
The Epistemological Grounds of the Conceptual Profile Theory

OPINION PAPER
The importance of morphodynamic processes at riffles usedas spawning grounds during the incubation time of nase(Chondrostoma nasus)
Christoph Hauer Æ Gunther Unfer ÆStefan Schmutz Æ Helmut Habersack
Received: 8 March 2006 / Revised: 23 August 2006 / Accepted: 15 September 2006 /Published online: 9 December 2006� Springer Science+Business Media B.V. 2006
Abstract An ecologically orientated flood pro-
tection project was implemented at the Austrian
lowland Sulm River. Habitat modelling was
conducted during a 3-year monitoring program
to assess the effects of river bed embankment
and the initiation of a new meander by con-
structing an initial side channel, and allowing
self-developing side erosion. Hydrodynamic and
physical habitat models were combined with
statistical methods. This study focused on the
necessity for including stability analysis when
modelling spawning grounds. The critical erosion
parameter of the cobbles at spawning grounds
was analysed using numerical modelling com-
bined with substrate maps of potential spawning
grounds of nase (Chondrostoma nasus). Due to
the specific characteristics of this reach of the
Sulm River, instability of the riffles can hamper
successful reproduction in nase. Spawning
grounds of Chondrostoma nasus and their sta-
bility must be guaranteed at least for the
duration of incubation. This morphodynamic
necessity should be incorporated in future res-
toration projects and when artificially restoring
spawning habitats.
Keywords Habitat modelling � Restoration
measures � River monitoring � Chondrostoma
nasus � Riffles � Spawning habitats
Introduction
Cyprinid habitat modelling studies are rare, but
even fewer studies have described the quantita-
tive effects of morphodynamic processes on
spawning ground quality. Habitat modelling has
generally focused on stable channels (Holzer
et al., 2002; Brodeur et al., 2004; Pasternack
et al., 2004; Gard, 2005), and omitted the effects
of sediment transport and stability of potential
spawning grounds. Knowledge of other species
requirements are more complete; for example,
the parameters of seasonal timing and homing
for salmonid spawning behaviour (Baril &
Magnan, 2002), and the physical characteristics
Handling editor: D. Ryder
C. Hauer � G. Unfer � S. Schmutz � H. HabersackDepartment of Water, Atmosphere and Environment,BOKU – University of Natural Resources & AppliedLife Sciences Vienna, Vienna, Austria
C. Hauer (&) � H. HabersackInstitute of Water Management, Hydrology andHydraulic Engineering, Muthgasse 18, 1190 Vienna,Austriae-mail: christoph.hauer @boku.ac.at
G. Unfer � S. SchmutzInstitute of Hydrobiology and Aquatic EcosystemManagement, Max-Emanuellstrasse 17, A-1180 Wien,Austria
Hydrobiologia (2007) 579:15–27
DOI 10.1007/s10750-006-0406-7
123

of successful spawning grounds (Ottaway et al.,
1981; Heggberget et al., 1988; Grost et al., 1991;
Elliot, 1994; Darchambeau & Poncin, 1997; Rubin
et al., 2005). In the case of cyprinid fish, however,
most studies deal with physiological and time-
related observations of the spawning act (Baras,
1994, 1995; Rinchard & Kestemont, 1996; Fred-
rich et al., 2003). Relatively few studies describe
the abiotic characteristics of the spawning habi-
tats (Keckeis, 1991; Maier et al., 1992; Stein, 1992;
Melcher, 1999).
The occurrence of riffles, which are used as
spawning grounds by hiders (salmonids) and
stickers (rheophilic cyprinids), was investigated
using a broad database relating to spatial scale
including bedform type and frequency by Emery
et al. (2004) and Sear (1996). Further studies on
historical river morphology maps were deemed
necessary for understanding river dynamics and
as a tool for successful restoration measures
(Hudson, 2002; Hohensinner et al., 2004). Previ-
ous studies have dealt with the geomorphologic
and hydraulic effects of such riffle restorations
(Pretty et al., 2003; Walker et al., 2004; Brooks
et al., 2005), including the movement and stability
of riffles (Wilkinson et al., 2004). Beyond such
morphodynamic processes, Kappesser et al.
(2002) defined a Riffle Stability Index (RSI)
which estimates the mobility of cobbles in riffles.
The effects of restored riffles as spawning
grounds, in accordance with the European Water
Framework Directive, have been discussed in the
scientific literature less for rheophilic cyprinids
than for salmonids (Rubin et al., 2004; Zeh &
Donni, 1994).
The present study therefore highlights the
needs of numerical models for morphodynamic
processes in restoration ecology and specifically
when dealing with spawning grounds of rheo-
philic cyprinids. Riffle sections play a major role
for reproduction of rheophilic cyprinids. Their
spawning grounds are characterized by adhesive
eggs on the bed sediments. For nase, we
hypothesised that the erosion of sediment
particles of the 2–6.3 cm size class at riffle
sections could mechanically damage the affixed
Chondrostoma nasus eggs and reduce spawning
success.
Methods
Study reach
The study reach is situated in the southern-east
part of Austria (–62950, 5181450; Gauss–Kruger/
national grid Austria). The Sulm River is formed
by two main tributaries originating from the
Central Alps and discharges into the Mur River,
a further tributary to the Danube. The biocenotic
description is defined as epipotamal (Muhar et al.,
1998) and barbel-region (Illies & Botoseanu,
1963). The riparian vegetation is defined based
on the main species by Otto (1981) as Salicetum
cinereae, Fraxino-Populetum, Salicetum albea for-
ests. After the confluence of the two tributaries
from the Central Alps, the Sulm River historically
resembled a meandering river (Muhar et al.,
1998). The bankfull discharge of the historical
Sulm was 40–45 m3 s–1 with an average slope of
0.0003 m m–1 (Habersack & Hauer, 2004). River
bed straightening in 1965 led to a reduction in
river sinuosity, with the slope increased to
0.001 m m–1 by meander cut-offs (Habersack &
Hauer, 2004). Within a further flood protection
project in 1999, the river channel was widened,
leading to an increased discharge profile of
80 m3 s–1. During the river widening and revital-
ization project, two small meanders were con-
structed. One of the meanders (Fig. 1) at river
station 12.960–13.356 km was modelled to analyse
the stability of the spawning habitats of the
rheophilic cyprinid nase (Fig. 1). The regulated
river bed was cut off and serves as a backwater in
low-flow situations. When the discharge increases
to more than 10 m3 s–1, the former river bed
functions as a flood channel (Fig. 1). This study
reach is located approximately 12.8 km upstream
of the mouth into the Mur River. Hydrological
data for the monitoring area were provided by the
Regional government of Styria (2001) and are
depicted in Table 1. The flow regime is indicated
as a pluvio-nivale 3 (i.e., the occurrence of three
equivalent flood peaks per annum is characteris-
tic; Mader et al., 1996).
