
Diversidad de hongos silvestres comestibles en un bosque de coniferas (Abies religiosa), en el...
115
UNIVERSIDBD BUTONOMB CHBPINGO / Programa Interdepartamental de Docencia, Investigaci6n y Servicio en Agroecologia /ni vERSIDAD DE HONGOS SILVESTRES COMESTIBLES EN UN BOSQUE DE CONIFERAS (Abies religiosa), EN EL ORIENTE DEL VALLE DE MEXICO /- TESIS PROFESIONBL Que como Requisito parcial para Obtener el Titulo de INGENIERO EN AGROECOLOGIA DIRECO ION ACADEMICA DEPro . DE SE.AVICIOSESCOLAAES ORCINA OE EXAME NiS PRESENT A: lwlioteca de Agrcec I gfa - .. _ -- C. LJ!ONZ4LEZ C4BRER4 / J41HE CHAPINGO, MEXICO 1997
Transcript of Diversidad de hongos silvestres comestibles en un bosque de coniferas (Abies religiosa), en el...

UNIVERSIDBD BUTONOMB CHBPINGO
/
Programa Interdepartamental de Docencia, Investigaci6n y Servicio en Agroecologia
/nivERSIDAD DE HONGOS SILVESTRES COMESTIBLES EN UN BOSQUE DE CONIFERAS (Abies religiosa), EN EL
ORIENTE DEL VALLE DE MEXICO /-
/~
TESIS PROFESIONBL Que como Requisito parcial
para Obtener el Titulo de
INGENIERO EN AGROECOLOGIA DIRECOION ACADEMICA DEPro. DE SE.AVICIOSESCOLAAES
ORCINA OE EXAMENiS fROfQ.siO~ALES
PRESENT A: lwlioteca de Agrcec I gfa
- ~ .. _ --
C. LJ!ONZ4LEZ C4BRER4/ J41HE
CHAPINGO, MEXICO
1997
~I====-====-== ·~~•4l=n=·~~~4~Q~•4~·~~s;;;:j"'

Esta tesis se realiz6 bajo Ia asesorfa del M. C. luis Villarreal Ruiz; ha sido revisada y aprobada por el siguiente jurado examinador.
I ) ' L ). I I /,__ ./1 / ( . / j / . ' /) --1- / F' jY'Iu,,_ ,1:"t / / ! -~··Ll~ , .J "f 1/. 'vc_ (<·J\f'{/--'{.t"' '' i-;""--- ------- -- ~· --------- -:.=...::..::..: :-.... -----;:- -·-- -------------------------
M.C. Maria Sol Robledo Y Monterrubio Presidenta
\
lng. Gerardo Noriega Altamirano Secretario -
------------------------- /~~-~ ----~------------------------------Bioi. E mu7'do Perez . Godinez
j Vocal
---------------------------~_- ' 0 _ o o d~L ______ _ Bioi. l~gentAa}J~
\./ Suplente

El prcscntc trabajo sc encucntra adscrito al proyccto CONABIO C066, cl reportc final de dicha
invcstigaci6n es " Los hongos silvcstrcs: Componcntcs de la Biodivcrsidad y alternativa para la
sustcntabilidad de los bosqucs templados de Mexico" , cuyo editor fuc el M . C. Luis Villarreal Rui z, en
agosto de 1996.
Agradczco a la CONABIO por el financiamicnto del prcscntc trabajo, asi como al M.C. Luis
Villarreal Ruiz, por su asesoria en los mucstrcos de campo y 1aboratorio, y por habcr estado dirigiendo cstc
documento aproximadamente 21 meses.

AGRADECIMIE 'TOS
AI sistema de bccas y scryicios sin costo, para genic de bajos rccursos de Ia Uni,·crsidad
Autonoma Chapingo, por habcrmc pcrmitido culminar mis cstudios.
AI Programa Intcrdepartarncntal de Doccncia, Invcstigaci6n y Scrvicio en Agroe{:ologia
(PlDISA) por habcrme dado una fom1aci6n integral en cl inicio de mi cancra profesional , en particular
al Coordinador General, Lie. Carlos Ferra Martinez por su apoyo moral y acaC: .11ico mostrado durante
mi estancia en Agroccologia.
A los profcsorcs de Agroecologia, Enrique Cruz Lopez y Juan Antonio Cruz Rodriguez, por el
apoyo brindado para Ia culminaci6n de Ia tcsis.
AI lng. Gerardo Noriega Altamirano, gracias por apoyarme en Ia culminaci6n del presentc
trabajo y sus valiosos consejos a lo largo de mi fonnaci6n profesional.
A las siguientes personas par sus valiosas ideas plasmadas en cl presente trabajo, asi como su
gran disposici6n para finalizarlo: M.C. Mariasol Robledo y Monterrubio; M.C. Edmundo Perez
Godinez, Bioi. Ileana Ebergenyi M agaloni y Ihg. Juan Vidal Bello.
AI M.C. Luis Villarreal Ruiz, Je agradesco por haber estado pendiente del anteproyecto de
tesis, realizaci6n de trabajo de campo y laboratorio y parte de Ia escritura final, gracias por habcm1e
iniciado en este fascinantc mundo de los hongos, y por habcrme fom1ado como profesionista.
Gracias aJ Dr. Christian Prat, investigador del Colcgio de Postgraduados y Director del
proyecto CP- ORSTOM, desarrollado en !a comunidad de Santa Catarina del Monte, y en especial a
Alfonso Marquez Ramos; por haber facilitado los datos de precipitaci6n.
A los habitantes de !a Comunidad de Santa Catarina del Monte, Municipio de Texcoco, por su
disposici6n para el desarrollo del trabajo; en especial al sefior Lucas Reyes Romero por su apoyo
durante los muestros en campo.

DEDICATORlA
Especialmente a mis padres Guillermina y Jayier; a los cuales les dcbo todo lo que soy en Ia Yida,
gracias por cl apoyo y Ia confianza depositada en mi.
Con carii'io a mis hermanos por habcrnos apoyado mutuamente durante toda nuestra infancia,
Bruno, Guillermo, Cam1ela, con especial carino a mis hem1anos Agustin y Marco.
A mis amigos durante mi estancia e'. la UniYersidad, Alberto, Zacs, Gerardo, Tom, Macario,
Colima, Betancurt y Omar.
A mis companeros y amigos de Ia especialidad, gracias por tantos momentos compartidos: Miguel
C, Everardo, Coyo, Max, Rusbel.
Con especial afecto a mis hermanas y amigas: Nonna, Lulu y Sandra, tambicn a mi amiga
Concepcion por su apoyo.
A Chela, Marta, Lupillo y Pedro, por habcrmc ayudado en esta dura etapa de tesista.

CONTENIDO
RESUMEN
Espanol-------------------------------
lngles--------·------
Pagina
·-------H
------------------1
INTRODUCCION ----------------------------------------- --------1
ANTECEDENTES ---------------------------------------------3 Los Hongos Silvestres Componentes de Ia Biodiversidad -----------------------3
lmportancia Antropogenica de Ia Biodiversidad---------------------------------5
Va lor Co mesti ble-----------------------------------------------------------------5
Va I or R e ere at i v o-------------------------------------------------------------------6
Va I or Espiritu a 1-------------------------------------------------------------------------------6
Valor Medici na 1------ --------------------------------------------------8 Valor Texti 1----------------------------------------------------------------------8
Organism os Indica do res-------------------------------------------------------------------8
I mportan cia B ioeti ca----- ----------------------------------------------8 Carateristicas Biol6gicas de los Macromicetos--------------------------------------------------------9
C i cl o de Vida de u n Macro m i cet o-------------------------------------------------------------------9
F ase So m atica-------------------------------------------------------------------9
F ase R e prod u cti v a ----------------------------------------------------------------------11
Form a ci 6 n de I ca rp6 foro ----------------------------------------------------------11
Es po ru I a ci 6 n ---------------------------------------------------------------------------1 3
Forma s de C reci m i e nt o ---------------------------------------------------------------------------------13
Hong os Hi pogeos----------------------------------------------------------------13
Hongos Epig eos------------------------------------------------------------------13
Tipos de Sustrato--------------- --------------------------------------------14
Lig n r colas-----------------------~-----------------------------------------------14
Cop r6fi I as---------------------------------------------------------------------------------------14
Fen f co I a s-------------------------------------------------------------------------------------------------14
T e rrf co I as----------------------------------------------------------------------------------------------14
Hum f co I as----------------------------------------------------------------------------------------------14
G ru pos E co 16g i cos--------------------------------------------------------------------------------------15
Saprobios--------------.,.----------------------------------------------------------15
Simbiontes Mutualistas-------------------------------------------15
A

End om i cor rizas-------- -- ---------------------------- ------------ ------- --- --- ---------------- -1 6
Ectom i corriz as---------- ------------- ------------------------------------------ ----------------1 6
Sim bio nte s Antagonist as------------------------------------------- --- --------------------------- ----19
Di stribuci6n G eog ratica de los Hongos-------- -- --------- --- ---------------------------------------- ------ 20
lmportancia en el Bosque--- ----------------------------------------- --------------------------- ----------------21
Al imento de Ani males Mic6f agos---- --- ------------------------ ----------------------------- ------21
Est a b iIi z adores-------------------------------------------- -------------- --------------------------------21
Distri buci6n Espacio-Temporal------- --------· ---------- ------ --- --- --------- -- ------------------------------21
Tie m po ------------------------------- ------------- --------------------------- ------------------------------21
Es p a c i o- ----------------------- ------------------- --------------------------------------- ------------------22
Factores que A feet an Ia Productividad de los Hongos------------------------------- -- -- ------------22
End 6 g enos-----------------------------------------------------------------------------------------------2 2
Ex 6 g enos-------- -------- ------------- ------------- ------------------- --------------------- -------------- 23
Si I v f co I as------------------------------ ----- ------------------------,------------------------ --------------24
Inter acci6 n con M aero hong os---------------------- -------------------------------------24
Ed ad de I Arbo I ado---------------------------------------------------------------------------2 5
Forra jeo por M icrofauna------------------------------- ------------- -----------------------26
Inter acci6 n con M i croo rg ani sm os------------------------------------------------------26
Re co I e c c i 6 n In tens iva-------------------------------- ------------------ --------------- ------2 6
Bases para e I U so Sus t e nta bl e-- ------------------------------.------------------------------------------------2 6
I nve ntari o y M o nitoreo eco 16g i co-------------------------------------------------------- ---------------- -----2 7
De te c c i 6 n -------- ---------------------------------------------------------- --------------------------------2 8
Eva I u a c i 6 n --------------------------- ---------------------------------------------- ------------------------28
Invest i g a c i 6 n------ ------------------------- ---------------------------------------------------------------29
Perspectivas de Conse rv aci6n 8 iol6g ica --------------------------------------------------------------------29
OBJETIVOS------- ------------------------------------------------------------ ---- -----------------------------------32
General----------------------------------------------------------------------------- -----------------------------------3 2
Part i cuI ares-------------------------- -------------------------------------------------------------- --------- ----- ----3 2
HI PO TES IS-----------------------------------------------------------------------------------------------------------3 3
MET 0 DOl OG lA------------------------ ----------------------------------------------------- -----------------------34
Sel ecci6n del Are a de Estud io---------------------------------------------------------------------------------34
In st a I a c i 6n de I o s C u ad r os--------------------------------------------------------------------------------------34
Toma de Datos en Campo------------------------------------------------------------------------------------- 35
Tom a de Datos en Labor atorio--------------------------------------------------------------------------------3 5
ld en ti fica ci 6 n de I as Es p e ci e s ________ : ______________ -----------------------------------------------------------3 6
B

Datos C I i mat icos---------------------------------------------------------------36
P races ami ent o de Ia Inform a ci 6 n-----------------------------------------------3 6
DESCRIPCION DEL AREA DE ESTUDI0-----------------------------------------------------39
Loca lizaci6n G eog rMi ca ------------------------------------------------------------3 9
S u st rat o G eo 16g i co------------------------------------------------------------::---------3 9
Fisi og rafi a------------------------------------------------------------------41
Sue I o ---------------------------------------------------------------------------------------------41
C I i m a __________________________ :_ --------------------------------------------------------4 2
Caract e rf st i cas Hid ro 16g i ca s------------------------------------------------------------------------------4 2
V e g eta c i 6 n ---------------------------------------------------------------------------------------------------------4 3
Faun a --------------------------------------------------------------------------------------------------------44
Esce n a rio C u It u ra I y So ci oe con 6 m i co -----------------------------------------------------------------------44
S e rv i c i o s ----------------------------------------------------------------------------------------------4 5
Fest i v id ad es ---------------------------------------------------------------------------------46
Uso del Suelo y Actividades Productivas-----------------------------------------------------------------46
Ag ri cu It u ra -------------------------------------------------------------------------------------------4 6
G an ad e ria ----------------------------------------------------------------------------------------------4 7
S i I v f cu It u ra ----------------------------------------------------------------------------------------------4 7
R e co I e cci6 n de Hong os--------------------------------------------------------------------4 7
Act i vi dad es Com pie menta ri as---------------------~--------------------------------------------------48
RESULT AD 0 S Y DISCUS 16 N ---------------------------------------------·-- ------------------------------------4 9
Especies de Hongos ldentificadas en el Bosque de Abies-----------------------------------------------49
S u st rat o de I as Es pe ci e s En con t rad as-----------------------------------------------------------------------52
I m porta n cia Ant ro pog en i ca ------------------------------·---------------------------------------------------------54
Import an cia E co 16g i ca -------------------------------------------------------------------------------------------57
Poblaci6n de Macromicetos en el Bosque de Abies-----------------------·-------------------------------59
Dive rsi dad en e I Bosque--------------------------------------------------------------------------59
T as a de D esco m pas i ci 6 n ------------------------------------------------------------------------------6 2
Es p e c i e s Abu n dante s ----------------------------------------------------------------------------------61
P rod u ct i vida d ---------------------------------------------------------------------------------------------6 7
Macro m i cet os Domin ant es-------------------------------------------------------------------------7 0
Especies de Mayor Liberaci6n de Nutrimentos---------------------------------"------------------71
Comportamiento de Ia Producci6n ante Factores Ambientales-------------------------------71
Period o de F ru ct ifi ca ci 6 n -----------------------------------------------------------------------------7 4
Rit m o de F ru ctifi ca ci 6 n de 1 as Es peci e s------------------------------------------------------------7 5
c

CONCLUSIONES
RECOMENDACIONES---
LITERATURA CONSULTADA
AN EX OS
------------------------78
--------------------- --------79
---------------------------------80
Anexo 1. Terminos Basicos de Ia Tesis.
Anexo 2. Hoja de registro de toma de datos en campo.
Anexo 3a y 3b. Hoja de registro de caracteristicas macromorfol6gicas.
Anexo 4. Oatos Climaticos, (A) . Datos de precipitaci6n; y (8) . Temperaturas
estimadas, (C). Producci6n en carp6foros totales.
Anexo 5. Macromicetos comestibles apreciados.
Anexo de Ia investigaci6n.
i nd ice de Figura s------------- - ------------------:------------------------------------------------- E
l n dice de C u ad ros ------------------------ --------------------------------------------------G
D

indice de Figuras Pagina
Figura 1. Representaci6n del ciclo de vida general de los macrohongos, con
requerimientos exogenos y endogenos en sus fases; modificado de Statemes (1993).--10
Figura 2. Localizaci6n del area de estudio, en Ia comunidad de Santa Catarina del Monte, en el
Oriente del Valle de Mexico. - · -----------------------------------------40
Figura 3. Bongos silvestrcs del bosque de Abies, a nivel de familia , en Ia comunidad de Santa
catarina del Monte, al Oriente del Valle de Mexico.-----------------------------------------------50
Figura 4. Porcentaje de las especies encontradas en cada tipo de sustrato, en el bosque
de Abies, en el Oriente del Valle de Mexico.-- ---------------------------------53
Figura 5. lmportancia antropogenica de los hongos estudiados en el bosque de Abies, en
el Oriente del Valle de Mexico.---- -------------------------------------------------55
Figura 6. lmportancia ecol6gica de los hongos estudiados en el bosque de Abies, en el
Oriente del Valle de Mexico.-------------------- -r-------------------------------------------58
Figura 7. Numero de carp6foros sanos y daf\ados de las especies silvestres en el bosque
de Abies, en el Oriente del Valle de Mexico.-----------------------------------------------62
Figura 8. Abundancia de especies en numero de carp6foros sanos en el bosque de Abies,
en Ia comunidad de Santa Catarina.-------- -----------------------------------------64
Figura 9. Abundancia de especies en peso fresco de carp6foros sanos en el bosque de
Abies, en Santa Catarina del Monte, localizada al Oriente del Valle de Mexico.------------65
Figura 10. Producci6n en peso fresco (g) en las parcel as de estudio durante 1995, en el
~ iente del Valle de Mexico.------------------- -----------------------------------------66
E

Figura 11. Producci6n en numero de carp6foros en las parcelas de estudio durante 1995,
en el Oriente del Valle de Mexico. --------------- -----66
Figura 12. Temperaturas medias estimadas para Ia zona de estudio, en grades oc durante
1995.-------------------------------------------------------------------71
Figura 13. Precipitaci6n ''luvial en mm durante 1995 y carpoforos totales en Ia zona de
estudio, localizada al Oriente del Valle de Mexico.---------------------------------------------72
F

fndice de Cuadros Pagina
Cuadro 1. Especies de hongos identificadas de Ia subdivision Ascomycotina, en el
Oriente del Valle de Mexico, en Ia comunidad de Santa Catarina del Monte, en el municipio de
Texcoco.- --------- ---------- --------- -------------------------------------------49
Cuad ro 2. Especies de hongos identificadas de Ia subd 'visi6n Basidiomycotina, en el
Oriente del Vall e de Mexico , en Ia comunidad de Santa Catarina del Monte, en el municip io de
T ex co co. ------------------------------ -------------------------- --------------··----------------------------4 9
Cuadro 3. Tipo de sustrato de los hongos silvestres estud iados, en el bosque de Abies, en
ia comunidad de Santa Catarina del Monte.------------------------------------------------------------52
Cuadro 4. lmportancia Antropogenica y ecol6gica de las especies de hongos estudiadas
en Ia comunidad de Santa Catarina del Monte, ubicada al Oriente del Valle de mexico.------54
Cuadra 5. Clasificaci6n de Ia toxicidad de macromicetos, modificado de Aroche et a!.
1 9 8 4 . -------------------------------------- --------------------------------------------------------- ------------------56
Cuadra 6. lmportancia ecol6gica de las especies silvestres encontradas en el bosque de
Abies----------- ---- ---- ------ ----------------------------------------------------------------------57.
Cuadra 7. Mementos de fruct ifi caci6n de las especies de macromicetos, en el bosque de
Abies , en Santa Catarina del Monte, al Oriente del Val le de Mexico.--------------------------------75
G

RESUMEN
El presente estudio, se realiz6 durante el ano de 1995, en Ia comunidad de Santa Catarina
del Monte, Mpio . de Texcoco, Edo . de Mexico; ubicada al Oriente del Val le de Mexico. Esta area
se eligi6 por los reportes existentes acerca de su gran conoci miento y riqueza de hongos
comestibles.
El objet ivo fue realizar un diagnostico sobre Ia diversidad y riqueza de especies de hongos
silvestres comestibles en un bosque de Abies religiosa, pensa ndo en un futuro man ejo sustenta ble ;
para ello se uti liz6 una area de muest reo de 0.5 hectareas, divid ida en 5 parcelas de 20 x 50
metros. Los muestreos se desarrolla ron de junio a diciembre, con intervalos de 15 dfas.
En el bosque de Abies, se registr6 una riqu eza de 30 especies de macromicetos , con un
fndice de diversidad de Shannon-Wiener (H') de 3.8, de las especies presen!es 28 (93.3%) son
comestibles, una es t6xica (3 .3%) y una (3 .3%) no ti ene valor alimenticio ; ecol6gicamente se
caracterizan : 56.6% son deg radadoras, 40% son simbiontes mutualistas y 3.3% son simbiontes
antagonistas; con respecto al sustrato , el domi nante es el terrfcola (45%), le sigue el humfcola
(31 %) , lign fcola (17%) yen menor proporci6n (3%) Ia especie parasita registrada sobre un hongo .
La epoca de fructificaci6n, fue de junio a noviembre con una cima en fructificaci6n y
producci6n (peso fresco y numero de carp6fo ros) durante el mes de agosto ; se registraron 1,178
carp6foros sanos que tota lizaron 63 ,23 kg/ha·1 de peso fresco. La tasa de descomposici6n
estimada fue del 6%.
La riqueza de especies detectada representa un almacen gen etico , disponible para
micologos y Ia industria de producci6n de hongos, no obstante es prudente continuar realizando
estudios de esta naturaleza , que son base para el disefio de un aprovechamiento sustentable .
H

SUMMARY
The present study was carried through the yea r of 1995, in the town of Santa Catarina Del
Monte, district of Texcoco, Mexico State , located at the orient of Valle de Mexico. This zone was
preferid by the reports about of its bigger kn owledge and wealth of edibles mushrooms.
The goa l was realize a diagnostic about the diversity and wealth of species of wild edibles
mushrooms in wildwood of Abies refigiosa , thinking in futu re sustained manag ement , by this was
used an area of 0.5, divided in 5 plots of 20 x 50 m. The sampling was carried of june to
december of 1995, with intervals of 15 days.
At wildwood of Abies, was registered a wealth of 30 species of macrofungi, with one index
of diversity of Shan non-Wiener (H') of 3.8; of the species 28 (93 .3%) are ed ibles, one is poison
(3 .3%) and other (3 .3%) without worth edible ; ecologyment were sorted 56 .6% as saprobes , 40 %
are mutual istic symbiosis and 3.3% are pathogens symbiosis, with relation to the su bstrate ,
dominant is the earthling (45%), next the species above the humus (31 %), woodlife (17%) and the
least (3%) the species pathogen rendered in a mushrooms.
The station of fruiting , was since june to no,vember with top in fruiting and production
(weight fresh and number of fruiting bodies) th rough the month of aug ust ; were registered 1,178
frui tingbodies without harm, that added 63 ,23 Kg/Ha 1 of weight fresh . The ratio of decomposition
considered was of 6%.
The wealth of species detected , represent on stock genetic, available for micologys and the
industry of production of mushrooms, however is prudent go on realizing studys alikes, that are
ground for the design of a sustained management.

