
Does Incubation Temperature Fluctuation Influence Hatchling Phenotypes in Reptiles? A Test Using...
12
Does Incubation Temperature Fluctuation Influence Hatchling Phenotypes in Reptiles? A Test Using Parthenogenetic Geckos Author(s): Sarah J. Andrewartha, Nicola J. Mitchell, and Peter B. Frappell Reviewed work(s): Source: Physiological and Biochemical Zoology, Vol. 83, No. 4 (July/August 2010), pp. 597-607 Published by: The University of Chicago Press Stable URL: http://www.jstor.org/stable/10.1086/652245 . Accessed: 17/01/2013 23:34 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. . The University of Chicago Press is collaborating with JSTOR to digitize, preserve and extend access to Physiological and Biochemical Zoology. http://www.jstor.org This content downloaded on Thu, 17 Jan 2013 23:34:36 PM All use subject to JSTOR Terms and Conditions
Transcript of Does Incubation Temperature Fluctuation Influence Hatchling Phenotypes in Reptiles? A Test Using...

Does Incubation Temperature Fluctuation Influence Hatchling Phenotypes in Reptiles? A TestUsing Parthenogenetic GeckosAuthor(s): Sarah J. Andrewartha, Nicola J. Mitchell, and Peter B. FrappellReviewed work(s):Source: Physiological and Biochemical Zoology, Vol. 83, No. 4 (July/August 2010), pp. 597-607Published by: The University of Chicago PressStable URL: http://www.jstor.org/stable/10.1086/652245 .
Accessed: 17/01/2013 23:34
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
The University of Chicago Press is collaborating with JSTOR to digitize, preserve and extend access toPhysiological and Biochemical Zoology.
http://www.jstor.org
This content downloaded on Thu, 17 Jan 2013 23:34:36 PMAll use subject to JSTOR Terms and Conditions

597
Does Incubation Temperature Fluctuation Influence Hatchling
Phenotypes in Reptiles? A Test Using Parthenogenetic Geckos
* Corresponding author; e-mail: [email protected].†Present address: School of Zoology, University of Tasmania, Hobart, Tasmania
7001, Australia.
Physiological and Biochemical Zoology 83(4):597–607. 2010. � 2010 by TheUniversity of Chicago. All rights reserved. 1522-2152/2010/8304-9116$15.00DOI: 10.1086/652245
Sarah J. Andrewartha1
Nicola J. Mitchell1,2,*Peter B. Frappell1,†
1Adaptational and Respiratory Physiology Laboratory,Department of Zoology, La Trobe University, Melbourne,Victoria 3086, Australia; 2School of Animal Biology,University of Western Australia, Crawley, Western Australia6009, Australia
Accepted 2/21/2010; Electronically Published 5/17/2010
ABSTRACT
Many lineages of parthenogenetic organisms have persistedthrough significant environmental change despite the con-straints imposed by their fixed genotype and limited evolu-tionary potential. The ability of parthenogens to occur sym-patrically with sexual relatives may in part be due to phenotypicplasticity in their responses to their environment, especiallywith respect to incubation temperature—a maternally selectedtrait. Here we measured the incubation temperatures selectedby two lineages of triploid parthenogenic geckos in the Het-eronotia binoei complex by allowing them to deposit clutchesalong a thermal gradient. The average nest temperature selectedwas 28.4�C, with no significant differences between partheno-genic races or individual clones. To investigate the effect ofnest-temperature variability on physiological and morpholog-ical traits, we incubated eggs from different races at one of fourincubation regimes ( , �3�, �5�, or �9�C). Embryos32� � 0�
incubated at constant 32�C developed faster than embryosreared under increasing extremes of diel temperature fluctua-tion (�3�, �5�C), and incubation at C was unsuc-32� � 9�
cessful. Incubation regime had no effect on the body size, pre-ferred substrate temperature, or mass-specific of hatchlings.Vo2
However, parthenogenic race had a significant effect on eggmass, tail length, snout-to-vent length, total length, and .Vo2
We conclude that developmental traits are strongly influencedby clonal genotypes in this parthenogenic complex but are wellbuffered against fluctuations in incubation temperature.
Introduction
The processes that promote phenotypic plasticity are critical tounderstanding how organisms adapt to their environment(Kingsolver and Huey 1998). Temperature has a profound effecton the development of ectotherms, and phenotypic traits thatare known to be sensitive to temperature include developmenttime, offspring size, morphology, sex, metabolic rates, and per-formance (Packard and Packard 1988; Pieau 1996; Shine et al.1997; Birchard 2004). The thermal environment can directlyinfluence the selection of phenotypes (Price et al. 2003), andmothers can theoretically attain high fitness by selecting ap-propriate incubation temperatures for their offspring (Jones etal. 1987; Mousseau and Fox 1998). Much of the work on in-cubation temperature as a maternal effect has been conductedon reptiles because both viviparous and oviparous species em-ploy behaviors (thermoregulation and nest-site selection) thatdirectly influence offspring phenotypes. For example, the Aus-tralian water dragon Physignathus lesueurii—a species with tem-perature-dependent sex determination—selects more shadednest sites in warmer parts of its range, and the result is thatnest temperatures and sex ratios are similar across a wide lat-itudinal range (Doody et al. 2006)
Reptile embryos generally develop successfully across a 5�–8�C range of constant temperatures but can develop across amuch broader range of temperatures provided that exposureto extreme temperatures is intermittent (Birchard 2004). Manysquamate reptiles (lizards, snakes, and amphisbaenians) nest inshallow soils where short-term fluctuations in egg temperatureare a consequence of daily variation in the cumulative solarradiation received and the degree of nest insulation. Selectionof nests based on average temperature is one mechanism bywhich females can influence the fitness of their offspring. Dailytemperature variation may have equally important conse-quences (Packard and Packard 1988; Shine 2004a), but theinfluence of temperature variation on phenotypic traits ispoorly understood. In part, this is due to an overwhelmingfocus on reaction norms whereby values for a particular mor-phological or physiological trait are systematically assessedacross a range of constant temperatures tolerated by embryos.However, recent studies have demonstrated that hatching traitssubject to selection are also responsive to the magnitude ofvariation in incubation temperature. For example, incubationof reptile eggs at fluctuating temperatures causes body size tobe smaller in snakes but improves locomotor performance inturtles and skinks relative to individuals incubated under con-stant conditions (Shine and Harlow 1996; Webb et al. 2001;Ashmore and Janzen 2003).