Additionally, the Pielach River is presented
based on the fact that the main parameters
(velocity, depth and substrate) for the study were
16 Hydrobiologia (2007) 579:15–27
123
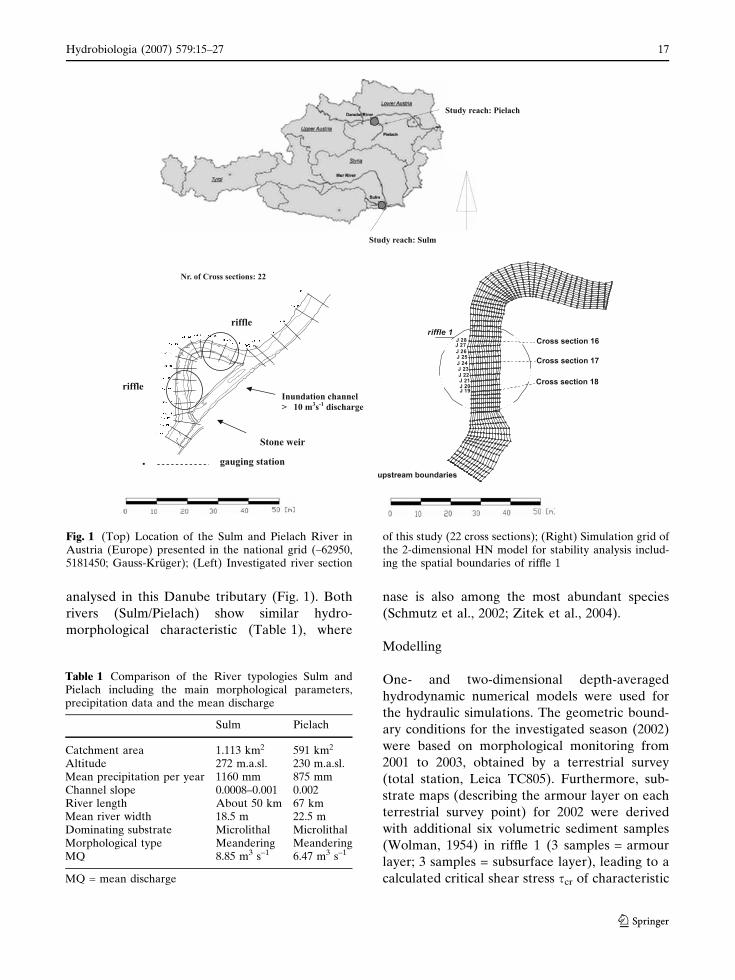
analysed in this Danube tributary (Fig. 1). Both
rivers (Sulm/Pielach) show similar hydro-
morphological characteristic (Table 1), where
nase is also among the most abundant species
(Schmutz et al., 2002; Zitek et al., 2004).
Modelling
One- and two-dimensional depth-averaged
hydrodynamic numerical models were used for
the hydraulic simulations. The geometric bound-
ary conditions for the investigated season (2002)
were based on morphological monitoring from
2001 to 2003, obtained by a terrestrial survey
(total station, Leica TC805). Furthermore, sub-
strate maps (describing the armour layer on each
terrestrial survey point) for 2002 were derived
with additional six volumetric sediment samples
(Wolman, 1954) in riffle 1 (3 samples = armour
layer; 3 samples = subsurface layer), leading to a
calculated critical shear stress scr of characteristic
Nr. of Cross sections: 22
upstream boundaries
J 19
J 20
J 21
J 23
J 22
J 24
J 25
J 26
J 27
J 28
Cross section 18
Cross section 17
Cross section 16
riffle 1
Inundation channel
> 10 m3s-1 discharge
Stone weir
riffle
riffle
. gauging station
Study reach: Pielach
Study reach: Sulm
Fig. 1 (Top) Location of the Sulm and Pielach River inAustria (Europe) presented in the national grid (–62950,5181450; Gauss-Kruger); (Left) Investigated river section
of this study (22 cross sections); (Right) Simulation grid ofthe 2-dimensional HN model for stability analysis includ-ing the spatial boundaries of riffle 1
Table 1 Comparison of the River typologies Sulm andPielach including the main morphological parameters,precipitation data and the mean discharge
Sulm Pielach
Catchment area 1.113 km2 591 km2
Altitude 272 m.a.sl. 230 m.a.sl.Mean precipitation per year 1160 mm 875 mmChannel slope 0.0008–0.001 0.002River length About 50 km 67 kmMean river width 18.5 m 22.5 mDominating substrate Microlithal MicrolithalMorphological type Meandering MeanderingMQ 8.85 m3 s–1 6.47 m3 s–1
MQ = mean discharge
Hydrobiologia (2007) 579:15–27 17
123
grain diameters with the formula of Meyer-Peter
& Muller (1949) in the selected riffles (Fig. 1).
scr ¼ 0:047 � ðqF � qWÞ � g � dm ð1Þ
where qF = density of sediment (2,665 kg m–3),
qW = density of water (1,000 kg m–3), dm = aver-
age diameter of sediment (m), g = gravity force
(9.81 m s–1).
Habitat modelling
To define the potential spawning grounds, a
third specific model (CASIMIR) was imple-
mented in the monitoring process. This model
has no separate module to calculate the hydrau-
lic conditions (Jorde, 1999). The geometric
boundaries and the one-dimensional (1D) sim-
ulated water surface were therefore necessary to
initiate the habitat modelling. To obtain the
two-dimensional velocity distribution from the
average value in the cross section, the habitat
model uses the formula of Darcy–Weissbach
(Schneider, 2001).
vm ¼1ffiffiffi
kp �
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
8 � g � Rhy � IE
p
ð2Þ
where vm = average velocity (m s–1), k = resis-
tance coefficient of Darcy–Weisbach (–), g =
gravity force (9.81 m s–1), Rhy = hydraulic radius
(m), IE = energy slope (m).
A possibility of using this equation for the
description of local currents consists of dividing
the wetted area in strips. This organization
becomes already pre-determined by the selected
vertical-gradual space resolution z which was
defined as 0.03 m.
mi ¼1ffiffiffiffi
ki
p �ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
8 � g � hi � IE
p
ð3Þ
To link the hydraulic simulation results with
suitability indices, the method of Bovee (1986)
was used. This method of multiplying suitabil-
ity indices was also utilized in the PHAB-
SIM model (Milhouse, 1989) and is shown in
Eq. 4.
(a) (b)
(c)
Station (m)
276
270271272273274275
0 10 20
)m(
noitavelE
8Trap Nr. 7 6 5 4 3 2 1
0
5
10
15
20
25
30
35
40
0246810121416182022
Cross sections
Wid
th [
m]
200120022003
Meander
-10
0
10
20
30
40
50
60
22 20 18 16 14 12 10 8 6 4 2
Cross sectionsagg
rad
atio
n /
ero
sio
n [
cm]
Morphological changes [cm]
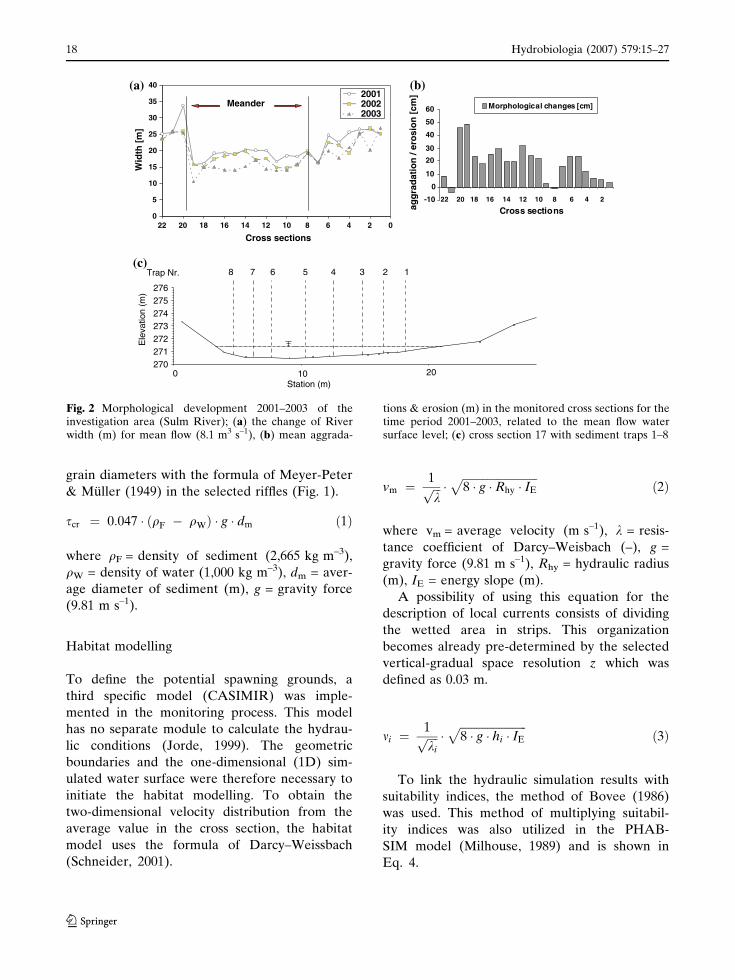
Fig. 2 Morphological development 2001–2003 of theinvestigation area (Sulm River); (a) the change of Riverwidth (m) for mean flow (8.1 m3 s–1), (b) mean aggrada-
tions & erosion (m) in the monitored cross sections for thetime period 2001–2003, related to the mean flow watersurface level; (c) cross section 17 with sediment traps 1–8
18 Hydrobiologia (2007) 579:15–27
123

SIges ¼ SId �SIv �SICI or general SIges ¼Y
I
i¼1
SIi
ð4Þ
where SId = Suitability Index depth, SIv = Suit-
ability Index velocity, SICl = Suitability Index
channel index, SIges = Suitability Index total,
SIi = Suitability index variable.