INTRODUCCI6N
Mexico posee alrededor de 2,000,000 km 2 de extensi6n territorial, en opini6n de
Rzedowski ( 1991), los bosques de encino y conifer as ocupan aproximadamente el 21 % de
esta superficie . Las variaciones en condiciones fisiograficas y altitudinales que caracterizan a
nuestro pafs, permiten que estos bosques presenten un mosaico vegetal rico en diversidad
- '1 rfstica y ecol6g ica (Cibrfan-Tovar eta/. 1995), Ia cual es superior a p~lses mas grandes en
extension territor ial, pero de menor diversidad en su fisiograffa; Groombridege (1992), senala
que un area con un mosaico de sistemas tendra mayor biodiversidad que una area clfmax.
De Ia biodiversidad existe un elemento poco estudiado, los macromicetos que
contribuyen a mantener Ia estabilidad de los ecos istemas forestales, debido a dos hechos: (a)
son los principales recicladores de nutrimentos (Dighton y Boddy, 1989; Duran-Degrau y
Lluvia, 1980); y (b) son organismos claves para el desarrollo y sobrevivencia de los arboles
(Le-Tacon, 1985; Stone, 1996).
Los macromicetos son importantes para Ia vida del hombre, ya que son aprovechados
mediante Ia recolecci6n por comunidades rurales asentadas en los bosques, constituyendose
en una fuente de ingreso mediante Ia venta y parte importante de Ia estrategia de subsistencia
(Villarreal y Perez-Moreno, 1989; Zamora-Martinez y Pascuai-Pola, 1995), para el resto de Ia
sociedad, radica en el uso como alimento natural (Kaul, 1987; Read eta/. 1989); como fuente
de inspiraci6n en actividades creativas: fotograffa, dibujo y pintura (Arnolds, 1988; Charles,
1974) .
Estimaciones de Guzman (1995), sobre los hongos que presumiblemente se
encuentran en el pals, resultan en 120,000 a 140,000 especies; solo se conocen 6,000
especies, que representan el 4 .5 a 6.6% de especies fungicas regist radas. Esto indica que
nuestro pals ha sido poco explorado con respecto a Ia flora micol6gica (Guzman, 1994;
Herrera, 1994), esta situaci6n rescata que los estudios en los macromicetos son escasos;
segun Conrado-Soto ( 1997), ocupamos el quinto Iugar mundial en riqueza de macromicetos
comestibles. Villarreal y Perez (1989), reportan 204 especies de hongos silvestres comestibles
en los bosques mexicanos; en este conocimiento etnico y rural se advierte un proceso de
degradaci6n, ocasionado por el avance de Ia modernidad y por los procesos de
transculturaci6n (Villarreal y Perez, 1989; Aroche eta/. 1984).

ANTECEDENTES
los Hongos Silvestres Componentes de Ia Biodiversidad
La palabra biodiversidad es una contracci6n de diversidad biol6gica, su uso es comun
como sin6nimo de riqueza de especies; en forma sencilla esta palabra incluye a todos los
seres vivos que habitan Ia tierra, desde los grandes mamiferos hasta los microorganismos
( ~roombridge, 1992). Los hongos forman parte de Ia diversidad microbiana y cuntribuyen
con el 4.2% de Ia biodivesidad global, estimada en 1. 7 millones de especies de seres vivos
que habitan el planeta (op. cit.). Hawksworth (1991), estima en 1.5 millones Ia diversidad del
rei no Fungi, de los cuales s61o se conoce el 5%.
Cuando se habla de Ia diversidad fungica de un ecosistema o un habitat especifico, se
refiere a Ia diversidad local o diversidad Alfa (Whittaker, 1972), Ia cual incluye riqueza de
especies y abundancia (Allen et a!. 1995; Bruns, 1995). Se reconoce que Ia diversidad en
nuestro planeta es variable, dependiendo de Ia regi6n, por lo general Ia diversidad de especies
es alta en zonas con abundante precipitaci6n, bajas altitudes y latitudes mfnimas,
caracteristicas que corresponden a las regiones tropicales, siguiendo en importancia las zonas
templadas. Actualmente se estima que alrededor del 7% de las areas terrestres del mundo
contienen 90% de las especies conocidas (Groombridge, 1992).
La perdida de biodiversidad atribuible a Ia extinci6n de especies es un fen6meno que
ocurre en forma natural en el planeta, sin embargo, actualmer1te las actividades
antropogenicas han provocado el incremento de este ritmo, sobre todo, si se compara con
cualquier otra epoca del pasado; este incremento se asocia a Ia expansi6n de las actividades
humanas (Groombrigde, 1992). Los efectos del desarrollo de Ia humanidad sobre Ia
biodiversidad se pueden dividir en dos tipos: (a).- directos: caza y recolecci6n; y (b).
indirectos: modificaci6n y destrucci6n del habitat.
En general las actividades humanas modifican el ambiente, reducen Ia abundancia de
especies y en casos extremos las extinguen cuando el habitat se torna inadecuado; de
conformidad con los calculos mas conservadores, se estima que se han extinguido 12.5
millones de especies y en el rei no Fungi se cita Ia citra del 8% (Groombridge, 1992). Es un
hecho que Ia perdida de Ia biodiversidad de nuestro planeta es irreversible, hoy en dia existe el
peligro de perder no s61o especies: sino Ia totalidad de los ecosistemas de los que depende Ia
raza humana (Mittermeier-Russell y Mittermeir, 1992).

El papel evolutivo de los hongos en el planeta se remonta al inicio de Ia vida, donde
sus ancestros hicieron posible Ia aparici6n de las primeras plantas terrestres, producto de Ia
asociaci6n simbi6tica mutualista entre una alga primitiva y un hongo endomicorrizico (Azc6n y
Barea, 1980; Hawksworth, 1991; Pirozynski, 1981 ); esto les permiti6 Ia dispersion al
proveerles ventajas adaptativas que les permitieron crecer y desarrollarse (Hawksworth,
1991). En Ia mitad del Cretacico aparecieron las ectomicorrizas, que al extenderse,
permitieron a las plantas colonizar desde los tr6picos hasta las regiones templadas, con
fluctuacionf'> de climas y sue los pobres (Hawksworth, 1991; Pirozynski, 1981).
En Ia actualidad los hongos, por su nutrici6n saprofitica son los mayores degradadores
de sustancias organicas en los ecosistemas terrestres (Alexopoulos, 1977; Dighton y Boddy,
1989; Duran-Degrau y Lluvia, 1980), seguidos en importancia por las bacterias; estos
organismos son los mas importantes componentes en el ciclo de nutrimentos, atribuible a su
metabolismo, ya que segregan enzimas que transforman los compuestos organicos complejos en
productos mas sencillos (Margalef, 1974; Mcnaughton y Wolf, 1984; Witkamp, 1966). Asi Ia
producci6n primaria neta que va directamente a los degradadores en los sistemas terrestres es
de 90 a 100%, en opini6n de Alexopoulos ( 1977); Christensen ( 1989) y Mcnaughton y Wolf
( 1984), si las bacterias, actinomycetes y los hongos, no realizaran su papel ecol6gico se
acumularian centimetros de materia organica, llegando a asfixiar todas las cosas vivas,
entonces Ia raza humana para sobrevivir deberfa apre_nder a vivir sobre estos restos organicos.
Un componente importante de Ia diversidad fungica Ia constituyen los hongos
superiores de las subdivisiones Ascomycotina y Basidiomycotina, que producen estructuras
reproductoras macrosc6picas; debido a su tamario visible a simple vista, se les conoce como
macrohongos (Chang y Miles, 1989; Kaul, 1987). Arnolds (1992) define a los macrohongos
como aquellos que producen carp6foros macrosc6picos, visibles individualmente, mas grandes
que 1 mm.
Aproximadamente 5,000 especies de macromicetos se asocian con arboles forestales
en las regiones boreales y templadas (Le-Tacon, 1985); existen macromi~etos en los bosques
tropicales, pero mal representados (Valenzuela eta!. 1981).
Los macrohongos se dividen en saprobios y simbiontes (Molina et a/. 1993): (a) los
saprobios, son los principales recicladores de nutrimentos en los ecosistemas forestales
(Arnolds, 1988; Dighton y Boddy; 1989); (b) los simbiontes son organismos clave para los
arboles forestales, ya que los mantas micorrfzicos incrementan Ia extensi6n del sistema
4

radicular, mejorando Ia nutric i6n vegetal e increm entando Ia res istencia a plagas y
enfermedades, lo que favorece el crecim iento y el desarrollo vegetal (Christensen, 1989;
Fogel, 1992; Hacskaylo, 1983; Le -Tacon, 1985; Mason et a!. 1983; Read, 1991; Stone,
1996).
Los bosques son fundamentales en varios procesos como Ia continuidad del ciclo del
agua, carbono, nitr6geno, entre otros elementos y sostienen una diversidad biol6gica necesaria
para Ia alimentaci6n humana, amen de ser fuente de bienes y servicios para el hombre
(Groom bridge, 1992), las actividades humanas afectan a Ia biosfera a escala global, un
ejemplo es el incremento de los niveles atmosfericos de C0 2 por Ia constante quema de
combustibl es f6s iles; con Ia destrucc i6n de los bosques se agudiza este problema, ya que ellos
de forma natural fijan carbono durante Ia foto sfntesis.
Como conclusion a lo expuesto, es evidente el papel que tienen los hongos en Ia
bi6sfera, participando en procesos biol6gicos y auxiliando a mantener Ia estabilidad ecologica.
lmportancia Antropogenica
Los macrohongos constituyen un elemento importante en Ia cultura del hombre, es
decir en el estilo de vida, asf Ia importancia de los hongos se identifica en: (a)- valor
comestible; (b)- valor recreative; (c)- valor espiritual;· (d)- valor medicinal; (e)- valor textil; (f)
organismos indicadores; y (g)- importancia bioetica.
Valor Comestible. A nivel mundial, Ia practica del aprovechamiento de los hongos silvestres
como alimento se remonta a los inicios del mismo hombre, uno de los registros escritos con
mayor antiguedad se localiza en Ia India, data alrededor de 500 af\os A·. C. (Kaul, 1987), Ia
domesticacion de hongos y su cultivo se remonta a 1 000 af\os en tal pafs (Chang y Miles,
1989). Kaul ( 1987), menciona que existen cerca de 2000 especies comestibles, solo 20 de
elias son cultivadas a nivel mundial, de ahf el valor de los hongos silvestres. Por otra parte, Ia
incapacidad para reproducir Ia simbiosis micorrfzica bajo condiciones controladas, salvo raras
excepciones esta se considera Ia principal razon del alto valor de ias especies silvestres
micorrfzicas que solo pueden recolectarse en bosques naturales (Stone, 1996).
En Mexico, los hongos silvestres comestibles se utilizan desde Ia epoca prehispanica,
asf los aztecas empleaban alrededor de 50 especies de hongos comestibles (Guzman, 1984).
Este recurso ha sido yes complementario a las dietas alimenticias de las comunidades rurales,
5

asi como fuente de ingresos por concepto de Ia venta durante Ia epoca de lluvias (Guzman,
1984; Villarreal y Perez, 1989; Zamora-Martinez y Pascuai-Pola, 1995).
En general a los que se dedican a recolectar hongos para Ia venta se les conoce como
"hongueros"; quienes pose en un amplio conocimiento em pi rico de los hongos; conocen
lugares donde fructifican, epoca, nombres vernaculos y los distinguen de las especies t6xicas
e incluso elaboran sistemas de clasificaci6n tradicional ! ~ispert et a!. 1984; Guzman, 1984;
Mapes et a!. 1981; A roche et a/. 1984). Algunos investigadores inform an de 204 especies de
hongos silvestres comestibles existentes en el pais, se advierte un proceso de degradaci6n del
conocimiento empirico, atribuible a Ia modernidad y a los procesos de transculturizaci6n que
padecen las etnias que habitan las regiones boscosas de Mexico (Aroche, et a/. 1984;
Villarreal y Perez, 1989.
Existen especies de hongos silvestres cuyo consume es significative, por lo que son
amplia e intensivamente recolectados (Read et a!. 1989); Ia recolecci6n de los carp6foros
constituye una actividad productiva que requiere de esfuerzo, asi un recolector cubre
alrededor de 11 .3 ha durante un periodo de 8 a 12 horas para acopiar 6 kg/dia (Zamora
Martinez y Pascuai-Pola, 1995); a partir de ahi aparecen los intermediaries de Ia
comercializaci6n, quienes obtienen los mayores bene,ficios econ6micos.
El uso de los hongos como alimento varia de regi6n a regi6n, existiendo clasificaciones
regionales y/o locales, que les estratifica en comestibles y no comestibles; existen especies
con bajos niveles de compuestos t6xicos, a las cuales con un tratamiento de remojo y/o
cocci6n son eliminados, por ejemplo Lactarius rufus en Finlandia al lavarse con agua a
temperatura ambiente queda libre de toxinas (Wasterlund y lngelot, 1981). En los bosques, el
numero de generos y especies venenosas es bajo, por ejemplo Garza et a!. (1985), reportan
un estudio realizado en un bosque de Quercus rysophylla, donde se encontr6 el 10% de
especies venenosas.
Existen factores de confusion para asignarles un valor monetario exacto a los hongos
silvestres, sin embargo se tienen estimaciones que muestran su valor, Norvell (1995),
menciona que los hongos silvestres comestibles y otros productos secundarios del bosque
ofrecen mas beneficios que los arboles mismos, esto en el marco de Ia vision del desarrollo
sustentable, asi se ha estimado que Ia recolecci6n de una sola especie (Cantarellus cibarius)
supera en valor econ6mico al valor de Ia madera producida en el mismo periodo; Zamora
Martinez y Pascuai-Pola ( 1995 l, reportan una producci6n anual valuada en 1 000 d61ares en
6

base a dos especies de hongos comestibles Morchel/a elata y Lyophyllum decastes
recolectadas en dos parcelas de Pinus montezumae, cuya dimension fue de 20 x 20 m2
por
sitio. Estas especies silvestres constituyen un potencial como fuente de alimento, pensando
en Ia futura domesticaci6n (Arnolds, 1988), cabe destacar que Ia diversidad actual de hongos
sirve como material genetico para obtener fenotipos con caraterfsticas ffsicas deseables por el
hombre.
Otro valor de los hongos es el cultivo en sustratos de "desecho" como esquilmos ie
trigo, coco, cebada, mafz, cafe, entre otros, proporcionando un producto con alto valor
nutritivo (Rambelli, 1985), al mismo tiempo que liberan los nutrimentos almacenados en estas
estructuras, caso contrario estos materiales son nocivos para el ambiente al arrojarse a rios,
lagos o terrenos desocupados. Delmas (1987a), reporta una producci6n mundial de
1,400,000 toneladas de hongos cultivados, estimando que Ia producci6n alcanzara los 2
millones de toneladas para el ano 2000.
Valor Recreativo. Los hongos en opinion de Arnolds (1988) y Charles (1974), son interesantes
y hermosos componentes del ambiente, ofrecen atracci6n a actividades creativas, como Ia
fotograffa, dibujo, pintura y arreglos florales; un ejemplo de Ia ultima actividad es realizado en
Ia UACH par Ia profesora Clara Alfaro-Maya (1996), quien diseno arreglos decorativos con
ejemplares de macromicetos en etapa de fructificaci6n. En los pafses desarrollados el
aprovechamiento de los hongos silvestres comestibles cumple una funci6n recreativa
(Arnolds, 1988; Markkula y Rantavaara, 1996), por ejemplo en Finlandia se recolectan hongos
como una actividad de tiempo libre en los bosques aledanos a las ciudades.
La importancia recreativa data de 195 7, a no en que se acun6 el termino mic6filo
(mycophilia), para describir a Ia gente que su vida gira en torno de los hongos, ya sea
estudiandolos (mic61ogos), o apreciandolos como aficionados, colecciona~do timbres postales,
Iibras, obras de artes, artesanfas, pinturas o fotograffas (Molina eta/. 1993).
Valor Espiritual. La importancia de los hongos tambien radica en el sentido mfstico que le han
conferido algunos pueblos, tal es el caso de Mexico, al respecto Dubovoy (1968) y Schultes
(1982), mencionan que se utilizaban en ceremonias magico-religiosas de pueblos indfgenas
con el fin de comunicarse con sus dioses, en esta comuni6n se les revelaba el porvenir y Ia
manera de resolver problemas, estos hongos eran llamados "Teonanacatl"; mas de 20
especies eran empleadas para este· proposito, en Ia epoca actual Guzman (1984), reporta una
iglesia en Chignahuapan, Puebla, dedicada al culto de un hongo "Nuestro Senor del
7

Honguito". Los testimonies del papel mistico que han jugado los hongos en el Mexico antiguo
han quedado en forma de esculturas de piedra, pinturas y c6dices (Gispert et a/. 1984;
Guzman, 1984).
Valor Medicinal. Otro uso reportado, pero poco documentado, es el medicina, Molina et a/.
( 1993), mencionan que los nativos americanos utilizaban esporas de hongos para reducir
inflamaciones e infecciones, tambien los consumfan como t6nicos y en preparados
combinados con plantas medicinales. En Ia medicina moe' .rna se esta acumulando evidencia
de que algunas especies de hongos macromicetos contienen estimulantes que fortalecen al
sistema inmunol6gico del cuerpo, por ejemplo en experimentos realizados con ratones se ha
logrado reducir los tum ores cancerigenos (op. cit.).
Valor Textil. Este uso es reciente, algunas especies constituyen una fuente natural de tintura
para tenir lana, seda u otras fibras (Rice y Beebee, 1980; Molina et a/. 1993), tambien se
puede elaborar tinta y papel de ellos (Molina eta/. 1993).
Organismos lndicadores. La presencia de varies grupos ecol6gicos de macrohongos sirven
para indicar el grade de algunos parametres ambientales tales como: Ia contaminaci6n del
aire, condiciones de suelo, tipo de manej-o y- gfadcr de- disturbio (Arnolds, 1988); lo anterior
puede ser util para hacer mas precisa Ia caracterizaci,6n de cierto habitat, delimitando tipos de
comunidades arb6reas presentes dentro del bosque (Arnolds, 1988; Bujakiewicz, 1982). Cada
tipo de bosque (encino, haya, confferas) tiene su micobiota caracterfstica, teniendo especies
exclusivas y sobrelapadas (Bujakiewicz, 1982; Hudson, 1980).
lmportancia Bioetica. Este motive vale por todos los demas, ya que conservar el almacen de
genes, es necesario para entender los procesos evolutivos; Ia humanidad es responsable de
que continuen existiendo las formas de vida que han tardado millones de anos en formarse
(Arnolds, 1988). Los mexicanos tenemos el compromise ineludible de asegurar Ia
perdurabilidad de Ia diversidad nacional (Rzedowski, 1991).
Con el desarrollo de Ia humanidad constantemente cambian las necesidades, formas
de vida sin valor aparente en el pasado hoy resultan recursos de valor, como ejemplo podemos
citar al petr61eo (Groom bridge, 1992). Con el constante aumento de Ia poblaci6n muchos
organismos no valorados pueden volverse importantes en un futuro cuahdo se descubra sus
valores, es decir los recursos no cteberian ser perdidos porque no sabemos su valor presente,
8

en opinion de Molina et a/. ( 1993) nuestra cultura actual es baja para percibir el valor
potencial de los hongos.
Los valores bioeticos son poderosos argumentos para conservar Ia biodiversidad,
aunque pueden ser de poca relevancia con Ia imperiosa necesidad de sob'revivir en un mundo
donde escasean los alimentos (Groom bridge, 1 992).
Conservar Ia divers1Jad biol6gica de los macrohongos, se justifica por motivos
culturales, econ6micos, el uso actual que presentan estos organismos al hombre, el uso
potencial a futuroy porque son necesarios para Ia existencia de Ia biosfera y Ia vida humana.
Caracterfsticas Biol6gicas de los Macromicetos
Los macromicetos forman un grupo de seres vivos, que poseen caraterfsticas
biol6gicas comunes, tales como: ciclo de vida, forma de crecimiento y grupos ecol6gicos.
Cicio de Vida de un Macromiceto. AI referirse a hongos silvestres comestibles al lector no
adentrado en el tema, le viene a Ia mente un champin6n (Agaricus bisporus), esto por ser Ia
especie mas cultivada y comercializada a nivel mundial (Delmas, 1987a), lo que Ia hace Ia
especie mas conocida. Este genero sirve para representar el ciclo de vida de los macromicetos
en general, el cual se divide en dos fases fundamelltales: somatica y reproductiva (Delmas,
1987b; Kaul, 1987), existiendo parametros ambientales y end6genos propios de cada fase; en
Ia naturaleza Ia sucesi6n climatica presenta condiciones 6ptimas para cada etapa en forma
cfclica, cerrandose asf el ciclo bio16gico (Kaul, 1987), como se exhibe en Ia Figura1 que
presenta el ciclo de vida general de los macrohongos.
Fase Somatica. Se inicia con Ia germinaci6n de una espora sobre un sustrato adecuado
que en presencia de condiciones ambientales propicias produce filamentos microsc6picos
llamados hifas (Singer, 1961 ); estas estructuras poseen un nucleo haploide.
Las hifas crecen individualmente, bifurcandose y ramificandose lateralmente por Ia
fusion con otras hifas, formando asf un micelio, el cual bajo condiciones 6ptimas presenta un
crecimiento circular; un ejemplo visible en los bosques son los llamados "cfrculos de hadas"
(Kent y Mcknight, 1987; Singer, 1961 ). Las partes apicales de una hifa estan constitufdas por
unB mas.a c.itop!asmatica que se mueve dentro de elias, este mov1'm1'er1to de! Ct't6piasma
activo es esencial para adquirir las fuentes organicas necesarias para un rapido crecimiento
(Wessels, 1991), este micelio primario es monocari6tico, el cual se fusiona con otro micelio
9

