Ecology and management of mahogany ( Swietenia macrophylla King) in the Chimanes Forest, Beni,...
26
Botanical Journal of the Linnean Society (1996), 122: 9-34. With 15 figures @ Is there a future for mahogany? Edited ly Andrew Madellan Ecology and management of mahogany (Swietenia rnacrophyZZa King) in the Chimanes Forest, Bed, Bolivia R.E. GULLISON, S.N. PANFIL, JJ. STROUSE AND S.P. HUBBELL Department of Ecology and Evolutionary Biology, Princeton Univen$, Princeton 08544-1 003, USA. Mahogany (Sruiclenia marrophrlla King) regenerates in areas of erosion on high terraces and in forest killed by flooding and deposition of alluvial sediments in the Chmanes Forest, Bolivia. These hydrological disturbancesare patchy, and only one of five stands of mahogany that we inventoried was regenerating. Mahogany survives these disturbances significantly better than the common tree species. The long time between disturbances appears to favour late maturation. Mahogany trees allocate little photosynthates to reproduction until they are very large emergents, at least 80 cm in diameter. The episodic nature of the regeneration sites means that mahogany stands are composed of one or a few cohorts, which are vulnerable to overharvesting, particularly with the current use of a minimum cutting diameter to regulate harvest. The delayed onset of fecundity means that the small trees that escape harvest are not very fecund, resulting in minimal seed input to logged forest. Only 7-9% of the gaps created by logging contain natural regeneration after 20 + yr. A successful management plan for mahogany would entail a monocyclic harvest, with a rotation age of 100 + years, the estimated time that it takes for trees to achieve commercial size in natural forest. Since the number of seed trees that will be left is small, they should be concentrated in sites that are likely to be conducive to natural regeneration, such as near rivers and flood damaged forest. Seed production will be maximized for a given basal area (opportunity cost to loggers)if trees c. 110cm dbh are selected as seed trees. The mahogany stocks in the Chimanes Forest are nearly exhausted, but the fmdings of this study could be used to help rebuild the mahogany populations, or to design management plans for the commercial species that have similar ecologies to mahogany. 01996 The Linnean Society of London ADDITIONAL KEY WORDS: -Amazon - conservation - demography - deposition - fecundity - flood - growth - life history - sustainable - tropical. CONTENTS Introduction ...... Forest and site description Previouslanduse ... Present land use ... Description of mahogany . . Morphology .... Phenology ..... Study area ....... .................... .................... .................... .................... .................... .................... .................... .................... 10 11 11 12 12 13 13 14 R.E. Gullison, current address: Renewable Resources Assessment Group, Centre for Environmental Technology, Imperial College of Science, Technology & Medicine, 8 Princes Gardens, London SW7 1NA S.N. Panfil, current address: Department of Botany, University of Georgia, Athens, GA 30602-7271, U.S.A. J.J. Strouse, current address: 2018 Master Drive, Baltimore, MD 21209, U S A . 9 00244074/96/090009+26 $18.00/0 01996 The Linnean Society of London
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Ecology and management of mahogany ( Swietenia macrophylla King) in the Chimanes Forest, Beni,...

Botanical Journal of the Linnean Society (1996), 122: 9-34. With 15 figures
@ I s there a future for mahogany? Edited ly Andrew Madellan
Ecology and management of mahogany (Swietenia rnacrophyZZa King) in the Chimanes Forest, B e d , Bolivia
R.E. GULLISON, S.N. PANFIL, JJ. STROUSE AND S.P. HUBBELL
Department of Ecology and Evolutionary Biology, Princeton Univen$, Princeton 08544-1 003, USA.
Mahogany (Sruiclenia marrophrlla King) regenerates in areas of erosion on high terraces and in forest killed by flooding and deposition of alluvial sediments in the Chmanes Forest, Bolivia. These hydrological disturbances are patchy, and only one of five stands of mahogany that we inventoried was regenerating. Mahogany survives these disturbances significantly better than the common tree species. The long time between disturbances appears to favour late maturation. Mahogany trees allocate little photosynthates to reproduction until they are very large emergents, at least 80 cm in diameter. The episodic nature of the regeneration sites means that mahogany stands are composed of one or a few cohorts, which are vulnerable to overharvesting, particularly with the current use of a minimum cutting diameter to regulate harvest. The delayed onset of fecundity means that the small trees that escape harvest are not very fecund, resulting in minimal seed input to logged forest. Only 7-9% of the gaps created by logging contain natural regeneration after 20 + yr. A successful management plan for mahogany would entail a monocyclic harvest, with a rotation age of 100 + years, the estimated time that it takes for trees to achieve commercial size in natural forest. Since the number of seed trees that will be left is small, they should be concentrated in sites that are likely to be conducive to natural regeneration, such as near rivers and flood damaged forest. Seed production will be maximized for a given basal area (opportunity cost to loggers) if trees c. 110 cm dbh are selected as seed trees. The mahogany stocks in the Chimanes Forest are nearly exhausted, but the fmdings of this study could be used to help rebuild the mahogany populations, or to design management plans for the commercial species that have similar ecologies to mahogany. 01996 The Linnean Society of London
ADDITIONAL KEY WORDS: -Amazon - conservation - demography - deposition - fecundity - flood - growth - life history - sustainable - tropical.
CONTENTS
Introduction . . . . . .
Forest and site description Previouslanduse . . . Present land use . . .
Description of mahogany . . Morphology . . . . Phenology . . . . .
Study area . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
10 11 11 12 12 13 13 14
R.E. Gullison, current address: Renewable Resources Assessment Group, Centre for Environmental Technology, Imperial College of Science, Technology & Medicine, 8 Princes Gardens, London SW7 1NA S.N. Panfil, current address: Department of Botany, University of Georgia, Athens, GA 30602-7271, U.S.A. J.J. Strouse, current address: 2018 Master Drive, Baltimore, MD 21209, USA.
9 00244074/96/090009+26 $18.00/0 01996 The Linnean Society of London

10 R. E. GULLISON E T A .
Life history . . . . . . . . . . . . . . . . . . . . Methods . . . . . . . . . . . . . . . . . . . . . .
Population structure . . . . . . . . . . . . . . , . . Post-harvest stand structure . . . . . . . . . . . . . . Spatial patterns and regeneration . . . . . . . . . . . . . Natural regeneration of logged populations . . . . . . . . . Fecundity . . . . . . . . . . . . . . . . . . . . Seed dispersal . . . . . . . . . . . , . . . . . . . Growth and mortality rates . . . . . . . . . . . . . .
Results . . . . . . . . . . . . . . . . . . . . . . . Population structure . . . . . . . . . . . . . . . . . Post-hawest stand structure . . . . . . . . . . . . . . Spatial patterns and regeneration . . . . . . . . . . . . . Natural regeneration of logged populations . . . . . . . . . Fecundity . . . . . . . . . . . . . . . . . . . . Seed dispersal . . . . . . . . . . . . . . . . . . . Seed mortality and germination . . . . . . . . . . . . . Growth and mortality rates . . . . . . . . . . . . . .
Discussion . . . . . . . . . . . . . . . . . . . . . . Hydrological disturbances and regeneration . . . . . . . . . Population structure . . . . . . . . . . . . . . . . . Life history and demography . . . . . . . . . . . . . . Effects of current management and recommendations for improvement
Conclusions . . . . . . . . . . . . . . . . . . . . . Acknowledgments . . . . . . . . . . . . . . . . . . . References . . . . . . . . . . . . . . . . . . . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
. . . . .
15 15 15 15 16 16 16 17 17 17 17 18 19 21 22 23 24 24 25 25 26 27 29 31 32 33
INTRODUCTION
High rates of deforestation in the Amazon and concern over the conservation status of some commercial timber species have drawn attention to the current poor understanding of the dynamics of Amazonian tree populations. The dynamics of these populations will be determined by the interplay of their life histories with the dynamics of the disturbance regimes that create regeneration sites (Clark, 1991a, b). The knowledge of these processes is of critical applied importance because it will form the basis of successful conservation and management plans.
The majority of research on neotropical forest disturbances has taken place in Mexico, Central America and the Caribbean. Various studies have identified the role in forest dynamics of treefall gaps (Lang & Knight, 1983; Hubbell & Foster, 1986; Brokaw, 1987; De Steven, 1988; Martinez-Ramos, Sarukhan & Pmero, 1988; Hartshorn, 1989), hurricanes (Crow, 1980; Frangi & Lugo, 1991) and fire (Lamb, 1966; Snook, 1993).
The agents of large scale forest disturbance in the Amazon differ from those in Central America. Fires are uncommon in Amazonian forest, at least in recent history (Uhl & Kauffman, 1990). Hurricanes are not present, but strong winds may cause considerable damage (Nelson et al., 1994; E. Ortiz, pers. comm.). Recent studies demonstrate the importance of river bend migration as a major disturbance in Amazonian forests (Salo et al., 1986; Kalliola et al., 199 1, 1992). Stream capture by rivers can change flooding and drainage patterns and result in conversion of forest to marsh along kilometre stretches 0. Terborgh, pers. comm.). There is also palynological and sediment evidence for catastrophic flooding events caused by periodic volcanic activity or changes in local or Andean precipitation regimes (Colinvaux et al., 1985; Frost & Miller, 1987).
Studies that relate Amazonian forest dynamics to these disturbance regimes are rare. The only example known to the authors is an excellent body of work on the life

MAHOGANY IN BOWIA 11
history characteristics of vegetation regenerating on primary successional habitats caused by the lateral migration of rivers (Foster, Arce & Wachter, 1986; Kalliola, Makinen & Salo, 1988; Kalliola et al., 1991; Salo & Kalliola, 1991).
Coupled with this lack of basic ecological knowledge is a noticeable absence of forests managed for sustained yield in the Amazon (Poore et al., 1989). While in itself not sufficient for good forest management, a sound ecological understanding of the population dynamics of Amazonian tree species is certainly a first step, and poor knowledge partially explains the current lack of successful forest management.
The current controversy over broad-leaved mahogany (Swietenia macrophylla King) illustrates how little is known about Amazonian tree populations. S. macrophylla (hereafter referred to as mahogany) occurs from Mexico to Bolivia (Pennington, 1981), but has been exhausted commercially in the northern parts of its range. Mahogany was proposed for listing on Appendix Two of the Committee for International Trade of Endangered Species in 1992, and again in 1994. Both attempts were unsuccessful. A successful listing would require producer and consumer countries to document all international trade in mahogany.
While the ecology of northern populations is well understood (Lamb, 1966; Snook, 1993), Virtually nothing is known about the ecology of Amazonian populations. Even basic inventories are lacking for mahogany over most of its range. An informed decision about the conservation status of mahogany depends heavily upon these data.
Here we present the results of a four year study on the ecology and management of mahogany in the Chimanes Forest, in the Bolivian Amazon. The goals of this study were (1) to identlfy the disturbances that lead to mahogany regeneration in the Chimanes Forest, (2) to obtain data on the basic demographic parameters and life history of the Chimanes populations, (3) to document the effects of current harvesting and management on mahogany, and (4) to use these data to suggest improvements to management.
