
In Vivo Analysis of Cell Proliferation and Apoptosis in the CNS

DYNAMIC INDUCTION OF THE LONG PENTRAXIN PTX3 IN THE CNSAFTER LIMBIC SEIZURES: EVIDENCE FOR A PROTECTIVE ROLE IN
SEIZURE-INDUCED NEURODEGENERATION
T. RAVIZZA,a D. MONETA,a B. BOTTAZZI,b G. PERI,b C. GARLANDA,b E. HIRSCH,c
G. J. RICHARDS,d A. MANTOVANIb;e and A. VEZZANIa*aLaboratory of Experimental Neurology, Department of Neuroscience, Mario Negri Institute for Pharmacological Research,
Via Eritrea 62, 20157 Milan, ItalybDepartment of Immunology and Cell Biology, Mario Negri Institute for Pharmacological Research, Via Eritrea 62,
20157 Milan, ItalycDepartment of Genetic, Biology and Biochemistry, University of Torino, Turin, ItalydPreclinical CNS Research, F. Ho¡mann-La Roche Ltd., CH-4070 Basel, Switzerland
eIstituto di Patologia Generale, University of Milano, Milan, Italy
AbstractöPentraxin 3, a prototypic long pentraxin, is induced by proin£ammatory signals in the brain. In£ammatorycytokines are rapidly induced in glia by epileptic activity. We show that pentraxin 3 immunoreactivity and mRNA areenhanced in the rat forebrain above undetectable control levels by limbic seizures with a dual pattern of induction. Within6 h from seizure onset, pentraxin 3 immunoreactivity was increased in astrocytes. Eighteen to 48 h later, speci¢c neuronalpopulations and leucocytes were strongly immunoreactive only in areas of neurodegeneration. This staining was abol-ished when neuronal cell loss, but not seizures, was prevented by blocking N-methyl-D-aspartate receptors. Pentraxin 33/3 mice had a more widespread seizure-related neuronal damage in the forebrain than their wild-type littermatesalthough both groups had similar epileptic activity.
Our results provide evidence that pentraxin 3 is synthesized in brain after seizures and may exert a protective role inseizure-induced neurodegeneration. ß 2001 IBRO. Published by Elsevier Science Ltd. All rights reserved.
Key words: acute phase proteins, cytokines, epilepsy, kainic acid, limbic system, pentraxins.
Pentraxins are a superfamily of proteins highly conservedin evolution (Goodman et al., 1996; Mantovani, 2000).The classic short pentraxins C-reactive protein (CRP)and serum amyloid P (SAP) component are mainly pro-duced in the liver in response to in£ammatory cytokines,in particular as a direct response to interleukin 6 (IL-6)stimulation (Baumann and Gauldie, 1994; Steel andWhitehead, 1994).
Long pentraxins are structurally related to, yet distinctfrom, classical short pentraxins. They are made of aC-terminal pentraxin domain coupled to diverse, unre-lated N-terminal portions (Goodman et al., 1996).They di¡er from short pentraxins in terms of gene orga-
nization, chromosomal localization, source of synthesisand function.
The ¢rst long pentraxin identi¢ed, pentraxin 3 (PTX3),was originally cloned as a gene highly induced by IL-1 inendothelial cells (Breviario et al., 1992; Introna et al.,1996) and other cell types like macrophages (VidalAlles et al., 1994) as well as, most recently, in thebrain (Polentarutti et al., 2000), and by tumor necrosisfactor-K (TNF-K) in ¢broblasts (Lee et al., 1993). It canbe secreted in a monomeric form but, when in solution, itassembles in 10^20 multimers (Bottazzi et al., 1997). Itspromoter contains a nuclear factor UB-sensitive elementthat seems essential for responses to IL-1 and TNF-K(Goodman et al., 1996; Basile et al., 1997).
Other long pentraxins were subsequently identi¢ed.These include guinea-pig apexin (Reid and Blobel,1994; Noland et al., 1994), rat neuronal pentraxin(NP1) (Schlimgen et al., 1995), human neuronal pen-traxin (NP2) (Hsu and Perin, 1995) and neuronal activityregulated pentraxin (NARP) (Tsui et al., 1996). Thesemembers of the long pentraxin family are constitutivelyexpressed in the CNS and have been proposed to have aneurobiological role. In particular, NP1 and NP2 may beinvolved in the uptake of synaptic material during syn-apse remodeling and NARP has been shown to promoteneurite outgrowth (Hsu and Perin, 1995; Schlimgen et
43
*Corresponding author. Tel. : +39-02-39014410; fax: +39-02-3546277.E-mail address: [email protected] (A. Vezzani).Abbreviations: CGP-40116, D-2-amino-4-methyl-5-phosphono-3-
pentenoic acid; EEG, electroencephalographic; GFAP, glial¢brillary acidic protein; (h)PTX3, (human) pentraxin 3; IL, inter-leukin; NARP, neuronal activity regulated pentraxin; NeuN,neuronal speci¢c nuclear protein; NP, neuronal pentraxin;PBS, phosphate-bu¡ered saline; SAP, serum amyloid P; SSC,saline sodium citrate; TRITC, tetramethyl-rhodamine isothiocya-nate.
NSC 5022 16-7-01
www.elsevier.com/locate/neuroscience
Neuroscience Vol. 105, No. 1, pp. 43^53, 2001ß 2001 IBRO. Published by Elsevier Science Ltd
Printed in Great Britain. All rights reservedPII: S 0 3 0 6 - 4 5 2 2 ( 0 1 ) 0 0 1 7 7 - 4 0306-4522 / 01 $20.00+0.00

al., 1995; Tsui et al., 1996; Kirkpatrick et al., 2000). Arole for NP2 in mediating the synaptic clustering ofK-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid(AMPA) glutamate receptors at a subset of excitatorysynapses has also been suggested (O'Brien et al., 1999).
We recently found that, di¡erently from neuronal longpentraxins, PTX3 is not expressed in the CNS but it ishighly inducible there in response to in£ammatory sig-nals (Polentarutti et al., 2000).
Limbic seizures have been shown to enhance the pro-duction of in£ammatory cytokines to a great extent inglia and these changes have functional consequences ontissue excitability (Vezzani et al., 1999; De Simoni et al.,2000; Vezzani et al., 2000).
In this study, we investigated whether PTX3 is in-duced in the rat CNS following epileptic activity, andin which cell populations, using immunocytochemistryand mRNA analysis. We also explored the functionalrole of this protein in seizures and the associated neuro-degenerative events using (3/3) mice lacking the PTX3gene.
These results provide the ¢rst unequivocal evidence,based on gene targeting, for a protective role of a longpentraxin in seizure-associated nerve cell loss.
EXPERIMENTAL PROCEDURES
Experimental animals
Male Sprague^Dawley rats (250^280 g) were purchased fromCharles River (Calco, Italy) and were housed at a constanttemperature (23³C) and relative humidity (60%) with a ¢xed12 h light^dark cycle and had free access to food and water.
PTX3 3/3 mice
PTX3-de¢cient mice were produced by homologous recombi-nation integrating an IRES-LacZ cassette followed by the PGK-neomycin resistance gene in exon 1, 71 bp downstream of the¢rst encoding ATG (C. Garlanda, E. Hirsch, B. Bottazzi, R.Nota, F. Riva, G. Peri, A. Vecchi and A. Mantovani, unpub-lished observations). Phenotypic analysis was performed on a129Sv-C57BL/6 mixed genetic background. In our experiments,we used 3/3 male adult mice (25 g) and their wild-type litter-mates as control group.
Procedures involving animals and their care were conductedin conformity with the institutional guidelines that are in com-pliance with national (4D.L. N.116, G.U., suppl. 40, 18-2-1992)and international laws and policies (EEC Council Directive 86/609, OJ L 358, 1, 12 December 1987; NIH Guide for the Careand Use of Laboratory Animals, US National Research Council1996). All e¡orts were made to minimize the number of animalsused and their su¡ering.
Injection of convulsants and evaluation of seizures
Kainic acid (Sigma-Aldrich, St. Louis, MO, USA) was dis-solved in saline and injected s.c. in rats at a dose of 12 mg/kg.Controls were rats injected with saline (1 ml/kg). Only the ratsshowing repeated episodes of severe generalized limbic seizureswith rearing and/or loss of postural control during 3 h afterconvulsant injection (about 80% of the rats) were used in thisstudy. This condition was required to select animals showingneuronal cell loss in various forebrain regions and experiencingsubsequent spontaneous and recurrent seizures, thus represent-ing a valuable model of human temporal lobe epilepsy (Sperk,1994).
