
Phenotypic and genetic differentiation between native and introduced plant populations
Upload
independentCategory
view
2download
0
Distribution and population dynamics of the introduced seaweed Grateloupiaturuturu (Halymeniaceae, Rhodophyta) along the Portuguese coast
RITA ARAUJO1*, JOSE VIOLANTE
1, RUI PEREIRA1, HELENA ABREU
1, FRANCISCO ARENAS1
AND ISABEL SOUSA-PINTO1,2
1CIMAR/CIIMAR – Interdisciplinary Centre for Marine and Environmental Research, University of Porto, Rua dos Bragas,
289, 4050-123 Porto, Portugal2Department of Botany, Faculty of Sciences, University of Porto, Rua do Campo Alegre, Porto, Portugal
ARAUJO R., VIOLANTE J., PEREIRA R., ABREU H., ARENAS F. AND SOUSA-PINTO I. 2011. Distribution and populationdynamics of the introduced seaweed Grateloupia turuturu (Halymeniaceae, Rhodophyta) along the Portuguese coast.Phycologia 50: 392–402. DOI: 10.2216/10-65.1
Despite the increasing number of coastal areas invaded over the last years by the introduced seaweed Grateloupia
turuturu, studies on this species are scarce worldwide, and its invasive capability and future impacts remain unexplored.In the present study, we describe the geographical distribution of the species after a few years of colonization of thePortuguese continental coastline. Additionally, we examine the structure and dynamics of one of the first population ofGrateloupia in our shores. Seasonal dynamics of standing biomass, density, size structure and inequality (sizevariability), together with the presence of fertile fronds were investigated during one year. Grateloupia was recorded inmidintertidal pools at 11 of the 36 localities sampled, two of which presenting an elevated abundance of fronds. Frondswere present during the entire year, but their size (length and biomass), density and size inequality varied seasonallyreaching highest values at the beginning of the summer. The annual cycle of the population studied included a slow-growth phase in winter, a fast-growth phase during spring-early summer when some density-dependent regulationappeared to occur, as well as a dieback phase at the end of summer. In terms of fertility, abundance of fronds bearingreproductive structures were seasonally variable although carposporophytic and tetrasporophytic fronds were recordedthroughout the year. The results of this work highlight some of the biological characteristics of Grateloupia turuturu thatmay determine its invasive nature and its possible impact on native ecosystems.
KEY WORDS: Geographical distribution, Grateloupia turuturu, Introduced species, Macroalgae, Population dynamics,Portugal
INTRODUCTION
Introductions of nonindigenous species are globally con-
sidered a threat to marine biodiversity, seriously impacting
native ecosystems worldwide (Carlton & Geller 1993; Bax et
al. 2001). In the European seas, more than 800 introduced
species have been reported, mostly in coastal areas
(Streftaris et al. 2005). Marine macroalgae are a significant
component of these introduced marine species, including
several highly prominent species that have caused signifi-
cant ecological and economic impact: e.g. Caulerpa taxifolia
(M. Vahl) C. Agardh, Codium fragile (Suringar) Hariot,
Sargassum muticum (Yendo) Fensholt or Undaria pinnati-
fida (Harvey) Suringar (Schaffelke et al. 2006). For most of
these introduced species there is insufficient information on
their current status and ecology in their new habitats,
preventing the prediction of their future impacts and
adoption of management politics.
The Portuguese coast is not as heavily invaded as other
European shores, and only 11 introduced species have been
recorded in this area (Araujo et al. 2009): Anotrichium
furcellatum (J. Agardh) Baldock, Antithamnionella spiro-
graphidis (Schiffner) E.M. Wollaston, Antithamnionella
ternifolia (J.D. Hooker & Harvey) Lyle, Antithamnion
densum (Suhr) M.A. Howe, Asparagopsis armata (Harvey)
[‘‘Falkenbergia rufolanosa’’ stage (Harvey) F. Schmitz],
Dasya sessilis Yamada, Grateloupia turuturu Yamada,
Gracilaria vermiculophyla (Ohmi) Papenfuss, Neosiphonia
harveyi (J. Bailey) M.-S. Kim, H.-G. Choi, Guiry & G.W.
Saunders, Sargassum muticum and Undaria pinnatifida.
Although the majority of these species were reported at low
local abundances for a few locations, some of them, e.g.
Antithamnionella ternifolia, Grateloupia turuturu and Sar-
gassum muticum, are widely spread along the Portuguese
coast (Araujo et al. 2009). Grateloupia turuturu, a recent
introduction to the Portuguese shores (Araujo et al. 2003),
is quickly becoming more abundant, and despite being
reported worldwide as an invasive species (Villalard-
Bohnsack & Harlin 1997; Verlaque et al. 2005), it is still
poorly studied.
Grateloupia turuturu is native to Japan and has been
reported on the Atlantic coasts of Europe and North
America and in the Mediterranean Sea (Cabioch et al. 1997;
Villalard-Bohnsack & Harlin 1997; Barbara & Cremades
2004). More recently it has been recorded in Australia
(Saunders & Withall 2006) and New Zealand (D’Archino et
al. 2007). Studies developed in France (Cabioch et al. 1997;
Simon et al. 2001), England (Cabioch et al. 1997) and the
United States (Harlin & Villalard-Bohnsack 2001; Villa-
lard-Bohnsack & Harlin 2001) had identified this species as
G. doryphora. However, Gavio & Fredericq (2002) using
comparative morphology and molecular analysis, showed* Corresponding author ([email protected]).
Phycologia (2011) Volume 50 (4), 392–402 Published 13 July 2011
392

that these previous works refer to misidentified specimens
of G. turuturu.
Grateloupia turuturu grows in both sheltered and exposed
areas (Villalard-Bohnsack & Harlin 1997; Simon et al.
2001), can tolerate wide variations of temperature and
salinity (Simon et al. 1999; Harlin & Villalard-Bohnsack
2001) and is well adapted to eutrophic conditions (Simon et
al. 2001). This species shows multiple recruitment strategies
(Harlin & Villalard-Bohnsack 2001) and fertile individuals
occur throughout the year in some locations (Harlin &
Villalard-Bohnsack 2001; Simon et al. 2001; Villalard-
Bohnsack & Harlin 2001).
The sources of introduction of this species are still not
clearly established although some evidence exists relating
Grateloupia turuturu’s introduction with aquaculture activ-
ities (in particular oyster farming), ship fouling and
ballasting (Villalard-Bohnsack & Harlin 1997) and short-
distance dispersal by stone-rafting and drifting of fertile
blades (Simon et al. 1999; Simon et al. 2001).