STUDY AREA
Forest and site description
The Chimanes Permanent Timber Production Forest (66" 00' to 67' 00' West and 14" 30' to 16" 00' South) is located in the Chimanes Region, in the state of Beni, Bolivia (Burniske, 1994). In addition to the Chimanes Permanent Timber Production Forest (hereafter referred to as the Chimanes Forest), the Chimanes Region contains a protected area, the Beni Biological Reserve, and two Indigenous Territories. The Chimanes Forest was officially established in 1986 with the granting of seven concessions, although logging has taken place for at least the last 20yr in this area.
The 449000ha Chimanes Forest contains small pockets of savanna, and is bounded by savanna to the south and east. The forest can be divided into five categories with aerial photography (Government of Bolivia, 1993). Non-flooded alluvial plains forest (Bal) is situated on high terraces and areas of low hills, and has a standing volume of 150-180m3 ha-' (all species, dbh > 40cm). Temporarily flooded alluvial plains forest (Ba2) is situated on low terraces and may be flooded during the rainy season. The standing volume is c. 100m3 ha-'. Transitional

12 R. E. GULLISON E7AL
TABLE 1. Description of the study sites where 100 ha plots were installed to study the stand structure of mahogany in the Chimanes Forest. Bal = non-flooded alluvial plains forest. Ba2 = temporarily flooded alluvial plains forest. Tree species diversity is for all trees >10 cm dbh. (Source:
del Agda, unpublished Bachelor's thesis) ~~ ~
Fatima/ Monte Grande Jaimanche Aguw Negras Cuberene
ForestType Bal
Altitude(m) 190
Ptot Size (ha) 1
No. of Species 85
Ba2 215
1
73
Four Most Iriaztca deltoids Poulsenia armuta
I&?f.mdeltOidea Ashocaryum sp. (Palmae) -76 (Palmae) -56
Lununia pann$m orobo parvijbra (Flacourtaceae) -52 (Myristicaceae) ~ 5 1
(Palmae) -44 (Annonaceae) 7 ~ 3 7
Abundant (Palmae) n=9S (Moraceae) w S 5 Species
Sacruiea exmhka UnOnl?tlSiSjlOribunda
Ba2 190
2.25 92
Ba2 170 1
66
Pseudolmedia h i s Unwnopsisjoribundu (Moraceae) -11 (Annonaceae) -106
Swatea emrhiza Ashocaryum sp. (Palmae) -97 (Palmae) -74
Guam mm@hyUa Pseudolmedia laeub (Meliaceae) nn=76 (Moraceae) n=73
Richoia sp. Rheedia sp. (Euphorbiaceae) -52 (Guttiferae) -32
temporarily flooded alluvial plains forest (Ba3) is similar to Ba2 but lower in height, and with a lower standing volume of c. 73 m3 had. The other two types of forest, low hill forest and mountain forest, are not of major importance in the area.
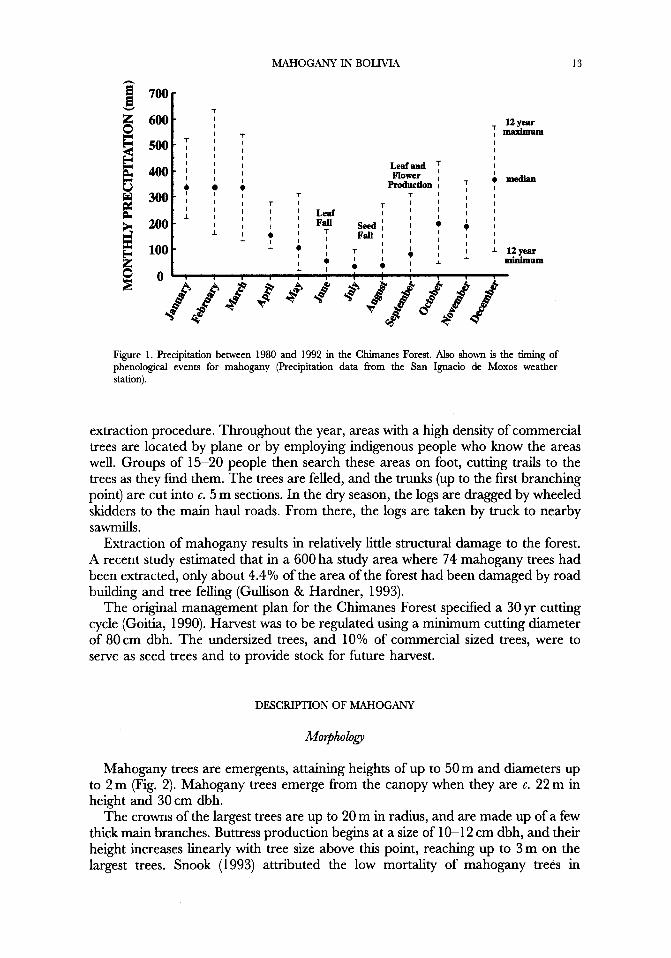
Botanical plots 1-2.25 ha in size have documented a tree diversity of 66-92 species > lOcm dbh (Table 1). Soils are derived from Tertiary and Late Quaternary Andean sediments, and are sandy/loam or clayish in nature (Government of Boiivia, 1993). The annual mean temperature is 26°C. Median precipitation at the two nearest weather stations (San Ignacio de Moxos and San Borja) is 202 1 and 23 16 mm yr-' for the period between 1980-1992 (Fig 1). July is the driest month of the dry season (defined as the months with median precipitation < = 100 mm), which lasts from May to September. The wettest months are December through March.
Prevbus Land use
Previous agricultural activity may have been recent enough to have influenced current forest structure and composition, and may be responsible for the relatively low tree species diversity of the Chimanes Forest. Pre-historic agricultural fields (camellones), typically 4.5 m wide and up to 200 m in length, are found on many of the savannas and throughout much of the Chimanes Forest. The cultures that created the camellones are poorly known, but are thought to date between 500-2300 B.P. (C. Erickson, pers. comm.). Camellones are present under mature forest in four of our six study sites, implying that much of the current forest was once either savanna or cleared for agriculture.
Present Land use
The seven timber concessions of the Chimanes Forest harvest mahogany almost exclusively (Goitia, 1990; Burniske, 1994). All the timber companies follow a similar
MAHOGANY IN BOLIVIA 13 h
W f 5 I I I I I I I I I
I I I I I I
T I I I I I
4 I T
12Y- l maximum I I
Figure 1 . Precipitation between 1980 and 1992 in the Chimanes Forest. Also shown is the timing of phenological events for mahogany (Precipitation data from the San Ignacio de Moxos weather station).
extraction procedure. Throughout the year, areas with a high density of commercial trees are located by plane or by employing indigenous people who know the areas well. Groups of 15-20 people then search these areas on foot, cutting trails to the trees as they find them. The trees are felled, and the trunks (up to the first branching point) are cut into c. 5 m sections. In the dry season, the logs are dragged by wheeled skidders to the main haul roads. From there, the logs are taken by truck to nearby sawmills.
Extraction of mahogany results in relatively little structural damage to the forest. A recent study estimated that in a 600 ha study area where 74 mahogany trees had been extracted, only about 4.4% of the area of the forest had been damaged by road building and tree felling (Gullison & Hardner, 1993).
The original management plan for the Chimanes Forest specified a 30yr cutting cycle (Goitia, 1990). Harvest was to be regulated using a minimum cutting diameter of 80cm dbh. The undersized trees, and 10% of commercial sized trees, were to serve as seed trees and to provide stock for future harvest.
DESCRIPTION OF MAHOGANY
Morphology
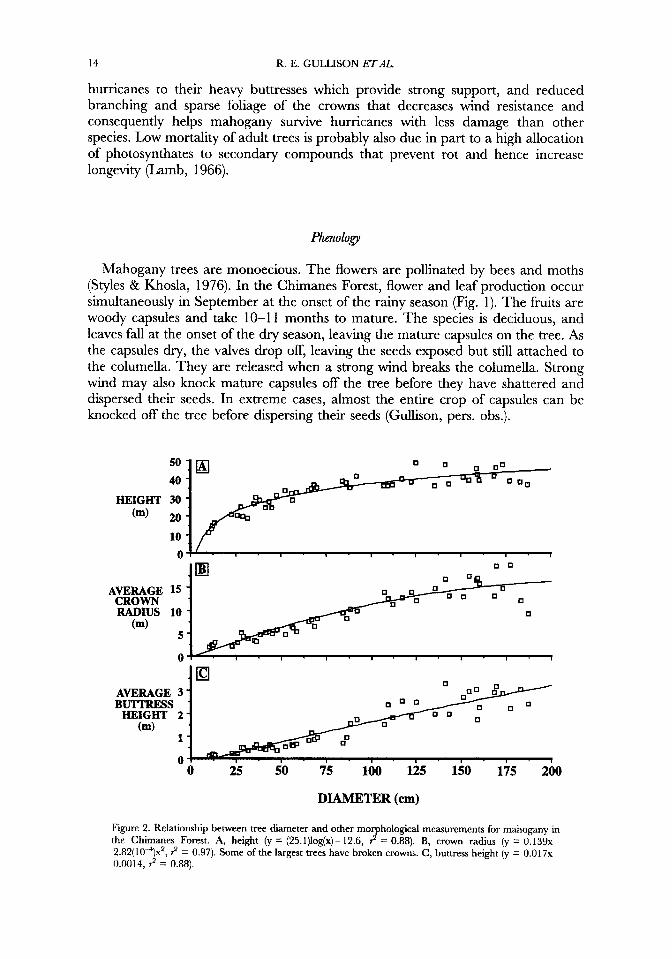
Mahogany trees are emergents, attaining heights of up to 50 m and diameters up to 2 m (Fig. 2). Mahogany trees emerge from the canopy when they are c. 22 m in height and 30cm dbh.
The crowns of the largest trees are up to 20 m in radius, and are made up of a few thick main branches. Buttress production begins at a size of 1&12 cm dbh, and their height increases linearly with tree size above this point, reaching up to 3 m on the largest trees. Snook (1993) attributed the low mortality of mahogany trees in
14 R. E. GULLISON ETAL.
hurricanes to their heavy buttresses which provide strong support, and reduced branching and sparse foliage of the crowns that decreases wind resistance and consequently helps mahogany survive hurricanes with less damage than other species. Low mortality of adult trees is probably also due in part to a high allocation of photosynthates to secondary compounds that prevent rot and hence increase longevity (Lamb, 1966).
Mahogany trees are monoecious. The flowers are pollinated by bees and moths (Styles & Khosla, 1976). In the Chimanes Forest, flower and leaf production occur simultaneously in September at the onset of the rainy season (Fig. 1). The fmits are woody capsules and take 10-1 1 months to mature. The species is deciduous, and leaves f d at the onset of the dry season, leaving the mature capsules on the tree. As the capsules dry, the valves drop off, leaving the seeds exposed but still attached to the columella. They are released when a strong wind breaks the columella. Strong wind may also knock mature capsules off the tree before they have shattered and dispersed their seeds. In extreme cases, almost the entire crop of capsules can be knocked off the tree before dispersing their seeds (Gullison, pers. obs.).