For assessing seizure susceptibility and neurodegeneration inPTX3 3/3 mice and their wild-type littermates, we used anintrahippocampal injection of 0,14 nmol kainic acid. This treat-ment was chosen to induce electroencephalographic (EEG) seiz-ures in 100% of the mice with 920% of mortality.
Mice were surgically implanted under deep equithesin anes-thesia (1% phenobarbital/4% chloral hydrate, 3 ml/kg, i.p.,Sigma) with unilateral cannula and bilateral hippocampalelectrodes under stereotaxic guidance as previously described(Vezzani et al., 2000). The coordinates from bregma for theimplantation of the electrodes were (mm): antero-posterior,31.9; lateral, þ 1.5, horizontal, 1.5 below dura. Cannula andelectrodes were connected to a multipin socket and secured tothe skull with acrylic dental cement. Animals were allowed 3^5days to recover from surgical procedures before the start of theexperiment.
EEG seizures consisted of high-frequency and/or multispikecomplexes and/or high-voltage synchronized spikes simulta-neously occurring in both hippocampi. They were quanti¢ed byreckoning the latency to the ¢rst seizure (onset) and the timespent in ictal activity and spiking (see Vezzani et al., 1999, 2000).
These mice were killed by transcardial perfusion (see below)48 h after the end of the experiment to assess seizure-inducedneurodegeneration.
Treatment with D-2-amino-4-methyl-5-phosphono-3-pentenoic acid(CGP-40116)
Rats were injected i.p. with 20 mg/kg CGP-40116, a selectivecompetitive antagonist of N-methyl-D-aspartate (NMDA) recep-tors, 3 h after seizure onset. This protocol prevents seizure-asso-ciated neuronal cell loss while allowing epileptic activity toendure for at least 3 h (Fujikawa et al, 1994).
Northern blot analysis
Rats were injected with kainic acid (n = 6, each experimentalgroup) and killed at various times after seizure induction (3 h to1 week). Controls were rats injected with saline and killed atcorresponding times (n = 4^6). Animals were killed by decapita-tion, their hippocampi were dissected out at 4³C and frozen at370³C until assay. Total mRNA was extracted according to theacid guanidino^phenol^chloroform method (Chomczynski andSacchi, 1987). Ten Wg of total mRNA was analyzed by electro-phoresis through 1% agarose/formaldehyde gels followed bynorthern blot transfer to Gene Screen Plus membranes (NEN,USA). Human full length cDNA for PTX3 was labelled by the
Table 1. Number of mice showing neuronal damage
Hippocampus Thalamus Amygdala Cortex Hypothalamus
Wild-type 12/14 4/14 1/14 0/14 0/143/3 14/14 8/14 3/14 4/14* 3/14
Fractions represent the number of mice showing degeneration in the various brain regions on the total number of mice. Two mice in the wild-type group had bilateral damage in the hippocampus while ¢ve mice showed bilateral degeneration in the PTX3 3/3 group.
*P6 0.05 by Fisher's test.
NSC 5022 16-7-01
T. Ravizza et al.44
Megaprime DNA labelling system (Amersham, UK) with[K-32P]dCTP (3000 Ci/mmol speci¢c activity).
The autoradiograms were scanned and quanti¢ed using acomputerized image analysis system (IBAS, Kontron-Zeiss) ac-cording to the procedure described by Masseroli et al. (1993).The signal associated with the presence of L-actin mRNA wasused as internal standard to normalize the PTX3 mRNA expres-sion.
In situ hybridization
A di¡erent group of rats was injected with kainic acid orsaline (n = 5) as previously described and killed by decapitation6 h (n = 5) and 24 h (n = 5) after injection.
Brain horizontal sections (14 Wm) were cut, thaw-mounted ongelatin-coated microscope slides then ¢xed in 4% paraformalde-hyde in phosphate-bu¡ered saline (PBS) (pH 7.4) for 20 minfollowed by three 5-min washes in PBS. Matching sectionsfrom control or kainate-treated rats (n = 5) were taken at a com-parable antero-posterior level of the hippocampus.
The procedure described by Polentarutti et al. (2000) wasfollowed.
Brie£y, oligodeoxyribonucleotide probes, prepared on a DNAsynthesizer (Genosys Biothec. Inc., TX, USA) complementaryto mouse PTX3 cDNA (nucleotides 178^237) were used. Theoligomers were labelled with [35S]K-thio-dATP (1300 Ci/mmol,NEN) by reaction with terminal deoxynucleotidyltransferaseand precipitated with ethanol/sodium chloride. The slices werehybridized at 42³C for 18 h with a 35S-labelled probe (3U105c.p.m.) in 5 ml hybridization bu¡er. The hybridization bu¡erconsisted of 50% formamide, 5Usaline sodium citrate (SSC)(1USSC is 150 mM NaCl, 15 mM sodium citrate, pH 7.2),500 Wg/ml salmon sperm DNA, 250 Wg/ml yeast tRNA, 1UDen-hart's solution (0.02% Ficoll, 0.02% polyvinylpyrrolidone, and0.02% bovine serum albumin), 10% dextran sulfate, and 20 mMdithiothreitol. At the end of the incubation, the slides werebrie£y rinsed twice in 1USSC, 50% formamide. They werewashed four times in 50% formamide in 2USSC (42³C, 15min), then rinsed in 1USSC and dipped brie£y in water. Thesections were then dipped in 70% ethanol, dried and exposed toLmax ¢lms for 20 weeks. The ¢lms were developed with a KodakD19 developer. The corresponding radiolabelled sense DNAwas used for excluding non-speci¢c hybridization of the probe.
Tissue preparation for immunocytochemistry
To assess the changes in the pattern of PTX3 immunoreactiv-ity after seizures, kainic acid-treated rats (n = 5, each experimen-tal group) and their time-matched controls (n = 3) were killed at3, 6, 18, 24, 48 h and 1 week after kainate injection.
Rats were deeply anaesthetized with equithesin and perfusedvia the ascending aorta with 250 ml PBS, 0.1 M, pH 7.4 fol-lowed by 500 ml of chilled paraformaldehyde (4%) in PBS aspreviously described (Schwarzer et al., 1996). After carefullyremoving the brains from the skull, they were post-¢xed in thesame ¢xative as above for 90 min at 4³C then transferred to 20%sucrose in PBS at 4³C for 24 h for cryoprotection. The brainswere then rapidly frozen by immersion in isopentane at 370³Cfor 3 min before being sealed into vials and stored at 370³Cuntil use.
Immunocytochemistry
Serial cryostat sections (40 Wm) were cut horizontally (Paxinosand Watson, 1986) from all brains. The ¢rst and second ratsections of each series of ¢ve were collected for staining ofPTX3.
To obtain a monoclonal antibody against human PTX3(hPTX3), recombinant hPTX3 puri¢ed from CHO 3.5 trans-fected cells (Bottazzi et al., 1997) was emulsi¢ed in Freund'scomplete adjuvant and Lewis rats were injected both s.c. andi.p. at 2 week intervals. Five days after the last immunization,spleen cells were recovered and fused with the mouse myeloma
cell line SP2/0 in polyethylene glycol 1550. Hybrids were thenselected in HAT (hypoxanthine, aminopterin, thymidine)medium, and after 2 weeks in culture they were screened fortheir reactivity against recombinant hPTX3 in direct enzyme-linked immunosorbent assay, western blot and immunostaining.Several hybrids positively reacted against hPTX3, which has82% sequence identity with mouse PTX3. We used MNB4IgG2a monoclonal antibody in this study since it was able tocross-react with murine PTX3 but it did not recognize the shortpentraxins CRP and SAP (Peri et al., 2000).
Brie£y, free-£oating brain sections were rinsed for 30 min in0.4% Triton X-100 in 100 mM PBS (pH 7.4) at 4³C followed by15 min in 3% fetal calf serum diluted in 0.1% Triton X-100^PBS.The primary antibody was diluted in 0.1% Triton X-100^PBScontaining 3% fetal calf serum and slices were incubated at 4³Covernight with the biotinylated primary monoclonal antibodyagainst hPTX3 (1:40).