Whereas a large amount of literature exists reporting
invasions by some marine macroalgal species like Sargas-
sum muticum (Arenas & Fernandez 2000; Staehr et al. 2000;
Plouguerne et al. 2006; Engelen & Santos 2009) or Caulerpa
taxifolia and Caulerpa racemosa (Piazzi et al. 2001; Balata
et al. 2004), studies on Grateloupia turuturu are insufficient
(Harlin & Villalard-Bohnsack 2001) despite its recently
noticed expansion range (D’Archino et al. 2007; Mathieson
et al. 2008) and its possible invasive nature (Villalard-
Bohnsack & Harlin 1997; Verlaque et al. 2005).
The overall impact of introduced species is directly linked
both with their final range of expansion and their local
abundances in the host communities (Parker et al. 1999).
To increase our capabilities for prediction of both the
spread and invasive nature of introduced species, detailed
and relevant information on the ecology of the species in its
new range of distribution is imperative. Moreover, this is
basic information for the implementation of preventive and
management actions.
The objectives of this work were to assess the current
geographical range of Grateloupia turuturu on the Portu-
guese coast and to investigate the structure and dynamics
(reflected in the seasonal variations of population param-
eters), including the different life-cycle phases, of the most
established population of this species in our shores. We
specifically studied the existence of temporal and small-
scale spatial variability in the frond length, biomass,
density, size distribution and size inequality of fronds. We
also examined the existence of significant temporal and
spatial differences in the reproductive status and relative
dominance of life-cycle phases in the population studied.
The biological characteristics of an introduced species are
important predictors of its invasive nature and consequent
impact in native communities.
MATERIAL AND METHODS
Study area
The study of the geographical distribution of Grateloupia
turuturu was conducted in the northern Portuguese coast
(Fig. 1). In this area the tidal regime is semidiurnal,
reaching 3.5–4 m, during spring tides and sea surface
temperature varies between 13 and 20uC during the year.
The study of the structure and dynamics of G. turuturu
populations was performed in rock pools situated in the
midintertidal level, shallower than 1 m and with size
between 1 m2 and 5 m2. Organisms living in these systems
experience unique fluctuations of environmental parame-
ters. Although environmental variations in rock pools are
already documented (Metaxas & Scheibling 1993), a brief
characterization of the environmental parameters fluctua-
tions experienced during low tide in rock pools of the
studied area was performed. From December 2008 to
November 2009, the temperature in one G. turuturu
dominated rock pool was registered hourly using data
loggers (Stow Away Tidbit Temp Logger). Mean annual
temperature was 16.07uC, ranging from a minimum of
8.2uC and a maximum of 28.62uC. Additionally, in three
different dates of sampling (June, August and November),
temperature, salinity and light (under and above canopy)
were registered hourly during the period of low tide in three
of the G. turuturu dominated rock pools (at each hour five
measures were done for salinity and temperature and 10
measures for light under and above canopy). Mean
temperature during low tide was 20uC with maximum
values of 28.9uC and minimum values of 13.7uC. During the
summer (August) temperature amplitude in pools during
low tide reached 7uC, whereas in winter (November)
temperature fluctuations were of 3uC. Mean salinity was
33.82%, with maximum values of 36.7% and minimum
values of 20% (during rainy days). Mean light below
canopy was 45.65 meinstein/m2 (mM), fluctuating between
273 mM and 0.6 mM, while above canopy a mean of
404.16 mM was registered, with maximum values of 886 mM
and minimum values of 15 mM. Light penetration was
reduced in November when compared with August. Mean
values registered above canopy were 760 mM in summer
and 229.03 mM in winter, while under canopy mean light
intensity was 75.67 mM in summer and 42.63 mM in winter.
Geographical distribution
To assess the geographical distribution of Grateloupia
turuturu on the Portuguese coast, 36 sites (all the
independent areas of the rocky shore) along a stretch of
250 km of the northern shore were visited from 2000 to
2007: sites with sparse distribution of fronds were visited
from May to September, but sites with well established
populations were visited at least once in each different
month (Fig. 1). This study was not extended further south
because the species was never reported at any other locality
further than our southernmost sampling locality (Buarcos)
(E. Berecibar, personal communication), with the exception
of a citation by L. Pereira (personal communication) in
Peniche (Fig. 1). At each locality, the intertidal area, from
high to low littoral (until 21 m mean low water), was
thoroughly surveyed (the shore was covered by foot and
visually surveyed) and G. turuturu specimens collected
whenever present. An evaluation of the abundance of
fronds at each site was done, distinguishing places with a
scarce distribution of fronds from localities with well
Araujo et al.: Geographical distribution and population dynamics of Grateloupia turuturu on the Portuguese coast 393

established populations. The relationship between popula-
tions density and distance from major ports was estimated
using the nonparametric Spearman correlation index.
Population structure and dynamics
The structure and dynamics of a population of Grateloupia
turuturu was studied at the most established population
(with higher abundance of fronds), located in Aguda
(41u039250N; 8u399240W). At this locality G. turuturu was
mainly found in intertidal rock pools.
From September 2004 to July 2005, population descrip-
tors such as frond length, biomass, density and size
distribution of fronds were studied bimonthly in three
randomly selected pools. Care was taken to avoid sampling
the same pool twice. In each pool, two 50 3 50 cm quadrats
were randomly chosen and destructively sampled. All the
Grateloupia turuturu specimens inside each quadrat were
scrapped, bagged and preserved in 4% formalin seawater.
In the laboratory, all the fronds collected were counted and
their length measured. Fronds biomass of G. turuturu was
estimated from a length-biomass relationship obtained by
measuring total length and dry weight of each of 127
fronds, collected between March and July. Fronds were
dried in an oven at 50uC for 48 h. The Gini coefficient (G)
was the size-structure descriptor selected for this study
(Bendel et al. 1989). This coefficient has been used in several
studies to describe the total amount of inequality in size of
plant populations (Santos 1995; Arenas & Fernandez 2000;
Damgaard & Weiner 2000). The coefficient ranges from a
minimum value of zero (when all the individuals have the
same size) to a theoretical maximum of one (when all
individuals except one have a size of zero) (Weiner &
Solbrig 1984).
Fig. 1. Study area with the sampling localities marked with numbers. Geographical distribution of Grateloupia turuturu is signalized withbold numbers and dot symbols filled in grey. The year G. turuturu was first observed in each location is included in brackets and theabundance of each population indicated. 1 – Insua de Caminha; 2 – Moledo; 3 – Vila Praia de Ancora; 4 – Forte do Cao; 5 – Montedor; 6 –Areosa; 7 – Viana do Castelo; 8 – Lima’s saltmarsh; 9 – Amorosa; 10 – Castelo do Neiva; 11 – Belinho; 12 – S. Bartolomeu do Mar; 13 – Riode Moinhos; 14 – Cepaes; 15 – Esposende; 16 – Cavado’s saltmarsh; 17 – Apulia; 18 – Agucadoura; 19 – Quiao; 20 – Povoa do Varzim; 21 –Vila do Conde; 22 – Mindelo; 23 – Praia dos Electricos; 24 – Angeiras; 25 – Cabo do Mundo; 26 – Leca da Palmeira; 27 – Foz do Douro;28 – Lavadores; 29 – Valadares; 30 – S. Felix da Marinha; 31 – Miramar; 32 – Aguda; 33 – Espinho; 34 – Ria de Aveiro; 35 – Barra deAveiro; 36 – Buarcos. The occurrence of the species in Peniche (L. Pereira, personal communication) is signalled. Main ports of the northernPortuguese coast are represented.
