
Does spawning behavior minimize exposure to environmental stressors for encapsulated gastropod...
12
Mar Biol (2007) 152:991–1002 DOI 10.1007/s00227-007-0748-3 123 RESEARCH ARTICLE Does spawning behavior minimize exposure to environmental stressors for encapsulated gastropod embryos on rocky shores? R. Przeslawski · A. R. Davis Received: 29 January 2007 / Accepted: 6 June 2007 / Published online: 13 July 2007 © Springer-Verlag 2007 Abstract Adults of motile intertidal invertebrates are able to seek shelter to avoid environmental stress associated with low tides, but embryos within egg masses are eVec- tively sessile for the duration of their encapsulation. Gastro- pod egg masses from 34 taxa on two rocky shores in SE Australia (34°3708S, 150°9203E and 34°3545S, 150°5320E) were surveyed over 2 years (June 2002–May 2004) to test the hypothesis that eggs are deposited in pat- terns that minimize exposure to environmental stress. Egg masses were expected to be predominantly deposited in shaded habitats not prone to environmental extremes. It was also anticipated that the deposition of egg masses in habi- tats exposed to UVR, desiccation, and/or extremes in tem- perature would occur when exposure to these abiotic factors was minimized. Among the taxa investigated, only four species spawned in full sun (Bembicium nanum, Nerita morio, Siphonaria zelandica and S. denticulata). Summer had the highest UVR index, water temperature, and air tem- perature as well as the lowest daytime tides. Univariate and multivariate analyses conWrmed that egg mass abundance was highest during summer, with no change in egg mass size. This study shows that those species depositing egg masses on the surfaces of rock platforms do not adjust the seasonal timing or macrohabitat location of their spawning to avoid physiologically stressful conditions, particularly UVR. Alternate reasons for the evolution of egg mass deposition behavior in apparently sub-optimal habitats are discussed, and it is almost certainly the complex interplay of a variety of highly species-speciWc factors that is respon- sible for the patterns observed. Introduction Intertidal environments are extremely dynamic, and organ- isms within them are often exposed to a range of potential stressors including desiccation, wave force, solar radiation, and marked Xuctuations in temperature and salinity (e.g., Gosselin and Chia 1995). These abiotic factors often oper- ate concurrently, and organisms in such environments may be synchronously exposed to multiple stressors during low tides (Newell 1970; HoVman et al. 2003; Przeslawski 2004). For intertidal organisms, an ability to withstand exposure to environmental stress is critical to survival (reviewed by Moran 1999; Przeslawski 2004). Embryos and larvae are considered the most vulnerable life stage of most organisms (Thorson 1950; Jackson and Strathmann 1981). Previous research has demonstrated that mortality and abnormality rates of marine invertebrate embryos and larvae increases following exposure to eco- logically realistic levels of ultraviolet radiation (UVR) (Adams and Shick 2001), desiccation (Pechenik 1978), fouling (Biermann et al. 1992), and extremes in salinity (Yaroslavtseva et al. 2001) or temperature (Ushakova 2003). Many intertidal organisms circumvent the negative eVects of environmental stressors by releasing gametes or larvae directly into the water column or brooding their Communicated by J.P. Grassle. R. Przeslawski (&) · A. R. Davis Institute for Conservation Biology, School of Biological Sciences, University of Wollongong, NorthWelds Avenue, Wollongong, NSW 2522, Australia e-mail: [email protected] Present Address: R. Przeslawski Department of Ecology and Evolution, Stony Brook University, 650 Life Sciences Building, Stony Brook, NY 11794-5245, USA
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Does spawning behavior minimize exposure to environmental stressors for encapsulated gastropod...

Mar Biol (2007) 152:991–1002
DOI 10.1007/s00227-007-0748-3RESEARCH ARTICLE
Does spawning behavior minimize exposure to environmental stressors for encapsulated gastropod embryos on rocky shores?
R. Przeslawski · A. R. Davis
Received: 29 January 2007 / Accepted: 6 June 2007 / Published online: 13 July 2007© Springer-Verlag 2007
Abstract Adults of motile intertidal invertebrates are ableto seek shelter to avoid environmental stress associatedwith low tides, but embryos within egg masses are eVec-tively sessile for the duration of their encapsulation. Gastro-pod egg masses from 34 taxa on two rocky shores in SEAustralia (34°37�08�S, 150°92�03�E and 34°35�45�S,150°53�20�E) were surveyed over 2 years (June 2002–May2004) to test the hypothesis that eggs are deposited in pat-terns that minimize exposure to environmental stress. Eggmasses were expected to be predominantly deposited inshaded habitats not prone to environmental extremes. It wasalso anticipated that the deposition of egg masses in habi-tats exposed to UVR, desiccation, and/or extremes in tem-perature would occur when exposure to these abioticfactors was minimized. Among the taxa investigated, onlyfour species spawned in full sun (Bembicium nanum, Neritamorio, Siphonaria zelandica and S. denticulata). Summerhad the highest UVR index, water temperature, and air tem-perature as well as the lowest daytime tides. Univariate andmultivariate analyses conWrmed that egg mass abundancewas highest during summer, with no change in egg masssize. This study shows that those species depositing egg
masses on the surfaces of rock platforms do not adjust theseasonal timing or macrohabitat location of their spawningto avoid physiologically stressful conditions, particularlyUVR. Alternate reasons for the evolution of egg massdeposition behavior in apparently sub-optimal habitats arediscussed, and it is almost certainly the complex interplayof a variety of highly species-speciWc factors that is respon-sible for the patterns observed.
Introduction
Intertidal environments are extremely dynamic, and organ-isms within them are often exposed to a range of potentialstressors including desiccation, wave force, solar radiation,and marked Xuctuations in temperature and salinity (e.g.,Gosselin and Chia 1995). These abiotic factors often oper-ate concurrently, and organisms in such environments maybe synchronously exposed to multiple stressors during lowtides (Newell 1970; HoVman et al. 2003; Przeslawski2004). For intertidal organisms, an ability to withstandexposure to environmental stress is critical to survival(reviewed by Moran 1999; Przeslawski 2004).
Embryos and larvae are considered the most vulnerablelife stage of most organisms (Thorson 1950; Jackson andStrathmann 1981). Previous research has demonstrated thatmortality and abnormality rates of marine invertebrateembryos and larvae increases following exposure to eco-logically realistic levels of ultraviolet radiation (UVR)(Adams and Shick 2001), desiccation (Pechenik 1978),fouling (Biermann et al. 1992), and extremes in salinity(Yaroslavtseva et al. 2001) or temperature (Ushakova2003). Many intertidal organisms circumvent the negativeeVects of environmental stressors by releasing gametes orlarvae directly into the water column or brooding their
Communicated by J.P. Grassle.
R. Przeslawski (&) · A. R. DavisInstitute for Conservation Biology, School of Biological Sciences, University of Wollongong, NorthWelds Avenue, Wollongong, NSW 2522, Australiae-mail: [email protected]
Present Address:R. PrzeslawskiDepartment of Ecology and Evolution, Stony Brook University, 650 Life Sciences Building, Stony Brook, NY 11794-5245, USA
123

992 Mar Biol (2007) 152:991–1002
oVspring. However, a considerable number of species, par-ticularly gastropods, enclose their embryos in benthic eggmasses where they remain for at least the early stages oftheir development (Thorson 1950). Whereas adults are ableto seek shelter to avoid stress associated with low tides,embryos within these egg masses cannot.
Encapsulated gastropod embryos rely on a suite ofmechanisms for protection against potentially harmfulenvironmental factors. Encapsulating structures themselvesmay reduce desiccation (Pechenik et al. 2003) and the rateof salinity change (Eyster 1986; Woods and DeSilets1997). Certain compounds may also guard against environ-mental stress; heat shock proteins protect embryos of thecephalaspid Melanochlamys diomedea from thermal stress(Podolsky and HoVmann 1998), and chemical sunscreenshave been identiWed in egg masses from a range of gastro-pods (Carefoot et al. 1998; Przeslawski et al. 2005a; but seeWraith et al. 2006). These mechanisms may be particularlyimportant for species that deposit egg masses on the uppersurfaces of rock platforms where they are exposed tonumerous stressors at low tide.
