
Diversity and antimicrobial activities of the fungal endophyte community associated with the...
13
Diversity and antimicrobial activities of the fungal endophyte community associated with the traditional Brazilian medicinal plant Solanum cernuum Vell. (Solanaceae) Mariana L.A. Vieira, Alice F.S. Hughes, Viviane B. Gil, Aline B.M. Vaz, Tânia M.A. Alves, Carlos L. Zani, Carlos A. Rosa, and Luiz H. Rosa Abstract: The diversity and antimicrobial activity of endophytic fungi associated with the Brazilian medicinal plant Sola- num cernuum Vell. were studied during summer and winter seasons. A total of 246 fungal isolates were obtained, including 225 filamentous fungi and 21 yeasts. They were identified by morphological, physiological, and molecular methods. Fifty- five different taxa represented by the phyla Ascomycota (33 taxa), Basidiomycota (21 taxa), and Zygomycota (one taxon) were identified. The most abundant taxa were closely related to Arthrobotrys foliicola, Colletotrichum gloeosporioides, Co- prinellus radians, Glomerella acutata, Diatrypella frostii, Phoma glomerata, Mucor sp., Phlebia subserialis, Phoma mori- cola, Phanerochaete sordida, and Colletotrichum sp. A total of 265 fungal extracts were screened and 64 (26.01%) displayed antimicrobial activities. Among these extracts, 18 (28.12%) presented antibacterial and antifungal activities, 42 (65.62%) displayed selective antibacterial activity, and four (6.25%) exhibited only antifungal activity. The best values of minimum inhibitory concentration were obtained from extracts of Cryptococcus rajasthanensis, Glomerella acutata, Lepto- sphaeria sp., and Phoma glomerata ranging from 7.8 to 15.62 μg/mL. This study is the first survey of the endophytic fungi community associated with S. cernuum, and our results show that they can represent a promising source of bioactive com- pounds. Key words: antimicrobial activity, bioprospecting, fungal endophytes, tropical medicinal plant. Résumé : La diversité et l’activité antimicrobienne des champignons endophytes associés à la plante médicinale brésilienne Solanum cernuum Vell. ont été étudiés au cours des saisons d’été et d’hiver. Un total de 246 isolats fongiques ont été obte- nus, dont 255 champignons filamenteux et 21 levures. Ils ont été identifiés par des méthodes morphologiques, physiologi- ques et moléculaires. Cinquante-cinq taxons différents représentés par les phylums Ascomycota (33 taxons), Basidiomycota (21 taxons) et Zygomycota (un taxon) ont été identifiés. Les espèces les plus abondantes étaient étroitement reliées à Arthro- botrys foliicola, Colletotrichum gloeosporioides, Coprinellus radians, Glomerella acutata, Diatrypella frostii, Phoma glome- rata, Mucor sp., Phlebia subserialis, Phoma moricola, Phanerochaete sordida et Colletotrichum sp. Un total de 265 extraits fongiques ont été criblés et 64 d’entre eux (26,01 %) possédaient une activité antimicrobienne. Parmi ces extraits, 18 (28,12 %) présentaient une activité antimicrobienne et antifongique. Les meilleures valeurs de concentrations inhibitrices mi- nimales ont été obtenues d’extraits de Cryptococcus rajasthanensis, Glomerella acutata, Leptosphaeria sp. et Phoma glome- rata, allant de 7,8 μg/mL à 15,62 μg/mL. Cette étude constitue la première vue d’ensemble de la communauté endophyte fongique associée à S. cernuum, et nos résultats montrent qu’elle peut représenter une source prometteuse de composés bioactifs. Mots‐clés : activité antimicrobienne, bioprospection, endophytes fongiques, plante médicinale tropicale. [Traduit par la Rédaction] Introduction Fungi have been considered a promising source of com- pounds with different biological activities, making them an attractive target for the discovery of new classes of bioactive natural products. Among the tropical fungi, endophytes have been studied as a source of several bioactive metabolites (Chomcheon et al. 2006; Rosa et al. 2010a). Endophytes are microorganisms that inhabit healthy plant tissues during at least one stage of their life cycle, and they do not cause any Received 13 February 2011. Revision received 29 September 2011. Accepted 3 October 2011. Published at www.nrcresearchpress.com/cjm on 19 December 2011. M.L.A. Vieira, A.F.S. Hughes, V.B. Gil, A.B.M. Vaz, C.A. Rosa, and L.H. Rosa. Departamento de Microbiologia, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Belo Horizonte, MG, P.O. Box 486, CEP 31270-901, Brazil. T.M.A. Alves and C.L. Zani. Laboratório de Química de Produtos Naturais, Centro de Pesquisas René Rachou, Fundação Oswaldo Cruz, Av. Augusto de Lima, 1715, Belo Horizonte, MG, 30190-002, Brazil. Corresponding author: Luiz H. Rosa (e-mail: [email protected]). 54 Can. J. Microbiol. 58: 54–66 (2012) doi:10.1139/W11-105 Published by NRC Research Press Can. J. Microbiol. Downloaded from www.nrcresearchpress.com by UNIVERSITY OF MISSISSIPPI on 02/28/12 For personal use only.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Diversity and antimicrobial activities of the fungal endophyte community associated with the...

Diversity and antimicrobial activities of the fungalendophyte community associated with thetraditional Brazilian medicinal plant Solanumcernuum Vell. (Solanaceae)
Mariana L.A. Vieira, Alice F.S. Hughes, Viviane B. Gil, Aline B.M. Vaz,Tânia M.A. Alves, Carlos L. Zani, Carlos A. Rosa, and Luiz H. Rosa
Abstract: The diversity and antimicrobial activity of endophytic fungi associated with the Brazilian medicinal plant Sola-num cernuum Vell. were studied during summer and winter seasons. A total of 246 fungal isolates were obtained, including225 filamentous fungi and 21 yeasts. They were identified by morphological, physiological, and molecular methods. Fifty-five different taxa represented by the phyla Ascomycota (33 taxa), Basidiomycota (21 taxa), and Zygomycota (one taxon)were identified. The most abundant taxa were closely related to Arthrobotrys foliicola, Colletotrichum gloeosporioides, Co-prinellus radians, Glomerella acutata, Diatrypella frostii, Phoma glomerata, Mucor sp., Phlebia subserialis, Phoma mori-cola, Phanerochaete sordida, and Colletotrichum sp. A total of 265 fungal extracts were screened and 64 (26.01%)displayed antimicrobial activities. Among these extracts, 18 (28.12%) presented antibacterial and antifungal activities, 42(65.62%) displayed selective antibacterial activity, and four (6.25%) exhibited only antifungal activity. The best values ofminimum inhibitory concentration were obtained from extracts of Cryptococcus rajasthanensis, Glomerella acutata, Lepto-sphaeria sp., and Phoma glomerata ranging from 7.8 to 15.62 µg/mL. This study is the first survey of the endophytic fungicommunity associated with S. cernuum, and our results show that they can represent a promising source of bioactive com-pounds.
Key words: antimicrobial activity, bioprospecting, fungal endophytes, tropical medicinal plant.
Résumé : La diversité et l’activité antimicrobienne des champignons endophytes associés à la plante médicinale brésilienneSolanum cernuum Vell. ont été étudiés au cours des saisons d’été et d’hiver. Un total de 246 isolats fongiques ont été obte-nus, dont 255 champignons filamenteux et 21 levures. Ils ont été identifiés par des méthodes morphologiques, physiologi-ques et moléculaires. Cinquante-cinq taxons différents représentés par les phylums Ascomycota (33 taxons), Basidiomycota(21 taxons) et Zygomycota (un taxon) ont été identifiés. Les espèces les plus abondantes étaient étroitement reliées à Arthro-botrys foliicola, Colletotrichum gloeosporioides, Coprinellus radians, Glomerella acutata, Diatrypella frostii, Phoma glome-rata, Mucor sp., Phlebia subserialis, Phoma moricola, Phanerochaete sordida et Colletotrichum sp. Un total de 265 extraitsfongiques ont été criblés et 64 d’entre eux (26,01 %) possédaient une activité antimicrobienne. Parmi ces extraits, 18(28,12 %) présentaient une activité antimicrobienne et antifongique. Les meilleures valeurs de concentrations inhibitrices mi-nimales ont été obtenues d’extraits de Cryptococcus rajasthanensis, Glomerella acutata, Leptosphaeria sp. et Phoma glome-rata, allant de 7,8 µg/mL à 15,62 µg/mL. Cette étude constitue la première vue d’ensemble de la communauté endophytefongique associée à S. cernuum, et nos résultats montrent qu’elle peut représenter une source prometteuse de composésbioactifs.
Mots‐clés : activité antimicrobienne, bioprospection, endophytes fongiques, plante médicinale tropicale.
