
Dual effect of nitric oxide donor on adjuvant arthritis
9
Dual effect of nitric oxide donor on adjuvant arthritis Adel A. Gomaa a, ⁎, Mohsen M. Elshenawy a , Noha A. Afifi b , Eman A. Mohammed c , Romany H. Thabit a a Department of Pharmacology, Faculty of Medicine, Assiut University, Assiut, Egypt b Department of Microbiology and Immunity, Faculty of Medicine, Assiut University, Assiut, Egypt c Department of Pathology, Faculty of Medicine, Assiut University, Assiut, Egypt abstract article info Article history: Received 28 October 2008 Received in revised form 4 January 2009 Accepted 15 January 2009 Keywords: Dual NO Dose Adjuvant arthritis The effect of medical use of NO donors on the pathogenesis of arthritis is still yet unclear. We investigated the effects of the NO donor, sodium nitroprusside (SNP), on the pathogenesis of adjuvant-induced arthritis in rats. Rats were given SNP intraperitoneally either from day 5 to day 14 (as a prophylactic protocol) or from day 16 to day 25 (as a therapeutic protocol) after inoculation of adjuvant. SNP administration, whether prophylactic or therapeutic, in doses of 0.1 and 1 mg/kg/d significantly aggravated pathogenesis of adjuvant arthritis in rats. SNP-treated rats showed significant (P b 0.05) increase in arthritis index, hind paw volume, ankle joint diameter and hyperalgesia compared with control adjuvant arthritic rats. However, in adjuvant rats given the smallest dose of SNP (0.01 mg/kg/d), arthritis index, volume of hind paws, ankle joint diameter, body weight loss, and hyperalgesia were significantly lower than that of control adjuvant rats. After 30 d of the induction of adjuvant arthritis, TNF alpha levels exhibited insignificant changes either in control adjuvant rats or in rats given SNP compared with control non adjuvant rats. IL-10 levels in adjuvant control rats and adjuvant rats given 1 mg or 0.1 mg/kg/d from day 15 to day 25 were significantly lower than that of control non adjuvant rats. Histopathology examination of ankle joint showed that large doses of SNP (1 mg or 0.1 mg/kg/d) increased the mononuclear cells infiltration and erosion of cartilage induced by adjuvant while the infiltration of the inflammatory cells in the synovium of adjuvant rats treated with 0.01 mg/kg/d was minimal and the pannus was inhibited with alleviation of erosion of articular cartilage. Prophylactic small dose of SNP improved the histological status more than the therapeutic small dose. The present work reveals that SNP administration, either prophylactic or therapeutic, was deleterious in higher doses. However, the smallest dose used 0.01 mg/kg/d attenuates joint inflammation, hyperalgesia and body weight loss in adjuvant arthritic rats. These results suggest that small dose of NO donor may exert partial protective effects while the safety of the clinical use of NO donors, in higher doses, in patients with rheumatoid arthritis is questioned. © 2009 Elsevier B.V. All rights reserved. 1. Introduction Cardiovascular diseases have become the main cause of excessive mortality of patients with rheumatoid arthritis [1]. Nitric oxide donors have been used for many years as a vasodilator and symptomatic treatment for cardiovascular diseases as angina pectoris, hypertension and congestive heart failure. Therefore, the nitrate medication may be needed for treatment of cardiovascular diseases in patients with rheumatoid arthritis. On basis of epidemiological studies, NO donor medication may accentuate bone sclerosis and contribute to disease progression if used in the presence of osteoarthritis [2]. In rheumatoid arthritis (RA), it has been demonstrated that serum and synovial fluid nitrite concentrations were significantly higher than control [3]. It has been suggested that endogenous production of NO is enhanced in proportion to the degree of inflammation in patients with RA owing to enhanced iNOS activity [4]. Also, in osteoarthritis, NO production was found to be higher [5]. It mediates many of the destructive effects of interleukin-1 (IL-1) and tumor necrosis factor-alpha (TNF-alpha) in the cartilage and inhibitors of NO synthesis have demonstrated retardation of clinical and histological signs and symptoms in experimentally induced osteoarthritis and other forms of arthritis [6]. It appears likely that the increase in NO associated with arthritis can be caused by pro-inflammatory cytokines and mechanical stress and molecular oxygen is required for production of NO that is associated with osteoarthritis and RA [7]. Further evidence of the deleterious effects of NO comes from the study of Nagy et al. who supported the NO inhibiting therapeutic strategies for the treatment of chronic inflammatory diseases such as RA and concluded that local inhibition of NO synthesis at the site of synovial inflammation may provide better therapeutic tool than systemic inhibition [8]. Their prior study revealed that overproduction of NO may perturb T cell activation, differentiation and effector response which may contribute in different ways to the pathogenesis of autoimmune diseases [9]. Contrary to the above studies, there also exists the conflicting notion that NO may be protective during an inflammatory process. It has been shown that NO prevents apoptosis in rheumatoid synovial cells by directly inhibiting caspase-3-activation [10] and the local International Immunopharmacology 9 (2009) 439–447 ⁎ Corresponding author. E-mail address: [email protected] (A.A. Gomaa). 1567-5769/$ – see front matter © 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.intimp.2009.01.009 Contents lists available at ScienceDirect International Immunopharmacology journal homepage: www.elsevier.com/locate/intimp
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Dual effect of nitric oxide donor on adjuvant arthritis

International Immunopharmacology 9 (2009) 439–447
Contents lists available at ScienceDirect
International Immunopharmacology
j ourna l homepage: www.e lsev ie r.com/ locate / in t imp
Dual effect of nitric oxide donor on adjuvant arthritis
Adel A. Gomaa a,⁎, Mohsen M. Elshenawy a, Noha A. Afifi b, Eman A. Mohammed c, Romany H. Thabit a
a Department of Pharmacology, Faculty of Medicine, Assiut University, Assiut, Egyptb Department of Microbiology and Immunity, Faculty of Medicine, Assiut University, Assiut, Egyptc Department of Pathology, Faculty of Medicine, Assiut University, Assiut, Egypt
⁎ Corresponding author.E-mail address: [email protected] (A.A. Gomaa).
1567-5769/$ – see front matter © 2009 Elsevier B.V. Aldoi:10.1016/j.intimp.2009.01.009
a b s t r a c t
a r t i c l e i n f oArticle history:
The effect of medical use of Received 28 October 2008Received in revised form 4 January 2009Accepted 15 January 2009Keywords:DualNODoseAdjuvant arthritis
NO donors on the pathogenesis of arthritis is still yet unclear. We investigated theeffects of the NO donor, sodium nitroprusside (SNP), on the pathogenesis of adjuvant-induced arthritis inrats. Rats were given SNP intraperitoneally either from day 5 to day 14 (as a prophylactic protocol) or fromday 16 to day 25 (as a therapeutic protocol) after inoculation of adjuvant. SNP administration, whetherprophylactic or therapeutic, in doses of 0.1 and 1 mg/kg/d significantly aggravated pathogenesis of adjuvantarthritis in rats. SNP-treated rats showed significant (Pb0.05) increase in arthritis index, hind paw volume,ankle joint diameter and hyperalgesia compared with control adjuvant arthritic rats. However, in adjuvantrats given the smallest dose of SNP (0.01 mg/kg/d), arthritis index, volume of hind paws, ankle joint diameter,body weight loss, and hyperalgesia were significantly lower than that of control adjuvant rats. After 30 d ofthe induction of adjuvant arthritis, TNF alpha levels exhibited insignificant changes either in control adjuvantrats or in rats given SNP compared with control non adjuvant rats. IL-10 levels in adjuvant control rats andadjuvant rats given 1 mg or 0.1 mg/kg/d from day 15 to day 25 were significantly lower than that of control nonadjuvant rats. Histopathology examination of ankle joint showed that large doses of SNP (1 mg or 0.1 mg/kg/d)increased themononuclear cells infiltration and erosion of cartilage induced by adjuvant while the infiltration ofthe inflammatory cells in the synovium of adjuvant rats treated with 0.01 mg/kg/d wasminimal and the pannuswas inhibited with alleviation of erosion of articular cartilage. Prophylactic small dose of SNP improved thehistological statusmore than the therapeutic small dose. Thepresentwork reveals that SNP administration, eitherprophylactic or therapeutic, was deleterious in higher doses. However, the smallest dose used 0.01 mg/kg/dattenuates joint inflammation, hyperalgesia and bodyweight loss in adjuvant arthritic rats. These results suggestthat small dose ofNOdonormayexert partial protective effectswhile the safety of the clinical use of NOdonors, inhigher doses, in patients with rheumatoid arthritis is questioned.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
Cardiovascular diseases have become the main cause of excessivemortality of patients with rheumatoid arthritis [1]. Nitric oxide donorshave been used for many years as a vasodilator and symptomatictreatment for cardiovascular diseases as angina pectoris, hypertensionand congestive heart failure. Therefore, the nitrate medication may beneeded for treatment of cardiovascular diseases in patients withrheumatoid arthritis. On basis of epidemiological studies, NO donormedication may accentuate bone sclerosis and contribute to diseaseprogression if used in the presence of osteoarthritis [2]. In rheumatoidarthritis (RA), it has been demonstrated that serum and synovial fluidnitrite concentrations were significantly higher than control [3]. It hasbeen suggested that endogenous production of NO is enhanced inproportion to the degree of inflammation in patients with RA owing toenhanced iNOS activity [4]. Also, in osteoarthritis, NO production wasfound to be higher [5]. It mediates many of the destructive effects of
l rights reserved.
interleukin-1 (IL-1) and tumor necrosis factor-alpha (TNF-alpha) inthe cartilage and inhibitors of NO synthesis have demonstratedretardation of clinical and histological signs and symptoms inexperimentally induced osteoarthritis and other forms of arthritis [6].