Interactions between mean temperatures and the magnitude
This content downloaded on Thu, 17 Jan 2013 23:34:36 PMAll use subject to JSTOR Terms and Conditions

598 S. J. Andrewartha, N. J. Mitchell, and P. B. Frappell
of temperature variation can influence phenotypes (Du andFeng 2008). Moreover, clutch effects (whether genetic or dueto the mothers’ thermal regime during embryogenesis) alsoinfluence hatchling traits (e.g., Jayne and Bennett 1990; Shineand Harlow 1996: Shine et al. 1997; Andrews et al. 2000; Ash-more and Janzen 2003), and complex interactions between ge-netic components (or other clutch effects), initial egg mass,mean temperature, and temperature fluctuation have been doc-umented in at least one study (Mullins and Janzen 2006). Otherstudies on reptiles, however, have found no effect of diel tem-perature fluctuation on hatchling traits (Andrews et al. 2000;Ashmore and Janzen 2003; Du and Ji 2003; Du et al. 2005).These variable responses to temperature fluctuation may bedue to differences in the thermal tolerance ranges of the speciesso far examined or to the confounding influence of maternaland genetic factors on phenotypic expression. Unraveling therelative importance of environmental influences on individualphenotypes may be most readily achieved in parthenogenicorganisms because variation within clones is attributable onlyto maternal and environmental factors.
Several squamate reptiles have evolved parthenogenetic lin-eages (White 1978); the best-studied example is the Heteronotiabinoei gecko complex that is broadly distributed across theAustralian arid zone (Moritz 1983; Strasburg and Kearney 2005;Strasburg et al. 2007). This complex consists of five sexual andtwo parthenogenic forms, some of which occur sympatrically.In a previous study of H. binoei, individuals hatched at largerbody sizes from warmer incubation temperatures (30�C) andhad higher rates of growth and survival (Kearney and Shine2004a), demonstrating that incubation temperature altershatchling phenotypes in a manner that should influence Dar-winian fitness. In this study we used offspring produced by thesame parthenogenic females as Kearney and Shine to assess (1)the mean and range of nest temperatures selected by each cloneand (2) whether the magnitude of diel fluctuations in incu-bation temperature affects morphological and physiologicaltraits of hatchlings, including their metabolic rates across abroad range of environmental temperatures.
Material and Methods
Study Species and Husbandry
The parthenogenetic races of Heteronotia binoei most likelyoriginated in the late Pleistocene (0.11–0.33 Ma), when Aus-tralian deserts were expanding and contracting in response to∼100-ka glacial cycles (Kearney et al. 2006; Byrne et al. 2008).Parthenogens were formed from an initial hybridization be-tween the CA6 and SM6 sexual species (the 3N1 and 3N2lineages) that later backcrossed with males of the progenitorspecies to form triploid clones of either race A (CA6/SM6/CA6) or race BC (CA6/SM6/SM6; Moritz 1983; Strasburg andKearney 2005). The high clonal diversity in both the A and BCraces suggest repeated hybridization events, but in general,mtDNA diversity within the parthenogenetic races is very lowand is similar to that of localized populations of the sexualspecies (Strasburg and Kearney 2005). In contrast, partheno-
gens exhibit high levels of heterozygosity in nuclear DNA mark-ers because of their triploid genome (Kearney and Shine 2004a,2004b). Although gene dosage may complicate the interpre-tation of physiological responses in triploids, a recent study oftriploid fishes suggests that one haplome is inactivated and thatgene expression in triploids is similar to that of diploid indi-viduals (Pala et al. 2008).
The reproductive physiology of sexual and parthenogenic H.binoei is broadly similar, but parthenogens have about 30%lower fecundity, producing fewer clutches than their sexualrelatives (Kearney and Shine 2005). During a typical breedingseason in the austral spring and summer, female H. binoeideposit several clutches of 1–2 eggs in an unattended nest site,usually in shallow soil or under bark or leaf litter (Bustard1969, 1970). Like other species in the subfamily Gekkonidae,the eggs of H. binoei have hard, calcareous shells that are rel-atively impermeable to water (Dunson 1982); hence, the waterpotential of nest sites should have no influence on offspringphenotype except indirectly via interactions between water po-tential and soil temperature.
A colony of wild-caught H. binoei parthenogens from centralAustralia that were subject to previous study at the Universityof Sydney (Kearney and Shine 2004a, 2004b, 2005) have beenmaintained in a controlled environment room at La Trobe Uni-versity (Melbourne, Australia) since 2003. Ten clone lines (cov-ering both races), each consisting of 1–10 females—whereclonal identity was confirmed by genetic typing using micro-satellite DNA markers (see Strasburg 2004)—were housedseparately on shallow sand within large plastic containers( ) and providedL # W # H p 400 mm # 260 mm # 120 mmwith shredded paper for cover. Sand temperatures in each boxvaried between about 21� and 44�C, depending on the prox-imity to a 200-mm-wide heating strip positioned beneath theback of each tub. Females were thus able to seek shelter, ther-moregulate, and select oviposition sites based on thermal cri-teria. Photoperiods were adjusted every 3 mo to approximatelocal day : night cycles, and females were fed twice weekly withcrickets and provided with water ad lib.
Thermal Environment of Nest Sites
Females were examined weekly for the presence of ovarianfollicles or shelled eggs that could be seen through the trans-lucent abdomen. In a pilot study in the 2003–2004 breedingseason, we recorded the position of eggs deposited in each tuband estimated the selected nest temperature from a thermalgradient previously measured with Thermacron iButton dataloggers (DS1921, �0.5�C precision, Maxim Integrated Prod-ucts, Sunnyvale, CA) in a representative container. In the 2004–2005 breeding season, we relocated heavily gravid females toindividual nesting containers with a cardboard cover positionedover half the sand surface to span the thermal gradient. A waterdish was positioned randomly on the uncovered sand. Femaleswere left in the nesting box until oviposition or were removedif eggs were reabsorbed. Eggs were located by carefully searchingthe sand and were removed and replaced with an iButton pro-
This content downloaded on Thu, 17 Jan 2013 23:34:36 PMAll use subject to JSTOR Terms and Conditions

Incubation of Clonal Geckos at Fluctuating Temperatures 599
grammed to record temperature every 5 min for at least 24 h.Egg mass (weighed in grams on a Mettler AE240 balance, ac-curate to four decimal places), mother, and clonal identity wererecorded, and mothers were returned to their clone box. Eggswere then transferred into individual 150-mL plastic jars halffilled with dry sand, covered with plastic cling film, and in-cubated under one of four thermal regimes.
Egg Incubation
In our pilot study in the 2003–2004 breeding season, femalesdid not nest in the front half of clone boxes where sand tem-peratures were cooler (∼21�–23�C). Clutches were usually lo-cated on or near the heating pad, where temperatures rangedbetween 29� and 44�C. Most clutches (39% of 69%) were po-sitioned where temperatures were approximately 32�–34�C. Asthese temperatures were similar to the mean preferred substratetemperature of 30.6�–31.2�C measured for adult H. binoei par-thenogens (Kearney and Shine 2004a), we selected 32�C as anindication of a preferred incubation temperature. Then, basedon the observation that H. binoei experience diel temperaturevariation up to 10�C during incubation (Bustard 1968, 1969),we established four experimental incubation regimes: one at aconstant 32�C, two regimes with moderate diel temperaturefluctuation ( C and C), and an extreme treat-32� � 3� 32� � 5�
ment of C (all sinusoidal cycles, zenith 1200 hours,32� � 10�
12L : 12D). Individual eggs were randomly assigned to an in-cubation regime; two eggs from a single mother were placedinto different treatments, and eggs from subsequent clutchesof the same individual were allocated to other treatments whenpossible. The incubation temperature of individual embryoswas estimated from half-hourly temperature records obtainedfrom iButtons positioned within the incubator inside sand-filled vials identical to those containing eggs. Incubators werechecked twice daily for hatchlings, and eggs that failed to de-velop (as judged by candling) were periodically removed.
