Societyof Edinburgh Earth Sciences 94 169ndash193
Daley AC Budd GE Caron JminusB Edgecombe GD and Collins D2009 The Burgess Shale anomalocaridid Hurdia and its significancefor early euarthropod evolution Science 323 1597ndash1600
Dunlop JA 2006 New ideas about the euchelicerate stemminuslineage In CDeltshev and P Stoev (eds) European Arachnology 2005 Acta Zoominuslogica Bulgarica (Supplement 1) 9ndash27
Dunlop JA and Selden PA 1997 The early history and phylogeny of thechelicerates In RA Fortey and RH Thomas (eds) Arthropod Relaminustionships Systematics Association Special Volume 55 221ndash235
Edgecombe GD and Ramskoumlld L 1999 Relationships of CambrianArachnata and the systematic position of Trilobita Journal of Paleonminustology 73 263ndash287
Farris JS Albert VA Kaumlllersjouml M Lipcomb D and Kluge AG1996 Parsimony jackknifing outperforms neighborminusjoining Cladistics12 99ndash124
GarciacuteaminusBellido DC and Collins D 2007 Reassessment of the genusLeanchoilia (Arthropoda Arachnomorpha) from the Middle CambrianBurgess Shale British Columbia Canada Palaeontology 50 693ndash709
GarciacuteaminusBellido DC Paterson JR Edgecombe GD Jago JB GehlingJG and Lee MSY 2009 The bivalved arthropods Isoxys and Tuzoiawith softminuspart preservation from the lower Cambrian Emu Bay ShaleLagerstaumltte (Kangaroo Island Australia) Palaeontology 52 1221ndash1241
Goloboff PA 1993 Estimating character weights during tree searchCladistics 9 83ndash91
Goloboff PA Farris JS and Nixon KC 2008 TNT a free program forphylogenetic analysis Cladistics 24 774ndash786
Harzsch S and Hafner G 2006 Evolution of eye development in arthrominuspods phylogenetic aspects Arthropod Structure amp Development 35319ndash340
Hendricks JR and Lieberman BS 2008 New phylogenetic insights intothe Cambrian radiation of arachnomorph arthropods Journal of Paleminusontology 82 585ndash594
Hou XminusG and Bergstroumlm J 1997 Arthropods of the Lower CambrianChengjiang fauna southwest China Fossils and Strata 45 1ndash116
Jago JB Zang W Sun X Brock GA Paterson JR and SkovstedCB 2006 A review of the Cambrian biostratigraphy of South Austraminuslia Palaeoworld 15 406ndash423
Liu Y Hou XminusG and Bergstroumlm J 2007 Chengjiang arthropod Leanminuschoilia illecebrosa (Hou 1987) reconsidered GFF 129 263ndash272
Luo H Hu S Chen L Zhang S and Tao Y 1999 Early CambrianChengjiang fauna from Kunming region China [in Chinese] 129 ppYunnan Science and Technology Press Kunming
Luo H Hu S Zhang S and Tao Y 1997 New occurrence of the EarlyCambrian Chengjiang fauna in Haikou Kunming Yunnan Provinceand study on Trilobitoidea Acta Geologica Sinica 71 122ndash132
Nixon KC 2002 WINCLADA Version 19908 Cornell University IthacaNY
Orr PJ Siveter DJ Briggs DEG Siveter DJ and Sutton MD 2000A new arthropod from the Silurian KonservatminusLagerstaumltte of Hereminusfordshire Proceedings of the Royal Society Series B Biological Sciminusences 267 1497ndash1504
Paterson JR and Brock GA 2007 Early Cambrian trilobites fromAngorichina Flinders Ranges South Australia with a new assemblagefrom the Pararaia bunyerooensis Zone Journal of Paleontology 81116ndash142
Paterson JR and Jago JB 2006 New trilobites from the Lower CambrianEmu Bay Shale Lagerstaumltte at Big Gully Kangaroo Island South Ausminustralia Memoirs of the Association of Australasian Palaeontologists 3243ndash57
Paterson JR Edgecombe GD GarciacuteaminusBellido DC Jago JB andGehling JG 2010 Nektaspid arthropods from the lower CambrianEmu Bay Shale Lagerstaumltte South Australia with a reassessment oflamellipedian relationships Palaeontology 53 377ndash402
Paterson JR Jago JB Gehling JG GarciacuteaminusBellido DC Edgecombe
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 399
GD and Lee MSY 2008 Early Cambrian arthropods from the EmuBay Shale Lagerstaumltte South Australia 319ndash325 In I Raacutebano RGozalo and D GarciacuteaminusBellido (eds) Advances in Trilobite ResearchCuadernos del Museo Geominero 9 319ndash325
Shultz JW 2007 A phylogenetic analysis of the arachnid orders based onmorphological characters Zoological Journal of the Linnean Society150 221ndash265
Stein M 2010 A new arthropod from the Early Cambrian of North Greenminusland with a ldquogreat appendagerdquominuslike antennula Zoological Journal ofthe Linnean Society 158 477ndash500
Stuumlrmer W and Bergstroumlm J 1978 The arthropod Cheloniellon from theDevonian Hunsruumlck Shale Palaumlontologische Zeitschrift 52 57ndash81
Sutton MD Briggs DEG Siveter DJ Siveter DJ and Orr PD2002 The arthropod Offacolus kingi (Chelicerata) from the Silurian ofHerefordshire England computer based morphological reconstrucminustions and phylogenetic affinity Proceedings of the Royal Society B Biminusological Sciences 269 1195ndash1203
Suzuki Y and Bergstroumlm J 2008 Respiration in trilobites a reevaluationGFF 130 211ndash229
Vannier J GarciacuteaminusBellido DC Hu XminusX and Chen AminusL 2009 Arthrominuspod visual predators in the early pelagic ecosystem evidence from the
Burgess Shale and Chengjiang biotas Proceedings of the Royal SocietyB Biological Sciences 276 2567ndash2574
Whittington HB 1974 Yohoia Walcott and Plenocaris n gen arthropodsfrom the Burgess Shale Middle Cambrian British Columbia Geologiminuscal Survey of Canada Bulletin 231 1ndash21
Wills MA Briggs DEG Fortey RA Wilkinson M and SneathPHA 1998 An arthropod phylogeny based on fossil and recent taxaIn GD Edgecombe (ed) Arthropod Fossils and Phylogeny 33ndash105Columbia University Press New York
Wilson LA 2006 Food for thought a morphological and taphonomic studyof fossilised digestive systems from Early to Middle Cambrian taxa275 pp Unpublished PhD thesis University of Cambridge Cambridge
Zhang XminusG and Hou X 2007 Gravitational constraints on the burial ofChengjiang fossils Palaios 22 448ndash453
Zhang XminusL Han J Zhang Z Liu H and Shu D 2004 Redescription ofthe Chengjiang arthropod Squamacula clypeata Hou and Bergstroumlm fromthe Lower Cambrian Southminuswest China Palaeontology 47 605ndash617
Zhao Y Zhu M Babcock LE Yuan L Parsley RL Peng J YangX and Wang Y 2005 Kaili Biota a taphonomic window on diversifiminuscation of metazoans from the basal Middle Cambrian Guizhou ChinaActa Geologica Sinica 79 751ndash765
400 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011

SAM P45904a b (unfigured) poorlyminuspreserved cephalic shield trunkwith wellminuspreserved midgut glands proximal part of telson SAMP45905ab (unfigured) cephalic shield lacking great appendages butexposing an eye trunk with incomplete telson SAM P45906a b (unfigminusured) great appendages with spinose projections oriented anteriorlytrunk preserved to end of midgut glands SAM P45908a b (unfigured)cephalic shield great appendages with flagella most of trunk (telsonmissing) showing midgut glands and exopod setae SAM P45909 (unminusfigured) small specimen with great appendages and series of mineralminusised midgut glands SAM P45910a b (unfigured) incomplete great apminuspendages oriented anteriorly trunk with complete series of midgutglands but indistinct appendages
Type locality Buck Quarry Big Gully (Fig 1)
Type horizon Emu Bay Shale (Cambrian Series 2 Stage 4)
DiagnosismdashAs for genus
DescriptionmdashCephalic shield 36 length of the trunk sagitminustally dorsal margin of cephalic shield gently convex in lateralview anteromedian margin inferred to be evenly rounded(based on comparison with lateral versus dorsal preservation ofBurgess Shale Leanchoilia and Alalcomenaeus) lacking an upminusturned median process Eye visible in SAM P54167 (Fig 4A)and P45905 relatively large (20 sagittal length of cephalon)exposed under anterolateral part of cephalic shield (Fig 4A2)
390 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Fig 4 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov SAM P45167 A P45167a A1 cephalic shield and most of trunkA2 detail of cephalic shield B P45167b A2 and B light from NE Overlap between adjacent tergites offset on left (Lol) and right (Rol) sides of bodyBoundaries between cephalic shield and anterior trunk tergites delimited in B by dashed lines Arrows mark positions of inferred dorsal transverse tendonsAbbreviations cn1ndash2 cephalic endopods cs cephalic shield gap peduncle of great appendage ol overlap between adjacent trunk tergites t1ndasht3 tergitesof trunk segments 1ndash3 tn trunk endopods
Fig 5 Leanchoiliid megacheiran arthropod Oestokerkus megacholix genet sp nov A SAM P 15531 B SAM P43630b Light from NE Abbreviaminustions cs cephalic shield ga great appendage gap peduncle of great apminuspendage mg midgut glands ol overlap between adjacent trunk tergites txtrunk exopods
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 391
Great appendage robust proportions of peduncle and disminustal spineminusbearing articles as in other members of Leanchoiliminusidae more commonly preserved with posterior orientation
but anteriorly oriented in SAM P45906 and SAM P45910Each of the three long spinose projections terminating at thesame length the projections evenly tapering distally with
392 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Fig 6 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov SAM P45168 A head and anterior part of trunk B Detail of spinoseprojections of great appendages and excretory organs (sensu Bruton and Whittington 1983) Abbreviations cs cephalic shield eo excretory organ gagreat appendage mg midgut glands tx trunk exopods
margins straight for much of their length or slightly curvedterminally distal spinose projection distinctly stouter at itsbase than the two more proximal projections and more markminusedly tapering along its length Dark irregularminusshaped low reminuslief lump on basal spineminusbearing article of great appendagesof SAM P45168 (eo in Fig 6) and SAM P45908 correminussponding to excretory organ of Bruton and Whittington(1983) Long flagella extending from each of the threespinose projections seen only in SAM P43631 (Figs 23A3) of similar narrow width along its preserved extent anminusnulations indistinguishable
Trunk of 11 segments of even length Maximum height oftrunk relative to length (excluding telson) ca 39 bodydeepest at tergite 4 Trunk tergite boundaries marked as narminusrow ridges in several specimens corresponding to narrowband of overlap between adjacent tergites (ol in Figs 2 3A14A1 B) Some specimens show tergites to be bisected by anarrow but sharp furrow of uncertain identity (eg possiblyimpression of dorsal transverse tendon cf Stein 2010 fig12B C E) (arrows in Figs 3A1 4B)
Exopods with fringe of long setae on cephalic and trunkappendages (Fig 2) setal fringe gradually shortening fromabout the fourth trunk segment Setae project ventrally bymore than 30 height of tergites in deepest part of trunk(Fig 2B) setae flattened narrow and parallelminussided alongmost of their length tapering near their distal ends to termiminusnate as a pointed tip (Figs 2B 3A3 arrows) Endopods exminusposed only in SAM P45167 (Fig 4) two pairs associatedwith cephalon (Fig 4B) those of anterior segments of trunkof similar morphology to those in cephalon all slender lackminusing obvious endites podomere boundaries unrecognisable
Midgut glands occupy a large extent of the dorsoventralheight of the trunk two midgut glands in cephalon of similarsize to those in each of the first seven segments of the trunk(Figs 3A1 5A 6A) all developed as radiating grooves emaminusnating from a globular structure at their ventral edge as in
Leanchoilia superlata (GarciacuteaminusBellido and Collins 2007 pl2 6) Foregut trace in SAM P43631 (Fig 2) indicates that gutcurves ventrally in anterior part of cephalon mouth regionnot preserved Terminal part of hindgut (in trunk segment 11)with closely spaced longitudinal lineations (Fig 2) that apminusparently correspond to folds in the gut wall hindgut termiminusnating at base of telson
Telson most completely preserved in holotype (SAMP43631 Figs 2B 3A2 B) In this specimen the telson likerest of body compressed in lateral aspect Telson shows meminusdian and marginal reinforcing ridges connecting a flat trianminusgular surface this surface appears to be partially broken inSAM P43631 showing underlying surface of other (left) halfof telson Lateral preservation of specimen implies that topcorresponds to an axial structure and bottom to a marginalone 30ndash35 angle in posterior edge of vane connecting thesetwo structures requires that in dorsal aspect telson would apminuspear forked and widening distally Five stout pointed dorminussally curved marginal spines attached to outer posterior edgeon each side of telson
Phylogenetic analysis ofLeanchoiliidaeDetermining the systematic position of Oestokerkus demandsan appraisal of the relationships of Leanchoiliidae as a wholeCladistic analysis including the bestminusknown members ofMegacheira including leanchoiliids has been conducted byCotton and Braddy (2004) and Vannier et al (2009) The taxominusnomic sampling used by Cotton and Braddy (2004) includedseveral megacheiran genera representative chelicerates anumber of taxa representing groups that have been proposedas possible chelicerate relatives (eg Sidneyia AglaspisCheloniellon) in other analyses (Dunlop and Selden 1997Hou and Bergstroumlm 1997) and members of the major lineagesof the Artiopoda sensu Hou and Bergstroumlm (1997) ie trilominusbiteminusallied lamellipedians Because artiopodan interminusrelationminusships have remained quite stable through a series of phylogenminusetic analyses (Edgecombe and Ramskoumlld 1999 Cotton andBraddy 2004 Hendricks and Lieberman 2008 Paterson et al2010) and artiopodans are now often regarded as less closelyrelated to Megacheira than is Chelicerata (Chen et al 2004Cotton and Braddy 2004 Dunlop 2006) we have reduced thesampling of Artiopoda in the present analysis To augment thediversity of ldquoshort great appendagerdquo arthropods Haikoucarisis added based on the description of H ercaiensis by Chen etal (2004) Offacolus (Orr et al 2000 Sutton et al 2002) hasbeen allied to Chelicerata and is included as a terminal Beminuscause the present analysis is principally concerned with relaminustionships of and within Leanchoiliidae sensu Liu et al (2007)we follow Vannier et al (2009) in sampling Alalcomenaeusand Leanchoilia at the species level Within Chelicerata Cotminuston and Braddy (2004) and Hendricks and Lieberman (2008)sampled ancestral states in Xiphosura by coding the Devonian
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 393
Fig 7 Reconstruction of Oestokerkus megacholix gen et sp nov in lateralview Total length and annulations on the flagella and positions of podominusmere boundaries of the frontal appendage are based on Leanchoilia superminuslata (Bruton and Whittington 1983)
synziphosurine Weinbergina Our coding for Xiphosura alsoemploys information from extant xiphosurans because theysupply more reliable information on the homology of the partsof the trunk appendages (Suzuki and Bergstroumlm 2008) and eyeanatomy (Harzsch and Hafner 2006) among other charactersystems than do fossils alone
Our character set extracts characters from the matrix ofCotton and Braddy (2004) that vary in the revised taxonomicsample reformulates some characters to accommodate reminusvised homology concepts and adds new characters especiallyfor leanchoiliids In total 32 characters are included (Table 1)In this analysis the primary homology of the ldquoshort great apminuspendagerdquo and chelicera is coded (as described by character 4)and the taxon sampling broadly constrains megacheirans aschelicerate relatives (as defended by Chen et al 2004 Cottonand Braddy 2004) rather than testing the alternative that theycould be stem group Euarthropoda (Budd 2002 Bergstroumlmand Hou 2005) A test of the two alternatives involves samminuspling of the entire diversity of euarthropods and a range ofnonminuseuarthropod outgroups an exercise beyond the scope ofthis study
Characters1 Mineralised cuticle (0) absent (1) present For definiminus
tion and discussion of this character see Cotton andBraddy (2004 character 30) and Paterson et al (2010character 1)
2 Curvature of anterior margin of cephalic shield (0) rounminusded or straight (1) pointed medially (2) with multiplespinose projections This character codes for variability inthe shape of the cephalic shield in Leanchoiliidae (state 0and 1 sensu GarciacuteaminusBellido and Collins [2007 textminusfigs10 and 9 respectively]) State 2 is added to accommodateOffacolus the median spine of a series that fringes the ceminusphalic margin is not convincingly homologised with themedian projection in leanchoiliids
3 Number of cephalic somitesappendages (0) antenna orfrontal appendage only (1) antenna or frontal appendageand two limb pairs (2) antenna or frontal appendage andthree limb pairs (3) antenna or frontal appendage and fourlimb pairs (4) antenna or frontal appendage and five limbpairs Coding for lamellipedians and Aglaspis is as discussedby Paterson et al (2010 character 2) Coding for Alalcomeminusnaeus cambricus (state 1) follows Briggs and Collins (1999)Leanchoilia illecebrosa (state 2) follows Liu et al (2007)and Leanchoilia persephone and Leanchoilia superlata (state1) follows GarciacuteaminusBellido and Collins (2007)
4 Form of first cephalic appendage (0) antenna (1) anteminusriorly directed raptorial appendage with reduced numberof podomeres each of the terminal podomeres bearing aspinose process on its distal margins a geniculation beminustween spinose podomeres and peduncle This characteris formulated to identify a homology between the frontalappendages of megacheirans and the chelicera of Cheliminus
cerata and state 1 corresponds to the putative apomorminusphic detail defended by Chen et al (2004) and Cottonand Braddy (2004) Vannier et al (2009 character 8)specify another shared property of ldquoshort great appendminusagesrdquo of the megacheiran (ss) type in their characterstate ldquobent at an anglerdquo referring to the geniculation beminustween articles of the peduncle or between the peduncleand distal spiniferous articles This property is sharedwith Chelicerata
5 Number of spineminusbearing articles on frontal appendage (0)three + movable finger (1) two + movable finger (2) one+ movable finger The number of spineminusbearing articles onthe frontal appendage (exclusive of the terminal movablefinger) varies from three in Yohoia (Whittington 1974 pl6 1ndash3 pl 7 3ndash6 pl 9 4ndash6) and Fortiforceps (Hou andBergstroumlm 1997 figs 31C D 32A 33B C E) to two inHaikoucaris (Chen et al 2004 fig 2A B) and Leanminuschoiliidae or only one the last being a chelicera
6 Long spinose projections on distal part of terminal threepodomeres of frontal appendage bearing a flagellum (0)absent (1) present The morphology of the great appendminusage in Leanchoiliidae is coded here We have not codedthe length of the spinose projections and the presence ofa flagellum as two independent characters Cotton andBraddyrsquos (2004) character 6 state 2 (ldquospinesrdquo longer thanthe entire podomere series) and character 8 state 1 (presminusence of an annulated flagellum) are thus grouped as a sinminusgle trait here
394 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Table 1 Characters coded for phylogenetic analysis Polymorphismindicated by ldquo+rdquo partial uncertainty by states separated by ldquordquo Abbreviminusations A Alalcomenaeus L Leanchoilia O Oestokerkus
Squamacula 0000minus minus minus2minus0 0110 00000 10minus minus0 1000 0
Cindarella 0030minus minus minus000 0010 00000 10minus minus0 11121 00
Kuamaia 000minus minus minus001 0011 10000 10minus minus0 11121 00
Kwanyinaspis 00 001 00112 1000 1000 1121 00
Olenoides 1020minus minus minus100 0011 00000 10minus minus0 01121 00
Retifacies 0020minus minus minus000 0010 00000 11000 1020 00
Aglaspis 10(23)0minus minus minus100 00101 00010 01001 0 00
Sidneyia 0000minus minus minus000 01102 00110 01101 00020 00
Emeraldella 0040minus minus minus2minus 0 00103 00110 01001 001(1+2)1 00
A cambricus 00111 10010 00001 00000 01110 01110 10
L illecebrosa 01211 11010 00001 01001 01210 11110 10
L persephone 00111 10010 00001 00000 01210 11110 10
L superlata 01111 11010 00001 01001 01210 11110 10
L hanceyi 00 00 0001 0000 01210
O megacholix 0011 100 00001 00000 01110 1110 0
Jianfengia 0021 0minus000 00005 00000 01000 010(12)0 0
Fortiforceps 00210 0minus00 00004 00000 01100 01230 20
Yohoia 00210 0minus000 00003 00011 01110 00010 (01)1
Haikoucaris 00211 0minus00 00003 00000 01(12)0 1010 0
Cheloniellon 0040minus minus minus100 00100 10010 01300 0030 00
Offacolus 02412 0minus200 0000 00010 01000 13minus minus 01
Xiphosura 00412 0minus100 10000 0010 01000 000minus minus 01
Eurypterida 00412 0minus100 10002 00010 01000 00minus minus minus 01
7 Length of flagellate great appendage (0) flagella termiminusnating at midlength of trunk (1) flagella extending to endof body Coding is applicable to Leanchoiliidae onlydistinguishing some Leanchoilia species that resembleAlalcomenaeus in having relative short great appendages(state 0) versus the longer appendages in L superlata andL illecebrosa
8 Position of lateral facetted eyes (0) ventral (1) dorsalsessile (2) absent Variation in Lamellipedia and severalother taxa coded herein follows Paterson et al (2010character 3)
9 Four or five ventral eyes arranged in a subtransverseband across head shield (0) absent (1) present A bandof ventral eyes is shared by the best known species ofLeanchoiliidae The distinction between three medianeyes in Alalcomenaeus cambricus (Briggs and Collins1999) and two in Leanchoilia illecebrosa (Liu et al2007) and both Leanchoilia superlata and L persephone(GarciacuteaminusBellido and Collins 2007) is uninformative forthe taxon sample here
10 Bulge in exoskeleton accommodating dropminusshaped venminustral eyes (0) absent (1) present This character followsEdgecombe and Ramskoumlld (1999 character 6) and Patminuserson et al (2010 character 5)
11 Dorsal median eyes (0) absent (1) present Coding for anabsence of dorsal median eyes in fossil taxa considers theabsence of either paired ocelli or a median eye tubercleand the character is scored as a chelicerate autapomorphy(see Cotton and Braddy 2004 character 24)
12 Width of doublure on cephalic shield (0) narrow or modminuserately wide (1) entire ventral side of head covered byexpansion of doublure The interpretation of state 1 inSidneyia and Squamacula is discussed by Cotton andBraddy (2004) and Paterson et al (2010 characters 12and 29)
13 Mouth covered by hypostome (0) absent (1) present Noldquoshort great appendagerdquo arthropods have been shown topossess a hypostome and a reconstruction of a smalllabrum in Alalcomenaeus (Briggs and Collins 1999) wasadmittedly conjectural The absence of a conspicuoushypostome contrasts with its ubiquitous presence in laminusmellipedians and several putative chelicerateminusallied taxathat have antennae (such as aglaspidids and Emeralminusdella) Chelicerates have a small epistomeminuslabrum (Bitschand Bitsch 2007) but like Cotton and Braddy (2004character 27 state 4) we code a hypostomal sclerite aslacking Kwanyinaspis has a large hypostome similar tothat found in aglaspidids (photograph of the holotype ofKwanyinaspis maotianshanensis shown to us by JavierOrtegaminusHernaacutendez [personal communication 2010])
14 Tergite articulations (0) extensive overlap of tergites (1)edge to edge articulations This character follows Edgeminuscombe and Ramskoumlld (1999 character 18) and Patersonet al (2010 character 20)
15 Number of trunk segments (0) nine (1) 11 (2) 12 (3)13 (4) 20 (5) 21 or 22 Coding is confined to taxa that do
not possess a pygidium because the precise number ofsomites associated with pygidia can sometimes not bedetermined with confidence Codings for Xiphosura andEurypterida use opisthosomal somite numbers followingShultz (2007 character 95)
16 Exoskeletal boundaries of anterior trunk segments (0)boundaries transverse or reflexed posterolaterally (1) reminusflexed anterolaterally This character follows Edgecombeand Ramskoumlld (1999 character 19) and Paterson et al(2010 character 21)
17 Trunk tergites with serrate lateral margins (0) absent (1)present Liu et al (2007 fig 2C) documented a row ofmarginal spines on the tergites of Leanchoilia illeceminusbrosa a featured shared by Leanchoilia superlata (egGarciacuteaminusBellido and Collins 2007 pl 1 7) but not otherterminals coded here
18 Differentiation in width of trunk tergites (0) trunk withgradational narrowing posteriorly (1) abrupt narrowingat boundary between prothorax and opisthothorax Anabrupt contraction in width between the boundary ofthe prominus and opisthothorax is observed in Emeraldellaand Sidneyia Yohoia has been reconstructed as having apronounced break in the shape of the trunk tergites beminushind segment 10 (Whittington 1974 textminusfig 2) thepoint at which the paratergal folds lose their spinose tipsand appendages are lacking Specimens preserved indorsal aspect (Whittington 1974 pl 10) suggest that thewidth of the opisthothorax is rather less abruptly narminusrowed and we code Yohoia as state 0
19 Appendages of posterior trunk segments (0) all trunktergites bearing appendages (1) one or more posteriortrunk segments lacking appendages Coding correspondsto Cotton and Braddyrsquos (2004) character 43 ldquopostabdominusmen of segments lacking appendagesrdquo
20 Paired tergal carinae (0) absent (1) present Liu et al(2007 265 fig 2F) recorded the presence of tergalcarinae in Leanchoilia illecebrosa shared with L superminuslata (eg Bruton and Whittington 1983 pl 17 106GarciacuteaminusBellido and Collins 2007 pl 1 5)
21 Pygidium (0) absent (1) present Coding of the pygiminusdium in many terminals herein was discussed by Paterminusson et al (2010 character 22)
22 Telson (0) absent (1) present Following Hendricks andLieberman (2008) a telson is a postminussegmental terminalstructure that lacks appendages Dunlop and Selden (1997)considered a telson to be absent in Cheloniellon but weidentify the conical projection posterior to the ultimate legpair and cerci (Stuumlrmer and Bergstroumlm 1978 figs 2 3 1011) as being most consistent with an identity as a telson
23 Telson shape (0) styliform (1) flapminuslike (2) lanceolate(3) short conical The outline of the telson in Haikouminuscaris ercaiensis is incompletely known but the mostcomplete specimen (Chen 2004 fig 472) constrains it aseither state 1 or state 2 A partial uncertainty coding isthus employed A unique state for Cheloniellon (state 3)is based on the interpretation in character 22
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 395
24 Telson fringed with spines (0) spines absent (1) spinespresent Presence of telson spines was introduced as a charminusacter by Hendricks and Lieberman (2008 character 34)
25 Paired flaps lateral to telson (0) absent (1) present Thischaracter describes the postminusventral plates of several taxa(see Edgecombe and Ramskoumlld 1999 character 25) Cotminuston and Braddy (2004 character 54) raised the possibilitythat the cerci of Cheloniellon could be homologous withpostminusventral plates (see also Hou and Bergstroumlm 1997 93)because they are similarly paired structures derived fromthe segment immediately anterior to the telson We havenot coded for this homology because the cerci of Chelominusniellon originate dorsally rather than ventrally
26 Midgut glands with submillimetric lamellae (0) absent(1) present Midgut glands in some stem group arthrominuspods and early crown group arthropods are metamericreniform organs with a characteristic lamellar structure(Butterfield 2002) Their morphology is identical inLeanchoilia and Oestokerkus notably including groovesthat radiate from a ventral median node (Figs 3A1 6A)The gut of Alalcomenaeus cambricus is more tubular(Briggs and Collins 1999 textminusfigs 10 11) and thoughan anterior six segments may display phosphatised latminuseral projections that likely represent gut diverticula (Wilminusson 2006) they have not been shown to have the detailedmorphology of Leanchoiliaminustype midgut glands Butterminusfield (2002 164) cited a figure from Bruton (1981 fig96) in support of ldquolamellarrdquo midgut glands in the cephaminuslon of Sidneyia Like Bruton (1981) in his description ofthe specimen we consider it likely that the structures inquestion are indeed gut diverticula but there is no basisfor identifying them more precisely as the reniformlamellar structures coded under this character
27 Exopod on first postminusantennal first postminusfrontal appendminusage segment (0) absent (1) present This character reminuslates to Cotton and Braddyrsquos (2004) character 3
28 Division of exopod (0) exopod an undivided flap (1)exopod differentiated into proximal and distal lobes (2)small flap with radiating lamellae (3) pediform exopoddivided into ca 6 articles Identification of the exopod inXiphosura follows Suzuki and Bergstroumlm (2008) Thischaracter groups conditions of the exopod shaft coded asindependent characters by Cotton and Braddy (2004)eg state 0 corresponds to their character 16 and state 1to their character 13 Our coding permits transformationbetween any two states without an extra implicit step
29 Exopod setae (0) short spiniform setae (1) long flatminustened tapering distally with slight separation (2)lamellar setae (flattened imbricating) (3) short finesetae A distinction is made between lamellipedian setaewhich are imbricated (state 2) and the pointed more disminustinctly separated and variably spiniform exopod setae ofleanchoiliids (state 1) This distinction is consistent withthe identification of megacheirans as nonminuslamellipedians(eg Hou and Bergstroumlm 1997 Bergstroumlm and Hou2005) Although Cotton and Braddy (2004 character 18)
regarded the form of the setae in Yohoia to be uncertainwe regard specimens figured by Whittington (1974 pl3 3) as conforming to the leanchoiliid state Squamaculadiffers from other taxa in its shorter spiniform setae(state 0) (Zhang et al 2004 textminusfig 1A B) Offacolushas longer spiniform projections on the distal articles ofthe exopod that have been variably identified as eitherspines or setae (Sutton et al 2002 fig 4b) but theirhomology with the setae that splay regularly along themargin of a flapminuslike exopod (eg states 0ndash3) is uncerminustain This and the following character for exopod setaeare left as ldquordquo for Offacolus A homology between thegill lamellae of Xiphosura and exopod setae of lamellipeminusdians has a long history (see Cotton and Braddy 2004character 18) We follow Suzuki and Bergstroumlm (2008)in identifying the xiphosuran gills as outgrowths of thebasis rather than of the exopod this perspective negates ahomology with the exopod setae of Lamellipedia Thepolymorphic coding for Emeraldella reflects the twoseparate states (1+2) displayed by the exopods in thetrunk (see Bruton and Whittington 1983 pl 8 45 47Edgecombe and Ramskoumlld 1999 fig 11)
30 Differentiation of setae along exopod (0) uniform orgradationally variable setae along length of exopod (1)lamellar setae confined to proximal lobe with distal lobebearing slender setae
31 Endopod segmentation (distal to coxa) (0) six or sevenpodomeres and terminal claw or telotarsal spurs (1)eight or more podomeres and terminal claw (2) ca 15podomeres State 1 is formulated to describe the segmenminustation of the leanchoiliid leg (GarciacuteaminusBellido and Collins2007 textminusfig 6 Liu et al 2007 fig 5) in contrast to thereduced number of podomeres in other euarthropods(Boxshall 2004) State 2 characterises Fortiforceps Apartial uncertainty coding is employed for Yohoia inwhich the precise number of podomeres is undeterminedand could be either state 0 or 1 (Whittington 1974 pl 74) but the spacing of the preserved podomere boundariesnegates the possibility of state 2
32 Endopods on anterior postminuscephalic segments (0) welldeveloped (1) rudimentary or lacking Following Cottonand Braddy (2004 character 11) the absence of trunkendopods in Yohoia (Whittington 1974) is coded as a puminustative homologue with the suppression of postminuscephalicendopods in Chelicerata
ResultsAs a convention for depicting the trees networks are rootedon Squamacula which separates artiopodan Lamellipedia(sensu Hou and Bergstroumlm 1997) from a group that includesMegacheira Chelicerata and Xenopoda all members ofwhich possess a telson
Under equal weights 14 shortest cladograms have a lengthof 77 steps with a Consistency Index of 064 and Retention Inminus
396 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
dex of 077 The strict consensus of these trees with supportvalues is shown in Fig 8 Leanchoiliidae is unambiguouslysupported in all 14 cladograms by four apomorphic characterstwo limbminusbearing cephalic segments posterior to the frontalappendage (character 3 state 1) long spinose projections onthe distal part of the terminal three podomeres of the frontalappendage that bear flagella (character 6 state 1) 11 trunksegments (character 15 state 1) and division of the exopodsinto two lobes (character 28 state 1) The arrangement of fouror five eyes in a subtransverse band across the cephalic shieldis unique to Leanchoiliidae (character 9 state 1) but is not anunambiguous autapomorphy of the group because the state isuncertain in close relatives notably Haikoucaris It is apominusmorphic for Leanchoiliidae only under particular optimisaminustions (Delayed Transformation)
The internal relationships of Leanchoiliidae are stableacross all 14 shortest cladograms Leanchoilia is united by thelanceolate telson (character 23 state 2) a unique unambiguminusous autapomorphy Within Leanchoilia a sister group relaminustionship between L superlata and L illecebrosa is well supminusported receiving a Bremer support of 3 and jackknife freminusquency of 86 These two species share four synapomorminusphies a medially pointed anterior cephalic margin (character2 state 1) flagella of the great appendage extending the lengthof the body (character 7 state 1) serrate lateral margins on thetrunk tergites (character 17 state 1) and paired tergal carinae(character 20 state 1) As such a species from Chengjiang (Lillecebrosa) and the Burgess Shale (L superlata) are moreclosely related to each other than are the two Burgess Shalecongeners a relationship that weakens the possibility that theoften cominusoccurring L superlata and L persephone could besexual dimorphs (GarciacuteaminusBellido and Collins 2007) ratherthan separate species The union of Leanchoilia and Oestominuskerkus is unambiguously supported by the shared midgutgland structure of both genera (character 26 state 1)
All shortest cladograms resolve Haikoucaris Yohoiaand Fortiforceps in that order as progressively less closelyrelated to Leanchoiliidae The union of these three taxa withLeanchoiliidae yields a clade that corresponds to Megacheirasensu Hou and Bergstroumlm (1997) apart from the ambiguousplacement of Jianfengia In half of the 14 shortest cladominusgrams Jianfengia is sister to the other megacheirans yieldminusing a monophyletic Megacheira ss In the remaining cladominusgrams Jianfengia is sister to Xenopoda sensu Hou andBergstroumlm (1997) Aglaspis and Chelicerata Haikoucarisand Leanchoiliidae are united by a frontal appendage comminusposed of two spineminusbearing articles in addition to the movminusable finger (character 5 state 1) Yohoia groups with thesetaxa based on spines fringing the telson (character 24 state1) and Fortiforceps is united with them at a broader levelbased on the flapminuslike telson shape (character 23 state 1)When Jianfengia is placed as sister to the other megacheiminusrans the entire clade is supported by the raptorial frontal apminuspendage (character 4 state 1) and the lack of a large hypominusstomal shield (character 13 state 0) In this set of cladominusgrams these features are nonminushomologous between Megaminus
cheira and Chelicerata whereas in some cladograms with thealternative resolution of Jianfengia as closer to cheliceratescharacter 4 is optimised as homologous in Megacheira andChelicerata
Reanalysis of the data with implied weights finds two optiminusmal trees (Fig 9) Relationships within Leanchoiliidae underequal weights are completely stable to reweighting as is theresolution of Haikoucaris as sister to Leanchoiliidae As suchthe character optimisations described above for the leanchoiminusliids are depicted the same in Fig 9 The main difference beminustween equal and implied weights involves the placement ofChelicerata Under equal weights (Fig 8) chelicerates are alminuslied to Xenopoda (here Emeraldella + Sidneyia + Chelominusniellon sensu clade L in Hou and Bergstroumlm 1997 fig 88)and Aglaspidida A clade composed of Aglaspidida Xenominuspoda and Chelicerata was similarly identified by Hou andBergstroumlm (1997 fig 88 their clade K) In our analysis thisgroup consistently finds support from one or more posteriortrunk segments lacking appendages (character 19 state 1) anddepending on alternative internal resolutions it is variably supminusported by dorsallyminuspositioned sessile eyes (character 8 state 1)and the absence of exopods on the first postminusantennalpostminuscheliceral limb (character 27 state 1) Under implied weightshowever chelicerates are more closely allied to Megacheirathan to aglaspidids or xenopodans (Fig 9)
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 397
2
582
76
Chelicera-morpha
Megacheira
62
2
53
50Leanchoiliidae
3
86
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
EurypteridaChelicerata
Fortiforceps
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia hanceyi
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
Fig 8 Strict consensus of 14 shortest cladograms (77 steps) based on charminusacter data in Table 1 Numbers above nodes are Bremer support those beminuslow nodes are jackknife frequencies gt50 Cheliceramorpha correspondsto a clade with the same composition in Cotton and Braddy (2004)
The major groupings in Fig 9 correspond closely to thoseidentified by Cotton and Braddy (2004) Notably the cladeuniting Aglaspis Xenopoda Offacolus Chelicerata Jianminusfengia and other Megacheira corresponds to their ldquoClade 3rdquo(Cotton and Braddy 2004 fig 8) for which they proposedthe name Cheliceramorpha The clade uniting CheloniellonAglaspis Sidneyia and Emeraldella in Fig 9 corresponds totheir ldquoClade 5rdquo and the clade that groups Megacheira andChelicerata is equivalent to their ldquoClade 4rdquo Their analysisdid not identify megacheirans as a monophyletic groupChelicerata was nested within the group and among taxaconsidered here Yohoia was resolved closer to cheliceratesthan to Leanchoiliidae In contrast Yohoia was regarded byBergstroumlm and Hou (2005) as more closely allied to Leanminuschoiliidae than to chelicerates a megacheiran grouping morecompatible with our cladograms The analysis of ldquogreat apminuspendage arthropodsrdquo by Vannier et al (2009) identified a