[Traduit par la Rédaction]
Introduction
Fungi have been considered a promising source of com-pounds with different biological activities, making them anattractive target for the discovery of new classes of bioactive
natural products. Among the tropical fungi, endophytes havebeen studied as a source of several bioactive metabolites(Chomcheon et al. 2006; Rosa et al. 2010a). Endophytes aremicroorganisms that inhabit healthy plant tissues during atleast one stage of their life cycle, and they do not cause any
Received 13 February 2011. Revision received 29 September 2011. Accepted 3 October 2011. Published atwww.nrcresearchpress.com/cjm on 19 December 2011.
M.L.A. Vieira, A.F.S. Hughes, V.B. Gil, A.B.M. Vaz, C.A. Rosa, and L.H. Rosa. Departamento de Microbiologia, Instituto de CiênciasBiológicas, Universidade Federal de Minas Gerais, Belo Horizonte, MG, P.O. Box 486, CEP 31270-901, Brazil.T.M.A. Alves and C.L. Zani. Laboratório de Química de Produtos Naturais, Centro de Pesquisas René Rachou, Fundação Oswaldo Cruz,Av. Augusto de Lima, 1715, Belo Horizonte, MG, 30190-002, Brazil.
Corresponding author: Luiz H. Rosa (e-mail: [email protected]).
54
Can. J. Microbiol. 58: 54–66 (2012) doi:10.1139/W11-105 Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIS
SISS
IPPI
on
02/2
8/12
For
pers
onal
use
onl
y.

apparent symptoms of disease or negative effects on theirhosts (Petrini 1991). Endophytic fungi can be found in al-most all plants, including trees, grasses, and herbaceousplants, and most of them belong to the Ascomycota phylumand its anamorphic stages (Huang et al. 2001).A variety of relationships may exist between endophytic
fungi and their hosts, ranging from mutualistic or symbioticinteractions to antagonistic or pathogenic (Schulz and Boyle2005). Because of what is supposed to be their contributionto the plant hosts, fungal endophytes may produce a plethoraof secondary metabolites that have a potential use in agricul-ture, medicine, and industry, such as novel antibiotics,immunosuppressants, antioxidants, and anticancer com-pounds. According to Strobel (2002), reasonable hypothesesshould govern the plant selection strategy for the discoveryof bioactive endophytic fungi; one of these includes plantsthat exhibit pharmacological activities. Recent studies havedemonstrated that endophytic fungi living in medicinal plantsexert a wide range of biological activities (Phongpaichit et al.2006; Huang et al. 2008; Rosa et al. 2010a).Brazil has a rich biodiversity of plant species with a long
tradition of herbal medicine, and many of them are used inthe form of crude extracts, infusions, or plasters to treat com-mon infections. Therefore, we selected the Brazilian medici-nal plant Solanum cernuum Vell. (Solanaceae), popularlyknown as “panacéia”, to study their endophytic fungal com-munities as a source of antimicrobial compounds. The leavesand stems of S. cernuum are commonly used to treat genito-urinary and skin infections, and its infusion has depurativeand diuretic properties (Carvalho 1989). In this work, we iso-lated the endophytic fungal community associated with leavesand stems of S. cernuum and screened them for their abilityto produce antibacterial and antifungal metabolites againstpathogenic microorganisms.
Materials and methods
Sample collection and isolation of fungal endophytesThe specimens of Solanum cernuum were collected in the
dry (winter) and rainy (summer) seasons of 2007 of the At-lantic Rain Forest, Brazil (19°52′S, 43°58′W). Identificationof S. cernuum was done by comparisons with the voucherspecimen deposited in the BHCB Herbarium (http://sciweb.nybg.org/science2/IndexHerbariorum.asp) of the Universi-dade Federal of Minas Gerais under the code BHCB 16876.The access of plant and fungi are according to the Brazilianbiological diversity rules.Four samples of apparently healthy leaves and stems were
randomly collected from each one of the 30 adult individualsof S. cernuum growing in the study area. Two leaves and twostems were collected from each of the 30 adult individuals ofS. cernuum. Three fragments of each leaf and stem wereplated, totaling 12 fragments per plant and 360 fragments atall. The same individuals were sampled during May and De-cember of 2007, corresponding to dry and rainy seasons, re-spectively. The leaves and stems were cut into pieces, placedin sterilized individual plastic bags, and stored for less than24 h at 10 °C prior to the isolation of endophytic fungi. Be-fore surface sterilization, the leaves and stems were cut into5-mm-long fragments using a flame-sterilized lamina in alaminar flow hood to avoid air contamination of the tissues
and all samples were separately processed. The fragmentswere surface-sterilized by immersion in Extran 2.5% (1 min),ethanol 70% (1 min), and 2% sodium hypochlorite (3 min),which was followed by washing with sterile distilled water(2 min) (Rosa et al. 2010a). Afterwards, they were plated onPetri plates containing supplemented with chloramphenicol(100 µg/mL), streptomycin sulphate (80 µg/mL), penicillin G(60 µg/mL), and benomil (2 µg/mL). The plates were incu-bated for up to 60 days at 28 °C, and the hyphal tip of thefungal colony growing out from the plant tissue was trans-ferred to PDA for filamentous fungi. Yeast colonies weretransferred to modified Sabouraud agar (glucose 2%, peptone1%, yeast extract 0.5%, agar 2%). After incubation at 28 °Cfor 10 days, culture purity was determined according to thecolony morphology. To test the effectiveness of surface steri-lization, 100 µL of the last rising water were plated on PDAand incubated at 28 °C. Long-term preservation of the fila-mentous fungal colonies was carried out in cryotubes with30% sterile glycerol at –80 °C and in sterile distilled waterat room temperature. Yeasts were maintained at –80 °C. Allendophytic fungal isolates were deposited into the CultureCollection of Microorganisms and Cells of the UniversidadeFederal de Minas Gerais.
Fungal identificationFilamentous fungal isolates were grouped into different
morphospecies according to their cultural characteristics: col-ony colour and texture, border type, and radial growth rateon PDA (Fröhlich et al. 2000). The protocol for the DNA ex-traction of filamentous fungi was conducted according toRosa et al. (2009). The internal transcribed spacer (ITS) do-mains of the rRNA gene were amplified with the universalprimers ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) andITS4 (5′-TCCTCCGCTTATTGATATGC-3′) (White et al.1990). Amplification of the ITS region was performed as de-scribed by Rosa et al. (2009). The yeasts were characterizedusing standard methods (Yarrow 1998), and their identifica-tion was carried out using the taxonomic keys of Kurtzmanand Fell (1998). The yeast identities were confirmed by se-quencing of the D1–D2 variable domains of the large-subunitrRNAgeneusing the primersNL1 (5′-GCATATCAATAAGCG-GAGGAAAAG-3′) and NL4 (5′-GGTCCGTGTTTCAA-GACGG-3′), as described by Lachance et al. (1999).Sequencing of filamentous fungi and yeasts was carried outusing an ET Dynamic Terminator kit in a MegaBACE1000/Automated 96 Capillary DNA sequencer (GE Health-care, USA). The nucleotide sequences and other related se-quences were aligned using the CLUSTALW softwarepackage (EMBL-EBI) (http://www.ebi.ac.uk/Tools/msa/clustalw2/). Consensus sequences were submitted to theGenBank database (see accession numbers in Tables 1 and 2).To identify the fungal species by DNA sequencing, the fol-
lowing preliminary criteria were used to interpret the sequen-ces of the GenBank database: for sequence identities >98%,the genus and species were accepted; for sequence identitiesbetween 95% and 97%, only the genus was accepted; for se-quence identities <95%, isolates were labelled as unknownspecies and identified in family, class, or order hierarchicallevels (Rosa et al. 2010b). However, the taxa that displayedidentities <97% or inclusive taxonomic level were submittedto phylogenetic inferences, which were estimated using
Vieira et al. 55
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIS
SISS
IPPI
on
02/2
8/12
For
pers
onal
use
onl
y.