Snorkelling observations on the habitat use of
the spawning ground by nase in the Pielach River
in 1997 were used to generate suitability indices
for this age-class (after Melcher, 1999; Table 2).
This function of multiplying suitability indices
allowed the areas of potential spawning grounds
in the 2002 season to be defined. The decision
to apply this method (multiplying suitability
indices) was based on the habitat demands of
this specific life-stage being well documented for
three parameters: velocity, depth and substrate
(Melcher, 1999). Moreover, the PHABSIM
method (Bovee, 1986) was an adequate tool
for predicting fish habitat. A number of studies
(Shuler & Nehring, 1993; Elliot et al., 1996;
Harby & Arnekleiv, 1994; Huusko & Yrjana,
1996) have used PHABSIM to evaluate the
effectiveness of stream restoration projects post-
construction. To gain further quantitative mod-
elling results, the method of weighted usable
area (WUA) (Bovee, 1986) was implemented in
the modeling process (Eq. 4).
WUA ¼X
n
i¼1
HSIi:Ai ð5Þ
where n = total number of grid cells (–),
HSIi = habitat suitability index (–), Ai = area of
single grid cells (m2).
Shear stress modelling
For simulating the stability of the potential
spawning grounds, the depth-averaged two-
dimensional hydrodynamic model was used. The
model was developed by Nujic (1999) and calcu-
lates the hydraulic conditions on a linear grid by a
finite volume approach. The convective flow is
based on the Upwind-scheme by Pironneau
(1989) and the discretisation of time is done by
an explicit Runge Kutta method in second order.
The shear stress was calculated for each node of
the grid with the following formula (Nujic, 2004):
s ¼ q � g � h � Ir ð6Þ
Ir ¼v2
k2str � h
43
ð7Þ
where q = density of water, v = velocity at node
(m s–1), kstr = roughness number by Strickler (1/
Manning n), h = water depth (m), g = gravity
force (9.81 m s–1).
The simulations were performed with the
parabolic eddy viscosity model and a constant
turbulent viscosity coefficient of 0.6.
Validation of initiation of sediment transport
For validating the simulated initiation of motions
of bedload transport, eight sediment traps were
installed in the river bed at cross section 17
(Fig. 1). The hydraulic efficiency of the traps
(diameter 0.4 m; depth 0.25 m) was analysed in
the laboratory. Several cases of filling 30, 60 and
90% were investigated by vertical velocity distri-
bution. The P.-EMS, developed by DELFT-
Hydraulics, was used to measure flow velocities.
Table 2 Suitability Indices (SI) of Chondrostoma nasus which were developed by Melcher (1999) by snorkelling at thePielach River
SI Very high High Low No SI
Water depth (cm) 15–30 30–45 45–90 <15, >90Current velocity (cm s–1) 100–110 80–120 70–80/130–140 <70, >140Substrate Microlithal Microlithal mesolithal pelal
Ad. Table 1: pelal (<0.063 cm), psammal (0.063–0.2 cm), akal (0.2–2 cm), microlithal (2–6.3 cm), mesolithal (6.3–20 cm),macrolithal (20–40 cm), megalithal >40 cm//very high (SI: 1–0.8), high (SI: 0.8–0.4), low (SI: 0.4–>0), no SI (SI: 0)
Hydrobiologia (2007) 579:15–27 19
123

The P.-EMS is capable of measuring velocity
components in a 2D-plane (Delft Hydraulics,
2006). Data were transported to data-acquisition
equipment (LabVIEW). The P.-EMS allows flow
velocity measuring within a range of 0–2.5 m s–1
(optional 0–5.0 m s–1) bi-axial in a four quadrant
range. Verticals were predefined according to
stationing of the installed sediment traps and
were probed by seven point sampling measures,
with main focus on the near-bottom flow pro-
cesses.
Hydrologic data
Hydrologic data for modelling the spawning
period of nase were taken from the gauge
Heimschuh, which was installed on a bridge
300 m upstream of the study area (–63067/
5181216; Gauss Kruger/national grid Austria).
The data were analysed for a two-month period in
2002 and 2001. The decision to run the modelling
with a 2-month time series was based on existing
correlations between spawning and water tem-
perature (Lelek & Penaz, 1963; Maier, 1993).
Depending on the weather conditions, spawning
in the study region can take place in April or
May. The hydrologic gauging data for 2001 were
also analysed for the defined time period to use a
different discharge data set for further interpre-
tation of possible erosion processes over spawn-
ing grounds.
Results
The calculation of the critical shear stress by the
formula of Meyer-Peter & Muller (1949) yielded
a certain classification of critical shear stress
values for the different sediment size classes
(Table 3). As the main available choritope type
in the study riffles the microlithal size class was
documented. The critical shear stress for this
substrate classification is defined between
15.21 N m–2 and 47.92 N m–2 (Table 3). Further
the small existing sediment size classes in the
possible spawning grounds akal (1.51–15.21 N m–2)
and mesolithal (47.92–152 N m–2) are listed.
In a second step, potential spawning grounds
were defined by analysing suitability curves of
nase (Melcher, 1999) (Table 2). The investigation
area features two riffles (Fig. 3) which are
suitable as nase spawning grounds; the use of
riffles as reproduction areas has been described in
other studies (Keckeis, 1991; Stein, 1992; Maier
et al., 1992). Both riffles contain the necessary
abiotic conditions for successful spawning (Fig. 3,
Table 2). Riffle 1 was selected for the numerical
stability analysis. It extends from nearby cross
sections 18 to 16 (Fig. 3).
The habitat model, which used the suitability
curves as biotic input, calculated the number
(WUAs) of potential spawning grounds of nase in
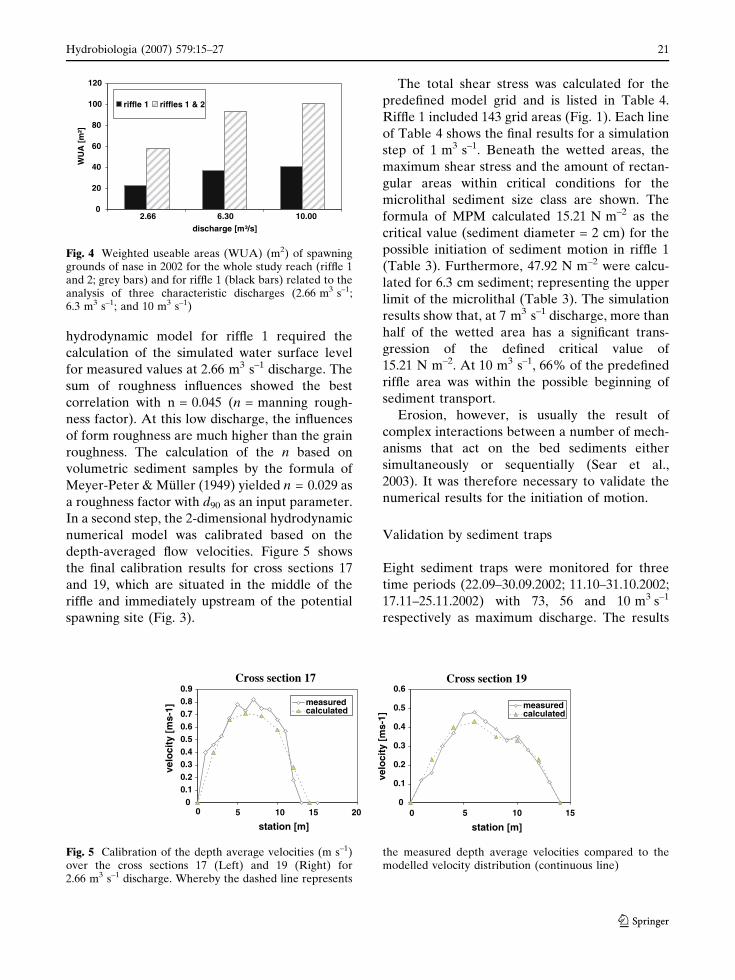
the 2002 season (Fig. 4). Figure 4 shows the total
weighted useable area of the investigation reach
and the specific values for riffle 1. The results
indicate an increase in potential spawning areas
(riffle sections) with increased discharge.
