
García-Bellido, D.C., Paterson, J.R., Edgecombe, G.D., Jago, J.B., Gehling, J.G. & Lee, M.S.Y.,...
21
THE BIVALVED ARTHROPODS ISOXYS AND TUZOIA WITH SOFT-PART PRESERVATION FROM THE LOWER CAMBRIAN EMU BAY SHALE LAGERSTA ¨ TTE (KANGAROO ISLAND, AUSTRALIA) by DIEGO C. GARCI ´ A-BELLIDO* , JOHN R. PATERSON , GREGORY D. EDGECOMBE à , JAMES B. JAGO§ , JAMES G. GEHLING – and MICHAEL S. Y. LEE** , – *Departamento de Paleontologı´a, Instituto de Geologı´a Econo ´ mica (CSIC-UCM), Facultad de CC. Geolo ´ gicas, Jose ´ Antonio Novais 2. Madrid 28040, Spain; e-mail [email protected] Division of Earth Sciences, School of Environmental and Rural Science, University of New England, Armidale, NSW 2351, Australia; e-mail [email protected] àDepartment of Palaeontology, Natural History Museum, Cromwell Road, London SW7 5BD, UK; e-mail [email protected] §School of Natural and Built Environments, University of South Australia, Mawson Lakes, SA 5095, Australia; e-mail [email protected] –South Australian Museum, North Terrace, Adelaide SA 5000, Australia; e-mail [email protected] **School of Earth and Environmental Sciences, University of Adelaide, Adelaide, SA 5005, Australia; e-mail [email protected] Typescript received 8 November 2008; accepted in revised form 12 February 2009 Abstract: Abundant material from a new quarry excavated in the lower Cambrian Emu Bay Shale (Kangaroo Island, South Australia) and, particularly, the preservation of soft- bodied features previously unknown from this Burgess Shale- type locality, permit the revision of two bivalved arthropod taxa described in the late 1970s, Isoxys communis and Tuzoia australis. The collections have also produced fossils belonging to two new species: Isoxys glaessneri and Tuzoia sp. Among the soft parts preserved in these taxa are stalked eyes, diges- tive structures and cephalic and trunk appendages, rivalling in quality and quantity those described from better-known Lagersta ¨tten, notably the lower Cambrian Chengjiang fauna of China and the middle Cambrian Burgess Shale of Canada. Key words: bivalved arthropods, Isoxys, Tuzoia, Cambrian, soft-body preservation, Emu Bay Shale, Australia. T he first fossils from the Emu Bay Shale came to light in 1952 (Sprigg et al. 1954; Sprigg 1955), soon followed by the discovery of the Emu Bay Shale Lagersta ¨tte site at Big Gully. Daily (1956) announced this find on the basis of very large articulated specimens of the trilobite Redli- chia, together with Lusiatops (later assigned to Estaingia) and Isoxys n. sp. Other researchers (M. Glaessner, M. Wade and B. McGowran) collected new soft-bodied mate- rial in late 1956 (B. McGowran, pers. comm., December 2007), but the first formal descriptions of non-mineral- ized taxa were not published until Glaessner (1979) described the bivalved arthropods Isoxys communis and Tuzoia australis, the palaeoscolecid priapulid Palaeoscolex antiquus, and two animals of uncertain affinities, Myosco- lex ateles and Vetustovermis planus. The report of Anomal- ocaris from the Emu Bay Shale (McHenry and Yates 1993) was followed by the recognition of two species in that genus (Nedin 1995a), one of which was formally named (A. briggsi Nedin, 1995). Nedin (1995b, 1999) also illustrated a few unmineralized trilobitomorphs from the Emu Bay Shale Lagersta ¨tte. Briggs and Nedin (1997) rein- terpreted Myoscolex as an Opabinia-like animal and called attention to the preservation of phosphatized muscle tis- sue in Myoscolex as the oldest known to date. Subsequent interpretation of Myoscolex as an annelid (Dzik 2004) has been judged ‘unlikely’ and ‘forced’ (Conway Morris and Peel 2008). The Emu Bay Shale Lagersta ¨tte remains Australia’s most informative Burgess Shale-type locality. The exca- vation of Buck Quarry, a new locality in the Emu Bay Shale at Big Gully, has produced better preserved soft- bodied structures of several previously named taxa (Pat- erson et al. 2008), as well as new diversity, including taxa only known so far from other Cambrian Lagersta ¨t- ten such as the Burgess Shale, or Chengjiang and Kaili in China. These new fossils are being studied as part of an Australian Research Council-sponsored South Australian Museum project, of which the present con- tribution is the first taxonomic paper in a forthcoming series. [Palaeontology, Vol. 52, Part 6, 2009, pp. 1221–1241] ª The Palaeontological Association doi: 10.1111/j.1475-4983.2009.00914.x 1221
Transcript of García-Bellido, D.C., Paterson, J.R., Edgecombe, G.D., Jago, J.B., Gehling, J.G. & Lee, M.S.Y.,...

THE BIVALVED ARTHROPODS ISOXYS AND TUZOIA
WITH SOFT-PART PRESERVATION FROM THE
LOWER CAMBRIAN EMU BAY SHALE LAGERSTATTE
(KANGAROO ISLAND, AUSTRALIA)
by DIEGO C. GARCIA-BELLIDO* , JOHN R. PATERSON� ,
GREGORY D. EDGECOMBE� , JAMES B. JAGO§ , JAMES G. GEHLING–
and MICHAEL S. Y. LEE**,–
*Departamento de Paleontologıa, Instituto de Geologıa Economica (CSIC-UCM), Facultad de CC. Geologicas, Jose Antonio Novais 2. Madrid 28040, Spain; e-mail
�Division of Earth Sciences, School of Environmental and Rural Science, University of New England, Armidale, NSW 2351, Australia; e-mail [email protected]
�Department of Palaeontology, Natural History Museum, Cromwell Road, London SW7 5BD, UK; e-mail [email protected]
§School of Natural and Built Environments, University of South Australia, Mawson Lakes, SA 5095, Australia; e-mail [email protected]
–South Australian Museum, North Terrace, Adelaide SA 5000, Australia; e-mail [email protected]
**School of Earth and Environmental Sciences, University of Adelaide, Adelaide, SA 5005, Australia; e-mail [email protected]
Typescript received 8 November 2008; accepted in revised form 12 February 2009
Abstract: Abundant material from a new quarry excavated
in the lower Cambrian Emu Bay Shale (Kangaroo Island,
South Australia) and, particularly, the preservation of soft-
bodied features previously unknown from this Burgess Shale-
type locality, permit the revision of two bivalved arthropod
taxa described in the late 1970s, Isoxys communis and Tuzoia
australis. The collections have also produced fossils belonging
to two new species: Isoxys glaessneri and Tuzoia sp. Among
the soft parts preserved in these taxa are stalked eyes, diges-
tive structures and cephalic and trunk appendages, rivalling
in quality and quantity those described from better-known
Lagerstatten, notably the lower Cambrian Chengjiang fauna
of China and the middle Cambrian Burgess Shale of Canada.
Key words: bivalved arthropods, Isoxys, Tuzoia, Cambrian,
soft-body preservation, Emu Bay Shale, Australia.
T he first fossils from the Emu Bay Shale came to light
in 1952 (Sprigg et al. 1954; Sprigg 1955), soon followed
by the discovery of the Emu Bay Shale Lagerstatte site at
Big Gully. Daily (1956) announced this find on the basis
of very large articulated specimens of the trilobite Redli-
chia, together with Lusiatops (later assigned to Estaingia)
and Isoxys n. sp. Other researchers (M. Glaessner, M.
Wade and B. McGowran) collected new soft-bodied mate-
rial in late 1956 (B. McGowran, pers. comm., December
2007), but the first formal descriptions of non-mineral-
ized taxa were not published until Glaessner (1979)
described the bivalved arthropods Isoxys communis and
Tuzoia australis, the palaeoscolecid priapulid Palaeoscolex
antiquus, and two animals of uncertain affinities, Myosco-
lex ateles and Vetustovermis planus. The report of Anomal-
ocaris from the Emu Bay Shale (McHenry and Yates
1993) was followed by the recognition of two species in
that genus (Nedin 1995a), one of which was formally
named (A. briggsi Nedin, 1995). Nedin (1995b, 1999) also
illustrated a few unmineralized trilobitomorphs from the
Emu Bay Shale Lagerstatte. Briggs and Nedin (1997) rein-
terpreted Myoscolex as an Opabinia-like animal and called
attention to the preservation of phosphatized muscle tis-
sue in Myoscolex as the oldest known to date. Subsequent
interpretation of Myoscolex as an annelid (Dzik 2004) has
been judged ‘unlikely’ and ‘forced’ (Conway Morris and
Peel 2008).
The Emu Bay Shale Lagerstatte remains Australia’s
most informative Burgess Shale-type locality. The exca-
vation of Buck Quarry, a new locality in the Emu Bay
Shale at Big Gully, has produced better preserved soft-
bodied structures of several previously named taxa (Pat-
erson et al. 2008), as well as new diversity, including
taxa only known so far from other Cambrian Lagerstat-
ten such as the Burgess Shale, or Chengjiang and Kaili
in China. These new fossils are being studied as part
of an Australian Research Council-sponsored South
Australian Museum project, of which the present con-
tribution is the first taxonomic paper in a forthcoming
series.
[Palaeontology, Vol. 52, Part 6, 2009, pp. 1221–1241]
ª The Palaeontological Association doi: 10.1111/j.1475-4983.2009.00914.x 1221
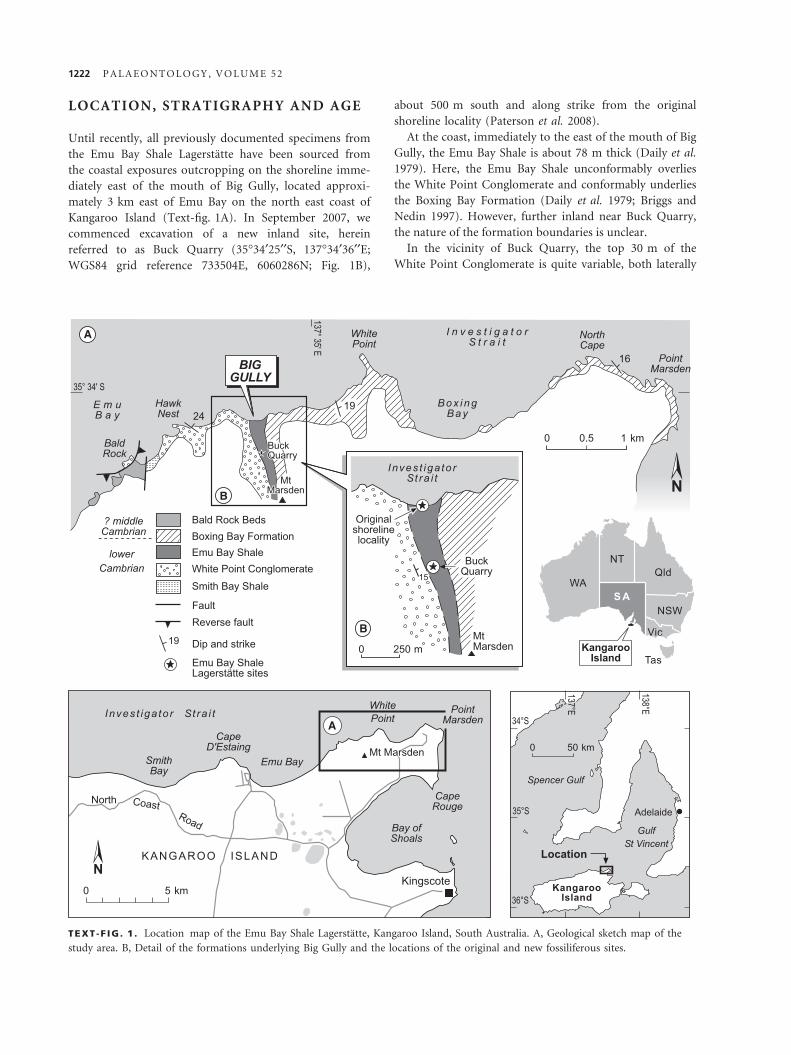
LOCATION, STRATIGRAPHY AND AGE
Until recently, all previously documented specimens from
the Emu Bay Shale Lagerstatte have been sourced from
the coastal exposures outcropping on the shoreline imme-
diately east of the mouth of Big Gully, located approxi-
mately 3 km east of Emu Bay on the north east coast of
Kangaroo Island (Text-fig. 1A). In September 2007, we
commenced excavation of a new inland site, herein
referred to as Buck Quarry (35�34¢25¢¢S, 137�34¢36¢¢E;
WGS84 grid reference 733504E, 6060286N; Fig. 1B),
about 500 m south and along strike from the original
shoreline locality (Paterson et al. 2008).
At the coast, immediately to the east of the mouth of Big
Gully, the Emu Bay Shale is about 78 m thick (Daily et al.
1979). Here, the Emu Bay Shale unconformably overlies
the White Point Conglomerate and conformably underlies
the Boxing Bay Formation (Daily et al. 1979; Briggs and
Nedin 1997). However, further inland near Buck Quarry,
the nature of the formation boundaries is unclear.
In the vicinity of Buck Quarry, the top 30 m of the
White Point Conglomerate is quite variable, both laterally
A
B
B
A
TEXT -F IG . 1 . Location map of the Emu Bay Shale Lagerstatte, Kangaroo Island, South Australia. A, Geological sketch map of the
study area. B, Detail of the formations underlying Big Gully and the locations of the original and new fossiliferous sites.
1222 P A L A E O N T O L O G Y , V O L U M E 5 2