monophyletic Leanchoiliidae but a polyphyletic Megacheira(to the exclusion of Haikoucaris and Jianfengia) Howevertheir resolution of Fortiforceps and even more so Yohoia asthe closest relatives of Leanchoiliidae is as in our trees (apartfrom the different position of Haikoucaris)
In summary a major controversy in the systematics of theldquoshort great appendagerdquo arthropods remains whether thechelicerates are nested within the group or are instead moreclosely allied to taxa conventionally identified as lamelliminuspedians specifically Xenopoda and Aglaspidida We findthat both of these results are compatible with the availabledata depending on character weighting protocols Noneminustheless the broader megacheiranminuschelicerate issue does notcloud the status of leanchoiliids as a clade or the identificaminustion of their closest relatives (ie Haikoucaris and Yohoia)and the internal relationships of Leanchoiliidae are stableand supported by compelling characters
398 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
Eurypterida
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
2 7 17 20
3
2
1 1 1
26
1
281563
1 1 1 1
5
1
24
1
23 19 20 27 32
28Clade 4
1 0
4 13 11 27 0
154 1 1
3 19 32Clade 3
2 3
2 28
0
3 1 121
1
22
Clade 5
1 0
19 27
1
25
15 28 301 1
2312
1 150 1 3 3
15 16 2923
18
1 1
10 16
1 1
1 8
1
14
3
3
28 30
1 0
5
15
1 1
1
1
22
Fortiforceps
23
1 1 0 1
1 1
Leanchoilia hanceyi
2
2
1
1
Fig 9 Characters optimised on one of two shortest cladograms with imminusplied weights using a concavity constant k = 3 Black squares are nonminushomoplastic changes white squares homoplastic changes all changes areunambiguous Branches without strict support collapsed after optimisingcharacters Numbers for characters (above the branches) and states (belowthe branches) correspond to Table 1 ldquoClades 3ndash5rdquo correspond to groups sonumbered by Cotton and Braddy (2004 fig 8)
AcknowledgementsAccess to the field area has been most kindly allowed by the Buck famminusily We thank our collaborators in Emu Bay Shale research Jim Jago(University of South Australia Mawson Lakes) Jim Gehling andMike Lee (South Australian Museum Adelaide) for tremendous supminusport and advice as well as Mike Gemmell Dennis Rice Allison DaleyAaron Camens John Laurie Glenn Brock Pierre Kruse and Trevorand Jenny Worthy (all South Australian Museum) for their assistancein the field and Natalie Schroeder MaryminusAnne Binnie and RondaAtkinson (all South Australian Museum) for organisational support inthe lab and in the field Javier OrtegaminusHernaacutendez (University of Camminusbridge Cambridge UK) advised us on Kwanyinaspis and the journalrsquosreferees provided useful feedback TNT is available courtesy of theWilli Hennig Society Funding has been provided by an AustralianResearch Council Linkage grant (LP0774959) to the University ofAdelaide with generous financial assistance from Beach Energy Ltdand the South Australian Museum SeaLink provided logistical supminusport DGBrsquos travel to Australia has been supported by grants from theSpanish Research Council (CSIC PA 1001105) the Complutense Uniminusversity of Madrid and the Spanish Ministry of Education and Science(CGL2006minus12245BTE CGL2009minus07073 and RYC2007minus00090)
ReferencesBergstroumlm J and Hou XminusG 2005 Early Palaeozoic lamellipedian arthrominus
pods In S Koenemann and RA Jenner (eds) Crustacea and Arthrominuspod Relationships Crustacean Issues 16 73ndash93
Bitsch J and Bitsch C 2007 The segmental organization of the head reminusgion in Chelicerata a critical review of recent studies and hypothesesActa Zoologica 88 317ndash335
Bousfeld EL 1995 A contribution to the natural classification of Lower andMiddle Cambrian arthropods food gathering and feeding mechanismsAmphipacifica 2 3ndash34
Boxshall GA 2004 The evolution of arthropod limbs Biological Reviews79 253ndash300
Bremer K 1994 Branch support and tree stability Cladistics 10 295ndash304Briggs DEG and Collins D 1999 The arthropod Alalcomenaeus cambricus
Simonetta from the Middle Cambrian Burgess Shale of British ColumbiaPalaeontology 42 953ndash977
Briggs DEG and Robison RA 1984 Exceptionally preserved nonminustrilobite arthropods and anomalocaridids from the Middle Cambrian ofUtah University of Kansas Paleontological Contributions 111 1ndash23
Briggs DEG Lieberman BS Hendricks JR Halgedahl SL andJerrard RD 2008 Middle Cambrian arthropods from Utah Journal ofPaleontology 82 238ndash254
Bruton DL 1981 The arthropod Sidneyia inexpectans Middle CambrianBurgess Shale British Columbia Philosophical Transactions of theRoyal Society of London Series B 295 619ndash656
Bruton DL and Whittington HB 1983 Emeraldella and Leanchoiliatwo arthropods from the Burgess Shale Middle Cambrian British Cominuslumbia Philosophical Transactions of the Royal Society of London Seminusries B 300 553ndash582
Budd GE 2002 A palaeontological solution to the arthropod head probminuslem Nature 417 271ndash275
Butterfield NJ 2002 Leanchoilia guts and the interpretation of threeminusdiminusmensional structures in Burgess Shaleminustype fossils Paleobiology 28155ndash171
Chen JminusY 2004 The Dawn of Animal World [in Chinese] 366 pp JiangScience and Technology Press Nanjing
Chen JminusY Waloszek D and Maas A 2004 A new ldquogreatminusappendagerdquo arminusthropod from the Lower Cambrian of China and homology of cheliceratechelicerae and raptorial anterominusventral appendages Lethaia 37 3ndash20
Cotton TJ and Braddy SJ 2004 The phylogeny of arachnomorph arthrominuspods and the origin of the Chelicerata Transactions of the Royal Societyof Edinburgh Earth Sciences 94 169ndash193
Daley AC Budd GE Caron JminusB Edgecombe GD and Collins D2009 The Burgess Shale anomalocaridid Hurdia and its significancefor early euarthropod evolution Science 323 1597ndash1600
Dunlop JA 2006 New ideas about the euchelicerate stemminuslineage In CDeltshev and P Stoev (eds) European Arachnology 2005 Acta Zoominuslogica Bulgarica (Supplement 1) 9ndash27
Dunlop JA and Selden PA 1997 The early history and phylogeny of thechelicerates In RA Fortey and RH Thomas (eds) Arthropod Relaminustionships Systematics Association Special Volume 55 221ndash235
Edgecombe GD and Ramskoumlld L 1999 Relationships of CambrianArachnata and the systematic position of Trilobita Journal of Paleonminustology 73 263ndash287
Farris JS Albert VA Kaumlllersjouml M Lipcomb D and Kluge AG1996 Parsimony jackknifing outperforms neighborminusjoining Cladistics12 99ndash124
GarciacuteaminusBellido DC and Collins D 2007 Reassessment of the genusLeanchoilia (Arthropoda Arachnomorpha) from the Middle CambrianBurgess Shale British Columbia Canada Palaeontology 50 693ndash709
GarciacuteaminusBellido DC Paterson JR Edgecombe GD Jago JB GehlingJG and Lee MSY 2009 The bivalved arthropods Isoxys and Tuzoiawith softminuspart preservation from the lower Cambrian Emu Bay ShaleLagerstaumltte (Kangaroo Island Australia) Palaeontology 52 1221ndash1241
Goloboff PA 1993 Estimating character weights during tree searchCladistics 9 83ndash91
Goloboff PA Farris JS and Nixon KC 2008 TNT a free program forphylogenetic analysis Cladistics 24 774ndash786
Harzsch S and Hafner G 2006 Evolution of eye development in arthrominuspods phylogenetic aspects Arthropod Structure amp Development 35319ndash340
Hendricks JR and Lieberman BS 2008 New phylogenetic insights intothe Cambrian radiation of arachnomorph arthropods Journal of Paleminusontology 82 585ndash594
Hou XminusG and Bergstroumlm J 1997 Arthropods of the Lower CambrianChengjiang fauna southwest China Fossils and Strata 45 1ndash116
Jago JB Zang W Sun X Brock GA Paterson JR and SkovstedCB 2006 A review of the Cambrian biostratigraphy of South Austraminuslia Palaeoworld 15 406ndash423
Liu Y Hou XminusG and Bergstroumlm J 2007 Chengjiang arthropod Leanminuschoilia illecebrosa (Hou 1987) reconsidered GFF 129 263ndash272
Luo H Hu S Chen L Zhang S and Tao Y 1999 Early CambrianChengjiang fauna from Kunming region China [in Chinese] 129 ppYunnan Science and Technology Press Kunming
Luo H Hu S Zhang S and Tao Y 1997 New occurrence of the EarlyCambrian Chengjiang fauna in Haikou Kunming Yunnan Provinceand study on Trilobitoidea Acta Geologica Sinica 71 122ndash132
Nixon KC 2002 WINCLADA Version 19908 Cornell University IthacaNY
Orr PJ Siveter DJ Briggs DEG Siveter DJ and Sutton MD 2000A new arthropod from the Silurian KonservatminusLagerstaumltte of Hereminusfordshire Proceedings of the Royal Society Series B Biological Sciminusences 267 1497ndash1504
Paterson JR and Brock GA 2007 Early Cambrian trilobites fromAngorichina Flinders Ranges South Australia with a new assemblagefrom the Pararaia bunyerooensis Zone Journal of Paleontology 81116ndash142
Paterson JR and Jago JB 2006 New trilobites from the Lower CambrianEmu Bay Shale Lagerstaumltte at Big Gully Kangaroo Island South Ausminustralia Memoirs of the Association of Australasian Palaeontologists 3243ndash57
Paterson JR Edgecombe GD GarciacuteaminusBellido DC Jago JB andGehling JG 2010 Nektaspid arthropods from the lower CambrianEmu Bay Shale Lagerstaumltte South Australia with a reassessment oflamellipedian relationships Palaeontology 53 377ndash402
Paterson JR Jago JB Gehling JG GarciacuteaminusBellido DC Edgecombe
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 399
GD and Lee MSY 2008 Early Cambrian arthropods from the EmuBay Shale Lagerstaumltte South Australia 319ndash325 In I Raacutebano RGozalo and D GarciacuteaminusBellido (eds) Advances in Trilobite ResearchCuadernos del Museo Geominero 9 319ndash325
Shultz JW 2007 A phylogenetic analysis of the arachnid orders based onmorphological characters Zoological Journal of the Linnean Society150 221ndash265
Stein M 2010 A new arthropod from the Early Cambrian of North Greenminusland with a ldquogreat appendagerdquominuslike antennula Zoological Journal ofthe Linnean Society 158 477ndash500
Stuumlrmer W and Bergstroumlm J 1978 The arthropod Cheloniellon from theDevonian Hunsruumlck Shale Palaumlontologische Zeitschrift 52 57ndash81
Sutton MD Briggs DEG Siveter DJ Siveter DJ and Orr PD2002 The arthropod Offacolus kingi (Chelicerata) from the Silurian ofHerefordshire England computer based morphological reconstrucminustions and phylogenetic affinity Proceedings of the Royal Society B Biminusological Sciences 269 1195ndash1203
Suzuki Y and Bergstroumlm J 2008 Respiration in trilobites a reevaluationGFF 130 211ndash229
Vannier J GarciacuteaminusBellido DC Hu XminusX and Chen AminusL 2009 Arthrominuspod visual predators in the early pelagic ecosystem evidence from the
Burgess Shale and Chengjiang biotas Proceedings of the Royal SocietyB Biological Sciences 276 2567ndash2574
Whittington HB 1974 Yohoia Walcott and Plenocaris n gen arthropodsfrom the Burgess Shale Middle Cambrian British Columbia Geologiminuscal Survey of Canada Bulletin 231 1ndash21
Wills MA Briggs DEG Fortey RA Wilkinson M and SneathPHA 1998 An arthropod phylogeny based on fossil and recent taxaIn GD Edgecombe (ed) Arthropod Fossils and Phylogeny 33ndash105Columbia University Press New York
Wilson LA 2006 Food for thought a morphological and taphonomic studyof fossilised digestive systems from Early to Middle Cambrian taxa275 pp Unpublished PhD thesis University of Cambridge Cambridge
Zhang XminusG and Hou X 2007 Gravitational constraints on the burial ofChengjiang fossils Palaios 22 448ndash453
Zhang XminusL Han J Zhang Z Liu H and Shu D 2004 Redescription ofthe Chengjiang arthropod Squamacula clypeata Hou and Bergstroumlm fromthe Lower Cambrian Southminuswest China Palaeontology 47 605ndash617
Zhao Y Zhu M Babcock LE Yuan L Parsley RL Peng J YangX and Wang Y 2005 Kaili Biota a taphonomic window on diversifiminuscation of metazoans from the basal Middle Cambrian Guizhou ChinaActa Geologica Sinica 79 751ndash765
400 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011

doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 391
Great appendage robust proportions of peduncle and disminustal spineminusbearing articles as in other members of Leanchoiliminusidae more commonly preserved with posterior orientation
but anteriorly oriented in SAM P45906 and SAM P45910Each of the three long spinose projections terminating at thesame length the projections evenly tapering distally with
392 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Fig 6 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov SAM P45168 A head and anterior part of trunk B Detail of spinoseprojections of great appendages and excretory organs (sensu Bruton and Whittington 1983) Abbreviations cs cephalic shield eo excretory organ gagreat appendage mg midgut glands tx trunk exopods
margins straight for much of their length or slightly curvedterminally distal spinose projection distinctly stouter at itsbase than the two more proximal projections and more markminusedly tapering along its length Dark irregularminusshaped low reminuslief lump on basal spineminusbearing article of great appendagesof SAM P45168 (eo in Fig 6) and SAM P45908 correminussponding to excretory organ of Bruton and Whittington(1983) Long flagella extending from each of the threespinose projections seen only in SAM P43631 (Figs 23A3) of similar narrow width along its preserved extent anminusnulations indistinguishable
Trunk of 11 segments of even length Maximum height oftrunk relative to length (excluding telson) ca 39 bodydeepest at tergite 4 Trunk tergite boundaries marked as narminusrow ridges in several specimens corresponding to narrowband of overlap between adjacent tergites (ol in Figs 2 3A14A1 B) Some specimens show tergites to be bisected by anarrow but sharp furrow of uncertain identity (eg possiblyimpression of dorsal transverse tendon cf Stein 2010 fig12B C E) (arrows in Figs 3A1 4B)
Exopods with fringe of long setae on cephalic and trunkappendages (Fig 2) setal fringe gradually shortening fromabout the fourth trunk segment Setae project ventrally bymore than 30 height of tergites in deepest part of trunk(Fig 2B) setae flattened narrow and parallelminussided alongmost of their length tapering near their distal ends to termiminusnate as a pointed tip (Figs 2B 3A3 arrows) Endopods exminusposed only in SAM P45167 (Fig 4) two pairs associatedwith cephalon (Fig 4B) those of anterior segments of trunkof similar morphology to those in cephalon all slender lackminusing obvious endites podomere boundaries unrecognisable
Midgut glands occupy a large extent of the dorsoventralheight of the trunk two midgut glands in cephalon of similarsize to those in each of the first seven segments of the trunk(Figs 3A1 5A 6A) all developed as radiating grooves emaminusnating from a globular structure at their ventral edge as in
Leanchoilia superlata (GarciacuteaminusBellido and Collins 2007 pl2 6) Foregut trace in SAM P43631 (Fig 2) indicates that gutcurves ventrally in anterior part of cephalon mouth regionnot preserved Terminal part of hindgut (in trunk segment 11)with closely spaced longitudinal lineations (Fig 2) that apminusparently correspond to folds in the gut wall hindgut termiminusnating at base of telson
Telson most completely preserved in holotype (SAMP43631 Figs 2B 3A2 B) In this specimen the telson likerest of body compressed in lateral aspect Telson shows meminusdian and marginal reinforcing ridges connecting a flat trianminusgular surface this surface appears to be partially broken inSAM P43631 showing underlying surface of other (left) halfof telson Lateral preservation of specimen implies that topcorresponds to an axial structure and bottom to a marginalone 30ndash35 angle in posterior edge of vane connecting thesetwo structures requires that in dorsal aspect telson would apminuspear forked and widening distally Five stout pointed dorminussally curved marginal spines attached to outer posterior edgeon each side of telson
Phylogenetic analysis ofLeanchoiliidaeDetermining the systematic position of Oestokerkus demandsan appraisal of the relationships of Leanchoiliidae as a wholeCladistic analysis including the bestminusknown members ofMegacheira including leanchoiliids has been conducted byCotton and Braddy (2004) and Vannier et al (2009) The taxominusnomic sampling used by Cotton and Braddy (2004) includedseveral megacheiran genera representative chelicerates anumber of taxa representing groups that have been proposedas possible chelicerate relatives (eg Sidneyia AglaspisCheloniellon) in other analyses (Dunlop and Selden 1997Hou and Bergstroumlm 1997) and members of the major lineagesof the Artiopoda sensu Hou and Bergstroumlm (1997) ie trilominusbiteminusallied lamellipedians Because artiopodan interminusrelationminusships have remained quite stable through a series of phylogenminusetic analyses (Edgecombe and Ramskoumlld 1999 Cotton andBraddy 2004 Hendricks and Lieberman 2008 Paterson et al2010) and artiopodans are now often regarded as less closelyrelated to Megacheira than is Chelicerata (Chen et al 2004Cotton and Braddy 2004 Dunlop 2006) we have reduced thesampling of Artiopoda in the present analysis To augment thediversity of ldquoshort great appendagerdquo arthropods Haikoucarisis added based on the description of H ercaiensis by Chen etal (2004) Offacolus (Orr et al 2000 Sutton et al 2002) hasbeen allied to Chelicerata and is included as a terminal Beminuscause the present analysis is principally concerned with relaminustionships of and within Leanchoiliidae sensu Liu et al (2007)we follow Vannier et al (2009) in sampling Alalcomenaeusand Leanchoilia at the species level Within Chelicerata Cotminuston and Braddy (2004) and Hendricks and Lieberman (2008)sampled ancestral states in Xiphosura by coding the Devonian
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 393
Fig 7 Reconstruction of Oestokerkus megacholix gen et sp nov in lateralview Total length and annulations on the flagella and positions of podominusmere boundaries of the frontal appendage are based on Leanchoilia superminuslata (Bruton and Whittington 1983)
synziphosurine Weinbergina Our coding for Xiphosura alsoemploys information from extant xiphosurans because theysupply more reliable information on the homology of the partsof the trunk appendages (Suzuki and Bergstroumlm 2008) and eyeanatomy (Harzsch and Hafner 2006) among other charactersystems than do fossils alone
Our character set extracts characters from the matrix ofCotton and Braddy (2004) that vary in the revised taxonomicsample reformulates some characters to accommodate reminusvised homology concepts and adds new characters especiallyfor leanchoiliids In total 32 characters are included (Table 1)In this analysis the primary homology of the ldquoshort great apminuspendagerdquo and chelicera is coded (as described by character 4)and the taxon sampling broadly constrains megacheirans aschelicerate relatives (as defended by Chen et al 2004 Cottonand Braddy 2004) rather than testing the alternative that theycould be stem group Euarthropoda (Budd 2002 Bergstroumlmand Hou 2005) A test of the two alternatives involves samminuspling of the entire diversity of euarthropods and a range ofnonminuseuarthropod outgroups an exercise beyond the scope ofthis study
Characters1 Mineralised cuticle (0) absent (1) present For definiminus
tion and discussion of this character see Cotton andBraddy (2004 character 30) and Paterson et al (2010character 1)
2 Curvature of anterior margin of cephalic shield (0) rounminusded or straight (1) pointed medially (2) with multiplespinose projections This character codes for variability inthe shape of the cephalic shield in Leanchoiliidae (state 0and 1 sensu GarciacuteaminusBellido and Collins [2007 textminusfigs10 and 9 respectively]) State 2 is added to accommodateOffacolus the median spine of a series that fringes the ceminusphalic margin is not convincingly homologised with themedian projection in leanchoiliids
3 Number of cephalic somitesappendages (0) antenna orfrontal appendage only (1) antenna or frontal appendageand two limb pairs (2) antenna or frontal appendage andthree limb pairs (3) antenna or frontal appendage and fourlimb pairs (4) antenna or frontal appendage and five limbpairs Coding for lamellipedians and Aglaspis is as discussedby Paterson et al (2010 character 2) Coding for Alalcomeminusnaeus cambricus (state 1) follows Briggs and Collins (1999)Leanchoilia illecebrosa (state 2) follows Liu et al (2007)and Leanchoilia persephone and Leanchoilia superlata (state1) follows GarciacuteaminusBellido and Collins (2007)
4 Form of first cephalic appendage (0) antenna (1) anteminusriorly directed raptorial appendage with reduced numberof podomeres each of the terminal podomeres bearing aspinose process on its distal margins a geniculation beminustween spinose podomeres and peduncle This characteris formulated to identify a homology between the frontalappendages of megacheirans and the chelicera of Cheliminus
cerata and state 1 corresponds to the putative apomorminusphic detail defended by Chen et al (2004) and Cottonand Braddy (2004) Vannier et al (2009 character 8)specify another shared property of ldquoshort great appendminusagesrdquo of the megacheiran (ss) type in their characterstate ldquobent at an anglerdquo referring to the geniculation beminustween articles of the peduncle or between the peduncleand distal spiniferous articles This property is sharedwith Chelicerata
5 Number of spineminusbearing articles on frontal appendage (0)three + movable finger (1) two + movable finger (2) one+ movable finger The number of spineminusbearing articles onthe frontal appendage (exclusive of the terminal movablefinger) varies from three in Yohoia (Whittington 1974 pl6 1ndash3 pl 7 3ndash6 pl 9 4ndash6) and Fortiforceps (Hou andBergstroumlm 1997 figs 31C D 32A 33B C E) to two inHaikoucaris (Chen et al 2004 fig 2A B) and Leanminuschoiliidae or only one the last being a chelicera
6 Long spinose projections on distal part of terminal threepodomeres of frontal appendage bearing a flagellum (0)absent (1) present The morphology of the great appendminusage in Leanchoiliidae is coded here We have not codedthe length of the spinose projections and the presence ofa flagellum as two independent characters Cotton andBraddyrsquos (2004) character 6 state 2 (ldquospinesrdquo longer thanthe entire podomere series) and character 8 state 1 (presminusence of an annulated flagellum) are thus grouped as a sinminusgle trait here
394 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Table 1 Characters coded for phylogenetic analysis Polymorphismindicated by ldquo+rdquo partial uncertainty by states separated by ldquordquo Abbreviminusations A Alalcomenaeus L Leanchoilia O Oestokerkus
Squamacula 0000minus minus minus2minus0 0110 00000 10minus minus0 1000 0
Cindarella 0030minus minus minus000 0010 00000 10minus minus0 11121 00
Kuamaia 000minus minus minus001 0011 10000 10minus minus0 11121 00
Kwanyinaspis 00 001 00112 1000 1000 1121 00
Olenoides 1020minus minus minus100 0011 00000 10minus minus0 01121 00
Retifacies 0020minus minus minus000 0010 00000 11000 1020 00
Aglaspis 10(23)0minus minus minus100 00101 00010 01001 0 00
Sidneyia 0000minus minus minus000 01102 00110 01101 00020 00
Emeraldella 0040minus minus minus2minus 0 00103 00110 01001 001(1+2)1 00
A cambricus 00111 10010 00001 00000 01110 01110 10
L illecebrosa 01211 11010 00001 01001 01210 11110 10
L persephone 00111 10010 00001 00000 01210 11110 10
L superlata 01111 11010 00001 01001 01210 11110 10
L hanceyi 00 00 0001 0000 01210
O megacholix 0011 100 00001 00000 01110 1110 0
Jianfengia 0021 0minus000 00005 00000 01000 010(12)0 0
Fortiforceps 00210 0minus00 00004 00000 01100 01230 20
Yohoia 00210 0minus000 00003 00011 01110 00010 (01)1
Haikoucaris 00211 0minus00 00003 00000 01(12)0 1010 0
Cheloniellon 0040minus minus minus100 00100 10010 01300 0030 00
Offacolus 02412 0minus200 0000 00010 01000 13minus minus 01
Xiphosura 00412 0minus100 10000 0010 01000 000minus minus 01
Eurypterida 00412 0minus100 10002 00010 01000 00minus minus minus 01
7 Length of flagellate great appendage (0) flagella termiminusnating at midlength of trunk (1) flagella extending to endof body Coding is applicable to Leanchoiliidae onlydistinguishing some Leanchoilia species that resembleAlalcomenaeus in having relative short great appendages(state 0) versus the longer appendages in L superlata andL illecebrosa
8 Position of lateral facetted eyes (0) ventral (1) dorsalsessile (2) absent Variation in Lamellipedia and severalother taxa coded herein follows Paterson et al (2010character 3)
9 Four or five ventral eyes arranged in a subtransverseband across head shield (0) absent (1) present A bandof ventral eyes is shared by the best known species ofLeanchoiliidae The distinction between three medianeyes in Alalcomenaeus cambricus (Briggs and Collins1999) and two in Leanchoilia illecebrosa (Liu et al2007) and both Leanchoilia superlata and L persephone(GarciacuteaminusBellido and Collins 2007) is uninformative forthe taxon sample here
10 Bulge in exoskeleton accommodating dropminusshaped venminustral eyes (0) absent (1) present This character followsEdgecombe and Ramskoumlld (1999 character 6) and Patminuserson et al (2010 character 5)
11 Dorsal median eyes (0) absent (1) present Coding for anabsence of dorsal median eyes in fossil taxa considers theabsence of either paired ocelli or a median eye tubercleand the character is scored as a chelicerate autapomorphy(see Cotton and Braddy 2004 character 24)
12 Width of doublure on cephalic shield (0) narrow or modminuserately wide (1) entire ventral side of head covered byexpansion of doublure The interpretation of state 1 inSidneyia and Squamacula is discussed by Cotton andBraddy (2004) and Paterson et al (2010 characters 12and 29)
13 Mouth covered by hypostome (0) absent (1) present Noldquoshort great appendagerdquo arthropods have been shown topossess a hypostome and a reconstruction of a smalllabrum in Alalcomenaeus (Briggs and Collins 1999) wasadmittedly conjectural The absence of a conspicuoushypostome contrasts with its ubiquitous presence in laminusmellipedians and several putative chelicerateminusallied taxathat have antennae (such as aglaspidids and Emeralminusdella) Chelicerates have a small epistomeminuslabrum (Bitschand Bitsch 2007) but like Cotton and Braddy (2004character 27 state 4) we code a hypostomal sclerite aslacking Kwanyinaspis has a large hypostome similar tothat found in aglaspidids (photograph of the holotype ofKwanyinaspis maotianshanensis shown to us by JavierOrtegaminusHernaacutendez [personal communication 2010])
14 Tergite articulations (0) extensive overlap of tergites (1)edge to edge articulations This character follows Edgeminuscombe and Ramskoumlld (1999 character 18) and Patersonet al (2010 character 20)
15 Number of trunk segments (0) nine (1) 11 (2) 12 (3)13 (4) 20 (5) 21 or 22 Coding is confined to taxa that do
not possess a pygidium because the precise number ofsomites associated with pygidia can sometimes not bedetermined with confidence Codings for Xiphosura andEurypterida use opisthosomal somite numbers followingShultz (2007 character 95)
16 Exoskeletal boundaries of anterior trunk segments (0)boundaries transverse or reflexed posterolaterally (1) reminusflexed anterolaterally This character follows Edgecombeand Ramskoumlld (1999 character 19) and Paterson et al(2010 character 21)
17 Trunk tergites with serrate lateral margins (0) absent (1)present Liu et al (2007 fig 2C) documented a row ofmarginal spines on the tergites of Leanchoilia illeceminusbrosa a featured shared by Leanchoilia superlata (egGarciacuteaminusBellido and Collins 2007 pl 1 7) but not otherterminals coded here
18 Differentiation in width of trunk tergites (0) trunk withgradational narrowing posteriorly (1) abrupt narrowingat boundary between prothorax and opisthothorax Anabrupt contraction in width between the boundary ofthe prominus and opisthothorax is observed in Emeraldellaand Sidneyia Yohoia has been reconstructed as having apronounced break in the shape of the trunk tergites beminushind segment 10 (Whittington 1974 textminusfig 2) thepoint at which the paratergal folds lose their spinose tipsand appendages are lacking Specimens preserved indorsal aspect (Whittington 1974 pl 10) suggest that thewidth of the opisthothorax is rather less abruptly narminusrowed and we code Yohoia as state 0
19 Appendages of posterior trunk segments (0) all trunktergites bearing appendages (1) one or more posteriortrunk segments lacking appendages Coding correspondsto Cotton and Braddyrsquos (2004) character 43 ldquopostabdominusmen of segments lacking appendagesrdquo
20 Paired tergal carinae (0) absent (1) present Liu et al(2007 265 fig 2F) recorded the presence of tergalcarinae in Leanchoilia illecebrosa shared with L superminuslata (eg Bruton and Whittington 1983 pl 17 106GarciacuteaminusBellido and Collins 2007 pl 1 5)
21 Pygidium (0) absent (1) present Coding of the pygiminusdium in many terminals herein was discussed by Paterminusson et al (2010 character 22)
22 Telson (0) absent (1) present Following Hendricks andLieberman (2008) a telson is a postminussegmental terminalstructure that lacks appendages Dunlop and Selden (1997)considered a telson to be absent in Cheloniellon but weidentify the conical projection posterior to the ultimate legpair and cerci (Stuumlrmer and Bergstroumlm 1978 figs 2 3 1011) as being most consistent with an identity as a telson
23 Telson shape (0) styliform (1) flapminuslike (2) lanceolate(3) short conical The outline of the telson in Haikouminuscaris ercaiensis is incompletely known but the mostcomplete specimen (Chen 2004 fig 472) constrains it aseither state 1 or state 2 A partial uncertainty coding isthus employed A unique state for Cheloniellon (state 3)is based on the interpretation in character 22
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 395
24 Telson fringed with spines (0) spines absent (1) spinespresent Presence of telson spines was introduced as a charminusacter by Hendricks and Lieberman (2008 character 34)
25 Paired flaps lateral to telson (0) absent (1) present Thischaracter describes the postminusventral plates of several taxa(see Edgecombe and Ramskoumlld 1999 character 25) Cotminuston and Braddy (2004 character 54) raised the possibilitythat the cerci of Cheloniellon could be homologous withpostminusventral plates (see also Hou and Bergstroumlm 1997 93)because they are similarly paired structures derived fromthe segment immediately anterior to the telson We havenot coded for this homology because the cerci of Chelominusniellon originate dorsally rather than ventrally
26 Midgut glands with submillimetric lamellae (0) absent(1) present Midgut glands in some stem group arthrominuspods and early crown group arthropods are metamericreniform organs with a characteristic lamellar structure(Butterfield 2002) Their morphology is identical inLeanchoilia and Oestokerkus notably including groovesthat radiate from a ventral median node (Figs 3A1 6A)The gut of Alalcomenaeus cambricus is more tubular(Briggs and Collins 1999 textminusfigs 10 11) and thoughan anterior six segments may display phosphatised latminuseral projections that likely represent gut diverticula (Wilminusson 2006) they have not been shown to have the detailedmorphology of Leanchoiliaminustype midgut glands Butterminusfield (2002 164) cited a figure from Bruton (1981 fig96) in support of ldquolamellarrdquo midgut glands in the cephaminuslon of Sidneyia Like Bruton (1981) in his description ofthe specimen we consider it likely that the structures inquestion are indeed gut diverticula but there is no basisfor identifying them more precisely as the reniformlamellar structures coded under this character
27 Exopod on first postminusantennal first postminusfrontal appendminusage segment (0) absent (1) present This character reminuslates to Cotton and Braddyrsquos (2004) character 3
28 Division of exopod (0) exopod an undivided flap (1)exopod differentiated into proximal and distal lobes (2)small flap with radiating lamellae (3) pediform exopoddivided into ca 6 articles Identification of the exopod inXiphosura follows Suzuki and Bergstroumlm (2008) Thischaracter groups conditions of the exopod shaft coded asindependent characters by Cotton and Braddy (2004)eg state 0 corresponds to their character 16 and state 1to their character 13 Our coding permits transformationbetween any two states without an extra implicit step
29 Exopod setae (0) short spiniform setae (1) long flatminustened tapering distally with slight separation (2)lamellar setae (flattened imbricating) (3) short finesetae A distinction is made between lamellipedian setaewhich are imbricated (state 2) and the pointed more disminustinctly separated and variably spiniform exopod setae ofleanchoiliids (state 1) This distinction is consistent withthe identification of megacheirans as nonminuslamellipedians(eg Hou and Bergstroumlm 1997 Bergstroumlm and Hou2005) Although Cotton and Braddy (2004 character 18)
regarded the form of the setae in Yohoia to be uncertainwe regard specimens figured by Whittington (1974 pl3 3) as conforming to the leanchoiliid state Squamaculadiffers from other taxa in its shorter spiniform setae(state 0) (Zhang et al 2004 textminusfig 1A B) Offacolushas longer spiniform projections on the distal articles ofthe exopod that have been variably identified as eitherspines or setae (Sutton et al 2002 fig 4b) but theirhomology with the setae that splay regularly along themargin of a flapminuslike exopod (eg states 0ndash3) is uncerminustain This and the following character for exopod setaeare left as ldquordquo for Offacolus A homology between thegill lamellae of Xiphosura and exopod setae of lamellipeminusdians has a long history (see Cotton and Braddy 2004character 18) We follow Suzuki and Bergstroumlm (2008)in identifying the xiphosuran gills as outgrowths of thebasis rather than of the exopod this perspective negates ahomology with the exopod setae of Lamellipedia Thepolymorphic coding for Emeraldella reflects the twoseparate states (1+2) displayed by the exopods in thetrunk (see Bruton and Whittington 1983 pl 8 45 47Edgecombe and Ramskoumlld 1999 fig 11)
30 Differentiation of setae along exopod (0) uniform orgradationally variable setae along length of exopod (1)lamellar setae confined to proximal lobe with distal lobebearing slender setae
31 Endopod segmentation (distal to coxa) (0) six or sevenpodomeres and terminal claw or telotarsal spurs (1)eight or more podomeres and terminal claw (2) ca 15podomeres State 1 is formulated to describe the segmenminustation of the leanchoiliid leg (GarciacuteaminusBellido and Collins2007 textminusfig 6 Liu et al 2007 fig 5) in contrast to thereduced number of podomeres in other euarthropods(Boxshall 2004) State 2 characterises Fortiforceps Apartial uncertainty coding is employed for Yohoia inwhich the precise number of podomeres is undeterminedand could be either state 0 or 1 (Whittington 1974 pl 74) but the spacing of the preserved podomere boundariesnegates the possibility of state 2
32 Endopods on anterior postminuscephalic segments (0) welldeveloped (1) rudimentary or lacking Following Cottonand Braddy (2004 character 11) the absence of trunkendopods in Yohoia (Whittington 1974) is coded as a puminustative homologue with the suppression of postminuscephalicendopods in Chelicerata
ResultsAs a convention for depicting the trees networks are rootedon Squamacula which separates artiopodan Lamellipedia(sensu Hou and Bergstroumlm 1997) from a group that includesMegacheira Chelicerata and Xenopoda all members ofwhich possess a telson
Under equal weights 14 shortest cladograms have a lengthof 77 steps with a Consistency Index of 064 and Retention Inminus
396 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
dex of 077 The strict consensus of these trees with supportvalues is shown in Fig 8 Leanchoiliidae is unambiguouslysupported in all 14 cladograms by four apomorphic characterstwo limbminusbearing cephalic segments posterior to the frontalappendage (character 3 state 1) long spinose projections onthe distal part of the terminal three podomeres of the frontalappendage that bear flagella (character 6 state 1) 11 trunksegments (character 15 state 1) and division of the exopodsinto two lobes (character 28 state 1) The arrangement of fouror five eyes in a subtransverse band across the cephalic shieldis unique to Leanchoiliidae (character 9 state 1) but is not anunambiguous autapomorphy of the group because the state isuncertain in close relatives notably Haikoucaris It is apominusmorphic for Leanchoiliidae only under particular optimisaminustions (Delayed Transformation)
The internal relationships of Leanchoiliidae are stableacross all 14 shortest cladograms Leanchoilia is united by thelanceolate telson (character 23 state 2) a unique unambiguminusous autapomorphy Within Leanchoilia a sister group relaminustionship between L superlata and L illecebrosa is well supminusported receiving a Bremer support of 3 and jackknife freminusquency of 86 These two species share four synapomorminusphies a medially pointed anterior cephalic margin (character2 state 1) flagella of the great appendage extending the lengthof the body (character 7 state 1) serrate lateral margins on thetrunk tergites (character 17 state 1) and paired tergal carinae(character 20 state 1) As such a species from Chengjiang (Lillecebrosa) and the Burgess Shale (L superlata) are moreclosely related to each other than are the two Burgess Shalecongeners a relationship that weakens the possibility that theoften cominusoccurring L superlata and L persephone could besexual dimorphs (GarciacuteaminusBellido and Collins 2007) ratherthan separate species The union of Leanchoilia and Oestominuskerkus is unambiguously supported by the shared midgutgland structure of both genera (character 26 state 1)
All shortest cladograms resolve Haikoucaris Yohoiaand Fortiforceps in that order as progressively less closelyrelated to Leanchoiliidae The union of these three taxa withLeanchoiliidae yields a clade that corresponds to Megacheirasensu Hou and Bergstroumlm (1997) apart from the ambiguousplacement of Jianfengia In half of the 14 shortest cladominusgrams Jianfengia is sister to the other megacheirans yieldminusing a monophyletic Megacheira ss In the remaining cladominusgrams Jianfengia is sister to Xenopoda sensu Hou andBergstroumlm (1997) Aglaspis and Chelicerata Haikoucarisand Leanchoiliidae are united by a frontal appendage comminusposed of two spineminusbearing articles in addition to the movminusable finger (character 5 state 1) Yohoia groups with thesetaxa based on spines fringing the telson (character 24 state1) and Fortiforceps is united with them at a broader levelbased on the flapminuslike telson shape (character 23 state 1)When Jianfengia is placed as sister to the other megacheiminusrans the entire clade is supported by the raptorial frontal apminuspendage (character 4 state 1) and the lack of a large hypominusstomal shield (character 13 state 0) In this set of cladominusgrams these features are nonminushomologous between Megaminus