It appears likely that the increase in NO associated with arthritiscan be caused by pro-inflammatory cytokines and mechanical stressand molecular oxygen is required for production of NO that isassociated with osteoarthritis and RA [7]. Further evidence of thedeleterious effects of NO comes from the study of Nagy et al. whosupported the NO inhibiting therapeutic strategies for the treatmentof chronic inflammatory diseases such as RA and concluded that localinhibition of NO synthesis at the site of synovial inflammation mayprovide better therapeutic tool than systemic inhibition [8]. Theirprior study revealed that overproduction of NO may perturb T cellactivation, differentiation and effector responsewhich may contributein different ways to the pathogenesis of autoimmune diseases [9].
Contrary to the above studies, there also exists the conflictingnotion that NO may be protective during an inflammatory process. Ithas been shown that NO prevents apoptosis in rheumatoid synovialcells by directly inhibiting caspase-3-activation [10] and the local

440 A.A. Gomaa et al. / International Immunopharmacology 9 (2009) 439–447
production of NO may be protective by the virtue of its ability toregulate the release of pro-inflammatory mediators [11]. In addition,nitric oxide donors were found to increase the production ofhyaluronic acid by synovial cells from patients with RA [12].
Other studies found that NO does not mediate the chronicinflammation and joint destruction which occur during the latterphase and the therapeutic administration of a selective inhibitor ofiNOS does not ameliorate the chronic inflammation and tissuedamage associated with adjuvant arthritis in rats [13]. Furthermore,it has been shown that NO has limited modulating effects in cartilagemetabolismwith evidence for both protective and deleterious effects[14] and no fundamental relationship between magnitude of NOproduction and arthritis susceptibility and severity suggesting thatNO has no effector role in arthritis [15]. Similarly, it has been foundthat the relationships between measures of arthritis disease activityand urinary and serum nitric oxide levels were not significant inrheumatoid patients [16].
Numerous studies have unanimously shown an associationbetween RA and impaired endothelium function [17]. Endothelialdysfunction is defined as loss of NO bioavailability in patients withchronic inflammatory conditions [18]. Some investigators havereported that activation of iNOS may lead to endothelial dysfunctionby depleting the bioavailability of tetrahydrobiopterin from endothe-lial nitric oxide synthase (e NOS) and subsequently uncouple eNOSresulting in production of superoxide anion rather than NO [19,20].More recently, Maki-Petaja et al. demonstrated an endothelialdysfunction and increased iNOS activity in rheumatoid patients [21].They suggested that inflammation is a key mediator in the process ofendothelial dysfunction possibly via activation of iNOS and increasedproduction of myeloperoxidase enzyme.
Interesting studies have implicated that NO has dual effects. It hasbeen reported that relatively low concentration of NO plays adefensive role in the immune system [22] and exerts anti-apoptoticeffects via cGMP [23] while higher concentration causes numerouspathological processes including inflammation [24], vascular damage[25] and apoptosis in various cell types [26]. Additionally, Kwak et al.showed that low concentration of SNP suppresses subsequent highconcentration SNP-induced apoptosis by inhibiting p38 kinase [27].
It is clear from the aforementioned reports that there is conflictingdata about the effect of NO donors and the effect of systemic use of NOdonors on pathogenesis of adjuvant-induced arthritis needs to beidentified. Therefore, we examine the effect of SNP as a representativeof nitric oxide donors on signs, symptoms, histopathology andcytokines in adjuvant-induced arthritis in rats.
2. Materials and methods
2.1. Animals
The experimental study was carried out using adult female albinorats of the Sprague–Dawley strain weighing between 160 and 200 g.The animals were acclimatized in a light- and temperature-controlledroom with a 12–12 h dark–light cycle. The rats were fed withcommercial pelleted rat feed and water was given ad libitum. Foodwas placed on the floor of the cage to facilitate access, as the painwhich accompanies adjuvant-induced arthritis renders the ratsimmobile and unable to use their hind limbs to obtain food from thecover mesh of the cage. The experimental protocol was approved bythe local ethical committee.
2.2. Reagents and drugs
Complete Freund's Adjuvant (CFA) was purchased from DifcoLaboratories, Detroit, Michigan, USA. Squalene was purchased fromMP Biomedicals, Inc. Sodium nitroprusside (SNP) was purchased fromSigma chemical, St. Louis, USA. SNP was freely dissolved in water.
2.3. Experimental induction of arthritis
In this study, adjuvant arthritis was induced in rats according topreviously described methods for the evaluation of rheumatoidarthritis. Based on preliminary experiments, the method of Trenthamet al. was modified by intradermal injection of 0.1 ml squalene beforeinoculation of CFA into a different site in the subplanter surface ofright hind paw to increase the sensitivity of rats used to CFA [28]. Ratswere divided into 8 groups (6 animals each). The first group (group I)served as normal control which received only 0.1 ml kg−1 saline. Eachrat in the other 7 groups received 0.1 ml of CFA and 0.1 ml of squalene.Rats in group II received intraperitoneally 0.1 ml of distilled water, thevehicle inwhich SNP was dissolved (Adjuvant arthritic control group).Treatment was initiated on day 5 to day 14 in three groups III, IV and Vwith SNP, given intraperitoneally, in doses of 1, 0.1 and 0.01 mg/kg/drespectively (prophylactic protocol). In groups VI, VII and VIII, SNPwasgiven i.p. as therapeutic protocol in doses of 1, 0.1 and 0.01 mg/kg/drespectively from day 16 to day 25. The day of inoculation wasregarded as day 0 while day 16 was the day in which oedema in thecontralateral, non-injected, hind paw was observed. Arthritis index,hind paw height, volume of paw oedema, body weight, rectaltemperature and pain threshold to pressure on hind paws, weremeasured daily from day 0 until day 30 after adjuvant inoculation. Atthe end of the study, the animals were sacrificed and blood wascollected. Blood samples were immediately centrifuged at 3000 rpmfor 10 min and serum samples were stored at −80 °C until assayed forTNF-alpha and IL-10. Specimens of ankle joints' tissues were alsoexamined for histopathology.
2.4. Arthritis index
Rats were evaluated daily for arthritis. The physical symptoms ofarthritis were judged by the following grading system [29]: 0 = normalpaws; 1 = erythema of toes; 2 = erythema and swelling of paws; 3 =swelling of ankles; 4 = complete swelling of thewhole leg and inabilityto bend it. The maximum achievable score is thus 16. Arthritis indexfor each rat was calculated by adding the four scores of individualpaws. A sensitized animal was considered to have arthritis when atleast one non-injected paw was inflamed [30].
2.5. Measurement of body weight and temperature in arthritic rats
Body weight for each rat was recorded before and daily afteradjuvant inoculation to assess food intake andweight gain throughoutthe period of arthritis. The difference between body weight in eachday and that of day 0 was calculated to determine the change in bodyweight in arthritic rats.
Body temperature, as an index of inflammation, was monitored forrats, before and daily after disease induction between 9:00 AM and11:00 AM, using a rectal thermometer.
2.6. Measurement of ankle diameter and paw volume changes
Changes in the ankle diameter of both ipsilateral (injected) andcontralateral (non-injected) hind paws, from the height on day 0, weredaily assessed using a Vernier scale [31].
Volumes of hind paws were measured before and daily afteradjuvant inoculation by using water displacement plethysmometry[32]. The changes of volumes of hind paws, from those of day 0, werecalculated.
