
Cultivation of Chondrus canaliculatus (C. Agardh) Greville (Gigartinales, Rhodophyta) in controlled...
13
Cultivation of Chondrus canaliculatus (C. Agardh) Greville (Gigartinales, Rhodophyta) in controlled environments Mario Edding Erika Fonck Roberto Acun ˜a Fadia Tala Received: 31 October 2006 / Accepted: 5 October 2007 / Published online: 27 October 2007 Ó Springer Science+Business Media B.V. 2007 Abstract Increased market demand for algal raw materials has stimulated research and development into new cultivation technologies, particularly in countries with economically important seaweed industries. Chondrus canaliculatus is a red alga endemic to the tem- perate Pacific coast of South America and produces a complex carrageenan. In Chile, this potential marine resource has been underutilized since the early 1990s, after it was replaced mainly by imports of Kappaphycus and Eucheum from Asia. The study of the cultivation of C. canaliculatus in outdoor tanks demonstrates the potential to produce large quantities of algal material due to the high productivity observed. The material produced may be important in Chile as feed for cultured algal herbivores such as Haliotis spp. Data were obtained on the key factors affecting the growth of this alga, including light, tem- perature, and nutrient concentrations. The results showed that, on a dry weight basis, productivity varied seasonally between 40 g m –3 wk –1 in autumn and 200 g m –3 wk –1 in spring, and the optimal stocking density in our experiments was 8 kg m –3 . Effects of self- shading were among the most important limiting factors in the culture. Discussion is presented on the interactions between temperature, pH, salinity, enrichment with N and P, and water exchange on the maximal growth of C. canaliculatus in this culture. Keywords Algae Chile Chondrus Cultivation Productivity Seaweed South East Pacific Introduction In Chile, various species of Phaeophyceae and Rhodophyta (Norambuena 1996; Buschmann et al. 2001; Anuario Estadı ´stico de Pesca 2005) have been exploited since the 1950s with production fluctuating between 74,000 and 410,850 wet metric tonnes over the last 15 years. Most of the production has been based on the exploitation of wild stocks used as M. Edding (&) E. Fonck R. Acun ˜a F. Tala Laboratorio de Bota ´nica Marina, Departamento de Biologı ´a Marina, Facultad de Ciencias del Mar, Universidad Cato ´lica del Norte, Casilla 117, Coquimbo, Chile e-mail: [email protected] 123 Aquacult Int (2008) 16:283–295 DOI 10.1007/s10499-007-9142-x
-
Upload
guadalajara -
Category
Documents
-
view
0 -
download
0
Transcript of Cultivation of Chondrus canaliculatus (C. Agardh) Greville (Gigartinales, Rhodophyta) in controlled...

Cultivation of Chondrus canaliculatus (C. Agardh)Greville (Gigartinales, Rhodophyta) in controlledenvironments
Mario Edding Æ Erika Fonck Æ Roberto Acuna Æ Fadia Tala
Received: 31 October 2006 / Accepted: 5 October 2007 / Published online: 27 October 2007� Springer Science+Business Media B.V. 2007
Abstract Increased market demand for algal raw materials has stimulated research and
development into new cultivation technologies, particularly in countries with economically
important seaweed industries. Chondrus canaliculatus is a red alga endemic to the tem-
perate Pacific coast of South America and produces a complex carrageenan. In Chile, this
potential marine resource has been underutilized since the early 1990s, after it was
replaced mainly by imports of Kappaphycus and Eucheum from Asia. The study of the
cultivation of C. canaliculatus in outdoor tanks demonstrates the potential to produce large
quantities of algal material due to the high productivity observed. The material produced
may be important in Chile as feed for cultured algal herbivores such as Haliotis spp. Data
were obtained on the key factors affecting the growth of this alga, including light, tem-
perature, and nutrient concentrations. The results showed that, on a dry weight basis,
productivity varied seasonally between 40 g m–3 wk–1 in autumn and 200 g m–3 wk–1 in
spring, and the optimal stocking density in our experiments was 8 kg m–3. Effects of self-
shading were among the most important limiting factors in the culture. Discussion is
presented on the interactions between temperature, pH, salinity, enrichment with N and P,
and water exchange on the maximal growth of C. canaliculatus in this culture.
Keywords Algae � Chile � Chondrus � Cultivation � Productivity � Seaweed �South East Pacific
Introduction
In Chile, various species of Phaeophyceae and Rhodophyta (Norambuena 1996; Buschmann
et al. 2001; Anuario Estadıstico de Pesca 2005) have been exploited since the 1950s with
production fluctuating between 74,000 and 410,850 wet metric tonnes over the last
15 years. Most of the production has been based on the exploitation of wild stocks used as
M. Edding (&) � E. Fonck � R. Acuna � F. TalaLaboratorio de Botanica Marina, Departamento de Biologıa Marina, Facultad de Ciencias del Mar,Universidad Catolica del Norte, Casilla 117, Coquimbo, Chilee-mail: [email protected]
123
Aquacult Int (2008) 16:283–295DOI 10.1007/s10499-007-9142-x

raw material for the production of phycocolloids, both in Chile and abroad. In recent years,
an indeterminate quantity of seaweeds has been continuously harvested from wild stocks as
fresh food for abalone (Haliotis spp.) in mass culture. In Chile, commercial-scale seaweed
cultivation is limited to Gracilaria chilensis. However, research into potential cultivation of
other species such as the brown macroalgae Lessonia trabeculata Villouta & Santelices and
Macrocystis pyrifera (Linnaeus) Agardh and the red macroalgae Gigartina skottsbergiiSetchell & Gardner, Callophyllis variegata (Bory de Saint Vincent) Kutzing and Chondr-acanthus chamissoi (C. Agardh) Kutzing is ongoing (Bulboa et al. 2005; Buschmann et al.
