
Dunkerque City air pollution particulate matter-induced cytotoxicity, oxidative stress and...
10
Toxicology in Vitro xxx (2005) xxx–xxx www.elsevier.com/locate/toxinvit 0887-2333/$ - see front matter 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.tiv.2005.09.012 ARTICLE IN PRESS Dunkerque City air pollution particulate matter-induced cytotoxicity, oxidative stress and inXammation in human epithelial lung cells (L132) in culture Guillaume Garçon a , Zeina Dagher a , Farid Zerimech b , Frédéric Ledoux c , Dominique Courcot d , Antoine Aboukais d , Emile Puskaric c , Pirouz Shirali a,¤ a Laboratoire de Recherche en Toxicologie Industrielle et Environnementale, Maison de la Recherche en Environnement Industriel de Dunkerque 2, 189A, Avenue Maurice Schumann, 59140 Dunkerque, France b Laboratoire de Biochimie et de Biologie Moléculaire, Hôpital Huriez, 01, Place de Verdun, 59045 Lille Cedex, France c Laboratoire Interdisciplinaire en Sciences de l’Environnement (ELICO UMR 8013), Maison de la Recherche en Environnement Naturel, 32, Avenue Foch, 62930 Wimereux, France d Laboratoire de Catalyse et Environnement (EA2598), Maison de la Recherche en Environnement Industriel de Dunkerque 1, 145, Avenue Maurice Schumann, 59140 Dunkerque, France Received 23 February 2005; accepted 26 September 2005 Abstract Exposure to urban airborne particulate matter (PM) has been associated with adverse health eVects. In this work, we focused our attention on the capacity of air pollution PM to induce cytotoxic, oxidative stress, and inXammatory responses in human epithelial lung cells (L132) in culture. PM were collected in Dunkerque, a French seaside city, and their physical and chemical characteristics were car- ried out. Their size distribution showed that 92.15% of the PM were equal or smaller than 2.5 and their speciWc surface area was 1 m 2 /g. Inorganic (i.e. Fe, Al, Ca, Na, K, Mg, Pb, etc.) and organic (i.e. VOC, PAH, etc.) chemicals were found in PM. Physical and chemical prop- erties of Dunkerque City’s PM suggested that much of the collected PM derived from wind-borne dust from the industrial complex and the heavy motor vehicle traYc. Their cytotoxicity, as evaluated by survival rate determination, lactate dehydrogenase activity, and mito- chondrial dehydrogenase activity showed concentration and time-dependent eVects in L132 cells (LC10 D 18.84 g PM/ml; LC50 D 75.36 g PM/ml). Moreover, in PM-exposed L132 cells, there were concentration- and time-dependent changes in lipid peroxida- tion, superoxide dismutase activity, 8-hydroxy-2-deoxyguanosine formation, and poly(ADP-ribosyl)ation, on the one hand, and in tumor necrosis factor-alpha secretion, inducible nitric oxide synthase activity, and nitric oxide release, on the other hand. Taken together, these Wndings suggested that oxidative stress and inXammatory responses proceeded cytotoxicity in PM-exposed L132 cells. 2005 Elsevier Ltd. All rights reserved. Keywords: Human lung epithelial cells (L132); Dunkerque City particulate matter 2.5; Physical and chemical characteristics; Cytotoxicity; Oxidative stress; InXammation 1. Introduction The health eVects of air pollution have been subject to intense study in recent years (for review see Brunekreef and Holgate, 2002). Correlation between increased exacerbation of respiratory diseases, cardiopulmonary morbidity, mor- tality and the levels of urban airborne particulate matter (PM) is now well established (Levy et al., 2000; Pope and Kanner, 1993; Schwartz et al., 1993). However, the increase in health risk occurs even when PM concentrations are below established air quality standard levels (Alfaro- Moreno et al., 2002). Epidemiological studies have also * Corresponding author. Tel.: +33 3 28237610; fax: +33 3 28237171. E-mail address: [email protected] (P. Shirali).
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Dunkerque City air pollution particulate matter-induced cytotoxicity, oxidative stress and...

Toxicology in Vitro xxx (2005) xxx–xxxwww.elsevier.com/locate/toxinvit
ARTICLE IN PRESS
Dunkerque City air pollution particulate matter-inducedcytotoxicity, oxidative stress and inXammationin human epithelial lung cells (L132) in culture
Guillaume Garçon a, Zeina Dagher a, Farid Zerimech b, Frédéric Ledoux c,Dominique Courcot d, Antoine Aboukais d, Emile Puskaric c, Pirouz Shirali a,¤
a Laboratoire de Recherche en Toxicologie Industrielle et Environnementale, Maison de la Recherche en Environnement Industriel de Dunkerque 2,189A, Avenue Maurice Schumann, 59140 Dunkerque, France
b Laboratoire de Biochimie et de Biologie Moléculaire, Hôpital Huriez, 01, Place de Verdun, 59045 Lille Cedex, Francec Laboratoire Interdisciplinaire en Sciences de l’Environnement (ELICO UMR 8013), Maison de la Recherche en Environnement Naturel,
32, Avenue Foch, 62930 Wimereux, Franced Laboratoire de Catalyse et Environnement (EA2598), Maison de la Recherche en Environnement Industriel de Dunkerque 1, 145,
Avenue Maurice Schumann, 59140 Dunkerque, France
Received 23 February 2005; accepted 26 September 2005
Abstract
Exposure to urban airborne particulate matter (PM) has been associated with adverse health eVects. In this work, we focused ourattention on the capacity of air pollution PM to induce cytotoxic, oxidative stress, and inXammatory responses in human epithelial lungcells (L132) in culture. PM were collected in Dunkerque, a French seaside city, and their physical and chemical characteristics were car-ried out. Their size distribution showed that 92.15% of the PM were equal or smaller than 2.5 and their speciWc surface area was 1 m2/g.Inorganic (i.e. Fe, Al, Ca, Na, K, Mg, Pb, etc.) and organic (i.e. VOC, PAH, etc.) chemicals were found in PM. Physical and chemical prop-erties of Dunkerque City’s PM suggested that much of the collected PM derived from wind-borne dust from the industrial complex andthe heavy motor vehicle traYc. Their cytotoxicity, as evaluated by survival rate determination, lactate dehydrogenase activity, and mito-chondrial dehydrogenase activity showed concentration and time-dependent eVects in L132 cells (LC10 D 18.84 �g PM/ml;LC50 D 75.36 �g PM/ml). Moreover, in PM-exposed L132 cells, there were concentration- and time-dependent changes in lipid peroxida-tion, superoxide dismutase activity, 8-hydroxy-2�-deoxyguanosine formation, and poly(ADP-ribosyl)ation, on the one hand, and intumor necrosis factor-alpha secretion, inducible nitric oxide synthase activity, and nitric oxide release, on the other hand. Taken together,these Wndings suggested that oxidative stress and inXammatory responses proceeded cytotoxicity in PM-exposed L132 cells. 2005 Elsevier Ltd. All rights reserved.
Keywords: Human lung epithelial cells (L132); Dunkerque City particulate matter 2.5; Physical and chemical characteristics; Cytotoxicity; Oxidativestress; InXammation
1. Introduction
The health eVects of air pollution have been subject tointense study in recent years (for review see Brunekreef and
* Corresponding author. Tel.: +33 3 28237610; fax: +33 3 28237171.E-mail address: [email protected] (P. Shirali).
0887-2333/$ - see front matter 2005 Elsevier Ltd. All rights reserved.doi:10.1016/j.tiv.2005.09.012
Holgate, 2002). Correlation between increased exacerbationof respiratory diseases, cardiopulmonary morbidity, mor-tality and the levels of urban airborne particulate matter(PM) is now well established (Levy et al., 2000; Pope andKanner, 1993; Schwartz et al., 1993). However, the increasein health risk occurs even when PM concentrations arebelow established air quality standard levels (Alfaro-Moreno et al., 2002). Epidemiological studies have also

2 G. Garçon et al. / Toxicology in Vitro xxx (2005) xxx–xxx
ARTICLE IN PRESS
been carried out using current air quality standards thatonly encompass PM size and concentration; these studiesdid never address the issue of PM composition (Brunekreefand Holgate, 2002). Therefore, PM-induced underlyingadverse mechanisms responsible for the increasing mortal-ity and morbidity have not been well described yet(Brunekreef and Holgate, 2002; Osornio-Vargas et al.,2003; Schwartz et al., 1996).
