
Molecular targets on mast cells and basophils for novel therapies

Digestive Diseases and Sciences, Vol. 40, No. 2 (Februa O, 1995), pp. 357-365
Distribution of Mast Cells in Human Ileocecal Region
STEFANO BACCI, DSc, MARIA SIMONETTA FAUSSONE-PELLEGRINI, DSc, BERND MAYER, PhD, and PAOLO ROMAGNOLI, MD
The number and histochemistry of mast cells were analyzed in surgical specimens of the ileocecal junction and neighboring intestinal segments. All the basophilic cells contained tryptase and some were immunoreactive for chymase, vasoactive intestinal polypeptide, or nitric oxide synthase. The medium density of mast cells per square millimeter was 31.90, 110.38, 72.83, 29.80, and 32.70, in the mucosa, submucosa, inner circular, outer circular, and longitudinal muscle layers, respectively. Mast cell density was higher at the ileocecal junction (for all layers together, 79.29 mast cells/mm a) than elsewhere (mast cells/mm2: ileum, 52.29; cecum, 59.22; cecocolonic junction, 54.65; ascending colon, 48.63). The differences among layers and among segments were significant and might be due to layer- and region-specific mast cell roles. Mast cell richness in the muscle coat, especially in the inner circular muscle layer, might be important in regulating its motility.
KEY WORDS: colon; gastrointestinal motility; histology; intestine; man; sphincters.
Mast cells (MC) are present in connective tissues of all mammals and their presence in the wall of the alimentary canal is well known (1-6). These cells secrete paracrine mediators (7, 8), ie, molecules that act in proximity of the cells that release them and are rapidly inactivated by extracellular enzymes (9). Since mediators released by MC may regulate mucosal se- cretion, absorption, and local blood flow (10), partic- ular emphasis has been given so far to the presence of MC in human intestinal mucosa and submucosa in normal and pathological conditions (11-22).
Some secretory products of MC may also be in- volved in the control of intestinal motility, directly or
Manuscript received January 31, 1994; accepted August 22, 1994. From the Department of Human Anatomy and Histology, Sec-
tion of Histology "E. Allara," University of Florence, Italy; and Institute of Pharmacology and Toxicology, University of Graz, Austria.
This research was supported by the Italian Ministry of Univer- sity, Science and Technology ("University Funds-40%" to P.R., "University Funds-60%" to M.S.F.-P. and P.R.) and Fonds zur F6rderung der Wissenschaftlichen Forschung in Osterreich (P 8836 to B.M.).
Address for reprint requests: Dr. P. Romagnoli, Department of Human Anatomy and Histology, Section of Histology "E. Allara," Viale G. Pieraccini 6, 1-50139 Firenze, Italy.
via the enteric nervous system (15, 23-28). Histamine, in particular, stimulates propulsive movements (29), whereas vasoactive intestinal polypeptide (VIP) and nitric oxide (NO) cause muscle relaxation (30, 31). Notwithstanding the possible MC influence on gut motility, only a little information is available on the presence of these cells within the intestinal muscle coat in normal and pathological conditions. Befus et al (5) described the presence of MC in the muscle coat of the human small and large bowel and stated that their density was much lower there than in the mucosa and submucosa.
Alterations in MC density within the muscle coat of the alimentary canal have been found in pathological conditions, as compared to healthy subjects. Numer- ous MC have been found in the intestine of rats sensitized against an antigen (32-34) or infested by nematodes (35), in sensitized guinea pigs (36), and, in man, in cases of hypertrophic esophagus (37) and jejunal infection by nematodes (38). The nature of basophilic cells in these conditions has not always been checked by tryptase histochemistry, which allows
Digestive Diseases aTwl Sciences, Vol. 40, No. 2 (Febma~ 1995) 0163.2110/95/(120t)-03575(17.50/0 © 1995 Plenum Publishing Corporation
357

for distinction between M C [which are rich in this enzyme (39)] and basophils [which are not (40)].
No information has been provided on the presence of MC in the muscle coat of the i leocecal junction. Since this junction is responsible for the regulat ion of intestinal content flow between the small and large bowel (41) and separates areas with different absorp- tive and secretory activities and different microflora, it would be of interest to know whether these cells are present in different numbers, or differently distrib- uted, in the ileocecal junction as compared with the adjacent ileum and colon. Fur the rmore , no a t tempt has been made until now to analyze the MC sepa- rately for each layer of the muscle coat, where an inner circular, an outer circular, and a longitudinal muscle layer, markedly different from each o ther in both morphology (42-44) and function (45, 46), are present. Since knowledge on the distr ibution of M C in baseline condit ions is a necessary premise for any study in pathology, we have addressed this issue in the uninvolved iteocecal junction removed from patients with colon cancer in remote locations and, for com- parison, in the neighboring intestinal segments. The analysis was extended by identifying some immuno- reactive molecules possibly contained in mast cells and important for the function of this region.
MATERIALS AND M E T H O D S
Reagents. Alcian blue and toluidine blue were from Gurr, Poole, England; tetramethylrhodamine isothiocya- hate-labeled avidin was purchased from Immunotech, Mar- seille, France; mouse anti-tryptase and anti-chymase mono- clonal primary antibodies were purchased from Chemicon International, Temecula, California; rabbit anti-VIP poly- clonal antibodies were from Amersham, Milan, Italy; rabbit anti-NO synthase polyclonal antibodies have been pro- duced by one of us (B.M.); goat anti-mouse or anti-rabbit polyclonal antibodies, alkaline phosphatase-linked Extravi- din, naphthol AS-BI phosphate, new fuchsin (triaminotri- tolylmethane), and all other reagents were from Sigma, Milan, Italy.