Spawning behavior may also minimize exposure ofencapsulated oVspring to potentially harmful environmen-tal factors. For example, a high proportion of marine gas-tropods in southeastern Australia deposit egg massesbeneath boulders in shallow pools where they are shieldedfrom UVR, desiccation, and other environmental extremes(BenkendorV and Davis 2004). Reducing reproductive out-put during episodes of heightened stress may also miminizerisks to oVspring. In addition to avoidance of environmentalstress, the underlying mechanisms driving the evolution ofegg mass deposition may also be the result of larval foodavailability (Ventura et al. 1997), predator avoidance(Spight 1977), adult physiology (Underwood 1979), or theoptimization of development rate (Hoegh-Guldberg andPearse 1995). These factors may vary at several temporaland spatial scales. For example, spawning patterns can varyamong seasons (Mileikovsky 1970), tidal cycles (Creese1980), or days (Mileikovsky 1970). Similarly, spawningpatterns can vary across latitudes (Collin 2003), regionssuch as headlands (BenkendorV and Davis 2004), habitatssuch as rock platforms, seagrass beds, or mudXats (Ben-kendorV and Davis 2004), and microhabitats such as cre-vices or puddles (Ocana and Emson 1999). It is presentlyunknown if embryonic exposure to environmental stress isavoided through the modiWcation of egg deposition behav-ior. This study focuses on the avoidance of environmentalstressors at the scales of season and habitat.
Egg mass size may also provide a degree of protection.IntraspeciWc variation in egg mass size is frequently appar-ent (authors’ personal observation), with smaller eggmasses containing fewer embryos (Hoagland 1986; Kasu-gai and Ikeda 2003). Thus, depositing smaller and fewer
egg masses during stressful periods may represent a bet-hedging strategy, reducing the number of encapsulatedembryos exposed to potentially harmful conditions. On theother hand, the lower surface to volume ratio of larger eggmasses may protect embryos against desiccation and ther-mal stress better than smaller egg masses.
Our overall aim was to determine if intertidal gastropodsreduce exposure to environmental stress for their encapsu-lated oVspring, an important consideration in the face of cli-mate change. We conducted surveys of intertidal gastropodegg masses, including both capsular and gelatinous forms(see Przeslawski 2004 for deWnitions), at two sites in south-eastern Australia over 2 years to determine whether egg-laying adults modify their use of spawning habitats inresponse to environmental stressors. We approached thisobjective in several ways; Wrst, we used data from a varietyof sources to conWrm that abiotic conditions vary annuallyin the degree to which they expose intertidal organisms tostressors. Second, we determined whether temporal pat-terns in the abundance, size, and species composition of theentire suite of egg masses was correlated to environmentalstress. Finally, we examined the relationship betweenreproductive output (egg mass abundance) and environ-mental conditions for targeted species. We predicted astrong negative correlation between abiotic stressors (expo-sure to UVR, temperature, and duration of aerial exposure)and the spawning frequency and size of egg masses forthose species laying on rock platforms. We anticipated aweaker correlation or no relationship between these envi-ronmental variables and the spawning of those species lay-ing egg masses beneath boulders.
Materials and methods
Rocky intertidal surveys were conducted at Bass Point(34°35�45�S, 150°53�20�E) and Bellambi (34°37�08�S,150°92�03�E) in southeastern Australia. These sites werechosen because they encompass both rock platforms andboulder Welds, have a high molluscan species diversity (Ben-kendorV and Davis 2002), and numerous egg masses havebeen found consistently at them (authors’ personal observa-tion). Hence these sites were not drawn from a large set ofpotential study locations; as such, ‘site’ was treated as a Wxedfactor in subsequent analyses. Egg masses at each site wererecorded Wve times during each austral season for 2 years(June 2002–May 2004) (Spring: September–November;Summer: December–February; Autumn: March–May; Win-ter: June–August). Visits to each site were chosen haphaz-ardly from days with low tides during daylight. Visits to asite were separated by at least a week so as to minimizerepeat counting of the same egg masses and to reduce distur-bance, particularly for egg masses beneath boulders.
123

Mar Biol (2007) 152:991–1002 993
Three surveys were undertaken at each site on each visit.Counts within quadrats of two sizes were made on the sur-face of rock platforms at haphazard positions along a 100-mtransect line running parallel to the shore along the mid-intertidal zone, approximately 200–800 mm above chartdatum, where egg masses were known to occur. The numberand species identiWcation of all egg masses (except Neritamorio) were recorded in a 50 £ 50 cm quadrat (n = 25). Asegg masses of N. morio were so numerous, the egg capsulesof this species were quantiWed within 25 £ 25 cm quadrats(n = 25) and positioned haphazardly on the same transect.The third survey involved examination of the undersides ofhaphazardly selected boulders (n = 25) in the mid-intertidalzone in which all egg masses were identiWed and recorded.Boulder volume was estimated by measuring maximumwidth, length and height of a boulder and averaged12,600 § 1,100 cm3 (mean § SEM, n = 75); boulder vol-ume did not vary signiWcantly between sites or times. Anegg mass was deWned as a ribbon, group of capsules, orgelatinous structure deposited by a single adult; this wasdetermined by the orientation of the encapsulating structure,similar developmental stage and/or the presence of a con-necting basal membrane. For surveys of N. morio, individ-ual egg capsules were counted, as it was not possible todetermine which groups of capsules a particular adult haddeposited. In addition, sizes of a random sample of eggmasses were recorded for Siphonaria denticulata, Bembi-cium nanum, Dolabrifera brazieri, and Mitra carbonaria, asthey were suYciently abundant. For gelatinous egg masses,length and width were recorded; and for capsular eggmasses, numbers of capsules were counted. We presentthese data as mean number of egg masses per quadrat oneach day of sampling in each season in each year. We alsoexamined the response of the entire egg mass assemblage toabiotic stressors with multiple regressions; the nature ofthese stressors is outlined below.
Egg masses were identiWed from previous studies (Rose1985a; Smith et al. 1989; Rudman 2007) or by identiWca-tion of adults spawning in situ (Table 1). Certain speciesdeposit egg masses that are indistinguishable in the Weldfrom other species (e.g., Dendrodoris fumata and D. nigra,Agnewia tritoniformis and Cronia contracta), and we werethus unable to identify these to species level (Table 1). Wealso recorded the tidal distribution of species from theavailable literature (Edgar 2001; Beesley et al. 1998; Rud-man 2007) in order to determine true intertidal specialistsand those species, which may live and also spawn subti-dally (Table 1).
Temporal variation in potential environmental stressorswas explored with ANOVAs. Air temperature (°C recordedfrom University of Wollongong station at 34°24�S,150°54�E), water temperature (°C recorded from Port Kem-bla at 34°30�S, 150°54�E) and tidal data (mm above or
below Chart Datum recorded for Port Kembla) wereacquired from the Bureau of Meteorology and the NationalTidal Facility for all days on which surveys were con-ducted; UVR data were obtained as monthly averagesof standard erythemal doses (SED) in Sydney (33°54�S,151°12�E) from the Australian Radiation Protection andNuclear Safety Agency. All variables were recorded at sta-tions within 25 km of both Weld sites, except for UVR data,which originated at a recording station approximately40 km north of Bellambi but was deemed comparable toconditions at our Weld sites because monthly indices areused to represent large-scale regional measurements. Assampling within seasons was haphazard, we elected to ana-lyze the factor time with eight levels, encompassing fourseasons over two years. Variances for low tides and temper-atures were heterogeneous (Cochran’s C test), and thesedata were log transformed to comply with assumptions ofANOVA. Tukey’s HSD tests were used to identify signiW-cant relationships a posteriori.
We also examined spatial and temporal variation inthe species composition of egg masses between sites,years, and seasons using non-metric multidimensionalscaling plots and permutational MANOVAs (Anderson2001). The relationships between environmental stres-sors and egg mass abundances on rock platforms andbeneath boulders were determined with multiple regres-sions on total abundance. Multiple regressions were alsoused to determine the relationship between egg mass sizeand stressors.
