
Ecological requirements of Ostracoda (Crustacea) in three types of springs in Turkey
9
Limnologica 36 (2006) 172–180 Ecological requirements of Ostracoda (Crustacea) in three types of springs in Turkey Okan Ku¨lko¨ylu¨og˘lu , Faruk Yılmaz Department of Biology, Faculty of Arts & Science, Abant I ˙ zzet Baysal University, Bolu-Turkey Received 10 November 2005; received in revised form 7 March 2006; accepted 14 March 2006 Abstract Seasonal occurrence of six ostracod species (Psychrodromus olivaceus, Potamocypris villosa, Ilyocypris inermis, Candona candida, Candona neglecta, and Cypridopsis vidua) varied from May 2003 to June 2004 in three types of spring (Limnocrene, Helocrene, Rheocrene). When I. inermis was only found in the limnocrene spring each month, P. olivaceus was common in all types of springs. The other four ostracods were found in the helocrene spring in fall, winter, and spring seasons. The unweighted pair group mean averages dendrogram separated three main clusters. The first cluster includes two species (C. vidua, C. candida) while I. inermis was found in the second cluster and P. olivaceus, C. neglecta, and P. villosa were found in the third. The occurrence of P. olivaceus and C. neglecta and P. villosa and C. candida was strongly correlated to each other (r ¼ 0.750 and 0.850, respectively). The Pearson correlation analyses showed a strong negative relationship between C. neglecta and water temperature (r ¼0.607), but other species did not show any significant correlation to any of the environmental variables used (P40.05). Canonical correspondence analyses (CCA) displayed all species closer to the center of the dendrogram, indicating high levels of species tolerances to environmental variables. Thus, the first axis of the CCA diagram explained 80% of the relationship between six species and five environmental variables. r 2006 Elsevier GmbH. All rights reserved. Keywords: Ecology; Springs; Ostracoda; Optima estimate; Tolerances; Turkey Introduction Hynes (1970) defined three types of spring: helocrene, limnocrene and rheocrene. In a helocrene spring, water seeps out off the ground slowly and is usually temporarily confined to small holes or ditches. Thus, these shallow springs are profoundly affected by seasonal and temporal changes. In limnocrene springs, however, water comes out of the ground and creates a pond at the source, before flowing out slowly. The pond is usually deeper than in helocrene springs, so that water is permanent in limnocrene springs. Conversely, there is no pond or pool formation in rheocrene spring, where water flows away on the streambed. Such physical differences amid these springs also cause differences in chemical and biological structure in each type, resulting different species assemblages. As regards, the observa- tion of the ostracods assemblage in the long term not only provides detailed knowledge about ecological and biological characteristics of these springs, but also provides information on the relationship between ecological conditions and species habitat preferences. Therefore, interpretation of such data results a better ARTICLE IN PRESS www.elsevier.de/limno 0075-9511/$ - see front matter r 2006 Elsevier GmbH. All rights reserved. doi:10.1016/j.limno.2006.03.003 Corresponding author. Tel.: +374 254 1226; fax: +374 253 4642. E-mail address: [email protected] (O. Ku¨lko¨ylu¨og˘lu).
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Ecological requirements of Ostracoda (Crustacea) in three types of springs in Turkey

ARTICLE IN PRESS
0075-9511/$ - se
doi:10.1016/j.lim
�CorrespondE-mail addr
Limnologica 36 (2006) 172–180
www.elsevier.de/limno
Ecological requirements of Ostracoda (Crustacea) in three types
of springs in Turkey
Okan Kulkoyluoglu�, Faruk Yılmaz
Department of Biology, Faculty of Arts & Science, Abant Izzet Baysal University, Bolu-Turkey
Received 10 November 2005; received in revised form 7 March 2006; accepted 14 March 2006
Abstract
Seasonal occurrence of six ostracod species (Psychrodromus olivaceus, Potamocypris villosa, Ilyocypris inermis,Candona candida, Candona neglecta, and Cypridopsis vidua) varied from May 2003 to June 2004 in three types ofspring (Limnocrene, Helocrene, Rheocrene). When I. inermis was only found in the limnocrene spring each month,P. olivaceus was common in all types of springs. The other four ostracods were found in the helocrene spring in fall,winter, and spring seasons. The unweighted pair group mean averages dendrogram separated three main clusters. Thefirst cluster includes two species (C. vidua, C. candida) while I. inermis was found in the second cluster and P. olivaceus,
C. neglecta, and P. villosa were found in the third. The occurrence of P. olivaceus and C. neglecta and P. villosa andC. candida was strongly correlated to each other (r ¼ 0.750 and 0.850, respectively). The Pearson correlation analysesshowed a strong negative relationship between C. neglecta and water temperature (r ¼ �0.607), but other species didnot show any significant correlation to any of the environmental variables used (P40.05). Canonical correspondenceanalyses (CCA) displayed all species closer to the center of the dendrogram, indicating high levels of species tolerancesto environmental variables. Thus, the first axis of the CCA diagram explained 80% of the relationship between sixspecies and five environmental variables.r 2006 Elsevier GmbH. All rights reserved.