Specimens. We have studied the ileocecal region of six patients, four males and two females, with ages ranging between 50 and 60 years, who had undergone right hemi- colectomy for nonstenosing carcinoma of the transverse colon. Neither sign of disease nor muscle hypertrophy were found macro- and microscopically in the ileocecal region, which was far beyond the limits of cancer invasion. Since the whole region was not available for investigation in all patients, the number of cases studied for each segment is indicated in parentheses. The surgical specimens were cut into segments as follows: (1) the last 3-5 cm of terminal ileum (prejunctional ileum; six cases); (2) ileocecal junction (five cases); (3) cecum (four cases); (4) cecocolonic junction (five cases); and (5) the first 3-5 cm of ascending colon (postjunctional colon; five cases). Surgery was performed in
BACCI ET AL
all cases with radical intention; Italian law and the ethical guidelines of the Italian National Medical Council were followed throughout the clinical and laboratory procedures.
Histochemistry. Blocks of tissue from each patient were fixed in Bouin's (47) and blocks from two patients in Mota's (2) and Carnoy's fluids (47) and embedded in paraplast. For bright-field microscopy, sections were stained either with 0.1% toluidine blue in 0.02 mol/liter phosphate buffer, pH 4.3, or 1% Alcian blue in 0.7 mol/liter HCI, pH 0.5. For fluorescence microscopy, sections were stained with an in- direct immunohistochemical method for tryptase, chymase (39, 48, 49), VIP, and nitric oxide synthase (NOs) either alone or preceded by tetramethylrhodamine isothiocya- nate-labeled avidin (50), which was diluted 1:400 in 1% albumin. Some sections were exposed to ultraviolet radia- tion to abolish autofluorescence before fluorochromization; other sections were stained without ultraviolet exposure. Some granulocytes in the mucosa fluoresced green even in unlabeled specimens and never fluoresced upon ultraviolet radiation. Therefore, that signal was interpreted as autofluorescence, presumably of eosinophils. Primary mouse monoclonal antibodies anti-tryptase and anti- chymase were diluted up to a protein concentration of 1.5 mg/ml and applied at 4 ° C (39-48). Primary rabbit poly- clonal antibody anti-VIP was diluted 1:1000 and applied at 4 ° C. Primary rabbit polyclonat antibody anti-NOs was di- luted 1:100 and applied at room temperature. All primary antibodies were diluted with Tris-buffered saline; 1% (w/v) albumin was added to anti-VIP and anti-NOs antibodies, 3% (v/v) Triton X-100 was added to anti-NOs antibodies. Anti-mouse or anti-rabbit polyclonal goat antibodies, as appropriate, were used as secondary antibodies. The sec- ondary antibodies were either fluorescein isothiocyanate- labeled or biotinylated and in turn revealed by Extravidin- alkaline phosphatase followed by enzyme histochemistry using naphthol AS-BI phosphate and new fuchsin (48). Fluorochrome-stained sections were examined with a Zeiss Axioskop microscope equipped for epifluorescence. The specificity of the immunostaining was tested by omitting the first antibody or substituting it with an irrelevant one.
Morphometry and Statistics. Mast cells were counted by one of the authors (S.B.) in sections from each tissue block, stained with toluidine blue, at a magnification of ×250. In some instances, MC were counted also upon staining with fluorescent avidin. In all cases, specimens fixed in Bouin's fluid were used for counts; when available, specimens fixed in Mota's fluid were also counted. The counts were separate for each layer (mucosa, submucosa, inner circular, outer circular, and longitudinal muscle layers). The MC of the myenteric plexus were counted together with those of the outer circular muscle layer. A total of 22 mm 2 were scanned; the scanned surface area for each segment and each layer of each patient ranged from 0.0375 to 0.725 mm 2. The number of MC per unit section surface area was indicated as MC density and expressed as number of cells per square millimeter. The data from each layer of each segment and each patient were pooled together and con- sidered to be a sample unit. For statistical analysis, MC densities were transformed into their logarithms, to keep the error variance low. Therefore, median (not mean) val- ues and their 95% confidence limits are indicated in the results; the confidence limits are asymmetrically distributed
358 Digestive Diseases and Sciences, Vol. 40, No. 2 (February 1995)
MAST CELLS IN HUMAN ILEOCECAL REGION
++:++< °.~+.+,+ , I i , , , I ~ +L
' +~>. b +,,,~,,W,.~ll - • m ++" •
• "+ ++
+ t
,,++. , .+*.,,+ .+,s, ~ , ~ , .+.~_. " +;_+ + + ~' q
~ " ' , - _ . • *+,,.,n + • • + +++
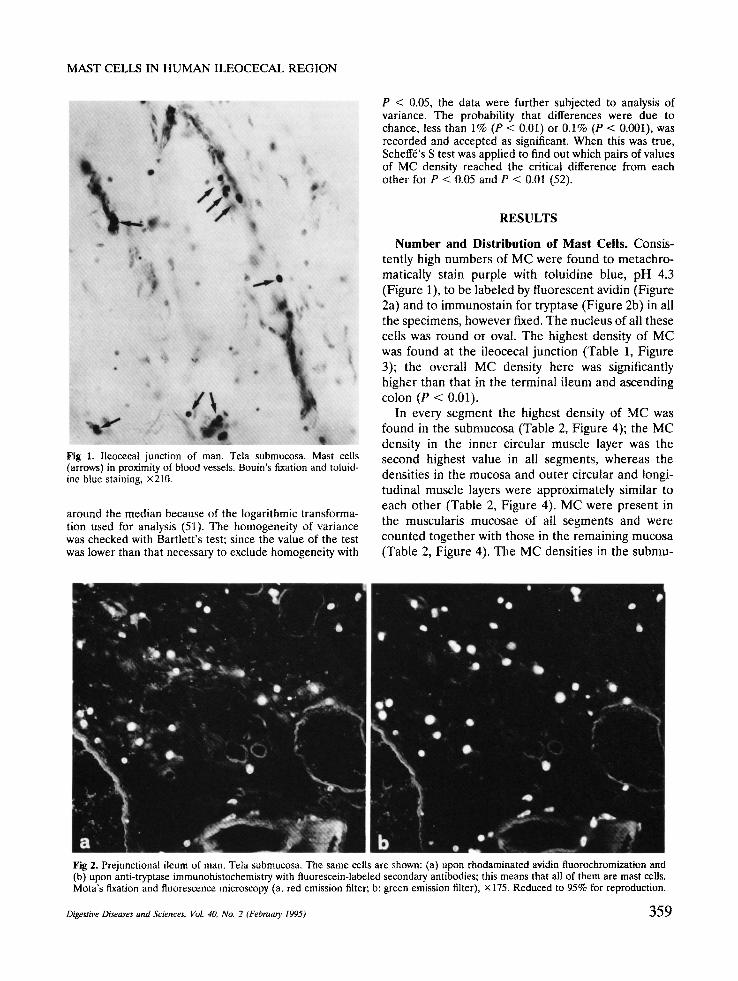
F i g 1. lleocecal junction of man. Tela submucosa. Mast cells (arrows) in proximity of blood vessels. Bouin's fixation and toluid- ine blue staining, ×210.