We also examined relationships between environmentalstressors and the abundances of egg masses for target spe-cies. Target species were deWned as those for which morethan Wve egg masses were found on more than Wve occa-sions or those species which contributed >1% of dissimilar-ity in egg mass composition between sites or seasons asdetermined by the SIMPER function in PRIMER v.5. Eightspecies fulWlled these criteria: B. nanum, N. morio, S. den-ticulata, and S. zelandica which spawn on rock platformsurfaces; and A. tritoniformis/C. contracta, C. eburnea, D.brazieri, and M. carbonaria which spawn beneath boulders(authors’ personal observation). For the multiple regres-sions, we used four quantitative abiotic variables; air tem-perature, water temperature, tidal height and UV radiation,as well as a qualitative variable, the phase of the moon (full,new and other), obtained from online database of theAustralian government department Geoscience Australia(http://www.ga.gov.au). This Wnal variable was incorporatedinto the analysis as an indicator variable (Zar 1998). Wefollowed the procedures outlined in Chatterjee and Price(2000), particularly the examination of residual plots, toassess the Wt of our data to the linear models. We detected asigniWcant outlier in the S. zelandica data set, which dispro-portionately modiWed the outcome of the analysis; accordingly,
123
994 Mar Biol (2007) 152:991–1002
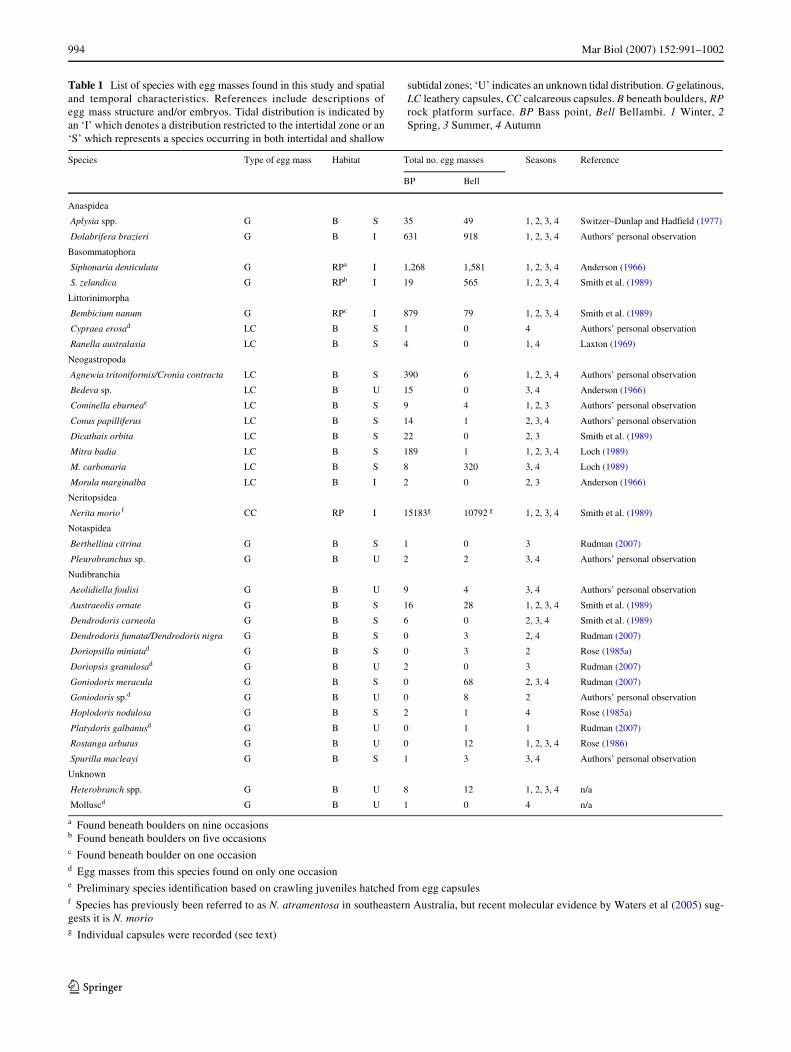
Table 1 List of species with egg masses found in this study and spatialand temporal characteristics. References include descriptions ofegg mass structure and/or embryos. Tidal distribution is indicated byan ‘I’ which denotes a distribution restricted to the intertidal zone or an‘S’ which represents a species occurring in both intertidal and shallow
subtidal zones; ‘U’ indicates an unknown tidal distribution. G gelatinous,LC leathery capsules, CC calcareous capsules. B beneath boulders, RProck platform surface. BP Bass point, Bell Bellambi. 1 Winter, 2Spring, 3 Summer, 4 Autumn
Species Type of egg mass Habitat Total no. egg masses Seasons Reference
BP Bell
Anaspidea
Aplysia spp. G B S 35 49 1, 2, 3, 4 Switzer–Dunlap and HadWeld (1977)
Dolabrifera brazieri G B I 631 918 1, 2, 3, 4 Authors’ personal observation
Basommatophora
Siphonaria denticulata G RPa I 1,268 1,581 1, 2, 3, 4 Anderson (1966)
S. zelandica G RPb I 19 565 1, 2, 3, 4 Smith et al. (1989)
Littorinimorpha
Bembicium nanum G RPc I 879 79 1, 2, 3, 4 Smith et al. (1989)
Cypraea erosad LC B S 1 0 4 Authors’ personal observation
Ranella australasia LC B S 4 0 1, 4 Laxton (1969)
Neogastropoda
Agnewia tritoniformis/Cronia contracta LC B S 390 6 1, 2, 3, 4 Authors’ personal observation
Bedeva sp. LC B U 15 0 3, 4 Anderson (1966)
Cominella eburneae LC B S 9 4 1, 2, 3 Authors’ personal observation
Conus papilliferus LC B S 14 1 2, 3, 4 Authors’ personal observation
Dicathais orbita LC B S 22 0 2, 3 Smith et al. (1989)
Mitra badia LC B S 189 1 1, 2, 3, 4 Loch (1989)
M. carbonaria LC B S 8 320 3, 4 Loch (1989)
Morula marginalba LC B I 2 0 2, 3 Anderson (1966)
Neritopsidea
Nerita morio f CC RP I 15183g 10792 g 1, 2, 3, 4 Smith et al. (1989)
Notaspidea
Berthellina citrina G B S 1 0 3 Rudman (2007)
Pleurobranchus sp. G B U 2 2 3, 4 Authors’ personal observation
Nudibranchia
Aeolidiella foulisi G B U 9 4 3, 4 Authors’ personal observation
Austraeolis ornate G B S 16 28 1, 2, 3, 4 Smith et al. (1989)
Dendrodoris carneola G B S 6 0 2, 3, 4 Smith et al. (1989)
Dendrodoris fumata/Dendrodoris nigra G B S 0 3 2, 4 Rudman (2007)
Doriopsilla miniatad G B S 0 3 2 Rose (1985a)
Doriopsis granulosad G B U 2 0 3 Rudman (2007)
Goniodoris meracula G B S 0 68 2, 3, 4 Rudman (2007)
Goniodoris sp.d G B U 0 8 2 Authors’ personal observation
Hoplodoris nodulosa G B S 2 1 4 Rose (1985a)
Platydoris galbanusd G B U 0 1 1 Rudman (2007)
Rostanga arbutus G B U 0 12 1, 2, 3, 4 Rose (1986)
Spurilla macleayi G B S 1 3 3, 4 Authors’ personal observation
Unknown
Heterobranch spp. G B U 8 12 1, 2, 3, 4 n/a
Molluscd G B U 1 0 4 n/a
a Found beneath boulders on nine occasionsb Found beneath boulders on Wve occasions c Found beneath boulder on one occasiond Egg masses from this species found on only one occasione Preliminary species identiWcation based on crawling juveniles hatched from egg capsulesf Species has previously been referred to as N. atramentosa in southeastern Australia, but recent molecular evidence by Waters et al (2005) sug-gests it is N. moriog Individual capsules were recorded (see text)
123
Mar Biol (2007) 152:991–1002 995
we removed it for the analyses presented here (Chatterjeeand Price 2000). Where data were heteroscedastic, we usedthe square root transformation, which has the advantage ofnormalizing data as well as improving heterogeneousvariances (Chatterjee and Price 2000). We calculated theDurbin Watson statistic (d) to assess for correlated errorsand examined estimated coeYcients for large changeswhen variables were added or deleted as evidence of multi-colinearity. In seeking the best explanatory model, weremoved terms from the regression model, paying closeattention to the adjusted r2 and the root error mean square(Chatterjee and Price 2000). We used JMP v.4 for ANOVAand multiple regression analyses, and PERMANOVA v.6(Anderson 2001) for permutational MANOVA tests.PRIMER v.5 was used for all other multivariate analyses.� = 0.05 in all statistical tests.