Environmental and/or anthropogenic aerosols are gen-erally heterogeneous mixtures that greatly vary in constitu-ent, particle size and chemical composition (Alfaro-Moreno et al., 2002; Harrison and Yin, 2000; Spurny,1998). Now, it is clearly evident that speciWc components ofthe air particulate alone, or as components of mixtures withother pollutants, are able to produce adverse biologicalresponses consistent with human morbidity Wndings (Levyet al., 2000; Schwartz et al., 1993, 1996; Spurny, 1998). Epi-demiological Wndings from several cities around the worldindicate that the greatest health risks correlate with smallerparticles (i.e. PM2.5 and PM0.1), which have the capacity toreach the distal regions of the lung after inhalation (Levyet al., 2000). Therefore, attention has turned to the compo-nents that are most likely to have toxic potential, and theWne (PM2.5) and the ultraWne (PM0.1) particles have beenidentiWed as potential mediators of some of the toxicity ofPM10 (Diociaiuti et al., 2001; Monn and Becker, 1999;Osornio-Vargas et al., 2003). Several studies have notablyhypothesized that particles of smaller size (i.e. PM2.5) aremore potent than larger (i.e. PM10) to induce cytotoxic and/or inXammatory responses in various lung models (Osor-nio-Vargas et al., 2003; Shukla et al., 2000; Spurny, 1998).
Experimental and/or epidemiological Wndings from sev-eral authors indicate that lung diseases are often associatedwith prooxidant/antioxidant imbalance and inXammatoryreaction, and there is increasing evidence that particulateair pollutants induce acute responses as well as exacerbateexisting inXammatory diseases in the lung (Becker et al.,2002; Ghio et al., 2002; Rahman and MacNee, 2000; Osor-nio-Vargas et al., 2003; Spurny, 1998). The generation ofoxidants by PM results not only in oxidative damage incells, but also in the activation of cell signaling pathwaysleading thereby to the activation of speciWc transcriptionfactors (e.g. nuclear factor kappa-B, NF-�B) and to thesubsequent secretion of proinXammatory mediators (e.g.,tumor necrosis factor-alpha, TNF-�; nitric oxide, NO)(Pozzi et al., 2003; Schins et al., 2002; Shukla et al., 2000).
In the present work, PM were collected in Dunkerque, aFrench seaside city located on the southern coast of theNorth-Sea, characterized by the proximity of industrialactivities (i.e. iron and steel industry, aluminum industry,oil reWnery, basic chemistry, pharmaceutical industry, planthealth production, food industry, etc.) and heavy motorvehicle traYc. Firstly, physical and chemical characteristicsfrom PM samples (i.e. size distribution, speciWc surfacearea, and inorganic and organic element composition) werecarried out. Secondly, the survival rates of L132 cellsexposed to collected PM were studied using the colony
forming method, and the cytotoxicity of the urban ambientair particles was evaluated through the determination oflactate dehydrogenase (LDH) activity in cell-free culturesupernatants, and mitochondrial dehydrogenase (MDH)activity in cultured cells. The L132 cells derive from normalhuman embryonic lung tissue, present epithelial morpho-logical characteristics, and exhibit typical features of pneu-mocytes (ErXe and Mellert, 1996). The use of L132 cells relyon their lung origin and their sensitivity to the eVects of PM(Shirali et al., 1994; Garçon et al., 2000). Thirdly, to contrib-ute to a better knowledge of the underlying adverse mecha-nisms involved in PM-induced cytotoxicity, we investigatedboth the oxidative stress and the inXammatory responses inPM-exposed L132 cells in culture, through the determina-tion of malondialdehyde (MDA) production, superoxidedismutase (SOD) activity, 8-hydroxy-2�-deoxyguanosine(8-OHdG) formation, poly(ADP-ribose) polymerase(PARP) activity, TNF-� secretion, NO release, and induc-ible nitric oxide synthase (iNOS) activity.
2. Materials and methods
2.1. Chemicals
Minimum essential medium (MEM) with Earle’s salts,fetal bovine serum (FBS), L-glutamin, and penicillin/strep-tomycin solution were provided by In Vitrogen-Life Tech-nologies (Cergy Pontoise, France). Cytotoxicity DetectionKit LDH and Cell Proliferation Reagent WST-1 were fromRoche Diagnostics (Neuilly-sur-Seine, France). RANSODreagent kit was purchased from Randox Laboratories(Montpellier-Frejorgues, France). Qiagen (Courtaboeuf,France) supplied the DNeasy Tissue Kit. Sigma-Aldrich(Saint-Quentin Fallavier, France) provided the nuclease P1and the alkaline phosphatase. Japan Institute for the Con-trol of Aging (Haruoka, Japan) supplied the 8-OHdGELISA kit. The PARP Inhibition Assay, the QuantikineHuman TNF-� Immunoassays, the Quantikine HumaniNOS Immunoassays, and the Total Nitric Oxide Assaywere from R&D Systems Europe (Abingdon, United King-dom). BCA protein reagent was from Pierce (IL, USA).
2.2. PM sampling, and PM physical and chemical characteristics
PM were collected in Dunkerque (51°04�N; 2°38�E), aFrench sea-side City located on the southern coast of theNorth-Sea, using high volume Sierra model 235 cascadeimpactor (Sierra Anderson, Smyna, GA, USA) (Garçonet al., 2002). Plates were mounted without any Wlters and noback up Wlter was used to maintain a constant aspirationXow rate (68 m3/h) for a period of 2 weeks. No back up Wlterwas used and the lowest stage was doubled to increase theeYciency of smallest particle (PM0.33) sampling. Collectionwas done continuously during 9 months, from January toSeptember, and impacting system was changed every 15days; two identical systems were used alternatively. Meteo-

G. Garçon et al. / Toxicology in Vitro xxx (2005) xxx–xxx 3
ARTICLE IN PRESS
rological data (i.e. wind speed, wind direction, temperature)were obtained from Meteo France. The collection urbanzone is characterized by the proximity of industrial activi-ties (i.e. iron and steel industry, aluminum industry, oilreWnery, basic chemistry, pharmaceutical industry, planthealth production, food industry, etc.), and a heavy motorvehicle traYc. After sampling, impacting plates were driedunder laminar Xow bench during 48 h, then PM wereremoved from the collection plates and stocked at 4 °C.About 2 g of PM have been collected by this way and care-fully homogenized.
PM size distribution: Scanning electron microscopy cou-pled with energy dispersive X-ray analysis (SEM–EDX;LeoTM 438 VP microscope and IXRF analysis system;Zeiss SAS, Le Pecq, France) was used to perform singleparticle analysis and also to determine PM size distribution.Prior to the analysis, PM were suspended in a n-hexanesolution using ultrasonic treatment, and then Wlteredthrough a nucleopore Wlter to obtain well-distributed anddispersed PM, without agglomerates, as suitable for theSEM–EDX analysis. After the Wlter carbon-coating, thedetermination of PM size has been done in the automaticmode with SEM operating conditions as follows: accelerat-ing voltage—25 keV; intensity—300 nA, and magniWca-tion—£5000. The minimal size of PM that can be observedwas 0.1 �m.
SpeciWc surface area: Adsorption data from the Bru-nauer Emmett Teller (BET) method using a Sorptomaticapparatus (Thermo Electron Corporation, Courtaboeuf,France) provided the speciWc surface area of PM. PM sam-ples were Wrst outgassed under vacuum at room tempera-ture, and the volume of pure nitrogen (N60) gas adsorbedto their surface, as measured at the liquid nitrogen tempera-ture (¡196 °C), was correlated to the total speciWc surfacearea of PM, including pores in the surface.
Inorganic element composition: The analytical procedureconsisted of the sample acidic total digestion of PM by aHNO3/HClO4/HF (4/1/1 v/v/v) mixture during 3 h at130 °C, the acid evaporation during 4 h at 170 °C, and thesubsequent analysis by inductively coupled plasma-atomicemission spectrometry (ICP-AES) using a Perkin–ElmerAAnalyst 600 (Perkin–Elmer Life Sciences France, Cour-taboeuf, France).
Organic compound composition: After a soxhlet extrac-tion with dichloromethane, organic compounds weredetected in PM using gas chromatography–mass spectrom-etry (GC–MS; Brüker EM40, Bruker, Wissembourg,France; Varian Saturn, Varian France, Coutaboeuf,France).