2.7. Analgesimetry
Using a Ugo basile analgesimeter (Ugo Basile Biological ResearchApparatus, Italy), a crescent pressure (in grams) was appliedseparately to the posterior paws until the animal displayed a reaction
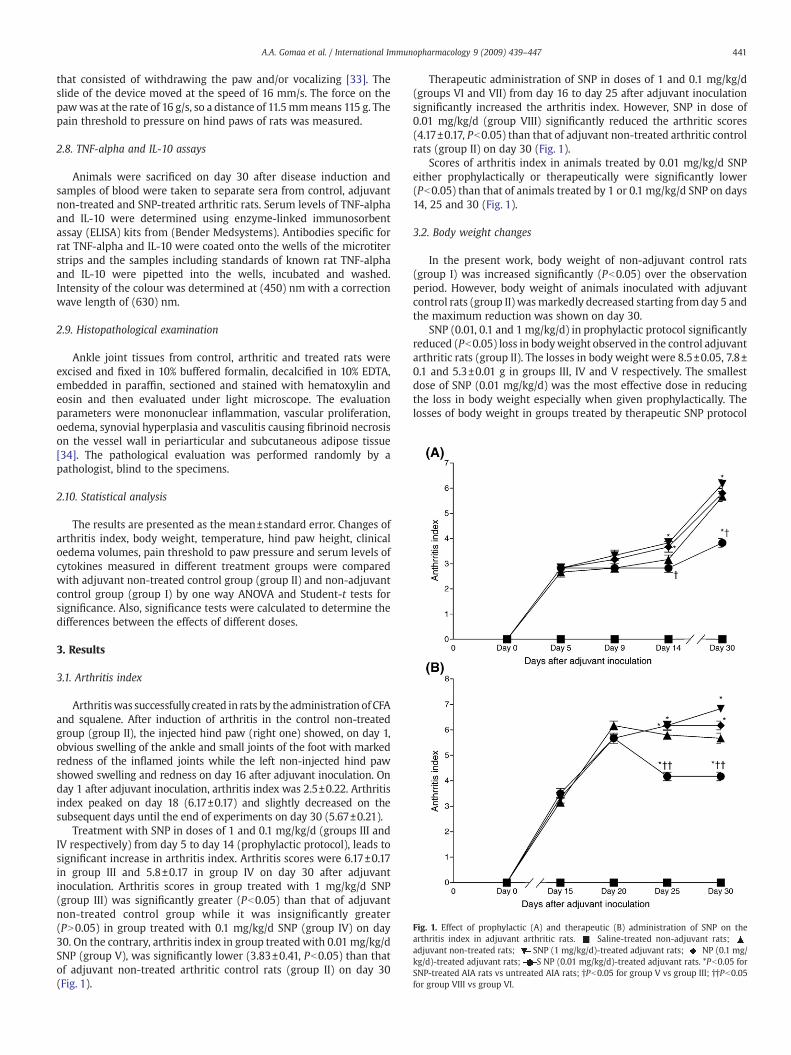
Fig. 1. Effect of prophylactic (A) and therapeutic (B) administration of SNP on thearthritis index in adjuvant arthritic rats. Saline-treated non-adjuvant rats;adjuvant non-treated rats; SNP (1 mg/kg/d)-treated adjuvant rats; NP (0.1 mg/kg/d)-treated adjuvant rats; S NP (0.01 mg/kg/d)-treated adjuvant rats. *Pb0.05 forSNP-treated AIA rats vs untreated AIA rats; †Pb0.05 for group V vs group III; ††Pb0.05for group VIII vs group VI.
441A.A. Gomaa et al. / International Immunopharmacology 9 (2009) 439–447
that consisted of withdrawing the paw and/or vocalizing [33]. Theslide of the device moved at the speed of 16 mm/s. The force on thepawwas at the rate of 16 g/s, so a distance of 11.5mmmeans 115 g. Thepain threshold to pressure on hind paws of rats was measured.
2.8. TNF-alpha and IL-10 assays
Animals were sacrificed on day 30 after disease induction andsamples of blood were taken to separate sera from control, adjuvantnon-treated and SNP-treated arthritic rats. Serum levels of TNF-alphaand IL-10 were determined using enzyme-linked immunosorbentassay (ELISA) kits from (Bender Medsystems). Antibodies specific forrat TNF-alpha and IL-10 were coated onto the wells of the microtiterstrips and the samples including standards of known rat TNF-alphaand IL-10 were pipetted into the wells, incubated and washed.Intensity of the colour was determined at (450) nm with a correctionwave length of (630) nm.
2.9. Histopathological examination
Ankle joint tissues from control, arthritic and treated rats wereexcised and fixed in 10% buffered formalin, decalcified in 10% EDTA,embedded in paraffin, sectioned and stained with hematoxylin andeosin and then evaluated under light microscope. The evaluationparameters were mononuclear inflammation, vascular proliferation,oedema, synovial hyperplasia and vasculitis causing fibrinoid necrosison the vessel wall in periarticular and subcutaneous adipose tissue[34]. The pathological evaluation was performed randomly by apathologist, blind to the specimens.
2.10. Statistical analysis
The results are presented as the mean±standard error. Changes ofarthritis index, body weight, temperature, hind paw height, clinicaloedema volumes, pain threshold to paw pressure and serum levels ofcytokines measured in different treatment groups were comparedwith adjuvant non-treated control group (group II) and non-adjuvantcontrol group (group I) by one way ANOVA and Student-t tests forsignificance. Also, significance tests were calculated to determine thedifferences between the effects of different doses.
3. Results
3.1. Arthritis index
Arthritiswas successfully created in rats by the administration of CFAand squalene. After induction of arthritis in the control non-treatedgroup (group II), the injected hind paw (right one) showed, on day 1,obvious swelling of the ankle and small joints of the foot with markedredness of the inflamed joints while the left non-injected hind pawshowed swelling and redness on day 16 after adjuvant inoculation. Onday 1 after adjuvant inoculation, arthritis index was 2.5±0.22. Arthritisindex peaked on day 18 (6.17±0.17) and slightly decreased on thesubsequent days until the end of experiments on day 30 (5.67±0.21).
Treatment with SNP in doses of 1 and 0.1 mg/kg/d (groups III andIV respectively) from day 5 to day 14 (prophylactic protocol), leads tosignificant increase in arthritis index. Arthritis scores were 6.17±0.17in group III and 5.8±0.17 in group IV on day 30 after adjuvantinoculation. Arthritis scores in group treated with 1 mg/kg/d SNP(group III) was significantly greater (Pb0.05) than that of adjuvantnon-treated control group while it was insignificantly greater(PN0.05) in group treated with 0.1 mg/kg/d SNP (group IV) on day30. On the contrary, arthritis index in group treated with 0.01 mg/kg/dSNP (group V), was significantly lower (3.83±0.41, Pb0.05) than thatof adjuvant non-treated arthritic control rats (group II) on day 30(Fig. 1).
Therapeutic administration of SNP in doses of 1 and 0.1 mg/kg/d(groups VI and VII) from day 16 to day 25 after adjuvant inoculationsignificantly increased the arthritis index. However, SNP in dose of0.01 mg/kg/d (group VIII) significantly reduced the arthritic scores(4.17±0.17, Pb0.05) than that of adjuvant non-treated arthritic controlrats (group II) on day 30 (Fig. 1).
Scores of arthritis index in animals treated by 0.01 mg/kg/d SNPeither prophylactically or therapeutically were significantly lower(Pb0.05) than that of animals treated by 1 or 0.1 mg/kg/d SNP on days14, 25 and 30 (Fig. 1).