Liberaci6n de esporas
L_
I
Desarrollo del carp6foro
Textura areno-limosa,
alto contenido de M.O . y pH acido
Reduci6n de
5 a 1 oc C
'r 0 a 40° C de temp. 6ptimo ae 25 a 35"
Figura 1. Representaci6n del ciclo de vida general de los macrohongos, con
requerimientos exogenos y endogenos en sus fases; modificado de Stamets ~ 1993).
sexualmente compatible {plasmogamia), despues del cual los nucleos retienen su
individualidad, asociandose en pares que pueden propagarse por un corto o indefinido tiempo,
dependiendo de Ia clase de micelio; en el caso de ascomicetos, esta fase tiene un perfqdo de
vida restringido, el cual es dependiente del micelio primario; en cambia en los basidiomicetos,
es posible un crecimiento vegetativo indefinido. Posterior a Ia plasmogamia se presenta Ia fase
10

dicari6tica (disponibilidad de genes), Ia cual es el requisito normal para Ia rep roducci6n sexua l
(Kaul, 1987).
Durante el proceso de desarrollo de Ia hifa de los hongos saprobios, se produce Ia
degradaci6n de los componentes organicos, y se realiza Ia nutrici6n del hongo, estos
nutrimentos son aprovechados como fuente de energfa y como proveedores de carbono para
Ia biosfntesis, los compuestos simples son transportados a traves d~ Ia membrana, los
compuestos complejos deben ser degradados a mon6meros fuera de Ia celula y esto se logra
por enzimas extracelu:ares que son liberadas a traves de las paredes (Kaul, 1987), estos
carbohidratos son empleados para producir mas micelio (Bruns, 1995).
En el caso de los hongos micorrfzicos, en el proceso de infecci6n las nuevas rafces
finas de Ia planta hospedera son rapidamente cubiertas por el micelio y tan pronto como
emergen, secretan enzimas que permiten Ia invasio n de las regiones intercelulares de Ia
corteza, asf una masa de hifas se vuelve una red de Hartig debido a cambios morfol6gicos
(Bruns, 1995; Hacskaylo, 1983).
El exito o abundancia de los micelios en el bosque es influenciada por el intervalo de
crecimiento, Ia tolerancia a extremos abi6ticos, Ia susceptibilidad al co!'sumo, Ia capacidad
para inhibir y para Ia producci6n de antibi6ticos; este se vuelve inactivo cuando las
condiciones ambientales son adversas, pero revive bajo condiciones favorables (Hudson,
1980),asf se conoce de algunos que tienen hasta 750 anos de edad (Arnolds, 1992).
Fase Reproductiva. Esta fase de los macromicetos, se diferencia en dos partes,
formaci6n del carp6foro y esporulaci6n.
Formaci6n del carp6foro. La capacidad de fructificaci6n es end6gena (Delmas, 1987b),
es decir, escrita en el programa genetico de las celulas del hongo; esta fase es
necesaria para renovar los caracteres de los organismos. Se caracteriza por Ia
aparici6n de cuerpos fructfferos propios de cada especie (!bar, 1980; Kent y Mcknight,
1987), los cuales son resultantes de Ia informacion hereditaria, interactuando con el
ambiente (Kaul, 1987).
La transformaci6n de crecimiento micelial a fase reproductiva ocurre por cambios de
varies factores ambientales, causados por Ia sucesi6n climatica, estos actuan como un
estfmulo transtornando el metabolismo (Delmas, 1987b; Kaul, 1987), para que exista
11

este cambia, se requiere un mfnimo lapso de crecimiento vegetat ive antes de que el
macrohongo pueda producir cuerpos fructfferos (Kaul, 1987) .
El cam bio fisiol6gico para el inicio de Ia fructificaci6n, se presenta cuando las hifas
dejan de crecer regularmente, los segmentos dejan de multiplicarse, se agrupa el
micelio y se destingue una nueva estructura que externamente se parece a una masa
algodonosa (Delmas, 1978b); las hifas emergen de esta agrupaci6n formando
primordios, los cuales son Ia etapa inicial de madurez de los carpofon . ..; ; el material
necesario para formar estas estructuras es obtenido de Ia desviacion masiva de
productos de Ia sfntesis hacia esa estructura; esta redistribuci6n de las reservas del
hongo es lo que permite formar las estructuras reproductoras (Kaul, 1987; Wessels,
1991). Los primeros carp6foros son los mejores desde el punto de vista biol6gico y de
crecimiento (Singer, 1961), Ia producci6n provoca que el micelio se agote, necesitando
un largo perfodo para recuperarse (Hawker, 195 7; Singer, 1961 l.
Delmas ( 1987b), indica que Ia formacion de los cuerpos fructfferos es fundamental para
realizar el proceso sexual de los macromicetos, en esta fase se comienza a guardar Ia
informacion genetica del hongo en estructuras reproductoras sexuales inmaduras,
denominadas esporas, que por Ia forma en que se encuentran dentro de los cuerpos
fructfferos, se dividen en dos grandes grupos (Duran-Degrau y Lluvfa, 1980): ( 1 l
Ascomicetos, las contienen en una estructura llamada asca, generalmente en numero de
8 esporas; y (2)- Basidiomicetos, se encuentran en el exterior de celulas llamadas
basidios, generalmente en numeros de 4 esporas.
La fructificaci6n se induce de dos formas: (a)- los factores ambientales alcanzan un
nivel acumulativo; y (b)- que alguno de ellos cause un dano al metabolismo, asf
tambien existe fructificacion cuando alguna condici6n atmosferica adversa amenaza Ia
supervivencia del micelio (Delmas, 1978b; Kaul, 1987). Los estfmulos ambientales de
Ia fructificaci6n varfan desde una cierta composicion de Ia atmosfera interna del
sustrato (C0 2,, 0 2 , etileno, vapor de agua, etc.) hasta cambios en Ia temperatura
optima del crecimiento micelial, Ia cual es de 25 a 30 oc, comunmente una reduccion
de 5 a 10 oc induce Ia fructificaci6n (Delmas, 1978b); condiciones ambientales
desfavorables provocan un desarrollo deficiente en los carp6foros (Hawker, 1957).
Esporulaci6n. Se inicia con Ia maduraci6n fisiol6gica del carpoforo, en este momenta
finaliza el almacenamiento de Ia informaci6n genetica del hongo en las esporas; estas
12

se forman dentro de los cuerpos fructfferos, especialmente en una capa llamada
himenio, para luego ser liberadas al madurar el carp6foro.
Las esporas son como "semillas" de los hongos, guardando el estado de dormancia
hasta que se presentan condiciones ambientales 6ptimas para activarse (Kent y
Mcknight, 1987; Kaul, 1987), las caracterfsticas de las esporas son propias del
genero, por lo que son importantes para Ia identificaci6n de los hongos (Kent y
Mcknight, 1987).
Las esporas de Ia mayorfa de macromicetos estan adaptadas basicamente para Ia
diseminaci6n por el viento y en algunos casos son transportadas por animales,
especialmente insectos y algunos mamfferos, el transporte lo realizan pasando por su
tracto digestive sin sufrir dafio (lbar, 1980; Singer, 1961).
Formas de Crecimiento. La reproducci6n sexual en los macromicetos ocurre por Ia formaci6n
de cuerpos fructfferos, mismos que pueden ser: (a)- hipogeos, aparecen bajo Ia tierra; y (b)
epigeos, aparecen sobre Ia tierra, en una area boscosa se localizan mas especies epigeas que
hip6geas (Fogel, 1992; Kaul, 1987).
Hongos Hipogeos. Su estudio no esta generalizado ya que son dificiles de localizar
(lbar, 1980), para estudiarlos es necesario excabar el area a una profundidad de
aproximadamente 10 em (Hunt y Trappe, 1985), tam bien se requieren periodos largos de
muestreo, porque no fructifican anualmente, llegando a transcurrir de 5 a 10 a nos para que se
rep ita un patr6n de fructificaci6n en algunas especies (Hunt y Trappe, 1985). La producci6n a
lo largo del aiio es constante sin tener cimas, Ia dispersi6n de las esporas depende de
animales mic6fagos por ello tienen una corta distancia de dispersi6n que los epigeos (Allen et
a!. 1995).
Hongos Epigeos. Su existencia es comun, su cuerpo fructifero se desarrolla sobre el
suelo, haciendolos visibles por sus variadas y coloridas formas; Ia producci6n tiene mas alti
bajos que en el caso de los hipogeos. Hay hongos epigeos que son similares en sus nichos,
desarrollo, funciones, pero no son identicos, asi cuando un hongo es removido, otra especie
con funciones similares coloniza ese nicho (Allen eta!. 1995); el endemismo a nivel de genero
es relativamente pequefio para estos hongos ya que al dispersar las esporas por el aire lo
hacen potencialmente sobre grandes distancias; asi existen especies en ambos hemisferios
13

continentales. En este grupo se encuentran casi Ia totalidad de los hongos silvestres
comestibles.
La sociabilidad es un caracter adicional para cada especie de los hongos epfgeos por lo
que pueden ser: solitaries, gregarios, formando arcos o creciendo en grupos o en forma
connada (Arnolds, 1988; Vogt et a!. 1992). La mayorfa de los hongos macrosc6picos
perduran de unos pocos dfas a varias semanas sin deterioro (boletos, rusulas, colibias),
algunos duran horas (pequenos coprinos) y otros tienen longevidad de hasta 4 meses
(polfporos) (Arnolds, 1992).
Tipos de Sustrato. Los hongos al ser organismos heterotr6ficos, son incapaces de sintetizar
sus propios alimentos; por lo que el micelio cumple una funci6n degradadora, esta estructura
se encuentra enterrada, penetra en Ia madera o en otros sustratos a los que degrada (Stone,
1996). De acuerdo con Ia presencia del micelio sobre el sustrato se les clasifica en:
Lignfcolas. Son aquellos hongos que degradan Ia madera en cualquiera de sus formas:
troncos, tocones de arboles y ramas. La descomposici6n de Ia madera es comun en los
suelos forestales, siendo este habitat explotado por los hongos (Bruns, 1995).
Copr6filas. Se localizan sobre excrementos de Ia macrofauna presente en el bosque,
ya sea de ganado o de fauna silvestre; Guzman-Davalos y Guzman ( 1979), indican que Ia
existencia de estas especies en zonas perturbadas esta condicionada a Ia presencia de ganado
en el bosque.
Fenfcolas. Un incendio en los suelos forestales modifica el pH a 8 o 9, lo que favorece
Ia colonizaci6n de especies llamadas fenfcolas; Ia recolonizaci6n es efectuada por estos
hongos ya que toleran condiciones de alcalinidad. Estas especies producen esporas que son
dispersadas por el viento, las cuales se encuentran en estado de dormancia hasta que un
detonador de calor las activa, generalmente temperaturas superiores a 50~ C (Hudson, 1980).
Terrfcolas. Se encuentran en los suelos forestales, aparentemente las especies
presentes en este tipo de sustrato son por lo general hongos micorrfzicos (Hudson, 1980).
Humfcolas. Se ubican sobre el mantillo forestal o cualquier tipo de materia organica en
descomposici6n (Hudson, 1980; Vogt eta/. 1992), tales como restos de plantas y animales
muertos; Ia mayorfa toleran distintos grados de acidez (Arnolds, 1988).
14

Grupos ecol6gicos. Los macromicetos por Ia forma de obtener los nutrimentos se dividen en
grupos ecol6gicos: saprobios, simbiontes mutualistas y antagonistas (Molina et a!. 1993),
aunque existen especies con capacidad de explotar varios habitats, asf especies de diferentes
grupos ecol6gicos !Iegan a fructificar en un mismo habitat (Arnolds, 1988); ademas, existe
especializaci6n en Ia forma de obtener los nutrimentos del sustrato dentro de un mismo grupo
ecol6gico, atribuido a mecanismos desarrollados para evitar Ia competencia entre especies
cercanas, evitando el uso de los mismos recursos en un nicho (Tanesaka et a/. 1993). asf
existe diversidad a nivel de microareas, con varios grupos ecol6gicos poblar ·o rafces
adyacentes o rafces secundarias, separadas unos pocos centimetres (Bruns, 1995).
Saprobios. Los hongos saprobios en los bosques son agentes degradadores del
mantilla forestal o cualquier otro tipo de materia organica muerta (Wasterlund y lngelot,
1981).
La funci6n ecol6gica de este grupo consiste en liberar los nutrimentos inmovilizados en
estructuras organicas (Dighton y Boddy, 1989); en este proceso de degradaci6n existe retorno
de carbo no en forma de di6xido a Ia atmosfera (Hudson, 1980). Sin este grupo de hongos el
ciclo de Ia materia organica no serfa tan eficiente, acumulandose los restos organicos en el
bosque (Arnolds, 1988).
La mayorfa de especies degradan celulosa, hemicelulosa y lignina, siendo esta ultima
utilizada casi exclusivamente por los basidiomycetes (Arnolds, 1992); del sustrato extraen
azucares simples, que se utilizan como fuente de carbone, de ellos Ia glucosa es virtualmente
utilizada por todos, siguiendo en importancia el almid6n (Hudson, 1'980). Estos hongos
tambien extraen minerales (P, F, y trazas de Manganese y Zinc) y vitaminas, las vitaminas
requeridas principalmente en mayor cantidad sonIa Tiamina y Ia Biotina (op. cit.).
Stone (1996), indica que este grupo es el mas numeroso; Dighton y Boddy (1989),
reportan que aproximadamente dos terceras partes de Ia biomasa microbial del mantilla
forestal y suelo de un bosque templado es de hongos saprobios. Este grupo inmoviliza y luego
recicla cantidades significantes de nutrimentos, lo cual reduce Ia lixiviaci6n del suelo (Molina
eta!. 1993).
Simbiontes Mutualistas. Los hongos y los vegetales forman una asociaci6n simbi6tica
mutualista conocida como micorriza, donde ambos organismos reciben beneficios.
15

Son reconocidos siete tipos de micorrizas: Ectomicorrizas (EM), arbuscular (AM),
ericoide, arbutiode, monotroide, orqufdeas y E-cepa (Allen eta/. 1995); de las cuales las de
mayor importancia ecol6gica debido a su predominancia son: Ia AM y EM (Allen eta!. 1995;
Azc6n y Barea, 1980; Le-Tacon, 1985).
La mayorfa de las micorrizas se localizan en las capas superficiales del suelo, en los
primeros 10 centimetres (Bruns, 1995; Read et a/. 1989); las micorrizas no siguen patrones
de Ia riqueza de las plantas, no se relacionan con Ia diversidac Je especies, ni correlacion con
gradientes latitudinales (Allen eta/. 1995).
En opinion de Azcon y Bare a ( 1980), las caraterfsticas de los principales grupos de
micorrfzas son:
Endomicorrizas. Son hongos asociadas a Ia mayorfa de las plantas de interes agricola,
se originaron simultaneamente con las plantas (Bruns, 1995; Pirozynski, 1980), hace
400 millones de a nos (Azc6n y Bare a, 1980). Este tipo de micorrfzas se caracterizan
porque el micelio del hongo no forma manto sabre Ia rafz y sus hifas penetran en el
interior de Ia celula de las rafces (Azc6n y Barea, 1980; Le-Tacon, 1985). Los tipos
contenidos en este grupo son: arbuscular (AM), ericoide, arbutiode, monotroide,
orqufdeas y E-cepa (Allen eta/. 1995).
Ectomicorrizas. Dentro de este grupo se incluyen los macromicetos, asociadas a
arboles forestales (Hacskaylo, 1983); Singer (1961), sefiala que casi Ia mitad de los
macromicetos de Europa y Norteamerica viven con este tipo de simbiosis. La mayorfa
de las plantas EM son simbiontes obligados (Allen eta!. 1995).
Surgieron en el periodo Jurasico y proliferaron del Cretacico al Terciario (Allen et a/.
1995), se caracterizan porque los filamentos subterraneos de los hongos no penetran
en las celulas radicales y por Ia presencia de un manto formado por el micelio en Ia
zona radicular (Azc6n y Barea, 1980; Le-Tacon, 1985).
El micelio normalmente es septado, el desarrollo del hongo en el interior de Ia corteza
es intercelular dando un aspecto de red, conocida como red de Hartig (Azc6n y Barea,
1980); se reproducen por esporas sexuales, las cuales son numerosas, pequefias y
mejor dispersadas por el viento en relaci6n con las AM, que son asexuales (Connell y
Lowman, 1989). El pH 6ptimo es de 3.3 a 5.5 para Ia mayorfa de estas especies
(Arnolds, 1988).
16

Se estiman de 5000 a 6000 especies EM a nivel mundial, agrupadas en 148 generos
(Bruns, 1995 Pirozynski, 1981 ). Su distribuci6n, generalmente esta delimitada por Ia
existencia de los bosques templados a boreales, proliferando en los bosques de
confferas, donde !Iegan a existir cientos de EM (Allen et a! 1995, Read, 1991), su
dominancia en este tipo de bosque se explica en que al abrirse un sitio en disturbio son
las que primero colonizan debido a su alto volumen de micelio y esporas que son
dispersadas por el viento con mayor eficiencia respecto a las AM, estas caracterfsticas
ffsicas les permiten c·...,,onizar nuevos habitats en relaci6n a las esporas de otros tipos,
que son incapaces de establecerse, debido a que el primer colonizador con su
crecimiento suprime a los demas (Connell y Lowman, 1989)
Las EM estan mal representadas en los bosques tropicales en opinion de Guzman 1984),
son comunes en plantas de las familias. Dipterocarpaceae, Fagaceae, Myrtaceae y
Leguminosae (Caesalpinioideae) (Connell y Lowman, 1989)
Los intervalos ambientales de las EM son estrechos para Ia mayorfa de las especies,
ademas exhiben amplia especificidad, asf se encuentran aquellas que son exclusivas de
un genera o familia a otras que son promiscuas (Allen eta/ 1995)
En los suelos forestales los nutrimentos disponibles para los vegetales se encuentran
en bajas concentraciones (Azc6n y Barea, 1980; Le-Tacon, 1985), asf el fosforo
soluble se encuentra en una concentraci6n de 1 a 5%, reduciendose rapidamente
debido a Ia extracci6n radicular, esto conduce a Ia existencia de una zona de
agotamiento, minimizandose este problema por Ia relaci6n simbi6tica existente entre el
arbol y el hongo; en esta simbiosis existe una ayuda mutua que contribuye al
desarrollo de ambos organismos, cada uno de ellos recibe un bene.ficio de Ia asociaci6n
(Christensen, 1989; Le-tacon, 1985, Vogt eta!. 1992)
Debido a que los mantos micorrfzicos extendidos y gruesos actuan como tejido
conductivo del arbol, extendiendose hasta 10 metros log ran mejoran Ia absorci6n de
agua, nutrimentos como: fosfatos, amonio y alanina, formas nitrogenadas: N
amoniacal (NH4 ), N nitrico (N03), aminoacidos), por consecuencia mejoran su
desarrollo y aumentan Ia resistencia a plagas y enfermedades (Allen et a/ 1995,
Christensen, 1989; Le-Tacon, 1985, Mason et a/. 1983, Read, 1991; Stone, 1996;
Vogt eta! 1992).
17