After three 5-min washes in PBS, immunoreactivity was testedby the avidin^biotin^peroxidase technique (Vectastain ABC kit,Vector Labs., USA). The sections were then reacted by incuba-tion with 0.4 mM 3P,3P-diaminobenzidine (DAB) (Sigma,Munich, Germany) in 50 mM Tris^HCl-bu¡ered saline (TBS),pH 7.4 and 0.01% H2O2, and the signal was ampli¢ed using
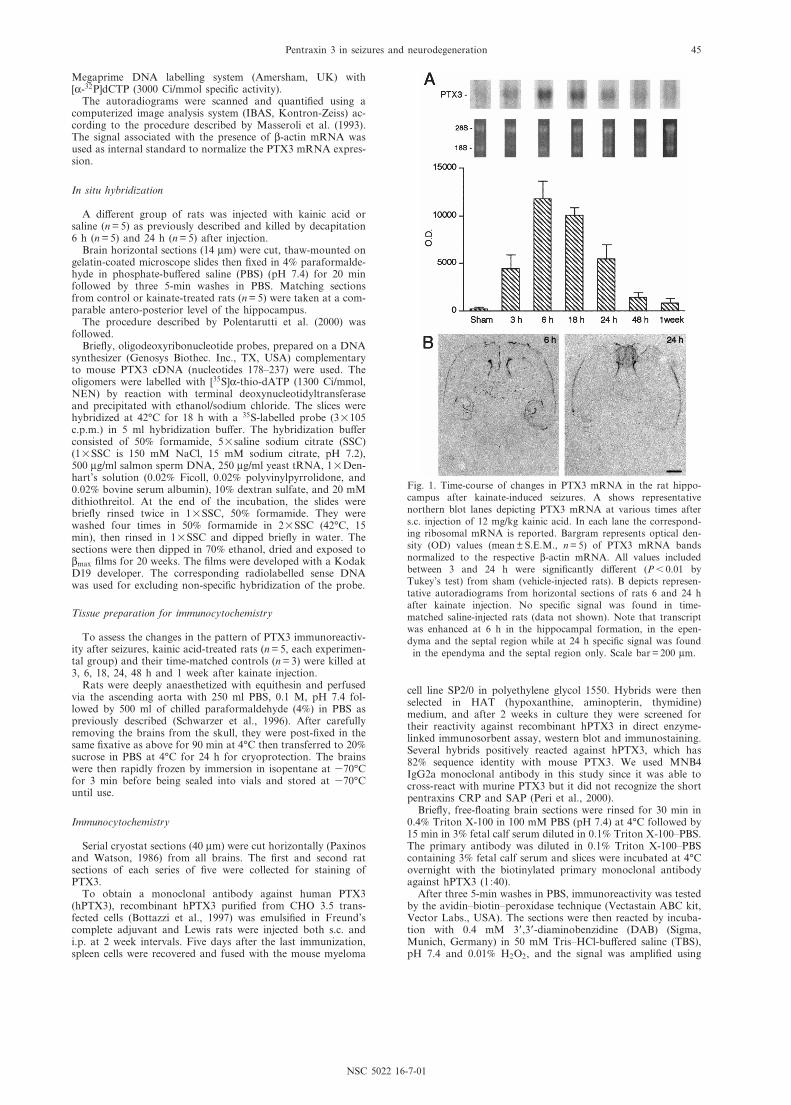
Fig. 1. Time-course of changes in PTX3 mRNA in the rat hippo-campus after kainate-induced seizures. A shows representativenorthern blot lanes depicting PTX3 mRNA at various times afters.c. injection of 12 mg/kg kainic acid. In each lane the correspond-ing ribosomal mRNA is reported. Bargram represents optical den-sity (OD) values (mean þ S.E.M., n = 5) of PTX3 mRNA bandsnormalized to the respective L-actin mRNA. All values includedbetween 3 and 24 h were signi¢cantly di¡erent (P6 0.01 byTukey's test) from sham (vehicle-injected rats). B depicts represen-tative autoradiograms from horizontal sections of rats 6 and 24 hafter kainate injection. No speci¢c signal was found in time-matched saline-injected rats (data not shown). Note that transcriptwas enhanced at 6 h in the hippocampal formation, in the epen-dyma and the septal region while at 24 h speci¢c signal was found
in the ependyma and the septal region only. Scale bar = 200 Wm.
NSC 5022 16-7-01
Pentraxin 3 in seizures and neurodegeneration 45
nickel ammonium. After DAB incubation, three 5-min washeswere done with TBS, then the slices were mounted onto gelatin-coated slides and dried overnight at room temperature. Theywere dehydrated and coverslipped the next day.
Control slices were prepared using the primary antibody pre-absorbed with the corresponding peptide (5 WM, 24 h, 4³C) andby incubating the slices without the primary antibody.
For double-immunostaining, the sections were ¢rst processedfor PTX3 immunoreactivity, as described above using a strept-avidin £uorescein isothiocyanate-conjugated secondary antibody(1:40, Pharmingen, San Diego, CA, USA). Then the slices werewashed three times for 5 min in PBS and incubated overnight at4³C with the primary antibody: rabbit anti-glial ¢brillary acidicprotein (GFAP; 1:500, Sigma-Aldrich, St. Louis, MO, USA), aselective marker of astrocytes, mouse anti-neuronal speci¢c
nuclear protein (NeuN) (1:1000, Chemicon, Temecula, CA,USA), a selective neuronal marker, or anti-rat biotinylatedCD45 (1:20, Cederlane, ON, Canada), a selective leucocytemarker. Fluorescence was then detected using, respectively, agoat anti-rabbit tetramethyl-rhodamine isothiocyanate(TRITC)-conjugated (1:40, Dako, Milan, Italy), a rabbit anti-mouse TRITC-conjugated (1:40, Dako, Milan, Italy) or astreptavidin TRITC-conjugated secondary antibody (1:40,Sigma).
Neuronal cell loss
Brain sections were taken at comparable antero-posterior andmedio-lateral levels in control and epileptic rats. Nissl stainingusing Cresyl Violet and Fluoro Jade were carried out in adjacent
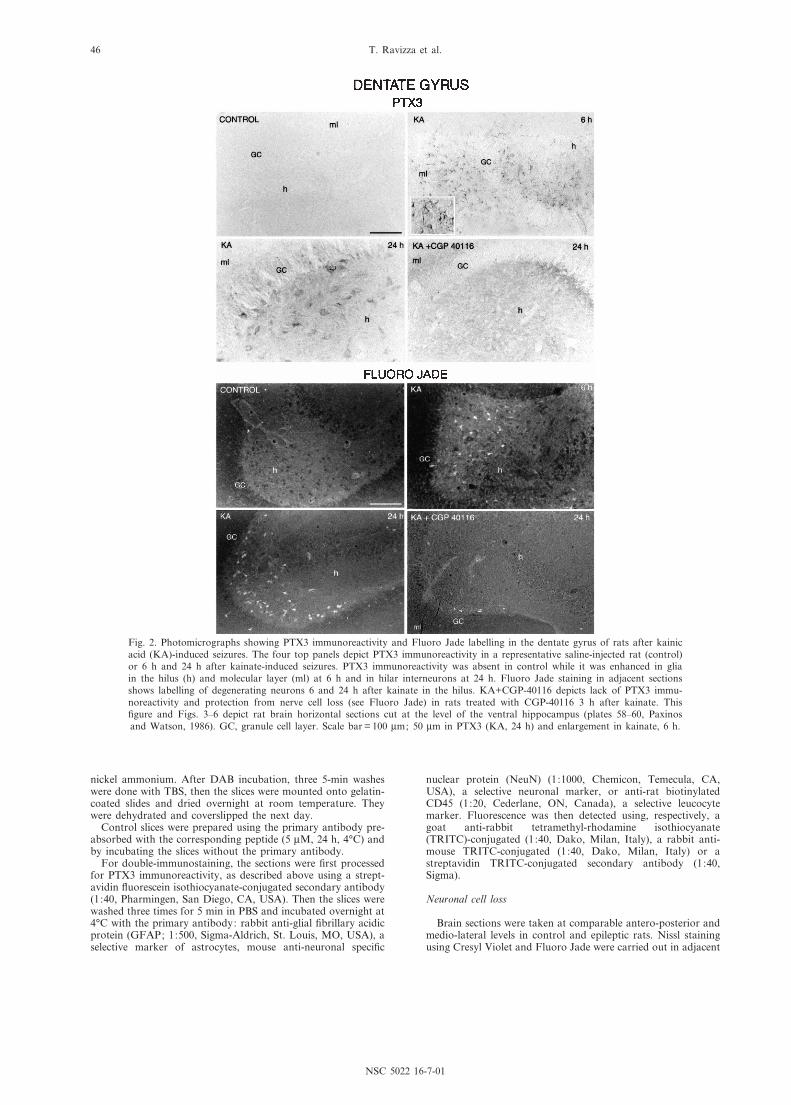
Fig. 2. Photomicrographs showing PTX3 immunoreactivity and Fluoro Jade labelling in the dentate gyrus of rats after kainicacid (KA)-induced seizures. The four top panels depict PTX3 immunoreactivity in a representative saline-injected rat (control)or 6 h and 24 h after kainate-induced seizures. PTX3 immunoreactivity was absent in control while it was enhanced in gliain the hilus (h) and molecular layer (ml) at 6 h and in hilar interneurons at 24 h. Fluoro Jade staining in adjacent sectionsshows labelling of degenerating neurons 6 and 24 h after kainate in the hilus. KA+CGP-40116 depicts lack of PTX3 immu-noreactivity and protection from nerve cell loss (see Fluoro Jade) in rats treated with CGP-40116 3 h after kainate. This¢gure and Figs. 3^6 depict rat brain horizontal sections cut at the level of the ventral hippocampus (plates 58^60, Paxinosand Watson, 1986). GC, granule cell layer. Scale bar = 100 Wm; 50 Wm in PTX3 (KA, 24 h) and enlargement in kainate, 6 h.