3.2. Body weight changes
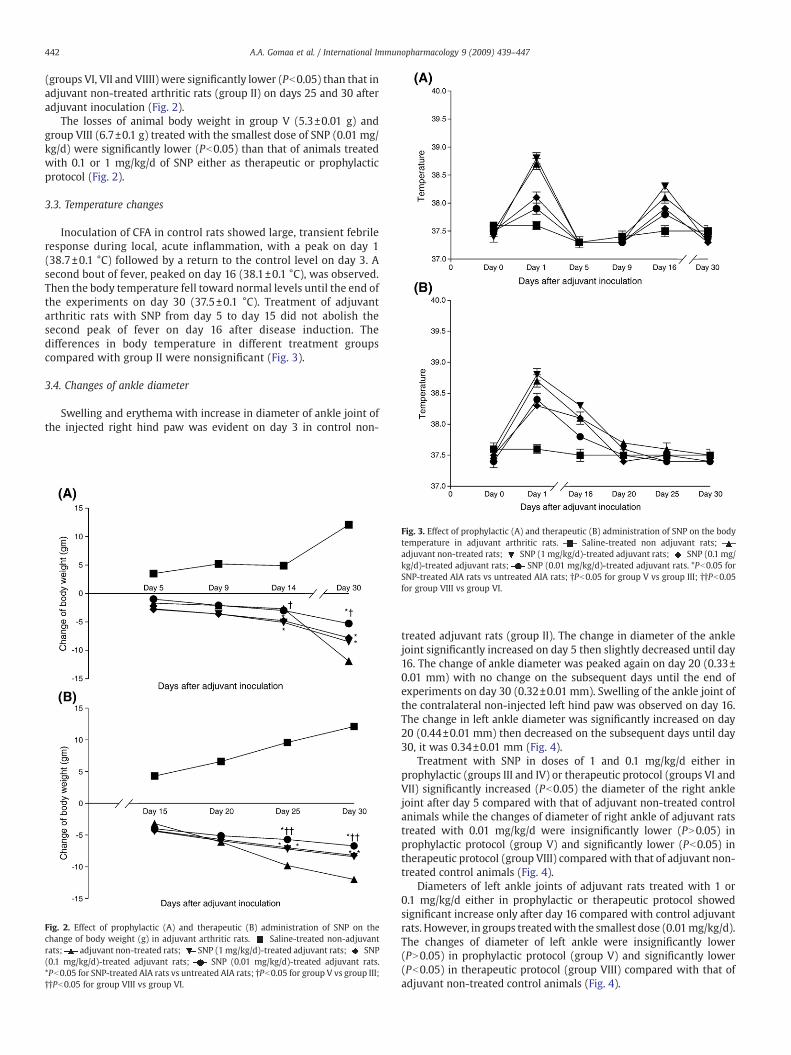
In the present work, body weight of non-adjuvant control rats(group I) was increased significantly (Pb0.05) over the observationperiod. However, body weight of animals inoculated with adjuvantcontrol rats (group II) wasmarkedly decreased starting from day 5 andthe maximum reduction was shown on day 30.
SNP (0.01, 0.1 and 1 mg/kg/d) in prophylactic protocol significantlyreduced (Pb0.05) loss in bodyweight observed in the control adjuvantarthritic rats (group II). The losses in body weight were 8.5±0.05, 7.8±0.1 and 5.3±0.01 g in groups III, IV and V respectively. The smallestdose of SNP (0.01 mg/kg/d) was the most effective dose in reducingthe loss in body weight especially when given prophylactically. Thelosses of body weight in groups treated by therapeutic SNP protocol
442 A.A. Gomaa et al. / International Immunopharmacology 9 (2009) 439–447
(groups VI, VII and VIIII) were significantly lower (Pb0.05) than that inadjuvant non-treated arthritic rats (group II) on days 25 and 30 afteradjuvant inoculation (Fig. 2).
The losses of animal body weight in group V (5.3±0.01 g) andgroup VIII (6.7±0.1 g) treated with the smallest dose of SNP (0.01 mg/kg/d) were significantly lower (Pb0.05) than that of animals treatedwith 0.1 or 1 mg/kg/d of SNP either as therapeutic or prophylacticprotocol (Fig. 2).
3.3. Temperature changes
Inoculation of CFA in control rats showed large, transient febrileresponse during local, acute inflammation, with a peak on day 1(38.7±0.1 °C) followed by a return to the control level on day 3. Asecond bout of fever, peaked on day 16 (38.1±0.1 °C), was observed.Then the body temperature fell toward normal levels until the end ofthe experiments on day 30 (37.5±0.1 °C). Treatment of adjuvantarthritic rats with SNP from day 5 to day 15 did not abolish thesecond peak of fever on day 16 after disease induction. Thedifferences in body temperature in different treatment groupscompared with group II were nonsignificant (Fig. 3).
3.4. Changes of ankle diameter
Swelling and erythema with increase in diameter of ankle joint ofthe injected right hind paw was evident on day 3 in control non-
Fig. 2. Effect of prophylactic (A) and therapeutic (B) administration of SNP on thechange of body weight (g) in adjuvant arthritic rats. Saline-treated non-adjuvantrats; adjuvant non-treated rats; SNP (1mg/kg/d)-treated adjuvant rats; SNP(0.1 mg/kg/d)-treated adjuvant rats; SNP (0.01 mg/kg/d)-treated adjuvant rats.*Pb0.05 for SNP-treated AIA rats vs untreated AIA rats; †Pb0.05 for group V vs group III;††Pb0.05 for group VIII vs group VI.
Fig. 3. Effect of prophylactic (A) and therapeutic (B) administration of SNP on the bodytemperature in adjuvant arthritic rats. Saline-treated non adjuvant rats;adjuvant non-treated rats; SNP (1 mg/kg/d)-treated adjuvant rats; SNP (0.1 mg/kg/d)-treated adjuvant rats; SNP (0.01 mg/kg/d)-treated adjuvant rats. *Pb0.05 forSNP-treated AIA rats vs untreated AIA rats; †Pb0.05 for group V vs group III; ††Pb0.05for group VIII vs group VI.
treated adjuvant rats (group II). The change in diameter of the anklejoint significantly increased on day 5 then slightly decreased until day16. The change of ankle diameter was peaked again on day 20 (0.33±0.01 mm) with no change on the subsequent days until the end ofexperiments on day 30 (0.32±0.01 mm). Swelling of the ankle joint ofthe contralateral non-injected left hind paw was observed on day 16.The change in left ankle diameter was significantly increased on day20 (0.44±0.01 mm) then decreased on the subsequent days until day30, it was 0.34±0.01 mm (Fig. 4).
Treatment with SNP in doses of 1 and 0.1 mg/kg/d either inprophylactic (groups III and IV) or therapeutic protocol (groups VI andVII) significantly increased (Pb0.05) the diameter of the right anklejoint after day 5 compared with that of adjuvant non-treated controlanimals while the changes of diameter of right ankle of adjuvant ratstreated with 0.01 mg/kg/d were insignificantly lower (PN0.05) inprophylactic protocol (group V) and significantly lower (Pb0.05) intherapeutic protocol (group VIII) compared with that of adjuvant non-treated control animals (Fig. 4).
Diameters of left ankle joints of adjuvant rats treated with 1 or0.1 mg/kg/d either in prophylactic or therapeutic protocol showedsignificant increase only after day 16 compared with control adjuvantrats. However, in groups treatedwith the smallest dose (0.01mg/kg/d).The changes of diameter of left ankle were insignificantly lower(PN0.05) in prophylactic protocol (group V) and significantly lower(Pb0.05) in therapeutic protocol (group VIII) compared with that ofadjuvant non-treated control animals (Fig. 4).
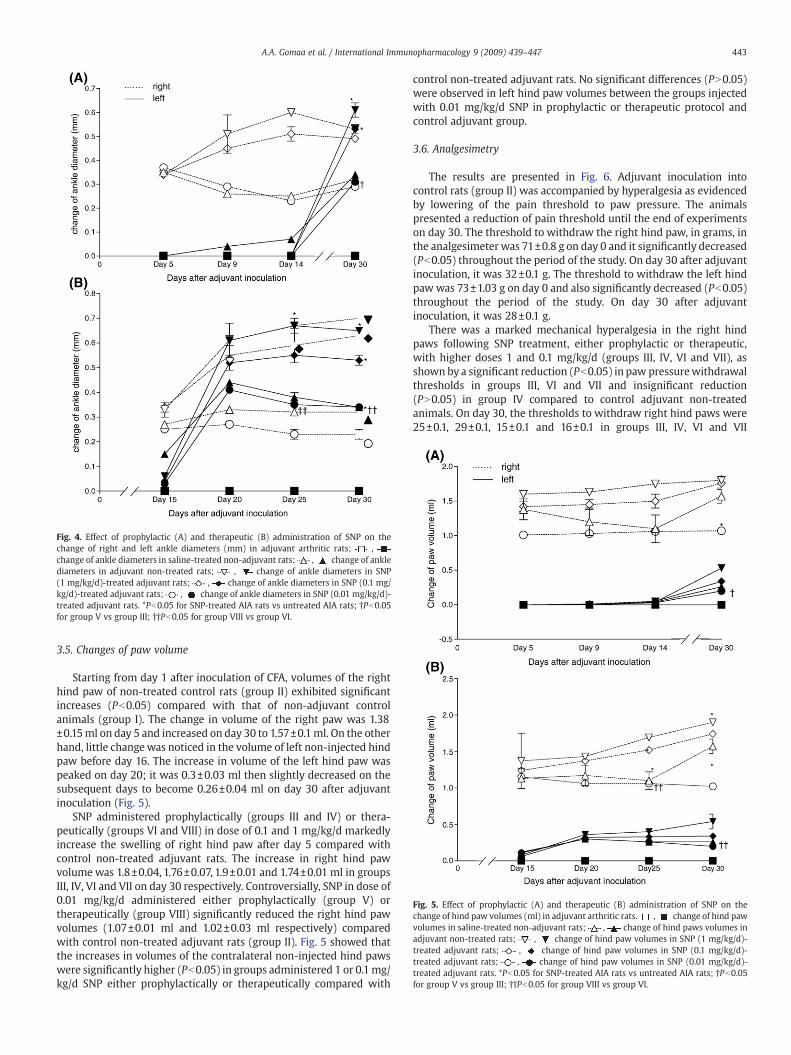
Fig. 4. Effect of prophylactic (A) and therapeutic (B) administration of SNP on thechange of right and left ankle diameters (mm) in adjuvant arthritic rats; ,change of ankle diameters in saline-treated non-adjuvant rats; , change of anklediameters in adjuvant non-treated rats; , change of ankle diameters in SNP(1 mg/kg/d)-treated adjuvant rats; , change of ankle diameters in SNP (0.1 mg/kg/d)-treated adjuvant rats; , change of ankle diameters in SNP (0.01 mg/kg/d)-treated adjuvant rats. *Pb0.05 for SNP-treated AIA rats vs untreated AIA rats; †Pb0.05for group V vs group III; ††Pb0.05 for group VIII vs group VI.