Results
We recorded egg masses from 34 gastropod species or taxo-nomic groups (Table 1). Only four species spawned on rockplatforms in full sun (B. nanum, N. morio, S. denticulata andS. zelandica). All of these egg masses were gelatinous,except for N. morio, which deposits calcareous capsules(Przeslawski 2004); and all had distributions restricted to theintertidal zone (Table 1). Egg masses of all other speciesexamined in this study were found exclusively beneath boul-ders, encompassing both capsular and gelatinous egg massesand both categories of tidal distribution (Table 1). Only B.nanum and S. denticulata had egg masses found in both habi-tats (Table 1), and then it was a very rare occurrence. Noneof the species laying egg masses under full sun conditionsshifted their spawning habitat to more protected microhabi-tats beneath boulders as abiotic conditions became morestressful. More species spawned in summer and autumn (24
species) than spring or winter (20 and 14 species, respec-tively). Egg masses from all four species spawning on rockplatforms were found in all seasons (Table 1), althoughabundances were lowest in winter, as detailed below.
Seasonal patterns in environmental stressors
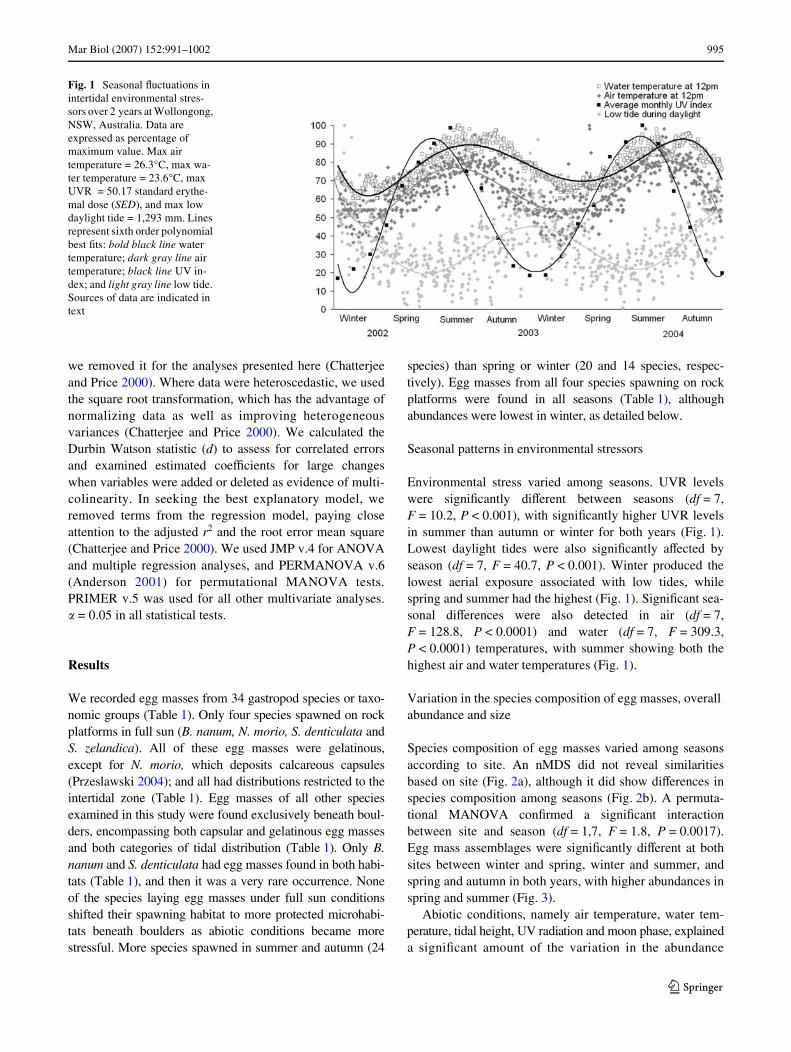
Environmental stress varied among seasons. UVR levelswere signiWcantly diVerent between seasons (df = 7,F = 10.2, P < 0.001), with signiWcantly higher UVR levelsin summer than autumn or winter for both years (Fig. 1).Lowest daylight tides were also signiWcantly aVected byseason (df = 7, F = 40.7, P < 0.001). Winter produced thelowest aerial exposure associated with low tides, whilespring and summer had the highest (Fig. 1). SigniWcant sea-sonal diVerences were also detected in air (df = 7,F = 128.8, P < 0.0001) and water (df = 7, F = 309.3,P < 0.0001) temperatures, with summer showing both thehighest air and water temperatures (Fig. 1).
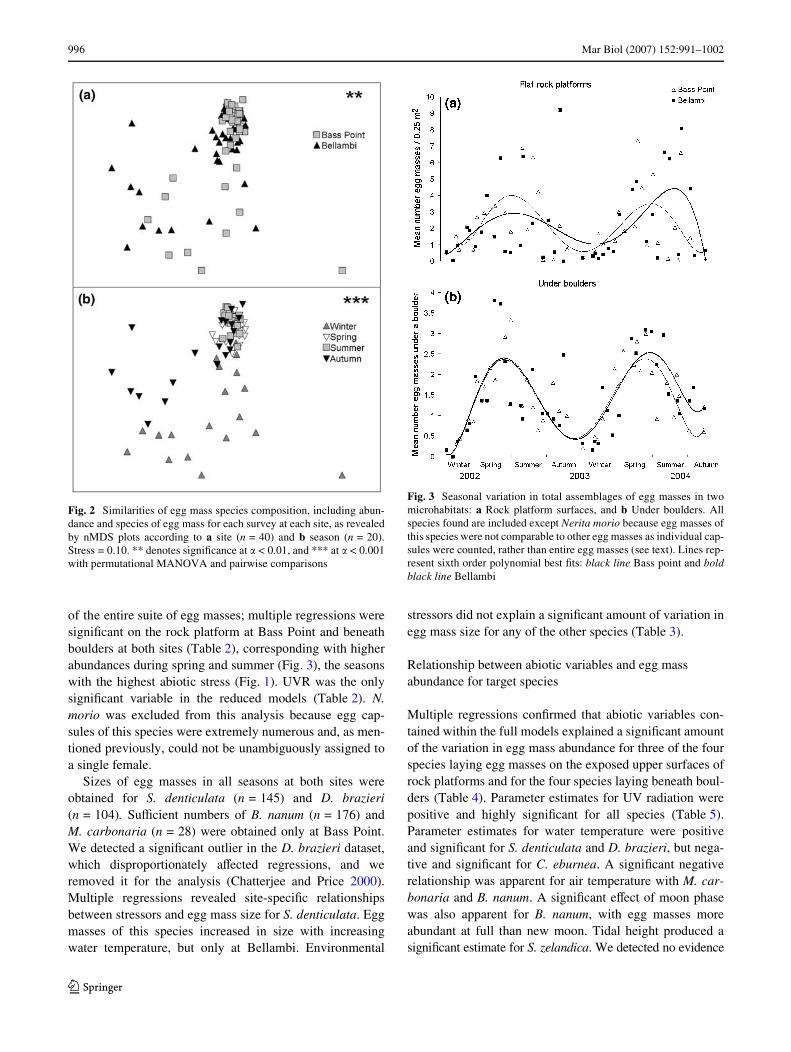
Variation in the species composition of egg masses, overall abundance and size
Species composition of egg masses varied among seasonsaccording to site. An nMDS did not reveal similaritiesbased on site (Fig. 2a), although it did show diVerences inspecies composition among seasons (Fig. 2b). A permuta-tional MANOVA conWrmed a signiWcant interactionbetween site and season (df = 1,7, F = 1.8, P = 0.0017).Egg mass assemblages were signiWcantly diVerent at bothsites between winter and spring, winter and summer, andspring and autumn in both years, with higher abundances inspring and summer (Fig. 3).
