
Public Hearing on Biological effects of ultraviolet radiation ...
Upload
independentCategory
view
0download
0
Planta (1992)188:143-154 P l a n t a
�9 Springer-Verlag 1992
Distribution and accumulation of ultraviolet-radiation-absorbing compounds in leaves of tropical mangroves Catherine E. Love lock 1., Barry F. Clough 1, and Ian E. W o o d r o w 2
1 Australian Institute of Marine Science, PMB 3 Townsville MC, and 2 Botany Department, James Cook University of North Queensland, Townsville, Q 4810, Australia
Received 30 January; accepted 20 April 1992
Abstract. Ultraviolet (UV)-absorbing phenolic com- pounds that have been shown to be protective against the damaging: effects of UV-B radiation (Tevini et al., 1991, Photochem. Photobiol. 53, 329-333) were found in the leaf epidermis of tropical mangrove tree species. These UV-absorbing phenolic compounds and leaf succulence function as selective filters, removing short and energetic wavelengths. A field survey showed that the concentra- tion of UV-absorbing compounds varied between spe- cies, between sites that would be experiencing similar levels of UV radiation, and between sun and shade leaves. Sun leaves have greater contents of phenolic com- pounds than shade leaves, and more saline sites have plants with greater levels in their leaves than less saline sites. In addition, increases in leaf nitrogen contents and quantum yields did not correlate with increasing levels of UV-absorbing compounds. It was concluded from these results that although UV-absorbing compounds form a UV-screen in the epidermis of mangrove leaves, UV radiation may not be the only factor influencing the accumulation of phenolic compounds, thus an experi- ment which altered the level of UV radiation incident on mangrove species was done. Near ambient levels of UV- A and UV-B radiation resulted in a greater content of UV-absorbing compounds in Bruguiera parviflora (Roxb.) Wight and Arn. ex Griff., but did not result in increases in B. gymnorrhiza (L.) Lamk or Rhizophora apiculata Blume. Total chlorophyll contents were lower in R. apiculata when it was grown under near-ambient levels of UV radiation than when it was grown under conditions of UV-A and UV-B depletion, but no dif- ferences were observed between the UV radiation treat- ments in the other two species. There was no difference in leaf morphology, carotenoid/chlorophyll ratios, or chlorophyll a/b ratios between UV treatments, although
* To whom correspondence should be addressed; FAX: 61 (77)796371 Abbreviations: UV = ultraviolet; HPLC = high-performance liquid chromatography; V = violaxanthin; A = antheraxanthin; Z= ze- axanthin
these varied among species; B. parviflora had the highest carotenoid/chlorophyll ratio and R. apiculata had the lowest. Thus it is proposed that differences in species response tu UV radiation may be influenced by their ability to dissipate excess visible solar radiation.
Key words: Carotenoid - Leaf (sun/shade) - Mangrove - Nitrogen nutrition - Phenolic compounds (UV absorb- ing) - Ultraviolet radiation
Introduction
Tropical mangroves receive high levels of ultraviolet (UV) radiation relative to plants inhabiting temperate regions (Frederick et al. 1989). The depletion of the atmospheric ozone layer and the expected associated increases in UV radiation at the earth's surface (Caldwell et al. 1989) may also increase the flux of UV radiation in mangrove environments. Thus, this study is directed towards assessing the mechanisms which may protect mangroves from UV radiation.
Ultraviolet radiation has been shown to be destructive to plants. Exposure of plants to enhanced UV-B radia- tion inhibits plant growth and depresses photosynthesis (see reviews by Caldwell 1981; Tevini and Teramura 1989). Reduced photosynthesis under enhanced UV-B radiation has been attributed to stomatal closure (Teramura 1983; Negash 1987), and to UV-B effects on protein synthesis and denaturation (Strid et al. 1990). It has been shown that UV-absorbing compounds (usually phenolic compounds) form a protective, UV-radiation screen within the leaf that could protect the chloroplasts and other UV-sensitive molecules from damaging UV-B radiation (Caldwell et al. 1983; Robberecht and Caldwell 1978; Les and Sheridan 1990).
