
The impact of distributional metrics in the quality of relational triples

DISTRIBUTIONAL RECORDS OF SHREWS (MAMMALIA, SORICOMORPHA, SORICIDAE)FROM NORTHERN CENTRAL AMERICA
WITH THE FIRST RECORD OF SOREX FROM HONDURAS
NEAL WOODMAN[Research Associate, Section of Mammals, Carnegie Museum of Natural History]
USGS Patuxent Wildlife Research Center, National Museum of Natural History, Smithsonian Institution, P.O. Box 37012, Washington, D.C. 20013-7012
JOHN O. MATSONDepartment of Biological Sciences
San Jose State University, One Washington Square, San Jose, California 95192-0100 [email protected]
TIMOTHy J. MCCARTHy*Scientific Preparator, Section of Mammals, Carnegie Museum of Natural History,
5800 Baum Boulevard, Pittsburgh, Pennsylvania 15206-3783
RALPH P. ECkERLIN[Research Associate, Section of Invertebrate Zoology, Carnegie Museum of Natural History]
Natural Sciences Division, Northern Virginia Community College, Annandale, Virginia [email protected]
WALTER BULMERNatural Sciences Division, Northern Virginia Community College, Annandale, Virginia 22003
NICTé ORDóñEz-GARzADepartment of Biological Sciences, Texas Tech University, Lubbock, Texas 79409-3131
*deceased, January 7, 2011
A N N A L S O F C A R N E G I E M U S E U MVOL. 80, NUMBER 3, PP. 207–237 15 JULy 2012
Twelve species of shrews (Soricomorpha, Soricidae) are currently known from northern Central American coun-tries of Guatemala and Honduras. This group includes two species of Sorex Linnaeus, 1758 (S. salvini Merriam, 1897; S. veraepacis Alston, 1877), a genus that was previ-ously known only as far south as central Guatemala (Junge and Hoffmann 1981; Matson and McCarthy 2005), and ten species of Cryptotis Pomel, 1848 [C. goodwini Jack-son, 1933; C. hondurensis Woodman and Timm, 1992; C. lacertosus Woodman, 2010; C. magnimanus Woodman
and Timm, 1999; C. mam Woodman, 2010; C. mayensis (Merriam, 1901); C. merriami Choate, 1970; C. oreoryc-tes Woodman, 2011a; C. orophilus (J.A. Allen, 1895); C. tropicalis (Merriam, 1895)], a genus whose distribution continues into the northern Andean highlands of South America (Choate 1970; Woodman 2002, 2003, 2010, 2011a; Woodman and Timm 1992, 1993, 1999; Woodman and Croft 2005). With the notable exception of C. may-ensis, most species of soricids in this region are typically confined to moist habitats at middle and upper elevations.
ABSTRACT
Short term surveys for small mammals in Guatemala and Honduras during 1992–2009 provided important new records for 12 taxa of shrews from 24 localities. These locality records expand the known geographic distributions for five species and for the genus Sorex Linnaeus, 1758: the geographic range of Cryptotis goodwini Jackson, 1933, now includes the Sierra de las Minas, Guatemala, and several isolated highlands in western Honduras; the known distribution of Cryptotis mayensis (Merriam, 1901) is increased with the first definite modern record for this shrew from Guatemala; Cryptotis merriami Choate, 1970, is now known to occur in the Sierra de las Minas and the Sierra del Merendón, Guatemala, as well as the isolated Sierra de Omoa and Montaña de La Muralla in Honduras, and its documented elevational range (600–1720 m) is expanded; records of Sorex veraepacis Alston, 1877, expand the known distribution of this species to include the Sierra de Yalijux, Guatemala; and discovery of Sorex salvini Merriam, 1897, at Celaque, Honduras (1825–3110 m), represents a considerable extension of the geographic range of the species, and it is the first record of the genus Sorex from Honduras. In addition, the first record of potential syntopy among C. goodwini, C. merriami, and Cryptotis orophilus (J.A. Allen, 1895), is reported at an elevation of 1430 m in the Sierra de Celaque, Honduras. Information associated with these records contributes substantially to knowledge of habitat use, elevational distributions, reproductive patterns, diet, and parasites of the species encountered. General patterns include the first evidence that Neotropical species of soricids have smaller litters than their temperate congeners.
kEy WORDS: food habits, Insectivora, parasites, pit fall, reproduction, shrews, trap success
INTRODUCTION

208 ANNALS OF CARNEGIE MUSEUM VOL. 80
Relative to many other small mammals, shrews can be notoriously difficult to capture using standard manufac-tured traps (Kirkland and Sheppard 1994). They are often under-represented or even lacking from short-term sur-veys upon which many of the known distributions of small mammals are based, particularly in understudied parts of the world such as Central America. The resulting paucity of specimens of shrews from northern Central America has seriously limited our understanding of species limits and distributions (Choate 1970; Hall 1981; Woodman and Timm 1992, 1993, 1999), which in turn inhibits our abil-ity to determine diversification and biogeographical pat-terns (Woodman 2005). Shrews are particularly important as members of high elevation small mammal faunas, yet we are only beginning to understand patterns of evolution and endemism exhibited by these animals. Documentation of shrews and the faunas in which they occur is especially important as the habitats where these animals live face the threat of potential extinction as a result of global warming (Colwell et al. 2008; McCain and Colwell 2011), as well as the traditional threats of encroachment by humans, live-stock, and invasive organisms (Fig. 1). In recent years (1992–2009), we have been involved in a series of research expeditions in Guatemala and Hondu-ras during which we employed pitfall traps in addition to standard manufactured traps to survey small mammals. Al-though the use of pitfall traps to obtain small mammals, espe-cially shrews, is not new (e.g., Buckner 1966), they remain underutilized by field mammalogists. Our methodology
increased captures of shrews relative to our earlier field experiences using traditional small mammal traps, a result that is clearly documented by relative capture rates. In our study of shrews from Central America, we also discovered two relevant, but previously unreported, specimens in nat-ural history collections. Based on the entirety of this work, we herein report on new records for ten species of shrews from this region. The new localities extend the known geo-graphical distributions for five of these species and include the first record of the genus Sorex from Honduras and the first modern record of C. mayensis from Guatemala. We provide information on reproduction, diet, and habitat for the species we encountered.
METHODS
During field inventories of small mammals in middle and upper montane humid forests in Guatemala and Hondu-ras during 1992–2009, we employed varying numbers of small, plastic pitfall containers (ca. 20 cm deep, 15 cm top inside diameter) in addition to combinations of live traps (Sherman LFA 3 x 3.5 x 9” traps) and snap traps (Museum Special mouse traps and Victor-style rat traps) to capture small mammals. Pitfalls generally were used without drift fencing. At one site (Chelemhá in 2007), we also unsuc-cessfully employed five one-gallon buckets with drift-fencing in pitfall arrays. All traps were checked twice per day. We use the term “trapnight” herein to refer to a single 24-hour period during which a trap or pitfall was open.
Fig. 1.—Landscape in the Sierra de Yalijux, Guatemala, illustrating a common pattern of deforestation of highland slopes with remnant patches of forest on the hilltops (photograph: NOG).

2012 WOODMAN ET AL.—DISTRIBUTIONAL RECORDS OF SHREWS 209
Voucher specimens were prepared as skins with skel-etons or as fluids, some with skulls removed and cleaned. Standard field measurements (with abbreviations in pa-rentheses) included: total length (TL), length of tail (LT), length of hind foot (HF), length of ear from notch (LE), and weight (WT). Length of head and body (HB) was sub-sequently calculated by subtracting LT from TL. These
measurements are presented in Table 1. For embryos, crown-to-rump length (CR) was recorded. All specimen measurements we report are in mm; weight is in g. Tis-sue samples and ectoparasites were preserved from most specimens. Common names used for shrews follow recent attempts to standardize the common names of mammals (Wilson and Cole 2000; Hutterer 2005; Reid 2009).
Table 1. Selected measurements of shrews from Guatemala and Honduras. Measurements are mean ± SD and range,with sample size in parentheses. Length measurements in millimeters and weight in grams.
Length of Head & Body
Length of Tail Tail as % ofHead & Body
Length of Hind Foot
Weight CondylobasalLength of Skull
Cryptotis goodwini 78 ± 7 26 ± 3 34 ± 4 14 ± 1 10.2 ± 2.4 20.0 ± 0.5
67–96 21–32 27–42 12–19 7.0–17.0 19.1–21.2
(n = 31) (n = 31) (n = 31) (n = 31) (n = 36) (n = 31)
Cryptotis lacertosus 82 ± 4 28 ± 2 34 ± 4 14 ± 1 14.7 ± 2.7 21.5 ± 0.6
75–87 24–30 28–40 12–15 9.8–16.5 20.8–22.8
(n = 8) (n = 8) (n = 8) (n = 8) (n = 7) (n = 8)
Cryptotis mam 75 ± 4 29 ± 2 38 ± 3 14 ± 1 8.4 ± 1.2 19.9 ± 0.4
64–81 22–32 33–44 11–16 6.5–10.5 18.8–20.4
(n = 31) (n = 31) (n = 31) (n = 31) (n = 12) (n = 22)
Cryptotis mayensis 69 20 48 10 – 19.0
(n = 1) (n = 1) (n = 1) (n = 1) (n = 1)
Cryptotis merriami 68 ± 6 27 ± 3 40 ± 4 12 ± 1 6.8 ± 0.8 19.6 ± 0.5
59–77 21–32 30–46 8–14 5.2–8.5 18.8–20.9
(n = 20) (n = 20) (n = 20) (n = 20) (n = 27) (n = 25)
Cryptotis oreoryctes 82 ± 5 30 ± 2 36 ± 4 15 ± 1 13.7 ± 2.5 21.1 ± 0.4
71–90 27–32 31–45 14–16 9.5–16.5 20.2–21.6
(n = 15) (n = 15) (n = 15) (n = 15) (n = 15) (n = 10)
Cryptotis orophila 67 23 34 12 5.6 17.3
(n = 1) (n = 1) (n = 1) (n = 1) (n = 1) (n = 1)
Cryptotis tropicalis 66 21 32 11 6.0 17.0
(n = 1) (n = 1) (n = 1) (n = 1) (n = 1) (n = 1)
Sorex salvini 63 ± 4 53 ± 1 84 ± 5 14 ± 1 5.3 ± 0.9 17.5, 18.5
60–68 52–54 78–89 13–16 4.2–6.5
(n = 4) (n = 4) (n = 4) (n = 4) (n = 4) (n = 2)
Sorex veraepacis chiapensis 68 ± 3 48 ± 3 71 ± 4 13 ± 1 6.8 ± 0.8 18.6 ± 0.5
64–41 45–54 66–80 12–14 5.5–8.0 17.9–19.4
(n = 10) (n = 10) (n = 10) (n = 10) (n = 10) (n = 9)
Sorex veraepacis ibarrai 75 ± 6 58 ± 4 78 ± 8 16 ± 1 9.5 ± 2.2 20.3 ± 0.4
59–89 38–68 43–101 10–21 5.3–14.7 19.5–21.1
(n = 125) (n = 125) (n = 125) (n = 115) (n = 125) (n = 21)
Sorex veraepacis veraepacis 72 ± 5 50 ± 3 70 ± 9 15 ± 1 7.4 ± 1.4 19.4 ± 0.5
58–80 42–58 57–97 13–16 6.0–11.0 18.5–20.0
(n = 30) (n = 30) (n = 30) (n = 30) (n = 16) (n = 13)
210 ANNALS OF CARNEGIE MUSEUM VOL. 80
Digestive tracts of a small number of individuals were preserved in 75% ethanol in the field for later study. One of us (RPE) subsequently made a complete inspection of the contents of the stomach and intestines. Although most prey remains were finely diced, resulting in a fine mush of mostly unidentifiable bits, a few remains retained di-agnostic characters, and these were identified to the nar-rowest taxonomic group possible for insight into the diets of Central American soricids. In addition, RPE collected and recorded endoparasites and ectoparasites from a ma-jority of individuals. Publications on parasites collected from shrews reported in this paper include Duszynski et al. (2003), Lewis and Eckerlin (2004), Eckerlin (2006), and Keirans and Eckerlin (2005). Capitalized vegetational associations are from the Hold-ridge classification of life zones (Holdridge 1947; MAGA 2001). Following the misguided ruling of the ICZN (2006), the genus Cryptotis is herein treated as masculine. Speci-mens referred to in this paper are deposited in the follow-ing systematic collections (abbreviations in parentheses): Natural History Museum, London (BMNH); Carnegie Museum of Natural History, Pittsburgh (CM); Florida Mu-seum of Natural History, Gainesville (FLMNH); Universi-ty of Michigan Museum of Zoology, Ann Arbor (UMMZ); Museo de la Fauna, Universidad Nacional Autónoma de Honduras, Tegucigalpa (UNAH); Museo de Historia Natu-ral, Universidad de San Carlos de Guatemala, Guatemala City (USAC); and National Museum of Natural History,
Washington (USNM). Uncataloged specimens are referred to by the field numbers of the collectors: John O. Matson (jom); Nicté Ordonez-Garza (nog); Nils Santos (ns); Ser-gio G. Pérez (sgp); and Timothy J. McCarthy (Tjm).
TRAPPING RESULTS
We captured a total of 280 individual soricids in pitfalls, Museum Special traps, and Sherman traps at 24 localities during field work in Guatemala and Honduras from 1992 through 2009. Most sites we visited had not previously been intensively surveyed for small mammals. Capture success of shrews varied throughout our field studies. A sample from our work, however, provides an example of the utility of using pitfalls in diversity surveys of small mammal communities or when targeting soricids in the Neotropics. In one subset of 23,882 trap-nights, we cap-tured 147 shrews. Capture rates varied by trap type, with pitfalls taking more than twice the number of shrews (np = 103 captures; 70% of captures) as Museum Specials (nms = 38; 26%) and many more than Sherman traps (nsh = 6; 4%). No shrews were taken in rat traps, which consequent-ly are not included in the total number of trapnights. When the trap effort (E = number of trapnights) by trap type is taken into account, trap success (t) was much higher for pitfalls [tp = np / Ep x 100 = 3%; where Ep = 3221 trap-nights (13% of total trapnights)] than for either Museum Specials (tms < 1%; Ems = 13,349 (56%)] or Shermans (tsh
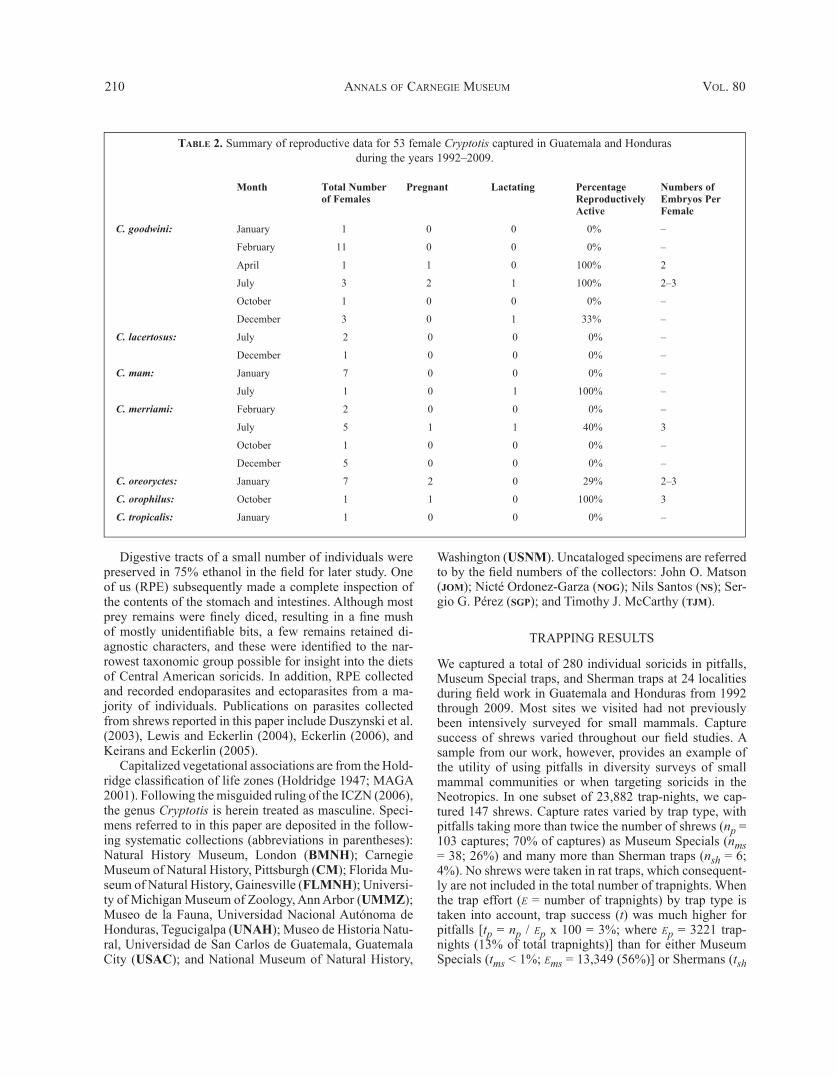
Table 2. Summary of reproductive data for 53 female Cryptotis captured in Guatemala and Hondurasduring the years 1992–2009.
Month Total Number of Females
Pregnant Lactating Percentage Reproductively Active
Numbers of Embryos Per Female
C. goodwini: January 1 0 0 0% –
February 11 0 0 0% –
April 1 1 0 100% 2
July 3 2 1 100% 2–3
October 1 0 0 0% –
December 3 0 1 33% –
C. lacertosus: July 2 0 0 0% –
December 1 0 0 0% –
C. mam: January 7 0 0 0% –
July 1 0 1 100% –
C. merriami: February 2 0 0 0% –
July 5 1 1 40% 3
October 1 0 0 0% –
December 5 0 0 0% –
C. oreoryctes: January 7 2 0 29% 2–3
C. orophilus: October 1 1 0 100% 3
C. tropicalis: January 1 0 0 0% –