and vertically, and is yet to be mapped and logged in
detail. It is essentially a moderately well-sorted, thin bed-
ded to massive, feldspathic to arkosic fine to medium
sandstone, which in places is quite micaceous. The rock is
well laminated at some levels. There is at least one lentic-
ular granule conglomerate horizon up to 1.5 m thick.
Sedimentary structures include tabular cross-bedding,
current parting lineation and wet sediment slumping.
At Buck Quarry, the Emu Bay Shale is about 60 m
thick, but is yet to be logged in detail. It comprises
mainly dark grey, sporadically pyritic, mudstones that at
some levels are highly fossiliferous. The newly collected
Lagerstatte material comes from approximately 7–12 m
above the base of the formation. Within the fossiliferous
interval are several coarse siltstone horizons, up to 20 cm
thick, that appear to have no internal structure, but have
planar sharp bases and may represent storm or sediment
gravity flow deposits. Some thin, fine grained sandstones,
occurring a few metres above the main fossiliferous hori-
zons, show wet sediment slumping that indicates a south-
erly depositional slope. Other units within the Emu Bay
Shale include well-sorted feldspathic sandstone horizons
up to 1 m thick and at least one conglomerate horizon,
which is a 1 to 2 m thick polymict conglomerate, at
about the middle of the formation, that is quite similar to
the polymict conglomerates within the White Point Con-
glomerate. At least one other fossiliferous horizon occurs
within mudstones just below this conglomerate. The dark
mudstones that contain the Lagerstatte appear to lens out
150–200 m south of Buck Quarry; they appear to be
replaced along strike by predominantly micaceous sand-
stones.
The basal part of the overlying Boxing Bay Formation
comprises laminated arkosic sandstones that contain trace
fossils and abundant, large-scale soft sediment slumping.
Detailed data on the faunal content and biostratigraphy
of the Emu Bay Shale have been documented previously
(e.g. Jell in Bengtson et al. 1990; Paterson and Jago 2006;
Jago et al. 2006a; Paterson et al. 2008) and will not be
reiterated herein. The Emu Bay Shale is correlated with
the traditional lower Cambrian (Series 2) Pararaia janeae
trilobite Zone of South Australia, equivalent to the early–
mid Canglangpuan Stage of China and the mid–late
Botoman of Siberia (Jago et al. 2006b; Paterson and
Brock 2007; Paterson et al. 2008).
PREVIOUS RESEARCH ON ISOXYS ANDTUZOIA
The genus Isoxys was originally described by Walcott
(1890) from carapaces recovered in the lower Cambrian
Chilhowee Group of Tennessee (USA). Several species
have been erected since 1890, mostly in the form of short
descriptions based on isolated carapaces. It is only in
recent years that evidence of its soft anatomy has come to
light, especially from the lower Cambrian Chengjiang taxa
(Shu et al. 1995; Vannier and Chen 2000) and the middle
Cambrian Burgess Shale (Garcıa-Bellido and Collins 2005;
Garcıa-Bellido et al. 2009). Discussions on the distribution
and palaeoecology of Isoxys have been offered by Williams
et al. (1996), Vannier and Chen (2000) and Vannier et al.
(2009). Research on Tuzoia is just as prolific, with over 25
nominal species, dating back to Walcott (1912). The
North American species were revised by Lieberman
(2003), who produced a list of 10 character states sug-
gested for distinguishing the various taxa, but concluding
that, because of multiple states of some of the characters,
the impossibility of ordering them and the lack of a plau-
sible outgroup, a phylogenetic analysis was premature.
After the extensive revision of the genus by Vannier et al.
(2007), which also reports the first occurrence of soft-body
preservation, fewer than ten species are presently accepted.
Based on similarities in carapace morphology and the eyes,
Isoxys and Tuzoia are considered to be closely related to
each other (Vannier et al. 2007), but their placement
within the Arthropoda remains ambiguous.
MATERIAL AND METHODS
Some of the fossils described here were from the collec-
tions of the South Australian Museum, material recovered
between the 1960s and 2007 at the mouth of Big Gully,
both from the wave-cut platform and from the cliff above
it. Most of the material, however, has been collected in
the field seasons of September 2007 and April and Octo-
ber 2008. Some fossils were prepared using a percussion
needle under a binocular microscope, revealing the parts
partially covered by matrix. Camera lucida drawings were
made of the specimens with the best preserved features. A
series of digital photographs of the fossils were taken, illu-
minated with a tungsten lamp at low and high angles to
the plane of the specimens, and the best ones selected for
illustration (usually the former, which enhances the relief
of these highly compressed fossils). Specimen numbers
refer to the palaeontological collections of the South
Australian Museum, Adelaide (prefix SAM P).
SYSTEMATIC PALAEONTOLOGY
Phylum ARTHROPODA Siebold and Stannius, 1845
Class, Order and Family UNCERTAIN
Genus ISOXYS Walcott, 1890
Type species. Isoxys chilhoweanus Walcott, 1890.
G A R C I A - B E L L I D O E T A L . : I S O X Y S A N D T U Z O I A F R O M E M U B A Y S H A L E ( A U S T R A L I A ) 1223

Other species. Isoxys acutangulus (Walcott, 1908); Isoxys carbo-
nelli Richter and Richter, 1927; Isoxys longissimus Simonetta and
Delle Cave, 1975; Isoxys communis Glaessner, 1979; Isoxys auritus
(Jiang, 1982 in Luo et al. 1982); Isoxys paradoxus Hou, 1987;
Isoxys bispinatus (Cui, 1991 in Huo et al. 1991); Isoxys zhurensis
Ivantsov, 1990; Isoxys volucris Williams et al. 1996; Isoxys curvi-
rostratus Vannier and Chen, 2000; Isoxys wudingensis Luo and
Hu, 2006 (in Luo et al. 2006). A considerable number of Isoxys
species have been described under open nomenclature from the
lower and middle Cambrian, mostly based on incomplete mate-
rial or single specimens.
Emended diagnosis. Arthropod with one pair of cephalic
appendages and a uniform series of at least 13 pairs of
biramous appendages. Long, narrow body covered almost
entirely by a bivalved, very thin unmineralized carapace.
Prominent, stalked, spherical to pear-shaped lateral eyes
protrude beyond the anterior margin of the carapace.
Each valve armed with prominent cardinal spines. Dorsal
outline straight or slightly projecting to form a weak to
well-developed cusp (small circular node may be present
below this cusp) anterior of mid-length. Ventral outline
semicircular, weakly preplete (valve is highest anterior to
its mid-length) to postplete (highest posterior to its mid-
length). Simple perimarginal features (very narrow to
more inflated rim). No flattened ⁄ concave marginal fea-
tures. Narrow to broad doublure may be present. Cara-
pace folded along the dorsal line (valves conjoined by a
narrow band of cuticle; absence of articulating hinge).
Internally, midgut glands may be present. External orna-
ment may be expressed as uniform micro-reticulation or
longitudinal striae. (Modified from Vannier and Chen
2000, p. 311).
Remarks. Isoxys usually occurs as empty carapaces,
mostly from the lower Cambrian, with some from the
middle Cambrian. It has been found in USA, western
Canada, Greenland, Spain, Siberia, south China, Austra-
lia and recently in France (Vannier et al. 2005). Isoxys
varies in size, with its overall length ranging from
c. 10 mm (I. carbonelli) to over 100 mm (I. longissimus,
I. paradoxus and, now, I. communis), although the length
of the valves, excluding the spines, usually varies from
11 mm (I. zhurensis) to 33 mm (I. auritus, I. acutangulus
and I. longissimus), but reaching over 60 mm in some of
the specimens of I. communis described herein. The
shape is especially variable with respect to the length,
thickness and curvature of the cardinal spines (Text-
fig. 2).
A
I. chilhoweanus I. acutangulus I. zhurensis
I. carbonelli
I. wudingaspisI. communisI. curvirostratus
I. longissimus
I. volucris I. bispinatus
I. paradoxus
I. glaessneriI. auritus
B C
E F
H
K
I
ML
D
G
J
TEXT -F IG . 2 . A–M, Outlines of Isoxys species described to date. They are drawn to the same scale and have been arranged
according to similarities in shape (modified from Vannier and Chen 2000 with information from the original descriptions). Specimens
oriented anterior to the left. Scale bar represents 5 mm.
1224 P A L A E O N T O L O G Y , V O L U M E 5 2