Tolerance of UV-B radiation has been shown to vary between genotypes (Flint et al. 1985; Murali et al. 1988; Larson et al. 1990; Teramura et al. 1990). This variation has been attributed to the varying thickness of leaves

144 C.E. Lovelock et al. : UV-absorbing compounds in leaves of tropical mangroves
( tha t a t t e n u a t e U V rad ia t ion ) , to different abil i t ies o f geno types to p r o d u c e U V - a b s o r b i n g c o m p o u n d s (Rob- be rech t a n d Ca ldwel l 1978; Caldwel l et al. 1983; Mura l i et al. 1988; Les and She r idan 1990), and also to differen- ces in l ea f pe rox idase act ivi t ies and ca ro t eno id conten ts (Mura l i et al. 1988). I t has also been shown tha t the c o m b i n a t i o n o f enhanced U V - B r ad i a t i on and o the r t r ea tmen t s tha t i m p o s e phys io log ica l stress, for example , h igh levels o f visible l ight (Cen and B o r n m a n 1990), wa te r deficits (Mura l i and T e r a m u r a 1986) and phos - p h a t e deficiencies (Mura l i and T e r a m u r a 1987), reduce the effects o f U V - B r a d i a t i o n re la t ive to t r ea tmen t s in which these a d d i t i o n a l stresses were no t imposed . This ind ica tes tha t a d d i t i o n a l phys io log ica l stress m a y in some way con t r i bu t e to p r o t e c t i o n f rom the effects o f U V rad i a t i on , e i ther by s t imula t ing the p r o d u c t i o n o f UV- a b s o r b i n g c o m p o u n d s ( shown by Cen and B o r n m a n (1990)), by resul t ing in a l t e ra t ions to l ea f m o r p h o l o g y , or t h r o u g h increased c a p a c i t y for o the r means o f p ro tec- t ion, for example an increase in the concen t r a t i on o f ca ro t eno ids .
M a n g r o v e species have high concen t r a t ions o f pheno l i c c o m p o u n d s in thei r leaves ( R o b e r t s o n 1988) and of ten have thick, succulent leaves (Ball et al. 1988). In the pas t , m o s t s tudies o f phenol ics in m a n g r o v e leaves have been done by ecologis ts wi th an interes t in the effects o f t ann ins on c o n s u m p t i o n o f l ea f l i t ter by an imals (e.g. P o o v a c h i r a n o n et al. 1986). Very litt le cons ide ra t ion has been given to a poss ib le phys io log ica l role for these c o m p o u n d s .
The a ims o f the exper imen t s descr ibed here were to : (i) inves t iga te pheno l i c c o m p o u n d s and leaf succulence (assoc ia ted with g rea te r leaf th ickness) in mangroves wi th a view to assessing thei r l ikely invo lvemen t in p r o v i d i n g p ro t ec t i on f rom U V rad ia t ion , (ii) de te rmine whe the r there are species differences in the accumula t i on o f pheno l i c c o m p o u n d s in m a n g r o v e leaves, a n d (iii) assess whe the r the l ight regimes u n d e r which leaves are g rown, site, o r n i t rogen nu t r i t i on cor re la te wi th leaf suc- culence a n d pheno l i c con ten t s o f m a n g r o v e leaves. A fi- nal expe r imen t was done to assess d i rec t ly the response o f m a n g r o v e s to differing levels o f U V rad ia t ion . O f pa r t i cu l a r in teres t was whe the r U V r a d i a t i o n s t imula tes the p ro tec t ive mechan i sms tha t have p rev ious ly been shown to occur in o the r p lants . T h a t is, whe the r there is: (i) inc reased p r o d u c t i o n a n d a c c u m u l a t i o n o f UV- a b s o r b i n g c o m p o u n d s , (ii) increased concen t r a t i on o f c a r o t eno id s , (iii) a change in l ea f m o r p h o l o g y , and (iv) whe the r the responses to U V rad i a t i on differ be tween species. Dif ferent levels o f U V r a d i a t i o n were o b t a i n e d in n a t u r a l sun l igh t by enclos ing p lan t s in g rowth cab ine ts m a d e ou t o f acryl ic sheet ing tha t e i ther e l imina ted U V - A and U V - B r a d i a t i o n ( f i l te red-out wave lengths shor te r t han 380 nm), or r e m o v e d wave leng ths shor te r than 300 nm.
Materials and methods
Distribution of phenolie compounds in mangrove leaves. Sun and shade leaves of Bruguiera parviflora (Roxb.) Wight and Arn. ex
Griff., B. gymnorrhiza (L.) Lamk, Rhizophora stylosa Griff. and Xylocarpus granatum Konig were collected at the mouth of the Daintree River (Lat. 16~ 146~ These were stained using a saturated solution of ferrous sulphate in water with 5% formal- dehyde, which selectively stains phenolic compounds (Johansen 1940). Leaf material was left in the stain for 48 h before being washed and stored in 70% ethanol. Leaf material was imbedded in paraffin before sectioning. Sections were cut 12 ktm thick, mounted and photographed.