around the median because of the logarithmic transforma- tion used for analysis (51). The homogeneity of variance was checked with Bartlett's test; since the value of the test was lower than that necessary to exclude homogeneity with
P < 0.05, the data were further subjected to analysis of variance. The probability that differences were due to chance, less than 1% (P < 0.01) or 0.1% (P < 0.001), was recorded and accepted as significant. When this was true, Scheff6's S test was applied to find out which pairs of values of MC density reached the critical difference from each other for P < 0.05 and P < 0.01 (52).
RESULTS
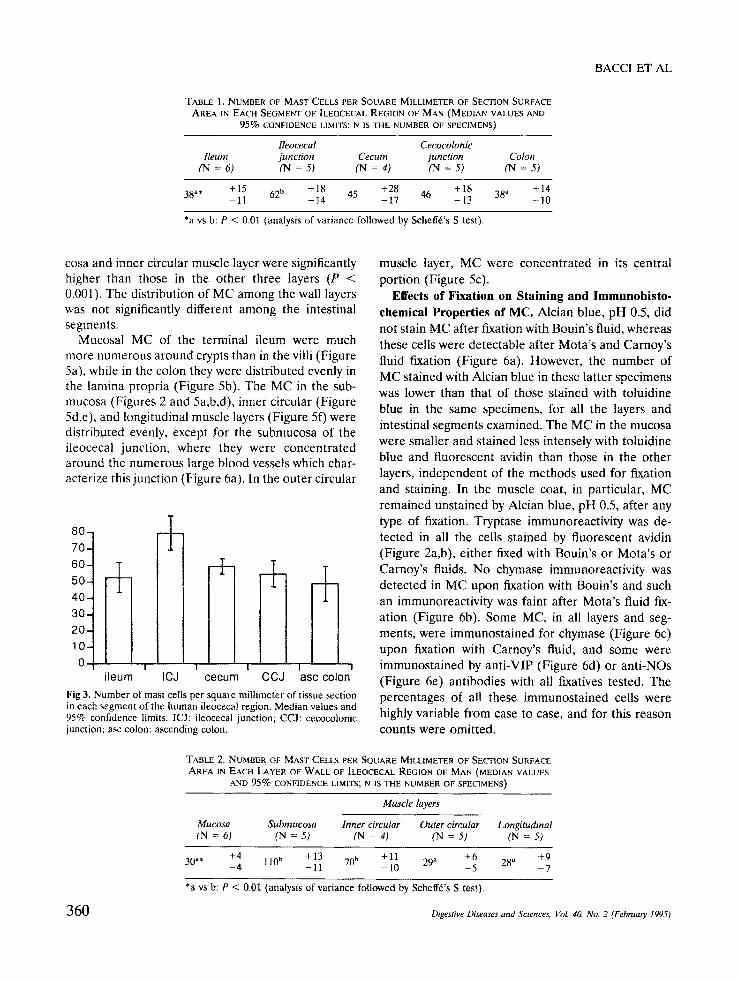
N u m b e r and Dis tr ibut ion of M a s t Cells . Consis- tently high numbers of MC were found to metachro- matically stain purple with toluidine blue, pH 4.3 (Figure 1), to be labeled by fluorescent avidin (Figure 2a) and to immunostain for tryptase (Figure 2b) in all the specimens, however fixed. The nucleus of all these cells was round or oval. The highest density of MC was found at the ileocecal junction (Table 1, Figure 3); the overall MC density here was significantly higher than that in the terminal ileum and ascending colon (P < 0.01).
In every segment the highest density of MC was found in the submucosa (Table 2, Figure 4); the MC density in the inner circular muscle layer was the second highest value in all segments, whereas the densities in the mucosa and outer circular and longi- tudinal muscle layers were approximately similar to each other (Table 2, Figure 4). MC were present in the muscularis mucosae of all segments and were counted together with those in the remaining mucosa (Table 2, Figure 4). The MC densities in the submu-
' # • e ~P @ o Q 41~ 0 ql •
.4 I1 ~ 'Ira, ", ql
~'++O • q l ' o * j O • Q +,u, b
• ' ~ • Ill • " + O O qlt " •
,it, ~@ ,,, • g Q l l + .. ,,
• 41, • +\ • • • l i t
° ++ e';w . . ~ o .~f.at, ~1t ~ "
+" b • " II
F i g 2. Prejunctional ileum of man. Tela submucosa. The same cells are shown: (a) upon rhodaminated avidin fluorochromization and (b) upon anti-tryptase immunohistochemistry with fluorescein-labeled secondary antibodies; this means that all of them are mast cells. Mota's fixation and fluorescence microscopy (a: red emission filter; b: green emission filter), × 175. Reduced to 95% for reproduction.