2012 WOODMAN ET AL.—DISTRIBUTIONAL RECORDS OF SHREWS 211
<< 1%; Esh = 7312 (31%)]. The greater success of pitfalls over snap-traps, however, is not guaranteed in all situa-tions. Six shrews we captured during 13 nights along the damp banks of a small stream at Campamento Don Tomás, Celaque, Honduras, in February, 1998, for example, were all taken in Museum Specials, and none was captured in a pitfall (2983 total trapnights: 1773 Museum Specials, 627 rat traps, 366 Shermans, 217 pitfalls). Pitfalls are not ap-propriate for all situations, such as areas with very shallow soils or along rock faces, and they generally require more time and effort to set up or move than more traditional trap types. Our relatively shallow pitfall traps typically captured only shrews and generally are not useful for ob-taining statistically meaningful samples of rodents or mar-supials. Despite these potential drawbacks, shrews remain poorly represented in systematic collections, and pitfalls are extremely useful for obtaining these animals in many common situations. In general, we had greatest success in capturing shrews when traps were placed in moister areas, such as the wet, sandy loam of river floodplains; damp stream-sides; poorly drained soils where there were abundant decaying tree falls; and shaded, mossy hillsides. Well-drained soils tended to produce few captures of shrews. Weather also played a role. At Campamento Naranjo, 2560 m, on Cerro Celaque, we collected nine individuals of two species (C. goodwini, S. salvini) in January and February, 1995, dur-ing typically cool, wet weather. In contrast, we captured no shrews at that site in February, 1998, during a regional drought resulting from a particularly strong El Niño event (Trenberth et al. 2002), despite similar trapping efforts (315 vs. 323 trapnights, respectively).
SPECIES ACCOUNTS
Cryptotis goodwini Jackson, 1933
Goodwin’s Broad-clawed Shrew was previously known from 41 specimens distributed among 18 highland locali-ties above ca. 1200 m in the Sierra Madre del Sur range of southern Chiapas, Mexico, to western Honduras. Although well documented in Guatemala, C. goodwini had been re-ported from only a single locality each in El Salvador and Honduras, and two localities in Chiapas (Hutterer 1980; Woodman and Timm 1999). Recent studies suggest that C. goodwini will prove to be a complex of several closely re-lated, cryptic species. Two of these species (C. lacertosus, C. oreoryctes) have already been recognized and described (Woodman 2010, 2011a). Pleistocene fossils of C. good-wini were reported from near Copán, Honduras (Woodman and Croft 2005). Our field work has provided 37 additional specimens from five localities. These records significantly expand the known distribution of C. goodwini into the Sierra de las Minas and the Sierra del Merendón in Guatemala, as well as the isolated Sierra de Omoa and Sierra de Celaque in western Honduras. In addition, a previously unreported
specimen (FLMNH 27718) from Parque Nacional de San-ta Barbara, Honduras, further expands the distribution of the species to the Montaña de Santa Barbara. The eleva-tional range of our specimens is 1620–2560 m, which is within the 1200–3355 m range reported by Woodman and Timm (1999). All of our specimens were taken in moist ar-eas along streams or in heavily wooded montane cloud for-est with deep leaf litter. Mosses and liverworts commonly were associated with traps where we caught this species. Among 20 females captured in January, February, April, July, October, and December, five exhibited evidence of recent reproduction (Table 2). A female taken 6 km NNW of San Lorenzo, Guatemala, on 15 April 1998, was preg-nant with two embryos (CR = 15). Among three females captured 10–12 July 2004 at Trifinio, Guatemala, one was lactating, and two were pregnant, one with two embryos (CR = 13) and one with three embryos (CR = 7). A female obtained on 11 December 1994 at Cusuco, Honduras, was lactating. Oocysts of the coccidian parasite Eimeria hondurensis Duszynski et al., 2003 (Apicomplexa, Coccidia, Eimeri-idae) were recovered from the intestinal tracts of C. good-wini from Cusuco and Celaque, Honduras, and from the female captured at 6 km NNW of San Lorenzo. Overall in-fection rate by this parasite was 73% of individuals tested. Oocysts of a second species, Eimeria whitakeri Upton and McAllister, 1991, were recovered from C. goodwini from Celaque and from the individual from 6 km NNW of San Lorenzo, with an overall infection rate of 50%; 18% of the tested shrews from these three localities hosted both parasites (Duszynski et al. 2003). A single female of the rare tick species Ixodes guatemalensis Kohls, 1956 (Acari, Ixodida, Ixodidae) was recovered from a C. goodwini from along the Camino a las Torres, Cerro Pinalón, Guatemala (Keirans and Eckerlin 2005).
New records (38).—GUATEMALA: Cerro Pinalón, 2700 m (CM 113275); 6 km NNW of San Lorenzo, 2200 m (CM 113276; USAC [SGP 500]); Trifinio, 2400 m (3 USAC [NOG 693, 720, 739]). HONDURAS: Cusuco, 1500–1600 m (CM 119693–119709); Celaque, 1430 m (CM 118519), 2075 m (CM 112878–112882), 2560 m (CM 119710–119716; 1 UNAH [JOM 6576]); above San José de los An-des, ca. 1900 m (FLMNH 27718) [Gazetteer localities 16, 17, 19, 23, 24, 25].
Cryptotis lacertosus Woodman, 2010
The recently described Muscular Broad-clawed Shrew is only known to occur in Subtropical Montane Wet Forest in the Sierra de los Cuchumatanes, western Guatemala. Its documented elevational distribution is 2680–3110 m (Woodman 2010). Our field work provided six specimens from two localities. Specimens from 5 km SW of San Ma-teo Ixtatán, the type locality for this species, were taken in a relatively closed-canopy cloud forest dominated by oaks (Quercus spp., Fagaceae), pines (Pinus spp., Pinaceae),

212 ANNALS OF CARNEGIE MUSEUM VOL. 80
and firs (Abies guatemalensis Rehder, Pinaceae), and hav-ing abundant mosses and downed trees. Reproductive bi-ology of C. lacertosus is unknown. None of three females captured in July and December 2005 was pregnant or oth-erwise showed evidence of reproductive activity (Table 2).
New records (6).—GUATEMALA: 5 km SW of San Ma-teo Ixtatán, 3110 m (USNM 569420, 569431, 569442, 569443, 569503); Yaiquich, 2680 m (USNM 569368) [Gazetteer localities 3, 4].
Cryptotis mam Woodman, 2010Fig. 2
The Mam Shrew, named for a Mayan indigenous group, occurs in mixed conifer forest above 2500 m in the Sierra de los Cuchumatanes, western Guatemala. The known ele-vational distribution is 2895–3350 m. Specimens from To-dos Santos Cuchumatán, previously referred to Jackson’s Broad-clawed Shrew, Cryptotis griseoventris Jackson, 1933, are now recognized as belonging to C. mam. The distribution of C. griseoventris (sensu stricto) is restricted to the Altiplanicie Central de Chiapas, Mexico (Woodman 2010). Our field work provided 12 specimens of C. mam from three localities. At Aldea el Rancho, we worked along streams that began along the base of a nearly vertical lime-stone escarpment. Dominant vegetation was conifers. In the Laguna Magdalena Reserve, our traplines were situ-ated along a fast-moving stream in a narrow limestone valley with a step-like series of falls. The stream entered a small pond (Laguna Magdalena) before pouring over a limestone bank and plunging down a vertical escarpment. The tops of the surrounding hills were covered with co-nifers, although grass and low vegetation dominated the lower hillsides and flat valley bottom. A number of shrews and mice were taken on or behind the heavy moss covering the moist, vertical faces of the falls, particularly adjacent to the pond. Trees at Laguna Magdalena and Aldea el Rancho
were gradually being removed for firewood and building materials, and both localities were heavily over-grazed by sheep. The number of buildings along the road at the entrance to the Laguna Magdalena Reserve increased dra-matically between our two visits, and, by 2008, grazing in the reserve had reduced the grass cover to stubble, re-ducing the diversity and numbers of small mammals we captured. Goldman (1951:295) noted similar human im-pacts on habitats near Todos Santos Cuchumatán as early as 1895–1896, and small mammal communities appear to have changed in composition, with reduced numbers and diversity since then (Woodman 2010). Reproductive biology of C. mam is mostly unknown. None of seven females captured in January was pregnant. A single female captured in July was lactating (Table 2). The lactating female was larger in both head-and-body length (81 mm) and mass (10.5 g) than any of the females (HB = 71 ± 2, 67–73; WT = 8.3 ± 1.0, 6.5–9.5, n = 7) or males (HB = 72 ± 6, 64–78; WT = 8.1 ± 1.3, 7.0–9.5, n = 4) captured in late December and January. Study of contents of stomachs and intestinal tracts of two females from Laguna Magdalena (USNM 569554, 570337) and two females from Aldea el Rancho (USNM 570313, 570314), captured in January, indicate that earth-worms (Oligochaeta) may be an important component of the diet of C. mam. Parts of earthworms and small amounts of plant matter were recovered from digestive tracts of all four individuals, and beetle (Coleoptera) fragments and seed coats were each found in one (Woodman 2010). Di-gestive tracts of two of these individuals appeared to be free of endoparasites. One, however, contained active tapeworms (Platyhelminthes, Cestoda), and another con-tained hymenolepid tapeworm eggs.
New records (12).—GUATEMALA: Puerto del Cielo, 3350 m (USNM 570248); Laguna Magdalena, 2925 m (USNM 569554, 569555, 570337, 570340); Aldea el Rancho, 3020 m (3 USAC [NOG 771, 804, 805]; USNM 570256, 570257, 570313, 570314) [Gazetteer localities 5, 6, 7].
Cryptotis mayensis (Merriam, 1901)
With the exception of an anomalous record from owl pel-lets in Guerrero, Mexico, the Mayan Shrew is restricted to the Yucatán Peninsula region of Mexico, Belize, and Gua-temala (Woodman and Timm 1993). Pleistocene fossils of C. mayensis are known from caves in Yucatán, Mexico (Woodman 1995). The only previous record of C. may-ensis from Guatemala is based on a collection of skeletal remains representing at least 104 individuals of unknown archeological age that were discovered in a Mayan vase in the ruins of Uaxactún (Murie 1935). A female C. mayensis (USAC 89) was captured 8 Au-gust 1988 by Enio B. Cano at Biotopo Cerro Cahuí, Petén. Cerro Cahuí is located along the northeast shore of Lago Petén-Itzá and elevations near here extend from above 100
Fig. 2.—Cryptotis mam [USAC (NOG 804)] from Aldea el Rancho, Gua-temala (photograph: JOM).