Hatchling Mass and Morphology
Newly hatched individuals were removed from incubators andimmediately weighed before being photographed on 1-mmgraph paper with a digital camera attached to a stereomicro-scope. Snout-to-vent and tail lengths (SVL and TL) were latermeasured from calibrated images using tpsDig 1.31 image anal-ysis software (F. J. Rohlf, State University of New York, StonyBrook).
Preferred Substrate Temperature
Hatchlings aged between 0 and 2 d old were placed at randomat either the hot (∼36�C) or cold (∼14.0�C) end of a 500-mmcustom-built aluminum block. The temperatures at each endof the block were controlled by thermostated water pumpedthrough copper heat exchange coils. The block was inset witha shallow groove large enough to allow hatchlings to turn (150mm wide by 150 mm high) and covered with clear Perspex to
allow viewing. The groove was divided into 10 divisions, andthe thermal gradient was calibrated on each day of use bymeasuring the temperature at positions 1 and 10 with a copper/constantan thermocouple. Following an initial acclimation pe-riod of 1 h, the position of the hatchling in the gradient wasrecorded every 5 min for 30 min, or until a consistent positionwas reached. Measurement of temperatures at positions 1–10within the groove showed that temperatures increased linearlywithin the gradient; hence, a linear equation was used to es-timate the substrate temperature at the modal position (Tpref)based on temperatures measured at positions 1 and 10.
Rate of Oxygen Consumption
Rates of oxygen consumption were determined for eachVo2
hatchling at a range of ambient temperatures (Ta) either beforeor after determination of Tpref (randomized; therefore, the timeof day that the measurements were made was also randomized),when the hatchling was between 0 and 2 d old. Each hatchlingwas placed into a small, darkened, water-jacketed chamber (in-ner dimensions: 50 mm long, 20 mm in diameter) throughwhich air flowed at a known rate of 40–60 mL min�1. Thechamber was first cooled to 17�C, and the hatchlings wereallowed 45 min to settle. The chamber was then sealed for 20min. Following the period that the chamber was sealed, thechamber was flushed and the excurrent gas dried (Drierite, W.A. Hammond Drierite Company, Xenia, OH) and scrubbedfree of CO2 (Dragersorb, Dragerwerk, Lubeck) before O2 frac-tional concentration was analyzed (ML206 gas analyzer, ADInstruments, Bella Vista, New South Wales). The chamber wasthen sequentially heated to 22�, 27�, 32�, and 37�C, allowing a45-min settling period between each increase in temperatureand a sealing time of 10–20 min (depending on temperature)to determine . The output was recorded by Powerlab at aVo2
rate of 10 Hz.was calculated from the time integral of the gas con-Vo2
centration curves multiplied by the flow and the reciprocal ofthe time during which the compartment was sealed (Frappelland Mortola 2000). Scrubbing CO2 from the excurrent gas takesinto account the respiratory exchange ratio, that is, the differ-ence between the number of moles of O2 consumed to CO2
produced (see Frappell et al. 1992, Appendix).
Data Analysis
Statistical procedures were performed in JMP (ver. 7.0.2, SASInstitute, Cary, NC) or Minitab (ver. 14, Minitab, State College,PA), and statistical significance was assumed at . DataP ! 0.05are presented as means � 1 SE. Egg masses and nest temper-atures were compared between races using restricted maximumlikelihood (REML) estimation of variance components, whichprovide reliable estimates of variance in unbalanced designs(Quinn and Keogh 2002). Race (A or BC) was treated as afixed effect, clone nested within race was treated as a randomeffect, and female was treated as a random factor nested withinrace and clone, with the dependent variable being either egg
This content downloaded on Thu, 17 Jan 2013 23:34:36 PMAll use subject to JSTOR Terms and Conditions

600 S. J. Andrewartha, N. J. Mitchell, and P. B. Frappell
Table 1: Reproductive output of parthenogenic Heteronotia binoei clones used in the study in 2004–2005
ParthenogeneticRace and Clone
Number ofFemalesin Colony
Number ofFemalesThat Reproduced
Number ofEggs Produced
AverageNumber ofClutchesper Female
Range ofOviposition Dates
A:7 10 10 38 2.2 October 19–February 710 1 1 6 3 October 25–December 2611 7 7 35 3.4 September 28–February 1717 1 1 3 2 December 16–January 2422 1 1 4 3 November 8–January 25
BC:4 8 6 9 1.2 December 5–January 2512 4 4 15 2.3 October 16–February 338 5 3 17 3.3 October 22–February 19
Table 2: REML analysis of the effect of race (fixed effect),clone (random effect nested within race), and females(random effect nested within race and clone) on egg massand selected nest temperatures of parthenogenicHeteronotia binoei
Source of Variation nTest or VarianceComponent
Variance(% of Total)
Egg mass:Race 2 F1, 6.12 p .381, NSClone (race) 8 9.852 # 10�4 26.4Female (race, clone) 33 1.545 # 10�4 4.1Residual 114 2.596 # 10�2 69.4
Nest temperature:Race 2 F1, 2.47 p .043, NSClone (race) 8 .764 4.5Female (race, clone) 32 �4.091 �24.1Residual 73 20.32 119.6
Note. Egg masses (g) and nest temperature (�C) were assessed for races
A and BC using 3–5 clones per race and 1–10 females per clone. F statistics
are given for the fixed effect race, and variance components are given for
random effects. NS p nonsignificant at .P ! 0.05
mass or nest temperature. In the analysis of egg mass, all eggswere treated as separate data points, whereas in analyses of nesttemperature, data from clutches of two eggs were only includedonce.
Separate REML analyses were performed to test for signifi-cant differences in egg mass and nest temperature amongclones. In these analyses, clones were treated as fixed factors,and females were treated as random factors nested withinclones. Tukey’s HSD tests were used for post hoc comparisonsamong clones. We used one-way ANOVA to assess the repeat-ability of traits measured for each female by computing intra-class correlation coefficients (ICCs) from the variance com-ponents of the ANOVA (Lessells and Boag 1987). Females forwhich we had a single measurement of either trait were ex-cluded from the ANOVA.