Fig. 5. Effect of prophylactic (A) and therapeutic (B) administration of SNP on thechange of hind paw volumes (ml) in adjuvant arthritic rats. , change of hind pawvolumes in saline-treated non-adjuvant rats; , change of hind paws volumes inadjuvant non-treated rats; , change of hind paw volumes in SNP (1 mg/kg/d)-treated adjuvant rats; , change of hind paw volumes in SNP (0.1 mg/kg/d)-treated adjuvant rats; , change of hind paw volumes in SNP (0.01 mg/kg/d)-treated adjuvant rats. *Pb0.05 for SNP-treated AIA rats vs untreated AIA rats; †Pb0.05for group V vs group III; ††Pb0.05 for group VIII vs group VI.
443A.A. Gomaa et al. / International Immunopharmacology 9 (2009) 439–447
3.5. Changes of paw volume
Starting from day 1 after inoculation of CFA, volumes of the righthind paw of non-treated control rats (group II) exhibited significantincreases (Pb0.05) compared with that of non-adjuvant controlanimals (group I). The change in volume of the right paw was 1.38±0.15ml on day 5 and increased on day 30 to 1.57±0.1ml. On the otherhand, little changewas noticed in the volume of left non-injected hindpaw before day 16. The increase in volume of the left hind paw waspeaked on day 20; it was 0.3±0.03 ml then slightly decreased on thesubsequent days to become 0.26±0.04 ml on day 30 after adjuvantinoculation (Fig. 5).
SNP administered prophylactically (groups III and IV) or thera-peutically (groups VI and VIII) in dose of 0.1 and 1 mg/kg/d markedlyincrease the swelling of right hind paw after day 5 compared withcontrol non-treated adjuvant rats. The increase in right hind pawvolume was 1.8±0.04, 1.76±0.07, 1.9±0.01 and 1.74±0.01 ml in groupsIII, IV, VI and VII on day 30 respectively. Controversially, SNP in dose of0.01 mg/kg/d administered either prophylactically (group V) ortherapeutically (group VIII) significantly reduced the right hind pawvolumes (1.07±0.01 ml and 1.02±0.03 ml respectively) comparedwith control non-treated adjuvant rats (group II). Fig. 5 showed thatthe increases in volumes of the contralateral non-injected hind pawswere significantly higher (Pb0.05) in groups administered 1 or 0.1mg/kg/d SNP either prophylactically or therapeutically compared with
control non-treated adjuvant rats. No significant differences (PN0.05)were observed in left hind paw volumes between the groups injectedwith 0.01 mg/kg/d SNP in prophylactic or therapeutic protocol andcontrol adjuvant group.
3.6. Analgesimetry
The results are presented in Fig. 6. Adjuvant inoculation intocontrol rats (group II) was accompanied by hyperalgesia as evidencedby lowering of the pain threshold to paw pressure. The animalspresented a reduction of pain threshold until the end of experimentson day 30. The threshold to withdraw the right hind paw, in grams, inthe analgesimeter was 71±0.8 g on day 0 and it significantly decreased(Pb0.05) throughout the period of the study. On day 30 after adjuvantinoculation, it was 32±0.1 g. The threshold to withdraw the left hindpawwas 73±1.03 g on day 0 and also significantly decreased (Pb0.05)throughout the period of the study. On day 30 after adjuvantinoculation, it was 28±0.1 g.
There was a marked mechanical hyperalgesia in the right hindpaws following SNP treatment, either prophylactic or therapeutic,with higher doses 1 and 0.1 mg/kg/d (groups III, IV, VI and VII), asshown by a significant reduction (Pb0.05) in paw pressurewithdrawalthresholds in groups III, VI and VII and insignificant reduction(PN0.05) in group IV compared to control adjuvant non-treatedanimals. On day 30, the thresholds to withdraw right hind paws were25±0.1, 29±0.1, 15±0.1 and 16±0.1 in groups III, IV, VI and VII

Fig. 6. Effect of prophylactic (A) and therapeutic (B) administration of SNP on painthreshold to pressure on hind paws in adjuvant arthritic rats. , pressure insaline-treated non-adjuvant rats; , pressure in adjuvant non-treated rats; ,
pressure in SNP (1 mg/kg/d)-treated adjuvant rats; , pressure in SNP(0.1 mg/kg/d)-treated adjuvant rats; , pressure in SNP (0.01 mg/kg/d)-treatedadjuvant rats. *Pb0.05 for SNP-treated AIA rats vs untreated AIA rats; †Pb0.05 for groupV vs group III; ††Pb0.05 for group VIII vs group VI.
Table 1Effect of i.p. administration of SNP on serum levels of TNF-α and IL-10 in adjuvantarthritic rats
Group Drug treatment Serum levels (picogram)
TNF-α IL-10
I Saline-treated (non-adjuvant) 30.7±2.3 345.6±64.4II Adjuvant arthritic (non-treated) 31.04±1.4 171.6±34*III SNP-treated [1 mg/kg/d] (prophylactic protocol) 33.4±6.8 248.3±71.1IV SNP-treated [0.1 mg/kg/d](prophylactic protocol) 32.6±2.4 225±58.6V SNP-treated [0.01 mg/kg/d] (prophylactic protocol) 31.2±3.7 249.4±52.5VI SNP-treated [1 mg/kg/d] (therapeutic protocol) 33.7±3.5 147.5±24.1*†VII SNP-treated [0.1 mg/kg/d] (therapeutic protocol) 31.8±5.6 159.6±38.8*VIII SNP-treated[0.01 mg/kg/d] (therapeutic protocol) 28.6±1.5 192.4±43.2*
Samples were taken from rats on day 30 after adjuvant inoculation.(*) Pb0.05 for SNP-treated rats vs saline-treated non adjuvant rats.(†) Pb0.05 for SNP-treated rats vs 0.01 mg/kg/d SNP-treated adjuvant arthritic rats(group II).
444 A.A. Gomaa et al. / International Immunopharmacology 9 (2009) 439–447
respectively. Hyperalgesia was significantly noticed (Pb0.05) in theleft non-injected hind paws in group VI but was insignificant (PN0.05)in groups III, IV and VII compared with control adjuvant rats (group II).The thresholds to withdraw left hind paws were 24±0.1, 26±0.1, 19±0.1 and 28±0.1 in groups III, IV, VI and VII respectively.
Contrarily, hyperalgesia in both right and left hind paws of ratstreated with SNP in a dose of 0.01 mg/kg/d, either prophylactic ortherapeutic (groups V and VIII), was significantly lower (Pb0.05)compared to adjuvant non-treated arthritic rats (group II). On day 30,the thresholds to withdraw right and left hind paws were 42±0.3and45±0.2 in group V; 45±0.1 and 41±0.1 in group VIII respectively.
3.7. Effect of SNP on serum TNF-alpha and IL-10 in adjuvant arthritic rats
As shown in Table 1, serum TNF-alpha level was insignificantly andslightly greater in adjuvant arthritic rats treated with 1 mg/kg/d SNPthan that of control adjuvant animals on day 30 after adjuvantinoculation. However there were no significant differences betweenthe serum levels of TNF-alpha in the different groups of theexperiment. Serum IL-10 level was significantly low (Pb0.05) inadjuvant non-treated control rats, (171.±34 pg) and therapeutic SNP-treated groups (group VI, VII, VIII) compared with non-adjuvantcontrol rats (345.6±64.4 pg).