[us 10 1
5
0
BUTTRESS AVERAGE HEIGHT 3 2 ~~, (m)
na 1
0 0 25 50 75 100 125 150 175 200
DIAMETER (cm)
Figure 2. Relationship between tree diameter and other morphological measurements for mahogany in the Chimanes Forest. A, height (y = (25.1)lodx)- 12.6, 2 = 0.88). B, crown radius (y = 0.139~- 2.82(104)x2, ? = 0.97). Some of the largest trees have broken crowns. C, buttress height (y = 0.017~- 0.0014, ? = 0.88).

MAHOGANY IN BOLIVIA 15
Laj2 histoy
Mahogany is considered a light demanding climax species. Mahogany seedlings and saplings are shade intolerant, exhibiting their fastest growth with overhead light and lateral shading (Lamb, 1966). They are intolerant of deep shade, but can survive in a suppressed state for years in partial shade.
Mahogany is adapted to regenerate after fires and hurricanes in Central America and Mexico (Lamb, 1966; Snook, 1993). Adult trees survive these ephemeral disturbances better than most other tree species in these communities. As a result, they can play a dominant role in post-disturbance regeneration.
A recent comparison among mahogany and two pioneer and one other climax species showed that although photosynthetic rates did not differ much between species, mahogany had slower height growth and higher rates of relative biomass increase than the other three species (Ramos & Grace, 1990). The authors concluded that differences in height growth were due to species differences in allocation. Given its requirement for episodic disturbances to regenerate, it seems reasonable to assume that mahogany allocates more of its photosynthates to structural support and to secondary compounds that will increase longevity relative to other species, at the expense of height growth. Negative correlations between growth rates and investment into secondary compounds have been documented among temperate tree species (Loehle, 1988).
METHODS
Population structure
Density and stand structure of mahogany were investigated in Bal and Ba2 forest types, which are the most common in the Chimanes Forest. Five plots of 1 km by 1 km (100 ha) were established (Table 1). The Bal plots (Fatima and Monte Grande) were located on a terrace 3 4 m above the current river flood plain. The three remaining sites were located in Ba2 forest. The Jaimanche plot was located approximately 1 km away from the Jaimanche River, in the Fatima concession. The Aguas Negras plot was placed so that it abutted a flooded savanna area. The Cuberene site was located next to the Cuberene River.
Within each plot, a central 1000 m transect was cut, and then 10 randomly located 500 m transects were cut perpendicular to this. The transects were searched for all mahogany > 2.5 cm dbh within 10 m each side of the transect, yielding a total area searched of 10 ha (10%) per plot. AU mahogany trees found within the transect were tagged, mapped and measured (dbh, estimate of height to first branching, and total height). In addition, one of the 10 transects was selected at random and searched for all mahogany seedlings and saplings (individuals < 2.5 cm dbh). The locations of any topographical features were noted in the Bal plots.
Post-hanlest stand structure
The timber companies logged three of the five plots (Fatima, Monte Grande, and Jaimanche) during or shortly after installation. We noted the fates of all mahogany

16 R. E. GULLISON l 9 A L
trees located in the general area of the plots. Trees were classified as ‘seed trees’ if they were marked to be retained for seed production. Trees were classified as ‘missed’ if they were of commercial size but had not been found by the workers that searched for the trees. Trees were classified as ‘standing rotten’ if they had been left standing after trial cuts with a chainsaw had revealed rot. Trees were classified as ‘to be felled’ if they were marked for felling, but because of logistical difficulties (primarily bad weather), had not been felled yet. ‘Felled’ trees were trees that had already been felled by the company. Trees that had died naturally and that had been removed by the companies were classified as ‘natural deaths’.
Spatial pattans and regeneration
To help identlfjr the disturbances that led to their recruitment, several mahogany populations were mapped in relation to their local topography. A 15 ha plot (300 m by 500m) was installed in the Bal forest (Chirizi) in the area of highest density of commercial trees. All mahogany trees > 2.5 cm dbh were mapped and measured, and the topographical features were noted.
Four plots were installed at the Cuberene site (Ba2 forest) because of the extensive regeneration found there. Three plots (Bolson #1-3-4, Bolson #5, and El Pichi) contained river bends, and the fourth plot (La Esperanza) was installed about 300 m from the river. Plots were installed by cutting a baseline parallel to the river, and then cutting perpendicular trails every 20m along the baseline to the river. AU mahoganies > 2.5 cm dbh were mapped and measured.
Natural regeneration of logged populations
Gullison and Hubbell (1992) found that many mahogany seedlings survive the felling and removal of parent trees. To determine whether these seedlings reach commercial size in Ba2 forest, 28 plots were installed in the area of old gaps created by mahogany removal 18-20 yr ago. The partially rotted stumps and branches were easily discernible, and each plot was located so that it bounded the old gap by 10 m on all sides. Each plot was systematically searched for all mahogany individuals > 2.5cm dbh.
The survey of the Chirizi plot described in the previous section provided an opportunity to examine natural regeneration in Bal forest because it included 1 1 old gaps formed by the felling of commercial mahogany trees 20-25yr ago.
Fecundig
The relationship between tree size and fecundity (the number of fruits or capsules produced by the tree in one year) was determined by counting the capsules on trees 5-175 cm dbh. Trees drop their leaves before the capsules open and the seeds disperse, so it was possible to get good counts from the ground with binoculars. Trees with many lianas that impeded viewing were excluded. The ground below the crowns was searched to detect any mature capsules that had fallen, and these were added to the count. The capsules of 106 trees were counted in 1992. Counts were not

MAHOGANY IN BOLIVIA 17
possible in 1993 because the logging companies felled many of the trees, and access was impossible to the remainder. In 1994, we counted the capsules in 89 trees. Only trees in intact forest were included for the study. Trees by roads had unusually high fecundities, perhaps due to increased lateral exposure to light. To convert the capsule counts to total number of seeds produced, the seeds in 88 capsules from a dozen trees were counted.
Seed dispersal
To measure seed dispersal, seeds were counted in 1 m by 80 m transects extending in eight compass directions from the base of five trees (99, 122, 144, 145, and 160 cm dbh). The seeds were counted twice over a four week period. Seeds were marked with a felt pen the first time to prevent double counting. The data are presented as the average number of seeds m-* for 4m intervals along the transects.
ofowth and mortalip rates
The growth and mortality of 1 17 mahogany trees 2.5-200 cm dbh were measured each year between 1992 and 1994. The growth and mortality data are presented as annual rates for doubling diameter categories. To determine the relative ability of mahogany to survive flooding and deposition, the mortality rate of the mahogany in B# 1-3-4 plot was compared to the mortality rates of the five most abundant species in this plot( > 5 cm dbh). B#1-3-4 experienced the most flooding and deposition of all the plots.
RESULTS
Population structure
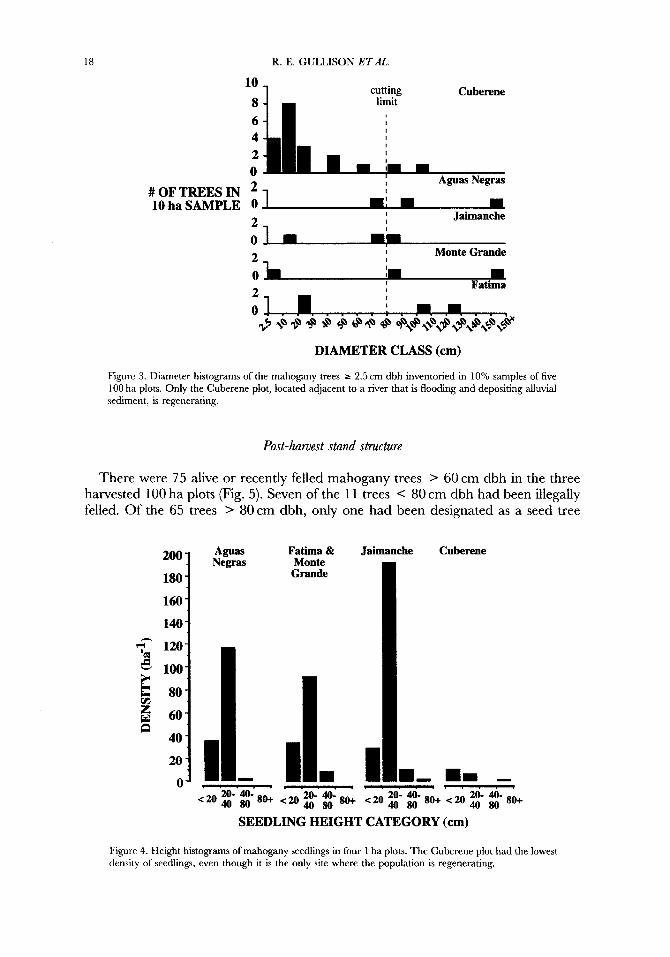
The density of mahogany trees > 80 cm dbh (legal commercial size) was 0.1-0.2 trees had on the five 100 ha plots (Fip 3). The density of trees < 80 cm dbh was similar among plots (0.1-0.2 trees ha- ), with the exception of the higher density of small trees in the Cuberene plot. Erosion gullies formed an average of 10.9% and 18.3% of the area of the transects of the Monte Grande and Fatima sites. There were no other distinct topographical features.
To determine whether a higher sampling intensity would have improved our estimate of the mean density of mahogany, we repeatedly calculated the standard error of the sample mean density as the data from an increasing number of transects was included. The standard error levelled off at a sampling intensity of 6-7%, demonstrating that our sampling intensity of 10% was more than adequate.
Seedling densities were also similar between plots (Fig. 9, averaging 174 seedlings had. The only exception was the Cuberene site, which despite the high density of individuals > 2.5 cm dbh, had a low density of 17 seedlings had.
18
200 - 180 - 160 - 140 - - 120- T4 100-
5; 801
E 60: n 40 - 20 -
0-
R. E. GULLISON E7AL.
lo 1 cutting Cuberene limit
I Aguas Negras
I! I I I Jaimanche
0
#OFTREESIN 2 1 I
10haSAMPLE 0 2
2 I
0
I
I
I Monte Grande
I I Fatima
0 1 = I
I I I
. .m. . . . . I . .=. .=. . . $5 \Q + 9 bp d 9 ++ d +\ff +Q@@\bp,%Q&j@
DIAMETER CLASS (cm)
Figure 3. Diameter histograms of the mahogany trees > 2.5 cm dbh inventoried in 10% samples of five 100 ha plots. Only the Cuberene plot, located adjacent to a river that is flooding and depositing alluvial sediment. is regenerating.
Post-haruest stand structure
There were 75 alive or recently felled mahogany trees > 60 cm dbh in the three harvested 100 ha plots (Fig. 5). Seven of the 1 1 trees < 80 cm dbh had been illegally felled. Of the 65 trees > 80cm dbh, only one had been designated as a seed tree
AEWW Fatima& Jaimanche Cuberene Nkras
< m I, I 20- ,-, 40-
40 80
Monte Grande
I I I, b . . . J
<20 g- g- 80+
SEEDLING HEIGHT CATEGORY (cm)
Figure 4. Height histograms of mahogany seedlings in four 1 ha plots. The Cuberene plot had the lowest density of seedlings, even though it is the only site where the population is regenerating.