Hereafter, data were pooled for races A and BC because ofthe low numbers of hatchlings available for many clones. x2
tests were used to analyze hatching success data across incu-bation regimes. One-way ANOVA with Tukey HSD post hoctests or general linear models (GLM) with egg mass as a co-variate were used to analyze data on incubation time, hatchlingmorphology, and preferred substrate temperature. dataVo2
were analyzed in a separate GLM with hatchling mass as acovariate. For morphological, , and Tpref measurements,Vo2
both eggs from two-egg clutches were included for analysis. Allnonsignificant interactions and factors were removed fromGLMs.
Results
Females produced between one and five clutches during the2004–2005 breeding season. Two clones (both race BC) thatproduced only a single egg were excluded from further analysis,and the reproductive output of the remaining eight clones isshown in Table 1. Some eggs were dented or broken whenremoved from the nest box and could not be used any longerin the study.
Egg mass did not differ between races, averaging 0.43 �
g (Table 2). However a separate analysis indicated signif-0.01
icant differences among clones ( , ), withF p 5.32 P p 0.0257, 16.6
females from clones 7, 10, and 38 producing larger eggs thanfemales from clone 11 (post hoc tests; Fig. 1). The ICC cal-culated from a one-way ANOVA with individual females asfactors was 0.32 ( , ), indicating that eggF p 2.98 P p 0.00124, 82
mass was repeatable within females.There were no differences in the nest temperature selected
in a thermal gradient by between-parthenogenic races (Table2), and the average nest temperature was 28.4�C (Fig. 2). Sim-ilarly, there were no significant differences among clones in theselected nest temperatures ( , ). The vari-F p 1.13 P p 0.437, 7.6
ance components of a one-way ANOVA, with individual fe-males as factors, produced an ICC of �0.1 ( ,F p 0.7321, 41
), indicating that nest temperatures selected by indi-P p 0.77vidual females were not repeatable.
Temperature logger records from incubators revealed thatthe mean temperature and extent of fluctuation about the mean
This content downloaded on Thu, 17 Jan 2013 23:34:36 PMAll use subject to JSTOR Terms and Conditions

Incubation of Clonal Geckos at Fluctuating Temperatures 601
Figure 1. Egg masses of Heteronotia binoei clones. Unfilled symbols are individual eggs, filled symbols are mean values for each clone line.Error bars are �1 SE. Clones not connected by the same symbol are significantly different.
varied slightly from the programmed cycles. The constant-temperature incubator averaged 31.6�C with a diel range of0.5�C. The cycling temperature incubators averaged 32.1� �
C, C, and C, but for simplicity, the2.8� 31.9� � 4.7� 31.6� � 9.2�
temperature treatments will be hereafter referred to as 32�Cconstant, C, C, and C.32� � 3� 32� � 5� 32� � 9�
No eggs hatched in the C treatment, and the eggs32� � 9�
incubated in this regime are excluded from further analysis.Hatching success in the other fluctuating regimes (52% in the
C and 66% in the C) was significantly lower32� � 3� 32� � 5�
than in the 32�C constant regime (76%; , ,2x p 8.17 df p 1). Increased fluctuation in incubation temperature andP ! 0.01
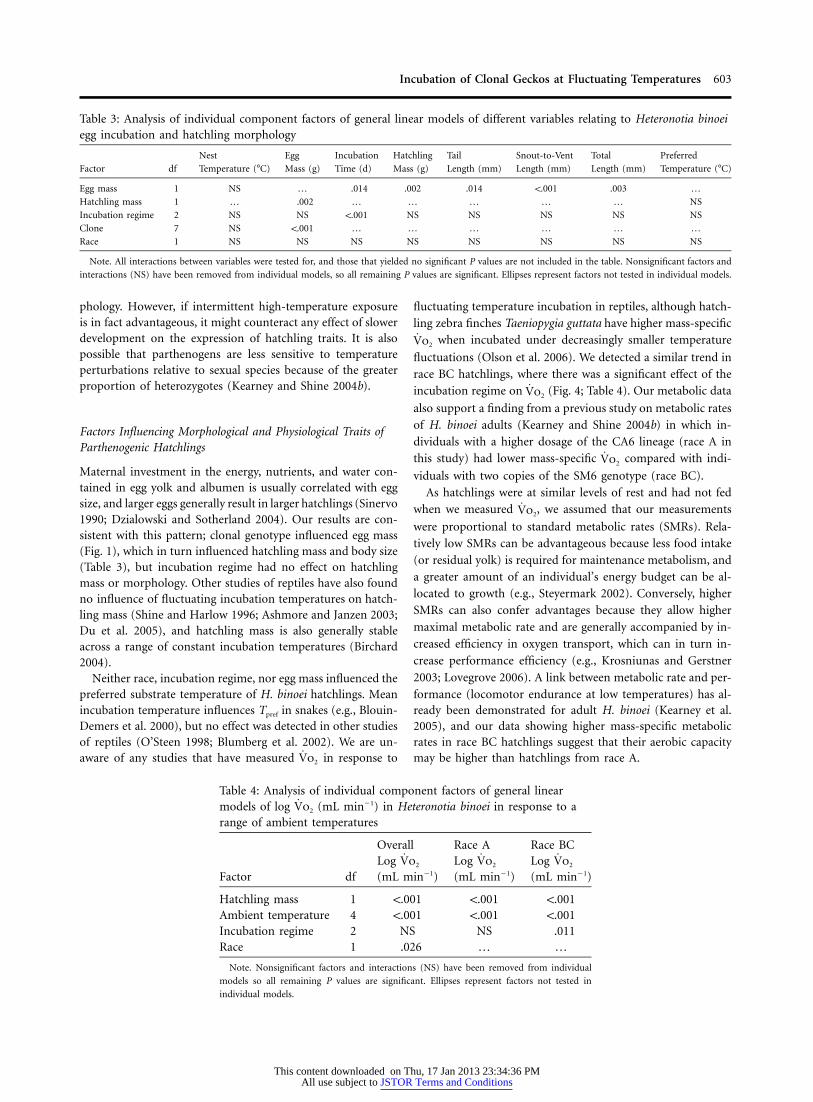
larger egg mass both extended incubation times, and theseeffects were independent (Fig. 3; Table 3). Egg mass had asignificant effect on hatchling mass, but hatchling mass wasnot influenced by race or incubation regime. Hatchling SVL,tail length, and consequently total length were only influencedby egg mass (Table 3).
There was no effect of race, incubation regime, or hatchlingmass on Tpref (Table 3), and most hatchlings selected a substratetemperature around 31�C (mean 31.3�C, interquartile range p30.5�–34.2� C, ). However, hatchling mass, incubationn p 43temperature, and race all affected hatchling (Table 4). WithVo2
increasing Ta, the mass-corrected of hatchlings increasedVo2
nonlinearly; the Q10 according to van ’t Hoff’s principle was3.31 between 17� and 22�C, 2.93 between 22� and 27�C, 2.01between 27� and 32�C, and 1.86 between 32� and 37�C. Hatch-lings from race A had an overall higher mass-specific metabolicrate than race BC (Fig. 4). Incubation temperature oscillationhad no effect on race A hatchling (Table 4) but did influ-Vo2
ence the of race BC hatchlings, where hatchlings from bothVo2
oscillating regimes had lower than hatchlings from the 32�CVo2
constant regime (Fig. 4).