There was no significant change (PN0.05) in serum levels of TNF-alpha in rats given SNP in a dose of 0.01 mg/kg/d either prophylacti-cally or therapeutically (groups V and VIII) compared with that insaline-treated adjuvant rats (group II). TNF-alpha levels were 31.2±3.7
and 28.6±1.5 pg in groups V and VIII respectively. Prophylactictreatment with 0.01 mg/kg/d SNP (group V) insignificantly increasedIL-10 levels (249.8±52.5, PN0.05) compared with those in saline-treated adjuvant rats (group II). However, there was significantdifferences between the IL-10 levels in adjuvant rats treatedtherapeutically with 1 mg/kg/d (147.5±24.1 pg) and 0.01 mg/kg/d(192.4±43.2 pg).
3.8. Histopathological examination
The histopathological examination of the left ankle joint revealednormal joint space, synovial lining, articular cartilage and subchondralbone in saline-treated non-adjuvant arthritic rats (Fig. 7A). On thecontrary, synovium of adjuvant-arthritic control rats was oedematousand thickened with a dense perivascular inflammatory infiltratecomposed of lymphocytes, plasma cells and macrophages filled thesynovial stroma. The vascularity was increased and the inflamed andhyperemic synovium crept over the articular cartilage forming apannus and causes erosion of the underlying cartilage (Fig. 7B).
In adjuvant arthritic rats treatedwith higher dose of SNP (1mg/kg/d)as prophylactic treatment, the examination showed severe destructionof the cartilage and subchondral bone with complete filling of the jointspace with pannus producing fibrous ankylosis of the joint (Fig. 7C).Therapeutic treatment of adjuvant rats with 1 mg/kg/d induced muchfibrovascular thickening of the synovium and excessive increase of theinflammatory cellular infiltrate, with scattered giant cells. The smallblood vessels are obstructed by endarteritis obliterans. Fibrocellularpannus was observed eroding the articular cartilage causing itsdestruction with penetration into the subchondral bone formingjuxta-articular erosions and subchondral cysts (Fig. 7D).
The infiltration of the inflammatory cells in the synovium ofadjuvant arthritic rats treated with the small dose of (0.01 mg/kg/d)SNP from day 5 to day 14 was minimal. The pannus was partlyinhibited with alleviation of the destruction of articular cartilagecompared with the adjuvant non-treated arthritic rats (Fig. 7E).Similarly, proliferation and infiltration of inflammatory cells in thesynovium of adjuvant arthritic rats treated with 0.01 mg/kg/d SNPfrom day 16 to day 25 were inhibited and the erosion of the articularcartilage was alleviated (Fig. 7F).
4. Discussion
Adjuvant arthritis in rat is an experimentalmodel that sharesmanyfeatures with human RA, such as swelling, cartilage degradation, andloss of joint function. It has been used for many years for evaluation ofanti-arthritic/anti-inflammatory agents [35]. In this model, ratsdevelop a chronic swelling in multiple joints, with influx ofinflammatory cells, erosion of joint cartilage and bone destructionafter inoculation of Complete Freund's Adjuvant (CFA). In this study,

Fig. 7. Representative histopathology of left ankle joints of (A) normal control rats (normal synovial lining, articular cartilage and subchondral bone), (B) adjuvant arthritic rats non-treated rats, (C) adjuvant arthritic rats treated with SNP (1 mg/kg/d) from day 5 to day 14, (D) adjuvant arthritic rats treated with SNP (1 mg/kg/d) from day 16 to day 25, (E) adjuvantarthritic rats treated with SNP (0.01 mg/kg/d) from day 5 to day 14, and (F) adjuvant arthritic rats treated with SNP (0.01 mg/kg/d) from day 16 to day 25. Note the increase in degreeof inflammation with cartilage and bone erosion by high dose of SNP and the marked inhibition of inflammatory cellular infiltration in the synovium with alleviation of articularcartilage erosion by the low dose of SNP.
445A.A. Gomaa et al. / International Immunopharmacology 9 (2009) 439–447
signs and symptoms of rheumatoid arthritis did not appear incontralateral non-injected hind paw after CFA inoculation so, themethod of induction of adjuvant arthritis wasmodified by intradermalinoculation of squalene in addition to CFA into the subplanter surfaceof right hind paw, to increase sensitivity of arthritic rats to CFA.Squalene was also used by others to potentiate the effect of CFA [36].
The present study demonstrated protective and deleterious effectsfor SNP administration on the local inflammatory infiltrate in theadjuvant arthritic rats. It was observed that the higher doses of SNP (1and 0.1 mg/kg/d) significantly exacerbated inflammatory processes inthe injected and non-injected hind paws of arthritic rats whetherthese doses were administered early (prophylactic) i.e. from day 5 today 14 or late (therapeutic) i.e. from day 16 to day 25. These dosesprovoked a significant increase in paw volume, ankle diameter,arthritis score and degree of cartilage erosion. On the contrary, this
study revealed that small dose of SNP (0.01mg/kg/d) was successful inreducing the inflammatory responses associated with induction ofarthritis by adjuvant and squalene. Prophylactic and therapeutic use of0.01 mg/kg/d SNP decreased significantly the arthritis index, pawvolume and ankle diameter of right hind paw of arthritic ratscompared with control adjuvant arthritic rats. Furthermore, smalldose of SNP reducedmarkedly the inflammatory cellular infiltration insynovium with alleviation of articular cartilage erosion.
High dose effects in the present study are in agreement with manyprevious studies. Topical application of nitroglycerin has beenreported to accelerate both cartilage degeneration and subchondralbone sclerosis in osteoarthritis [37,38]. Lane et al. demonstrated thatNO donormedicationmay accentuate bone sclerosis and contribute todisease progression if used in the presence of osteoarthritis [2]. Otherstudies reported that SNP mediate chondrocyte death and it has been

446 A.A. Gomaa et al. / International Immunopharmacology 9 (2009) 439–447
viewed as NO-mediated form of chondrocyte apoptosis. Kim et al. inline with Del Carlo and Loeser, reported that high doses of SNP inducechondrocyte death [39,40]. It was believed that NO was the solemediator of SNP-induced chondrocyte death and peroxynitrite, areactionproduct of NOand superoxide anion, or theprimarybyproductsof the decomposition of SNP might contribute to its cytotoxicity [41].
Patients with rheumatoid arthritis have documented evidence forincreased endogenous NO synthesis [8] and increased iNOS activity incomparison with control [21] supporting the hypothesis that over-production of NO may be important in pathogenesis of RA. Directinvolvement of NOergic mechanisms in the development of adjuvantarthritis was also demonstrated by many investigators [42,43]. More-over, Vuolteenaho et al. and Nagy et al. suggested that NO mediatesmany of destructive effects of IL-1 and TNF-alpha in the cartilage andthat inhibitors of NO synthesis have demonstrated retardation of clinicaland histopathological signs and symptoms in experimentally inducedosteoarthritis and other forms of arthritis [6,9].
However, controversies regarding NO role and NO donors' effecthave arisen. Laitao et al. demonstrated that locally applied isosorbidedinitrate decreased bone resorption in experimental periodontitis inrats suggesting a local anti-inflammatory effect of isosorbide [44]. Inrecent observation, nitroglycerin has been shown to stimulate cellproliferation and osteoblastic differentiation of human bone marrow-derived mesenchymal stem cells through a direct release of NO [45].SNP was effective in stimulation of synovial cells from patients withRA and increasing hyaluronic acid production [12]. In addition, nitratewas found to exert a protective effect upon hypochlorus acid-inducedchondrocyte toxicity suggesting that NO has a novel cytoprotectiverole in inflamed joints [46].
Agreeing with such reports, our results showed that the low dose ofSNP (0.01 mg/kg/d) used in this study has possible anti-inflammatoryeffects. Therefore, our findings suggest that SNP produces dual effects(which is dose dependent).Many studies support thepresentfindings. Ithas been reported that lowconcentration of SNP suppresses subsequenthigh concentration SNP-induced apoptosis [27]. Furthermore, Kim et al.reported that SNP in low concentrations protects chondrocytes fromtoxic effect of the higher concentration [39]. Consistent with the notionof adual effects ofNOdonors, several experiments have shown that localapplication of drugs generating a lowNO concentration reduces incisionpain through activationof guanylate cyclasewhiledrugs generatinghighNO concentration intensify pain via guanylate cyclase-independentmechanism [47,48]. With respect to joint pain there is only one clinicalstudy showing that transdermal nitroglycerin provided partial sympto-matic pain relief in patient with painful shoulders [49]. Consistent withthese data ourwork demonstrated that high doses of theNOdonor, SNPintensified the hyperalgesia induced by CFA in arthritic rats while thesmall dose of SNP reduced the hyperalgesia. The precise mechanism ofhow NO is affecting pain transduction pathways remains unclear.However, Duarte et al. reported that NO-promoted analgesia wasblocked by cGMP inhibitor [50]. The analgesic effect of sildenafil, aphosphodiesterase-5-inhibitor working via increasing cGMP produc-tion,waspotentiated byNOdonors. Therefore, NOdonor lowdose couldbe protective and analgesic through activation of cGMP.