Isoxys communis Glaessner, 1979
Plates 1–2; Text-figures 2H, 3, 4I
1979 Isoxys communis Glaessner, pp. 22–23, fig. 1.
1995b Isoxys communis Glaessner, 1979; Nedin, pp. 71–72,
p. 102, pl. 4, fig. 5; pl. 10, fig. 1–2; table 6.1,
fig. 6.2.
1996 Isoxys communis Glaessner, 1979; Williams et al.,
fig. 7.3–7.4.
2008 Isoxys communis Glaessner, 1979; Paterson et al.,
fig. 2E.
Types. Holotype (SAM P21001), a carapace, and three figured
paratypes (SAM P21002, 21004, 21006).
Material. Over 120 specimens, most preserved as part and coun-
terpart, from which only five per cent preserve the entire length
of both cardinal spines, but 20 and 25 per cent preserve the
anterior and posterior cardinal spines, respectively.
Emended diagnosis. Large postplete carapace (valve high-
est posterior of mid-length) with slightly convex anterior
third of dorsal line. Valve L:H ratio generally over 2.
Strong marginal concavity under anterior cardinal spine,
but posterior concavity almost disappears with straight
posteroventral margin. Stout, upcurving anterior cardinal
spine up to twice as long as the straight to lightly droopy,
usually slim, posterior cardinal spine. Faintly reticulated
valve with swollen latero-admarginal ridge defining a
doublure occasionally present. A pair of large eyes pro-
trudes forward from the anterior margin of the carapace.
Evidence of single cephalic appendages and several trunk
appendages. Midgut glands occasionally present in up to
seven trunk segments.
Description
Carapace, eyes and appendages. The total length of the carapace
of I. communis – including cardinal spines – exceeds 100 mm in
the largest specimens (over 104 mm in a specimen with an
incomplete posterior spine). The valve length – measured as the
distance between the changes in slope at the base of cardinal
spines, see Text-figure 4I – ranges from 12.5 mm (Pl. 1, fig. 4)
to 67 mm (Pl. 1, fig. 2), with an average of 30 mm, and has a
height between 5.5 and 28 mm. The valve L:H ratio is an aver-
age of 2.15, with a maximum of 2.91 and a minimum of 1.33,
and with a very good fit (R2 = 0.9183, Text-fig. 3A). The great-
est height of the valve is generally behind the mid-length (post-
plete) or near it (amplete), with an occasional preplete
specimen. The anterior cardinal spine is approximately 145–200
per cent of the length of the posterior one, and together they
constitute about 40–50 per cent of the total length of the cara-
pace. The length of the cardinal spines in complete specimens
varies from 6 to 27 mm for the anterior one, and 3.5 to
19.4 mm in the posterior one. In a growth series, the anterior
and posterior spines show positive allometry, increasing in
length of about one-third and one-fourth, respectively, when
compared with that of the valve, and have fit values of c. 0.8
(Text-fig. 3B–C). When the texture of the carapace is visible, a
fine reticulate pattern is obvious, with polygons 0.5–1 mm across
(Pl. 1, figs 5–7). Well-preserved specimens also show evidence of
an incomplete perimarginal ridge (Pl. 1, figs 2–4). Most speci-
mens in the SAM collections are preserved in lateral orientation,
but some are preserved in dorsal aspect (Pl. 1, figs 7–8), but not
in the ‘butterfly’ position observed for other Isoxys species (e.g.
Briggs et al. 1994, fig. 102; Shu et al. 1995, fig. 1C; Williams
et al. 1996, fig. 6.5). One specimen shows a faint median line
that may correspond to a weak hinge. The carapace is strongly
concave under the anterior spine where the eyes are located
(Pl. 1, figs 2–5; Pl. 2, fig. 2).
The most conspicuous feature in the specimens of I. commu-
nis with soft parts is the presence of a pair of large, spherical
eyes (Pl. 1, figs 10–11; Pl. 2, figs 2–3). They were probably
stalked, but only very faint traces of this remain. The eyes are 2–
5 mm in diameter, with an average of 3.25 mm, and protrude
forward from beneath the strongly concave frontal edge of the
carapace, just under the anterior cardinal spine. About half of
the specimens in the collections preserve eyes, but the quality of
preservation varies, from a pair of well-defined circular struc-
tures to faint round traces in the appropriate position. None
preserve definitive evidence of ommatidia (a pitted surface is
occasionally developed; Pl. 1, fig. 10).
A single specimen in the collection shows a cephalic append-
age, which is almost 9 mm long and 1 mm thick, originating
from the anterior part of the body, just under the insertion of
the eyes (Pl. 2, figs 2–3). The appendage is curved upwards, but
there are no traces of the podomere boundaries, which makes
the determination of number of segments in this appendage
impossible.
A few specimens preserve faint impressions of the trunk
appendages under the carapace, mostly in the form of long, nar-
row lines or grooves (Pl. 2, figs 2–3). These would correspond
to the endopods observed in other Isoxys species.
Digestive system. The only recognizable feature in the alimentary
canal of Isoxys communis is serially repeated, three-dimensional
structures (Pl. 2, figs 4–6), which have been described in some
other Burgess Shale arthropods, particularly Leanchoilia, as mid-
gut glands or hepatopancreas (Butterfield 2002; Garcıa-Bellido
and Collins 2007), as well as in an unknown arthropod from the
Chinese Kaili Lagerstatte (Zhu et al. 2004). These metameric
ovoid structures are typically 1 by 1.8 mm in size, and in the
best-preserved specimen, are multilobular, with the largest lobes
measuring 1.5 mm in diameter (Pl. 2, figs 4–5). Midgut glands
are present in seven trunk segments and present a granular tex-
ture (Pl. 2, figs 4–5), perhaps replicating the internal micro-
spheres recognized in thin sections of the midgut glands of
Leanchoilia (Butterfield 2002). These microspheres have been
regarded as enzyme or lipid vesicles (Butterfield 2002) or as pos-
sible bacteria (Vannier and Chen 2002).
Two specimens show an oval-shaped black fibrous material
obliquely positioned in the central area of the carapace (Pl. 2,
figs 7–9). This material is arranged in wide bundles of thin fibres
G A R C I A - B E L L I D O E T A L . : I S O X Y S A N D T U Z O I A F R O M E M U B A Y S H A L E ( A U S T R A L I A ) 1225