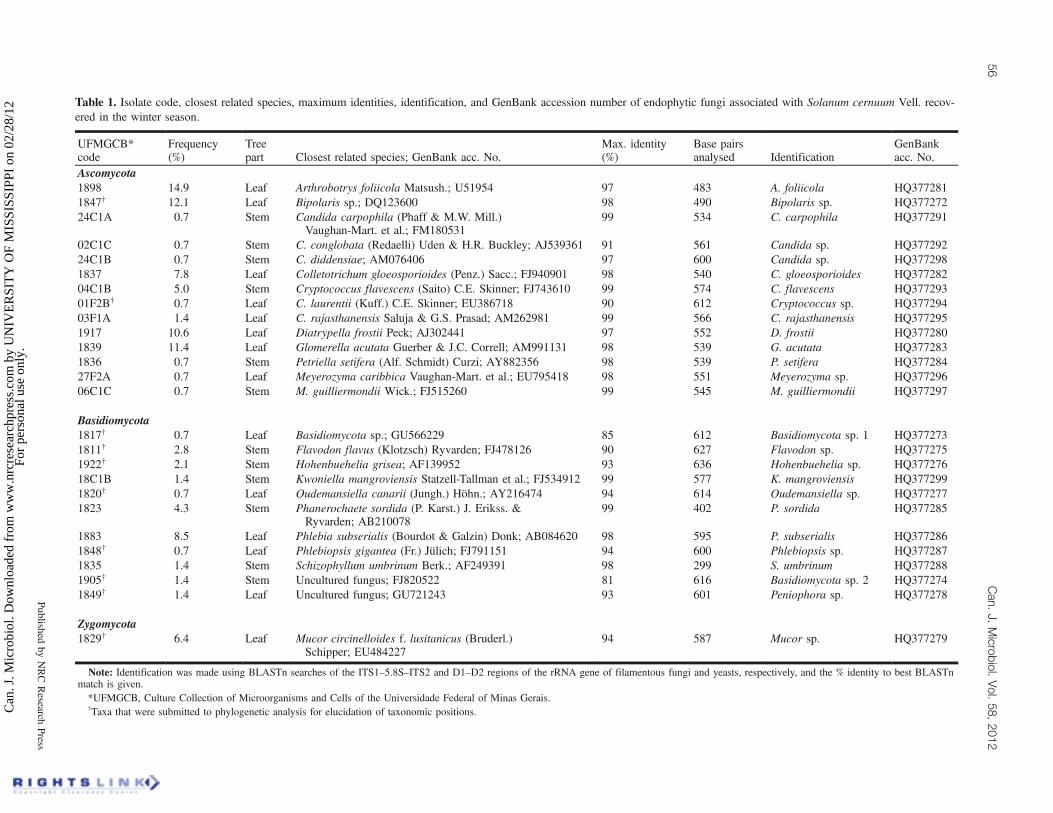
Table 1. Isolate code, closest related species, maximum identities, identification, and GenBank accession number of endophytic fungi associated with Solanum cernuum Vell. recov-ered in the winter season.
UFMGCB*code
Frequency(%)
Treepart Closest related species; GenBank acc. No.
Max. identity(%)
Base pairsanalysed Identification
GenBankacc. No.
Ascomycota1898 14.9 Leaf Arthrobotrys foliicola Matsush.; U51954 97 483 A. foliicola HQ3772811847† 12.1 Leaf Bipolaris sp.; DQ123600 98 490 Bipolaris sp. HQ37727224C1A 0.7 Stem Candida carpophila (Phaff & M.W. Mill.)
Vaughan-Mart. et al.; FM18053199 534 C. carpophila HQ377291
02C1C 0.7 Stem C. conglobata (Redaelli) Uden & H.R. Buckley; AJ539361 91 561 Candida sp. HQ37729224C1B 0.7 Stem C. diddensiae; AM076406 97 600 Candida sp. HQ3772981837 7.8 Leaf Colletotrichum gloeosporioides (Penz.) Sacc.; FJ940901 98 540 C. gloeosporioides HQ37728204C1B 5.0 Stem Cryptococcus flavescens (Saito) C.E. Skinner; FJ743610 99 574 C. flavescens HQ37729301F2B† 0.7 Leaf C. laurentii (Kuff.) C.E. Skinner; EU386718 90 612 Cryptococcus sp. HQ37729403F1A 1.4 Leaf C. rajasthanensis Saluja & G.S. Prasad; AM262981 99 566 C. rajasthanensis HQ3772951917 10.6 Leaf Diatrypella frostii Peck; AJ302441 97 552 D. frostii HQ3772801839 11.4 Leaf Glomerella acutata Guerber & J.C. Correll; AM991131 98 539 G. acutata HQ3772831836 0.7 Stem Petriella setifera (Alf. Schmidt) Curzi; AY882356 98 539 P. setifera HQ37728427F2A 0.7 Leaf Meyerozyma caribbica Vaughan-Mart. et al.; EU795418 98 551 Meyerozyma sp. HQ37729606C1C 0.7 Stem M. guilliermondii Wick.; FJ515260 99 545 M. guilliermondii HQ377297
Basidiomycota1817† 0.7 Leaf Basidiomycota sp.; GU566229 85 612 Basidiomycota sp. 1 HQ3772731811† 2.8 Stem Flavodon flavus (Klotzsch) Ryvarden; FJ478126 90 627 Flavodon sp. HQ3772751922† 2.1 Stem Hohenbuehelia grisea; AF139952 93 636 Hohenbuehelia sp. HQ37727618C1B 1.4 Stem Kwoniella mangroviensis Statzell-Tallman et al.; FJ534912 99 577 K. mangroviensis HQ3772991820† 0.7 Leaf Oudemansiella canarii (Jungh.) Höhn.; AY216474 94 614 Oudemansiella sp. HQ3772771823 4.3 Stem Phanerochaete sordida (P. Karst.) J. Erikss. &
Ryvarden; AB21007899 402 P. sordida HQ377285
1883 8.5 Leaf Phlebia subserialis (Bourdot & Galzin) Donk; AB084620 98 595 P. subserialis HQ3772861848† 0.7 Leaf Phlebiopsis gigantea (Fr.) Jülich; FJ791151 94 600 Phlebiopsis sp. HQ3772871835 1.4 Stem Schizophyllum umbrinum Berk.; AF249391 98 299 S. umbrinum HQ3772881905† 1.4 Stem Uncultured fungus; FJ820522 81 616 Basidiomycota sp. 2 HQ3772741849† 1.4 Leaf Uncultured fungus; GU721243 93 601 Peniophora sp. HQ377278
Zygomycota1829† 6.4 Leaf Mucor circinelloides f. lusitanicus (Bruderl.)
Schipper; EU48422794 587 Mucor sp. HQ377279
Note: Identification was made using BLASTn searches of the ITS1–5.8S–ITS2 and D1–D2 regions of the rRNA gene of filamentous fungi and yeasts, respectively, and the % identity to best BLASTnmatch is given.*UFMGCB, Culture Collection of Microorganisms and Cells of the Universidade Federal of Minas Gerais.†Taxa that were submitted to phylogenetic analysis for elucidation of taxonomic positions.
56Can.
J.Microbiol.
Vol.58,
2012
Publishedby
NRCResearch
Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIS
SISS
IPPI
on
02/2
8/12
For
pers
onal
use
onl
y.

Table 2. Isolate code, closest related species, maximum identities, identification, and GenBank accession number of endophytic fungi associated with Solanum cernuum Vell. recoveredin the summer season.
UFMGCB*code
Frequency(%) Part Closest related species; GenBank acc. No.
Max. identity(%)
Base pairsanalysed Identification
GenBankacc. No.