Digestive Diseases and Sciences, Vol. 40, No. 2 (Febn,a~ 1995) 359
BACCI ET AL
TABLE l, NUMBER OF MAST CELLS PER SQUARE MILLIMETER OF SECTION SURFACE AREA 1N EACH SEGMENT OF ILEOCECAL REGION OF MAN (MEDIAN VALUES AND
95°~ CONFIDENCE LIMITS; N IS THE NUMBER OF SPECIMENS)
Heocecal Cecocolonic Reum junction Cecum junction Colon
(N = 6) (N = 5) (N = 4) (N = 5) (N = 5)
38a. + 15 62 ~ + 18 45 +28 46 + 18 38 . +14 - 1 1 - 1 4 - 1 7 - 1 3 - 1 0
*a vs b: P < 0.01 (analysis of variance followed by Scheff6"s S test),
cosa and inner circular muscle layer were significantly higher than those in the other three layers (P < 0.001). The distribution of MC among the wall layers was not significantly different among the intestinal segments.
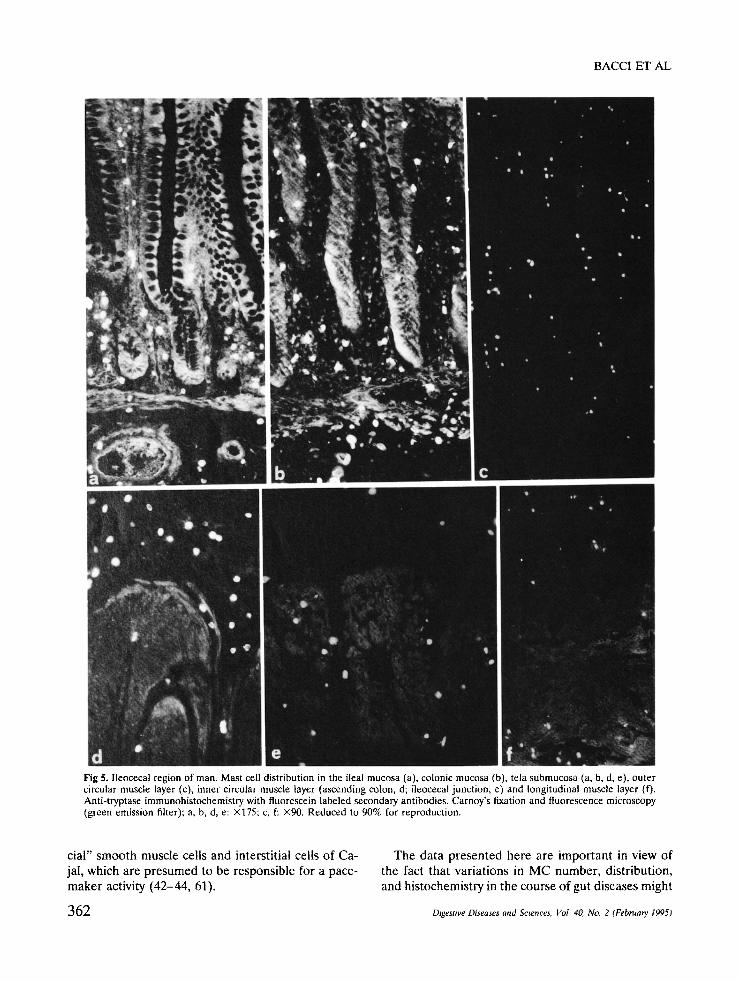
Mucosal MC of the terminal ileum were much more numerous around crypts than in the villi (Figure 5a), while in the colon they were distributed evenly in the lamina propria (Figure 5b). The MC in the sub- mucosa (Figures 2 and 5a,b,d), inner circular (Figure 5d,e), and longitudinal muscle layers (Figure 5f) were distributed evenly, except for the submucosa of the ileocecal junction, where they were concentrated around the numerous large blood vessels which char- acterize this junction (Figure 6a). In the outer circular
80-
70-
60-
50-
40-
30-
20-
10-
0
+ ....... !
+ II U
I I I I i leum ICJ cecum CCJ asc colon
Fig 3. Number of mast cells per square mil l imeter of tissue section in each segment of the human ileocecal region. Median values and 95% confidence limits. ICJ: ileocecal junction; CCJ: cecocolonic junction; asc colon: ascending colon,
muscle layer, MC were concentrated in its central portion (Figure 5c).