Keywords: Ecology; Springs; Ostracoda; Optima estimate; Tolerances; Turkey
Introduction
Hynes (1970) defined three types of spring: helocrene,limnocrene and rheocrene. In a helocrene spring, waterseeps out off the ground slowly and is usuallytemporarily confined to small holes or ditches. Thus,these shallow springs are profoundly affected byseasonal and temporal changes. In limnocrene springs,however, water comes out of the ground and creates apond at the source, before flowing out slowly. The pond
e front matter r 2006 Elsevier GmbH. All rights reserved.
no.2006.03.003
ing author. Tel.: +374 254 1226; fax: +374 253 4642.
ess: [email protected] (O. Kulkoyluoglu).
is usually deeper than in helocrene springs, so that wateris permanent in limnocrene springs. Conversely, there isno pond or pool formation in rheocrene spring, wherewater flows away on the streambed. Such physicaldifferences amid these springs also cause differences inchemical and biological structure in each type, resultingdifferent species assemblages. As regards, the observa-tion of the ostracods assemblage in the long term notonly provides detailed knowledge about ecological andbiological characteristics of these springs, but alsoprovides information on the relationship betweenecological conditions and species habitat preferences.Therefore, interpretation of such data results a better

ARTICLE IN PRESSO. Kulkoyluoglu, F. Yılmaz / Limnologica 36 (2006) 172–180 173
understanding of the effects of the environmentalfactors (e.g. anthropogenic factors) on the ecologicalrequirements of the species.
Springs are unique aquatic environments because ofhaving almost constant physical and chemical condi-tions all year around (Hynes, 1970; Kulkoyluoglu, 1999;Kulkoyluoglu & Vinyard, 1998, 2000; Sarkka, Levonen,& Makela, 1997). Such stable characteristics providespecies-specific conditions, in which it is expected to seecertain kinds of species diversity and life cycles. There-fore, any changes in species assemblages (e.g. presenceor absence) may be interpreted as the effects of externalfactors (i.e. anthropogenic factors) or species introduc-tion to these habitats. On the other hands, speciesdiversity can also be dependant on the types of springssince each type has its own specific conditions (Sarkkaet al., 1997). However, due to lack of knowledge on bothcharacteristics of springs and species habitat prefer-ences, interpretation of species presence and absence(binary data) may not be meaningful (Kulkoyluoglu &Vinyard, 1998).
Class Ostracoda is diverse and a widespread group inthe world but one in which most species are sensitive tothe changes in environmental conditions. Therefore,they can be used as bioindicators to determine waterquality, spatial and temporal changes in environment(Kulkoyluoglu, 2004; Kulkoyluoglu & Dugel, 2004;Kulkoyluoglu & Vinyard, 1998, 2000; Mezquita, Grif-fiths, Domınguez, & Lozano-Quilis, 2001; Ruiz, Gonza-lez-Regalado, Borrego, Abad, & Pendon, 2004).Although the idea of using ostracods as bioindicatorspecies is critical, it requires detailed knowledge of therelationship between ecology of species and their spatialdistribution. Without adequate knowledge, the conceptof using ostracods as bioindicators may result inspurious conclusions.
Almost nothing is known about the ecologicalpreferences of spring-dwelling ostracods in Turkey.Therefore, this study aims (1) to contribute the knowl-edge on the ecology of these ostracods in three types ofspring, (2) to outline their seasonal differences in eachspring, and (3) to focus on the importance of ostracodsas bioindicators.
Material and methods
The three springs studied are located around LakeAladag (40136026400N 31140051400E, at 1347m), 30 kmsouth of Bolu (Fig. 1). Among the three springs sampledmonthly from May 2003 to June 2004, spring one wasdefined as rheocrene, spring two was limnocrene andspring three was helocrene (Table 1). The area aroundthe springs is dominated with pine trees (Pinus sylvestris)and annual plants, and is a well known with touristattraction that introduces a continuous anthropogenic
influence. Each spring has its own environmentalcharacteristics; both rheocrene and limnocrene springswere present all year around, but the helocrene springoccurred only from October 2003 to the end of June2004, and dried out during rest of the study period.Because of harsh winter conditions during the months(January–March) of the year 2004, sampling was notpossible. There was no macrophyte growth in theshallow (ca. 15 cm of depth) and stagnant helocrenespring (70 cm in diameter), where bottom substrateconsists of 70% silt and 30% coarse sand. In contrast,although we did not measure the actual water current,rheocrene spring was slowly flowing at about laminarrate, where the source of the spring bottom includes60% very coarse sand, 30% silt and 10% fine gravelcovered by water-cress and some other macrophytes.The limnocrene spring has about 15 cm of depth with50 cm of diameter. The spring bottom compromiseabout 60% silt, 30% sand and 10% of clay particlescovered with about 50% of macrophytes.
Monthly samples were taken from each spring with aMuller hand net (250 mm mesh size) and were fixedin 70% alcohol in situ. Then after, material was filteredunder tap water with standardized sieves (0.25:0.50:1.0:2.0mm in mesh size, respectively). Ostracod samplesand other invertebrates were separated from each otherunder a stereomicroscope and fixed again in 70% ofalcohol. Specimen dissection was done in lactophenolsolution and mounted in a permanent slide. Thetaxonomic investigation was done based on adultindividuals soft body parts and valve morphologyfollowing taxonomic keys of Meisch (2000) and theindex of bibliography of nonmarine ostracods of Kempf(1997).