Absorptanee of UV by fresh epidermal pieces. The epidermal absorp- tion spectra of leaves was assessed in the mangrove species, Bru- guiera parviflora, B. gymnorrhiza and Rhizophora stylosa which were growing in 100 % seawater in natural sunlight at the Australian Institute of Marine Science (Lat. 19~ Long. 147~ Using a scalpel, six epidermal pieces were peeled from six leaves of each species. These epidermal pieces were mounted in a small holder and clamped in a quartz spectrophotometer cuvette to ensure that they did not move during measurement of absorptance. An initial mea- surement of the absorption spectrum from 250 to 750 nm in distilled water using a distilled-water blank was made using a scanning spectrophotometer (model U-3200; Hitachi, Tokyo, Japan). Epi- dermal pieces were then washed in 100 % methanol for 2 h while still in the cuvette to remove UV-absorbing compounds (Robberecht and Caldwell 1978; Caldwell et al. 1983). During this period the methanol was changed half-hourly and stored. The four half-hourly extracts for each epidermal peel were combined and the absorption spectrum of the extracted pigment was measured. After the 2-h extraction the epidermal pieces were then rescanned in distilled water. The difference between the initial and final integrated values represents the amount of pigment removed. Values are expressed as a proportion of the initial absorptance since epidermal thickness and the exact area of epidermis placed in the light path was not mea- sured.
Using a spectrophotometer to measure the spectral qualities of the epidermis of leaves is less than ideal because it does not account for light reflected by the epidermis (Robberecht and Caldwell 1978). The preferred equipment, consisting of an integrating sphere attach- ed to a monochromator via a fibre-optic cable (Robberecht and Caldwell 1978) was not available for use in the experiments de- scribed in this paper. Hence, the epidermal absorption characteris- tics presented here are not absolute. However, they are adequate to allow comparisons between the mangrove species.
Effect of sun, shade, site, and species on the phenolic contents of leaves. Leaves from sun and shade environments of mangrove species Bruguiera parviflora, B. gymnorrhiza, Ceriops tagal (Perr.) C.B. Robinson, Rhizophora stylosa, Lumnitzera racemosa Willd. and Xylocarpus granatum were collected at four sites, three in the Daintree River and one in Bowling Green Bay (Lat. 19~ Long. 147~ close to the Australian Institute of Marine Science (Table 1). All six species were not present at each site. At each site, two trees of each species present were selected and five replicate leaves from both sun and shade conditions were collected. Leaves were weighed and their area measured using a planimeter (Li-Cor, Lin- coln, Neb., USA). They were then dried for 7 d at 70 ~ C and reweighed. The dried leaves were ground to a fine powder in an agate ball mill (KHD Humboldt, Wedag, FRG). 0.1 g of the dried powder was assayed for phenolic content using the Folin-Denis colormetric assay for soluble phenolics (Allen et al. 1974), with improvements recommended by Mole and Waterman (1987). Tan- nic acid was used as a standard. A preliminary test of variance within the trees sampled (i.e. between the five leaves of any tree) showed that variation between individual leaves from any one tree, in the sun or shade, was small. Therefore, the five replicate leaves for each tree were pooled for analysis. Leaf succulence was cal- culated as : (leaf fresh weight-leaf dry weight)/leaf area.
Nitrogen nutrition and phenolic accumulation. Seedlings of Rhizo- phora scvlosa were grown for 18 months in nutrient culture in the shadehouse at the Australian Institute of Marine Science (see

C.E. Lovelock et al. : UV-absorbing compounds in leaves of tropical mangroves 145
Table 1. Distribution of mangrove species at selected sites in nor th Queensland
Site Location and site description Species present
1 Daintree River mouth, South Rhizophora stylosa - High rainfall ( > 2 m ' yr-1) Bruguiera parviflora - Substrate salinity: approx. 350 mmol NaC1 B. 9ymnorrhiza
Xylocarpus granatum
B. gymnorrhiza B. parviflora X. granatum
Daintree River, 6 km up-fiver - High rainfall ( > 2 m" y r - 1) - Substrate salinity: 100-200 mmol NaC1
Daintree River mouth creek, Nor th - High rainfall ( > 2 m - yr -1) - Substrate salinity: approx. 350 mmol NaC1
Bowling Green Bay Nor th - Low rainfall (1-1.4 m . y r - 1 ) - Substrate salinity: > 350 mmol NaC1
R. stylosa B. #ymnorrhiza X. granatum
R. stylosa Ceriops tagal Lumnitzera racemosa
Clough 1984). Nutrient solutions were adjusted to 25% seawater. Nitrogen was supplied as bo th ammonium and nitrate to make up nitrogen levels to 0, 5, 10, 20 or 180 mg �9 1-1. Maximum visible light levels within the shadehouse were approx 1000~mol quan- ta �9 m -2 �9 s -1.