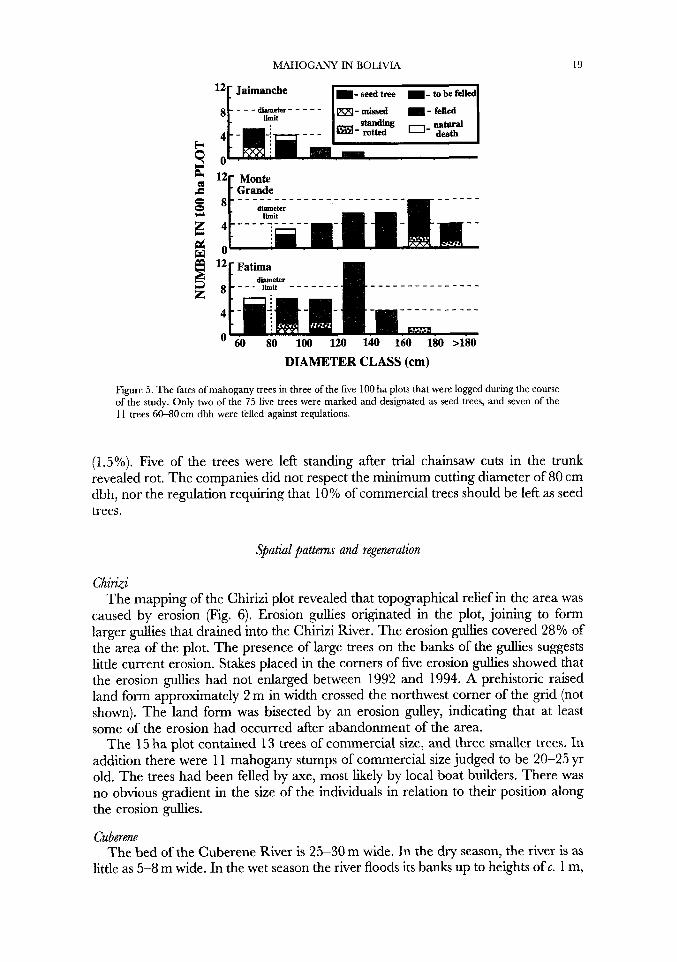
MAHOGANY IN BOLIVIA
121 Jaimanche m-seedtree --tobefell
19
2 12 Monte F Grande
DIAMETER CLASS (cm)
Figure 5 . The fates of mahogany trees in three of the five 100 ha plots that were logged during the course of the study. Only two of the 75 live trees were marked and designated as seed trees, and seven of the 11 trees 60-80 cm dbh were felled against regulations.
(1.5%). Five of the trees were left standing after trial chainsaw cuts in the trunk revealed rot. The companies did not respect the minimum cutting diameter of 80 cm dbh, nor the regulation requiring that 10% of commercial trees should be left as seed trees.
Spatial patterns and regeneration
Chi&$ The mapping of the Chirizi plot revealed that topographical relief in the area was
caused by erosion (Fig. 6). Erosion gullies originated in the plot, joining to form larger gullies that drained into the Chirizi River. The erosion gullies covered 28% of the area of the plot. The presence of large trees on the banks of the gullies suggests little current erosion. Stakes placed in the corners of five erosion gulhes showed that the erosion gullies had not enlarged between 1992 and 1994. A prehistoric raised land form approximately 2 m in width crossed the northwest corner of the grid (not shown). The land form was bisected by an erosion gulley, indicating that at least some of the erosion had occurred after abandonment of the area.
The 15 ha plot contained 13 trees of commercial size, and three smaller trees. In addition there were 1 1 mahogany stumps of commercial size judged to be 20-25 yr old. The trees had been felled by axe, most likely by local boat builders. There was no obvious gradient in the size of the individuals in relation to their position along the erosion gullies.
Cuberene The bed of the Cuberene River is 25-30 m wide. Xn the dry season, the river is as
little as 5-8 m wide. In the wet season the river floods its banks up to heights of c. 1 m,

20 R. E. GULLISON ETAL.
as evident by water marks on trees. The forest is heterogeneous, with no clear successional gradients with increasing distance from the river. Small differences in elevation determine whether a patch of forest is flooded or not.
Regeneration was present in three of the four plots in this area (Figs 7, 8). Only the unflooded Pichi plot had no individuals in the smaller size classes. There appear to be at least three different microhabitats where the smaller size classes occur. First, a few seedlings were present in the primary successional habitat created by river bend migration. Second, some of the seedlings were located along the banks of the river bends where presumably the extra light permits survival and growth. Many of these seedlings were of bad form and overgrown by vines. The third and most important microhabitat where regeneration occurred was low in areas on the plots that experienced annual flooding and deposition of alluvial sediment. Flooding and deposition had opened up large areas of forest by killing all size classes of trees. Flooding can reach areas quite distant from the river, as demonstrated in the Esperanza plot that is 3 0 N 0 0 m distant.
Terrace 4m
Depth of Erosion Gullies
t
Erosion 100 m ~ t e g o r y - Gulley
0 -Stump
- Diameter
(cm) 0 50.t 0 10-25 * 2.5 - 10
Figure 6. A map of the 15 ha plot at Chirizi, located between the Fatima and Monte Grande 100 ha plots. A; the plot is located on a high terrace that is cut by erosion gulhes that network together. Where the gullies leave the west side of the plot they have attained a depth of 4m. The erosion gullies cover 28% of the area of the plot. B, the sizes and locations of live trees, and the locations of old stumps, axe shown in relation to the erosion gullies. The high density of trees and stumps in this plot is associated with the large area of past erosion.
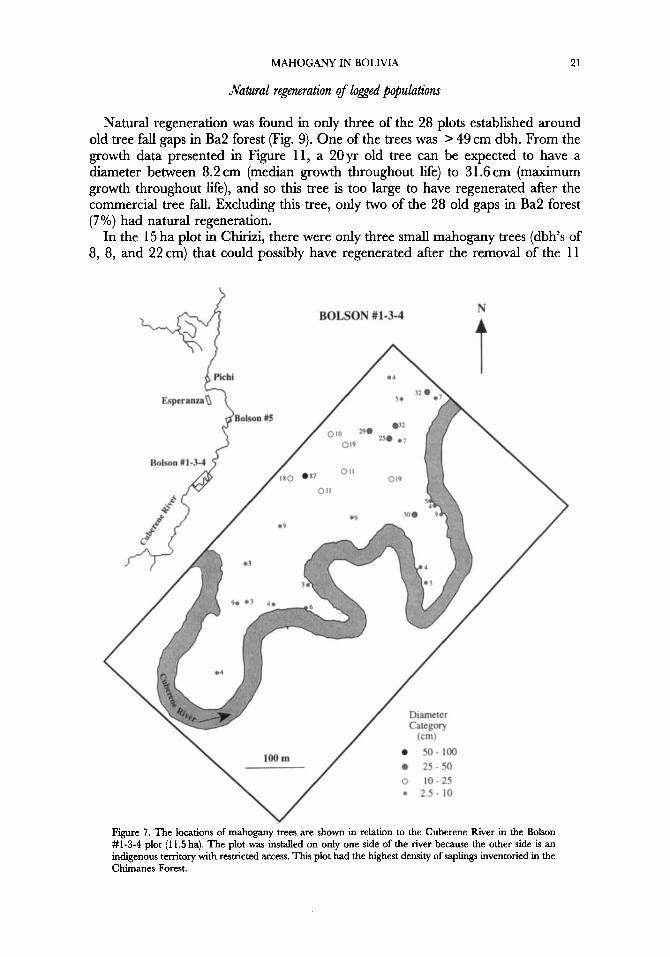
MAHOGANY IN BOLIVIA 21
Natural regaeration of logged popuhtions
Natural regeneration was found in only three of the 28 plots established around old tree fall gaps in Ba2 forest (Fig. 9). One of the trees was > 49 cm dbh. From the growth data presented in Figure 1 1, a 20yr old tree can be expected to have a diameter between 8.2 cm (median growth throughout Me) to 31.6 cm (maximum growth throughout Me), and so this tree is too large to have regenerated after the commercial tree f d . Excluding this tree, only two of the 28 old gaps in Ba2 forest (7%) had natural regeneration.
In the 15 ha plot in Chirizi, there were only three small mahogany trees (dbh’s of 8, 8, and 22 cm) that could possibly have regenerated after the removal of the 11
N A BOLSON #1-3-4
Diameter
25-50 O 10-25 \ / * 2.5- 10
Figure 7. The locations of mahogany trees are shown in relation to the Cuberene River in the Bolson #1-3-4 plot (1 1.5 ha). The plot was installed on only one side of the river because the other side is an indigenous territory with restricted access. This plot had the highest density of saplings inventoried in the Chimanes Forest.
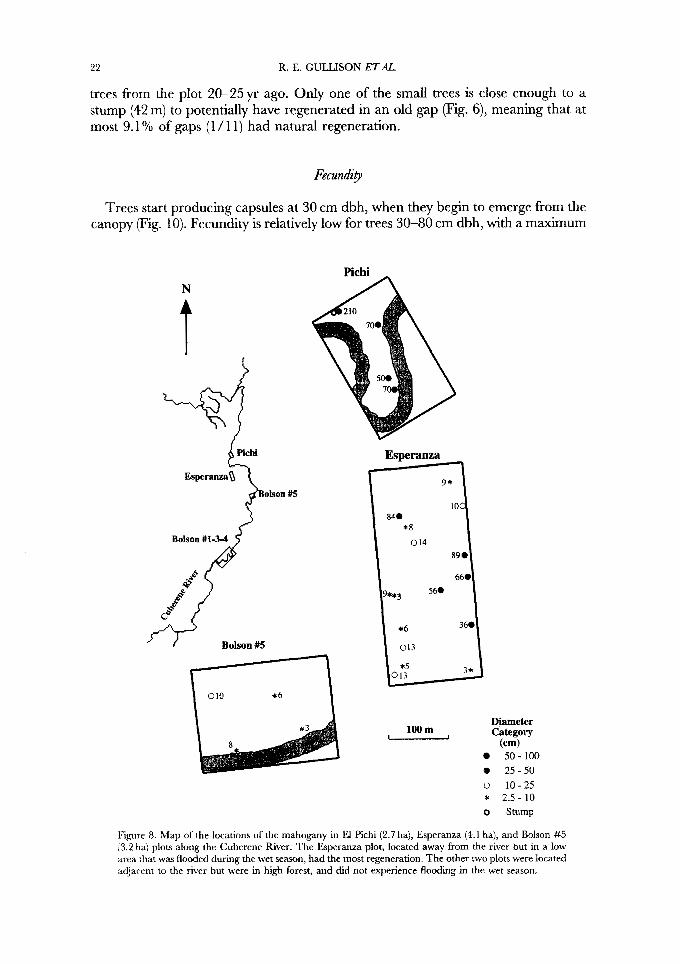
22 R. E. GULLISON ETAL.
c
trees from the plot 20-25yr ago. Only one of the s m d trees is close enough to a stump (42 m) to potentially have regenerated in an old gap (Fig. 6), meaning that at most 9.1 YO of gaps (1 / 1 1) had natural regeneration.
9*
1oc 840
*8
0 1 4 89 0
6 6 0
9**3 56.
*6 36.