Discussion
Quantification of lifetime fitness requires survival to sexualmaturity and the production of viable offspring, but in the
absence of these metrics, hatchling phenotypes can be usefulindicators of the fitness of an individual or a genotype (Arnold1983; Deeming 2004; Kingsolver and Huey 2008). In this studywe found that none of the morphological or physiological traitsof hatchlings we assessed were affected by magnitude of thedaily fluctuation in incubation temperature apart from a sig-nificant influence of the incubation regime on the mass-specific
of race BC parthenogens (Table 4; Fig. 4B). However, theVo2
magnitude of the diel temperature fluctuation did influencedevelopment times and hatching success, and embryonic mor-tality was lower and development times were faster with de-creasing temperature fluctuation. The most significant deter-minant of the hatchling phenotype was clonal genotype, whichaffected egg mass and consequently affected hatchling body sizeand .Vo2
Few studies of reptiles exist in which females have been al-lowed to select nest sites along a thermal gradient, but a typicalresponse in the field is for females to select incubation tem-peratures that are intermediate in the nonlethal range for de-velopment (Birchard 2004; Blouin-Demers et al. 2004). Thisstudy was no exception; the average nest temperature selected(28.4�C; Fig. 2) was similar to the optimum constant incubationtemperature for hatchling growth and survival (30�C; Kearneyand Shine 2004a). This pattern supports the prediction of Mar-tin and Huey (2008) that preferred temperatures should beslightly below the optimal temperature determined by a fitnessfunction. However, the nest temperatures selected by femalesin each clone (26�–30�C; Fig. 2) were lower than we expected,based on the modal nest temperature selected in 2003–2004(32�–34�C). Consequently, the incubation regimes that we em-ployed—fluctuating about a mean of 32�C—were near the up-per end of the spectrum of incubation temperatures toleratedby Heteronotia binoei, whereas a previous study of the effectsof constant temperature on the morphological and phenotypictraits of parthenogens encompassed the cooler end of the de-velopmental spectrum (24�–30�C; Kearney and Shine 2004a).
In this study, embryos subjected to the most extreme treat-
This content downloaded on Thu, 17 Jan 2013 23:34:36 PMAll use subject to JSTOR Terms and Conditions

602 S. J. Andrewartha, N. J. Mitchell, and P. B. Frappell
Figure 2. Nest temperatures selected by Heteronotia binoei clones. The measured oviposition site temperatures for the eight clones in the studyare shown by unfilled circles, and filled circles indicate the mean (�1 SE) for each clone.
Figure 3. Time to hatching (d g�1 egg mass) for races A and BCfollowing incubation at either constant or fluctuating temperatures.Error bars are �1 SE. Paired symbols represent statistically responsesto incubation regimes for each race.
ment ( C) spent more than 4 h of each daily cycle at32� � 9�
temperatures above 40�C, which was lethal exposure becauseno eggs hatched. However, hatching success in the constant32�C treatment (76%) was higher than the 45%–69% recordedfor parthenogenetic races of H. binoei incubated at constanttemperatures between 24� and 30�C (Kearney and Shine 2004a).Further, although hatching success at C and32� � 3� 32� �
C (52%–66%) was significantly lower than at constant 32�C,5�
these values are similar to hatching rates from constant-temperature incubation between 24� and 30�C (Kearney andShine 2004a), suggesting that our moderately fluctuating in-cubation regimes were not especially stressful. Analysis of tem-perature records from three H. binoei nests in central Australia(measured over 44 d in September and October) showed dailyoscillations of 3.5�, 6.6�, and 9.3�C (M. R. Kearney, unpublisheddata), indicating that our �3�C and �5�C treatments wererealistic approximations of diel temperature variation experi-enced in natural nests.
Daily fluctuations in temperature should have no influenceon development times over and above that of the daily meanprovided that (1) development rate and temperature are linearlyrelated over the range of temperatures experienced by the em-bryos and (2) there is no hysteresis in the action of temperatureon development rate (Georges et al. 1994). Because we foundthat development times increased with greater fluctuationaround the same mean temperature, it is likely that develop-ment rates and temperature were not linearly related across therange of temperatures to which we exposed the embryos. Sim-ilarly, the disparate effects of increasing temperature fluctuationreported in other studies—either increasing (Ashmore and Jan-zen 2003; Du and Ji 2003) or decreasing development times(Shine and Harlow 1996; Shine 2004b)—could be explainedby nonlinearity in temperature/development-rate relationship(for a recent review, see Georges et al. 2005). We used the
program DEVARA (Dallwitz and Higgins 1992) to estimate theparameters of a nonlinear development-rate function for par-thenogenetic H. binoei races. This program is specifically de-signed to accommodate data from experiments in which in-cubation temperatures vary (e.g., Mitchell et al. 2008), and ourfitted model suggests that the development rate of H. binoeipeaks at 2.25% d�1 at 35�C and falls rapidly to 0% d�1 by 40�C(Fig. 5). Given that embryos in the C and C32� � 3� 32� � 5�
treatment were exposed to temperatures at which developmentrates are nonlinear and in fact decline, it is notable that wefound few associated affects on hatchling physiology or mor-
This content downloaded on Thu, 17 Jan 2013 23:34:36 PMAll use subject to JSTOR Terms and Conditions
Incubation of Clonal Geckos at Fluctuating Temperatures 603
Table 3: Analysis of individual component factors of general linear models of different variables relating to Heteronotia binoeiegg incubation and hatchling morphology
Factor df
Nest
Temperature (�C)
Egg
Mass (g)
Incubation
Time (d)
Hatchling
Mass (g)
Tail
Length (mm)
Snout-to-Vent
Length (mm)
Total
Length (mm)
Preferred
Temperature (�C)
Egg mass 1 NS … .014 .002 .014 !.001 .003 …
Hatchling mass 1 … .002 … … … … … NS
Incubation regime 2 NS NS !.001 NS NS NS NS NS
Clone 7 NS !.001 … … … … … …
Race 1 NS NS NS NS NS NS NS NS
Note. All interactions between variables were tested for, and those that yielded no significant P values are not included in the table. Nonsignificant factors and
interactions (NS) have been removed from individual models, so all remaining P values are significant. Ellipses represent factors not tested in individual models.