In the present experiments, it was observed that, in adjuvant non-treated arthritic rats (group II), therewas significant loss of bodyweightsin arthritic animals while the bodyweight in saline non-adjuvant group(group I) increased gradually over the study duration. This is inagreement with others who reported that body weight loss is anindication of abnormal conditions [51,52]. Koufany et al. also demon-strated that in all arthritic rats, body weight decreased progressively asarthritis settled [53]. This study demonstrated that SNP treatmentsignificantly reduced the loss in body weight observed in the adjuvantnon-treated arthritic animals and the smallest dose 0.01 mg/kg/d wasthe most effective in reducing the decrease in body weight especiallywhen given prophylactically. Weight loss can be explained by thedecrease of food intake observed throughout the period of the study due
to immobility accompanying hyperalgesia which was less in animalstreated with the low dose of SNP. To our knowledge, no study reportedthe effect of NO donors on the body weight loss of arthritic animals.
Considering the contribution of TNF-alpha and IL-10 to adjuvantarthritis severity, the present study showed that serum TNF-α level inadjuvant arthritic non-treated rats was slightly higher than that ofnormal rats with no arthritis, on day 30 after induction of arthritis. Thedifferences in TNF-α levels between the groups were insignificant.This observation is consistent with Philippe et al. who reported thatthe systemic TNF-α concentration had significantly increased 6 h afteradjuvant injection, peaked at 12 h, returned to near controlconcentrations on day 2, and increased slightly until day 20 [30].Serum IL-10 levels in control adjuvant rats were significantly lowercompared with those in saline-treated non-adjuvant rats. This isconsistent with the fact that IL-10 has potent anti-inflammatoryactivity [54]. This study demonstrated that serum IL-10 levels on day30 after adjuvant inoculation, in rats given SNP therapeutically, weresignificantly reduced compared with those in saline control rats.However, IL-10 levels in animals given the lower dose, 0.01 mg/kg/d,were higher than those in animals given the higher dose, 1 mg/kg/d.
In this study, our histological observations revealed that theadministration of large doses of SNP in arthritic rats was associatedwith the increase in the destruction of the inflamed joint.We speculatethat this observed effect may be due to the increase of the formation ofperoxynitrite in the synovium where peroxynitrite produces cellularinjury and necrosis. However, the small dose of SNP reduced theinflammatory cellular infiltration in the synovium with alleviation ofthe articular cartilage erosion. We proposed that the protective effectof the small dose is secondary to a reduced formation of peroxynitritein the synovium,where small dose of SNPmayexert negative feed backregulation on iNOS gene expression.[55,56].
In conclusion, the results presented in this study reveal that systemicuse of NOdonor, SNP,may produce either exacerbation or attenuation ofadjuvant arthritis in rats depending on administered dose. The smallestdose of SNP has protective effect against inflammatory processes,hyperalgesia and weight loss in adjuvant arthritic rats. Also, small dosereduces the serum TNF-alpha and insignificantly increases serum IL-10while higher doses have the reverse actions. These findings suggest thatnitrite medication in large dose contributes to disease progression ofrheumatoid arthritis but low dose may exert protective effects againstthe pathogenesis of the disease.
References
[1] Wallberg-Jonsson S, Ohman ML, Dahlqvist SR. Cardiovascular morbidity andmortality in patients with seropositive rheumatoid arthritis in northern Sweden.J Rheumatol 1997;24:445–51.
[2] Lane NE,Williams EN, Hung YY, H ochbergMC, Cummings SR, Nevitt MC. Associationof nitrate use with risk of new radiographic features of hip osteoarthritis in elderlywhite women: the study of osteoporotic fractures. Arthritis Rheum 2003;49:752–8.
[3] Jacob T, Morrell M, Manzi S, Verdile V, Simmons RL, Peitzman A. Elevated nitrites ininflammatory joint disease: nitric oxide in the arthritides. Arthritis Rheum1992;35:R9–15.
[4] Yki-Jarvinen H, Bergholm R, Leirisalo-Repo M. Increased inflammatory activityparallels increased basal nitric oxide production and blunted response to nitricoxide in vivo in rheumatoid arthritis. Ann Rheum Dis 2003;62:630–4.
[5] Scher JU, Pillinger MH, Abramson SB. Nitric oxide synthases and osteoarthritis.Curr Rheum Reports 2007;9:9–15.
[6] Vuolteenaho K, Moilanen T, Knowles RG, Moilanen E. The role of nitric oxide inosteoarthritis. Scand J Rheumatol 2007;36:247–58.
[7] Fermor B, Christensen SE, Youn I, Cernancec JM, Davies CM, Weinberg JB. Oxygen,nitric oxide and articular cartilage. Eur cells Mterials 2007;13:56–65.
[8] Nagy G, Clark JM, Buzas E, Gorman C, Pasztoi M, Koncz A, et al. Nitric oxideproduction of T lymphocytes is increased in rheumatoid arthritis. Immunol Lett2008;118:55–8.
[9] Nagy G, Clark JM, Buzas EI, Gorman CL, Cope AP. Nitric oxide, chronic inflammationand autoimmunity. Immunol Lett 2007;111:1–5.
[10] Migita K, Yamasaki S, Kita M, Ida H, Shibatomi K. Nitric oxide protects culturedrheumatoid synovial cells from Fas-induced apoptosis by inhibiting caspase-3.Immunology 2001;103:362–7.
[11] Paul-Clark MJ, Gilory DW, Willis D, Willoughby DA, Tomlinson A. Nitric oxidesynthase inhibitors have opposite effects on acute inflammation depending ontheir route of administration. J Immunol 2001;166:1169–77.

447A.A. Gomaa et al. / International Immunopharmacology 9 (2009) 439–447
[12] Chenevier-Gobeaux C, Morin-Robinet S, Lemarechal H, Poiraudeau S, Ekindjian JC,Borderie D. Effects of pro- and anti-inflammatory cytokines and nitric oxide donorsonhyaluronic acid synthesis by synovial cells frompatientswith rheumatoid arthritis.Clin Sci 2004;107:291–6.
[13] Fletcher DS, Widmer WR, Luell S, Christen A, Orevillo C, Shah S, et al. Therapeuticadministration of a selective inhibitor of nitric oxide synthase does not amelioratethe chronic inflammation and tissue damage associated with adjuvant-inducedarthritis in rats. J Pharmacol Exp Ther 1998;284:714–21.
[14] Jang D, Murrell GA. Nitric oxide in arthritis. Free Radic Biol Med 1998;24:1511–9.[15] Miletie T, Kovacevie V, ujie V, Stanojevic S, Mitie K, Lazarevic Macanovic M, et al.
Reactive oxygen species (ROS), but not nitric oxide (NO), contribute to straindifferences in the susceptibility to experimental arthritis in rats. Immunobiology2007;212:95–105.
[16] Weinberg JB, Fermor B, Guilak F. Nitric oxide synthase and cyclooxygenaseinteractions in cartilage and meniscus: relationships to joint physiology, arthritis,tissue repair. Subcell Biochem 2007;42:31–62.
[17] Hansel S, Lssig G, Pistrosch F, Passauer J. Endothelial dysfunction in young patientswith long-term rheumatoid arthritis and low disease activity. Atherosclerosis2003;170:177–80.
[18] Vaudo G, Marchesi S, Gerli R. Endothelial dysfunction in young patients withrheumatoid arthritis and low disease activity. Ann Rheum Dis 2004;63:31–5.
[19] Gunnett CA, LundDD,McDowell AK, Faraci FM, Heistad DD.Mechanisms of induciblenitric oxide synthase-mediated vascular dysfunction. Arterioscler Thromb Vasc Biol2005;25:1617–22.
[20] Xu J, Xie Z, Reece R, Pimental D, Zou MH. Uncoupling of endothelial nitric oxidesynthase by hypochlorus acid: role of NAD(P)H oxidase-derived superoxide andperoxynitrite. Arterioscler Thromb Vasc Biol 2006;26:2688–95.