2.3. Cells and culture conditions
The cell line we used originates from the normal lung tis-sue of a human embryo, and is deposited under the designa-tion L132 in the American type culture collection (ATCC;ATCC number: CCL-5). The morphology of these cells isepithelial, and they exhibit typical features of pneumocytes
(ErXe and Mellert, 1996). L132 cells were cultured in sterileplastic Xasks (Corning; Fisher ScientiWc Labosi SAS, Elan-court, France), in MEM with Earle’s salts, containing: 5%(v/v) FBS, 1% (v/v) L-glutamin (200 mM), 1% (v/v) penicil-lin (10 000 IU/ml) and 1% (v/v) streptomycin (10 000 UG/ml) (In Vitrogen-Life Technologies). Exponentially grow-ing cells were maintained at 37 °C, in a humidiWed atmo-sphere containing 5% CO2. All the L132 cells we used in thestudy of the cytotoxicity, on the one hand, and in the studyof the oxidative stress and the inXammatory response, onthe other hand, derived from the same initial cell culture.
2.4. Sampling of culture supernatants and cells for the study of the cytotoxicity
Colony forming method: Survival rates of L132 cellsexposed to PM were determined using the colony formingmethod (Shirali et al., 1995). BrieXy, 100 proliferating cellswere seeded in culture dishes (60 £ 15 mm; Corning; FisherScientiWc Labosi SAS) to determine cell viability after 10days of incubation at 37 °C, in a humidiWed atmospherecontaining 5% CO2, in the continuous presence of PM with-out renewing the culture media. Accordingly, 10 culturedishes were chosen at random as control cells (i.e. incubatedonly with cultured media) and Wve culture dishes per PMconcentration were designated at random as exposed cells(i.e. incubated with culture media containing increasingconcentrations of PM: 18.84, 37.68, 56.52, 75.36 or150.72�g PM/ml). Both the LC10 and LC50 values werecalculated comparing the number of crystal violet-stainedcell colonies in exposed culture dishes with the number innon-exposed culture dishes.
Measurement of released LDH activity: Released LDHactivities were evaluated in cell-free culture supernatants(Boland et al., 2001). L132 cells were seeded in 96-well cul-ture plates (Costar; Fisher ScientiWc Labosi SAS) at a den-sity of 1 £ 104 cells/200 �l culture media containing 18.84,37.68, 56.52 or 75.36�g PM/ml and incubated at 37 °C in ahumidiWed atmosphere containing 5% CO2. Non-exposedcells were used as negative controls. Accordingly, 16 repli-cates were chosen at random as control cells (i.e. incubatedonly with cultured media) and eight replicates per PM con-centration were designated at random as exposed cells (i.e.incubated with culture media containing 18.84, 37.68, 56.52or 75.36�g PM/ml). After 24, 48 or 72 h of incubation, 100-�l-aliquots of culture supernatants were collected cell-freeand transferred in new 96-well culture plates to allow themeasurement of extracellular LDH activity. The colorimet-ric assay for the quantiWcation of LDH activity releasedfrom the cytosol of damaged cells was performed accordingto the manufacturer’s recommendations (CytotoxicityDetection Kit LDH, Roche).
Measurement of MDH activity: MDH activities weredetermined in cultured cells (Takenouchi and Munekata,1995). We used the experimental design previouslydescribed for the evaluation of released LDH. Non-exposedcells were used as negative controls. After 24, 48 or 72 h of

4 G. Garçon et al. / Toxicology in Vitro xxx (2005) xxx–xxx
ARTICLE IN PRESS
incubation, the colorimetric assay for the quantiWcation ofcell proliferation and/or cell viability, based on the cleavageof the tetrazolium salt WST-1 by MDH in viable cells, wasrealized (Cell Proliferation Reagent WST-1, Roche).
2.5. Sampling of culture supernatants and cells for the study of oxidative stress and inXammatory response
Cell exposure: Depending on the incubation time, L132cells were seeded at diVerent density (i.e. 3 £ 106, 1.5 £ 106
or 0.75 £ 106 cells/20 ml of culture media, respectively) inculture plastic Xasks (Corning; Fisher ScientiWc LabosiSAS) and incubated at 37 °C, in a humidiWed atmospherecontaining 5% CO2, for 24 h. Culture supernatants wereremoved to eliminate non-adherent cells. Only living L132cells were also incubated in the continuous presence of PMat their LC10 (i.e. 18.84 �g PM/ml) or LC50 (i.e.75.36 �g PM/ml) for 24, 48 or 72 h of incubation, withoutrenewing the culture media. Both the LC10 and the LC50values were previously calculated owing to the colonyforming method. Non-exposed cells were used as negativecontrols. Accordingly, for each incubation time, 10 cultureXasks were chosen at random as control cells (i.e. incubatedonly with cultured media) and Wve culture Xasks per PMconcentration were designated at random as exposed cells(i.e. incubated with culture media containing 18.84 or75.36 �g PM/ml).
Sampling of culture supernatants and cells: After 24, 48 or72 h of incubation, 1-ml-aliquots of culture supernatantswere collected and immediately frozen at ¡80 °C for thedetermination of TNF-�, NO, or total protein contents.Adherent cells were removed and centrifuged (500g, 10 min,4 °C). Cell pellets were washed twice with 10-ml-aliquots ofsterile PBS (In Vitrogen-Life Technologies) and cell-ali-quots were collected and quickly frozen at ¡80 °C until thefurther determination of MDA concentration, SOD activ-ity, 8-OHdG concentration, PARP activity, iNOS activity,and total protein content.
Study of oxidative stress: Lipid peroxidation was evalu-ated in cell lysates using a high performance liquid chroma-tography MDA assay (Garçon et al., 2000). Total SODactivities were assessed in cell lysates using the RANSODreagent kit (Randox Laboratories) (Garçon et al., 2001a).8-OHdG concentrations were determined using the methodoriginally described by Toyokuni et al. (1997) and modiWedby Garçon et al. (2001b). BrieXy, after DNA extraction(DNeasyTissue Kit, Qiagen) and digestion (nuclease P1and alkaline phosphatase, Sigma-Aldrich), the formation ofthe oxidative DNA adduct 8-OHdG was measured usingthe 8-OHdG ELISA kit (Japan Institute for the Control ofAging). PARP activities were studied in cell lysates usingthe PARP Inhibition Assay (R&D Systems Europe)according to the manufacturer’s instructions.
Study of inXammatory response: TNF-� concentrations incollected cell culture supernatants were determined using acommercially available enzyme immunoassay (QuantikineHuman TNF-� Immunoassay, R&D Systems Europe).
iNOS activities in cell lysates were evaluated in cell lysatesusing the Quantikine Human iNOS Immunoassay (R&DSystems Europe), according to the manufacturer’s recom-mendations. Total NO concentrations in collected cell cul-ture supernatants were studied using the Griess reactionafter the enzymatic conversion of nitrate to nitrite by nitratereductase (total nitric oxide assay, R&D Systems Europe).
Total protein contents in both the cell lysates and thecollected cell culture supernatants were determined withbicinchoninic reagent using the BCA protein reagent(Pierce) (Smith et al., 1985).
2.6. Statistical analysis
Results were expressed as mean values and ranges (mini-mum value–maximum value). Data from cell culturesexposed to increasing concentrations of PM were comparedwith those from non-exposed cell cultures. Statistical analy-ses were performed by the Mann–Whitney U test (Soft-ware: SPSS for Windows, v10.05, 2000; Paris, France).Statistically signiWcant diVerences were reported with p val-ues <0.05.
3. Results
3.1. PM physical and chemical characteristics
Fig. 1 shows the size distribution results of collected PMas depicted as cumulative frequencies (%) and relative fre-quencies (%). Accordingly, 64.24%, 92.15%, and 98.49% ofPM total number were equal or smaller than 1 �m, 2.5 �m,and 5 �m, respectively. The highest number of collected PMwere also detected in size classes included in PM with ageometric diameter inferior or equal to 2.5 �m: 0–0.5 �m(33.63%), 0.5–1.0 �m (30.61%), 1.0–1.5 �m (14.33%), 1.5–2.0 �m (8.69%), and 2.0–2.5�m (4.89%). Their speciWc sur-face area was 1 m2/g. Table 1 shows the inorganic elementsfound in collected PM. Among them, Fe, Al, Ca, Na, K,
Fig. 1. The size distribution of the particle matter (PM) collected in Dun-kerque, a French seaside city located on the southern coast of the North-Sea, using high volume cascade impactor, is depicted as cumulative fre-quency (%; curve) and relative frequency (%; histogram). PM size distri-bution has been determined using scanning electron microscopy.