2001, 2004; Edding et al. 1990; Edding and Tala 2003; Westermeier et al. 2006).
Most carrageenan is extracted from Kappaphycus alvarezii (Doty) ex P. C. Silva to
produce j carrageenan, and Eucheuma denticulatum (Burman) Collins and Hervey, to
produce i carrageenan. Both these species are cultured predominantly along the coasts of
the Philippines and Indonesia (Trono 1993; McHugh 2003). Chondrus crispus Stackhouse,
the original source of carrageenans (j and k forms), is still used to a limited extent
(McHugh 2003). Chile has imported raw Eucheuma seaweed for carrageenan processing,
80% of which is used by the food industry. This genus appears to be replacing the local
production of ‘‘lichen gomosus’’, composed of Ahnfeltiopsis furcellata (C. Agardh) P. C.
Silva and DeCew, and Chondrus canaliculatus (C. Agardh) Greville. The national demand
for carrageenophytic algae has undergone an important increase since 1995, mainly due to
the establishment of processing plants to extract the colloids, producing a total of 1586 tons
in 2004 (Anuario Estadıstico de Pesca 2005).
C. canaliculatus is a well-known marine alga endemic to the Pacific temperate coast of
South America; it is a potentially important carrageenan-producing species occurring from
the Chinchas Islands, Peru (14�S) (Dawson et al. 1964; Acleto and Zuniga 1998) to Chiloe-
Chile (41�S) (Ramirez and Santelices 1991). The binomial taxonomy of this species is
under revision, however, since the cystocarpic morphology (Arakaki et al. 1997) and the
phylogenetic analysis based on rbcL sequencing (Hommersand et al. 1999) demonstrated
that the species does not conform to Chondrus. Until a new designation of the binomial is
agreed upon, the commonly used name, C. canaliculatus, will be used throughout this
paper. This alga grows on protected shores, forming perennial holdfasts in inter- and
subtidal areas in at least 17 known locations throughout Chile (Ramirez and Santelices
1991; Acuna 2000). Some C. canaliculatus populations diminished due to uncontrolled
human exploitation during the 1980s (Vasquez and Westermeier 1993). Carrageenan yields
from these plants are 50.5% dry wt for cystocarpic plants, and 54.4% for tetrasporic plants,
with contents of 16% and 13% of 3- and 6-anhydrogalactoses, respectively (Ayala and
Matsuhiro 1986). Alkali-modified carrageenans improves the yield of 3,6-anhydrogalac-
tose by 25% (Matsuhiro and Zalungo 1983). The presence of k, i, and j carrageenan in all
phases of its life cycle were described by Matsuhiro and Rivas (1993), while Martinez
(1998) found that the proportion of j and i carrageenan could be modified by changes in
salinity and temperature. Tapia et al. (1987) provided data on its growth in underwater
containers, with a maximum daily growth rate of 3.48%. Vega and Meneses (1999)
described the seasonal and spatial (i.e., intertidal versus subtidal stands) variations in the
productivity and reproduction of C. canaliculatus under field conditions in a population at
Puerto Aldea, in north-central Chile. Our previous experience with mass cultures of marine
algae in outdoor tanks (Edding et al. 1987; Edding 1995), as well as that of other authors
(e.g., Bidwell et al. 1985; Braud and Armat 1996) led us to examine the feasibility of
cultivating C. canaliculatus in the laboratory and in outdoor tanks as a way of insuring the
long-term stability of supply and price of carrageenan-containing raw material. The
advantages of seaweed farming would be: (a) independence from fluctuating climatic
284 Aquacult Int (2008) 16:283–295
123

conditions, (b) independence from the labour-intensive seaweed collecting, (c) increased
purity of raw material, and (d) the possibility of choosing seaweed strains with high
carrageenan yields and carrageenan having desired compositions and properties. How the
environmental factors may effect the growth and productivity of C. canaliculatus is critical
for the determination of the parameters affecting its culture.
Materials and methods
Algal material
An initial stock biomass of C. canaliculatus, mainly vegetative plants, was collected from
a natural population located at Puerto Aldea (30�160S; 71�300W) at a depth of 5 m. This
location is the site of a fishing village at the southern end of Tongoy Bay, 62 km south of
Coquimbo, Chile. Here, the mountains protect the study site from the prevailing S-SW
winds except during winter when the winds blow from the north, generating heavy surf.
Algal biomass was transported to the laboratory in thermal coolers, and cleaned of her-
bivores and epiphytes. This material was maintained in a greenhouse holding pond, which
received a continuous flow of fresh seawater, and was used at our laboratory to supply algal
biomass for different experiments.
Laboratory experiments
Several experiments were designed to observe the response of Chondrus canaliculatus in a
controlled environment to environmental changes to which the algae might be exposed
during outdoor tank cultivation. The relative growth rate of C. canaliculatus was evaluated
under different light and temperature regimes, using 250 ml Erlenmeyer flasks containing
about 4 g wet weight in 50 ml of filtered (0.45 lm) seawater enriched with Provasoli
medium (Starr and Zeikus 1987). The pH was adjusted to 8.08 using TRIS buffer, and the
culture medium was replaced daily. The cultures were arranged on a light-temperature
gradient table to obtain combinations among temperatures of 8.6, 14.3, 20.7, 26.7�C and
photon flux densities of 54, 127, and 216 lmol m–2 s–1. All tests were done using three
replicates. Neutral light filters were used to provide a gradient of light from 40 W cool
white fluorescent lamps. The intensity of photosynthetic active radiation (PAR) was
determined with a Li-250 radiometer (LI-COR Inst., USA). All systems were allowed to
acclimate for a week to the test conditions, and then daily growth was estimated by
measuring biomass increments following the methods of Edding (1995).