394 Phycologia, Vol. 50 (4), 2011

To test the existence of temporal and small-scale spatial
variability in the population descriptors studied, univariate
analysis of variance (two-way ANOVA) was performed.
The assumption of homogeneity of variances was tested
using Cochran’s test and, when appropriate, data transfor-
mation was performed (Underwood 1997). Analysis in-
cluded two factors: Time (fixed, with six levels correspond-
ing to each sampling date) and Pool (random, with three
levels).
Reproduction
Grateloupia turuturu exhibits a haploid-diploid life cycle,
including two independent isomorphic reproductive phases
[tetrasporophyte (diploid) and gametophyte (haploid)] and
a third reproductive stage (carposporophyte), developed
within the female gametophyte.
To study the reproductive dynamics of Grateloupia
turuturu, the reproductive status of all the fronds collected
in the 50 3 50 cm plots was determined for all the sampling
occasions. All the new fronds below 5 cm were considered
recruits, but fronds smaller than 5 cm growing from older
damaged fronds were not included in this category.
However, this study was unable to distinguish between
fronds formed from new and old crusts. Female gameto-
phytes/carposporophytes were identified by naked eye,
whenever the fronds were obviously mature. In the case
of tetrasporophytic fronds or when the identification of the
reproductive status was not obvious, cross-sections of the
blade were observed under a dissecting microscope. To test
for temporal and small-scale spatial significant variations in
the reproductive status (number of reproductive and
nonreproductive fronds) and in the life-cycle phase (number
of carposporophytic and tetrasporophytic fronds), univar-
iate analysis of variance (three-way ANOVA) was per-
formed. The assumption of homogeneity of variances was
tested using Cochran’s test and, when appropriate, data
transformation was performed (Underwood 1997). Analy-
sis included three factors: Reproductive status or Life-cycle
phase (fixed, with two levels), Time (fixed, with six levels
corresponding to each sampling date) and Pool (random,
with three levels). Student – Newman – Keuls (SNK) tests
were used for a posteriori comparisons of means. ANOVA
was performed using the GMAV5 software (University of
Sydney).
RESULTS
Geographical distribution
On the Portuguese coast, Grateloupia turuturu occurs
mainly in midintertidal rock pools of both sheltered and
exposed rocky shores, although, in some locations, it was
also found at the low littoral level. G. turuturu was found at
11 of the 36 surveyed localities (Fig. 1). At two of these
localities [Aguda (32) and Foz do Douro (27)] this species
presented elevated abundances (. 25% of percentage cover)
in midintertidal pools. In one locality [Buarcos (36)] the
species was present at intermediate abundances (5–25% of
percentage cover), and at all the other localities sampled,
fronds were sparsely distributed along the shore (, 5% of
percentage cover). The geographical distribution of G.
turuturu on the Portuguese coast was not continuous along
the rocky shore, although suitable substratum (rocky shore)
for its establishment was available along the surveyed
coastline. Instead, G. turuturu populations were mainly
found nearby the main shipping ports of the northern coast
of Portugal (Fig. 1). However, the distance from main ports
(port of Viana do Castelo, port of Leixoes, port of Aveiro
and port of Figueira da Foz) was not significantly
correlated with the density of G. turuturu populations
(Table 1).
Population structure and dynamics
Grateloupia turuturu’s fronds showed a wide variety of
morphological forms from lanceolate (simple or divided) to
fronds with marginal proliferations (Figs 2–5). Blade size
varied seasonally with maximum blade length (72 cm) in
May and July. Frond length was much lower at all the
sampling dates, varying from a minimum of 2.03 cm 6 0.06
(mean 6 SE) in March to a maximum of 10.99 cm 6 0.34
(mean 6 SE) in September. Length and biomass were
positively correlated for G. turuturu fronds and biomass
was calculated, for each frond, based on the following
equation: Ln B 5 27.6206 + 1.9072*Ln L (r 5 0.937, P ,
0.001), where B is frond biomass and L is frond length (N 5
127). Except for the first sampling date, September, the
studied population was dominated by fronds of small size
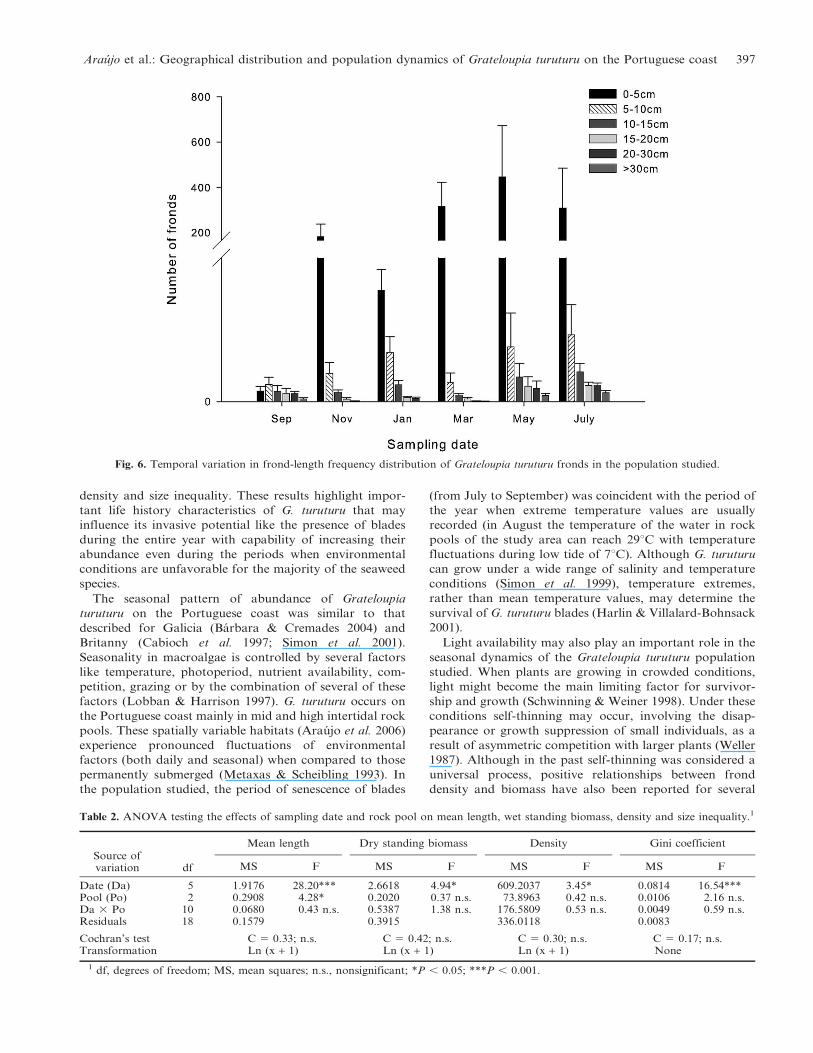
(length , 5 cm) (Fig. 6). This dominance was stronger
between March and July when the percentage of small
fronds averaged 82.16% 6 5.96. During this period the
largest fronds (. 10 cm) were also recorded, representing
around 8% of the population fronds. All the population
descriptors studied differed significantly among sampling
dates (Table 2), and in the case of mean frond length, pool
was also a significant factor (Table 2). No significant
interactions Time 3 Pool were found for any of the
variables studied (Table 2). The annual cycle of the G.