NSC 5022 16-7-01
T. Ravizza et al.46
sections of controls and epileptic rats to assess neurodegenera-tion and the correct position of the electrodes and the injectiontrack. For each mouse, four coronal sections were analyzed atthe level of the septal pole of the hippocampus including theinjection site (plates 281^301, Sidman et al., 1971) and four atthe level of the temporal pole (plates 335^343).
Fluoro Jade labelling was carried out as described bySchmued et al. (1997). Brie£y, sections were dried and rehy-drated in absolute ethanol (3 min), 75% ethanol (3 min), 50%ethanol (3 min) and distilled water (1 min). Then, they wereincubated for 15 min in 0.06% potassium permanganate, washedfor 1 min in distilled water and transferred to 0.001% FluoroJade staining solution. After staining, the sections were rinsedthree times for 1 min in distilled water, dried, immersed inxylene and coverslipped.
Assessment of seizure-induced neurotoxicity in PTX3 3/3 miceand their littermates control
We adopted a `severity score' for assessing the pattern ofFluoro Jade-positive neurons 48 h after kainic acid, rangingfrom 0 (absence of neurodegenerating neurons) to 5.5 (maximalneurodegeneration pattern). The score of 1 was given to miceshowing only CA3 degeneration restricted to the injected hippo-campus, a score of 2 was given when at least two regions in theinjected hippocampus showed degenerating neurons (i.e. CA3and CA1 or CA3 and hilus), a score of 3 was given whenCA1 and CA3 pyramidal cells together with hilar interneuronswere simultaneously a¡ected in the injected hippocampus. Whenthe damage (as de¢ned by scores 2 and 3) was bilateral, butrestricted to the hippocampus, the respective scores wereincreased by 0.5. When the damage (as de¢ned by 2 and 3)was either unilateral or bilateral, but also involving extrahippo-campal regions, the severity score was increased by 2 (see Fig. 7and Table 1). The score was given to each mouse by two inde-pendent investigators unaware of the identity of the experimen-tal groups.
Statistical analysis of data
Data are the means þ S.E.M. (n = number of animals). Thee¡ects of treatments were analyzed by one-way analysis of var-iance followed by Tukey's test for unconfounded means, byStudent's t-test or by Fisher's test. In all instances P6 0.05was considered statistically signi¢cant.
RESULTS
PTX3 mRNA
Northern blot analysis of PTX3 mRNA in the rathippocampus clearly showed that the transcript wasnot expressed at measurable levels in control tissue(Fig. 1A). After kainate seizures, mRNA levels rapidlyrose, reaching a peak between 6 and 18 h, then slowlyreturning to control level within 1 week. Analysis of NP1mRNA in the same samples showed that this long pen-traxin was expressed in control tissue (Schlimgen et al.,1995) but its transcript did not change after seizures(data not shown).
In situ hybridization analysis of PTX3 mRNA (Fig. 1B)typically showed that 6 h after kainate, the transcriptwas expressed in the hippocampal formation, in theependyma of the lateral ventricle and in the ventral tha-lamic nuclei, while at 24 h no speci¢c signal wasobserved except in the ependyma and the septal region(n = 5, each experimental group). No speci¢c signal wasfound in the controls (saline-injected rats) both at 6 and24 h (data not shown).
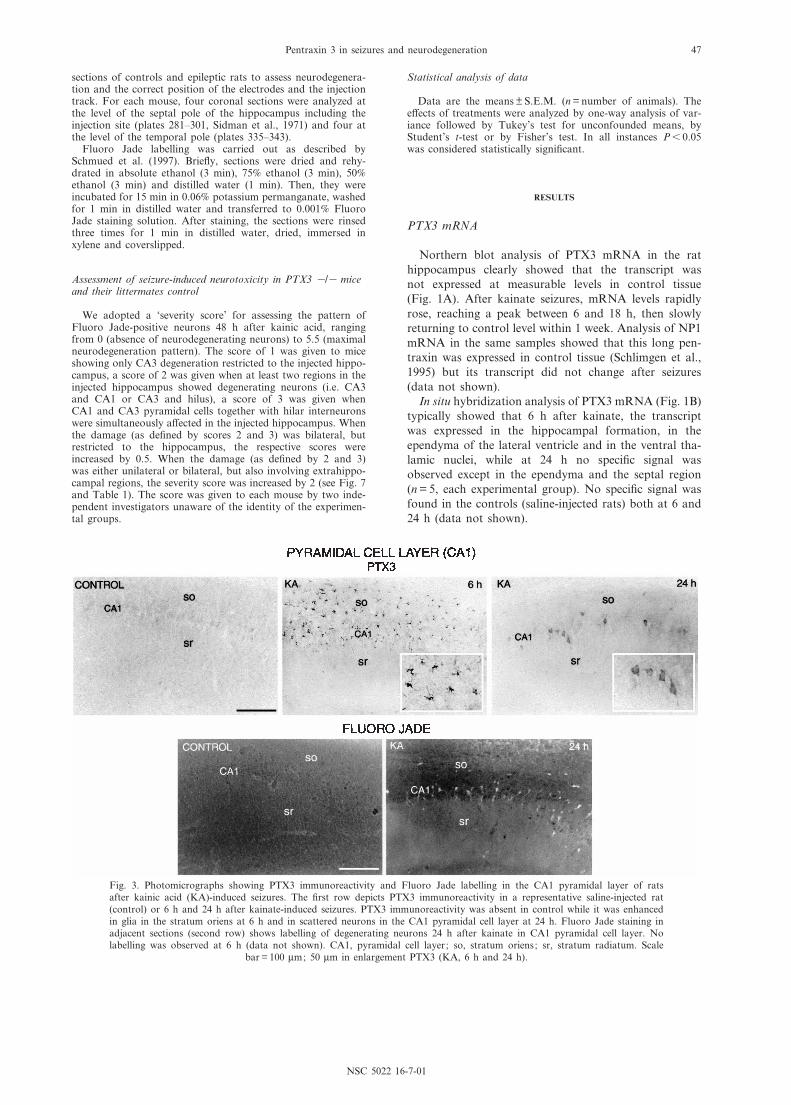
Fig. 3. Photomicrographs showing PTX3 immunoreactivity and Fluoro Jade labelling in the CA1 pyramidal layer of ratsafter kainic acid (KA)-induced seizures. The ¢rst row depicts PTX3 immunoreactivity in a representative saline-injected rat(control) or 6 h and 24 h after kainate-induced seizures. PTX3 immunoreactivity was absent in control while it was enhancedin glia in the stratum oriens at 6 h and in scattered neurons in the CA1 pyramidal cell layer at 24 h. Fluoro Jade staining inadjacent sections (second row) shows labelling of degenerating neurons 24 h after kainate in CA1 pyramidal cell layer. Nolabelling was observed at 6 h (data not shown). CA1, pyramidal cell layer; so, stratum oriens; sr, stratum radiatum. Scale
bar = 100 Wm; 50 Wm in enlargement PTX3 (KA, 6 h and 24 h).
NSC 5022 16-7-01
Pentraxin 3 in seizures and neurodegeneration 47
Immunohistochemistry
The changes in the immunocytochemical pattern ofPTX3 at various time points after kainate-induced seiz-ures were compared with the pattern of neuronal cell lossusing adjacent brain slices. Neurodegeneration wasassessed by Fluoro Jade in the dentate gyrus, hippocam-pus proper, layer III of the entorhinal cortex (Figs. 2^6),piriform cortex, periventricular thalamic nuclei and lat-eral amygdala (data not shown).
Six hours after kainate seizures, neuronal cell loss wasrestricted to the hippocampus (hilar interneurons) and/orlayer III neurons of the entorhinal cortex while temporaland piriform cortex, periventricular thalamic nuclei andamygdala were involved in the damage from 18 honwards (see below).