and closely resembles the phosphatized muscle fibres of the co-
occurring Myoscolex ateles (see Briggs and Nedin 1997, e.g.
fig. 8.6).
Comparison with other species. The carapace of Isoxys communis
is average in terms of cardinal spine length (Text-fig. 2), but
stands out in valve length, well above most other species. Its
overall length is also greater than most Isoxys species, the largest
specimens of I. communis rivalling the holotype of I. longissimus
from the Burgess Shale (105 mm, Simonetta and Delle Cave
1975, p. 6, fig. 6F) in maximum length. Although cardinal spines
are very useful as diagnostic qualitative characters, we believe
that they should be employed with caution, as they may not be
preserved in their whole length because of their fragility. Thus, it
might be better to exclude them as a quantitative character from
size comparisons between Isoxys species, and preferably base this
on more frequently preserved parameters such as valve length
and height. By that measure, most Isoxys species have L:H ratios
around two, with I. longissimus nearing three (Garcıa-Bellido
et al. 2009), and I. zhurensis (Ivantsov 1990) as little as 1.5.
Based on general outline, the closest species is I. curvirostratus
(Text-fig. 2G), but the Chinese species has shorter cardinal
spines, especially the posterior one, and a striated ornament very
different from the reticulated pattern of I. communis. This retic-
ulated pattern is only shared with I. auritus (Vannier and Chen
2000), which has a finer reticulation (0.2–0.35 mm in diameter),
other species having smooth, dimpled or striated ornaments.
This reticulation only becomes apparent in the best-preserved
specimens and when the true texture of the carapace is visible,
because a calcite layer tends to develop on internal moulds of
the valves, especially in the original outcrop at the wave-cut
platform (Pl. 2, fig. 9; Pl. 3, fig. 6; Pl. 4, fig. 6).
The eyes of I. acutangulus (Garcıa-Bellido et al. 2009),
I. auritus (Shu et al. 1995, fig. 4A), I. curvirostratus (Vannier and
Chen 2000, figs 3A, 4D) and Isoxys sp. (Briggs et al. 2008, fig.
14) do not differ significantly in shape or position from those in
I. communis, but are considerably smaller (1–2.7 mm in diame-
ter) than those of the latter (up to 5 mm). This could be par-
tially attributable to the larger size of I. communis.
Regarding the appendages, the cephalic appendage preserved
in I. communis (Pl. 2, figs 2–3) is reminiscent of those described
in the Burgess Shale’s I. acutangulus (Garcıa-Bellido et al. 2009)
and Chengjiang’s I. auritus (Vannier et al. 2009) as well as a
small specimen assigned to the latter species with question (Hu
2005). However, it is slightly shorter than the ones in those spe-
cies, and there is no evidence of the robust spines of the Cana-
dian species or the numerous thin spines displayed by the
Chinese material. The trunk appendages of I. communis are posi-
tioned similarly to those in the species mentioned above, where
the rarely preserved endopods in the Burgess Shale material also
appear to be thin and elongate. However, we lack evidence, as
yet, of the more commonly recognizable paddle-like exopods,
fringed with a series of setae, as are found in I. acutangulus and
the Chinese taxa (I. auritus and I. curvirostratus, Vannier and
Chen 2000).
The size of the midgut glands in I. communis falls within the
range described for I. acutangulus (Garcıa-Bellido et al. 2009) of
0.5–2.6 mm in diameter, and likewise have a granular texture.
However, that species has midgut glands present on up to 10 (vs
seven) trunk segments and are disposed in pairs.
Occurrence. Wave-cut platform, coast cliff and Buck Quarry,
lower Cambrian Emu Bay Shale, Kangaroo Island, South Australia.
Isoxys glaessneri sp. nov.
Plate 3; Text-figures 2E, 3
Derivation of name. In honour of Martin F. Glaessner, the first
author to publish on the Emu Bay Shale Lagerstatte.
Holotype. SAM P14651 (Pl. 3, fig. 1).
Paratypes. SAM P14654, P14667, P14668, P14710, P14715,
P14724, P14728, P14729, P14740, P14992, P14995, P15091,
P15100, P15113, P15116, P15119, P15129, P15181, P15197,
P19994, P41724, most preserved as part and counterpart.
Diagnosis. Small species with smooth carapace. Dorsal
valve line straight or slightly domed in the centre and
semicircular ventral outline; valve L:H ratio generally
below two and weakly preplete to amplete (valve is high-
est around its mid-length), with faint or absent doublure.
Marginal concavities under cardinal spines subequal.
Combined length of cardinal spines 16–26 per cent of
total length, with anterior spine more heavily built and
EX PLANATION OF PLATE 1
Figs 1–11. Isoxys communis Glaessner, 1979, Emu Bay Shale, Big Gully, Kangaroo Island, South Australia. 1, SAM P21001 A, holotype;
both cardinal spines are incomplete, and it presents the characteristic pink hue of specimens collected from the type locality. 2,
SAM P14634, largest specimen in the collections, posterior spine lost. 3, SAM P14704 B, well-preserved small specimen with eyes
(arrow) and a strong lateral ridge, running parallel to valve margin, possibly defining the edge of the doublure. 4, SAM P15078 B,
smallest specimen. 5, SAM P14716 A, almost complete lateral specimen showing the fine reticulate ornament. 6, SAM P14706,
detail of the fine reticulate ornament of the valves. 7, SAM P15291B, dorsoventrally compressed complete specimen. 8, SAM
P14801, incomplete dorsally preserved specimen, with evidence of a faint hinge line. 9, SAM P14643, detail of droopy posterior
cardinal spine. 10, SAM P14658 A, detail of very well-preserved eye, faintly pitted texture may not correspond to ommatidia. 11,
SAM P14655A, detail of anterior region with two eyes preserved. All specimens oriented with anterior to the right, except Figure
1. All scale bars represent 5 mm.
1226 P A L A E O N T O L O G Y , V O L U M E 5 2

GARCIA-BELLIDO et al., Isoxys communis
1 2
3 4
5 7
6
9 10 11
8
PLATE 1
A
B
C
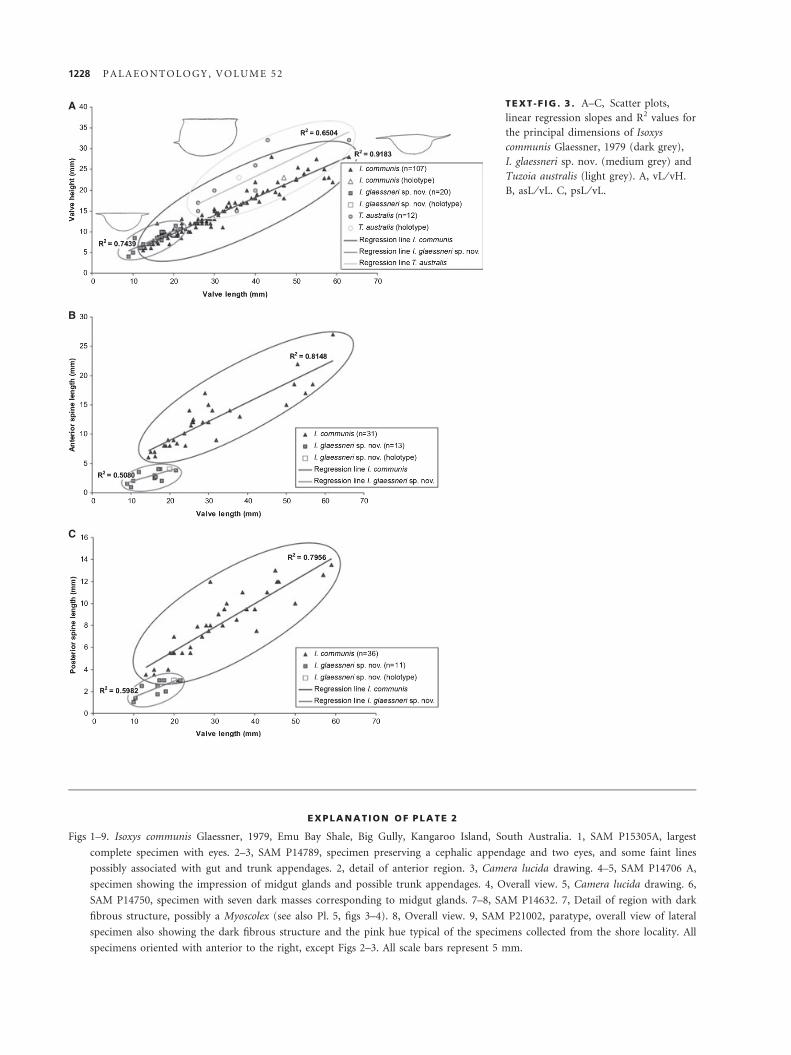
TEXT -F IG . 3 . A–C, Scatter plots,
linear regression slopes and R2 values for
the principal dimensions of Isoxys
communis Glaessner, 1979 (dark grey),
I. glaessneri sp. nov. (medium grey) and
Tuzoia australis (light grey). A, vL ⁄ vH.
B, asL ⁄ vL. C, psL ⁄ vL.
EX PLANATION OF PLATE 2
Figs 1–9. Isoxys communis Glaessner, 1979, Emu Bay Shale, Big Gully, Kangaroo Island, South Australia. 1, SAM P15305A, largest
complete specimen with eyes. 2–3, SAM P14789, specimen preserving a cephalic appendage and two eyes, and some faint lines
possibly associated with gut and trunk appendages. 2, detail of anterior region. 3, Camera lucida drawing. 4–5, SAM P14706 A,
specimen showing the impression of midgut glands and possible trunk appendages. 4, Overall view. 5, Camera lucida drawing. 6,
SAM P14750, specimen with seven dark masses corresponding to midgut glands. 7–8, SAM P14632. 7, Detail of region with dark
fibrous structure, possibly a Myoscolex (see also Pl. 5, figs 3–4). 8, Overall view. 9, SAM P21002, paratype, overall view of lateral
specimen also showing the dark fibrous structure and the pink hue typical of the specimens collected from the shore locality. All
specimens oriented with anterior to the right, except Figs 2–3. All scale bars represent 5 mm.
1228 P A L A E O N T O L O G Y , V O L U M E 5 2

GARCIA-BELLIDO et al., Isoxys communis
1
2 3
ca
ta?
rele
asg? rv
lv
ta rv
mgg
4 5
76
9
8
PLATE 2