The effect of salinity on photosynthesis was evaluated in algae incubated in a walk-in
chamber, with a controlled temperature (18�C), PAR of 100 lmol m–2 s–1 and a photo-
period of 12:12 h (light:dark). Seawater salinities of 20, 25, 30, 32.7, and 42.5 psu were
established by dilution with distilled water for salinities lower than 34 psu and removing
H2O by freezing to obtain greater salinity. Plants (4 g wet weight per replicate; n = 3) were
maintained in culture for 96 h at the different salinities, and then transferred to an ambient
salinity of 34 psu for 48 h, after which productivity was measured as changes in oxygen by
using the Winkler method with light/dark bottles following the methods of Strickland and
Parsons (1972).
Aquacult Int (2008) 16:283–295 285
123

Outdoor tank cultures
Culture experiments were done from austral spring 1994 to spring 1995 using 1000 l
rectangular fiberglass tanks. The tank design was the same as that which we previously
used for Gracilaria culture (Ambler et al. 1988; Edding 1995). Tanks received a contin-
uous flow of 50 lm sand-filtered seawater from La Herradura Bay (30�S), and continuous
aeration (Edding 1995).
The rates of productivity at different algal densities were observed over annual cycles.
The densities used for the algal culture were 2.1, 4.2, 8.4, and 15.5 kg m–3 (n = 2), and
the production was measured each 15 days. The increase in biomass was removed after
each measurement, determined by weighing, thus leaving the same initial amount of algae
in the tank at the beginning of each measurement period. The biomass produced was dried
in an oven at 65�C to obtain dry weights (dw), and the productivity was expressed as
g dw m–3 wk–1.
Productivity was analyzed in relation to seasonal variations in light, temperature, pH,
salinity, and nutrient concentrations in the incoming water. Water temperatures were
measured using a handheld thermometer both in the tanks and in incoming water three
times per day (09:00, 12:00, and 18:00 h). At the same time, the surfaces and underwater
PAR radiation were measured in the tanks using a Li-Cor quantum meter as cited above.
The pH was measured with a Digi-Sense meter (Cole Palmer Co., USA), and the salinity
was measured by conductivity (ECT-5 meter, Toho Dentan Co.). Seasonal variations
(n = 3) of nitrogen and phosphorus (Kjeldahl method; AOAC 2000) in the tissues of
C. canaliculatus, and nitrate, nitrite, ammonium, and phosphate were measured in triplicate
in the water tanks (Strickland and Parsons 1972).
In addition, analyses of carrageenan for each treatment (n = 3) were made by aqueous
extraction (Ayala and Matsuhiro 1986) at the end of the experiment.
The effect of N and P concentrations (n = 5, 25, 125, 625, and 3125 lM) at a ratio of
about 15:1 on productivity were recorded during three summer weeks (December 94 to
January 95), together with tissue content of these nutrients as described above. The
experiment was carried out in 1000 l tanks with an algal biomass density of 15.5 kg m–3.
The nutrient sources used were reagent-grade NaNO3 and KH2PO4. In addition, analyses of
carrageenans were made for each treatment (n = 3) at the end of each experiment. The
occurrence of epiphytism was recorded by harvesting and identifying epiphytes from the
cultured plants over a period of 30 days in each tank treatment.
The effect on productivity of changing rates of water flow through tanks was measured
in two control tanks with a flow rate of 425 l h–1 (10.2 vol d–1), two tanks with 208 l h–1
(5 vol d–l), and two tanks with 42 l h–l (1 vol d–l). Variations in temperature, pH, salinity,
and tissue N and P was measured in these tanks as described above.
Data analyses
One-way analysis of variance (ANOVA) or Kruskal–Wallis tests were performed,
depending on the normality and heterocedasticity of variances. Percentage values were
angularity transformed. The null hypothesis was rejected at a values = 0.05. A Tukey
multiple range test was used to detect significant differences within treatments. A two-way
ANOVA test was used to evaluate differences measured in the cross-gradient tests with
light and temperature in the laboratory. When the number of data points differed between
experiments, a multiple comparison procedure (Dunn’s method) was applied.
286 Aquacult Int (2008) 16:283–295
123

Results
Laboratory experiments
Algae in the test cultures always presented a healthy appearance, and those in optimal
conditions grew well throughout the test period. The growth rate increased with increasing
temperature (at day 10) of culture under some of the test conditions (Table 1). No growth
was detected with the combination of lower temperature and high range of irradiance. At
the higher temperature, there was no statistically significant difference in growth attrib-
utable to an increase in light level (Table 1). In the second phase of the experiment (at day
14), there was no growth at the lowest (8.6�C) and highest (26.7�C) temperatures
(Table 1). No effect of light on growth was detected at 14.3�C and no growth was observed
at the highest light intensity. At 20.7�C, growth was observed at all light intensities with no
significant differences attributable to the different light intensities (Table 1).