A total of seven different variables were measured instudy site: dissolved oxygen (DO (mg/L)), percentoxygen saturation (%S), salinity (Sal (ppt)), watertemperature (t (w) (1C)), electrical conductivity (EC(mS/cm)), pH, and redox potential (Eh (mV)). YSI-85DO-meter was used to measure DO, (%S), Sal, t 1C, andEC, while Eh and pH were measured with Hanna HI-98150 pH/ORP meter. Coordinates and elevation weredetermined with a geographical positioning system(GPS Garmin-45XL).
Species–environmental relationship was examined byusing canonical correspondence analysis (CCA) wherefive environmental variables with six species found in 12months were used. Also, to delete arch-effect andmulticolinearity, rare species were downweighted andtaxa encountered less than two were excluded from theanalyses (Ter Braak 1986, 1995). Species estimatedoptimum (uk) and tolerance (tk) values were calculatedfollowing the application of De Almeida and Gil (2001),Birks, Line, Juggins, Stevenson, and Ter Braak (1990),Ter Braak and Barendregt (1986), and Kulkoyluogluand Dugel (2004). By calculating the tolerance and

ARTICLE IN PRESS
Fig. 1. Locations of three springs (%) around Lake Aladag (Bolu, Turkey). Spring one (Spr.1) is rheocrene, spring two (Spr. 2) is
limnocrene, and spring three (Spr. 3) is helocrene. Scale bar ¼ 60m.
O. Kulkoyluoglu, F. Yılmaz / Limnologica 36 (2006) 172–180174
estimated optimum values of a species, it is possible toreconstruct past historical environmental conditions in aparticular habitat (Birks, 1995). Taxonomic assemblagesof six species were examined with a clustering analysis ofunweighted pair group mean averages (UPGMA). Allstatistical analyses were conducted using the multi-variate statistical package (MVSP) program version 3.1(Kovach, 1998) and SPSS program version 11.01.
Results
Six ostracod species (Psychrodromus olivaceus, Pota-
mocypris villosa, Ilyocypris inermis, Candona candida,Candona neglecta, and Cypridopsis vidua) were found inall three kinds of springs. Of these, there was acontinuous occurrence of I. inermis in the limnocrenespring for each month, and P. olivaceus was common in
all three springs. The remaining four species wererecovered from the helocrene spring in fall, winter, andspring seasons. Based on the species binary (presence/absence) data, UPGMA dendrogram separated threemain clusters. Two species (C. vidua and C. candida)were found together in the first cluster, while secondcluster comprised only one non-swimming species,I. inermis, and the third cluster had three other species(P. olivaceus, C. neglecta, and P. villosa), of which thefirst two are also non-swimmers. The Pearson correla-tion analyses showed a strong negative relationshipbetween C. neglecta and water temperature (r ¼ �0.607,Po0.05), when other species did not show anysignificant correlation to any of those environmentalvariables used (P40.05). However, seasonal occurrenceof P. olivaceus and C. neglecta (r ¼ 0.750) and P. villosa
and C. candida (r ¼ 0.850) was strongly related. CCA,explaining 80% of the relationship between six species

ARTICLE IN PRESS
Table 1. Monthly measurements (e.g., May-03 represents May 2003) of environmental variables at three stations (springs)
Date DO %S Eh EC TDS pH Sal t(w) Numbers of specimen
Male Female
Rheocrene spring
May-03 12.06 124.30 �43.60 90.00 60.30 7.75 0.00 13.00 10 Cc
Jun-03 6.68 69.40 �67.10 75.10 50.32 8.03 0.00 17.00
Jul-03 4.33 44.60 �36.30 138.90 93.06 7.67 0.10 15.30
Aug-03 7.37 72.00 125.20 83.88 0.10 11.70
Sep-03 6.31 61.30 �32.40 124.70 83.55 7.71 0.10 13.60
Oct-03 5.28 50.00 �37.10 118.10 79.13 7.56 0.10 12.50 5 Po 2 Pv, 14 Po
Nov-03 5.84 55.70 �37.60 121.20 81.20 7.54 0.10 11.80
Dec-03 10.09 86.00 �12.90 95.00 63.65 7.20 0.00 7.50 6 Po
Apr-04 7.58 75.60 �34.20 124.90 83.68 7.63 0.10 13.50 1 Po
May-04 6.05 63.30 �41.70 73.10 48.98 7.74 0.00 14.90 1 Po
Jun-04 7.72 77.80 �4.20 120.10 80.47 7.05 0.10 13.80 1 Po 1 Po
Mean 7.21 70.91 �34.71 109.66 73.47 7.59 0.06 13.15
Limnocrene spring
Jun-03 5.18 64.10 �75.40 131,40 88.04 8,30 0.10 15.30 42 Ii
Jul-03 4.52 41.10 �5.40 137.10 91.86 7.49 0.10 17.60 30 Ii
Aug-03 5.80 57.40 127.90 85.69 0.10 13.30 21 Ii
Sep-03 4.96 52.00 �39.20 127.80 85.63 7.62 0.10 16.60 9 Ii
Oct-03 4.86 50.40 �27.40 126.80 84.96 7.24 0.10 11.60 39 Ii, 2 Po
Nov-03 5.09 52.10 �38.20 87.50 58.63 7.57 0.00 11.30 7 Ii, 2Po