slightly longer (100–140 per cent) than posterior cardinal
spine. Cardinal spines straight or posterior one slightly
tilted downwards. A pair of large eyes protrudes from
below the anterior margin of the carapace.
Description. The total length of the carapace ranges from 12 to
28 mm. Valve length ranges from 9 to 21.6 mm, with a mean
length of 15.7 mm. Valve height varies from 4 to 11.4 mm.
These values follow a linear regression of R2 = 0.7439 (Text-
fig. 3A), with L:H ratio averaging 1.87, with a maximum of 2.25
and a minimum of 1.24. The anterior cardinal spine is 1–4 mm
long, while the posterior varies from 1 to 3 mm in length. The
scatter graphs of cardinal spine length against valve length show
a regression line with a poor R2 value of c. 0.51 and 0.60 (Text-
fig. 3B–C). The valve occasionally shows a faint perimarginal line
(Pl. 3, figs 2, 5, 9). The only soft-part feature recognized to date
in this species is the presence of eyes. Seven of the specimens in
the type collection preserve them; they are 2 mm in diameter,
except in the smallest specimen, where they have a diameter of
1 mm (Pl. 3, figs 3–4).
Comparison with other species. This species of Isoxys is among
the smallest in the genus, very similar in valve length to I. volu-
cris from Greenland (excluding cardinal spines), and slightly lar-
ger than I. zhurensis from Siberia and I. carbonelli from Spain.
However, the type and only specimen of the latter seems to be
partially covered by matrix (Richter and Richter 1927, fig. 1).
The outline of I. glaessneri sp. nov., with subequal cardinal
spines, straight dorsal line and almost identical anteroposterior
symmetry, is considerably different from the co-occurring
I. communis (Text-fig. 2). Of the species known to date, the clos-
est in general shape are I. chilhoweanus from the United States,
I. zhurensis from Siberia and I. auritus from China, the former
two having less concavity under the spines and the latter having
a densely reticulated texture. The regression line in valve L:H
values for I. glaessneri follows the same trend of that in I. com-
munis (Text-fig. 3A), but only overlaps in the lowest range of
the latter, extending beyond it into lower values. Specimens of
I. glaessneri and I. communis in the area of size overlap consis-
tently differ in cardinal spine length and shape, as well as valve
outline and texture, the smallest specimens of the latter already
showing their characteristic reticulate pattern (Pl. 1, figs 3–4).
The scatter points for cardinal spine length to valve length values
in I. glaessneri do not overlap with those of I. communis, and
their regression lines have shallower slopes than those in the lar-
ger species (Text-figs 3B–C), suggesting that the spines grow
slower during ontogeny. The smooth texture of I. glaessneri,
present in all its representatives, is also shared with many of the
other species in the genus, but the lack of texture in some of the
latter could be a taphonomic artefact.
The other species with soft-part preservation, I. auritus (Shu
et al. 1995) and I. curvirostratus (Vannier and Chen, 2000) from
the lower Cambrian of China, as well as I. acutangulus (Garcıa-
Bellido et al. 2009), also have large subspherical eyes, similar to
those in I. glaessneri.
Occurrence. Wave-cut platform, coast cliff and Buck Quarry,
lower Cambrian Emu Bay Shale, Kangaroo Island, South
Australia.
Class UNCERTAIN
Order TUZOIDA Simonetta and Delle Cave, 1975
Family TUZOIIDAE Raymond, 1935
Genus TUZOIA Walcott, 1912
Type species. Tuzoia retifera Walcott 1912, middle Cambrian
Burgess Shale, British Columbia, Canada.
Other species. More than 20 different species have been described
since the genus was established. However, Vannier et al. (2007)
recognized only seven valid species, suggesting that many previ-
ously described species were based on misinterpretation of diag-
nostic marginal and frill spines because of compaction and the
different burial orientations. The valid species are Tuzoia retifera
Walcott, 1912; T. burgessensis Resser, 1929; T. canadensis Resser,
1929; T. polleni Resser, 1929; T. australis Glaessner, 1979; T. gun-
theri Robison and Richards, 1981; T. bispinosa Yuan and Zhao,
1999, and two distinct Tuzoia spp. (Chlupac and Kordule 2002;
Vannier et al. 2007). A new species – T. tylodesa – has since been
described from the early Cambrian of China (Luo et al. 2006).
Surprisingly, no Tuzoia specimens have yet been described from
the lower Cambrian Chengjiang fauna.
Diagnosis. See Vannier et al. (2007, pp. 458–459).
Remarks. Tuzoia is almost always found in the form of
empty carapaces, with most occurrences from the middle
EX PLANATION OF PLATE 3
Figs 1–10. Isoxys glaessneri sp. nov., Emu Bay Shale, Big Gully, Kangaroo Island, South Australia. 1, SAM P14651, holotype, lateral
specimen. 2, SAM P14728, paratype, largest complete specimen. 3–4, SAM P 14667, paratype, smallest specimen, with pair of
eyes. 3, Overall view. 4, Camera lucida drawing showing slightly uplifted posterior cardinal spine. 5, SAM P14994, paratype, small
specimen with well preserved cardinal spines. 6, SAM P14668, paratype, lateral specimen partially covered by a thin, pinkish layer
of calcite. 7, SAM P14654, paratype, specimen with slightly uplifted posterior cardinal spine and preserving one eye (arrow). 8,
SAM P15129, paratype, lateral specimen showing low L:H ratio. The reddish stain under the cardinal spine on the left might be
mistaken for an eye, but morphology of the cardinal spines shows that this is the posterior region. 9, SAM P15197, paratype,
lateral specimen with a slightly uplifted posterior cardinal spine. 10, SAM P14992, paratype, lateral specimen preserving an eye
(arrow). Specimens 2–7 oriented anterior to the left. All scale bars represent 5 mm.
1230 P A L A E O N T O L O G Y , V O L U M E 5 2

GARCIA-BELLIDO et al., Isoxys glaessneri
1 2
3 4
5
7
6
9 10
8
PLATE 3

Cambrian of North America (Canada and USA), two
from North and South China and an unnamed species
from Bohemia. However, the genus originated in the early
Cambrian, from which there are three species, from
North America (Canada and USA) and Australia and an
unnamed species from South China. Tuzoia varies consid-
erably in size, valve length ranging from 15 mm in the
smallest specimens of T. bispinosa (Vannier et al. 2007) to
an estimated 180 mm in Tuzoia sp. from Bohemia
(Chlupac and Kordule 2002). The overall outline is more
constant than in Isoxys, but varies in the concavity under
the anterior cardinal process, size of the reticulation and,
particularly, the number and stoutness of spines in the
valve margins and lateral ridge, from the almost bare
T. australis (described below) to the extremely spinose
T. canadensis (Vannier et al. 2007, fig. 25.3).
Tuzoia australis Glaessner, 1979
Plate 4; Plate 5, figures 1–5; Text-figures 3A, 4A–F, H
1979 Tuzoia australis Glaessner, pp. 23–24, fig. 2.
1995b Tuzoia australis Glaessner, 1979; Nedin, p. 72, 103;
pl. 4, fig. 6; pl. 10, fig. 3–4; table 6.1, fig. 6.2.
2007 Tuzoia australis Glaessner, 1979; Vannier et al.,
p. 466; fig. 26.
2008 Tuzoia australis Glaessner, 1979; Paterson et al.,
fig. 2E.
Types. Holotype (SAM F17451), a carapace, and one figured
paratype (SAM P21007).
Material. Sixteen complete or almost complete specimens, most
preserved as part and counterpart, and some fragments.
Emended diagnosis. Small species of Tuzoia with postplete
carapace, and only one marginal spine (posteroventral)
and no evidence of dorsal or ridge spines. Dorsal line
straight or slightly convex. Posterior margin usually sur-
passing posterior cardinal process, which is never the case
with anterior region. Low relief lateral ridge parallel to
dorsal line and located half way, or slightly lower, down
the valve. Carapace length from 22 to 63 mm, and L:H
ratio of 1.73 (n = 9). Cardinal processes subequal; pos-
teroventral spine medium sized, pointed and directed
backwards. Anterior and posterior concavities under car-
dinal processes variable between specimens, but similar in
size and shape. Average a angle of c. 23 degrees. Irregular
reticulate pattern, with average maximum size of poly-
gons 1.53 mm across.
Description. The holotype of Tuzoia australis (Pl. 4, fig. 1) is
36 mm in length and 23 mm in height; the specimens in the
collection have an average length of 37.2 mm and average height
of 22.2 mm (see Text-fig. 4H), although they range from 15 to
36 mm in height. The regression line for the twelve specimens
where both these parameters were available has an R2 value of
0.6504 (Text-fig. 3A). The L:H ratio varies between 1.34 and
2.37. Values of a could only be measured in seven specimens
(Text-fig. 4A–F, plus the paratype in Glaessner 1979, fig. 2B),
and they range between 20 and 25 degrees. The general outline
is consistent (Text-fig. 4), but some specimens (Text-fig. 4D–E)
show a slight bulge in the margin between the posterior process
and postero-ventral spine. There is an odd specimen (SAM P
15142), which, oriented based on the typical characters of the
species (postplete outline and posterior margin extending well
beyond the posterior cardinal process), has its marginal spine
located in the anterior margin (Text-fig. 4C). The angle between
the dorsal line and the insertion of the spine is 28 degrees, also
falling outside the typical a value of the species. We cannot
ascertain whether this small specimen is malformed or that this
is related to ontogeny, but not observed in an even smaller spec-
imen (Text-fig. 4F).
The collection has two types of specimens in dorsal view:
those with valves splayed out, a.k.a. ‘butterfly’ configuration
(Pl. 4, fig. 3; Pl. 5, fig. 3) – where the edge of the valves corre-
sponds to the ventral margins – and dorsoventrally compressed
(dvc) specimens (Pl. 4, fig. 4; Pl. 5, fig. 4) – where the edge of
the valves corresponds to the lateral ridge – very rarely co-occur-
ring, as in a specimen of Tuzoia canadensis (Vannier et al. 2007,
fig. 14.2). The latter would be expected to show possible spines
in the lateral ridge if these had been present. The ridge is notice-
able in both dvc and lateral specimens (Pl. 4, figs 2–3, 5; Pl. 5,
figs 1–2), and it is low in relief and located slightly above the
insertion point of the posteroventral spine, which in turn is
about two-thirds down the height of the valve.
EX PLANATION OF PLATE 4
Figs 1–6. Tuzoia australis Glaessner, 1979, Emu Bay Shale, Big Gully, Kangaroo Island, South Australia. 1, SAM P17451A, holotype,
lateral specimen (see Text-fig 4A). Anterior cardinal process and posterior ventral spine of underlying valve indicate that the
valves are slightly displaced. 2, SAM P14679, incomplete lateral specimen, with eyes. 3, SAM P14674A, specimen in ‘butterfly’
configuration, splayed along its dorsal hinge (see Text-fig 4B). Mid-anterior region (arrow) with faint traces of dark fibrous
material, possibly Myoscolex remains. 4, SAM P14677A, specimen with eyes (arrows), dorsoventrally compressed and lacking a
split along its dorsal hinge. 5, SAM P15142A, small, lateral specimen, with ventral spine in anterior margin of valve (see Text-
fig 4C). 6, SAM P14697A, ‘butterfly’ specimen partially covered by a calcite layer, detail of eyes (arrows). Notice small areas of
dark fibrous material over the calcite (see also counterpart on Pl. 5, fig. 3). All specimens oriented anterior to the right. All scale
bars represent 5 mm.
1232 P A L A E O N T O L O G Y , V O L U M E 5 2

