
Diversity Covering AMA1-MSP119 Fusion Proteins as Malaria Vaccines
13
Published Ahead of Print 19 February 2013. 2013, 81(5):1479. DOI: 10.1128/IAI.01267-12. Infect. Immun. and Clemens H. M. Kocken David R. Cavanagh, Anthony A. Holder, Alan W. Thomas Walraven, Nicole van der Werff, Marjolein van der Eijk, Roberto Rodriguez Garcia, Vanessa Riasat, Vanessa Bart W. Faber, Sumera Younis, Edmond J. Remarque, Proteins as Malaria Vaccines Fusion 19 Diversity Covering AMA1-MSP1 http://iai.asm.org/content/81/5/1479 Updated information and services can be found at: These include: SUPPLEMENTAL MATERIAL Supplemental material REFERENCES http://iai.asm.org/content/81/5/1479#ref-list-1 at: This article cites 50 articles, 16 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on June 10, 2014 by guest http://iai.asm.org/ Downloaded from on June 10, 2014 by guest http://iai.asm.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Diversity Covering AMA1-MSP119 Fusion Proteins as Malaria Vaccines

Published Ahead of Print 19 February 2013. 2013, 81(5):1479. DOI: 10.1128/IAI.01267-12. Infect. Immun.
and Clemens H. M. KockenDavid R. Cavanagh, Anthony A. Holder, Alan W. Thomas Walraven, Nicole van der Werff, Marjolein van der Eijk,Roberto Rodriguez Garcia, Vanessa Riasat, Vanessa Bart W. Faber, Sumera Younis, Edmond J. Remarque, Proteins as Malaria Vaccines
Fusion19Diversity Covering AMA1-MSP1
http://iai.asm.org/content/81/5/1479Updated information and services can be found at:
These include:
SUPPLEMENTAL MATERIAL Supplemental material
REFERENCEShttp://iai.asm.org/content/81/5/1479#ref-list-1at:
This article cites 50 articles, 16 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from
on June 10, 2014 by guest
http://iai.asm.org/
Dow
nloaded from

Diversity Covering AMA1-MSP119 Fusion Proteins as MalariaVaccines
Bart W. Faber,a Sumera Younis,a Edmond J. Remarque,a Roberto Rodriguez Garcia,a Vanessa Riasat,a Vanessa Walraven,a
Nicole van der Werff,a Marjolein van der Eijk,a David R. Cavanagh,b Anthony A. Holder,c Alan W. Thomas,a Clemens H. M. Kockena
Biomedical Primate Research Centre, Department of Parasitology, Rijswijk, The Netherlandsa; Institute of Immunology and Infection Research, University of Edinburgh,Edinburgh, United Kingdomb; Division of Parasitology, MRC National Institute for Medical Research, London, United Kingdomc
To overcome polymorphism in the malaria vaccine candidate Plasmodium falciparum apical membrane antigen 1 (PfAMA1),fusion protein chimeras comprised of three diversity-covering (DiCo) PfAMA1 molecules (D1, D2, and D3) and two allelic vari-ants of the C-terminal 19-kDa region of merozoite surface protein 1 (MSP119) (variants M1 and M2) were generated. A mixtureof fusion proteins (D1M1/D2M2D3) and the D1M1D2M2D3 fusion were compared to a single-unit mixture (D1/D2/D3/M1) inan immunological study in groups of rabbits. Following immunization, titers of antibodies (Abs) against four naturally occur-ring PfAMA1 alleles were high for all groups, as were growth inhibition assay (GIA) levels against two antigenically distinct labo-ratory parasite strains. Fusion of AMA1 to MSP119 did not suppress levels of antibodies against the AMA1 component. In addi-tion, the breadth of antibody responses was unaffected. Anti-AMA1 antibodies were largely responsible for parasite growthinhibition, as shown in reversal-of-inhibition experiments by adding competing AMA1 antigen. For all groups, titration of theMSP119 antigen into the GIA led to only a small decrease in parasite inhibition, although titers of antibodies against MSP119
were increased 15-fold for the groups immunized with fusion proteins. GIA with affinity-purified anti-MSP119 antibodiesshowed that the 50% inhibitory concentrations of the anti-MSP119 antibody preparations were in the same order of magnitudefor all animals tested, leading to the conclusion that fusing MSP119 to PfAMA1 leads to a small but significant increase in func-tional antibody levels. This study shows that combination of multiple vaccine candidates in fusion proteins may lead to im-proved characteristics of the vaccine.
Over the last few years, a reduction in the morbidity and mor-tality of malaria has been observed (1). This has been attrib-
uted to the increased efforts to implement classical methods oftransmission reduction, such as the use of insecticide-treated bednets and indoor spraying, improved diagnosis, and prompt treat-ment of malaria patients. Despite these efforts, malaria still causesat least 800,000 deaths per year (2, 3) and is still one of the dead-liest infectious diseases.
Many hurdles lie ahead in the battle against malaria, for exam-ple, the increasing parasite resistance to first-line drugs andemerging resistance to insecticides of the mosquito vectors. Addi-tional tools, such as an effective malaria vaccine, will be indispens-able to achieve the goal of local elimination (4) and subsequenteradication of malaria.
Vaccine approaches targeting various stages of the complexparasite life cycle are being investigated. Encouraging results havebeen obtained with the preerythrocytic vaccine RTS,S (5), whichhas entered phase III clinical evaluation (www.malariavaccine.org), with a reduction of severe disease in approximately 50% ofchildren. Other promising developments in this area include thepossible use of attenuated sporozoites as a vaccine (6, 7).
The two potential vaccines mentioned above are preerythro-cytic, aiming to prevent the parasite reaching full maturation inthe liver and thus providing sterile immunity. In that respect, theobserved protection against severe forms of malaria provided byRTS,S immunization is an unexpected result for which no plausi-ble explanation has been found (8). Given this moderate efficacy,the short lifetime of protection (9, 10), and the lower levels ofantibodies against blood-stage antigens in RTS,S-vaccinated chil-dren (11), combination of these preerythrocytic vaccines with
blood-stage vaccines is highly desirable, as blood-stage immunitywill further reduce the risk of complicated malarial disease.
A number of promising blood-stage vaccine candidates, manyof which have been identified as merozoite surface antigens, havebeen defined. These include merozoite surface protein 1 (MSP1),MSP2, MSP3, RESA, GLURP, EBA175, and apical membrane an-tigen 1 (AMA1). Most of these antigens have entered clinical phaseI evaluation. New, promising vaccine candidates, e.g., Rh5, arealso being pursued (12).
Both AMA1 and MSP1 are essential components of the asexualblood-stage merozoite (13, 14), while AMA1 has also been iden-tified as a sporozoite protein, possibly involved in invasion of theliver cell (15–17), while MSP1 is also produced during liver-stagedevelopment (18). The vaccine potential of both of these proteinshas recently been reviewed (19, 20). More recent data show that aheterologously produced Plasmodium knowlesi AMA1 (PkAMA1)protein vaccine can protect monkeys from otherwise lethal ma-laria challenge (21), while a clinical trial performed in Mali sug-
Received 13 November 2012 Returned for modification 9 December 2012Accepted 13 February 2013
Published ahead of print 19 February 2013
Editor: J. H. Adams
Address correspondence to Bart W. Faber, [email protected].
B.W.F., S.Y., and E.J.R. contributed equally to this article.
Supplemental material for this article may be found at http://dx.doi.org/10.1128/IAI.01267-12.
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/IAI.01267-12
May 2013 Volume 81 Number 5 Infection and Immunity p. 1479–1490 iai.asm.org 1479
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

gests that children immunized with the AMA1 protein from the3D7 line may be protected from challenge with the homologousparasite (22).
Both MSP1 and AMA1 antigens are polymorphic. MSP1 isessentially dimorphic, although some parts of this large moleculeare much more variable (23). The 19-kDa C-terminal part ofMSP1 (MSP119) that is a leading vaccine candidate is relativelyconserved, and variability is restricted to 4 to 6 amino acid resi-dues (24).
Plasmodium falciparum AMA1 (PfAMA1) is highly polymor-phic (25). Over 15% of the residues can change without having anobvious effect on function in invasion (25; E. J. Remarque, unpub-lished observations). However, with few exceptions, only one ortwo alternative amino acids are found at each of the polymorphicpositions, making the protein essentially bi- or trimorphic (26).Whether this bi- or trimorphism is the result of molecular re-straints or indicative of limited selective immune pressure on theindividual residues is currently unknown. The fact that three res-idues located at the edge of a hydrophobic cleft potentially in-volved in receptor binding have greater diversity (3, 3, and 6 vari-ants, respectively) (25, 27) points to the latter suggestion.Interestingly, most polymorphic residues are located on one faceof the molecule (27). In an attempt to overcome problems associ-ated with polymorphism in relation to vaccine development (24),we have designed three artificial PfAMA1 proteins (diversity-cov-ering [DiCo] proteins) that elicit high titers of antibodies towardmultiple P. falciparum strains when administered as a vaccinemixture in rabbits (25) and are functional, as measured by in vitrogrowth inhibition assays (GIAs). The DiCo AMA1 mixture hasbeen produced under current good manufacturing practice(cGMP), and a clinical trial (phase Ia/b) is scheduled to start at thebeginning of 2013.
A combination of multiple antigens from the same or differentstages is considered by many to be the ultimate malaria vaccine,and combination vaccines consisting of a mixture of two antigenshave entered clinical trials, for example, MSP142 and 2 alleles ofAMA1 (28). Combination of components of subunit vaccines isnot straightforward, as unwanted immunological interactions be-tween the components may occur. Anticipating these potentialproblems and at the same time minimizing the costs of produc-tion, we have combined the smallest immunologically relevantparts of AMA1 (domains I and II) and MSP119 in single-unit fu-sion proteins, showing that there are vaccine-related benefits (29).Here we have combined the DiCo strategy with the fusion strategyand developed fusion proteins comprising DiCo modules (25)and two forms of the MSP119 module. These fusions were ex-pected to induce high levels of functional anti-AMA1 and anti-MSP119 antibodies, resulting in high levels of parasite growth in-hibition in vitro. Both AMA1 and MSP119 modules were expectedto play a role in overcoming the polymorphism present in P. fal-ciparum: the DiCo proteins by their design and the MSP119 mod-ules because they cover the observed polymorphism and the mol-ecule is relatively well conserved (29).