Dec-03 9.62 76.90 �22.00 74.40 49,85 7.62 0.00 5.70 12 Ii
Apr-04 8.71 89.10 �35.30 61.10 40.94 7.64 0.00 16.40 1 Po 57 Ii, 1Po
May-04 2.98 32.40 �45.50 121.70 81.54 7.66 0.10 17.10 69 Ii
Jun-04 5.27 54.10 �3.20 132.40 88.71 7.04 0.10 16.40 1 Po 42 Ii, 9 Po
Mean 5.70 56.96 �32.40 112.81 75.58 7.58 0.07 14.13
Helocrene spring
Oct-03 5.34 50.70 �31.0 139.50 93.47 7.52 0.10 12.70 19 Cc 14 Cc, 36 Pv, 3Cv
Nov-03 5.19 43.40 �16.3 263.50 176.55 7.24 0.10 5.30 14 Cn 24 Cn, 9 Po, 9 Pv
Dec-03 10.83 89.10 �59.0 124.40 83.35 8.03 0.10 3.80 7 Cc, 4 Po 14 Cn, 4 Cv, 14Pv, 14 Po
Apr-04 8.94 80.50 �52.6 148.90 99.76 7.97 0.10 10.40 11 Cn, 5 Po 29 Cn, 4 Pv, 30 Po
May-04 3.65 42.50 �26.5 148.50 99.50 6.46 0.10 13.40 2 Cn, 1 Pv, 1 Po
Jun-04 5.28 72.00 �121.8 110.00 73.70 9.00 0.10 17.50 1 Cn 10 Cn, 3 Pv
Mean 6.54 63.03 �51.2 155.80 104.39 7.70 0.10 10.52
Abbreviations and variables (with respective units) include: (DO) dissolved oxygen, (t(w)) water temperature, (%S) percent oxygen saturation, pH,
(Eh) redox potential, (Sal) salinity, (EC) electrical conductivity, (TDS) total dissolved solids, (Cn) Candona neglecta, (Cc) C. candida, (Ii) Ilyocypris
inermis, (Cv) Cypridopsis vidua, (Po) Psychrodromus olivaceus, (Pv) Potamocypris villosa.
O. Kulkoyluoglu, F. Yılmaz / Limnologica 36 (2006) 172–180 175
and five environmental variables, showed all specieslocated closer to the center of the dendrogram. This mayimply high tolerance levels of ostracods to environ-mental variables, suggesting cosmopolitan characteris-tics of these species (but see discussion below). Althoughresults indicate similar occurrences among species,differences can be identified between ecological toler-ances and requirements of some species.
Discussion
Species were clustered in UPGMA dendrogram(Fig. 2) although they are in different taxonomic groups.For example, C. vidua and C. candida were found
together in the first cluster even though they belongto different genera. Similarly, although the secondcluster included only one rare non-swimming species,Ilyocypris inermis, three species belonging to threegenera (P. olivaceus, C. neglecta, and P. villosa) werefound in the third cluster, of which the first two are alsonon-swimmers. According to classic ecological view,complete competitors cannot coexist (Hardin, 1960). Ifthis is correct, the occurrence of ostracod species mayimply that the species in each cluster do not completewith each other, even though they were encountered inthe same place at the same time. This also suggests thatthe ecological preferences of each species may varydepending on species-specific characters (Kulkoyluoglu,2003; Kulkoyluoglu & Vinyard, 2000).

ARTICLE IN PRESS
UPGMA
Jaccard's Coefficient
P. villosa
P. olivaceus
C. neglecta
I. inermis
C. candida
C. vidua
0.04 0.2 0.36 0.52 0.68 0.84 1
I
II
III
Fig. 2. Unweighted pair group mean average (UPGMA) analysis places the species into three main clusters.
CCA joint plot
Axis 2
Axis 1
M03
J03Jl03 Ag03
S03
O03
N03
D03 A04M04
J04
CcIi Pv Po
-2.4
-4.9
-7.3
2.4
4.9
7.3
9.8
12.2
-2.4-4.9-7.3 2.4 4.9 7.3 9.8 12.2
DO
T(w)
EH
EC
pH
Cn Cv
Fig. 3. Canonical correspondence analysis (CCA) shows relationship between six species (triangle) and five environmental variables
(arrows) within 11 months (square). Explaining of abbreviations and variables see Table 1; e.g., M03 – May 2003 (month and year),
etc.
O. Kulkoyluoglu, F. Yılmaz / Limnologica 36 (2006) 172–180176
For example, C. vidua a cosmopolitan species isknown to have high tolerance levels to differentenvironmental variables (Kulkoyluoglu, 2000). Indeed,in the CCA ordination diagram, the species was locatedcloser to the center (Fig. 3) and did not show anysignificant correlation to the variables used in this study(Table 2). However, occurrence of this species can berelated to saline environments (Mezquita, Tapia, &Roca, 1999) or in some cases destruction of aquatichabitats (Kulkoyluoglu, 2005). Scharf (1998) empha-
sized that the disappearance of C. vidua in LakeArendsee was coincidental with the destruction ofsubmersed macrophytes by pollution. During our study,we did not observe any macrophyte growth in thehelocerene spring where conductivity and salinity valueswere found between the minimum and maximum rangesreported earlier in the literature (Kulkoyluoglu, 2000).