013
Fecund+
3* *5
Trees start producing capsules at 30 cm dbh, when they begin to emerge from the canopy (Fig. 10). Fecundity is relatively low for trees 30-80 cm dbh, with a maximum
N
Esperanzaa
Olson #5 f Bolson # I 3 4
c $
J.jY Bolson #5
Diameter 100 m Category -
(cm) 50-100 2 5 - 5 0
0 10-2.5 * 2.5- 10 o Stump
Figure 8. Slap of the locations of the mahogany in El Pichi (2.7 ha), Esperanza (4.1 ha), and Bolson #5 l3.2 ha) plors along the Cuberene River. The Esperanza plot, located away from the river but in a low area that was flooded during the wet season, had the most regeneration. The other two plots were located adjacent to the river but were in high forest, and did not experience flooding in the wet season.
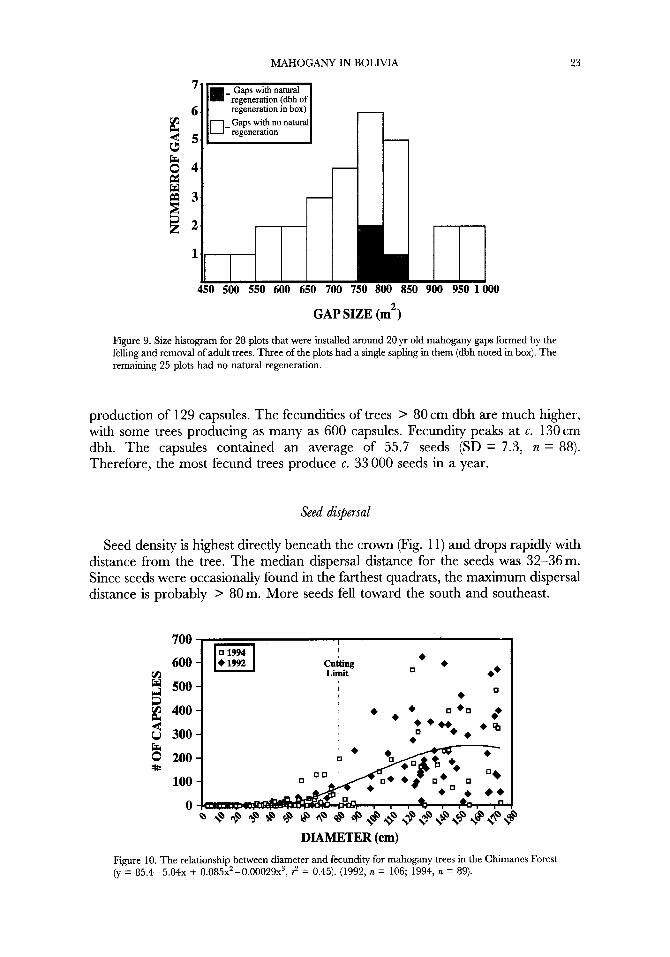
MAHOGANY IN BOLIVIA
regeneration in box)
450 500 550 600 650 700 750 800 850 900 950 lo00
23
GAP SIZE (mL)
Figure 9. Size histogram for 28 plots that were installed around 20 yr old mahogany gaps formed by the felling and removal of adult trees. Three of the plots had a single sapling in them (dbh noted in box). The remaining 25 plots had no natural regeneration.
production of 129 capsules. The fecundities of trees > 80 cm dbh are much higher, with some trees producing as many as 600 capsules. Fecundity peaks at c. 130 cm dbh. The capsules contained an average of 55.7 seeds (SD = 7.3, n = 88). Therefore, the most fecund trees produce c. 33 000 seeds in a year.
Seed dispersal
Seed density is highest directly beneath the crown (Fig. 1 1) and drops rapidly with distance from the tree. The median dispersal distance for the seeds was 32-36m. Since seeds were occasionally found in the farthest quadrats, the maximum dispersal distance is probably > 80m. More seeds fell toward the south and southeast.
s DIAMETER (cm)
Figure 10. The relationship between diameter and fecundity for mahogany trees in the Chimanes Forest (y = 85.4-5.04~ + 0.085~'-0.00029x3, 12 = 0.45). (1992, n = 106; 1994, n = 89).
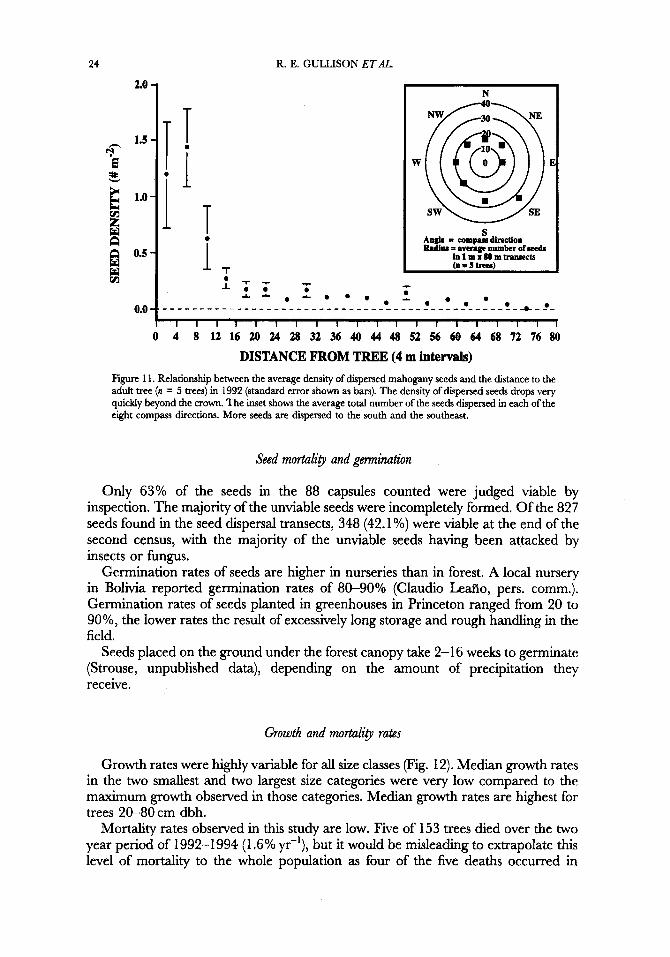
24
2.0
1.5 T
% 3
1.0 m E n a 0.5
0.0
R. E. GULLISON ETAL.
N
w@ sw SE
0 4 8 1 2 1 6 2 0 2 4 2 8 3 2 3 6 4 0 4 4 4 8 5 2 5 6 6 0 6 4 6 8 7 2 7 6 8 0
DISTANCE FROM TREE (4 m intervals) Figure 1 1. Relationship between the average density of dispersed mahogany seeds and the distance to the adult tree (n = 5 trees) in 1992 (standard error shown as bars). The density of dispersed seeds drops very quickly beyond the crown. The inset shows the average total number of the seeds dispersed in each of the eight compass directions. More seeds are dispersed to the south and the southeast.
Seed mortulip and germination
Only 63% of the seeds in the 88 capsules counted were judged viable by inspection. The majority of the unviable seeds were incompletely formed. Of the 827 seeds found in the seed dispersal transects, 348 (42.1 %) were viable at the end of the second census, with the majority of the unviable seeds having been attacked by insects or fungus.
Germination rates of seeds are higher in nurseries than in forest. A local nursery in Bolivia reported germination rates of 80-90% (Claudio Leaflo, pers. comm.). Germination rates of seeds planted in greenhouses in Princeton ranged from 20 to 90%, the lower rates the result of excessively long storage and rough handling in the field.
Seeds placed on the ground under the forest canopy take 2-16 weeks to germinate (Strouse, unpublished data), depending on the amount of precipitation they receive.
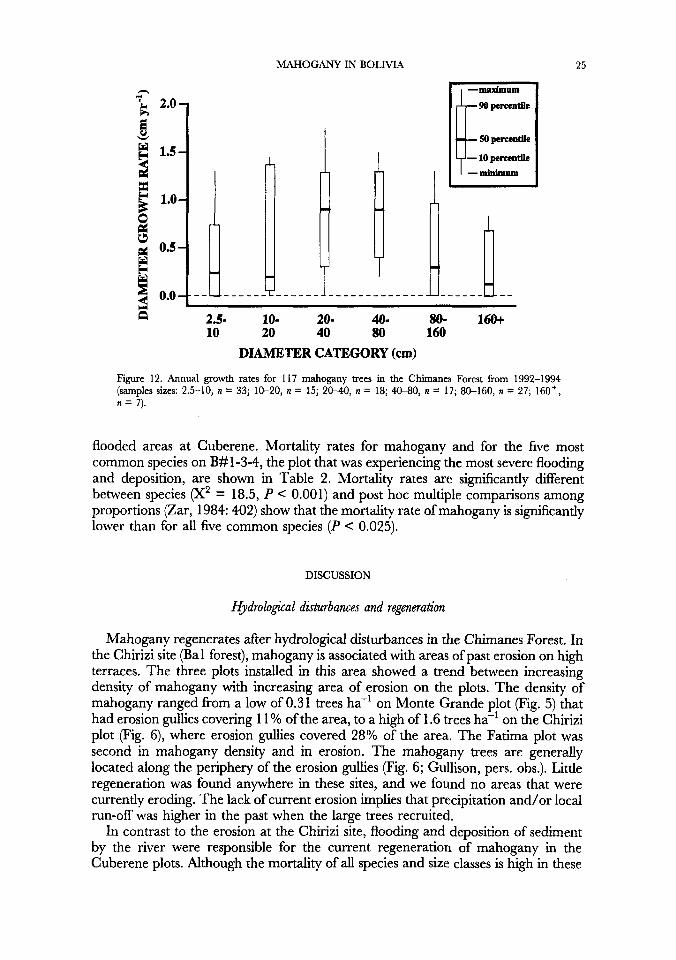
Growth and rnortalig rates
Growth rates were highly variable for all size classes (Fig. 12). Median growth rates in the two smallest and two largest size categories were very low compared to the maximum growth observed in those categories. Median growth rates are highest for trees 2&80cm dbh.
Mortality rates observed in this study are low. Five of 153 trees died over the two year period of 1992-1994 (1.6% yf'), but it would be misleading to extrapolate this level of mortality to the whole population as four of the five deaths occurred in
MAHOGANY IN BOLIVIA 25
n
1 2.0
W 8 -9Operrmtile
- 50 percentile
- 10 percentile --minimnm
DIAMETER CATEGORY (cm)
Figure 12. Annual growth rates for 117 mahogany trees in the Chimanes Forest from 1992-1994 (samples sizes: 2.5-10, n = 33; 10-20, n = 15; 2 M 0 , n = 18; 4W0, n = 17; 80-160, n = 27; 160+, n = 7).