2012 WOODMAN ET AL.—DISTRIBUTIONAL RECORDS OF SHREWS 213
m to just above 350 m. This specimen represents the first undoubtedly modern specimen of the Mayan Shrew from Guatemala, and it represents the southwestern limit of the species’ known distribution.
New record (1).—GUATEMALA: Cerro Cahuí (USAC 89) [Gazetteer Locality 1].
Cryptotis merriami Choate, 1970
Merriam’s Shrew previously was known from 42 speci-mens from 17 localities distributed through highlands at elevations ranging from ca. 900 m to 1650 m in the Alti-planicie Central de Chiapas, Mexico, to northern Nicara-gua, with disjunct populations in the Tilarán Highlands of northern Costa Rica (Woodman and Timm 1993; Wood-man 2000). In Chiapas, C. merriami is represented only by skeletal remains from owl pellets (Choate 1970; Woodman and Timm 1993). Pleistocene fossils of this species were reported from caves near Copán, Honduras (Woodman and Croft 2005). Our field work resulted in an additional 29 specimens from seven localities in Guatemala and three localities in Honduras. These records include specimens from Sierra de las Minas, Sierra del Merendón, and Cerro Seja in the Si-erra de Santa Cruz. Specimens from Trifinio, near the base of Cerro Montecristo, amplify the known distribution of this species into the southeastern mountains of Guatemala. Our new records in Honduras extend its distribution to the isolated Sierra de Omoa and Montaña de La Muralla. We captured C. merriami at localities that range in elevation from 600 to 1750 m, thereby expanding both the lower and upper elevational limits documented for this species. All of our specimens were found in direct association with humid vegetation at mid- to high-elevations. At 1620 m at Cusuco, Honduras, C. merriami and C. goodwini were relatively abundant and sympatric. The only other locality where we recorded these two species together is at the Visitors Center for Celaque National Park, Honduras. In Guatemala, a single C. merriami (UMMZ 117845) pre-viously was captured in 1970 at Finca Concepción. Dur-ing a short stay there in 2007, we captured a male and a female in a small patch of secondary forest surrounded by cornfields. A second pair was taken in a large patch of dis-turbed hardwood forest bordering pasture on a hill behind Hotel Country Delights, Guatemala, and a third pair in sec-ondary growth in a former coffee plantation at El Limo, Guatemala. These records suggest some tolerance for habit disturbance and an ability to survive in small patches of disturbed and secondary forest. Of 13 females from the months of February, July, Oc-tober, and December, only two showed signs of recent reproductive activity (Table 2). A pregnant female carry-ing three embryos (CR = 10 mm) was captured on 20 July 2007 at Hotel Country Delights (USNM 570132), and a lactating female was recorded 11 July 1994 at Cerro Pozo de Agua, Guatemala (CM 119078). Sample sizes are low
for this species, but females average nearly the same size in July (HB = 69 ± 4, 63–73; WT = 7.3 ± 0.4, 7.0–8.0, n = 5) and December (HB = 66 ± 7, 61–74, n = 3; WT = 7.0 ± 0.5, 6.2–7.5, n = 5). Males likewise exhibit little variation in size between July (HB = 69 ± 8, 60–74; WT = 6.5 ± 0.5, 6.0–7.0, n = 3) and December (HB = 67 ± 4, 62–72, n = 5; WT = 6.6 ± 0.7, 5.2–7.2, n = 8), although males average lower in mass than females from the same month. Analysis of the contents of digestive tracts of three in-dividuals from Finca Concepción (USNM 570112), Hotel Country Delights (USNM 570132), and El Limo (USNM 570049) yielded only one with identifiable remains. The individual from El Limo contained tracheal tubes and frag-ments of a chitinous exoskeleton from a large insect. The shrew from Hotel Country Delights contained mammal hairs, probably from self-grooming. The digestive tracts of two of these individuals yielded no obvious endoparasites, but the third contained a roundworm (Nematoda). In addi-tion, oocysts of the coccidian parasite Eimeria honduren-sis were recovered from intestinal tracts of individual C. merriami from Cusuco and from Celaque (Duszynski et al. 2003).
New records (29).—GUATEMALA: Finca Concepción, 1290 m (USNM 570108, 570112); Hotel Country Delights, 1640–1665 m (USNM 570122, 570132); El Limo, 1475 m (USNM 570049, 570125); El Trifinio, 1650 m, 1750 m (2 USAC [NGO 666, 667]); Cerro Seja, 1020 m (CM 119079); Río Bobos, 600 m (CM 118752); Cerro Pozo de Agua, 1200 m (CM 119078). HONDURAS: Cusuco, 1620 m (CM 119717–119731); Celaque, 1430 m (CM 112883, 118520); Sendero el Pizote, 1480 m (CM 118521) [Gazet-teer localities 14, 15, 18, 19, 20, 21, 22, 23, 24, 26].
Cryptotis oreoryctes Woodman, 2011a
The recently described Yalijux Shrew occurs above 2000 m in Subtropical Montane Rain Forest in the Sierra de Yalijux, central Guatemala. Our field work yielded 15 individuals of this species from two localities. The only other known specimens of this shrew are four individuals collected in the nineteenth century near Cobán (Woodman 2011b). Those obtained in January 2007 at the Chelemhá Cloud Forest Re-serve were captured on a steep, moist, north-facing slope with abundant downed trees and mosses in a cloud forest dominated by oaks and pines (Woodman 2011a). Reproductive biology is poorly documented. Among seven females captured in January, two were pregnant, one with two embryos (CM 120099) and one (with obviously-worn dentition—USNM 569878) with three embryos (Ta-ble 2). Testes of males captured in January ranged in size from 2.5 x 2 mm to 10 x 6.5 mm.
New records (15).—GUATEMALA: Finca Chinaux, 2042–2190 m (CM 120103–120108); Chelemhá, 2090 m (CM 120097–120102; USNM 569854, 569877, 569878) [Gazet-teer localities 12, 13].

214 ANNALS OF CARNEGIE MUSEUM VOL. 80
Cryptotis orophilus (J.A. Allen, 1895)
The Central American Least Shrew has a geographic range from northern El Salvador and western Honduras to the Central Valley of Costa Rica (Choate 1970; Woodman and Timm 1993), and its known elevational distribution is ca. 1150–1985 m. In El Salvador, the species is known on the basis of two specimens from one locality and a series of owl pellet remains from a second locality. In Honduras, it was previously known from five specimens from four lo-calities (Choate 1970; Woodman and Croft 2005). In Gua-temala, C. orophilus is probably replaced by the morpho-logically-similar C. tropicalis, but the distributional limits of both species are essentially unknown. Pleistocene fos-sils of C. orophilus were reported from a cave near Copán, Honduras (Woodman and Croft 2005). We captured a female C. orophilus on 12 October 1992 in a pitfall set in the floodplain of the Río Arcágual near the Visitors Center, 1430 m, Celaque National Park, Hondu-ras. This individual was pregnant with three embryos (CR = 5 mm). Two days later, we captured a female C. good-wini (CM 118519) and a female C. merriami (CM 118520) at the same general locality where the C. orophilus was taken, and on 18 February 1998, we captured a second fe-male C. merriami (CM 112883). This is the first record of sympatry (and potential syntopy) of these three species, although their captures were temporally segregated.
New record (1).—HONDURAS: Celaque, 1430 m (CM 118522) [Gazetteer Locality 24].
Cryptotis tropicalis (Merriam, 1895)Fig. 3
The Tropical Least Shrew has a geographic range from southern Chiapas through Guatemala (Choate 1970;
Woodman and Timm 1993). Its elevational distribution is poorly known, but specimens have been recorded from ca. 975–1580 m. We captured a non-reproductive female in a pitfall trap set in dense secondary growth in an overgrown coffee plantation 3.5 km north of La Trinidad, Guatemala, on 5 January 2009. This was the only shrew taken at the local-ity, where we spent only a single night and accumulated only 149 trapnights (21 pitfall trapnights). The nature of the vegetation at this locality suggests that this species may have a relatively high tolerance for habitat disturbance or early succession plant associations.
New record (1).—GUATEMALA: 3.5 km N La Trinidad, 1580 m (USNM 570435) [Gazetteer Locality 2].
Sorex salvini Merriam, 1897
Salvin’s Shrew (Sorex salvini) formerly was considered to be one of the six subspecies of Sorex saussurei Merriam, 1892, that were discontinuously distributed at elevations of ca. 1825–3110 m in highlands from southwestern Du-rango and southern Coahuila, Mexico, to western Guate-mala (Hall 1981). We concur with Hutterer (2005) that the two Guatemalan subspecies (S. saussurei godmani Mer-riam, 1897, and S. saussurei salvini Merriam, 1897) refer to the same taxon, but S. s. salvini has priority, and this is the name that should be used for the Guatemalan populations. Recent taxonomic work (Carraway 2007) on the Mexican forms of S. saussurei resulted in the northernmost subspe-cies, S. saussurei saussurei, being subdivided among three geographically-overlapping taxa: a monotypic S. saussurei (sensu stricto) endemic to central Mexico; S. mediopua Carraway, 2007; and S. veraecrucis altoensis Carraway, 2007. Three former subspecies of S. saussurei from south-ern Mexico (S. s. cristobalensis Jackson, 1925; S. s. oaxa-cae Jackson, 1925; S. s. veraecrucis Jackson, 1925) were all recognized by Carraway (2007) as subspecies of S. veraecrucis Jackson, 1925. Unfortunately, the taxonomic status of the Guatemalan subspecies (S. saussurei salvini) was not addressed, although its status is relevant to the tax-onomy of the Mexican taxa. Carraway (2007) showed S. veraecrucis cristobalensis is distributed in both the Altip-lanicie Central of northern Chiapas and the Sierra Madre de Chiapas in southern Chiapas. The bases of these two high-land regions converge in Guatemala (see Biogeography), and it makes biogeographical sense that, if these two dis-junct Chiapan populations belong to the same subspecies, then they are likely to be conspecific with Guatemalan S. saussurei salvini as well. If so, then S. saussurei salvini and S. veraecrucis are conspecific, and the correct name for all four of these subspecies is S. salvini (hence, S. salvini cris-tobalensis, S. salvini oaxacae, S. salvini salvini, S. salvini veraecrucis), because that name has priority under the rules of zoological nomenclature. The alternative is that S. salvini and S. veraecrucis are distinct species. Either way, S. salvini is the correct species name for the Guatemalan populations.
Fig. 3.—Cryptotis tropicalis (USNM 570435) from 3.5 km N La Trini-dad, Guatemala (photograph: NW).

2012 WOODMAN ET AL.—DISTRIBUTIONAL RECORDS OF SHREWS 215
There are few previous records of S. salvini from Gua-temala. Jackson (1928) reported five specimens from three localities: one from Todos Santos Cuchumatán, Huehu-etenango (as S. saussurei godmani: USNM 77023); two from 9000 feet on Volcán Santa María, Quezaltenango (as S. saussurei godmani: USNM 77044, 77045); and two from 10,200 feet at Calel, Totonicapán (USNM 77035, 77071). Carleton and Huckaby (1975) captured two S. salvini (UMMZ 117849, 117850) in pine-oak cloud forest (Subtropical Lower Montane Wet Forest) with ferns and moss-covered fallen trunks and branches at 2950 m at Yai-quich in the northern Sierra de los Cuchumatanes, Huehu-etenango. The relative rarity of this species, or at least the difficulty in capturing it, is emphasized by the few speci-mens we obtained during our extensive and wide-ranging field work during 1992–2009. Of the total of 280 individ-ual shrews we captured, four are S. salvini. We captured a single, reproductively inactive female S. salvini in a pitfall on 31 January 1995 near Campamento Naranjo, 2560 m, Celaque National Park, Honduras. This is the first record of the genus Sorex in Honduras, and it represents a sig-nificant extension to the east and to the south of the previ-ously known distributional limit in the Sierra de las Minas of Guatemala (Matson and McCarthy 2005). In January 2003, we obtained one male and one female S. salvini at Finca Chinaux, Guatemala, compared with 20 S. veraepa-cis ibarrai and six Cryptotis oreoryctes. In June 2006, we captured a female S. salvini in a pitfall trap in the lower portion of disturbed cloud forest at Finca La Chingada on the northwest slope of Volcán de Zunil, Guatemala. We also took a S. veraepacis chiapensis at that locality, as well as six S. v. chiapensis at nearby Bosque Ojo de Agua in early July 2006. The documented elevational distribution of S. salvini is 2560–3110 m. None of the three females we captured in January and June was reproductively active (Table 3). The digestive
tract of the S. salvini from Finca la Chingada (USNM 569591) contained numerous fragments (legs, terga, etc.) of insect exoskeleton, possibly from a coleopteran. No en-doparasites were encountered in the digestive tract of this shrew.
New records (4).—GUATEMALA: Finca la Chingada, 2720 m (USNM 569591); Finca Chinaux, 2042–2190 m (CM 120109, 120110). HONDURAS: Celaque, 2560 m (CM 119732) [Gazetteer localities 10, 12, 24].
Sorex veraepacis Alston, 1877
The Verapaz Shrew includes four subspecies (S. veraepa-cis chiapensis Jackson 1925, S. v. ibarrai Matson and Mc-Carthy, 2005, S. v. mutabilis Merriam, 1898, and S. v. ve-raepacis) that were formerly considered to be distributed discontinuously in highlands from central Guerrero, Mex-ico, to the Sierra de las Minas, in east-central Guatemala (Hall 1981; Matson and McCarthy 2005). Recently, some populations of S. v. mutabilis from Guerrero and Oaxaca were recognized as a separate species, S. ixtlanensis Car-raway, 2007. This action, however, has left the eastern and northeastern limits of S. v. mutabilis in question, as Car-raway (2007:80) was unable to determine whether relevant populations in Guerrero, Oaxaca, and Veracruz were S. v. mutabilis or S. ixtlanensis. In Mexico, the documented ele-vational range of S. veraepacis is 1120–3110 m (Carraway 2007). In Guatemala, the known elevational range was 2200–3050 m (Hall 1981; Matson and McCarthy 2005), but we found S. v. ibarrai as low as 1475 m.
Sorex veraepacis chiapensis Jackson 1925
The Chiapan Verapaz Shrew is known to occur in the Alti-planicie Central de Chiapas, the Sierra Madre de Chiapas,
Table 3. Summary of reproductive data for 97 female Sorex captured in Guatemala during the years 1996–2009.
Month Total Number of Females
Pregnant Lactating Percentage Reproductively Active
Numbers of Embryos Per Female
S. salvini: January 2 0 0 0% –
June 1 0 0 0% –
S. veraepacis chiapensis: July 8 0 3 38% –
S. veraepacis ibarrai: January 24 3 1 17% 1–2
February 2 0 0 0% –
April 26 2 10 46% 2
May 16 1 6 44% 2
July 7 0 3 43% –
S. veraepacis veraepacis January 6 0 0 0% –
July 5 2 2 80% 2