turuturu population studied included different development
phases. During winter (November–March), no significant
changes in biomass, density and Gini coefficient were
recorded (Fig. 7). Minimum values of standing biomass
(1.56 6 0.4 g/m2), density (640 6 141.18 fronds/m2) and
Gini coefficient (0.35 6 0.02) were recorded during this
period (Fig. 7). From March to early summer, a growth
period occurred with significant increases in biomass and
size inequalities. No significant differences were found for
plant density among dates (probably due to the high
variability among quadrats), but the number of plants
Table 1. Spearman correlation between the distance from mainports and population density (Pden) in each of the localitiessampled.
Valid Spearman T (N 2 2) P
Port Viana do Castelo& Pden 11 0.526091 1.85586 0.096448
Port Leixoes & Pden 11 20.104062 20.31389 0.760758Port Aveiro & Pden 11 20.460655 21.55700 0.153896Port Figueira da Foz
& Pden 11 20.526091 21.85586 0.096448
Araujo et al.: Geographical distribution and population dynamics of Grateloupia turuturu on the Portuguese coast 395

progressively increased until May when the maximum
densities were found (maximum mean density 2182.7 6
1126.41 fronds/m2, with a single plot maximum of 7692
fronds/m2). Maximum mean standing biomass was found in
July (13 6 1.46 g/m2), when some of the other population
descriptors like density and size inequality appeared to be
already declining (Fig. 7). Later in the summer (September)
the population was maintained by few old big fronds
(Fig. 6). During this period minimum values of density and
Gini coefficient were recorded simultaneously with low
levels of standing biomass (Fig. 7).
Reproduction
Fronds with reproductive structures were found throughout
the year and were dominant among those larger than 5 cm
long, except for March and May when no significant
differences were found between the number of reproductive
and nonreproductive fronds (Fig. 8) (Table 3). Neverthe-
less, two reproductive peaks were recorded: one in winter
(January) and another one at the beginning of the summer
(July) (Fig. 8). These peaks of abundance of reproductive
fronds were not coincident with the peaks of incorporation
of smallest size individuals (presumably new recruits) into
the population (Figs 6, 8). Mature tetrasporophytes and
female gametophytes/carposporophytes were recorded at
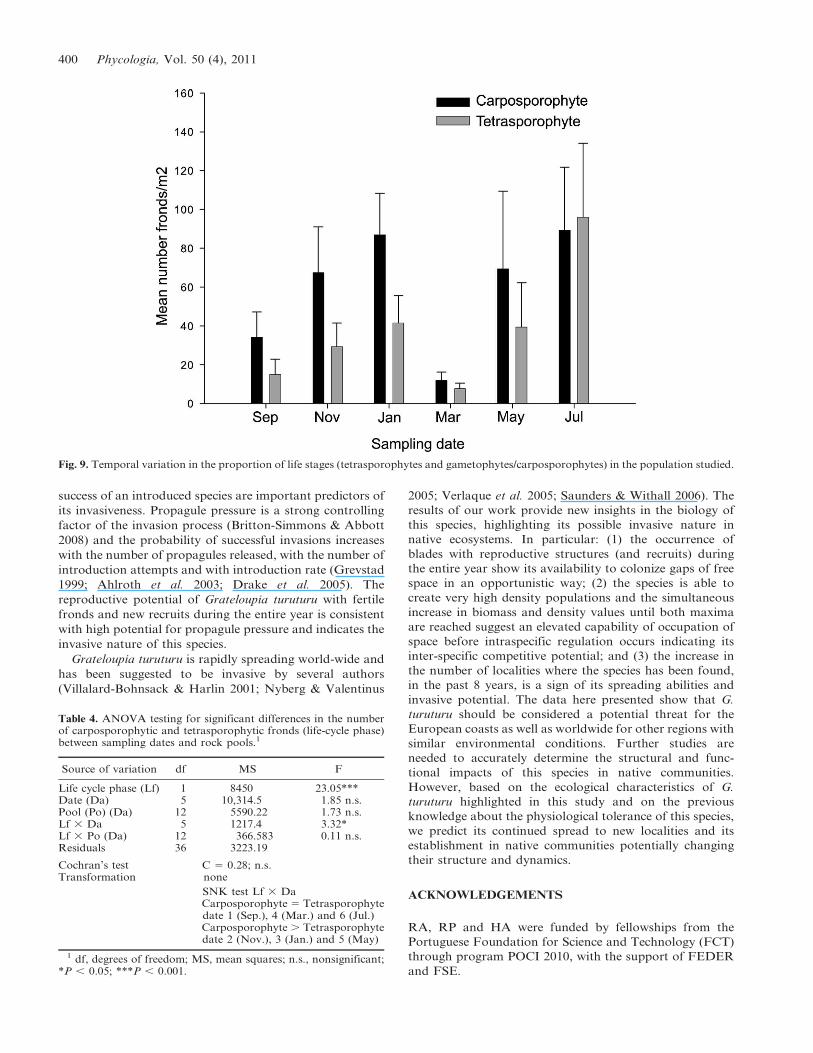
all sampling dates (Fig. 9), but male gametophytes were
never found throughout the study. Carposporophytic
fronds outnumbered the tetrasporophytic ones at all dates,
except for September, March and July (Fig. 9) (Table 4).
DISCUSSION
In this study Grateloupia turuturu was found in 30% of the
visited localities (11 out of 36), occupying rocky shores
along a 200 km stretch of the Portuguese coast. Dispersal of
G. turuturu to new locations occurs by two main processes:
(1) primary remote dispersal events often favoured by
human activities and characterized by spatial discontinuity
between donor and new established population and (2)
secondary marginal dispersal events to adjacent locations
(Simon et al. 2001). The geographical distribution of the
Portuguese populations, grouped around the main com-
mercial harbours in the area (Viana do Castelo, Leixoes,
Aveiro, Figueira da Foz and Peniche) and separated by 10s
of kms, suggest the occurrence of both primary and
secondary dispersal modes. Shipping related activities
would originate founder populations from where G.
turuturu would disperse into neighbour suitable localities.