We describe the design, production, and preliminary stabilitystudies on proteins comprised of the three DiCo proteins (de-noted modules D1, D2, and D3) (25) and two forms of MSP119
(denoted M1 and M2). Two of these, the D1M1D2M2D3 chimeraand a combination of smaller fusions (D1M1 and D2M2D3), werecompared immunologically to a mixture of separate modules (D1,D2, D3, and M1) and to the mixture of D1 and D2 to examine
whether or not addition of the third DiCo protein is needed toobtain a broad coverage of response to the polymorphism presentin PfAMA1.
MATERIALS AND METHODSParasite strains. P. falciparum lines 3D7 and FCR3 were maintained invitro using standard culture techniques in an atmosphere of 5% CO2, 5%O2, and 90% N2. Parasite line identity was verified by AMA1 gene se-quence analysis.
Construction and selection of expression clones for fusion proteins.There are two common types of (Pf)MSP119 sequence: Wellcome (M1;GenBank accession no. X02919), which is of the Q-KNG type and whichdiffers at four positions from the sequence of 3D7 (M2; GenBank acces-sion no. XM_001352134), which is of the E-TSR type. M1 also has adouble cysteine replacement, a modification used to increase immunoge-nicity (29, 30). M1 and M2 were produced according to published meth-ods (31).
Three artificial (Pf)AMA1 DiCo sequences (D1 to D3) optimized forPichia pastoris expression were purchased (DNA20, Menlo Park, CA). Thesequence coding for the AMA1 FVO prodomain (amino acids 25 to 96;GenBank accession no. AJ277646), followed by the artificial DiCo aminoacid sequences described before (amino acids 97 to 545) (25) with oneadditional change (K376R) at an identified proteolytic site (32, 33), wasexpressed in P. pastoris. The FVO AMA1 variant with this replacement(K376R) showed reduced proteolysis when expressed in P. pastoris (B. W.Faber, unpublished results).
The small fusion protein (D1M1) was generated using a PCR-basedstrategy to generate two overlapping sequences and then fuse them. Thefirst PCR used a forward primer comprising an EcoRI restriction sitesequence (with extra overhang), followed by a 20-bp overlap of the FVOAMA1 prodomain sequence (nucleotides [nt] 76 to 96); the reverseprimer comprised 15 bp complementary to the DNA sequence for the endof D1 (nt 1620 to 1635), followed by 16 bp complementary to the start ofM1, using D1 as the template. The second PCR used a forward primercomprising the last 15 bp coding for the last part of D1, followed by thefirst 16 bp coding for M1; the reverse primer comprised 16 bp comple-mentary to the end of M1 and a thymidine and two adenines to serve as astop codon, followed by the complement of an XbaI restriction site (plusextra overhang), with M1 as the template. The resulting products of thefirst and second PCRs were used in a further PCR with the forward primerof the first PCR and the reverse primer of the second PCR, to generate thefusion product.
To construct the trimer (D2M2D3), a combination of fusion PCR andmolecular cloning was used. Two smaller modules (D2M2 and M2D3)were made by fusion PCR in a fashion similar to that described for D1M1.D2M2 is a fusion product of D2 (amino acids 97 to 442) and M2 (aminoacids 1607 to 1702 of the 3D7 line) with an EcoRI restriction site at thestart of the product and a NotI restriction site at the end. The secondmodule (M2D3) is a fusion product of M2 and D3 (amino acids 106 to442) with a NotI restriction site at the start of the product and an XbaIrestriction site at the end in frame with the vector-encoded hexahistidinetag. The final product (D2M2D3) was obtained by digestion of bothsmaller clones with BtsI and XbaI, isolating the small insert from theM2D3 clone and inserting that in the D2M2 vector, also digested withBtsI/XbaI. The BtsI site is unique in the M2 module.
The pentameric fusion (D1M1D2M2D3) was generated by insertionof a D1M1 module (amino acids 97 to 442; EcoRI/NotI) into a pPicZ� Avector (Invitrogen, Leek, The Netherlands) containing the D2M2D3module in the NotI-XbaI site, starting at amino acid 106 of the D2 mod-ule. These clones were constructed with appropriate restriction sites andstarting points and were cloned in frame with the vector-encoded hexa-histidine tag.
Final constructs were used to transform Escherichia coli DH5� cells.Plasmids were isolated, and the sequences were checked (Baseclear, Le-iden, The Netherlands) and subsequently used to transform P. pastoris
Faber et al.
1480 iai.asm.org Infection and Immunity
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

KM71H following the manufacturer’s instructions. A clone positive foreach construct was selected following small-scale cultivation (10 ml). Ta-ble S1 in the supplemental material gives an overview of the amino acidsequences of the proteins used throughout this study.
Protein production. Fermentation used either 3- or 7-liter fermentors(Applikon, Schiedam, The Netherlands) with initial starting volumes of 1and 2 liters, respectively. Fermentation was at 30°C and pH 6.0, essentiallyas described in the manufacturer’s fermentation manual for P. pastorisMuts cells (Invitrogen), but with slight modifications in the air/oxygensparging regime and one-fifth of the amounts of salts being used. Duringbatch phase, air was sparged at 1 liter per minute per liter starting volumeof medium; during glycerol feed, oxygen was sparged to 20% saturation;and during methanol induction, air was sparged at 0.5 liter per min perliter starting volume, using oxygen to maintain saturation at 20%. Batchphase was normally for 24 h, followed by a fed-batch phase for 20 h andinduction for 24 h. After the induction phase, the medium was adjusted topH 7.8 and cooled to 15°C. Cells were removed by centrifugation (5,000 �g, 25 min, 4°C), and the supernatant was first filtered across a 0.22-�m-pore-size hollow-fiber filter using a QuixStand system (GE Healthcare,Diegem, Belgium) and then loaded (8 ml/min) on an immobilized metalaffinity chromatography column charged with nickel or copper (chelatingSepharose fast flow; GE Healthcare, Diegem, Belgium) equilibrated with20 mM sodium phosphate, pH 7.8, 500 mM NaCl, and 5 mM imidazole.The column was washed (20 mM sodium phosphate, pH 7.8, 500 mMNaCl, 10 mM imidazole), and then bound protein was eluted (20 mMsodium phosphate, pH 7.8, 500 mM NaCl, 100 mM imidazole). Bufferreplacement was achieved by 3 consecutive concentration-dilution steps(1:15) in phosphate-buffered saline (PBS), pH 7.4, using Amicon concen-trators with 10-kDa-cutoff filters (Millipore, Etten-Leur, The Nether-lands). Other PfAMA1 allele proteins (amino acids 25 to 545) were pro-duced according to published methods (34). Protein concentrations weredetermined using micro-bicinchoninic acid (Pierce, Rockford, IL). Thelevel of purity of the proteins and the percentage of cleavage of the pro-teins were estimated from the SDS-polyacrylamide gels (nonreduced andreduced, respectively) using a ChemiDOC densitometer with QuantityOne software (Bio-Rad, Veenendaal, The Netherlands).
Antigenicity and identification of products. The antigenicity of ex-pressed proteins was confirmed by Western blotting using rabbit serumraised against the complete FVO AMA1 ectodomain and MSP119 Well-come (29), rat monoclonal antibody (MAb) 4G2 recognizing a reduction-sensitive conserved inhibitory epitope (35) and rat MAb 58F8 recog-nizing a linear epitope at the N terminus of AMA1 (36), and the anti-MSP119 mouse MAb 12.10 that also recognizes a reduction-sensitiveepitope (37). Alkaline phosphatase (AP)-conjugated secondary antibodies(anti-rat or anti-mouse, as appropriate) were used to detect the primary an-tibodies using BCIP (5-bromo-4-chloro-3-indolylphosphate)–nitroblue tet-razolium as the substrate.
Rabbit immunizations. Rabbits were housed and immunized, andblood was sampled by Biogenes GmbH, Berlin, Germany, according tonational animal welfare regulations. Four groups of eight rabbits wereimmunized on days 0, 28, and 56 with a mixture of Pichia-expressed D1and D2 (25 �g each), a mixture of D1, D2, and D3 (17 �g each) plus 50 �gM1, a mixture of D1M1 and D2M2D3 (15 and 35 �g, respectively), or 50�g D1M1D2M2D3. The novel adjuvant CoVaccine HT (Protherics Med-icines Development Ltd., a BTG International Group Company, London,United Kingdom) was used to enhance the immune response. CoVaccineHT is an oil-in-water emulsion with sucrose fatty acid sulfate esters as theactive ingredient (38). Vaccine formulations were prepared according tothe manufacturer’s instructions. Antisera obtained 2 weeks after the thirdimmunization (day 70) were tested for reactivity by enzyme-linked im-munosorbent assay (ELISA), and functional capacity was tested by in vitroGIAs. No adverse effects of immunization were observed.
ELISA. ELISA was performed in duplicate on serum samples in 96-well flat-bottom microtiter plates (Greiner, Alphen a/d Rijn, The Nether-lands) coated with 500 ng/ml purified AMA1 or MSP119 antigen accord-
ing to published methods (25). The secondary antibody was anti-rabbitIgG conjugated to alkaline phosphatase (Pierce, Rockford, IL). Samplesfor a standard curve were included on each plate, and the titers of theexperimental samples were calculated using a four-parameter fit. Titerswere expressed either as arbitrary units (AU; 1 AU is the reciprocal dilu-tion at which an optical density of 1 is achieved) or as a protein concen-tration by including polyclonal antibody standards of known concentra-tion (39).
IgG purification. Antibodies used for growth inhibition assays were pu-rified on protein G columns (Sigma, St. Louis, MO) using standard protocols,exchanged into RPMI 1640 medium using Amicon Ultra-15 concentrators(30-kDa cutoff; Millipore), filter sterilized, and stored at �20°C until use. IgGconcentrations were determined using a NanoDrop ND-1000 spectropho-tometer (NanoDrop Technologies, Wilmington, DE).
Antigen-specific IgG was isolated on immobilized M1 and the AMA1FVO variant. The proteins were coupled to CNBr-activated Sepharose(Invitrogen, Leek, The Netherlands) according to the manufacturer’s in-structions, and then columns of these were used to isolate antigen-specificantibodies from the protein G-purified total IgG. Bound specific antibod-ies were eluted using 250 mM glycine, pH 3.0, and collected in 1-mlfractions into 1 ml 1.0 M Tris-HCl, pH 8.0, to quickly readjust the pH, andthen a buffer exchange procedure was performed.