GARCIA-BELLIDO et al., Tuzoia australis
1
2
3
4
5 6
PLATE 4

The reticulate pattern covers the whole carapace and has a
variable mesh size, with the smallest polygons occurring along
the margins, and the largest on the central region, particularly
above the lateral ridge, where they range from 0.8 to 2 mm in
diameter –averaging 1.53 mm – but reach up to 4 mm in one of
the largest specimens (Pl. 5, fig. 2). Polygons vary in number of
sides from 4 to 8, with most having five or six sides.
Three specimens of Tuzoia australis preserve a pair of eyes
(Pl. 4, figs 2, 4, 6), two in dorsal view and one in lateral view.
The eyes are oval, 3–4 by 2.5 mm in size, and abut the carapace
or are even partially hidden under it. By comparison with extant
analogues, these large, stalked eyes were probably compound
eyes, but no ommatidia can be recognized in our fossils.
As in Isoxys communis (see above), some specimens (Pl. 4, fig.
3; Pl. 5, figs 3–4) show oval patches of a thin layer of dark
fibrous material. These are all dorsal specimens, and the patch is
placed in the centre of the anterior region, crossing obliquely
from one valve to the other. The other dvc specimen (Pl. 5, fig.
4) also shows evidence of this material, in the form of very thin
grooves only obvious under microscope with very low angle light.
The fact that this structure is never bilaterally symmetrical and
that it is absent in all lateral specimens leads us to believe that it
is not part of the original internal anatomy of the animal, but
rather a possible superimposed feature. As with I. communis, the
patches on T. australis more closely resemble the phosphatized
muscle fibres of Myoscolex ateles (cf. Briggs and Nedin 1997).
Comparison with other species. Our data on Tuzoia australis
(Text-fig. 3A) indicate that it falls in the lower third of the size
range of the genus (Vannier et al. 2007, fig. 26), beside the
smallest specimens of T. burgessensis, T. bispinosa and T. retifera
from the middle Cambrian Burgess Shale. The L:H ratio average
of T. australis (1.73) falls slightly above the ratios of all the other
species (1.3–1.6, according to Vannier et al. 2007), making it the
shallowest in the genus. The general outline differs from the rest
in that it has only one spine in its posterior margin and none
dorsally, ventrally (see Text-fig. 4G) or in the lateral ridge. The
concavity under the anterior process (‘notch’ in Vannier et al.
2007) is also weak or almost nonexistent. The average a angle,
however, falls within the typical values (20–40 degrees) indicated
by Vannier et al. (2007), albeit on the lower end (22.8 degrees).
Regarding the soft-part features, the only possible comparison
can be made with the three specimens from the Burgess Shale
reported by Vannier et al. (2007). They describe a specimen with
an eye stalk (T. burgessensis) and two with a pair of eyes (Tuzoia
sp.) among more than 330 specimens examined. This makes the
ratio of specimens with eyes in T. australis (25 per cent) consid-
erably higher than that found by Vannier et al. (2007). However,
their eyes are larger (5–5.5 mm in diameter) than those in
T. australis, and the eyes in one of the specimens of Tuzoia sp.
from the Burgess Shale (Vannier et al. 2007, fig. 23.4) are pre-
served with a thin reticulate pattern reminiscent of the visual
cortex of compound eyes in many extant crustaceans. The eyes
in the Australian species also lack the long stalks of the Cana-
dian material (up to 15 mm; Vannier et al. 2007, figs 23.1–4,
23.10 and 24.1, 3), looking more like those in their other speci-
men (Vannier et al. 2007, figs 23.6–7 and 24.2). Vannier et al.
(2007, fig. 23.5) describe a fine 1.5-mm long structure appearing
from the base of the eye stalk, which they consider segmented
and interpret as a possible fragment of antenna or antennule.
One of the specimens of Tuzoia sp. with soft parts in Vannier
et al. (2007, p. 456) presents ‘enigmatic linear or fibrous struc-
tures’, which are described as ‘thin and parallel lines preserved as
faint imprints’. Unfortunately, these are barely recognizable in the
corresponding illustration (Vannier et al. 2007, fig. 24.1). How-
ever, they interpret them as possible appendage setae. If these
enigmatic lines correspond to the same structure, the Australian
specimens show that they are not related to trunk appendages.
The other Tuzoia sp. with eyes shows evidence of putative midgut
glands (Vannier et al. 2007, figs 23.8–9), resembling those
described in Leanchoilia (Garcıa-Bellido and Collins 2007) and
Isoxys (see above), which have not been recognized in T. australis.
Occurrence. Wave-cut platform, coast cliff and Buck Quarry,
lower Cambrian Emu Bay Shale, Kangaroo Island, South Australia.
Tuzoia sp.
Plate 5, figures 6–7; Plate 6
Material. Six specimens, all but one fragmentary, and three pre-
served with part and counterpart.
Description. Large species of Tuzoia with unknown marginal or
fringe spines. Dorsal line is straight, as preserved in a partially
complete lateral specimen (Pl. 5, fig. 6). The most complete
specimen has a vL of 135 mm and a vH of 62 mm (Pl. 6, fig.
1), but the species might have reached a larger size, as a frag-
ment – oriented according to the position of the preserved
EX PLANATION OF PLATE 5
Figs 1–5. Tuzoia australis Glaessner, 1979. Emu Bay Shale, Big Gully, Kangaroo Island, South Australia. 1, SAM P15292B, almost
complete lateral specimen (see Text-fig 4E). 2, SAM P14719B, lateral specimen with strongly displaced valves outlined by dashed
lines (see Text-fig 4D). 3, SAM P14697B, ‘butterfly’ specimen with dark fibrous structure which may correspond to possible
remains of a Myoscolex ateles obliquely superposed over its mid-anterior region. 4, SAM P15136A, dvc specimen with dark fibrous
structure material obliquely superposed over its mid-anterior region. 5, SAM P14674B, detail of reticulate pattern adjacent to the
lateral ridge. 6–7, Tuzoia sp., Emu Bay Shale, Big Gully, Kangaroo Island, South Australia.
Figs 6–7. Tuzoia sp. 6, SAM P14983A, most complete specimen in lateral aspect, with dorsal and anterior margins preserved. 7, SAM
P15147A, detail of regularly tessellated reticulate pattern. Specimens oriented anterior to the right, except 1–2.
All scale bars represent 5 mm.
1234 P A L A E O N T O L O G Y , V O L U M E 5 2

GARCIA-BELLIDO et al., Tuzoia australis and Tuzoia sp.
1 2
3 4
5
76
PLATE 5

ventral margin – is more than 100 mm long by 62 mm high
(Pl. 6, fig. 5). The only preserved cardinal process (anterior) is
small with a slight concavity under it. The anterior margin, how-
ever, extends well beyond the anterior cardinal process, which is
also evident in the ‘butterfly’ specimen (Pl. 6, fig. 1). The alter-
native interpretation would be that this is the posterior margin,
but there is no evidence of marginal spines. The reticulate pat-
tern is very dense and uniform, with maximum size of polygons
1.1–2.0 mm across and an average of 1.58 mm. The mesh size
does not diminish close to the lateral ridge (Pl. 6, figs 3–4), but
does so slightly in the marginal areas (Pl. 6, figs 1, 6). The most
complete specimen shows a 5 mm wide depression along the
whole margin of its preserved side, including the area behind
the eyes (Pl. 6, figs 1–2), which may correspond to a doublure.
Only one specimen preserves eyes, and these are located medially
but slightly displaced from the hinge line (Pl. 6, fig. 1). These
oval eyes are 9 by 7 mm and have a 2-mm thick cortex in their
anterior surface (Pl. 6, fig. 2).
Comparison with other species. This form of Tuzoia falls within
the top of the range for the genus, among the largest specimens
of T. burgessensis and T. retifera (Vannier et al. 2007, fig. 26),
almost reaching the preserved 140 mm of Tuzoia sp. from Bohe-
mia (Chlupac and Kordule 2002, fig. 1). However, it should be
noted that the holotypes, figured in Vannier et al. (2007, figs
16.1 and 17.1), of T. guntheri (L = 77.3 mm, H = 58.6 mm) and
T. polleni (L = 65 mm, H = 57.5mm) have been erroneously
placed in their H:L plot (Vannier et al. 2007, fig. 26), and are,
thus, noticeably smaller than Tuzoia sp. from Buck Quarry. The
dimensions of the smallest, yet incomplete, specimen of this spe-
cies (H > 44 mm) are greater than the largest T. australis
(H = 36 mm), and the regular pattern of the former is consider-
ably different from the irregular reticulation seen in the latter
(see Pl. 5, figs 5, 7). However, variation in number of sides of
the polygons is equivalent to that of other species (4–8), with
hexagons also being the most common. The eyes in Tuzoia sp.
were more than twice as large to those in Tuzoia australis and
show evidence of a visual cortex, but were located in a similar
position, at least in dorsoventral specimens (Pl. 4, fig. 6; Pl. 6,
fig. 1). They also lacked the long stalks present in the Canadian
species (Vannier et al. 2007). The most complete specimen of
Tuzoia sp. shows a marginal depression around its whole perim-
eter that has never been recognized in T. australis. In the light of
the quality and quantity of soft-bodied evidence shown here for
both Australian species of Tuzoia, the presence of presumed
antennae should be regarded with caution, while the statement
in the generic diagnosis that it has ‘eye stalks at least three times
longer than the eye itself’ (Vannier et al. 2007, p. 458) should be
removed altogether.
Occurrence. Buck Quarry, lower Cambrian Emu Bay Shale, Kan-
garoo Island, South Australia.
MODE OF LIFE
The presence of large stalked eyes in Isoxys probably indi-
cates an active predatory habit. They resemble the eyes of
some Burgess Shale arthropods thought to have been pre-
dators, such as Opabinia and Odaraia (Briggs and Whit-
tington 1985), with good vision for locating and pursuing
their prey. It is worth noting that Odaraia has been
regarded as having the largest eyes of any Cambrian
arthropod relative to its size, even including Anomalocaris
(Briggs and Whittington 1985). The new data show that
Isoxys, including both the Australian and Canadian taxa
(Garcıa-Bellido et al. 2009), can now be recognized as
one of the Cambrian arthropods with the largest eyes in
relation to its body size, with those of I. communis
exceeding 18 per cent of the valve length in a number of
specimens. The large eyes in Tuzoia sp. from the Burgess
Shale and in the two Australian taxa also seem to point
to the ability to detect prey or predators.
The frontal appendage evident from Canadian, Chi-
nese and Australian Isoxys had an obvious raptorial
function and is likely to have been used for both captur-
ing prey and carrying food to the mouth region in a
manner similar to the pedipalps of fossil and recent
arachnids (Vannier et al. 2009).
Midgut glands are known in some extant arthropods
(e.g. chelicerates and remipede crustaceans) where they
are used in the digestion and absorption of food, and the
storage of food obtained infrequently. Recognition of the
midgut glands in Isoxys acutangulus as an integral part of
its alimentary system points to a similar function. The
presence of metameric midgut glands in Leanchoilia and
other fossil arthropods has been taken as evidence that
they were carnivores or scavengers (Butterfield 2002; Van-
nier and Chen 2002; Zhu et al. 2004), and the same
would apply to Isoxys, and possibly Tuzoia.
EX PLANATION OF PLATE 6
Figs 1–6. Tuzoia sp., Emu Bay Shale, Big Gully, Kangaroo Island, South Australia. 1, SAM P15333B, most complete specimen
(‘butterfly’ configuration), with explanatory drawing in bottom left corner. Notice the doublure, particularly around the margin
of the left valve. 2, SAM P15333A, detail of the anterior region with the pair of eyes, showing a cortical area (arrows). 3–4, SAM
P14652B, large fragment preserving part of the lateral ridge. 3, Overall view. 4, Detail of lateral ridge and surrounding
homogeneously small tessellate pattern. 5–6, SAM P15148B, large fragment preserving part of the marginal pattern. 5, Overall
view. 6, Detail of margin with polygons becoming smaller closer to valve edge. Specimen in 1–2 oriented anterior to the right,
unknown in others. All scale bars represent 5 mm.
1236 P A L A E O N T O L O G Y , V O L U M E 5 2