cheira and Chelicerata whereas in some cladograms with thealternative resolution of Jianfengia as closer to cheliceratescharacter 4 is optimised as homologous in Megacheira andChelicerata
Reanalysis of the data with implied weights finds two optiminusmal trees (Fig 9) Relationships within Leanchoiliidae underequal weights are completely stable to reweighting as is theresolution of Haikoucaris as sister to Leanchoiliidae As suchthe character optimisations described above for the leanchoiminusliids are depicted the same in Fig 9 The main difference beminustween equal and implied weights involves the placement ofChelicerata Under equal weights (Fig 8) chelicerates are alminuslied to Xenopoda (here Emeraldella + Sidneyia + Chelominusniellon sensu clade L in Hou and Bergstroumlm 1997 fig 88)and Aglaspidida A clade composed of Aglaspidida Xenominuspoda and Chelicerata was similarly identified by Hou andBergstroumlm (1997 fig 88 their clade K) In our analysis thisgroup consistently finds support from one or more posteriortrunk segments lacking appendages (character 19 state 1) anddepending on alternative internal resolutions it is variably supminusported by dorsallyminuspositioned sessile eyes (character 8 state 1)and the absence of exopods on the first postminusantennalpostminuscheliceral limb (character 27 state 1) Under implied weightshowever chelicerates are more closely allied to Megacheirathan to aglaspidids or xenopodans (Fig 9)
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 397
2
582
76
Chelicera-morpha
Megacheira
62
2
53
50Leanchoiliidae
3
86
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
EurypteridaChelicerata
Fortiforceps
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia hanceyi
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
Fig 8 Strict consensus of 14 shortest cladograms (77 steps) based on charminusacter data in Table 1 Numbers above nodes are Bremer support those beminuslow nodes are jackknife frequencies gt50 Cheliceramorpha correspondsto a clade with the same composition in Cotton and Braddy (2004)
The major groupings in Fig 9 correspond closely to thoseidentified by Cotton and Braddy (2004) Notably the cladeuniting Aglaspis Xenopoda Offacolus Chelicerata Jianminusfengia and other Megacheira corresponds to their ldquoClade 3rdquo(Cotton and Braddy 2004 fig 8) for which they proposedthe name Cheliceramorpha The clade uniting CheloniellonAglaspis Sidneyia and Emeraldella in Fig 9 corresponds totheir ldquoClade 5rdquo and the clade that groups Megacheira andChelicerata is equivalent to their ldquoClade 4rdquo Their analysisdid not identify megacheirans as a monophyletic groupChelicerata was nested within the group and among taxaconsidered here Yohoia was resolved closer to cheliceratesthan to Leanchoiliidae In contrast Yohoia was regarded byBergstroumlm and Hou (2005) as more closely allied to Leanminuschoiliidae than to chelicerates a megacheiran grouping morecompatible with our cladograms The analysis of ldquogreat apminuspendage arthropodsrdquo by Vannier et al (2009) identified a
monophyletic Leanchoiliidae but a polyphyletic Megacheira(to the exclusion of Haikoucaris and Jianfengia) Howevertheir resolution of Fortiforceps and even more so Yohoia asthe closest relatives of Leanchoiliidae is as in our trees (apartfrom the different position of Haikoucaris)
In summary a major controversy in the systematics of theldquoshort great appendagerdquo arthropods remains whether thechelicerates are nested within the group or are instead moreclosely allied to taxa conventionally identified as lamelliminuspedians specifically Xenopoda and Aglaspidida We findthat both of these results are compatible with the availabledata depending on character weighting protocols Noneminustheless the broader megacheiranminuschelicerate issue does notcloud the status of leanchoiliids as a clade or the identificaminustion of their closest relatives (ie Haikoucaris and Yohoia)and the internal relationships of Leanchoiliidae are stableand supported by compelling characters
398 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
Eurypterida
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
2 7 17 20
3
2
1 1 1
26
1
281563
1 1 1 1
5
1
24
1
23 19 20 27 32
28Clade 4
1 0
4 13 11 27 0
154 1 1
3 19 32Clade 3
2 3
2 28
0
3 1 121
1
22
Clade 5
1 0
19 27
1
25
15 28 301 1
2312
1 150 1 3 3
15 16 2923
18
1 1
10 16
1 1
1 8
1
14
3
3
28 30
1 0
5
15
1 1
1
1
22
Fortiforceps
23
1 1 0 1
1 1
Leanchoilia hanceyi
2
2
1
1
Fig 9 Characters optimised on one of two shortest cladograms with imminusplied weights using a concavity constant k = 3 Black squares are nonminushomoplastic changes white squares homoplastic changes all changes areunambiguous Branches without strict support collapsed after optimisingcharacters Numbers for characters (above the branches) and states (belowthe branches) correspond to Table 1 ldquoClades 3ndash5rdquo correspond to groups sonumbered by Cotton and Braddy (2004 fig 8)
AcknowledgementsAccess to the field area has been most kindly allowed by the Buck famminusily We thank our collaborators in Emu Bay Shale research Jim Jago(University of South Australia Mawson Lakes) Jim Gehling andMike Lee (South Australian Museum Adelaide) for tremendous supminusport and advice as well as Mike Gemmell Dennis Rice Allison DaleyAaron Camens John Laurie Glenn Brock Pierre Kruse and Trevorand Jenny Worthy (all South Australian Museum) for their assistancein the field and Natalie Schroeder MaryminusAnne Binnie and RondaAtkinson (all South Australian Museum) for organisational support inthe lab and in the field Javier OrtegaminusHernaacutendez (University of Camminusbridge Cambridge UK) advised us on Kwanyinaspis and the journalrsquosreferees provided useful feedback TNT is available courtesy of theWilli Hennig Society Funding has been provided by an AustralianResearch Council Linkage grant (LP0774959) to the University ofAdelaide with generous financial assistance from Beach Energy Ltdand the South Australian Museum SeaLink provided logistical supminusport DGBrsquos travel to Australia has been supported by grants from theSpanish Research Council (CSIC PA 1001105) the Complutense Uniminusversity of Madrid and the Spanish Ministry of Education and Science(CGL2006minus12245BTE CGL2009minus07073 and RYC2007minus00090)
ReferencesBergstroumlm J and Hou XminusG 2005 Early Palaeozoic lamellipedian arthrominus
pods In S Koenemann and RA Jenner (eds) Crustacea and Arthrominuspod Relationships Crustacean Issues 16 73ndash93
Bitsch J and Bitsch C 2007 The segmental organization of the head reminusgion in Chelicerata a critical review of recent studies and hypothesesActa Zoologica 88 317ndash335
Bousfeld EL 1995 A contribution to the natural classification of Lower andMiddle Cambrian arthropods food gathering and feeding mechanismsAmphipacifica 2 3ndash34
Boxshall GA 2004 The evolution of arthropod limbs Biological Reviews79 253ndash300
Bremer K 1994 Branch support and tree stability Cladistics 10 295ndash304Briggs DEG and Collins D 1999 The arthropod Alalcomenaeus cambricus
Simonetta from the Middle Cambrian Burgess Shale of British ColumbiaPalaeontology 42 953ndash977
Briggs DEG and Robison RA 1984 Exceptionally preserved nonminustrilobite arthropods and anomalocaridids from the Middle Cambrian ofUtah University of Kansas Paleontological Contributions 111 1ndash23
Briggs DEG Lieberman BS Hendricks JR Halgedahl SL andJerrard RD 2008 Middle Cambrian arthropods from Utah Journal ofPaleontology 82 238ndash254
Bruton DL 1981 The arthropod Sidneyia inexpectans Middle CambrianBurgess Shale British Columbia Philosophical Transactions of theRoyal Society of London Series B 295 619ndash656
Bruton DL and Whittington HB 1983 Emeraldella and Leanchoiliatwo arthropods from the Burgess Shale Middle Cambrian British Cominuslumbia Philosophical Transactions of the Royal Society of London Seminusries B 300 553ndash582
Budd GE 2002 A palaeontological solution to the arthropod head probminuslem Nature 417 271ndash275
Butterfield NJ 2002 Leanchoilia guts and the interpretation of threeminusdiminusmensional structures in Burgess Shaleminustype fossils Paleobiology 28155ndash171
Chen JminusY 2004 The Dawn of Animal World [in Chinese] 366 pp JiangScience and Technology Press Nanjing
Chen JminusY Waloszek D and Maas A 2004 A new ldquogreatminusappendagerdquo arminusthropod from the Lower Cambrian of China and homology of cheliceratechelicerae and raptorial anterominusventral appendages Lethaia 37 3ndash20
Cotton TJ and Braddy SJ 2004 The phylogeny of arachnomorph arthrominuspods and the origin of the Chelicerata Transactions of the Royal Societyof Edinburgh Earth Sciences 94 169ndash193
Daley AC Budd GE Caron JminusB Edgecombe GD and Collins D2009 The Burgess Shale anomalocaridid Hurdia and its significancefor early euarthropod evolution Science 323 1597ndash1600
Dunlop JA 2006 New ideas about the euchelicerate stemminuslineage In CDeltshev and P Stoev (eds) European Arachnology 2005 Acta Zoominuslogica Bulgarica (Supplement 1) 9ndash27
Dunlop JA and Selden PA 1997 The early history and phylogeny of thechelicerates In RA Fortey and RH Thomas (eds) Arthropod Relaminustionships Systematics Association Special Volume 55 221ndash235
Edgecombe GD and Ramskoumlld L 1999 Relationships of CambrianArachnata and the systematic position of Trilobita Journal of Paleonminustology 73 263ndash287
Farris JS Albert VA Kaumlllersjouml M Lipcomb D and Kluge AG1996 Parsimony jackknifing outperforms neighborminusjoining Cladistics12 99ndash124
GarciacuteaminusBellido DC and Collins D 2007 Reassessment of the genusLeanchoilia (Arthropoda Arachnomorpha) from the Middle CambrianBurgess Shale British Columbia Canada Palaeontology 50 693ndash709
GarciacuteaminusBellido DC Paterson JR Edgecombe GD Jago JB GehlingJG and Lee MSY 2009 The bivalved arthropods Isoxys and Tuzoiawith softminuspart preservation from the lower Cambrian Emu Bay ShaleLagerstaumltte (Kangaroo Island Australia) Palaeontology 52 1221ndash1241
Goloboff PA 1993 Estimating character weights during tree searchCladistics 9 83ndash91
Goloboff PA Farris JS and Nixon KC 2008 TNT a free program forphylogenetic analysis Cladistics 24 774ndash786
Harzsch S and Hafner G 2006 Evolution of eye development in arthrominuspods phylogenetic aspects Arthropod Structure amp Development 35319ndash340
Hendricks JR and Lieberman BS 2008 New phylogenetic insights intothe Cambrian radiation of arachnomorph arthropods Journal of Paleminusontology 82 585ndash594
Hou XminusG and Bergstroumlm J 1997 Arthropods of the Lower CambrianChengjiang fauna southwest China Fossils and Strata 45 1ndash116
Jago JB Zang W Sun X Brock GA Paterson JR and SkovstedCB 2006 A review of the Cambrian biostratigraphy of South Austraminuslia Palaeoworld 15 406ndash423
Liu Y Hou XminusG and Bergstroumlm J 2007 Chengjiang arthropod Leanminuschoilia illecebrosa (Hou 1987) reconsidered GFF 129 263ndash272
Luo H Hu S Chen L Zhang S and Tao Y 1999 Early CambrianChengjiang fauna from Kunming region China [in Chinese] 129 ppYunnan Science and Technology Press Kunming
Luo H Hu S Zhang S and Tao Y 1997 New occurrence of the EarlyCambrian Chengjiang fauna in Haikou Kunming Yunnan Provinceand study on Trilobitoidea Acta Geologica Sinica 71 122ndash132
Nixon KC 2002 WINCLADA Version 19908 Cornell University IthacaNY
Orr PJ Siveter DJ Briggs DEG Siveter DJ and Sutton MD 2000A new arthropod from the Silurian KonservatminusLagerstaumltte of Hereminusfordshire Proceedings of the Royal Society Series B Biological Sciminusences 267 1497ndash1504
Paterson JR and Brock GA 2007 Early Cambrian trilobites fromAngorichina Flinders Ranges South Australia with a new assemblagefrom the Pararaia bunyerooensis Zone Journal of Paleontology 81116ndash142
Paterson JR and Jago JB 2006 New trilobites from the Lower CambrianEmu Bay Shale Lagerstaumltte at Big Gully Kangaroo Island South Ausminustralia Memoirs of the Association of Australasian Palaeontologists 3243ndash57
Paterson JR Edgecombe GD GarciacuteaminusBellido DC Jago JB andGehling JG 2010 Nektaspid arthropods from the lower CambrianEmu Bay Shale Lagerstaumltte South Australia with a reassessment oflamellipedian relationships Palaeontology 53 377ndash402
Paterson JR Jago JB Gehling JG GarciacuteaminusBellido DC Edgecombe
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 399
GD and Lee MSY 2008 Early Cambrian arthropods from the EmuBay Shale Lagerstaumltte South Australia 319ndash325 In I Raacutebano RGozalo and D GarciacuteaminusBellido (eds) Advances in Trilobite ResearchCuadernos del Museo Geominero 9 319ndash325
Shultz JW 2007 A phylogenetic analysis of the arachnid orders based onmorphological characters Zoological Journal of the Linnean Society150 221ndash265
Stein M 2010 A new arthropod from the Early Cambrian of North Greenminusland with a ldquogreat appendagerdquominuslike antennula Zoological Journal ofthe Linnean Society 158 477ndash500
Stuumlrmer W and Bergstroumlm J 1978 The arthropod Cheloniellon from theDevonian Hunsruumlck Shale Palaumlontologische Zeitschrift 52 57ndash81
Sutton MD Briggs DEG Siveter DJ Siveter DJ and Orr PD2002 The arthropod Offacolus kingi (Chelicerata) from the Silurian ofHerefordshire England computer based morphological reconstrucminustions and phylogenetic affinity Proceedings of the Royal Society B Biminusological Sciences 269 1195ndash1203
Suzuki Y and Bergstroumlm J 2008 Respiration in trilobites a reevaluationGFF 130 211ndash229
Vannier J GarciacuteaminusBellido DC Hu XminusX and Chen AminusL 2009 Arthrominuspod visual predators in the early pelagic ecosystem evidence from the
Burgess Shale and Chengjiang biotas Proceedings of the Royal SocietyB Biological Sciences 276 2567ndash2574
Whittington HB 1974 Yohoia Walcott and Plenocaris n gen arthropodsfrom the Burgess Shale Middle Cambrian British Columbia Geologiminuscal Survey of Canada Bulletin 231 1ndash21
Wills MA Briggs DEG Fortey RA Wilkinson M and SneathPHA 1998 An arthropod phylogeny based on fossil and recent taxaIn GD Edgecombe (ed) Arthropod Fossils and Phylogeny 33ndash105Columbia University Press New York
Wilson LA 2006 Food for thought a morphological and taphonomic studyof fossilised digestive systems from Early to Middle Cambrian taxa275 pp Unpublished PhD thesis University of Cambridge Cambridge
Zhang XminusG and Hou X 2007 Gravitational constraints on the burial ofChengjiang fossils Palaios 22 448ndash453
Zhang XminusL Han J Zhang Z Liu H and Shu D 2004 Redescription ofthe Chengjiang arthropod Squamacula clypeata Hou and Bergstroumlm fromthe Lower Cambrian Southminuswest China Palaeontology 47 605ndash617
Zhao Y Zhu M Babcock LE Yuan L Parsley RL Peng J YangX and Wang Y 2005 Kaili Biota a taphonomic window on diversifiminuscation of metazoans from the basal Middle Cambrian Guizhou ChinaActa Geologica Sinica 79 751ndash765
400 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011

Great appendage robust proportions of peduncle and disminustal spineminusbearing articles as in other members of Leanchoiliminusidae more commonly preserved with posterior orientation
but anteriorly oriented in SAM P45906 and SAM P45910Each of the three long spinose projections terminating at thesame length the projections evenly tapering distally with
392 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Fig 6 Leanchoiliid megacheiran arthropod Oestokerkus megacholix gen et sp nov SAM P45168 A head and anterior part of trunk B Detail of spinoseprojections of great appendages and excretory organs (sensu Bruton and Whittington 1983) Abbreviations cs cephalic shield eo excretory organ gagreat appendage mg midgut glands tx trunk exopods
margins straight for much of their length or slightly curvedterminally distal spinose projection distinctly stouter at itsbase than the two more proximal projections and more markminusedly tapering along its length Dark irregularminusshaped low reminuslief lump on basal spineminusbearing article of great appendagesof SAM P45168 (eo in Fig 6) and SAM P45908 correminussponding to excretory organ of Bruton and Whittington(1983) Long flagella extending from each of the threespinose projections seen only in SAM P43631 (Figs 23A3) of similar narrow width along its preserved extent anminusnulations indistinguishable
Trunk of 11 segments of even length Maximum height oftrunk relative to length (excluding telson) ca 39 bodydeepest at tergite 4 Trunk tergite boundaries marked as narminusrow ridges in several specimens corresponding to narrowband of overlap between adjacent tergites (ol in Figs 2 3A14A1 B) Some specimens show tergites to be bisected by anarrow but sharp furrow of uncertain identity (eg possiblyimpression of dorsal transverse tendon cf Stein 2010 fig12B C E) (arrows in Figs 3A1 4B)
Exopods with fringe of long setae on cephalic and trunkappendages (Fig 2) setal fringe gradually shortening fromabout the fourth trunk segment Setae project ventrally bymore than 30 height of tergites in deepest part of trunk(Fig 2B) setae flattened narrow and parallelminussided alongmost of their length tapering near their distal ends to termiminusnate as a pointed tip (Figs 2B 3A3 arrows) Endopods exminusposed only in SAM P45167 (Fig 4) two pairs associatedwith cephalon (Fig 4B) those of anterior segments of trunkof similar morphology to those in cephalon all slender lackminusing obvious endites podomere boundaries unrecognisable
Midgut glands occupy a large extent of the dorsoventralheight of the trunk two midgut glands in cephalon of similarsize to those in each of the first seven segments of the trunk(Figs 3A1 5A 6A) all developed as radiating grooves emaminusnating from a globular structure at their ventral edge as in
Leanchoilia superlata (GarciacuteaminusBellido and Collins 2007 pl2 6) Foregut trace in SAM P43631 (Fig 2) indicates that gutcurves ventrally in anterior part of cephalon mouth regionnot preserved Terminal part of hindgut (in trunk segment 11)with closely spaced longitudinal lineations (Fig 2) that apminusparently correspond to folds in the gut wall hindgut termiminusnating at base of telson
Telson most completely preserved in holotype (SAMP43631 Figs 2B 3A2 B) In this specimen the telson likerest of body compressed in lateral aspect Telson shows meminusdian and marginal reinforcing ridges connecting a flat trianminusgular surface this surface appears to be partially broken inSAM P43631 showing underlying surface of other (left) halfof telson Lateral preservation of specimen implies that topcorresponds to an axial structure and bottom to a marginalone 30ndash35 angle in posterior edge of vane connecting thesetwo structures requires that in dorsal aspect telson would apminuspear forked and widening distally Five stout pointed dorminussally curved marginal spines attached to outer posterior edgeon each side of telson
Phylogenetic analysis ofLeanchoiliidaeDetermining the systematic position of Oestokerkus demandsan appraisal of the relationships of Leanchoiliidae as a wholeCladistic analysis including the bestminusknown members ofMegacheira including leanchoiliids has been conducted byCotton and Braddy (2004) and Vannier et al (2009) The taxominusnomic sampling used by Cotton and Braddy (2004) includedseveral megacheiran genera representative chelicerates anumber of taxa representing groups that have been proposedas possible chelicerate relatives (eg Sidneyia AglaspisCheloniellon) in other analyses (Dunlop and Selden 1997Hou and Bergstroumlm 1997) and members of the major lineagesof the Artiopoda sensu Hou and Bergstroumlm (1997) ie trilominusbiteminusallied lamellipedians Because artiopodan interminusrelationminusships have remained quite stable through a series of phylogenminusetic analyses (Edgecombe and Ramskoumlld 1999 Cotton andBraddy 2004 Hendricks and Lieberman 2008 Paterson et al2010) and artiopodans are now often regarded as less closelyrelated to Megacheira than is Chelicerata (Chen et al 2004Cotton and Braddy 2004 Dunlop 2006) we have reduced thesampling of Artiopoda in the present analysis To augment thediversity of ldquoshort great appendagerdquo arthropods Haikoucarisis added based on the description of H ercaiensis by Chen etal (2004) Offacolus (Orr et al 2000 Sutton et al 2002) hasbeen allied to Chelicerata and is included as a terminal Beminuscause the present analysis is principally concerned with relaminustionships of and within Leanchoiliidae sensu Liu et al (2007)we follow Vannier et al (2009) in sampling Alalcomenaeusand Leanchoilia at the species level Within Chelicerata Cotminuston and Braddy (2004) and Hendricks and Lieberman (2008)sampled ancestral states in Xiphosura by coding the Devonian
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 393
Fig 7 Reconstruction of Oestokerkus megacholix gen et sp nov in lateralview Total length and annulations on the flagella and positions of podominusmere boundaries of the frontal appendage are based on Leanchoilia superminuslata (Bruton and Whittington 1983)
synziphosurine Weinbergina Our coding for Xiphosura alsoemploys information from extant xiphosurans because theysupply more reliable information on the homology of the partsof the trunk appendages (Suzuki and Bergstroumlm 2008) and eyeanatomy (Harzsch and Hafner 2006) among other charactersystems than do fossils alone
Our character set extracts characters from the matrix ofCotton and Braddy (2004) that vary in the revised taxonomicsample reformulates some characters to accommodate reminusvised homology concepts and adds new characters especiallyfor leanchoiliids In total 32 characters are included (Table 1)In this analysis the primary homology of the ldquoshort great apminuspendagerdquo and chelicera is coded (as described by character 4)and the taxon sampling broadly constrains megacheirans aschelicerate relatives (as defended by Chen et al 2004 Cottonand Braddy 2004) rather than testing the alternative that theycould be stem group Euarthropoda (Budd 2002 Bergstroumlmand Hou 2005) A test of the two alternatives involves samminuspling of the entire diversity of euarthropods and a range ofnonminuseuarthropod outgroups an exercise beyond the scope ofthis study
Characters1 Mineralised cuticle (0) absent (1) present For definiminus
tion and discussion of this character see Cotton andBraddy (2004 character 30) and Paterson et al (2010character 1)
2 Curvature of anterior margin of cephalic shield (0) rounminusded or straight (1) pointed medially (2) with multiplespinose projections This character codes for variability inthe shape of the cephalic shield in Leanchoiliidae (state 0and 1 sensu GarciacuteaminusBellido and Collins [2007 textminusfigs10 and 9 respectively]) State 2 is added to accommodateOffacolus the median spine of a series that fringes the ceminusphalic margin is not convincingly homologised with themedian projection in leanchoiliids
3 Number of cephalic somitesappendages (0) antenna orfrontal appendage only (1) antenna or frontal appendageand two limb pairs (2) antenna or frontal appendage andthree limb pairs (3) antenna or frontal appendage and fourlimb pairs (4) antenna or frontal appendage and five limbpairs Coding for lamellipedians and Aglaspis is as discussedby Paterson et al (2010 character 2) Coding for Alalcomeminusnaeus cambricus (state 1) follows Briggs and Collins (1999)Leanchoilia illecebrosa (state 2) follows Liu et al (2007)and Leanchoilia persephone and Leanchoilia superlata (state1) follows GarciacuteaminusBellido and Collins (2007)
4 Form of first cephalic appendage (0) antenna (1) anteminusriorly directed raptorial appendage with reduced numberof podomeres each of the terminal podomeres bearing aspinose process on its distal margins a geniculation beminustween spinose podomeres and peduncle This characteris formulated to identify a homology between the frontalappendages of megacheirans and the chelicera of Cheliminus
cerata and state 1 corresponds to the putative apomorminusphic detail defended by Chen et al (2004) and Cottonand Braddy (2004) Vannier et al (2009 character 8)specify another shared property of ldquoshort great appendminusagesrdquo of the megacheiran (ss) type in their characterstate ldquobent at an anglerdquo referring to the geniculation beminustween articles of the peduncle or between the peduncleand distal spiniferous articles This property is sharedwith Chelicerata
5 Number of spineminusbearing articles on frontal appendage (0)three + movable finger (1) two + movable finger (2) one+ movable finger The number of spineminusbearing articles onthe frontal appendage (exclusive of the terminal movablefinger) varies from three in Yohoia (Whittington 1974 pl6 1ndash3 pl 7 3ndash6 pl 9 4ndash6) and Fortiforceps (Hou andBergstroumlm 1997 figs 31C D 32A 33B C E) to two inHaikoucaris (Chen et al 2004 fig 2A B) and Leanminuschoiliidae or only one the last being a chelicera
6 Long spinose projections on distal part of terminal threepodomeres of frontal appendage bearing a flagellum (0)absent (1) present The morphology of the great appendminusage in Leanchoiliidae is coded here We have not codedthe length of the spinose projections and the presence ofa flagellum as two independent characters Cotton andBraddyrsquos (2004) character 6 state 2 (ldquospinesrdquo longer thanthe entire podomere series) and character 8 state 1 (presminusence of an annulated flagellum) are thus grouped as a sinminusgle trait here
394 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Table 1 Characters coded for phylogenetic analysis Polymorphismindicated by ldquo+rdquo partial uncertainty by states separated by ldquordquo Abbreviminusations A Alalcomenaeus L Leanchoilia O Oestokerkus
Squamacula 0000minus minus minus2minus0 0110 00000 10minus minus0 1000 0
Cindarella 0030minus minus minus000 0010 00000 10minus minus0 11121 00
Kuamaia 000minus minus minus001 0011 10000 10minus minus0 11121 00
Kwanyinaspis 00 001 00112 1000 1000 1121 00
Olenoides 1020minus minus minus100 0011 00000 10minus minus0 01121 00
Retifacies 0020minus minus minus000 0010 00000 11000 1020 00
Aglaspis 10(23)0minus minus minus100 00101 00010 01001 0 00
Sidneyia 0000minus minus minus000 01102 00110 01101 00020 00
Emeraldella 0040minus minus minus2minus 0 00103 00110 01001 001(1+2)1 00
A cambricus 00111 10010 00001 00000 01110 01110 10
L illecebrosa 01211 11010 00001 01001 01210 11110 10
L persephone 00111 10010 00001 00000 01210 11110 10
L superlata 01111 11010 00001 01001 01210 11110 10
L hanceyi 00 00 0001 0000 01210
O megacholix 0011 100 00001 00000 01110 1110 0
Jianfengia 0021 0minus000 00005 00000 01000 010(12)0 0
Fortiforceps 00210 0minus00 00004 00000 01100 01230 20
Yohoia 00210 0minus000 00003 00011 01110 00010 (01)1
Haikoucaris 00211 0minus00 00003 00000 01(12)0 1010 0
Cheloniellon 0040minus minus minus100 00100 10010 01300 0030 00
Offacolus 02412 0minus200 0000 00010 01000 13minus minus 01
Xiphosura 00412 0minus100 10000 0010 01000 000minus minus 01
Eurypterida 00412 0minus100 10002 00010 01000 00minus minus minus 01
7 Length of flagellate great appendage (0) flagella termiminusnating at midlength of trunk (1) flagella extending to endof body Coding is applicable to Leanchoiliidae onlydistinguishing some Leanchoilia species that resembleAlalcomenaeus in having relative short great appendages(state 0) versus the longer appendages in L superlata andL illecebrosa
8 Position of lateral facetted eyes (0) ventral (1) dorsalsessile (2) absent Variation in Lamellipedia and severalother taxa coded herein follows Paterson et al (2010character 3)
9 Four or five ventral eyes arranged in a subtransverseband across head shield (0) absent (1) present A bandof ventral eyes is shared by the best known species ofLeanchoiliidae The distinction between three medianeyes in Alalcomenaeus cambricus (Briggs and Collins1999) and two in Leanchoilia illecebrosa (Liu et al2007) and both Leanchoilia superlata and L persephone(GarciacuteaminusBellido and Collins 2007) is uninformative forthe taxon sample here
10 Bulge in exoskeleton accommodating dropminusshaped venminustral eyes (0) absent (1) present This character followsEdgecombe and Ramskoumlld (1999 character 6) and Patminuserson et al (2010 character 5)
11 Dorsal median eyes (0) absent (1) present Coding for anabsence of dorsal median eyes in fossil taxa considers theabsence of either paired ocelli or a median eye tubercleand the character is scored as a chelicerate autapomorphy(see Cotton and Braddy 2004 character 24)
12 Width of doublure on cephalic shield (0) narrow or modminuserately wide (1) entire ventral side of head covered byexpansion of doublure The interpretation of state 1 inSidneyia and Squamacula is discussed by Cotton andBraddy (2004) and Paterson et al (2010 characters 12and 29)
13 Mouth covered by hypostome (0) absent (1) present Noldquoshort great appendagerdquo arthropods have been shown topossess a hypostome and a reconstruction of a smalllabrum in Alalcomenaeus (Briggs and Collins 1999) wasadmittedly conjectural The absence of a conspicuoushypostome contrasts with its ubiquitous presence in laminusmellipedians and several putative chelicerateminusallied taxathat have antennae (such as aglaspidids and Emeralminusdella) Chelicerates have a small epistomeminuslabrum (Bitschand Bitsch 2007) but like Cotton and Braddy (2004character 27 state 4) we code a hypostomal sclerite aslacking Kwanyinaspis has a large hypostome similar tothat found in aglaspidids (photograph of the holotype ofKwanyinaspis maotianshanensis shown to us by JavierOrtegaminusHernaacutendez [personal communication 2010])
14 Tergite articulations (0) extensive overlap of tergites (1)edge to edge articulations This character follows Edgeminuscombe and Ramskoumlld (1999 character 18) and Patersonet al (2010 character 20)
15 Number of trunk segments (0) nine (1) 11 (2) 12 (3)13 (4) 20 (5) 21 or 22 Coding is confined to taxa that do
not possess a pygidium because the precise number ofsomites associated with pygidia can sometimes not bedetermined with confidence Codings for Xiphosura andEurypterida use opisthosomal somite numbers followingShultz (2007 character 95)
16 Exoskeletal boundaries of anterior trunk segments (0)boundaries transverse or reflexed posterolaterally (1) reminusflexed anterolaterally This character follows Edgecombeand Ramskoumlld (1999 character 19) and Paterson et al(2010 character 21)
17 Trunk tergites with serrate lateral margins (0) absent (1)present Liu et al (2007 fig 2C) documented a row ofmarginal spines on the tergites of Leanchoilia illeceminusbrosa a featured shared by Leanchoilia superlata (egGarciacuteaminusBellido and Collins 2007 pl 1 7) but not otherterminals coded here
18 Differentiation in width of trunk tergites (0) trunk withgradational narrowing posteriorly (1) abrupt narrowingat boundary between prothorax and opisthothorax Anabrupt contraction in width between the boundary ofthe prominus and opisthothorax is observed in Emeraldellaand Sidneyia Yohoia has been reconstructed as having apronounced break in the shape of the trunk tergites beminushind segment 10 (Whittington 1974 textminusfig 2) thepoint at which the paratergal folds lose their spinose tipsand appendages are lacking Specimens preserved indorsal aspect (Whittington 1974 pl 10) suggest that thewidth of the opisthothorax is rather less abruptly narminusrowed and we code Yohoia as state 0
19 Appendages of posterior trunk segments (0) all trunktergites bearing appendages (1) one or more posteriortrunk segments lacking appendages Coding correspondsto Cotton and Braddyrsquos (2004) character 43 ldquopostabdominusmen of segments lacking appendagesrdquo
20 Paired tergal carinae (0) absent (1) present Liu et al(2007 265 fig 2F) recorded the presence of tergalcarinae in Leanchoilia illecebrosa shared with L superminuslata (eg Bruton and Whittington 1983 pl 17 106GarciacuteaminusBellido and Collins 2007 pl 1 5)
21 Pygidium (0) absent (1) present Coding of the pygiminusdium in many terminals herein was discussed by Paterminusson et al (2010 character 22)
22 Telson (0) absent (1) present Following Hendricks andLieberman (2008) a telson is a postminussegmental terminalstructure that lacks appendages Dunlop and Selden (1997)considered a telson to be absent in Cheloniellon but weidentify the conical projection posterior to the ultimate legpair and cerci (Stuumlrmer and Bergstroumlm 1978 figs 2 3 1011) as being most consistent with an identity as a telson
23 Telson shape (0) styliform (1) flapminuslike (2) lanceolate(3) short conical The outline of the telson in Haikouminuscaris ercaiensis is incompletely known but the mostcomplete specimen (Chen 2004 fig 472) constrains it aseither state 1 or state 2 A partial uncertainty coding isthus employed A unique state for Cheloniellon (state 3)is based on the interpretation in character 22
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 395
24 Telson fringed with spines (0) spines absent (1) spinespresent Presence of telson spines was introduced as a charminusacter by Hendricks and Lieberman (2008 character 34)
25 Paired flaps lateral to telson (0) absent (1) present Thischaracter describes the postminusventral plates of several taxa(see Edgecombe and Ramskoumlld 1999 character 25) Cotminuston and Braddy (2004 character 54) raised the possibilitythat the cerci of Cheloniellon could be homologous withpostminusventral plates (see also Hou and Bergstroumlm 1997 93)because they are similarly paired structures derived fromthe segment immediately anterior to the telson We havenot coded for this homology because the cerci of Chelominusniellon originate dorsally rather than ventrally
26 Midgut glands with submillimetric lamellae (0) absent(1) present Midgut glands in some stem group arthrominuspods and early crown group arthropods are metamericreniform organs with a characteristic lamellar structure(Butterfield 2002) Their morphology is identical inLeanchoilia and Oestokerkus notably including groovesthat radiate from a ventral median node (Figs 3A1 6A)The gut of Alalcomenaeus cambricus is more tubular(Briggs and Collins 1999 textminusfigs 10 11) and thoughan anterior six segments may display phosphatised latminuseral projections that likely represent gut diverticula (Wilminusson 2006) they have not been shown to have the detailedmorphology of Leanchoiliaminustype midgut glands Butterminusfield (2002 164) cited a figure from Bruton (1981 fig96) in support of ldquolamellarrdquo midgut glands in the cephaminuslon of Sidneyia Like Bruton (1981) in his description ofthe specimen we consider it likely that the structures inquestion are indeed gut diverticula but there is no basisfor identifying them more precisely as the reniformlamellar structures coded under this character
27 Exopod on first postminusantennal first postminusfrontal appendminusage segment (0) absent (1) present This character reminuslates to Cotton and Braddyrsquos (2004) character 3
28 Division of exopod (0) exopod an undivided flap (1)exopod differentiated into proximal and distal lobes (2)small flap with radiating lamellae (3) pediform exopoddivided into ca 6 articles Identification of the exopod inXiphosura follows Suzuki and Bergstroumlm (2008) Thischaracter groups conditions of the exopod shaft coded asindependent characters by Cotton and Braddy (2004)eg state 0 corresponds to their character 16 and state 1to their character 13 Our coding permits transformationbetween any two states without an extra implicit step
29 Exopod setae (0) short spiniform setae (1) long flatminustened tapering distally with slight separation (2)lamellar setae (flattened imbricating) (3) short finesetae A distinction is made between lamellipedian setaewhich are imbricated (state 2) and the pointed more disminustinctly separated and variably spiniform exopod setae ofleanchoiliids (state 1) This distinction is consistent withthe identification of megacheirans as nonminuslamellipedians(eg Hou and Bergstroumlm 1997 Bergstroumlm and Hou2005) Although Cotton and Braddy (2004 character 18)
regarded the form of the setae in Yohoia to be uncertainwe regard specimens figured by Whittington (1974 pl3 3) as conforming to the leanchoiliid state Squamaculadiffers from other taxa in its shorter spiniform setae(state 0) (Zhang et al 2004 textminusfig 1A B) Offacolushas longer spiniform projections on the distal articles ofthe exopod that have been variably identified as eitherspines or setae (Sutton et al 2002 fig 4b) but theirhomology with the setae that splay regularly along themargin of a flapminuslike exopod (eg states 0ndash3) is uncerminustain This and the following character for exopod setaeare left as ldquordquo for Offacolus A homology between thegill lamellae of Xiphosura and exopod setae of lamellipeminusdians has a long history (see Cotton and Braddy 2004character 18) We follow Suzuki and Bergstroumlm (2008)in identifying the xiphosuran gills as outgrowths of thebasis rather than of the exopod this perspective negates ahomology with the exopod setae of Lamellipedia Thepolymorphic coding for Emeraldella reflects the twoseparate states (1+2) displayed by the exopods in thetrunk (see Bruton and Whittington 1983 pl 8 45 47Edgecombe and Ramskoumlld 1999 fig 11)
30 Differentiation of setae along exopod (0) uniform orgradationally variable setae along length of exopod (1)lamellar setae confined to proximal lobe with distal lobebearing slender setae
31 Endopod segmentation (distal to coxa) (0) six or sevenpodomeres and terminal claw or telotarsal spurs (1)eight or more podomeres and terminal claw (2) ca 15podomeres State 1 is formulated to describe the segmenminustation of the leanchoiliid leg (GarciacuteaminusBellido and Collins2007 textminusfig 6 Liu et al 2007 fig 5) in contrast to thereduced number of podomeres in other euarthropods(Boxshall 2004) State 2 characterises Fortiforceps Apartial uncertainty coding is employed for Yohoia inwhich the precise number of podomeres is undeterminedand could be either state 0 or 1 (Whittington 1974 pl 74) but the spacing of the preserved podomere boundariesnegates the possibility of state 2
32 Endopods on anterior postminuscephalic segments (0) welldeveloped (1) rudimentary or lacking Following Cottonand Braddy (2004 character 11) the absence of trunkendopods in Yohoia (Whittington 1974) is coded as a puminustative homologue with the suppression of postminuscephalicendopods in Chelicerata