0
10
20
30
40
50
60
70
80
90
100
PM geometric diameter (µm)
Cum
ulat
ive
and
rela
tive
freq
uenc
ies
(%)
0 1 2 3 4 5 6 7 8 9 10

G. Garçon et al. / Toxicology in Vitro xxx (2005) xxx–xxx 5
ARTICLE IN PRESS
Mg, Pb, and Ti were the most abundantly present. In col-lected PM, there were not only inorganic elements usuallyassociated with natural environment (i.e. Ca, Na, Mg, Ti,Sr, etc.), but also so-called anthropogenic elements (i.e. Fe,Al, Pb, Mn, Zn, Ba, Cu, Cr, Ni, Cd, etc.). Table 2 shows thelist of the organic compounds detected in PM samples. Inparticular, various volatile organic compounds (VOC) andpolycyclic aromatic hydrocarbons (PAH) were coated ontocollected PM. The concentrations of the PAH-coated ontocollected PM are presented in the Table 2.
3.2. Cytotoxicity of PM
PM induced a concentration-dependent cytotoxic eVectin proliferating L132 cells, as determined using the colony
Table 1Inorganic compounds detected in PM
Inorganic compounds Concentrations (% w/w)
Fe 7.84Al 5.83Ca 4.95Na 1.88K 0.97Mg 0.81Pb 0.80Ti 0.51Mn 0.352Zn 0.184Ba 0.064Cu 0.037Cr 0.032V 0.030Ni 0.029Sr 0.022Li 0.011Mo 0.004Cd <0.001Co <0.0006
Table 2Organic compounds detected in PM
The concentrations of the polycyclic aromatic hydrocarbons (�g/g) arespeciWed.
Cycloheptene Benzo(b+k)Xuoranthene (6.55 �g/g)Benzene Benzo(a)pyrene (7.90 �g/g)Dimethylbenzene Phenanthrene (9.94 �g/g)Ethenylethylbenzene Pyrene (4.03 �g/g)Ethylmethylbenzene Benzene dicarboxylique acidStyrene DibutylphtalateToluene Dihydrobenzopyranone by-productsTriethylbenzene DiphenyletherDimethylbinaphtalene DodecanolDimethylnaphtalene Hexadeanoic acidEthylnaphtalene NonanolIndane Tetradecanoic acidMethyllindane BenzonitrileMethylnaphtalene ChloronitrobenzeneTrimethylnaphtalene MethylpentanamideNaphtalene MethylpyridinChrysene (9.54 �g/g) ParaYns (>C16)Fluoranthene (1.45 �g/g)
forming method (Fig. 2). The LC10 and LC50 values were18.84 and 75.36�g PM/ml, respectively. Moreover, PM gen-erated concentration and time-dependent increases ofreleased LDH activity (Fig. 3). Statistically signiWcantincreases of extracellular LDH activities were observed inL132 cells 72 h after their exposure to 56.52 or 75.36�g PM/ml (p < 0.001, respectively). The control values for releasedLDH did not diVer depending on the incubation time (24–72 h), indicating the background of cell death. At least, PM
Fig. 2. Survival rates (%) of L132 cells 10 days after their incubation in thecontinuous presence of increasing concentrations of collected particlematter (PM; 18.84, 37.68, 56.52, 75.36 or 150.72 �g PM/ml) without renew-ing the culture media. Non-exposed cells were used as negative controls.These values are depicted as mean values and ranges (minimum value–maximum value) of 10 cell cultures for negative controls and Wve cell cul-tures for every PM concentrations. Lethal concentrations at 10% and 50%(LC10 and LC50, respectively) were calculated comparing the number ofcrystal violet-stained cell colonies in exposed culture dishes with the num-ber in non-exposed culture dishes.
y = -0.6345 + 100r² = 0.9907
PM concentration (µg/ml)
Via
bilit
y(%
)0
25
50
75
100
0 25 50 75 100 125 150 175
125
LC10(18.84 µg/ml)
LC50(75.36 µg/ml)
Fig. 3. Released lactate dehydrogenase (LDH) activity (U of absorbency)in cell-free culture supernatants of L132 cells 24, 48 or 72 h after theirincubation in the continuous presence of increasing concentrations of par-ticle matter (PM; 18.84, 37.68, 56.52 or 75.36 �g PM/ml) without renewingthe culture media. Non-exposed cells were used as negative controls.These values are depicted as mean values and ranges (minimum value–maximum value) of 16 replicates for negative controls and eight replicatesfor every PM concentrations (a D not signiWcant and d D p < 0.001; Wilco-xon rank-sum test).
6 G. Garçon et al. / Toxicology in Vitro xxx (2005) xxx–xxx
ARTICLE IN PRESS
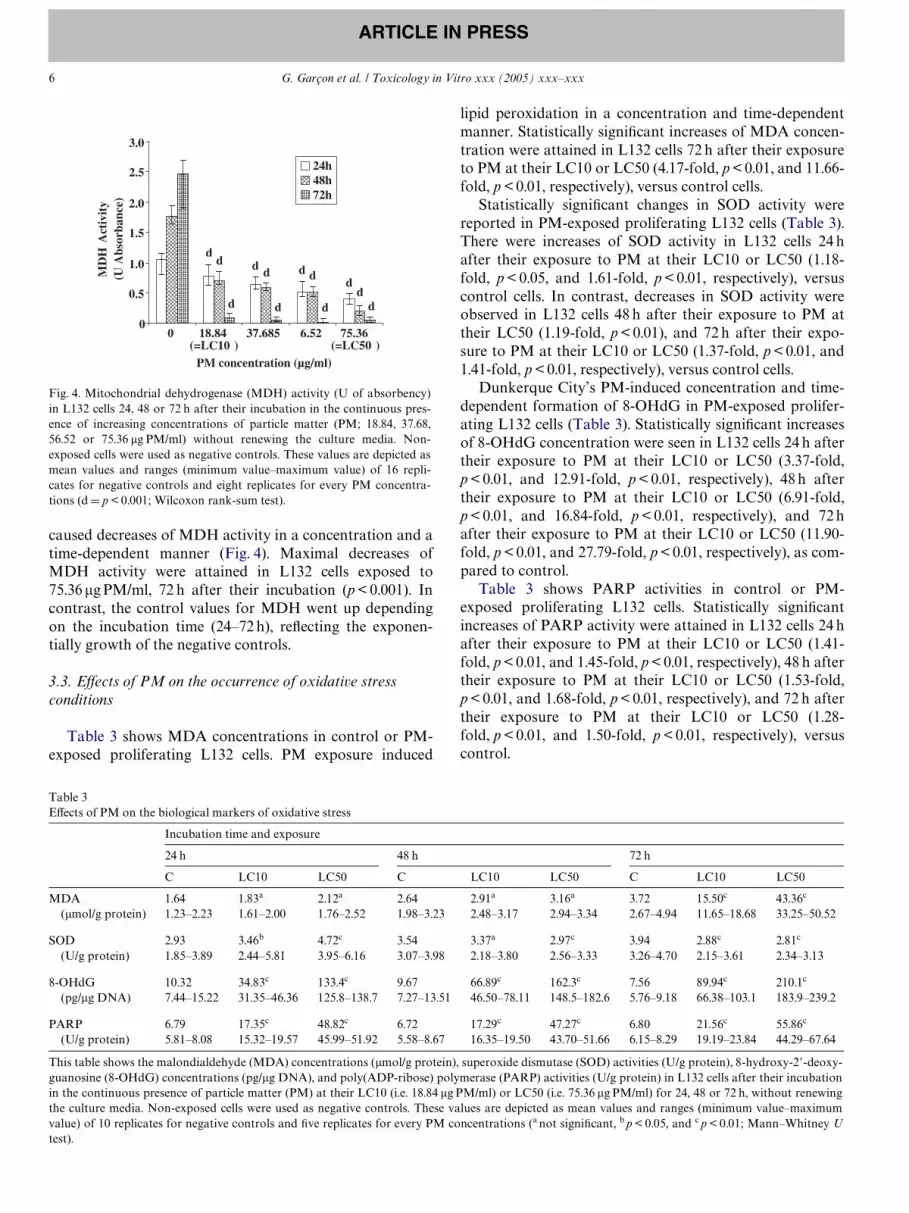
caused decreases of MDH activity in a concentration and atime-dependent manner (Fig. 4). Maximal decreases ofMDH activity were attained in L132 cells exposed to75.36 �g PM/ml, 72 h after their incubation (p < 0.001). Incontrast, the control values for MDH went up dependingon the incubation time (24–72 h), reXecting the exponen-tially growth of the negative controls.