A leaf from the second leaf pair on the primary shoot of each of the four plants grown at each nitrogen level was harvested, weighed, and its area measured (Li-Cor planimeter). A 10-cm z disc was then punched from the leaf and used to determine the apparent quantum yield of photosynthesis on an incident-light basis. This was done in a Leaf Disc Electrode system (Hansetech, Kings Lynn, Norfolk, UK) illuminated with a tungsten halogen projector lamp (Sylvania, Tokyo, Japan) from which the infrared component was removed by a Schott infrared filter (Schott AG, Mainz, FRG) and passed through a Schott cold mirror. Neutral-density filters (Schott) were used to adjust the light levels. Carbon dioxide was supplied to the leaf disc by passing a humidified stream of 5% COz in air (prepared by CIG, Townsville, Australia), at a flow of 5 ml �9 m i n - 1 into the chamber (Bj6rkman and Demmig 1987). Prior to measure- ments the instrument was set to zero using a stream of nitrogen gas, and calibrated by injecting 1 ml of air from a gas-tight syringe into the closed chamber. Before measurements began on a leaf disc it was exposed to approx. 160 ~tmol quanta �9 m -2 �9 s -1 for 20 min. The leaf disc was then placed in darkness to measure a dark-respiration rate. Light levels were then increased in five steps to approx. 70 ~tmol quanta �9 m -z - s -1 with a final measurement at 160 lamol quanta �9 m - 2. s - 1. The chamber was closed and a measurement of the Oz evolution made once a stable rate of 02 evolution was reached at each new light level. The chamber was then reopened and flushed with the stream of 5 % COz in air until the 02 concentration in the chamber had returned to normal (usually not longer than 2 min). The leaf disc was then dried at 70 ~ C for one week, ground and assayed for soluble phenolics (see above). Of the ground tissue, 0.1 g was also used to determine carbon, nitrogen and hydrogen contents of leaves (CHN-600 Determinator; Leco Corp., St. Joseph, Mich., USA).
Growth cabinets. Four plants from the mangrove species Rhizophora apiculata, Bruguiera gymnorrhiza and B. parviflora (all from the family Rhizophoraceae) were grown between July and November in 100 % seawater nutrient solutions (Clough 1984), in environments with either ambient or depleted UV-radiat ion using natural sunlight as the light source. The two UV radiation treatments (depleted UV, or near-ambient UV radiation levels) were obtained by using ven- tilated growth cabinets made out of acrylic sheeting that either removed UV radiation (Shinkolite; Mitsui, Tokyo, Japan), filtering out wavelengths less than 380 nm, or allowed much of the UV radiation to penetrate the cabinet, filtering out wavelengths less than 300 nm (Acropoly; supplied by Cadillac Plastics, Sydney,
Australia). Each growth cabinet was ventilated using two electric fans that each exhausted 1 m 3- min -1 (part no. 508-021; Radio- Spares Components, Perth, W.A., Australia). After four months under these conditions the three youngest, fully expanded leaves from each plant were used for analysis.
Sampling procedure. Four discs (1.37 cm 2 ) w e r e punched from the leaf, frozen in liquid nitrogen and stored at - 8 0 ~ C until further analysis. The remainder of the leaf was weighed and its area mea- sured (Li-Cor planimeter). Leaves were dried at 70 ~ C for a week to determine leaf dry weights. Succulence was determined as de- scribed above. Measurements of leaf area included the area of the leaf discs punched when the leaves were harvested.
Pigment analysis. Pigments from leaves were extracted three times in 100% acetone (described in Lovelock 1992). Total carotenoid contents (i.e. neoxanthin + violaxanthin + antheraxanthin + lu- tein + zeaxanthin + [3 carotene), contents of xanthophyll-cycle com- ponents (i.e. violaxanthin (V) + antheraxanthin (A) + zeaxanthin (Z)) and chlorophyll levels were determined by high-performance liquid chromatography (HPLC) (Juhler and Cox 1990). Quantifica- tion of the UV-absorbing compounds was also obtained by HPLC. The UV-absorbing compounds within leaves were eluted on a 25-cm-long C-8 column (Brownlee; Applied Biosystems, Foster City, Calif., USA) with a C-8 guard column preceding it with the detector set at 340 nm using a gradient modified for mangroves from Tevini et al. (1991). The gradient was linearly developed from 20% to 49% solvent B (5% ortho-phosphoric acid, 16% methanol and 18% tetrahydrofuran in water) in solvent A (5% ortho- phosphoric acid, 6% methanol, and 10% tetrahydrofuran in water) over 10 min with a flow rate of 1 ml �9 m i n - 1. The gradient of Tevini et al. (1991) was developed over 14 min.
Statistical analysis. Statistical analysis was done using the comput- ing packages STATISTIX (NH Analytical Software, Roseville, Minn., USA) and the general linear model procedure (GLM) of SAS (SAS Institute Inc., Cary, N. C., USA). Differences in the proport ion of UV-absorbing compounds leached from mangrove leaves of different species was assessed using a one-way analysis of variance. Significance was assessed using an F-ratio test at P = 0.05. The least significant difference (LSD), defined as the standard error of the difference between two means multiplied by the critical t-value (i.e. LSD = tcrlt*(S/lf~)), was used for comparison between means.