ResultsAs a convention for depicting the trees networks are rootedon Squamacula which separates artiopodan Lamellipedia(sensu Hou and Bergstroumlm 1997) from a group that includesMegacheira Chelicerata and Xenopoda all members ofwhich possess a telson
Under equal weights 14 shortest cladograms have a lengthof 77 steps with a Consistency Index of 064 and Retention Inminus
396 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
dex of 077 The strict consensus of these trees with supportvalues is shown in Fig 8 Leanchoiliidae is unambiguouslysupported in all 14 cladograms by four apomorphic characterstwo limbminusbearing cephalic segments posterior to the frontalappendage (character 3 state 1) long spinose projections onthe distal part of the terminal three podomeres of the frontalappendage that bear flagella (character 6 state 1) 11 trunksegments (character 15 state 1) and division of the exopodsinto two lobes (character 28 state 1) The arrangement of fouror five eyes in a subtransverse band across the cephalic shieldis unique to Leanchoiliidae (character 9 state 1) but is not anunambiguous autapomorphy of the group because the state isuncertain in close relatives notably Haikoucaris It is apominusmorphic for Leanchoiliidae only under particular optimisaminustions (Delayed Transformation)
The internal relationships of Leanchoiliidae are stableacross all 14 shortest cladograms Leanchoilia is united by thelanceolate telson (character 23 state 2) a unique unambiguminusous autapomorphy Within Leanchoilia a sister group relaminustionship between L superlata and L illecebrosa is well supminusported receiving a Bremer support of 3 and jackknife freminusquency of 86 These two species share four synapomorminusphies a medially pointed anterior cephalic margin (character2 state 1) flagella of the great appendage extending the lengthof the body (character 7 state 1) serrate lateral margins on thetrunk tergites (character 17 state 1) and paired tergal carinae(character 20 state 1) As such a species from Chengjiang (Lillecebrosa) and the Burgess Shale (L superlata) are moreclosely related to each other than are the two Burgess Shalecongeners a relationship that weakens the possibility that theoften cominusoccurring L superlata and L persephone could besexual dimorphs (GarciacuteaminusBellido and Collins 2007) ratherthan separate species The union of Leanchoilia and Oestominuskerkus is unambiguously supported by the shared midgutgland structure of both genera (character 26 state 1)
All shortest cladograms resolve Haikoucaris Yohoiaand Fortiforceps in that order as progressively less closelyrelated to Leanchoiliidae The union of these three taxa withLeanchoiliidae yields a clade that corresponds to Megacheirasensu Hou and Bergstroumlm (1997) apart from the ambiguousplacement of Jianfengia In half of the 14 shortest cladominusgrams Jianfengia is sister to the other megacheirans yieldminusing a monophyletic Megacheira ss In the remaining cladominusgrams Jianfengia is sister to Xenopoda sensu Hou andBergstroumlm (1997) Aglaspis and Chelicerata Haikoucarisand Leanchoiliidae are united by a frontal appendage comminusposed of two spineminusbearing articles in addition to the movminusable finger (character 5 state 1) Yohoia groups with thesetaxa based on spines fringing the telson (character 24 state1) and Fortiforceps is united with them at a broader levelbased on the flapminuslike telson shape (character 23 state 1)When Jianfengia is placed as sister to the other megacheiminusrans the entire clade is supported by the raptorial frontal apminuspendage (character 4 state 1) and the lack of a large hypominusstomal shield (character 13 state 0) In this set of cladominusgrams these features are nonminushomologous between Megaminus
cheira and Chelicerata whereas in some cladograms with thealternative resolution of Jianfengia as closer to cheliceratescharacter 4 is optimised as homologous in Megacheira andChelicerata
Reanalysis of the data with implied weights finds two optiminusmal trees (Fig 9) Relationships within Leanchoiliidae underequal weights are completely stable to reweighting as is theresolution of Haikoucaris as sister to Leanchoiliidae As suchthe character optimisations described above for the leanchoiminusliids are depicted the same in Fig 9 The main difference beminustween equal and implied weights involves the placement ofChelicerata Under equal weights (Fig 8) chelicerates are alminuslied to Xenopoda (here Emeraldella + Sidneyia + Chelominusniellon sensu clade L in Hou and Bergstroumlm 1997 fig 88)and Aglaspidida A clade composed of Aglaspidida Xenominuspoda and Chelicerata was similarly identified by Hou andBergstroumlm (1997 fig 88 their clade K) In our analysis thisgroup consistently finds support from one or more posteriortrunk segments lacking appendages (character 19 state 1) anddepending on alternative internal resolutions it is variably supminusported by dorsallyminuspositioned sessile eyes (character 8 state 1)and the absence of exopods on the first postminusantennalpostminuscheliceral limb (character 27 state 1) Under implied weightshowever chelicerates are more closely allied to Megacheirathan to aglaspidids or xenopodans (Fig 9)
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 397
2
582
76
Chelicera-morpha
Megacheira
62
2
53
50Leanchoiliidae
3
86
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
EurypteridaChelicerata
Fortiforceps
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia hanceyi
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
Fig 8 Strict consensus of 14 shortest cladograms (77 steps) based on charminusacter data in Table 1 Numbers above nodes are Bremer support those beminuslow nodes are jackknife frequencies gt50 Cheliceramorpha correspondsto a clade with the same composition in Cotton and Braddy (2004)
The major groupings in Fig 9 correspond closely to thoseidentified by Cotton and Braddy (2004) Notably the cladeuniting Aglaspis Xenopoda Offacolus Chelicerata Jianminusfengia and other Megacheira corresponds to their ldquoClade 3rdquo(Cotton and Braddy 2004 fig 8) for which they proposedthe name Cheliceramorpha The clade uniting CheloniellonAglaspis Sidneyia and Emeraldella in Fig 9 corresponds totheir ldquoClade 5rdquo and the clade that groups Megacheira andChelicerata is equivalent to their ldquoClade 4rdquo Their analysisdid not identify megacheirans as a monophyletic groupChelicerata was nested within the group and among taxaconsidered here Yohoia was resolved closer to cheliceratesthan to Leanchoiliidae In contrast Yohoia was regarded byBergstroumlm and Hou (2005) as more closely allied to Leanminuschoiliidae than to chelicerates a megacheiran grouping morecompatible with our cladograms The analysis of ldquogreat apminuspendage arthropodsrdquo by Vannier et al (2009) identified a
monophyletic Leanchoiliidae but a polyphyletic Megacheira(to the exclusion of Haikoucaris and Jianfengia) Howevertheir resolution of Fortiforceps and even more so Yohoia asthe closest relatives of Leanchoiliidae is as in our trees (apartfrom the different position of Haikoucaris)
In summary a major controversy in the systematics of theldquoshort great appendagerdquo arthropods remains whether thechelicerates are nested within the group or are instead moreclosely allied to taxa conventionally identified as lamelliminuspedians specifically Xenopoda and Aglaspidida We findthat both of these results are compatible with the availabledata depending on character weighting protocols Noneminustheless the broader megacheiranminuschelicerate issue does notcloud the status of leanchoiliids as a clade or the identificaminustion of their closest relatives (ie Haikoucaris and Yohoia)and the internal relationships of Leanchoiliidae are stableand supported by compelling characters
398 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
Eurypterida
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
2 7 17 20
3
2
1 1 1
26
1
281563
1 1 1 1
5
1
24
1
23 19 20 27 32
28Clade 4
1 0
4 13 11 27 0
154 1 1
3 19 32Clade 3
2 3
2 28
0
3 1 121
1
22
Clade 5
1 0
19 27
1
25
15 28 301 1
2312
1 150 1 3 3
15 16 2923
18
1 1
10 16
1 1
1 8
1
14
3
3
28 30
1 0
5
15
1 1
1
1
22
Fortiforceps
23
1 1 0 1
1 1
Leanchoilia hanceyi
2
2
1
1
Fig 9 Characters optimised on one of two shortest cladograms with imminusplied weights using a concavity constant k = 3 Black squares are nonminushomoplastic changes white squares homoplastic changes all changes areunambiguous Branches without strict support collapsed after optimisingcharacters Numbers for characters (above the branches) and states (belowthe branches) correspond to Table 1 ldquoClades 3ndash5rdquo correspond to groups sonumbered by Cotton and Braddy (2004 fig 8)
AcknowledgementsAccess to the field area has been most kindly allowed by the Buck famminusily We thank our collaborators in Emu Bay Shale research Jim Jago(University of South Australia Mawson Lakes) Jim Gehling andMike Lee (South Australian Museum Adelaide) for tremendous supminusport and advice as well as Mike Gemmell Dennis Rice Allison DaleyAaron Camens John Laurie Glenn Brock Pierre Kruse and Trevorand Jenny Worthy (all South Australian Museum) for their assistancein the field and Natalie Schroeder MaryminusAnne Binnie and RondaAtkinson (all South Australian Museum) for organisational support inthe lab and in the field Javier OrtegaminusHernaacutendez (University of Camminusbridge Cambridge UK) advised us on Kwanyinaspis and the journalrsquosreferees provided useful feedback TNT is available courtesy of theWilli Hennig Society Funding has been provided by an AustralianResearch Council Linkage grant (LP0774959) to the University ofAdelaide with generous financial assistance from Beach Energy Ltdand the South Australian Museum SeaLink provided logistical supminusport DGBrsquos travel to Australia has been supported by grants from theSpanish Research Council (CSIC PA 1001105) the Complutense Uniminusversity of Madrid and the Spanish Ministry of Education and Science(CGL2006minus12245BTE CGL2009minus07073 and RYC2007minus00090)
ReferencesBergstroumlm J and Hou XminusG 2005 Early Palaeozoic lamellipedian arthrominus
pods In S Koenemann and RA Jenner (eds) Crustacea and Arthrominuspod Relationships Crustacean Issues 16 73ndash93
Bitsch J and Bitsch C 2007 The segmental organization of the head reminusgion in Chelicerata a critical review of recent studies and hypothesesActa Zoologica 88 317ndash335
Bousfeld EL 1995 A contribution to the natural classification of Lower andMiddle Cambrian arthropods food gathering and feeding mechanismsAmphipacifica 2 3ndash34
Boxshall GA 2004 The evolution of arthropod limbs Biological Reviews79 253ndash300
Bremer K 1994 Branch support and tree stability Cladistics 10 295ndash304Briggs DEG and Collins D 1999 The arthropod Alalcomenaeus cambricus
Simonetta from the Middle Cambrian Burgess Shale of British ColumbiaPalaeontology 42 953ndash977
Briggs DEG and Robison RA 1984 Exceptionally preserved nonminustrilobite arthropods and anomalocaridids from the Middle Cambrian ofUtah University of Kansas Paleontological Contributions 111 1ndash23
Briggs DEG Lieberman BS Hendricks JR Halgedahl SL andJerrard RD 2008 Middle Cambrian arthropods from Utah Journal ofPaleontology 82 238ndash254
Bruton DL 1981 The arthropod Sidneyia inexpectans Middle CambrianBurgess Shale British Columbia Philosophical Transactions of theRoyal Society of London Series B 295 619ndash656
Bruton DL and Whittington HB 1983 Emeraldella and Leanchoiliatwo arthropods from the Burgess Shale Middle Cambrian British Cominuslumbia Philosophical Transactions of the Royal Society of London Seminusries B 300 553ndash582
Budd GE 2002 A palaeontological solution to the arthropod head probminuslem Nature 417 271ndash275
Butterfield NJ 2002 Leanchoilia guts and the interpretation of threeminusdiminusmensional structures in Burgess Shaleminustype fossils Paleobiology 28155ndash171
Chen JminusY 2004 The Dawn of Animal World [in Chinese] 366 pp JiangScience and Technology Press Nanjing
Chen JminusY Waloszek D and Maas A 2004 A new ldquogreatminusappendagerdquo arminusthropod from the Lower Cambrian of China and homology of cheliceratechelicerae and raptorial anterominusventral appendages Lethaia 37 3ndash20
Cotton TJ and Braddy SJ 2004 The phylogeny of arachnomorph arthrominuspods and the origin of the Chelicerata Transactions of the Royal Societyof Edinburgh Earth Sciences 94 169ndash193
Daley AC Budd GE Caron JminusB Edgecombe GD and Collins D2009 The Burgess Shale anomalocaridid Hurdia and its significancefor early euarthropod evolution Science 323 1597ndash1600
Dunlop JA 2006 New ideas about the euchelicerate stemminuslineage In CDeltshev and P Stoev (eds) European Arachnology 2005 Acta Zoominuslogica Bulgarica (Supplement 1) 9ndash27
Dunlop JA and Selden PA 1997 The early history and phylogeny of thechelicerates In RA Fortey and RH Thomas (eds) Arthropod Relaminustionships Systematics Association Special Volume 55 221ndash235
Edgecombe GD and Ramskoumlld L 1999 Relationships of CambrianArachnata and the systematic position of Trilobita Journal of Paleonminustology 73 263ndash287
Farris JS Albert VA Kaumlllersjouml M Lipcomb D and Kluge AG1996 Parsimony jackknifing outperforms neighborminusjoining Cladistics12 99ndash124
GarciacuteaminusBellido DC and Collins D 2007 Reassessment of the genusLeanchoilia (Arthropoda Arachnomorpha) from the Middle CambrianBurgess Shale British Columbia Canada Palaeontology 50 693ndash709
GarciacuteaminusBellido DC Paterson JR Edgecombe GD Jago JB GehlingJG and Lee MSY 2009 The bivalved arthropods Isoxys and Tuzoiawith softminuspart preservation from the lower Cambrian Emu Bay ShaleLagerstaumltte (Kangaroo Island Australia) Palaeontology 52 1221ndash1241
Goloboff PA 1993 Estimating character weights during tree searchCladistics 9 83ndash91
Goloboff PA Farris JS and Nixon KC 2008 TNT a free program forphylogenetic analysis Cladistics 24 774ndash786
Harzsch S and Hafner G 2006 Evolution of eye development in arthrominuspods phylogenetic aspects Arthropod Structure amp Development 35319ndash340
Hendricks JR and Lieberman BS 2008 New phylogenetic insights intothe Cambrian radiation of arachnomorph arthropods Journal of Paleminusontology 82 585ndash594
Hou XminusG and Bergstroumlm J 1997 Arthropods of the Lower CambrianChengjiang fauna southwest China Fossils and Strata 45 1ndash116
Jago JB Zang W Sun X Brock GA Paterson JR and SkovstedCB 2006 A review of the Cambrian biostratigraphy of South Austraminuslia Palaeoworld 15 406ndash423
Liu Y Hou XminusG and Bergstroumlm J 2007 Chengjiang arthropod Leanminuschoilia illecebrosa (Hou 1987) reconsidered GFF 129 263ndash272
Luo H Hu S Chen L Zhang S and Tao Y 1999 Early CambrianChengjiang fauna from Kunming region China [in Chinese] 129 ppYunnan Science and Technology Press Kunming
Luo H Hu S Zhang S and Tao Y 1997 New occurrence of the EarlyCambrian Chengjiang fauna in Haikou Kunming Yunnan Provinceand study on Trilobitoidea Acta Geologica Sinica 71 122ndash132
Nixon KC 2002 WINCLADA Version 19908 Cornell University IthacaNY
Orr PJ Siveter DJ Briggs DEG Siveter DJ and Sutton MD 2000A new arthropod from the Silurian KonservatminusLagerstaumltte of Hereminusfordshire Proceedings of the Royal Society Series B Biological Sciminusences 267 1497ndash1504
Paterson JR and Brock GA 2007 Early Cambrian trilobites fromAngorichina Flinders Ranges South Australia with a new assemblagefrom the Pararaia bunyerooensis Zone Journal of Paleontology 81116ndash142
Paterson JR and Jago JB 2006 New trilobites from the Lower CambrianEmu Bay Shale Lagerstaumltte at Big Gully Kangaroo Island South Ausminustralia Memoirs of the Association of Australasian Palaeontologists 3243ndash57
Paterson JR Edgecombe GD GarciacuteaminusBellido DC Jago JB andGehling JG 2010 Nektaspid arthropods from the lower CambrianEmu Bay Shale Lagerstaumltte South Australia with a reassessment oflamellipedian relationships Palaeontology 53 377ndash402
Paterson JR Jago JB Gehling JG GarciacuteaminusBellido DC Edgecombe
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 399
GD and Lee MSY 2008 Early Cambrian arthropods from the EmuBay Shale Lagerstaumltte South Australia 319ndash325 In I Raacutebano RGozalo and D GarciacuteaminusBellido (eds) Advances in Trilobite ResearchCuadernos del Museo Geominero 9 319ndash325
Shultz JW 2007 A phylogenetic analysis of the arachnid orders based onmorphological characters Zoological Journal of the Linnean Society150 221ndash265
Stein M 2010 A new arthropod from the Early Cambrian of North Greenminusland with a ldquogreat appendagerdquominuslike antennula Zoological Journal ofthe Linnean Society 158 477ndash500
Stuumlrmer W and Bergstroumlm J 1978 The arthropod Cheloniellon from theDevonian Hunsruumlck Shale Palaumlontologische Zeitschrift 52 57ndash81
Sutton MD Briggs DEG Siveter DJ Siveter DJ and Orr PD2002 The arthropod Offacolus kingi (Chelicerata) from the Silurian ofHerefordshire England computer based morphological reconstrucminustions and phylogenetic affinity Proceedings of the Royal Society B Biminusological Sciences 269 1195ndash1203
Suzuki Y and Bergstroumlm J 2008 Respiration in trilobites a reevaluationGFF 130 211ndash229
Vannier J GarciacuteaminusBellido DC Hu XminusX and Chen AminusL 2009 Arthrominuspod visual predators in the early pelagic ecosystem evidence from the
Burgess Shale and Chengjiang biotas Proceedings of the Royal SocietyB Biological Sciences 276 2567ndash2574
Whittington HB 1974 Yohoia Walcott and Plenocaris n gen arthropodsfrom the Burgess Shale Middle Cambrian British Columbia Geologiminuscal Survey of Canada Bulletin 231 1ndash21
Wills MA Briggs DEG Fortey RA Wilkinson M and SneathPHA 1998 An arthropod phylogeny based on fossil and recent taxaIn GD Edgecombe (ed) Arthropod Fossils and Phylogeny 33ndash105Columbia University Press New York
Wilson LA 2006 Food for thought a morphological and taphonomic studyof fossilised digestive systems from Early to Middle Cambrian taxa275 pp Unpublished PhD thesis University of Cambridge Cambridge
Zhang XminusG and Hou X 2007 Gravitational constraints on the burial ofChengjiang fossils Palaios 22 448ndash453
Zhang XminusL Han J Zhang Z Liu H and Shu D 2004 Redescription ofthe Chengjiang arthropod Squamacula clypeata Hou and Bergstroumlm fromthe Lower Cambrian Southminuswest China Palaeontology 47 605ndash617
Zhao Y Zhu M Babcock LE Yuan L Parsley RL Peng J YangX and Wang Y 2005 Kaili Biota a taphonomic window on diversifiminuscation of metazoans from the basal Middle Cambrian Guizhou ChinaActa Geologica Sinica 79 751ndash765
400 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011

margins straight for much of their length or slightly curvedterminally distal spinose projection distinctly stouter at itsbase than the two more proximal projections and more markminusedly tapering along its length Dark irregularminusshaped low reminuslief lump on basal spineminusbearing article of great appendagesof SAM P45168 (eo in Fig 6) and SAM P45908 correminussponding to excretory organ of Bruton and Whittington(1983) Long flagella extending from each of the threespinose projections seen only in SAM P43631 (Figs 23A3) of similar narrow width along its preserved extent anminusnulations indistinguishable
Trunk of 11 segments of even length Maximum height oftrunk relative to length (excluding telson) ca 39 bodydeepest at tergite 4 Trunk tergite boundaries marked as narminusrow ridges in several specimens corresponding to narrowband of overlap between adjacent tergites (ol in Figs 2 3A14A1 B) Some specimens show tergites to be bisected by anarrow but sharp furrow of uncertain identity (eg possiblyimpression of dorsal transverse tendon cf Stein 2010 fig12B C E) (arrows in Figs 3A1 4B)
Exopods with fringe of long setae on cephalic and trunkappendages (Fig 2) setal fringe gradually shortening fromabout the fourth trunk segment Setae project ventrally bymore than 30 height of tergites in deepest part of trunk(Fig 2B) setae flattened narrow and parallelminussided alongmost of their length tapering near their distal ends to termiminusnate as a pointed tip (Figs 2B 3A3 arrows) Endopods exminusposed only in SAM P45167 (Fig 4) two pairs associatedwith cephalon (Fig 4B) those of anterior segments of trunkof similar morphology to those in cephalon all slender lackminusing obvious endites podomere boundaries unrecognisable
Midgut glands occupy a large extent of the dorsoventralheight of the trunk two midgut glands in cephalon of similarsize to those in each of the first seven segments of the trunk(Figs 3A1 5A 6A) all developed as radiating grooves emaminusnating from a globular structure at their ventral edge as in
Leanchoilia superlata (GarciacuteaminusBellido and Collins 2007 pl2 6) Foregut trace in SAM P43631 (Fig 2) indicates that gutcurves ventrally in anterior part of cephalon mouth regionnot preserved Terminal part of hindgut (in trunk segment 11)with closely spaced longitudinal lineations (Fig 2) that apminusparently correspond to folds in the gut wall hindgut termiminusnating at base of telson
Telson most completely preserved in holotype (SAMP43631 Figs 2B 3A2 B) In this specimen the telson likerest of body compressed in lateral aspect Telson shows meminusdian and marginal reinforcing ridges connecting a flat trianminusgular surface this surface appears to be partially broken inSAM P43631 showing underlying surface of other (left) halfof telson Lateral preservation of specimen implies that topcorresponds to an axial structure and bottom to a marginalone 30ndash35 angle in posterior edge of vane connecting thesetwo structures requires that in dorsal aspect telson would apminuspear forked and widening distally Five stout pointed dorminussally curved marginal spines attached to outer posterior edgeon each side of telson
Phylogenetic analysis ofLeanchoiliidaeDetermining the systematic position of Oestokerkus demandsan appraisal of the relationships of Leanchoiliidae as a wholeCladistic analysis including the bestminusknown members ofMegacheira including leanchoiliids has been conducted byCotton and Braddy (2004) and Vannier et al (2009) The taxominusnomic sampling used by Cotton and Braddy (2004) includedseveral megacheiran genera representative chelicerates anumber of taxa representing groups that have been proposedas possible chelicerate relatives (eg Sidneyia AglaspisCheloniellon) in other analyses (Dunlop and Selden 1997Hou and Bergstroumlm 1997) and members of the major lineagesof the Artiopoda sensu Hou and Bergstroumlm (1997) ie trilominusbiteminusallied lamellipedians Because artiopodan interminusrelationminusships have remained quite stable through a series of phylogenminusetic analyses (Edgecombe and Ramskoumlld 1999 Cotton andBraddy 2004 Hendricks and Lieberman 2008 Paterson et al2010) and artiopodans are now often regarded as less closelyrelated to Megacheira than is Chelicerata (Chen et al 2004Cotton and Braddy 2004 Dunlop 2006) we have reduced thesampling of Artiopoda in the present analysis To augment thediversity of ldquoshort great appendagerdquo arthropods Haikoucarisis added based on the description of H ercaiensis by Chen etal (2004) Offacolus (Orr et al 2000 Sutton et al 2002) hasbeen allied to Chelicerata and is included as a terminal Beminuscause the present analysis is principally concerned with relaminustionships of and within Leanchoiliidae sensu Liu et al (2007)we follow Vannier et al (2009) in sampling Alalcomenaeusand Leanchoilia at the species level Within Chelicerata Cotminuston and Braddy (2004) and Hendricks and Lieberman (2008)sampled ancestral states in Xiphosura by coding the Devonian
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 393
Fig 7 Reconstruction of Oestokerkus megacholix gen et sp nov in lateralview Total length and annulations on the flagella and positions of podominusmere boundaries of the frontal appendage are based on Leanchoilia superminuslata (Bruton and Whittington 1983)
synziphosurine Weinbergina Our coding for Xiphosura alsoemploys information from extant xiphosurans because theysupply more reliable information on the homology of the partsof the trunk appendages (Suzuki and Bergstroumlm 2008) and eyeanatomy (Harzsch and Hafner 2006) among other charactersystems than do fossils alone
Our character set extracts characters from the matrix ofCotton and Braddy (2004) that vary in the revised taxonomicsample reformulates some characters to accommodate reminusvised homology concepts and adds new characters especiallyfor leanchoiliids In total 32 characters are included (Table 1)In this analysis the primary homology of the ldquoshort great apminuspendagerdquo and chelicera is coded (as described by character 4)and the taxon sampling broadly constrains megacheirans aschelicerate relatives (as defended by Chen et al 2004 Cottonand Braddy 2004) rather than testing the alternative that theycould be stem group Euarthropoda (Budd 2002 Bergstroumlmand Hou 2005) A test of the two alternatives involves samminuspling of the entire diversity of euarthropods and a range ofnonminuseuarthropod outgroups an exercise beyond the scope ofthis study
Characters1 Mineralised cuticle (0) absent (1) present For definiminus
tion and discussion of this character see Cotton andBraddy (2004 character 30) and Paterson et al (2010character 1)
2 Curvature of anterior margin of cephalic shield (0) rounminusded or straight (1) pointed medially (2) with multiplespinose projections This character codes for variability inthe shape of the cephalic shield in Leanchoiliidae (state 0and 1 sensu GarciacuteaminusBellido and Collins [2007 textminusfigs10 and 9 respectively]) State 2 is added to accommodateOffacolus the median spine of a series that fringes the ceminusphalic margin is not convincingly homologised with themedian projection in leanchoiliids
3 Number of cephalic somitesappendages (0) antenna orfrontal appendage only (1) antenna or frontal appendageand two limb pairs (2) antenna or frontal appendage andthree limb pairs (3) antenna or frontal appendage and fourlimb pairs (4) antenna or frontal appendage and five limbpairs Coding for lamellipedians and Aglaspis is as discussedby Paterson et al (2010 character 2) Coding for Alalcomeminusnaeus cambricus (state 1) follows Briggs and Collins (1999)Leanchoilia illecebrosa (state 2) follows Liu et al (2007)and Leanchoilia persephone and Leanchoilia superlata (state1) follows GarciacuteaminusBellido and Collins (2007)
4 Form of first cephalic appendage (0) antenna (1) anteminusriorly directed raptorial appendage with reduced numberof podomeres each of the terminal podomeres bearing aspinose process on its distal margins a geniculation beminustween spinose podomeres and peduncle This characteris formulated to identify a homology between the frontalappendages of megacheirans and the chelicera of Cheliminus
cerata and state 1 corresponds to the putative apomorminusphic detail defended by Chen et al (2004) and Cottonand Braddy (2004) Vannier et al (2009 character 8)specify another shared property of ldquoshort great appendminusagesrdquo of the megacheiran (ss) type in their characterstate ldquobent at an anglerdquo referring to the geniculation beminustween articles of the peduncle or between the peduncleand distal spiniferous articles This property is sharedwith Chelicerata
5 Number of spineminusbearing articles on frontal appendage (0)three + movable finger (1) two + movable finger (2) one+ movable finger The number of spineminusbearing articles onthe frontal appendage (exclusive of the terminal movablefinger) varies from three in Yohoia (Whittington 1974 pl6 1ndash3 pl 7 3ndash6 pl 9 4ndash6) and Fortiforceps (Hou andBergstroumlm 1997 figs 31C D 32A 33B C E) to two inHaikoucaris (Chen et al 2004 fig 2A B) and Leanminuschoiliidae or only one the last being a chelicera
6 Long spinose projections on distal part of terminal threepodomeres of frontal appendage bearing a flagellum (0)absent (1) present The morphology of the great appendminusage in Leanchoiliidae is coded here We have not codedthe length of the spinose projections and the presence ofa flagellum as two independent characters Cotton andBraddyrsquos (2004) character 6 state 2 (ldquospinesrdquo longer thanthe entire podomere series) and character 8 state 1 (presminusence of an annulated flagellum) are thus grouped as a sinminusgle trait here
394 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Table 1 Characters coded for phylogenetic analysis Polymorphismindicated by ldquo+rdquo partial uncertainty by states separated by ldquordquo Abbreviminusations A Alalcomenaeus L Leanchoilia O Oestokerkus
Squamacula 0000minus minus minus2minus0 0110 00000 10minus minus0 1000 0
Cindarella 0030minus minus minus000 0010 00000 10minus minus0 11121 00
Kuamaia 000minus minus minus001 0011 10000 10minus minus0 11121 00
Kwanyinaspis 00 001 00112 1000 1000 1121 00
Olenoides 1020minus minus minus100 0011 00000 10minus minus0 01121 00
Retifacies 0020minus minus minus000 0010 00000 11000 1020 00
Aglaspis 10(23)0minus minus minus100 00101 00010 01001 0 00
Sidneyia 0000minus minus minus000 01102 00110 01101 00020 00
Emeraldella 0040minus minus minus2minus 0 00103 00110 01001 001(1+2)1 00
A cambricus 00111 10010 00001 00000 01110 01110 10
L illecebrosa 01211 11010 00001 01001 01210 11110 10
L persephone 00111 10010 00001 00000 01210 11110 10
L superlata 01111 11010 00001 01001 01210 11110 10
L hanceyi 00 00 0001 0000 01210
O megacholix 0011 100 00001 00000 01110 1110 0
Jianfengia 0021 0minus000 00005 00000 01000 010(12)0 0
Fortiforceps 00210 0minus00 00004 00000 01100 01230 20
Yohoia 00210 0minus000 00003 00011 01110 00010 (01)1
Haikoucaris 00211 0minus00 00003 00000 01(12)0 1010 0
Cheloniellon 0040minus minus minus100 00100 10010 01300 0030 00
Offacolus 02412 0minus200 0000 00010 01000 13minus minus 01
Xiphosura 00412 0minus100 10000 0010 01000 000minus minus 01
Eurypterida 00412 0minus100 10002 00010 01000 00minus minus minus 01
7 Length of flagellate great appendage (0) flagella termiminusnating at midlength of trunk (1) flagella extending to endof body Coding is applicable to Leanchoiliidae onlydistinguishing some Leanchoilia species that resembleAlalcomenaeus in having relative short great appendages(state 0) versus the longer appendages in L superlata andL illecebrosa
8 Position of lateral facetted eyes (0) ventral (1) dorsalsessile (2) absent Variation in Lamellipedia and severalother taxa coded herein follows Paterson et al (2010character 3)
9 Four or five ventral eyes arranged in a subtransverseband across head shield (0) absent (1) present A bandof ventral eyes is shared by the best known species ofLeanchoiliidae The distinction between three medianeyes in Alalcomenaeus cambricus (Briggs and Collins1999) and two in Leanchoilia illecebrosa (Liu et al2007) and both Leanchoilia superlata and L persephone(GarciacuteaminusBellido and Collins 2007) is uninformative forthe taxon sample here
10 Bulge in exoskeleton accommodating dropminusshaped venminustral eyes (0) absent (1) present This character followsEdgecombe and Ramskoumlld (1999 character 6) and Patminuserson et al (2010 character 5)
11 Dorsal median eyes (0) absent (1) present Coding for anabsence of dorsal median eyes in fossil taxa considers theabsence of either paired ocelli or a median eye tubercleand the character is scored as a chelicerate autapomorphy(see Cotton and Braddy 2004 character 24)
12 Width of doublure on cephalic shield (0) narrow or modminuserately wide (1) entire ventral side of head covered byexpansion of doublure The interpretation of state 1 inSidneyia and Squamacula is discussed by Cotton andBraddy (2004) and Paterson et al (2010 characters 12and 29)
13 Mouth covered by hypostome (0) absent (1) present Noldquoshort great appendagerdquo arthropods have been shown topossess a hypostome and a reconstruction of a smalllabrum in Alalcomenaeus (Briggs and Collins 1999) wasadmittedly conjectural The absence of a conspicuoushypostome contrasts with its ubiquitous presence in laminusmellipedians and several putative chelicerateminusallied taxathat have antennae (such as aglaspidids and Emeralminusdella) Chelicerates have a small epistomeminuslabrum (Bitschand Bitsch 2007) but like Cotton and Braddy (2004character 27 state 4) we code a hypostomal sclerite aslacking Kwanyinaspis has a large hypostome similar tothat found in aglaspidids (photograph of the holotype ofKwanyinaspis maotianshanensis shown to us by JavierOrtegaminusHernaacutendez [personal communication 2010])
14 Tergite articulations (0) extensive overlap of tergites (1)edge to edge articulations This character follows Edgeminuscombe and Ramskoumlld (1999 character 18) and Patersonet al (2010 character 20)
15 Number of trunk segments (0) nine (1) 11 (2) 12 (3)13 (4) 20 (5) 21 or 22 Coding is confined to taxa that do
not possess a pygidium because the precise number ofsomites associated with pygidia can sometimes not bedetermined with confidence Codings for Xiphosura andEurypterida use opisthosomal somite numbers followingShultz (2007 character 95)
16 Exoskeletal boundaries of anterior trunk segments (0)boundaries transverse or reflexed posterolaterally (1) reminusflexed anterolaterally This character follows Edgecombeand Ramskoumlld (1999 character 19) and Paterson et al(2010 character 21)
17 Trunk tergites with serrate lateral margins (0) absent (1)present Liu et al (2007 fig 2C) documented a row ofmarginal spines on the tergites of Leanchoilia illeceminusbrosa a featured shared by Leanchoilia superlata (egGarciacuteaminusBellido and Collins 2007 pl 1 7) but not otherterminals coded here
18 Differentiation in width of trunk tergites (0) trunk withgradational narrowing posteriorly (1) abrupt narrowingat boundary between prothorax and opisthothorax Anabrupt contraction in width between the boundary ofthe prominus and opisthothorax is observed in Emeraldellaand Sidneyia Yohoia has been reconstructed as having apronounced break in the shape of the trunk tergites beminushind segment 10 (Whittington 1974 textminusfig 2) thepoint at which the paratergal folds lose their spinose tipsand appendages are lacking Specimens preserved indorsal aspect (Whittington 1974 pl 10) suggest that thewidth of the opisthothorax is rather less abruptly narminusrowed and we code Yohoia as state 0
19 Appendages of posterior trunk segments (0) all trunktergites bearing appendages (1) one or more posteriortrunk segments lacking appendages Coding correspondsto Cotton and Braddyrsquos (2004) character 43 ldquopostabdominusmen of segments lacking appendagesrdquo
20 Paired tergal carinae (0) absent (1) present Liu et al(2007 265 fig 2F) recorded the presence of tergalcarinae in Leanchoilia illecebrosa shared with L superminuslata (eg Bruton and Whittington 1983 pl 17 106GarciacuteaminusBellido and Collins 2007 pl 1 5)
21 Pygidium (0) absent (1) present Coding of the pygiminusdium in many terminals herein was discussed by Paterminusson et al (2010 character 22)
22 Telson (0) absent (1) present Following Hendricks andLieberman (2008) a telson is a postminussegmental terminalstructure that lacks appendages Dunlop and Selden (1997)considered a telson to be absent in Cheloniellon but weidentify the conical projection posterior to the ultimate legpair and cerci (Stuumlrmer and Bergstroumlm 1978 figs 2 3 1011) as being most consistent with an identity as a telson
23 Telson shape (0) styliform (1) flapminuslike (2) lanceolate(3) short conical The outline of the telson in Haikouminuscaris ercaiensis is incompletely known but the mostcomplete specimen (Chen 2004 fig 472) constrains it aseither state 1 or state 2 A partial uncertainty coding isthus employed A unique state for Cheloniellon (state 3)is based on the interpretation in character 22
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 395
24 Telson fringed with spines (0) spines absent (1) spinespresent Presence of telson spines was introduced as a charminusacter by Hendricks and Lieberman (2008 character 34)
25 Paired flaps lateral to telson (0) absent (1) present Thischaracter describes the postminusventral plates of several taxa(see Edgecombe and Ramskoumlld 1999 character 25) Cotminuston and Braddy (2004 character 54) raised the possibilitythat the cerci of Cheloniellon could be homologous withpostminusventral plates (see also Hou and Bergstroumlm 1997 93)because they are similarly paired structures derived fromthe segment immediately anterior to the telson We havenot coded for this homology because the cerci of Chelominusniellon originate dorsally rather than ventrally
26 Midgut glands with submillimetric lamellae (0) absent(1) present Midgut glands in some stem group arthrominuspods and early crown group arthropods are metamericreniform organs with a characteristic lamellar structure(Butterfield 2002) Their morphology is identical inLeanchoilia and Oestokerkus notably including groovesthat radiate from a ventral median node (Figs 3A1 6A)The gut of Alalcomenaeus cambricus is more tubular(Briggs and Collins 1999 textminusfigs 10 11) and thoughan anterior six segments may display phosphatised latminuseral projections that likely represent gut diverticula (Wilminusson 2006) they have not been shown to have the detailedmorphology of Leanchoiliaminustype midgut glands Butterminusfield (2002 164) cited a figure from Bruton (1981 fig96) in support of ldquolamellarrdquo midgut glands in the cephaminuslon of Sidneyia Like Bruton (1981) in his description ofthe specimen we consider it likely that the structures inquestion are indeed gut diverticula but there is no basisfor identifying them more precisely as the reniformlamellar structures coded under this character
27 Exopod on first postminusantennal first postminusfrontal appendminusage segment (0) absent (1) present This character reminuslates to Cotton and Braddyrsquos (2004) character 3
28 Division of exopod (0) exopod an undivided flap (1)exopod differentiated into proximal and distal lobes (2)small flap with radiating lamellae (3) pediform exopoddivided into ca 6 articles Identification of the exopod inXiphosura follows Suzuki and Bergstroumlm (2008) Thischaracter groups conditions of the exopod shaft coded asindependent characters by Cotton and Braddy (2004)eg state 0 corresponds to their character 16 and state 1to their character 13 Our coding permits transformationbetween any two states without an extra implicit step
29 Exopod setae (0) short spiniform setae (1) long flatminustened tapering distally with slight separation (2)lamellar setae (flattened imbricating) (3) short finesetae A distinction is made between lamellipedian setaewhich are imbricated (state 2) and the pointed more disminustinctly separated and variably spiniform exopod setae ofleanchoiliids (state 1) This distinction is consistent withthe identification of megacheirans as nonminuslamellipedians(eg Hou and Bergstroumlm 1997 Bergstroumlm and Hou2005) Although Cotton and Braddy (2004 character 18)