Another species occurred in the first UPGMAdiagram was a bisexual species, C. candida. LikeC. vidua, this species has been commonly reported from

ARTICLE IN PRESS
Table 2. Pearson correlation analyses showing significant relationships between six species and seven environmental variables
DO (%)S Eh EC pH Sal t(w) Cc Cn Cv Ii Pv Po
DO 1.000
11
(%)S 0.948** 1.000
11 11
Eh 0.040 �0.014 1.000
10 10 10
EC �0.721* �0.789** 0.171 1.000
11 11 10 11
pH 0.428 0.455 �0.471 �0.416 1.000
10 10 10 10 10
Sal �0.812** �0.827** 0.248 0.834** �0.415 1.000
11 11 10 11 10 11
t(w) �0.630* �0.382 0.069 0.155 �0.147 0.401 1.000
11 11 10 11 10 11 11
Cc 0.019 0.033 0.021 0.004 �0.251 0.086 �0.077 1.000
11 11 10 11 10 11 11 11
Cn 0.474 0.328 0.264 �0.069 0.269 �0.270 �0.607* �0.282 1.000
11 11 10 11 10 11 11 11 11
Cv �0.160 �0.222 �0.072 0.546 �0.020 0.223 �0.191 �0.189 0.321 1.000
11 11 10 11 10 11 11 11 11 11
Ii �0.419 �0.345 0.206 �0.095 �0.254 0.208 0.452 0.002 �0.023 �0.485 1.000
11 11 10 11 10 11 11 11 11 11 11
Pv 0.054 �0.075 0.028 0.115 �0.185 0.145 �0.427 0.850** 0.101 �0.048 0.031 1.000
11 11 10 11 10 11 11 11 11 11 11 11
Po 0.473 0.349 0.440 �0.274 0.152 �0.170 �0.478 0.207 0.750** �0.134 0.270 0.503 1.000
11 11 10 11 10 11 11 11 11 11 11 11 11
For abbreviations, see Table 1.
*Po0.05, **Po0.01.
O. Kulkoyluoglu, F. Yılmaz / Limnologica 36 (2006) 172–180 177
variety of aquatic habitats in Turkey. C. candida wasgenerally found in the undisturbed springs in Finland,where it preferred cool waters (Sarkka et al., 1997).Although the species was recorded as euchronal,occurring all year around, in both natural and manmadelakes (Keyser & Nagorskaya, 1998; Kulkoyluoglu, 2004;Yılmaz & Kulkoyluoglu, 2006), it was encountered onlytwice in both helocrene and rheocrene springs. Owing toits cool water preference, the species was called asoligothermophilic (Hiller, 1972). During our study, thespecies was also found at cool (ca. 13 1C) watertemperature (Table 1), confirming previous records.
Although its geographical distribution is rare inTurkey, I. inermis was frequently recorded fromlimnocrene spring where the minimum and maximumwater temperature ranged between 5.7 and 17.6 1C(average 14.13 1C). The CCA diagram (Fig. 3) displayeda positive relationship between the occurrence ofI. inermis and two variables (redox potential andtemperature), and a negative correlation to DO andpH. This implies that I. inermis can be found in waterswith lower oxygen content and low to moderate pH.Among the six species, I. inermis had the highest optima(uk ¼ 14.08) and the lowest tolerance (tk ¼ 2.54) valuesto water temperature (Table 3). This species was
reported from cold waters with less conductivity,corresponding to low salinity (Meisch, 2000; Mezquitaet al., 1999), whereas the Pearson correlation analysis(Table 2) failed to show any significant relationship tothose variables used in the present study. Overall, resultssuggest that the ecological tolerance levels of I. inermis
seem to be larger than previously known. However, dueto the rare occurrence of the species, ecological data isinsufficient to be certain of this idea at the moment.