Such increases for potential spawning areas are
limited by the initiation of motion of sediment
particles. The calibration of the two-dimensional
Table 3 Critical shear stress values based on the formulaof Meyer-Peter & Muller for characteristic sediment sizediameters
Choriotope Sediment size Critical shear stress(N m–2) (after MPM)
Pelal <0.063 mm <0.48Psammal 0.063–2 mm 0.48–1.52Akal 2–20 mm 1.52–15.21Microlithal 20–63 mm 15.21–47.92Mesolithal 63–200 mm 47.92–152Macrolithal 200–400 mm 152–304.28Megalithal >400 mm >304.28
MPM: Meyer-Peter & Muller formula
269.4
269.6
269.8
270
270.2
270.4
270.6
270.8
0246810121416182022
Cross sections
]ls.a.m[
noitavele
minimum channel
water surfaceelevation
riffle
Q = 1 m3s-1
riffle 2
Fig. 3 Longitudinal profile of the study reach (SulmRiver) with potential spawning areas (riffle sections)including the bed elevation and the water surface levelfor 1 m3 s–1
20 Hydrobiologia (2007) 579:15–27
123
hydrodynamic model for riffle 1 required the
calculation of the simulated water surface level
for measured values at 2.66 m3 s–1 discharge. The
sum of roughness influences showed the best
correlation with n = 0.045 (n = manning rough-
ness factor). At this low discharge, the influences
of form roughness are much higher than the grain
roughness. The calculation of the n based on
volumetric sediment samples by the formula of
Meyer-Peter & Muller (1949) yielded n = 0.029 as
a roughness factor with d90 as an input parameter.
In a second step, the 2-dimensional hydrodynamic
numerical model was calibrated based on the
depth-averaged flow velocities. Figure 5 shows
the final calibration results for cross sections 17
and 19, which are situated in the middle of the
riffle and immediately upstream of the potential
spawning site (Fig. 3).
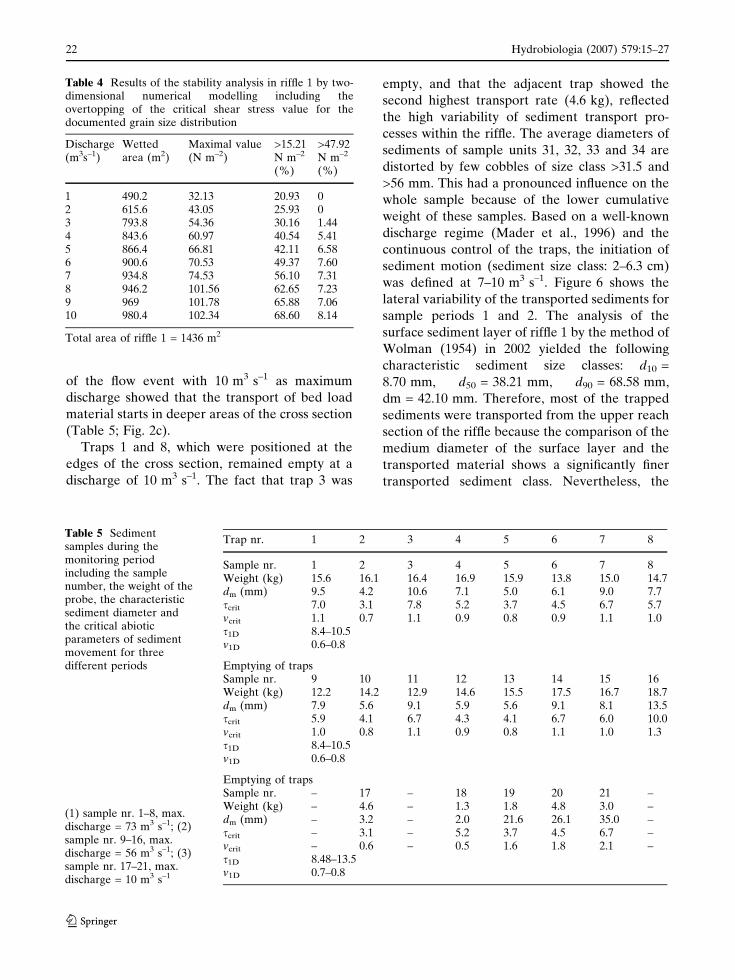
The total shear stress was calculated for the
predefined model grid and is listed in Table 4.
Riffle 1 included 143 grid areas (Fig. 1). Each line
of Table 4 shows the final results for a simulation
step of 1 m3 s–1. Beneath the wetted areas, the
maximum shear stress and the amount of rectan-
gular areas within critical conditions for the
microlithal sediment size class are shown. The
formula of MPM calculated 15.21 N m–2 as the
critical value (sediment diameter = 2 cm) for the
possible initiation of sediment motion in riffle 1
(Table 3). Furthermore, 47.92 N m–2 were calcu-
lated for 6.3 cm sediment; representing the upper
limit of the microlithal (Table 3). The simulation
results show that, at 7 m3 s–1 discharge, more than
half of the wetted area has a significant trans-
gression of the defined critical value of
15.21 N m–2. At 10 m3 s–1, 66% of the predefined
riffle area was within the possible beginning of
sediment transport.
Erosion, however, is usually the result of
complex interactions between a number of mech-
anisms that act on the bed sediments either
simultaneously or sequentially (Sear et al.,
2003). It was therefore necessary to validate the
numerical results for the initiation of motion.
Validation by sediment traps
Eight sediment traps were monitored for three
time periods (22.09–30.09.2002; 11.10–31.10.2002;
17.11–25.11.2002) with 73, 56 and 10 m3 s–1
respectively as maximum discharge. The results
0
20
40
60
80
100
120
2.66 6.30 10.00
discharge [m³/s]
]²m[
AU
W
riffle 1 riffles 1 & 2
Fig. 4 Weighted useable areas (WUA) (m2) of spawninggrounds of nase in 2002 for the whole study reach (riffle 1and 2; grey bars) and for riffle 1 (black bars) related to theanalysis of three characteristic discharges (2.66 m3 s–1;6.3 m3 s–1; and 10 m3 s–1)
Cross section 17 Cross section 19
00 05 510 1015 15 20
0.10.20.30.40.50.60.70.80.9
station [m]
velo
city
[m
s-1]
measuredcalculated
0
0.1
0.2
0.3
0.4
0.5
0.6
station [m]
]1-sm[
yticolev
measuredcalculated
Fig. 5 Calibration of the depth average velocities (m s–1)over the cross sections 17 (Left) and 19 (Right) for2.66 m3 s–1 discharge. Whereby the dashed line represents
the measured depth average velocities compared to themodelled velocity distribution (continuous line)
Hydrobiologia (2007) 579:15–27 21
123
of the flow event with 10 m3 s–1 as maximum
discharge showed that the transport of bed load
material starts in deeper areas of the cross section
(Table 5; Fig. 2c).