Comparison of leaf phenolic concentrations and succulence between tree species and position in the canopy (sun/shade) was assessed using analysis of variance on a site-by-site basis because of the unbalanced design of the experiment (not all species appear at every site). Species and position in the canopy were represented as
146 C.E. Lovelock et al. : UV-absorbing compounds in leaves of tropical mangroves
crossed factors, with trees nested within the crossing of species and canopy-position factors. The position of leaves within canopies and species were considered fixed effects and trees as a random effect. Comparison between sites and sun/shade used data for R. stylosa over sites 1, 3 and 4. The design was the same as for species and position-in-the-canopy comparisons, except site replaced the species as one of the fixed effects. Significance of effects was assessed using an F-ratio test at P = 0.05; LSD was calculated as described above.
Relationships between leaf nitrogen and each of phenolic con- tents, quantum yield and leaf specific weight were investigated using regression analysis, as was the relationship between specific weights of leaves and their soluble phenolic contents. Significance of the independent variable (leaf nitrogen content or specific weight of leaves) was assessed using a t-test at a significance level P = 0.05.
In the growth-cabinet experiments, analysis of variance of the plant-response variables (e.g. pigment concentrations and leaf mor- phological characteristics) with UV treatment and species used a design in which species and UV treatments were represented as crossed factors, with trees nested within the crossing of species and UV treatment, and with leaves nested within trees. Significance of effects was assessed using F-ratio tests. Logarithmic transforma- tions of the data were made where the variability in the data was not constant, in order to satisfy the constant variance requirement of analysis-of-variance models.
Results