In vitro GIA. The effect of purified IgG antibodies on parasite growthand/or erythrocyte invasion was evaluated in vitro in triplicate using 96-well flat-bottomed plates (Greiner, Alphen a/d Rijn, The Netherlands),starting with synchronized mature P. falciparum schizonts at a startingparasitemia of 0.2 to 0.4%, a hematocrit of 2.0%, and a final volume of 100�l containing 10% normal human serum and 20 �g ml�1 gentamicin inRPMI 1640. After 40 to 42 h, cells were resuspended, and 50 �l was trans-ferred into 200 �l ice-cold PBS. The cells were then centrifuged, the su-pernatant was removed, and the plates were frozen. Inhibition of parasitegrowth was estimated using the Plasmodium lactate dehydrogenase assayas previously described (15). Parasite growth inhibition, reported as apercentage, was calculated as follows: 100 � [(Aexperimental � Abackground)/(Acontrol � Abackground) � 100], where A represents the absorbance. Neg-ative-control IgG was isolated from rabbits that had been immunized withadjuvant alone. Positive-control IgG, isolated from the pooled sera of 100rabbits that had been immunized with a mixture of 7 different AMA1molecules, was used at a standard concentration that has reproduciblybeen shown to yield an inhibition of 70%, to facilitate comparisons be-tween experiments. This standard anti-AMA-1 IgG fraction (BG98) canbe obtained through the European Malaria Reference Reagent Repositoryat www.malariaresearch.eu.
Reversal of growth inhibition. AMA1 and MSP119 antigens were buf-fer exchanged against RPMI 1640 (Gibco, Paisley, United Kingdom) bymeans of a repeated concentration/dilution procedure using Amicon ul-trafiltration devices (Invitrogen, Breda, The Netherlands). Protein con-centrations were estimated by use of a NanoDrop ND-1000 spectropho-tometer. Antigens were serially diluted from 50 to 0 �g/ml and incubatedwith protein G-purified IgG elicited against the different antigen combi-nations and fusions in incomplete culture medium (total volume, 48 �l/well) for 1 h at 37°C in 96-well half-area flat-bottomed plates (Gibco). Inthe absence of the antigen, the concentration of IgG used gave 70 to 80%growth inhibition in a standard GIA.
A parasitized erythrocyte suspension was prepared and added to theplate so that the final concentration of the culture had the same para-sitemia and hematocrit levels as those in the GIA, and the assay procedurewas continued as described above.
Statistical analysis. All statistical analyses were performed using the Rstatistics package, version 2.6 (R Development Core Team [cran.r-project.org/manuals.html]).
IgG titers were compared by analysis of variance (ANOVA), usinglog-transformed IgG titers as the dependent variable and vaccine antigenas the independent variable. GIA titers were analyzed with nonlinearmixed-effect (NLME) models (23) to allow the simultaneous comparison
Diversity Covering AMA1-MSP119 Fusions
May 2013 Volume 81 Number 5 iai.asm.org 1481
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from
of GIA titers at various IgG concentrations while correcting for pseu-doreplication (22). Student’s t test was used to assess the statistical signif-icance of between-group differences. Tukey’s honest-significant-differ-ence (HSD) test was used to compare pairwise IgG titers between differentcoating antigens.
P values of �0.05 were considered statistically significant. All plotswere prepared with the R statistical package.
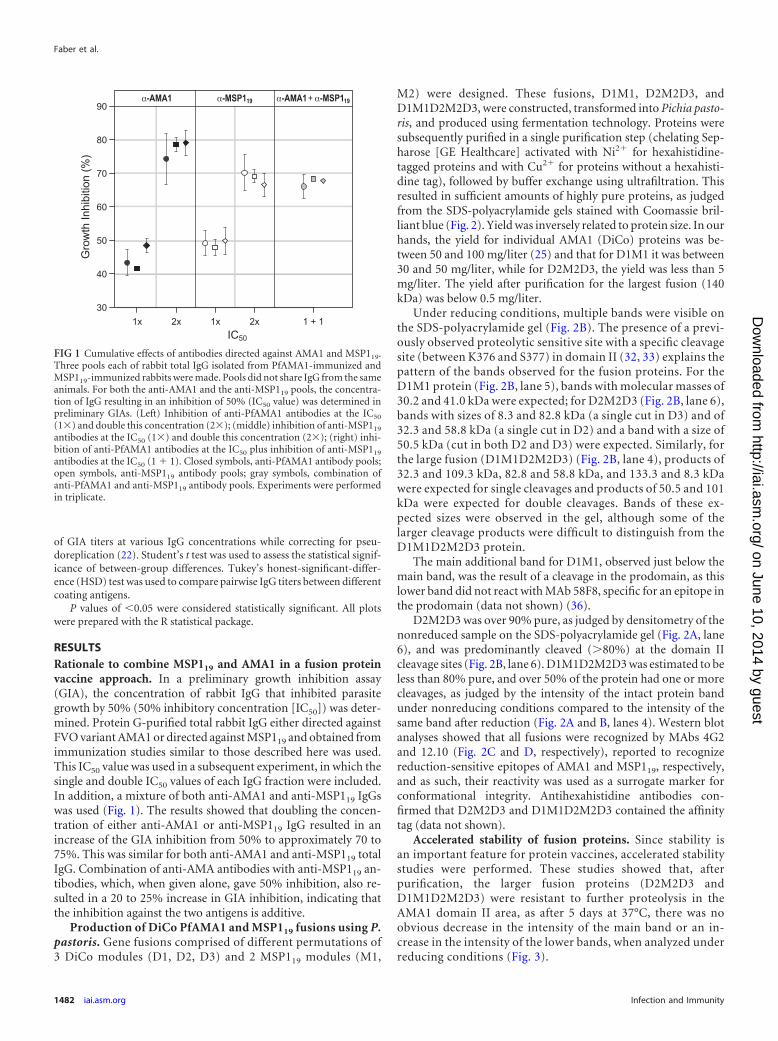
RESULTSRationale to combine MSP119 and AMA1 in a fusion proteinvaccine approach. In a preliminary growth inhibition assay(GIA), the concentration of rabbit IgG that inhibited parasitegrowth by 50% (50% inhibitory concentration [IC50]) was deter-mined. Protein G-purified total rabbit IgG either directed againstFVO variant AMA1 or directed against MSP119 and obtained fromimmunization studies similar to those described here was used.This IC50 value was used in a subsequent experiment, in which thesingle and double IC50 values of each IgG fraction were included.In addition, a mixture of both anti-AMA1 and anti-MSP119 IgGswas used (Fig. 1). The results showed that doubling the concen-tration of either anti-AMA1 or anti-MSP119 IgG resulted in anincrease of the GIA inhibition from 50% to approximately 70 to75%. This was similar for both anti-AMA1 and anti-MSP119 totalIgG. Combination of anti-AMA antibodies with anti-MSP119 an-tibodies, which, when given alone, gave 50% inhibition, also re-sulted in a 20 to 25% increase in GIA inhibition, indicating thatthe inhibition against the two antigens is additive.
Production of DiCo PfAMA1 and MSP119 fusions using P.pastoris. Gene fusions comprised of different permutations of3 DiCo modules (D1, D2, D3) and 2 MSP119 modules (M1,
M2) were designed. These fusions, D1M1, D2M2D3, andD1M1D2M2D3, were constructed, transformed into Pichia pasto-ris, and produced using fermentation technology. Proteins weresubsequently purified in a single purification step (chelating Sep-harose [GE Healthcare] activated with Ni2� for hexahistidine-tagged proteins and with Cu2� for proteins without a hexahisti-dine tag), followed by buffer exchange using ultrafiltration. Thisresulted in sufficient amounts of highly pure proteins, as judgedfrom the SDS-polyacrylamide gels stained with Coomassie bril-liant blue (Fig. 2). Yield was inversely related to protein size. In ourhands, the yield for individual AMA1 (DiCo) proteins was be-tween 50 and 100 mg/liter (25) and that for D1M1 it was between30 and 50 mg/liter, while for D2M2D3, the yield was less than 5mg/liter. The yield after purification for the largest fusion (140kDa) was below 0.5 mg/liter.
Under reducing conditions, multiple bands were visible onthe SDS-polyacrylamide gel (Fig. 2B). The presence of a previ-ously observed proteolytic sensitive site with a specific cleavagesite (between K376 and S377) in domain II (32, 33) explains thepattern of the bands observed for the fusion proteins. For theD1M1 protein (Fig. 2B, lane 5), bands with molecular masses of30.2 and 41.0 kDa were expected; for D2M2D3 (Fig. 2B, lane 6),bands with sizes of 8.3 and 82.8 kDa (a single cut in D3) and of32.3 and 58.8 kDa (a single cut in D2) and a band with a size of50.5 kDa (cut in both D2 and D3) were expected. Similarly, forthe large fusion (D1M1D2M2D3) (Fig. 2B, lane 4), products of32.3 and 109.3 kDa, 82.8 and 58.8 kDa, and 133.3 and 8.3 kDawere expected for single cleavages and products of 50.5 and 101kDa were expected for double cleavages. Bands of these ex-pected sizes were observed in the gel, although some of thelarger cleavage products were difficult to distinguish from theD1M1D2M2D3 protein.
The main additional band for D1M1, observed just below themain band, was the result of a cleavage in the prodomain, as thislower band did not react with MAb 58F8, specific for an epitope inthe prodomain (data not shown) (36).
D2M2D3 was over 90% pure, as judged by densitometry of thenonreduced sample on the SDS-polyacrylamide gel (Fig. 2A, lane6), and was predominantly cleaved (�80%) at the domain IIcleavage sites (Fig. 2B, lane 6). D1M1D2M2D3 was estimated to beless than 80% pure, and over 50% of the protein had one or morecleavages, as judged by the intensity of the intact protein bandunder nonreducing conditions compared to the intensity of thesame band after reduction (Fig. 2A and B, lanes 4). Western blotanalyses showed that all fusions were recognized by MAbs 4G2and 12.10 (Fig. 2C and D, respectively), reported to recognizereduction-sensitive epitopes of AMA1 and MSP119, respectively,and as such, their reactivity was used as a surrogate marker forconformational integrity. Antihexahistidine antibodies con-firmed that D2M2D3 and D1M1D2M2D3 contained the affinitytag (data not shown).