Los mantas micorrizicos pueden coexistir entre elias formando redes o puentes
miceliares entre arboles vecinos, circulando sustancias y elementos de uno a otro;
mantienen al hongo anclado al suelo y son capaces de acelerar Ia degradaci6n de
compuestos organicos Jiberando nitr6geno, f6sforo y minerales insolubles, poniendolos
a disposici6n de los arboles, explicable por tener enzimas degradadoras extracelulares
como polifenol oxidasas, fosfatosas, celulasas y proteinasas (Allen et a/. 1995;
Christensen, 1989; Dighton y Boddy, 1989; Le-Tacon, 1985; Read, 1991; Read eta/.
1989; Stone, 1996).
El arbol proporciona los glucidos que no puede sintetizar el hongo, como azucares
simples: sacarosa, fructuosa y glucosa (Le-Tacon, 1985; Vogt et a/. 1992); de las
sustancias fotosintetizadas se desvfan a las micorrizas entre el 10 y 40% (Hacskaylo,
1983; Le-Tacon, 1985). ademas el factor M (principia activo no identificado,
producido por las rafces) estimula Ia germinaci6n de esporas y el crecimiento del
micelio en hongos EM (Hacskaylo, 1983; Kaul, 1987).
Existen sustancias producidas en cantidades bajas por el metabolismo del hongo
llamadas auxinas y citocininas, basicas para el establecimiento y mantenimiento de Ia
simbiosis (Hacskaylo, 1983). a quienes se les atribuye: (a)- reducci6n de Ia actividad
meristematica; (b)- formaci6n de Ia red de Hartig; (c)- retraso de Ia maduraci6n de las
celulas de Ia corteza; (d)- elasticidad de Ia pared celular del hongo, lo cual resulta en
una permeabilidad de Ia membrana para regular los rangos de traslocaci6n de iones y
componentes; (d)- incremento de las rafces laterales; y (e)- retraso de Ia senecencia del
tejido radicular.
Tambien transitan en las micorrizas otros compuestos mas complejos, se trata de
vitaminas y sustancias de crecimiento, algunas de las cuales estimulan el desarrollo del
hongo, siendo estas ultimas indispensables para Ia formaci6n de los 6rganos de
reproducci6n (Le-Tacon, 1985); el sitio de trasferencia de los productos absorbidos
hacia el arbol es Ia red de Hartig.
Existen estudios que revelan en forma numerica Ia importancia de esta simbiosis en los
sistemas forestales: (a) en un bosque de abeto (Pseudotsuga merziessl) el retorno de
materia organica al suelo par rafces finas y micorrizadas varia de 78 a 84% del retorno
total al arbol y de casi 80% del retorno de N y P (Dighton y Boddy, 1989; Read,
1991 ); (b) las hifas de macromicetos tienen cantidades mas altas de elementos que el
18

mantilla: 2.9 a 9.4 en P, 1.8 a 8.2 en N y 2-3 en F (Le-T aeon, 1 985); estos
nutrimentos son liberados en forma soluble a su muerte y en vida por excreci6n
(Christensen, 1989; Dighton y Boddy, 1989); y (c) del 10 al 20% de los fotosintatos
son reciclados directamente por los hongos micorrfzicos (Christen~en, 1989).
El establecimiento de micorrizas depende de Ia disponibilidad de azucares exudados
alredor de las rafces, Ia colonizaci6n es directamente a los carbohidratos del interior de
las rafces, de esta 1 anera Ia concentraci6n interna del arbol tiene una influencia
directa sobre el establecimiento y mantenimiento (Hacskaylo, 1983; Vogt eta/. 1992).
La persistencia de Ia asociaci6n no depende unicamente del huesped y el hongo, sino
tambien de los organismos del suelo, donde cada componente esta sujeto a Ia
influencia del ambiente (Mason et a/. 1983). Se presume que el hongo produce
hormonas que auxilian a mantener Ia relaci6n simbi6tica (Hacskaylo, 1983).
La fructificaci6n de los hongos micorrfzicos es menos regular anualmente que Ia de los
hongos saprobios (Villeneuve, 1989). una producci6n alta no pueqe ser esperada hasta
que las rafces de los arboles hayan alcanzado un suficiente nivel de nutrici6n, que se
puede lograr en 10 af\os (Wasterlund y lngelot, 1981). La visualizaci6n de un
carp6foro es relacionada con Ia existencia de un manto micorrfzico (Deacon et a/.
1983), un decremento de ellos indica una disminuci6n de rafces micorrizadas (Arnolds,
1 988); Ia perdida de una especie asociada con un tipo de arbol dominante en el
sistema es fatal (Allen eta/. 1995).
Las estructuras reproductivas sexuales de los carp6foros, poseen un alto porcentaje de
nitr6geno que al ser dispersadas por el viento o macrofauna se estableceran en el sitio,
entonces mucho nitr6geno sera removido del Iugar (Dighton y Boddy, 1989).
Simbiontes Antagonistas. Con respecto a los dos grupos referiqos, son escasas las
especies de macromicetos que obtienen los nutrimentos, atacando y matando los tejidos vivos
donde se desarrollan, pudiendo causar ia muerte de todo el organismo. Dos parasitos
conocidos son: Heterobasidion annosum y Armillaria ostoyae (Molina eta/. 1993).
Del tema expuesto se concluye que al ser pocas las especies de macromicetos del
grupo de los simbiontes antagonistas, existen escazos estudios y publicaciones que tratan
sobre este tema; del grupo de los saprobios se tiene un poco mas de conocimiento; el grupo
de los simbiontes mutualistas el mas estudiado.
19

Distribuci6n Geografica de los Hongos
La mayorfa de los hongos macrosc6picos se ubican en los bosques templados, de alta
montana y boreales, cuoy ecosistemas son propicios para su desarrollo y fructificaci6n (lbar,
1980; Singer, 1961; Valenzuela eta/. 1981); las condiciones 6ptimas varfan para las diferentes
especies, pero se mantienen en intervalos ambientales de estos.
Los hongos son organismos individuales, con sus re laciones proplas con el ambiente,
por lo que t ienen requerimientos especfficos que trascienden Ia necesid:_,d por un huesped
(Allen et a!. 1995); Ia disponibilidad de materia organica es el factor determinante de su
presencia (Valenzuela eta!. 1981; Villeneuve eta!. 1989).
Existen macromicetos en los bosques tropicales pero en una menor proporci6n
(Valenzuela et a!. 1981), probablemente debido a Ia exhuberante vegetaci6n que lim ita nichos
apropiados para el establecimiento de los hongos; los hongos comestibles de los bosques
tropicales son pequenos y poco carnosos, en comparaci6n con los otros bosques (Guzman
Davalos y Guzman, 1979). Comparando Ia funga de los bosques de confferas templados a
boreales, con los bosques tropicales, estos ultimos poseen muchas especies pero poco
frecuentes, mientras que los primeros tienen pocas pero con alta frecuencia (Villeneuve et a!.
1989). Garza et a!. (1985), reportan para un bosque de Quercus rysophylla los hongos en
orden de importancia: micorrfzicos, lignfcolas y saprobios.
Existen en Ia naturaleza especies que poseen requerimientos ambientales excluyentes,
por ejemplo: Hygrophorus marzuolus crece sobre Ia nieve, los generos Morchella y Gyromitra,
se desarrollan cuando Ia nieve se funde (Stone, 1996) y el genero Tulostoma, que tolera
condiciones de aridez (!bar, 1980).
Los bosques poseen mecanismos naturales que les permiten conservar Ia diversidad de
macromicetos, tales como: (a) - interacciones inter e intra especfficas; y (b)- disturbios
naturales como incendios, sequfas, heladas, entre otros. Los disturbios propician parches
en bosques para Ia colonizaci6n de los hongos y como el sistema nunca alcanza el equilibrio,
se evita Ia exclusion; tambien existen disturbios menores tales como: excavaci6n,
excrementos, esqueletos o Ia formaci6n de nuevas rafces, asf Bruns ( 1995) encontr6 que siete
diferentes tipos de bosque mostraron un volumen anual creciente de 42 a 49%. Las
interacciones se desarrollan por bacterias del suelo y mic6fagos invertebrados con los hongos
macromicetos, previniendo Ia exc(usi6n e incrementando Ia diversidad (op. cit). Se concluye
que en un bosque natural el potencial para mantener Ia diversidad fungica nunca es nulo, pero
20

Ia intervencion humana puede contribuir a Ia creacion de zonas perturbadas, donde no se
conserven las especies .
lmportancia en el Bosque
Los macrohongos tambien son importantes en los bosques, al ser fuente de alimento
para animales micofagos y como estabilizadores del ecosistema.
Alimento de Animales Micofagos. Los carpoforos de los hongos silvestr s poseen un alto valor
alimenticio, por contener aminoacidos, protefnas y azucares; son parte en Ia nutrici6n de
pequef\os mamfferos que basan su alimentacion en estas estructuras reproductivas,
deniminandoles por este hecho animales micofagos, entre los que tenemos conejos, tuzas y
ardillas (Allen eta/. 1995; Christensen, 1989).
Estabilizadores. Los macromicetos micorrfzicos auxilian a determinar Ia riqueza de las plantas;
en los bosques las extensivas redes de hongos EM pueden controlar Ia riqueza de especies
arboreas o arbustivas ya que al colonizar un sitio no dejan espacios libres y solo podra
colonizar una planta que pueda asociarse a las EM (Allen eta/. 1995).
Distribuci6n Espacio-Temporal
La fructificacion de los hongos macromiceto~ varia en el tiempo y el espacio (Dighton
eta/. 1986; Fogel, 1 992; Vogt eta/. 1992), Hunt y Trappe (1985) reportan que el registro
de especies nuevas en un estudio, nose estabiliza si se incrementa el periodo de observacion
y el area de muestreo.
Tiempo. Existe sucesi6n fungica con el desarrollo del sitio, presentandose especies
caracterfsticas propias de bosques j6venes o etapa temprana, especies traslapadas y
especies propias de bosques que se encuentran en Ia etapa final de Ia sucesion o etapa tardfa
(Arnolds, 1988; Bujakiewicz, 1 982; Vogt et a/. 1992). Pocas especies fructifican af\o con
af\o, Ia mayoria tienen un patron de de 5 anos, se conocen especies de 1 0 af\os (Arnolds,
1991 ); Ia fructificacion de especies en un afio donde presentan su informacion genetica
completa forma y tamafio; dirige a caracterizar a los afios en "buenos". y "malos" (Arnolds,
1992; Wasterlund y lngelot, 1981).
Otra causa de Ia estacionalidad en el tiempo, son las condiciones ambientales propias
de cada estacion, existiendo variaciones en temperatura, humedad y actividad de otros
21

microorganismos, resultando en liberaci6n de nutrimentos, con variaciones en cal idad y
cantidad (Bruns, 1995; Read, 1991 ).
Espacio. La etapa temprana de una sucesi6n ecol6gica es comun en arboles j6venes, se
identifica en rafces salientes que son colonizadas, pero es desplazada en pocos anos por Ia
etapa tardfa, Ia cual hace su aparici6n sobre las rafces pr6ximas al tronco hasta lograr Ia
expulsion; Ia estacionalidad en Ia colonizaci6n se atribuye a que Ia etapa tardfa parece requerir
ma~ carbohidratos y nitr6geno del huesped, colonizando Ia rafz hasta que puede curilplir los
requerimientos nutricionales (Arnolds, 1988; Bruns, 1995).
Factores que Afectan Ia Productividad de los Hongos
Diversos autores concluyen que Ia productividad de los hongos silvestres esta
relacionada a factores: end6genos, ex6genos y silvfcolas y sus interrelaciones, los cuales
influyen antes y durante el perfodo de crecimiento (Arnolds, 1992; Chac?n y Guzman, 1995;
Charles, 1974; Delmas, 1987a; Hunt y Trappe, 1985; Vogt eta!. 1992), asf si el ano anterior
fue seco puede retrasar Ia fructificaci6n; si existi6 alta produccion, entonces se agotan las
reservas y disminuye Ia producci6n actual; los carpoforos solo se desarrollan cuando las
condiciones ambientales son 6ptimas, se reconoce que para cada especie estas condiciones
son diferentes (Bills et a!. 1986; Hudson, 1980; Mason et a!. 1983; Stone, 1996), aunque
todas dentro de un estrecho intervale (bosques tempi ados a boreales). No hay diferencias
fundamentales en Ia fructificacion in situ e in vitro (Delmas, 1978b).
De las especies que fructifican, Ia producci6n es dominada por algunas de elias (Bills et
a!. 1986; Kalames y Silver, 1988); Hunt y Trappe ( 1985), reportan que 7 y 4 especies (a no 1
y 2) totalizaron el 77.6% y 91.3% de peso seco total.
End6genos. La producci6n es controlada por factores end6genos, es decir, por Ia informacion
genetica (Delmas, 1978b; Kaul, 1987); el potencial genetico de cada hongo es el que dicta
bajo que estfmulos fructificar, forma del carp6foro y tamafio maximo.
Los carp6foros s61o se desarrollan si el micelio tiene reservas adecuadas (Hudson,
1980; Kaul, 1987; Wessels, 1991 ); Ia nutrici6n es probablemente el segundo factor en
importancia para Ia reproducci6n (Kaul, 1987), los hongos en general acumulan C, N, K, F,
sulfuros, P, Mg y en menor proporci6n, Hierro, Zinc, Cobre, Manganese y carbohidratos,
algunas especies requieren vitaminas (biotinas, tiaminas y piridoxina) y compuestos conocidos
22

como sustancias de crecimiento, para formar los 6rganos de reproducci6n (Kaul, 1987; Le-
Tacon, 1985; Vogt eta!. 1992).
Ex6genos. El hongo para iniciar Ia fase de fructificaci6n requiere que los factores ambientales
sean 6ptimos para su metabolismo. Los principales factores son precipitaci6n y temperatura.
El metabolismo del hongo requiere agua para absorber los elementos nutritivos en
soluci6n, durante Ia fase ve !:1etativa (Kaul, 1987), estos conocimientos generales sobre los
macromicetos, aunados al hecho de que aparece una maxima producci6n de carp6foros en
tiempo de lluvia, nos revela que el agua y Ia temperatura son los principales factores
ambientales de Ia producci6n (Hudson, 1980; Hunt y Trappe, 1985; Wasterlund y lngelbt,
1981 ).
Existen estudios que reconocen en terminos numericos estos dos factores como
principales: (a)- Hudson ( 1980), encntr6 que el inicio de Ia producci6n de hongos en un
bosque de hayas, es a 50% de humedad relativa y 4°C de temperatura; (b)- Chacon y Guzman
(1995), afirman que Ia precipitaci6n y Ia temperatura son los factores mas importantes,
surgiendo carp6foros al exceder Ia precipitaci6n los 40 mm en un bosque subtropical; (c)- Kaul
(1987), reporta que un 70% de humedad relativa es el limite mas inferior para Ia
fructificaci6n; (d) Zamora-Martinez y Pascual-Pol a ( 1'995), senalan que emergen carp6foros en
un bosque de Pino montezumae cuando Ia precipitaci6n excede los 100 mm; (e)- Norvell
(1995), refiere que Ia temperatura y Ia precipitaci6n durante Ia esta.ci6n de crecimiento
explican de 19 a 42% del comportamiento de Ia biomasa y de 24 a 48% el numero de
carp6foros; (f)- Hunt y Trappe (1985), encontraron una correlaci6n lineal entre biomasa de
carp6foros hipogeos, temperatura y precipitaci6n, con un valor de r2 de 0.685 y (g)- Vogt et
a!. ( 1992), encontraron que Ia producci6n ocurre de 2 a 3 semanas despues de una abundante
lluvia, que provoc6 una alta humedad en el suelo.
De los estudios anteriores, se deduce que se requiere de un minimo de humedad para
que inicie Ia fructificaci6n, aunque un exceso de agua tambien es desfavorable para Ia mayoria
de los hongos (le-Tacon, 1985). En un ano seco s61o las especies tolerantes a Ia sequfa
forman carp6foros {Polyporus spp.) mientras que en un ano humedo fructifican principalmente
las especies higr6filas es decir las carnosas (Kaul, 1987; Wasterlund y lngelot, 1981).
Wasterlund y lngelot ( 1981), reportan que los hongos micorrfzicos son mas
dependientes de Ia humedad que los saprobios, asf observaron en un af\o humedo hongos
23

micorrizicos quienes comprendieron aproximadamente el 50% de Ia producci6n, disminuyendo
a 5% y 10% en un afio seco. Con respecto a Ia temperatura Kaul ( 1987), reporta que Ia
mayorfa de los carp6foros se desarrollan de 10 a 40° C, con un 6ptimo de 25 a 35°C.
El crecimiento vegetative se desarrolla en forma optima con una concentraci6n baja de
oxfgeno, requiriendo un nivel mas alto en Ia fase de fructificaci6n; con respecto al C0 2 Ia fase
reproductiva requiere un nivel mas bajo que el crecimiento micelial. Los factores anteriores
s61o son problema en condiciones de cultivo intf'lSO (Kaul, 1987). El factor luz es necesario
en Ia reproducci6n, ya que induce Ia formaci6n de estructuras sexuales, orientandolas en Ia
dispersion de esporas (op. cit.).
Las condiciones edaficas que estimulan Ia formaci6n de carp6foros y su funci6n de
descomposici6n son: Ia textura areno-limosa, baja densidad, alto contenido de materia
organica y pH acido (Vogt eta!. 1992; Zamora-Martinez y Pascuai-Pola, 1995).
Caracterfsticas tales como el olor, color y forma de una misma especie son
modificadas porIa influencia de Ia latitud y altitud (lbar, 1980; Stone, 1996). En general, Ia
riqueza de especies de los bosques templados decrece hacia el norte, con el incremento de Ia
latitud, esto por pasar de climas templados a climas articos, provocando el decremento de
sustratos organicos y de plantas vasculares (Villeneuve eta/. 1989).
AI ser 6ptimos los factores ex6genos las especies con carp6foros chicos fructifican
primero, si continuan las condiciones fructificaran las especies con carp6foros grandes, ya que
las primeras requieren menos nutrientes (Deacon eta!. 1983; Hudson, 1980).
Silvfcolas. Existen factores dentro del bosque que afectan Ia producci6n, tales como Ia
interacci6n con macrohongos, edad del arbolado, forrajeo por microfauna, interacci6n con
microorganismos y recolecci6n intensiva.
lnteraccion con Macrohongos. Cambios en aparici6n de especies o volumen, debido a
Ia presencia de macrohongos, se concibe como interacci6n, Wasterlund y lngelot (1981)
reportan un ejemplo de interacci6n, comparando Ia abundancia de Hypho!oma spp. y
Armillariella me/lea, observaron que donde una es abundante, Ia otra se encuentra en baja
densidad, esto podria ser un ejemplo de competencia por nutrimentos.
24

Edad del Arbolado. La edad del arbolado determinu Ia calidad y cantidad de
nutrimentos, Ia vegetaci6n y las especies fungicas presentes (Dighton et a!. 1986; Hudson,
1980; Hunt y Trappe, 1985; Schoonmaker y Mckee, 1988; Vogt eta!. 1992). Por lo expuesto
Frankland ( 1992) basndose en el estrato arboreo clasifica a los bosques en: j6venes,
maduros y viejos.
Joven. Se caracteriza por una baja diversidad de hongos, los cuales no son especfficos
de un huesped, pudiendo asociarse con casi todas las especies arboreas {Fran',,and,
1992). Kalames y Silver { 1988), opinan que estos bosques poseen baja producci6n de
hongos y bajo porcentaje de deterioro de carp6foros.
Maduro. Se distingue por un dose! casi cerrado y por Ia existencia de Ia maxima
diversidad de macrohongos {Frankland, 1992). El dose! incrementa Ia humedad
indirectamente, reduce Ia radiaci6n y el movimiento del afre, evita el calor extremoso
del verano y el frio del invierno, manteniendo las condiciones favorables por periodos
mas largos, lo que resulta en una producci6n mas prolongada. Lo anterior explica
existen altos valores de producci6n en estos bosques{Dighton et a/. 1986; Hudson,
1980; Hunt y Trappe, 1985; Vogt eta!. 1992; Wasterlund y lngelot, 1981).
Para Estonia Kalames y Silver {1988), que Ia producci6n de los bosques maduros
aumenta dos o tres veces mas que en los pinos j6venes, senalan que el deterioro de
los carp6foros es casi 1 /3 a 1/2 de Ia producci6n, estas perdidas existen a pesar de
tener un intervalo de muestreo de 10 dfas. La producci6n de hongos no comestibles,
en los bosques maduros es de 1/2 a 1 /4 de los hongos comestibles, pero su numero
es dos veces mas grande que los hongos comestibles {op. cit.).
Viejo. Frankland {1992), caracteriza a estos bosques por Ia especificidad de los
macromicetos hacia cierto huesped.
Los bosques viejos son los mejores sitios de producci6n {Wasterlund y lngelot, 1981),
tal vez porque el crecimiento de pastos se reduce; otro hecho que puede influir es que
Ia biomasa total del bosque se incrementa con Ia sucesi6n {Dighton y Boddy, 1989,
Frankland, 1992). En estos bosques decrece Ia diversidad de especies de arboles
{Schoonmaker y Mckee, 1988).
25