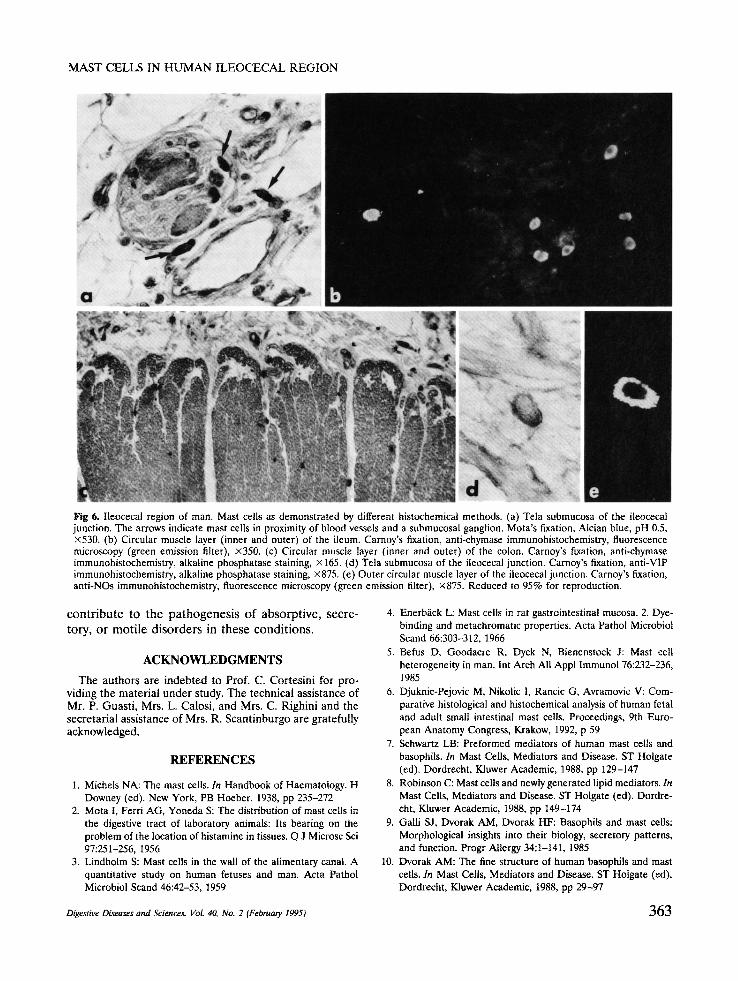
Effects of Fixation on Staining and lmmunohisto- chemical Properties of MC. Alcian blue, pH 0.5, did not stain MC after fixation with Bouin's fluid, whereas these cells were detectable after Mota's and Carnoy's fluid fixation (Figure 6a). However, the number of MC stained with Alcian blue in these latter specimens was lower than that of those stained with toluidine blue in the same specimens, for all the layers and intestinal segments examined. The MC in the mucosa were smaller and stained less intensely with toluidine blue and fluorescent avidin than those in the other layers, independent of the methods used for fixation and staining. In the muscle coat, in particular, MC remained unstained by Alcian blue, pH 0.5, after any type of fixation. Tryptase immunoreactivity was de- tected in all the cells stained by fluorescent avidin (Figure 2a,b), either fixed with Bouin's or Mota's or Carnoy's fluids. No chymase immunoreactivity was detected in MC upon fixation with Bouin's and such an immunoreactivity was faint after Mota's fluid fix- ation (Figure 6b). Some MC, in all layers and seg- ments, were immunostained for chymase (Figure 6c) upon fixation with Carnoy's fluid, and some were immunostained by anti-VIP (Figure 6d) or anti-NOs (Figure 6e) antibodies with all fixatives tested. The percentages of all these immunostained cells were highly variable from case to case, and for this reason counts were omitted.
TABLE 2. NUMBER OF MAST CELLS PER SQUARE MILLIMETER OF SECTION SURFACE AREA IN EACH LAYER OF WALL OF ILEOCECAL REGION OF MAN (MEDIAN VALUES
AND 95% CONFIDENCE LIMITS; N IS THE NUMBER OF SPECIMENS)
Muscle layers
Mucosa Submucosa Inner circular Outer circular Longitudinal (N = 6) (N =5) (N = 4) (N = 5) (N =5)
30"* +4 110b +13 70 b +11 29 . ~-6 28 . +9 - 4 - 1 1 - 1 0 - 5 - 7
*a vs b: P < 0.01 (analysis of variance followed by Scheff6's S test),
360 Digestive Diseases and Sciences, Vol. 40, No, 2 (February 1995)

MAST CELLS IN HUMAN ILEOCECAL REGION
60
4O
r I I I I I
ileum ICJ cecum CCJ asc, colon
Fig 4. Number of mast cells per square millimeter of tissue section in each layer and segment of the human ileocecat region. Median values and 95% confidence limits. ICJ: ileocecal junction; CCJ: cecocolonic junction; asc. colon: ascending colon. Empty column: mucosa; hatched column: submucosa; black column: inner circular muscle layer; gray column: outer circular muscle layer; lightly stippled column: longitudinal muscle layer.
DISCUSSION
In this research, detailed information was obtained on the number and distribution of MC in the ileocecal region of man. We found significant differences in MC density among the segments of this region and among the layers of its wall. In particular, MC were present in higher numbers at the ileocecal junction and in the submucosa than elsewhere, Within the muscle coat they were most numerous in the inner portion of the circular muscle layer than elsewhere in the muscle. The relative differences among segments and layers observed here should be considered inde- pendent of the preoperatory conditions, anesthesia, and surgical procedures, although it cannot be ex- cluded that these factors have influenced the absolute values of MC density.
The presence of unsegmented nuclei and intense tryptase immunoreactivity in all the cells labeled by avidin indicates that all basophilic cells observed were mast ceils and not basophilic granulocytes (7, 53). On the contrary, many basophilic cells were identified as basophilic granulocytes in several alimentary canal diseases, such as hypertrophic esophagus (37). There- fore, the presence of these latter cells in the wall of the alimentary canal can be related to these diseases.
The consistency of the results of toluidine blue staining, fluorescent avidin fluorochromization, and tryptase immunostaining in each specimen, as well as the consistency of the results between the specimens fixed in Bouin's and those in Mota's fluid, give us confidence that our counts are representative of MC densities in the tissue. The utility of tryptase immu- nolabeling to circumvent problems in labeling MC with basic dyes after fixation in formaldehyde-based
Digestive Diseases and Sciences, VoL 40, No. 2 (February 1995)
fluids, like Bouin's, has been stated in previous re- ports (54). Here we show that the same is true for fluorescent avidin. Preservation of the MC of the human intestine by formol based fixatives, such as Bouin's, has been reported by several authors (24, 54, 55). Rather than impairing the fixation of these cells, the use of formol based fixatives influences their staining properties, in particular with toluidine blue at low pH (11, 24). Only Carnoy's fluid allowed good preservation of chymase immunoreactivity; the use- fulness of Carnoy's fluid for this purpose had already been recognized (39). All fixatives used allowed for VIP and NOs immunostaining.