There are three species (P. villosa, C. neglecta, andP. olivaceus) clustered in the third UPGMA diagram.While two species (P. villosa and C. neglecta) were onlyencountered in helocrene spring, the bisexual popula-tions of P. olivaceus were reported from all three typesof springs. Roca and Baltanas (1993) stated thatC. neglecta is a cold stenothermal and oligothermophilicspecies. Meisch (2000) pinpointed that this species canwithstand increased water temperature, and can inhabithypoxic conditions, where oxygen concentration dropsto as low as 3mgL�1 in summer. The distribution ofC. neglecta is well known in Holarctic regions with itswide tolerance to cold and organically rich habitats(Mischke, 2001). This finding is partially supported withthe present study that C. neglecta showed highestoptimum (uk ¼ 9.08) levels to DO (Table 3). According
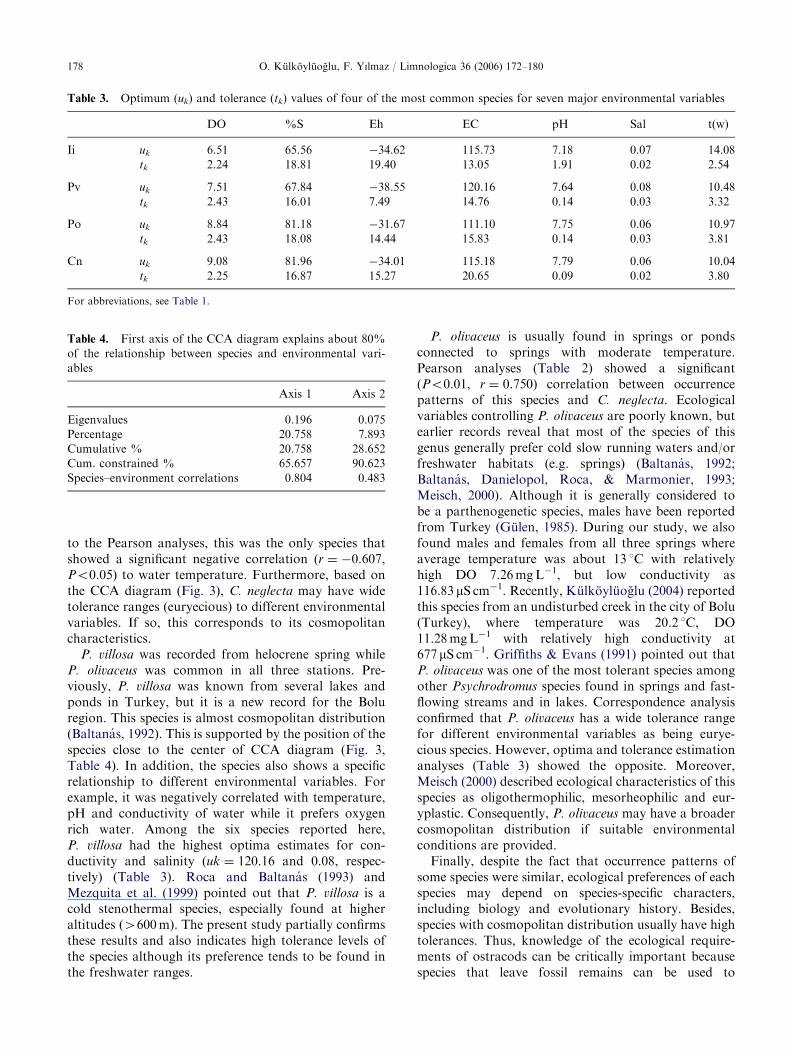
ARTICLE IN PRESS
Table 4. First axis of the CCA diagram explains about 80%
of the relationship between species and environmental vari-
ables
Axis 1 Axis 2
Eigenvalues 0.196 0.075
Percentage 20.758 7.893
Cumulative % 20.758 28.652
Cum. constrained % 65.657 90.623
Species–environment correlations 0.804 0.483
Table 3. Optimum (uk) and tolerance (tk) values of four of the most common species for seven major environmental variables
DO %S Eh EC pH Sal t(w)
Ii uk 6.51 65.56 �34.62 115.73 7.18 0.07 14.08
tk 2.24 18.81 19.40 13.05 1.91 0.02 2.54
Pv uk 7.51 67.84 �38.55 120.16 7.64 0.08 10.48
tk 2.43 16.01 7.49 14.76 0.14 0.03 3.32
Po uk 8.84 81.18 �31.67 111.10 7.75 0.06 10.97
tk 2.43 18.08 14.44 15.83 0.14 0.03 3.81
Cn uk 9.08 81.96 �34.01 115.18 7.79 0.06 10.04
tk 2.25 16.87 15.27 20.65 0.09 0.02 3.80
For abbreviations, see Table 1.
O. Kulkoyluoglu, F. Yılmaz / Limnologica 36 (2006) 172–180178
to the Pearson analyses, this was the only species thatshowed a significant negative correlation (r ¼ �0.607,Po0.05) to water temperature. Furthermore, based onthe CCA diagram (Fig. 3), C. neglecta may have widetolerance ranges (euryecious) to different environmentalvariables. If so, this corresponds to its cosmopolitancharacteristics.
P. villosa was recorded from helocrene spring whileP. olivaceus was common in all three stations. Pre-viously, P. villosa was known from several lakes andponds in Turkey, but it is a new record for the Boluregion. This species is almost cosmopolitan distribution(Baltanas, 1992). This is supported by the position of thespecies close to the center of CCA diagram (Fig. 3,Table 4). In addition, the species also shows a specificrelationship to different environmental variables. Forexample, it was negatively correlated with temperature,pH and conductivity of water while it prefers oxygenrich water. Among the six species reported here,P. villosa had the highest optima estimates for con-ductivity and salinity (uk ¼ 120.16 and 0.08, respec-tively) (Table 3). Roca and Baltanas (1993) andMezquita et al. (1999) pointed out that P. villosa is acold stenothermal species, especially found at higheraltitudes (4600m). The present study partially confirmsthese results and also indicates high tolerance levels ofthe species although its preference tends to be found inthe freshwater ranges.