Abiotic conditions, namely air temperature, water tem-perature, tidal height, UV radiation and moon phase, explaineda signiWcant amount of the variation in the abundance
Fig. 1 Seasonal Xuctuations in intertidal environmental stres-sors over 2 years at Wollongong, NSW, Australia. Data are expressed as percentage of maximum value. Max air temperature = 26.3°C, max wa-ter temperature = 23.6°C, max UVR = 50.17 standard erythe-mal dose (SED), and max low daylight tide = 1,293 mm. Lines represent sixth order polynomial best Wts: bold black line water temperature; dark gray line air temperature; black line UV in-dex; and light gray line low tide. Sources of data are indicated in text
123
996 Mar Biol (2007) 152:991–1002
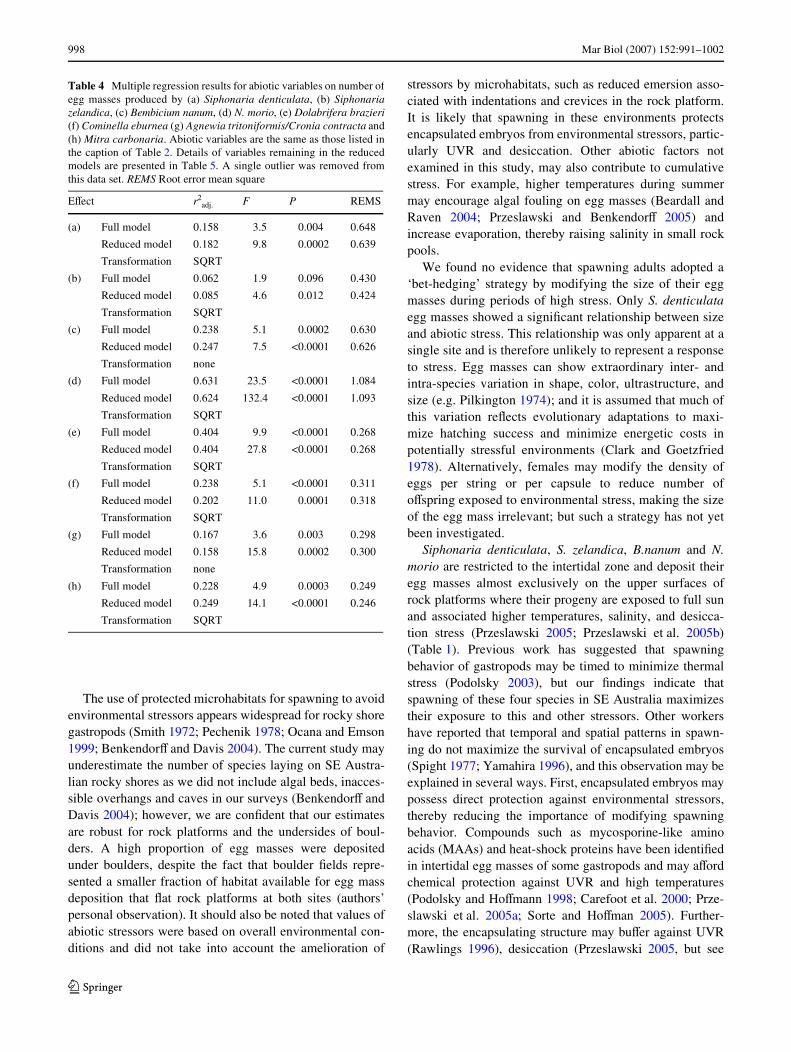
of the entire suite of egg masses; multiple regressions weresigniWcant on the rock platform at Bass Point and beneathboulders at both sites (Table 2), corresponding with higherabundances during spring and summer (Fig. 3), the seasonswith the highest abiotic stress (Fig. 1). UVR was the onlysigniWcant variable in the reduced models (Table 2). N.morio was excluded from this analysis because egg cap-sules of this species were extremely numerous and, as men-tioned previously, could not be unambiguously assigned toa single female.
Sizes of egg masses in all seasons at both sites wereobtained for S. denticulata (n = 145) and D. brazieri(n = 104). SuYcient numbers of B. nanum (n = 176) andM. carbonaria (n = 28) were obtained only at Bass Point.We detected a signiWcant outlier in the D. brazieri dataset,which disproportionately aVected regressions, and weremoved it for the analysis (Chatterjee and Price 2000).Multiple regressions revealed site-speciWc relationshipsbetween stressors and egg mass size for S. denticulata. Eggmasses of this species increased in size with increasingwater temperature, but only at Bellambi. Environmental
stressors did not explain a signiWcant amount of variation inegg mass size for any of the other species (Table 3).
Relationship between abiotic variables and egg mass abundance for target species
Multiple regressions conWrmed that abiotic variables con-tained within the full models explained a signiWcant amountof the variation in egg mass abundance for three of the fourspecies laying egg masses on the exposed upper surfaces ofrock platforms and for the four species laying beneath boul-ders (Table 4). Parameter estimates for UV radiation werepositive and highly signiWcant for all species (Table 5).Parameter estimates for water temperature were positiveand signiWcant for S. denticulata and D. brazieri, but nega-tive and signiWcant for C. eburnea. A signiWcant negativerelationship was apparent for air temperature with M. car-bonaria and B. nanum. A signiWcant eVect of moon phasewas also apparent for B. nanum, with egg masses moreabundant at full than new moon. Tidal height produced asigniWcant estimate for S. zelandica. We detected no evidence
Fig. 2 Similarities of egg mass species composition, including abun-dance and species of egg mass for each survey at each site, as revealedby nMDS plots according to a site (n = 40) and b season (n = 20).Stress = 0.10. ** denotes signiWcance at � < 0.01, and *** at � < 0.001with permutational MANOVA and pairwise comparisons
Fig. 3 Seasonal variation in total assemblages of egg masses in twomicrohabitats: a Rock platform surfaces, and b Under boulders. Allspecies found are included except Nerita morio because egg masses ofthis species were not comparable to other egg masses as individual cap-sules were counted, rather than entire egg masses (see text). Lines rep-resent sixth order polynomial best Wts: black line Bass point and boldblack line Bellambi
123

Mar Biol (2007) 152:991–1002 997
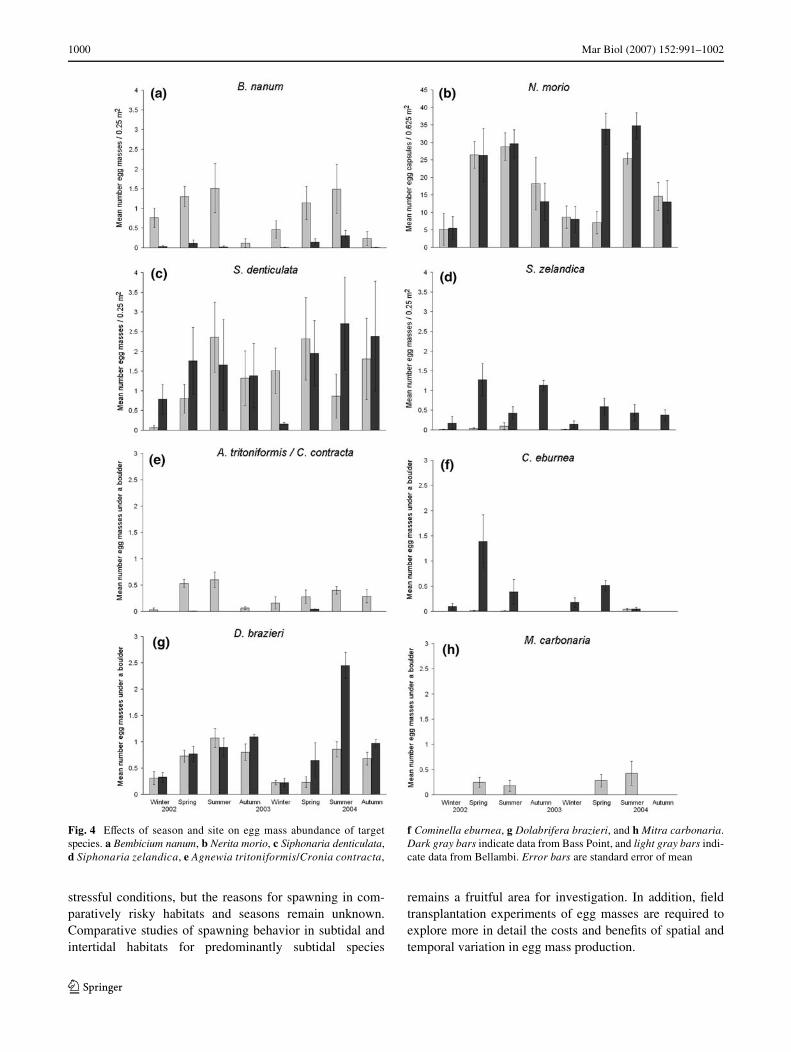
of multicollinearity in the data sets; and autocorrelation, asrevealed by the Durbin-Watson statistic, was marginallysigniWcant for two of the four analyses on rock platforms,but highly signiWcant for three species beneath boulders.These results are supported by the fact that egg massesof most target species were found most frequently in springand summer (Fig. 4), corresponding with stressful seasons(Fig. 1).