El numero de especies y de carp6foros se incrementa hasta llegar a un nivel maximo
hasta que el dose! se cierra, luego existe un decremento de Ia diversidad (Vogt et a!.
1992). Una explicaci6n es por Ia sucesi6n, Ia cual determina Ia cobertura del pasta;
en una fase terminal de Ia sucesi6n climatica casi no existe pasto, se ha encontrado
una correlaci6n negativa entre pastos y Ia abundancia de carp6foros de macrohongos
(Arnolds, 1988).
En el inicio de una sucesi6n artificial, iniciada por una tala total,' se ve Ia dominancia
de hierbas, despues de arbustos y a los 20-30 anos dominan las confferas; a los 40
anos de Ia plantaci6n el estrato corresponde a confferas en 49%, arbustos con 33% y
hierbas en 18% (Dighton y Boddy, 1989).
Forrajeo por Microfauna. Existe microfauna mic6faga que se alimenta del micelio de
macromicetos, tales como: hormigas, escarabajos, colembolos, is6podos, lombrices y
dfpteros (Hudson, 1980; Kendrick, 1992), esto aumenta Ia tasa de descomposici6n de los
carp6foros, disminuyendo Ia producci6n comercial. En Africa Kendrick (1992) reporta un caso
contrario, una simbiosis entre Boletus y un insecta que resulta en macromicetos de alto valor
comercial como Termitomyees spp. en Africa.
lnteracci6n con Microorganismos. los macromicetos reciben efectos estimulantes de
otros microorganismos, tales como microhongos, bacterias y actinomicetos, lo que
probablemente resulta en un aumento de Ia producci6n (Kaul ( 1987).
Recolecci6n lntensiva. La recolecci6n intensiva constituye una fuente de Ia
disminuci6n de carp6foros, sin embargo esto parece no alterar Ia producci6n (Arnolds, 1988;
Wasterlund y lngelot, 1981). Norvell ( 1995) report a que despues de 1 0 a nos de realizar una
recolecci6n intensiva de Cantharellus cibarius, no se afect6 Ia productividad; por ella una
prohibici6n general de recolectarlos serfa inadecuada e indeseable para proteger a los
macromicetos (Arnolds, 1988).
Bases para el Uso Sustentable
El deterioro de Ia productividad ecol6gica y Ia persistentia de Ia pobreza en
todo el mundo cada dia es mas preocupante por ella es factible cuestionar el modelo de
desarrollo actual; desde Ia perspectiva ecol6gica el capital natural de los bosques y su
productividad son elementos fundamentales a considerar en el desarrollo para garantizar el
26

bienestar de hoy y asegurar el de manana, caso contrario nuestro almacen de recursos
naturales continuara en franca desminuci6n.
Lo expuesto nos conduce a proteger Ia producci6n ecol6gica asegurando para el largo
plazo Ia supervivencia de Ia biodiversidad, incluyendo al hombre, el aprovechamiento
sustentable de los macromicetos se encuadra en esta arista.
Para enfilar hacia un uso sustentable de los macrohonr lS silvestres, es necesario
como primer paso realizar un inventario y monitoreo ecol6gico de Ia zona a mane jar.
lnventario y Monitoreo Ecol6gico. Dentro de una Mycetation, termino propuesto por Arnolds
(1992), para referir todos los tipos de hongos presentes en un habitat, se distinguen los
macrohongos y lo microhongos; esta clasificaci6n es subjetiva, creada por el investigador para
satisfacer los objetivos de su estudio.
AI estudiar una mycetation a nivel alpha (nivel del habitat) de macromicetos, los
estudios se basan en Ia aparici6n de carpoforos, lo cual tiene dificultades: no todas las
especies son de fructificacion epigea (Bruns, 1995), no revelan una idea completa de los
eventos que ocurren bajo Ia tierra (Mason et a/. 1983); mas cuando no son una estimaci6n
exacta del volumen y de Ia actividad del micelio, P?Seen un grado de correlacion aceptable
(Arnolds, 1988; 1992; Mason eta/. 1983).
El monitoreo de hongos silvestres comestibles puede ser definido, como el registro
repetido de los datos que interesan al objetivo del muestreo (Arnolds, 1992); el prop6sito del
monitoreo determina que informacion deber ser colectada (Molina eta/. 1993): Por los datos
que se recolectan se subdivide el monitoreo en tres tipos: (1 )- deteccion, (2)- evaluacion y (3)
investigaci6n (op. cit.).
Dos condiciones deben ser respetadas, sin importar de que simple o complejo es un
monitoreo: (a)- las observaciones deben ser repetidas cfclicamente; y (b)- deben compararse
los resultados con una referencia, estableciendose asf Ia diferencia. Si Ia informacion obtenida
es abierta, es decir resultado de Ia cooperaci6n de Ia gente que se involucra de alguna
manera con los bosques, propietarios, recolectores, visit<mtes e investigr:~riorP.s. sP. or:~rr:~nti7r:~
que Ia informacion obtenida sera verfdica, una vez obtenida Ia informaci6n debe integrarse en
un banco de datos.
27

El monitoreo se realiza en una area representativa llamada "area minima". donde las
parcelas se ubican cubriendo Ia variacion de los microhabitats (Arnolds, 1992), es decir deben
colocarse parcelas en vegetaci6n conservada y lugares abiertos (Cifuentes-Bianco et a/.
1986). debe intentarse que contenga de un 90-95% de las especies fungicas (Fogel, 1992).
Arnolds (1992) establece que el area minima en un estudio de macromicetos en un bosque,
debe ser de cientos de metros.
C 'lndes numeros de cuadrantes chicos son utilizados en estudios donde Ia meta es
obtener el maximo registro de especies y cuadrantes grandes para obtener Ia produccion y
abundancia de las especies (Fogel, 1992). Arnolds (1992) menciona que un estudio de una
comunidad fungica debe tener como numero mfnimo cinco parcelas.
Detecci6n. El prop6sito es documentar Ia producci6n de hongos silvestres y Ia
variabilidad en el espacio y tiempo, es decir, como se comporta Ia producci6n a lo largo del
aiio. En los sitios de manejo de Ia poblaci6n se cuantifica Ia diversidad, que incluye riqueza o
numero de especies y Ia totalidad o abundancia de los individuos (Allen et a/. 1995; Bruns,
1995). Tambien estudia Ia composici6n de especies vegetales, estructura del bosque,
productividad y datos generales del suelo.
Como el inicio de las lluvias es el elemento mas fuerte determinante para el inicio de Ia
epoca de fructificaci6n, este monitoreo debera ser ajustado con el inicio de una abundante
precipitaci6n (Molina et a/. 1993). Los muestreos deberan ser repetidos a lo largo del periodo
de fructificaci6n para obtener resultados completos, ademas, los sitios deberan ser visitados
sucesivamente a lo largo de varios a nos para determinar Ia variaci6n anual; Hering ( 1966).
concluye que al menos ocho aiios son necesarios para caracterizar Ia funga de un sitio.
Tambien se analiza Ia producci6n a lo largo de los aiios con respecto a Ia variaci6n
anual que presente Ia composici6n de especies vegetales, estructura del bosque y edad para
determinar las condiciones 6ptimas para Ia fructificaci6n (Molina et a!. 1993).
Evaluaci6n. Se inicia cuando en el estudio de detecci6n, se observa una declinaci6n
de especies o de Ia producci6n, realizando evaluaciones adicionales para determinar Ia causa
de este efecto y Ia durabilidad en el tiempo (Molina eta!. 1993). Un monitoreo de evaluaci6n
auxilia para conocer las causas de Ia declinaci6n, en este analisis se anexa al monitoreo el
efecto de Ia recolecci6n, remoci6n, tipo de disturbio o cambios en el habitat.
28

lnvestigaci6n. Es el metoda deta!lado de monitoreo, realiza en los sitios de muestreo
intensas observaciones; el objetivo principal es comprender el papel ecol6gico de las
comunidades fungicas en el bosque y observar el comportamiento a largo plazo.
En este metoda se investiga el papel de los hongos en el ecosistema, que papel juegan
en Ia regeneraci6n de los bosques, en los ciclos de nutrimentos 'y como afectan Ia
productividad forestal (Molina eta/. 1993).
En cualquiera de los tipos de monitoreo, para Ia descripci6n tax6nomica se exige
recolectar varios ejemplares de diferente tamano y grado de desarrollo, ya que un s61o
individuo de cada especie es insuficiente para fines cientificos, excepto para las especies de
gran tamano (Cifuentes-Bianco eta/. 1986). Una identificaci6n taxon6mica debe ser realizada
por personas preparadas, las cuales deben poseer un buen entrenamiento y experiencia, asi se
garantiza que toda Ia informaci6n que esta conectada a una especie es veridica (Molina et a/.
1993); Christensen (1989), opina que Ia incorrecta determinacion taxon6mica es un problema
actual presente en los registros de especies.
Mantener un control del monitoreo puede ser dificil en areas • donde el acceso no
puede ser limitado, coordinarse con las autoridades pertinentes es esencial para mantener Ia
integridad de estos estudios (Molina eta/. 1993).
Perspectivas de Conservaci6n Biol6gica. Sin importar el tipo de recurso a aprovechar, el primer
paso para hacer un manejo racional y sostenido, es conocerlos y cuantificarlos (Groombridge,
1992). El exito para el manejo de los hongos silvestres comestibles es conocer Ia biologfa de
estos, Ia importancia en el bosque y que constituyen parte de una actividad econ6mica que
proporciona beneficios econ6micos a Ia poblaci6n (Molina et at. 1993).
Es un hecho que el conocimiento biol6gico de los macrohongos en nuestro pais es
incipiente, hay que diseiiar un plan a largo plazo para conocer Ia micobiota nacional, estos
estudios exigen inversion y son necesarios para mantener Ia pro~ucci6n de hongos
silvestres para futuras generaciones (Molina eta/. 1993).
La gente del bosque interesada exclusivamente en producci6n de madera, esta
reconociendo Ia importancia ecol6gica de los macromicetos, asi como su valor econ6mico
como producto del bosque cultivado (Read eta!. 1989).
29

Para poder disefiar un plan de manejo racional en una zona ~onde se desarrollen
hongos, se deben conocer los aspectos biol6gicos de estas especies, donde destacan (Molina
eta!. 1993):
a)- Que especies fructifican yen que cantidades.
b)- Que cantidades pueden ser recolectadas sin afectar el funcionamiento del sistema.
c)- Como el manejo del bosque afecta a las diferentes especies de hongos.
d)- Como incrementar Ia producci6n de biomasa debido al manejo forestal.
e)- Que especies estan en peli ._ ·o local de extinci6n.
En los bosques del pais cuando se han cumplido las medidas anteriores, se siguen los
siguientes lineamientos generales, adecuandolos a Ia especie que se quiera proteger; Arnolds
{1992), reconoce que no existe un manejo que beneficie a todos los grupos ecol6gicos, como
ejemplo del manejo especial lo son las Morchelias, que fructifican despt.~es de un incendio y
los Cantarelos, que fructifican bajo condiciones en disturbio (Molina eta/. 1993).
Los lineamientos generales para el manejo de los bosques con proposito de un
aprovechamiento sustentable de los macrohongos comestibles, son:
(a)- Para un maximo aprovechamiento, se deben recoger los carp6foros en un intervale corto
para evitar que los ejemplares sean encontrados en estado de senecencia, pudrici6n o que
sean consumidos por gusanos o pequenos mamfferos; tambien existe perdida si se recogen los
carp6foros en estado juvenil debido a que no alcanzan Ia biomasa maxima, (Arnolds, 1992;
Vogt eta!. 1992).
(b)- Hay que evitar dejar grandes claros en los aprovechamientos forestales, deben explotarse
en baja intensidad, dejando ahf los desperdicios de Ia extracci6n de madera, Norvell { 1985) y
Molina eta!. { 1993) encontraron que este tipo de manejo de baja intensidad es favorable para
Ia fructificaci6n de las especies micorrfzicas; por su parte Wasterlund e lngeli:it ( 1981)
concluyen que un bosque con desechos organicos es mas productive.
(c)- Debe respetarse Ia vida silvestre en los bosques, ya que el regreso de Ia mycobiota en
zonas taladas y cortadas depende en parte de Ia dispersion de los animales (Schoonmaker y
Mckee, 1988), esto corre peligro par el constante deterioro que estan sufriendo los bosques,
acortando el habitat de Ia fauna. La mycobiota es excepcionalmente considerada en Ia
planeaci6n del manejo del bosque (Arnolds, 1988).
30

d)· Deben elaborarse !eyes que prohiban recolectar las especies en peligro de extinci6n de una
regi6n, imponiendo medidas restrictivas en las especies comestibles, ya que pueden servir
como fuente futura de alimento (Groombridge, 1992; Norvell, 1985). Estas !eyes pueden
aplicarse a nivel de los intermediarios de Ia producci6n de hongos silvestres, prohibiendoles
comprar especies en peligro de extinci6n y a nivel de recolectores, sancionando a quien
infringa esta norma.
Para conservar Ia diversidad de los hongos silvestres comest:oles, deben crearse
algunas areas de tamano suficiente donde no exista alteraci6n. El terrer estos habitats es
necesario ya que existen especies incapaces de persistir en un pequeno habitat fragmentado
(Groombridge, 1992; Molina et a/. 1993), para ser sustentable econ6micamente estos
espacios pueden funcionar como parques abiertos al publico, con Ia prohibici6n de no causar
dano, un ejemplo son los Parques Nacionales de Washington que cobran a las familias una
cuota para poder permitirles que tomen 50 Iibras de hongos, autorizando solo dos visitas al
ano (Molina eta/. 1993).
31


















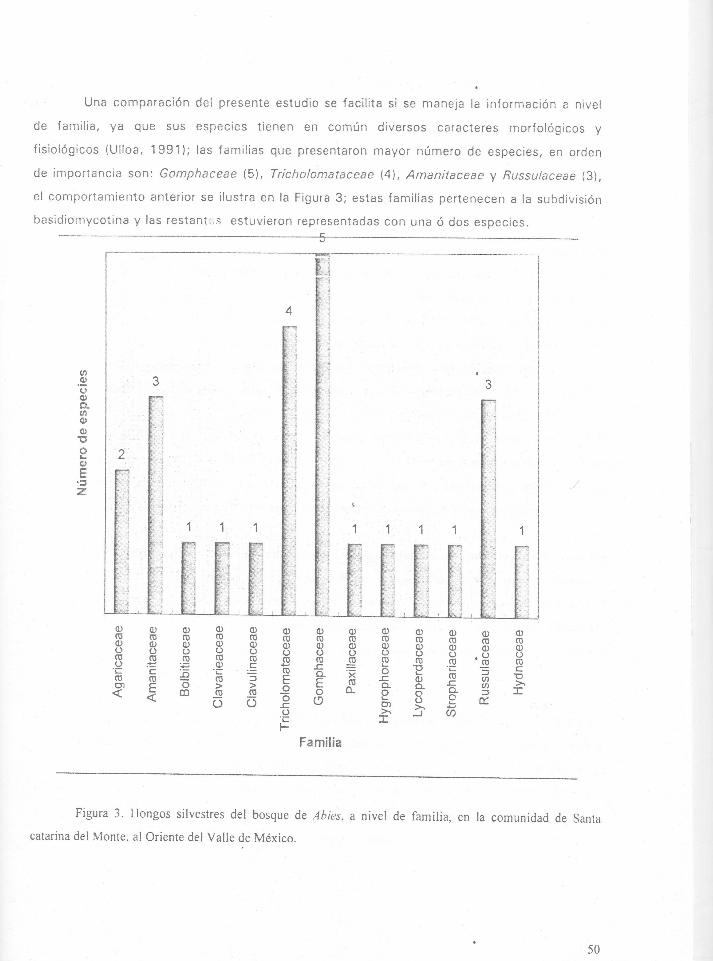




















producci6n comercial de cultivo reduce los gastos en el control ambiental, d isminuyendo el
costo del producto.
La riqueza encontrada en el presente trabajo corre peligro debido a Ia explotaci6n del . .
bosque observada en Ia zona, Ia cual ya ha sido advertida por Gonzalez (1982) y un equipo
interdisciplinario de Ia UACH (An6nimo, 1986), que reportan que sobre los bosques de Ia zona
existe una fuerte presi6n por Ia extracci6n de madera, lena, hierbas medicinales, musgo y
tierra, se debe disenar un plan de manejo integral del bosque, para proteg '=: r esta diversidad,
este debe estar sustentado en tres bases: (a)- traer beneficia social a Ia zona ; (b)- ser rentable
econ6micamente; y (c) - ser racionalmente ecol6gico.
Macromicetos Dominantes. Una especie se considera dominante si presenta un mfnimo de 5%
de Ia producci6n, estimada en alguna medida cuanti f icable, como forma o numero, de acuerdo
con Hering (1966) . Como ya se dijo en este trabajo se eva lu6 en numero de carp6foros y en
peso fresco, considerando en forma separada estos parametres las especies domiantes varian,
ya que existe una correlaci6n baja entre numero y biomasa; al respecto Hunt y Trappe ( 1987),
reportan una r2 de s61o 0.15.
Las especies dominantes respecto a numero de carp6foros fueron: Gomphus f/occosus
con 187 carp6foros, Pholiota sp. con 187 carp6foros: Collybia dryophila con 175 carp6foros y
Lactarius scrobicu/atus con 139 carp6foros, como se observa en Ia Figura 7 . En Ia Figura 8 se
aprecia que dichas especies representaron el 58.4% de toda Ia producci6n en numero de
carp6foros
Considerando a las especies dominantes en peso fresco en el area representativa del
bosque de Abies fueron: Lactarius scrobiculatus, Gomphus floccosus, Lactarius salmonicolor
y Sarcodon scabrosus, estas cuatro especies constituyeron el 69.6% de toda Ia productividad
en peso, como se observa en Ia Figura 9.
Se observa que Ia comunidad fungica estudiada posee pocas especies dominantes y
muchas especies escasas o raras, resultados que corroboran lo propuesto por Margalef
(1974), como una constante en Ia naturaleza.
El hecho de que el 64.6 % (40.8 k/ Ha-1) de Ia producci6n de hongos comestibles sea
aportada por s61o tres especies, es una citra util en un aprovechamiento comercial, ya que Ia
empresa obtendrfa este volumen solo dedicandose a recolectar estas tres especies.
70
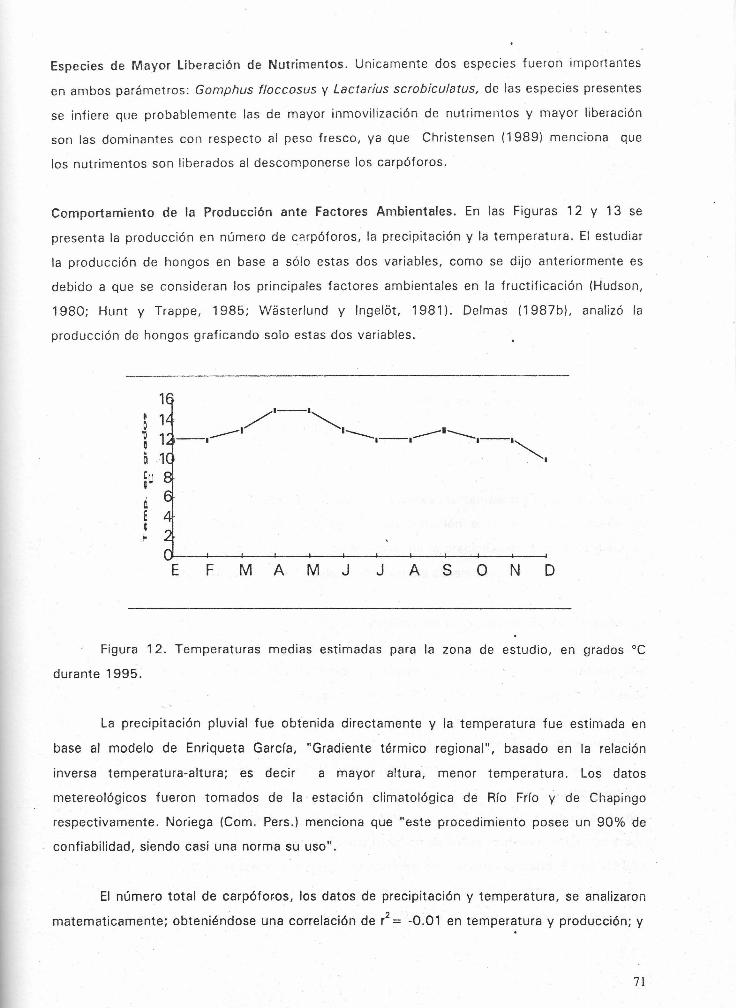
Especies de Mayor Liberaci6n de Nutrimentos. Unicamente dos especies fueron importantes
en ambos parametros: Gomphus floccosus y Lactarius scrobiculatus, de las especies presentes
se infiere que probablemente las de mayor inmovilizaci6n de nutrimentos y mayor liberaci6n
son las dominantes con respecto al peso fresco, ya que Christensen ( 1989) menciona que
los nutrimentos son liberados al descomponerse los carp6foros.
Comportamiento de Ia Producci6n ante Factores Ambientales. En las Figuras 12 y 13 se
presenta Ia producci6n en numero de Ci'lrp6foros, Ia precipitaci6n y Ia temperatura. El estudiar
Ia producci6n de hongos en base a s61o estas dos variables, como se dijo anteriormente es
debido a que se consideran los principales factores ambientales en Ia fructificaci6n (Hudson,
1980; Hunt y Trappe, 1985; Wasterlund y lngelot, 1981). Delmas (1987b), analiz6 Ia
producci6n de hongos graficando solo estas dos variables .
,/1--1~1 I --·--- --.--.-- --·--· ~.
E F M A M J J A s 0 N D
Figura 12. Temperaturas medias estimadas para Ia zona de estudio, en grados oc durante 1995.
La precipitaci6n pluvial fue obtenida directamente y Ia temperatura fue estimada en
base al modelo de Enriqueta Garcfa, "Gradiente termico regional", basado en Ia relaci6n
inversa temperatura-altura; es decir a mayor altura, menor temperatura. Los datos
metereol6gicos fueron tornados de Ia estaci6n climatol6gica de Rfo Frfo y de Chapingo
respectivamente. Noriega (Com. Pers.) menciona que "este procedimiento posee un 90% de
confiabilidad, siendo casi una norma su uso".
El numero total de carp6foms, los datos de precipitaci6n y temperatura, se analizaron
matematicamente; obteniendose una correlaci6n de r2 = -0.01 en temperatura y producci6n; y
71