3.3. EVects of PM on the occurrence of oxidative stress conditions
Table 3 shows MDA concentrations in control or PM-exposed proliferating L132 cells. PM exposure induced
Fig. 4. Mitochondrial dehydrogenase (MDH) activity (U of absorbency)in L132 cells 24, 48 or 72 h after their incubation in the continuous pres-ence of increasing concentrations of particle matter (PM; 18.84, 37.68,56.52 or 75.36 �g PM/ml) without renewing the culture media. Non-exposed cells were used as negative controls. These values are depicted asmean values and ranges (minimum value–maximum value) of 16 repli-cates for negative controls and eight replicates for every PM concentra-tions (d D p < 0.001; Wilcoxon rank-sum test).
lipid peroxidation in a concentration and time-dependentmanner. Statistically signiWcant increases of MDA concen-tration were attained in L132 cells 72 h after their exposureto PM at their LC10 or LC50 (4.17-fold, p < 0.01, and 11.66-fold, p < 0.01, respectively), versus control cells.
Statistically signiWcant changes in SOD activity werereported in PM-exposed proliferating L132 cells (Table 3).There were increases of SOD activity in L132 cells 24 hafter their exposure to PM at their LC10 or LC50 (1.18-fold, p < 0.05, and 1.61-fold, p < 0.01, respectively), versuscontrol cells. In contrast, decreases in SOD activity wereobserved in L132 cells 48 h after their exposure to PM attheir LC50 (1.19-fold, p < 0.01), and 72 h after their expo-sure to PM at their LC10 or LC50 (1.37-fold, p < 0.01, and1.41-fold, p < 0.01, respectively), versus control cells.
Dunkerque City’s PM-induced concentration and time-dependent formation of 8-OHdG in PM-exposed prolifer-ating L132 cells (Table 3). Statistically signiWcant increasesof 8-OHdG concentration were seen in L132 cells 24 h aftertheir exposure to PM at their LC10 or LC50 (3.37-fold,p < 0.01, and 12.91-fold, p < 0.01, respectively), 48 h aftertheir exposure to PM at their LC10 or LC50 (6.91-fold,p < 0.01, and 16.84-fold, p < 0.01, respectively), and 72 hafter their exposure to PM at their LC10 or LC50 (11.90-fold, p < 0.01, and 27.79-fold, p < 0.01, respectively), as com-pared to control.
Table 3 shows PARP activities in control or PM-exposed proliferating L132 cells. Statistically signiWcantincreases of PARP activity were attained in L132 cells 24 hafter their exposure to PM at their LC10 or LC50 (1.41-fold, p < 0.01, and 1.45-fold, p < 0.01, respectively), 48 h aftertheir exposure to PM at their LC10 or LC50 (1.53-fold,p < 0.01, and 1.68-fold, p < 0.01, respectively), and 72 h aftertheir exposure to PM at their LC10 or LC50 (1.28-fold, p < 0.01, and 1.50-fold, p < 0.01, respectively), versuscontrol.
Table 3EVects of PM on the biological markers of oxidative stress
This table shows the malondialdehyde (MDA) concentrations (�mol/g protein), superoxide dismutase (SOD) activities (U/g protein), 8-hydroxy-2�-deoxy-guanosine (8-OHdG) concentrations (pg/�g DNA), and poly(ADP-ribose) polymerase (PARP) activities (U/g protein) in L132 cells after their incubationin the continuous presence of particle matter (PM) at their LC10 (i.e. 18.84 �g PM/ml) or LC50 (i.e. 75.36 �g PM/ml) for 24, 48 or 72 h, without renewingthe culture media. Non-exposed cells were used as negative controls. These values are depicted as mean values and ranges (minimum value–maximumvalue) of 10 replicates for negative controls and Wve replicates for every PM concentrations (a not signiWcant, b p < 0.05, and c p < 0.01; Mann–Whitney Utest).
Incubation time and exposure
24 h 48 h 72 h
C LC10 LC50 C LC10 LC50 C LC10 LC50
MDA(�mol/g protein)
1.64 1.83a 2.12a 2.64 2.91a 3.16a 3.72 15.50c 43.36c
1.23–2.23 1.61–2.00 1.76–2.52 1.98–3.23 2.48–3.17 2.94–3.34 2.67–4.94 11.65–18.68 33.25–50.52
SOD(U/g protein)
2.93 3.46b 4.72c 3.54 3.37a 2.97c 3.94 2.88c 2.81c
1.85–3.89 2.44–5.81 3.95–6.16 3.07–3.98 2.18–3.80 2.56–3.33 3.26–4.70 2.15–3.61 2.34–3.13
8-OHdG(pg/�g DNA)
10.32 34.83c 133.4c 9.67 66.89c 162.3c 7.56 89.94c 210.1c
7.44–15.22 31.35–46.36 125.8–138.7 7.27–13.51 46.50–78.11 148.5–182.6 5.76–9.18 66.38–103.1 183.9–239.2
PARP(U/g protein)
6.79 17.35c 48.82c 6.72 17.29c 47.27c 6.80 21.56c 55.86c
5.81–8.08 15.32–19.57 45.99–51.92 5.58–8.67 16.35–19.50 43.70–51.66 6.15–8.29 19.19–23.84 44.29–67.64

G. Garçon et al. / Toxicology in Vitro xxx (2005) xxx–xxx 7
ARTICLE IN PRESS
3.4. EVects of PM on the development of the inXammatory response
Dunkerque City’s PM-induced TNF-� secretion byL132 cells in a concentration- and time-dependent manner(Table 4). SigniWcant increases of TNF-� concentrationwere determined in the culture supernatants of L132 cells24 h after their exposure to PM at their LC50 (6.05-fold,p < 0.01), 48 h after their exposure to PM at their LC10(2.16-fold, p < 0.01) and LC50 (10.03-fold, p < 0.01), and72 h after their exposure to PM at their LC10 (8.72-fold,p < 0.01) and LC50 (19.10-fold, p < 0.01), as compared tocontrol.
Table 4 shows iNOS activities in control or PM-exposedproliferating L132 cells. There were statistically signiWcantincreases of iNOS activity in L132 cells 24 h after theirexposure to PM at their LC10 or LC50 (2.37-fold, p < 0.01,and 2.81-fold, p < 0.01, respectively), 48 h after their expo-sure to PM at their LC10 or LC50 (2.55-fold, p < 0.01, and3.30-fold, p < 0.01, respectively), and 72 h after their expo-sure to PM at their LC10 or LC50 (3.60-fold, p < 0.01, and8.88-fold, p < 0.01, respectively), versus control.
The releases of NO in the culture supernatants of controlor PM-exposed proliferating L132 cells are shown in Table4. Statistically signiWcant increases of NO concentrationwere seen in L132 cells 24 h after their exposure to PM attheir LC10 or LC50 (2.55-fold, p < 0.01, and 7.18-fold,p < 0.01, respectively), 48 h after their exposure to PM attheir LC10 or LC50 (2.57-fold, p < 0.01, and 7.04-fold,p < 0.01, respectively), and 72 h after their exposure to PMat their LC10 or LC50 (3.53-fold, p < 0.01, and 8.21-fold,p < 0.01, respectively), as compared to control.
4. Discussion
In the present work, we focused our attention on thecytotoxicity, the oxidative stress, and the inXammatoryresponses induced by Dunkerque City air pollution PM inL132 cells.