regarded the form of the setae in Yohoia to be uncertainwe regard specimens figured by Whittington (1974 pl3 3) as conforming to the leanchoiliid state Squamaculadiffers from other taxa in its shorter spiniform setae(state 0) (Zhang et al 2004 textminusfig 1A B) Offacolushas longer spiniform projections on the distal articles ofthe exopod that have been variably identified as eitherspines or setae (Sutton et al 2002 fig 4b) but theirhomology with the setae that splay regularly along themargin of a flapminuslike exopod (eg states 0ndash3) is uncerminustain This and the following character for exopod setaeare left as ldquordquo for Offacolus A homology between thegill lamellae of Xiphosura and exopod setae of lamellipeminusdians has a long history (see Cotton and Braddy 2004character 18) We follow Suzuki and Bergstroumlm (2008)in identifying the xiphosuran gills as outgrowths of thebasis rather than of the exopod this perspective negates ahomology with the exopod setae of Lamellipedia Thepolymorphic coding for Emeraldella reflects the twoseparate states (1+2) displayed by the exopods in thetrunk (see Bruton and Whittington 1983 pl 8 45 47Edgecombe and Ramskoumlld 1999 fig 11)
30 Differentiation of setae along exopod (0) uniform orgradationally variable setae along length of exopod (1)lamellar setae confined to proximal lobe with distal lobebearing slender setae
31 Endopod segmentation (distal to coxa) (0) six or sevenpodomeres and terminal claw or telotarsal spurs (1)eight or more podomeres and terminal claw (2) ca 15podomeres State 1 is formulated to describe the segmenminustation of the leanchoiliid leg (GarciacuteaminusBellido and Collins2007 textminusfig 6 Liu et al 2007 fig 5) in contrast to thereduced number of podomeres in other euarthropods(Boxshall 2004) State 2 characterises Fortiforceps Apartial uncertainty coding is employed for Yohoia inwhich the precise number of podomeres is undeterminedand could be either state 0 or 1 (Whittington 1974 pl 74) but the spacing of the preserved podomere boundariesnegates the possibility of state 2
32 Endopods on anterior postminuscephalic segments (0) welldeveloped (1) rudimentary or lacking Following Cottonand Braddy (2004 character 11) the absence of trunkendopods in Yohoia (Whittington 1974) is coded as a puminustative homologue with the suppression of postminuscephalicendopods in Chelicerata
ResultsAs a convention for depicting the trees networks are rootedon Squamacula which separates artiopodan Lamellipedia(sensu Hou and Bergstroumlm 1997) from a group that includesMegacheira Chelicerata and Xenopoda all members ofwhich possess a telson
Under equal weights 14 shortest cladograms have a lengthof 77 steps with a Consistency Index of 064 and Retention Inminus
396 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
dex of 077 The strict consensus of these trees with supportvalues is shown in Fig 8 Leanchoiliidae is unambiguouslysupported in all 14 cladograms by four apomorphic characterstwo limbminusbearing cephalic segments posterior to the frontalappendage (character 3 state 1) long spinose projections onthe distal part of the terminal three podomeres of the frontalappendage that bear flagella (character 6 state 1) 11 trunksegments (character 15 state 1) and division of the exopodsinto two lobes (character 28 state 1) The arrangement of fouror five eyes in a subtransverse band across the cephalic shieldis unique to Leanchoiliidae (character 9 state 1) but is not anunambiguous autapomorphy of the group because the state isuncertain in close relatives notably Haikoucaris It is apominusmorphic for Leanchoiliidae only under particular optimisaminustions (Delayed Transformation)
The internal relationships of Leanchoiliidae are stableacross all 14 shortest cladograms Leanchoilia is united by thelanceolate telson (character 23 state 2) a unique unambiguminusous autapomorphy Within Leanchoilia a sister group relaminustionship between L superlata and L illecebrosa is well supminusported receiving a Bremer support of 3 and jackknife freminusquency of 86 These two species share four synapomorminusphies a medially pointed anterior cephalic margin (character2 state 1) flagella of the great appendage extending the lengthof the body (character 7 state 1) serrate lateral margins on thetrunk tergites (character 17 state 1) and paired tergal carinae(character 20 state 1) As such a species from Chengjiang (Lillecebrosa) and the Burgess Shale (L superlata) are moreclosely related to each other than are the two Burgess Shalecongeners a relationship that weakens the possibility that theoften cominusoccurring L superlata and L persephone could besexual dimorphs (GarciacuteaminusBellido and Collins 2007) ratherthan separate species The union of Leanchoilia and Oestominuskerkus is unambiguously supported by the shared midgutgland structure of both genera (character 26 state 1)
All shortest cladograms resolve Haikoucaris Yohoiaand Fortiforceps in that order as progressively less closelyrelated to Leanchoiliidae The union of these three taxa withLeanchoiliidae yields a clade that corresponds to Megacheirasensu Hou and Bergstroumlm (1997) apart from the ambiguousplacement of Jianfengia In half of the 14 shortest cladominusgrams Jianfengia is sister to the other megacheirans yieldminusing a monophyletic Megacheira ss In the remaining cladominusgrams Jianfengia is sister to Xenopoda sensu Hou andBergstroumlm (1997) Aglaspis and Chelicerata Haikoucarisand Leanchoiliidae are united by a frontal appendage comminusposed of two spineminusbearing articles in addition to the movminusable finger (character 5 state 1) Yohoia groups with thesetaxa based on spines fringing the telson (character 24 state1) and Fortiforceps is united with them at a broader levelbased on the flapminuslike telson shape (character 23 state 1)When Jianfengia is placed as sister to the other megacheiminusrans the entire clade is supported by the raptorial frontal apminuspendage (character 4 state 1) and the lack of a large hypominusstomal shield (character 13 state 0) In this set of cladominusgrams these features are nonminushomologous between Megaminus
cheira and Chelicerata whereas in some cladograms with thealternative resolution of Jianfengia as closer to cheliceratescharacter 4 is optimised as homologous in Megacheira andChelicerata
Reanalysis of the data with implied weights finds two optiminusmal trees (Fig 9) Relationships within Leanchoiliidae underequal weights are completely stable to reweighting as is theresolution of Haikoucaris as sister to Leanchoiliidae As suchthe character optimisations described above for the leanchoiminusliids are depicted the same in Fig 9 The main difference beminustween equal and implied weights involves the placement ofChelicerata Under equal weights (Fig 8) chelicerates are alminuslied to Xenopoda (here Emeraldella + Sidneyia + Chelominusniellon sensu clade L in Hou and Bergstroumlm 1997 fig 88)and Aglaspidida A clade composed of Aglaspidida Xenominuspoda and Chelicerata was similarly identified by Hou andBergstroumlm (1997 fig 88 their clade K) In our analysis thisgroup consistently finds support from one or more posteriortrunk segments lacking appendages (character 19 state 1) anddepending on alternative internal resolutions it is variably supminusported by dorsallyminuspositioned sessile eyes (character 8 state 1)and the absence of exopods on the first postminusantennalpostminuscheliceral limb (character 27 state 1) Under implied weightshowever chelicerates are more closely allied to Megacheirathan to aglaspidids or xenopodans (Fig 9)
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 397
2
582
76
Chelicera-morpha
Megacheira
62
2
53
50Leanchoiliidae
3
86
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
EurypteridaChelicerata
Fortiforceps
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia hanceyi
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
Fig 8 Strict consensus of 14 shortest cladograms (77 steps) based on charminusacter data in Table 1 Numbers above nodes are Bremer support those beminuslow nodes are jackknife frequencies gt50 Cheliceramorpha correspondsto a clade with the same composition in Cotton and Braddy (2004)
The major groupings in Fig 9 correspond closely to thoseidentified by Cotton and Braddy (2004) Notably the cladeuniting Aglaspis Xenopoda Offacolus Chelicerata Jianminusfengia and other Megacheira corresponds to their ldquoClade 3rdquo(Cotton and Braddy 2004 fig 8) for which they proposedthe name Cheliceramorpha The clade uniting CheloniellonAglaspis Sidneyia and Emeraldella in Fig 9 corresponds totheir ldquoClade 5rdquo and the clade that groups Megacheira andChelicerata is equivalent to their ldquoClade 4rdquo Their analysisdid not identify megacheirans as a monophyletic groupChelicerata was nested within the group and among taxaconsidered here Yohoia was resolved closer to cheliceratesthan to Leanchoiliidae In contrast Yohoia was regarded byBergstroumlm and Hou (2005) as more closely allied to Leanminuschoiliidae than to chelicerates a megacheiran grouping morecompatible with our cladograms The analysis of ldquogreat apminuspendage arthropodsrdquo by Vannier et al (2009) identified a
monophyletic Leanchoiliidae but a polyphyletic Megacheira(to the exclusion of Haikoucaris and Jianfengia) Howevertheir resolution of Fortiforceps and even more so Yohoia asthe closest relatives of Leanchoiliidae is as in our trees (apartfrom the different position of Haikoucaris)
In summary a major controversy in the systematics of theldquoshort great appendagerdquo arthropods remains whether thechelicerates are nested within the group or are instead moreclosely allied to taxa conventionally identified as lamelliminuspedians specifically Xenopoda and Aglaspidida We findthat both of these results are compatible with the availabledata depending on character weighting protocols Noneminustheless the broader megacheiranminuschelicerate issue does notcloud the status of leanchoiliids as a clade or the identificaminustion of their closest relatives (ie Haikoucaris and Yohoia)and the internal relationships of Leanchoiliidae are stableand supported by compelling characters
398 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
Eurypterida
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
2 7 17 20
3
2
1 1 1
26
1
281563
1 1 1 1
5
1
24
1
23 19 20 27 32
28Clade 4
1 0
4 13 11 27 0
154 1 1
3 19 32Clade 3
2 3
2 28
0
3 1 121
1
22
Clade 5
1 0
19 27
1
25
15 28 301 1
2312
1 150 1 3 3
15 16 2923
18
1 1
10 16
1 1
1 8
1
14
3
3
28 30
1 0
5
15
1 1
1
1
22
Fortiforceps
23
1 1 0 1
1 1
Leanchoilia hanceyi
2
2
1
1
Fig 9 Characters optimised on one of two shortest cladograms with imminusplied weights using a concavity constant k = 3 Black squares are nonminushomoplastic changes white squares homoplastic changes all changes areunambiguous Branches without strict support collapsed after optimisingcharacters Numbers for characters (above the branches) and states (belowthe branches) correspond to Table 1 ldquoClades 3ndash5rdquo correspond to groups sonumbered by Cotton and Braddy (2004 fig 8)
AcknowledgementsAccess to the field area has been most kindly allowed by the Buck famminusily We thank our collaborators in Emu Bay Shale research Jim Jago(University of South Australia Mawson Lakes) Jim Gehling andMike Lee (South Australian Museum Adelaide) for tremendous supminusport and advice as well as Mike Gemmell Dennis Rice Allison DaleyAaron Camens John Laurie Glenn Brock Pierre Kruse and Trevorand Jenny Worthy (all South Australian Museum) for their assistancein the field and Natalie Schroeder MaryminusAnne Binnie and RondaAtkinson (all South Australian Museum) for organisational support inthe lab and in the field Javier OrtegaminusHernaacutendez (University of Camminusbridge Cambridge UK) advised us on Kwanyinaspis and the journalrsquosreferees provided useful feedback TNT is available courtesy of theWilli Hennig Society Funding has been provided by an AustralianResearch Council Linkage grant (LP0774959) to the University ofAdelaide with generous financial assistance from Beach Energy Ltdand the South Australian Museum SeaLink provided logistical supminusport DGBrsquos travel to Australia has been supported by grants from theSpanish Research Council (CSIC PA 1001105) the Complutense Uniminusversity of Madrid and the Spanish Ministry of Education and Science(CGL2006minus12245BTE CGL2009minus07073 and RYC2007minus00090)
ReferencesBergstroumlm J and Hou XminusG 2005 Early Palaeozoic lamellipedian arthrominus
pods In S Koenemann and RA Jenner (eds) Crustacea and Arthrominuspod Relationships Crustacean Issues 16 73ndash93
Bitsch J and Bitsch C 2007 The segmental organization of the head reminusgion in Chelicerata a critical review of recent studies and hypothesesActa Zoologica 88 317ndash335
Bousfeld EL 1995 A contribution to the natural classification of Lower andMiddle Cambrian arthropods food gathering and feeding mechanismsAmphipacifica 2 3ndash34
Boxshall GA 2004 The evolution of arthropod limbs Biological Reviews79 253ndash300
Bremer K 1994 Branch support and tree stability Cladistics 10 295ndash304Briggs DEG and Collins D 1999 The arthropod Alalcomenaeus cambricus
Simonetta from the Middle Cambrian Burgess Shale of British ColumbiaPalaeontology 42 953ndash977
Briggs DEG and Robison RA 1984 Exceptionally preserved nonminustrilobite arthropods and anomalocaridids from the Middle Cambrian ofUtah University of Kansas Paleontological Contributions 111 1ndash23
Briggs DEG Lieberman BS Hendricks JR Halgedahl SL andJerrard RD 2008 Middle Cambrian arthropods from Utah Journal ofPaleontology 82 238ndash254
Bruton DL 1981 The arthropod Sidneyia inexpectans Middle CambrianBurgess Shale British Columbia Philosophical Transactions of theRoyal Society of London Series B 295 619ndash656
Bruton DL and Whittington HB 1983 Emeraldella and Leanchoiliatwo arthropods from the Burgess Shale Middle Cambrian British Cominuslumbia Philosophical Transactions of the Royal Society of London Seminusries B 300 553ndash582
Budd GE 2002 A palaeontological solution to the arthropod head probminuslem Nature 417 271ndash275
Butterfield NJ 2002 Leanchoilia guts and the interpretation of threeminusdiminusmensional structures in Burgess Shaleminustype fossils Paleobiology 28155ndash171
Chen JminusY 2004 The Dawn of Animal World [in Chinese] 366 pp JiangScience and Technology Press Nanjing
Chen JminusY Waloszek D and Maas A 2004 A new ldquogreatminusappendagerdquo arminusthropod from the Lower Cambrian of China and homology of cheliceratechelicerae and raptorial anterominusventral appendages Lethaia 37 3ndash20
Cotton TJ and Braddy SJ 2004 The phylogeny of arachnomorph arthrominuspods and the origin of the Chelicerata Transactions of the Royal Societyof Edinburgh Earth Sciences 94 169ndash193
Daley AC Budd GE Caron JminusB Edgecombe GD and Collins D2009 The Burgess Shale anomalocaridid Hurdia and its significancefor early euarthropod evolution Science 323 1597ndash1600
Dunlop JA 2006 New ideas about the euchelicerate stemminuslineage In CDeltshev and P Stoev (eds) European Arachnology 2005 Acta Zoominuslogica Bulgarica (Supplement 1) 9ndash27
Dunlop JA and Selden PA 1997 The early history and phylogeny of thechelicerates In RA Fortey and RH Thomas (eds) Arthropod Relaminustionships Systematics Association Special Volume 55 221ndash235
Edgecombe GD and Ramskoumlld L 1999 Relationships of CambrianArachnata and the systematic position of Trilobita Journal of Paleonminustology 73 263ndash287
Farris JS Albert VA Kaumlllersjouml M Lipcomb D and Kluge AG1996 Parsimony jackknifing outperforms neighborminusjoining Cladistics12 99ndash124
GarciacuteaminusBellido DC and Collins D 2007 Reassessment of the genusLeanchoilia (Arthropoda Arachnomorpha) from the Middle CambrianBurgess Shale British Columbia Canada Palaeontology 50 693ndash709
GarciacuteaminusBellido DC Paterson JR Edgecombe GD Jago JB GehlingJG and Lee MSY 2009 The bivalved arthropods Isoxys and Tuzoiawith softminuspart preservation from the lower Cambrian Emu Bay ShaleLagerstaumltte (Kangaroo Island Australia) Palaeontology 52 1221ndash1241
Goloboff PA 1993 Estimating character weights during tree searchCladistics 9 83ndash91
Goloboff PA Farris JS and Nixon KC 2008 TNT a free program forphylogenetic analysis Cladistics 24 774ndash786
Harzsch S and Hafner G 2006 Evolution of eye development in arthrominuspods phylogenetic aspects Arthropod Structure amp Development 35319ndash340
Hendricks JR and Lieberman BS 2008 New phylogenetic insights intothe Cambrian radiation of arachnomorph arthropods Journal of Paleminusontology 82 585ndash594
Hou XminusG and Bergstroumlm J 1997 Arthropods of the Lower CambrianChengjiang fauna southwest China Fossils and Strata 45 1ndash116
Jago JB Zang W Sun X Brock GA Paterson JR and SkovstedCB 2006 A review of the Cambrian biostratigraphy of South Austraminuslia Palaeoworld 15 406ndash423
Liu Y Hou XminusG and Bergstroumlm J 2007 Chengjiang arthropod Leanminuschoilia illecebrosa (Hou 1987) reconsidered GFF 129 263ndash272
Luo H Hu S Chen L Zhang S and Tao Y 1999 Early CambrianChengjiang fauna from Kunming region China [in Chinese] 129 ppYunnan Science and Technology Press Kunming
Luo H Hu S Zhang S and Tao Y 1997 New occurrence of the EarlyCambrian Chengjiang fauna in Haikou Kunming Yunnan Provinceand study on Trilobitoidea Acta Geologica Sinica 71 122ndash132
Nixon KC 2002 WINCLADA Version 19908 Cornell University IthacaNY
Orr PJ Siveter DJ Briggs DEG Siveter DJ and Sutton MD 2000A new arthropod from the Silurian KonservatminusLagerstaumltte of Hereminusfordshire Proceedings of the Royal Society Series B Biological Sciminusences 267 1497ndash1504
Paterson JR and Brock GA 2007 Early Cambrian trilobites fromAngorichina Flinders Ranges South Australia with a new assemblagefrom the Pararaia bunyerooensis Zone Journal of Paleontology 81116ndash142
Paterson JR and Jago JB 2006 New trilobites from the Lower CambrianEmu Bay Shale Lagerstaumltte at Big Gully Kangaroo Island South Ausminustralia Memoirs of the Association of Australasian Palaeontologists 3243ndash57
Paterson JR Edgecombe GD GarciacuteaminusBellido DC Jago JB andGehling JG 2010 Nektaspid arthropods from the lower CambrianEmu Bay Shale Lagerstaumltte South Australia with a reassessment oflamellipedian relationships Palaeontology 53 377ndash402
Paterson JR Jago JB Gehling JG GarciacuteaminusBellido DC Edgecombe
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 399
GD and Lee MSY 2008 Early Cambrian arthropods from the EmuBay Shale Lagerstaumltte South Australia 319ndash325 In I Raacutebano RGozalo and D GarciacuteaminusBellido (eds) Advances in Trilobite ResearchCuadernos del Museo Geominero 9 319ndash325
Shultz JW 2007 A phylogenetic analysis of the arachnid orders based onmorphological characters Zoological Journal of the Linnean Society150 221ndash265
Stein M 2010 A new arthropod from the Early Cambrian of North Greenminusland with a ldquogreat appendagerdquominuslike antennula Zoological Journal ofthe Linnean Society 158 477ndash500
Stuumlrmer W and Bergstroumlm J 1978 The arthropod Cheloniellon from theDevonian Hunsruumlck Shale Palaumlontologische Zeitschrift 52 57ndash81
Sutton MD Briggs DEG Siveter DJ Siveter DJ and Orr PD2002 The arthropod Offacolus kingi (Chelicerata) from the Silurian ofHerefordshire England computer based morphological reconstrucminustions and phylogenetic affinity Proceedings of the Royal Society B Biminusological Sciences 269 1195ndash1203
Suzuki Y and Bergstroumlm J 2008 Respiration in trilobites a reevaluationGFF 130 211ndash229
Vannier J GarciacuteaminusBellido DC Hu XminusX and Chen AminusL 2009 Arthrominuspod visual predators in the early pelagic ecosystem evidence from the
Burgess Shale and Chengjiang biotas Proceedings of the Royal SocietyB Biological Sciences 276 2567ndash2574
Whittington HB 1974 Yohoia Walcott and Plenocaris n gen arthropodsfrom the Burgess Shale Middle Cambrian British Columbia Geologiminuscal Survey of Canada Bulletin 231 1ndash21
Wills MA Briggs DEG Fortey RA Wilkinson M and SneathPHA 1998 An arthropod phylogeny based on fossil and recent taxaIn GD Edgecombe (ed) Arthropod Fossils and Phylogeny 33ndash105Columbia University Press New York
Wilson LA 2006 Food for thought a morphological and taphonomic studyof fossilised digestive systems from Early to Middle Cambrian taxa275 pp Unpublished PhD thesis University of Cambridge Cambridge
Zhang XminusG and Hou X 2007 Gravitational constraints on the burial ofChengjiang fossils Palaios 22 448ndash453
Zhang XminusL Han J Zhang Z Liu H and Shu D 2004 Redescription ofthe Chengjiang arthropod Squamacula clypeata Hou and Bergstroumlm fromthe Lower Cambrian Southminuswest China Palaeontology 47 605ndash617
Zhao Y Zhu M Babcock LE Yuan L Parsley RL Peng J YangX and Wang Y 2005 Kaili Biota a taphonomic window on diversifiminuscation of metazoans from the basal Middle Cambrian Guizhou ChinaActa Geologica Sinica 79 751ndash765
400 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011

synziphosurine Weinbergina Our coding for Xiphosura alsoemploys information from extant xiphosurans because theysupply more reliable information on the homology of the partsof the trunk appendages (Suzuki and Bergstroumlm 2008) and eyeanatomy (Harzsch and Hafner 2006) among other charactersystems than do fossils alone
Our character set extracts characters from the matrix ofCotton and Braddy (2004) that vary in the revised taxonomicsample reformulates some characters to accommodate reminusvised homology concepts and adds new characters especiallyfor leanchoiliids In total 32 characters are included (Table 1)In this analysis the primary homology of the ldquoshort great apminuspendagerdquo and chelicera is coded (as described by character 4)and the taxon sampling broadly constrains megacheirans aschelicerate relatives (as defended by Chen et al 2004 Cottonand Braddy 2004) rather than testing the alternative that theycould be stem group Euarthropoda (Budd 2002 Bergstroumlmand Hou 2005) A test of the two alternatives involves samminuspling of the entire diversity of euarthropods and a range ofnonminuseuarthropod outgroups an exercise beyond the scope ofthis study
Characters1 Mineralised cuticle (0) absent (1) present For definiminus
tion and discussion of this character see Cotton andBraddy (2004 character 30) and Paterson et al (2010character 1)
2 Curvature of anterior margin of cephalic shield (0) rounminusded or straight (1) pointed medially (2) with multiplespinose projections This character codes for variability inthe shape of the cephalic shield in Leanchoiliidae (state 0and 1 sensu GarciacuteaminusBellido and Collins [2007 textminusfigs10 and 9 respectively]) State 2 is added to accommodateOffacolus the median spine of a series that fringes the ceminusphalic margin is not convincingly homologised with themedian projection in leanchoiliids
3 Number of cephalic somitesappendages (0) antenna orfrontal appendage only (1) antenna or frontal appendageand two limb pairs (2) antenna or frontal appendage andthree limb pairs (3) antenna or frontal appendage and fourlimb pairs (4) antenna or frontal appendage and five limbpairs Coding for lamellipedians and Aglaspis is as discussedby Paterson et al (2010 character 2) Coding for Alalcomeminusnaeus cambricus (state 1) follows Briggs and Collins (1999)Leanchoilia illecebrosa (state 2) follows Liu et al (2007)and Leanchoilia persephone and Leanchoilia superlata (state1) follows GarciacuteaminusBellido and Collins (2007)
4 Form of first cephalic appendage (0) antenna (1) anteminusriorly directed raptorial appendage with reduced numberof podomeres each of the terminal podomeres bearing aspinose process on its distal margins a geniculation beminustween spinose podomeres and peduncle This characteris formulated to identify a homology between the frontalappendages of megacheirans and the chelicera of Cheliminus
cerata and state 1 corresponds to the putative apomorminusphic detail defended by Chen et al (2004) and Cottonand Braddy (2004) Vannier et al (2009 character 8)specify another shared property of ldquoshort great appendminusagesrdquo of the megacheiran (ss) type in their characterstate ldquobent at an anglerdquo referring to the geniculation beminustween articles of the peduncle or between the peduncleand distal spiniferous articles This property is sharedwith Chelicerata
5 Number of spineminusbearing articles on frontal appendage (0)three + movable finger (1) two + movable finger (2) one+ movable finger The number of spineminusbearing articles onthe frontal appendage (exclusive of the terminal movablefinger) varies from three in Yohoia (Whittington 1974 pl6 1ndash3 pl 7 3ndash6 pl 9 4ndash6) and Fortiforceps (Hou andBergstroumlm 1997 figs 31C D 32A 33B C E) to two inHaikoucaris (Chen et al 2004 fig 2A B) and Leanminuschoiliidae or only one the last being a chelicera
6 Long spinose projections on distal part of terminal threepodomeres of frontal appendage bearing a flagellum (0)absent (1) present The morphology of the great appendminusage in Leanchoiliidae is coded here We have not codedthe length of the spinose projections and the presence ofa flagellum as two independent characters Cotton andBraddyrsquos (2004) character 6 state 2 (ldquospinesrdquo longer thanthe entire podomere series) and character 8 state 1 (presminusence of an annulated flagellum) are thus grouped as a sinminusgle trait here
394 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Table 1 Characters coded for phylogenetic analysis Polymorphismindicated by ldquo+rdquo partial uncertainty by states separated by ldquordquo Abbreviminusations A Alalcomenaeus L Leanchoilia O Oestokerkus
Squamacula 0000minus minus minus2minus0 0110 00000 10minus minus0 1000 0
Cindarella 0030minus minus minus000 0010 00000 10minus minus0 11121 00
Kuamaia 000minus minus minus001 0011 10000 10minus minus0 11121 00
Kwanyinaspis 00 001 00112 1000 1000 1121 00
Olenoides 1020minus minus minus100 0011 00000 10minus minus0 01121 00
Retifacies 0020minus minus minus000 0010 00000 11000 1020 00
Aglaspis 10(23)0minus minus minus100 00101 00010 01001 0 00
Sidneyia 0000minus minus minus000 01102 00110 01101 00020 00
Emeraldella 0040minus minus minus2minus 0 00103 00110 01001 001(1+2)1 00
A cambricus 00111 10010 00001 00000 01110 01110 10
L illecebrosa 01211 11010 00001 01001 01210 11110 10
L persephone 00111 10010 00001 00000 01210 11110 10
L superlata 01111 11010 00001 01001 01210 11110 10
L hanceyi 00 00 0001 0000 01210
O megacholix 0011 100 00001 00000 01110 1110 0
Jianfengia 0021 0minus000 00005 00000 01000 010(12)0 0
Fortiforceps 00210 0minus00 00004 00000 01100 01230 20
Yohoia 00210 0minus000 00003 00011 01110 00010 (01)1
Haikoucaris 00211 0minus00 00003 00000 01(12)0 1010 0
Cheloniellon 0040minus minus minus100 00100 10010 01300 0030 00
Offacolus 02412 0minus200 0000 00010 01000 13minus minus 01
Xiphosura 00412 0minus100 10000 0010 01000 000minus minus 01
Eurypterida 00412 0minus100 10002 00010 01000 00minus minus minus 01
7 Length of flagellate great appendage (0) flagella termiminusnating at midlength of trunk (1) flagella extending to endof body Coding is applicable to Leanchoiliidae onlydistinguishing some Leanchoilia species that resembleAlalcomenaeus in having relative short great appendages(state 0) versus the longer appendages in L superlata andL illecebrosa
8 Position of lateral facetted eyes (0) ventral (1) dorsalsessile (2) absent Variation in Lamellipedia and severalother taxa coded herein follows Paterson et al (2010character 3)
9 Four or five ventral eyes arranged in a subtransverseband across head shield (0) absent (1) present A bandof ventral eyes is shared by the best known species ofLeanchoiliidae The distinction between three medianeyes in Alalcomenaeus cambricus (Briggs and Collins1999) and two in Leanchoilia illecebrosa (Liu et al2007) and both Leanchoilia superlata and L persephone(GarciacuteaminusBellido and Collins 2007) is uninformative forthe taxon sample here
10 Bulge in exoskeleton accommodating dropminusshaped venminustral eyes (0) absent (1) present This character followsEdgecombe and Ramskoumlld (1999 character 6) and Patminuserson et al (2010 character 5)
11 Dorsal median eyes (0) absent (1) present Coding for anabsence of dorsal median eyes in fossil taxa considers theabsence of either paired ocelli or a median eye tubercleand the character is scored as a chelicerate autapomorphy(see Cotton and Braddy 2004 character 24)
12 Width of doublure on cephalic shield (0) narrow or modminuserately wide (1) entire ventral side of head covered byexpansion of doublure The interpretation of state 1 inSidneyia and Squamacula is discussed by Cotton andBraddy (2004) and Paterson et al (2010 characters 12and 29)
13 Mouth covered by hypostome (0) absent (1) present Noldquoshort great appendagerdquo arthropods have been shown topossess a hypostome and a reconstruction of a smalllabrum in Alalcomenaeus (Briggs and Collins 1999) wasadmittedly conjectural The absence of a conspicuoushypostome contrasts with its ubiquitous presence in laminusmellipedians and several putative chelicerateminusallied taxathat have antennae (such as aglaspidids and Emeralminusdella) Chelicerates have a small epistomeminuslabrum (Bitschand Bitsch 2007) but like Cotton and Braddy (2004character 27 state 4) we code a hypostomal sclerite aslacking Kwanyinaspis has a large hypostome similar tothat found in aglaspidids (photograph of the holotype ofKwanyinaspis maotianshanensis shown to us by JavierOrtegaminusHernaacutendez [personal communication 2010])
14 Tergite articulations (0) extensive overlap of tergites (1)edge to edge articulations This character follows Edgeminuscombe and Ramskoumlld (1999 character 18) and Patersonet al (2010 character 20)
15 Number of trunk segments (0) nine (1) 11 (2) 12 (3)13 (4) 20 (5) 21 or 22 Coding is confined to taxa that do
not possess a pygidium because the precise number ofsomites associated with pygidia can sometimes not bedetermined with confidence Codings for Xiphosura andEurypterida use opisthosomal somite numbers followingShultz (2007 character 95)
16 Exoskeletal boundaries of anterior trunk segments (0)boundaries transverse or reflexed posterolaterally (1) reminusflexed anterolaterally This character follows Edgecombeand Ramskoumlld (1999 character 19) and Paterson et al(2010 character 21)
17 Trunk tergites with serrate lateral margins (0) absent (1)present Liu et al (2007 fig 2C) documented a row ofmarginal spines on the tergites of Leanchoilia illeceminusbrosa a featured shared by Leanchoilia superlata (egGarciacuteaminusBellido and Collins 2007 pl 1 7) but not otherterminals coded here
18 Differentiation in width of trunk tergites (0) trunk withgradational narrowing posteriorly (1) abrupt narrowingat boundary between prothorax and opisthothorax Anabrupt contraction in width between the boundary ofthe prominus and opisthothorax is observed in Emeraldellaand Sidneyia Yohoia has been reconstructed as having apronounced break in the shape of the trunk tergites beminushind segment 10 (Whittington 1974 textminusfig 2) thepoint at which the paratergal folds lose their spinose tipsand appendages are lacking Specimens preserved indorsal aspect (Whittington 1974 pl 10) suggest that thewidth of the opisthothorax is rather less abruptly narminusrowed and we code Yohoia as state 0
19 Appendages of posterior trunk segments (0) all trunktergites bearing appendages (1) one or more posteriortrunk segments lacking appendages Coding correspondsto Cotton and Braddyrsquos (2004) character 43 ldquopostabdominusmen of segments lacking appendagesrdquo
20 Paired tergal carinae (0) absent (1) present Liu et al(2007 265 fig 2F) recorded the presence of tergalcarinae in Leanchoilia illecebrosa shared with L superminuslata (eg Bruton and Whittington 1983 pl 17 106GarciacuteaminusBellido and Collins 2007 pl 1 5)
21 Pygidium (0) absent (1) present Coding of the pygiminusdium in many terminals herein was discussed by Paterminusson et al (2010 character 22)
22 Telson (0) absent (1) present Following Hendricks andLieberman (2008) a telson is a postminussegmental terminalstructure that lacks appendages Dunlop and Selden (1997)considered a telson to be absent in Cheloniellon but weidentify the conical projection posterior to the ultimate legpair and cerci (Stuumlrmer and Bergstroumlm 1978 figs 2 3 1011) as being most consistent with an identity as a telson
23 Telson shape (0) styliform (1) flapminuslike (2) lanceolate(3) short conical The outline of the telson in Haikouminuscaris ercaiensis is incompletely known but the mostcomplete specimen (Chen 2004 fig 472) constrains it aseither state 1 or state 2 A partial uncertainty coding isthus employed A unique state for Cheloniellon (state 3)is based on the interpretation in character 22
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 395
24 Telson fringed with spines (0) spines absent (1) spinespresent Presence of telson spines was introduced as a charminusacter by Hendricks and Lieberman (2008 character 34)
25 Paired flaps lateral to telson (0) absent (1) present Thischaracter describes the postminusventral plates of several taxa(see Edgecombe and Ramskoumlld 1999 character 25) Cotminuston and Braddy (2004 character 54) raised the possibilitythat the cerci of Cheloniellon could be homologous withpostminusventral plates (see also Hou and Bergstroumlm 1997 93)because they are similarly paired structures derived fromthe segment immediately anterior to the telson We havenot coded for this homology because the cerci of Chelominusniellon originate dorsally rather than ventrally
26 Midgut glands with submillimetric lamellae (0) absent(1) present Midgut glands in some stem group arthrominuspods and early crown group arthropods are metamericreniform organs with a characteristic lamellar structure(Butterfield 2002) Their morphology is identical inLeanchoilia and Oestokerkus notably including groovesthat radiate from a ventral median node (Figs 3A1 6A)The gut of Alalcomenaeus cambricus is more tubular(Briggs and Collins 1999 textminusfigs 10 11) and thoughan anterior six segments may display phosphatised latminuseral projections that likely represent gut diverticula (Wilminusson 2006) they have not been shown to have the detailedmorphology of Leanchoiliaminustype midgut glands Butterminusfield (2002 164) cited a figure from Bruton (1981 fig96) in support of ldquolamellarrdquo midgut glands in the cephaminuslon of Sidneyia Like Bruton (1981) in his description ofthe specimen we consider it likely that the structures inquestion are indeed gut diverticula but there is no basisfor identifying them more precisely as the reniformlamellar structures coded under this character
27 Exopod on first postminusantennal first postminusfrontal appendminusage segment (0) absent (1) present This character reminuslates to Cotton and Braddyrsquos (2004) character 3
28 Division of exopod (0) exopod an undivided flap (1)exopod differentiated into proximal and distal lobes (2)small flap with radiating lamellae (3) pediform exopoddivided into ca 6 articles Identification of the exopod inXiphosura follows Suzuki and Bergstroumlm (2008) Thischaracter groups conditions of the exopod shaft coded asindependent characters by Cotton and Braddy (2004)eg state 0 corresponds to their character 16 and state 1to their character 13 Our coding permits transformationbetween any two states without an extra implicit step
29 Exopod setae (0) short spiniform setae (1) long flatminustened tapering distally with slight separation (2)lamellar setae (flattened imbricating) (3) short finesetae A distinction is made between lamellipedian setaewhich are imbricated (state 2) and the pointed more disminustinctly separated and variably spiniform exopod setae ofleanchoiliids (state 1) This distinction is consistent withthe identification of megacheirans as nonminuslamellipedians(eg Hou and Bergstroumlm 1997 Bergstroumlm and Hou2005) Although Cotton and Braddy (2004 character 18)
regarded the form of the setae in Yohoia to be uncertainwe regard specimens figured by Whittington (1974 pl3 3) as conforming to the leanchoiliid state Squamaculadiffers from other taxa in its shorter spiniform setae(state 0) (Zhang et al 2004 textminusfig 1A B) Offacolushas longer spiniform projections on the distal articles ofthe exopod that have been variably identified as eitherspines or setae (Sutton et al 2002 fig 4b) but theirhomology with the setae that splay regularly along themargin of a flapminuslike exopod (eg states 0ndash3) is uncerminustain This and the following character for exopod setaeare left as ldquordquo for Offacolus A homology between thegill lamellae of Xiphosura and exopod setae of lamellipeminusdians has a long history (see Cotton and Braddy 2004character 18) We follow Suzuki and Bergstroumlm (2008)in identifying the xiphosuran gills as outgrowths of thebasis rather than of the exopod this perspective negates ahomology with the exopod setae of Lamellipedia Thepolymorphic coding for Emeraldella reflects the twoseparate states (1+2) displayed by the exopods in thetrunk (see Bruton and Whittington 1983 pl 8 45 47Edgecombe and Ramskoumlld 1999 fig 11)
30 Differentiation of setae along exopod (0) uniform orgradationally variable setae along length of exopod (1)lamellar setae confined to proximal lobe with distal lobebearing slender setae
31 Endopod segmentation (distal to coxa) (0) six or sevenpodomeres and terminal claw or telotarsal spurs (1)eight or more podomeres and terminal claw (2) ca 15podomeres State 1 is formulated to describe the segmenminustation of the leanchoiliid leg (GarciacuteaminusBellido and Collins2007 textminusfig 6 Liu et al 2007 fig 5) in contrast to thereduced number of podomeres in other euarthropods(Boxshall 2004) State 2 characterises Fortiforceps Apartial uncertainty coding is employed for Yohoia inwhich the precise number of podomeres is undeterminedand could be either state 0 or 1 (Whittington 1974 pl 74) but the spacing of the preserved podomere boundariesnegates the possibility of state 2
32 Endopods on anterior postminuscephalic segments (0) welldeveloped (1) rudimentary or lacking Following Cottonand Braddy (2004 character 11) the absence of trunkendopods in Yohoia (Whittington 1974) is coded as a puminustative homologue with the suppression of postminuscephalicendopods in Chelicerata
ResultsAs a convention for depicting the trees networks are rootedon Squamacula which separates artiopodan Lamellipedia(sensu Hou and Bergstroumlm 1997) from a group that includesMegacheira Chelicerata and Xenopoda all members ofwhich possess a telson
Under equal weights 14 shortest cladograms have a lengthof 77 steps with a Consistency Index of 064 and Retention Inminus