Ascomycota2231† 6.5 Stem Arthrobotrys foliicola Matsush.; U51954 96 516 Arthrobotrys sp. HM9971162183 0.8 Leaf A. foliicola; U51954 98 562 A. foliicola HM7751802141 7.3 Stem Bipolaris setariae Shoemaker; GU290228 97 545 B. setariae HM9971262154 0.8 Stem Botryosphaeria dothidea (Moug.: Fr.) Ces. & De Not.; GQ870286 97 532 B. dothidea HM9971272201 2.4 Stem Colletotrichum gloeosporioides (Penz.) Sacc.; GU066701 99 503 C. gloeosporioides HM9971282190† 8.9 Stem Colletotrichum sp.; EF672325 98 403 Colletotrichum sp. HQ37727101F1A 0.8 Leaf Candida carpophila; FM180531 99 587 C. carpophila HQ3772902163† 1.6 Stem Curvularia sp.; GQ184733 98 559 Curvularia sp. HM7751802143† 0.8 Leaf Dothideomycetes sp.; EU680503 98 541 Dothideomycetes sp. HM9971132177 0.8 Stem Edenia gomezpompae M.C. González et al.; EF565744 98 507 E. gomezpompae HM9971292193† 1.6 Leaf Eutypella scoparia (Schwein.) Ellis & Everh.; EU436688 96 554 Eutypella sp. HM9971302242† 4.0 Leaf Leptosphaeria sp.; HM008629 99 472 Leptosphaeria sp. HM9971152246† 2.4 Leaf Fungal endophyte; FJ466716 93 472 Chaetosphaeriaceae
sp.HM997125
2147† 7.3 Stem Fusarium equiseti (Corda) Sacc.; GQ505690 96 511 Fusarium sp. HM9971142171 3.2 Stem F. oxysporum Schltdl.; GU445378 97 527 F. oxysporum HM9971312175† 0.8 Leaf Petriella sp.; EU315012 99 528 Petriella sp. HM9971172241 10.5 Leaf Phoma moricola Sacc.; GQ352491 99 480 P. moricola HM9971322151† 10.5 Leaf Phoma sp.; FJ176472 98 474 Phoma sp. HM99711828C2A 1.6 Stem Meyerozyma guilliermondii Wick.; FJ972213 99 545 M. guilliermondii HQ377289
Basidiomycota2214† 0.8 Stem Basidiomycete sp.; AY605710 96 615 Meruliaceae sp. HM9971192130 4.0 Leaf Cercospora viticola (Ces.) Sacc.; EU520251 98 641 C. viticola HM9971332139† 13.7 Leaf Coprinellus radians (Desm.) Vilgalys et al.; AB470820 99 573 C. radians HM9971202239† 0.8 Stem Flavodon flavus (Klotzsch) Ryvarden; FJ010207 88 632 Basidiomycetes sp. HM9971222215† 0.8 Leaf Hapalopilus salmonicolor (Berk. & M.A. Curtis) Pouzar;
AY98649985 519 Basidiomycetes sp. HM997123
2212 1.6 Leaf Oudemansiella canarii (Jungh.) Höhn.; AY216474 98 611 O. canarii HQ5341012178 3.2 Stem Phanerochaete sordida (P. Karst.) J. Erikss. & Ryvarden;
AB21007897 594 P. sordida HM997134
2216 0.8 Stem Phlebia subserialis (Bourdot & Galzin) Donk; AB084620 97 588 P. subserialis HM9971352195† 0.8 Leaf Uncultured fungus; FJ820522 85 527 Polyporales sp. HM9971242192† 0.8 Leaf Uncultured root-associated fungus; EU144601 95 650 Coprinaceae sp. HM997121
Note: Identification was made using BLASTn searches of the ITS1–5.8S–ITS2 and D1–D2 regions of the rRNA gene of filamentous fungi and yeasts, respectively, and the % identity to best BLASTnmatch is given.*UFMGCB, Culture Collection of Microorganisms and Cells of the Universidade Federal of Minas Gerais.†Taxa that were submitted to phylogenetic analysis for elucidation of taxonomic positions.
Vieira
etal.
57
Publishedby
NRCResearch
Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIS
SISS
IPPI
on
02/2
8/12
For
pers
onal
use
onl
y.

MEGA version 4.0 (Tamura et al. 2007). The maximumcomposite likelihood model was used to estimate evolution-ary distance with bootstrap values calculated from 1000 rep-licate runs.
Species abundance, richness, evenness, and spatialanalysesSix healthy leaves and six stems of each plant (a total of
360 fragments) were plated on Petri plates containing PDA.The percentage abundance of each taxon was calculated ac-cording to the following formula: percentage abundance oftaxon A = (occurrence of taxon A × 100)/occurrence of alltaxa. These values were used to determine the prevalence ofeach taxon in comparison with the total fungal communityobtained from the leaves of S. cernuum. For measurement ofthe species diversity, we used the richness, evenness, anddominance categories given by three indexes: (i) ShannonH = –∑ni/n ln (ni/n), (ii) Simpson’s index = 1 – sum[(ni/n)2],and (iii) Dominance (D) = sum[(ni/n)2], where ni is thenumber of individuals of the taxon i and n is the total num-ber of individuals. Species richness refers to the number ofspecies in a community, and species dominance refers tothe contribution of individuals. The similarities among fun-gal communities from different seasons were estimated us-ing a Bray–Curtis (B) measure (Krebs 1999) and aSorensen coefficient (QS). The Bray–Curtis index is repre-sented by the formula: BCij = Si + Sj – 2Cij/Si + Sj, whereCij is the sum of minimum abundances of the various spe-cies (abundance at the site where the species is the rarest)and Si and Sj are the total number of specimens captured atboth sites. The Bray–Curtis values range is 0–1, where 0means that two communities do not share any species and1 means that the communities contain all the same speciesat identical frequency. The Sorensen coefficient (QS) analy-sis represented by the formula: QS = 2C/(A + B), where Aand B are the species numbers in samples A and B, respec-tively, and C is the number of species shared by the twosamples. The Sorensen coefficient puts more weight onjoint occurrences than on mismatches and is expressed withvalues between 0 (no similarity) and 1 (absolute similarity).All results were obtained with 95% confidence, and thebootstrap values were calculated from 1000 interactions.The index calculations were carried out using the computerprogramme PAST version 1.90 (Ryan et al. 1995).
Preparation of fungal and plant extracts for antimicrobialassaysEndophytic filamentous fungi and yeasts were previously
grown, respectively, on PDA and Sabouraud agar at 28 °Cfor 14 days at 70%–80% humidity for solid state fermentationtechnique. Afterwards, seven plugs (0.5 mm in diameter) ofculture agar medium, along with the fungal growth weretransferred to 1.5 mL tubes, each containing 1 mL of metha-nol. The tube content was then macerated one time, incu-bated at 10 °C for 48 h, and filtrated. To compare theantimicrobial activity among endophytic fungal extracts andS. cernuum, leaves and stems of each plant specimens werecollected, macerated with 10 mL of methanol, and incubatedat 10 °C for 48 h; this was the procedure used for the extrac-tion of fungal metabolites (Vaz et al. 2009). The solvent ofthe organic phases was completely evaporated until dryness
in a vacuum centrifuge at 35 °C. Crude extract stock solu-tions at a concentration of 10 mg/mL were prepared in tripli-cate and stored at –20 °C. These solutions were diluted withH2O–MeOH (9:1), and the final concentration of 100 µg/mLwas used in the antimicrobial assays.
Evaluation of antimicrobial activityPlant and fungal extracts were screened for their antimicro-
bial activity using the agar diffusion method according toRosa et al. (2003) against the potentially pathogenic bacteriaEscherichia coli ATCC 25922, Bacteroides fragilis ATCC25285, Staphylococcus aureus ATCC 12600, the opportunis-tic yeasts Candida albicans ATCC 18804, and Candida kru-sei ATCC 2159. For the agar diffusion assay, yeast targetswere previously grown on Sabouraud agar at 28 °C for 48 h,and bacteria was grown on BHI (Difco/USA) plus agar at 2%and incubated at 35 °C for 24 h. Colonies of the anaerobeB. fragilis were grown on BHI agar supplemented withhemin (0.05 mg/mL), menadione (0.01 mg/mL), and yeastextract (5 mg/mL) and incubated in an anaerobic chamber(Forma Scientific Company, Marietta, Ohio, USA) at 37 °Cfor 48 h in an atmosphere of 85% N2, 10% H2, and 5% CO2.Inocula of the target microorganisms were adjusted to theMcFarland No. 1 scale in optical density for yeasts, corre-sponding to 3 × 108 CFU/mL, and the McFarland 0.5 scalein optical density for bacteria, corresponding to 1 × 108 to2 × 108 CFU/mL. The inoculum concentration was con-firmed by a spectrophotometric reading at 580 and 626 nmfor yeasts and bacteria, respectively. Yeasts (1 mL) were ino-culated using a swab onto a plate containing Sabouraud agar.Bacterial inocula (1 mL) were plated on BHI agar, supple-mented with hemin, menadione, and yeast extract as de-scribed above, and then B. fragilis was seeded onto thesurface of the assay plates. The extracts at the concentrationof 100 µg/mL were applied to disc blanks on the Petri disheswith the target microorganisms, which were incubated for24–48 h at 35 °C and 28 °C for bacteria and yeasts, respec-tively. Bacteroides fragilis assay plates were incubated anae-robically at 37 °C for 48 h as previously described. Afterincubation, inhibition zones were observed, measured, andrecorded. Amphotericin B (Sigma, USA) and chlorampheni-col (Sigma, USA), both at 100 µg/mL, were used as positivecontrols for yeasts and bacteria, respectively. Solvent (H2O–MeOH, 9:1) and extract of the culture medium were used asnegative controls. All agar diffusion assays were performedin triplicate.
Minimum inhibitory concentration (MIC) determinationThirty-nine methanolic extracts produced from endophytic
fungi isolated during the winter season and 38 from thesummer season were selected according to their antimicrobialactivities for the determination of the minimum inhibitoryconcentration (MIC). The target microorganisms used for theMIC assays were the same as those selected for the diffusionagar method. The MIC of crude extracts was determined bythe microdilution method according to the protocols againstbacteria and yeasts of the National Committee for ClinicalLaboratory Standards (2004). After incubation, the lowestconcentration of extracts that inhibited the visible growth ofmicroorganisms was recorded as the MIC. All MIC assayswere performed in duplicate.
58 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIS
SISS
IPPI
on
02/2
8/12
For
pers
onal
use
onl
y.