Our immunohistochemical results show that the MC of the human ileocecal region are heterogeneous for the content of several mediators. This may indi- cate that subtypes of MC are present in this region, as elsewhere (39, 49), all of which are tryptase-positive and only some are chymase-, NOs-, or VIP-positive, but the observed heterogeneity might depend also on different maturative or secretory states of these cells. For the mucosa and submucosa, the finding of chy- mase-positive MC confirms previous data found in literature (56); no previous data were found for the MC located in the muscle coat. The findings that some MC can be immunolabeled for VIP and NOs are also new. These findings are in agreement with pharmacological data (57, 58) that indicate a release of NO and VIP by MC.
The presence of these mediators in the MC of the ileocecal region might fit well with the biological roles of all these substances, and the extremely high num- ber of these cells in the submucosa and inner circular muscle layer, especially at the ileocecal junction, might be related to layer- and region-specific roles. In fact, some mediators generated by tryptase activity can produce smooth muscle contraction and in- creased vasopermeability (59), and some generated by chymase activity can cause vasoconstriction (60), while VIP and NO might be involved in muscle coat relaxation (30, 31).
In particular, MC-released mediators, regulating blood vessel caliber, may determine variations in the degree of the submucosal swelling, and, consequently, contribute to the gating function at the level of the ileocecal junction. Furthermore, MC-released medi- ators influencing muscle coat motile behavior may have an important role in regulating gut movements and, more specifically, ileocecal junction opening and closing. The high density of MC in the inner circular muscle layer, especially at the ileocecal junction, may be correlated with the presence in this layer of "spe-
361
BACCI ET AL
Q
D • •
$
• "t" • ~
1 ,m
d " e f : . . , Fig 5. Ileocecal region of man. Mast cell distribution in the ileal mucosa (a), colonic mucosa (b), tela submucosa (a, b, d, e), outer circular muscle layer (c), inner circular muscle layer (ascending colon, d; ileocecal junction, e) and longitudinal muscle layer (f). Anti-tryptase immunohistochemistry with fluorescein labeled secondary antibodies. Carnoy's fixation and fluorescence microscopy (green emission filter): a, b, d, e: ×175; c, f: ×90. Reduced to 90% for reproduction.
cial" smooth muscle cells and interstitial cells of Ca- jal, which are presumed to be responsible for a pace- maker activity (42-44, 61).
362
The data presented here are important in view of the fact that variations in MC number, distribution, and histochemistry in the course of gut diseases might
Digestive Diseases and Sciences', Vol. 40, No. 2 (February 1995)
MAST CELLS IN H U M A N I L E O C E C A L R E G I O N
Fig 6, Ileocecal region of man. Mast cells as demonstrated by different histochemical methods. (a) Tela submucosa of the ileocecat junction. The arrows indicate mast cells in proximity of blood vessels and a submucosal ganglion. Mota's fixation, Alcian blue, pH 0.5, x530. (b) Circular muscle layer (inner and outer) of the ileum. Carnoy's fixation, anti-chymase immunohistochemistry, fluorescence microscopy (green emission filter), x350. (c) Circular muscle layer (inner and outer) of the colon. Carnoy's fixation, anti-chymase immunohistochemistry, alkaline phosphatase staining, x 165. (d) Tela submucosa of the ileocecal junction. Carnoy's fixation, anti-VIP immunohistochemistry, alkaline phosphatase staining, ×875. (e) Outer circular muscle layer of the ileocecal junction. Carnoy's fixation, anti-NOs immunohistochemistry, fluorescence microscopy (green emission filter), x875. Reduced to 95% for reproduction.
contribute to the pathogenesis of absorptive, secre- tory, or motile disorders in these conditions.
ACKNOWLEDGMENTS
The authors are indebted to Prof. C. Cortesini for pro- viding the material under study. The technical assistance of Mr. P. Guasti, Mrs. L Calosi, and Mrs. C. Righini and the secretarial assistance of Mrs. R. Scantinburgo are gratefully acknowledged.