396 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
dex of 077 The strict consensus of these trees with supportvalues is shown in Fig 8 Leanchoiliidae is unambiguouslysupported in all 14 cladograms by four apomorphic characterstwo limbminusbearing cephalic segments posterior to the frontalappendage (character 3 state 1) long spinose projections onthe distal part of the terminal three podomeres of the frontalappendage that bear flagella (character 6 state 1) 11 trunksegments (character 15 state 1) and division of the exopodsinto two lobes (character 28 state 1) The arrangement of fouror five eyes in a subtransverse band across the cephalic shieldis unique to Leanchoiliidae (character 9 state 1) but is not anunambiguous autapomorphy of the group because the state isuncertain in close relatives notably Haikoucaris It is apominusmorphic for Leanchoiliidae only under particular optimisaminustions (Delayed Transformation)
The internal relationships of Leanchoiliidae are stableacross all 14 shortest cladograms Leanchoilia is united by thelanceolate telson (character 23 state 2) a unique unambiguminusous autapomorphy Within Leanchoilia a sister group relaminustionship between L superlata and L illecebrosa is well supminusported receiving a Bremer support of 3 and jackknife freminusquency of 86 These two species share four synapomorminusphies a medially pointed anterior cephalic margin (character2 state 1) flagella of the great appendage extending the lengthof the body (character 7 state 1) serrate lateral margins on thetrunk tergites (character 17 state 1) and paired tergal carinae(character 20 state 1) As such a species from Chengjiang (Lillecebrosa) and the Burgess Shale (L superlata) are moreclosely related to each other than are the two Burgess Shalecongeners a relationship that weakens the possibility that theoften cominusoccurring L superlata and L persephone could besexual dimorphs (GarciacuteaminusBellido and Collins 2007) ratherthan separate species The union of Leanchoilia and Oestominuskerkus is unambiguously supported by the shared midgutgland structure of both genera (character 26 state 1)
All shortest cladograms resolve Haikoucaris Yohoiaand Fortiforceps in that order as progressively less closelyrelated to Leanchoiliidae The union of these three taxa withLeanchoiliidae yields a clade that corresponds to Megacheirasensu Hou and Bergstroumlm (1997) apart from the ambiguousplacement of Jianfengia In half of the 14 shortest cladominusgrams Jianfengia is sister to the other megacheirans yieldminusing a monophyletic Megacheira ss In the remaining cladominusgrams Jianfengia is sister to Xenopoda sensu Hou andBergstroumlm (1997) Aglaspis and Chelicerata Haikoucarisand Leanchoiliidae are united by a frontal appendage comminusposed of two spineminusbearing articles in addition to the movminusable finger (character 5 state 1) Yohoia groups with thesetaxa based on spines fringing the telson (character 24 state1) and Fortiforceps is united with them at a broader levelbased on the flapminuslike telson shape (character 23 state 1)When Jianfengia is placed as sister to the other megacheiminusrans the entire clade is supported by the raptorial frontal apminuspendage (character 4 state 1) and the lack of a large hypominusstomal shield (character 13 state 0) In this set of cladominusgrams these features are nonminushomologous between Megaminus
cheira and Chelicerata whereas in some cladograms with thealternative resolution of Jianfengia as closer to cheliceratescharacter 4 is optimised as homologous in Megacheira andChelicerata
Reanalysis of the data with implied weights finds two optiminusmal trees (Fig 9) Relationships within Leanchoiliidae underequal weights are completely stable to reweighting as is theresolution of Haikoucaris as sister to Leanchoiliidae As suchthe character optimisations described above for the leanchoiminusliids are depicted the same in Fig 9 The main difference beminustween equal and implied weights involves the placement ofChelicerata Under equal weights (Fig 8) chelicerates are alminuslied to Xenopoda (here Emeraldella + Sidneyia + Chelominusniellon sensu clade L in Hou and Bergstroumlm 1997 fig 88)and Aglaspidida A clade composed of Aglaspidida Xenominuspoda and Chelicerata was similarly identified by Hou andBergstroumlm (1997 fig 88 their clade K) In our analysis thisgroup consistently finds support from one or more posteriortrunk segments lacking appendages (character 19 state 1) anddepending on alternative internal resolutions it is variably supminusported by dorsallyminuspositioned sessile eyes (character 8 state 1)and the absence of exopods on the first postminusantennalpostminuscheliceral limb (character 27 state 1) Under implied weightshowever chelicerates are more closely allied to Megacheirathan to aglaspidids or xenopodans (Fig 9)
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 397
2
582
76
Chelicera-morpha
Megacheira
62
2
53
50Leanchoiliidae
3
86
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
EurypteridaChelicerata
Fortiforceps
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia hanceyi
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
Fig 8 Strict consensus of 14 shortest cladograms (77 steps) based on charminusacter data in Table 1 Numbers above nodes are Bremer support those beminuslow nodes are jackknife frequencies gt50 Cheliceramorpha correspondsto a clade with the same composition in Cotton and Braddy (2004)
The major groupings in Fig 9 correspond closely to thoseidentified by Cotton and Braddy (2004) Notably the cladeuniting Aglaspis Xenopoda Offacolus Chelicerata Jianminusfengia and other Megacheira corresponds to their ldquoClade 3rdquo(Cotton and Braddy 2004 fig 8) for which they proposedthe name Cheliceramorpha The clade uniting CheloniellonAglaspis Sidneyia and Emeraldella in Fig 9 corresponds totheir ldquoClade 5rdquo and the clade that groups Megacheira andChelicerata is equivalent to their ldquoClade 4rdquo Their analysisdid not identify megacheirans as a monophyletic groupChelicerata was nested within the group and among taxaconsidered here Yohoia was resolved closer to cheliceratesthan to Leanchoiliidae In contrast Yohoia was regarded byBergstroumlm and Hou (2005) as more closely allied to Leanminuschoiliidae than to chelicerates a megacheiran grouping morecompatible with our cladograms The analysis of ldquogreat apminuspendage arthropodsrdquo by Vannier et al (2009) identified a
monophyletic Leanchoiliidae but a polyphyletic Megacheira(to the exclusion of Haikoucaris and Jianfengia) Howevertheir resolution of Fortiforceps and even more so Yohoia asthe closest relatives of Leanchoiliidae is as in our trees (apartfrom the different position of Haikoucaris)
In summary a major controversy in the systematics of theldquoshort great appendagerdquo arthropods remains whether thechelicerates are nested within the group or are instead moreclosely allied to taxa conventionally identified as lamelliminuspedians specifically Xenopoda and Aglaspidida We findthat both of these results are compatible with the availabledata depending on character weighting protocols Noneminustheless the broader megacheiranminuschelicerate issue does notcloud the status of leanchoiliids as a clade or the identificaminustion of their closest relatives (ie Haikoucaris and Yohoia)and the internal relationships of Leanchoiliidae are stableand supported by compelling characters
398 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
Eurypterida
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
2 7 17 20
3
2
1 1 1
26
1
281563
1 1 1 1
5
1
24
1
23 19 20 27 32
28Clade 4
1 0
4 13 11 27 0
154 1 1
3 19 32Clade 3
2 3
2 28
0
3 1 121
1
22
Clade 5
1 0
19 27
1
25
15 28 301 1
2312
1 150 1 3 3
15 16 2923
18
1 1
10 16
1 1
1 8
1
14
3
3
28 30
1 0
5
15
1 1
1
1
22
Fortiforceps
23
1 1 0 1
1 1
Leanchoilia hanceyi
2
2
1
1
Fig 9 Characters optimised on one of two shortest cladograms with imminusplied weights using a concavity constant k = 3 Black squares are nonminushomoplastic changes white squares homoplastic changes all changes areunambiguous Branches without strict support collapsed after optimisingcharacters Numbers for characters (above the branches) and states (belowthe branches) correspond to Table 1 ldquoClades 3ndash5rdquo correspond to groups sonumbered by Cotton and Braddy (2004 fig 8)
AcknowledgementsAccess to the field area has been most kindly allowed by the Buck famminusily We thank our collaborators in Emu Bay Shale research Jim Jago(University of South Australia Mawson Lakes) Jim Gehling andMike Lee (South Australian Museum Adelaide) for tremendous supminusport and advice as well as Mike Gemmell Dennis Rice Allison DaleyAaron Camens John Laurie Glenn Brock Pierre Kruse and Trevorand Jenny Worthy (all South Australian Museum) for their assistancein the field and Natalie Schroeder MaryminusAnne Binnie and RondaAtkinson (all South Australian Museum) for organisational support inthe lab and in the field Javier OrtegaminusHernaacutendez (University of Camminusbridge Cambridge UK) advised us on Kwanyinaspis and the journalrsquosreferees provided useful feedback TNT is available courtesy of theWilli Hennig Society Funding has been provided by an AustralianResearch Council Linkage grant (LP0774959) to the University ofAdelaide with generous financial assistance from Beach Energy Ltdand the South Australian Museum SeaLink provided logistical supminusport DGBrsquos travel to Australia has been supported by grants from theSpanish Research Council (CSIC PA 1001105) the Complutense Uniminusversity of Madrid and the Spanish Ministry of Education and Science(CGL2006minus12245BTE CGL2009minus07073 and RYC2007minus00090)
ReferencesBergstroumlm J and Hou XminusG 2005 Early Palaeozoic lamellipedian arthrominus
pods In S Koenemann and RA Jenner (eds) Crustacea and Arthrominuspod Relationships Crustacean Issues 16 73ndash93
Bitsch J and Bitsch C 2007 The segmental organization of the head reminusgion in Chelicerata a critical review of recent studies and hypothesesActa Zoologica 88 317ndash335
Bousfeld EL 1995 A contribution to the natural classification of Lower andMiddle Cambrian arthropods food gathering and feeding mechanismsAmphipacifica 2 3ndash34
Boxshall GA 2004 The evolution of arthropod limbs Biological Reviews79 253ndash300
Bremer K 1994 Branch support and tree stability Cladistics 10 295ndash304Briggs DEG and Collins D 1999 The arthropod Alalcomenaeus cambricus
Simonetta from the Middle Cambrian Burgess Shale of British ColumbiaPalaeontology 42 953ndash977
Briggs DEG and Robison RA 1984 Exceptionally preserved nonminustrilobite arthropods and anomalocaridids from the Middle Cambrian ofUtah University of Kansas Paleontological Contributions 111 1ndash23
Briggs DEG Lieberman BS Hendricks JR Halgedahl SL andJerrard RD 2008 Middle Cambrian arthropods from Utah Journal ofPaleontology 82 238ndash254
Bruton DL 1981 The arthropod Sidneyia inexpectans Middle CambrianBurgess Shale British Columbia Philosophical Transactions of theRoyal Society of London Series B 295 619ndash656
Bruton DL and Whittington HB 1983 Emeraldella and Leanchoiliatwo arthropods from the Burgess Shale Middle Cambrian British Cominuslumbia Philosophical Transactions of the Royal Society of London Seminusries B 300 553ndash582
Budd GE 2002 A palaeontological solution to the arthropod head probminuslem Nature 417 271ndash275
Butterfield NJ 2002 Leanchoilia guts and the interpretation of threeminusdiminusmensional structures in Burgess Shaleminustype fossils Paleobiology 28155ndash171
Chen JminusY 2004 The Dawn of Animal World [in Chinese] 366 pp JiangScience and Technology Press Nanjing
Chen JminusY Waloszek D and Maas A 2004 A new ldquogreatminusappendagerdquo arminusthropod from the Lower Cambrian of China and homology of cheliceratechelicerae and raptorial anterominusventral appendages Lethaia 37 3ndash20
Cotton TJ and Braddy SJ 2004 The phylogeny of arachnomorph arthrominuspods and the origin of the Chelicerata Transactions of the Royal Societyof Edinburgh Earth Sciences 94 169ndash193
Daley AC Budd GE Caron JminusB Edgecombe GD and Collins D2009 The Burgess Shale anomalocaridid Hurdia and its significancefor early euarthropod evolution Science 323 1597ndash1600
Dunlop JA 2006 New ideas about the euchelicerate stemminuslineage In CDeltshev and P Stoev (eds) European Arachnology 2005 Acta Zoominuslogica Bulgarica (Supplement 1) 9ndash27
Dunlop JA and Selden PA 1997 The early history and phylogeny of thechelicerates In RA Fortey and RH Thomas (eds) Arthropod Relaminustionships Systematics Association Special Volume 55 221ndash235
Edgecombe GD and Ramskoumlld L 1999 Relationships of CambrianArachnata and the systematic position of Trilobita Journal of Paleonminustology 73 263ndash287
Farris JS Albert VA Kaumlllersjouml M Lipcomb D and Kluge AG1996 Parsimony jackknifing outperforms neighborminusjoining Cladistics12 99ndash124
GarciacuteaminusBellido DC and Collins D 2007 Reassessment of the genusLeanchoilia (Arthropoda Arachnomorpha) from the Middle CambrianBurgess Shale British Columbia Canada Palaeontology 50 693ndash709
GarciacuteaminusBellido DC Paterson JR Edgecombe GD Jago JB GehlingJG and Lee MSY 2009 The bivalved arthropods Isoxys and Tuzoiawith softminuspart preservation from the lower Cambrian Emu Bay ShaleLagerstaumltte (Kangaroo Island Australia) Palaeontology 52 1221ndash1241
Goloboff PA 1993 Estimating character weights during tree searchCladistics 9 83ndash91
Goloboff PA Farris JS and Nixon KC 2008 TNT a free program forphylogenetic analysis Cladistics 24 774ndash786
Harzsch S and Hafner G 2006 Evolution of eye development in arthrominuspods phylogenetic aspects Arthropod Structure amp Development 35319ndash340
Hendricks JR and Lieberman BS 2008 New phylogenetic insights intothe Cambrian radiation of arachnomorph arthropods Journal of Paleminusontology 82 585ndash594
Hou XminusG and Bergstroumlm J 1997 Arthropods of the Lower CambrianChengjiang fauna southwest China Fossils and Strata 45 1ndash116
Jago JB Zang W Sun X Brock GA Paterson JR and SkovstedCB 2006 A review of the Cambrian biostratigraphy of South Austraminuslia Palaeoworld 15 406ndash423
Liu Y Hou XminusG and Bergstroumlm J 2007 Chengjiang arthropod Leanminuschoilia illecebrosa (Hou 1987) reconsidered GFF 129 263ndash272
Luo H Hu S Chen L Zhang S and Tao Y 1999 Early CambrianChengjiang fauna from Kunming region China [in Chinese] 129 ppYunnan Science and Technology Press Kunming
Luo H Hu S Zhang S and Tao Y 1997 New occurrence of the EarlyCambrian Chengjiang fauna in Haikou Kunming Yunnan Provinceand study on Trilobitoidea Acta Geologica Sinica 71 122ndash132
Nixon KC 2002 WINCLADA Version 19908 Cornell University IthacaNY
Orr PJ Siveter DJ Briggs DEG Siveter DJ and Sutton MD 2000A new arthropod from the Silurian KonservatminusLagerstaumltte of Hereminusfordshire Proceedings of the Royal Society Series B Biological Sciminusences 267 1497ndash1504
Paterson JR and Brock GA 2007 Early Cambrian trilobites fromAngorichina Flinders Ranges South Australia with a new assemblagefrom the Pararaia bunyerooensis Zone Journal of Paleontology 81116ndash142
Paterson JR and Jago JB 2006 New trilobites from the Lower CambrianEmu Bay Shale Lagerstaumltte at Big Gully Kangaroo Island South Ausminustralia Memoirs of the Association of Australasian Palaeontologists 3243ndash57
Paterson JR Edgecombe GD GarciacuteaminusBellido DC Jago JB andGehling JG 2010 Nektaspid arthropods from the lower CambrianEmu Bay Shale Lagerstaumltte South Australia with a reassessment oflamellipedian relationships Palaeontology 53 377ndash402
Paterson JR Jago JB Gehling JG GarciacuteaminusBellido DC Edgecombe
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 399
GD and Lee MSY 2008 Early Cambrian arthropods from the EmuBay Shale Lagerstaumltte South Australia 319ndash325 In I Raacutebano RGozalo and D GarciacuteaminusBellido (eds) Advances in Trilobite ResearchCuadernos del Museo Geominero 9 319ndash325
Shultz JW 2007 A phylogenetic analysis of the arachnid orders based onmorphological characters Zoological Journal of the Linnean Society150 221ndash265
Stein M 2010 A new arthropod from the Early Cambrian of North Greenminusland with a ldquogreat appendagerdquominuslike antennula Zoological Journal ofthe Linnean Society 158 477ndash500
Stuumlrmer W and Bergstroumlm J 1978 The arthropod Cheloniellon from theDevonian Hunsruumlck Shale Palaumlontologische Zeitschrift 52 57ndash81
Sutton MD Briggs DEG Siveter DJ Siveter DJ and Orr PD2002 The arthropod Offacolus kingi (Chelicerata) from the Silurian ofHerefordshire England computer based morphological reconstrucminustions and phylogenetic affinity Proceedings of the Royal Society B Biminusological Sciences 269 1195ndash1203
Suzuki Y and Bergstroumlm J 2008 Respiration in trilobites a reevaluationGFF 130 211ndash229
Vannier J GarciacuteaminusBellido DC Hu XminusX and Chen AminusL 2009 Arthrominuspod visual predators in the early pelagic ecosystem evidence from the
Burgess Shale and Chengjiang biotas Proceedings of the Royal SocietyB Biological Sciences 276 2567ndash2574
Whittington HB 1974 Yohoia Walcott and Plenocaris n gen arthropodsfrom the Burgess Shale Middle Cambrian British Columbia Geologiminuscal Survey of Canada Bulletin 231 1ndash21
Wills MA Briggs DEG Fortey RA Wilkinson M and SneathPHA 1998 An arthropod phylogeny based on fossil and recent taxaIn GD Edgecombe (ed) Arthropod Fossils and Phylogeny 33ndash105Columbia University Press New York
Wilson LA 2006 Food for thought a morphological and taphonomic studyof fossilised digestive systems from Early to Middle Cambrian taxa275 pp Unpublished PhD thesis University of Cambridge Cambridge
Zhang XminusG and Hou X 2007 Gravitational constraints on the burial ofChengjiang fossils Palaios 22 448ndash453
Zhang XminusL Han J Zhang Z Liu H and Shu D 2004 Redescription ofthe Chengjiang arthropod Squamacula clypeata Hou and Bergstroumlm fromthe Lower Cambrian Southminuswest China Palaeontology 47 605ndash617
Zhao Y Zhu M Babcock LE Yuan L Parsley RL Peng J YangX and Wang Y 2005 Kaili Biota a taphonomic window on diversifiminuscation of metazoans from the basal Middle Cambrian Guizhou ChinaActa Geologica Sinica 79 751ndash765
400 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011

7 Length of flagellate great appendage (0) flagella termiminusnating at midlength of trunk (1) flagella extending to endof body Coding is applicable to Leanchoiliidae onlydistinguishing some Leanchoilia species that resembleAlalcomenaeus in having relative short great appendages(state 0) versus the longer appendages in L superlata andL illecebrosa
8 Position of lateral facetted eyes (0) ventral (1) dorsalsessile (2) absent Variation in Lamellipedia and severalother taxa coded herein follows Paterson et al (2010character 3)
9 Four or five ventral eyes arranged in a subtransverseband across head shield (0) absent (1) present A bandof ventral eyes is shared by the best known species ofLeanchoiliidae The distinction between three medianeyes in Alalcomenaeus cambricus (Briggs and Collins1999) and two in Leanchoilia illecebrosa (Liu et al2007) and both Leanchoilia superlata and L persephone(GarciacuteaminusBellido and Collins 2007) is uninformative forthe taxon sample here
10 Bulge in exoskeleton accommodating dropminusshaped venminustral eyes (0) absent (1) present This character followsEdgecombe and Ramskoumlld (1999 character 6) and Patminuserson et al (2010 character 5)
11 Dorsal median eyes (0) absent (1) present Coding for anabsence of dorsal median eyes in fossil taxa considers theabsence of either paired ocelli or a median eye tubercleand the character is scored as a chelicerate autapomorphy(see Cotton and Braddy 2004 character 24)
12 Width of doublure on cephalic shield (0) narrow or modminuserately wide (1) entire ventral side of head covered byexpansion of doublure The interpretation of state 1 inSidneyia and Squamacula is discussed by Cotton andBraddy (2004) and Paterson et al (2010 characters 12and 29)
13 Mouth covered by hypostome (0) absent (1) present Noldquoshort great appendagerdquo arthropods have been shown topossess a hypostome and a reconstruction of a smalllabrum in Alalcomenaeus (Briggs and Collins 1999) wasadmittedly conjectural The absence of a conspicuoushypostome contrasts with its ubiquitous presence in laminusmellipedians and several putative chelicerateminusallied taxathat have antennae (such as aglaspidids and Emeralminusdella) Chelicerates have a small epistomeminuslabrum (Bitschand Bitsch 2007) but like Cotton and Braddy (2004character 27 state 4) we code a hypostomal sclerite aslacking Kwanyinaspis has a large hypostome similar tothat found in aglaspidids (photograph of the holotype ofKwanyinaspis maotianshanensis shown to us by JavierOrtegaminusHernaacutendez [personal communication 2010])
14 Tergite articulations (0) extensive overlap of tergites (1)edge to edge articulations This character follows Edgeminuscombe and Ramskoumlld (1999 character 18) and Patersonet al (2010 character 20)
15 Number of trunk segments (0) nine (1) 11 (2) 12 (3)13 (4) 20 (5) 21 or 22 Coding is confined to taxa that do
not possess a pygidium because the precise number ofsomites associated with pygidia can sometimes not bedetermined with confidence Codings for Xiphosura andEurypterida use opisthosomal somite numbers followingShultz (2007 character 95)
16 Exoskeletal boundaries of anterior trunk segments (0)boundaries transverse or reflexed posterolaterally (1) reminusflexed anterolaterally This character follows Edgecombeand Ramskoumlld (1999 character 19) and Paterson et al(2010 character 21)
17 Trunk tergites with serrate lateral margins (0) absent (1)present Liu et al (2007 fig 2C) documented a row ofmarginal spines on the tergites of Leanchoilia illeceminusbrosa a featured shared by Leanchoilia superlata (egGarciacuteaminusBellido and Collins 2007 pl 1 7) but not otherterminals coded here
18 Differentiation in width of trunk tergites (0) trunk withgradational narrowing posteriorly (1) abrupt narrowingat boundary between prothorax and opisthothorax Anabrupt contraction in width between the boundary ofthe prominus and opisthothorax is observed in Emeraldellaand Sidneyia Yohoia has been reconstructed as having apronounced break in the shape of the trunk tergites beminushind segment 10 (Whittington 1974 textminusfig 2) thepoint at which the paratergal folds lose their spinose tipsand appendages are lacking Specimens preserved indorsal aspect (Whittington 1974 pl 10) suggest that thewidth of the opisthothorax is rather less abruptly narminusrowed and we code Yohoia as state 0
19 Appendages of posterior trunk segments (0) all trunktergites bearing appendages (1) one or more posteriortrunk segments lacking appendages Coding correspondsto Cotton and Braddyrsquos (2004) character 43 ldquopostabdominusmen of segments lacking appendagesrdquo
20 Paired tergal carinae (0) absent (1) present Liu et al(2007 265 fig 2F) recorded the presence of tergalcarinae in Leanchoilia illecebrosa shared with L superminuslata (eg Bruton and Whittington 1983 pl 17 106GarciacuteaminusBellido and Collins 2007 pl 1 5)
21 Pygidium (0) absent (1) present Coding of the pygiminusdium in many terminals herein was discussed by Paterminusson et al (2010 character 22)
22 Telson (0) absent (1) present Following Hendricks andLieberman (2008) a telson is a postminussegmental terminalstructure that lacks appendages Dunlop and Selden (1997)considered a telson to be absent in Cheloniellon but weidentify the conical projection posterior to the ultimate legpair and cerci (Stuumlrmer and Bergstroumlm 1978 figs 2 3 1011) as being most consistent with an identity as a telson
23 Telson shape (0) styliform (1) flapminuslike (2) lanceolate(3) short conical The outline of the telson in Haikouminuscaris ercaiensis is incompletely known but the mostcomplete specimen (Chen 2004 fig 472) constrains it aseither state 1 or state 2 A partial uncertainty coding isthus employed A unique state for Cheloniellon (state 3)is based on the interpretation in character 22
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 395
24 Telson fringed with spines (0) spines absent (1) spinespresent Presence of telson spines was introduced as a charminusacter by Hendricks and Lieberman (2008 character 34)
25 Paired flaps lateral to telson (0) absent (1) present Thischaracter describes the postminusventral plates of several taxa(see Edgecombe and Ramskoumlld 1999 character 25) Cotminuston and Braddy (2004 character 54) raised the possibilitythat the cerci of Cheloniellon could be homologous withpostminusventral plates (see also Hou and Bergstroumlm 1997 93)because they are similarly paired structures derived fromthe segment immediately anterior to the telson We havenot coded for this homology because the cerci of Chelominusniellon originate dorsally rather than ventrally
26 Midgut glands with submillimetric lamellae (0) absent(1) present Midgut glands in some stem group arthrominuspods and early crown group arthropods are metamericreniform organs with a characteristic lamellar structure(Butterfield 2002) Their morphology is identical inLeanchoilia and Oestokerkus notably including groovesthat radiate from a ventral median node (Figs 3A1 6A)The gut of Alalcomenaeus cambricus is more tubular(Briggs and Collins 1999 textminusfigs 10 11) and thoughan anterior six segments may display phosphatised latminuseral projections that likely represent gut diverticula (Wilminusson 2006) they have not been shown to have the detailedmorphology of Leanchoiliaminustype midgut glands Butterminusfield (2002 164) cited a figure from Bruton (1981 fig96) in support of ldquolamellarrdquo midgut glands in the cephaminuslon of Sidneyia Like Bruton (1981) in his description ofthe specimen we consider it likely that the structures inquestion are indeed gut diverticula but there is no basisfor identifying them more precisely as the reniformlamellar structures coded under this character
27 Exopod on first postminusantennal first postminusfrontal appendminusage segment (0) absent (1) present This character reminuslates to Cotton and Braddyrsquos (2004) character 3
28 Division of exopod (0) exopod an undivided flap (1)exopod differentiated into proximal and distal lobes (2)small flap with radiating lamellae (3) pediform exopoddivided into ca 6 articles Identification of the exopod inXiphosura follows Suzuki and Bergstroumlm (2008) Thischaracter groups conditions of the exopod shaft coded asindependent characters by Cotton and Braddy (2004)eg state 0 corresponds to their character 16 and state 1to their character 13 Our coding permits transformationbetween any two states without an extra implicit step
29 Exopod setae (0) short spiniform setae (1) long flatminustened tapering distally with slight separation (2)lamellar setae (flattened imbricating) (3) short finesetae A distinction is made between lamellipedian setaewhich are imbricated (state 2) and the pointed more disminustinctly separated and variably spiniform exopod setae ofleanchoiliids (state 1) This distinction is consistent withthe identification of megacheirans as nonminuslamellipedians(eg Hou and Bergstroumlm 1997 Bergstroumlm and Hou2005) Although Cotton and Braddy (2004 character 18)
regarded the form of the setae in Yohoia to be uncertainwe regard specimens figured by Whittington (1974 pl3 3) as conforming to the leanchoiliid state Squamaculadiffers from other taxa in its shorter spiniform setae(state 0) (Zhang et al 2004 textminusfig 1A B) Offacolushas longer spiniform projections on the distal articles ofthe exopod that have been variably identified as eitherspines or setae (Sutton et al 2002 fig 4b) but theirhomology with the setae that splay regularly along themargin of a flapminuslike exopod (eg states 0ndash3) is uncerminustain This and the following character for exopod setaeare left as ldquordquo for Offacolus A homology between thegill lamellae of Xiphosura and exopod setae of lamellipeminusdians has a long history (see Cotton and Braddy 2004character 18) We follow Suzuki and Bergstroumlm (2008)in identifying the xiphosuran gills as outgrowths of thebasis rather than of the exopod this perspective negates ahomology with the exopod setae of Lamellipedia Thepolymorphic coding for Emeraldella reflects the twoseparate states (1+2) displayed by the exopods in thetrunk (see Bruton and Whittington 1983 pl 8 45 47Edgecombe and Ramskoumlld 1999 fig 11)
30 Differentiation of setae along exopod (0) uniform orgradationally variable setae along length of exopod (1)lamellar setae confined to proximal lobe with distal lobebearing slender setae
31 Endopod segmentation (distal to coxa) (0) six or sevenpodomeres and terminal claw or telotarsal spurs (1)eight or more podomeres and terminal claw (2) ca 15podomeres State 1 is formulated to describe the segmenminustation of the leanchoiliid leg (GarciacuteaminusBellido and Collins2007 textminusfig 6 Liu et al 2007 fig 5) in contrast to thereduced number of podomeres in other euarthropods(Boxshall 2004) State 2 characterises Fortiforceps Apartial uncertainty coding is employed for Yohoia inwhich the precise number of podomeres is undeterminedand could be either state 0 or 1 (Whittington 1974 pl 74) but the spacing of the preserved podomere boundariesnegates the possibility of state 2
32 Endopods on anterior postminuscephalic segments (0) welldeveloped (1) rudimentary or lacking Following Cottonand Braddy (2004 character 11) the absence of trunkendopods in Yohoia (Whittington 1974) is coded as a puminustative homologue with the suppression of postminuscephalicendopods in Chelicerata
ResultsAs a convention for depicting the trees networks are rootedon Squamacula which separates artiopodan Lamellipedia(sensu Hou and Bergstroumlm 1997) from a group that includesMegacheira Chelicerata and Xenopoda all members ofwhich possess a telson
Under equal weights 14 shortest cladograms have a lengthof 77 steps with a Consistency Index of 064 and Retention Inminus
396 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
dex of 077 The strict consensus of these trees with supportvalues is shown in Fig 8 Leanchoiliidae is unambiguouslysupported in all 14 cladograms by four apomorphic characterstwo limbminusbearing cephalic segments posterior to the frontalappendage (character 3 state 1) long spinose projections onthe distal part of the terminal three podomeres of the frontalappendage that bear flagella (character 6 state 1) 11 trunksegments (character 15 state 1) and division of the exopodsinto two lobes (character 28 state 1) The arrangement of fouror five eyes in a subtransverse band across the cephalic shieldis unique to Leanchoiliidae (character 9 state 1) but is not anunambiguous autapomorphy of the group because the state isuncertain in close relatives notably Haikoucaris It is apominusmorphic for Leanchoiliidae only under particular optimisaminustions (Delayed Transformation)
The internal relationships of Leanchoiliidae are stableacross all 14 shortest cladograms Leanchoilia is united by thelanceolate telson (character 23 state 2) a unique unambiguminusous autapomorphy Within Leanchoilia a sister group relaminustionship between L superlata and L illecebrosa is well supminusported receiving a Bremer support of 3 and jackknife freminusquency of 86 These two species share four synapomorminusphies a medially pointed anterior cephalic margin (character2 state 1) flagella of the great appendage extending the lengthof the body (character 7 state 1) serrate lateral margins on thetrunk tergites (character 17 state 1) and paired tergal carinae(character 20 state 1) As such a species from Chengjiang (Lillecebrosa) and the Burgess Shale (L superlata) are moreclosely related to each other than are the two Burgess Shalecongeners a relationship that weakens the possibility that theoften cominusoccurring L superlata and L persephone could besexual dimorphs (GarciacuteaminusBellido and Collins 2007) ratherthan separate species The union of Leanchoilia and Oestominuskerkus is unambiguously supported by the shared midgutgland structure of both genera (character 26 state 1)
All shortest cladograms resolve Haikoucaris Yohoiaand Fortiforceps in that order as progressively less closelyrelated to Leanchoiliidae The union of these three taxa withLeanchoiliidae yields a clade that corresponds to Megacheirasensu Hou and Bergstroumlm (1997) apart from the ambiguousplacement of Jianfengia In half of the 14 shortest cladominusgrams Jianfengia is sister to the other megacheirans yieldminusing a monophyletic Megacheira ss In the remaining cladominusgrams Jianfengia is sister to Xenopoda sensu Hou andBergstroumlm (1997) Aglaspis and Chelicerata Haikoucarisand Leanchoiliidae are united by a frontal appendage comminusposed of two spineminusbearing articles in addition to the movminusable finger (character 5 state 1) Yohoia groups with thesetaxa based on spines fringing the telson (character 24 state1) and Fortiforceps is united with them at a broader levelbased on the flapminuslike telson shape (character 23 state 1)When Jianfengia is placed as sister to the other megacheiminusrans the entire clade is supported by the raptorial frontal apminuspendage (character 4 state 1) and the lack of a large hypominusstomal shield (character 13 state 0) In this set of cladominusgrams these features are nonminushomologous between Megaminus
cheira and Chelicerata whereas in some cladograms with thealternative resolution of Jianfengia as closer to cheliceratescharacter 4 is optimised as homologous in Megacheira andChelicerata
Reanalysis of the data with implied weights finds two optiminusmal trees (Fig 9) Relationships within Leanchoiliidae underequal weights are completely stable to reweighting as is theresolution of Haikoucaris as sister to Leanchoiliidae As suchthe character optimisations described above for the leanchoiminusliids are depicted the same in Fig 9 The main difference beminustween equal and implied weights involves the placement ofChelicerata Under equal weights (Fig 8) chelicerates are alminuslied to Xenopoda (here Emeraldella + Sidneyia + Chelominusniellon sensu clade L in Hou and Bergstroumlm 1997 fig 88)and Aglaspidida A clade composed of Aglaspidida Xenominuspoda and Chelicerata was similarly identified by Hou andBergstroumlm (1997 fig 88 their clade K) In our analysis thisgroup consistently finds support from one or more posteriortrunk segments lacking appendages (character 19 state 1) anddepending on alternative internal resolutions it is variably supminusported by dorsallyminuspositioned sessile eyes (character 8 state 1)and the absence of exopods on the first postminusantennalpostminuscheliceral limb (character 27 state 1) Under implied weightshowever chelicerates are more closely allied to Megacheirathan to aglaspidids or xenopodans (Fig 9)
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 397
2
582
76
Chelicera-morpha
Megacheira
62
2
53
50Leanchoiliidae
3
86
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
EurypteridaChelicerata
Fortiforceps
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia hanceyi
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
Fig 8 Strict consensus of 14 shortest cladograms (77 steps) based on charminusacter data in Table 1 Numbers above nodes are Bremer support those beminuslow nodes are jackknife frequencies gt50 Cheliceramorpha correspondsto a clade with the same composition in Cotton and Braddy (2004)
The major groupings in Fig 9 correspond closely to thoseidentified by Cotton and Braddy (2004) Notably the cladeuniting Aglaspis Xenopoda Offacolus Chelicerata Jianminusfengia and other Megacheira corresponds to their ldquoClade 3rdquo(Cotton and Braddy 2004 fig 8) for which they proposedthe name Cheliceramorpha The clade uniting CheloniellonAglaspis Sidneyia and Emeraldella in Fig 9 corresponds totheir ldquoClade 5rdquo and the clade that groups Megacheira andChelicerata is equivalent to their ldquoClade 4rdquo Their analysisdid not identify megacheirans as a monophyletic groupChelicerata was nested within the group and among taxaconsidered here Yohoia was resolved closer to cheliceratesthan to Leanchoiliidae In contrast Yohoia was regarded byBergstroumlm and Hou (2005) as more closely allied to Leanminuschoiliidae than to chelicerates a megacheiran grouping morecompatible with our cladograms The analysis of ldquogreat apminuspendage arthropodsrdquo by Vannier et al (2009) identified a
monophyletic Leanchoiliidae but a polyphyletic Megacheira(to the exclusion of Haikoucaris and Jianfengia) Howevertheir resolution of Fortiforceps and even more so Yohoia asthe closest relatives of Leanchoiliidae is as in our trees (apartfrom the different position of Haikoucaris)
In summary a major controversy in the systematics of theldquoshort great appendagerdquo arthropods remains whether thechelicerates are nested within the group or are instead moreclosely allied to taxa conventionally identified as lamelliminuspedians specifically Xenopoda and Aglaspidida We findthat both of these results are compatible with the availabledata depending on character weighting protocols Noneminustheless the broader megacheiranminuschelicerate issue does notcloud the status of leanchoiliids as a clade or the identificaminustion of their closest relatives (ie Haikoucaris and Yohoia)and the internal relationships of Leanchoiliidae are stableand supported by compelling characters
398 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
Eurypterida
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
2 7 17 20
3
2
1 1 1
26
1
281563
1 1 1 1
5
1
24
1
23 19 20 27 32
28Clade 4
1 0
4 13 11 27 0
154 1 1
3 19 32Clade 3
2 3
2 28
0
3 1 121
1
22
Clade 5
1 0
19 27
1
25
15 28 301 1
2312
1 150 1 3 3
15 16 2923
18
1 1
10 16
1 1
1 8
1
14
3
3
28 30
1 0
5
15
1 1
1
1
22
Fortiforceps
23
1 1 0 1
1 1
Leanchoilia hanceyi
2
2
1
1
Fig 9 Characters optimised on one of two shortest cladograms with imminusplied weights using a concavity constant k = 3 Black squares are nonminushomoplastic changes white squares homoplastic changes all changes areunambiguous Branches without strict support collapsed after optimisingcharacters Numbers for characters (above the branches) and states (belowthe branches) correspond to Table 1 ldquoClades 3ndash5rdquo correspond to groups sonumbered by Cotton and Braddy (2004 fig 8)