Accelerated stability of fusion proteins. Since stability isan important feature for protein vaccines, accelerated stabilitystudies were performed. These studies showed that, afterpurification, the larger fusion proteins (D2M2D3 andD1M1D2M2D3) were resistant to further proteolysis in theAMA1 domain II area, as after 5 days at 37°C, there was noobvious decrease in the intensity of the main band or an in-crease in the intensity of the lower bands, when analyzed underreducing conditions (Fig. 3).
FIG 1 Cumulative effects of antibodies directed against AMA1 and MSP119.Three pools each of rabbit total IgG isolated from PfAMA1-immunized andMSP119-immunized rabbits were made. Pools did not share IgG from the sameanimals. For both the anti-AMA1 and the anti-MSP119 pools, the concentra-tion of IgG resulting in an inhibition of 50% (IC50 value) was determined inpreliminary GIAs. (Left) Inhibition of anti-PfAMA1 antibodies at the IC50
(1�) and double this concentration (2�); (middle) inhibition of anti-MSP119
antibodies at the IC50 (1�) and double this concentration (2�); (right) inhi-bition of anti-PfAMA1 antibodies at the IC50 plus inhibition of anti-MSP119
antibodies at the IC50 (1 � 1). Closed symbols, anti-PfAMA1 antibody pools;open symbols, anti-MSP119 antibody pools; gray symbols, combination ofanti-PfAMA1 and anti-MSP119 antibody pools. Experiments were performedin triplicate.
Faber et al.
1482 iai.asm.org Infection and Immunity
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

The only fusion protein that contains the AMA1 FVO prodo-main, D1M1, appeared to be the least stable protein. After incu-bation at 37°C for 5 days, the prodomain was completely removed.Removal of the prodomain of the protein led to further degrada-
tion, illustrated by additional bands appearing below the first deg-radation band (Fig. 3). Overall, the stability of the purified pro-teins varied between fermentations.
Immunogenicity of AMA1-MSP119 fusion proteins. Rabbits
FIG 2 SDS-PAGE analysis of the proteins used throughout the study. (A) Nonreduced proteins (20 �g/lane); (B) reduced proteins (20 �g/lane); (C) Westernblot (nonreduced, 4 �g/lane) with rat MAb 4G2, recognizing a conserved reduction-sensitive epitope on PfAMA1; (D) Western blot (nonreduced, 4 �g/lane)with mouse MAb 12.10, recognizing a reduction-sensitive epitope on MSP119. Lanes 1, DiCo1; lanes 2, DiCo2; lanes 3, DiCo3; lanes 4, D1M1D2M2D3; lanes 5,D1M1; lanes 6, D2M2D3; lanes 7, M1. The sizes of standard marker proteins are indicated.
FIG 3 Accelerated stability study of the proteins used. Proteins were incubated for 5 days at 37°C. (A) SDS-PAGE of nonreduced proteins; (B) SDS-PAGE ofreduced proteins. Lanes 1 and 2, M1 at times zero and 5 days, respectively; lanes 3 and 4, D1M1 at times zero and 5 days, respectively; lanes 5 and 6, D2M2D3 attimes zero and 5 days, respectively; lanes 7 and 8 at D1M1D2M2D3, times zero and 5 days, respectively. Twenty micrograms of protein was used per lane. Thesizes of standard marker proteins are indicated.
Diversity Covering AMA1-MSP119 Fusions
May 2013 Volume 81 Number 5 iai.asm.org 1483
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

were immunized with mixtures of (fusion) proteins formulated inCoVaccine HT. Immunization was with a mixture of D1 and D2,a mixture of D1, D2, and D3 combined with M1, a mixture ofD1M1 and D2M2D3, and D1M1D2M2D3.
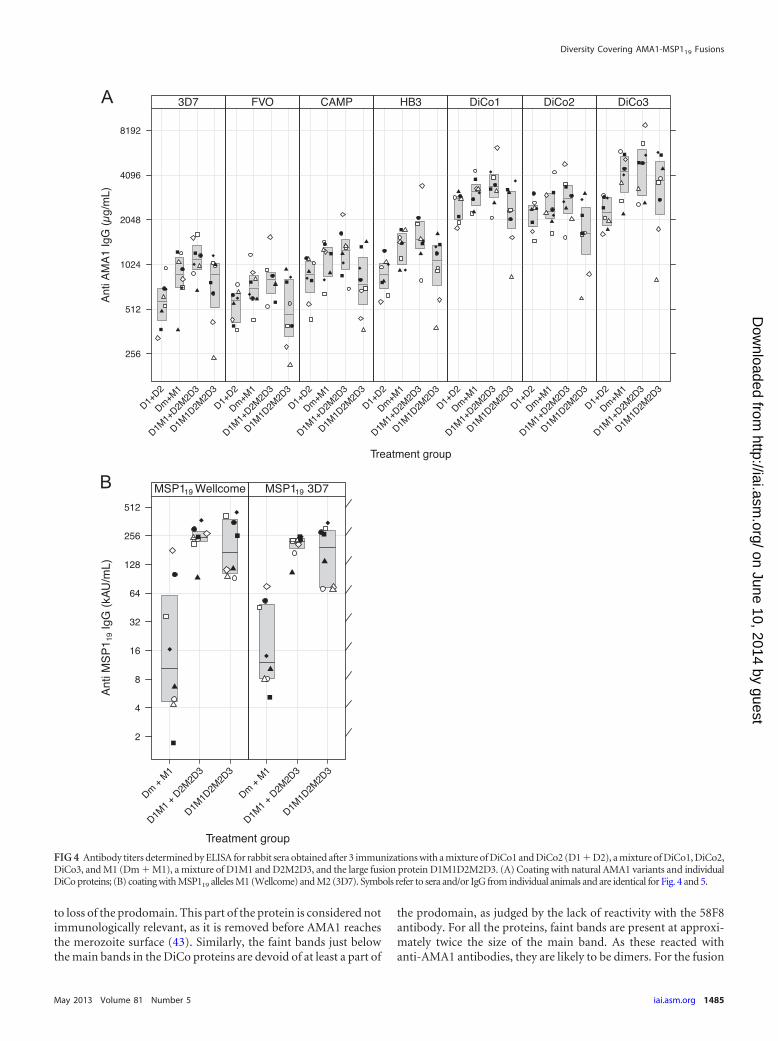
Titers of serum antibodies against AMA1 proteins (amino ac-ids 97 to 545) from the FVO, HB3, 3D7, and CAMP lines (aminoacids 25 to 545) (Fig. 4A), as well as against both MSP119 modules(Fig. 4B), were determined.
For all groups, a significantly higher titer of antibody againstthe DiCo proteins compared to that against each of the naturalalleles was observed (Student’s t test, P � 0.0001 in all cases). Thegroup immunized with the large fusion protein (D1M1D2M2D3)showed a statistically significantly lower titer of antibody againstDiCo2 than the D1M1/D2M2D3 group (P � 0.034, Tukey’s HSDtest). No statistically significant difference was observed in com-parison with the D1/D2/D3/M1 group (P � 0.11, Tukey’s HSDtest), although, on average, a 1.6-fold lower titer was observed.
The D1/D2 group had, as expected, lower titers of antibodiesagainst DiCo3 (Fig. 4A). Although the levels were 1.8- and 2.0-foldlower than those in the D1/D2/D3/M1 and D1M1/D2M2D3groups, respectively, the difference was statistically significantonly in the latter group (P � 0.024). The difference from theD1M1D2M2D3 group was lower (1.4-fold) and not statisticallysignificant. Notably, compared to the D1/D2/D3/M1 group, theD1/D2 group showed statistically significantly lower titers againstthe 3D7 and HB3 alleles. Lower titers were also observed againstthe FVO and CAMP alleles, but these were not statistically signif-icant. A similar picture emerged after comparison of the D1/D2group with the D1M1/D2M2D3 group, while no significant dif-ference was observed when the D1/D2 group was compared to theD1M1D2M2D3 group.
ELISA titers of antibodies against M1 and M2 were low for thegroup that received the D1/D2/D3/M1 mixture. No statisticallysignificant difference between the titers against M1 and M2 couldbe observed (P � 0.4705). Rabbits immunized with fusion pro-teins (D1M1/D2M2D3 or D1M1D2M2D3) showed 15- to 20-foldhigher titers of antibodies against MSP119 (P � 0.0001). Again, nostatistically significant differences between the titers against M1and M2 were observed for these groups.
Functionality of antibodies. In vitro GIAs were carried out withtwo P. falciparum strains using protein G-purified total IgG (Fig. 5).Results show that IgG isolated from rabbits immunized with the mix-ture of three DiCos and M1 (D1/D2/D3/M1) was highly inhibitoryagainst both strains. This was also evident for IgG against the combi-nation of the fusion proteins (D1M1-D2M2D3) and for the largefusion (D1M1D2M2D3). Inhibition levels at 6 mg/ml of the latterIgG were statistically significantly lower than those for the D1/D2/D3/M1 and the D1M1/D2M2D3 groups (P�0.0369 and P�0.0200,respectively) for the FCR3 strain, while they were not significantlydifferent for the 3D7 strain. Total IgG from the D1/D2 group ap-peared to have a slightly lower inhibitory capacity than total IgGsfrom the D1/D2/D3/M1 and the D1M1/D2M2D3 groups (Fig. 5),but this was not statistically significant either for the FCR3 strain orfor the 3D7 strain.