For both test periods, statistical analysis (two-way ANOVA) showed significant dif-
ferences (P \ 0.05) only due to the temperature factor; there was no significant difference
(P [ 0.05) due to the light factor nor to the light/temperature interaction.
Figure 1 shows the effects of salinity on net photosynthesis by C. canaliculatus plants.
Photosynthesis differed significantly (one-way ANOVA, P \ 0.05) among the salinities.
Intermediate salinities (30 and 32.7 psu) showed higher photosynthesis, and both lower
and higher salinities depressed photosynthetic activity.
Outdoor tank cultures
Figure 2a shows that water temperatures over the culture period varied from a winter low
of 12�C to a summer high of 23�C, with only slight differences between incoming water
and tank water. In both cases, an increase in temperature was observed during the daily
measurements in each season (Fig. 2a). Irradiance values were highly variable during the
day and within each season (Kruskal–Wallis, P \ 0.05). The average irradiance reached at
noon was similar in both spring and summer (Fig. 2a). The pH in the tanks remained fairly
constant over time, with increases registered towards the end of day; in both inflow and
tank water (Fig. 2b). A significant (Kruskal–Wallis, P \ 0.05) rise in pH of the water was
detected in the water in the tanks when compared with the incoming water. There were no
significant differences (Kruskal–Wallis, P [ O.O5) in salinity between the incoming and
Table 1 C. canaliculatus growth rate at different combinations of light and temperature in laboratoryexperiments. The algae were acclimated for 1 week before the measurements began
8.6�C 14.3�C 20.7�C 26.7�C
Growth rate at day 10 (% day–1)
54 lmol m–2s–1 1.23 ± 1.23 2.07 ± 0.04 3.69 ± 0.29 3.10 ± 2.98
127 lmol m–2s–1 0.0 1.18 ± 0.48 3.59 ± 1.38 3.17 ± 1.66
216 lmol m–2s–1 0.0 0.71 ± 0.47 3.69 ± 0.25 4.16 ± 1.20
Growth rate at day 14 (% day–1)
54 lmol m–2s–1 0.0 0.14 ± 0.10 0.71 ± 0.04 0.0
127 lmol m–2s–1 0.0 0.12 ± 0.12 0.93 ± 0.65 0.0
216 lmol m–2s–1 0.0 0.0 0.72 ± 0.01 0.0
Aquacult Int (2008) 16:283–295 287
123

tank water (Fig. 2b). However, differences in salinity were observed (Kruskal–Wallis,
P \ 0.05) during spring 1995, when the salinity was higher than in spring 1994 and
summer 1995 (Fig. 2b).
The PO4 level in water was significantly different (one-way ANOVA, P \ 0.05) among
seasons, with higher values in autumn and in the spring of 1995 (Fig. 2c), whereas the
tissue phosphorus levels were significantly higher (one-way ANOVA, P \ 0.05) only in
the spring of 1995 (Fig. 2c). The main N source in the tank water was nitrate, followed by
lower values for ammonium (2 lM) and nitrite (0.5 lM). Seawater nitrates remained fairly
constant over the entire study period (one-way ANOVA, P [ 0.05), with only slight
decreases in autumn (Fig. 2c). Ammonium differed among seasons (Kruskal–Wallis,
P \ 0.05), with higher values during spring 1994 and decreasing markedly during the other
seasons (Fig. 2c). The tissue N levels showed a significant (one-way ANOVA, P \ 0.05)
increase during the year of culture, from spring 1994 to spring 1995 (Fig. 2c). Ratios for
N:P were similar when comparing the water with the plant tissues.
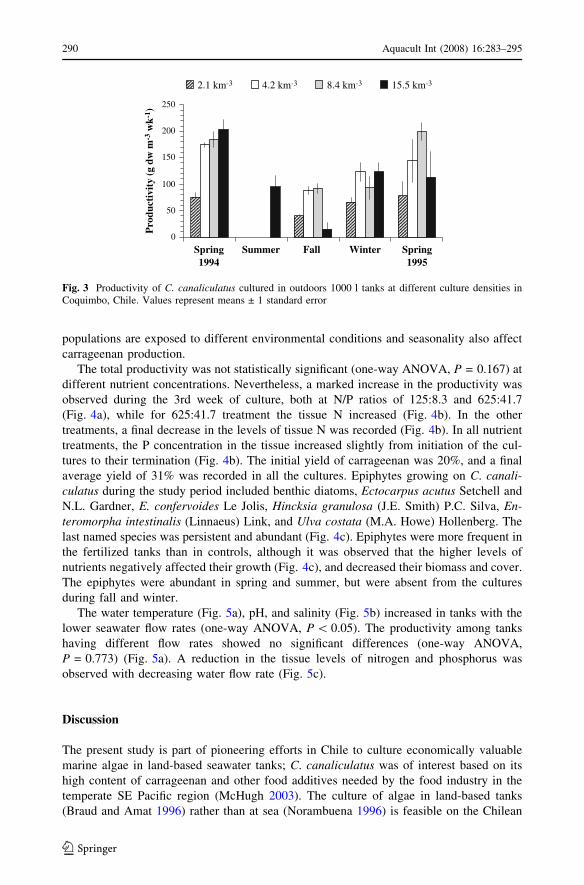
Productivity of C. canaliculatus varied significantly (two-way ANOVA, P \ 0.001)
with season and culture density (Fig. 3). In overall terms, the productivity showed a
seasonal trend, with maximum values in spring 1994 and 1995, decreasing during summer
and autumn 1995, with an increase in biomass beginning again toward winter. During the
summer, it was impossible to quantify the productivity of Chondrus in the smaller density
tanks due to the high degree of epiphytism occurring on the plants. During spring 1994, a
significant (one-way ANOVA, P \ 0.05) depression in productivity was found when
compared with the other densities (Fig. 3). Almost equal productivities (one-way ANOVA,
P \ 0.05) were obtained in autumn with tank loadings of 4.2 and 8.4 kg m–3, which were
both higher than when the initial loading was 15.5 kg m–3 (Fig. 3). Productivity was not
significantly different among the four tank loading levels during the winter (one-way
ANOVA, P = 0.059) (Fig. 3). During spring 1995, high productivity was apparent at a
density of 8.4 kg m–3, although a statistically significant difference could not be detected
among the different algal densities (one-way ANOVA, P = 0.166). Over a 112-day period
during winter–spring 1995, the biomass of C. canaliculatus doubled after the first 12 weeks
(tank loading 8.4 kg m–3). The culture productivity fluctuated between a maximum dry
weight of 195 g m–3 wk–1 reached in the fifth week, and a minimum of 54 g m–3 wk–1 in the
14th week, with an average dry weight value of 128 g m–3 wk–1.