Table 4: Analysis of individual component factors of general linearmodels of log (mL min�1) in Heteronotia binoei in response to aVo2
range of ambient temperatures
Factor df
OverallLog Vo2
(mL min�1)
Race ALog Vo2
(mL min�1)
Race BCLog Vo2
(mL min�1)
Hatchling mass 1 !.001 !.001 !.001Ambient temperature 4 !.001 !.001 !.001Incubation regime 2 NS NS .011Race 1 .026 … …
Note. Nonsignificant factors and interactions (NS) have been removed from individual
models so all remaining P values are significant. Ellipses represent factors not tested in
individual models.
phology. However, if intermittent high-temperature exposureis in fact advantageous, it might counteract any effect of slowerdevelopment on the expression of hatchling traits. It is alsopossible that parthenogens are less sensitive to temperatureperturbations relative to sexual species because of the greaterproportion of heterozygotes (Kearney and Shine 2004b).
Factors Influencing Morphological and Physiological Traits ofParthenogenic Hatchlings
Maternal investment in the energy, nutrients, and water con-tained in egg yolk and albumen is usually correlated with eggsize, and larger eggs generally result in larger hatchlings (Sinervo1990; Dzialowski and Sotherland 2004). Our results are con-sistent with this pattern; clonal genotype influenced egg mass(Fig. 1), which in turn influenced hatchling mass and body size(Table 3), but incubation regime had no effect on hatchlingmass or morphology. Other studies of reptiles have also foundno influence of fluctuating incubation temperatures on hatch-ling mass (Shine and Harlow 1996; Ashmore and Janzen 2003;Du et al. 2005), and hatchling mass is also generally stableacross a range of constant incubation temperatures (Birchard2004).
Neither race, incubation regime, nor egg mass influenced thepreferred substrate temperature of H. binoei hatchlings. Meanincubation temperature influences Tpref in snakes (e.g., Blouin-Demers et al. 2000), but no effect was detected in other studiesof reptiles (O’Steen 1998; Blumberg et al. 2002). We are un-aware of any studies that have measured in response toVo2
fluctuating temperature incubation in reptiles, although hatch-ling zebra finches Taeniopygia guttata have higher mass-specific
when incubated under decreasingly smaller temperatureVo2
fluctuations (Olson et al. 2006). We detected a similar trend inrace BC hatchlings, where there was a significant effect of theincubation regime on (Fig. 4; Table 4). Our metabolic dataVo2
also support a finding from a previous study on metabolic ratesof H. binoei adults (Kearney and Shine 2004b) in which in-dividuals with a higher dosage of the CA6 lineage (race A inthis study) had lower mass-specific compared with indi-Vo2
viduals with two copies of the SM6 genotype (race BC).As hatchlings were at similar levels of rest and had not fed
when we measured , we assumed that our measurementsVo2
were proportional to standard metabolic rates (SMRs). Rela-tively low SMRs can be advantageous because less food intake(or residual yolk) is required for maintenance metabolism, anda greater amount of an individual’s energy budget can be al-located to growth (e.g., Steyermark 2002). Conversely, higherSMRs can also confer advantages because they allow higher
maximal metabolic rate and are generally accompanied by in-
creased efficiency in oxygen transport, which can in turn in-
crease performance efficiency (e.g., Krosniunas and Gerstner
2003; Lovegrove 2006). A link between metabolic rate and per-
formance (locomotor endurance at low temperatures) has al-ready been demonstrated for adult H. binoei (Kearney et al.2005), and our data showing higher mass-specific metabolicrates in race BC hatchlings suggest that their aerobic capacitymay be higher than hatchlings from race A.
This content downloaded on Thu, 17 Jan 2013 23:34:36 PMAll use subject to JSTOR Terms and Conditions

604 S. J. Andrewartha, N. J. Mitchell, and P. B. Frappell
Figure 4. Mass-specific versus ambient temperature in race A andVo2
race BC hatchlings following incubation at either constant or fluctu-ating temperatures. Error bars are �1 SE.
Figure 5. Development rate of parthenogenetic Heteronotia binoei em-bryos as a function of incubation temperature. Data from this studyare indicated by filled circles, and horizontal lines show the extent ofthe temperature fluctuation. Other data points are all from constant-temperature incubation: unfilled squares are data from Kearney andShine (2004a), and unfilled triangles are unpublished data (N. J. Mitch-ell). The solid line is a hypothetical embryonic development-rate func-tion for parthenogenetic H. binoei fitted to the combined data usingthe program DEVARA (Dallwitz and Higgins 1992) with starting pa-rameters , , , , and , where b1b p 2.2 b p 20 b p 35 b p 6 b p 0.41 2 3 4 5
is the maximum development rate, b2 is the lower temperature at whichthe development rate approaches 0, and b3 is the temperature at themaximum development rate. Parameters b4 and b5, which control theasymmetry and steepness of the curve, were fixed at 6 and 0.4, re-spectively, which is recommended when data at extreme temperaturesare scarce (Dallwitz and Higgins 1992). All data points were weightedequally.
Could Phenotypic Plasticity Explain the Persistence ofHeteronotia binoei Parthenogens?
Several authors have suggested that phenotypic plasticity mayexplain the persistence of parthenogens through environmentalchange, given that the alternative mechanism–adaptation–canonly occur via mutation and not genetic recombination (Schei-ner 1993; Dybdahl and Kane 2005). Parthenogenic H. binoeihave survived temperature changes during the late Pleistoceneof around 10�C (Byrne et al. 2008)—presumably by trackingfavorable climates and by exploiting plasticity in their responsesto thermal environments (sensu Price et al. 2003). In this study,females within a single clone selected nest temperatures acrossat least a 10�C range (e.g., clones 7 and 11; Fig. 2). Whetherthese laboratory-generated data reflect within-clone differencesin the temperatures of natural nest sites is unknown, but giventhat the latitudinal range of some clones spans at least 150 km(M. R. Kearney, unpublished data), marked incubation tem-perature variation is likely. Hence, landscape-scale variation innest temperatures could potentially generate phenotypic vari-ability in this system because hatchling traits, such as body
mass and running speed, and posthatchling growth rates andsurvival are influenced by temperature (Kearney and Shine2004a, 2004b). Conversely, our data imply that microhabitat-scale variation in nest irradiation or insulation would not trans-late into marked differences in phenotypes (with the exceptionof incubation duration), but experiments where the mean tem-perature is closer to the preferred nest temperature of ∼28�Cmay be more revealing.
As a final cautionary note, it is worth recognizing that phe-notypes at hatching may not be a reliable indicator of futurefitness. In tuatara Sphenodon punctatus, for example, hatchlingsincubated in natural nests in which temperatures varied dailyand seasonally hatched at body sizes similar to conspecificsreared at constant temperatures, but under identical posthatch-ing conditions, they grew more slowly and were the smallestcohort at 10 mo of age (Nelson et al. 2004, 2006). However,at 5 yr of age, naturally incubated males were larger and heavierthan males incubated at constant temperatures, reversing theearlier trend (Gruber 2007). While these effects could be at-tributable to differences in temperature variability or wateravailability during incubation, the example illustrates why thisparthenogenic model (where confounding effects of clutch, sex,and water potential can be controlled) is an ideal system for
This content downloaded on Thu, 17 Jan 2013 23:34:36 PMAll use subject to JSTOR Terms and Conditions

Incubation of Clonal Geckos at Fluctuating Temperatures 605
evaluating the effects of variable incubation temperatures onphenotypic traits. The rapid attainment of sexual maturity intriploid H. binoei (1–2 yr; Bustard 1970) means that the long-term influences of the incubation environment on lifetime fit-ness will be relatively easy to assess.