Traps 1 and 8, which were positioned at the
edges of the cross section, remained empty at a
discharge of 10 m3 s–1. The fact that trap 3 was
empty, and that the adjacent trap showed the
second highest transport rate (4.6 kg), reflected
the high variability of sediment transport pro-
cesses within the riffle. The average diameters of
sediments of sample units 31, 32, 33 and 34 are
distorted by few cobbles of size class >31.5 and
>56 mm. This had a pronounced influence on the
whole sample because of the lower cumulative
weight of these samples. Based on a well-known
discharge regime (Mader et al., 1996) and the
continuous control of the traps, the initiation of
sediment motion (sediment size class: 2–6.3 cm)
was defined at 7–10 m3 s–1. Figure 6 shows the
lateral variability of the transported sediments for
sample periods 1 and 2. The analysis of the
surface sediment layer of riffle 1 by the method of
Wolman (1954) in 2002 yielded the following
characteristic sediment size classes: d10 =
8.70 mm, d50 = 38.21 mm, d90 = 68.58 mm,
dm = 42.10 mm. Therefore, most of the trapped
sediments were transported from the upper reach
section of the riffle because the comparison of the
medium diameter of the surface layer and the
transported material shows a significantly finer
transported sediment class. Nevertheless, the
Table 4 Results of the stability analysis in riffle 1 by two-dimensional numerical modelling including theovertopping of the critical shear stress value for thedocumented grain size distribution
Discharge(m3s–1)
Wettedarea (m2)
Maximal value(N m–2)
>15.21N m–2
(%)
>47.92N m–2
(%)
1 490.2 32.13 20.93 02 615.6 43.05 25.93 03 793.8 54.36 30.16 1.444 843.6 60.97 40.54 5.415 866.4 66.81 42.11 6.586 900.6 70.53 49.37 7.607 934.8 74.53 56.10 7.318 946.2 101.56 62.65 7.239 969 101.78 65.88 7.0610 980.4 102.34 68.60 8.14
Total area of riffle 1 = 1436 m2
Table 5 Sedimentsamples during themonitoring periodincluding the samplenumber, the weight of theprobe, the characteristicsediment diameter andthe critical abioticparameters of sedimentmovement for threedifferent periods
(1) sample nr. 1–8, max.discharge = 73 m3 s–1; (2)sample nr. 9–16, max.discharge = 56 m3 s–1; (3)sample nr. 17–21, max.discharge = 10 m3 s–1
Trap nr. 1 2 3 4 5 6 7 8
Sample nr. 1 2 3 4 5 6 7 8Weight (kg) 15.6 16.1 16.4 16.9 15.9 13.8 15.0 14.7dm (mm) 9.5 4.2 10.6 7.1 5.0 6.1 9.0 7.7scrit 7.0 3.1 7.8 5.2 3.7 4.5 6.7 5.7vcrit 1.1 0.7 1.1 0.9 0.8 0.9 1.1 1.0s1D 8.4–10.5v1D 0.6–0.8
Emptying of trapsSample nr. 9 10 11 12 13 14 15 16Weight (kg) 12.2 14.2 12.9 14.6 15.5 17.5 16.7 18.7dm (mm) 7.9 5.6 9.1 5.9 5.6 9.1 8.1 13.5scrit 5.9 4.1 6.7 4.3 4.1 6.7 6.0 10.0vcrit 1.0 0.8 1.1 0.9 0.8 1.1 1.0 1.3s1D 8.4–10.5v1D 0.6–0.8
Emptying of trapsSample nr. – 17 – 18 19 20 21 –Weight (kg) – 4.6 – 1.3 1.8 4.8 3.0 –dm (mm) – 3.2 – 2.0 21.6 26.1 35.0 –scrit – 3.1 – 5.2 3.7 4.5 6.7 –vcrit – 0.6 – 0.5 1.6 1.8 2.1 –s1D 8.48–13.5v1D 0.7–0.8
22 Hydrobiologia (2007) 579:15–27
123
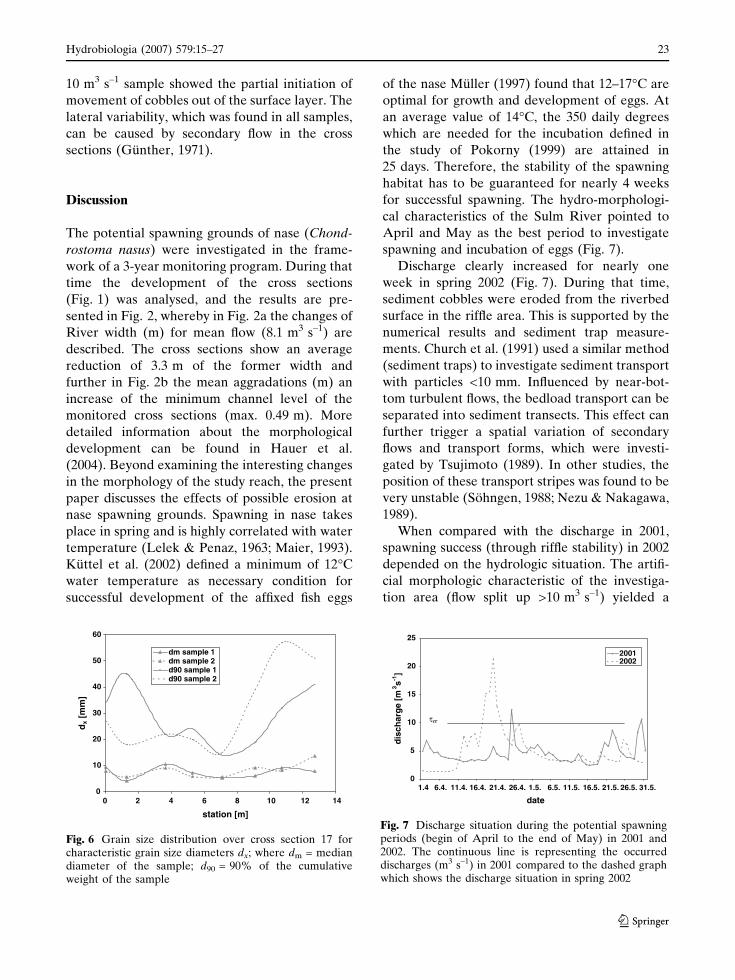
10 m3 s–1 sample showed the partial initiation of
movement of cobbles out of the surface layer. The
lateral variability, which was found in all samples,
can be caused by secondary flow in the cross
sections (Gunther, 1971).
Discussion
The potential spawning grounds of nase (Chond-
rostoma nasus) were investigated in the frame-
work of a 3-year monitoring program. During that
time the development of the cross sections
(Fig. 1) was analysed, and the results are pre-
sented in Fig. 2, whereby in Fig. 2a the changes of
River width (m) for mean flow (8.1 m3 s–1) are
described. The cross sections show an average
reduction of 3.3 m of the former width and
further in Fig. 2b the mean aggradations (m) an
increase of the minimum channel level of the
monitored cross sections (max. 0.49 m). More
detailed information about the morphological
development can be found in Hauer et al.
(2004). Beyond examining the interesting changes
in the morphology of the study reach, the present
paper discusses the effects of possible erosion at
nase spawning grounds. Spawning in nase takes
place in spring and is highly correlated with water
temperature (Lelek & Penaz, 1963; Maier, 1993).
Kuttel et al. (2002) defined a minimum of 12�C
water temperature as necessary condition for
successful development of the affixed fish eggs
of the nase Muller (1997) found that 12–17�C are
optimal for growth and development of eggs. At
an average value of 14�C, the 350 daily degrees
which are needed for the incubation defined in
the study of Pokorny (1999) are attained in
25 days. Therefore, the stability of the spawning
habitat has to be guaranteed for nearly 4 weeks
for successful spawning. The hydro-morphologi-
cal characteristics of the Sulm River pointed to
April and May as the best period to investigate
spawning and incubation of eggs (Fig. 7).