500 ----------------------------- ----
450 .
400
350
300
250
200
150
100
50
0 -~,
precipiuci6n , / ____ _/
E F M A M A s 0 N '\)
MESES
Figura 13. Precipitaci6n pluvial en mm dura11te 1995 y carpoforos totales en Ia zona
de estudio, localizada al Oriente del Valle de Mexico .
r2 = 0. 73 en precipitaci6n y producci6n. Como se observa, Ia temperatura no correlacion6
para nada con respecto a Ia producci6n, pero Ia precipitaci6n sf tuvo un valor medio de
correlaci6n, esto ultimo indica que el comportamiento de Ia precipitaci6n a lo largo del ano,
influy6 en 73% para que Ia producci6n siguiera Ia tendencia observada.
Moreno-Zarate ( 1990), en un estudio desarrollado en Ia region de Santa Catarina del
Monte en dos vegetaciones dominadas por: Abies religiosa y Pinus hartwegii, encontr6 una
relaci6n directa y positiva entre los factores climaticos, precipitaci6n y temperatura, con
respecto a Ia producci6n observada en Ia zona. Villarreai-Ruiz ( 1994), en bosques del Cofre de
Perote , Veracruz, reporta que de ambos parametres, Ia precipitaci6n fue Ia que mejor
correlacion6 con Ia producci6n y !a relaci6n temperatura-producci6n fue menos evidente;
tambien menciona que el inicio de Ia fructificaci6n esta precedido de un incremento en las
temperaturas, contrastando esto ultimo con el presente estudio donde Ia fructificaci6n fue
precedida por una disminuci6n de Ia temperatura. Como se puede observar al comparar ambos
estudios con el presente, existe un comportamiento unico de estos factores ambientales con
respecto a Ia producci6n, llegando incluso a comportarse de manera contra ria. Fogel ( 1981),
disen6 un modelo para expl!car Ia producci6n en base a dos correlaciones:
biomasa/temperatura y temperatura/precipitaci6n, al aplicarlo observ6 un buen ajuste visual,
72

aunque Ia predicci6n fue mas baja que lo obse rvado , ante este resultado, el autor reconoce Ia
necesidad de incorporar otros factores al modelo, citando como ejemplo Ia condici6n del sitio.
Otros autores (Hunt y Trappe, 1985; Vogt et a/. 1992; Christensen, 1989) tambie n
reconocen que Ia producci6n no solo es afectada por factores ambientales sino que existen
muchos mas, siendo los mas importantes : reservas del micelio, mic6fagos presentes, biomasa
y edad de Ia vegetaci6n. Ante todo lo anterior, el comportamiento de Ia producci6n con
respecto a las vari ables ambientales aquf analizadas, puede servir solo como datos en el
ajuste de un futuro modelo que incluya a todos los factores que presumiblemente afectan Ia
producci6n de los carp6foros.
Analizando las tres graticas se describe Ia producci6n de hongos, el inicio de Ia
producci6n corresponde a un declive de Ia temperatura en mayo, fecha que registr6 una
precipitaci6n de 56.4 mm; Ia cual garantiza en un suelo franco una humedad a capacidad de
campo y alta humedad atmosferica. La fructificaci6n continua en una relaci6n inversa al
decline de Ia temperatura, es decir, mientras mas desciende Ia temperatura, esta aumenta;
durante este perfodo el contenido de humedad sigue Ia misma tendencia, dandose Ia cima de
producci6n y precipitaci6n (218 .9 mm) en el mes de agosto, fecha que registr6 las mas bajas
temperaturas durante este lapso ( 12.2° C). Continua un descenso vertical de Ia producci6n
simultaneamente con un ligero aumento en Ia ter;nperatura y un brusco descenso de Ia
precipitaci6n; despues Ia temperatura retoma su nivel anterior (baja) y existe una precipitaci6n
parecida a Ia del inicio de Ia fructificaci6n, en este punto se da una producci6n similar a Ia del
inicio de Ia fructificaci6n; sigue disminuyendo Ia temperatura (Ia humedad se mantiene)
concomitantemente con Ia producci6n, esta ultima se registra como cero en diciembre, mes
que registro las temperaturas mas bajas del aiio .
En base a las graticas se destacan varios hechos: (a), De'sde el inicio de Ia
fructificaci6n hasta un poco mas de Ia mitad, Ia lfnea de Ia producci6n fue ascendente
simultanea con ia precipitaci6n y en forma inversa con Ia temperatura, el comportamiento de
estos factores ambientales fue 6ptimo para Ia fructificaci6n; (b) el brusco decline de Ia
producci6n puede ser causado por agotamiento del micelio que fructific6 abundantemente 3
meses (junio-agosto), lo anterior por el hecho que al inicio del declive, Ia temperatura aumento
solo un poco y aunque Ia precipitaci6n descendi6 bruscamente, no es 16gico que los niveles de
humedad hayan descendido tambien bruscamente; (c) En los lapsos junio y octubre, meses
con temperaturas y precipitaci6n ·similares (alrededor de 90 mm y 12° C), se obtuvo una
producci6n similar y (d) Ia producci6n a final del aiio con una constante de humedad y una
73

temperatura de 12° C, va disminuyendo conforme lo hace Ia temperatura. Hunt y Trape
(1987) mencionan Ia existencia de especies con un metabolismo adaptado a temperaturas
baj~s para fructificar, por lo que se espera que Ia produci6n descienda conforme Ia
temperatura se aleja del nivel antes mencionado, ya que cada vez son menos las especies
adaptadas a este intervale de temperatura .
En general, Ia producci6n de hongos silvestres con respecto a ambos factores
ambientales se relacion6 visualmente con Ia temperatura, ya cuando ocurrieron cambios en Ia
grafica de Ia temperatura, se observan cambios en Ia producci6n, ver figuras Figura 12 y 13;
existi6 relaci6n pero teniendo como condici6n Ia existencia de una precipitaci6n suficiente
(mayor de de 61 .3 mm) para mantener al suelo a una capacidad de campo. Para concluir
sabre los niveles que deben alcanzar estos factores para que exista fructificaci6n, (CUanto
durara? o (que Ia interrumpira?, hacen falta mas estudios de Ia zona realizados en otros alios
ya que no se puede generalizar, debido a que los fen6menos y condiciones no se repitieron en
el afio.
Una conclusi6n parcial sabre el bosque de Abies, es que existira fructificaci6n a una
temperatura de 11.8 a 12.72° C, teniendo como condici6n una precipitaci6n mayor a 61.3
mm. Comparando estos niveles que deben alcanzar los principales factores ambientales para
que exista producci6n, con los trabajos reportados 'en Ia literatura (factores end6genos) se
observa que cada tipo de bosque, tiene sus factores ambientales propios; lo que refuerza Ia
idea ya mencionada, es necesario estudiar cad a tipo de bosque ya que ·los resultados no se
pueden generalizar.
Periodo de fructificaci6n. La distribuci6n estacional en el bosque de Abies fue de junio a
noviembre, este intervalo es de 6 meses, como se observa en el Cuadro 7.
El mes de agosto registr6 una fructificaci6n maxima del 83% de las especies, tam bien
en esta fecha se obtuvo Ia cima de Ia producci6n de carp6foros en peso fresco. Estos
registros son utiles en un estudio cientiffco para conocer Ia fecha de cuando ,o..o®r .o.b.W.r:\W
mas carp6foros, por consiguiente mas cepas y en una explotaci6n comercial para ver cuando
Ia empresa requerira toda su capacidad instalada.
Como se observa Gomphus floccosus fue Ia unica especie presente durante toda Ia
epoca de fructificaci6n; Agaricus silvicola, Lactarius scrobicu/atus y Lycoperdon umbrinum
74

Cuadro 7 . Momentos de fructificaci6n de las especies de macromicetos , en el bosque de Abies, en Santa Catarina del Monte, al Oriente del Valle de Mexico .
HONG OS SIL VES TRES JUNlO JULIO AGOS10 SEP11E MBRE OC1UBR< NOVIEMBRE
Agaricus silvaticus .! t .! A. silvicola ~ ~ ~ ~
Amani ta fu/va ~ ~ . A. gemmata ~ ~
A. rub escens ~ C/avariadelphus truncatus .! .! Clavulina cristata .! Clitocybe gibbs .! ~ .! .! Col/ybis dryophils .! .! .! ~
Gomphu'i floccosus .! .! .! ~ .! .! Gyromtr -'l• infula .! Helve/Is crisps ~ Hygrophoropsis aurantiaca .! .! .! Hygrophorus chrysodon .! .! H ypomyces lactifluorum .! ~ ' . Laccaria lace era ~ ~ Lactarius sa lmonicolor .! ~ ~ ~
L. scrobiculatus .! ~ .! ~ .! Lycoperdon umbrinum .! .! .! .! .! Morchella esculenta .! Pholiota sp. ~ .! Psatirella spadices .! Raman's botrytis .! R. 1/sva .! R. sanguinea .! .! R. stric ta ~ Russula brevipes .! ~ Sarcodan scabrosus .! ~ Sarcosphaera eximia .! ~ Tricholoma flavoviren s .! .! ~
fueron especies que estuvieron presentes 5 meses. Este intervalo de aparici6n nos muestra
que poseen gran amplitud ecol6gica para fructificar; debido a su capacidad adaptativa a las
diversas condiciones ambientales observadas, por lo que estas especies serfan al menos
teoricamente mas faciles de domesticar.
Las especies de los generos Gomphus y Lactarius revisten mas esfuerzo de
domesticaci6n al ser especies micorrfzicas debido a Ia complejidad para reproducir el estado
simbi6tico in vitro e inducir su fructificaci6n (Stone, 1996); sin embargo, est as especies
dentro del grupo micorrfzico aquf estudiado, son las especies mas faciles de domesticar, ya
que present an un gran intervalo de condiciones apropiadas para fructificar.
Ritmo de Fructificaci6n de las Especies. Las especies del presente trabajo se clasificaron de Ia
siguiente manera, de acuerdo con Villarreai-Rufz ( 1994):
A- 3% con fructificaci6n temprana corta.
Pho!iota sp.
8- 20% temprana prolongada.
Co!lybia dryophi!a, Gomphus floccosus, Hygrophoropsis aurantiaca, Lactarius
salmonicolor, L. scrobiculatus, Lycoperdon umbrinum.
75.

lilJiioteca de A£rcacolegfa C- 33% fructificaci6n corta a mediados de Ia estaci6n.
Amanita gemmata, A. rubescens, Clavulina cristata, He/vella crispa, Laccaria
/accata, Psatirella spadicea, Ramaria botrytis, R. flava, R. sanguinea, R.
stricta.
0- 20% con fructificaci6n tardfa prolongada.
Agaricus silvaticus, A. si/vicola, C/itocybe gibba, Hypomyces lactif/uorum,
Sarcosphaera eximia, Tricholoma f/avovirens
E- 23% con fructificaci6n tardfa corta.
Amanita fulva, Clavariadelphus truncatus, Gyromitra infula, Hygrophorus
chrysodon, Morchella esculenta, Russula brevipes, Sarcodon scabrosus.
Actualmente Ia gente que habita los bosques de Ia regi6n ("hongueros", leiiadores) y
que aprovecha a los hongos en forma tradicional, consideran que Ia producci6n ha disminufdo.
Exis~en muchas causas probables no fundamentadas cientfficamente: sequfas, extracci6n de
arboles, manejo forestal, pr<kticas agrfcolas y Ia recolecci6n (Guzman y Villarreal, 1984;
Norvell, 1995). La ultima practica es considerada injustamente como Ia principal causa
(Norvell, 1995). por gente que no posee un conocimiento micol6gico basico, mas como ya se
mencion6, no existe impacto por Ia recolecci6n al menos en el corto plazo de tres aiios
(Arnolds, 1988; Norvell, 1995; Wasterlund y lngelot, 1981).
Sobre el impacto de Ia recolecci6n a largo plazo, s61o existe el estudio de Norvell
( 1995). quien concluye que no existe efecto por Ia recolecci6n despues de 1 0 a nos de
evaluaci6n; Se requieren estudios similares para poder concluir que no existe impacto a largo
plazo.
AI recolectar no se aprovechan todos los carp6foros, ya que algunos de ellos se
encuentran en estado de descomposici6n. En este sentido Mildh ( 1978). en Finlandia reporta
que 70% de Ia producci6n noes aprovechada por el ataque de larvas, Kalames y Silver (1988)
en Estonia reportan de 1/5 a 1/2 de hongos inapropiados para el consumo humano; Villarreai
Ruiz (1966) menciona en Michoacan una descomposici6n del 56%; y el presente trabajo 6%.
La existencia de algunos carp6foros en estado de descomposici6n que !Iegan a su madurez
fisiol6gica puede ser una explicaci6n del por que no existe efecto a corto plaza, debido a dos
hechos: (a)- los carp6foros producen millones de esporas (Kendrick, 1992). dispersandose asf
miles de elias provenientes de los carp6foros no recolectados y (b)- al recolectar queda el
micelio, este seguira degradando; nutriendose y fructificando, como se dijo anteriormente
existen registros de micelios de hasta 750 aiios de longevidad (Arnolds, 1992).
76

Lo anteriormente expuesto nos da idea, que es mas factible que Ia perdida de
diversidad fungica, sea ocasionada por Ia mala explotaci6n forestal y las practicas agrfcolas;
actividades que si afectan al micelio.
77

CONCLUSIONES
1- La riqueza de especies de hongos identificados en el presente trabajo (30 especies), fue
superior a trabajos similares desarrollados en Ia misma zona; con respecto. a Ia producci6n esta
fue menor, total izando 63.2 kg .
2- El bosque de Abies estudiado present6 una sucesi6n de especies fungicas, con momentos
de fructificaci6n yuxtapuestos, totalizando una amplia epoca de fructificaci6n; de Junio a
Noviembre con una cima en Ia producci6n y fructificaci6P gn el mes de agosto .
3- Las especies estudiadas pertenecen en su mayorla al grupo ecol6gico de los degradadores,
en 57%, al grupo mutualista un 33% y el antagonista con 3%.
4- La abundancia en el area medida en numero de carp6foros, fue dominada por pocas
especies: Collybia dryophi/a, Gomphus ffoccosus, Pho/iota sp. y Lactarius scrobicu/atus; y en
peso fresco: Gomphus ffoccosus, Lactarius salmonico/or, L. scrobiculatus y Sacordon
scabrosus .
5.- Las especies con mayor importancia ecol6gica, en cuanto a liberaci6n de nutrimentos son
Gomphus floccosus y Lactarius scrobiculatus.
6.- Existi6 fructificaci6n a temperaturas ambientales de 11.8 a 12.72° C, teniendo como
condici6n una precipitaci6n mayor a 61.3 mm.
7- La producci6n correlacion6 visualmente con Ia temperatura y numericamente con Ia
precipitaci6n (correlaci6n producci6n y precipitaci6n = . 73)
6- El bosque de Abies de Ia comunidad, present6 microhabitats, ocasionados por cambios en
Ia vegetaci6n y las condiciones topograticas.
78

RECOMENDACIONES
- Se requiere mas investigaci6n como Ia del presente trabajo, en el area, Ia cual debe abarcar
varios aiios.
- La industria regional de producci6n de hongos, debe intentar domesticar especies de este
almacen genetico, ya que existe Ia garantfa de que Ia cepa estara adaptada a Ia zona,
disminuyendo el gasto por el control ambiental, lo que harfa competitive este producto.
- Es necesario realizar investigaciones micol6gicas que cubran cada tipo de bosque del pafs,
ante Ia fuerte presi6n que existe sobre ellos como fuente de diversos recursos.
- Un programa de aprovechamiento por los pobladores del recurso hongos silvestres del
bosque, debe estar sustentado en un manejo integral que incluira especies de diferentes
hongos de importancia comercial; el primer paso en este aprovechamiento es el conocimiento
de esta investigaci6n por parte de los pobladores.
- Para evitar Ia perdida de diversidad de macromicetos, es necesario un manejo integral del
bosque con Ia finalidad de conservar el equilibria ecol6gico, 6ptimo para Ia fructificaci6n de los
hongos en el area de estudio.
79

LITERATURA CIT ADA
AINSWORTH and BISBY • S. 1963. Dictionary of the fungi. Ainsworth, G. C. Ed. Surrey.
ALEXOPOULOS, C. J. 1977. lntroducci6n a Ia micologfa. Universitaria, Buenos Aires.
ALFARO-MAYA, C. 1996. Los hongos comestibles y su uso en arreglos decorativos. Goegraffa
Agricola (Estudios de Ia agricultura Mexicana), UA( '-l.
ALLEN, E.B., M.F. ALLEN, D.J. HELM, J.M. TRAPPE, R. MOLINA and E. RINC6N. 1995.
Patterns and regulation of mycorrhizal plant and fungal diversity. Plant and Soil 170: 47-62.
ANONIMO. 1986. Caracterizaci6n de una comunidad del sotomontano de Ia Sierra de Rio Frio (Santa
Catarina del Monte, Texcoco, Mexico). Equipo interdisciplinario. UACH, Chapingo.
ARNOLDS, E. 1988. The changin macromycete flora in the Netherlands. Trans. Br. Mycol.
Soc. 90 (3): 391-406.
ARNOLDS, E. 1992. The analysis and classification of fungal communities with special reference
to macrofungi. In: W. Winterhoff (Ed.). Fungi in Vegetation Science. 7-47. Kluwer Academic
Publishers, Bristol.
AROCHE, R., J. CIFUENTES, F. LOREA, P. FUENTES, J. BONAVIDES, H. GALICIA, E.
MENENDEZ, 0. AGUILAR, y V. VALENZUELA. 1984. Macromicetos t6xicos y comestibles de
una regi6n comunal del Valle de Mexico. Bal. Soc. Mex. Mic.19: 291-318.
ASTEINZA-BILBAO, G. 1993. Impacto ambiental, productivo y econ6mico de Ia desertificaci6n en Ia
porci6n Oriental del Valle de Mexico. In: Delafuente, J., R. Ortega y M. Samano (Coord.). Agricultura y
agronomfa en Mexico. 500 ai'ios, UACH, Chapingo.
AZCON-G. C. y J. M. BAREA. 1980. Micorrizas. lnvestigaci6n y Ciencia. 4 7: 8-16.
BILLS, G. F., G. I. HOLTZMAN, and 0. K. MILLER, JR. 1986. Comparison of ectomycorrhizal
basidiomycte communities in red spruce versus northern hardwood forests of West Virginia.
Can. J. Bot. 64:760-768.
80

BOHUS, G. and BABOS, M. 1973 . Data to the role of terricolous macrofungi in deciduous forest
ecosystems. Miko/6giai Kozlemenyel. 2:77-79.
BRUNS, T. D. 1995. Thoughts on the processes that maintain local especies diversity
de ectomycorrhizal fungi. Planta and Soil. 170 : 63-73.
BUJAKIEWICZ, A. 1982. Macromycetes as an element of forest structure on the Babia G6ra
Massif. In: Struktur and Dynamik Von Waldern. J. Cramer, Vad;: z.
CIBRIAN-TOVAR, D., J. T. MENDEZ-MONTIEL, R. CAMPO-BOLANOS, H. 0. YATES-Illy J.
E. FLORES-LARA. 1995. lnsectos forestales de Mexico/ forest insects of Mexico. UACH,
Chapingo.
CIFUENTES-BLANCO, J. , M. VILLEGAS y L. PEREZ-RAMiREZ. 1986. Horgos. In: Lot, A. y F.
Chiang (Camps.). Manual de herbaria. Consejo Nacional Para Ia Flora de Mexico, Mexico, D.F.
CLARKE, R. 1986. The Handbook of Ecological Monitoring. Clarendon Press, Oxford .
CONNELL, J. H. and M. D. LOWMAN. 1989. Low-diversity tropical rain forests: some possible
mechanisms for their existence. Am. Nat. 134 (1 ): 88- 119. '
CONRADO SOTO. 1997. Hongos silvestres comestibles de Mexico. In . Camara nacional de Comercio de
Guadalajara (Secci6n agronegocios). I ciclo de conferencias sobre producci6n comercial de hongos. 19 al
22 de febrero de 1997, Guadalajara.
CHAC6N, S. y G. GUZMAN. 1995. Observations on the phenology of t~n fungal especies in
the subtropical forests at Xalapa, Mexico. In: Mycol. Res. 99 (1):54-56.
CHANG, S. T. and P. G. MILES. 1989. Edible mushrooms and their cultivation. CRC, Boca
Raton.
CHRISTENSEN, M. 1989. A view of fungal ecology. Mycologia. 81(1):1-19.
DEACON, J. A., S. J. DONALDSON and F. T. LAST. 1983. Sequences and interactions of
mycorrhizal fungi on birch. In: Hawksworth, D. L. (ed.). Frontiers in mycology. Honorary and
81

genera l lectures from the Fourth International Mycological Congress, Regensburg. 1990.
CAB . International, Gran Bretana.
DELMAS, J. 1987a. Cultivation of new species of edible fungi present state and prospects.
Indian Mushroom Sciences 2: XXVII-XXXVIII.
DELMAS, J. 1987b. Fruiting requirements of fungi under natural and artificial condit ,-·ms.
Indian Mushroom Science 2: 219-229.
DIGHTON, J. and l. BODDY. 1989. Role of fungi in nitrogen, phosphorus and sulphur cycling
in temperate forest ecosystems. In: Nitrogen, phosphorus and sulphur utilization by fungi.
Boddy, L., R. Marchant yD. J. Read. (Eds .). British Mycological Society, Cambridge .
DIGHTON, J., J. M. POSKITT and D. M. HOWARD. 1986. Changes in ocurrence of
basidiomycete fruitbodies during forest stand development: with specific reference to
mycorrhizal species. Trans. Br. Mycol. Soc. 87 ( 1): 163-171.
DIGHTON, J . . J. M. POSKITT, and D. M. HOWARD. 1986. Changes in ocurrence of
Basidiomycete fruitbodies during forest stand development: with specific reference to '
mycorrhizal species. Trans. Br. Mycol. Soc. 87(1):163-171.
DUBOVOY. C. 1986. Conocimiento de los hongos en el Mexico antiguo. Bole tin lnformativo
de Ia Sociedad Mexicana de Micologfa. 2: 16-24.
DURAN-DEGRAU, N. yR. P. LLUVIA. 1980. los hongos. JOVER. Barcelona.
FERNANDEZ, D., V. M. BANDALA, L. MONTOYA, y G. GUZMAN. 1994. Contribuci6n al
conocimiento de los hongos del Bosque Tropical de Ia Mancha en Veracruz. In. Sociedad Mexicana de
Micologia y Universidad de Guanajuato. 5 Congreso Nacional de Micologia. 27 a 30 de noviembre de 1994.
Programas y memorias, Guanajuato.
FIERROS-LOPEZ, M. y L. GUZMAN-DAVALOS. 1994. Introducci6n al conocimiento de los
macromicetos de Ia Sierra de Quila, Jalisco. In. Sociedad Mexicana de Micologia y Universidad de
Guanajuato. 5 Congreso Nacional de Micolo~ia. 27 a 30 de noviembre de 1994. 'programas y memorias,
Guanajuato.
82