Acknowledgments
We thank Michael Kearney for donating the gecko colony andfor advice on husbandry, Eva Suric and Tobie Cousipectous foranimal care, Mark Blacket for genotyping the mothers, LyndalHorne and Joanne Avraam for assistance conducting some ex-periments, and Jon Green for guidance on statistics. Brian Tay-lor is gratefully acknowledged for building the incubators andtemperature-control devices for these experiments. We alsothank Ray Huey and an anonymous reviewer for their helpfulcomments on the manuscript. S.J.A. was supported by an Aus-tralian Postgraduate Award and N.J.M. was the recipient of aLa Trobe University Postdoctoral Fellowship. Experiments wereapproved by the La Trobe University Animal Ethics Committee(AEC 99/42[L]V17), and the gecko colonies were maintainedunder permits 10003633 and 10003634 from the Victorian De-partment of Sustainability and Environment. S.J.A. and N.J.Mcontributed equally to this study.
Literature Cited
Andrews R.M., T. Mathies, and D.A. Warner. 2000. Effect ofincubation temperature on morphology, growth, and sur-vival of juvenile Sceloporus undulatus. Herpetol Monogr 14:420–431.
Arnold S.J. 1983. Morphology, performance and fitness. AmZool 23:347–361.
Ashmore G.M. and F.J. Janzen. 2003. Phenotypic variation insmooth softshell turtles (Apalone mutica) from eggs incu-bated in constant versus fluctuating temperatures. Oecologia134:182–188.
Birchard G.F. 2004. Effects of incubation temperature. Pp. 103–123 in D.C. Deeming, ed. Reptilian Incubation: Environ-ment, Evolution and Behaviour. Nottingham UniversityPress, Nottingham.
Blouin-Demers G., K.J. Kissner, and P.J. Weatherhead. 2000.Plasticity in preferred substrate temperature of young snakesin response to temperature during development. Copeia2000:841–845.
Blouin-Demers G., P.J. Weatherhead, and J.R. Row. 2004. Phe-notypic consequences of nest site selection in black rat snakes(Elaphe obseleta). Can J Zool 82:449–456.
Blumberg M.S., S.J. Lewis, and G. Sokoloff. 2002. Incubationtemperature modulates post-hatching thermoregulatory be-havior in the Madagascar ground gecko, Paroedura pictus. JExp Biol 205:2777–2784.
Bustard H.R. 1968. The ecology of the Australian gecko Het-
eronotia binoei in northern New South Wales. J Zool (Lond)156:483–497.
———. 1969. The micro-environment of a natural lizard nest.Copeia 1969:536–539.
———. 1970. The population ecology of the Australian gek-konid lizard Heteronotia binoei in an exploited forest. J Zool(Lond) 162:31–42.
Byrne M., D. K. Yeates, L. Joseph, M. Kearney, J. Bowler, M.A. J. Williams, S. Cooper, et al. 2008. Birth of a biome:insights into the assembly and maintenance of the Australianarid zone biota. Mol Ecol 17:4398–4417.
Dallwitz M.J. and J.P. Higgins. 1992. User’s guide to DEVAR:a computer program for estimating development rate as afunction of temperature. Commonwealth Scientific and In-dustrial Research Organization, Canberra.
Deeming D.C. 2004. Post-hatching phenotypic effects of in-cubation in reptiles. Pp. 229–252 in D.C. Deeming, ed. Rep-tilian Incubation: Environment, Evolution and Behaviour.Nottingham University Press, Nottingham.
Doody J.S., F. Guarino, A. Georges, B. Corey, G. Murray, andM.W. Ewert. 2006. Nest site choice compensates for climateeffects on sex ratios in a lizard with environmental sex de-termination. Evol Ecol Res 20:307–330.
Du W. and J. Feng. 2008 Phenotypic effects of thermal meanand fluctuations on embryonic development and hatchlingtraits in a lacertid lizard, Takydromus serpentrionalis. J ExpZool A 309:139–146.
Du W. and X. Ji. 2003. The effects of incubation thermal en-vironments on size, locomotor performance and early growthof hatchling soft-shelled turtles, Pelodiscus sinensis. J ThermBiol 28:279–286.
Du W., L. Shou, J. Shen, and Y. Lu. 2005. Influence of fluc-tuating incubation temperatures on hatchling traits in a Chi-nese skink, Eumeces chinensis. Herpetol J 15:139–142.
Dunson W.A. 1982. Low water vapor conductance of hard-shelled eggs of the gecko lizards Hemidactylus and Lepido-dactylus. J Exp Zool 219:377–379.
Dybdahl M.F. and S. L. Kane. 2005. Adaptation vs. phenotypicplasticity in the success of a clonal invader. Ecology 86:1592–1601.
Dzialowski E.M. and P.R. Sotherland. 2004. Maternal effects ofegg size on emu Dromaius novaehollandiae egg compositionand hatchling phenotype. J Exp Biol 207:597–606.
Frappell P.B., C. Lanthier, R.V. Baudinette, and J.P. Mortola.1992. Metabolism and ventilation in acute hypoxia: a com-parative analysis in small mammalian species. Am J Physiol262:R1040–R1046.
Frappell P.B. and J.P. Mortola. 2000. Respiratory function in anewborn marsupial with skin gas exchange. Respir Physiol120:35–45.
Georges A., K. Beggs, J.E. Young, and J.S. Doody. 2005. Mod-elling development of reptile embryos under fluctuating tem-perature regimes. Physiol Biochem Zool 78:18–30.
Georges A., C. Limpus, and R. Stoutjesdijk. 1994. Hatchlingsex in the marine turtle Caretta caretta is determined by
This content downloaded on Thu, 17 Jan 2013 23:34:36 PMAll use subject to JSTOR Terms and Conditions

606 S. J. Andrewartha, N. J. Mitchell, and P. B. Frappell
proportion of development at a temperature, not daily du-ration of exposure. J Exp Zool 270:432–444.
Gruber M.A.M. 2007. Conservation of Tuatara (Sphenodon):An Evaluation of the Survival and Growth of ArtificiallyIncubated, Head-Started Juveniles. BS thesis. Victoria Uni-versity of Wellington.
Jayne B.C. and A.F. Bennett. 1990. Scaling of speed and en-durance in garter snakes: a comparison of cross-sectionaland longitudinal allometries. J Zool (Lond) 220:257–277.