216 ANNALS OF CARNEGIE MUSEUM VOL. 80
and the Sierra Madre de Guatemala. The reported eleva-tional range is 1690–2895 m in Chiapas (Matson and Mc-Carthy 2005; Carraway 2007) and 2300–3050 m in Gua-temala (Matson and McCarthy 2005). We had ten captures of this shrew at three cloud forest localities in Guatemala with an elevational distribution of 2720–3100 m. All of our records of S. v. chiapensis were in the month of July. Among the eight females we captured, none was pregnant, but three were lactating (Table 3). This pattern is similar to that observed in July for S. v. ibarrai (see fol-lowing account). Analysis of the contents of the digestive tract of an in-dividual from Finca la Chingada (USNM 569674) yielded numerous insect parts, including the wing of a hemip-teran. Digestive tracts of three individuals from Bosque Zunil (USNM 569688, 569689, 569696) each contained fragments of insects, including one coleopteran. One of these also contained a complete insect larva, possibly a wireworm (Elateridae). Among digestive tracts from four individuals captured at Bosque Ojo de Agua (USNM 569709–569711, 569733), one contained nothing identi-fiable, three had fragments of insects, and one contained some plant debris. All eight individuals were infected with tapeworms; two also contained hymenolepid eggs.
New records (10).—GUATEMALA: Bosque Ojo de Agua, 3100 m (USNM 569709–569711, 569732, 569733, 569747); Finca la Chingada, 2720 m (USNM 569674); Bosque Zunil, 3000 m (USNM 569696), 3100 m (USNM 569688, 569689) [Gazetteer localities 9, 10, 11].
Sorex veraepacis ibarrai Matson and McCarthy, 2005Fig. 4
Ibarra’s Verapaz Shrew was known previously from 2200–2800 m in the Sierra de las Minas (Matson and McCar-thy 2005). We had 129 captures of this species from five montane forest localities with an elevational distribution of 1475–2800 m. In general, this species is the most easily captured shrew in Guatemala. It was relatively common at many
sites where we encountered it. In our 1998 surveys of small mammals at Cerro Pinalón and 6 km NNW of San Lorenzo in the Sierra de las Minas, Guatemala, S. veraepacis ibar-rai was the second most common species after Peromys-cus grandis Goodwin, 1932, representing 19% (ntot = 176 total captures) and 11% (ntot = 208) of all captures of small mammals at the two sites, respectively (Matson and Mc-Carthy 2005). This shrew was the fifth most common of 15 species at Chelemhá, with 8% of captures (ntot = 179 captures; Etot = 3208 trapnights), and it was the fifth most common of nine species at El Limo (9%; ntot =79 captures; Etot = 1494 trapnights). Reproduction in S. v. ibarrai occurs in at least four of the first six months of the year, with a possible peak in April through July (Table 3). Among 75 females captured in the months of January, April, May, and July, a total of 35% evidenced recent reproductive activity, with preg-nant or lactating individuals in each of those four months (Table 2). In January, however, only 17% of females were reproductively active, whereas in April, May, and July, the percentage increased to 43–46%. The typical number of embryos carried by females is two (in five of six pregnant females), although we observed one case in which a fe-male had a single embryo. Females captured in January average larger in mass (WT = 10.0 ± 2.0, 7.0–14.7, n = 24) than those taken in April (WT = 8.7 ± 1.7, 6.0–11.0, n = 26), May (WT = 7.4 ± 2.0, 5.3–13.5, n = 16), or July (WT = 8.6 ± 1.2, 7.0–10.0, n = 7). Males exhibit the same general pattern, averaging heaviest in January (WT = 11.6 ± 1.6, 8.5–14.6, n = 25) and lighter in April (WT = 9.5 ± 1.9, 7.0–12.0, n = 8), May (WT = 8.6 ± 1.6, 6.1–11.0, n = 9), and July (WT = 10.8 ± 1.4, 9.5–13.0, n = 6). Like females, males average lightest in May, but they are rela-tively heavier in July. In any single month, males average heavier than females from the same month. Size differ-ences through the year may reflect the presence of differ-ent age cohorts or seasonal differences in resources. Digestive tracts of four individuals from El Limo (USNM 570050, 570051, 570069, 570070) all contained fragments of chitinous insect exoskeleton, and, in one case, antennae and legs. One of these shrews also had plant debris in both the stomach and the small intestine. Among the same four individuals, one showed no evidence of internal parasites. The other three, however, all had both tapeworms and nematodes. Sorex veraepacis ibarrai is the only known host of the hystrichopsyllid flea (Siphonap-tera, Hystrichopsyllidae) Hystrichopsylla guatemalensis Lewis and Eckerlin, 2004 (Eckerlin, 2006). Prevalence of infestation by this flea varied from 9% of potential hosts at 6 km NNW of San Lorenzo to 29% at Cerro Pinalón (Lewis and Eckerlin 2004).
New records (129).—GUATEMALA: Finca Chinaux, 2042–2190 m (CM 120125–120144); Chelemhá, 2090 m (CM 120111–120123; USNM 569840, 569841, 569855, 569860, 569892, 569907, 569908, 569923, 569929, 569942, 569950, 569973–569976); Cerro Pinalón, 2560–
Fig. 4.—Sorex veraepacis ibarrai (USNM 570021) from El Limo, Gua-temala (photograph: NOG).

2012 WOODMAN ET AL.—DISTRIBUTIONAL RECORDS OF SHREWS 217
2800 m, (USAC 3735, 3736; 6 uncatalogued USAC [NS 28; SGP 300, 400, 401, 426, 438]), 2560 m (CMNH 113277–113281), 2700–2800 m (CMNH 113282–113294; 6 USAC [JOM 6860; SGP 714, 715, 724; TJM 9705, 9709]); 6 km NNW of San Lorenzo, 2200 m (CMNH 113295–113317; 19 USAC [JOM 6743, 6744, 6745, 6773, 6774; SGP 430, 477–480, 488, 499, 501–503, 508, 509, 684, 696]); El Limo, 1475 m (USNM 570021, 570026, 570027, 570050, 570051, 570069, 570070) [Gazetteer localities 12, 13, 16, 17, 18].
Sorex veraepacis veraepacis Alston, 1877Fig. 5
Alston’s Verapaz Shrew previously was reported to oc-cur at 2840–3080 m in the Sierra de los Cuchumatanes of central-western Guatemala and western Sierra de Santa Cruz in central Guatemala (Matson and McCarthy 2005). We had 16 captures from six localities with an elevational distribution of 2680–3160 m. E.W. Nelson and E.A. Goldman had relatively high numbers of captures of this shrew (22 individuals) in the Sierra de los Cuchumatanes of Guatemala during their survey in December 1895–January 1896 (Jackson 1928), even though they did not employ pitfall traps. In contrast, our work at four cloud forest localities in the Sierra de los Cuchumatanes yielded only eight specimens of S. v. ve-raepacis. Unlike the abundant S. v. ibarrai, S. v. veraepa-cis generally was captured in equal or lesser numbers than sympatric broad-clawed Cryptotis in the Sierra de los Cu-chumatanes. Only at El Retiro, where Cryptotis appeared to be absent, did we catch S. v. veraepacis in some num-bers, but even at this locality it was only the fifth most abundant taxon in a documented community of six species of small mammals. All of our captures of S. v. veraepacis were during ei-ther January or July. Among six females from January, none exhibited evidence of recent reproductive activity (Table 3). Among five captures of females in July, two were pregnant and two were lactating. Both pregnant fe-males had two embryos, which appears to be the modal number among Guatemalan subspecies of S. veraepacis (Table 2). Females captured in January averaged smaller than females taken in July for both HB (January, 69 ± 5, 59–75, n = 6; July, 75 ± 3, 71–80, n = 5) and WT (January, 6.7 g ± 0.8, 6.0–8.0, n = 6; July, 8.9 g ± 1.6, 7.0–11.0, n = 5). The heaviest females were the two pregnant individuals (10.0–11.0 g), but the remaining three July females also were heavier (7.0– 8.5 g) than most of the January females. This is distinct from the pattern observed among S. v. ibar-rai (see previous account). The contents of the digestive tract of one individual from Aldea el Rancho (USNM 570294) were mostly too finely chopped-up to identify, although there was one un-digested fragment of a plant leaf, and a few mammal hairs, the latter probably from grooming.
New records (16).—GUATEMALA: 5 km SW San Mateo Ixtatán, 3110 m (USNM 569418, 569419, 569440, 569441); Yaiquich, 2680 m (USNM 569375); Puerto del Cielo, 3350 m (1 USAC [NOG 823]; Laguna Magdalena, 2925 m (USNM 570338); Aldea el Rancho, 3020 m (1 USAC [NOG 806]; USNM 570293, 570294); El Retiro, 3030 m (USNM 570378, 570397, 570398), 3160 m (USNM 570232, 570408, 570426) [Gazetteer localities 3, 4, 5, 6, 7, 8].
CO-OCCURRENCES OF SHREWS
Although the few records of species like C. orophilus, C. tropicalis, and S. salvini indicate that sampling was prob-ably insufficient to capture all species at each locality, we can make some generalizations regarding potential sym-patry of species of shrews in the highlands of Guatemala and Honduras. As many as four species might potentially occur in the elevation range 500–2000 m: (1) C. merriami; (2) either C. tropicalis (in Guatemala) or C. orophilus (in Honduras); (3) C. goodwini; and (4) S. v. ibarrai (in Gua-temala; Table 4). Above 2000 m, as many as three species may be found: (1) one of three subspecies of S. veraepa-cis (in Guatemala); (2) S. salvini; and (3) a broad-clawed Cryptotis (C. goodwini, C. lacertosus, C. mam, C. oreo-ryctes). In highlands of central Honduras, C. hondurensis and the broad-clawed C. magnimanus also may be pres-ent (Woodman and Timm 1992, 1999). At localities where shrews were present, our work revealed the presence of from one to four species (see Gazetteer); individual trap-ping sites within localities (each site based at a distinct elevation) yielded only one to three species (Table 4). Cryptotis mayensis, whose lowland habitat and dis-tribution are distinct from most other species in Central America and Mexico, has never been found occurring with any other species of shrew. We exclude that species and the record of C. goodwini from San José de los Andes (FLMNH 27718) from further discussion, because neither record represents our field efforts. Among the remaining 24 localities, there are several that include more than one trapping site, yielding a total of 30 sites where we obtained shrews. We recorded only a single species at fewer than
Fig. 5.—Sorex veraepacis veraepacis (USAC [NOG 806]) from el Rancho, Guatemala (photograph: NOG).

218 ANNALS OF CARNEGIE MUSEUM VOL. 80
half of the localities (42%) and at a slight majority of sites (53%); two species were recorded at half of the localities (50%) and most of the remaining sites (40%); and three species were found at one locality (4%) and two sites (7%). One locality (4%) yielded four species. A number of factors may be responsible for the low numbers of species recorded at most localities and trapping sites. Most im-portant was our focus on higher elevation montane habits,
thereby lowering the likelihood of capturing species with greater affinity for lower and middle elevations. Other factors may include differences in sampling intensity, weather conditions, annual and multi-annual population fluctuations, and our inability to recognize relevant varia-tion within habitats. There is a distinct possibility, how-ever, that the low numbers of species—and the relatively low numbers of captures—provides a reasonably accurate
Table 4. Elevational distributions of shrews reported in this study. Localities are arranged in order by lowest elevation sampled at that locality. Some localities have more than one trapping site. Locality numbers are as reported in the Gazetteer of Localities. Horizontal lines are elevational
limits from the literature (see individual species accounts). Abbreviations for species: Cgo, Cryptotis goodwini; Cla, C. lacertosus; Cmm, C. mam; Cmy, C. mayensis; Cme, C. merriami; Cor, C. orophila; Coy, C. oreoryctes; Ctr, C. tropicalis; Svc, Sorex veraepacis chiapensis; Svi, S. v.
ibarrai; Svv, S. v. veraepacis; Ss, S. salvini.
Lowest Species Of Shrews PresentLocality Elevation Cmy Cme Ctr Cor Cgo Svi Coy Ss Svc Svv Cla Cmm 5 3350 — Svv Cmm 8 3160 Svv 3 3110 — Svv Cla 9, 11 3100 Svc 8 3030 Svv 7 3020 Svv Cmm 11 3000 Svc 6 2925 Svv Cmm 2895 —10 2720 Ss Svc16 2700 Cgo Svi 4 2680 Svv Cla 16, 24 2560 Cgo Svi Ss19 2400 Cgo 2300 —17 2200 Cgo Svi 13 2090 Svi Coy24 2075 Cgo 12 2040 Svi Coy Ss 1985 —25 1900 Cgo 19 1650 Cme 15 1640 Cme 2 1580 Ctr23 1500 Cme Cgo 26 1480 Cme 18 1475 Cme Svi 24 1430 Cme Cor Cgo 14 1290 Cme 22 1200 Cme — 1150 — 20 1020 Cme 975 — 21 600 Cme 1 100 Cmy

2012 WOODMAN ET AL.—DISTRIBUTIONAL RECORDS OF SHREWS 219
accounting of presence, absence, and abundance at most sites. That this may be true is supported by the complete-ness of the patterns in Table 4, which emphasizes the el-evation limits of taxa, but which also shows that specific subsets of species that may act as regional morphological equivalents of each other do not occur at the same local-ity, such as C. orophilus and C. tropicalis; the four species of broad-clawed Cryptotis (C. goodwini, C. lacertosus, C. mam, C. oreoryctes); and the three subspecies of S. ve-raepacis. In Guatemala, no site included more than one species of Cryptotis, yet at one of the five Honduran sites, we found two species of Cryptotis occurring syntopically, and, at another site, we found three. The two localities where more than two species of shrews were encountered were Finca Chinaux in the Si-erra de Yalijux, Guatemala, and Celaque in the Sierra de Celaque, Honduras. We captured a total of 29 shrews of three taxa over eight nights at Finca Chinaux. These in-cluded six C. oreoryctes, two S. salvini, and 21 S. v. ibar-rai. At Chelemhá, the only other site we sampled in the Sierra de Yalijux, we captured 37 individuals of only two species during two trips for a total of 13 nights: nine C. oreoryctes and 28 S. v. ibarrai. The species missing at Chelemhá, S. salvini, is infrequently captured and poorly represented in systematic collections. Although Chelemhá is at a higher elevation than Finca Chinaux, it is within the elevational range of S. salvini, and that species probably occurs there, but in very low numbers, and it is difficult to capture. At Celaque, Honduras, we captured one C. goodwini, two C. merriami, and one C. orophilus at 1430 m; five C. goodwini at 2075 m; and seven C. goodwini and one S. salvini at 2560 m. The lowest site represents the first record of either syntopy or sympatry of the three species of Cryptotis. Neither C. merriami nor C. orophilus is known to occur above 2000 m elevation, so they probably are not present at the two higher trapping sites. Similarly, S. sal-vini is not known from below 2000 m in Guatemala, so our trapping records from these three sites probably are accurate reflections of the compositions of the shrew com-munities. Fox and Kirkland (1992; Kirkland 1991) hypothesized an “assembly rule” for communities of shrews in eastern North America, which commonly contain up to six species. They noted that these assemblages consist of individual species of shrews that can be grouped into three size class-es based on mass (Kirkland 1991): small (mean = 3–4 g); medium (5–7 g); large (12–20 g). In each size class, there is a maximum of two species, one of which is common, the other uncommon. The size classes presumably function to lessen competition among shrews and promote coexis-tence in the same general habitat. Different size groupings of shrews in Asia, for example, have been shown to have different foraging modes (epigeal vs. hypogeal) and to ex-ploit different proportions of prey of different size classes (Churchfield and Sheftel 1994; Churchfield et al. 1999). Fox and Kirkland’s (1992) assembly rule predicts that
each of these size groups would be filled with one species before a second species of any size class would be pres-ent (Kirkland 1991). If a similar assembly rule exists for the less diverse Central American shrew communities, we would expect each of the species in an association of three or fewer species to belong to a different size class similar to those defined by Kirkland (1991). At Finca Chinaux, the three species represent three distinct size classes (Table 1), although the groupings are somewhat different than those based on eastern North American shrews. The smallest species, S. salvini (mean = 5.3 g for the species—Table 1; 6.0 g for two individuals from Sierra de Yalijux), would be classed as a member of Kirkland’s (1991) “medium” body size group; S. v. ibar-rai (9.5 g; 10.8 g for 49 individuals from Sierra de Yali-jux) falls into the gap between the “medium” and “large” categories; and C. oreoryctes (13.7 g) fits into the “large” grouping. The distribution of size classes at this site ap-pears to conform to the expectations of the assembly rule. At the lowest elevation at Celaque, there are two spe-cies from the “medium” size class, C. merriami (5.8 g for 27 from Guatemala and Honduras; two individuals from Celaque average 6.7 g) and C. orophilus (5.8 g for 14 individuals from Honduras and Costa Rica; 5.6 g for the individual from Celaque), and one species from the gap between the “medium” and “large” categories, C. good-wini (8.9 g for 14 individuals from Celaque). The presence of two species in the same size class (i.e. C. merriami, C. orophilus), without a member of a third size class present, violates the assumptions of the assembly rule. Size variation in the Sierra de Yalijux may function in part to limit competition, but, if so, the relationship is com-plex. The magnitude of seasonal variation in mean size of S. v. ibarrai (which is lower among species of Cryptotis), may permit this species to vary its prey focus through the year, and differences in mean mass between sexes may serve to limit the overlap in food resource use between fe-males and males. Both patterns may complicate size rela-tionships among species, but probably are one reason that S. v. ibarrai is dominant numerically where it occurs. For C. oreoryctes, the highly derived forearm adaptations that permit it to exploit a hypogeal feeding guild (Woodman
Table 5. Summary of reproductive data for all female shrews captured in Guatemala and Honduras during the
years 1992–2008.
Total Percentage Number of ReproductivelyMonth Females Pregnant Lactating Active
January 50 5 1 12%February 15 0 0 0%April 27 3 10 48%May 16 1 6 44%June 1 0 0 0%July 31 5 11 52%October 3 1 0 33%December 9 0 1 11%