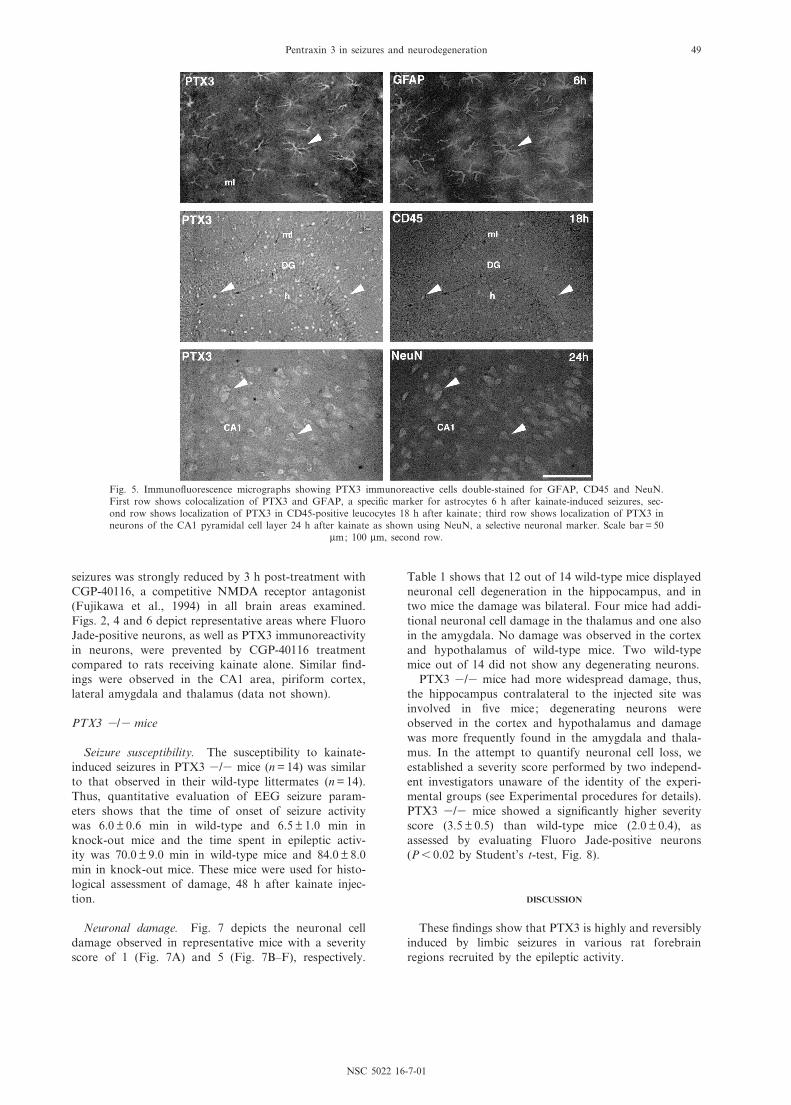
Figs. 2^4 depict PTX3 immunoreactivity 6 h and 24 hafter seizures in the hippocampal formation compared toadjacent Fluoro Jade-stained sections. In dentate gyrus(Fig. 2), CA1 (Fig. 3) and CA3 (Fig. 4), the pattern ofPTX3 immunoreactivity consistently shifted from glia(6 h) to neurons (24 h). Double-immunostaining experi-ments showed that PTX3 was colocalized with GFAP inastrocytes at 6 h (Fig. 5) and with NeuN in neurons at
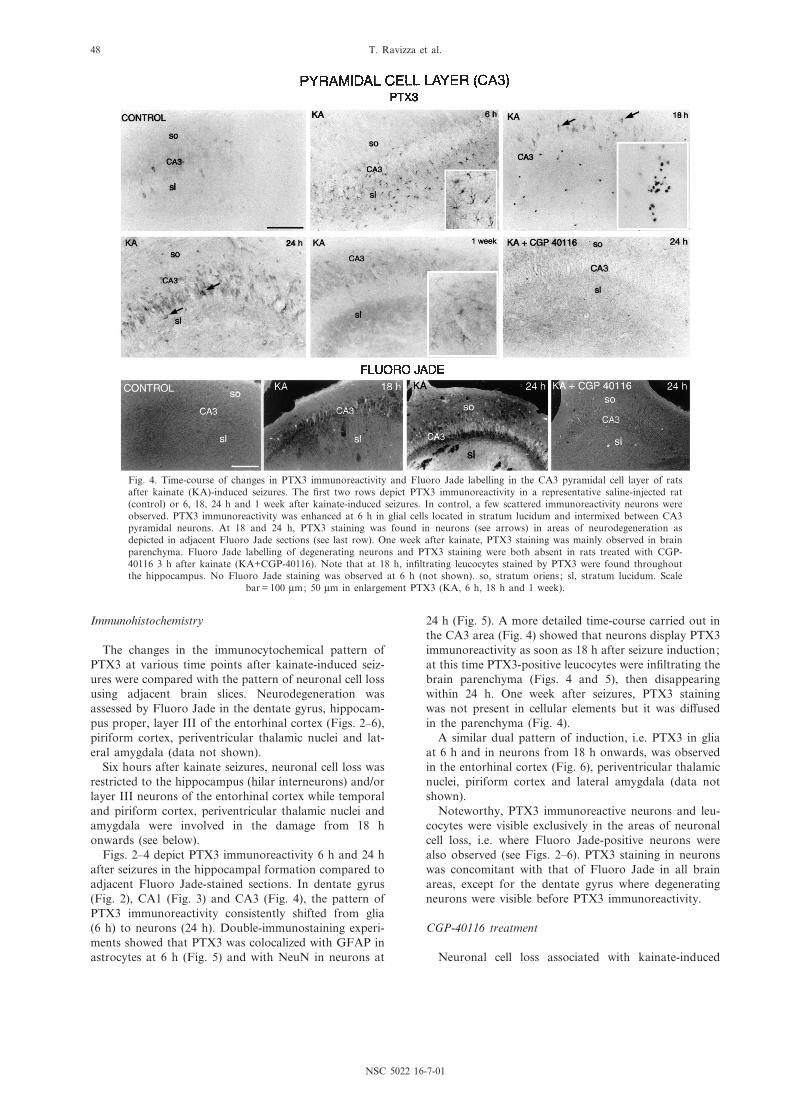
24 h (Fig. 5). A more detailed time-course carried out inthe CA3 area (Fig. 4) showed that neurons display PTX3immunoreactivity as soon as 18 h after seizure induction;at this time PTX3-positive leucocytes were in¢ltrating thebrain parenchyma (Figs. 4 and 5), then disappearingwithin 24 h. One week after seizures, PTX3 stainingwas not present in cellular elements but it was di¡usedin the parenchyma (Fig. 4).
A similar dual pattern of induction, i.e. PTX3 in gliaat 6 h and in neurons from 18 h onwards, was observedin the entorhinal cortex (Fig. 6), periventricular thalamicnuclei, piriform cortex and lateral amygdala (data notshown).
Noteworthy, PTX3 immunoreactive neurons and leu-cocytes were visible exclusively in the areas of neuronalcell loss, i.e. where Fluoro Jade-positive neurons werealso observed (see Figs. 2^6). PTX3 staining in neuronswas concomitant with that of Fluoro Jade in all brainareas, except for the dentate gyrus where degeneratingneurons were visible before PTX3 immunoreactivity.
CGP-40116 treatment
Neuronal cell loss associated with kainate-induced
Fig. 4. Time-course of changes in PTX3 immunoreactivity and Fluoro Jade labelling in the CA3 pyramidal cell layer of ratsafter kainate (KA)-induced seizures. The ¢rst two rows depict PTX3 immunoreactivity in a representative saline-injected rat(control) or 6, 18, 24 h and 1 week after kainate-induced seizures. In control, a few scattered immunoreactivity neurons wereobserved. PTX3 immunoreactivity was enhanced at 6 h in glial cells located in stratum lucidum and intermixed between CA3pyramidal neurons. At 18 and 24 h, PTX3 staining was found in neurons (see arrows) in areas of neurodegeneration asdepicted in adjacent Fluoro Jade sections (see last row). One week after kainate, PTX3 staining was mainly observed in brainparenchyma. Fluoro Jade labelling of degenerating neurons and PTX3 staining were both absent in rats treated with CGP-40116 3 h after kainate (KA+CGP-40116). Note that at 18 h, in¢ltrating leucocytes stained by PTX3 were found throughoutthe hippocampus. No Fluoro Jade staining was observed at 6 h (not shown). so, stratum oriens; sl, stratum lucidum. Scale
bar = 100 Wm; 50 Wm in enlargement PTX3 (KA, 6 h, 18 h and 1 week).