Epidemiological studies have been carried out using cur-rent air quality standards that only encompass particle sizeand concentration; these studies did not address the issue ofparticle composition (Osornio-Vargas et al., 2003). Hence,in this work, the main physical and chemical characteristicsof Dunkerque City’s PM were determined. Size distributionresults of collected PM showed that 92.15% of PM totalnumber were equal or smaller than 2.5�m, thereby suggest-ing that they are human respirable. According to epidemio-logical Wndings from several cities around the world, thegreatest health risks correlate with smaller PM which havethe ability to reach the distal regions of the lung after inha-lation (Levy et al., 2000). Components such as transitionmetals, VOC, and PAH are known to be associated withPM, either through adherence to the core particle or as anintegral component of the particle (Dreher et al., 1997;Monn and Becker, 1999; Nel et al., 2001). A total of 20inorganic elements were found in Dunkerque City’s PM.Among them, Fe, Al, Ca, Na, Pb, Mg, and Mn were themost abundantly present and constitute not only inorganicelements usually associated with natural environment (i.e.Ca, Mg, Na), but also so-called anthropogenic elements (i.e.Al, Mn, Pb, Zn). The speciWc surface area (i.e. 1 m2/g) of thePM under study showed the adsorption of various chemi-cals. Accordingly, VOC and PAH-coated onto PM werenotably detected. It is also interesting to report that,according to the considered PAH, PAH concentrations inDunkerque City’s PM were equal or smaller than thosemeasured in PM samples collected in two locations in Paris(Baulig et al., 2004). Taken together, the results arising fromthe physical and chemical characterization of DunkerqueCity’s PM suggest that much of the collected PM derivedfrom wind-borne dust from the industrial complex and theheavy diesel motor vehicle traYc. However, Baulig et al.(2004) reported that the physical and chemical properties ofparticles revealed to be unsatisfactory to predict their levelsof biological reactivity. Probably, the diVerent signalingpathways elicited by each particle component (inorganicelements, organic compounds, etc.) interact and/or
Table 4EVects of PM on the biological markers of inXammation
This table shows the tumor necrosis factor-alpha (TNF-�) concentrations (pg/mg protein), inducible nitric oxide synthase (iNOS) activities (U/g protein),and nitric oxide (NO) concentrations (�mol/g protein) in L132 cells and cell culture supernatants after their incubation in the continuous presence of par-ticle matter (PM) at their LC10 (i.e. 18.84 �g PM/ml) or LC50 (i.e. 75.36 �g PM/ml) for 24, 48 or 72 h, without renewing the culture media. Non-exposedcells were used as negative controls. These values are depicted as mean values and ranges (minimum value–maximum value) of 10 replicates for negativecontrols and Wve replicates for every PM concentrations (a not signiWcant and c p < 0.01; Mann–Whitney U test).
Incubation time and exposure
24 h 48 h 72 h
C LC10 LC50 C LC10 LC50 C LC10 LC50
TNF-�(pg/mg protein)
2.55 2.49a 15.47c 2.70 5.85c 27.15c 2.40 20.90c 45.80c
2.09–3.09 2.23–2.72 13.30–18.28 2.32–3.28 4.29–7.11 25.21–29.55 2.19–2.64 19.66–22.95 41.29–49.15
iNOS(U/mg protein)
10.76 25.56c 30.25c 12.54 31.99c 41.43c 14.56 52.46c 129.4c
7.95–16.53 20.04–27.75 29.60–34.36 8.17–19.79 23.58–39.80 30.56–57.46 10.15–26.07 40.84–59.41 93.00–155.8
NO(�mol/g protein)
6.79 17.35c 48.82c 6.72 17.29c 47.28c 6.80 21.57c 55.86c
5.81–8.08 15.32–19.57 46.00–51.92 5.58–8.67 16.35–19.50 43.71–51.66 6.15–8.29 19.20–23.84 44.29–67.64

8 G. Garçon et al. / Toxicology in Vitro xxx (2005) xxx–xxx
ARTICLE IN PRESS
converge to give a similar cellular response. Hence, to betterevaluate the adverse health eVects of PM exposure, it willbe of great interest to consider not only the physical andchemical characteristics of PM but also to determine theirtoxicological reactivity in target cells.
We showed that Dunkerque City’s PM lead to concen-tration-dependent cytotoxicity in proliferating L132 cells,as determined using the colony forming method. PM-induced concentration-dependent and time-dependentincreases of released LDH activity. Statistically signiWcantincreases of extracellular LDH activity were observed inL132 cells 72 h after their exposure to the highest PM con-centrations, thereby suggesting late alterations of mem-brane integrity and/or permeability. In contrast, PMseemed to early aVect the MDH activity, which is a measureof both the cell toxicity and the cell proliferation, since theformazan conversion depends on the amount as well as themitochondrial metabolic competence of the cells. Indeed,PM caused decreases of MDH activity in a concentrationand a time-dependent manner, and maximal decrease ofMDH activity was attained in L132 cells exposed to LC50,72 h after their incubation. Hence, these results partly corre-late to the observed concentration and time-dependentgrowth alteration assessed through the survival rate analy-sis. Taken together, these results showed that the PM understudy induced a global cytotoxicity in the considered lungin vitro system.
Thereafter, we investigated the capacity of DunkerqueCity’s PM to induce oxidative stress conditions and inXam-matory response in this lung cell target. Firstly, we foundthat in vitro exposure to these PM induce lipid peroxida-tion in a concentration and a time-dependent manner. Thisresult is consistent with the data about LDH activity sug-gesting the occurrence of signiWcant alteration of mem-brane Xuidity or integrity only 72 h after cell exposure toPM. SigniWcant changes in SOD activity (i.e. initiallyincreases then decreases) were reported in PM-exposedL132 cells, versus control cells. Accordingly, it has alreadybeen described that the reactive oxygen intermediates(ROI) generated in oxidative stress conditions could alterproteins and thereby damage some antioxidant enzymes(De Zwart et al., 1999; Gilks et al., 1998; Mates and San-chez-Jimenez, 1999). The literature also indicated that SODis not particularly protected against ROI and that this pro-cess could contribute to its inactivation. Since SOD is prin-cipally a protective enzyme, its depletion or lowered activitycan lead to adverse eVects through the accumulation ofROI in cells and oxidative damages in biological macro-molecules (Garçon et al., 2001a,b; Escobar et al., 1996).Among them, DNA is oxidized to a number of damagedproducts in vitro and one of these products, 8-OHdG, hasbeen suggested for a marker of oxidative damage in DNAin vitro and in vivo (Tokiwa et al., 1999). We showed thatPM induced a concentration and time-dependent forma-tion of 8-OHdG in L132 cells. According to Shi et al.(2003), PM are able to generate hydroxyl radicals andtherefore to induce formation of 8-OHdG. Such an exces-
sive production of oxidative DNA adducts seemed to causea concentration and time-dependent activation of PARP inPM-exposed cells. PARP activation and poly(ADP-ribose)polymerization are also proposed to be important for theregulation of many cellular processes such as DNA repair,cell death, chromatin functions, and genomic stability (Her-ceg and Wang, 2001). The activation of PARP is an imme-diate cellular reaction to DNA strand breakage as inducedby alkylating agents, ionizing radiation or oxidants (Bou-chard et al., 2003; Burkle, 2001; Nicoletti and Stella, 2003).Accordingly, recent data in the literature reported thatsome of the main covalent metals (i.e. Fe, Al, Pb, Mn, Zn)detected in Dunkerque City’s PM could be Wrmly involvedin redox systems, which can lead to the initiation of radicalreactions (Becker et al., 2002; Garçon et al., 2001a,b; Ghioand Devlin, 2001; Hetland et al., 2001; Shi et al., 2003). Theassociations between metals in PM soluble fractions andlung responses have been observed in in vivo and in vitrosystems (Hetland et al., 2001). Ghio and Devlin (2001) havesuggested that the disparate eVects of PM extracts seen intheir study resulted from physical or chemical diVerencesother than metal content and catalyzed oxidants cannot beexcluded. In addition, it is now well known that the meta-bolic activation by enzyme-catalyzed reactions of the VOCand/or the PAH-coated onto PM could result in the exces-sive production of ROI capable of interfering with cellhomeostasis (Garçon et al., 2001a,b; Liden et al., 2003).