P. olivaceus is usually found in springs or pondsconnected to springs with moderate temperature.Pearson analyses (Table 2) showed a significant(Po0.01, r ¼ 0.750) correlation between occurrencepatterns of this species and C. neglecta. Ecologicalvariables controlling P. olivaceus are poorly known, butearlier records reveal that most of the species of thisgenus generally prefer cold slow running waters and/orfreshwater habitats (e.g. springs) (Baltanas, 1992;Baltanas, Danielopol, Roca, & Marmonier, 1993;Meisch, 2000). Although it is generally considered tobe a parthenogenetic species, males have been reportedfrom Turkey (Gulen, 1985). During our study, we alsofound males and females from all three springs whereaverage temperature was about 13 1C with relativelyhigh DO 7.26mgL�1, but low conductivity as116.83 mS cm�1. Recently, Kulkoyluoglu (2004) reportedthis species from an undisturbed creek in the city of Bolu(Turkey), where temperature was 20.2 1C, DO11.28mgL�1 with relatively high conductivity at677 mS cm�1. Griffiths & Evans (1991) pointed out thatP. olivaceus was one of the most tolerant species amongother Psychrodromus species found in springs and fast-flowing streams and in lakes. Correspondence analysisconfirmed that P. olivaceus has a wide tolerance rangefor different environmental variables as being eurye-cious species. However, optima and tolerance estimationanalyses (Table 3) showed the opposite. Moreover,Meisch (2000) described ecological characteristics of thisspecies as oligothermophilic, mesorheophilic and eur-yplastic. Consequently, P. olivaceus may have a broadercosmopolitan distribution if suitable environmentalconditions are provided.
Finally, despite the fact that occurrence patterns ofsome species were similar, ecological preferences of eachspecies may depend on species-specific characters,including biology and evolutionary history. Besides,species with cosmopolitan distribution usually have hightolerances. Thus, knowledge of the ecological require-ments of ostracods can be critically important becausespecies that leave fossil remains can be used to

ARTICLE IN PRESSO. Kulkoyluoglu, F. Yılmaz / Limnologica 36 (2006) 172–180 179
reconstruct past historical conditions, estimate currenthabitat qualities, as well as predict future changes inaquatic habitats. Such an approach corresponds withthe idea of using ostracods as indicator species.However, this requires an understanding of (i) ecologicalrequirements and tolerance levels of individual speciesand (ii) the occurrence patterns (seasonality) of speciesin particular habitats. Yet, these are both poorly knownfor many ostracod species. Consequently, the value ofostracods as indicator species is not fully understoodenough and requires more detailed studies.
Acknowledgments
We thank to Dr. Ian P. Wilkinson (British GeologicalSurvey) for his kind help and suggestions on the firstdraft of this paper. Also, Ismail OZCAN and BugraS-ENGUL (Abant Izzet Baysal University) were thankedfor their help in the field study. FY appreciates to hisparent Emin and Rabiye YILMAZ for their continuoussupport during this study. Part of this study wassupported by Turkish Scientific and Technical ResearchCouncil (TUBITAK Project No. TBAG: 103T028).
References
De Almeida, S. E. P., & Gil, M. C. P. (2001). Ecology of
freshwater diatoms from the central region of Portugal.
Cryptogamie Algologie, 22, 109–126.
Baltanas, A. (1992). A contribution to the knowledge of the
cypridid ostracode fauna (Crustacea, Ostracoda, Cypridi-
dae) on the Iberian Peninsula, and a comparison with
adjacent areas. Archiv fur Hydrobiologie, 3, 419–452.
Baltanas, A., Danielopol, D. L., Roca, J. R., & Marmonier, P.
(1993). Psychrodromus betharrami n. sp. (Crustacea, Os-
tracoda): Morphology, ecology and biogeography. Zoolo-
gischer Anzeiger, 231, 39–57.
Birks, H. J. B. (1995). Quantitative palaeoenvironmental
reconstructions. In D. Maddy, & J. S. Brew (Eds.),
Statistical modelling of quaternary science data. QRA
technical guide no. 5 (pp. 161–254). Cambridge: Quaternary
Research Association.
Birks, H. J. B., Line, J. M., Juggins, S., Stevenson, A. C., &
Ter Braak, C. J. F. (1990). Diatoms and pH reconstruction.
Philosophical Transactions A of The Royal Society, London,
327, 263–278.
Griffiths, H. I., & Evans, J. G. (1991). Some freshwater
ostracods (Crustacea:Ostracoda) from South Wales. Fresh-
water Forum, 1, 64–72.
Gulen, D. (1985). Bisexual Ostracoda (Crustacea) populations
in Anatolia, Istanbul University Fen Fak. Biyoloji Mec-
muası (Seri B), 50, 81–86.
Hardin, G. (1960). The competitive exclusion principal.
Science, 131, 1292–1297.
Hiller, D. (1972). Untersuchungen zur Biologie und Okologie
Limnischer Ostracoden aus der Umgebung von Hamburg.
Archiv fur Hydrobiologie, 40(Suppl.), 400–497.
Hynes, H. B. N. (1970). The ecology of running waters.
Liverpool: Liverpool University Press.
Kempf, E. K. (1997). Index and bibliography of Nonmarine
Ostracoda Part 6. Sonderveroffentlichungen des Geolo-
gischen Instituts der Universitat zu Koln, 109, 1–143.