NSC 5022 16-7-01
T. Ravizza et al.48
seizures was strongly reduced by 3 h post-treatment withCGP-40116, a competitive NMDA receptor antagonist(Fujikawa et al., 1994) in all brain areas examined.Figs. 2, 4 and 6 depict representative areas where FluoroJade-positive neurons, as well as PTX3 immunoreactivityin neurons, were prevented by CGP-40116 treatmentcompared to rats receiving kainate alone. Similar ¢nd-ings were observed in the CA1 area, piriform cortex,lateral amygdala and thalamus (data not shown).
PTX3 3/3 mice
Seizure susceptibility. The susceptibility to kainate-induced seizures in PTX3 3/3 mice (n = 14) was similarto that observed in their wild-type littermates (n = 14).Thus, quantitative evaluation of EEG seizure param-eters shows that the time of onset of seizure activitywas 6.0 þ 0.6 min in wild-type and 6.5 þ 1.0 min inknock-out mice and the time spent in epileptic activ-ity was 70.0 þ 9.0 min in wild-type mice and 84.0 þ 8.0min in knock-out mice. These mice were used for histo-logical assessment of damage, 48 h after kainate injec-tion.
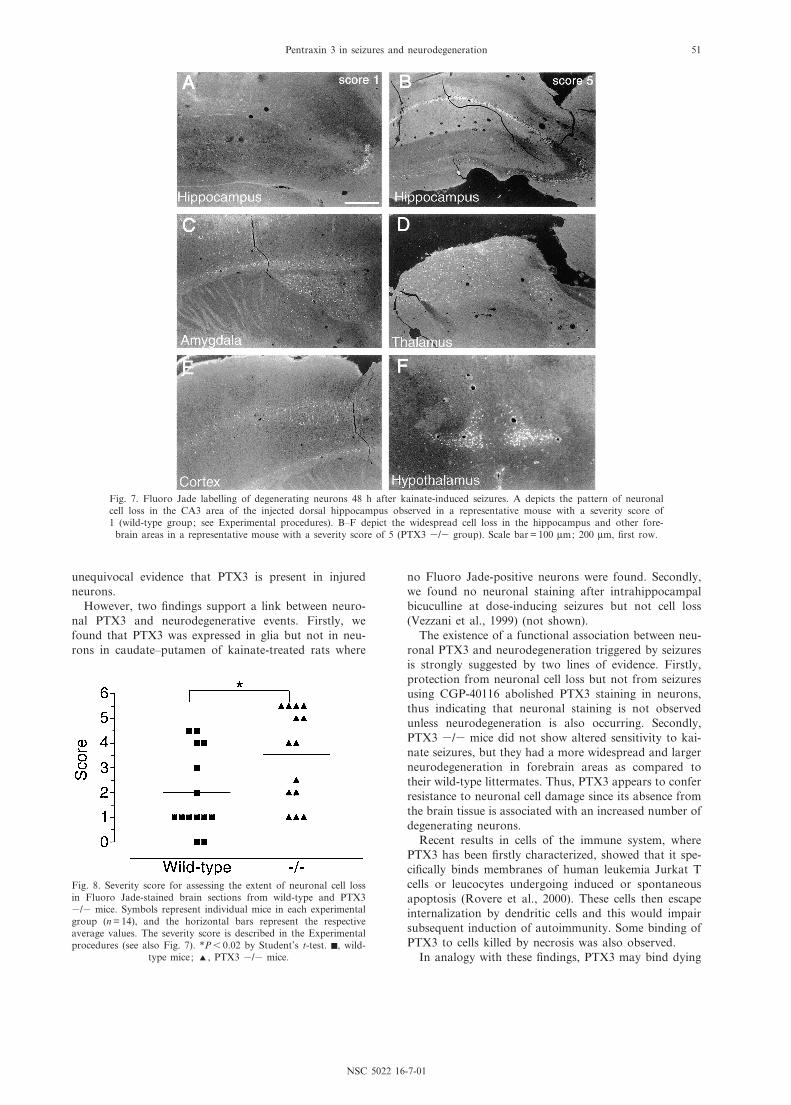
Neuronal damage. Fig. 7 depicts the neuronal celldamage observed in representative mice with a severityscore of 1 (Fig. 7A) and 5 (Fig. 7B^F), respectively.
Table 1 shows that 12 out of 14 wild-type mice displayedneuronal cell degeneration in the hippocampus, and intwo mice the damage was bilateral. Four mice had addi-tional neuronal cell damage in the thalamus and one alsoin the amygdala. No damage was observed in the cortexand hypothalamus of wild-type mice. Two wild-typemice out of 14 did not show any degenerating neurons.
PTX3 3/3 mice had more widespread damage, thus,the hippocampus contralateral to the injected site wasinvolved in ¢ve mice; degenerating neurons wereobserved in the cortex and hypothalamus and damagewas more frequently found in the amygdala and thala-mus. In the attempt to quantify neuronal cell loss, weestablished a severity score performed by two independ-ent investigators unaware of the identity of the experi-mental groups (see Experimental procedures for details).PTX3 3/3 mice showed a signi¢cantly higher severityscore (3.5 þ 0.5) than wild-type mice (2.0 þ 0.4), asassessed by evaluating Fluoro Jade-positive neurons(P6 0.02 by Student's t-test, Fig. 8).
DISCUSSION
These ¢ndings show that PTX3 is highly and reversiblyinduced by limbic seizures in various rat forebrainregions recruited by the epileptic activity.
Fig. 5. Immuno£uorescence micrographs showing PTX3 immunoreactive cells double-stained for GFAP, CD45 and NeuN.First row shows colocalization of PTX3 and GFAP, a speci¢c marker for astrocytes 6 h after kainate-induced seizures, sec-ond row shows localization of PTX3 in CD45-positive leucocytes 18 h after kainate; third row shows localization of PTX3 inneurons of the CA1 pyramidal cell layer 24 h after kainate as shown using NeuN, a selective neuronal marker. Scale bar = 50
Wm; 100 Wm, second row.
NSC 5022 16-7-01
Pentraxin 3 in seizures and neurodegeneration 49

NP1 mRNA was not modi¢ed by seizures in the sameanimals, thus indicating that only speci¢c long pentrax-ins are rapidly upregulated by neuronal activity. In thisrespect, a transient induction of NARP mRNA has alsobeen reported after electroshock-induced seizures andLTP in the hippocampus and cortex (Tsui et al., 1996).
In agreement with recent evidence (Polentarutti et al.,2000), PTX3 was not constitutively expressed in brain,except for a few immunoreactive neurons scattered in thehippocampus and entorhinal cortex. This is a major dif-ference with the other long pentraxins so far identi¢ed,which are expressed in neurons under physiological con-ditions (Hsu and Perin, 1995; Schlimgen et al., 1995;Tsui et al., 1996).
Double-immunostaining using speci¢c markers clearlyidenti¢ed three di¡erent cell types in brain tissue showingPTX3 immunostaining after seizures, namely astrocytes,leucocytes and neurons.
PTX3 was present in each cell population within aspeci¢c time frame. Thus, during the ¢rst 18 h fromseizure induction, PTX3 was observed in astrocytes andleucocytes while at later time points it was predomi-nantly present in neurons.
The presence of this protein in glia coincides with themaximal induction of its mRNA (3^18 h), while the tran-script level was lower or barely measurable when neu-rons were predominantly immunostained (24^48 h). Thissuggests that PTX3 is synthesized by glia after seizureswhile its presence in neurons may re£ect PTX3 bindingor uptake rather than enhanced synthesis. PTX3 indeedbinds discrete membrane domains in Jurkat cell linesundergoing apoptosis in a late phase of programmedcell death (Rovere et al., 2000), as well as the comple-ment component C1q (Bottazzi et al., 1997) that is con-stitutively expressed in CNS neurons (Morgan andGasque, 1996).
A major observation is that neuronal PTX3 was foundonly in CNS regions where seizure-induced neurodegen-eration occurred. Thus, neuronal cells stained by PTX3antibody were observed in brain areas and neuronal pop-ulations overlapping with those positive to Fluoro Jade,a well established marker of degenerating neurons(Schmued et al., 1997).