Jones J.S., J.A. Coyne, and L. Partridge. 1987. Estimation ofthe thermal niche of Drosophila melanogaster using atemperature-sensitive mutation. Am Nat 130:83–90.
Kearney M., M.J. Blacket, J.L. Strasburg, and C. Moritz. 2006.Waves of parthenogenesis in the desert: evidence for theparallel loss of sex in a grasshopper and a gecko from Aus-tralia. Mol Ecol 15:1743–1748.
Kearney M. and R. Shine. 2004a. Developmental success, sta-bility, and plasticity in closely related parthenogenetic andsexual lizards (Heternotia, Gekkonidae). Evolution 58:1560–1572.
———. 2004b. Morphological and physiological correlates ofhybrid parthenogenesis. Am Nat 164:803–813.
———. 2005. Lower fecundity in parthenogenetic geckos thansexual relatives in the Australian arid zone. J Evol Biol 18:609–618.
Kearney M., R. Wahl, and K. Autumn. 2005. Increased capacityfor sustained locomotion at low temperature in partheno-genetic geckos of hybrid origin. Physiol Biochem Zool 78:316–324.
Kingsolver J.G. and R.B. Huey. 1998. Evolutionary analyses ofmorphological and physiological plasticity in thermally var-iable environments. Am Zool 38:545–560.
———. 2008. Size, temperature, and fitness: three rules. EvolEcol Res 10:251–268.
Krosniunas E.H. and G.E. Gerstner. 2003. A model of vertebrateresting metabolic rate: balancing energetics and O2 transportin system design. Respir Physiol Neurobiol 134:93–113.
Lessells, C.M., and P.T. Boag. 1987. Unrepeatable repeatabilities:a common mistake. Auk 104:116–121.
Lovegrove B.G. 2006. The power of fitness in mammals: per-ceptions from the African slipstream. Physiol Biochem Zool79:224–236.
Martin T.L. and R.B. Huey. 2008. Why “suboptimal” is optimal:Jensen’s inequality and ectotherm thermal preferences. AmNat 171:E102–E118.
Mitchell N.J., M.R. Kearney, N.J. Nelson, and W.P. Porter. 2008.Predicting the fate of a living fossil: how will global warmingaffect sex determination and hatching phenology in tuatara?Proc R Soc B 275:2185–2193.
Moritz C. 1983. Parthenogenesis in the endemic Australianlizard Heteronotia binoei (Gekkonidae). Science 220:735–737.
Mousseau T.A. and C.W. Fox, eds. 1998. Maternal Effects asAdaptations. Oxford University Press, New York.
Mullins M.A. and F.J. Janzen. 2006. Phenotypic effects of ther-mal means and variances on smooth softshell turtle (Apalonemutica) embryos and hatchlings. Herpetologica 62:27–36.
Nelson N., M. Thompson, S. Pledger, S. Keall, and C. Daugh-erty. 2004. Egg mass determines hatchling size, and incu-bation temperature influences post-hatching growth of tu-atara, Sphenodon punctatus. J Zool (Lond) 263:1–11.
———. 2006. Performance of juvenile tuatara depends on age,clutch, and incubation regime. J Herpetol 40:399–403.
Olson C.R., C.M. Vleck, and D. Vleck. 2006. Periodic coolingof bird eggs reduces embryonic growth efficiency. PhysiolBiochem Zool 79:927–936.
O’Steen S. 1998. Embryonic temperature influences juveniletemperature choice and growth rate in snapping turtles Che-lydra serpentina. J Exp Biol 201:439–449.
Packard G.C. and M.J. Packard. 1988. The physiological ecologyof reptilian eggs and embryos. Pp. 523–605 in C. Gans andR.B. Huey, eds. The Biology of the Reptilia. Vol. 16. Liss,Washington.
Pala I., M.M. Coelho, and M. Schartl. 2008. Dosage compen-sation by gene-copy silencing in a triploid hybrid fish. CurrBiol 18:1344–1348.
Pieau C. 1996. Temperature variation and sex determinationin reptiles. BioEssays 18:19–26.
Price T., A. Qvarnstrom, and D. Irwin. 2003. The role of phe-notypic plasticity in driving genetic evolution. Proc R Soc B270:1433–1440.
Quinn G.P. and M.J. Keogh. 2002. Experimental Design andData Analysis for Biologists. Cambridge University Press,Cambridge.
Scheiner S.M. 1993. Genetics and evolution of phenotypic plas-ticity. Annu Rev Ecol Syst 24:35–68.
Shine R. 2004a. Adaptive consequences of developmental plas-ticity. Pp. 187–210 in D.C. Deeming, ed. Reptilian Incuba-tion: Environment, Evolution and Behaviour. NottinghamUniversity Press, Nottingham.
———. 2004b. Seasonal shifts in nest temperature can modifythe phenotypes of hatchling lizards, regardless of overallmean incubation temperature. Funct Ecol 18:43–49.
Shine R., M.J. Elphick, and P.S. Harlow. 1997. The influenceof natural incubation environments on the phenotypic traitsof hatchling lizards. Ecology 78:2559–2568.
Shine R. and P.S. Harlow. 1996. Maternal manipulation of off-spring phenotypes via nest-site selection in an oviparous liz-ard. Ecology 77:1808–1817.
Sinervo B. 1990. The evolution of maternal investment in liz-ards: an experimental and comparative analysis of egg sizeand its effects on offspring performance. Evolution 44:279–294.
Steyermark A.C. 2002. A high standard metabolic rate con-strains juvenile growth. Zoology 105:147–151.
Strasburg J.L. 2004. Eight highly polymorphic microsatellite locifor the Australian gecko Heteronotia binoei. Mol Ecol Notes4:456–458.
Strasburg J.L. and M. Kearney. 2005. Phylogeography of sexualHeteronotia binoei (Gekkonidae) in the Australian arid zone:climatic cycling and repetitive hybridization. Mol Ecol 14:2755–2772.
Strasburg J.L., M. Kearney, C. Moritz, and A.R. Templeton.
This content downloaded on Thu, 17 Jan 2013 23:34:36 PMAll use subject to JSTOR Terms and Conditions

Incubation of Clonal Geckos at Fluctuating Temperatures 607
2007. Comparing phylogeography with distribution mod-eling: multiple Pleistocene range expansions in a parthe-nogenetic gecko from the Australian arid zone. PLoS ONE8:e760.
Webb J.K., G.P. Brown, and R. Shine. 2001. Body size, locomotor
speed and antipredator behaviour in a tropical snake (Tropi-donophis mairii, Colubridae): the influence of incubation en-vironments and genetic factors. Funct Ecol 15:561–569.
White M.J.D. 1978. Modes of Speciation. W.H. Freeman, SanFrancisco.
This content downloaded on Thu, 17 Jan 2013 23:34:36 PMAll use subject to JSTOR Terms and Conditions




