Determination of relative contribution of MSP119 andAMA1 antibodies to in vitro growth inhibition activity. Titra-tion of the AMA1 3D7 protein into GIAs executed with the 3D7parasite strain resulted in a significant reversal of inhibition for allgroups. Residual levels of inhibition at the maximum amount ofantigen used (50 �g/ml) varied between groups. For the total IgG
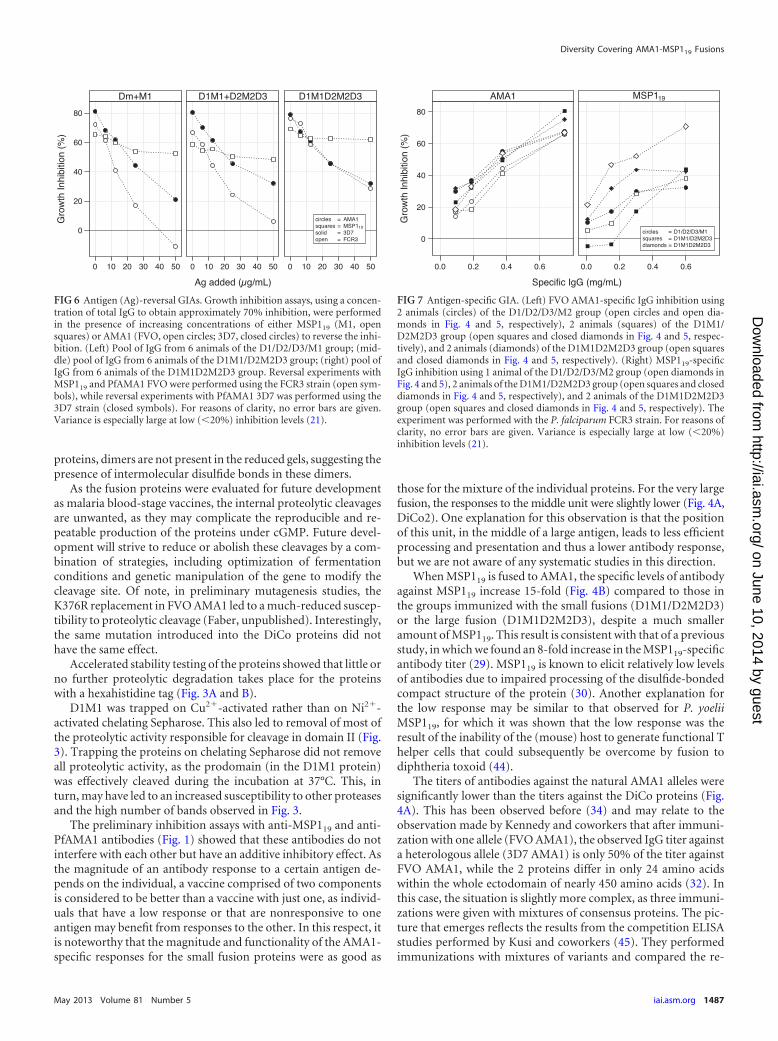
isolated from the rabbits in the D1/D2/D3/M1 group, the residualinhibition was 20% (Fig. 6). For the D1M1/D2M2D3 group andthe D1M1D2M2D3 group, the residual level was 30%.
Similar reversal-of-inhibition experiments with the FCR3strain, using FVO AMA1 as the reversing antigen (FVO AMA1 isidentical to FCR3 AMA1, apart from one amino acid in the prodo-main), resulted in complete reversal of inhibition for the D1/D2/D3/M1 group, in approximately 10% residual inhibition for theD1M1/D2M2D3 group, and in approximately 30% inhibition forthe D1M1D2M2D3 group.
Reversal of the inhibition with the MSP119 (M1, Wellcome)protein showed much higher residual inhibition levels for allgroups. Using the 3D7 strain, 50 �g/ml MSP119 reversed inhibi-tion from 65% to 52% for the IgG raised against the mixture ofDiCo proteins plus MSP119 (D1/D2/D3/M1) and from 60% to50% and 70% to 60% for the small-fusion-protein group (D1M1-D2M2D3) and the large-fusion-protein group (D1M1D2M2D3),respectively.
To confirm the contribution of anti-MSP119 antibodies to thein vitro inhibition, two IgG fractions from each immunizationgroup were used to isolate MSP119-specific antibodies. The se-lected samples were among those with the highest levels of in vitroinhibition. Isolation was done by affinity purification either onMSP119 (M1) coupled to Sepharose or on FVO AMA1 coupled toSepharose as a control. Both affinity-purified fractions were as-sessed for their inhibitory capacity in a GIA using the FCR3 strain(Fig. 7). The specific inhibitory activity of the anti-AMA1 antibod-ies was higher than that for the anti-MSP119 antibodies, illustratedby the IC50, the value at which 50% inhibition was observed. Foranti-AMA1 antibodies, the IC50 value was estimated to be 0.4mg/ml and was fairly consistent between samples. For the anti-MSP119 antibodies, the IC50 value was difficult to estimate, due tothe higher variance between the samples, but these were certainlymuch higher than the values found for anti-AMA1 antibodies.
DISCUSSION
The small fusion protein D1M1, comprised of the full-lengthectodomain of AMA1 (DiCo1) and MSP119 (Wellcome), ex-pressed reasonably well in Pichia pastoris. With approximately30 mg/liter, the yield was slightly lower than the yield of theAMA1 unit alone (33), and optimization of the fermentationconditions for D1M1 resulted in an increased yield (40). The yieldof the D1M1D2M2D3 fusion product was very low at 0.5 mg/liter,too low to produce vaccine at a large scale, for which a greater than100-fold increase in yield would need to be achieved. The yield ofthe D2M2D3 antigen/protein was also low at 5 mg/liter. Preliminaryexperiments with a P. pastoris strain overexpressing native proteindisulfide isomerase (41) have resulted in a 2.4-fold increase in thelevel of production of this protein (Faber, unpublished). This im-provement, in combination with optimization of the fermentationconditions, may lead to yields that are sufficiently high to allow cGMPproduction and phase I clinical evaluation.
The proteins were purified in a simple robust purification pro-cedure, in accordance with the methodology that was used beforefor the FVO AMA1 protein that was produced at cGMP level andsubsequently evaluated in the clinic (33, 42). The purity of theproducts was judged to be sufficient for the studies described here,as all products showed a single main band following nonreducingSDS-PAGE, with the exception of the D1M1 product with twobands, one of the expected size and one slightly smaller, likely due
Faber et al.
1484 iai.asm.org Infection and Immunity
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from
to loss of the prodomain. This part of the protein is considered notimmunologically relevant, as it is removed before AMA1 reachesthe merozoite surface (43). Similarly, the faint bands just belowthe main bands in the DiCo proteins are devoid of at least a part of
the prodomain, as judged by the lack of reactivity with the 58F8antibody. For all the proteins, faint bands are present at approxi-mately twice the size of the main band. As these reacted withanti-AMA1 antibodies, they are likely to be dimers. For the fusion
FIG 4 Antibody titers determined by ELISA for rabbit sera obtained after 3 immunizations with a mixture of DiCo1 and DiCo2 (D1 � D2), a mixture of DiCo1, DiCo2,DiCo3, and M1 (Dm � M1), a mixture of D1M1 and D2M2D3, and the large fusion protein D1M1D2M2D3. (A) Coating with natural AMA1 variants and individualDiCo proteins; (B) coating with MSP119 alleles M1 (Wellcome) and M2 (3D7). Symbols refer to sera and/or IgG from individual animals and are identical for Fig. 4 and 5.
Diversity Covering AMA1-MSP119 Fusions
May 2013 Volume 81 Number 5 iai.asm.org 1485
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from
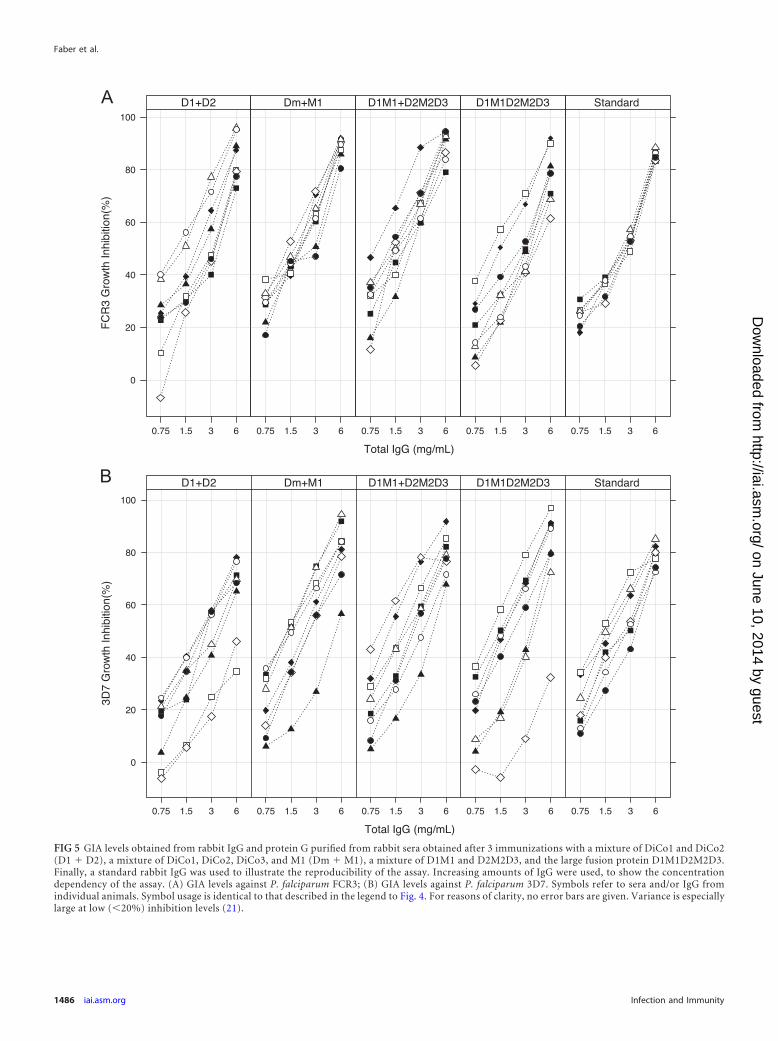
FIG 5 GIA levels obtained from rabbit IgG and protein G purified from rabbit sera obtained after 3 immunizations with a mixture of DiCo1 and DiCo2(D1 � D2), a mixture of DiCo1, DiCo2, DiCo3, and M1 (Dm � M1), a mixture of D1M1 and D2M2D3, and the large fusion protein D1M1D2M2D3.Finally, a standard rabbit IgG was used to illustrate the reproducibility of the assay. Increasing amounts of IgG were used, to show the concentrationdependency of the assay. (A) GIA levels against P. falciparum FCR3; (B) GIA levels against P. falciparum 3D7. Symbols refer to sera and/or IgG fromindividual animals. Symbol usage is identical to that described in the legend to Fig. 4. For reasons of clarity, no error bars are given. Variance is especiallylarge at low (�20%) inhibition levels (21).
Faber et al.
1486 iai.asm.org Infection and Immunity
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from
proteins, dimers are not present in the reduced gels, suggesting thepresence of intermolecular disulfide bonds in these dimers.