Results
A total of 246 isolates of fungal endophytes were recov-ered from 360 samples of stems and leaves of S. cernuum,225 of which were filamentous fungi and 21 were yeasts.The process for surface sterilization of leaves and steamswas efficient and inhibited the growth of epiphytic micro-organisms. During winter, a total of 122 (46.03%) endophyticfungi were recovered, which included 104 filamentous fungiand 18 yeasts. During summer, 124 (46.79%) endophyticfungi were obtained, which included 121 filamentous fungiand three yeasts.All filamentous fungal isolates were grouped into 78 dif-
ferent morphospecies (35 were from winter and 43 werefrom summer seasons) based on their colony macro- andmicro-morphology characteristics. According to previousstudies about the diversity and taxonomy of fungal endophytecommunities in tropical ecosystems (Vaz et al. 2009; Rosa etal. 2010a), most of the endophytic fungi did not produceconidia or spores when cultured on common mycologicalmedia, such as potato dextrose, cornmeal, malt extract, Sabo-uraud dextrose, yeast extract, and sucrose agars. For this rea-son, at least 50% of the isolates of each differentmorphospecies were selected for molecular identification. Us-ing molecular analysis, 17 filamentous fungi and nine yeasttaxa were identified as endophytic fungi of S. cernuum inthe winter group (Table 1), and 27 filamentous fungi andtwo yeasts were identified in the summer group (Table 2).These fungi were represented by the phyla Ascomycota, Basi-diomycota, and Zygomycota. The endophytic fungi wereclosely related to the taxa Arthrobotrys foliicola, Colletotri-chum gloeosporioides, Glomerella acutata, Diatrypella fros-tii, Mucor sp., Phlebia subserialis, Phanerochaete sordida,which were most frequent in the winter season; in contrast,the endophyte fungi closely related to the taxa Coprinellusradians, Phoma glomerata, Phoma moricola, and Colletotri-chum sp. were most frequent in the summer season. The gen-era Candida, Colletotrichum, Meyerozyma, Oudemansiella,Petriella, Phanerochaete, and Phlebia were found in boththe winter and summer seasons. Twenty-seven taxa were notidentified at species level (11 of the winter group and 16 ofthe summer group) using the molecular techniques. Eighteentaxa were identified only at the genus level, three to the fam-ily, three to the class, and three to the phylum levels. Thesefungi ranged from 2% to 17% of nucleotide differences whencompared with their nearest relatives (Figs. 1a and 1b). Thediversity index values were similar between the fungal com-munities obtained in the winter (Shannon H = 2.75, Simp-son = 0.92, and D = 0.08) and summer (Shannon H = 2.92,Simpson = 0.93, and D = 0.07) seasons. However, the taxacomposition was variable, and specific taxa occurred in bothseasons, which was confirmed by values of 0.29 for the Sor-ensen and 0.12 for the Bray–Curtis similarity indexes. Thespecies found in both seasons were closely related to Bipola-ris sp., Candida carpophila, Colletotrichum gloeosporioides,Phanerochaete sordida, Phlebia subserialis, Phoma mori-cola, Phoma sp., and Meyerozyma guilliermondii.A total of 64 extracts (26.01%) out of the 246 produced
displayed antimicrobial activity against at least one target mi-croorganism, with inhibition zones ranging from 2.4 to14 mm in diameter (Table 3); of those 64 extracts, 33
(51.56%) were produced by fungi obtained in the winter sea-son and 31 (48.43%) in the summer. The plant extracts ob-tained in summer season displayed antimicrobial activityagainst S. aureus and C. krusei. Among the bioactive ex-tracts, 18 (28.1%) isolates showed broad antimicrobial spec-trum (antibacterial and antifungal activities). Forty-two(65.6%) isolates displayed only antibacterial activity, andfour (6.2%) exhibited only antifungal activity. The antibacte-rial activity detected was higher in the extracts of endophytesobtained in the winter season; in contrast, the majority of theendophytes collected in the summer season showed antifun-gal activity. Some extracts of endophyte taxa showed thebest halo diameter values of antimicrobial activity againstthe microbial targets; among them Meyerozyma guilliermon-dii UFMGCB 06C1C inhibited C. krusei (8.8 mm); Colleto-trichum gloeosporioides UFMGCB 1854 (10.48 mm) andUFMGCB 1856 (10.48) and Kwoniella mangroviensisUFMGCB 18C1B inhibited E. coli (10.8 mm); Bipolaris sp.UFMGCB 1847 (10.82 mm) and Dothideomycetes sp.UFMGCB 2219 inhibited B. fragilis (11.8 mm); Mucor sp.UFMGCB 1838 inhibited S. aureus (13.1 mm); Petriella se-tifera UFMGCB 1836 (13.79) and Schizophyllum umbrinumUFMGCB 1835 inhibited S. aureus (14 mm).Forty-two active extracts were submitted to MIC determi-
nation, ranging from 7.8 to >500 µg/mL against the targetmicroorganisms (Table 3). The best MIC values against thebacteria (7.8 µg/mL) were obtained with the extracts of taxaCryptococcus rajasthanensis UFMGCB 03F1A and Colleto-trichum gloeosporioides UFMGCB 2142 against E. coli,Glomerella acutata UFMGCB 1840 against S. aureus, andCoprinellus radians UFMGCB 2139 against B. fragilis. Ad-ditionally, the taxa Cryptococcus sp. UFMGCB 01F1A (MICvalue of 6.25 µg/mL against C. albicans) and Fusarium sp.UFMGCB 2147 (MIC value of 15.62 µg/mL against C. kru-sei) produced the best MIC values against the yeast targets.
DiscussionMost of the fungal genera obtained as endophytes in
S. cernuum have already been described as endophytes ofplants from tropical and temperate ecosystems (Higgins et al.2007; Huang et al. 2008; Vaz et al. 2009; Rosa et al. 2010a).However, almost one third of the endophyte species showedlow similarity with the sequences of known fungal species.Arthrobotrys foliicola was the taxon most frequently foundin S. cernuum. Some species of Arthrobotrys have been re-ported as endophytes, such as Arthrobotrys conoides, whichis associated with the bark of Crataeva magna (Nalini et al.2005), and Arthrobotrys oligospora and Arthrobotrys folii-cola, both from Oryza sativa (Zakaria et al. 2010). The sec-ond most frequent species was Colletotrichumgloeosporioides, which is a known fungal endophyte. Colle-totrichum species (including C. gloeosporioides) are foundinhabiting several plant hosts (Marcelino et al. 2008; Rosa etal. 2010a). The taxon Glomerella acutata (teleomorph stageof Colletotrichum acutatum) was the third most frequent fun-gus, which is an endophyte fungus able to infect a widerange of plant hosts (mosses, bryophytes, and angiosperms)living in both tropical and cold ecosystems (Higgins et al.2007). The fourth most frequent endophyte of S. cernuumwas Coprinellus radians. Coprinellus (Psathyrellaceae) is a
Vieira et al. 59
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIS
SISS
IPPI
on
02/2
8/12
For
pers
onal
use
onl
y.

basidiomycetous genus and includes more than 40 definedspecies (Redhead et al. 2001), of which some species havebeen obtained as endophytes of Theobroma cacao in CostaRica and Ecuador (Márquez et al. 2008; Thomas et al.2008). The other taxa recovered from S. cernuum, includingDiatrypella frostii, Phoma glomerata, Mucor sp., Phlebiasubserialis, and Phoma moricola, are also found as endo-phytes in different plant species (Glawe 1983; Qi et al.2009; Abreu et al. 2010; Sim et al. 2010). Thirteen taxawere obtained in low frequencies with an abundance ≤0.37%,
which represent the minor components of this fungal com-munity. The presence of different basidiomycetous fungi as-sociated with S. cernuum (genera Cercospora, Coprinellus,Flavodon, Hohenbuehelia, Kwoniella, Oudemansiella, Penio-phora, Phanerochaete, Phlebia, Phlebiopsis, and Schizophyl-lum) are noteworthy because reports on the Basidiomycotaspecies as endophytes are scarce.A total of 27 endophyte taxa recovered (11 from the winter
and 16 from the summer seasons) showed low similarity withsequences of fungi deposited in the GenBank database. These
Fig. 1. Phylogenetic analysis of filamentous fungi recovered in winter (a) and summer (b) that were recovered from Solanum cernuum Vell.and their nearest relatives. The tree was constructed based on the rRNA gene sequences (ITS1–5.8S–ITS2) by using the maximum compositelikelihood model. The tree was rooted with Trametes versicolor (AF042324) as an outgroup.
60 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIS
SISS
IPPI
on
02/2
8/12
For
pers
onal
use
onl
y.