REFERENCES
1. Michels NA: The mast cells. In Handbook of Haematology. H Downey (ed). New York, PB Hoeber, 1938, pp 235-272
2. Mota I, Ferri AG, Yoneda S: The distribution of mast cells in the digestive tract of laboratory animals: Its bearing on the problem of the location of histamine in tissues. Q J Microsc Sci 97:251-256, 1956
3. Lindholm S: Mast cells in the wall of the alimentary canal. A quantitative study on human fetuses and man. Acta Pathol Microbiol Scand 46:42-53, 1959
4. Enerb~ick L: Mast cells in rat gastrointestinal mucosa. 2. Dye- binding and metachromatic properties. Acta Pathol Microbiol Scand 66:303-312, 1966
5. Befus D, Goodacre R, Dyck N, Bienenstock J: Mast cell heterogeneity in man. lnt Arch All Appl Immunol 76:9_32-236, 1985
6. Djuknic-Pejovic M, Nikolic 1, Rancic G, Avramovic V: Com- parative histological and histochemical analysis of human fetal and adult small intestinal mast cells. Proceedings, 9th Euro- pean Anatomy Congress, Krakow, 1992, p 59
7. Schwartz LB: Preformed mediators of human mast cells and basophils. In Mast Cells, Mediators and Disease. ST Holgate (ed). Dordrecht, Kluwer Academic, 1988, pp 129-147
8. Robinson C: Mast cells and newly generated lipid mediators. In Mast Cells, Mediators and Disease. ST Holgate (ed). Dordre- cht, Kluwer Academic, 1988, pp 149-174
9. Galli S J, Dvorak AM, Dvorak HF: Basophils and mast cells: Morphological insights into their biology, secretory patterns, and function. Progr Allergy 34:1-141, 1985
10. Dvorak AM: The fine structure of human basophils and mast cells. In Mast Cells, Mediators and Disease. ST Holgate (ed). Dordrecht, Kluwer Academic, 1988, pp 29-97
Digestive Diseases and Sciences, Vol. 40, No. 2 (February 1995) 363

11. Strobel S, Miller HR, Ferguson A: Human intestinal mucosal mast cells: Evaluation of fixation and staining techniques. J Clin Pathol 34:851-858, 1981
12. Kosnai I, Kuitunen P, Savilahti E, Sipponen P: Mast cells and eosinophils in the jejunal mucosa of patients with intestinal cow's milk allergy and celiac disease of childhood. J Pediatr Gastroenterol Nutr 3:368-372, 1984
13. Fox CC, Dvorak AM, Peters SP, Kagey-Sobotka A, Lichten- stein LM: Isolation and characterization of human intestinal mucosal mast cells. J Immunol 135:483-491, 1985
14. Wingren U, Hallert C, Norrby K, Enerb~ick L: Histamine and mucosal mast cells in gluten enteropathy. Agents Actions 18:266-268, 1986
15. Barrett KE, Metcalfe DD: Mucosal mast cells and lgE. In Immunology of the Gastrointestinal Tract and Liver. MF Hey- worth. AL Jones (eds). New York, Raven Press, 1988, pp 65-92
16. Enerb~ick L: Mucosal mast cells in the rat and in man. Int ~u-ch Allergy Appl lmmunol 82:249-255, 1987
17. Befus AD, Dyck N, Goodacre R, Bienenstock J: Mast cells from the human intestinal lamina propria. Isolation, histo- chemical subtypes, and functional characterization. J Immunol 138:2604-2610, 1987
18. Ferguson J, Thompson RP, Greaves MW: Intestinal mucosal masl ceils: enumeration in urticaria pigmentosa and systemic mastocytosis. Br J Dermatol 119:573-578, 1988
19. Pearce FL: Mast cell heterogeneity. In Mast Ceils, Mediators and Disease. ST Holgate (ed). Dordrecht, Kluwer Academic, 1988, pp 175-193
20. Baum AC, Bhatia P, Miner PB: Increased colonic mucosal mast cells associated with severe watery diarrhea and micro- scopic colitis. Dig Dis Sci 34:1462-1465, 1989
21. Pearce FL, Boulos PB, Lau HY, Liu WL, Tainsh KR: Func- tional heterogeneity of human mast cells. Int Arch Allergy Appl Immunot 94:239-240, 1991
22. King T, Biddle W, Bhatia P, Moore J, Miner PB: Colonic mucosal mast cell distribution at line of demarcation of active ulcerative colitis. Dig Dis Sci 37:490-495, 1992
23. Frieling T, Cooke H J, Wood JD: Histamine receptors on submucous neurons in guinea pig colon. Am J Physiot 264:G74-G80, 1988
24. Stead R, Dixon M, Bramwell N, Riddell R, Bienenstock J: Mast ceils are closely apposed to nerves in the human gastro- intestinal mucosa. Gastroenterology 97:575-585, 1989
9_5. Arizono N, Matsuda S, Hattori T, Kojima T, Maeda T, Galli S J: Anatomical variation in mast cell nerve associations in the rat small intestine, heart, lung, and skin. Similarities of distance between neural processes and mast cells, eosinophils, or plasma cells in the jejunal lamina propria. Lab Invest 62:626- 634, 1990
26. Bienenstock J, Macqueen G, Sestini P, Marshall JS, Stead RH, Perdue MH: Mast cell/nerve interaction in vitro and in vivo. Am Rev Respir Dis 143:$55-$58, 1991
27. Wood JD: Communication between minibrain in gut and en- teric immune system. News Physiol Sci (NIPS) 6:64-69, 1991
28. Wood JD: Histamine signals in enteric neuroimmune interac- tions. Ann NY Acad Sci 664:275-283, 1991
29. Sarna SK, Otterson MF, Cowtes VE, Sethi AK, Telford GL: In vivo motor response to gut inflammation. In Immunomodula- tion of Motility. W Snape, SM Collins (eds). Boca Raton, Florida, CRC, 1991, pp 83-93
30. Grider JR: Identification of neurotransmitters regulating intes-
BACCI E T A L
tinal peristaltic reflex in humans. Gastroenterology 97:1414- 1419, 1989
31. Sanders KM, Ward SM: Nitric oxide as a mediator of nona- drenergic noncholinergic neurotransmission. Am J Physiol 262:G379-G391, 1992
32. Vermillion DL, Ernst PB, Scicchitano R, Collins SM: Antigen induced contraction of jejunal smooth muscle in sensitized rats. Am J Physiol 255:G701-G708, 1988