FOGEL, R. 1981. Quantification of sporocarp produced by hypogeous fungi. In: Wicklow, D.T.
y G.C. Carroll (Eds.) . The fungal community: its organizat ion and role in ecosystems . Marcel
Decker, New York.
FRANCO-L6PEZ, J.,G. DELACRUZ-AGUERO, A. CRUZ-G6MEZ, A. ROCHA-RAMiREZ, N.
NAVARRETE-SALGADO, G. FLORES MARTiNEZ, E. KATO-MIRANDA, S. SANCHEZ-COLiN, L.G.
ABARCA-ARENAS, C. M. BEDIA-SAMANEZ y !.WINFIELD-AGUILAR. 1985. Manual de
ecologfa. Trillas. D.F.
FRANKLAND, J. C. 1992. Mechanism in fungal succession. In: Wicklow, D.T. y G.C. Carroll
(Eds.). The fungal community: its organization and role in ecosystems. Marcel Decker, New
York.
GARCiA, E. t 978. Los climas del valle de Mexico segun el sistema de clasificaci6n climatica de Koeppen
modificado porIa autora. CP/ SARH, Montecillo.
GARZA, F., J. GARCiA, y J. CASTILLO. 1985. Macromicetos asociados al bosque de Quercus
rysophy/la en algunas localidades del centro del estado de Nuevo Leon. Rev. Mex. Mic. 1 :423-
437.
GIS PERT, M ., 0. NAVA y J. CIFUENTES. 1984. Estudio comparative del saber tradicional de
los hongos en dos comunidades de Ia Sierra del Ajusco. Bol. Soc. Mex. Mic. 19:253-264.
GONZALEZ, J. 1982. Notas sobre Ia etnomicologfa nahuatl. Bol. Soc. Mex. Mic. 17: 181-186.
GONZALEZ, R. 1981. Ecologfa humana y etnobotanica de un pueblo campesino de Ia Sierra
Nevada, Mex., Santa Catarina del Monte. Tesis de Licenciatura. Fac. de Ciencias, UNAM,
Mexico, D.F.
GONZALEZ-LOERA, J. yD. LOPEZ-VELAZQUEZ. t99la. Estudio ecol6gico de Ia interacci6n
vegetaci6n-suelo-recuperaci6n en Ia zona erosionada del municipio de Texcoco, Edo. de Mexico. In: Anaya
Perez, M. A., J. A. Castellanos- Suarez y J. Ocampo, L. (Comps.). Segundo foro de investigaci6n y servicio
del Oriente del estado de Mexico. UACH, Chapingo.
83

GONZALEZ-LOERA, J. y D. L6PEZ-VELAZOUEZ. 1991 b. Los recursos vegetales silvestres en
el Municipio de Texcoco, Mexico. In: Ortega-Paczka, R., G. Pa lomino-Hasbach, F. Castillo
Gonzalez, V .A . Gonzalez-Hernandez y M. Livera-Murioz (Eds.). Avances en el estudio de los
recursos Fitogeneticos de Mexico. Sociedad Mexicana de Fitogenetica, A.C . (SOMEFI).
Chapingo.
GROOMBRIDGE, B. (ED.). 1992. Global biodiversity. Status of the earth's living resources.
Chapm ·n& Hall, Londres.
GUZMAN, G. 1984. El uso de los hongos en Mesoamerica . Ciencia y Desarrollo. 59: 17-27.
GUZMAN, G. 1994. Las colecciones de hongos en Mexico y su problemati~a en Ia biodiversidad
del pais. Bot. Soc. Bot. Mexico. 55: 35-37.
GUZMAN, G., V. M. BANDALA, L. MONTOYA, S. CHACON, y G. CARRION. 1994. La diversidad
fUngica en el estado de Veracruz. Base de datos de las especies registradas. ln . Sociedad Mexicana de
Micologia y Universidad de Guanajuato. 5 Congreso Nacional de Micologia. 27 a 30 de noviembre de 1994.
Programas y memorias, Guanajuato.
GUZMAN, G. 1995. La diversidad de hongos en Mexico. Ciencias. 39, julio-septiembre: 52-
57.
GUZMAN, G. y l. VILLARREAL. 1984. Estudio sobre los hongos, lfquenes y mixomicetos del
Cafre de Perote, Veracruz, I. lntroducci6n a Ia micoflora de Ia regi6n. Bol; Soc. Mex. Mic. 19:
107-124.
GUZMAN-DAVALOS, L y G. GUZMAN. 1979. Estudio ecol6gico comparative entre los hongos
(macromicetos) de los bosques tropicales y los de confferas del sureste de Mexico. Bol. Soc.
Mex. Mic. 13:89-125.
HACSKAYLO, E. 1983. Researching the potential of forest tree mycorrhizae. In: Atkinson, D.,
K. K. S. Bhat, M. P. Coutts, P. A. Mason, D. J. Read (Eds.). Tree root systems and their
mycorrhizas. Martinus Nijhoff I Dr. W. Junk. Publishers, The Hague.
HAWKER, L. E. 1957. The physiology of reproduction in fungi. The syndics of the Cambridge
university. Gran Bretaria.
84

HAWKSWORTH, D.L. 1991. The fungal dimension of biodiversity: magnitude, significance, ·
and conservation. Mycological Research 95:641-655.
HAWKSWORTH, D.L., B.C . SUTION and G.C. AINSWORTH. 1983. Ainsworth & Bisby 's
dictionary of the fungi. Commonwealth Mycological Institute C.A.B, Kew.
HERING, T. F. 1966. The terricolous higher fungi four lake district woodland. Trans. Br.
mycol. Soc. 49: 369-383.
HERRERA, T. 1994. Perspectivas de Ia investigaci6n en micologfa. Bol. Soc. Bot. Mexico. 55:
39-44.
HUDSON, H. J. 1980. Fungal saprophytism.(Sec. edt.). The Camelot. Southampton. Gran
Bretafia.
HUNT, G.A. and J.M. TRAPPE. 1987. Seasonal hypogeous sporocarp production in a western
Oregon Douglas-fir stand. Can. J. Bot. 65: 438-445.
IBAR, L. 1980. C6mo ... buscar, conocer, guisar y con~ervar setas. AEDOS, Barcelona.
INEGI. 1990a. Estado de Mexico. Resultados definitivos. Datos por localidad (lntegraci6n territorial). XI
Censo General de poblaci6n y vivienda, 1990, Aguascalientes.
INEGI. I990b. Texcoco. Cuademo de informaci6n basica para Ia planeaci6n municipal, Aguascalientes.
KALAMEES, K. and S. SILVER. 1988. Fungal productivity of pine healths in North-West
Estonia. Acta Bot. Fennica 136: 95-98.
KAUL, T.N. 1987. Fruiting in larger fungi. A review. Indian Mushrooms Sciences 2: 230-253.
KENDRICK, B. 1992. The fifth Kingdom (Sec. Ed.). Mycologue Publications: Ontario.
LARGENT, D. 1973. How to identify mushrooms (to genus) using only macroscopic features.
Mad River, Eureka.
85

LARGENT, 0; 1977. How to identify mushrooms to genus 1: Macroscopics features . Mad
River, Eureka .
LARGENT, O.L. and H.O. THIERS. 1977. How to identify mushrooms to Genus II : Field
identification of Genera. Mad River, Eureka.
LARGENT, O.L., D. JOHONSON and R. WATLING. 1980. How to identify mushrooms to
Genus Ill: microscopic features. Mad River, Eureka.
LE-TACON, F. 1985. Las micorrizas, una cooperaci6n entre plantas y hongos. Mundo
cientffico. 49:777-784.
MAPES, C., G. GUZMAN y J . CABALLERO, 1991. Etnomicologfa Purepecha: El conocimiento
y uso de los hongos en Ia cuenca de Patzcuaro, Michoacan. Dir. Gral. de Culturas Populares,
Soc. Mex. de Micologfa, lnst. de Biologfa, UNAM, Mexico, D.F.
MARGALEF, R. 1974. Ecologfa. Omega., Barcelona
MARiA-ARiAS, R., J. GARCiA, y F. GARZA. 1994. Estudio de macromycetes del Cerro "el Potosi",
Mpio. De Galeana, N. L. In. Sociedad Mexicana de Micol6gia y Universidad de Guanajuato. 5 Congreso
Nacional de Micologia. 27 a 30 de noviembre de 1994. Programas y memorias, Guanajuato.
MARKKULA, M. and A. RANTAVAARA. 1996. Consumption of mushrooms and some other
wild food products in f inland. Http://www.geirik.is/Nsfs/markkula.htm.
MARTiNEZ-SALDANA, T. y A, ROMERO-CONTRERAS. 1991. La consol idaci6n del estado en
el Imperio Texcocano. In: Anaya- Perez, M.A., J. A. Castellanos- Suarez y J. Ocampo, L. (Comps.).
Segundo foro de investigaci6n y servicio del Oriente del estado de Mexico. UACH, Chapingo.
MASON, P. A., J. WILSON, F. T. LAST and C. WALKER. 1983. The concept of succesion in
relation to the spread of sheathing mycorrhizal fungi on inoculated tree seedlings growing in
unsterile soils. In: Atkinson, D., K. K. S. Bhat, M. P. Coutts, P. A. Mason, D. J. Read (Eds.).
Tree root systems and their mycorrhizas. Martinus Nijhoff I Dr. W. Junk. Publishers, The
Hague.
86

MCNAUGHTON, S. J. and l. l. WOLF. 1984. Ecologfa general. Omega, Barcelona.
MILDH, U. 1978. The organization for collecting forest mushroom in Finland. Karstenia 18
(Sup!.): 106-107.
MITIERMEIER-RUSSELL, A. y G. D. MITIERMEIR, C. 1992. La importancia de Ia diversidad
biol6rica de Mexico . In: Sarukhan, J . y R. Dirzo (Comp.). Mexico ante los retos de Ia
biodiversidad. CONABIO, Mexico.
MOLINA, R., T. O'DELL, D. LUOMA, M. AMARANTHUS, M. CASTELLANO, and K. RUSSELL.
1993. Biology, ecology, and social aspects of wild edible mushrooms in the forests of the
Pacific Northwest: a preface to maintaing commercial harvest . Gen.Tech.Rep.PNW-GTR-
309.Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research
Station.
MORENO-ZARATE, C. 1990. Los hongos comestibles: Un componente de Ia productividad del
bosque en Santa Catarina del Monte, Mexico. Tesis de Maestrfa en Ciencias. Colegio de
Postgraduados, Montecillo, Mexico.
NORVELL, l. 1995. Loving the chanterelle to death? the ten-year Oregon chanterelle project.
Mcllvainea 12 (1): 6-25.
ORTIZ-SOLORIO, C. A. y H. E. CUANALO-DE LA CERDA. 1977. Levantamiento fisiografico del area
de influencia de Chapingo. Para Ia cartografia de tierras erosionadas. UACH, Chapingo.
ORTIZ-SOLORIO, M. D. M., M.D. C. GUTIERREZ-CASTORENA y M.D. C. OLMEDO
BOLANOS. 1993. Modificaciones en el paisaje inducidas por el hombre en Ia Subcuenca del Rio Texcoco.
In: Delafuente, J., R. Ortega y M. Samano (Coords.). Agricultura y agronomia en Mexico. 500 anos. UACH,
Chapingo.
PEREZ-SILVA, E. 1971. Algunos macromicetos de Chiapas (Mexico). Bol. Soc. Mex. Mic.
5:23-35.
PIELOU, E. C. 1996. Shannon's formula as a measure of especific diversity: its use
and misuse. Am. Nat. 100 (914): 463- 465.
87

PIROZYNSKI. K. A. 1981. Interactions between fungi and plants through the ages. Can. J.
Bot. 59: 1824- 1827.
RAMBELU, A. 1985. Manual on mushroom cultivation. FAO, Roma.
READ, D. J. 1991. Mycorrhizas in ecosystem- nature's response to the "law of the
minimu-n" In: Hawksworth, D. L. (ed.). Frontiers in mycology. Honorary and general lec.'"'res
from the Fourth International Mycological Congress, Regensburg. 1990. CAB. International,
Gran Bretafia.
READ, D. J ., J. R. LEAKE y A . R. LANGDALE. 1989. The nitrogen nutrition of mycorrhizal
fungi and their host plants. In : Boddy, L., R. Marchant y D. J . Read (Eds.). Nitrogen,
phosphorus and sulphur utilization by fungi. British Mycological Society, Gran Bretafia.
REYES-VILLEGAS, J. L. 1992. Evaluaci6n de Ia edofauna en 6 diferentes tecnicas de roturaci6n del
tepetate en Santa Catarina del Monte. Texcoco. Edo de Mexico. Tesis de Jicenciatura. UACH, Chapingo.
RICE, M, and D. BEEBEE, 1980. Mushromms for color. MAD RIVER, Eureka.
RZEDOWSKI, J. 1991. Diversidad y origenes de Ia flora fanerogamica de Mexico. Acta
Botanica Mexicana. 14:3-21.
SAS INSTITUTE. 1982. SAS users's guide. SAS Institute Inc., Raleigh.
SCHOONMAKER, P. and A. MCKEE. 1988. Species composition and diversity during
secondary succession of coniferous forest in the Western Cascade Mountains of Oregon. In:
Forest Science. 34 (4) :960-979.
SHULTES, R. E. 1983. Plantas alucinogenas . Medica/Mexicana, Mexico, D.F.
SERRA TO, C. M. A., NAVARRETE, C. y M. PEREZ, V. 1991. Composici6n de ofrendas en panteones de
algunas comunidades del municipio de Texcoco, Mexico, en 1990. In : Anaya- Perez, M. A., J. A.
Castellanos- Suarez y J. Ocampo, L. (Comps.) . Segundo foro de investigaci6n y servicio del Oriente del
estado de Mexico. UACH, Chapingo.
88

SINGER, R. 1961. Mushrooms and truffles. lnterscience, Gran Bretana.
SOSA-RAMIREZ, M. L. S., S. RANGEL-CALDERON, y R. VALENZUELA-GARZA. 1994. Lista
preliminar de los macromicetos de Ia regi6n Centro-Norte del estado de Hidalgo. ln. Sociedad Mexicana de
Micologia y Universidad de Guanajuato. 5 Congreso Nacional de Micologia. 27 a 30 de noviembre de 1994.
Programas y memorias, Guanajuato.
SPP. 1983. Estados Unidos Mexicanos. E 14831. Chalco. Mexico y Distrito Federal. Carta edafo16gica.
Mexico, D.F.
SPP.1981. Nomenclator del estado de Mexico.SPP. Mexico, D. F.
STAMETS, C. 1993. Growing gourmet & Medicinal Mushrooms. Ten Speed. Berkefey.
STATES, J. S. 1981. Useful criteria in the description of fungal communites. In: Wicklow,
D.T. y G.C. Carroll (Eds.). The fungal community: Its organization and role in ecosystems.
Marcel Decker, New York.
STONE, w. 1996. A short mushroom primer. The fungus kingdom.
http://www ./clark .edul-wstone /stuff /primer .html.
SUTTON, B. y P. HARMON. 1991. Fundamentos de ecologfa. Limusa. Mexico, D.F.
TANESAKA, E., H. MASUDA and K. KINUGAWA. 1993. Wood degrading ability of
basidiomycetes that are wood decomposers, litter decomposers or mycorrhizal symbionts.
Myco/ogia. 85 (3):347-354.
TERMORSHUIZEN, A. J. Y A. P. SCHAFFERS. 1987. Ocurrence of carpophores of
ectomycorrhizal fungi in selected stand of Pinus sy/vestris in the Netherlands in relation to
stand vitality and air pollution. In: Plant and Soil. 104: 209-217.
UllOA, M. 1991. Diccionario ilustrado de micologfa. UNAM, D.F.
VALENZUElA, R., G. GUZMAN y J. CASTillO. 1981. Descripciones de especies de .
macromicetos poco conocidas en Mexico, con discusiones sobre su ecologfa y distribuci6n. Bo/.
Soc. Mex. Mic. 15: 67-120.
89

v ARELA, F. 1994. Para que conservar y para que . ln. Sociedad Mexicana de Micologia y Universidad de
Guanajuato. 5 Congreso Nacional de Micologia. 27 a 30 de noviembre de 1994. Programas y memorias,
Guanajuato.
VAZQUEZ-LOPEZ, G., LOPEZ, S. J. C. y CIFUENTES, B. J. 1994. Macromicetos del municipio de
Guanajuato. ln. Sociedad Mexicana de Micologia y Universidad de Guanajuato. 5 Congreso Nacional de
Micologia. 27 a 30 de noviembre de 1994. Prograrnas y memorias, Guanajuato.
VAZQUEZ-MENDOZA, S. y V ALENZUELA.GARZA. 1994. Estudio fungistico de los macromicetos de
Ia Sierra Norte del estado de Puebla. ln. Sociedad Mexicana de Micologia y Universidad de Guanajuato. 5
Congreso Nacional de Mico1ogia. 27 a 30 de noviembre de I 994. Program as y memorias, Guanajuato.
VILLARREAL-RUIZ, L. 1994. Analisis ecol6gico-silvfcola de Ia productividad natural de hongos
comestibles silvestres en los bosques del Cofre de Perote, Veracruz. Tesis de Maestrfa en
Ciencias. Colegio de Postgraduados, Montecillo, Mexico.
VILLARREAL, l. y J. PEREZ-MORENO. 1989. los hongos comestibles silvestres de Mexico,
un enfoque integral. Mico/. Neotrop. Ap/. 2: 77-114.
VILLARREAL-RUIZ, L. y F. CASTILLO-GONZALEZ. 1994. Proyecto: los hongos silvestres:
componentes de Ia biodiversidad y alternativa para Ia sustentabilidad de los bosques
templados de Mexico. CONABIO (C066). IREGEP. Colegio de Postgraduados. Montecillo, Edo
de Mexico.
VILLARREAL-RUIZ, L. (Ed.). 1996. los hongos silvestres: Componentes de Ia biodiversidad y
alternativa para Ia sustentabilidad de los bosques templados de Mexico. lnforme Final
Proyecto-CONABIO C066. lnstituto de Recursos Geneticos y Productividad. Colegio de
Postgraduados en Ciencias Agrfcolas, Montecillo, Edo.de Mexico.
VILLENEUVE, N., M. M. GRANDTNER, y J. A. FORTIN. 1989. Frequency and diversity of
ectomycorrhizal and saprophytic macrofungi in the laurentide Mountains of Quebec. Can. J.
Bot. 67: 2616- 2629.
90

VOGT, A. K., J. BLOOMFIELD, J. F. AMMIRATI and S. R. AMMIRATI. 1992. Sporocarp
production by Basidiomycetes, with emphasis on forest ecosystems. ln : Carroll , G. C. yD. T.
Wicklow (Eds.). The fungal community: its organization and role in thE( ecosystem (second
edition) . Marcel Dekkey. New York.
WASTERLUND, I. and INGELOT. 1981. Fru itbody production of larger fungi in some young
Swedish forest whit special reference to logging waste. Forest Eco/. Manage. 3: 269-294.
WESSELS, J. G. H. 1991. Fungal growth and development: a molecular perspective. In:
Hawksworth, D. L. (ed.) . Frontiers in mycology. Honorary and general lectures from the
Fourth International Mycological Congress, Regensburg. 1990. CAB. International, Gran
Bretafia.
WHITTAKER, R. H. 1972. Evolution and measurement of species diversity . Taxon . 21 (2/3):
213-251.
ZAMORA-MARTiNEZ, M.C. and C. NIETO DE PASCUAL-POLA. 1995. Natural production of
wild edible mushrooms in the southwestern rural territory of Mexico City, Mexico. Forest
Ecology and Management 72:13-20.
91