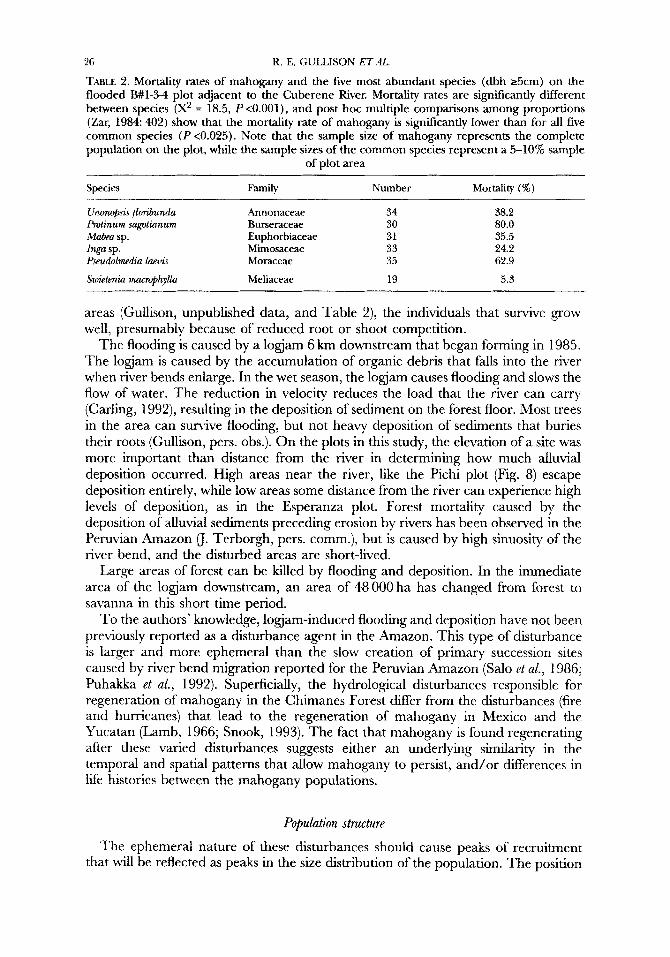
flooded areas at Cuberene. Mortality rates for mahogany and for the five most common species on B# 1-3-4, the plot that was experiencing the most severe flooding and deposition, are shown in Table 2. Mortality rates are significantly different between species (X2 = 18.5, P < 0.001) and post hoc multiple comparisons among proportions (Zar, 1984: 402) show that the mortality rate of mahogany is significantly lower than for all five common species (P < 0.025).
DISCUSSION
Hydrological disturbances and regeneration
Mahogany regenerates after hydrological disturbances in the Chimanes Forest. In the Chirizi site (Bal forest), mahogany is associated with areas of past erosion on high terraces. The three plots installed in this area showed a trend between increasing density of mahogany with increasing area of erosion on the plots. The density of mahogany ranged from a low of 0.3 1 trees ha-' on Monte Grande plot (Fig. 5) that had erosion gullies covering 1 1 % of the area, to a high of 1.6 trees ha-' on the Chirizi plot (Fig. 6), where erosion gullies covered 28% of the area. The Fatima plot was second in mahogany density and in erosion. The mahogany trees are generally located along the periphery of the erosion gullies (Fig. 6; Gullison, pers. obs.). Little regeneration was found anywhere in these sites, and we found no areas that were currently eroding. The lack of current erosion implies that precipitation and/or local run-off was higher in the past when the large trees recruited.
In contrast to the erosion at the Chirizi site, flooding and deposition of sediment by the river were responsible for the current regeneration of mahogany in the Cuberene plots. Although the mortality of all species and size classes is high in these
26 R. E. GULLISON ETAL.
TABLE 2. Mortality rates of mahogany and the five most abundant species (dbh 25cm) on the flooded B#1-34 plot adjacent to the Cuberene River. Mortality rates are significantly different between species (X2 = 18.5, P <0.001), and post hoc multiple comparisons among proportions (Zar, 1984 402) show that the mortality rate of mahogany is significantly lower than for all five common species ( P <0.025). Note that the sample size of mahogany represents the complete population on the plot, while the sample sizes of the common species represent a 5-10% sample
of plot area
Species Family Number Mortality (%)
UnonopSisflorihundn Annonaceae 34
Inga sp. Mimosaceae 33
Sriidenia onarrOphj.ll0 Meliaceae 19
Protinurn sagotianurn Burseraceae 30 Mabe0 sp. Euphorbiaceae 31
Pseudolmedia laeii,zs Moraceae 35
38.2 80.0 35.5 24.2 62.9
5.3
areas (Gullison, unpublished data, and Table 2), the individuals that survive grow well, presumably because of reduced root or shoot competition.
The flooding is caused by a logjam 6 km downstream that began forming in 1985. The logjam is caused by the accumulation of organic debris that falls into the river when river bends enlarge. In the wet season, the logjam causes flooding and slows the flow of water. The reduction in velocity reduces the load that the river can carry (Carling, 1992), resulting in the deposition of sediment on the forest floor. Most trees in the area can survive flooding, but not heavy deposition of sediments that buries their roots (Gullison, pers. obs.). On the plots in this study, the elevation of a site was more important than distance from the river in determining how much alluvial deposition occurred. High areas near the river, like the Pichi plot (Fig. 8) escape deposition entirely, while low areas some distance from the river can experience high levels of deposition, as in the Esperanza plot. Forest mortality caused by the deposition of alluvial sediments preceding erosion by rivers has been observed in the Peruvian Amazon 0. Terborgh, pers. comm.), but is caused by high sinuosity of the river bend, and the disturbed areas are short-lived.
Large areas of forest can be killed by flooding and deposition. In the immediate area of the logjam downstream, an area of 48000ha has changed from forest to savanna in this short time period.
To the authors' knowledge, logjam-induced flooding and deposition have not been previously reported as a disturbance agent in the Amazon. This type of disturbance is larger and more ephemeral than the slow creation of primary succession sites caused by river bend migration reported for the Peruvian Amazon (Salo et al., 1986; Puhakka et al., 1992). Superficially, the hydrological disturbances responsible for regeneration of mahogany in the Chimanes Forest differ from the disturbances (fire and hurricanes) that lead to the regeneration of mahogany in Mexico and the Yucatan (Lamb, 1966; Snook, 1993). The fact that mahogany is found regenerating after these varied disturbances suggests either an underlying similarity in the temporal and spatial patterns that allow mahogany to persist, and/or differences in life histories between the mahogany populations.
Population structure
The ephemeral nature of these disturbances should cause peaks of recruitment that will be reflected as peaks in the size distribution of the population. The position

MAHOGANY IN BOLIVIA 27
of the peak or peaks will depend on the time since they regenerated (Clark, 199 1 a). Few of the plots have enough trees to demonstrate the size distributions well. Two of the harvest plots do, Fatima and Monte Grande, and they have unimodal size distributions with the modes considerably above the minimum cutting diameter (Fig. 5). The size distributions for trees to be harvested over larger areas also show multiple peaks (Fig. 13). These distributions support the hypothesis that these stands have undergone episodic regeneration.
The overall density of mahogany in the Chimanes Forest may be lower than the density of 0.1-0.2 trees had that we found in our plots, as the plots were installed in areas that were known to contain mahogany. The low density of mahogany in the Chimanes Forest (Fig. 3) may be due to the age of the stands, or to low seed input at the time of regeneration. It seems unlikely that density dependent processes are regulating the population at such a low level, given that densities of up to 6-8 trees ha-' > 80 cm dbh occasionally occur in the Chimanes Forest (Gullison, pers. obs.), and that the density of mahogany is low compared to other regions. For example, Verissimo et al. (1 995), in the state of Para, Brasil, report densities of 1 .O commercial tree ha-', while Lamb (1 966) gives densities of large mahogany emergents in the Yucatan Peninsula of Mexico ranging 2.2-22 trees ha-'.
Lij% history and demograply
The temporal allocation of photosynthates between growth and reproduction (the life history) will determine how a tree species responds demographically to the regeneration sites created by the local disturbance regime (Stearns, 1976; Bell, 1980; Clark, 1991b). Here we interpret the demographic parameters of mahogany
50 -
40 - & 3 30-
20 -
10 -
n I
5 z
DIAMETER CATEGORY (cm) Figure 13. Diameter distribution of 245 mahogany trees inventoried prior to harvest by the Programa Chimanes forestry staff. The census area was not recorded. Multiple peaks are present in the distribution, suggesting several regeneration events. The current management plan is poorly followed, with the forestry department's own data showing that only 3.7% of commercial trees were left as seed trees, instead of the required 10%. These seed trees, and the 10 smaller trees, will supposedly provide the next harvest. (Data provided by Programa Chimanes/Subprograma Forestal).
28 R. E. GULLISON ETAL.
observed in this study in light of the observation that mahogany regenerates after large scale disturbances. Ideally, experimental manipulations of investment into growth or reproduction are used to demonstrate tradeoffs, but this is diacult for long lived organisms such as trees. Instead, we are limited to examining and interpreting the variation present in these parameters in natural populations (for an example see Eis, Garman & Ebel, 1965).
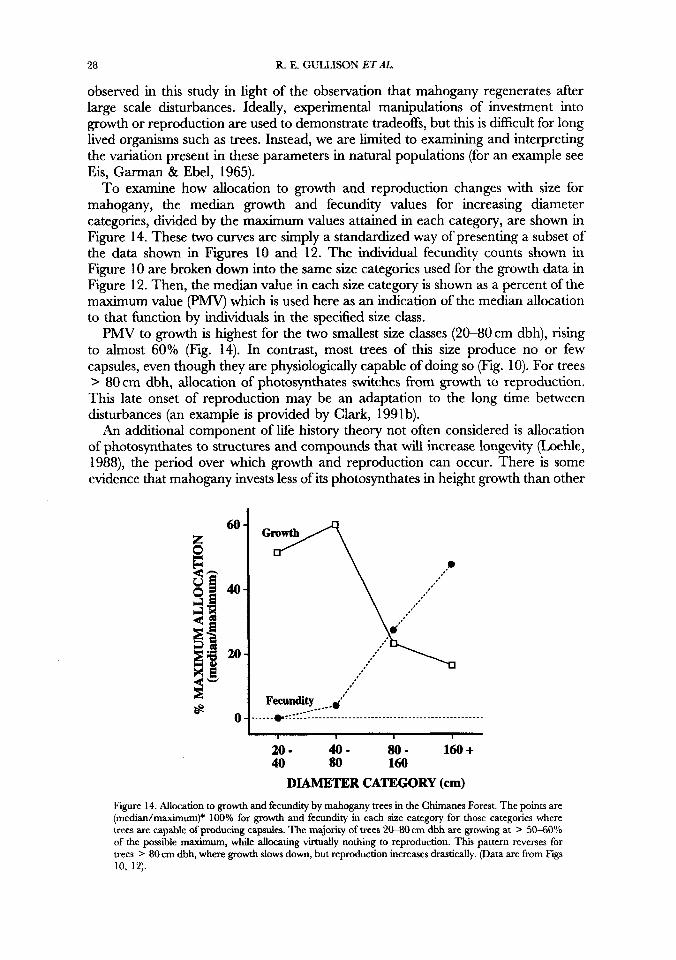
To examine how allocation to growth and reproduction changes with size for mahogany, the median growth and fecundity values for increasing diameter categories, divided by the maximum values attained in each category, are shown in Figure 14. These two curves are simply a standardized way of presenting a subset of the data shown in Figures 10 and 12. The individual fecundity counts shown in Figure 10 are broken down into the same size categories used for the growth data in Figure 12. Then, the median value in each size category is shown as a percent of the maximum value (PMV) which is used here as an indication of the median allocation to that function by individuals in the specified size class.
Ph4Y to growth is highest for the two smallest size classes (20-80cm dbh), rising to almost 60% (Fig. 14). In contrast, most trees of this size produce no or few capsules, even though they are physiologically capable of doing so (Fig. 10). For trees > 80cm dbh, allocation of photosynthates switches from growth to reproduction. This late onset of reproduction may be an adaptation to the long time between disturbances (an example is provided by Clark, 1991b).