Since the experimental conditions required for FluoroJade staining were not compatible for double-immuno-staining with PTX3 antibody, we cannot provide
Fig. 6. Photomicrographs showing PTX3 immunoreactivity and Fluoro Jade labelling in the entorhinal cortex of rats afterkainic acid (KA)-induced seizures. The four top panels depict PTX3 immunoreactivity in a representative saline-injected rat(control) or 6 h and 24 h after kainate-induced seizures. KA+CGP-40116 depicts lack of PTX3 immunoreactivity and protec-tion from nerve cell loss (see Fluoro Jade) in rats treated with CGP-40116 3 h after kainate. PTX3 immunoreactivity waspresent in control in a few scattered neurons in layer III of the entorhinal cortex (Ent III), while it was enhanced in the samelayer in glia at 6 h and in neurons at 24 h. Fluoro Jade staining in adjacent sections (last row) shows labelling of degenerat-ing neurons 24 h after KA in layer III. No labelling was observed at 6 h (data not shown). EntM, medial entorhinal cortex.
Scale bar = 50 Wm in enlargements; 100 Wm, second and third row; 200 Wm, ¢rst row.
NSC 5022 16-7-01
T. Ravizza et al.50
unequivocal evidence that PTX3 is present in injuredneurons.
However, two ¢ndings support a link between neuro-nal PTX3 and neurodegenerative events. Firstly, wefound that PTX3 was expressed in glia but not in neu-rons in caudate^putamen of kainate-treated rats where
no Fluoro Jade-positive neurons were found. Secondly,we found no neuronal staining after intrahippocampalbicuculline at dose-inducing seizures but not cell loss(Vezzani et al., 1999) (not shown).
The existence of a functional association between neu-ronal PTX3 and neurodegeneration triggered by seizuresis strongly suggested by two lines of evidence. Firstly,protection from neuronal cell loss but not from seizuresusing CGP-40116 abolished PTX3 staining in neurons,thus indicating that neuronal staining is not observedunless neurodegeneration is also occurring. Secondly,PTX3 3/3 mice did not show altered sensitivity to kai-nate seizures, but they had a more widespread and largerneurodegeneration in forebrain areas as compared totheir wild-type littermates. Thus, PTX3 appears to conferresistance to neuronal cell damage since its absence fromthe brain tissue is associated with an increased number ofdegenerating neurons.
Recent results in cells of the immune system, wherePTX3 has been ¢rstly characterized, showed that it spe-ci¢cally binds membranes of human leukemia Jurkat Tcells or leucocytes undergoing induced or spontaneousapoptosis (Rovere et al., 2000). These cells then escapeinternalization by dendritic cells and this would impairsubsequent induction of autoimmunity. Some binding ofPTX3 to cells killed by necrosis was also observed.
In analogy with these ¢ndings, PTX3 may bind dying
Fig. 7. Fluoro Jade labelling of degenerating neurons 48 h after kainate-induced seizures. A depicts the pattern of neuronalcell loss in the CA3 area of the injected dorsal hippocampus observed in a representative mouse with a severity score of1 (wild-type group; see Experimental procedures). B^F depict the widespread cell loss in the hippocampus and other fore-
brain areas in a representative mouse with a severity score of 5 (PTX3 3/3 group). Scale bar = 100 Wm; 200 Wm, ¢rst row.
Fig. 8. Severity score for assessing the extent of neuronal cell lossin Fluoro Jade-stained brain sections from wild-type and PTX33/3 mice. Symbols represent individual mice in each experimentalgroup (n = 14), and the horizontal bars represent the respectiveaverage values. The severity score is described in the Experimentalprocedures (see also Fig. 7). *P6 0.02 by Student's t-test. F, wild-
type mice; R, PTX3 3/3 mice.
NSC 5022 16-7-01
Pentraxin 3 in seizures and neurodegeneration 51

neurons (apoptotic and/or necrotic) and rescue themfrom otherwise irreversible damage. PTX3 was observedin subsets of neurons within an apparently homogenouspopulation of cells (i.e. CA1 or CA3). It is thereforepossible that PTX3 staining occurs only in those cellsin a speci¢c time window during the degeneration pro-cess (Rovere et al., 2000). This mechanism is compatiblewith the evidence that neurodegeneration induced byseizures is a progressive and delayed process, includingboth apoptosis and necrosis (Schreiber and Baudry,1995), which can be blocked even hours after the initialinsult (Fujikawa et al., 1994).
We thus suggest that seizures induced PTX3 synthesisin glia; PTX3 is then released and subsequently bindsseizure-challenged neurons. This may occur by passivedi¡usion of PTX3 into the brain parenchyma or itsactive transport, i.e. by in¢ltrating leucocytes. The earlyglial production of PTX3 after seizures may play a rolein attracting leucocytes from the vasculature in the areasof neuronal cell loss (Akiyama et al., 1994).
Central administration of IL-1 in mice has been shownto induce PTX3 mRNA in the brain (Polentarutti et al.,2000). Since IL-1L is highly induced in glia within 2 hfrom the onset of seizures (Vezzani et al., 1999; DeSimoni et al., 2000), it is likely that this cytokine isinvolved in the early production of PTX3 in astrocytes.Six to 18 h after kainate seizures, IL-1L immunoreactiveglia was indeed found in brain areas showing PTX3staining (data not shown).
The mechanisms underlying the action of PTX3 inneurons remain to be elucidated. Since proposed mech-anisms of neurodegeneration focus generally on the trig-gering of toxic biochemical pathways by an increasedintracellular concentration of Ca2� (Lipton and Rosen-berg, 1994; Schreiber and Baudry, 1995), binding ofPTX3 to neurons may confer neuroprotection by inhib-iting Ca2� in£ux or intracellular Ca2� mobilization.
Alternatively, PTX3 may induce the production ofanti-in£ammatory molecules in the targeted cells whichin turn a¡ord protection from neurodegeneration. In thisrespect, IL-10 and IL-1 receptor antagonists have beenshown to be neuroprotective in various experimentalconditions (Rothwell, 1991).
Involvement of long pentraxins in the mechanisms ofneuronal cell loss has recently been supported by theevidence that overexpression of NP1 in cerebellar granulecells may play a permissive role in apoptotic cell deathinduced by potassium deprivation (De Gregorio et al.,2001).
An interesting aspect of the pentraxin family thatappears to be important for their cellular function isthat they bind to sugars in a calcium-dependent manner(Emsley et al., 1997). Using this lectin-like property, SAPis believed to coat molecules present in the basementmembrane and protect them from proteolytic degrada-tion (Emsley et al., 1997); NARP, similar to plant lec-tins, promotes neuronal dendritic outgrowth (Tsui et al.,1996) most likely by binding speci¢c sugar acceptor siteson the surface of neurons. Finally, evidence that gluta-mate receptor function can be selectively modi¢ed byplant lectins provides a further clue for understandinghow pentraxins might function at a molecular level. Inthis respect, recent ¢ndings have shown that NP2 isselectively enriched at some excitatory synapses in thehippocampus and it is associated with AMPA receptorclustering (O'Brien et al., 1999). Some of these mecha-nisms of action may be shared by PTX3 and mediate itse¡ects on neurons.
In summary, the dynamic expression of PTX3 in thebrain, its rapid upregulation by neuronal activity and theincreased seizure-induced neurodegeneration in PTX33/3 mice strongly suggest that this long pentraxin hasa functional role in seizure-related events. Long pentrax-ins thus represent a novel family of molecules expressedin the brain which appear to have biological functionsunder physiological and/or pathophysiological condi-tions.
AcknowledgementsöThis work was supported by MURST,projects Nuovi marcatori per la diagnostica delle patologieimmunoin¢amamtorie and Co¢nanziamento, by CNR, TargetProject Biotechnology and by FIRS. We are grateful to FeliceDe Ceglie for his dedicated and skillful contribution in ¢guresettings. We also acknowledge Dr. Marisa Aliprandi for con-tributing to part of this study and Dr. Robert Schwarcz (Uni-versity of Maryland, Baltimore, MD, USA) for his precioussuggestions during experimental planning.
REFERENCES
Akiyama, H., Tooyama, I., Kondo, H., Ikeda, K., Kimura, H., McGeer, E.G., McGeer, P.L., 1994. Early response of brain resident microglia tokainic acid-induced hippocampal lesions. Brain Res. 635, 257^268.
Basile, A., Sica, A., D'Aniello, E., Breviario, F., Garrido, G., Castellano, M., Mantovani, A., Introna, M., 1997. Characterization of the promoterfor the human long pentraxin PTX3: Role of NF-UB in tumor necrosis factor-alpha and interleukin-1beta regulation. J. Biol. Chem. 272,8172^8178.
Baumann, H., Gauldie, J., 1994. The acute phase response. Immunol. Today 15, 74^80.Bottazzi, B., Vouret-Craviari, V., Bastone, A., De Gioia, L., Matteucci, C., Peri, G., Sprea¢co, F., Pausa, M., D'Ettorre, C. et al., 1997. Multimer
formation and ligand recognition by the long pentraxin PTX3. Similarities and di¡erences with the short pentraxins C-reactive protein andserum amyloid P component. J. Biol. Chem. 272, 32817^32823.