The dispersal capability of G. turuturu is unknown but it
seems to be able to establish new populations and spread
along the shoreline in relative short periods of time.
However, the lack of a significant correlation between the
distance from main ports and the density of G. turuturu
populations suggests that other factors than the dispersal
characteristics of the species might account for the
geographical patterns found. The establishment of an
introduced species in a new environment is also dependent
on the interactive effects of factors like the environmental
and biological conditions encountered at the recipient
community and the biological traits of the introduced
species (Romanuk & Kolasa 2005; Beisner et al. 2006).
Invasive seaweeds commonly exhibit a group of charac-
teristics that confer them opportunistic, stress tolerant and/
or biological competent capabilities and consequent capa-
bility for invasiveness (Valentine et al. 2007). The estab-
lishment of an invasive species in native communities is
facilitated by the occurrence of disturbance events that
release space for new colonizers, but life history character-
istics of the invader are also determinant for its introduc-
tion and persistence in native communities (Valentine et al.
2007). The annual cycle of the population of Grateloupia
turuturu studied included two growing phases: a slow
growing winter phase, characterized by little changes in
mean length, standing biomass, density and size inequality
of fronds and a fast-growing spring phase, when the
population simultaneously increased its standing biomass,
Figs 2–5. Herbarium specimens collected along the Portuguesecoast.
Fig. 2. Specimen collected in Leca da Palmeira (August 2004;PO-Algae 3216).Fig. 3. Specimen collected in Foz do Douro (August 2004; PO-Algae 3213).Fig. 4. Specimen collected in Angeiras (October 2004; PO-Algae3220).Fig. 5. Specimen collected in Buarcos (July 2000; PO-Algae2336).
396 Phycologia, Vol. 50 (4), 2011
density and size inequality. These results highlight impor-
tant life history characteristics of G. turuturu that may
influence its invasive potential like the presence of blades
during the entire year with capability of increasing their
abundance even during the periods when environmental
conditions are unfavorable for the majority of the seaweed
species.
The seasonal pattern of abundance of Grateloupia
turuturu on the Portuguese coast was similar to that
described for Galicia (Barbara & Cremades 2004) and
Britanny (Cabioch et al. 1997; Simon et al. 2001).
Seasonality in macroalgae is controlled by several factors
like temperature, photoperiod, nutrient availability, com-
petition, grazing or by the combination of several of these
factors (Lobban & Harrison 1997). G. turuturu occurs on
the Portuguese coast mainly in mid and high intertidal rock
pools. These spatially variable habitats (Araujo et al. 2006)
experience pronounced fluctuations of environmental
factors (both daily and seasonal) when compared to those
permanently submerged (Metaxas & Scheibling 1993). In
the population studied, the period of senescence of blades
(from July to September) was coincident with the period of
the year when extreme temperature values are usually
recorded (in August the temperature of the water in rock
pools of the study area can reach 29uC with temperature
fluctuations during low tide of 7uC). Although G. turuturu
can grow under a wide range of salinity and temperature
conditions (Simon et al. 1999), temperature extremes,
rather than mean temperature values, may determine the
survival of G. turuturu blades (Harlin & Villalard-Bohnsack
2001).
Light availability may also play an important role in the
seasonal dynamics of the Grateloupia turuturu population
studied. When plants are growing in crowded conditions,
light might become the main limiting factor for survivor-
ship and growth (Schwinning & Weiner 1998). Under these
conditions self-thinning may occur, involving the disap-
pearance or growth suppression of small individuals, as a
result of asymmetric competition with larger plants (Weller
1987). Although in the past self-thinning was considered a
universal process, positive relationships between frond
density and biomass have also been reported for several
Fig. 6. Temporal variation in frond-length frequency distribution of Grateloupia turuturu fronds in the population studied.
Table 2. ANOVA testing the effects of sampling date and rock pool on mean length, wet standing biomass, density and size inequality.1
Source ofvariation df
Mean length Dry standing biomass Density Gini coefficient
MS F MS F MS F MS F
Date (Da) 5 1.9176 28.20*** 2.6618 4.94* 609.2037 3.45* 0.0814 16.54***Pool (Po) 2 0.2908 4.28* 0.2020 0.37 n.s. 73.8963 0.42 n.s. 0.0106 2.16 n.s.Da 3 Po 10 0.0680 0.43 n.s. 0.5387 1.38 n.s. 176.5809 0.53 n.s. 0.0049 0.59 n.s.Residuals 18 0.1579 0.3915 336.0118 0.0083
Cochran’s test C 5 0.33; n.s. C 5 0.42; n.s. C 5 0.30; n.s. C 5 0.17; n.s.Transformation Ln (x + 1) Ln (x + 1) Ln (x + 1) None
1 df, degrees of freedom; MS, mean squares; n.s., nonsignificant; *P , 0.05; ***P , 0.001.
Araujo et al.: Geographical distribution and population dynamics of Grateloupia turuturu on the Portuguese coast 397

seaweeds species (Santos 1995; Scrosati & DeWreede 1997;
Scrosati 2000). In the studied population, at the end of the
fast growing period, an increase in biomass was followed by
a decrease in density caused by the reduction in the number
of smaller blades. Size inequality also decreased because the
disappearance of small fronds (that were dominant in the
population) reduced the variability in frond size. These
findings are in agreement with density regulation by self-
thinning, when biomass and density were simultaneously at
their maximum although the decrease in the number of
small size blades can also be related with a diminishment in
the recruitment success. Interactions between species are
major structuring agents of seaweed communities (Chap-
man 1986), and in many cases disturbance events in native
assemblages create the opportunity for alien species to
establish at high densities (Valentine & Johnson 2003,
2004). G. turuturu was capable of simultaneously increase
its frond biomass and density without intraspecific regula-
tion, which might be determinant for its competitive
dominance when patches of bare space are available for
colonization.
Fronds of Grateloupia turuturu bearing reproductive
structures were found during the entire year but their
abundances varied seasonally. The relative abundance of
gametophytic and tetrasporophytic fronds is an important
descriptor of populations structure for seaweeds with
different life-history phases (Scrosati & Mudge 2004a).
Carposporophytic fronds were more abundant than tetra-
sporophytic fronds in three of the sampling occasions.
Similar patterns of gametophytic dominance were found for
other seaweed species (Scrosati & Mudge 2004a, b), but the
opposite trend of life-phase dominance was also described
for some species (Carmona & Santos 2006). In general,
Gigartinales seem to have a dominance of gametophytes
(Scrosati & Mudge 2004a, b; Thornber & Gaines 2004)
while tetrasporophytes are the most abundant life phase in
Fig. 7. Temporal variation in a) mean length (cm, mean 6 standard error, n 5 6), b) dry standing biomass (gm22, mean 6 standard error, n5 6), c) frond density (number fronds m22, mean 6 standard error, n 5 6) and d) frond size inequality (mean 6 standard error, n 5 6) forGrateloupia turuturu fronds in the population studied.