An additional component of life history theory not often considered is allocation of photosynthates to structures and compounds that will increase longevity (Loehle, 1988), the period over which growth and reproduction can occur. There is some evidence that mahogany invests less of its photosynthates in height growth than other
I 1 I I I
20 - 40- 80- 160 + 40 80 160
DIAMETER CATEGORY (cm) Figure 14. Allocation to growth and fecundity by mahogany trees in the Chimanes Forest. The points are (median/maximum)* 100% for growth and fecundity in each size category for those categories where trees are capable of producing capsules. The majority of trees 2W30 em dbh are growing at > 50-60°/o of the possible maximum, while allocating virtually nothing to reproduction. This pattern reverses for trees > 80 cm dbh, where growth slows down, but reproduction increases drastically. (Data are from Figs 10, 12).

MAHOGANY IN BOLIVIA 29
early successional species, and may invest instead in secondary compounds to decrease rot (Ramos & Grace, 1990). Snook (1993) suggests that mahogany invests in buttresses to increase its ability to survive through hurricanes. Adaptation to episodic disturbances would reward allocation of photosynthates to increasing longevity for two reasons. First, the return time of the disturbances that permit regeneration could be very long and unpredictable. Therefore, the longer a tree survived, the higher the probability of it experiencing the disturbance necessary for regeneration. Second, the episodic disturbances that create regeneration sites for mahogany may destroy the seed and seedling banks (Snook, 1993; Gullison, pers. obs.). Reproductive trees must survive through the disturbances to ensure that their seeds and seedlings are present after the disturbance. Snook (1 993) has shown that mahogany trees survive fires and hurricanes better than other species. Similarly, we have observed that mahogany trees survive flooding better than the common species in the Cuberene area (Table 2).
In summary, mahogany is a long lived, late maturing species that regenerates after large scale hydrological disturbances in the Chimanes Forest. This leads to the development of stands composed of one or a few cohorts, very different from the ‘reverse-j’ shaped size distribution of a species that readily regenerates in tree fill gaps or under closed canopy. The frequency and magnitude of past large scale flooding and erosion events in the Chimanes Forest, and their role in influencing forest dynamics in the Chimanes Forest, is still poorly understood.
Eflects of current management and recommendations for improvement
Haruest &dub The time for mahogany trees to reach commercial size in natural forest can be
estimated from the growth data in Figure 12. If a tree grows at the maximum rate throughout its life, it will take 52yr to reach commercial size, while growing at the median growth rate it will take 148yr. The actual time to achieve commercial size should fall within this range. Gullison and Hubbell (1992) estimated from growth rings that the existing commercial trees took on average 105 yr to reach the 80 cm size limit. The strong seasonality in the Chimanes Forest, and some preliminary evidence from scarring growth windows (cuts made to the cambium in consecutive years which identlftr when growth rings are formed), suggest that these growth rings are annual.
The growth rates of mahogany appear remarkably consistent across its range. Lamb (1966) reports average growth rates of 3.6 to 9.1 mm yr-’, nearly identical to the median growth rates found in this study of 2.6 to 9.Omm yr-’. Snook (1993) found a similar growth rate of 2.0-10.9mm yr-’.
Unless silvicultural treatments are undertaken to increase growth, it should be assumed that it will take somewhere between 105 and 148 years for mahogany trees to reach commercial size in natural forest.
The slow growth rates and episodic nature of regeneration events make it clear that the current management plan for mahogany of a 30yr polycyclic harvest is inappropriate. Even if the companies obeyed the 80 cm minimum diameter and left 10% of trees > 80 cm dbh as required, harvest would often result in the removal of entire stands, with no or too few survi\;ing trees to permit subsequent harvests, or to provide postharvest seed input to the site.

30 R. E. G L U S O N ETAL.
The episodic nature and large scale of the disturbances that mahogany requires to regenerate mean that a monocyclic harvest is more appropriate for this species. The rotation age would be longer than the current 30yr cutting cycle, more likely to be on the order of 100 + years. At least at current stand density, it is unlikely that the reduced annual harvest volume that this would imply would be economically viable, so the companies would need to augment their harvest with other timber species (R. Rice, pers. comm.) or have much larger concessions.
Seed trees
Size. Under the current management plan, the population of seed trees is composed of the 10% of trees > 80 cm dbh that is left by the companies, and all trees < 80 cm dbh. This results in a large decrease in seed input to logged forest, as trees < 80 cm produce relatively few seeds (Fig. 10) and the reduction of trees > 80 cm is so great. The reduction in seed input will be even greater when the smaller trees are illegally felled and < 10% of commercial trees are not left, as is happening in the Chimanes Forest (Figs 5, 13).
The impact of harvest on seed input into the three harvest plots (Monte Grande, Fatima and Jaimanche) can be estimated by multiplying the pre- and post harvest diameter distributions by their estimated fecundity from the regression in Figure 10. Combined pre-harvest fecundity (n = 75 trees) on the three sites was 13 489 capsules. After harvest, only six healthy trees (seed trees and trees missed by the company) and five trees left standing because of heart rot remained. Assuming that the rotted trees do not deviate from the fecundity regression, postharvest capsule production is 1492, or 1 1.1 YO of preharvest values. The proportion of total seeds produced by obviously inferior (rotted) trees increases from 6.2% before harvest to 60% after harvest.
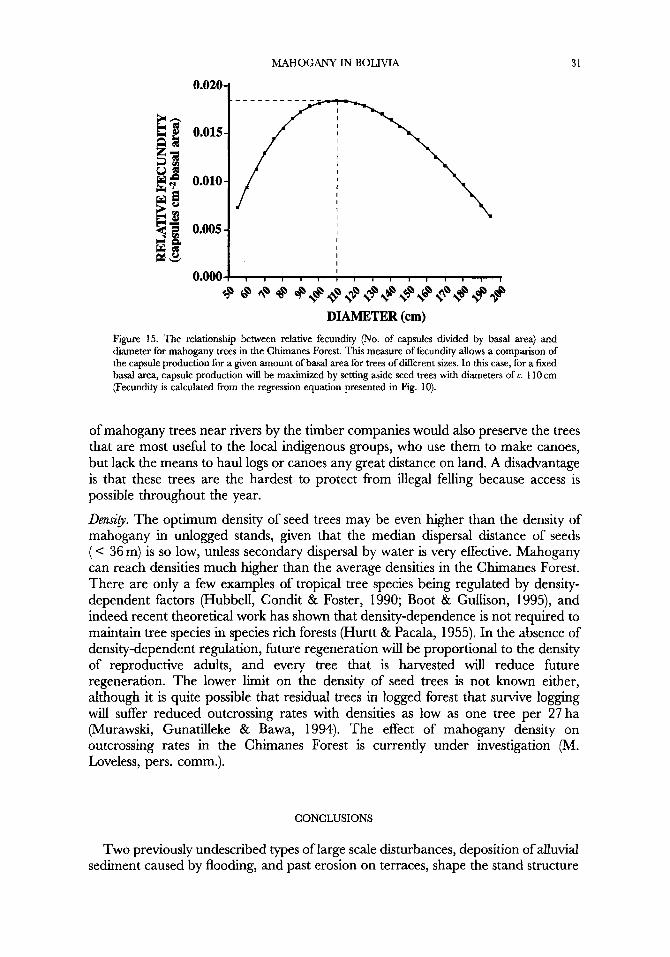
Postharvest fecundity can be increased by requiring companies to leave larger seed trees. Requiring a percentage of total basal area of the commercial stand to be preserved, rather than a percentage of the number of trees, will eliminate the incentive to leave the smallest trees possible. The size of seed trees should be chosen to maximize fecundity for a given amount of basal area (opportunity cost to loggers). For mahogany in the Chimanes Forest, this occurs between 90 and 130 cm dbh (Fig. 15). Smaller trees in this range are preferable because of their higher reproductive value.
Location. The existing management plan for seed trees does not account for the localized nature of regeneration sites. Seed trees are left at a uniform low density throughout the forest, rather than being concentrated in areas with the highest potential for regeneration. If the number of seed trees is limited, there is little point in leaving seed trees in logged forest where regcncration rarely occurs. Not only do seedlings that survive harvest in general not survive until commercial size (Fig. 9), but the post harvest density (0.036 trees ha-') results in a spacing of trees so much greater than median dispersal distances (spacing of 527 m between seed trees vs median seed dispersal distance of 36m) that it seems unlikely that they could colonize most regeneration sites should they appear.
Seed trees should be concentrated in areas that are most suitable for regeneration. Our results suggest that suitable sites are near rivers, particularly active rivers that are flooding, or in zones where the forest has been recently killed by floods. These areas can be identified easily with satellite photos. An additional benefit is that secondary water dispersal of seeds should be greater for trees near rivers. Reducing the felling
MAHOGANY IN BOLIVIA
0.020l 31
o . o o o ! , I I I , I I , I I , , I , I
9 9 4Q eo +,$$10@@.5P@Q\9+@@@$@ DIAMETER (cm)
Figure 15. The relationship between relative fecundity (No. of capsules divided by basal area) and diameter for mahogany trees in the Chimanes Forest. This measure of fecundity allows a comparison of the capsule production for a given amount of basal area for trees of different sizes. In this case, for a fixed basal area, capsule production will be maximized by setting aside seed trees with diameters of c. 1 10 cm (Fecundity is calculated from the regression equation presented in Fig. 10).
of mahogany trees near rivers by the timber companies would also preserve the trees that are most useful to the local indigenous groups, who use them to make canoes, but lack the means to haul logs or canoes any great distance on land. A disadvantage is that these trees are the hardest to protect from illegal felling because access is possible throughout the year.
Densip. The optimum density of seed trees may be even higher than the density of mahogany in unlogged stands, given that the median dispersal distance of seeds ( C 36 m) is so low, unless secondary dispersal by water is very effective. Mahogany can reach densities much higher than the average densities in the Chimanes Forest. There are only a few examples of tropical tree species being regulated by density- dependent factors (Hubbell, Condit & Foster, 1990; Boot & Gullison, 1995), and indeed recent theoretical work has shown that density-dependence is not required to maintain tree species in species rich forests (Hurtt & Pacala, 1955). In the absence of density-dependent regulation, future regeneration will be proportional to the density of reproductive adults, and every tree that is harvested will reduce future regeneration. The lower limit on the density of seed trees is not known either, although it is quite possible that residual trees in logged forest that survive logging will suffer reduced outcrossing rates with densities as low as one tree per 27ha (Murawski, Gunatilleke & Bawa, 1994). The effect of mahogany density on outcrossing rates in the Chimanes Forest is currently under investigation (M. Loveless, pers. comm.).
CONCLUSIONS
Two previously undescribed types of large scale disturbances, deposition of alluvial sediment caused by flooding, and past erosion on terraces, shape the stand structure

32 R. E. GULLISON ETAL.
and population dynamics of mahogany in the Chimanes Forest. Mahogany regenerates after these episodic disturbances, and its stands are made up of one or a few cohorts. The long time between disturbances favours late maturation. Mahogany trees allocate their photosynthates to growth until they reach 80cm dbh, at which point they increase allocation to reproduction. Unfortunately, this coincides with the minimum cutting diameter, and the postharvest seed production is substantially reduced. The polycyclic selective harvesting is misguided given the even-aged stands. This study shows why management plans must be based on a sound understanding of the ecology of the target species.