220 ANNALS OF CARNEGIE MUSEUM VOL. 80
and Stephens 2010; Woodman 2011a) are probably more important than size for limiting interspecific competition for food resources. For this species, large size may assist it in handling the larger prey (e.g., earthworms) that hy-pogeal foraging makes available (Churchfield and Sheftel 1994; Churchfield et al. 1999). At Celaque, the one spe-cies that is most different in size (C. goodwini) is also the most divergent morphologically (Woodman 2010, 2011a), and, like C. oreoryctes, it probably specializes to a greater degree on hypogeal prey. The two species that are clos-est in size at this locality (C. merriami, C. orophilus) are the most similar morphologically (they can be difficult to distinguish in the hand), and both are more adapted for preying on epigeal invertebrates. Limiting competition between these two species may require them to forage in different microhabitats.
REPRODUCTION IN CENTRAL AMERICAN SHREWS
Data on reproductive patterns of Mexican and Central American soricids are scarce and scattered. A pregnant Cryptotis berlandieri Baird, 1857, was reported from Tamaulipas in July, and lactating females (n = 2) were present in June and July; a pregnant Cryptotis mexicanus (Coues, 1877) was documented in Veracruz in December, and a lactating female in October; pregnant Cryptotis mag-nus (Merriam, 1895) were noted in Oaxaca in May (n = 1) and October (n = 1); pregnant Cryptotis nigrescens in Costa Rica and Panama were reported in May and July (n = 3) and a single lactating female in March (Alvarez 1963; Hall and Dalquest 1963; Robertson and Rickart 1975; Woodman and Timm 1993). The most complete data avail-able for a Neotropical shrew are from Cryptotis meriden-sis (J.A. Allen, 1895) Thomas, 1898, in the Venezuelan Andes, which appears to have the potential to reproduce at any time of the year (Woodman and Díaz de Pascual 2004). Pregnant females of this species were present in all months of the year except July and December. The largest proportion of pregnant females was in April (23% of April females). Females were lactating in all months of the year, with the highest proportion in July (36% of July females). Reproductive information for the species of soricids we captured in Guatemala and Honduras is incomplete and mostly from the first half of the year (Tables 2–3). Our most extensive dataset is for S. veraepacis ibarrai, for which we have information from 75 females representing five months of the year. These data indicate that some fe-males are pregnant or lactating in January, April, May, and July, but that a minority of the female population is repro-ductively active at any given time. The relative numbers of pregnant and lactating females suggest an increased level of reproduction in April through July, at least in contrast to the first two months of the year. This pattern is supported by scarce data available from S. v. chiapensis and S. v. ve-raepacis, which also show females reproductively active
in July (Table 2). Our data for Cryptotis are less complete, owing to the lower numbers of females captured. Overall, those data indicate that female C. goodwini can be repro-ductively active in April and December; C. mam and C. merriami in July; C. oreoryctes in January; and C. orophi-lus in October (Table 3). The generalized pattern for all Soricidae combined (Table 5) suggests that reproductively active shrews are present in January, April, May, July, October, and Decem-ber. The only month (for which we have data) in which there appears to be a lull in reproduction is February. Peak reproductive activity appears to be in April through July, with a tapering off toward October. If representative, this suggests that shrews may be timing most births and lacta-tion to coincide with rainy season, which lasts from ap-proximately mid-May through October. Arthropod abun-dance is positively associated with rainfall (Bergallo and Magnusson 1999), as is availability of earthworms (Frago-so and Lavelle 1992), hence the peak of reproduction may be timed to coincide with increased availability of food resources. The pattern should be considered with caution, however, as it is heavily biased by data from a single spe-cies, S. v. ibarrai. It also combines data from two distinct genera and eight species, and different species of shrews can exhibit distinct reproductive patterns in the same re-gion (Yu et al. 2001), as do Neotropical montane species of rodents (Heideman and Bronson 1992; Vázquez et al. 2000). In fact, the different patterns of seasonal size varia-tion for S. v. ibarrai (females and males largest in January, smallest in May) and S. v. veraepacis (females largest in July, smallest in January; insufficient data for males) sug-gests different reproductive peaks in these two subspecies, if the pattern reflects the presence of different age cohorts at different times of the year,. The number of embryos in Sorex from Guatemala and Honduras ranges from one to two, with a modal value of two offspring. In Cryptotis, the range is two to three off-spring (Table 3). These are atypically small potential litter sizes for species in these two genera compared to species in northern North America. Sorex cinereus Kerr, 1792, for example, is reported to average seven young per litter in Ohio and Manitoba and eight young per litter in Ontario, with an overall range of 3–12 young per litter (Buckner 1966; Forsyth 1976; Innes et al. 1990). Sorex arcticus Kerr, 1792, averages seven young per litter in Ohio and Wisconsin and eight young per litter in Minnesota (range = 5–9; Buckner 1966; Baird et al. 1983). Large numbers of young also are typical for S. fumeus G.M. Miller, 1895 (2–8 young), S. hoyi Baird, 1857 (5–8), and S. palustris Rich-ardson, 1828 (3–10; Whitaker and Hamilton 1998). Whita-ker (1974) reports a mean of five embryos (range: 2–7) for Cryptotis parvus (Say, 1823) in North America. Simi-larly, Mock and Conaway (1976) calculated a mean of five young (range: 1–9, n = 327 litters) for a captive colony of that species. Among Neotropical soricids, however, small-er litters appear to be more typical. Three embryos were recorded for a female C. berlandieri, from Tamaulipas,

2012 WOODMAN ET AL.—DISTRIBUTIONAL RECORDS OF SHREWS 221
Mexico; a female C. mexicanus from Veracruz, Mexico; two female C. magnus from Oaxaca, Mexico; and a female C. nigrescens from Costa Rica. Two C. nigrescens from Panama each contained a single embryo (Alvarez 1963; Hall and Dalquest 1963; Robertson and Rickart 1975; Woodman and Timm 1993). A litter size of three pups is typical for Venezuelan C. meridensis, the known range be-ing from two to four embryos per female (Woodman and Díaz de Pascual 2004).
BIOGEOGRAPHY
The biogeography of northern Central America relies to a large extent on the geological history of the region and the resulting topography, which has been addressed in detail by Donnelly et al. (1990) and Savage (2002). This influ-ence can be seen in the distributions of a wide variety of animals (MacVean and Schuster 1981; Savage 1982; Wake 1987; Crother et al. 1992; Hernández-Baños et al. 1995; Sullivan et al. 1997; Campbell 1999; Watson and Peter-son 1999; Sullivan et al. 2000; Castoe et al. 2003). Most soricids in Central America (with the notable exception
of C. mayensis) tend to be middle- to high-elevation spe-cies with a requirement for moist habitats, so the interplay between topography and climate, particularly rainfall, has a profound influence on their current distributions. Past events that had the most relevance to current distributions probably include Pleistocene environmental changes that at times promoted active alpine glaciation in the highest parts of Guatemala and Costa Rica as well as associated lowering of climatic zones, redistribution of plant species, and restructuring of vegetational communities throughout Central America (Markgraf 1989; Bush and Colinvaux 1990; Bush et al. 1992; Leyden et al. 1993; Islebe and Hooghiemstra 1997). During these times, some low- and middle-elevation gaps that currently act as barriers for highland mammals might have been effectively removed. Throughout southern and central Guatemala, numerous isolated mountain ranges occur that are connected at low to middle elevations (ca. 700–1200 m) and may have been occupied in the past by species that now occur only at high-er elevations (Fig. 6). The Altiplanicie Central of northern Chiapas extends southeast by way of such “middle eleva-tions” into western Guatemala. These middle elevations
Fig. 6.—Map of northern Central America illustrating major topographical features. The contour line represents 1000 m elevation.

222 ANNALS OF CARNEGIE MUSEUM VOL. 80
provide a connection, not only with the Guatemalan high-lands, but with the Sierra Madre of southern Chiapas. In intervening west-central Guatemala, the extensive Sierra de los Cuchumatanes in Huehuetenango and Quiche de-partments has its own middle elevation connections to the east via the Sierra de Chuacús in Baja Verapaz to the Sierra de las Minas in northern El Progreso and Zacapa, and to the Sierra de Yalijux in AltaVerapaz. The Sierra de las Minas is the last extensive mountain range to the east that reaches elevations above 3000 m. This highland is separated from the Sierra de Merendón along the border with Honduras by the Río Motagua Valley. In addition to its low elevation, the Río Motagua Valley rapidly becomes xeric upstream, resulting from a rain shadow effect of the Sierra de las Minas, which intercepts moisture-laden east-ern trade winds. The dryness of the valley extends upslope a considerable distance on both sides of the river, providing
an effective barrier to dispersal between the Sierra de las Minas and both the Sierra del Merendón to the east and the Sierra Madre de Guatemala to the south. The Sierra de los Cuchumatanes also has mid-elevation connections south to the Sierra Madre de Guatemala, which continues east al-most to the borders with El Salvador and Honduras, but is effectively separated from mountains in those countries by extensive lowlands. Within El Salvador and Honduras, the principal highlands are lower and more isolated. The re-sulting maze of highlands and low, dry valleys establishes a series of broadly distributed, but widely separated habitat islands for moist, highland-adapted small mammals. The discontinuous distributions of shrews through this portion of northern Central America holds interest at both the species and genus levels. The distribution of Sorex ter-minates in the discontinuous ranges of mountains and hills in western Honduras, whereas the distribution of Cryptotis
Fig. 7.—Map of northern Central America showing the locations of sampling localities in Guatemala and Honduras. Numbers on the map key to locality numbers in the Gazetteer of Localities.

2012 WOODMAN ET AL.—DISTRIBUTIONAL RECORDS OF SHREWS 223
continues into South America. Does this distribution rep-resent dispersal to the east, or is it the vicariant remnant of a larger, more southern distribution that contracted? Of the two species of Sorex in Guatemala, S. veraepacis typically is the most common shrew locally, it is more continuously-distributed, and it has a lower elevational limit than S. sal-vini, which is rare and appears to be discontinuously dis-tributed. Yet, S. veraepacis has not been found further east in Central America than the Sierra de las Minas, whereas the locally less common S. salvini, which is now known from Honduras, is the southeasternmost representative of the genus in the New World. The two species have been reported sympatrically, if not syntopically, at several lo-calities (Jackson 1928; Carleton and Huckaby 1975). The ecological adaptations or historical events that distinguish these two species and permit the less common species to have a greater eastward distribution remain to be discov-ered. Although our work has expanded the known distribu-tion of the broad-clawed Cryptotis, these taxa, like S. sal-vini and S. veraepacis, are restricted to higher elevations (>1400 m), and the group reaches the southern limits of its distribution in western Honduras. In contrast, C. merriami is widely distributed at mid-elevations in northern Guate-mala and Honduras, and C. orophilus occurs in Honduras, but not Guatemala, where it probably is essentially re-placed by C. tropicalis. The southern distributions of both C. merriami and C. orophilus are discontinuous, spanning
the lowlands of the Nicaraguan gap, to terminate in Costa Rica. The complex distributions of these taxa continue to be worked out, but their individual biogeographical histo-ries will require further understanding of the taxonomic relationships of the individual species, as well as a com-prehensive phylogenetic framework for both genera. More-over, in-depth ecological and life-history profiles for each species are essential to understanding geographical limits. These types of information gain added urgency given the high proportion of soricid species that are restricted to moist, high-elevation habitats and are thus potentially vul-nerable to the effects of environmental changes associated with human-induced habitat destruction and global warm-ing (Colwell et al. 2008; McCain and Colwell 2011). Knowledge of soricids and other small mammals in Gua-temala and Honduras would benefit greatly by relatively inexpensive long-term pitfall trapping at individual sites, such as the substantial studies carried out successfully by Díaz de Pascual and collaborators in Venezuela (Díaz de Pascual 1993, 1994; Durant et al. 1994; Díaz de Pascual and de Ascenção 2000).
Gazetteer of Localities
Localities in the following list are divided between Gua-temala and Honduras. They are further organized by topo-graphical features and are listed approximately north to south and west to east. The locality numbers key to numbers
Fig. 8.—View of camp and secondary vegetation at 3.5 km N La Trinidad, Guatemala, in January 2009 (photograph: NW).