398 Phycologia, Vol. 50 (4), 2011

Gelidiales (Melo & Neushul 1993; Carmona & Santos
2006). For Cryptonemiales like G. turuturu these patterns of
dominance are not studied. The imbalance in the gameto-
phyte-tetrasporophyte ratio found in this study might be
related with different requirements of the two life phases
that enhance their abundance under particular conditions.
Distinct physiological responses between life phases were
described for different species for parameters like the
resistance to desiccation (Scrosati & Mudge 2004a),
mechanical properties (Carrington et al. 2001), density of
antifouling structures (Plouguerne et al. 2007), vegetative
growth rate and spore release (Carmona & Santos 2006)
and algal quality and production of defense compounds
(Verges et al. 2008). Recent works have shown that
fecundity and fertility of different life-cycle phases and the
interactive effects of environmental conditions can be
determinant for the gametophyte-tetrasporophyte ratio
found in seaweed populations (Thornber & Gaines 2004;
Fierst et al. 2005). For G. turuturu the ratio between
different life-history stages has not been studied elsewhere;
therefore we do not know the importance of this factor in
the invasive process.
In the population studied the periods of highest
abundances of reproductive fronds and of small size
individuals (presumably recruits) were not temporally
coincident. This decoupling could result from failures in
recruitment due to environmental conditions (e.g. UV
radiation and sedimentation) or grazing, both being also
seasonally variable (Diaz-Pulido & McCook 2003; Schiel et
al. 2006; Zacher et al. 2007; Navarro et al. 2008). Survival
of recruits might be enhanced by mild environmental
conditions experienced in spring and autumn and reduced
by extreme environmental conditions during summer and
winter. Additionally, seasonality might also affect the
reproductive potential of fertile blades. Seasonal differences
in relative fecundity, spore release and germination were
found for the tetrasporophytic phase of Gelidium robustum,
although reproductive fronds were present over the entire
year (Melo & Neushul 1993). The abundance of Grateloupia
turuturu’s recruits might also result from the use of other
recruitment strategies besides the formation of fronds by
spore germination. New fronds can also arise from older
crusts, from older damaged fronds and from new crusts
produced by older crusts or fronds (Harlin & Villalard-
Bohnsack 2001). The reproductive output and germination
Fig. 8. Temporal variation in the proportion of fronds (above 5 cm long) bearing reproductive structures in the population studied.
Table 3. ANOVA testing for significant differences in the numberof reproductive and nonreproductive fronds (reproductive status)between sampling dates and rock pools.1
Source ofvariation df MS F
Reproductivestatus (Rs)
1 154.448 246.68***
Date (Da) 5 2.303 1.62 n.s.Pool (Po) (Da) 12 1.4252 1.26 n.s.Rs 3 Da 5 4.9928 7.397**Rs 3 Po (Da) 12 0.6261 0.56 n.s.Residuals 36 1.1268
Cochran’s test C 5 0.2; n.s.Transformation Ln (x + 1)
SNK test Rs 3 DaN reproductive 5 N nonreproductivedate 4 (Mar.) and 5 (May)N reproductive . N nonreproductivedate 1 (Sep.), 2 (Nov.), 3 (Jan.) and 6 (Jul.)
1 df, degrees of freedom; MS, mean squares; n.s., nonsignificant;**P , 0.01; ***P , 0.001.
Araujo et al.: Geographical distribution and population dynamics of Grateloupia turuturu on the Portuguese coast 399
success of an introduced species are important predictors of
its invasiveness. Propagule pressure is a strong controlling
factor of the invasion process (Britton-Simmons & Abbott
2008) and the probability of successful invasions increases
with the number of propagules released, with the number of
introduction attempts and with introduction rate (Grevstad
1999; Ahlroth et al. 2003; Drake et al. 2005). The
reproductive potential of Grateloupia turuturu with fertile
fronds and new recruits during the entire year is consistent
with high potential for propagule pressure and indicates the
invasive nature of this species.
Grateloupia turuturu is rapidly spreading world-wide and
has been suggested to be invasive by several authors
(Villalard-Bohnsack & Harlin 2001; Nyberg & Valentinus
2005; Verlaque et al. 2005; Saunders & Withall 2006). The
results of our work provide new insights in the biology of
this species, highlighting its possible invasive nature in
native ecosystems. In particular: (1) the occurrence of
blades with reproductive structures (and recruits) during
the entire year show its availability to colonize gaps of free
space in an opportunistic way; (2) the species is able to
create very high density populations and the simultaneous
increase in biomass and density values until both maxima
are reached suggest an elevated capability of occupation of
space before intraspecific regulation occurs indicating its
inter-specific competitive potential; and (3) the increase in
the number of localities where the species has been found,
in the past 8 years, is a sign of its spreading abilities and
invasive potential. The data here presented show that G.
turuturu should be considered a potential threat for the
European coasts as well as worldwide for other regions with
similar environmental conditions. Further studies are
needed to accurately determine the structural and func-
tional impacts of this species in native communities.
However, based on the ecological characteristics of G.
turuturu highlighted in this study and on the previous
knowledge about the physiological tolerance of this species,
we predict its continued spread to new localities and its
establishment in native communities potentially changing
their structure and dynamics.
ACKNOWLEDGEMENTS
RA, RP and HA were funded by fellowships from the
Portuguese Foundation for Science and Technology (FCT)
through program POCI 2010, with the support of FEDER
and FSE.
Table 4. ANOVA testing for significant differences in the numberof carposporophytic and tetrasporophytic fronds (life-cycle phase)between sampling dates and rock pools.1
Source of variation df MS F
Life cycle phase (Lf) 1 8450 23.05***Date (Da) 5 10,314.5 1.85 n.s.Pool (Po) (Da) 12 5590.22 1.73 n.s.Lf 3 Da 5 1217.4 3.32*Lf 3 Po (Da) 12 366.583 0.11 n.s.Residuals 36 3223.19
Cochran’s test C 5 0.28; n.s.Transformation none
SNK test Lf 3 DaCarposporophyte 5 Tetrasporophytedate 1 (Sep.), 4 (Mar.) and 6 (Jul.)Carposporophyte . Tetrasporophytedate 2 (Nov.), 3 (Jan.) and 5 (May)
1 df, degrees of freedom; MS, mean squares; n.s., nonsignificant;*P , 0.05; ***P , 0.001.
Fig. 9. Temporal variation in the proportion of life stages (tetrasporophytes and gametophytes/carposporophytes) in the population studied.
400 Phycologia, Vol. 50 (4), 2011

REFERENCES
AHLROTH P., ALATALO R.V., HOLOPAINEN A., KUMPULAINEN T. &SUHONEN J. 2003. Founder population size and number of sourcepopulations enhance colonization success in waterstriders.Oecologia 137: 617–620.