As the fusion proteins were evaluated for future developmentas malaria blood-stage vaccines, the internal proteolytic cleavagesare unwanted, as they may complicate the reproducible and re-peatable production of the proteins under cGMP. Future devel-opment will strive to reduce or abolish these cleavages by a com-bination of strategies, including optimization of fermentationconditions and genetic manipulation of the gene to modify thecleavage site. Of note, in preliminary mutagenesis studies, theK376R replacement in FVO AMA1 led to a much-reduced suscep-tibility to proteolytic cleavage (Faber, unpublished). Interestingly,the same mutation introduced into the DiCo proteins did nothave the same effect.
Accelerated stability testing of the proteins showed that little orno further proteolytic degradation takes place for the proteinswith a hexahistidine tag (Fig. 3A and B).
D1M1 was trapped on Cu2�-activated rather than on Ni2�-activated chelating Sepharose. This also led to removal of most ofthe proteolytic activity responsible for cleavage in domain II (Fig.3). Trapping the proteins on chelating Sepharose did not removeall proteolytic activity, as the prodomain (in the D1M1 protein)was effectively cleaved during the incubation at 37°C. This, inturn, may have led to an increased susceptibility to other proteasesand the high number of bands observed in Fig. 3.
The preliminary inhibition assays with anti-MSP119 and anti-PfAMA1 antibodies (Fig. 1) showed that these antibodies do notinterfere with each other but have an additive inhibitory effect. Asthe magnitude of an antibody response to a certain antigen de-pends on the individual, a vaccine comprised of two componentsis considered to be better than a vaccine with just one, as individ-uals that have a low response or that are nonresponsive to oneantigen may benefit from responses to the other. In this respect, itis noteworthy that the magnitude and functionality of the AMA1-specific responses for the small fusion proteins were as good as
those for the mixture of the individual proteins. For the very largefusion, the responses to the middle unit were slightly lower (Fig. 4A,DiCo2). One explanation for this observation is that the positionof this unit, in the middle of a large antigen, leads to less efficientprocessing and presentation and thus a lower antibody response,but we are not aware of any systematic studies in this direction.
When MSP119 is fused to AMA1, the specific levels of antibodyagainst MSP119 increase 15-fold (Fig. 4B) compared to those inthe groups immunized with the small fusions (D1M1/D2M2D3)or the large fusion (D1M1D2M2D3), despite a much smalleramount of MSP119. This result is consistent with that of a previousstudy, in which we found an 8-fold increase in the MSP119-specificantibody titer (29). MSP119 is known to elicit relatively low levelsof antibodies due to impaired processing of the disulfide-bondedcompact structure of the protein (30). Another explanation forthe low response may be similar to that observed for P. yoeliiMSP119, for which it was shown that the low response was theresult of the inability of the (mouse) host to generate functional Thelper cells that could subsequently be overcome by fusion todiphtheria toxoid (44).
The titers of antibodies against the natural AMA1 alleles weresignificantly lower than the titers against the DiCo proteins (Fig.4A). This has been observed before (34) and may relate to theobservation made by Kennedy and coworkers that after immuni-zation with one allele (FVO AMA1), the observed IgG titer againsta heterologous allele (3D7 AMA1) is only 50% of the titer againstFVO AMA1, while the 2 proteins differ in only 24 amino acidswithin the whole ectodomain of nearly 450 amino acids (32). Inthis case, the situation is slightly more complex, as three immuni-zations were given with mixtures of consensus proteins. The pic-ture that emerges reflects the results from the competition ELISAstudies performed by Kusi and coworkers (45). They performedimmunizations with mixtures of variants and compared the re-
FIG 6 Antigen (Ag)-reversal GIAs. Growth inhibition assays, using a concen-tration of total IgG to obtain approximately 70% inhibition, were performedin the presence of increasing concentrations of either MSP119 (M1, opensquares) or AMA1 (FVO, open circles; 3D7, closed circles) to reverse the inhi-bition. (Left) Pool of IgG from 6 animals of the D1/D2/D3/M1 group; (mid-dle) pool of IgG from 6 animals of the D1M1/D2M2D3 group; (right) pool ofIgG from 6 animals of the D1M1D2M2D3 group. Reversal experiments withMSP119 and PfAMA1 FVO were performed using the FCR3 strain (open sym-bols), while reversal experiments with PfAMA1 3D7 was performed using the3D7 strain (closed symbols). For reasons of clarity, no error bars are given.Variance is especially large at low (�20%) inhibition levels (21).
FIG 7 Antigen-specific GIA. (Left) FVO AMA1-specific IgG inhibition using2 animals (circles) of the D1/D2/D3/M2 group (open circles and open dia-monds in Fig. 4 and 5, respectively), 2 animals (squares) of the D1M1/D2M2D3 group (open squares and closed diamonds in Fig. 4 and 5, respec-tively), and 2 animals (diamonds) of the D1M1D2M2D3 group (open squaresand closed diamonds in Fig. 4 and 5, respectively). (Right) MSP119-specificIgG inhibition using 1 animal of the D1/D2/D3/M2 group (open diamonds inFig. 4 and 5), 2 animals of the D1M1/D2M2D3 group (open squares and closeddiamonds in Fig. 4 and 5, respectively), and 2 animals of the D1M1D2M2D3group (open squares and closed diamonds in Fig. 4 and 5, respectively). Theexperiment was performed with the P. falciparum FCR3 strain. For reasons ofclarity, no error bars are given. Variance is especially large at low (�20%)inhibition levels (21).
Diversity Covering AMA1-MSP119 Fusions
May 2013 Volume 81 Number 5 iai.asm.org 1487
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

sponses to these to the responses to sequential immunization withdifferent single variants. This led, in both cases, to a strong re-sponse to the epitopes that are shared between the proteins usedfor immunization. This could be explained by assuming that themore often that an epitope is “seen” by the immune system, thestronger that the response to that epitope will be. The observationthat the natural AMA1 alleles induce a lower response than theindividual DiCo proteins is in agreement with this explanation, asthe conserved or shared epitopes of the DiCo proteins have beenseen at least three times. For the natural alleles, the number of(specific) epitopes that have been seen will inevitably be lower,leading to lower antibody titers overall.
In general, the GIA inhibition by total IgG obtained after im-munization with a mixture of antigens (D1/D2/D3/M1) was sim-ilar to that obtained after immunization with the small-fusion-protein mixture (D1M1/D2M2D3) and the large fusion protein(D1M1D2M2D3). However, IgG against the latter showed slightlylower inhibition of the FVO strain, while no significant differencewas observed for the 3D7 strain. As AMA1 FVO differs in only 9amino acids from the DiCo2 protein (25), the lower titer of anti-body against the DiCo2 allele may explain this observation.
From the reversal-of-inhibition studies (Fig. 6), it is clear thatthe major part of the inhibition observed in the GIA can be attrib-uted to anti-AMA1 antibodies in all groups. Although not com-pletely saturated, the antibody-mediated inhibition falls, in a con-centration-dependent way, to a very low level after addition of 50�g/ml AMA1. Due to the relatively high variance at low inhibitionlevels in this assay (21), it was not possible to accurately determinethe level of the residual activity attributable to anti-MSP119 anti-bodies. On the other hand, the reversal of inhibition by titration ofMSP119 was very low and saturated at 25 �g/ml. It should be notedthat the relation between the concentration of IgG and the level ofinhibition is not linear. For assays where inhibition was between20% and 80%, each doubling of the IgG concentration resulted ina 20% increase in inhibition (Fig. 1). If this relationship holds true,then the reversal of inhibition by MSP119 from 70% to �60%represents a significant concentration of IgG that equates with anestimated level of inhibition of between 15 and 30% in the absenceof anti-AMA1 antibodies. The residual levels of inhibition for theFVO and 3D7 strains of 0% and 20%, respectively, for the D1/D2/D3/M2 group, 10% and 30%, respectively, for the D1M2-DM2D3group, and 30% and 30%, respectively, for the D1M2DM2D3group are over similar ranges. There seemed to be a trend towardslightly higher residual inhibition in the groups containing fusionproteins (middle and right-hand panels versus left-hand panel inFig. 6); however, the 15-fold difference in anti-MSP119 antibodytiter between the groups is not clearly visible from the reversal-of-inhibition experiments.
The GIAs with affinity-purified anti-MSP119 antibodies con-firm that anti-MSP119 antibodies have inhibitory activity in vitro.The IC50 value for anti-MSP119 antibodies was higher than theIC50 value of the anti-AMA1 antibodies, so more anti-MSP119
antibodies were needed to achieve similar levels of inhibition. Theobservation that there was no (large) difference between the IC50
values of the MSP119-specific antibodies derived from the differ-ent immunization groups (Fig. 7) shows that the 15-fold increaseof the antibody titer is a significant functional improvement of thevaccine.
The samples selected for the affinity purification GIAs showedhigh levels of inhibition in the assay (Fig. 7), because these were
still available after the GIA reversal experiments that had con-sumed large amounts of antibodies of the samples with low levelsof inhibition. As shown previously (46) and confirmed by the datain Fig. 4 and 5, there was a correlation between (anti-AMA1)antibody levels and GIA activity. Furthermore, the reversal-of-inhibition experiments showed that the contribution of the anti-MSP119 antibodies to overall inhibition was not high. Therefore,the chance that the other animals from these groups would haveantibody profiles that differ substantially from the ones describedis low, although it cannot be formally excluded.
Analysis of the ELISA titers against the different AMA1 allelesshowed significantly lower titers against two of these variants(3D7, HB3) for the D1/D2 group than the D1/D2/D3/M1 group(Fig. 4A, 3D7 and HB3). Although this did not translate to statis-tically significantly lower levels of inhibition in GIA, we concludethat use of two of the DiCo proteins was insufficient to reach thesame breadth of response achieved with three DiCo proteins. Thisis consistent with the results obtained in studies where naturalalleles were combined (32, 46). Including both variants of MSP119
in the vaccine does not lead to an improved immunogenicity pro-file with respect to coverage. However, if the DiCo principle ap-plies, use of a combination of two or more engineered MSP119
modules may still be beneficial, as the combination could targetthe immune response to epitopes of choice, enabling the fine spec-ificity that is deemed necessary to make an MSP119-based vaccinework (47–49).
In previous studies (25, 46), we have shown that purified IgGisolated from rabbits immunized with a mixture of three DiCoAMA1 proteins, to overcome polymorphism in AMA1, has anenhanced capacity to inhibit different laboratory strains of P. fal-ciparum in an in vitro growth inhibition assay. This study showsthat small fusion proteins with MSP119 and DiCo modules (i.e.,D1M1 and D2M2D3) do not compromise this enhanced capacity.With very large fusions, e.g., D1M1D2M2D3, this capacity is ap-parently compromised, as responses toward the central compo-nent were diminished.