Discussion
Our Wndings conWrm that the majority of southeastern Aus-tralian rocky shore gastropods laid egg masses beneathboulders where encapsulated embryos are sheltered fromextreme environmental conditions, and most of these egg
masses are produced in spring and summer. Our predictionthat those species depositing egg masses on the surface ofrock platforms would modify their spawning behavior inresponse to abiotic stressors was not supported. These spe-cies continued to lay egg masses in full sun and had higheregg mass abundances in spring and summer, coincidingwith the most physiologically stressful period. Clearly thespawning peak for these species coincides with periods ofhigh environmental stress, and Weld observations on B.nanum and S. denticulata show that rock platform surfacesare not optimal habitats for developing embryos (Przeslaw-ski 2005; Przeslawski et al. 2005b). In addition, egg-masseswill be synchronously exposed to physiological extremesfrom a multitude of stressors. Interactions among stressorscan further elevate mortality in early life history stages(HoVman et al. 2003; Przeslawski et al. 2005b).
Table 3 Multiple regression results for abiotic variables on size of egg masses of (a) B. nanum, (b) Siphonaria denticulata, (c) Dolabriferabrazieri, and (d) Mitra carbonaria. Abiotic variables are the same as those listed in the caption of Table 2
n/a not applicablea SigniWcant parameter included in reduced model was water temperatureb A single outlier was removed from this data set (see text)
EVect Bass point Bellambi
r2adj. F P REMS r2
adj. F P REMS
(a) Full model 0.0121 1.5 0.1945 0.6522 n/a
Transformation SQRT
(b) Full model 0.0284 2.0 0.0993 0.9179 0.0563 3.1 0.0163 1.0009
Reduced modela n/a 0.0564 9.6 0.0023 1.0008
Transformation None None
(c) Full model 0.0253 1.7 0.1632 7.4614 0.0500 2.3 0.0602 5.6184
Transformation None None
(d) Full model 0.0228 1.2 0.3553 2.4642 n/a
Transformation SQRT
Table 2 Multiple regression results for abiotic variables on meannumber of all egg masses per survey on (a) rock platform surfaces(Excluding N. morio as egg capsules from this species could not beunambiguously assigned to an individual adult (see text for further de-tails)) and (b) undersides of boulders. Abiotic variables included four
quantitative variables; air temperature, water temperature, tidal heightand UV radiation, as well as a qualitative variable, the phase of the moon(full, new and other). Reduced models include signiWcant parametersfrom whole-model multiple regression; this was only UVR in all caseswhere full model was signiWcant. REMS, Root error mean square
EVect Bass point Bellambi
r2adj. F P REMS r2
adj. F P REMS
(a) Full model 0.2260 3.8 0.0108 1.7915 0.0198 1.2 0.3295 2.4629
Reduced model 0.2746 15.8 0.0003 1.7344 n/a
Transformation None None
(b) Full model 0.5143 11.3 <0.0001 0.2941 0.4456 8.8 <0.0001 0.3066
Reduced model 0.5106 41.6 <0.0001 0.2952 0.4596 34.2 <0.0001 0.3028
Transformation SQRT SQRT
123
998 Mar Biol (2007) 152:991–1002
The use of protected microhabitats for spawning to avoidenvironmental stressors appears widespread for rocky shoregastropods (Smith 1972; Pechenik 1978; Ocana and Emson1999; BenkendorV and Davis 2004). The current study mayunderestimate the number of species laying on SE Austra-lian rocky shores as we did not include algal beds, inacces-sible overhangs and caves in our surveys (BenkendorV andDavis 2004); however, we are conWdent that our estimatesare robust for rock platforms and the undersides of boul-ders. A high proportion of egg masses were depositedunder boulders, despite the fact that boulder Welds repre-sented a smaller fraction of habitat available for egg massdeposition that Xat rock platforms at both sites (authors’personal observation). It should also be noted that values ofabiotic stressors were based on overall environmental con-ditions and did not take into account the amelioration of
stressors by microhabitats, such as reduced emersion asso-ciated with indentations and crevices in the rock platform.It is likely that spawning in these environments protectsencapsulated embryos from environmental stressors, partic-ularly UVR and desiccation. Other abiotic factors notexamined in this study, may also contribute to cumulativestress. For example, higher temperatures during summermay encourage algal fouling on egg masses (Beardall andRaven 2004; Przeslawski and BenkendorV 2005) andincrease evaporation, thereby raising salinity in small rockpools.
We found no evidence that spawning adults adopted a‘bet-hedging’ strategy by modifying the size of their eggmasses during periods of high stress. Only S. denticulataegg masses showed a signiWcant relationship between sizeand abiotic stress. This relationship was only apparent at asingle site and is therefore unlikely to represent a responseto stress. Egg masses can show extraordinary inter- andintra-species variation in shape, color, ultrastructure, andsize (e.g. Pilkington 1974); and it is assumed that much ofthis variation reXects evolutionary adaptations to maxi-mize hatching success and minimize energetic costs inpotentially stressful environments (Clark and Goetzfried1978). Alternatively, females may modify the density ofeggs per string or per capsule to reduce number ofoVspring exposed to environmental stress, making the sizeof the egg mass irrelevant; but such a strategy has not yetbeen investigated.
Siphonaria denticulata, S. zelandica, B.nanum and N.morio are restricted to the intertidal zone and deposit theiregg masses almost exclusively on the upper surfaces ofrock platforms where their progeny are exposed to full sunand associated higher temperatures, salinity, and desicca-tion stress (Przeslawski 2005; Przeslawski et al. 2005b)(Table 1). Previous work has suggested that spawningbehavior of gastropods may be timed to minimize thermalstress (Podolsky 2003), but our Wndings indicate thatspawning of these four species in SE Australia maximizestheir exposure to this and other stressors. Other workershave reported that temporal and spatial patterns in spawn-ing do not maximize the survival of encapsulated embryos(Spight 1977; Yamahira 1996), and this observation may beexplained in several ways. First, encapsulated embryos maypossess direct protection against environmental stressors,thereby reducing the importance of modifying spawningbehavior. Compounds such as mycosporine-like aminoacids (MAAs) and heat-shock proteins have been identiWedin intertidal egg masses of some gastropods and may aVordchemical protection against UVR and high temperatures(Podolsky and HoVmann 1998; Carefoot et al. 2000; Prze-slawski et al. 2005a; Sorte and HoVman 2005). Further-more, the encapsulating structure may buVer against UVR(Rawlings 1996), desiccation (Przeslawski 2005, but see
Table 4 Multiple regression results for abiotic variables on number ofegg masses produced by (a) Siphonaria denticulata, (b) Siphonariazelandica, (c) Bembicium nanum, (d) N. morio, (e) Dolabrifera brazieri(f) Cominella eburnea (g) Agnewia tritoniformis/Cronia contracta and(h) Mitra carbonaria. Abiotic variables are the same as those listed inthe caption of Table 2. Details of variables remaining in the reducedmodels are presented in Table 5. A single outlier was removed fromthis data set. REMS Root error mean square
EVect r2adj. F P REMS
(a) Full model 0.158 3.5 0.004 0.648
Reduced model 0.182 9.8 0.0002 0.639
Transformation SQRT
(b) Full model 0.062 1.9 0.096 0.430
Reduced model 0.085 4.6 0.012 0.424
Transformation SQRT
(c) Full model 0.238 5.1 0.0002 0.630
Reduced model 0.247 7.5 <0.0001 0.626
Transformation none
(d) Full model 0.631 23.5 <0.0001 1.084
Reduced model 0.624 132.4 <0.0001 1.093
Transformation SQRT
(e) Full model 0.404 9.9 <0.0001 0.268