300
400
500
600
20 25 30 32.7 45
Salinity (psu)
Pho
tosy
nthe
sis
(um
ol O
2 g-
1 h-
1 )
Fig. 1 Net photosynthesis of C. canaliculatus at different salinities. Values represent means ± 1 standarderror
288 Aquacult Int (2008) 16:283–295
123

Over the test period, the carrageenan yield was greater in the culture system containing
8.4 kg m–3, with an average carrageenan content of 26%, followed by the culture at
15.5 kg m–3 with a carrageenan yield of 21%, and at 4.2 kg m–3 with an average of 15%
carrageenan. In the system containing 2.1 kg m–3, the yield of carrageenan could not be
determined due to the high degree of contamination of the culture by epiphytes. The
variability in carrageenan yield can be explained by the quality of the plants, since
0
5
10
15
20
25
Wat
er t
empe
ratu
re (
°C)
0
1000
2000
3000 PF
D (µm
ol m-2 s -1)
a
6.5
7.5
8.5
9.5
pH
34
35
36
37
Salinity (psu)
Inflow water pH Tank water pH
Inflow water salinity Tank water salinityb o o
0
2
4
6
8
10
12
Spring1994
Summer Fall Winter Spring1995
Wat
er n
utri
ents
(µM
)
0
10
20
30
40
50
Tissue N
and P (m
g g-1)
NO3 NO2 NH4PO4 Tissue N Tissue P
c
T° inflow water T° tank water PFD
Fig. 2 Seasonal variations in (a) photon flux density and water temperature in the tanks, (b) salinity and pHof inflow and tank water, (c) nutrient in the incoming water and in C. canaliculatus tissues. For photon flux,water temperature, pH, and salinity, the points within each season represent the averages of the dailymeasurements taken at 9:00, 12:00 and 18:00 h. Values represent means ± 1 standard error
Aquacult Int (2008) 16:283–295 289
123
populations are exposed to different environmental conditions and seasonality also affect
carrageenan production.
The total productivity was not statistically significant (one-way ANOVA, P = 0.167) at
different nutrient concentrations. Nevertheless, a marked increase in the productivity was
observed during the 3rd week of culture, both at N/P ratios of 125:8.3 and 625:41.7
(Fig. 4a), while for 625:41.7 treatment the tissue N increased (Fig. 4b). In the other
treatments, a final decrease in the levels of tissue N was recorded (Fig. 4b). In all nutrient
treatments, the P concentration in the tissue increased slightly from initiation of the cul-
tures to their termination (Fig. 4b). The initial yield of carrageenan was 20%, and a final
average yield of 31% was recorded in all the cultures. Epiphytes growing on C. canali-culatus during the study period included benthic diatoms, Ectocarpus acutus Setchell and
N.L. Gardner, E. confervoides Le Jolis, Hincksia granulosa (J.E. Smith) P.C. Silva, En-teromorpha intestinalis (Linnaeus) Link, and Ulva costata (M.A. Howe) Hollenberg. The
last named species was persistent and abundant (Fig. 4c). Epiphytes were more frequent in
the fertilized tanks than in controls, although it was observed that the higher levels of
nutrients negatively affected their growth (Fig. 4c), and decreased their biomass and cover.
The epiphytes were abundant in spring and summer, but were absent from the cultures
during fall and winter.
The water temperature (Fig. 5a), pH, and salinity (Fig. 5b) increased in tanks with the
lower seawater flow rates (one-way ANOVA, P \ 0.05). The productivity among tanks
having different flow rates showed no significant differences (one-way ANOVA,
P = 0.773) (Fig. 5a). A reduction in the tissue levels of nitrogen and phosphorus was
observed with decreasing water flow rate (Fig. 5c).
Discussion
The present study is part of pioneering efforts in Chile to culture economically valuable
marine algae in land-based seawater tanks; C. canaliculatus was of interest based on its
high content of carrageenan and other food additives needed by the food industry in the
temperate SE Pacific region (McHugh 2003). The culture of algae in land-based tanks
(Braud and Amat 1996) rather than at sea (Norambuena 1996) is feasible on the Chilean
0
50
100
150
200
250
Spring1994
Summer Winter Spring1995
Pro
duct
ivit
y (g
dw
m-3
wk-
1 )
2.1 km-3 4.2 km-3 8.4 km-3 15.5 km-3
Fall
Fig. 3 Productivity of C. canaliculatus cultured in outdoors 1000 l tanks at different culture densities inCoquimbo, Chile. Values represent means ± 1 standard error
290 Aquacult Int (2008) 16:283–295
123

coast since there are no major climatic variations between the seasons that would interrupt
the culture process. Also, tank cultures lend themselves to controlled regimes of light and
nutrients, and proper management of the water quality control epiphytes (Bouarab et al.