Discharge clearly increased for nearly one
week in spring 2002 (Fig. 7). During that time,
sediment cobbles were eroded from the riverbed
surface in the riffle area. This is supported by the
numerical results and sediment trap measure-
ments. Church et al. (1991) used a similar method
(sediment traps) to investigate sediment transport
with particles <10 mm. Influenced by near-bot-
tom turbulent flows, the bedload transport can be
separated into sediment transects. This effect can
further trigger a spatial variation of secondary
flows and transport forms, which were investi-
gated by Tsujimoto (1989). In other studies, the
position of these transport stripes was found to be
very unstable (Sohngen, 1988; Nezu & Nakagawa,
1989).
When compared with the discharge in 2001,
spawning success (through riffle stability) in 2002
depended on the hydrologic situation. The artifi-
cial morphologic characteristic of the investiga-
tion area (flow split up >10 m3 s–1) yielded a
00 2 4 6 8 10 12 14
10
20
30
40
50
60
station [m]
dx
]m
m[
dm sample 1dm sample 2d90 sample 1d90 sample 2
Fig. 6 Grain size distribution over cross section 17 forcharacteristic grain size diameters dx; where dm = mediandiameter of the sample; d90 = 90% of the cumulativeweight of the sample
0
5
10
15
20
25
1.4 6.4. 11.4. 16.4. 21.4. 26.4. 1.5. 6.5. 11.5. 16.5. 21.5. 26.5. 31.5.
date
m[e
grahcsi
d3 s
1 -]
20012002
τcr
Fig. 7 Discharge situation during the potential spawningperiods (begin of April to the end of May) in 2001 and2002. The continuous line is representing the occurreddischarges (m3 s–1) in 2001 compared to the dashed graphwhich shows the discharge situation in spring 2002
Hydrobiologia (2007) 579:15–27 23
123

specific morphodynamic situation for riffle 1.
During high flow events (bankfull discharge) the
meander section examined in this study acted as a
sediment trap (Fig. 2a, b), depositing material in
the riffle section. This material will be eroded at
discharges of approximately mean flow. Such
non-uniform sediment transport processes pro-
hibit the build up of a well-developed armour
layer (Parker & Sutherland, 1990). This was also
supported by the detailed documentation of the
morphological development over the monitoring
period (Hauer et al., 2004). Where they described
the development of the bottom shear stress at
bankfull discharge which was reduced from
76 N m–2 (cross section 22) to 10 N m–2 (cross
section 18) and is therefore responsible for
aggradation in the study reach. Armouring is
crucial for the stability of bed materials of aquatic
habitats in river systems (Sear et al., 2003).
Armouring greatly reduces bed material loads,
limits scour, and provides spawning sites for fish
and secure substrate for invertebrates and benthic
algae (Sear et al., 2003). These benefits to the
ecosystem are lost if the armour is removed
during a major flood, gravel mining or dredging.
In the area investigated during this study, the
artificial meander functioned as a sediment trap.
This prohibited the build-up of the necessary
armour layer in the spawning riffles. Deposition
and superposition of bed load at higher dis-
charges, caused by the reduced sediment trans-
port capacity, and erosion processes, which start
very early at mean discharge made the entire
reach very unstable for spawning of nase. Thus,
the restored river section was still morphologi-
cally dynamic with habitat changes expected
through time. This study shows that short-term
effects (3 years) of river restoration on nase
(Chondrostoma nasus) and their spawning
grounds (riffle sites) should be interpreted with
caution. When restoring spawning grounds, long-
term morphological development is an important
issue. The functionality of a restored spawning
area has to be maintained for future generations
of nase, which might carry out homing migrations
to their place of birth. Generally homing brings
an individual fish back to an environment which is
known to be suitable for reproduction (Wootton,
1992). The ability to home to particular spawning
locations is well documented for salmonid species
(e.g., Hasler & Scholz, 1983; Quinn, 1993).
Although evidence of homing during spawning
migrations in non-salmonid species is sparse
(Lucas et al., 1998), studies recently carried out
in Austria clearly supported homing migrations of
barbels and nase (Wiesner et al., 2006).
The European Water Frame Directive aims to
achieve a good ecological status for the rivers of
Member States. This requires healthy fish popu-
lation structures to guarantee sustainability for
this part of the aquatic ecosystem. Fish spawning
grounds and their stability must be guaranteed at
least for 4 weeks (Chondrostoma nasus). This
morphodynamic necessity must be incorporated
in future restoration projects and when artificially
restoring spawning grounds.
Acknowledgements The author wishes to thank DIKaroline Maierhofer and DI Novak Irene for support,DI Rudolf Hornich from the Regional Government ofStyria for funding the monitoring.
References
Baras, E., 1994. Constraints imposed by high-densities onbehavioral spawning strategies in the barbell, Barbusbarbus. Folia Zoological 43(3): 255–266.
Baras, E., 1995. Thermal related variations of seasonal anddaily spawning periodicity in barbus barbus. Journalof Fish Biology 46: 915–917.
Baril, M. & P. Magnan, 2002. Seasonal timing and dielactivity of lacustrine brook charr, Salvelinus fontinalis,spawning in a lake outlet. Environmental Biology ofFishes 64: 175–181.
Bovee, K. D., 1986. Development and evaluation ofhabitat suitability criteria for use in the instream flowincremental methology . Biological report 86, US Fishand Wildlife Service.
Brodeur, P., M. Mingelbier & J. Morin, 2004. Impact ofwater discharge on fish reproduction measured using2D numerical habitat in the St Lawrence River,Canada. Proceeding of the Fifth International Sym-posium in Ecohydraulics, 237–241.
Brooks, A. J., T. Haeusler, I. Reinfels & S. Williams, 2005.Hydraulic microhabitats and the distribution of macr-oinvertebrate assemblages in riffles. Freshwater Biol-ogy 50: 331–334.
Church, M., J. F. Wolcott & W. K. Fletcher, 1991. A test ofequal mobility in fluvial sediment transport – behav-iour of the sand fraction. Water Resources Research27: 2941–2951.
Darchambeau, F. & P. Poncin, 1997. Field observation ofthe spawning behaviour of European grayling. Journalof Fish Biology 51: 1066–1068.
24 Hydrobiologia (2007) 579:15–27
123

Delft Hydraulics, 2006. Technical Documentation of theP.-EMS. Delft Hydraulics Laboratories, NL.
Elliot, C. R. N., D. J. Willis & M. C. Acreman, 1996.Application of the physical habitat simulation(PHABSIM) model as an assessment tool for riverinehabitat restoration techniques. In Leclerc, M., et al.(eds), Ecohydraulics 2000, Proceedings of the SecondIAHR International Symposium on Habitat Hydrau-lics. Quebec, Canada. Volume B, 607–618.
Elliot, J. M., 1994. Quantitative Ecology and the BrownTrout. Oxford University Press, Oxford.
Emery, J. C., A. M. Gurnell, C. Geog, N. J. Clifford & G.E. Petts, 2004. Characteristics and controls of gravel-bed riffles: an analysis of data from the river habitatsurvey. Water and Environmental Journal 19: 210–216.
Fredrich, F., S. Ohmann, B. Curio & F. Kirschbaum, 2003.Spawning migrations of the chub in the River Spree,Germany. Journal of Fish Biology 63: 710–723.
Gard, M., 2005. Variability in flow-habitat relationships asa function of transect number for PHABSIM model-ling. River Research and Application 21: 1013–1019.
Grost, R. T., Hubert & W. A. Weschke, 1991. Descriptionof brown trout redds in a mountain stream. Transac-tion of the Atlantic Fisheries Society 120: 582–588.
Gunther, A., 1971. Die kritische mittlere Sohlschubspan-nung bei Geschiebemischungen unter Berucksichti-gung der Deckschichtbildung und derturbulenzbedingten Schubspannungsschwankungen,Mitt. Nr. 3 der VAW der ETH Zurich.