In vitro exposure to collected PM caused signiWcantincreases of TNF-� secretion and NO generation in a con-centration and time-dependent manner, suggesting the pro-gressive development of an inXammatory response. Inaddition to their oxidative eVects, ROI have been impli-cated in the regulation of NF-�B, and therefore, of TNF-�(Blackwell and Christman, 1997). This cell-derived cytokinesometimes restores normal homeostasis. At other times,however, because of its overproduction or its aberrantrelease, it appears to act as an ampliWcation factor in thecascade of acute inXammatory events (Luster et al., 1999).Stimulated cells are also known to release large quantitiesof NO as a direct consequence of iNOS activation by TNF-� (Rao, 2000). Accordingly, we showed that PM inducedincreases of iNOS activity in a concentration and time-dependent manner in PM-exposed cells. Accordingly, otherauthors have demonstrated that in vitro exposure to someof the metals (i.e. Fe, Al, Pb, Mn, Zn), VOC or PAHdetected in Dunkerque City’s PM resulted in cytokinereleases (Alfaro-Moreno et al., 2002; Becker et al., 1996;Bayram et al., 1998; Dye et al., 1999; Garçon et al., 2000,2001a,b; Liden et al., 2003; Monn and Becker, 1999; Osor-nio-Vargas et al., 2003; Pozzi et al., 2003; Shukla et al.,2000; Takizawa et al., 1999). In all these studies, the induc-tion of excessive cytokine secretion, and, therefore, thedevelopment of an inXammatory response might rely onthe capacity of such air pollutants to generate ROI, whichhave been Wrmly implicated in the stringent regulation ofNF-�B, and therefore, of the cytokine network (Winyardand Blake, 1997). It has been proposed that eVects of PM-

G. Garçon et al. / Toxicology in Vitro xxx (2005) xxx–xxx 9
ARTICLE IN PRESS
exposure should be related to relationship between theirphysical and chemical characteristics, and their potency toinduce inXammatory eVects that aggravate according totime (Hetland et al., 2001). Schaumann et al. (2004) con-clude that environmentally relevant concentrations ofPM2.5 from a smelter area induced distinct airway inXam-mation in healthy subjects with a selective inXux of mono-cytes and increased generation of oxidant radicals. Theysupported the hypothesis that the higher concentration oftransition metals in these PM2.5 might be responsible forthis increased inXammation.
In conclusion, the physical and chemical properties ofDunkerque City’s PM suggest that much of the collectedPM derived from wind-borne dust from the industrial com-plex, and the heavy diesel motor vehicle traYc. The highconcentrations of transition metals (i.e. Fe, Al, Pb, Mn, Zn)and the presence of organic compounds (i.e. VOC andPAH) found in the PM under study might be Wrmlyinvolved in the generation of oxidative stress conditions,and, therefore, in the development of an inXammatoryresponse. Taken together, the present Wndings indicate thatoxidative stress and inXammatory responses proceededcytotoxicity in Dunkerque City’s PM-exposed L132 cells.
Acknowledgements
The research described in this article has been supportedby grants from the University of Littoral-Opale Coast, theFranco-British Interreg IIIA European Program (FrenchConvention no. 24), and the Regional Council of Nord/Pas-de-Calais (Convention no. 04 06 0092).
References
Alfaro-Moreno, E., Martinez, L., García-Cuellar, C., Bonner, J.C., Mur-ray, J.C., Rosas, I., Ponce-de-Leon Rosales, S., Miranda, J., Osornio-Vargas, A.R., 2002. Biological eVects induced in vitro by PM10 fromthree diVerent zones of Mexico City. Environ. Health Perspect. 110,715–720.
Baulig, A., Poirault, J.J., Ausset, P., Schins, R., Shi, T., Baralle, D., Dorlh-ene, P., Meyer, M., Lefevre, R., Baeza-Squiban, A., Marano, F., 2004.Physicochemical characteristics and biological activities of seasonalatmospheric particulate matter sampling in two locations of Paris.Environ. Sci. Technol. 38, 5985–5992.
Bayram, H., Devalia, J.L., Sapsford, R.J., Ohtoshi, T., Miyabara, Y., Sagai,M., Davies, R.J., 1998. The eVect of diesel exhaust particles on cell func-tion and release of inXammatory mediators from human bronchial epi-thelial cells in vitro. Am. J. Respir. Cell. Mol. Biol. 18, 441–448.
Becker, S., Soukup, J.M., Gilmour, M.I., Devlin, R.B., 1996. Stimulation ofhuman and rat alveolar macrophages by urban air particulates: eVecton oxidant radical generation and cytokine production. Toxicol. Appl.Pharmacol. 141, 637–648.
Becker, S., Soukup, J.M., Gallagher, J.E., 2002. DiVerential particulate airpollution induced oxidant stress in human granulocytes monocytesand alveolar macrophages. Toxicol. in Vitro 16, 209–218.
Blackwell, T.S., Christman, J.W., 1997. The role of nuclear factor-�B incytokine gene regulation. Am. J. Respir. Cell. Mol. Biol. 17, 3–9.
Boland, S., Baeza-Squiban, A., Bonvallot, V., Houcine, O., Pain, C., Meyer,M., Marano, F., 2001. Similar eVects induced by diesel exhaust particlesfrom a representative diesel vehicle recovered from Wlters and standardreference material 1650. Toxicol. in Vitro 15, 379–385.
Bouchard, V., Rouleau, M., Poirier, G., 2003. PARP-1, a determinant ofcell in response to DNA damage. Exp. Hematol. 31, 446–454.
Brunekreef, B., Holgate, S., 2002. Air pollution and health. Lancet 360,1233–1242.
Burkle, A., 2001. Poly(ADP-ribosyl)ation, a DNA damage-driven proteinmodiWcation and regulator of genomic instability. Cancer Lett. 163, 1–5.
De Zwart, L.L., Meerman, J.H.N., Commandeur, J.N.M., Vermeulen, N.P.,1999. Biomarkers of free radical damage applications in experimentalanimals and in humans. Free Radical. Biol. Med. 26, 202–226.
Diociaiuti, M., Balduzzi, M., De Berardis, B., Cattani, G., Stacchini, G.,Ziemacki, G., Marconi, A., Paoletti, L., 2001. The two PM(2.5) andPM(2.5–1.0) coarse fractions: evidence of diVerent biological activity.Environ. Res. 86, 254–262.
Dreher, K.L., Jaskot, R.H., Lehmann, J.R., Richards, J.H., McGee, J.K.,Ghio, A.J., Costa, D.L., 1997. Soluble transition metals mediate resid-ual oil Xy ash induced acute lung injury. J. Toxicol. Environ. Health 50,285–305.
Dye, J.A., Adler, K.B., Richards, J.H., Dreher, K.L., 1999. Role of solublemetals in oil Xy ash-induced airway epithelial injury and cytokine geneexpression. Am. J. Physiol. 277, L498–L510.
ErXe, V., Mellert, W., 1996. Human cell line LC5 and its use US Patent5582967 dated December 10, 1996.
Escobar, J.A., Rubio, M.A., Lissi, E.A., 1996. SOD and catalase inactiva-tion by singlet oxygen and peroxyl radicals. Free Radical. Biol. Med.20, 285–290.
Garçon, G., Shirali, P., Garry, S., Fontaine, M., Zerimech, F., Martin, A.,Hannothiaux, M.H., 2000. Polycyclic aromatic hydrocarbons-coatedonto Fe2O3 particles: assessment of cellular membrane damage andantioxidant system disruption in human epithelial lung cells (L132) inculture. Toxicol. Lett. 117, 25–35.
Garçon, G., Zerimech, F., Hannothiaux, M.H., Gosset, P., Martin, A.,Marez, T., Shirali, P., 2001a. Antioxidant defense disruption by polycy-clic aromatic hydrocarbons-coated onto Fe2O3 particles in human lungcells (A549). Toxicology 166, 129–137.
Garçon, G., Garry, S., Gosset, P., Zerimech, F., Martin, A., Hannothiaux,M.H., Shirali, P., 2001b. Benzo(a)pyrene-coated onto Fe2O3 particles-induced lung tissue injury: role of free radicals. Cancer Lett. 167, 7–15.
Garçon, G., Ledoux, F., Hannothiaux, M.H., Zerimech, F., Puskaric, E.,Shirali, P., 2002. Urban particulate air pollution and evaluation of itstoxicity on human pulmonary cells in culture. In: 6th InternationalAerosol Conference Taiwan (Abstract).
Ghio, A.J., Devlin, R.B., 2001. InXammatory lung injury after bronchialinstillation of air pollution particles. Am. J. Respir. Crit. Care. Med.164, 704–708.