AcknowledgementsAccess to the field area has been most kindly allowed by the Buck famminusily We thank our collaborators in Emu Bay Shale research Jim Jago(University of South Australia Mawson Lakes) Jim Gehling andMike Lee (South Australian Museum Adelaide) for tremendous supminusport and advice as well as Mike Gemmell Dennis Rice Allison DaleyAaron Camens John Laurie Glenn Brock Pierre Kruse and Trevorand Jenny Worthy (all South Australian Museum) for their assistancein the field and Natalie Schroeder MaryminusAnne Binnie and RondaAtkinson (all South Australian Museum) for organisational support inthe lab and in the field Javier OrtegaminusHernaacutendez (University of Camminusbridge Cambridge UK) advised us on Kwanyinaspis and the journalrsquosreferees provided useful feedback TNT is available courtesy of theWilli Hennig Society Funding has been provided by an AustralianResearch Council Linkage grant (LP0774959) to the University ofAdelaide with generous financial assistance from Beach Energy Ltdand the South Australian Museum SeaLink provided logistical supminusport DGBrsquos travel to Australia has been supported by grants from theSpanish Research Council (CSIC PA 1001105) the Complutense Uniminusversity of Madrid and the Spanish Ministry of Education and Science(CGL2006minus12245BTE CGL2009minus07073 and RYC2007minus00090)
ReferencesBergstroumlm J and Hou XminusG 2005 Early Palaeozoic lamellipedian arthrominus
pods In S Koenemann and RA Jenner (eds) Crustacea and Arthrominuspod Relationships Crustacean Issues 16 73ndash93
Bitsch J and Bitsch C 2007 The segmental organization of the head reminusgion in Chelicerata a critical review of recent studies and hypothesesActa Zoologica 88 317ndash335
Bousfeld EL 1995 A contribution to the natural classification of Lower andMiddle Cambrian arthropods food gathering and feeding mechanismsAmphipacifica 2 3ndash34
Boxshall GA 2004 The evolution of arthropod limbs Biological Reviews79 253ndash300
Bremer K 1994 Branch support and tree stability Cladistics 10 295ndash304Briggs DEG and Collins D 1999 The arthropod Alalcomenaeus cambricus
Simonetta from the Middle Cambrian Burgess Shale of British ColumbiaPalaeontology 42 953ndash977
Briggs DEG and Robison RA 1984 Exceptionally preserved nonminustrilobite arthropods and anomalocaridids from the Middle Cambrian ofUtah University of Kansas Paleontological Contributions 111 1ndash23
Briggs DEG Lieberman BS Hendricks JR Halgedahl SL andJerrard RD 2008 Middle Cambrian arthropods from Utah Journal ofPaleontology 82 238ndash254
Bruton DL 1981 The arthropod Sidneyia inexpectans Middle CambrianBurgess Shale British Columbia Philosophical Transactions of theRoyal Society of London Series B 295 619ndash656
Bruton DL and Whittington HB 1983 Emeraldella and Leanchoiliatwo arthropods from the Burgess Shale Middle Cambrian British Cominuslumbia Philosophical Transactions of the Royal Society of London Seminusries B 300 553ndash582
Budd GE 2002 A palaeontological solution to the arthropod head probminuslem Nature 417 271ndash275
Butterfield NJ 2002 Leanchoilia guts and the interpretation of threeminusdiminusmensional structures in Burgess Shaleminustype fossils Paleobiology 28155ndash171
Chen JminusY 2004 The Dawn of Animal World [in Chinese] 366 pp JiangScience and Technology Press Nanjing
Chen JminusY Waloszek D and Maas A 2004 A new ldquogreatminusappendagerdquo arminusthropod from the Lower Cambrian of China and homology of cheliceratechelicerae and raptorial anterominusventral appendages Lethaia 37 3ndash20
Cotton TJ and Braddy SJ 2004 The phylogeny of arachnomorph arthrominuspods and the origin of the Chelicerata Transactions of the Royal Societyof Edinburgh Earth Sciences 94 169ndash193
Daley AC Budd GE Caron JminusB Edgecombe GD and Collins D2009 The Burgess Shale anomalocaridid Hurdia and its significancefor early euarthropod evolution Science 323 1597ndash1600
Dunlop JA 2006 New ideas about the euchelicerate stemminuslineage In CDeltshev and P Stoev (eds) European Arachnology 2005 Acta Zoominuslogica Bulgarica (Supplement 1) 9ndash27
Dunlop JA and Selden PA 1997 The early history and phylogeny of thechelicerates In RA Fortey and RH Thomas (eds) Arthropod Relaminustionships Systematics Association Special Volume 55 221ndash235
Edgecombe GD and Ramskoumlld L 1999 Relationships of CambrianArachnata and the systematic position of Trilobita Journal of Paleonminustology 73 263ndash287
Farris JS Albert VA Kaumlllersjouml M Lipcomb D and Kluge AG1996 Parsimony jackknifing outperforms neighborminusjoining Cladistics12 99ndash124
GarciacuteaminusBellido DC and Collins D 2007 Reassessment of the genusLeanchoilia (Arthropoda Arachnomorpha) from the Middle CambrianBurgess Shale British Columbia Canada Palaeontology 50 693ndash709
GarciacuteaminusBellido DC Paterson JR Edgecombe GD Jago JB GehlingJG and Lee MSY 2009 The bivalved arthropods Isoxys and Tuzoiawith softminuspart preservation from the lower Cambrian Emu Bay ShaleLagerstaumltte (Kangaroo Island Australia) Palaeontology 52 1221ndash1241
Goloboff PA 1993 Estimating character weights during tree searchCladistics 9 83ndash91
Goloboff PA Farris JS and Nixon KC 2008 TNT a free program forphylogenetic analysis Cladistics 24 774ndash786
Harzsch S and Hafner G 2006 Evolution of eye development in arthrominuspods phylogenetic aspects Arthropod Structure amp Development 35319ndash340
Hendricks JR and Lieberman BS 2008 New phylogenetic insights intothe Cambrian radiation of arachnomorph arthropods Journal of Paleminusontology 82 585ndash594
Hou XminusG and Bergstroumlm J 1997 Arthropods of the Lower CambrianChengjiang fauna southwest China Fossils and Strata 45 1ndash116
Jago JB Zang W Sun X Brock GA Paterson JR and SkovstedCB 2006 A review of the Cambrian biostratigraphy of South Austraminuslia Palaeoworld 15 406ndash423
Liu Y Hou XminusG and Bergstroumlm J 2007 Chengjiang arthropod Leanminuschoilia illecebrosa (Hou 1987) reconsidered GFF 129 263ndash272
Luo H Hu S Chen L Zhang S and Tao Y 1999 Early CambrianChengjiang fauna from Kunming region China [in Chinese] 129 ppYunnan Science and Technology Press Kunming
Luo H Hu S Zhang S and Tao Y 1997 New occurrence of the EarlyCambrian Chengjiang fauna in Haikou Kunming Yunnan Provinceand study on Trilobitoidea Acta Geologica Sinica 71 122ndash132
Nixon KC 2002 WINCLADA Version 19908 Cornell University IthacaNY
Orr PJ Siveter DJ Briggs DEG Siveter DJ and Sutton MD 2000A new arthropod from the Silurian KonservatminusLagerstaumltte of Hereminusfordshire Proceedings of the Royal Society Series B Biological Sciminusences 267 1497ndash1504
Paterson JR and Brock GA 2007 Early Cambrian trilobites fromAngorichina Flinders Ranges South Australia with a new assemblagefrom the Pararaia bunyerooensis Zone Journal of Paleontology 81116ndash142
Paterson JR and Jago JB 2006 New trilobites from the Lower CambrianEmu Bay Shale Lagerstaumltte at Big Gully Kangaroo Island South Ausminustralia Memoirs of the Association of Australasian Palaeontologists 3243ndash57
Paterson JR Edgecombe GD GarciacuteaminusBellido DC Jago JB andGehling JG 2010 Nektaspid arthropods from the lower CambrianEmu Bay Shale Lagerstaumltte South Australia with a reassessment oflamellipedian relationships Palaeontology 53 377ndash402
Paterson JR Jago JB Gehling JG GarciacuteaminusBellido DC Edgecombe
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 399
GD and Lee MSY 2008 Early Cambrian arthropods from the EmuBay Shale Lagerstaumltte South Australia 319ndash325 In I Raacutebano RGozalo and D GarciacuteaminusBellido (eds) Advances in Trilobite ResearchCuadernos del Museo Geominero 9 319ndash325
Shultz JW 2007 A phylogenetic analysis of the arachnid orders based onmorphological characters Zoological Journal of the Linnean Society150 221ndash265
Stein M 2010 A new arthropod from the Early Cambrian of North Greenminusland with a ldquogreat appendagerdquominuslike antennula Zoological Journal ofthe Linnean Society 158 477ndash500
Stuumlrmer W and Bergstroumlm J 1978 The arthropod Cheloniellon from theDevonian Hunsruumlck Shale Palaumlontologische Zeitschrift 52 57ndash81
Sutton MD Briggs DEG Siveter DJ Siveter DJ and Orr PD2002 The arthropod Offacolus kingi (Chelicerata) from the Silurian ofHerefordshire England computer based morphological reconstrucminustions and phylogenetic affinity Proceedings of the Royal Society B Biminusological Sciences 269 1195ndash1203
Suzuki Y and Bergstroumlm J 2008 Respiration in trilobites a reevaluationGFF 130 211ndash229
Vannier J GarciacuteaminusBellido DC Hu XminusX and Chen AminusL 2009 Arthrominuspod visual predators in the early pelagic ecosystem evidence from the
Burgess Shale and Chengjiang biotas Proceedings of the Royal SocietyB Biological Sciences 276 2567ndash2574
Whittington HB 1974 Yohoia Walcott and Plenocaris n gen arthropodsfrom the Burgess Shale Middle Cambrian British Columbia Geologiminuscal Survey of Canada Bulletin 231 1ndash21
Wills MA Briggs DEG Fortey RA Wilkinson M and SneathPHA 1998 An arthropod phylogeny based on fossil and recent taxaIn GD Edgecombe (ed) Arthropod Fossils and Phylogeny 33ndash105Columbia University Press New York
Wilson LA 2006 Food for thought a morphological and taphonomic studyof fossilised digestive systems from Early to Middle Cambrian taxa275 pp Unpublished PhD thesis University of Cambridge Cambridge
Zhang XminusG and Hou X 2007 Gravitational constraints on the burial ofChengjiang fossils Palaios 22 448ndash453
Zhang XminusL Han J Zhang Z Liu H and Shu D 2004 Redescription ofthe Chengjiang arthropod Squamacula clypeata Hou and Bergstroumlm fromthe Lower Cambrian Southminuswest China Palaeontology 47 605ndash617
Zhao Y Zhu M Babcock LE Yuan L Parsley RL Peng J YangX and Wang Y 2005 Kaili Biota a taphonomic window on diversifiminuscation of metazoans from the basal Middle Cambrian Guizhou ChinaActa Geologica Sinica 79 751ndash765
400 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011

24 Telson fringed with spines (0) spines absent (1) spinespresent Presence of telson spines was introduced as a charminusacter by Hendricks and Lieberman (2008 character 34)
25 Paired flaps lateral to telson (0) absent (1) present Thischaracter describes the postminusventral plates of several taxa(see Edgecombe and Ramskoumlld 1999 character 25) Cotminuston and Braddy (2004 character 54) raised the possibilitythat the cerci of Cheloniellon could be homologous withpostminusventral plates (see also Hou and Bergstroumlm 1997 93)because they are similarly paired structures derived fromthe segment immediately anterior to the telson We havenot coded for this homology because the cerci of Chelominusniellon originate dorsally rather than ventrally
26 Midgut glands with submillimetric lamellae (0) absent(1) present Midgut glands in some stem group arthrominuspods and early crown group arthropods are metamericreniform organs with a characteristic lamellar structure(Butterfield 2002) Their morphology is identical inLeanchoilia and Oestokerkus notably including groovesthat radiate from a ventral median node (Figs 3A1 6A)The gut of Alalcomenaeus cambricus is more tubular(Briggs and Collins 1999 textminusfigs 10 11) and thoughan anterior six segments may display phosphatised latminuseral projections that likely represent gut diverticula (Wilminusson 2006) they have not been shown to have the detailedmorphology of Leanchoiliaminustype midgut glands Butterminusfield (2002 164) cited a figure from Bruton (1981 fig96) in support of ldquolamellarrdquo midgut glands in the cephaminuslon of Sidneyia Like Bruton (1981) in his description ofthe specimen we consider it likely that the structures inquestion are indeed gut diverticula but there is no basisfor identifying them more precisely as the reniformlamellar structures coded under this character
27 Exopod on first postminusantennal first postminusfrontal appendminusage segment (0) absent (1) present This character reminuslates to Cotton and Braddyrsquos (2004) character 3
28 Division of exopod (0) exopod an undivided flap (1)exopod differentiated into proximal and distal lobes (2)small flap with radiating lamellae (3) pediform exopoddivided into ca 6 articles Identification of the exopod inXiphosura follows Suzuki and Bergstroumlm (2008) Thischaracter groups conditions of the exopod shaft coded asindependent characters by Cotton and Braddy (2004)eg state 0 corresponds to their character 16 and state 1to their character 13 Our coding permits transformationbetween any two states without an extra implicit step
29 Exopod setae (0) short spiniform setae (1) long flatminustened tapering distally with slight separation (2)lamellar setae (flattened imbricating) (3) short finesetae A distinction is made between lamellipedian setaewhich are imbricated (state 2) and the pointed more disminustinctly separated and variably spiniform exopod setae ofleanchoiliids (state 1) This distinction is consistent withthe identification of megacheirans as nonminuslamellipedians(eg Hou and Bergstroumlm 1997 Bergstroumlm and Hou2005) Although Cotton and Braddy (2004 character 18)
regarded the form of the setae in Yohoia to be uncertainwe regard specimens figured by Whittington (1974 pl3 3) as conforming to the leanchoiliid state Squamaculadiffers from other taxa in its shorter spiniform setae(state 0) (Zhang et al 2004 textminusfig 1A B) Offacolushas longer spiniform projections on the distal articles ofthe exopod that have been variably identified as eitherspines or setae (Sutton et al 2002 fig 4b) but theirhomology with the setae that splay regularly along themargin of a flapminuslike exopod (eg states 0ndash3) is uncerminustain This and the following character for exopod setaeare left as ldquordquo for Offacolus A homology between thegill lamellae of Xiphosura and exopod setae of lamellipeminusdians has a long history (see Cotton and Braddy 2004character 18) We follow Suzuki and Bergstroumlm (2008)in identifying the xiphosuran gills as outgrowths of thebasis rather than of the exopod this perspective negates ahomology with the exopod setae of Lamellipedia Thepolymorphic coding for Emeraldella reflects the twoseparate states (1+2) displayed by the exopods in thetrunk (see Bruton and Whittington 1983 pl 8 45 47Edgecombe and Ramskoumlld 1999 fig 11)
30 Differentiation of setae along exopod (0) uniform orgradationally variable setae along length of exopod (1)lamellar setae confined to proximal lobe with distal lobebearing slender setae
31 Endopod segmentation (distal to coxa) (0) six or sevenpodomeres and terminal claw or telotarsal spurs (1)eight or more podomeres and terminal claw (2) ca 15podomeres State 1 is formulated to describe the segmenminustation of the leanchoiliid leg (GarciacuteaminusBellido and Collins2007 textminusfig 6 Liu et al 2007 fig 5) in contrast to thereduced number of podomeres in other euarthropods(Boxshall 2004) State 2 characterises Fortiforceps Apartial uncertainty coding is employed for Yohoia inwhich the precise number of podomeres is undeterminedand could be either state 0 or 1 (Whittington 1974 pl 74) but the spacing of the preserved podomere boundariesnegates the possibility of state 2
32 Endopods on anterior postminuscephalic segments (0) welldeveloped (1) rudimentary or lacking Following Cottonand Braddy (2004 character 11) the absence of trunkendopods in Yohoia (Whittington 1974) is coded as a puminustative homologue with the suppression of postminuscephalicendopods in Chelicerata
ResultsAs a convention for depicting the trees networks are rootedon Squamacula which separates artiopodan Lamellipedia(sensu Hou and Bergstroumlm 1997) from a group that includesMegacheira Chelicerata and Xenopoda all members ofwhich possess a telson
Under equal weights 14 shortest cladograms have a lengthof 77 steps with a Consistency Index of 064 and Retention Inminus
396 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
dex of 077 The strict consensus of these trees with supportvalues is shown in Fig 8 Leanchoiliidae is unambiguouslysupported in all 14 cladograms by four apomorphic characterstwo limbminusbearing cephalic segments posterior to the frontalappendage (character 3 state 1) long spinose projections onthe distal part of the terminal three podomeres of the frontalappendage that bear flagella (character 6 state 1) 11 trunksegments (character 15 state 1) and division of the exopodsinto two lobes (character 28 state 1) The arrangement of fouror five eyes in a subtransverse band across the cephalic shieldis unique to Leanchoiliidae (character 9 state 1) but is not anunambiguous autapomorphy of the group because the state isuncertain in close relatives notably Haikoucaris It is apominusmorphic for Leanchoiliidae only under particular optimisaminustions (Delayed Transformation)
The internal relationships of Leanchoiliidae are stableacross all 14 shortest cladograms Leanchoilia is united by thelanceolate telson (character 23 state 2) a unique unambiguminusous autapomorphy Within Leanchoilia a sister group relaminustionship between L superlata and L illecebrosa is well supminusported receiving a Bremer support of 3 and jackknife freminusquency of 86 These two species share four synapomorminusphies a medially pointed anterior cephalic margin (character2 state 1) flagella of the great appendage extending the lengthof the body (character 7 state 1) serrate lateral margins on thetrunk tergites (character 17 state 1) and paired tergal carinae(character 20 state 1) As such a species from Chengjiang (Lillecebrosa) and the Burgess Shale (L superlata) are moreclosely related to each other than are the two Burgess Shalecongeners a relationship that weakens the possibility that theoften cominusoccurring L superlata and L persephone could besexual dimorphs (GarciacuteaminusBellido and Collins 2007) ratherthan separate species The union of Leanchoilia and Oestominuskerkus is unambiguously supported by the shared midgutgland structure of both genera (character 26 state 1)
All shortest cladograms resolve Haikoucaris Yohoiaand Fortiforceps in that order as progressively less closelyrelated to Leanchoiliidae The union of these three taxa withLeanchoiliidae yields a clade that corresponds to Megacheirasensu Hou and Bergstroumlm (1997) apart from the ambiguousplacement of Jianfengia In half of the 14 shortest cladominusgrams Jianfengia is sister to the other megacheirans yieldminusing a monophyletic Megacheira ss In the remaining cladominusgrams Jianfengia is sister to Xenopoda sensu Hou andBergstroumlm (1997) Aglaspis and Chelicerata Haikoucarisand Leanchoiliidae are united by a frontal appendage comminusposed of two spineminusbearing articles in addition to the movminusable finger (character 5 state 1) Yohoia groups with thesetaxa based on spines fringing the telson (character 24 state1) and Fortiforceps is united with them at a broader levelbased on the flapminuslike telson shape (character 23 state 1)When Jianfengia is placed as sister to the other megacheiminusrans the entire clade is supported by the raptorial frontal apminuspendage (character 4 state 1) and the lack of a large hypominusstomal shield (character 13 state 0) In this set of cladominusgrams these features are nonminushomologous between Megaminus
cheira and Chelicerata whereas in some cladograms with thealternative resolution of Jianfengia as closer to cheliceratescharacter 4 is optimised as homologous in Megacheira andChelicerata
Reanalysis of the data with implied weights finds two optiminusmal trees (Fig 9) Relationships within Leanchoiliidae underequal weights are completely stable to reweighting as is theresolution of Haikoucaris as sister to Leanchoiliidae As suchthe character optimisations described above for the leanchoiminusliids are depicted the same in Fig 9 The main difference beminustween equal and implied weights involves the placement ofChelicerata Under equal weights (Fig 8) chelicerates are alminuslied to Xenopoda (here Emeraldella + Sidneyia + Chelominusniellon sensu clade L in Hou and Bergstroumlm 1997 fig 88)and Aglaspidida A clade composed of Aglaspidida Xenominuspoda and Chelicerata was similarly identified by Hou andBergstroumlm (1997 fig 88 their clade K) In our analysis thisgroup consistently finds support from one or more posteriortrunk segments lacking appendages (character 19 state 1) anddepending on alternative internal resolutions it is variably supminusported by dorsallyminuspositioned sessile eyes (character 8 state 1)and the absence of exopods on the first postminusantennalpostminuscheliceral limb (character 27 state 1) Under implied weightshowever chelicerates are more closely allied to Megacheirathan to aglaspidids or xenopodans (Fig 9)
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 397
2
582
76
Chelicera-morpha
Megacheira
62
2
53
50Leanchoiliidae
3
86
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
EurypteridaChelicerata
Fortiforceps
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia hanceyi
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
Fig 8 Strict consensus of 14 shortest cladograms (77 steps) based on charminusacter data in Table 1 Numbers above nodes are Bremer support those beminuslow nodes are jackknife frequencies gt50 Cheliceramorpha correspondsto a clade with the same composition in Cotton and Braddy (2004)
The major groupings in Fig 9 correspond closely to thoseidentified by Cotton and Braddy (2004) Notably the cladeuniting Aglaspis Xenopoda Offacolus Chelicerata Jianminusfengia and other Megacheira corresponds to their ldquoClade 3rdquo(Cotton and Braddy 2004 fig 8) for which they proposedthe name Cheliceramorpha The clade uniting CheloniellonAglaspis Sidneyia and Emeraldella in Fig 9 corresponds totheir ldquoClade 5rdquo and the clade that groups Megacheira andChelicerata is equivalent to their ldquoClade 4rdquo Their analysisdid not identify megacheirans as a monophyletic groupChelicerata was nested within the group and among taxaconsidered here Yohoia was resolved closer to cheliceratesthan to Leanchoiliidae In contrast Yohoia was regarded byBergstroumlm and Hou (2005) as more closely allied to Leanminuschoiliidae than to chelicerates a megacheiran grouping morecompatible with our cladograms The analysis of ldquogreat apminuspendage arthropodsrdquo by Vannier et al (2009) identified a
monophyletic Leanchoiliidae but a polyphyletic Megacheira(to the exclusion of Haikoucaris and Jianfengia) Howevertheir resolution of Fortiforceps and even more so Yohoia asthe closest relatives of Leanchoiliidae is as in our trees (apartfrom the different position of Haikoucaris)
In summary a major controversy in the systematics of theldquoshort great appendagerdquo arthropods remains whether thechelicerates are nested within the group or are instead moreclosely allied to taxa conventionally identified as lamelliminuspedians specifically Xenopoda and Aglaspidida We findthat both of these results are compatible with the availabledata depending on character weighting protocols Noneminustheless the broader megacheiranminuschelicerate issue does notcloud the status of leanchoiliids as a clade or the identificaminustion of their closest relatives (ie Haikoucaris and Yohoia)and the internal relationships of Leanchoiliidae are stableand supported by compelling characters
398 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
Eurypterida
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
2 7 17 20
3
2
1 1 1
26
1
281563
1 1 1 1
5
1
24
1
23 19 20 27 32
28Clade 4
1 0
4 13 11 27 0
154 1 1
3 19 32Clade 3
2 3
2 28
0
3 1 121
1
22
Clade 5
1 0
19 27
1
25
15 28 301 1
2312
1 150 1 3 3
15 16 2923
18
1 1
10 16
1 1
1 8
1
14
3
3
28 30
1 0
5
15
1 1
1
1
22
Fortiforceps
23
1 1 0 1
1 1
Leanchoilia hanceyi
2
2
1
1
Fig 9 Characters optimised on one of two shortest cladograms with imminusplied weights using a concavity constant k = 3 Black squares are nonminushomoplastic changes white squares homoplastic changes all changes areunambiguous Branches without strict support collapsed after optimisingcharacters Numbers for characters (above the branches) and states (belowthe branches) correspond to Table 1 ldquoClades 3ndash5rdquo correspond to groups sonumbered by Cotton and Braddy (2004 fig 8)
AcknowledgementsAccess to the field area has been most kindly allowed by the Buck famminusily We thank our collaborators in Emu Bay Shale research Jim Jago(University of South Australia Mawson Lakes) Jim Gehling andMike Lee (South Australian Museum Adelaide) for tremendous supminusport and advice as well as Mike Gemmell Dennis Rice Allison DaleyAaron Camens John Laurie Glenn Brock Pierre Kruse and Trevorand Jenny Worthy (all South Australian Museum) for their assistancein the field and Natalie Schroeder MaryminusAnne Binnie and RondaAtkinson (all South Australian Museum) for organisational support inthe lab and in the field Javier OrtegaminusHernaacutendez (University of Camminusbridge Cambridge UK) advised us on Kwanyinaspis and the journalrsquosreferees provided useful feedback TNT is available courtesy of theWilli Hennig Society Funding has been provided by an AustralianResearch Council Linkage grant (LP0774959) to the University ofAdelaide with generous financial assistance from Beach Energy Ltdand the South Australian Museum SeaLink provided logistical supminusport DGBrsquos travel to Australia has been supported by grants from theSpanish Research Council (CSIC PA 1001105) the Complutense Uniminusversity of Madrid and the Spanish Ministry of Education and Science(CGL2006minus12245BTE CGL2009minus07073 and RYC2007minus00090)
ReferencesBergstroumlm J and Hou XminusG 2005 Early Palaeozoic lamellipedian arthrominus
pods In S Koenemann and RA Jenner (eds) Crustacea and Arthrominuspod Relationships Crustacean Issues 16 73ndash93
Bitsch J and Bitsch C 2007 The segmental organization of the head reminusgion in Chelicerata a critical review of recent studies and hypothesesActa Zoologica 88 317ndash335
Bousfeld EL 1995 A contribution to the natural classification of Lower andMiddle Cambrian arthropods food gathering and feeding mechanismsAmphipacifica 2 3ndash34
Boxshall GA 2004 The evolution of arthropod limbs Biological Reviews79 253ndash300
Bremer K 1994 Branch support and tree stability Cladistics 10 295ndash304Briggs DEG and Collins D 1999 The arthropod Alalcomenaeus cambricus
Simonetta from the Middle Cambrian Burgess Shale of British ColumbiaPalaeontology 42 953ndash977
Briggs DEG and Robison RA 1984 Exceptionally preserved nonminustrilobite arthropods and anomalocaridids from the Middle Cambrian ofUtah University of Kansas Paleontological Contributions 111 1ndash23
Briggs DEG Lieberman BS Hendricks JR Halgedahl SL andJerrard RD 2008 Middle Cambrian arthropods from Utah Journal ofPaleontology 82 238ndash254
Bruton DL 1981 The arthropod Sidneyia inexpectans Middle CambrianBurgess Shale British Columbia Philosophical Transactions of theRoyal Society of London Series B 295 619ndash656
Bruton DL and Whittington HB 1983 Emeraldella and Leanchoiliatwo arthropods from the Burgess Shale Middle Cambrian British Cominuslumbia Philosophical Transactions of the Royal Society of London Seminusries B 300 553ndash582
Budd GE 2002 A palaeontological solution to the arthropod head probminuslem Nature 417 271ndash275
Butterfield NJ 2002 Leanchoilia guts and the interpretation of threeminusdiminusmensional structures in Burgess Shaleminustype fossils Paleobiology 28155ndash171
Chen JminusY 2004 The Dawn of Animal World [in Chinese] 366 pp JiangScience and Technology Press Nanjing
Chen JminusY Waloszek D and Maas A 2004 A new ldquogreatminusappendagerdquo arminusthropod from the Lower Cambrian of China and homology of cheliceratechelicerae and raptorial anterominusventral appendages Lethaia 37 3ndash20
Cotton TJ and Braddy SJ 2004 The phylogeny of arachnomorph arthrominuspods and the origin of the Chelicerata Transactions of the Royal Societyof Edinburgh Earth Sciences 94 169ndash193
Daley AC Budd GE Caron JminusB Edgecombe GD and Collins D2009 The Burgess Shale anomalocaridid Hurdia and its significancefor early euarthropod evolution Science 323 1597ndash1600
Dunlop JA 2006 New ideas about the euchelicerate stemminuslineage In CDeltshev and P Stoev (eds) European Arachnology 2005 Acta Zoominuslogica Bulgarica (Supplement 1) 9ndash27
Dunlop JA and Selden PA 1997 The early history and phylogeny of thechelicerates In RA Fortey and RH Thomas (eds) Arthropod Relaminustionships Systematics Association Special Volume 55 221ndash235
Edgecombe GD and Ramskoumlld L 1999 Relationships of CambrianArachnata and the systematic position of Trilobita Journal of Paleonminustology 73 263ndash287
Farris JS Albert VA Kaumlllersjouml M Lipcomb D and Kluge AG1996 Parsimony jackknifing outperforms neighborminusjoining Cladistics12 99ndash124
GarciacuteaminusBellido DC and Collins D 2007 Reassessment of the genusLeanchoilia (Arthropoda Arachnomorpha) from the Middle CambrianBurgess Shale British Columbia Canada Palaeontology 50 693ndash709
GarciacuteaminusBellido DC Paterson JR Edgecombe GD Jago JB GehlingJG and Lee MSY 2009 The bivalved arthropods Isoxys and Tuzoiawith softminuspart preservation from the lower Cambrian Emu Bay ShaleLagerstaumltte (Kangaroo Island Australia) Palaeontology 52 1221ndash1241
Goloboff PA 1993 Estimating character weights during tree searchCladistics 9 83ndash91
Goloboff PA Farris JS and Nixon KC 2008 TNT a free program forphylogenetic analysis Cladistics 24 774ndash786
Harzsch S and Hafner G 2006 Evolution of eye development in arthrominuspods phylogenetic aspects Arthropod Structure amp Development 35319ndash340
Hendricks JR and Lieberman BS 2008 New phylogenetic insights intothe Cambrian radiation of arachnomorph arthropods Journal of Paleminusontology 82 585ndash594
Hou XminusG and Bergstroumlm J 1997 Arthropods of the Lower CambrianChengjiang fauna southwest China Fossils and Strata 45 1ndash116
Jago JB Zang W Sun X Brock GA Paterson JR and SkovstedCB 2006 A review of the Cambrian biostratigraphy of South Austraminuslia Palaeoworld 15 406ndash423
Liu Y Hou XminusG and Bergstroumlm J 2007 Chengjiang arthropod Leanminuschoilia illecebrosa (Hou 1987) reconsidered GFF 129 263ndash272
Luo H Hu S Chen L Zhang S and Tao Y 1999 Early CambrianChengjiang fauna from Kunming region China [in Chinese] 129 ppYunnan Science and Technology Press Kunming
Luo H Hu S Zhang S and Tao Y 1997 New occurrence of the EarlyCambrian Chengjiang fauna in Haikou Kunming Yunnan Provinceand study on Trilobitoidea Acta Geologica Sinica 71 122ndash132
Nixon KC 2002 WINCLADA Version 19908 Cornell University IthacaNY
Orr PJ Siveter DJ Briggs DEG Siveter DJ and Sutton MD 2000A new arthropod from the Silurian KonservatminusLagerstaumltte of Hereminusfordshire Proceedings of the Royal Society Series B Biological Sciminusences 267 1497ndash1504
Paterson JR and Brock GA 2007 Early Cambrian trilobites fromAngorichina Flinders Ranges South Australia with a new assemblagefrom the Pararaia bunyerooensis Zone Journal of Paleontology 81116ndash142
Paterson JR and Jago JB 2006 New trilobites from the Lower CambrianEmu Bay Shale Lagerstaumltte at Big Gully Kangaroo Island South Ausminustralia Memoirs of the Association of Australasian Palaeontologists 3243ndash57
Paterson JR Edgecombe GD GarciacuteaminusBellido DC Jago JB andGehling JG 2010 Nektaspid arthropods from the lower CambrianEmu Bay Shale Lagerstaumltte South Australia with a reassessment oflamellipedian relationships Palaeontology 53 377ndash402
Paterson JR Jago JB Gehling JG GarciacuteaminusBellido DC Edgecombe
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 399
GD and Lee MSY 2008 Early Cambrian arthropods from the EmuBay Shale Lagerstaumltte South Australia 319ndash325 In I Raacutebano RGozalo and D GarciacuteaminusBellido (eds) Advances in Trilobite ResearchCuadernos del Museo Geominero 9 319ndash325
Shultz JW 2007 A phylogenetic analysis of the arachnid orders based onmorphological characters Zoological Journal of the Linnean Society150 221ndash265
Stein M 2010 A new arthropod from the Early Cambrian of North Greenminusland with a ldquogreat appendagerdquominuslike antennula Zoological Journal ofthe Linnean Society 158 477ndash500
Stuumlrmer W and Bergstroumlm J 1978 The arthropod Cheloniellon from theDevonian Hunsruumlck Shale Palaumlontologische Zeitschrift 52 57ndash81
Sutton MD Briggs DEG Siveter DJ Siveter DJ and Orr PD2002 The arthropod Offacolus kingi (Chelicerata) from the Silurian ofHerefordshire England computer based morphological reconstrucminustions and phylogenetic affinity Proceedings of the Royal Society B Biminusological Sciences 269 1195ndash1203
Suzuki Y and Bergstroumlm J 2008 Respiration in trilobites a reevaluationGFF 130 211ndash229
Vannier J GarciacuteaminusBellido DC Hu XminusX and Chen AminusL 2009 Arthrominuspod visual predators in the early pelagic ecosystem evidence from the
Burgess Shale and Chengjiang biotas Proceedings of the Royal SocietyB Biological Sciences 276 2567ndash2574
Whittington HB 1974 Yohoia Walcott and Plenocaris n gen arthropodsfrom the Burgess Shale Middle Cambrian British Columbia Geologiminuscal Survey of Canada Bulletin 231 1ndash21
Wills MA Briggs DEG Fortey RA Wilkinson M and SneathPHA 1998 An arthropod phylogeny based on fossil and recent taxaIn GD Edgecombe (ed) Arthropod Fossils and Phylogeny 33ndash105Columbia University Press New York
Wilson LA 2006 Food for thought a morphological and taphonomic studyof fossilised digestive systems from Early to Middle Cambrian taxa275 pp Unpublished PhD thesis University of Cambridge Cambridge
Zhang XminusG and Hou X 2007 Gravitational constraints on the burial ofChengjiang fossils Palaios 22 448ndash453
Zhang XminusL Han J Zhang Z Liu H and Shu D 2004 Redescription ofthe Chengjiang arthropod Squamacula clypeata Hou and Bergstroumlm fromthe Lower Cambrian Southminuswest China Palaeontology 47 605ndash617
Zhao Y Zhu M Babcock LE Yuan L Parsley RL Peng J YangX and Wang Y 2005 Kaili Biota a taphonomic window on diversifiminuscation of metazoans from the basal Middle Cambrian Guizhou ChinaActa Geologica Sinica 79 751ndash765
400 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
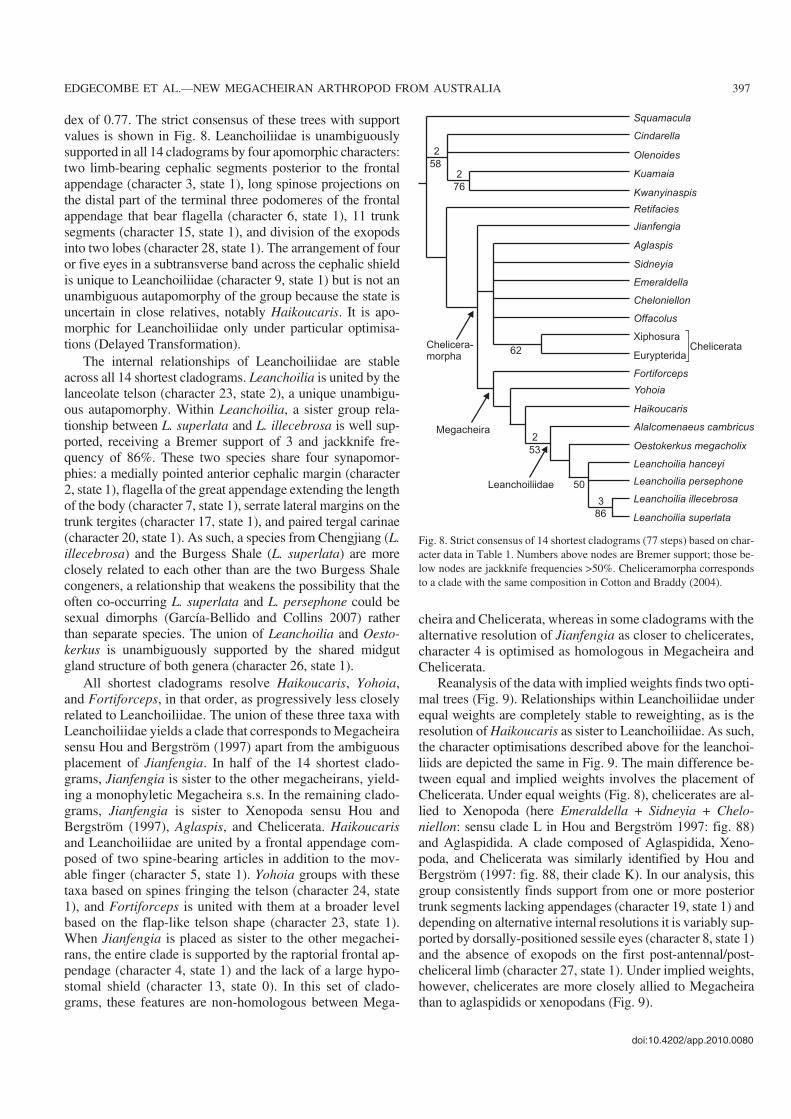
dex of 077 The strict consensus of these trees with supportvalues is shown in Fig 8 Leanchoiliidae is unambiguouslysupported in all 14 cladograms by four apomorphic characterstwo limbminusbearing cephalic segments posterior to the frontalappendage (character 3 state 1) long spinose projections onthe distal part of the terminal three podomeres of the frontalappendage that bear flagella (character 6 state 1) 11 trunksegments (character 15 state 1) and division of the exopodsinto two lobes (character 28 state 1) The arrangement of fouror five eyes in a subtransverse band across the cephalic shieldis unique to Leanchoiliidae (character 9 state 1) but is not anunambiguous autapomorphy of the group because the state isuncertain in close relatives notably Haikoucaris It is apominusmorphic for Leanchoiliidae only under particular optimisaminustions (Delayed Transformation)
The internal relationships of Leanchoiliidae are stableacross all 14 shortest cladograms Leanchoilia is united by thelanceolate telson (character 23 state 2) a unique unambiguminusous autapomorphy Within Leanchoilia a sister group relaminustionship between L superlata and L illecebrosa is well supminusported receiving a Bremer support of 3 and jackknife freminusquency of 86 These two species share four synapomorminusphies a medially pointed anterior cephalic margin (character2 state 1) flagella of the great appendage extending the lengthof the body (character 7 state 1) serrate lateral margins on thetrunk tergites (character 17 state 1) and paired tergal carinae(character 20 state 1) As such a species from Chengjiang (Lillecebrosa) and the Burgess Shale (L superlata) are moreclosely related to each other than are the two Burgess Shalecongeners a relationship that weakens the possibility that theoften cominusoccurring L superlata and L persephone could besexual dimorphs (GarciacuteaminusBellido and Collins 2007) ratherthan separate species The union of Leanchoilia and Oestominuskerkus is unambiguously supported by the shared midgutgland structure of both genera (character 26 state 1)
All shortest cladograms resolve Haikoucaris Yohoiaand Fortiforceps in that order as progressively less closelyrelated to Leanchoiliidae The union of these three taxa withLeanchoiliidae yields a clade that corresponds to Megacheirasensu Hou and Bergstroumlm (1997) apart from the ambiguousplacement of Jianfengia In half of the 14 shortest cladominusgrams Jianfengia is sister to the other megacheirans yieldminusing a monophyletic Megacheira ss In the remaining cladominusgrams Jianfengia is sister to Xenopoda sensu Hou andBergstroumlm (1997) Aglaspis and Chelicerata Haikoucarisand Leanchoiliidae are united by a frontal appendage comminusposed of two spineminusbearing articles in addition to the movminusable finger (character 5 state 1) Yohoia groups with thesetaxa based on spines fringing the telson (character 24 state1) and Fortiforceps is united with them at a broader levelbased on the flapminuslike telson shape (character 23 state 1)When Jianfengia is placed as sister to the other megacheiminusrans the entire clade is supported by the raptorial frontal apminuspendage (character 4 state 1) and the lack of a large hypominusstomal shield (character 13 state 0) In this set of cladominusgrams these features are nonminushomologous between Megaminus
cheira and Chelicerata whereas in some cladograms with thealternative resolution of Jianfengia as closer to cheliceratescharacter 4 is optimised as homologous in Megacheira andChelicerata
Reanalysis of the data with implied weights finds two optiminusmal trees (Fig 9) Relationships within Leanchoiliidae underequal weights are completely stable to reweighting as is theresolution of Haikoucaris as sister to Leanchoiliidae As suchthe character optimisations described above for the leanchoiminusliids are depicted the same in Fig 9 The main difference beminustween equal and implied weights involves the placement ofChelicerata Under equal weights (Fig 8) chelicerates are alminuslied to Xenopoda (here Emeraldella + Sidneyia + Chelominusniellon sensu clade L in Hou and Bergstroumlm 1997 fig 88)and Aglaspidida A clade composed of Aglaspidida Xenominuspoda and Chelicerata was similarly identified by Hou andBergstroumlm (1997 fig 88 their clade K) In our analysis thisgroup consistently finds support from one or more posteriortrunk segments lacking appendages (character 19 state 1) anddepending on alternative internal resolutions it is variably supminusported by dorsallyminuspositioned sessile eyes (character 8 state 1)and the absence of exopods on the first postminusantennalpostminuscheliceral limb (character 27 state 1) Under implied weightshowever chelicerates are more closely allied to Megacheirathan to aglaspidids or xenopodans (Fig 9)
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 397
2
582
76
Chelicera-morpha
Megacheira
62
2
53
50Leanchoiliidae
3
86
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
EurypteridaChelicerata
Fortiforceps
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia hanceyi