Breviario, F., D'Aniello, E.M., Golay, J., Peri, G., Bottazzi, B., Bairoch, A., Saccone, S., Marzella, R., Predazzi, V., Rocchi, M. et al., 1992.Interleukin-1-inducible genes in endothelian cells. Cloning of a new gene related to C-reactive protein and serum amyloid P component. J. Biol.Chem. 267, 22190^22197.
Chomczynski, P., Sacchi, N., 1987. Single-step method of RNA isolation by acid guanidinium thiocyanate^phenol^chloroform extraction. Ann.Biochem. 162, 156^159.
De Gregorio, N., Gasull, T., Trullas, R., 2001. Overexpression of neuronal pentraxin 1 is involved in neuronal death evoked by low K� incerebellar granule cells. J. Biol. Chem. 276, 796^803.
NSC 5022 16-7-01
T. Ravizza et al.52

De Simoni, M.G., Perego, C., Ravizza, T., Moneta, D., Conti, M., Marchesi, F., DeLuigi, A., Garattini, S., Vezzani, A., 2000. In£ammatorycytokines and related genes are induced in the rat hippocampus by limbic status epilepticus. Eur. J. Neurosci. 12, 2623^2633.
Emsley, J., White, H.E., O'Hara, B.P., Oliva, G., Srinivasan, N., Tickle, I.J., Blundell, T.L., Pepys, M.B., Wood, S.P., 1997. Structure ofpentameric human serum amyloid P component. Nature 367, 338^345.
Fujikawa, D.G., Daniels, A.H., Kim, J.S., 1994. The competitive NMDA receptor antagonist CGP 40116 protects against status epilepticus-induced neuronal damage. Epilepsy Res. 17, 207^219.
Goodman, A.R., Cardozo, T., Abagyan, R., Altmeier, A., Wisniewski, H.-G., Vilcekt, J., 1996. Long pentraxins: an emerging group of proteinswith diverse functions. Cytokine Growth Factor Rev. 7, 191^202.
Hsu, Y.C., Perin, M.S., 1995. Human neuronal pentraxin II (NPTX2): conservation, genomic structure, and chromosomal localization. Genomics28, 220^227.
Introna, M., Vidal Alles, V., Castellano, M., Picardi, G., De Gioia, L., Bottazzi, B., Peri, G., Breviario, F., Salmona, M., De Gregorio, L.,Mantovani, A. et al., 1996. Cloning of mouse ptx3, a new member of the pentraxin gene family expressed at extrahepatic sites. Blood 87, 1862^1872.
Kirkpatrick, L.L., Matzuk, M.M., Dodds, D'N.C., Perin, M.S., 2000. Biochemical interactions of the neuronal pentraxins. J. Biol. Chem. 275,17786^17792.
Lee, G.W., Lee, T.H., Vilcek, J., 1993. TSG-14, a tumor necrosis factor- and IL-1-inducible protein, is a novel member of the pentaxin family ofacute phase proteins. J. Immunol. 150, 1804^1812.
Lipton, S.A., Rosenberg, P.A., 1994. Excitatory amino acids as a ¢nal common pathway for neurologic disorders. New Engl. J. Med. 330, 613^622.
Mantovani, A., 2001. Pentraxins. In: Encyclopedia of Molecular Medicine. J. Wiley and Sons, NY, in press.Masseroli, M., Messori, A., Bendotti, C., Ponti, M., Forloni, G., 1993. Automatic quantitative evaluation of autoradiographic band ¢lms by
computerized image analysis. Life Sci. 53, 331^336.Morgan, B.P., Gasque, P., 1996. Expression of complement in the brain: role in health and disease. Immunol. Today 17, 461^466.Noland, T.D., Friday, B.B., Puliti, M., Salepicco, S., Mosci, P., Bistoni, F., 1994. The sperm acrosomal matrix contains a novel member of the
pentraxin family of calcium-dependent binding protein. J. Biol. Chem. 269, 32607^32614.O'Brien, R.J., Xu, D., Petralia, R.S., Steward, O., Huganir, R.L., Worley, P., 1999. Synaptic clustering of AMPA receptors by the extracellular
immediate-early gene product narp. Neuron 23, 309^323.Paxinos, G., Watson, C., 1986. The Rat Stereotaxic Atlas. Academic Press, New York.Peri, G., Introna, M., Corradi, D., Iacuitti, G., Signorini, S., Avanzini, F., Pizzetti, F., Maggioni, A.P., Moccetti, T., Metra, M. et al., 2000. PTX3,
a prototypical long pentraxin, is an early indicator of acute myocardial infarction in humans. Circulation 102, 636^641.Polentarutti, N., Bottazzi, B., Di Santo, E., Blasi, E., Agnello, D., Ghezzi, P., Introna, M., Bartfai, T., Richards, G., Mantovani, A., 2000.
Inducible expression of the long pentraxin PTX3 in the central nervous system. J. Neuroimmunol. 106, 87^94.Reid, M.S., Blobel, C.P., 1994. Apexin, an acrosomal pentraxin. J. Biol. Chem. 269, 32615^32620.Rothwell, N.J., 1991. Functions and mechanisms of interleukin 1 in the brain. Trends Pharmacol. Sci. 12, 430^436.Rovere, P., Peri, G., Vallinoto, C., Bondanza, A., Bottazzi, B., Doni, A., Garlanda, C., Zimmermann, V.S., Fascio, U., Sabbadini, M.G. et al.,
2000. The long pentraxin PTX3 binds to human apoptotic cells and regulates their clearance by professional antigen presenting dendritic cells.Blood 96, 4300^4306.
Schlimgen, A.K., Helms, J.A., Vogel, H., Perin, M.S., 1995. Neuronal pentraxin, a secreted protein with homology to acute phase proteins of theimmune system. Neuron 14, 519^526.
Schmued, L.C., Albertson, C., Slikker, W., 1997. Fluoro-Jade: a novel £uorochrome for the sensitive and reliable histochemical localization ofneuronal degeneration. Brain Res. 751, 37^46.
Schreiber, S.S., Baudry, M., 1995. Selective neuronal vulnerability in the hippocampus^a role for gene expression? Trends Neurosci. 18, 446^451.Schwarzer, C., Sperk, G., Samanin, R., Rizzi, M., Gariboldi, M., Vezzani, A., 1996. Neuropeptides-immunoreactivity and their mRNA expression
in kindling: functional implications for limbic epileptogenesis. Brain Res. Rev. 22, 27^50.Sidman, R.L., Angevine, J.B., Taber Pierce, E.T., 1971. Atlas of the Mouse Brain and Spinal Cord. Harvard University Press, Cambridge.Sperk, G., 1994. Kainic acid seizures in the rat. Prog. Neurobiol. 42, 1^32.Steel, D.M., Whitehead, A.S., 1994. The major acute phase reactants: C-reactive protein, serum amyloid A protein. Immunol. Today 15, 81^88.Tsui, C.C., Copeland, N.G., Gilbert, D.J., Jenkis, N.A., Barnes, C., Worley, P.F., 1996. Narp, a novel member of the pentraxin family, promotes
neurite outgrowth and is dynamically regulated by neuronal activity. J. Neurosci. 15, 2463^2478.Vezzani, A., Conti, M., De Luigi, A., Ravizza, T., Moneta, D., Marchesi, F., De Simoni, M.G., 1999. Interleukin-1beta immunoreactivity and
microglia are enhanced in the rat hippocampus by focal kainate application: functional evidence for enhancement of electrographic seizures.J. Neurosci. 19, 5054^5065.
Vezzani, A., Moneta, D., Conti, M., Richichi, C., Ravizza, T., De Luigi, A., De Simoni, M.G., Sperk, G., Andell-Jonsson, S., Lundkvist, J. et al.,2000. Powerful anticonvulsant action of IL-1 receptor antagonist on intracerebral injection and astrocytic overexpression in mice. Proc. Natl.Acad. Sci. USA 97, 11534^11539.
Vidal Alles, V., Bottazzi, B., Peri, G., Golay, J., Introna, M., Mantovani, A., 1994. Inducible expression of PTX3, a new member of the pentraxinfamily, in human mononuclear phagocytes. Blood 84, 3483^3493.
(Accepted 12 April 2001)
NSC 5022 16-7-01
Pentraxin 3 in seizures and neurodegeneration 53
Copyright © 2022 FDOKUMEN




