Ghio, A.J., Silbajoris, R., Carson, J.L., Samet, J.M., 2002. Biological eVectsof oil Xy ash. Environ. Health Perspect. 110, 89–94.
Gilks, C.B., Price, K., Wright, J.L., Churg, A., 1998. Antioxidant geneexpression in rat lung after exposure to cigarette smoke. Am. J. Pathol.152, 269–278.
Harrison, R.M., Yin, J., 2000. Particulate matter in the atmosphere: whichparticle properties are important for its eVects on health? Sci. TotalEnviron. 249, 85–101.
Herceg, Z., Wang, Z.Q., 2001. Functions of poly(ADP-ribose)polymerase(PARP) in DNA repair, genomic integrity and cell death. Mutat. Res.477, 97–110.
Hetland, R.B., Myhre, O., Lag, M., Hongve, D., Schwarze, P.E., Refsnes,M., 2001. Importance of soluble metals and reactive oxygen species forcytokine release induced by mineral particles. Toxicology 165, 133–144.
Levy, J.I., Hammit, J.K., Spengler, J.D., 2000. Estimating the mortalityimpacts of particulate matter: what can be learned from between-studyvariability? Environ. Health Perspect. 108, 109–117.
Liden, J., Ek, A., Palmberg, L., Okret, S., Larsson, K., 2003. Organic dustactivates NF-�B in lung epithelial cells. Respir. Med. 97, 882–892.
Luster, M.I., Simeonova, P.P., Gallucci, R., Matheson, J., 1999. Tumornecrosis factor-� and toxicology. Crit. Rev. Toxicol. 29, 491–511.
Mates, J.M., Sanchez-Jimenez, F., 1999. Antioxidant enzymes and theirimplications in pathophysiologic processes. Front Biosci. 4, d339–d345.

10 G. Garçon et al. / Toxicology in Vitro xxx (2005) xxx–xxx
ARTICLE IN PRESS
Monn, C., Becker, S., 1999. Cytotoxicity and induction of proinXamma-tory cytokines from human monocytes exposed to Wne (PM2.5) andcoarse particles (PM10–2.5) in outdoor and indoor air. Toxicol. Appl.Pharmacol. 155, 245–252.
Nel, A.E., Diaz-Sanchez, D., Li, N., 2001. The role of particulate pollutantsin pulmonary inXammation and asthma: evidence for the involvementof organic chemicals and oxidative stress. Curr. Opin. Pulm. Med. 7,20–26.
Nicoletti, V.G., Stella, A.M.G., 2003. Role of PARP under stress condi-tions: cell death or protection? Neurochem. Res. 28, 187–194.
Osornio-Vargas, A.R., Bonner, J.C., Alfaro-Moreno, E., Martinez, L., Gar-cia-Cuellar, C., Ponce-de-Leon Rosales, S., Miranda, J., Rosas, I., 2003.ProinXammatory and cytotoxic eVects of Mexico city air pollution par-ticulate matter in vitro are dependent on particle size and composition.Environ. Health Perspect. 111, 1289–1293.
Pope, C.A., Kanner, R.E., 1993. Acute eVects of PM10 pollution on pulmo-nary function of smokers with mild to moderate chronic obstructivepulmonary disease. Am. Rev. Respir. Dis. 147, 1336–1340.
Pozzi, R., De Berardis, B., Paoletti, L., Guastadisegni, C., 2003. InXamma-tory mediators induced by coarse (PM2.5–1.0) and Wne (PM2.5) urbanair particles in RAW 2647 cells. Toxicology 183, 243–254.
Rahman, I., MacNee, W., 2000. Oxidative stress and regulation of glutathi-one in lung inXammation. Eur. Respir. J. 16, 534–554.
Rao, K.M., 2000. Molecular mechanisms regulating iNOS expression invarious cell types. J. Toxicol. Environ. Health Part. B Crit. Rev. 3, 27–58.
Schins, R.P., Knaapen, A.M., Weishaupt, C., Winzer, A., Borm, P.J.A.,2002. Cytotoxic and inXammatory eVects of coarse and Wne particulatematter in macrophages and epithelial cells. Ann. Occup. Hyg. 46, 203–206.
Schaumann, F., Borm, P.J.A., Herbrich, A., Knoch, J., Pitz, M., Schins,R.P.F., Luettig, B., Hohlfeld, J.M., Heinrich, J., Krug, N., 2004.Metal-rich ambient particles (particulate matter 2.5) cause airwayinXammation in healthy subjects. Am. J. Respir. Crit. Care. Med. 170,898–903.
Schwartz, J., Slater, D., Larson, T.V., Pierson, W.E., Koenig, J.Q., 1993.Particulate air pollution and hospital emergency room visits forasthma in Seattle. Am. Rev. Respir. Dis. 147, 826–831.
Schwartz, J., Dockery, D.W., Neas, L.M., 1996. Is daily mortality associ-ated specially with Wne particles? J. Air Waste Manag. Assoc. 46, 927–939.
Shi, T., Knaapen, A.M., Begerow, J., Birmili, W., Borm, P.J.A., Schins,R.P.F., 2003. Temporal variations of hydroxyl radical generation and8-hydroxy-2�-deoxyguanosine formation by coarse and Wne particulatematter. Occup. Environ. Med. 60, 315–321.
Shirali, P., Teissier, E., Marez, T., Hildebrand, H.F., Haguenoer, J.M., 1994.EVect of �Ni3S2 on arachidonic acid metabolites in cultured humanlung cells (L132 cell line). Carcinogenesis 15, 759–762.
Shirali, P., Gosset, P., Marez, T., Hachimi, A., Muller, J.F., Haguenoer,J.M., 1995. Cytotoxicity evaluation of hematite (Fe2O3) benzo(a)pyreneand pyrene and analysis of particles by Laser Microprobe Mass Ana-lyser (in-vitro). Cell Biol. Toxicol. 11, 215.
Shukla, A., Timblin, C., BeruBe, K., Gordon, T., McKinney, W., Driscoll, K.,Vacek, P., Mossman, B.T., 2000. Inhaled particulate matter causesexpression of nuclear factor (NF)-kB-related genes and oxidant-depen-dent NF-kB activation in vitro. Am. J. Respir. Cell. Mol. Biol. 23, 182–187.
Smith, P.K., Krohn, R.I., Hermanson, G.T., Mallia, A.K., Gartner, F.H.,Provenzano, M.D., Fujimoto, E.K., Goeke, N.M., Olson, B.J., Klenk,D.C., 1985. Measurement of protein using bicinchoninic acid. Anal.Biochem. 150, 76–85.
Spurny, K.R., 1998. On the physics chemistry and toxicology of ultraWneanthropogenic atmospheric aerosols (UAAA): new advances. Toxicol.Lett. 97, 253–261.
Takenouchi, T., Munekata, E., 1995. Trophic eVects of substance P and �-amyloid peptide on dibutyryl cyclic AMP-diVerentiated human leuke-mic (HL-60) cells. Life Sci. 56, 479–484.
Takizawa, H., Ohtoshi, T., Kawasaki, S., Kohyama, T., Desaki, M., Kas-ama, T., Kobayashi, K., Nakahara, K., Yamamoto, K., Matsushima,K., Kudoh, S., 1999. Diesel exhaust particles induce NF-kB activationin human bronchial epithelial cells in-vitro: importance in cytokinetranscription. J. Immunol. 162, 4705–4711.
Tokiwa, H., Sera, N., Nakanishi, Y., Sagai, M., 1999. 8-Hydroxy-deoxy-guanosine formed in human lung tissues and the association with dieselexhaust particles. Free Radical Biol. Med. 27, 1251–1258.
Toyokuni, S., Tanaka, T., Hattori, Y., Nishiyama, Y., Yoshida, A., Uchida,K., Hiai, H., Ochi, H., Osawa, T., 1997. Quantitative immunohisto-chemical determination of 8-hydroxy-2�-deoxyguanosine by a mono-clonal antibody N451: its application to ferric nitrilotriacetate-inducedrenal carcinogenesis model. Lab. Invest. 76, 365–374.
Winyard, P.G., Blake, D.R., 1997. Antioxidant redox-regulated transcrip-tion factors and inXammation. Adv. Pharmacol. 38, 403–421.




