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
Fig 8 Strict consensus of 14 shortest cladograms (77 steps) based on charminusacter data in Table 1 Numbers above nodes are Bremer support those beminuslow nodes are jackknife frequencies gt50 Cheliceramorpha correspondsto a clade with the same composition in Cotton and Braddy (2004)
The major groupings in Fig 9 correspond closely to thoseidentified by Cotton and Braddy (2004) Notably the cladeuniting Aglaspis Xenopoda Offacolus Chelicerata Jianminusfengia and other Megacheira corresponds to their ldquoClade 3rdquo(Cotton and Braddy 2004 fig 8) for which they proposedthe name Cheliceramorpha The clade uniting CheloniellonAglaspis Sidneyia and Emeraldella in Fig 9 corresponds totheir ldquoClade 5rdquo and the clade that groups Megacheira andChelicerata is equivalent to their ldquoClade 4rdquo Their analysisdid not identify megacheirans as a monophyletic groupChelicerata was nested within the group and among taxaconsidered here Yohoia was resolved closer to cheliceratesthan to Leanchoiliidae In contrast Yohoia was regarded byBergstroumlm and Hou (2005) as more closely allied to Leanminuschoiliidae than to chelicerates a megacheiran grouping morecompatible with our cladograms The analysis of ldquogreat apminuspendage arthropodsrdquo by Vannier et al (2009) identified a
monophyletic Leanchoiliidae but a polyphyletic Megacheira(to the exclusion of Haikoucaris and Jianfengia) Howevertheir resolution of Fortiforceps and even more so Yohoia asthe closest relatives of Leanchoiliidae is as in our trees (apartfrom the different position of Haikoucaris)
In summary a major controversy in the systematics of theldquoshort great appendagerdquo arthropods remains whether thechelicerates are nested within the group or are instead moreclosely allied to taxa conventionally identified as lamelliminuspedians specifically Xenopoda and Aglaspidida We findthat both of these results are compatible with the availabledata depending on character weighting protocols Noneminustheless the broader megacheiranminuschelicerate issue does notcloud the status of leanchoiliids as a clade or the identificaminustion of their closest relatives (ie Haikoucaris and Yohoia)and the internal relationships of Leanchoiliidae are stableand supported by compelling characters
398 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
Eurypterida
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
2 7 17 20
3
2
1 1 1
26
1
281563
1 1 1 1
5
1
24
1
23 19 20 27 32
28Clade 4
1 0
4 13 11 27 0
154 1 1
3 19 32Clade 3
2 3
2 28
0
3 1 121
1
22
Clade 5
1 0
19 27
1
25
15 28 301 1
2312
1 150 1 3 3
15 16 2923
18
1 1
10 16
1 1
1 8
1
14
3
3
28 30
1 0
5
15
1 1
1
1
22
Fortiforceps
23
1 1 0 1
1 1
Leanchoilia hanceyi
2
2
1
1
Fig 9 Characters optimised on one of two shortest cladograms with imminusplied weights using a concavity constant k = 3 Black squares are nonminushomoplastic changes white squares homoplastic changes all changes areunambiguous Branches without strict support collapsed after optimisingcharacters Numbers for characters (above the branches) and states (belowthe branches) correspond to Table 1 ldquoClades 3ndash5rdquo correspond to groups sonumbered by Cotton and Braddy (2004 fig 8)
AcknowledgementsAccess to the field area has been most kindly allowed by the Buck famminusily We thank our collaborators in Emu Bay Shale research Jim Jago(University of South Australia Mawson Lakes) Jim Gehling andMike Lee (South Australian Museum Adelaide) for tremendous supminusport and advice as well as Mike Gemmell Dennis Rice Allison DaleyAaron Camens John Laurie Glenn Brock Pierre Kruse and Trevorand Jenny Worthy (all South Australian Museum) for their assistancein the field and Natalie Schroeder MaryminusAnne Binnie and RondaAtkinson (all South Australian Museum) for organisational support inthe lab and in the field Javier OrtegaminusHernaacutendez (University of Camminusbridge Cambridge UK) advised us on Kwanyinaspis and the journalrsquosreferees provided useful feedback TNT is available courtesy of theWilli Hennig Society Funding has been provided by an AustralianResearch Council Linkage grant (LP0774959) to the University ofAdelaide with generous financial assistance from Beach Energy Ltdand the South Australian Museum SeaLink provided logistical supminusport DGBrsquos travel to Australia has been supported by grants from theSpanish Research Council (CSIC PA 1001105) the Complutense Uniminusversity of Madrid and the Spanish Ministry of Education and Science(CGL2006minus12245BTE CGL2009minus07073 and RYC2007minus00090)
ReferencesBergstroumlm J and Hou XminusG 2005 Early Palaeozoic lamellipedian arthrominus
pods In S Koenemann and RA Jenner (eds) Crustacea and Arthrominuspod Relationships Crustacean Issues 16 73ndash93
Bitsch J and Bitsch C 2007 The segmental organization of the head reminusgion in Chelicerata a critical review of recent studies and hypothesesActa Zoologica 88 317ndash335
Bousfeld EL 1995 A contribution to the natural classification of Lower andMiddle Cambrian arthropods food gathering and feeding mechanismsAmphipacifica 2 3ndash34
Boxshall GA 2004 The evolution of arthropod limbs Biological Reviews79 253ndash300
Bremer K 1994 Branch support and tree stability Cladistics 10 295ndash304Briggs DEG and Collins D 1999 The arthropod Alalcomenaeus cambricus
Simonetta from the Middle Cambrian Burgess Shale of British ColumbiaPalaeontology 42 953ndash977
Briggs DEG and Robison RA 1984 Exceptionally preserved nonminustrilobite arthropods and anomalocaridids from the Middle Cambrian ofUtah University of Kansas Paleontological Contributions 111 1ndash23
Briggs DEG Lieberman BS Hendricks JR Halgedahl SL andJerrard RD 2008 Middle Cambrian arthropods from Utah Journal ofPaleontology 82 238ndash254
Bruton DL 1981 The arthropod Sidneyia inexpectans Middle CambrianBurgess Shale British Columbia Philosophical Transactions of theRoyal Society of London Series B 295 619ndash656
Bruton DL and Whittington HB 1983 Emeraldella and Leanchoiliatwo arthropods from the Burgess Shale Middle Cambrian British Cominuslumbia Philosophical Transactions of the Royal Society of London Seminusries B 300 553ndash582
Budd GE 2002 A palaeontological solution to the arthropod head probminuslem Nature 417 271ndash275
Butterfield NJ 2002 Leanchoilia guts and the interpretation of threeminusdiminusmensional structures in Burgess Shaleminustype fossils Paleobiology 28155ndash171
Chen JminusY 2004 The Dawn of Animal World [in Chinese] 366 pp JiangScience and Technology Press Nanjing
Chen JminusY Waloszek D and Maas A 2004 A new ldquogreatminusappendagerdquo arminusthropod from the Lower Cambrian of China and homology of cheliceratechelicerae and raptorial anterominusventral appendages Lethaia 37 3ndash20
Cotton TJ and Braddy SJ 2004 The phylogeny of arachnomorph arthrominuspods and the origin of the Chelicerata Transactions of the Royal Societyof Edinburgh Earth Sciences 94 169ndash193
Daley AC Budd GE Caron JminusB Edgecombe GD and Collins D2009 The Burgess Shale anomalocaridid Hurdia and its significancefor early euarthropod evolution Science 323 1597ndash1600
Dunlop JA 2006 New ideas about the euchelicerate stemminuslineage In CDeltshev and P Stoev (eds) European Arachnology 2005 Acta Zoominuslogica Bulgarica (Supplement 1) 9ndash27
Dunlop JA and Selden PA 1997 The early history and phylogeny of thechelicerates In RA Fortey and RH Thomas (eds) Arthropod Relaminustionships Systematics Association Special Volume 55 221ndash235
Edgecombe GD and Ramskoumlld L 1999 Relationships of CambrianArachnata and the systematic position of Trilobita Journal of Paleonminustology 73 263ndash287
Farris JS Albert VA Kaumlllersjouml M Lipcomb D and Kluge AG1996 Parsimony jackknifing outperforms neighborminusjoining Cladistics12 99ndash124
GarciacuteaminusBellido DC and Collins D 2007 Reassessment of the genusLeanchoilia (Arthropoda Arachnomorpha) from the Middle CambrianBurgess Shale British Columbia Canada Palaeontology 50 693ndash709
GarciacuteaminusBellido DC Paterson JR Edgecombe GD Jago JB GehlingJG and Lee MSY 2009 The bivalved arthropods Isoxys and Tuzoiawith softminuspart preservation from the lower Cambrian Emu Bay ShaleLagerstaumltte (Kangaroo Island Australia) Palaeontology 52 1221ndash1241
Goloboff PA 1993 Estimating character weights during tree searchCladistics 9 83ndash91
Goloboff PA Farris JS and Nixon KC 2008 TNT a free program forphylogenetic analysis Cladistics 24 774ndash786
Harzsch S and Hafner G 2006 Evolution of eye development in arthrominuspods phylogenetic aspects Arthropod Structure amp Development 35319ndash340
Hendricks JR and Lieberman BS 2008 New phylogenetic insights intothe Cambrian radiation of arachnomorph arthropods Journal of Paleminusontology 82 585ndash594
Hou XminusG and Bergstroumlm J 1997 Arthropods of the Lower CambrianChengjiang fauna southwest China Fossils and Strata 45 1ndash116
Jago JB Zang W Sun X Brock GA Paterson JR and SkovstedCB 2006 A review of the Cambrian biostratigraphy of South Austraminuslia Palaeoworld 15 406ndash423
Liu Y Hou XminusG and Bergstroumlm J 2007 Chengjiang arthropod Leanminuschoilia illecebrosa (Hou 1987) reconsidered GFF 129 263ndash272
Luo H Hu S Chen L Zhang S and Tao Y 1999 Early CambrianChengjiang fauna from Kunming region China [in Chinese] 129 ppYunnan Science and Technology Press Kunming
Luo H Hu S Zhang S and Tao Y 1997 New occurrence of the EarlyCambrian Chengjiang fauna in Haikou Kunming Yunnan Provinceand study on Trilobitoidea Acta Geologica Sinica 71 122ndash132
Nixon KC 2002 WINCLADA Version 19908 Cornell University IthacaNY
Orr PJ Siveter DJ Briggs DEG Siveter DJ and Sutton MD 2000A new arthropod from the Silurian KonservatminusLagerstaumltte of Hereminusfordshire Proceedings of the Royal Society Series B Biological Sciminusences 267 1497ndash1504
Paterson JR and Brock GA 2007 Early Cambrian trilobites fromAngorichina Flinders Ranges South Australia with a new assemblagefrom the Pararaia bunyerooensis Zone Journal of Paleontology 81116ndash142
Paterson JR and Jago JB 2006 New trilobites from the Lower CambrianEmu Bay Shale Lagerstaumltte at Big Gully Kangaroo Island South Ausminustralia Memoirs of the Association of Australasian Palaeontologists 3243ndash57
Paterson JR Edgecombe GD GarciacuteaminusBellido DC Jago JB andGehling JG 2010 Nektaspid arthropods from the lower CambrianEmu Bay Shale Lagerstaumltte South Australia with a reassessment oflamellipedian relationships Palaeontology 53 377ndash402
Paterson JR Jago JB Gehling JG GarciacuteaminusBellido DC Edgecombe
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 399
GD and Lee MSY 2008 Early Cambrian arthropods from the EmuBay Shale Lagerstaumltte South Australia 319ndash325 In I Raacutebano RGozalo and D GarciacuteaminusBellido (eds) Advances in Trilobite ResearchCuadernos del Museo Geominero 9 319ndash325
Shultz JW 2007 A phylogenetic analysis of the arachnid orders based onmorphological characters Zoological Journal of the Linnean Society150 221ndash265
Stein M 2010 A new arthropod from the Early Cambrian of North Greenminusland with a ldquogreat appendagerdquominuslike antennula Zoological Journal ofthe Linnean Society 158 477ndash500
Stuumlrmer W and Bergstroumlm J 1978 The arthropod Cheloniellon from theDevonian Hunsruumlck Shale Palaumlontologische Zeitschrift 52 57ndash81
Sutton MD Briggs DEG Siveter DJ Siveter DJ and Orr PD2002 The arthropod Offacolus kingi (Chelicerata) from the Silurian ofHerefordshire England computer based morphological reconstrucminustions and phylogenetic affinity Proceedings of the Royal Society B Biminusological Sciences 269 1195ndash1203
Suzuki Y and Bergstroumlm J 2008 Respiration in trilobites a reevaluationGFF 130 211ndash229
Vannier J GarciacuteaminusBellido DC Hu XminusX and Chen AminusL 2009 Arthrominuspod visual predators in the early pelagic ecosystem evidence from the
Burgess Shale and Chengjiang biotas Proceedings of the Royal SocietyB Biological Sciences 276 2567ndash2574
Whittington HB 1974 Yohoia Walcott and Plenocaris n gen arthropodsfrom the Burgess Shale Middle Cambrian British Columbia Geologiminuscal Survey of Canada Bulletin 231 1ndash21
Wills MA Briggs DEG Fortey RA Wilkinson M and SneathPHA 1998 An arthropod phylogeny based on fossil and recent taxaIn GD Edgecombe (ed) Arthropod Fossils and Phylogeny 33ndash105Columbia University Press New York
Wilson LA 2006 Food for thought a morphological and taphonomic studyof fossilised digestive systems from Early to Middle Cambrian taxa275 pp Unpublished PhD thesis University of Cambridge Cambridge
Zhang XminusG and Hou X 2007 Gravitational constraints on the burial ofChengjiang fossils Palaios 22 448ndash453
Zhang XminusL Han J Zhang Z Liu H and Shu D 2004 Redescription ofthe Chengjiang arthropod Squamacula clypeata Hou and Bergstroumlm fromthe Lower Cambrian Southminuswest China Palaeontology 47 605ndash617
Zhao Y Zhu M Babcock LE Yuan L Parsley RL Peng J YangX and Wang Y 2005 Kaili Biota a taphonomic window on diversifiminuscation of metazoans from the basal Middle Cambrian Guizhou ChinaActa Geologica Sinica 79 751ndash765
400 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011

The major groupings in Fig 9 correspond closely to thoseidentified by Cotton and Braddy (2004) Notably the cladeuniting Aglaspis Xenopoda Offacolus Chelicerata Jianminusfengia and other Megacheira corresponds to their ldquoClade 3rdquo(Cotton and Braddy 2004 fig 8) for which they proposedthe name Cheliceramorpha The clade uniting CheloniellonAglaspis Sidneyia and Emeraldella in Fig 9 corresponds totheir ldquoClade 5rdquo and the clade that groups Megacheira andChelicerata is equivalent to their ldquoClade 4rdquo Their analysisdid not identify megacheirans as a monophyletic groupChelicerata was nested within the group and among taxaconsidered here Yohoia was resolved closer to cheliceratesthan to Leanchoiliidae In contrast Yohoia was regarded byBergstroumlm and Hou (2005) as more closely allied to Leanminuschoiliidae than to chelicerates a megacheiran grouping morecompatible with our cladograms The analysis of ldquogreat apminuspendage arthropodsrdquo by Vannier et al (2009) identified a
monophyletic Leanchoiliidae but a polyphyletic Megacheira(to the exclusion of Haikoucaris and Jianfengia) Howevertheir resolution of Fortiforceps and even more so Yohoia asthe closest relatives of Leanchoiliidae is as in our trees (apartfrom the different position of Haikoucaris)
In summary a major controversy in the systematics of theldquoshort great appendagerdquo arthropods remains whether thechelicerates are nested within the group or are instead moreclosely allied to taxa conventionally identified as lamelliminuspedians specifically Xenopoda and Aglaspidida We findthat both of these results are compatible with the availabledata depending on character weighting protocols Noneminustheless the broader megacheiranminuschelicerate issue does notcloud the status of leanchoiliids as a clade or the identificaminustion of their closest relatives (ie Haikoucaris and Yohoia)and the internal relationships of Leanchoiliidae are stableand supported by compelling characters
398 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011
Squamacula
Cindarella
Olenoides
Kuamaia
Kwanyinaspis
Retifacies
Jianfengia
Aglaspis
Sidneyia
Emeraldella
Cheloniellon
Offacolus
Xiphosura
Eurypterida
Yohoia
Haikoucaris
Alalcomenaeus cambricus
Oestokerkus megacholix
Leanchoilia persephone
Leanchoilia illecebrosa
Leanchoilia superlata
2 7 17 20
3
2
1 1 1
26
1
281563
1 1 1 1
5
1
24
1
23 19 20 27 32
28Clade 4
1 0
4 13 11 27 0
154 1 1
3 19 32Clade 3
2 3
2 28
0
3 1 121
1
22
Clade 5
1 0
19 27
1
25
15 28 301 1
2312
1 150 1 3 3
15 16 2923
18
1 1
10 16
1 1
1 8
1
14
3
3
28 30
1 0
5
15
1 1
1
1
22
Fortiforceps
23
1 1 0 1
1 1
Leanchoilia hanceyi
2
2
1
1
Fig 9 Characters optimised on one of two shortest cladograms with imminusplied weights using a concavity constant k = 3 Black squares are nonminushomoplastic changes white squares homoplastic changes all changes areunambiguous Branches without strict support collapsed after optimisingcharacters Numbers for characters (above the branches) and states (belowthe branches) correspond to Table 1 ldquoClades 3ndash5rdquo correspond to groups sonumbered by Cotton and Braddy (2004 fig 8)
AcknowledgementsAccess to the field area has been most kindly allowed by the Buck famminusily We thank our collaborators in Emu Bay Shale research Jim Jago(University of South Australia Mawson Lakes) Jim Gehling andMike Lee (South Australian Museum Adelaide) for tremendous supminusport and advice as well as Mike Gemmell Dennis Rice Allison DaleyAaron Camens John Laurie Glenn Brock Pierre Kruse and Trevorand Jenny Worthy (all South Australian Museum) for their assistancein the field and Natalie Schroeder MaryminusAnne Binnie and RondaAtkinson (all South Australian Museum) for organisational support inthe lab and in the field Javier OrtegaminusHernaacutendez (University of Camminusbridge Cambridge UK) advised us on Kwanyinaspis and the journalrsquosreferees provided useful feedback TNT is available courtesy of theWilli Hennig Society Funding has been provided by an AustralianResearch Council Linkage grant (LP0774959) to the University ofAdelaide with generous financial assistance from Beach Energy Ltdand the South Australian Museum SeaLink provided logistical supminusport DGBrsquos travel to Australia has been supported by grants from theSpanish Research Council (CSIC PA 1001105) the Complutense Uniminusversity of Madrid and the Spanish Ministry of Education and Science(CGL2006minus12245BTE CGL2009minus07073 and RYC2007minus00090)
ReferencesBergstroumlm J and Hou XminusG 2005 Early Palaeozoic lamellipedian arthrominus
pods In S Koenemann and RA Jenner (eds) Crustacea and Arthrominuspod Relationships Crustacean Issues 16 73ndash93
Bitsch J and Bitsch C 2007 The segmental organization of the head reminusgion in Chelicerata a critical review of recent studies and hypothesesActa Zoologica 88 317ndash335
Bousfeld EL 1995 A contribution to the natural classification of Lower andMiddle Cambrian arthropods food gathering and feeding mechanismsAmphipacifica 2 3ndash34
Boxshall GA 2004 The evolution of arthropod limbs Biological Reviews79 253ndash300
Bremer K 1994 Branch support and tree stability Cladistics 10 295ndash304Briggs DEG and Collins D 1999 The arthropod Alalcomenaeus cambricus
Simonetta from the Middle Cambrian Burgess Shale of British ColumbiaPalaeontology 42 953ndash977
Briggs DEG and Robison RA 1984 Exceptionally preserved nonminustrilobite arthropods and anomalocaridids from the Middle Cambrian ofUtah University of Kansas Paleontological Contributions 111 1ndash23
Briggs DEG Lieberman BS Hendricks JR Halgedahl SL andJerrard RD 2008 Middle Cambrian arthropods from Utah Journal ofPaleontology 82 238ndash254
Bruton DL 1981 The arthropod Sidneyia inexpectans Middle CambrianBurgess Shale British Columbia Philosophical Transactions of theRoyal Society of London Series B 295 619ndash656
Bruton DL and Whittington HB 1983 Emeraldella and Leanchoiliatwo arthropods from the Burgess Shale Middle Cambrian British Cominuslumbia Philosophical Transactions of the Royal Society of London Seminusries B 300 553ndash582
Budd GE 2002 A palaeontological solution to the arthropod head probminuslem Nature 417 271ndash275
Butterfield NJ 2002 Leanchoilia guts and the interpretation of threeminusdiminusmensional structures in Burgess Shaleminustype fossils Paleobiology 28155ndash171
Chen JminusY 2004 The Dawn of Animal World [in Chinese] 366 pp JiangScience and Technology Press Nanjing
Chen JminusY Waloszek D and Maas A 2004 A new ldquogreatminusappendagerdquo arminusthropod from the Lower Cambrian of China and homology of cheliceratechelicerae and raptorial anterominusventral appendages Lethaia 37 3ndash20
Cotton TJ and Braddy SJ 2004 The phylogeny of arachnomorph arthrominuspods and the origin of the Chelicerata Transactions of the Royal Societyof Edinburgh Earth Sciences 94 169ndash193
Daley AC Budd GE Caron JminusB Edgecombe GD and Collins D2009 The Burgess Shale anomalocaridid Hurdia and its significancefor early euarthropod evolution Science 323 1597ndash1600
Dunlop JA 2006 New ideas about the euchelicerate stemminuslineage In CDeltshev and P Stoev (eds) European Arachnology 2005 Acta Zoominuslogica Bulgarica (Supplement 1) 9ndash27
Dunlop JA and Selden PA 1997 The early history and phylogeny of thechelicerates In RA Fortey and RH Thomas (eds) Arthropod Relaminustionships Systematics Association Special Volume 55 221ndash235
Edgecombe GD and Ramskoumlld L 1999 Relationships of CambrianArachnata and the systematic position of Trilobita Journal of Paleonminustology 73 263ndash287
Farris JS Albert VA Kaumlllersjouml M Lipcomb D and Kluge AG1996 Parsimony jackknifing outperforms neighborminusjoining Cladistics12 99ndash124
GarciacuteaminusBellido DC and Collins D 2007 Reassessment of the genusLeanchoilia (Arthropoda Arachnomorpha) from the Middle CambrianBurgess Shale British Columbia Canada Palaeontology 50 693ndash709
GarciacuteaminusBellido DC Paterson JR Edgecombe GD Jago JB GehlingJG and Lee MSY 2009 The bivalved arthropods Isoxys and Tuzoiawith softminuspart preservation from the lower Cambrian Emu Bay ShaleLagerstaumltte (Kangaroo Island Australia) Palaeontology 52 1221ndash1241
Goloboff PA 1993 Estimating character weights during tree searchCladistics 9 83ndash91
Goloboff PA Farris JS and Nixon KC 2008 TNT a free program forphylogenetic analysis Cladistics 24 774ndash786
Harzsch S and Hafner G 2006 Evolution of eye development in arthrominuspods phylogenetic aspects Arthropod Structure amp Development 35319ndash340
Hendricks JR and Lieberman BS 2008 New phylogenetic insights intothe Cambrian radiation of arachnomorph arthropods Journal of Paleminusontology 82 585ndash594
Hou XminusG and Bergstroumlm J 1997 Arthropods of the Lower CambrianChengjiang fauna southwest China Fossils and Strata 45 1ndash116
Jago JB Zang W Sun X Brock GA Paterson JR and SkovstedCB 2006 A review of the Cambrian biostratigraphy of South Austraminuslia Palaeoworld 15 406ndash423
Liu Y Hou XminusG and Bergstroumlm J 2007 Chengjiang arthropod Leanminuschoilia illecebrosa (Hou 1987) reconsidered GFF 129 263ndash272
Luo H Hu S Chen L Zhang S and Tao Y 1999 Early CambrianChengjiang fauna from Kunming region China [in Chinese] 129 ppYunnan Science and Technology Press Kunming
Luo H Hu S Zhang S and Tao Y 1997 New occurrence of the EarlyCambrian Chengjiang fauna in Haikou Kunming Yunnan Provinceand study on Trilobitoidea Acta Geologica Sinica 71 122ndash132
Nixon KC 2002 WINCLADA Version 19908 Cornell University IthacaNY
Orr PJ Siveter DJ Briggs DEG Siveter DJ and Sutton MD 2000A new arthropod from the Silurian KonservatminusLagerstaumltte of Hereminusfordshire Proceedings of the Royal Society Series B Biological Sciminusences 267 1497ndash1504
Paterson JR and Brock GA 2007 Early Cambrian trilobites fromAngorichina Flinders Ranges South Australia with a new assemblagefrom the Pararaia bunyerooensis Zone Journal of Paleontology 81116ndash142
Paterson JR and Jago JB 2006 New trilobites from the Lower CambrianEmu Bay Shale Lagerstaumltte at Big Gully Kangaroo Island South Ausminustralia Memoirs of the Association of Australasian Palaeontologists 3243ndash57
Paterson JR Edgecombe GD GarciacuteaminusBellido DC Jago JB andGehling JG 2010 Nektaspid arthropods from the lower CambrianEmu Bay Shale Lagerstaumltte South Australia with a reassessment oflamellipedian relationships Palaeontology 53 377ndash402
Paterson JR Jago JB Gehling JG GarciacuteaminusBellido DC Edgecombe
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 399
GD and Lee MSY 2008 Early Cambrian arthropods from the EmuBay Shale Lagerstaumltte South Australia 319ndash325 In I Raacutebano RGozalo and D GarciacuteaminusBellido (eds) Advances in Trilobite ResearchCuadernos del Museo Geominero 9 319ndash325
Shultz JW 2007 A phylogenetic analysis of the arachnid orders based onmorphological characters Zoological Journal of the Linnean Society150 221ndash265
Stein M 2010 A new arthropod from the Early Cambrian of North Greenminusland with a ldquogreat appendagerdquominuslike antennula Zoological Journal ofthe Linnean Society 158 477ndash500
Stuumlrmer W and Bergstroumlm J 1978 The arthropod Cheloniellon from theDevonian Hunsruumlck Shale Palaumlontologische Zeitschrift 52 57ndash81
Sutton MD Briggs DEG Siveter DJ Siveter DJ and Orr PD2002 The arthropod Offacolus kingi (Chelicerata) from the Silurian ofHerefordshire England computer based morphological reconstrucminustions and phylogenetic affinity Proceedings of the Royal Society B Biminusological Sciences 269 1195ndash1203
Suzuki Y and Bergstroumlm J 2008 Respiration in trilobites a reevaluationGFF 130 211ndash229
Vannier J GarciacuteaminusBellido DC Hu XminusX and Chen AminusL 2009 Arthrominuspod visual predators in the early pelagic ecosystem evidence from the
Burgess Shale and Chengjiang biotas Proceedings of the Royal SocietyB Biological Sciences 276 2567ndash2574
Whittington HB 1974 Yohoia Walcott and Plenocaris n gen arthropodsfrom the Burgess Shale Middle Cambrian British Columbia Geologiminuscal Survey of Canada Bulletin 231 1ndash21
Wills MA Briggs DEG Fortey RA Wilkinson M and SneathPHA 1998 An arthropod phylogeny based on fossil and recent taxaIn GD Edgecombe (ed) Arthropod Fossils and Phylogeny 33ndash105Columbia University Press New York
Wilson LA 2006 Food for thought a morphological and taphonomic studyof fossilised digestive systems from Early to Middle Cambrian taxa275 pp Unpublished PhD thesis University of Cambridge Cambridge
Zhang XminusG and Hou X 2007 Gravitational constraints on the burial ofChengjiang fossils Palaios 22 448ndash453
Zhang XminusL Han J Zhang Z Liu H and Shu D 2004 Redescription ofthe Chengjiang arthropod Squamacula clypeata Hou and Bergstroumlm fromthe Lower Cambrian Southminuswest China Palaeontology 47 605ndash617
Zhao Y Zhu M Babcock LE Yuan L Parsley RL Peng J YangX and Wang Y 2005 Kaili Biota a taphonomic window on diversifiminuscation of metazoans from the basal Middle Cambrian Guizhou ChinaActa Geologica Sinica 79 751ndash765
400 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011

AcknowledgementsAccess to the field area has been most kindly allowed by the Buck famminusily We thank our collaborators in Emu Bay Shale research Jim Jago(University of South Australia Mawson Lakes) Jim Gehling andMike Lee (South Australian Museum Adelaide) for tremendous supminusport and advice as well as Mike Gemmell Dennis Rice Allison DaleyAaron Camens John Laurie Glenn Brock Pierre Kruse and Trevorand Jenny Worthy (all South Australian Museum) for their assistancein the field and Natalie Schroeder MaryminusAnne Binnie and RondaAtkinson (all South Australian Museum) for organisational support inthe lab and in the field Javier OrtegaminusHernaacutendez (University of Camminusbridge Cambridge UK) advised us on Kwanyinaspis and the journalrsquosreferees provided useful feedback TNT is available courtesy of theWilli Hennig Society Funding has been provided by an AustralianResearch Council Linkage grant (LP0774959) to the University ofAdelaide with generous financial assistance from Beach Energy Ltdand the South Australian Museum SeaLink provided logistical supminusport DGBrsquos travel to Australia has been supported by grants from theSpanish Research Council (CSIC PA 1001105) the Complutense Uniminusversity of Madrid and the Spanish Ministry of Education and Science(CGL2006minus12245BTE CGL2009minus07073 and RYC2007minus00090)
ReferencesBergstroumlm J and Hou XminusG 2005 Early Palaeozoic lamellipedian arthrominus
pods In S Koenemann and RA Jenner (eds) Crustacea and Arthrominuspod Relationships Crustacean Issues 16 73ndash93
Bitsch J and Bitsch C 2007 The segmental organization of the head reminusgion in Chelicerata a critical review of recent studies and hypothesesActa Zoologica 88 317ndash335
Bousfeld EL 1995 A contribution to the natural classification of Lower andMiddle Cambrian arthropods food gathering and feeding mechanismsAmphipacifica 2 3ndash34
Boxshall GA 2004 The evolution of arthropod limbs Biological Reviews79 253ndash300
Bremer K 1994 Branch support and tree stability Cladistics 10 295ndash304Briggs DEG and Collins D 1999 The arthropod Alalcomenaeus cambricus
Simonetta from the Middle Cambrian Burgess Shale of British ColumbiaPalaeontology 42 953ndash977
Briggs DEG and Robison RA 1984 Exceptionally preserved nonminustrilobite arthropods and anomalocaridids from the Middle Cambrian ofUtah University of Kansas Paleontological Contributions 111 1ndash23
Briggs DEG Lieberman BS Hendricks JR Halgedahl SL andJerrard RD 2008 Middle Cambrian arthropods from Utah Journal ofPaleontology 82 238ndash254
Bruton DL 1981 The arthropod Sidneyia inexpectans Middle CambrianBurgess Shale British Columbia Philosophical Transactions of theRoyal Society of London Series B 295 619ndash656
Bruton DL and Whittington HB 1983 Emeraldella and Leanchoiliatwo arthropods from the Burgess Shale Middle Cambrian British Cominuslumbia Philosophical Transactions of the Royal Society of London Seminusries B 300 553ndash582
Budd GE 2002 A palaeontological solution to the arthropod head probminuslem Nature 417 271ndash275
Butterfield NJ 2002 Leanchoilia guts and the interpretation of threeminusdiminusmensional structures in Burgess Shaleminustype fossils Paleobiology 28155ndash171
Chen JminusY 2004 The Dawn of Animal World [in Chinese] 366 pp JiangScience and Technology Press Nanjing
Chen JminusY Waloszek D and Maas A 2004 A new ldquogreatminusappendagerdquo arminusthropod from the Lower Cambrian of China and homology of cheliceratechelicerae and raptorial anterominusventral appendages Lethaia 37 3ndash20
Cotton TJ and Braddy SJ 2004 The phylogeny of arachnomorph arthrominuspods and the origin of the Chelicerata Transactions of the Royal Societyof Edinburgh Earth Sciences 94 169ndash193
Daley AC Budd GE Caron JminusB Edgecombe GD and Collins D2009 The Burgess Shale anomalocaridid Hurdia and its significancefor early euarthropod evolution Science 323 1597ndash1600
Dunlop JA 2006 New ideas about the euchelicerate stemminuslineage In CDeltshev and P Stoev (eds) European Arachnology 2005 Acta Zoominuslogica Bulgarica (Supplement 1) 9ndash27
Dunlop JA and Selden PA 1997 The early history and phylogeny of thechelicerates In RA Fortey and RH Thomas (eds) Arthropod Relaminustionships Systematics Association Special Volume 55 221ndash235
Edgecombe GD and Ramskoumlld L 1999 Relationships of CambrianArachnata and the systematic position of Trilobita Journal of Paleonminustology 73 263ndash287
Farris JS Albert VA Kaumlllersjouml M Lipcomb D and Kluge AG1996 Parsimony jackknifing outperforms neighborminusjoining Cladistics12 99ndash124
GarciacuteaminusBellido DC and Collins D 2007 Reassessment of the genusLeanchoilia (Arthropoda Arachnomorpha) from the Middle CambrianBurgess Shale British Columbia Canada Palaeontology 50 693ndash709
GarciacuteaminusBellido DC Paterson JR Edgecombe GD Jago JB GehlingJG and Lee MSY 2009 The bivalved arthropods Isoxys and Tuzoiawith softminuspart preservation from the lower Cambrian Emu Bay ShaleLagerstaumltte (Kangaroo Island Australia) Palaeontology 52 1221ndash1241
Goloboff PA 1993 Estimating character weights during tree searchCladistics 9 83ndash91
Goloboff PA Farris JS and Nixon KC 2008 TNT a free program forphylogenetic analysis Cladistics 24 774ndash786
Harzsch S and Hafner G 2006 Evolution of eye development in arthrominuspods phylogenetic aspects Arthropod Structure amp Development 35319ndash340
Hendricks JR and Lieberman BS 2008 New phylogenetic insights intothe Cambrian radiation of arachnomorph arthropods Journal of Paleminusontology 82 585ndash594
Hou XminusG and Bergstroumlm J 1997 Arthropods of the Lower CambrianChengjiang fauna southwest China Fossils and Strata 45 1ndash116
Jago JB Zang W Sun X Brock GA Paterson JR and SkovstedCB 2006 A review of the Cambrian biostratigraphy of South Austraminuslia Palaeoworld 15 406ndash423
Liu Y Hou XminusG and Bergstroumlm J 2007 Chengjiang arthropod Leanminuschoilia illecebrosa (Hou 1987) reconsidered GFF 129 263ndash272
Luo H Hu S Chen L Zhang S and Tao Y 1999 Early CambrianChengjiang fauna from Kunming region China [in Chinese] 129 ppYunnan Science and Technology Press Kunming
Luo H Hu S Zhang S and Tao Y 1997 New occurrence of the EarlyCambrian Chengjiang fauna in Haikou Kunming Yunnan Provinceand study on Trilobitoidea Acta Geologica Sinica 71 122ndash132
Nixon KC 2002 WINCLADA Version 19908 Cornell University IthacaNY
Orr PJ Siveter DJ Briggs DEG Siveter DJ and Sutton MD 2000A new arthropod from the Silurian KonservatminusLagerstaumltte of Hereminusfordshire Proceedings of the Royal Society Series B Biological Sciminusences 267 1497ndash1504
Paterson JR and Brock GA 2007 Early Cambrian trilobites fromAngorichina Flinders Ranges South Australia with a new assemblagefrom the Pararaia bunyerooensis Zone Journal of Paleontology 81116ndash142
Paterson JR and Jago JB 2006 New trilobites from the Lower CambrianEmu Bay Shale Lagerstaumltte at Big Gully Kangaroo Island South Ausminustralia Memoirs of the Association of Australasian Palaeontologists 3243ndash57
Paterson JR Edgecombe GD GarciacuteaminusBellido DC Jago JB andGehling JG 2010 Nektaspid arthropods from the lower CambrianEmu Bay Shale Lagerstaumltte South Australia with a reassessment oflamellipedian relationships Palaeontology 53 377ndash402
Paterson JR Jago JB Gehling JG GarciacuteaminusBellido DC Edgecombe
doi104202app20100080
EDGECOMBE ET ALmdashNEW MEGACHEIRAN ARTHROPOD FROM AUSTRALIA 399
GD and Lee MSY 2008 Early Cambrian arthropods from the EmuBay Shale Lagerstaumltte South Australia 319ndash325 In I Raacutebano RGozalo and D GarciacuteaminusBellido (eds) Advances in Trilobite ResearchCuadernos del Museo Geominero 9 319ndash325
Shultz JW 2007 A phylogenetic analysis of the arachnid orders based onmorphological characters Zoological Journal of the Linnean Society150 221ndash265
Stein M 2010 A new arthropod from the Early Cambrian of North Greenminusland with a ldquogreat appendagerdquominuslike antennula Zoological Journal ofthe Linnean Society 158 477ndash500
Stuumlrmer W and Bergstroumlm J 1978 The arthropod Cheloniellon from theDevonian Hunsruumlck Shale Palaumlontologische Zeitschrift 52 57ndash81
Sutton MD Briggs DEG Siveter DJ Siveter DJ and Orr PD2002 The arthropod Offacolus kingi (Chelicerata) from the Silurian ofHerefordshire England computer based morphological reconstrucminustions and phylogenetic affinity Proceedings of the Royal Society B Biminusological Sciences 269 1195ndash1203
Suzuki Y and Bergstroumlm J 2008 Respiration in trilobites a reevaluationGFF 130 211ndash229
Vannier J GarciacuteaminusBellido DC Hu XminusX and Chen AminusL 2009 Arthrominuspod visual predators in the early pelagic ecosystem evidence from the
Burgess Shale and Chengjiang biotas Proceedings of the Royal SocietyB Biological Sciences 276 2567ndash2574
Whittington HB 1974 Yohoia Walcott and Plenocaris n gen arthropodsfrom the Burgess Shale Middle Cambrian British Columbia Geologiminuscal Survey of Canada Bulletin 231 1ndash21
Wills MA Briggs DEG Fortey RA Wilkinson M and SneathPHA 1998 An arthropod phylogeny based on fossil and recent taxaIn GD Edgecombe (ed) Arthropod Fossils and Phylogeny 33ndash105Columbia University Press New York
Wilson LA 2006 Food for thought a morphological and taphonomic studyof fossilised digestive systems from Early to Middle Cambrian taxa275 pp Unpublished PhD thesis University of Cambridge Cambridge
Zhang XminusG and Hou X 2007 Gravitational constraints on the burial ofChengjiang fossils Palaios 22 448ndash453
Zhang XminusL Han J Zhang Z Liu H and Shu D 2004 Redescription ofthe Chengjiang arthropod Squamacula clypeata Hou and Bergstroumlm fromthe Lower Cambrian Southminuswest China Palaeontology 47 605ndash617
Zhao Y Zhu M Babcock LE Yuan L Parsley RL Peng J YangX and Wang Y 2005 Kaili Biota a taphonomic window on diversifiminuscation of metazoans from the basal Middle Cambrian Guizhou ChinaActa Geologica Sinica 79 751ndash765
400 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011

GD and Lee MSY 2008 Early Cambrian arthropods from the EmuBay Shale Lagerstaumltte South Australia 319ndash325 In I Raacutebano RGozalo and D GarciacuteaminusBellido (eds) Advances in Trilobite ResearchCuadernos del Museo Geominero 9 319ndash325
Shultz JW 2007 A phylogenetic analysis of the arachnid orders based onmorphological characters Zoological Journal of the Linnean Society150 221ndash265
Stein M 2010 A new arthropod from the Early Cambrian of North Greenminusland with a ldquogreat appendagerdquominuslike antennula Zoological Journal ofthe Linnean Society 158 477ndash500
Stuumlrmer W and Bergstroumlm J 1978 The arthropod Cheloniellon from theDevonian Hunsruumlck Shale Palaumlontologische Zeitschrift 52 57ndash81
Sutton MD Briggs DEG Siveter DJ Siveter DJ and Orr PD2002 The arthropod Offacolus kingi (Chelicerata) from the Silurian ofHerefordshire England computer based morphological reconstrucminustions and phylogenetic affinity Proceedings of the Royal Society B Biminusological Sciences 269 1195ndash1203
Suzuki Y and Bergstroumlm J 2008 Respiration in trilobites a reevaluationGFF 130 211ndash229
Vannier J GarciacuteaminusBellido DC Hu XminusX and Chen AminusL 2009 Arthrominuspod visual predators in the early pelagic ecosystem evidence from the
Burgess Shale and Chengjiang biotas Proceedings of the Royal SocietyB Biological Sciences 276 2567ndash2574
Whittington HB 1974 Yohoia Walcott and Plenocaris n gen arthropodsfrom the Burgess Shale Middle Cambrian British Columbia Geologiminuscal Survey of Canada Bulletin 231 1ndash21
Wills MA Briggs DEG Fortey RA Wilkinson M and SneathPHA 1998 An arthropod phylogeny based on fossil and recent taxaIn GD Edgecombe (ed) Arthropod Fossils and Phylogeny 33ndash105Columbia University Press New York
Wilson LA 2006 Food for thought a morphological and taphonomic studyof fossilised digestive systems from Early to Middle Cambrian taxa275 pp Unpublished PhD thesis University of Cambridge Cambridge
Zhang XminusG and Hou X 2007 Gravitational constraints on the burial ofChengjiang fossils Palaios 22 448ndash453
Zhang XminusL Han J Zhang Z Liu H and Shu D 2004 Redescription ofthe Chengjiang arthropod Squamacula clypeata Hou and Bergstroumlm fromthe Lower Cambrian Southminuswest China Palaeontology 47 605ndash617
Zhao Y Zhu M Babcock LE Yuan L Parsley RL Peng J YangX and Wang Y 2005 Kaili Biota a taphonomic window on diversifiminuscation of metazoans from the basal Middle Cambrian Guizhou ChinaActa Geologica Sinica 79 751ndash765
400 ACTA PALAEONTOLOGICA POLONICA 56 (2) 2011