224 ANNALS OF CARNEGIE MUSEUM VOL. 80
shown on Figure 7. Dates represent times when sites were sampled. Species of soricids encountered at each locality are provided in brackets.
GuatemalaPetén Lowlands
1. Cerro Cahuí, 100–300 m (17º00’18”N; 89º43’00”W); Petén Dept. (8 August 1988). This site is based on a single museum record from a locality not sampled by members of our team. It is located along the northeastern border of Lake Petén-Itzá and is within the Subtropical Moist Forest life zone. [C. mayensis]
Guatemalan Extension of the Altiplanicie Central de Chiapas
2. 3.5 km N La Trinidad, 1580 m (16º03’30”N, 91º42’51”W; Fig. 8); Huehuetenango Dept. (5 January 2009). We spent a single night at this locality, setting out 124 Museum Specials, four Victor rat traps, and 21 pitfalls in dense secondary growth in an overgrown coffee plan-tation. We captured eight rodents and our only specimen of the Tropical Small-eared Shrew. The locality is within
the Subtropical Lower Montane Moist Forest life zone. [C. tropicalis]
Sierra de los Cuchumatanes
3. 5 km SW of San Mateo Ixtatán, 3110 m (15º48’36”N, 91º30’20”W; Fig. 9); Huehuetenango Dept. (27–30 July 2005; 30 December 2005 to 2 January 2006). This is the type locality for Cryptotis lacertosus. Our camp was along a dirt road at the head of a trail leading to the Cerro Bobí radio towers (Matson et al. in press). The north-facing slope held a healthy, closed-canopy cloud forest, whereas the south-facing slope was cut over and had a very open canopy. The remnant forest here contained a mixture of oak, fir, and some pine associated with deep leaf litter and abundant ferns and mosses. The dense understory had fallen logs in abundance. The site is within the Subtropical Montane Wet Forest life zone. [C. lacertosus, S. v. veraepacis]
4. Yaiquich, 2680 m (15º45’44”N, 91º30’10”W; Figs. 10–11); 6 km NW of Santa Eulalia, Huehuetenango Dept. (22–25 July 2005). This is near the type locality for the Mayan Deermouse, Peromyscus mayensis Carleton and Huckaby, 1975. The remnant forest along the hillsides was still be-ing logged, and a few large (ca. 60 cm diameter) oaks and
Fig. 9.—View from Cerro Bobí of base camp 5 km southwest of San Mateo Ixtatán, Guatemala, in January 2006. To the left of the tents and truck was a large patch of uncut, healthy cloud forest where we ran traplines (photograph: NOG).

2012 WOODMAN ET AL.—DISTRIBUTIONAL RECORDS OF SHREWS 225
pines remained among the scattered stumps of still larger (ca. 120 cm diameter) oaks and firs. Ferns and mosses were abundant, and the ground had deep leaf litter. There were numerous fallen trunks and branches. The valley below the forest was dominated by meadow and pasture (Matson et al., in press). This site is within the Subtropical Montane Wet Forest life zone. [C. lacertosus, S. v. veraepacis]
5. Puerto del Cielo, 3350 m (15º33’17”N, 91º36’00”W; Fig. 12); 5 km N Todos Santos Cuchumatán, Huehuetenango Dept. (2–5 January 2005; 17 July 2008). We visited this lo-cality twice (Matson et al. in press). Vegetation is predomi-nantly secondary scrub in a cut-over fir forest with scattered pines. There were many limestone outcrops. On the ground, a 10–15-cm-deep humus layer was overlain by ca. 5–15 cm layer of moss. Our 650 trapnights in 2005 provided only one shrew. In 2008, we set 99 Museum Specials, 20 pitfalls, and eight Victor rat traps. This effort yielded seven rodents and one C. mam. This locality is within the Subtropical Montane Wet Forest life zone. [C. mam, S. v. veraepacis]
6. Laguna Magdalena, 2925 m (15º32’06”N, 91º23’33”W; Figs. 13–14); 22 km NNE of Chiantla, Huehuetenango Dept. (4 January 2006, 9–11 January 2008). We visited this locality twice (Matson et al. in press). The Laguna Magdalena Reserve is named for a small pond (Laguna Magdalena) at the end of a fast-moving stream in a narrow limestone valley with a step-like series of moss-covered falls 1–2 m in height. As late as 2000, there were only one or two houses in the area, and forest covered the slopes.
Since then, an improved dirt road was constructed, and by January 2006, about 40 families were living along the road at the entrance to the reserve. Both inside and outside the reserve, trash littered the stream and meadow, and sheep, pigs, and dogs were abundant. The tops of the surrounding hills were still covered with conifers, but the hillsides were over-grazed pasture, and the remaining forest was being slowly removed for firewood and building materials. Grass in the flat valley bottom and steep hillsides leading to the pond was thick. Runways of the Guatemalan Vole, Mi-crotus guatemalensis Merriam, 1898, were evident in the grassy areas, and six individuals were taken in one night of trapping. When we returned January 2008, the valley was strongly overgrazed. There was no obvious evidence of voles in the valley bottoms, and we captured only a single individual (on the hillside above the lagoon) in three nights of trapping. The locality is within the Subtropical Montane Wet Forest life zone. [C. mam, S. v. veraepacis]
7. Aldea el Rancho, 3020 m (15º29’22”N, 91º33’16”W; Fig. 15); 6.5 km SE of Todos Santos Cuchumatán, in the Reserva Forestal Todos Santos Cuchumatán, Huehu-etenango Dept. (31 December 2004–2 January 2005; 6–8 January 2008). We visited this site twice, working along La Maceta [also spelled as Maseta] Ecological Trail, a steep, rocky path leading through an open forest of pine with fir, cypress (Juniperus standleyi Steyermark and Cu-pressus lusitanica Miller, Cupressaceae), and monkey paw trees (Chiranthodendron pentadactylon Larreategui, Mal-vaceae) to a nearly vertical limestone escarpment. A num-
Fig. 10.—Road near Yaiquich, Guatemala, in July 2005, lined with neat stacks of freshly-cut firewood awaiting pick-up (photograph: JOM).

226 ANNALS OF CARNEGIE MUSEUM VOL. 80
Fig. 12.—Camp at Puerto del Cielo, Guatemala, in January 2005 (photograph: JOM).
Fig. 11.—Camp at Yaiquich, Guatemala, in July 2005 (photograph: WB).

2012 WOODMAN ET AL.—DISTRIBUTIONAL RECORDS OF SHREWS 227
Fig. 13.—Entrance to the Laguna Magdalena Reserve, Guatemala, in January 2008. The stream feeding the laguna is in the lower right of the photograph (photograph: NW).
Fig. 14.—Laguna Magdalena, Guatemala, in January 2006. (photograph: JOM).

228 ANNALS OF CARNEGIE MUSEUM VOL. 80
ber of trees had been harvested for lumber and firewood, and logging continued while we were there. We captured three C. mam and one S. v. veraepacis during our first visit in 2005. In 2008, we obtained four C. mam and two S. v. veraepacis in 1123 trapnights (575 Museum Specials, 241 Shermans, 50 rat traps, 257 pitfalls). All of the shrews were taken in mossy areas or along a small, intermittent stream. The locality is within the Subtropical Montane Wet Forest life zone. [C. mam, S. v. veraepacis]
Montaña Cuilco
8. El Retiro, 2950 m, 3030 m, 3160 m (15º29’N, 91º55’W; Figs. 16–17); 9–10 km NNE Cuilco, Huehuetenango Dept. Montaña Cuilco and Montaña Peña Blanca together form an isolated highland complex that is bordered by the Cuilco and Selegua river valleys southwest of the Sierra de los Cuchu-matanes. Our locality of El Retiro actually comprises three trapping sites: 9 km NNE Cuilco, 2950 m (15º29’00”N, 91º55’51”W), where we worked in July 2008; 9.5 km NNE Cuilco, 3030 m (15º29’09”N, 91º55’20”W), where we were located in December 2008 and January 2009; and 10 km NNE Cuilco, 3160 m (15º29’22”N, 91º55’16”W), where we had traplines in July 2008 and January 2009. All of the sites are within an extensive stand of mixed forest com-prised of pine, cypress, and hardwoods that is controlled by the local community. Mosses and hardwoods were more abundant at the highest elevation, pine and cypress at 3030 m. Although lumbering primarily consists of extraction of individual trees, at least two small parcels had been clear-cut. Portions of the area surrounding the stand are a patch-work of forest and open grassland, at least some of which is a result of clear-cutting. Locals told us that there had been “two types” of squirrels in the forest, but now the squirrels were gone. A fire burned through the area about ten years prior to our visit, possibly coinciding with the widespread wildfires that occurred in northern Central America as a re-sult of unusually dry conditions during the El Niño year of 1998. Despite three visits and at least 1627 trapnights (in-cluding 310 pitfall nights) at this locality, we captured only six S. v. veraepacis: none were taken at 2950 m; three were captured in January 2009 at 3030 m; and one was captured in July 2008 and two in January 2009 at 3160 m. El Retiro is within the Subtropical Montane Wet Forest life zone. [S. v. veraepacis]
Sierra Madre de Guatemala
9. Bosque Ojo de Agua, 3100 m (15º05’23”N, 91º36’08”W); 5 km ENE Cabricán, Quetzaltenango Dept. (6–8 July 2006). This site is in a 56-ha section of forest managed by the vil-lage of Cabricán. The open forest was dominated by pine and fir with scattered oak and cypress, an understory of bushes, ferns, and mosses, and a fairly deep accumulation of leaf litter. The sound of chain saws was heard every day. The village of Calel, where Nelson and Goldman collected
in 1895 (Goldman 1951), was visible from higher elevations in the forest. This locality is within the Subtropical Montane Moist Forest life zone. [S. veraepacis chiapensis]
10. Finca la Chingada, 2720 m (14º45’07”N, 91º27’47”W); 4 km SE Zunil, Quetzaltenango Dept. (30 June to 3 July 2006). Finca La Chingada and the two Bosque Zunil sites (Locality 11) are at different elevations along the western slope of Volcán Zunil that we worked in June and July 2006. Vegetation was primarily secondary cloud forest with a few large (ca. 30 m tall, 60–70 cm diameter) remaining trees interspersed with meadows of tussock grass. The ground in the forest had deep leaf litter with abundant moss-covered rocks. Trees were actively being cut and removed for fire-wood while we were there. This locality is within the Sub-tropical Lower Montane Wet Forest life zone. [S. salvini, S. veraepacis chiapensis]
11. Bosque Zunil, 3000 m, 3100 m (14º43’N, 91º28’W); 6–6.5 km SE of Zunil, Quetzaltenango Dept. (2–3 July 2006). This locality comprises two sites about 1½ hours’ hike up a ridge on the western slope of Volcán Zunil from Finca la Chingada (Locality 10): 6 km SE of Zunil, 3000 m (14º43’14”N, 91º28’51”W), where we worked 2–3 July; and 6.5 km SE of Zunil, 3100 m (14º43’55”N, 91º28’09”W), where we worked 3 July. The cloud forest at 3000 m was similar to that at Finca La Chingada in general aspect, and, as at that site, trees were actively being cut and removed for firewood. Cloud forest at 3100 m was in better condition, with abundant pine, fir, cypress, oaks, and occa-sional monkey paw. Trees were covered in epiphytes, and the ground had abundant areas of deep moss. The locality is within the Subtropical Lower Montane Wet Forest life zone. [S. veraepacis chiapensis]
Sierra de Yalijux
12. Finca Chinaux, 2042–2190 m (15º23’47”N, 90º06’17”W); Alta Verapaz Dept. (6–13 January 2003). Campamento de los Helechos is the informal name given to our campsite at 2042 m on the western side of the Si-erra de Yalijux, from which we worked up the north slope of the mountains. The locality is within the Subtropical Montane Rain Forest life zone. [C. oreoryctes, S. salvini, S. veraepacis ibarrai]
13. Chelemhá, 2090 m (15º23’09”N, 90º03’44”W; Fig. 18); Alta Verapaz Dept. (9 to 14 January 2003; 31 Decem-ber 2006 to 6 January 2007). This locality is in the private Chelemhá Cloud Forest Reserve, managed by the conser-vation organization, Unión para Proteger el Bosque Nu-boso. The cloud forest is dominated by oaks and pines that form a mostly closed canopy at ca. 20 m, with occasional emergents reaching to ca. 25 m, and a subcanopy at ca. 5 m. Tree ferns, vines, epiphytes, mosses, lycopods, and liverworts abounded. The floor of the forest had abundant downed, mossy tree trunks and branches. Leaf litter was

2012 WOODMAN ET AL.—DISTRIBUTIONAL RECORDS OF SHREWS 229
Fig. 15.—Beginning of the La Maceta Ecological Trail, Aldea el Rancho, Guatemala, in January 2008 (photograph: NW).
Fig. 16.—Logging camp at El Retiro, 9 km NNE Cuilco (2950 m), Guatemala, in July 2008 (photograph: WB).

230 ANNALS OF CARNEGIE MUSEUM VOL. 80
Fig. 17.—Overlooking base camp (to the left of the road) at El Retiro, 9.5 km NNE Cuilco (3030 m), Guatemala, in January 2009 (photograph: NW).
Fig. 18.—Chelemhá Cloud Forest Reserve, Sierra de Yalijux, Guatemala, in January 2007 (photograph: NOG).

2012 WOODMAN ET AL.—DISTRIBUTIONAL RECORDS OF SHREWS 231
Fig. 19.—View south from the house at Finca La Concepción, Baja Verapaz, Guatemala, in July 2007. The small patch of forest on the slope behind the house marks where C. merriami was captured. The intervening cornfields represent the dominant vegetation at this locality (photograph: NW).
Fig. 20.—Hotel Country Delights, Baja Verapaz, Guatemala, in July 2007. Cryptotis merriami was captured in the small patch of forest on the hill behind the hotel (photograph: NW).

232 ANNALS OF CARNEGIE MUSEUM VOL. 80
Fig. 21.—View south from the Sierra de Yalijux to the Sierra de las Minas, Guatemala (photograph: NOG).
Fig. 22.—Secondary forest along the stream at El Limo, Zacapa, Guatemala, in July 2007 (photograph: NOG).