ANEXO 1
Terminos Basicos de Ia Tesis.
Carp6foro. Organa de los macrohongos visible a simple vista, con caraterfsticas fisicas propias
de Ia especie; formado por el micelio en Ia fase reproductiva para dispersar las estructuras
reproductoras .
Crecimiento Vegetative. Lapso en que el mi '3lio acumula reservas nutricionales, termina al
presentarse condiciones ambientales 6ptimas para Ia fructificaci6n.
Especies Abundantes. Son macrohongos que poseen porcentajes de Ia producci6n, estimada
en alguna medida cuantificable; superiores a Ia producci6n promedio.
Especies Dominantes. Especies que contribuyen con un mfnimo del 5% de Ia producci6n
estimada en cualquier medida cuantificable, como forma, cobertura o numero.
Espora. Estructura reproductiva de los macrohongos contenidas dentro de los carp6foros en
una capa llamada himenio, las esporas son como las "semillas" de los hongos.
Factores End6genos. Es Ia informacion genetica que contiene cada especie de macrohongos
que es Ia que dicta bajo que condiciones fructificar, tamaf\o y forma del carp6foro.
Factores Ex6genos. Son factores ambientales variables en su nivel por Ia sucesi6n climatica,
tales como: humedad, temperatura, luz, pH, densidad del suelo, entre otros.
Factores Silvlcolas. Son los factores bioticos con los cuales el micelio esta relacionado;
especies arboreas presentes, cobertura del dosel, forrajeo por microfauna, recolecci6n y
interacci6n con microorganismos.
Fase Reproductiva. Se inicia cuando se presentan condiciones ambientales 6ptimas, teniendo
como requisito que el micelio posea reservas nutricionales, se divide en dos partes: (a)
formaci6n de las estructuras reproductoras carp6foro, y (b) liberaci6n de esporas.
Fase Somf.tica. Etapa del ciclo de vida de los macromicetos que inicia con Ia germinaci6n de . las esporas o formaci6n de las hifas, las cuales en conjunto forman el micelio; este fase es de
92

nutrici6n, continua hasta que se presentan condiciones ambientales 6ptimas para Ia fase de
reproducci6n.
Fructificaci6n. Formaci6n de las estructuras de dispersion de las esporas, denominadas
carp6foros.
Hifa. Filamento microsc6pico originado por Ia germinaci6n de Ia espora; degrada el sustrato
organico en que se encuentra.
Hongos Comestibles. Carp6foros que poseen un alto valor alimenticio, por contener
aminoacidos, grasas, protefnas y azucares; libres de compuestos t6xicos que danan de alguna
forma el metabolismo del hombre.
Hongos Epigeos. Macromicetos que producen sus estructuras reproductoras sabre el suelo.
Hongos Hipogeos. Especies que producen estructuras reproductoras enterradas.
Hongos silvestres. Macrohongos presentes en Ia naturaleza.
Hongos sin Valor Alimenticio. Carp6foros que poseen bajo porcentaje de protefnas, grasas,
vitaminas y azucares o cuyo sabor es desagradable.
Hongos T6xicos. Especies que alteran el metabolismo del hombre, al contener sustancias
t6xicas.
Macrohongos. Hongos que poseen estructuras reproductoras macrosc6picas mas grandes de
un milimetro, visibles a simple vista.
Macrohongos Saprobios. Especies que degradan todo tipo de materia organica muerta, liberan
los nutrimentos inmovilizados y los dejan de forma asimilable para los veg'etales.
Macrohongos Simbiontes Antagonistas. Organismos que obtienen sus nutrimentos atacando
y matando los tejidos vivos donde se desarrollan.
93

Macrohongos Simbiontes Mutualistas. Especies que forman una relaci6n simbiotica con los
vegetales, ambos organismos reciben beneficios mutuos que contribuyen a su desarrollo y
sobrevivencia.
Micelio. Grupo de hifas que se han fusionado lateralmente y frontalmente, formando una masa
citoplasmatica que a simple vista tiene una apariencia algodonosa.
Micofagos. Pequeiios mamiferos como tuzas y conejos que basan su alimentaci6n en el
consumo de los carp6foros de los macrohongos.
Mic6filas. Personas que su vida gira alrededor de los hongos, ya sea estudiandolos o
utilizandolos de alguna forma.
Periodo de Fructificaci6n. Lapso en que se presenta Ia fructificaci6n de las especies de
macrohongos en una area determinada.
Recolecci6n. Toma y aprovechamiento del cuerpo reproductive de los macrohongos presentes en
Ia naturaleza.
Sustrato. Es el Iugar en donde se ha establecido el micelio de una especie de macromiceto,
anclandose, degradando y nutriendose.
Tasa de Oescomposici6n. Porcentaje de carp6foros que presentan algun signo de deterioro,
ocasionado por daiios ambientales, de organismos o como resultado de su propia madurez.
94
\
_ .. - .. ~
Lahoraturio de ltecursos <;cneticos en llungos
I.O CALIUAD: COORDENAUAS (GPS): N
ALTITUD: TIPO DE VEGETACION: 0
NO. UE CUADRANTE: NO. DE INVENTARIO: FECHA:
NO. DE NO. DE PESO FRESCO PESO FRESCO SOCIABILIDAD CLA~ E ESPECIE CARPOFOROS CARPOFOROS (gJ (gJ SUST .' GRU . OBSERVACIOHES
CARPO ORCS CARPO ORCS TROF.' IND. AGRUP. 1 COEF.ABUHD .' SAN OS AGUSAHADOS SAN OS AGUSANADOS
-
~ I . 3 :J Jj
:;)
~ :J :J :J :<::
I
:)
'.J :J :::;
Jj .. ~ ()
0 o= ·-0 r: -i
' < ..:l I'.. (
..
l a ·• r...i....i.:a;Lr=...•--"-""' -=-c= -- --==

Vl ell (_)
·oo 0
~ 0
§ .... (_)
"' E
~ ';:J Vl
·c: C)
u r: 5 C)
"0 0 .... -Vl
·c:n 0 .... (J
"0 ell '0' ;r:
i l"'i 0 X ~ z <
( : ( J I I (; I () l l L l l ( ) \ r (; V 4. l l IJ 4. l l () \ IN\IIIIJI() l l[ I)[CIJI.>\()\ CLNLriU)\ l' J)I){)J )(JCII\'IIlAI J
l.alHlfattll·lo de L)(.-.:un~ Ccnetlu:JS en liuuws Formato para toma de datos macromorfol6gicos de Boletales
Basado en Hernandez-Munoz ( 1995)
Nom. cientifico Colec1or No. _________ _ Localidad Coordenadas (GPS) __ -,-__ Allitud m s n.m. Vegeucion Asoc.-::--- --Suslnto _ ___ fecba No. de Foto ____ _ l'll.t:O : Tam.ano y forma _______ _ M .. gcu Superficie seca 0 humeda 0 cerosa 0 aceitos.t 0 viscosa 0 glutinos.t 0 Co lo • ---,--,-~- --------,----------------------Or namentacion (forma, tanuiio, color) _ _ ____ _
Ck ras ____________ _ ___________________ _
Conlexto: tamaiio nun Color (i_cambia?), __________ _ _ Consistcnca _______ _ _ __: ____________________ _
olor sabor _ _ --==-----=:-----=----TUBOS: umano union : libres 0 subadb. 0 adb. 0 decurr. 0 color: ______________ __________________ _
POROS: tamaiio (de uno solo o nUmero por mml), _ _ __ -::=-----------form.a: circLtl.ar 0 a.ogulares 0 bolc:tinoides 0 irregularesO colo~camb~?), ______________________________________ __ Ch ras. ___________________ ___ _______________ ___ ~---
l.STIPrn::unuiio superficie: seca 0 bumeda 0 cerosa tlaceitosa 0 viscosa 0 giLltinosa 0 posicion : : cent. 0 excent. 0 lat. 0 fomu. : cilin . 0 cllv. 0 subcllv.Oalcu.Oventric. 0 radic.O fi.stuloso 0 Uulbo redon.OcLiv. 0 obuc. 0 emarg. 0 bendi. (J GlANDUlAS 0 Color ____________________________________________________________ ___
Om amelltacion (fomu., unurlo, color)
Mlcelio (color):---c--:-------Contexto: consistencia _______ _______________ _ colo~~camba?) olor _________ _
sabor oLras_ --::::-- -;:::;:----;::;:--= ANLLLO 0 VI:LO 0 posicion: apic. 0 subapic. 0 cent. 0 inf .O mov. 0 cort . 0 tipo: fibril 0 cscam.O ~le 0 doble 0 colgante 0 subpcronado 0 peronado 0 Color _________________________________________________________ __
COLOR DE LA ESPORADA ___ :---:--::-::-.,---.,----,-PRUEBAS MACROQUfMICAS
KEACTIVO PARTE lJTlLIZADA REACCION (COLOR) TIEMPO
C{)Lt61{) ()t V () § T 6 VA U U A I ; {) § IN\ Ill I Jl () l >t ( ; [(:I JI)UJ\ (;1:"-1[ IU:()\ l' l i i .'( JI >I I( :11\'11 >AIl
I dbVf'alu-lo de llt.>e:UfS()S u~nellt:~ en IIUl&H •\ Formato para toma de datos macromorfol6gicos de Agaricales en general
Basado en Herruindez-Mui\oz ( 1995)
Nom. cieutifico --- -------Colector Localidad
--- ------- --- - No. ____ _ Coordenadas (GPS)
Ahitud _____ m s.n.m. Vegetacion Asoc. SuSirato Fecha No. de Foto Pll.£0: Tamaiio y fomu ___ _
Margen : ___ =--- ---.,-----Superficie: seca 0 bumeda 0 ceros.t 0 acei10sa 0 viscosa 0 glutinosa 0 lligr<ifano 0 Color _ _ ___ ______ _ _ _ ______ _ Omamcnucion (forma, tamaiio, color) ______ ____ ___________ _
CHras ____________ ~~---=~-----------Contexto : tam.aiio mm Col01ii.cambia?) consistencia olor sabor AMINAS: libres 0 subadberidas 0 adb. 0 sinu. 0 unci. 0 subdec. 0 - decu~. 0 -frecuencia : separadas 0 poco separadas 0 jw11as 0 muy juntas 0 forma : estrecba 0 ancba 0 ventric. 0 Borde _______ ---------Sabor Color: lateral _ _____ _______ _ borde ___ _ _ _______________ _
CHras~·~~-------------------~---------£STIPIT£:tamado superficie___,,_-----,,--- --=--posicion : : cent. 0 excent. 0 lat. 0 fomu. : filifO cilin. 0 clav. 0 subclav 0 aten.O
ventric. 0 radic.O Bulbo: redon.Ocl.av. 0 oblic. 0 em.ug. 0 bendi 0 Color Omamentacion (forma, tamado, color) ____________ --:---
Contexto: consistenca. _________________________ _
color~,---------=~-----------------------------------------------buecoO fistulo'so 0 olor _______________________ _:_
sabor otras'--=-----,=---::=----::=-ANILLO 0 VELO 0 posicion: apic. 0 subapic. 0 cent. 0 inf.O mov. 0 cortin. 0 tipo: fibrii.O cscam.O simple. 0 doble 0 colgante 0 subperonado 0 peronado 0 Color (del aniUo en ambos lados), ___ _ _________ ~----------COLOR D£ LA ESPORADA _____ _ ______________ _
PRliEBAS MACROQUIMICAS REACTIVO PARTE lJflLIZADA REACCION (COLOR) TlEMPO
---- ------

Vl
13 "@) 0
~ 0 E 0 ... u re E
~ ;;; ·c:: (J
u <'::
~ u
'"0 0 ... ;;; "t:I; (J .... u
'"0 <'C
"0' ::r: .:: I"')
0 ;.< ~ z. ..(
C()LC(31() ()L ()()~T6124UU4()()~ I~UiflJJ() ( ){ l!LUJVS()S CI)OilffiC()S Y VIJ()()I J4:JI\-'I()AU
1 ai.KJI"atul() de Ve.:unos €cneucos en ll()luws Formato para toma de datos macromorfo16gicos de Russula
Basado en Hern.indez-MunO:z (1995)
Nom cientifico Col <:c tor No. l.ocahd• d---------- ------ Coordenadas (GPSl_-:-_ --------Ahrtud m s.n.m. Vegetacion ________ _ A soc Su>trato Fecha _________ _ No. de Foto
PILlO: TanWio y fornu _______ -:-:-- --------------
Margen--==--- --=--Sup erfi .: ie : seca 0 burueda 0 cerosa 0 aceitosa 0 viscosa 0 glutinosa 0 Colu r __ -:-:---:--::----------,---,-------------- - -------Omamentacion (forma, tamaiio, color) ___ _________________ _
Ckr• s _______________________ ~:-:---------------------------------------Contexto : tam.ano mm Color{j,c.mbia?) ______________ _ Con>ist eucia _ _______________ _______ _ Cut icula _______ ____________ ____________ ___
u lor sabor _____ =-----==-------LAI\IINAS: subadb. 0 adb . 0 sinu. 0 unci. 0 sub<lecurt. 0 decurr. 0 frecueucia: sepuadlls 0 poco separadas 0 juntas 0 muy juntas 0 fornu estrecha 0 a.ncba 0 ventricosa 0 Borde. _______________ _
S• Lur Color (lateral y del borde), _____________ _
Otra s ~--·-----------------------c:-lS nrrrl:tamaiio superficie ______ _
posicion : CCIII. OexcC:nt. 0 lAteral 0 forma : cilin. 0 c~v. 0 subclav. 0 atC1l . 0 veutric . 0 radic. 0 Color _____ _ _ _____ ________ _
Onumentacioo (forma, tamailo, color). _____________________ _ Coute xto: coosistencu, _____________ _ _ __________ __ _ co lor ________________________________________________________________ _
olor _ _ _ __________ _ sobor __ otras. _ __________ ___
CO LOR 0 [ LA ESPORAOA:-:::-:-=c---:---,-----..,--:--=----------PRUEBAS MACROQUIMICAS
REACTIVO PARTE LJTILIZADA REACClON (COLOR) TIEMPO reS04 contexto cstipitc 1'<$04 laminas h no l contexto est ipite Feno l laminas KOtt contexto estipite f-. : l ll l;iminas An li in ii coutex1o est ipite An d111 a laminas
C {) I . l: ( ) I () f ) l () () S f ( ·. 1.! A l l IJ A l l () \ INUIJIJJ(J U : VC:U.!VSOS UM"IH.(Ji ,- I'I.>(X.t!C, JIVII ,.u;
I diU l"iiCH"II • de! Lli~HII"\4'1> • =~~nNIU>\ 1!11 I ~"11!1>\ Formato para toma de datos macromorfol6gicos de AmaniliJ
Basado en Hern.indez-Munoz(l'l95) Nom crmtifico Coloctor ----- No Loc.1hd.ad CoordaudaS(G~-------AJtitud m s n m Vcgcta66n ASOC ______ ___ _
Sustrato Focha No de Foto ____ _
PI LEO: Tamaiio y forma __ _ Margen liso 0 lig estriado D estnado D estnado-tuber Lfb31ida obscoCiln Superficie. seca 0 humeda 0 cerosa 0 aceitosa 0 vrsc.osa [J glutrnosa 0 Color Omar"nm= ta=o..,on- . .,..p.,..a-=-rch=e-n'"',-=-erc:n-cb-:-ra:-n:-o-:s-:-o'D...-co:--:s:-tra:-:-s-.D...-7fa:-n~n:-os:-:-:o•o.....-::-granuloso --crt.liriloso D escamas alniro-truncad.as 0 rorucas 0 anguJ . 0 pi ram 0 afilad.as 0 &rreg 0 (tanui>o, color) Otras Conte-~-o-ta-~~--------mmCo-l'o_r _ _ __________________ _
ol or sabor ______ _
LAMINAS: libres 0 subadhend.as 0 Color (borde) Sabor '--------------~C~o~lo-r~(l~at-~~~-y-d~cl~bo.-r'de~)~~~~--------------------
cnras, ____________________________________________________ __
ESTIPITE:umano superfioe forma filif Ocilin D clav 0 subdav 0 altfl 0 verllnc 0 radJc"lQ"R"fi:;SI-:-:Uir;:o:;:-so::TDr----Bulbo rcrlon .Odav 0 obhc 0 emarg 0 ht!ndi. 0 Color On~ma--~--o~oo~(~fu-m-~--.ta~n-un~--o.-co-.1o-r~) --------------------------------------
Contexto consistmcia color --------- --- -- ----------------
olor sabo_r_______________________________ -ou.5------------------------------
ANILLO 0 YELO 0 posrc .: aprc . Osubaprc. 0 CC11t 0 inf 0 mov 0 tJpo: fibrilOescarn O"mple 0 doble Ocolgarrte 0 subperon. 0 peron 0 Color (de arullo, ell ambos I ados), ________ _ ____ _______ ___
VOL VA: fugaz Ofarinosa 0 escam 0 fibril 0 zooada 0 constnc:uO saco membr ocread.a 0 acampanad.a 0 cotumada O .peronad.a 0 circunc 0 margm simple 0 doble 0 Unioo con cl estiprte solo ert Ia base 0 hasta parte media 0 hasta aprce bulb 0
COLOR DE LA ESPORAOA ________________ ___ _
REACTIVO KOH 5',. KOH5% FcS04
PRUEBAS MACROQUII\UCAS
PARTE LJflLIZADA qJICutiS
contexto 13minas
REACClON (COLOR) TIEMPO

Anexo 4. Datos climaticos.
A. Datos de precipitaci6n (mm) ENERO 30.5 FEBRERO 0 MARZO 13.4 ABRIL 14.5 MAYO 65.4 JUNIO 118.2 JULIO 91.4 AGOSTO 218.9 SEPTIEMBRE 58,5 OCTUBRE 65.4 NOVIEMBRE 61.3 DICIEMBRE 65.3
Anexo B. ENERO FEBRERO MARZO ABRIL MAYO JUNIO JULIO
Temperaturas estimadas (·c) 11.63
AGOSTO SEPTIEMBRE OCTUBRE NOVIEMBRE DICIEMBRE
11.86 13.06 14.58 15.05 12.72 12.27 12.20 12.6 12.32 11.8 10.26
Anexo C. ENERO FEBRERO MARZO ABRIL
Producci6n en carp6foros· totales 0
MAYO JUNIO JULIO AGOSTO SEPTIEMBRE OCTUBRE NOVIEMBRE DICIEMBRE
0 0 0 0 181 378 483 130 60 18 0
-. '.

ANEXO 5. Macromicetos comestibles aprcciados.
Del estudio se presentan las especies comestibles apreciadas para su venta por los recolectores
f
I i ~ ~
.I
I '
f ~ ..
I .~· 4 • •
f .. ) .,.,; I ( ~ .. '),
I ,
2. Gomphus jloccosus
3.Helvella crispa
4. Morchella escu/enta
5. Ram aria jlava

Anexo de Ia investigaci6n
-GP INS:~,~:~EIN~NA~Z~ E :,~~:~~E~C~~~~~~~LAS INSTITUTO DE RECURSOS GENETICOS Y PRODUCTIVIDAD
PROGRAMA DE GENETICA
Bioi. Jorge Gonzalez Loera Coordinador' General del P.I.D.I.S.A. de Ia UACH PRESENTE
En atenci6n a su amable carte de mayo 29 de 1995, en Ia cual solicita mi colaboraci6n para dirigir Ia tes is de licenciatura del C. Jaime Gonzalez Cabrera, estudiante del sexto ario (segundo semestre) de Ia carrara de lngeniero en Agroecologia , le informo que acepto fungir como director de tesis .
Par tal motive le comunico que estara adscrito al proyecto CONABIO C066 , comprometiendome a sufragar los gastds de pasaje al area de estudio, asi como brindar las facilidades necesarias para Ia impresi6n final de Ia tesis y el page del tiraje de Ia misma. de acuerdo con los requisites minimos de titulaci6n solicitados par Ia UACH. Es conveniente aclarar que par razones administrativas , el page del tiraje de Ia tesis tendra una vigencia de 12 meses. a partir del primero de junio.
Sabre Ia solicitud para que el C. Gonzalez-Cabrera desarrolle su Servicio Social en el Laboratorio de Recursos Geneticos Fung icos del Programa de Genet ica del IREGEP , le comunico que no existe ningun inconveniete al respecto . par lo que en breve se haran los tramites necesarios para su formal izaci6n .
Sin otro particular me despido con un cord ial saluda.
M. e Ruiz Invest Adjunto
Laboratorio de Recursos Geneticos Fung icos
dei iREGEP
c. p. Dr. Manuel U vera Murioz. Di rector del IREGEP. c.p .. Dr. Jorge Rodrig uez Alcazar, Coordinador de Ens erianz.a del IR EG EP. c.p. Dr. Jo se Moli na Galan . Vocal Doc ente del Prog rama de Genetica del IREGEP .
;:;,;c .p. Jaime Gonzalez Cabrera . ~studiante del PI D. I SA, de Ia UACH.
C:metera Me:Uco-Texcoco .Km 35.5 56230 Morut:e illo, Texcoco, Ed.o. de Mexico Td. y Fax (595) 45265; Fu (595) 45713




