ARAUJO R., BARBARA I., SANTOS G., RANGEL M. & SOUSA-PINTO I.2003. Fragmenta Chorologica Occidentalia. Anales JardinBotanico Madrid 60: 405–409.
ARAUJO R., SOUSA-PINTO I., BARBARA I. & QUINTINO V. 2006.Macroalgal communities of intertidal rock pools in the northwestcoast of Portugal. Acta Oecologica 30: 192–202.
ARAUJO R., BARBARA I., TIBALDO M., BERECIBAR E., DIAZ TAPIA
P., PEREIRA R., SANTOS R. & SOUSA-PINTO I. 2009. Checklist ofbenthic marine algae and cyanobacteria of northern Portugal.Botanica Marina 52: 24–46.
ARENAS F. & FERNANDEZ C. 2000. Size structure and dynamics ina population of Sargassum muticum (Phaeophyceae). Journal ofPhycology 36: 1012–1020.
BALATA D., PIAZZI L. & CINELLI F. 2004. A comparison amongassemblages in areas invaded by Caulerpa taxifolia and C.racemosa on a subtidal Mediterranean rocky bottom. MarineEcology 25: 1–13.
BARBARA I. & CREMADES J. 2004. Grateloupia lanceola versusGrateloupia turuturu (Gigartinales, Rhodophyta) en la PenınsulaIberica. Anales Jardin Botanico Madrid 61: 103–118.
BAX N., CARLTON J.T., MATHEWA-AMOS A., HAEDRICH R.L.,HOWARTH F.G., PURCELL J.E., RIESER A. & GRAY A. 2001. Thecontrol of biological invasions in world’s oceans. ConservationBiology 5: 1234–1246.
BEISNER B.E., HOVIUS J., HAYWARD A., KOLASA J. & ROMANUK
T.N. 2006. Environmental productivity and biodiversity effectson invertebrate community invasibility. Biological Invasions 8:655–664.
BENDEL R.B., HIGGINS S.S., TEBERG J.E. & PYKE D.A. 1989.Comparison of skewness coefficient, coefficient of variation, andGini coefficient as inequality measures within populations.Oecologia 78: 394–400.
BRITTON-SIMMONS K.H. & ABBOTT K.C. 2008. Short and long-term effects of disturbance and propagule pressure on abiological invasion. Journal of Ecology 96: 68–77.
CABIOCH J., CASTRICFEY A., LHARDYHALOS M. & RIO A. 1997.Grateloupia doryphora and Grateloupia filicina var luxurians(Rhodophyta, Halymeniaceae) from the coasts of Brittany(France). Cryptogamie Algologie 18: 117–137.
CARLTON J.T. & GELLER J.B. 1993. Ecological roulette: the globaltransport of nonindigenous marine organisms. Science 5117:78–82.
CARMONA R. & SANTOS R. 2006. Is there an ecophysiologicalexplanation for the gametophyte-tetrasporophyte ratio inGelidium sesquipedale (Rhodophyta)? Journal of Phycology 42:259–269.
CARRINGTON E., GRACE S.P. & CHOPIN T. 2001. Life historyphases and the biomechanical properties of the red alga Chondruscrispus (Rhodophyta). Journal of Phycology 37: 699–704.
CHAPMAN A.R.O. 1986. Population and community ecology ofseaweeds. Advances in Marine Biology 23: 1–161.
DAMGAARD C. & WEINER J. 2000. Describing inequality in plantsize or fecundity. Ecology 81: 1139–1142.
D’ARCHINO R., NELSON W.A. & ZUCCARELLO G.C. 2007. Invasivemarine red alga introduced to New Zealand waters: first recordof Grateloupia turuturu (Halymeniaceae, Rhodophyta). NewZeland Journal of Marine and Freshwater Research 40: 599–604.
DIAZ-PULIDO G. & MCCOOK L.J. 2003. Relative roles of herbivoryand nutrients in the recruitment of coral-reef seaweeds. Ecology84: 2026–2033.
DRAKE J.M., BAGGENSTOS P. & LODGE D.M. 2005. Propagulepressure and persistence in experimental populations. BiologyLetters 1: 480–483.
ENGELEN A. & SANTOS R. 2009. Which demographic traitsdetermine population growth in the invasive brown seaweedSargassum muticum? Journal of Ecology 97: 675–684.
FIERST J., TERHORST C., KUBLER J.E. & DUDGEON S. 2005.Fertilization success can drive patterns of phase dominance incomplex life histories. Journal of Phycology 41: 238–249.
GAVIO B. & FREDERICQ S. 2002. Grateloupia turuturu (Halymenia-ceae, Rhodophyta) is the correct name of the non-native speciesin the Atlantic known as Grateloupia doryphora. EuropeanJournal of Phycology 37: 349–359.
GREVSTAD F.S. 1999. Experimental invasions using biologicalcontrol introductions: the influence of release size on the chanceof population establishment. Biological Invasions 1: 313–323.
HARLIN M.M. & VILLALARD-BOHNSACK M. 2001. Seasonaldynamics and recruitment strategies of the invasive seaweedGrateloupia doryphora (Halymeniaceae, Rhodophyta) in Narra-gansett Bay and Rhode Island Sound, Rhode Island, USA.Phycologia 40: 468–474.
LOBBAN C.S. & HARRISON P.J. 1997. Seaweed ecology andphysiology. Cambridge University Press, Cambridge, UK. 359 pp.
MATHIESON A.C., DAWES C.J., PEDERSON J., GLADYCH R.A. &CARLTON J.T. 2008. The Asian red seaweed Grateloupia turuturu(Rhodophyta) invades the Gulf of Maine. Biological Invasions10: 985–988.
MELO R.A. & NEUSHUL M. 1993. Life-history and reproductivepotential of the agarophyte Gelidium robustum in California.Hydrobiologia 261: 223–229.
METAXAS A. & SCHEIBLING R.E. 1993. Community structure andorganization of tidepools. Marine Ecology Progress Series 98:187–198.
NAVARRO N.P., MANSILLA A. & PALACIOS M. 2008. UVB effectson early developmental stages of commercially importantmacroalgae in southern Chile. Journal of Applied Phycology 20:897–906.
NYBERG C.D. & VALENTINUS I. 2005. Can species traits be used topredict marine macroalgal introductions? Biological Invasions 7:265–279.
PARKER I.M., SIMBERLOFF D., LONSDALE W.M., GOODELL K.,WONHAM M., KAREIVA P.M., WILLIAMSON M.H., VON HOLLE
B., MOYLE P.B., BYERS J.E. & GOLDWASSER L. 1999. Impact:toward a framework for understanding the ecological effects ofinvaders. Biological Invasions 1: 3–19.
PIAZZI L., CECCHERELLI G. & CINELLI F. 2001. Threat tomacroalgal diversity: effects of the introduced green algaCaulerpa recemosa in the Mediterranean. Marine EcologyProgress Series 210: 149–159.