taxa showed different levels of sequence similarity with taxaidentified in the levels of genus (Arthrobotrys, Bipolaris,Candida, Colletotrichum, Cryptococcus, Curvularia, Meyero-zyma, Eutypella, Flavodon, Fusarium, Hohenbuehelia, Lep-tosphaeria, Mucor, Oudemansiella, Petriella, Phlebiopsis,Phoma, and Peniophora), family (Chaetosphaeriaceae,Meruliaceae, and Coprinaceae), class (Basidiomycetes andDothideomycetes), order (Polyporales), or phyla (Basidio-mycota), which suggests that these endophytic fungi could
represent new fungal species. However, more morphologicaland phylogenetic taxonomic studies will be necessary to de-termine the correct identification of these fungal species aswell as describe whether or not they are new taxa.Population fluctuation of bioactive endophytic fungi dur-
ing the winter and summer seasons was detected in thisstudy. The number of bioactive taxa was similar between thetwo seasons. However, the antibacterial activity was observedmore in the winter; in contrast, the antifungal activity was
Fig. 1 (concluded).
Vieira et al. 61
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIS
SISS
IPPI
on
02/2
8/12
For
pers
onal
use
onl
y.

Table 3. Antimicrobial activity crude extracts obtained from endophytic fungi isolated from Solanum cernuum Vell. against yeasts and bacterial targets.
Bacteria Yeasts
Ec Sa Bf Ck Ca
UFMGCBcode* Endophytic taxa
Inhibitionzone (mm)
MIC(µg/mL)
Inhibitionzone (mm)
MIC(µg/mL)
Inhibitionzone (mm)
MIC(µg/mL)
Inhibitionzone (mm)
MIC(µg/mL)
Inhibitionzone (mm)
MIC(µg/mL)
Winter1825 Bipolaris sp. — — — — 7.34 125 — — — —1827 Bipolaris sp. — — 4.55 NA — — — —1847 Bipolaris sp. — — 10.82 NA — — — — — —1850 Bipolaris sp. — — 7.32 NA — — — — — —24C1A Candida carpophila 9 >500 — — — — — — — —1837 Colletotrichum gloeosporioides — — 7.87 NA — — — — — —1851 C. gloeosporioides — — 9.70 31.2 6.79 >500 — — — —1852 C. gloeosporioides — — 9.75 NA — — — — — —1854 C. gloeosporioides — — 10.48 >500 — — — — — —1856 C. gloeosporioides — — 10.48 >500 — — — — — —03F1A Cryptococcus rajasthanensis 7.56 7.8 — — — — — — — —26F2A C. rajasthanensis 6.33 31.2 — — — — — — — —01F2A Cryptococcus sp. 8.34 125 7.52 >500 — — — — — —1834 Glomerella acutata — — 7.40 NA — — — —1839 G. acutata — — — — 6.54 >500 — — — —1840 G. acutata — — 7.16 7.8 6.68 62.5 — — — —1860 G. acutata — — — — 6.53 250 — — — —18C1A Kwoniella mangroviensis 2.45 250 6.72 250 — — — — — —18C1B K. mangroviensis 10.8 >500 6.44 >500 — — — — — —1829 Mucor sp. — — 7.40 250 — — — —1830 Mucor sp. — — 7.03 NA — — — —1833 Mucor sp. — — 4.57 >500 — — — —1838 Mucor sp. — — 13.10 125 — — — — — —1842 Mucor sp. — — — — 4.68 NA — — — —1820 Oudemansiella sp. — — — — 7.05 >500 — — — —1849 Peniophora sp. — — 9.94 31.25 — — — — — —1836 Petriella setifera — — 13.79 250 — — — — — —1822 Phanerochaete sordida — — — — 7.16 NA — — — —1855 P. sordida — — — — 6.66 NA — — — —1873 P. sordida — — — — 6.68 62.5 — — — —1848 Phlebiopsis sp. — — 10.39 62.5 — — — — — —06C1C Meyerozyma guilliermondii 5.60 >500 — — — — 8.85 125 7.65 NA1835 Schizophyllum umbrinum — — 14.02 250 6.59 >500 — — — —Plant extracts Solanum cernuum leaves — — — — — — — — — —
S. cernuum stems — — — — — — — — — —
62Can.
J.Microbiol.
Vol.58,
2012
Publishedby
NRCResearch
Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIS
SISS
IPPI
on
02/2
8/12
For
pers
onal
use
onl
y.

Table 3 (concluded).
Bacteria Yeasts
Ec Sa Bf Ck Ca
UFMGCBcode* Endophytic taxa
Inhibitionzone (mm)
MIC(µg/mL)
Inhibitionzone (mm)
MIC(µg/mL)
Inhibitionzone (mm)
MIC(µg/mL)
Inhibitionzone (mm)
MIC(µg/mL)
Inhibitionzone (mm)
MIC(µg/mL)
Summer2215 Basidiomycetes sp. 7.31 15.6 — — 2.44 >5002239 Basidiomycetes sp. 7.6 15.6 — — — — — — — —2157 Bipolaris setariae 7.23 125 — — 7.40 >500 — — 2.47 >5002181 B. setariae — — 7.92 NA — — — — 2.64 NA2154 Botryosphaeria dothidea — — — — 6.97 125 7.09 125 4.69 >5002246 Chaetosphaeriaceae sp. — — — — 7.18 NA 2.39 NA2142 Colletotrichum gloeosporioides 7.6 7.8 — — — — — — — —2192 Coprinaceae sp. 7.57 125 — — — — — — — —2139 Coprinellus radians — — — — 7.38 7.8 — — 2.46 >5002155 C. radians 7.38 >500 — — — — — — — —2176 C. radians 7.2 NA — — — — — — — —2187 C. radians — — — — — — 7.81 NA 2.60 NA01F1A Cryptococcus sp. — — — — — — 7.23 62.5 — —2163 Curvularia sp. 6.85 >500 — — — — — — — —2219 Dothideomycetes sp. 11.84 125 — — 3.95 NA2194 Edenia gomezpompae — — 6.75 NA 7.51 NA — — 4.75 NA2193 Eutypella sp. — — 7.69 NA — — — — 2.56 NA2171 Fusarium oxysporum 6.81 NA — — — — — — — —2248 F. oxysporum — — 8.78 NA — — — — 2.93 NA2140 Fusarium sp. 7.71 125 — — — — — — — —2147 Fusarium sp. — — — — 7.21 NA — — 2.40 15.62188 Fusarium sp. — — — — — 7.10 NA 2.37 NA2242 Leptosphaeria sp. 6.72 125 — — — — — — — —2214 Meruliaceae sp. 7.87 NA — — — — 2.62 NA2206 Phanerochaete sordida — — — — 7.50 NA — — 2.50 1252197 Phoma moricola — — — — 7.76 250 — — 2.59 >5002241 P. moricola 7.69 >500 8.12 >500 — — — — 2.71 NA2237 P moricola — — — — 7.25 NA — — 2.42 NA2238 Phoma sp. 7.6 62.5 — — — — — — — —28C2A Meyerozyma guilliermondii 6.6 250 4.35 NA — — 6.95 250 — —2195 Polyporales sp. — — — — — — 8.46 NA 2.82 NAPlant extracts Solanum cernuum leaves — — 7.4 250 — — — — — —
S. cernuum stems — — — — — — 7.9 125 — —Controlsdrugs
Amphotericin B — — — — — — 9.36 0.5 8.72 0.25–1
Chloramphenicol 10.95 2–8 15.71 2–16 10.14 3–15 — — — —Note: Ec, Escherichia coli; Sa, Staphylococcus aureus; Bf, Bacteroides fragilis; Ca, Candida albicans; Ck, Candida krusei. NA, not assayed.*UFMGCB, Culture Collection of Microorganisms and Cells of the Universidade Federal of Minas Gerais.
Vieira
etal.
63
Publishedby
NRCResearch
Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIS
SISS
IPPI
on
02/2
8/12
For
pers
onal
use
onl
y.