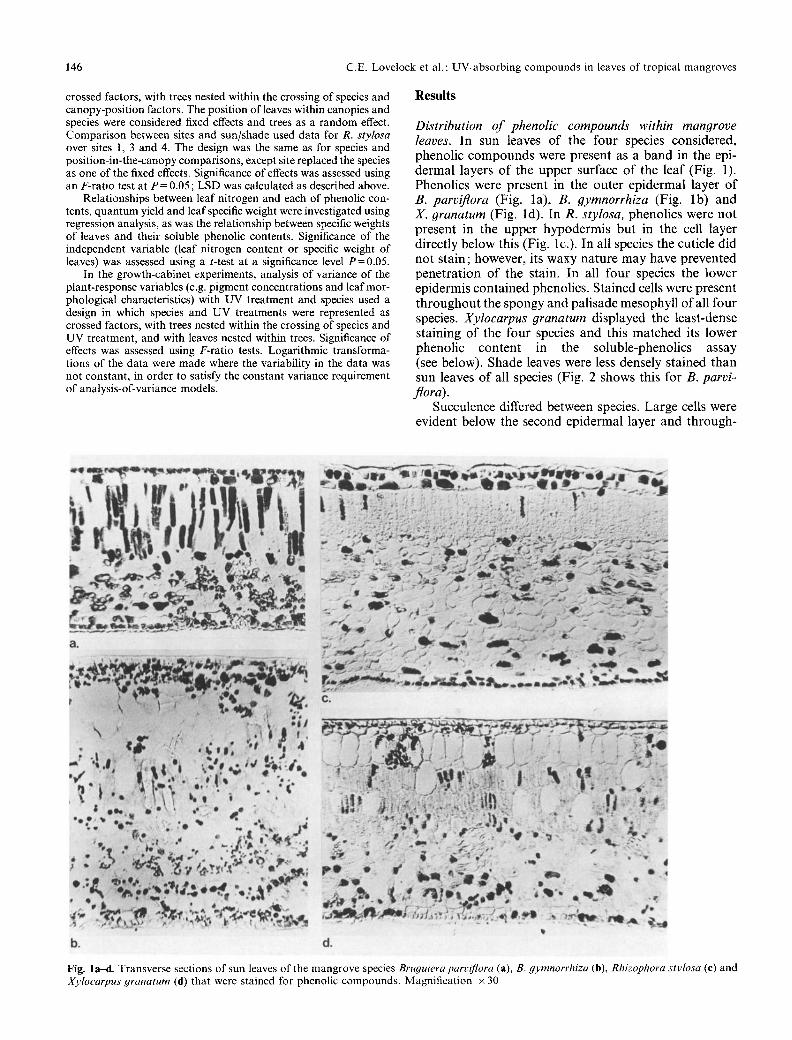
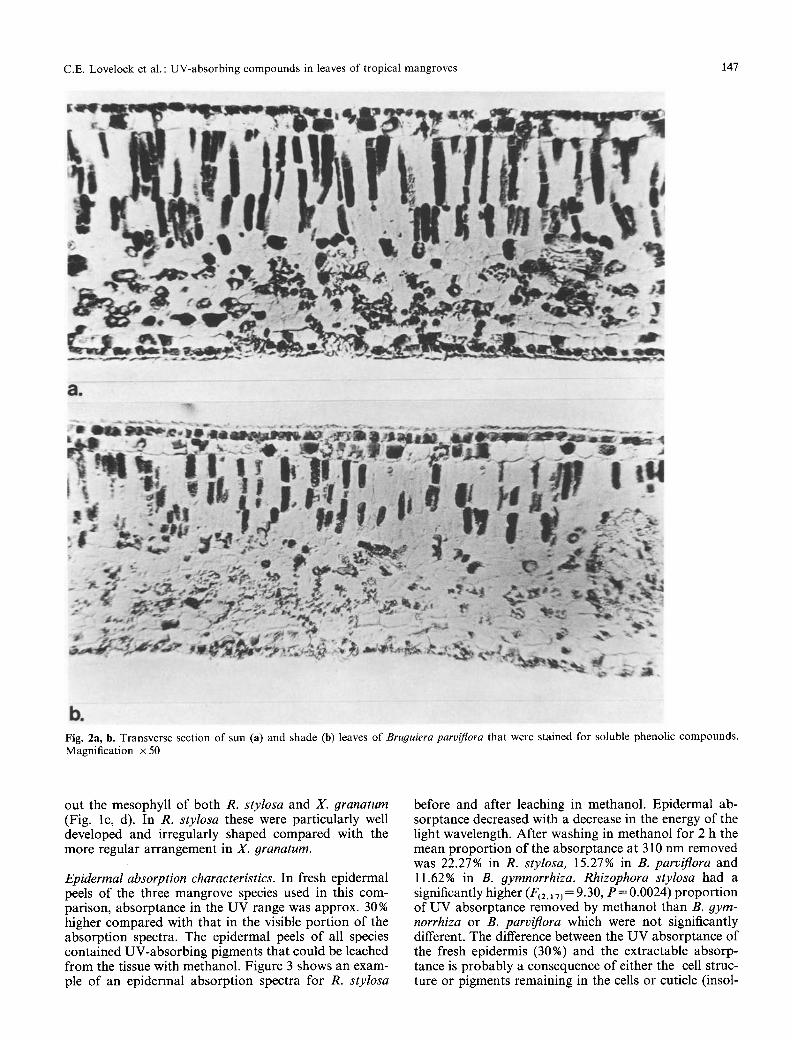
D&tribution of phenolic compounds within mangrove leaves. In sun leaves o f the four species cons idered , phenol ic c o m p o u n d s were presen t as a band in the epi- de rma l layers o f the u p p e r surface o f the leaf (Fig. 1). Phenol ics were present in the ou te r ep ide rma l layer o f B. parviflora (Fig. la ) , B. 9ymnorrhiza (Fig. lb ) and X. granatum (Fig. ld) . In R. stylosa, phenol ics were no t p resen t in the u p p e r h y p o d e r m i s bu t in the cell layer d i rec t ly be low this (Fig. lc.). In all species the cuticle d id no t s ta in ; however , its waxy na tu r e m a y have p reven ted p e n e t r a t i o n o f the stain. In all four species the lower ep idermis con ta ined phenol ics . S ta ined cells were presen t t h r o u g h o u t the spongy and pa l i sade mesophy l l o f all four species. Xyloearpus granatum disp layed the leas t -dense s ta ining o f the four species and this ma t ched its lower phenol ic con ten t in the so luble-phenol ics assay (see below). Shade leaves were less densely s ta ined than sun leaves o f all species (Fig. 2 shows this for B. parvi- flora).
Succulence differed between species. La rge cells were evident be low the second ep ide rma l layer and th rough-
Fig. l a d . Transverse sections of sun leaves of the mangrove species Bruguiera parvi)qora (a), B. gymnorrhiza (b), Rhizophora stylosa (e) and Xylocarpus #ranatum (d) that were stained for phenolic compounds. Magnification • 30
C.E. Lovelock et al. : UV-absorbing compounds in leaves of tropical mangroves 147
Fig. 2a, b. Transverse section of sun (a) and shade (b) leaves of Bruguiera parviflora that were stained for soluble phenolic compounds. Magnification x 50
out the mesophyll of both R. stylosa and X. granatum (Fig. lc, d). In R. stylosa these were particularly well developed and irregularly shaped compared with the more regular arrangement in X. granatum.