GARCIA-BELLIDO et al., Tuzoia sp.
1 2
3
5
4 6
PLATE 6

The reticulation recognized in Tuzoia and I. communis
is similar to that found in a variety of other Cambrian
fossil arthropods, ranging from the anomalocaridid Hur-
dia (Delle Cave and Simonetta 1991, fig. 22E–F), the ole-
nelloid trilobite Wanneria walcottana (see Lieberman
1999, fig. 11.1) to the Chengjiang arthropod Retifacies
(see Hou and Bergstrom 1997, figs 48–50), as well as
some recent ostracodes (see Vannier et al. 2007). This
reticulation resembles that found in pelagic ostracodes,
which has been suggested to constitute a structural adap-
tation to produce the most resistant construction for the
lightest possible carapace with a minimum metabolic
investment, together with an enhancement of hydrody-
namic capabilities, reducing drag and retarding sinking
(Vannier and Chen 2000; Vannier et al. 2007). Because of
the presence of reticulation in benthic organisms such as
Retifacies or Wanneria, this second line of argument
might not be as robust as the indications coming from
the very low occurrence ratio of specimens with soft-parts
against empty carapaces or exuviae, and from the wide,
circumequatorial distribution of the genus for both the
early and middle Cambrian. On the other hand, the
higher proportion of Tuzoia with soft parts (mainly in
the form of eyes) in the Emu Bay Shale than in any other
locality where it occurs, including the Burgess Shale, may
suggest that T. australis could have led a nektonic life,
but closer to the seafloor than other species in the genus.
The presence in both I. communis and T. australis
(Pl. 2, figs 7–9; Pl. 5, figs 3–4) of large irregular patches
of dark fibrous material similar to the phosphatized mus-
cle fibres of Myoscolex ateles is taken as evidence that this
material is indeed remains of Myoscolex rather than inter-
nal structures that are identical in Isoxys and Tuzoia.
Myoscolex might have scavenged the carcasses of the biv-
alved arthropods once they reached the ocean bottom or
used their empty exuviae as a hiding place in an other-
wise barren seafloor.
In both Tuzoia and Isoxys, specimens in completely
open ‘butterfly’ configuration often show splitting of car-
dinal spines (Pl. 4, fig 3; Briggs et al. 1994, pl. 102; Van-
nier and Chen 2000, figs 2F–G; Hou et al. 2004, fig.
16.15b), often extending along part of the dorsal hinge,
and never preserve soft parts, whereas dvc specimens and
those in barely splayed ‘butterfly’ configuration show an
A B C
D E F
G H
vsL
vHvL eD psL vH
vL
eDasL
α
I
TEXT -F IG . 4 . A–F, H, Scaled outlines of Tuzoia australis, Emu Bay Shale, Big Gully, Kangaroo Island, South Australia. A, SAM
P17451A, holotype (see Pl. 4, fig. 1). B, SAM P14674, a large specimen in ‘butterfly’ configuration (see Pl. 4, fig. 3). C, SAM P15142,
small specimen, lacking ventral posterior spine, yet with a spine originating from the anterior ventral margin of the valve (see Pl. 4,
fig. 5). D, SAM P14719, large specimen with partially disarticulated valves (see Pl. 5, fig. 2). E, SAM P15292, almost complete large
specimen (see Pl. 5, fig. 1). F, SAM P15134, smallest specimen. G, Generalized outline of Tuzoia (modified from Vannier et al. 2007,
fig. 1.2). H, Outline of Tuzoia australis with the dimensions referred to in the text. I, Outline of Isoxys communis with the dimensions
referred to in the text. Specimens oriented anterior to the right. Scale bar represents 5 mm.
1238 P A L A E O N T O L O G Y , V O L U M E 5 2

intact hinge line and often preserve eyes and other soft-
bodied features (Pl. 4, figs 4, 6; Pl. 6, figs 1–2; Vannier
et al. 2007, figs 23.1, 23.6, 24.1–2; Garcıa-Bellido et al.
submitted). Some lateral specimens of Tuzoia are also
partially disarticulated along the dorsal line, displacing
the valves (Pl. 4, fig. 1; Pl. 5, fig. 2), and never preserve
eyes. This seems to suggest that, as with extant and fossil
arthropods (McNamara and Rudkin 1984; Garcıa-Bellido
and Collins 2004), these two bivalved arthropods weak-
ened their exoskeleton along a pre-established line enough
to allow them to exit their exuviae during the process of
moulting. Thus, specimens in completely splayed ‘butter-
fly’ configuration constitute empty moults (exuviae),
while dvc specimens correspond to either carcasses or
specimens buried alive.
Acknowledgements. Funding for this research was provided by an
Australian Research Council Linkage grant (LP0774959), with
generous financial assistance from the South Australian Museum
and Beach Petroleum Ltd. We are sincerely grateful to the Buck
family for access to the field area. We thank Jean Vannier and
an anonymous referee for their detailed and constructive
criticism of the manuscript. Thanks also to Natalie Schroeder
for photography, plus her assistance in the field, together with
Dennis Rice, Ronda Atkinson, Mike Gemmell, Leonie Feutrill,
Trevor and Jenny Worthy and John Laurie; SeaLink for logistical
support; and Dean Oliver for drafting Text-figure 1. DCGB also
thanks Sue Mikkelsen (SAM), the Spanish Research Council, the
Complutense University in Madrid and the Spanish Ministry of
Education and Science for financial support.
REFERENCES
B E N G T S O N , S., C ON W A Y M OR R I S , S., C O OP E R , B. J.,
J E L L , P. A. and R UN N E G A R , B. N. 1990. Early Cambrian
fossils from South Australia. Memoirs of the Association of
Australasian Palaeontologists, 9, 1–364.
B R I G G S , D. E. G. and N E DI N , C. 1997. The taphonomy and
affinities of the problematic fossil Myoscolex from the Lower
Cambrian Emu Bay Shale. Journal of Paleontology, 71, 22–32.
—— and W HI T TI N G T ON , H. B. 1985. Modes of life of
arthropods from the Burgess Shale, British Columbia. Trans-
actions of the Royal Society of Edinburgh, 76, 149–160.
—— E R W I N , D. H. and CO L L I E R , F. J. 1994. The fossils of
the Burgess Shale. Smithsonian Institution Press, Washington
and London, 238 pp.
—— L I E BE R M A N , B. S., H E N DR I CK S , J. R., HA L G E -
DA H L , S. L. and J A R R A R D, R. D. 2008. Middle Cambrian
arthropods from Utah. Journal of Paleontology, 82, 238–254.
B UT T E R F I E L D , N. J. 2002. Leanchoilia guts and the interpre-
tation of the three-dimensional structures in Burgess Shale-
type fossils. Paleobiology, 28, 155–171.
C H L UP A C , I. and KO R DU L E , V. 2002. Arthropods of Bur-
gess Shale type from the Middle Cambrian of Bohemia (Czech
Republic). Bulletin of the Czech Geological Survey, 77, 167–182.
C ON W A Y M OR R I S , S. and PE E L , J. S. 2008. The earliest
annelids: Lower Cambrian polychaetes from the Sirius Passet
Lagerstatte, Peary Land, North Greenland. Acta Palaeontologica
Polonica, 53, 137–148.
D A I L Y , B. 1956. The Cambrian in South Australia. 91–147. In
R OD G E R S , J. (ed.). El Sistema Cambrico, su paleogeografıa y
el problema de su base. 20th International Geological Congress,
Mexico, 1956. 762 pp.
—— M I L N E S , A. R., T W I D A L E , C. R. and B OU R N E , J. A.
1979. Geology and geomorphology. 1–38. In T Y L E R , M. J.,
L I N G , J. K. and T W I D A L E , C. R. (eds). Natural history of
Kangaroo Island. Royal Society of South Australia, Adelaide,
184 pp.
D E L L E CA V E , L. and S I M O N E T T A , A. M. 1991. Early Pal-
aeozoic arthropods and problems of arthropod phylogeny;
with some notes on taxa of doubtful affinities. 189–244. In
S I M ON E T T A , A. M. and CO N W A Y M O R R I S , S. (eds).
The early evolution of the Metazoa and the significance of prob-
lematic taxa. Cambridge University Press, Cambridge, 296 pp.
D Z I K , J. 2004. Anatomy and relationships of the early Cam-
brian Myoscolex. Zoologica Scripta, 33, 57–69.
G A R C I A - B E L L I D O, D. C. and C O L LI N S , D. H. 2004.
Moulting arthropod caught in the act. Nature, 429, 40.
—— —— 2005. Isoxys with soft parts from the Middle Cambrian
Burgess Shale. 50. In PE N G, S. C., Z HU , M. Y., L I , G. X.
and V A N I TE N , H. (eds). Acta Micropalaeontologica Sinica
(Supplement). Nanjing Institute of Geology and Palaeontolo-
gy, Nanjing, China, 223 pp.
—— —— 2007. Reassessment of the Genus Leanchoilia (Arthro-
poda, Arachnomorpha) from the Burgess Shale, British
Columbia, Canada. Palaeontology, 50, 603–709.
—— V A N N I E R , J. and C OL L I N S , D. 2009. Soft-part preser-
vation in two species of the arthropod Isoxys from the middle
Cambrian Burgess Shale of British Columbia, Canada. Acta
Palaeontologica Polonica, 54, doi: 10.4202/pp.2009.0024.
G L A E S S N E R, M. F. 1979. Lower Cambrian Crustacea and
annelid worms from Kangaroo Island, South Australia. Alche-
ringa, 3, 21–31.
H O U, X. G. 1987. Early Cambrian large bivalved arthropods
from Chengjiang, eastern Yunnan. Acta Palaeontologica Sinica,
26, 286–298.
—— and B E R G S T R O M , J. 1997. Arthropods of the Lower
Cambrian Chengjiang fauna, southwest China. Fossils and
Strata, 45, 1–116.
—— A L D R I D G E , R. J., B E R G S T R OM , J., S I V E T E R , D. J.,
S I V E TE R , D. J. and F E N G , X. H. 2004. The Cambrian
fossils of Chengjiang, China: the flowering of early animal life.
Blackwell Publishing, Oxford, 233 pp.
H U, S. X. 2005. Taphonomy and palaeoecology of the early
Cambrian Chengjiang biota from Eastern Yunnan, China.
Berliner palaobiologische Abhandlungen, 7, 1–197.
H UO , S. C., S HU , D. and CU I , Z. L. 1991. Cambrian Bradori-
ida of China. Geological Publishing House, Beijing, 249 pp.
I V A N T S OV , A. I. 1990. Pervye hakhodki fillokarid v nizhnem
Kembrii Iakutii. Palaeontologicheskii Zhurnal, 1990, 130–132.
[In Russian].
J A GO , J. B., PA T E R S O N , J. R., A L E X A N D E R , E. M. and
G E HL I N G , J. G. 2006a. Fossiliferous Cambrian localities on
G A R C I A - B E L L I D O E T A L . : I S O X Y S A N D T U Z O I A F R O M E M U B A Y S H A L E ( A U S T R A L I A ) 1239