Reduced model 0.404 27.8 <0.0001 0.268
Transformation SQRT
(f) Full model 0.238 5.1 <0.0001 0.311
Reduced model 0.202 11.0 0.0001 0.318
Transformation SQRT
(g) Full model 0.167 3.6 0.003 0.298
Reduced model 0.158 15.8 0.0002 0.300
Transformation none
(h) Full model 0.228 4.9 0.0003 0.249
Reduced model 0.249 14.1 <0.0001 0.246
Transformation SQRT
123

Mar Biol (2007) 152:991–1002 999
Rawlings 1999), and changes in salinity and temperature(Pechenik 1979; Pechenik 1982; Woods and DeSilets1997). Second, spawning behavior may simply reXect adultreproductive physiology. Spawning of many gastropods isthermally regulated (Underwood 1979), and it may be thatreproductive output is generally higher in warmer seasonsdespite increased potential for embryonic exposure to envi-ronmental stress. Third, seasonal variation in egg massabundance and size may reXect conditions that maximizedevelopmental rate. Highest temperatures occur in summermonths, and rock pools in which S. denticulata, B. nanumand N. morio spawn can reach 35°C in summer during lowtides (authors’ pers. obs.). Developmental rate is directlyrelated to temperature up to a threshold (reviewed byPalmer 1994). Rapid development reduces the time spent invulnerable life stages (Havenhand 1993) and may be of par-ticular importance to species that complete all or most ofpre-metamorphic development encapsulated. However, riskof mortality upon leaving the capsule may also be very high(Hunt and Scheibling 1997), and the four species hererelease pelagic larvae (BenkendorV and Davis 2004 andreferences therein). A more plausible beneWt of a fasterdevelopment rate may be to hasten the development of pro-tective mechanisms (Podolsky 2003), the beneWts of which
may counteract the negative eVects of abiotic stressors dur-ing summer months. Fourth, the timing and placement ofegg masses may reXect predation pressure, which may behigher in areas sheltered from stressors such as the under-sides of boulders. By spawning on rock platforms, femalesmay reduce predation risk for their oVspring (Spight 1977;Ocana and Emson 1999). This may be particularly impor-tant during warmer seasons, as benthic invertebrate preda-tors increase their feeding rate during warmer months(Sanford 2002). Fifth, spawning behavior may reXect opti-mal conditions for later larval stages, rather than encapsu-lated embryos. Summer coincides with the peak period ofphytoplankton abundance (Underwood 1979), and optimalconditions for larval recruitment and/or food availabilitymay thus outweigh risks associated with spawning in springor summer. A Wnal alternative is that egg masses in theintertidal zone represent only a small proportion of the totalreproductive output of many predominantly subtidal spe-cies. This is demonstrably incorrect for the four species lay-ing predominantly in full sun environments and for severalother species listed in Table 1; they are all restricted to theintertidal zone. In conclusion, those species laying onexposed surfaces do not reduce their reproductive outputnor change their spawning habitat under physiologically
Table 5 Parameter estimates (§ standard errors) and their signiWcance for abiotic variables retained in the reduced multiple regression models for egg mass-es of rocky shore gastropods. Phase of the moon (full, new and other) was represented by two indicator variables (IV). Evi-dence for autocorrelation was tested with the Durbin–Watson statistic, d
Parameter estimates for retained variables
Rock platform species
B. nanum N. morio S. denticulata S. zelandica
Intercept 1.060(0.420)* 0.095(0.289) ¡0.829(0.570) ¡0.194(0.174)
IVmoon1 ¡0.664(0.235)**
IVmoon2 ¡0.455(0.188)*
UV radiation 0.013(0.003)*** 0.054(0.004)*** 0.009(0.003)*** 0.004(0.002)*
Air temperature ¡0.052(0.025)*
Water temperature 0.067(0.031)*
Low tide 0.001(0.0003)**
Durbin–Watson d 1.59 n/a 2.12 1.59
P value 0.024 0.640 0.027
Under boulder species
A. tritoniformis D. brazieri M. carbonaria C. eburnea
C. contracta
Intercept ¡0.088(0.079) ¡0.785(0.239)** 0.214(0.150) 0.762(0.283)**
IVmoon1
IVmoon2
UV radiation 0.005(0.001)*** 0.005(0.001)*** 0.006(0.001)*** 0.005(0.001)***
Air temperature ¡0.026(0.010)**
Water temperature 0.071(0.012)*** ¡0.047(0.015)**
Low tide
Durbin–Watson d n/a 1.40 1.03 1.23
P value 0.0017 <0.0001 0.0001
n/a not applicable
*P < 0.05; **P < 0.01; ***P < 0.001
123
1000 Mar Biol (2007) 152:991–1002
stressful conditions, but the reasons for spawning in com-paratively risky habitats and seasons remain unknown.Comparative studies of spawning behavior in subtidal andintertidal habitats for predominantly subtidal species
remains a fruitful area for investigation. In addition, Weldtransplantation experiments of egg masses are required toexplore more in detail the costs and beneWts of spatial andtemporal variation in egg mass production.
Fig. 4 EVects of season and site on egg mass abundance of targetspecies. a Bembicium nanum, b Nerita morio, c Siphonaria denticulata,d Siphonaria zelandica, e Agnewia tritoniformis/Cronia contracta,
f Cominella eburnea, g Dolabrifera brazieri, and h Mitra carbonaria.Dark gray bars indicate data from Bass Point, and light gray bars indi-cate data from Bellambi. Error bars are standard error of mean
123

Mar Biol (2007) 152:991–1002 1001
Acknowledgments The authors are grateful to K. BenkendorV,B. Rudman , P. Middlefart, and D. Beechey for assistance in speciesidentiWcation. M. Byrne and several anonymous reviewers provided valu-able comments on this manuscript. This represents contribution 279from the Ecology and Genetics group at the University of Wollongong.
References
Adams NL, Shick JM (2001) Mycosporine-like amino acids preventUVB-induced abnormalities during early development of thegreen sea urchin Strongylocentrotus droebachiensis. Mar Biol138:267–280
Anderson DT (1966) Further observations on the life histories of lit-toral gastropods in New South Wales. Proc Lin Soc NSW 90:242–251
Anderson MJ (2001) A new method for non-parametric multivariateanalysis of variance. Aust Ecol 26:32–46
Beardall J, Raven JA (2004) The potential eVects of global climatechange on microalgal photosynthesis, growth and ecology. Phyc-ologia 43:26–40
Beesley PL, Ross GLB, Wells A (1998) Mollusca: the southern synthe-sis, Part B. CSIRO Publishing, Melbourne
BenkendorV K, Davis AR (2002) Identifying hotspots of molluscanspecies richness on rocky intertidal reefs. Biodivers Conserv11:1959–1973
BenkendorV K, Davis AR (2004) Gastropod egg mass deposition on atemperate, wave-exposed coastline in New South Wales, Austra-lia: implications for intertidal conservation. Aquat Conserv14:263–280
Biermann C, Schinner G, Strathmann R (1992) InXuence of solarradiation, microalgal fouling, and current on deposition site andsurvival of embryos of a dorid nudibranch gastropod. Mar EcolProg Ser 86:205–215
Carefoot TH, Harris M, Taylor BE, Donovan D, Karentz D (1998) My-cosporine-like amino acids: possible UV protection in eggs of thesea hare Aplysia dactylomela. Mar Biol 130:389–396
Carefoot TH, Karentz D, Pennings S, Young C (2000) Distribution ofmycosporine-like amino acids in the sea hare Aplysia dactylo-mela: eVects of diet on amounts and types sequestered over timein tissues and spawn. Comp Biochem Phys C 126:91–104
Chatterjee S, Price B (2000) Regression analysis by example, 3rd edn.Wiley, New York
Clark K, Goetzfried A (1978) Zoogeographic inXuences on develop-ment patterns of North Atlantic sacoglossa and nudibranchia,with a discussion of factors aVecting egg size and number. J Mol-lus Stud 44:283–294
Collin R (2003) Worldwide patterns in mode of development in Calyp-traied gastropods. Mar Ecol Prog Ser 247:103–122
Creese RG (1980) Reproductive cycles and fecundities of two speciesof Siphonaria (Mollusca: Pulmonata) in South-eastern Australia.Aust J Mar Freshw Res 31:37–47