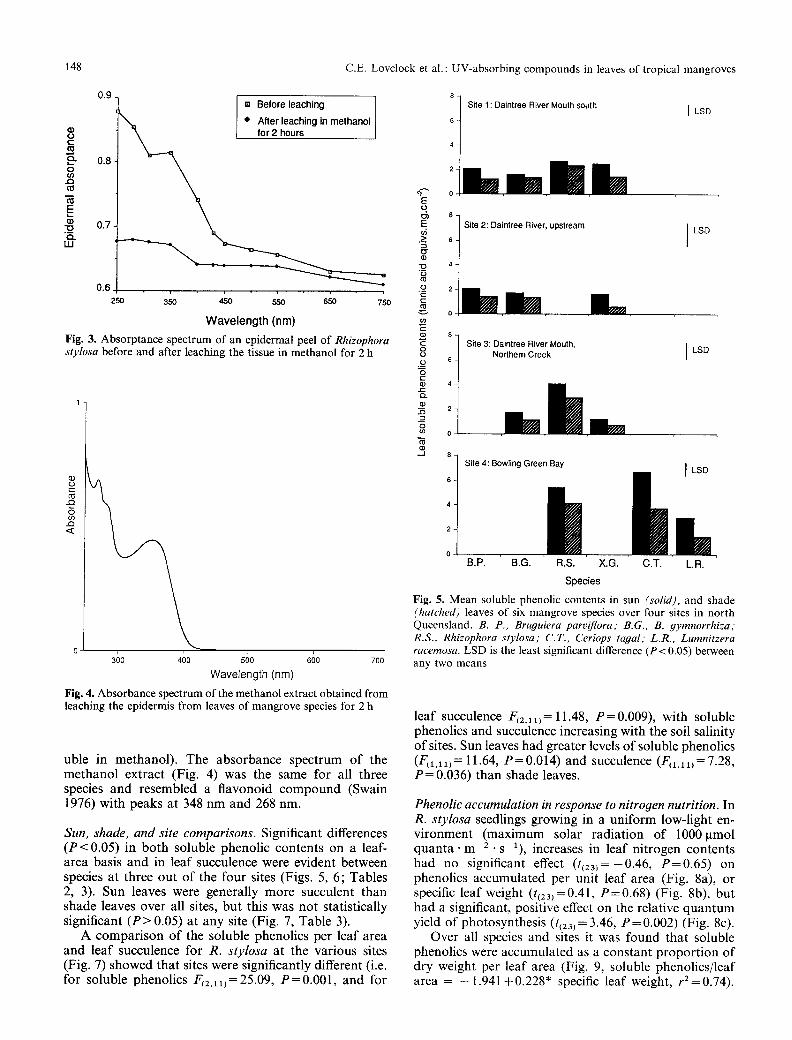
Epidermal absorption character&tics. In fresh epidermal peels of the three mangrove species used in this com- parison, absorptance in the UV range was approx. 30% higher compared with that in the visible port ion of the absorption spectra. The epidermal peels of all species contained UV-absorbing pigments that could be leached from the tissue with methanol. Figure 3 shows an exam- ple of an epidermal absorption spectra for R. stylosa
before and after leaching in methanol. Epidermal ab- sorptance decreased with a decrease in the energy of the light wavelength. After washing in methanol for 2 h the mean proport ion of the absorptance at 310 nm removed was 22.27% in R. stylosa, 15.27% in B. parviflora and 11.62% in B. 9ymnorrhiza. Rhizophora stylosa had a significantly higher (F~z,l 7) = 9.30, P = 0.0024) proport ion of UV absorptance removed by methanol than B. gym- norrhiza or B. parviflora which were not significantly different. The difference between the UV absorptance of the fresh epidermis (30%) and the extractable absorp- tance is probably a consequence of either the cell struc- ture or pigments remaining in the cells or cuticle (insol-
148 C.E. Lovelock et al. : UV-absorbing compounds in leaves of tropical mangroves
0.9 [] Before leaching �9 After leaching in methanol
~ for2 hours
- 0.8
-o 0.7.
0.6 250 350 4~50 550 S50 750
Wavelength (nm)
Fig. 3. Absorptance spectrum of an epidermal peel of Rhizophora stylosa before and after leaching the tissue in methanol for 2 h
o E .Q O o3 ..Q
<
I
0 ] i 300 400 500 600 7(30
Wavelength (nm)
Fig. 4. Absorbance spectrum of the methanol extract obtained from leaching the epidermis from leaves of mangrove species for 2 h
uble in methanol). The absorbance spectrum of the methanol extract (Fig. 4) was the same for all three species and resembled a flavonoid compound (Swain 1976) with peaks at 348 nm and 268 nm.
Sun, shade, and site comparisons. Significant differences ( P < 0.05) in both soluble phenolic contents on a leaf- area basis and in leaf succulence were evident between species at three out o f the four sites (Figs. 5, 6; Tables 2, 3). Sun leaves were generally more succulent than shade leaves over all sites, but this was not statistically significant ( P > 0.05) at any site (Fig. 7, Table 3).
A compar ison of the soluble phenolics per leaf area and leaf succulence for R. stylosa at the various sites (Fig. 7) showed that sites were significantly different (i.e. for soluble phenolics F(2,ii)=25.09, P=0 .001 , and for
E o t)') E
(3
r "E-
u)
o o o
8
o
_J
Site 1 : Daintree River Mouth south
'lil=mlm 0.
8- Site 2: Daintree River, upstream
6-
4
0
S Site 3: Daintree River Mouth,
Northem Creek 6
4
2
0
8 - Site 4: Bowling Green Bay
6 -
4-
2-
o- B.P. B.G. R.S. X.G. C.T. L.R.