only observed in the summer season. One explanation of thissituation might be that the endophyte assemblage depends onthe physiological status of the host plant, which can showvariation across the seasons due to factors like water and nu-trient availability, photoperiod, temperature, and others.These variations may be responsible for differences in thebiological activities of the endophytic fungal communities ofS. cernuum. Methanolic extracts from leaves and stems ofS. cernuum obtained during winter and summer were also as-sayed against the target microorganisms. Plant samples col-lected in winter did not exhibit antimicrobial activity; incontrast, in the summer, leaves and stems of S. cernuum dis-played antifungal and antibacterial activities against S. aureusand C. krusei, respectively.The taxa Kwoniella mangroviensis UFMGCB 18C1B, Mu-
cor sp. UFMGCB 1838, Schizophyllum umbrinum UFMGCB1835, Dothideomycetes sp. UFMGCB 2219, and Meyero-zyma guilliermondii UFMGCB 06C1C showed the best in-hibitory halos against the target microorganisms. Few Mucortaxa have been reported as endophytes and as producers ofbioactive molecules; however, some antitumoral activitieshave already been detected in an isolate, Mucor sp. (Huanget al. 2001), and in strains of Mucor rouxianus (Miao et al.2009). The Schizophyllum genus includes mainly saprophyticspecies, which occur in a wide range of substrata, althoughthey are not particularly active in the degradation of woodworldwide (Cooke 1961). However, some studies have de-scribed the Schizophyllum species as endophytes of someplants species, such as Elaeis guineensis, Theobroma cacao,and Theobroma gileri (Evans et al. 2003; Crozier et al. 2006;Rungjindamai et al. 2008). Additionally, the Schizophyllumtaxa have recently attracted significant attention due to theirbiological activities, specifically antitumoral activity(Zjawiony 2004). According to Cooke (1961), Schizophyllumumbrinum, which was found as endophyte in S. cernuum, is aneotropical taxon, and our study described for the first timethat this taxon is an endophyte and a source of antimicrobialcompounds. The class Dothideomycetes includes pathogenic,endophytic, or epiphytic taxa, and some works have de-scribed the potential of endophytic Dothideomycetes as pro-ducers of bioactive compounds. The extract of endophyticDothideomycetes sp., recovered from the medicinal plantLeea rubra, shows cytotoxicity against the Vero cell line(Chomcheon et al. 2006). Additionally, an endophytic isolateof Dothideomycetes sp., recovered from the medicinal plantTylophora indica, shows antifungal activity against the plantpathogens Fusarium oxysporum and Sclerotinia sclerotiorum(Kumar et al. 2011).Glomerella acutata, Colletotrichum gloeosporioides, Fusa-
rium sp., Phoma sp., Coprinellus radians, Cryptococcus ra-jasthanensis, and Cryptococcus sp. showed the best valuesof MIC against the bacteria and yeasts used as targets. Thecomplex Colletotrichum and Glomerella species have beendescribed as prolific endophytic bioactive fungi that presentantimicrobial (Liu et al. 2001) and cytotoxic (Rosa et al.2010a) activities. Endophytic Fusarium species are able toproduce compounds with antimicrobial (Vaz et al. 2009) andcytotoxic (Rosa et al. 2010a) activities. The species Phomamoricola, detected as an endophyte of S. cernuum, was alsoreported to be an endophyte of the medicinal plant Garciniaparvifolia, and this species is able to produce antimicrobial
compounds that are active against Listeria monocytogenesand Agromyces lapidis (Sim et al. 2010).In this study, we detected five endophytic yeasts able to
produce antimicrobial compounds (Candida carpophila,Cryptococcus rajasthanensis, Cryptococcus sp., Kwoniellamangroviensis, and Meyerozyma guilliermondii). This repre-sents interesting results, because few studies of taxonomyand antimicrobial activity of tropical endophytic yeasts havebeen reported. Among them, Vaz et al. (2009) reported theantimicrobial activities of the endophytic taxa of yeasts Rho-dotorula mucilaginosa, Bensingtonia sp., and Candida para-psilosis that are associated with tropical orchids; they showedantibacterial activity against E. coli, S. aureus, Bacillus cer-eus, and Salmonella typhimurium. In addition, we discovereda new habitat for the yeast Kwoniella mangroviensis, whichwas isolated from mangrove habitats in Florida and the Baha-mas Islands and from the bark of cork oak trees in Spain(Statzell-Tallman et al. 2008).Cryptococcus rajasthanensis, Colletotrichum gloeospor-
ioides, Coprinellus radians, Cryptococcus sp., and Fusariumsp. showed the best MIC values against the target microor-ganisms. These MIC values are comparable to those ofknown antibacterial and antifungal drugs; for example, chlor-amphenicol has activity against E. coli (MIC 2–8 µg/mL) andS. aureus (MIC 2–16 µg/mL), amphotericin B has activityagainst C. albicans (MIC 0.25–1 µg/mL), and fluconazole(MIC 0.25–2 µg/mL) has activity against C. krusei (NationalCommittee for Clinical Laboratory Standards 2004). Theseextracts are potential candidates for further studies, whichcan serve as sources of new antimicrobial compounds.The Brazilian medicinal plant S. cernuum showed a di-
verse endophytic fungal community that include taxa that areable to produce antimicrobial compounds, which justifiestheir selection for bioassay-oriented chemical investigationsto isolate and characterize prototype molecules for new drugdevelopment. Additionally, the results obtained in this worksuggest that endophytic fungal communities living in associ-ation with ethnobotanical Brazilian flora could be potentsources of active natural products.
AcknowledgementsThis work received support from the Fundação of Amparo
the Pesquisa of Minas Gerais (FAPEMIG) and Conselho Na-cional de Desenvolvimento Científico e Tecnológico (CNPq).
ReferencesAbreu, L.M., Almeida, A.R., Salgado, M., and Pfenning, L.H. 2010.
Fungal endophytes associated with the mistletoe Phoradendronperrottettii and its host tree Tapirira guianensis.Mycol. Prog. 9(4):559–566. doi:10.1007/s11557-010-0663-8.
Carvalho, L.D.F. 1989. Espécies de Solanum (Seção Cernuum eLepidotum) usadas na medicina popular brasileira. Rev. Bras.Farmacogn. 70: 7–10. doi:10.1590/S0102-695X2007000400012.
Chomcheon, P., Sriubolmas, N., Wiyakrutta, S., Ngamrojanavanich,N., Chaichit, N., Mahidol, C., et al. 2006. Cyclopentenones,scaffolds for organic syntheses produced by the endophytic fungusmitosporic Dothideomycete sp. LRUB20. J. Nat. Prod. 69(9):1351–1353. doi:10.1021/np060148h. PMID:16989533.
Cooke, W.M. 1961. The genus Schizophyllum. Mycologia, 53(6):575–599. doi:10.2307/3756459.
Crozier, J., Thomas, S.E., Aime, M.C., Evans, H.C., and Holmes, K.
64 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIS
SISS
IPPI
on
02/2
8/12
For
pers
onal
use
onl
y.

A. 2006. Molecular characterization of fungal endophyticmorphospecies isolated from stems and pods of Theobromacacao. Plant Pathol. 55(6): 783–791. doi:10.1111/j.1365-3059.2006.01446.x.
Evans, H.C., Holmes, K.A., and Thomas, S.E. 2003. Endophytes andmycoparasites associated with an indigenous forest tree, Theo-broma gileri, in Ecuador and preliminary assessment of theirpotential as biocontrol agents of cocoa diseases. Mycol. Prog. 2(2):149–160. doi:10.1007/s11557-006-0053-4.
Fröhlich, J., Hyde, K.D., and Petrini, O. 2000. Endophytic fungiassociated with palms. Mycol. Res. 104(10): 1202–1212. doi:10.1017/S095375620000263X.
Glawe, D.A. 1983. Observations on the anamorph of Diatrypellafrostii. Mycologia, 75(5): 913–915. doi:10.2307/3792786.
Higgins, K.L., Arnold, A.E., Miadlikowska, J., Sarvate, S.D., andLutzoni, F. 2007. Phylogenetic relationship, host affinity, andgeographic structure of boreal and artic endophytes from threemajor plant lineages. Mol. Phylogenet. Evol. 42(2): 543–555.doi:10.1016/j.ympev.2006.07.012.
Huang, Y., Wang, J., Li, G., Zheng, Z., and Su, W. 2001. Antitumorand antifungal activities in endophytic fungi isolated frompharmaceutical plants Taxus mairei, Cephalataxus fortunei andTorreya grandis. FEMS Immunol. Med. Microbiol. 31(2): 163–167. doi:10.1111/j.1574-695X.2001.tb00513.x. PMID:11549424.
Huang, W.Y., Cai, Y.Z., Hyde, K.D., Corke, H., and Sun, M. 2008.Biodiversity of endophytic fungi associated with 29 traditionalChinese medicinal plants. Fungal Divers. 33: 61–75.
Kimura, M. 1980. A simple method for estimating evolutionary ratesof base substitutions through comparative studies of nucleotidesequences. J. Mol. Evol. 16(2): 111–120. doi:10.1007/BF01731581. PMID:7463489.
Krebs, C. 1999. Ecological methodology. 2nd ed. Addison-WesleyEducational Publishers, Inc., Menlo Park, Calif., USA.
Kumar, S., Kaushik, N., Edrada-Ebel, R., Ebel, R., and Proksch, P.2011. Isolation, characterization, and bioactivity of endophyticfungi of Tylophora indica. World J. Microbiol. Biotechnol. 27(3):571–577. doi:10.1007/s11274-010-0492-6.