2001). These considerations can collectively lead to higher productivity rates in tank
cultures in different seasons.
Our study clearly demonstrated the dependence of C. canaliculatus on environmental
factors. Growth of the vegetative plants in response to light and temperature variation was
0:05:0
.3
25:1.
7
125:8
.3
625:4
1.7
3125
:208.3
1º week
3º week0
50
100
150
200
250
Pro
duct
ivit
y(g
dw
m-2
wk-
1 )
0
4
8
12
16
20
0:0 5:0.3
25:1.
7
125:8
.3
625:4
1.7
3125
:208.3
Tis
sue
leve
l (m
g g
dw-1
)
Ni Nf Pi Pf
0
20
40
60
80
0:0 5:0.3
25:1.
7
125:8
.3
625:4
1.7
3125
:208.3
Nutrients (N:P µM)
Dis
lodg
ed e
piph
ytes
(%
)
0
1
2
3
4
5
Biom
ass (%dw
)
Ectocarpales Ulvales Epiphyte biomass
a
b
c
Fig. 4 C. canaliculatus (a)productivity, (b) tissue nitrogen(Ni = initial N; Nf = final N) andphosphorous (Pi = initial P;Pf = final P), and (c) epiphytechanges during cultivation inoutdoor tanks with differentnutrient concentrations. Theexcess production of C.canaliculatus was removedweekly. Values representmeans ± 1 standard error
Aquacult Int (2008) 16:283–295 291
123

similar to that of carposporic plants of this species cultured under similar conditions
(Acuna 2000). The results have shown good growth of the plants at 14�C and 20�C, at
lower and middle irradiance levels. Generally, the higher (environmental) temperature
ranges proved lethal for some seaweed (Breeman 1988; Martinez 1999), and the lowest
ranges produced decreases in growth and productivity (Martinez 1998; Pang et al. 2005).
Changes in pH in the culture water have been related to the inorganic carbon availability
in the seawater, and are active in controlling the balance among inorganic ionic forms of
carbon in the water (Zou et al. 2004). Thus, productivity should be limited both by the
ionic species of carbon available and the amounts of inorganic carbon in the water. Craigie
100
200
300
400
Pro
duct
ivit
y(g
dw m
-3w
k-1 )
16
17
18
19
20
Tem
perature (°C)
Productivity Temperaturea
8.0
8.5
9.0
9.5
10.0
pH
34.2
34.4
34.6
34.8
35.0
Salinity (psu)
pH Salinityb
0
5
10
15
20
25
Inflow 10.2 1
Water exchange (volumes per day)
Tis
sue
N (
mg
g dw
-1)
3
4
5
Tissue P
(mg g dw
-1)
N Pc
5
Fig. 5 C. canaliculatus (a)weekly productivity and tankwater temperature in cultivationwith different rates of waterexchange, (b) tank water salinity,and pH variations, and (c) tissuenitrogen and phosphorus. Valuesrepresent means ± 1 standarderror
292 Aquacult Int (2008) 16:283–295
123

and Shacklock (1995) reported that adding CO2 or H2CO3 to cultures attenuated the pH
change and maintained the inorganic C availability in C. crispus culture. Salinity is also a
factor that must to be taken into account in open-air tank cultures of C. canaliculatus, as
the present results showed that both low and high salinity values depressed its photo-
synthesis. Continuous replacement of water in the tanks helps prevents changes in salinity,
where there may be dilution of the water by rain, or increases in salinity due to
evaporation.
Nutrient dynamics play a key role in obtaining a high-quality product, both in its
individual appearance, and in the management of epiphytes. Addition of high concentra-
tions of nutrients produced a decline in productivity in the first weeks, but also reduced
contamination by epiphytes, which appeared to be more affected than C. canaliculatus.
Manipulation of nutrient concentrations may be a useful method for controlling epiphytism
in these tank cultures in the future, although both photosynthesis and carrageenan content
could decline under these conditions (Chopin and Floc’h 1992). This supposition was not
apparent in the present study, however, as the carrageenan yield from C. canaliculatusremained unchanged in spite of the different nutrient treatments. Also, it is important to
mention that the carrageenan yield obtained by Ayala and Matsuhiro (1986) was based on
plants collected in April and December in the same locality as this work. The carrageenan
yield obtained in this work can be explained by the life history of the algae and the
environmental conditions plants have in the cultivation tanks.