[21] Maki-Petaja KM, Cheriyan J, Booth AD, Hall FC, Brown J, Wallace SML, et al.Inducible nitric oxide synthase is increased in patients with rheumatoid arthritisand contributes to endothelial dysfunction. Int J Cardiol 2008;129:399–405.
[22] Hobbs AJ, Higgs A, Moncada S. Inhibition of nitric oxide synthase as a potentialtherapeutic target. Annu Rev Pharmacol Toxicol 1999;39:191–220.
[23] Dimmeler S, Haendeler J, Nehls M, Zeiher A. Suppression of apoptosis by nitricoxide via inhibition of interleukin-1beta-converting enzyme (1CE)-like andcysteine protease protein (CPPP)-32-like proteases. J Exp Med 1997;185:601–7.
[24] Bogdan C. Nitric oxide and the immune response. Nat Immunol 2001:907–16.[25] Luoma JS, Stralin P, Marklund SL, Hiltunen TP, Sarkioja T, Yla-Hertluala S.
Expression of SOD and iNOS in macrophages and smooth muscle cells in humanand rabbit atherosclerotic lesions: colocalization with epitopes characteristic ofoxidized LDL and peroxynitrite-modified proteins. Atheroscler Thromb Vasc Biol1998;18:157–67.
[26] Arstall MA, Sawyer DM, Fukazawa R, Kelly RA. Cytokine-mediated apoptosis incardiac myocytes: the role of inducible nitric oxide synthase induction andperoxynitrite generation. Circ Res 1999;85:829–40.
[27] Kwak HJ, Park KM, Lee S, Lim HJ, Go SH, Eom SM, et al. Preconditioning with lowconcentration NO attenuates subsequent NO-induced apoptosis in vascularsmooth muscle cells via HO-1-dependent mitochondrial death pathway. ToxicolAppl Pharmacol 2006;217:176–84.
[28] Trentham DE, Townes AS, Kang AH. Autoimmunity to type II collagen: anexperimental model of arthritis. J Exp Med 1977;146:857–68.
[29] Wooley PH, Luthra HS, Stuart JM, David CS. Type II collagen-induced arthritis inmice. Major histocompatibility complex (I region) linkage and antibodyresponse. J Exp Med 1981;154:688–700.
[30] Philippe L, Pascale GP, Corinne G, Karim B, Bernard T, Patrick N, et al. Relationsbetween functional, inflammatory, and degenerative parameters during adjuvantarthritis in rats. Am J Physiol 1997;273:R1550–6.
[31] Mary RL, Geetha T, Varalakshmi P. Effect of Vernonia cinerea less flower extract inadjuvant-induced arthritis. Gen Pharmaclol 1998;31:601–6.
[32] David LB, Elizabeth AK, Michael FJ, Chih-Hung L, Shripad SB, Michael W, et al. Anti-inflammatory effects of ABT-702, a novel non-nucleoside adenosine kinaseinhibitor, in rat adjuvant arthritis. J Pharmacol Exp Ther 2001;296:495–500.
[33] Andersen ML, Tufik S. Altered sleep and behavioral patterns of arthritic rats. SleepRes Online 2000;3:161–7.
[34] McCartney-Francis NL, Chan J, Wahl SM. Inflammatory joint disease: clinical,histological, and molecular parameters of acute and chronic inflammation andtissue destruction. In: Winyard PG, Willoughby DA, editors. Inflammationprotocols. New Jersey: Humana Press; 2003. p. 147–59. G.J. Dusting. Nitric oxidein coronary artery disease: roles in atherosclerosis, myocardial.
[35] Watnich AS. The adjuvant arthritic rat: a tool for discovering antiarthritic drugs.In: Rosenthale ME, Mansmann Jr H, editors. Immunopharmacology. New York:Spectrum; 1975. p. 90–105.
[36] Santos L, Tipping PG. Attenuation of adjuvant arthritis in rats by treatment withoxygen radical scavengers. Immunol Cell Biol 1994;72:747–9.
[37] CakeMA,AppleyardRC,ReadRA,GhoshP, SwainMV,MurrellGC. Topical administrationof the nitric oxide donor glyceryl trinitrate modifies the structural and biomechanicalproperties of ovine articular cartilage. Osteoarthr Cartil 2003;11:872–8.
[38] Cake MA, Read RA, Appleyard RC, Hwa SY, Ghosh P. The nitric oxide donor glyceryltrinitrate increases subchondral bone sclerosis and cartilage degenerationfollowing ovine meniscectomy. Osteoarthr Cartil 2004;12:974–81.
[39] KimHA, Lee KB, Bae SC. Themechanism of low-concentration sodiumnitroprusside-mediated protection of chondrocyte death. Arthritis Res Therapy 2005;7:R 526–35.
[40] Del Carlo M, Loeser RF. Nitric oxide-mediated chondrocyte cell death requires thegeneration of additional reactive oxygen species. Arthritis Rheum 2002;46:394–403.
[41] Del Carlo M, loeser RF. Increased oxidative stress with aging reduces chondrocytesurvival: correlation with intracellular glutathione levels. Arthritis Rheum 2003;48:3419–30.
[42] Afonso V, Champy R, Mitrovic D, Collin P, Lomri A. Reactive oxygen species andsuperoxide dismutases: role in joint diseases. Jt Bone Spine 2007;74:324–9.
[43] Belgolovykh LA, Dubikov AI, Medved EE. Nitroxide synthase activities at differentstages of adjuvant arthritis. Bull Exp Biol Med 2007;143(5):638–41.
[44] Laitao RF, Rocha FA, Chaves HV, Lima V, Cunha FQ, Ribeiro RA, et al. Locally appliedisosorbide decreases bone resorption in experimental periodontitis in rats. JPeriodontol 2004;75:1227–32.
[45] Huang L, Qui N, Zhang C, Wei HY, Li YL, Zhou HH, et al. Nitroglycerin enhancesproliferation and osteoblastic differentiation in human mesenchymal stem cellsvia nitric oxide pathway. Acta Pharmacol Sin 2008;29:580–6.
[46] Whiteman M, Rose P, Siau JL, Halliwell B. Nitrite-mediated protection againsthypochlorous acid-induced chondrocyte toxicity: a novel cytoprotective role ofnitric oxide in the inflamed joint? Arthritis Rheum 2003;48:3140–50.
[47] Prado WA, Schiavon VF, Cunha FQ. Dual effect of local application of nitric oxidedonors in a model of incision pain in rats. Eur J Pharmacol 2002;441:57–65.
[48] Rocha JCS, Peixoto MEB, Jancer S, Cunha FQ, Ribeiro RA, Rocha FAC. Dual effect ofnitric oxide in articular inflammatory pain in zymosan-induced arthritis in rats. Br JPharmacol 2002;136:588–96.
[49] Berrazueta JR, Losada A, Poveda J, Ochoteco A, Riestra A, Salas E, et al. Successfultreatment of shoulder pain syndrome due to supraspinatus tendonitis withtransdermal nitroglycerin. A double blind study. Pain 1995;66:63–7.
[50] Duarte IDG, Lorenzetti BB, Ferreira SH. Peripheral analgesia and activation of thenitric oxide–cyclic GMP pathway. Eur J Pharmacol 1990;186:289–93.
[51] Calvino B, Crepon-BernardMA, Le Bars D. Parallel clinical and behavioral studies ofadjuvant-induced arthritis in the rat: possible relationship with “chronic pain”.Behav Brain Res 1987;24:11–29.
[52] Dardick SJ, Basbaum AI, Levine JD. The contribution of pain to disability inexperimentally induced arthritis. Arthritis Rheum 1989;29:1017–22.
[53] Koufany M, Moulin D, Bianchi A, Muresan M, Sebillaud S, Netter P, et al. Anti-inflammatory effect of antidiabetic thiazolidinediones prevents bone resorptionrather than cartilage changes in experimental polyarthritis. Arthritis Res Therapy2008;10:R6–15.
[54] Fiorentino DF, Zlotinik A, Mosmann TR, HowardM, O'Garra A. Interleukin-10 inhibitscytokine production by activated macrophages. J Immunol 1991;147:3815–22.
[55] Hancock CM, Riegger-Krugh C. Modulation of pain in osteoarthritis: role of nitricoxide. Clin J Pain 2008;24:353–65.
[56] Chang K, Lee SJ, Cheong I, Billiar TR, Chung HT, Han JA, et al. Nitric oxide suppressesinducible nitric oxide synthase expression by inhibiting post-translation modifi-cation of 1KappaB. Exp Mol Med 2004;36:311–24.




