Habersack, H. & C. Hauer, 2004. Monitoring okologischorientierter Hochwasserschutzmaßnahmen an derSulm/Stmk., Arbeitspakete Flussmorphologie undHochwasserschutz. Studie im Auftrag der Steiermar-kischen Landesregierung, Graz und des Bundesmin-isterium fur Land und Forstwirtschaft, Umwelt undWasserwirtschaft, Wien. .
Harby, A. & J. V. Arnekleiv, 1994. Biotope improvementanalysis in the River Dallaa with the River SystemSimulator. In Proceedings of the first InternationalSymposium on Habitat Hydraulics. Trondheim, Nor-way, 619–630.
Hasler, A. D. & A. T. Scholz, 1983. Olfactory Imprintingand Homing in Salmon. Springer Verlag.
Hauer, C., H. Habersack, S. Schmutz, G. Unfer, K.Maierhofer & I. Novak, 2004. The effects of morpho-dynamic processes on the habitat quality of therheophilous cyprinid nase (Chondrostoma nasus) ina restored Austrian lowland river. Proceedings of theFifth International Conference on Ecohydraulics -Aquatic Habitats, 798–802.
Heggberget, T. G., L. P. Hansen & T. F. Naesje, 1988.Within-River spawning migration of Atlantic Salmon(Salmo salar). Canadian Journal of fisheries andAquatic sciences 45: 1691–1698.
Hohensinner, S., H. Habersack, G. Zauner & M. Jung-wirth, 2004. Reconstruction of the characteristics of anatural alluvial river-floodplain system and hydro-morphological changes following human modifica-tions: The Danube River (1812–1991). RiverResearch and Applications 20: 25–41.
Holzer, G., A. Muller, A. Peter & M. Schneider, 2002.Fischereiliches Gutachten uber die Aarebaggerung inThun. Kastanienbaum, EAWAG.
Hudson, P. F., 2002. Pool-riffle morphology in an activelymigrating alluvial channel: the Lower MississippiRiver. Physical Geography 23: 154–169.
Huusko, A. & T. Yrjana, 1996. Effects of instreamenhancement structures on brown trout habitat avail-ability in a channelized boreal river: a PHABSIM –approach. In Leclerc, M., et al. (eds), Ecohydraulics2000, Proceedings of the Second IAHR InternationalSymposium on Habitat Hydraulics. Quebec, Canada.Volume B, 619–630.
Illies, J. & L. Botoseanu, 1963. Problemes et methodes dela classification et de la zonation ecologique des eauxcourantes, considerees surtout du point de vue faun-istique. Mitteilungen. Internationale Vereinigung furTheoretische und Angewandte Limnologie 12: 1–57.
Jorde, K., 1999. Das Simulationsmodell CASIMIR alsHilfsmittel zur Festlegung okologisch begrundeterMindestwasserregelung. Tagungsband ProblemkreisPflichtwasserabgabe, 21 – 23 Juni, Graz, SchriftreiheEuronatur.
Kappesser, G. B., 2002. A Riffle Stability Index to evaluatesediment loading to streams. Journal of the AmericanWater Resources Association 38: 1069–1081.
Keckeis, H., 1991. Fortpflanzungsbiologie und okologischeKennzeichnung von Laichgebieten der Nase (Chond-rostoma nasus) in der Donau. Workshop Biologie undGefahrdung heimischer Kleinfischarten, Innsbruck.
Kuttel, S., A. Peter & A. Wuest, 2002. Temperaturprafer-enzen und –limiten von Fischarten SchweizerischerFließgewasser. Beitrag aus dem Rhone Revital-isierungsprojekt.
Lelek, L. & M. Penaz, 1963. Spawning of Chondrostomanasus in the Brumovka River. Folia Zoologica 12:121–134.
Lucas, M. C., T. J. Thom, A. Duncan & O. Slavic, 1998.Coarse fish migration; occurrence, causes and impli-cations. R&D Technical Report W152, University ofDurham.
Mader, H., T. Steidl & R. Wimmer, 1996. Klimatologisch-hydrologische Typisierung der osterreichischen Fli-eßgewasser. Umweltbundesamt, Monographien,Wien.
Maier, K., M. Turcsany, M. Krieg & C. Tinguely, 1992.Untersuchungen an einem Laichplatz der Nase(Chondrostroma nasus) im Unterlauf der Sense(Schweiz, Kt. Bern). DGL, erweiterte Zusammenfas-sung der Jahrestagung 1992, Band 1: 258–263.
Maier, K., 1993. Erfassung und Katalogisierung der wich-tigsten Laichgebiete der Nase (Chondostroma nasus)in den schweizerischen Rheinflussen. BUWAL, Sek-tion Fischerei. Bern.
Melcher, A., 1999. Biotische Habitatmodellierung im Zugeeines Gewasserbetreuungskonzeptes anhand der Leb-ensraumanspruche der Nase (Chondrostoma nasus).Abteilung fur Hydrobiologie, BOKU Wien.
Meyer -Peter, E. & P. Muller, 1949. Formulas for bed –load transport. International Association of HydraulicResearch, 2nd Meeting, Stockholm.
Hydrobiologia (2007) 579:15–27 25
123

Milhouse, R. T., 1989. Physikal Habitat Simulation SystemReference Manual - Vers 2¢, Instream Flow Informa-tion Paper No. 26. U.S. Department of the Interior,Fish and Wildlife Service.
Muhar, S., M. Kainz, M. Kaufmann & M. Schwarz, 1998.Ausweisung flusstypspezifisch erhaltener Fließgewas-ser in Osterreich. BMLF, Wien.
Muller, R., 1997. Vorlesungsskript Fischkunde in derSchweiz 1997/98.
Nezu, I. & H. Nakagawa, 1989. Self forming mechanism oflongitudinal sand rigdes and troughs in fluvial openchannel flows. XXIII Congress. IAHR, Ottawa.
Nujic, M., 1999. Praktischer Einsatz eines hochgenauenVerfahrens fur die Berechnung von tiefengemitteltenStromungen. Mitteilungen des Institutes fur Wasser-wesen der Universitat der Bundeswehr Munchen, Nr.64.
Nujic, M., 2004. Erganzungen zu HYDRO_AS–2D, Einzweidimensionales Stromungsmodell fur die wasser-wirtschaftliche Praxis.
Ottaway, E. M., P. A. Carling, A. Clare & N. A. Reader,1981. Observations on the structure of brown trout,Salmo trutta Linnaeus, redds. Journal of Fish Biology19: 593–607.
Otto, H., 1981. Auwalder im steirischen Mur und Ra-abgebiet im Rahmen der Erfassung SchutzenswerterBiotope der Steiermark.- ed. Amt der Steiermarkis-chen Landesregierung, Graz, Austria maps. Flood-plain biodiversity, vegetation classification, mapping,Mur River, Raab River, floodplain restoration.
Parker, G. & A. J. Sutherland, 1990. Fluvial Armor.Journal of Hydraulic Research 28: 529–544.
Pasternack, G. B., C. L. Wang & J. E. Merz, 2004.Application of a 2D hydrodynamic model to design ofreach-scale spawning gravel replenishment on theMokelumne River, California. River Research andApplications 20: 205–225.
Petts, G. E., 1994. Impounded Rivers Perspectives forEcological Management. John Wiley and Sons, NewYork.
Pironneau, P., 1989. Finite Element Methods for Fluids.Masson, Paris.
Pokorny, B., 1999. Untersuchungen zur Drift und Habita-tauswahl der fruhen Entwicklungsstadien der NaseChondrostoma nasus an der Pielach. Diploma thesis.Institut fur Wasservorsorge, Gewasserokologie undAbfallwirtschaft, Abteilung fur Hydrobiologie, Fis-chereiwirtschaft und Aquakultur Wien.
Pretty, J. L., S. S. C. Harrison, D. J. Sheperd, C. Smith, A.G. Hildrew & R. D. Hey, 2003. River rehabilitationand fish populations: assessing the benefit of instreamstructures. Journal of Applied Ecology 40: 251–265.