We have previously shown that there are clear benefits fromuse of the combination of AMA1 (FVO) and MSP119 antigens in asingle polypeptide (29). In the current study, we have shown thatfusion of MSP119 to DiCo modules leads to 15-fold higherMSP119-specific antibody titers (Fig. 4B) with a specific inhibitorycapacity (IC50 value) (Fig. 7) similar to that achieved with theantibody induced by a separate MSP119 antigen, although the lev-els of IgG were still low compared with those elicited against theAMA1 component.
It has recently been shown that antibodies against MSP119 mayact via a different mechanism than antibodies against AMA1: fol-lowing internalization of the antibody into the freshly invadedcell, it can prevent development of the parasite (50). Moreover,anti-MSP119 antibodies have been implicated in antibody-depen-dent cell-mediated immunity (51), implying that part of the po-tential inhibitory capacity of the anti-MSP119 antibodies is notmeasured in the GIA, since immune cells are absent in this assay.The antibodies may also have complement-dependent activities.
In summary, these studies show that the combination of thesmaller fusion proteins (D1M1 and D2M2D3) makes a potentmalaria vaccine candidate. We expect that it will be feasible toproduce these proteins in sufficient quantity and quality undercGMP, especially since we recently confirmed that the hexahisti-dine tag containing the D1M1 product has a satisfactory acceler-
Faber et al.
1488 iai.asm.org Infection and Immunity
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

ated stability profile (Faber, unpublished). The immunologicaldata presented here show that there is added value in the combi-nation of these fusion proteins over that of the single AMA1 andMSP119 components as well as over that of the existing DiCo pro-tein mixtures currently under development.
ACKNOWLEDGMENTS
Funding was provided by EMVDA grant LSHP-CT-2007-037506,E(M)VI, and BPRC. Work in the A. A. Holder laboratory was funded bythe United Kingdom MRC (U117532067).
We thankfully acknowledge Henk van Westbroek, who created thefigures.
REFERENCES1. Mendis K, Rietveld A, Warsame M, Bosman A, Greenwood B, Werns-
dorfer WH. 2009. From malaria control to eradication: the WHO per-spective. Trop. Med. Int. Health 14:802– 809.
2. World Health Organization. 2010. World malaria report: 2010. WorldHealth Organization, Geneva, Switzerland.
3. Murray CJ, Rosenfeld LC, Lim SS, Andrews KG, Foreman KJ,Haring D, Fullman N, Naghavi M, Lozano R, Lopez AD. 2012.Global malaria mortality between 1980 and 2010: a systematic analysis.Lancet 379:413– 431.
4. Bhattarai A, Ali AS, Kachur SP, Martensson A, Abbas AK, Khatib R,Al-Mafazy AW, Ramsan M, Rotllant G, Gerstenmaier JF, Molteni F,Abdulla S, Montgomery SM, Kaneko A, Bjorkman A. 2007. Impact ofartemisinin-based combination therapy and insecticide-treated nets onmalaria burden in Zanzibar. PLoS Med. 4:e309. doi:10.1371/journal.pmed.0040309.
5. Gordon DM, McGovern TW, Krzych U, Cohen JC, Schneider I, La-Chance R, Heppner DG, Yuan G, Hollingdale M, Slaoui M, Hauser P,Voet P, Sadoff JC, Ballou WR. 1995. Safety, immunogenicity, and effi-cacy of a recombinantly produced Plasmodium falciparum circumsporo-zoite protein-hepatitis B surface antigen subunit vaccine. J. Infect. Dis.171:1576 –1585.
6. Mueller AK, Labaied M, Kappe SH, Matuschewski K. 2005. Geneticallymodified Plasmodium parasites as a protective experimental malaria vac-cine. Nature 433:164 –167.
7. Hoffman SL, Billingsley PF, James E, Richman A, Loyevsky M, Li T,Chakravarty S, Gunasekera A, Li M, Stafford R, Ahumada A, EpsteinJE, Sedegah M, Reyes S, Richie TL, Lyke KE, Edelman R, Laurens M,Plowe CV, Sim BK. 2010. Development of a metabolically active, non-replicating sporozoite vaccine to prevent Plasmodium falciparum ma-laria. Hum. Vaccin. 6:97–106.
8. Agnandji ST, Lell B, Soulanoudjingar SS, Fernandes JF, Abossolo BP,Conzelmann C, Methogo BG, Doucka Y, Flamen A, Mordmuller B,Issifou S, Kremsner PG, Sacarlal J, Aide P, Lanaspa M, Aponte JJ,Nhamuave A, Quelhas D, Bassat Q, Mandjate S, Macete E, Alonso P,Abdulla S, Salim N, Juma O, Shomari M, Shubis K, Machera F, HamadAS, Minja R, Mtoro A, Sykes A, Ahmed S, Urassa AM, Ali AM,Mwangoka G, Tanner M, Tinto H, D’Alessandro U, Sorgho H, Valea I,Tahita MC, Kabore W, Ouedraogo S, Sandrine Y, Guiguemde RT,Ouedraogo JB, Hamel MJ, Kariuki S, Odero C, et al. 2011. First resultsof phase 3 trial of RTS,S/AS01 malaria vaccine in African children. N. Engl.J. Med. 365:1863–1875.
9. Alonso PL, Sacarlal J, Aponte JJ, Leach A, Macete E, Aide P, SigauqueB, Milman J, Mandomando I, Bassat Q, Guinovart C, Espasa M,Corachan S, Lievens M, Navia MM, Dubois MC, Menendez C,Dubovsky F, Cohen J, Thompson R, Ballou WR. 2005. Duration ofprotection with RTS,S/AS02A malaria vaccine in prevention of Plasmo-dium falciparum disease in Mozambican children: single-blind extendedfollow-up of a randomised controlled trial. Lancet 366:2012–2018.
10. Scheller LF, Azad AF. 1995. Maintenance of protective immunity againstmalaria by persistent hepatic parasites derived from irradiated sporozo-ites. Proc. Natl. Acad. Sci. U. S. A. 92:4066 – 4068.
11. Bejon P, Cook J, Bergmann-Leitner E, Olotu A, Lusingu J, MwacharoJ, Vekemans J, Njuguna P, Leach A, Lievens M, Dutta S, von SeidleinL, Savarese B, Villafana T, Lemnge MM, Cohen J, Marsh K, Corran PH,Angov E, Riley EM, Drakeley CJ. 2011. Effect of the pre-erythrocyticcandidate malaria vaccine RTS,S/AS01E on blood stage immunity inyoung children. J. Infect. Dis. 204:9 –18.
12. Douglas AD, Williams AR, Illingworth JJ, Kamuyu G, Biswas S, Good-man AL, Wyllie DH, Crosnier C, Miura K, Wright GJ, Long CA, OsierFH, Marsh K, Turner AV, Hill AV, Draper SJ. 2011. The blood-stagemalaria antigen PfRH5 is susceptible to vaccine-inducible cross-strainneutralizing antibody. Nat. Commun. 2:601. doi:10.1038/ncomms1615.
13. Triglia T, Healer J, Caruana SR, Hodder AN, Anders RF, Crabb BS,Cowman AF. 2000. Apical membrane antigen 1 plays a central role inerythrocyte invasion by Plasmodium species. Mol. Microbiol. 38:706 –718.
14. Sanders PR, Kats LM, Drew DR, O’Donnell RA, O’Neill M, Maier AG,Coppel RL, Crabb BS. 2006. A set of glycosylphosphatidyl inositol-anchored membrane proteins of Plasmodium falciparum is refractory togenetic deletion. Infect. Immun. 74:4330 – 4338.
15. Silvie O, Franetich JF, Charrin S, Mueller MS, Siau A, Bodescot M,Rubinstein E, Hannoun L, Charoenvit Y, Kocken CH, Thomas AW,Van Gemert GJ, Sauerwein RW, Blackman MJ, Anders RF, Pluschke G,Mazier D. 2004. A role for apical membrane antigen 1 during invasion ofhepatocytes by Plasmodium falciparum sporozoites. J. Biol. Chem. 279:9490 –9496.
16. Lasonder E, Ishihama Y, Andersen JS, Vermunt AM, Pain A, SauerweinRW, Eling WM, Hall N, Waters AP, Stunnenberg HG, Mann M. 2002.Analysis of the Plasmodium falciparum proteome by high-accuracy massspectrometry. Nature 419:537–542.
17. Bozdech Z, Llinas M, Pulliam BL, Wong ED, Zhu J, DeRisi JL. 2003.The transcriptome of the intraerythrocytic developmental cycle of Plas-modium falciparum. PLoS Biol. 1:E5. doi:10.1371/journal.pbio.0000005.
18. Szarfman A, Walliker D, McBride JS, Lyon JA, Quakyi IA, Carter R.1988. Allelic forms of gp195, a major blood-stage antigen of Plasmodiumfalciparum, are expressed in liver stages. J. Exp. Med. 167:231–236.
19. Remarque EJ, Faber BW, Kocken CH, Thomas AW. 2008. Apical mem-brane antigen 1: a malaria vaccine candidate in review. Trends Parasitol.24:74 – 84.
20. Holder AA. 2009. The carboxy-terminus of merozoite surface protein 1:structure, specific antibodies and immunity to malaria. Parasitology 136:1445–1456.
21. Mahdi Abdel Hamid M, Remarque EJ, van Duivenvoorde LM, van derWerff N, Walraven V, Faber BW, Kocken CH, Thomas AW. 2011.Vaccination with Plasmodium knowlesi AMA1 formulated in the noveladjuvant co-vaccine HT protects against blood-stage challenge in rhesusmacaques. PLoS One 6:e20547. doi:10.1371/journal.pone.0020547.
22. Thera MA, Doumbo OK, Coulibaly D, Laurens MB, Ouattara A, KoneAK, Guindo AB, Traore K, Traore I, Kouriba B, Diallo DA, Diarra I,Daou M, Dolo A, Tolo Y, Sissoko MS, Niangaly A, Sissoko M, Takala-Harrison S, Lyke KE, Wu Y, Blackwelder WC, Godeaux O, VekemansJ, Dubois MC, Ballou WR, Cohen J, Thompson D, Dube T, Soisson L,Diggs CL, House B, Lanar DE, Dutta S, Heppner DG, Jr, Plowe CV.2011. A field trial to assess a blood-stage malaria vaccine. N. Engl. J. Med.365:1004 –1013.