2012 WOODMAN ET AL.—DISTRIBUTIONAL RECORDS OF SHREWS 233
about 10 cm deep, overlying an often deep, but rocky soil. The reserve is the type locality for Cryptotis oreoryctes. In 2006–2007, we worked primarily along established trails on a north-facing slope. Howler Monkeys [Alouatta pal-liata (Gray, 1849)] were heard and occasionally seen here. The locality is within the Subtropical Montane Rain Forest life zone. [C. oreoryctes, S. veraepacis ibarrai]
Río Polochic / Río Panimá Divide
14. Finca Concepción, 1290 m (15º15’54”N, 90º07’33”W; Fig. 19); 3 km S of Tucurú, Baja Verapaz Dept. (18–19 July 2007). This former coffee plantation is the type local-ity for Peromyscus grandis Goodwin, 1932. At the time of our visit, the land had been almost entirely converted to cultivation of corn. There were only a few, very small patches of secondary forest left. We were told the planta-tion, which was purportedly owned by a vice minister of agriculture, was for sale. The locality is within the Sub-tropical Moist Forest life zone. [C. merriami]
Quebrada La Cruz Valley
15. Hotel Country Delights, 1640–1665 m (15º14’08”N, 90º15’13”W; Fig. 20); a hotel and restaurant 9 km S of Pasmolón (2 km W of Purulha), between km 166 and km 167 on highway CA-14, BajaVerapaz Dept. (20 July 2007). Quebrada La Cruz is a tributary of Río Sulín, which flows into Río Panimá, itself a tributary of Río Matanzas. We set lines totaling 46 Museum Specials, 30 Shermans, three Victor rat traps, and 37 pitfalls in a small, disturbed island of mostly secondary forest bordering pasture on the slope of a small hill behind the restaurant and guest houses. The forest contained pines, cypress, oaks, and tree ferns. Bro-meliads and other epiphytes were abundant, and ground cover included several species of ferns, mosses, and liver-worts. A single night of trapping yielded two shrews and three rodents. The locality is within the Subtropical Wet Forest life zone. [C. merriami]
Fig. 23.—Overlooking cloud forest at Trifinio, Guatemala, in July 2004 (photograph: NOG).

234 ANNALS OF CARNEGIE MUSEUM VOL. 80
Sierra de las Minas
16. Cerro Pinalón, 2560 m, 2700–2800 m (ca. 15º05’N, 89º56’W; Fig. 21); Municipio San Agustín Acasaguastlán, El Progreso Dept. (2–12 May 1998). This locality compris-es two general trapping sites, La Cabaña and Camino a las Torres. La Cabaña, 2560 m (15º04’50”N, 89º56’32”W), is a ranger station and visitors center that included a sleeping cabin with bunks, kitchen, and solar-powered lighting. It is in well-preserved cloud forest, and Resplendent Quetzals (Pharomachrus mocinno de la Llave, 1832) and Howler Monkeys were heard and occasionally seen, and a Coati [Nasua narica (Linnaeus, 1766)] frequented camp. The team ran multiple traplines here throughout our stay (2–12 May 1998), but captured only S. v. ibarrai. Camino a las Tor-res, 2700–2800 m (15º04’54”N, 89º55’59”W), comprises a series of traplines set along a road running to transmission towers at the top of the mountain, which is the highest point in this part of the Sierra de las Minas. These traplines were run 4–12 May 1998, and they obtained C. goodwini and S. v. ibarrai, as well as a weasel (Mustela frenata Lichtenstein, 1831). A particularly strong El Niño event took place dur-ing 1998 (Trenberth et al. 2002), and forest fires were burn-ing at lower elevations. Despite this event, temperatures at the ranger station on 5 May ranged from 4º–26ºC, and frost coated the ground on some mornings. The locality is within the Subtropical Lower Montane Rain Forest life zone. [C. goodwini, S. veraepacis ibarrai]
17. 6 km NNW of San Lorenzo, 2200 m (15º08’09”N, 89º41’10”W); [ca. 15.5 km NNW of Río Hondo] Zacapa Dept. (8–17 April 1998). It was quite dry during the El Niño year of 1998 (Trenberth et al. 2002), and forest fires burn-ing at lower elevations impeded our arrival at this locality. The upper slopes here contained healthy stands of cloud forest with large numbers of lianas, lichens, and epiphytes. Our traplines ran through cloud forest and upslope into elfin forest and ericaceous shrub, which also capped the summit of the cordillera. We captured two C. goodwini and 42 S. v. ibarrai here. A kinkajou [Potos flavus (Scheber, 1774)] passed through camp each night. The locality is within the Subtropical Lower Montane Rain Forest life zone. [C. goodwini, S. veraepacis ibarrai]
18. El Limo, 1475 m (15º11’22”N, 89º25’08”W; Fig. 22); ca. 9.5 km NW of Gualán, Zacapa Dept. (11–15 July 2007). This site was on a former hillside coffee plantation that at the time supported an early successional secondary forest with abundant tree ferns, Cecropia, and patches of a red-flowered Heliconia. A few large diameter trees included oak, pine, and liquidambar. Bromeliads and other epiphytes were present, but not in the abundance typical of cloud for-est or humid forest. The ground typically had about 5–10 cm of humus overlying a well-drained, sandy soil. A 3-m-wide, sandy-bottomed stream passed through the property. The locality is within the Subtropical Wet Forest life zone. [C. merriami, S. veraepacis ibarrai]
Cerro Montecristo
19. Trifinio, 1650–1750 m, 2400 m (ca. 14º28’N, 89º21’W; Fig. 23); Chiquimula Dept. (4–7, 10–14 July 2004). The Trifinio-Fraternidad Biosphere Reserve pro-tects an 11,984-ha cloud forest surrounding Punto Trifinio, where the borders of El Salvador, Guatemala, and Hon-duras meet at 2418 m on Cerro Montecristo. Our team worked at two sites on the Guatemalan portion of the mountain. On 4–7 July, 2004, we worked at 1650–1750 m elevation near the community of Caserío El Duraznal (14°30’23”N, 89°22’49”W), Aldea Santa Rosalía, Esqui-pulas, where we captured only C. merriami. From 10 to 14 July 2004, our traplines were based at 2400 m at Punto Trifinio (14°25’10”N, 89°21’ 22”W), Aldea Los Planes, Concepción Las Minas, near the summit of Cerro Mon-tecristo. At the higher site, we captured only C. goodwini. The locality is within the Subtropical Wet Forest life zone. [C. goodwini, C. merriami]
Sierra de Santa Cruz
20. Cerro Seja, 1020 m (15º41’21”N, 89º08’23”W); 4.0 km S, 8.35 km W of La Libertad, Izabal Dept. (29–31 July 1994). The locality is within the Subtropical Moist Forest life zone. [C. merriami]
Sierra del Merendón
21. Río Bobos, 600 m (15º21’07”N, 88º43’10”W): 13.45 km S, 12.4 km E Morales, Izabal Dept. (11–15 December 1992). The Sierra del Merendón is also referred to as the Sierra del Espíritu Santo. Our team worked along the Río Bobos in a remant patch of forest in the vicinity of a hydro-electric dam (15º21’53”N, 88º43’32”W). The locality is within the Subtropical Wet Forest life zone. [C. merriami]
22. Cerro Pozo de Agua, 1200 m (15º21’52”N, 88º41’24”W); 12.4 km S, 15.2 km E of Morales, Izabal Dept. (8–13 July 1994). The locality is within the Sub-tropical Wet Forest life zone. [C. merriami]
HondurasSierra de Omoa
23. Cusuco, 1500–1620 m (ca. 15º30’N, 88º13’W); ca. 0.3 km SW of the Visitors Center, Cusuco National Park, Cor-tés Dept. (3–11 December 1994; 16 February 1998). The Sierra de Omoa appears to be a northern extension of the Sierra del Merendón. Our team worked along the Danto Trail (Sendero de Danto), which runs along Río Cusuco for part of its length, and along the Quetzal Trail (Sen- dero de Quetzal). The locality is within the Subtropical Wet Forest life zone. [C. goodwini, C. merriami]

2012 WOODMAN ET AL.—DISTRIBUTIONAL RECORDS OF SHREWS 235
Sierra de Celaque
24. Celaque, 1430 m, 2075 m, 2560 m (14º33’N, 88º39’W); Celaque National Park, ca. 11 km SW of Gracias, Lempira Dept. (11–15 October 1992; 19 January–5 February 1995; 2–19 February 1998). This locality comprises three sites on the eastern slope of Cerro de las Minas, also known as Montaña de Celaque: the Visitors Center; Campamen-to Don Tomás; and Campamento Naranjo. The Celaque National Park Visitors Center, 1430 m (14º33’29”N, 88º38’30”W), was sampled 11–15 October 1992 and 17–19 February 1998. Habitats worked here included the floodplain of the Río Arcagual. This site provided the first record of possible sympatry among C. goodwini, C. mer-riami, and C. orophilus. Campamento Don Tomás, 2075 m (14º32’40”N, 88º39’40”W), is an established camp three to five hours’ hike up from the Visitors Center. A small wooden shelter sat in a hectare-sized former cornfield that had subsequently grown up in a monoculture of alder (Al-nus). Surrounding this was good cloud forest. This site, which was visited 2–15 February 1998, yielded eight C. goodwini, all of which were taken along a small stream lined with mosses and liverworts. In the El Niño year of 1998, this stream was reduced to a mere trickle. A Caco-mistle [Bassariscus sumichrasti (Saussure, 1860)] passed through camp on 5 February 1998. Campamento Naranjo, 2560 m (14º33’N, 88º40’W), is located near the top of the eastern slope of Cerro de las Minas. We worked here in January and February 2005, during which we obtained the first record of Sorex salvini from Honduras, along with eight Cryptotis goodwini. When we visited again 4–14 February 1998, during an El Niño year, we obtained no shrews. The locality is within the Subtropical Lower Mon-tane Moist Forest life zone. [C. goodwini, C. merriami, C. orophilus, S. salvini]
Montaña de Santa Barbara
25. Above San José de los Andes, ca. 1900 m (ca. 14º54’N, 88º06’W); Parque Nacional de Santa Barbara, Santa Bar-bara Dept. This locality is based on a museum specimen taken on the eastern slopes of Montaña de Santa Barbara, and it was not visited by members of our team. The local-ity is within the Subtropical Lower Montane Wet Forest life zone. [C. goodwini]
Montaña de La Muralla
26. Sendero el Pizote, 1480 m (15º05’54”N, 86º44’W); La Muralla National Park, Olancho Dept. (19–21 October 1992). El Pizote is a primary hiking trail on Montaña de La Muralla. The locality is within the Subtropical Lower Montane Wet Forest life zone. [C. merriami]
ACKNOWLEDGMENTS
For various forms of assistance, we thank J. Bucklin, J.A. Campbell, G.A. Cruz D., L. Barahona, E. Fernández, S.R. Flores R., Fundación Ecologista, H. Galeano, FUNDAECO, L.L. Hodges, J.A. Ibarra, G. Ibarra U., H. Lanier, S. Midence, J. Monzón S., B. Myton, A. Novack, A.J. Paredes, S.G. Pérez C., V. Roth, M. Roth, J.C. Schuster, E.N. Smith, S.P. Thorn, O. Valdéz, and J.R. Valdéz. E. Cano and J.J. Polisar kindly provided access to specimen records; L. Wilkins assisted information re-quests. Special thanks to P.D. Jenkins, R. Portela Miguez, D. Hills, and R. Harbord (BMNH) for access to registers, literature, and specimens, and J. Thackary for access to archives in the BMNH Library. Field work was supported by M. Graham Netting Fund; O’Neil Field Research Fund; National Geographic Society (#6105-98 to TJM); the R.K. Mellon North American Mammal Research Institute of the Carnegie Museum of Natu-ral History; U.S. Food and Drug Administration (to RPE); The Section of Mammals, Carnegie Museum of Natural History; The Department of Biological Sciences, San Jose State University (to JOM); Northern Vir-ginia Community College Professional Development Fund (to RPE); The Nature Conservancy; Campmor Inc.; and the United States Geological Survey. We are grateful to the Departamento de Biología, Universidad Nacional Autónoma de Honduras, Cooperación Hondureña de Forestales (COHDEFOR; V. Archaga, T. García), and Consejo Nacional de Areas Protegidas, Guatemala (CONAP; O.F. Lara, M. García de Solórzano, F. Herrera, E. Flores), for providing collection permits and other valuable assistance. The Unión para Proteger el Bosque Nuboso (UPROBON) graciously granted permission to work in Chelemhá Natural Reserve. Robert P. Reynolds and two anonymous reviewers provided helpful com-ments on previous versions of this manuscript. Any use of trade, product, or firm names is for descriptive purposes only and does not imply en-dorsement by the U.S. government.
LITERATURE CITED
ALVAREz, T. 1963. The recent mammals of Tamaulipas, México. University of Kansas Publications, Museum of Natural History, 14:363–473.
BAIRD, D.D., R.M. TIMM, AND G.E. NORDqUIST. 1983. Reproduction in the arctic shrew, Sorex arcticus. Journal of Mammalogy, 64:298–301.
BERGALLO, H.G., AND W.E. MAGNUSSON. 1999. Effects of climate and food availability on four rodent species in southeastern Brazil. Journal of Mammalogy, 80:472–486.
BUCkNER, C.H. 1966. Populations and ecological relationships of shrews in tamarack bogs of southeastern Manitoba. Journal of Mammalogy, 47:181–194.
BUSH, M.B., AND P.A. COLINVAUx. 1990. A pollen record of a complete glacial cycle from lowland Panama. Journal of Vegetation Science, 1:105–118.
BUSH, M.B., D.R. PIPERNO, P.A. COLINVAUx, P.E. DE OLIVEIRA, L.A. kRISSEk, M.C. MILLER, AND W.E. ROWE. 1992. A 14 300-yr paleo-ecological profile of a lowland tropical lake in Panama. Ecological Monographs, 62:251–275.
CAMPBELL, J.A. 1999. Distribution patterns of amphibians in Middle America. Pp. 111–210, in Patterns of Distribution of Amphibians: A Global Perspective (W.E. Duellman, ed.). The Johns Hopkins University Press, Baltimore
CARLETON, M.D., AND D.G. HUCkABy. 1975. A new species of Peromyscus from Guatemala. Journal of Mammalogy, 56:444–451.
CARRAWAy, L.N. 2007. Shrews (Eulypotyphla: Soricidae) of Mexico. Monographs of the Western American Naturalist, 3:1–91.
CASTOE, T.A., P.T. CHIPPINDALE, J.A. CAMPBELL, L.k. AMMERMAN, AND C.L. PARkINSON. 2003. Molecular systematics of the Middle American jumping pitvipers (Genus Atropoides) and phylogeogra-phy of the Atropoides nummifer complex. Herpetologica, 59:420–431.
CHOATE, J.R. 1970. Systematics and zoogeography of Middle American shrews of the genus Cryptotis. University of Kansas Publications,