The seasonal pattern of productivity of C. canaliculatus followed that reported for other
algae from temperate coasts, with maxima in spring and minima toward winter (Edding
et al. 1987). The continuous production observed for C. canaliculatus cultivated in tanks
could reach a mean of 6.5 wet Kg year–1 m–3 (128 g dw m–3 wk–1) at the most favorable
density. Similar productivity values were found for Gracilaria chilensis (125 g dw m–3 wk–1,
Edding 1995), Chondracanthus chamissoi (160 g dw m–3 wk–1, M. Edding, unpublished)
and Ulva sp. (92 g dw m–3 wk–1) (J. Macchiavello, personal communication) cultivated in
the same tank system. This observation was carried out at the experimental level, the next
step being to consider economic evaluation of the model at the pilot scale. The productivity
of C. canaliculatus was in the same range as the values observed in tank culture of
C. crispus in Canada (Craigie and Shacklock 1995). The climatic conditions in Canada,
however, included low water temperatures between December and April, resulting in very
low rates of production, during which time the culture had to be suspended (Craigie and
Shacklock 1995). In Chile, this type of culture could be carried out all year, since the
normal summer–winter temperature variation in the seawater seldom exceeds 10�C, not
going below a minimum of 11�C. Algal production in tanks may be severely altered,
however, by climatic and atmospheric changes that accompany the El Nino southern
oscillation (ENSO) phenomenon, which occurs periodically on the Chilean coast, and often
produces mass mortalities of naturally occurring algal populations (Glantz 1998; Edding 1999).
Different culture objectives can be obtained by varying the nutrient inputs to the cul-
tures. Concentrations of 125 N:8.3 P stimulate productivity, whereas higher concentrations
could be used in spring and summer to control epiphytism. Future research should include
scaling up of the culture and verification of the optimal conditions, as well as the selection
of strains and clones providing optimum production of biomass and carrageenans. A good
measure of success was achieved in this area by Craigie and Schaklock (1995) in the
culture of C. crispus in Canada; these authors selected clones having optimal growth,
desirable types and yields of carrageenans, and favorable colour mutants. Factors such as
carbon supply, pest control, and disease prevention also need to be considered in futures
Aquacult Int (2008) 16:283–295 293
123

studies on C. canaliculatus in tank culture. Chondrus canaliculatus biomass needs to be
evaluated in practical terms for its potential use in the carrageenan industry, for the culture
of marine herbivores such as Haliotis spp., and even for human consumption.
Acknowledgements This study was made possible by support from Multiexport Algas SA (Chile). Wethank Mr. Danilo Peralta for expert technical help and acknowledge technical assistance from the studentsRoberto Silva, Carmen Rivera, Susana Espana, and Fungyi Chow. Finally, we would like to thank JulioNazar, Lautaro Moreno, and Dr. Louis DiSalvo for helpful reviews of this manuscript, as well as theanonymous reviewers for their comments.
References
Acleto C, Zuniga R (1998) Introduccion a las Algas. Editorial Escuela Nueva. Lima, Peru. 383 ppAcuna R (2000) Variacion en el crecimiento de Chondrus canaliculatus (Rhodophyta: Gigartinales)
provenientes de distintas poblaciones. Thesis marine biology, Universidad Catolica del Norte, Co-quimbo, Chile. 109 pp
Ambler R, Edding M, Rodrıguez A, Escobar S (1988) Excrecion de amonio por Argopecten purpuratus y suutilizacion por Gracilaria sp.: una posibilidad de policultivo. Gayana Botanica (Chile) 45:309–315
AOAC (2000) Official methods of analysis of AOAC international, vols 1 and 2, 17th edn. AOAC Inter-national, Gaithersburg
Anuarlo Estadistico de Pesca (2005) Servicio Nacional de Pesca. Ministerio de Economia. Fomento yReconstruccion, Chile
Arakaki N, Ramırez MR, Cordoba C (1997) Desarrollo morfologico y taxonomıa de Chondrus canaliculatus(C.Ag.) Grevil1e (Rhodophyta, Gigartinaceae) de Peru y Chile. Bol Mus Nac Hist Nat 46:7–22
Ayala H, Matsubiro B (1986) Carrageenans from tetrasporic and cystocarpic Chondrus canaliculatus.Phytochemistry 25:1895–1897
Bidwell R, McLachlan J, Lloyd N (1985) Tank cultivation of Irish moss, Chondrus crispus Stackh. Bot Mar28:87–98
Bouarab K, Potin P, Weinberger F, Correa J, Kloareg B (2001) The Chondrus crispus–Acrochaeteoperculata host–pathogen association, a novel model in glycobiology and applied phycopathology.J Appl Phycol 13(2):185–193
Braud J, Amat M (1996) Chondrus crispus (Gigartinaceae, Rhodophyta) tank cultivation: optimizing carboninput by a fixed pH and use of salt water well. Hydrobiology 326/327:335–340
Breeman AM (1988) Relative importance of temperature and other factors in determining geographicalboundaries of seaweeds: experimental and phenological evidence. Helgol Meeresunters 42:199–241
Bulboa C, Macchiavello J, Oliveira E, Fonck E (2005) First attempt to cultivate the carrageenan producingseaweed Chondracanthus chamissoi (Rhodophyta; Gigartinales): in Northern Chile. Aquac Res36:1069–1074