PLOUGUERNE E., LE LANN K., CONNAN S., JECHOUX G.,DESLANDES E. & STIGER-POUVREAU V. 2006. Spatial andseasonal variation in density, reproductive status, length andphenolic content of the invasive brown macroalga Sargassummuticum (Yendo) Fensholt along the coast of Western Brittany(France). Aquatic Botany 85: 337–344.
PLOUGUERNE E., TREPOS R., JECHOUX G., LENNON J.F.,DESLANDES E. & STIGER-POVREAU V. 2007. An investigation ofthe presence and variations in abundance of UV-absorbingstructures in Grateloupia turuturu (Yamada) (Halymeniaceae,Rhodophyta) from Brittany (France). Cryptogamie Algologie 28:159–167.
ROMANUK T.N. & KOLASA J. 2005. Resource limitation, biodiver-sity and competitive effects interact to determine the invisibilityof rock pool microcosms. Biological Invasions 7: 711–722.
SANTOS R. 1995. Size structure and inequality in a commercialstand of the seaweed Gelidium sesquipedale. Marine EcologyProgress Series 119: 253–263.
SAUNDERS G.W. & WITHALL R.D. 2006. Collections of the invasivespecies Grateloupia turuturu (Halymeniales, Rhodophyta) fromTasmania, Australia. Phycologia 45: 711–714.
SCHAFFELKE B., SMITH J.E. & HEWITT C.L. 2006. Introducedmacroalgae - a growing concern. Journal of Applied Phycology18: 529–541.
SCHIEL D.R., WOOD S.A., DUNMORE R.A. & TAYLOR D.I. 2006.Sediment on rocky intertidal reefs: effects on early post-settlement stages of habitat-forming seaweeds. Journal ofExperimental Marine Biology and Ecology 331: 158–172.
SCHWINNING S. & WEINER J. 1998. Mechanisms determining thedegree of size asymmetry in competition among plants. Oecologia113: 447–455.
Araujo et al.: Geographical distribution and population dynamics of Grateloupia turuturu on the Portuguese coast 401

SCROSATI R. 2000. The interspecific biomass-density relationshipfor terrestrial plants: where do clonal red seaweeds stand andwhy? Ecology Letters 3: 191–197.
SCROSATI R. & DEWREEDE R.E. 1997. Dynamics of the biomass-density relationship and frond biomass inequality for Mazzaellacornucopiae (Gigartinaceae, Rhodophyta): implications for theunderstanding of frond interactions. Phycologia 36: 506–516.
SCROSATI R. & MUDGE B. 2004a. Persistence of gametophytepredominance in Chondrus crispus (Rhodophyta, Gigartinaceae)from Nova Scotia after 12 years. Hydrobiologia 519: 215–218.
SCROSATI R. & MUDGE B. 2004b. Effects of elevation, waveexposure, and year on the proportion of gametophytes andtetrasporophytes in Mazzaella parksii (Rhodophyta, Gigartina-ceae) populations. Hydrobiologia 520: 199–205.
SIMON C., AR GALL E., LEVAVASSEUR G. & DESLANDES E. 1999.Effects of short-term variations of salinity and temperature onthe photosynthetic response of the red alga Grateloupiadoryphora from Brittany (France). Botanica Marina 42: 437–440.
SIMON C., AR GALL E. & DESLANDES E. 2001. Expansion of the redalga Grateloupia doryphora along the coasts of Brittany (France).Hydrobiologia 443: 23–29.
STAEHR P.A., PEDERSEN M.F., THOMSEN M.S., WERNBERG T. &KRAUSE-JENSEN D. 2000. Invasion of Sargassum muticum inLimfjorden (Denmark) and its possible impact on the indigenousmacroalgal community. Marine Ecology Progress Series 207:79–88.
STREFTARIS N., ZENETOS A. & PAPATHANASSIOU E. 2005.Globalization in marine ecosystems: the story of non-indigenousmarine species across European seas. Oceanographic MarineBiological Annual Review 43: 419–453.
THORNBER C.S. & GAINES S.D. 2004. Population demographics inspecies with biphasic life cycles. Ecology 85: 1661–1664.
UNDERWOOD A.J. 1997. Experiments in ecology: their logical designand interpretation using analysis of variance. Cambridge Univer-sity Press, Cambridge, UK. 499 pp.
VALENTINE J.P. & JOHNSON C.R. 2003. Establishment of theintroduced kelp Undaria pinnatifida in Tasmania depends ondisturbance to native algal assemblages. Journal of ExperimentalMarine Biology and Ecology 295: 63–90.
VALENTINE J.P. & JOHNSON C.R. 2004. Establishment of theintroduced kelp Undaria pinnatifida following dieback of thenative macroalga Phyllospora comosa in Tasmania, Australia.Marine and Freshwater Research 55: 223–230.
VALENTINE J.P., MAGIEROWSKI R.H. & JOHNSON C.R. 2007.Mechanism of invasion: establishment, spread and persistence ofintroduced seaweeds populations. Botanica Marina 50: 351–360.
VERGES A., PAUL N.A. & STEINBERG P.D. 2008. Sex and life-history stage alter herbivore responses to a chemically defendedred alga. Ecology 89: 1334–1343.
VERLAQUE M., BRANNOCK P.M., KOMATSU T., VILLALARD-BOHNSACK M. & MARSTON M. 2005. The genus Grateloupia C.Agardh (Halymeniaceae, Rhodophyta) in the Than Lagoon(France, Mediterranean): a case study of marine plurispecificintroductions. Phycologia 44: 477–496.
VILLALARD-BOHNSACK M. & HARLIN M.M. 1997. The appearanceof Grateloupia doryphora (Halymeniaceae, Rhodophyta) on thenortheast coast of North America. Phycologia 36: 324–328.
VILLALARD-BOHNSACK M. & HARLIN M.M. 2001. Grateloupiadoryphora (Halymeniaceae, Rhodophyta) in Rhode Island waters(USA): geographical expansion, morphological variations andassociated algae. Phycologia 40: 372–380.
WEINER J. & SOLBRIG O.T. 1984. The meaning and measurementof size hierarchies in plant-populations. Oecologia 61: 334–336.
WELLER D.E. 1987. Self-thinning exponent correlated withallometric measures of plant geometry. Ecology 68: 813–821.
ZACHER K., ROLEDA M.Y., HANELT D. & WIENCKE C. 2007. UVeffects on photosynthesis and DNA in propagules of threeAntarctic seaweeds (Adenocystis utricularis, Monostroma hariotiiand Porphyra endiviifolium). Planta 225: 1505–1516.
Received 27 August 2010; accepted 19 January 2011
Associate editor: Lisandro Benedetti-Cecchi
402 Phycologia, Vol. 50 (4), 2011
Copyright © 2022 FDOKUMEN




