236 ANNALS OF CARNEGIE MUSEUM VOL. 80
Museum of Natural History, 19:195–317.CHURCHFIELD, S., V.A. NESTERENkO, AND E.A. SHVARTS. 1999. Food
niche overlap and ecological separation among six species of coex-isting forest shrews (Insectivora: Soricidae) in the Russian Far East. Journal of Zoology, 248:349–359.
CHURCHFIELD, S., AND B.I. SHEFTEL. 1994. Food niche overlap and eco-logical separation in a multi-species community of shrews in the Siberian arctic. Journal of Zoology, 234:105–124.
COLWELL, R.k., G. BREHM, C.L. CARDELúS, A.C. GILMAN, AND J.T. LONGINO. 2008. Global warming, elevational range shifts, and lowland biotic attrition in the wet tropics. Science, 322:258–261.
CROTHER, B.J., J.A. CAMPBELL, AND D.M. HILLIS. 1992. Phylogeny and historical biogeography of the palm pitvipers, genus Bothriechis: biochemical and morphological evidence. Pp. 1–20, in Biology of the Pitvipers (J.A. Campbell and E.D. Brodie, Jr., eds.). Selva, Tyler, Texas.
DíAz DE PASCUAL, A. 1993. Caracterización del habitat de algunas espe-cies de pequeños mamíferos de la selva nublada de Monte Zerpa, Mérida. Ecotropicos, 6:1–9.
———. 1994. The rodent community of the Venezuelan Andes. Polish Ecological Studies, 20:155–161.
DíAz DE PASCUAL, A., AND A.A. DE ASCENçãO. 2000. Diet of the cloud forest shrew Cryptotis meridensis (Insectivora: Soricidae) in the Venezuelan Andes. Acta Theriologica, 45:13–24.
DONNELLy, T.W., G.S. HORNE, R.C. FINCH, AND E. LOPéz-RAMOS. 1990. Northern Central America; the Maya and Chortis blocks. Pp. 37–765, in The Caribbean Region. (G. Dengo and J.E. Case, eds.). The Geology of North America, Volume H. Geological Society of America, Boulder, Colorado.
DURANT, P., A. DíAz AND A. DíAz DE PASCUAL. 1994. Pequeños mamíferos alto-andinos Mérida-Venezuela. Revista Forestal Latinoamericana, 14/94:103–131.
DUSzyNSkI, D.W., R.P. ECkERLIN, AND T.J. MCCARTHy. 2003. Eimeria species from Cryptotis shrews (Insectivora: Soricidae) with descrip-tion of a new species. Journal of Parasitology, 89:974–977.
ECkERLIN, R.P. 2006. Checklist of the fleas (Siphonaptera) of Guatemala. Pp. 453–456, in Biodiversidad de Guatemala (E.B. Cano, ed.). Universidad del Valle de Guatemala, Guatemala City.
FORSyTH, D.J. 1976. A field study of growth and development of nest-ling masked shrews (Sorex cinereus). Journal of Mammalogy, 57:708–721.
FOx, B.J., AND G.L. kIRkLAND, JR. 1992. An assembly rule for functional groups applied to North American soricid communities. Journal of Mammalogy, 73:491–503.
FRAGOSO, C., AND P. LAVELLE. 1992. Earthworm communities of tropical rain forests. Soil Biology and Biochemistry, 24:1397–1408.
GOLDMAN, E.A. 1951. Biological investigations in Mexico. Smithsonian Miscellaneous Collections, 115:1–476.
GOODWIN, G.G. 1932. Three new Reithrodontomys and two new Peromyscus from Guatemala. American Museum Novitates, 560:1–5.
HALL, E.R. 1981. Mammals of North American, Second Edition. John Wiley and Sons, New York.
HALL, E.R., AND W.W. DALqUEST. 1963. The mammals of Veracruz. University of Kansas Publications, Museum of Natural History, 14:165–362.
HEIDEMAN, P.D., AND F.H. BRONSON. 1992. A pseudoseasonal reproduc-tive strategy in a tropical rodent, Peromyscus nudipes. Journal of Reproduction and Fertility, 95:57–67.
HERNáNDEz-BAñOS, B.E., A.T. PETERSON, A.G. NAVARRO-SIGUENzA, AND B.P. ESCALANTE-PLIEGO. 1995. Bird faunas of humid montane forests of Mesoamerica: biogeographic patterns and priorities for conservation. Bird Conservation International, 5:252–277.
HOLDRIDGE, L.R. 1947. Determination of world plant formations from simple climatic data. Science, 105:367–368.
HUTTERER, R. 1980. A record of Goodwin’s shrew, Cryptotis goodwini, from Mexico. Mammalia, 44:413.
———. 2005. Order Soricomorpha. Pp. 220–311, in Mammal Species of the World: A Taxonomic and Geographic Reference, Third Edition
(D.E. Wilson and D.M. Reeder, eds.). Johns Hopkins University Press, Baltimore.
ICzN [INTERNATIONAL COMMISSION ON zOOLOGICAL NOMENCLATURE]. 2006. Opinion 2164 (Case 3328). Didelphis Linnaeus, 1758 (Mammalia, Didelphidae): gender corrected to feminine, and Cryptotis Pomel, 1848 (Mammalia, Soricidae): gender fixed as masculine. Bulletin of Zoological Nomenclature, 63:282–283.
INNES, D.G.L., J.F. BENDELL, B.J. NAyLOR, AND B.A. SMITH. 1990. High densities of the Masked Shrew, Sorex cinereus, in Jack Pine plantations in Northern Ontario. American Midland Naturalist, 124:330-341.
ISLEBE, G.A., AND H. HOOGHIEMSTRA. 1997. Vegetation and climate history of montane Costa Rica since the last glacial. Quaternary Science Reviews, 16:589–604.
JACkSON, H.H.T. 1928. A taxonomic review of the American long-tailed shrews. North American Fauna, 51:1–238.
JUNGE, J.A., AND R.S. HOFFMANN. 1981. An annotated key to the long-tailed shrews (genus Sorex) of the United States and Canada, with notes on Middle American Sorex. Occasional Paper of the Museum of Natural History, The University of Kansas, 94:1–48.
kEIRANS, J.E., AND R.P. ECkERLIN. 2005. Description of the male and redescription of the female of Ixodes (Ixodes) guatemalensis and description of the male, nymph and larva of Ixodes (Afrixodes) moreli and redescription of the female (Acari: Ixodida: Ixodidae). International Journal of Acarology 31:259–268.
kIRkLAND, G.L., JR. 1991. Competition and coexistence in shrews (Insectivora, Soricidae). In The Biology of the Soricidae (J.S. Findley and T.L. Yates, eds.). Special Publication, The Museum of Southwestern Biology, University of New Mexico, 1:15–22.
kIRkLAND, G.L., JR., AND P.k. SHEPPARD. 1994. Proposed standard pro-tocol for sampling small mammal communities. In Advances in the Biology of Shrews (J.F. Merritt, G.L. Kirkland, Jr., and R.K. Rose, eds.). Special Publication of Carnegie Museum of Natural History, 18:277–283.
LEWIS, R.E., AND R.P. ECkERLIN. 2004. A new species of Hystrichopsylla Taschenberg, 1880 (Siphonaptera: Hystrichopsyllidae) from Guatemala. Proceedings of the Entomological Society of Washington, 106:757–760.
LEyDEN, B.W., M. BRENNER, D.A. HODELL, AND J.H. CURTIS. 1993. Late Pleistocene climate in the Central American lowlands. Geophysical Monographs, 78:165–178.
MAGA [MINISTERIO DE AGRICULTURA, GANADERíA y ALIMENTACIóN]. 2001. Mapa de zonas de vida Holdridge. República de Guatemala. Ministerio de Agricultura, Ganadería y Alimentación. Unidad de Políticas e Información Estratégica, Guatemala City, Guatemala.
MARkGRAF, V. 1989. Palaeoclimates in Central and South America since 18,000 BP based on pollen and lake-level records. Quaternary Science Reviews, 8:1–24.
MATSON, J.O., AND T.J. MCCARTHy. 2005. A new subspecies of Verapaz shrew (Sorex veraepacis) from Guatemala. In Advances in the Biology of Shrews II (J.F. Merritt, S. Churchfield, R. Hutterer, and B.I. Sheftel, eds.). Special Publication of the International Society of Shrew Biologists, 1:63–70.
MATSON, J.O., N. ORDóñEz-GARzA, W. BULMER, AND R.P. ECkERLIN. 2012. Small mammal communities in the Sierra de los Cuchumatanes, Huehuetenango, Guatemala. Mastozoología Neotropical, 19:71–84.
MACVEAN, C., AND J.C. SCHUSTER. 1981. Altitudinal distribution of passalid beetles (Coleoptera, Passalidae) and Pleistocene dispersal on the volcanic chain of northern Central America. Biotropica, 13:29–38.
MCCAIN, C.M., AND R.k. COLWELL. 2011. Assessing the threat to mon-tane biodiversity from discordant shifts in temperature and precipi-tation in a changing environment. Ecology Letters, 14:1236–1245.
MOCk, O.B., AND C.H. CONAWAy. 1976. Reproduction of the least shrew (Cryptotis parva) in captivity. Pp. 59–74, in The Laboratory Animal in the Study of Reproduction (Th. Anikatzides, S. Erichsen, and A. Spiegel, eds.). Gustav Fischer Verlag, New York.
MURIE, A. 1935. Mammals from Guatemala and British Honduras. Miscellaneous Publication of the Museum of Zoology, University

2012 WOODMAN ET AL.—DISTRIBUTIONAL RECORDS OF SHREWS 237
of Michigan, 26:1–30. REID, F.A. 2009. A Field Guide to the Mammals of Central America
and Southeast Mexico. Second edition. Oxford University Press, New York.
ROBERTSON, P.B., AND E.A. RICkART. 1975. Cryptotis magna. Mammalian Species, 61:1–2.
SAVAGE, J.M. 1982. The enigma of the Central American herpetofauna: dispersals or vicariance? Annals of the Missouri Botanical Garden, 69:464–597.
———. 2002. The Amphibians and Reptiles of Costa Rica. University of Chicago Press, Chicago.
SULLIVAN, J.k., E. ARELLANO, AND D. ROGERS. 2000. Comparative phy-logeography of Mesoamerican highland rodents: concerted versus independent response to past climatic fluctuations. The American Naturalist, 155:755–768.
SULLIVAN, J.k., J.A. MARkERT, AND C.W. kILPATRICk. 1997. Phylogeography and molecular systematics of the Peromyscus aztecus species group (Rodentia: Muridae) inferred using parsimony and likelihood. Systematic Biology, 46:426–440.
TRENBERTH, k.E., J.M. CARON, D.P. STEPANIAk, AND S. WORLEy. 2002. Evolution of El Niño–Southern Oscillation and global atmospheric surface temperatures. Journal of Geophysical Research, 107(D8, 4065): 17pp (doi: 10.1029/2000JD000298). Available from http://www.agu.org/
VázqUEz, L.B., R.A. MEDELLíN, AND G.N. CAMERON. 2000. Population and community ecology of small rodents in montane forest of west-ern Mexico. Journal of Mammalogy, 81:77–85.
WAkE, D.B. 1987. Adaptive radiation of salamanders in Middle American cloud forests. Annals of the Missouri Botanical Garden, 74:242–264.
WATSON, D.M., AND A.T. PETERSON. 1999. Determinants of diversity in a naturally fragmented landscape: humid montane forest avifaunas of Mesoamerica. Ecography, 22:582–589.
WHITAkER, J.O., JR. 1974. Cryptotis parva. Mammalian Species, 43:1–8.WHITAkER, J.O., JR., AND W.J. HAMILTON, JR. 1998. Mammals of the
Eastern United States, Third Edition. Cornell University Press, Ithaca.
WILSON, D.E., AND F.R. COLE. 2000. Common Names of Mammals of the World. Smithsonian Institution Press, Washington.
WOODMAN, N. 1995. Morphological variation between Pleistocene and Recent samples of Cryptotis (Insectivora: Soricidae) from the Yucatan Peninsula, Mexico. Journal of Mammalogy, 76:223–231.
———. 2000. Cryptotis merriami Choate in Costa Rica: syntopy with Cryptotis nigrescens (Allen) and possible character displacement. Caribbean Journal of Science, 36:289–299.
———. 2002. A new species of small-eared shrew from Colombia and
Venezuela (Mammalia: Soricomorpha: Soricidae: Genus Cryptotis). Proceedings of the Biological Society of Washington, 115:249–272.
———. 2003. A new small-eared shrew of the Cryptotis nigrescens-group from Colombia (Mammalia: Soricomorpha: Soricidae). Proceedings of the Biological Society of Washington, 116:853–872.
———. 2005. Evolution and biogeography of Mexican small-eared shrews of the Cryptotis mexicana-group (Insectivora: Soricidae). Pp. 523–534, in Contribuciones Mastozoológicas en Homenaje a Bernardo Villa (V. Sánchez-Cordero and R.A. Medellín, eds.). Instituto de Biología and Instituto de Ecología, Universidad Nacional Autónoma de México, and CONABIO, Mexico City.
———. 2010. Two new shrews (Soricidae) from the western highlands of Guatemala. Journal of Mammalogy, 91:566–579.
———. 2011a. Patterns of morphological variation among semi-fosso-rial shrews in the highlands of Guatemala, with the description of a new species (Mammalia, Soricomorpha, Soricidae). Zoological Journal of the Linnean Society, 163:1267–1288.
———. 2011b. Nomenclatural notes and identification of small-eared shrews (Mammalia: genus Cryptotis) from Cobán, Guatemala, in The Natural History Museum, London. Proceedings of the Biological Society of Washington, 124:249–258.
WOODMAN, N., AND D.A. CROFT. 2005. Fossil shrews from Honduras and their significance for late glacial evolution in body size (Mammalia: Soricidae: Cryptotis). Fieldiana: Geology, 51:1–30.
WOODMAN, N., AND A. DíAz DE PASCUAL. 2004. Cryptotis meridensis. Mammalian Species, 761:1–5.
WOODMAN, N., AND R.B. STEPHENS. 2010. At the foot of the shrew: manus morphology distinguishes closely-related Cryptotis good-wini and Cryptotis griseoventris (Mammalia, Soricidae) in Central America. Biological Journal of the Linnean Society, 99:118–134.
WOODMAN, N., AND R.M. TIMM. 1992. A new species of small-eared shrew, Genus Cryptotis (Insectivora: Soricidae), from Honduras. Proceedings of the Biological Society of Washington, 105:1–12.
———. 1993. Intraspecific and interspecific variation in the Cryptotis nigrescens species complex of small-eared shrews (Insectivora: Soricidae), with the description of a new species from Colombia. Fieldiana: Zoology, 74:1–30.
———. 1999. Geographic variation and evolutionary relationships among broad-clawed shrews of the Cryptotis goldmani-group (Mammalia: Insectivora; Soricidae). Fieldiana: Zoology, 91:1–35.
yU, H.-T., T.-W. CHENG, AND W.-H. CHOU. 2001. Seasonal activity and reproduction of two syntopic white-toothed shrews (Crocidura attenuata and C. kurodai) from a subtropical montane forest in central Taiwan. Zoological Studies, 40:163–169.

Copyright © 2022 FDOKUMEN



