the north coast of Kangaroo Island. 21–24. In J A G O, J. B.
and Z A N G , W. L. (eds). South Australia 2006. XI Interna-
tional Conference of the Cambrian Stage Subdivision Working
Group. Field Guide. Geological Society of Australia, South
Australian Division, Adelaide, 59 pp.
—— Z A N G , W., S UN , X., BR O CK , G. A., P A T E R S O N ,
J. R. and S KO V S T E D , C. B. 2006b. A review of the Cambrian
biostratigraphy of South Australia. Palaeoworld, 15, 406–423.
L I E BE R M A N , B. S. 1999. Systematic revision of the Olenelloi-
dea (Trilobita, Cambrian). Bulletin of the Yale University
Peabody Museum of Natural History, 45, 1–150.
—— 2003. A new soft-bodied fauna: The Pioche Formation of
Nevada. Journal of Paleontology, 77, 674–690.
L U O, H. L., J I A N G , Z. W., W U , X. C., S O N G, X. L. and
OU Y A N G , L. 1982. The Sinian-Cambrian boundary in east-
ern Yunnan, China. People’s Publishing House of Yunnan,
China, 265 pp.
—— F U , X. P., HU , S. H., L I , Y., C H E N , L. Z., Y O U, T. and
L I U , Q. 2006. New bivalved arthropods from the early Cam-
brian Guanshan fauna in the Kunming and Wuding area. Acta
Palaeontologica Sinica, 45, 460–472.
M C HE N R Y , B. and Y A T E S , A. 1993. First report of the enig-
matic metazoan Anomalocaris from the Southern Hemisphere
and a trilobite with preserved appendages from the Early
Cambrian of Kangaroo Island, South Australia. Records of the
South Australian Museum, 26, 77–86.
M C N A M A R A , K. J. and R U DK I N , D. M. 1984. Techniques
of trilobite exuviation. Lethaia, 17, 153–163.
N E D I N , C. 1995a. The Emu Bay Shale, a Lower Cambrian fos-
sil Lagerstatten, Kangaroo Island, South Australia. Memoirs of
the Association of Australasian Palaeontologists, 18, 31–40.
—— 1995b. The palaeontology and palaeoenvironment of the Early
Cambrian Emu Bay Shale, Kangaroo Island, South Australia.
Unpublished PhD thesis, University of Adelaide, 207 pp.
—— 1999. Anomalocaris predation on nonmineralized and min-
eralized trilobites. Geology, 27, 987–990.
P A T E R S ON , J. R. and B R OC K , G. A. 2007. Early Cambrian
trilobites from Angorichina, Flinders Ranges, South Australia,
with a new assemblage from the Pararaia bunyerooensis Zone.
Journal of Paleontology, 81, 116–142.
—— and J A G O , J. B. 2006. New trilobites from the Lower
Cambrian Emu Bay Shale Lagerstatte at Big Gully, Kangaroo
Island, South Australia. Memoirs of the Association of Austral-
asian Palaeontologists, 32, 43–57.
—— —— GE H L I N G, J. G., G A R C I A - B E L L I D O , D. C.,
E DG E C OM B E , G. D. and L E E , M. S. Y. 2008. Early Cam-
brian arthropods from the Emu Bay Shale Lagerstatte, South
Australia. 319–325. In R A BA N O, I., G OZ A L O , R. and
G A R CI A - BE L L I DO , D. (eds). Advances in trilobite
research. Instituto Geologico y Minero de Espana, Madrid, 448
pp.
R A Y M ON D, P. E. 1935. Leanchoilia and other Mid-Cambrian
Arthropoda. Bulletin of the Museum of Comparative Zoology,
Harvard University, 76, 205–230.
R E S S E R, C. E. 1929. New Lower and Middle Cambrian Crus-
tacea. Proceedings of the U.S. National Museum, 76, 1–18.
R I C H T E R , R. and R I C HT E R , E. 1927. Eine Crustacee (Isoxys
carbonelli n. sp.) in den Archaeocyathus-Bildungen der Sierra
Morena und ihre Stratigraphische Beurteilung. Senckenbergi-
ana, 9, 188–195.
R O BI S O N , R. A. and R I C HA RD S , B. C. 1981. Larger bivalve
arthropods from the Middle Cambrian of Utah. Paleontological
Contributions of the University of Kansas, 106, 1–19.
S H U , D. G., Z H A N G , X. L. and G E Y E R , G. 1995. Anatomy
and systematic affinities of the Lower Cambrian bivalved
arthropod Isoxys auritus. Alcheringa, 19, 333–342.
S I E BO L D, K. T. E. von and S T A N N I U S , H. F. 1845.
Lehrbuch der vergleichenden Anatomie der wirbellosen Tiere.
Von Veit, Berlin.
S I M O N E T TA , A. M. and DE L L E CA V E , L. F. 1975. The
Cambrian non Trilobite arthropods from the Burgess Shale of
British Columbia. A study of their comparative morphology,
taxonomy and evolutionary significance. Paleontographia
Italica, 69 (New Series 39), 1–37.
S P R I G G , R. C. 1955. The Point Marsden Cambrian beds, Kan-
garoo Island, South Australia. Transactions of the Royal Society
of South Australia, 78, 165–168.
—— C A M P A N A , B. and K I N G , D. 1954. Kingscote map sheet,
geological atlas of South Australia, 1:253,440 series. Geological
Survey of South Australia, Adelaide.
V A N N I E R , J. and C H E N , J. Y. 2000. The Early Cambrian
colonization of pelagic niches exemplified by Isoxys (Artho-
poda). Lethaia, 33, 295–311.
—— —— 2002. Digestive system and feeding mode in Cambrian
naraoiid arthropods. Lethaia, 35, 107–120.
—— G A R CI A - BE LL I DO , D. C., H U, S. X. and C H E N , A.
L. 2009. Arthropod visual predators in the early pelagic
ecosystem: evidence from the Burgess Shale and Chengjiang
biotas. Proceedings of the Royal Society B, 276, 2567–2574.
—— W I L L I A M S , M., A L V A R O , J. J., V I Z C A I N O , D.,
M O N CE R E T , S. and M ON C E R E T , E. 2005. New Early
Cambrian bivalved arthropods from southern France. Geologi-
cal Magazine, 142, 1–13.
—— C A R O N , J.-B., Y U A N , J. L., B R I G G S , D. E. G., C OL -
L I N S , D., Z HA O, Y. L. and Z HU , M. Y. 2007. Tuzoia:
morphology and lifestyle of a large bivalved arthropod of the
Cambrian seas. Journal of Paleontology, 81, 445–471.
W A L C O TT , C. D. 1890. The fauna of the Lower Cambrian or
Olenellus Zone. Reports of the U.S. Geological Survey, 10, 509–
763.
—— 1908. Mount Stephen rocks and fossils. The Canadian
Alpine Journal, 1, 232–248.
—— 1912. Cambrian Geology and Paleontology II. No. 6.
Middle Cambrian Branchiopoda, Malacostraca, Trilobita, and
Merostomata. Smithsonian Miscellaneous Collections, 57, 145–229.
W I L L I A M S , M., S I V E T E R , D. J. and P E E L , J. S. 1996. Isoxys
(Arthropoda) from the early Cambrian Sirius Passet Lagerstatte,
North Greenland. Journal of Paleontology, 70, 947–954.
Y U A N , J. L. and Z H A O, Y. L. 1999. Tuzoia (bivalved arthro-
pods) from the Lower-Middle Cambrian Kaili Formation of
Taijiang, Guizhou. Acta Palaeontologica Sinica, 38 (Suppl.),
88–93. [In Chinese, with English abstract].
Z H U, M. Y., V A N N I E R , J., V A N I TE N , H. and ZH A O ,
Y. L. 2004. Direct evidence for predation on trilobites in the
Cambrian. Proceedings of the Royal Society of London B (Sup-
pl.), 271, S277–S280.
1240 P A L A E O N T O L O G Y , V O L U M E 5 2

APPENDIX
List of symbols used in explanatory figures and text-figures:
as Anterior spine
ca Cephalic appendage
e Stalked eye
g Gut
H Height
l Prefix indicating left side
L Length
mgg Midgut glands
ps Posterior spine
r Prefix indicating right side
ta Trunk appendage
v Carapace valve
Hachures show breaks in slope, the solid line at the upper edge
of the break, the hachures directed down-slope from this line.
G A R C I A - B E L L I D O E T A L . : I S O X Y S A N D T U Z O I A F R O M E M U B A Y S H A L E ( A U S T R A L I A ) 1241