Quinn, T. P., 1993. A review of homing and straying ofwild and hatchery produced salmon. Fisheries Re-search 18: 29–44.
Regional Government of Styria, 2001. Broschure, Hoch-wasserschutz Sulm – Heimschuh.
Rinchard, J. & P. Kestemont, 1996. Comparative study ofreproductive biology in single- and multiple-spawnercyprinid fish. 1. Morphological and historical features.Journal of Fish Biology 49: 883–894.
Rubin, J. F., C. Glimsater & T. Jarvi, 2004. Characteristicsand rehabilitation of the spawning habitats of the seatrout, Salmo trutta, in Gotland (Sweden). FisheriesManagement and Ecology 11: 15–22.
Rubin, J. F., C. Glimsater & T. Jarvi, 2005. Spawningcharacteristics of the anadromous brown trout in asmall Swedish stream. Journal of Fish Biology 66:107–121.
Schmutz, S., A. Zitek, S. Zobl, M. Jungwirth, N. Knopf, E.Kraus, T. Bauer & T. Kaufmann, 2002. Integratedapproach to the conservation and restoration ofDanube salmon, Hucho hucho, populations in Aus-tria. In Collares-Pereira, M. J., I. G. Cowx & M. M.Coelho (eds). Freshwater Fish Conservation – Op-tions for the Future. Fishing News Book, Oxford, 157–171.
Schneider, M., 2001. Habitat und Abflussmodellierung mitunscharfen Berechnungsansatzen. Mitteilungen desInstituts fur Wasserbau, Universitat Stuttgart, Heft108.
Sear, D. A., 1996. Sediment transport processes in pool-riffle sequences. Earth Surface and Landforms 21:241–262.
Sear, D. A., D. Malcolm, D. Newson & C. R. Thorne,2003. Guidebook of Applied Fluvial Geomorphology.R&D Technical Report FD1914.
Shuler, S. W. & R. P. Nehring, 1993. Using the physicalhabitat simulation model to evaluate a stream habitatenhancement project. River 4: 175–193.
Sohngen, B., 1988. Flussmorphologie und Stabilitat desGewasserbettes. Verkehrswasserbauliche Ausspr-achetage der BAW, Karlsruhe.
Spindler, T., 1988. Okologie der Brutfische in der Donaubei Wien. PhD, University of Vienna.
Stein, H., 1992. Fischlaichplatze an Fließgewassern, Krite-rien zur Untersuchung, Identifizierung und Bewer-tung im Rahmen von Beweissicherungen. Seminar furSachverstandige der Binnenfischerei. Bonn.
Tsujimoto, R., 1989. Longitudinal Stripes of AlternateSorting due to Cellular Secondary Currents, XXIII.Congress. IAHR, Ottawa,.
Walker, D. R., R. G. Millar & R. W. Newburry, 2004.Energy profiles across constructed riffles. Journal ofHydraulic Engineering – ASCE 130: 199–207.
Wiesner, C., M. Jungwirth, S. Schmutz, G. Unfer & A.Zitek, 2006. Importance of Connectivity in the Dan-ube River Catchment. In DWA-Deutsche Vereini-gung fur Wasserwirtschaft, Abwasser und Abfall e.V.:Internationales DWA-Symposium zur Wasserwirts-chaft – Durchgangigkeit von Gewassern fur dieaquatische Fauna, DWA-Themen, 142–150.
Wilkinson, S. N., R. J. Keller & I. D. Rutherford, 2004.Phase-shifts in shear stress as an explanation for themaintenance of pool-riffle sequences. Earth SurfaceProcesses and Landforms 29: 737–753.
Wolman, M. G., 1954. A method of sampling coarse river-bed material: transactions of the American Geophys-ical Union. U.S. Geological Survey Ask USGS, 951–956.
Wootton, J. R., 1992. The Ecology of Teleost Fishes.Chapman & Hall, London.
26 Hydrobiologia (2007) 579:15–27
123

Zeh, M., & Donni W., 1994. Restoration of spawninggrounds for trout and grayling in the River High-Rhine. Aquatic Sciences 56: 59–69.
Zitek, A., G. Unfer, C. Wiesner, D. Fleischanderl & S.Muhar, 2004. Monitoring okologisch orientierter Ho-chwasserschutzmaßnahmen. Studie im Auftrag derSteiermarkischen Landesregierung, Graz und des BMfur Land- und Forstwirtschaft, Umwelt und Wasser-wirtschaft, Wien.
Christoph Hauer is a Research ass-itant in Department of Water,Atmosphere and Environment,BOKU – University of NaturalResources & Applied Life SciencesVienna, Vienna, Austria. Hisresearch expertise in hydrodynamicnumerical modelling: Application of1D, 2D and 3D models in terms of
river restoration. Habitat modelling: Application ofhydrodynamic numerical models for habitat analysesincluding morphodynamic processes. Development of aconceptual model. Fishecological monitoring: Fish popu-lation surveys, evaluating ecological integrity based on fish.Ecomorphology: Leitbild development, monitoring ofnear-natural hydrologic engineering projects. Restorationof running waters: Theoretical concepts of running waterrestoration, methodological developments, referencedefinition and restoration evalution.
Gunther Unfer is a Research assis-tant in Department of Water,Atmosphere and Environment,BOKU – University of Natural Re-sources & Applied Life SciencesVienna, Vienna, Austria. His re-search expertise in projectmanage-
ment. Fishecology: fishmigration, population ecology,ecology of native species. Fishecological monitoring:Functionality of fish migration aids, fish population sur-verys, evaluating ecological integrity based on fish. Fish-eries: Ecologically oriented fisheries concepts, fish farming.Ecomorphology: Leitbild develompment, monitoring ofnear-natural hydrologic engineering projects. Specialmethodological expertise: Electrofihing, radio telemetry,fish tagging, fish traps, diving.
Stefan Schmutz is a Faculty memberin Department of Water, Atmo-sphere and Environment, BOKU –University of Natural Resources &Applied Life Sciences Vienna, Vien-na, Austria. His research expertise inAutecology of native fish species:
Migratory behaviour (spawning runs, migrations of juve-niles, larval and juvenile drift). Radio telemetry, devel-opment of field sampling devices (automatic driftsampler). Habitat preference (methodological develop-
ments, multivariate habitat models). Development of fish-based methods to assessing the ecological integrity of run-ning waters: Standardisation of field sampling protocols,theoretical assessment concepts, assessment criteria, fishcoenotic reference conditions, historic fish fauna, referencemodels, assessment applications, development of an Aus-trian-wide Fish and Habitat Database (HaFiDat). Resto-ration of running waters: Theoretical concepts of runningwater restoration, methodological developments, refer-ence definition, restoration evaluation. Fish-based assess-ment of human impacts in running waters: Riverengineering, fish migration barriers, water abstraction,hydro peaking, flushing of impoundments (development ofmultivariate statistical habitat models for assessing flowand physical habitat alterations). Fish migration devices:Development and evaluation of nature-like fish passes,downstream fish passage solutions. Fisheries managementof running waters: Impact of stocking on autochthonouspopulations, effects of fishing regulations.
Helmut Habersack is a Facultymember in Department of Water,Atmosphere and Environment,BOKU – University of NaturalResources & Applied Life SciencesVienna, Vienna, Austria. His re-search expertise in River morphol-ogy: Monitoring and documentationof the morphological development inAustrian rivers. Sediment transportmeasurements: Development and
application of measuring techniques for bed load andsuspended load transport. Flood analyses: Documentationand analyses of the catastrophic flood events 2002 and2005. Development of guidelines. Hydrodynamic model-ling: Application of 1D, 2D and 3D models in terms ofriver restoration. Ecomorphology: Leitbild development,monitoring of near-natural hydrologic engineering pro-jects. Restoration of running waters: Theoretical conceptsof running water restoration, methodological develop-ments, reference definition, and restoration evaluation.
Hydrobiologia (2007) 579:15–27 27
123
Copyright © 2022 FDOKUMEN




