23. Tanabe K, Mackay M, Goman M, Scaife JG. 1987. Allelic dimorphism ina surface antigen gene of the malaria parasite Plasmodium falciparum. J.Mol. Biol. 195:273–287.
24. Takala SL, Plowe CV. 2009. Genetic diversity and malaria vaccine design,testing and efficacy: preventing and overcoming ‘vaccine resistant ma-laria’. Parasite Immunol. 31:560 –573.
25. Remarque EJ, Faber BW, Kocken CH, Thomas AW. 2008. A diversity-covering approach to immunization with Plasmodium falciparum apicalmembrane antigen 1 induces broader allelic recognition and growth inhi-bition responses in rabbits. Infect. Immun. 76:2660 –2670.
26. Chesne-Seck ML, Pizarro JC, Normand BV, Collins CR, BlackmanMJ, Faber BW, Remarque EJ, Kocken CH, Thomas AW, Bentley GA.2005. Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites. Mol. Biochem. Para-sitol. 144:55– 67.
27. Bai T, Becker M, Gupta A, Strike P, Murphy VJ, Anders RF, BatchelorAH. 2005. Structure of AMA1 from Plasmodium falciparum reveals aclustering of polymorphisms that surround a conserved hydrophobicpocket. Proc. Natl. Acad. Sci. U. S. A. 102:12736 –12741.
28. Ellis RD, Wu Y, Martin LB, Shaffer D, Miura K, Aebig J, Orcutt A,Rausch K, Zhu D, Mogensen A, Fay MP, Narum DL, Long C, Miller L,Durbin AP. 2012. Phase 1 study in malaria naive adults of BSAM2/Alhydrogel(R)�CPG 7909, a blood stage vaccine against P. falciparummalaria. PLoS One 7:e46094. doi:10.1371/journal.pone.0046094.
29. Faber BW, Remarque EJ, Morgan WD, Kocken CH, Holder AA,
Diversity Covering AMA1-MSP119 Fusions
May 2013 Volume 81 Number 5 iai.asm.org 1489
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

Thomas AW. 2007. Malaria vaccine-related benefits of a single proteincomprising Plasmodium falciparum apical membrane antigen 1 domainsI and II fused to a modified form of the 19-kilodalton C-terminal fragmentof merozoite surface protein 1. Infect. Immun. 75:5947–5955.
30. Hensmann M, Li C, Moss C, Lindo V, Greer F, Watts C, Ogun SA,Holder AA, Langhorne J. 2004. Disulfide bonds in merozoite surfaceprotein 1 of the malaria parasite impede efficient antigen processing andaffect the in vivo antibody response. Eur. J. Immunol. 34:639 – 648.
31. Morgan WD, Birdsall B, Frenkiel TA, Gradwell MG, Burghaus PA, SyedSE, Uthaipibull C, Holder AA, Feeney J. 1999. Solution structure of anEGF module pair from the Plasmodium falciparum merozoite surfaceprotein 1. J. Mol. Biol. 289:113–122.
32. Kennedy MC, Wang J, Zhang Y, Miles AP, Chitsaz F, Saul A, Long CA,Miller LH, Stowers AW. 2002. In vitro studies with recombinant Plas-modium falciparum apical membrane antigen 1 (AMA1): production andactivity of an AMA1 vaccine and generation of a multiallelic response.Infect. Immun. 70:6948 – 6960.
33. Faber BW, Remarque EJ, Kocken CH, Cheront P, Cingolani D, Xhon-neux F, Jurado M, Haumont M, Jepsen S, Leroy O, Thomas AW. 2008.Production, quality control, stability and pharmacotoxicity of cGMP-produced Plasmodium falciparum AMA1 FVO strain ectodomain ex-pressed in Pichia pastoris. Vaccine 26:6143– 6150.
34. Kusi KA, Faber BW, Thomas AW, Remarque EJ. 2009. Humoral im-mune response to mixed PfAMA1 alleles; multivalent PfAMA1 vaccinesinduce broad specificity. PLoS One 4:e8110. doi:10.1371/journal.pone.0008110.
35. Kocken CH, van der Wel AM, Dubbeld MA, Narum DL, van de RijkeFM, van Gemert GJ, van der Linde X, Bannister LH, Janse C, WatersAP, Thomas AW. 1998. Precise timing of expression of a Plasmodiumfalciparum-derived transgene in Plasmodium berghei is a critical determi-nant of subsequent subcellular localization. J. Biol. Chem. 273:15119 –15124.
36. Narum DL, Thomas AW. 1994. Differential localization of full-lengthand processed forms of PF83/AMA-1 an apical membrane antigen of Plas-modium falciparum merozoites. Mol. Biochem. Parasitol. 67:59 – 68.
37. McBride JS, Heidrich HG. 1987. Fragments of the polymorphic Mr185,000 glycoprotein from the surface of isolated Plasmodium falcip-arum merozoites form an antigenic complex. Mol. Biochem. Parasitol.23:71– 84.
38. Hilgers LA, Blom AG. 2006. Sucrose fatty acid sulphate esters as novelvaccine adjuvant. Vaccine 24(Suppl 2):S2– 81–S2-82.
39. Remarque EJ, Roestenberg M, Younis S, Walraven V, van der Werff N,Faber BW, Leroy O, Sauerwein R, Kocken CH, Thomas AW. 2012.Humoral immune responses to a single allele PfAMA1 vaccine in healthymalaria-naive adults. PLoS One 7:e38898. doi:10.1371/journal.pone.0038898.
40. Martens S, Borchert S-O, Faber BW, Cornelissen G, Luttmann R. 2011.Fully automated production of potential malaria vaccines with Pichia pas-toris in integrated processing. Eng. Life Sci. 11:429 – 435.
41. Tsai CW, Duggan PF, Shimp RL, Jr, Miller LH, Narum DL. 2006.Overproduction of Pichia pastoris or Plasmodium falciparum protein di-sulfide isomerase affects expression, folding and O-linked glycosylation ofa malaria vaccine candidate expressed in P. pastoris. J. Biotechnol. 121:458 – 470.
42. Roestenberg M, Remarque E, de Jonge E, Hermsen R, Blythman H,Leroy O, Imoukhuede E, Jepsen S, Ofori-Anyinam O, Faber B, KockenCH, Arnold M, Walraven V, Teelen K, Roeffen W, de Mast Q, BallouWR, Cohen J, Dubois MC, Ascarateil S, van der Ven A, Thomas A,Sauerwein R. 2008. Safety and immunogenicity of a recombinant Plas-modium falciparum AMA1 malaria vaccine adjuvanted with Alhydrogel,Montanide ISA 720 or AS02. PLoS One 3:e3960. doi:10.1371/journal.pone.0003960.
43. Howell SA, Withers-Martinez C, Kocken CH, Thomas AW, BlackmanMJ. 2001. Proteolytic processing and primary structure of Plasmodiumfalciparum apical membrane antigen-1. J. Biol. Chem. 276:31311–31320.
44. Stanisic DI, Martin LB, Liu XQ, Jackson D, Cooper J, Good MF. 2003.Analysis of immunological nonresponsiveness to the 19-kilodalton frag-ment of merozoite surface protein 1 of Plasmodium yoelii: rescue bychemical conjugation to diphtheria toxoid (DT) and enhancement of im-munogenicity by prior DT vaccination. Infect. Immun. 71:5700 –5713.
45. Kusi KA, Faber BW, van der Eijk M, Thomas AW, Kocken CH,Remarque EJ. 2011. Immunization with different PfAMA1 alleles in se-quence induces clonal imprint humoral responses that are similar to re-sponses induced by the same alleles as a vaccine cocktail in rabbits. Malar.J. 10:40. doi:10.1186/1475-2875-10-40.
46. Kusi KA, Faber BW, Riasat V, Thomas AW, Kocken CH, Remarque EJ.2010. Generation of humoral immune responses to multi-allele PfAMA1vaccines; effect of adjuvant and number of component alleles on thebreadth of response. PLoS One 5:e15391. doi:10.1371/journal.pone.0015391.
47. Corran PH, O’Donnell RA, Todd J, Uthaipibull C, Holder AA, CrabbBS, Riley EM. 2004. The fine specificity, but not the invasion inhibitoryactivity, of 19-kilodalton merozoite surface protein 1-specific antibodies isassociated with resistance to malarial parasitemia in a cross-sectional sur-vey in The Gambia. Infect. Immun. 72:6185– 6189.
48. Ngoundou-Landji J, Nwuba RI, Anumudu CI, Odaibo AB, MatondoMaya WD, Awobode HO, Okafor CM, Morenikeji OA, Asinobi A,Nwagwu M, Holder AA, Ntoumi F. 2010. Fine specificity of anti-MSP119antibodies and multiplicity of Plasmodium falciparum merozoite surfaceprotein 1 types in individuals in Nigeria with sub-microscopic infection.Malar. J. 9:287. doi:10.1186/1475-2875-9-287.
49. Okech BA, Corran PH, Todd J, Joynson-Hicks A, Uthaipibull C, Eg-wang TG, Holder AA, Riley EM. 2004. Fine specificity of serum antibod-ies to Plasmodium falciparum merozoite surface protein, PfMSP-1(19),predicts protection from malaria infection and high-density parasitemia.Infect. Immun. 72:1557–1567.
50. Arnot DE, Cavanagh DR, Remarque EJ, Creasey AM, Sowa MP, Mor-gan WD, Holder AA, Longacre S, Thomas AW. 2008. Comparativetesting of six antigen-based malaria vaccine candidates directed towardmerozoite-stage Plasmodium falciparum. Clin. Vaccine Immunol. 15:1345–1355.
51. McIntosh RS, Shi J, Jennings RM, Chappel JC, de Koning-Ward TF,Smith T, Green J, van Egmond M, Leusen JH, Lazarou M, van deWinkel J, Jones TS, Crabb BS, Holder AA, Pleass RJ. 2007. The impor-tance of human FcgammaRI in mediating protection to malaria. PLoSPathog. 3:e72. doi:10.1371/journal.ppat.0030072.
Faber et al.
1490 iai.asm.org Infection and Immunity
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from




