Kurtzman, C.P., and Fell, J.W. 1998. The yeast, a taxonomic study.4th ed. Elsevier, New York, USA.
Lachance, M.A., Bowles, J.M., Starmer, W.T., and Barker, J.S. 1999.Kodamaea kakaduensis and Candida tolerans, two new ascomy-cetous yeast species from Australian hibiscus flowers. Can. J.Microbiol. 45(2): 172–177. PMID:10380650.
Liu, C.H., Zou, W.X., Lu, H., and Tan, R.X. 2001. Antifungal activityof Artemisia annua endophyte cultures against phytopathogenicfungi. J. Biotechnol. 88(3): 277–282. doi:10.1016/S0168-1656(01)00285-1. PMID:11434973.
Marcelino, J., Giordano, R., Gouli, S., Gouli, V., Parker, B.L.,Skinner, M., et al. 2008. Colletotrichum acutatum var. fioriniae(teleomorph: Glomerella acutata var. fioriniae var. nov.) infectionof a scale insect. Mycologia, 100(3): 353–374. doi:10.3852/07-174R. PMID:18751543.
Márquez, S.S., Bills, F.G., and Zabalgogeazcoa, I. 2008. Diversityand structure of the fungal endophytic assemblages from towsympatric coastal grasses. Fungal Divers. 33: 87–100.
Miao, Z., Wang, Y., Yu, X., Guo, B., and Tang, K. 2009. A newendophytic taxane production fungus from Taxus chinensis. Appl.Biochem. Microbiol. 45(1): 81–86. doi:10.1134/S0003683809010141.
Nalini, M.S., Mahesh, B., Tejesvi, M.V., Prakash, H.S., Subbaiah, V.,Kini, K.R., and Shetty, H.S. 2005. Fungal endophytes from thethree leaved caper, Crataeva magna (Lour.) (Capparidaceae).Mycopathologia, 159(2): 245–249. doi:10.1007/s11046-004-5497-y. PMID:15770450.
National Committee for Clinical Laboratory Standards. 2004.Performance standards for antimicrobial susceptibility testing.Fourteenth Informational Supplement NCCLS documents M100-S14. NCCLS, Wayne, Penn., USA.
Petrini, O. 1991. Fungal endophytes of tree leaves. In Microbialecology of leaves. Edited by N.J. Andrews and S.S. Hirano.Springer, New York, USA.
Phongpaichit, S., Rungjindamai, N., Rukachaisirikul, V., andSakayaroj, J. 2006. Antimicrobial activity in cultures of endophy-tic fungi isolated from Garcinia species. FEMS Immunol. Med.Microbiol. 48(3): 367–372. doi:10.1111/j.1574-695X.2006.00155.x.PMID:17052267.
Qi, F.H., Jing, Z.X., Wang, Y.G., and Zhan, Y. 2009. Fungalendophytes from Acer ginnalaMaxim: isolation, identification andtheir yield of gallic acid. Lett. Appl. Microbiol. 49(1): 98–104.doi:10.1111/j.1472-765X.2009.02626.x. PMID:19413766.
Redhead, S.A., Vilgalys, R., Moncalvo, J.M., Johnson, J., andHopple, J.S. 2001. Coprinus Pers., and the disposition of Coprinusspecies sensu lato. Taxon, 50(1): 203–241. doi:10.2307/1224525.
Rosa, L.H., Machado, K.M., Jacob, C.C., Capelari, M., Rosa, C.A.,and Zani, C.L. 2003. Screening of Brazilian basidiomycetes forantimicrobial activity. Mem. Inst. Oswaldo Cruz, 98(7): 967–974.doi:10.1590/S0074-02762003000700019. PMID:14762527.
Rosa, L.H., Vaz, A.B.M., Caligiorne, R.B., Campolina, S., and Rosa,C.A. 2009. Endophytic fungi associated with the Antarctic GrassDeschampsia antarctica Desv. (Poaceae). Polar Biol. 32(2): 161–167. doi:10.1007/s00300-008-0515-z.
Rosa, L.H., Gonçalves, V.N., Caligiorne, R.B., Alves, T.M.A.,Rabello, A., Sales, P.A., et al. 2010a. Leishmanicidal, trypanoci-dal, and cytotoxic activities of endophytic fungi associated withbioactive plants in Brazil. Braz. J. Microbiol. 41(2): 420–430.doi:10.1590/S1517-83822010000200024.
Rosa, L.H., de Lourdes Almeida Vieira, M., Santiago, I.F., and Rosa,C.A. 2010b. Endophytic fungi community associated with thedicotyledonous plant Colobanthus quitensis (Kunth) Bartl.(Caryophyllaceae) in Antarctica. FEMS Microbiol. Ecol. 73(1):178–189. doi:10.1111/j.1574-6941.2010.00872.x. PMID:20455944.
Rungjindamai, N., Pinruan, U., Choeyklin, R., Hattori, T., and Jones,E.B.G. 2008. Molecular characterization of basidiomycetousendophytes isolated from leaves, rachis and petioles of the oilpalm, Elaeis guineensis, in Thailand. Fungal Divers. 33: 139–161.
Ryan, G.S., Harper, D.A.T., and Whalley, J.S. 1995. PALSTAT,statistics for Palaeontologist. Chapman & Hall, London, UK.
Schulz, B., and Boyle, C. 2005. The endophytic continuum. Mycol.Res. 109(6): 661–686. doi:10.1017/S095375620500273X. PMID:16080390.
Sim, J.H., Khoo, C.H., Lee, L.H., and Cheah, Y.K. 2010. Moleculardiversity of fungal endophytes isolated from Garcinia mangostanaand Garcinia parvifolia. J. Microbiol. Biotechnol. 20(4): 651–658.doi:10.4014/jmb.0909.09030. PMID:20467234.
Statzell-Tallman, A., Belloch, C., and Fell, J.W. 2008. Kwoniellamangroviensis gen. nov., sp.nov. (Tremellales, Basidiomycota), ateleomorphic yeast from mangrove habitats in the Florida Ever-glades and Bahamas. FEMS Yeast Res. 8(1): 103–113. doi:10.1111/j.1567-1364.2007.00314.x. PMID:17961172.
Strobel, G.A. 2002. Rainforest endophytes and bioactive products.Crit. Rev. Biotechnol. 22(4): 315–333. doi:10.1080/07388550290789531. PMID:12487423.
Tamura, K., Dudley, J., Nei, M., and Kumar, S. 2007. MEGA4:molecular evolutionary genetics analysis (MEGA) softwareversion 4.0. Mol. Biol. Evol. 24(8): 1596–1599. doi:10.1093/molbev/msm092. PMID:17488738.
Thomas, S.E., Crozier, J.M., Aime, C., Evans, H.C., and Holmes, K.
Vieira et al. 65
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIS
SISS
IPPI
on
02/2
8/12
For
pers
onal
use
onl
y.

A. 2008. Molecular characterisation of fungal endophyticmorphospecies associated with the indigenous forest tree,Theobroma gileri, in Ecuador. Mycol. Res. 112(7): 852–860.doi:10.1016/j.mycres.2008.01.008. PMID:18534836.
Vaz, A.B.M., Mota, R.C., Bomfim, M.R.Q., Vieira, M.L.A., Zani, C.L., Rosa, C.A., and Rosa, L.H. 2009. Antimicrobial activity ofendophytic fungi associated with Orchidaceae in Brazil. Can. J.Microbiol. 55(12): 1381–1391. doi:10.1139/W09-101. PMID:20029530.
White, T.J., Bruns, T.D., and Lee, S.B. 1990. Amplification anddirect sequencing of fungal ribosomal RNA genes for phyloge-netics. In PCR protocols: a guide to methods and applications.
Edited by N.A. Innis, J. Gelfand, and J. Sninsky. Academic Press,San Diego, Calif., USA.
Yarrow, D. 1998. Methods for the isolation, maintenance andidentification of yeast. In The yeasts: a taxonomic study. Editedby C.P. Kurtzman and J.W. Fell, Elsevier, Amsterdam.
Zakaria, Z.A., Sufian, A.S., Ramasamy, K., Ahmat, N., Sulaiman, M.R., Arifah, A.K., et al. 2010. In vitro antimicrobial activity ofMuntingia calabura extracts and fractions. Afr. J. Microbiol. Res.4: 304–308.
Zjawiony, J.K. 2004. Biologically active compounds from Aphyllo-phorales (polypore) fungi. J. Nat. Prod. 67(2): 300–310. doi:10.1021/np030372w. PMID:14987072.
66 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIS
SISS
IPPI
on
02/2
8/12
For
pers
onal
use
onl
y.




