33. Fargeas M J, Theodourou V, Fioramonti J, Bueno L: Relation- ship between mast cell degranulation and jejunal myoelectric alterations in intestinal anaphylaxis in rats. Gastroenterology 102:157-162, 1992
34. Scott RB, Maric M: Mediation of anaphylaxis-induced jejunal circular smooth muscle contractions in rats. Dig Dis Sci 38:396-402, 1993
35. Marzio L, Blennherhassett P, Vermillion D, Chiverton S, Col- lins S: Distribution of mast cells in intestinal muscle of nema- tode sensitized rats. Am J Physiot 262:G477-G482, 1992
36. Barnette MS, Grous M: Characterization of the antigen in- duced contraction of colonic smooth muscle from sensitized guinea pigs. Am J Physiol 262:GI44-G149, 1992
37. Tung HN, Schulze-Delrieu K, Shirazi S: Infiltration of hyper- trophic esophageal smooth muscle by mast cells and basophils. J Submicrosc Cytol Pathol 25:93-102, 1993
38. Gustowska L, Ruitenberg El, Elgersma A, Kociecka W: In- crease of mucosal mast cells in the jejunum of patients infected with TrichineUa spiralis. Int Arch Allergy Appl Immunol 71:304-308, 1983
39. Irani AM, Schechter NM, Craig SS, De Blois G, Schwartz LB: Two types of human mast cells that have distinct neutral protease composition. Proc Natl Acad Sci USA 83:4464-4468, 1986
40. Galli S J: New insights into the riddle of mast cells. Microen- vironmental regulation of mast cell development and pheno- typic heterogeneity. Lab Invest 62:5-33, 1990
41. Phillips SF, Quigley EM, Kumar D, Karnath PS: Motility of the ileocolonic junction. Gut 29:390-406, 1988
42. Faussone-Pellegrini MS, Cortesini C: Some ultrastructural fea- tures of the muscular coat of human small intestine. Aeta Anat 115:47-68, 1983
43. Faussone-Pellegrini MS, Cortesini C, Pantalone D: Neuromus- colar structures specific to the submucosal border of the human colonic circular muscle layer. Can J Physiol Pharmacol 68:1437-1446, 1990
44. Faussone-Pellegrini MS, Pantalone D, Cortesini C: Morpho- logical evidence for a cecocolonic junction in man and func- tional implications. Acta Anat 146:22-30, 1993
45. Couturier D, Roze D, Couturier-Turpin MH, Debray C: Elec- tromyography of the colon in situ: A n experimental study in man and in the rabbit. Gastroenterology 56:317-322, 1969
46. Dapoigny M, Trolese JF, Bommelaer G, Tournur R: Myoelec- tric spiking activity of right colon, left colon, and rectosigmoid of healthy humans. Dig Dis Sci 33:1007-1012, 1988
47. Pearse AG: Histochemistry. Theoretical and Applied. Edin- burgh, Churchill Livingstone, 1980
48. Polak JM, Van Noorden S: Immunocytochemistry. Modern Methods and Application. Bristol, Wright, 1986
49. Irani AM, Bradford TR, Kepley CL, Schechter NM, Schwartz LB: Detection of MCT and MCTC types of human mast cells by immunohistochemistry using new monoclonal anti-tryptase and anti-chymase antibodies. J Histochem Cytochem 37:1509- 1515, t989
364 Digestive Diseases and Sciences, VoL 40, No. 2 (February 1995)

MAST CELLS IN HUMAN ILEOCECAL REGION
50. Tharp MD, Seelig LL, Tigelaar RE, Bergstresser PR: Conju- gated avidin binds to mast cell granules. J Histochem Cyto- chem 33:27-32, 1985
51. Murray GD. Statistical aspects of research methodology. Br J Surg 78:777-781, 1991
52. Kirk RE: Experimental Design: Procedures for the Behavioral Sciences. Monterey, Brooks-Cole, 1982
53. Hunt LW, Colby TW, Weiler DA, Sur S, Butterfield JH: Immunofluorescent staining for mast cells in idiopathic pulmo- nary fibrosis. Quantification and evidence for extracellular release of mast cell tryptase. Mayo Clin Proc 67:941-948, 1992
54. Huntley JF, Newlands GF, Gibson S, Ferguson A, Miller HR: Histochemical demonstration of chymotrypsin like serine es- terases in mucosal mast cells in four species including man. J Clin Pathol 38:375-384, 1985
55. Eliakim F, Gilead L, Ligumsky M, Okon E, Rachmilewitz D, Razin E: Histamine and chondroitin sulfate E proteoglycan released by cultured human colonic mucosa: indication for possible presence of E mast cells. Proc Natl Acad Sci USA 83:461-464, 1986
56. Irani AM, Craig SS, De Blois G, Elson CO, Schechter NM, Schwartz LB: Deficiency of the tryptase positive, chymase
negative mast cell type in gastrointestinal mucosa of patients with defective T lymphocyte function. J Immunol 138:4381- 4386, 1987
57. Masini E, Salvemini D, Pistelli A, Mannaioni PF, Vane JR: Rat mast cells synthetize a nitric oxide like-factor which modulates the release of histamine. Agents Actions 33:61-63, 1991
58. Goetzl E J, Finch RJ, Peterson KE, Sreedharan P, Turch CW: Mast cell and basophil mediation of lymphocyte functions. In Mast Cells, Mediators and Disease. ST Holgate (ed). Dordre- cht, Kluwer Academic, 1988, pp 247-253
59. Reilly CF, Tewksbury DA, Schechter NM, Travis J: Rapid conversion of angiotensin I to angiotensin II by neutrophils and mast cells proteinases. J Biol Chem 257:8619-869-5, 1982
60. Schwartz LB, Kawahara MS, Hugli TE, Vik D, Fearon DT, Austen KF: Generation of C3a anaphylatoxin from human C3 by human mast cell tryptase. J Immunol 130:18991-18996, 1983
61. Faussone-Pellegrini MS, Cortesini C, Romagnoli P: Sull'- ultrastruttura della tunica muscolare della porzione cardiale dell'esofago e dello stomaco umano con particolare riferi- mento alle cosiddette cellute interstiziali del Cajal. Arch Ital Anat Embriol 82:157-177, 1977
Digestive Diseases and Sciences, VoL 40, No. 2 (Febn,ary 1995) 365
Copyright © 2022 FDOKUMEN




