It is too late to manage for the sustained production of mahogany in the Chimanes Forest. One company finished its supply of mahogany three years ago, and two others admit to haLing only a one or two year supply left. AU three companies are in the process of buying illegally felled mahogany trees from the Multiethnic Indigenous Territory located in the middle of the Chimanes Forest. A management plan that included a much lower cut of mahogany, the harvest of other commercial species, and the concentration of seed trees in areas conducive to mahogany regeneration would have been more appropriate. Luckily the density of mahogany is low, and little structural damage has been done to the forest even though the harvest levels are unsustainable at the species level (Gullison & Hardner, 1993).
The same larger scale hydrological disturbances that structure mahogany populations may determine the population dynamics of many other tree species. If this is so, the principles from this study can help design management plans for future commercial species in the Chimanes Forest should there be political will to do so.
ACLWOWLEDGMENTS
Thanks to Jhonny Galloso, Valentin and Rudolf0 Garcia, Lucio and Joselo Isita, Raul Gonzales, Claudio Leafio, Gregory Dicum, Jared Hardner, Corine Vrie- sendorp, and The Grand Chimanes Council for valuable help in the field. Dick Rice and Hector Claure handled the logistics of the project. Henry Horn, Lyn Loveless, Jake Overton, Laura Snook, Kristina Stinson, Corine Vriesendorp, John Terborgh and two anonymous reviewers read and commented upon previous drafts of this paper. Ruth Silva translated the paper into Spanish. Steve Pacala, Don Stratton, Andy Dobson and Henry Horn have provided sound advice and supervision throughout the project. The following institutions or companies have provided financial assistance: National Science Foundation and the Office of Forestry, Environment and Natural Resources, Bureau of Science and Technology, of the U.S. Agency for International Development under NSF grant No. BSR-9100058; BOLFOR Bolivia’s Sustainable Forest Management Project funded by USAID and the Government of Bolivia through the Ministry of Sustainable Development and the Environment; Thompson Mahogany Company; Industria Maderera ‘San Francisco’ S.R.L.; Hermann Miller, Inc.; the U.S.D.A. Forest Service, US. Agency for International Development/Bolivia; International Tropical Timber Organiza- tion PD 88/90; and the National Science and Engineering Research Council (Canada).

MAHOGANY IN BOLIVIA 33
REFERENCES
Bell G. 1980. The costs of reproduction and their consequences. American Naturalist 116: 45-76. Boot RGA, Gullison RE. 1995. Approaches to developing sustainable extraction systems for tropical forest
products. Ecologizal Applications 5: 896-903. Brokaw NVL. 1987. Gap-phase regeneration of three pioneer tree species in a tropical forest.Jouma1 $Ecology 75:
9-19. Burniske GR. 1994. Final Evaluation - Phase 1. Consemation, m a n a g m t , util&ation, integral and sustained use oftheforests
o f the Chimanes Region o f thz Deparhmt o f E l Beni, Bolivia. Yokohama: International Tropical Timber Organization.
Carling PA. 1992. In-stream hydraulics and sediment transport. In: Calow P, Petts GE, eds. 7he rivers handbook: hydrologicel and ecologkal principles. London: Blackwell Scientific Publications, 10 1-1 25.
Clark JS. 1991a. Disturbance and population structure on the shifting mosaic landscape. Ecology 72: 1 1 19-1 137.
Clark JS. 1991b. Disturbance and life history on the shifting mosaic landscape. Ecolagy 72: 1102-1 118. Colinvaux PA, Miller MC, Liu K, Steinitz-Kannan My Frost 1. 1985. Discovery of permanent Amazon
lakes and hydraulic disturbance in the upper Amazon Basin. Nature 313: 4245. Crow TR. 1980. A rain forest chronicle: a 30-year record of change in structure and composition at El Verde,
Puerto Rico. Bwhopica 12: 42-55. De Steven D. 1988. Light gaps and long-term seedling performance of a Neotropical canopy tree (Optoyx
panamasis, Leguminosae). 30~ml of Tropical Ecology 4: 407-41 1. del Aguila M. 1995. Tree diversity in the Bosque Chimanes, Beni, Bolivia. Unpublished Bachelor’s thesis.
University of Trinidad, Bolivia. Eis S, Garman EH, Ebel LF. 1965. Relation between cone production and diameter increment of douglas fir
(Pseudotsqa menriesii (Mirb.). Francon), grand fir (Abies grandis Dougl.) and western white pine (Enus monticola Dougl.). CanadianJoumal O f B o h y 43: 1553-1559.
Foster RB, Arce BJ, Wachter TS. 1986. Dispersal and the sequential plant communities in Amazonian Peru flood plain. In: Estrada A, Fleming TH, eds. Fn@uores and seed dispersal. Dordrecht: Dr W. Junk Publishers,
FrangiJL, Lug0 AE. 1991. Hurricane damage to a flood plain forest in the Luquillo Mountains of Puerto Rico.
Frost I, Miller MC. 1987. Late Holocene flooding in the Ecuadorian rain forest. Freshwater Biology 18:
Goita L. 1990. Infom tecnico-econaico: pryecto de acciones forestales iniciales (preliminar). Unpublished project report. Government of Bolivia. 1993. Consmation, management, haruesting, and integrated and sustained use offorests in the
Chimanes Repion, Beni, Bolivia. International Tropical Timber Organization Project Proposal PD 33/93. Yokohama.
Gullison RE, Hubbell SP. 1992. Regeneracion natural de la mara (SWietenia macrophylla) en El Bosque Chimanes, Bolivia. Ecologia en Bolivia 19: 43-56.
Gullison RE, Hardnerg. 1993. The effects of road design and harvest intensity on forest damage caused by selective logging: empirical results and a simulation model from the Bosque Chimanes, Bolivia. Forest Ecology and M a n a g m t 59: 1-14.
Hartshorn GS. 1989. Gap-phase dynamics and tropical tree species richness. In: Holm-Nielsen LB, Nielsen IC, Balsev H. Tropical Forarts: botanical &namics, speciation, and diversi&. London: Academic Press Limited, 65-73.
Hubbell SP, Foster RB. 1986. Canopy gaps and the dynamics of a Neotropical forest. In: Crawley MJ, ed. Plant Ecolosy. Oxford: Blackwell Scientific Publishers, 77-95.
Hubbell SP, Condit R, Foster R. 1990. Presence and absence of density dependence in a neotropical tree community. Philosophical Trmactiom ofthe R y a l So&& OfLondon 3 330: 269-281.
Hurtt GC, Pacala SW. 1995. The consequences of recruitment limitation: reconciling chance, history, and competitive differences between plants. Journal of %oretical Biology 176: 1-12.
Kalliola R, Makinen Y, Salo J. 1988. Muse0 de Historia Natural Jar,& Prado’No. 19. Regenmeracwn natural de selum en la Amazonia P m a n a 2: Autecolagia de a@nm esfiecies sucesionales. Lima: Universidad Nacional Mayor de San Marcos.
Kallioli R, Salo J, Puhakka My Rajasilta M. 1991. New site formation and colonizing vegetation in primary succession on the western Amazon floodplains. Journal OfEcolasy 79: 877-901.
Kalliola R, Salo J, Puhakka M, Rajasilta My Hame T, Neller RJ, Rasanen ME, Danjoy Arias WA. 1992. Upper Amazon channel migration: implications of vegetation perturbance and succession using bitemporal Landsat MSS images. NatuatunuirsenSchaJien 79: 75-79.
565-572.
Biotropica 23: 324-335.
443453.
Lamb FB. 1966. Mahogaany oftropical America: its ecolosy and m a n a g m t . Ann Arbor: University of Michigan Press. Lang GE, Kuight IM. 1983. Tree growth, mortality, recruitment, and canopy gap formation during a 10-year
Loehle C. 1988. Tree lie history strategies: the role of defences. CanadianJoumal ofForest Research 18: 20S222. period in a tropical moist forest. Ecolosy 64: 1075-1080.

34 R. E. GULLISON ETAL.
Martinez-Run08 M, Spruiihan J, Pinero D. 1988. The demography of tropical trees in the context of forest gap dynamics: the case of the Asfrocarywn mexiGanum at Los Tuxtlas Tropical Rain Forest. In: Davy AJ, Hutchings MJ, Watkinson AR eds. P h t Population Ecotogv. OxFord Blackwell Scientific Publishers, 293-31 3.
Murawski DA, Gnnatilleke IAUN, BawP KS. 1994. The effects of selective logging on inbreeding in Shorea mgistq!~5d (Diptocarpaceae) from Sri Lanka. C o n 1 6 Binlogp 8: 997-1002.
Nelson BW, Kaps V, AdamsJB, Oliveira WJ, Bra- OPG, do Amaral IL. 1994. Forest disturbance by large blowdowns in the Brazilian Amazon. Ecology 75: 853458.
Pemhgton TD. 1981. Flora Nmtropica Monograph No. 28: Melkeae. New York New York Botanical Garden. Poore D, Burgess P, Palmer J, Rietbergen, Synnott T. 1989. No timber without trees: suctainabilip in the tropical
foresf. London: Earthscan. Puhakka M, Kalliola R, Rajasilta M, Salo J. 1992. River types, site evolution and successional vegetation
patterns in Peruvian Amazonia. Journal ofBWgeogra& 19: 651465. Rfmos J, Grace J. 1990. The effects of shade on the gas exchange of seedlings of four tropical trees from Mexico.
Functional EEology 4: 667477. Salo J, W o l a R, Hnkkinen I, Makinen Y, Niemela P, Puhakka M, Coley PD. 1986. River dynamics
and the diversity of Amazon lowland forest. ScimU 322: 254-258. Salo J, W o l a R. 1991. River dynamics and natural forest regeneration in the Peruvian Amazon. In: Gomez-
Pompa A, Whitmore TC, Hadley M, eds. Rainforest resew& and management. Park Ridge: Parthenon Publishing Group, 245-256.
Snook LK. 1993. Stand dynamics of mahogany (SWietenia macrophylla King) and associated species after fire and humcane in the tropical forests of the Yucatan Peninsula, Mexico. Doctoral Dissertation, Yale School of Forestry and Environmental Studies. Ann Arbor, M I University Microfilms International #9317535, 254 pp.
Stearns S. 1976. Life-history tactics: a review of the ideas. n2e &rtcrb Revim OfBiology 51: 3 4 7 . Styles BT, Khosla PK. 1976. Cytology and reproductive biology of Meliaceae. In: Burley J, Styles BT, eds.
Tropud trees: variatim, br+ and conmuntion. London: Academic Press Inc. Ltd., 61457. Uhl C, Ka-JB. 1990. Deforestation, fire susceptibility, and potential tree responses to fire in Eastern
Amazon. Ecology 71: 437-449. Verissimo A, Barreto P, Tnrifo R, Uhl C. 1995. Extraction of a high-value natural resource from Amazonia:
the case of mahogany. Forest Ecologv and Managm2mf 72: 3940. Zar JH. 1984. BiOstatufi~d A M ~ s ~ s . Englewood Cliffs: Prentice-Hall.