Buschmann A, Correa J, Westermeier R, Hernandez M, Norambuena R (2001) Red algal farming in Chile: areview. Aquaculture 194:203–220
Buschmann AH, Varela D, Cifuentes M, Henriquez L, Westermeier R, Correa J (2004) Experimental indoorcultivation of the carragenophytic red alga Gigartina skottsbergii. Aquaculture 241(1–4):357–370
Chopin T, Floc’h J-Y (1992) Eco-physio1ogical and biochemical study of two of the most contrasting formsof Chondrus crispus (Rhodophyta, Gigartinales). Mar Ecol Prog Ser 81:185–195
Craigie J, Schaklock P (1995) Culture of Irish Moss. In: Boghen AD (ed) Cold-water aquaculture in AtlanticCanada. The Canadian Institute for Research on Regional Development, Canada, pp 363–390
Dawson EY, Acleto C, Foldvik N (1964) The seaweeds of Peru. Nova Hedwig 13:1–111Edding M (1995) Cultivo de Gracilaria en estanques, In: Alvea1 K, Ferrario E, Oliveira y E. Sar E (eds)
Manual de Metodos Ficologicos. Universidad de Concepcion, Concepcion, pp 577–591Edding M (1999) El fenomeno de El Nino y sus efectos en la biologıa de las algas marinas. Seminario Taller
Estado del conocimiento sobre ENSO en el norte de Chile. Proyecto monitoreo regional del impacto delos eventos ‘‘El Nino’’ sobre los recursos biologicos y sus usos en America Latina. Universidad Catolicadel Norte, Coquimbo, Chile, 25 y 26 de Octubre. Abstract
Edding M, Tala F (2003) Development of techniques for the cultivation of Lessonia trabeculata ViIlouta etSantelices (Phaeophyceae: Laminariales) in Chile. Aquac Res 34:507–515
Edding M, Black H, Macchiavello J (1987) Culture of Graclaria sp. in outdoor tanks: productivity.Hydrobiology 151/152:369–373
294 Aquacult Int (2008) 16:283–295
123

Edding M, Venegas M, Orrego P, Fonck E (1990) Culture and growth of Lessonia trabeculata juvenilesporophytes in northern Chile. Hydrobiology 204/205:361–366
Glantz M (1998) Corrientes de Cambio: El Impacto de ‘‘El Nino’’ sobre el Clima y la Sociedad. CambridgeUniversity Press, Valparaıso, p 141
Hommersand M, Fredericq S, Freshwater D, Hughey J (1999) Recent developments in the systematics ofGigartinaceae (Gigartinales, Rhodophyta) based on rbcL sequence analysis and morphological evidence.Phycol Res 47:139–151
Martınez G (1998) Efecto de la temperatura y salinidad sobre el crecimiento y la sıntesis de carragenanos enlas fases reproductivas de Chondrus canaliculatus (e. Agardh) Greville (Rhodophyta: Gigartinales).Thesis marine biology, Universidad Catolica del Norte, Coquimbo, Chile. 69 pp
Martınez EA (1999) Latitudinal differences in thermal tolerance among microscopic sporophytes of theKelp Lessonia nigrescens (Phaeophyta: Laminariales) Pac. Science 54(1):74–81
Matsuhiro B, Rivas P (1993) Second derivative Fourier transforms infra red spectra of seaweeds galactans.J Appl Phycol 5:45–51
Matsuhiro B, Zalungo A, (1983) Colorimetric determination of 3,6-anhydrogalactosa in polysaccharidesfrom red seaweeds. Carbohydr Res 118:276–279
McHugh DJ (2003) A guide to the seaweed industry. FAO fisheries technical paper no. 441. Food andAgriculture Organization of the United Nations, Rome, 105 pp
Neori A, Chopin T, Troell M, Buschmann AH, Kraemer GP, Halling C, Shpigel M, Yarish C (2004)Integrated aquaculture: rationale, evolution and state of the art emphasizing seaweed biofiltration inmodern mariculture. Aquaculture 231:361–391
Norambuena R (1996) Recent trends of seaweeds production in Chile. Hydrobiology 326/327:371–379Pang SJ, Chen LT, Zhuang DG, Fei XG, Sun JZ (2005) Cultivation of the brown alga Hizikia fitsiformis
(Harvey) Okamura: enhanced seedling production in tumbled culture. Aquaculture 245(1–4):321–329Ramırez ME, Santelices B (1991) Catalogo de las algas marinas bentonicas de la costa temperada del
Pacifico de Sudamerica. Facultad de Ciencias Biologicas, Pontificia Universidad Catolica de Chile.Monografıas Bio1ogicas 5, 437 pp
Starr R, Zeikus J (1987) UTEX—the culture collection of algae at the University of Texas at Austin.J Phycol 29:1–106
Strickland J, Parsons T (1972) A practica1 handbook of seawater analysis. Fisheries Research Board ofCanada. 310 pp
Tapia L, Dıaz M, Velasquez C (1987) Growth of Chondrus canaliculatus (Rhodophyta, Gigartinales) insubmerged containers. Rev Lat Acuacu1 32:7–14
Trono G (1993) Eucheuma and Kappaphycus: taxonomy and cultivation. In: Ohno M, Critchley A (eds)Seaweed cultivation and marine ranching. Kanagawa International Fisheries Training Center, JapanInternational Cooperation Agency, Yokosuka, pp 75–88
Vasquez J, Westermeier R (1993) Limiting factors in optimizing seaweed yield in Chile. Hydrobiology 260/261:313–320
Vega J, Meneses I (1999) Seasonal and spatial monitoring of productivi1y and of reproduction of Chondruscanaliculatus (Gigartinales Rhodophyta) from Chile. Bot Mar 44:571–581
Westermeier R, Patino L, Piell MI, Maier I, Mueller D (2006) A new approach to kelp mariculture in Chile:production of free-floating sporophytes seedlings from gametophyte cultures of Lessonia trabeculataand Macrocystis pyrifera. Aquac Res 37:164–171
Zou D, Xia J, Yang Y (2004) Photosynthetic use of exogenous inorganic carbon in the agarophyte Grac-ilarla lemaneiformis (Rhodophyta). Aquaculture 237(1–4):421–431
Aquacult Int (2008) 16:283–295 295
123




