Edgar GJ (2001) Australian marine life: the plants and animals of tem-perate waters. New Holland Publishers, Chatswood
Eyster L (1986) The embryonic capsules of nudibranch molluscs: lit-erature review and new studies on albumen and capsule wallultrastructure. Am Malacol Bull 4:205–216
Gosselin LA, Chia FS (1995) Characterizing temperate rocky shoresfrom the perspective of an early juvenile snail: the main threatsto survival of newly hatched Nucella emarginata. Mar Biol122:625–635
Havenhand J (1993) Egg to juvenile period, generation time, and theevolution of larval type in marine invertebrates. Mar Ecol ProgSer 97:247–260
Hoagland KE (1986) Patterns of encapsulation and brooding in theCalyptraeidae (Prosobranchia: Mesogastropoda). Am MalacolBull 4:173–183
Hoegh-Guldberg O, Pearse JS (1995) Temperature, food availability,and the development of marine invertebrate larvae. Am Zool35:415–425
HoVman JR, Hansen LJ, Klinger T (2003) Interactions between UVradiation and temperature limit inferences from single-factorexperiments. J Phycol 39:268–272
Hunt HL, Scheibling RE (1997) Role of early post-settlement mortalityin recruitment of benthic marine invertebrates. Mar Ecol Prog Ser155:269–301
Jackson GA, Strathmann RR (1981) Larval mortality from oVshoremixing as a link between precompetent and competent periods ofdevelopment. Am Nat 118:16–26
Kasugai T, Ikeda Y (2003) Description of the egg mass of the pygmycuttleWsh, Idiosepius paradoxus (Cephalopoda: Idiosepiidae),with special reference to its multiple gelatinous layers. Veliger46:105–110
Laxton JH (1969) Reproduction in some New Zealand Cymatiidae(Gastropoda: Prosobranchia). Zool J Linn Soc Lond 48:237–253
Loch I (1989) Some light on dark Mitres. Aust Shell News 65:4–5Mileikovsky SA (1970) Seasonal and daily dynamics of pelagic larvae
of marine shelf bottom invertebrates in nearshore waters of Kan-kalaksha Bay (White Sea). Mar Biol 5:180–194
Moran AL (1999) Size and performance of juvenile marine inverte-brates: potential contrasts between intertidal and subtidal benthichabitats. Am Zool 39:304–312
Newell RC (1970) Biology of intertidal animals. American ElsevierPublishing, New York
Ocana TMJ, Emson RH (1999) Maturation, spawning and develop-ment in Siphonaria pectinata linnaeus (Gastropoda: Pulmonata)at Gibraltar. J Moll Stud 65:185–193
Palmer AR (1994) Temperature sensitivity, rate of development, andtime to maturity: geographic variation in laboratory-reared Nucel-la and a cross-phyletic overview. In: Wilson Jr WH, Stricker SA,Shinn GL (eds) Reproduction and development of marine inver-tebrates. Johns Hopkins University Press, Baltimore, pp 177–194
Pechenik J (1978) Adaptation to intertidal development: studies onNassarius obsoletus. Biol Bull 154:282–291
Pechenik J (1979) Role of encapsulation in invertebrate life histories.Am Nat 114:870
Pechenik J (1982) Ability of some gastropod egg capsules to protectagainst low-salinity stress. J Exp Mar Biol Ecol 3:195–208
Pechenik J, Marsden ID, Pechenik O (2003) EVects of temperature,salinity, and air exposure on development of the estuarine pulmo-nate gastropod Amphibola crenata. J Exp Mar Biol Ecol 292:159–176
Pilkington M (1974) The eggs and hatching of some New ZealandProsobranch molluscs. J R Soc N Z 4:411–431
Podolsky RD (2003) Integrating development and environment tomodel reproductive performance in natural populations of anintertidal gastropod. Integr Comp Biol 43:450–458
Podolsky RD, HoVmann GE (1998) Embryo development during tide-related thermal stress: evidence of a protective role for heat-shockproteins. Am Nat 38:186
Przeslawski R (2004) A review of the eVects of environmental stresson embryonic development within intertidal gastropod egg mass-es. Moll Res 24:43–63
Przeslawski R (2005) Combined eVects of solar radiation and desicca-tion on the mortality and development of encapsulated embryosof rocky shore gastropods. Mar Ecol Prog Ser 298:169–177
Przeslawski R, BenkendorV K (2005) The role of surface fouling in thedevelopment of encapsulated gastropod embryos. J Moll Stud71:75–83
123

1002 Mar Biol (2007) 152:991–1002
Przeslawski R, BenkendorV K, Davis AR (2005a) A quantitative sur-vey of mycosporine-like amino acids (MAAs) in intertidal eggmasses from temperate rocky shores. J Chem Ecol 31:2417–2438
Przeslawski R, Davis AR, BenkendorV K (2005b) Synergies, climatechange and the development of rocky shore invertebrates. GlobChange Biol 11:515–522
Rawlings TA (1996) Shields against ultraviolet radiation: an additionalprotective role for the egg capsules of benthic marine gastropods.Mar Ecol Prog Ser 136:81–95
Rawlings TA (1999) Adaptations to physical stresses in the intertidalzone: the egg capsules of neogastropod molluscs. Am Zool39:240–243
Rose RA (1985a) The spawn and development of 29 NSW Opistho-branchs (Mollusca: Gastropoda). Proc Linn Soc NSW 108:23–36
Rose RA (1985b) The spawn and embryonic development of colourvariants of Dendrodoris nigra Stimpson (Mollusca: Nudibran-chia). J Malacol Soc Aust 7:75–88
Rose RA (1986) Direct development in Rostanga arbutus (Angas)(Mollusca: Nudibranchia) and the eVects of temperature andsalinity on embryos reared in the laboratory. J Malacol Soc Aust7:141–154
Rudman WB (2007) Sea slug forum. Australian museum. http://www.seaslugforum.net (accessed 17 May 2007)
Sanford E (2002) The feeding, growth, and energetics of two rockyintertidal predators (Pisaster ochraceus and Nucella canaliculata)under water temperatures simulating episodic upwelling. J ExpMar Biol Ecol 273:199–218
Smith B, Black JH, Sheperd SA (1989) Molluscan egg masses and cap-sules. In: Shepherd SA, Thomas IM (eds) Marine invertebrates ofsouthern Australia, part 2. Southern Australia Government Print-ing Division, Adelaide, pp 841–891
Smith BJ (1972) Don’t kill the young. Survival 2:86–87Sorte CJB, HoVman GE (2005) Thermotolerance and heat-shock
protein expression in Northeastern PaciWc Nucella species withdiVerent biogeographic ranges. Mar Biol 146:985–993
Spight TM (1977) Do intertidal snails spawn in the right places? Evo-lution 31:682–691
Switzer-Dunlap M, HadWeld MG (1977) Observations on develop-ment, larval growth and metamorphosis of four species ofAplysiidae (Gastropoda: Opisthobranchia) in laboratory culture.J Exp Mar Biol Ecol 29:245–261
Thorson G (1950) Reproductive and larval ecology of marine bottominvertebrates. Biol Rev 25:1–45
Underwood AJ (1979) The ecology of intertidal gastropods. Adv MarBiol 16:111–210
Ushakova O (2003) Combined eVect of salinity and temperature onSpirorbis spirorbis L. and Circeus spirillum L. larvae from theWhite Sea. J Exp Mar Biol Ecol 296:23–33
Ventura CRR, Falcao APC, Santos JS, Fiori CS (1997) Reproductivecycle and feeding periodicity in the starWsh Astropecten brasiliensisin the Cabo Frio upwelling ecosystem (Brazil). Invertebr ReprodDev 31:135–141
Waters JM, King TM, O’Loughlin PM, Spencer HG (2005) Phylogeo-graphical disjunction in abundant high-dispersal littoral gastro-pods. Mol Ecol 14:483–494
Woods HA, DeSilets RJ (1997) Egg-mass gel of Melanochlamys diom-edea (Bergh) protects embryos from low salinity. Biol Bull193:341–349
Wraith J, Przeslawski R, Davis AR (2006) UV-induced mortality inencapsulated intertidal embryos: are MAAs an eVective sun-screen? J Chem Ecol 32:993–1004
Yamahira K (1996) The role of intertidal egg deposition on survival ofthe puVer, Takifugu niphobles (Jordan et Snyder), embryos. J ExpMar Biol Ecol 98:291–306
Yaroslavtseva LM, Sergeeva EP, Kulikova VA (2001) The eVective-ness of the protection of embryos and larvae within egg massesof the gastropod Epheria turrita against changes in salinity anddesiccation. Russ J Mar Biol 27:8–13
Zar JH (1998) Biostatistical analysis. Prentice Hall, Upper SaddleRiver
123




