Keyser, D., & Nagorskaya, L. (1998). Ostracods in the vicinity
of Minsk, Belarus. Mitteilungen aus dem Hamburgischen
Zoologischen Museum und Institut, 95, 115–131.
Kovach, W. (1998). Multi-variate statistical package: Version
3.01, Pentraeth.
Kulkoyluoglu, O. (1999). Taxonomy, ecology and biogeo-
graphic distribution of spring water Ostracoda (Crustacea)
in Nevada. Ph.D. dissertation, University of Nevada, Reno,
USA.
Kulkoyluoglu, O. (2000). The importance of cosmopolitan and
indicator species of Ostracoda (Crustacea) in Turkey based
on some water parameters. In Sinop water product
conference proceedings (pp. 421–437).
Kulkoyluoglu, O. (2003). Ecology of freshwater Ostracoda
(Crustacea) from lakes and reservoirs in Bolu-Turkey.
Journal of Freshwater Ecology, 18, 343–347.
Kulkoyluoglu, O. (2004). On the usage of ostracoda (Crusta-
cea) as bioindicator species in the different aquatic habitats
in the Bolu region, Turkey. Ecological Indicators, 4,
139–147.
Kulkoyluoglu, O. (2005). On the relationship between ecology
and phenology of freshwater ostracods in Lake Golkoy
(Bolu, Turkey). Aquatic Ecology, 39, 295–304.
Kulkoyluoglu, O., & Dugel, M. (2004). Ecology ad spatio-
temporal patterns of Ostracoda (Crustacea) from Lake
Golcuk (Bolu, Turkey). Archiv fur Hydrobiologie, 160,
67–83.
Kulkoyluoglu, O., & Vinyard, G. L. (1998). New bisexual form
of Cavernocypris subterranea (Wolf, 1920) (Crustacea,
Ostracoda) from Idaho. Great Basin Naturalist, 58, 380–385.
Kulkoyluoglu, O., & Vinyard, G. L. (2000). The distribution
and the ecology of the freshwater Ostracoda (Crustacea)
collected from springs of Nevada, Idaho, and Oregon: A
preliminary study. Western North American Naturalist, 60,
291–303.
Meisch, C. (2000). Freshwater Ostracoda of western and
central Europe. Sußwasserfauna von Mitteleuropa, 8, I-xii,
1–522. Heidelberg: Spektrum Akademischer Verlag.
Mezquita, F., Tapia, G., & Roca, R. C. (1999). Ostracoda
from springs on the eastern Iberian Peninsula: Ecology,
biogeography and paleolimnological implications. Palaeo-
geography, Palaeoclimatology, Paleoecology, 148, 65–85.
Mezquita, F., Griffiths, H. I., Domınguez, M. I., & Lozano-
Quilis, M. A. (2001). Ostracoda (Crustacea) as ecological
indicators: A case study from the Iberian Mediterranean
brooks. Archiv fur Hydrobiologie, 150, 545–560.
Mischke, S. (2001). Mid and late Holocene palaeoenvironment
of the lakes Eastern Juyanze and Sogo Nur in NW China,
based on ostracod species assemblages and shell chemistry.
Berliner Geowissenschaftliche Abhandlungen, E 35.
Roca, J. R., & Baltanas, A. (1993). Ecology and distribution of
Ostracoda in Pyrenean springs. Journal of Crustacean
Biology, 13, 165–174.
Ruiz, F., Gonzalez-Regalado, M. L., Borrego, J., Abad, M.,
& Pendon, J. G. (2004). Ostracoda and foraminifera as

ARTICLE IN PRESSO. Kulkoyluoglu, F. Yılmaz / Limnologica 36 (2006) 172–180180
short-term tracers of environmental changes in very
polluted areas: The Odiel Estuary (SW Spain). Environ-
mental Pollution, 129, 49–61.
Sarkka, J., Levonen, L., & Makela, J. (1997). Meiofauna of
springs in Finland in relation to environmental factors.
Hydrobiologia, 347, 139–150.
Scharf, B. W. (1998). Eutrophication history of Lake Arendsee
(Germany). Palaeogeography, Palaeoclimatology, Palaeoe-
cology, 140, 85–96.
Ter Braak, C. J. F. (1986). Canonical correspondance analysis:
A new eigenvector technique for multivariate direct
gradient analysis. Ecology, 67, 1167–1179.
Ter Braak, C. J. F. (1995). Ordination. In R. H. G. Jongman,
C. J. F. Ter Braak, & O. F. R. Van Tongeren (Eds.), Data
analysis in community and landscape ecology. Cambridge:
Cambridge University Press.
Ter Braak, C. J. F., & Barendregt, L. G. (1986). Weighted
averaging of species indicator values: Its efficiency in
environmental calibration. Mathematical Biosciences, 78,
57–72.
Yılmaz, F., & Kulkoyluoglu, O. (2006). Tolerance, optimum
ranges, and ecological requirements of freshwater Ostraco-
da (Crustacea) in Lake Aladag (Bolu, Turkey). Ecological
Research, 21, 165–173.




