Species
LSD
LSD
LSD
I Rll
Fig. 5. Mean soluble phenolic contents in sun (solid), and shade (hatched) leaves of six mangrove species over four sites in north Queensland. B. P., Bruguiera parviflora; B.G., B. gymnorrhiza," R.S., Rh&ophora stylosa; C.T., Ceriops tapal; L.R., Lumnitzera racemosa. LSD is the least significant difference (P< 0.05) between any two means
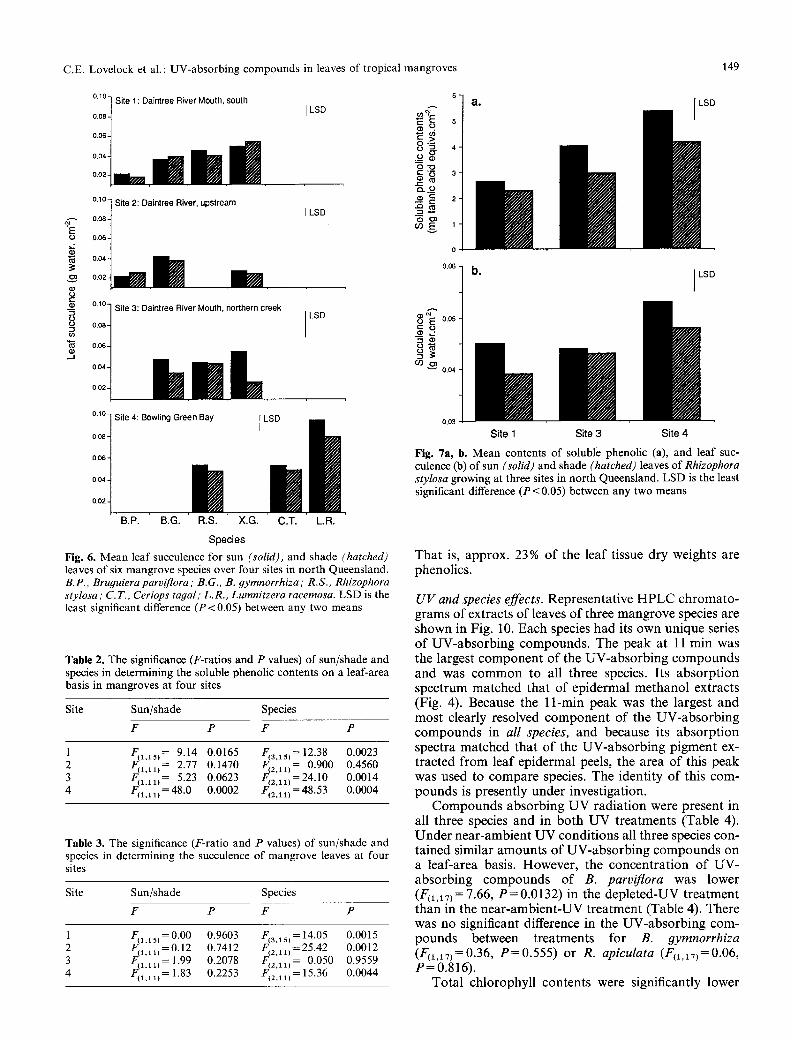
leaf succulence F(2 ,11 )= 11.48, P = 0.009), with soluble phenolics and succulence increasing with the soil salinity of sites. Sun leaves had greater levels of soluble phenolics (F(i,u) = 11.64, P = 0.014) and succulence (F(1.ii)= 7.28, P = 0.036) than shade leaves.
Phenolic accumulation in response to nitropen nutrition. In R. stylosa seedlings growing in a uniform low-light en- vironment (maximum solar radiation of 1000gmol quan ta , m -2 - s - i ) , increases in leaf nitrogen contents had no significant effect ( t{23)=-0.46, P=0 .6 5 ) on phenolics accumulated per unit leaf area (Fig. 8a), or specific leaf weight (t~23)=0.41, P=0 .68 ) (Fig. 8b), but had a significant, positive effect on the relative quantum yield of photosynthesis (t(23)= 3.46, P = 0.002) (Fig. 8c).
Over all species and sites it was found that soluble phenolics were accumulated as a constant proport ion of dry weight per leaf area (Fig. 9, soluble phenolics/leaf area = - 1.941 +0.228" specific leaf weight, rZ= 0.74).
C.E. Lovelock et al. : UV-absorbing compounds in leaves of tropical mangroves 149
%
i1) o
o= r
.J
0.10] Site 1 : Daintree River Mouth, south 0.08
0.06
0.04 - ~ 0.02 -
0.060"0e0"10 t Site 2: Daintree River, upstream
0.04 - ~
0.02 -
~176 1 Site 3: Daintree River Mouth, northern creek
J 0.08
0.06
0 . 0 4 - ~
0.02-
0.10-
0.08 -
0.06 -
0.04 -
6 LSD uJ
~ 4
o"O
O..._~ _ ~ 8
LSD ~ ~ o
0.06
LSD
S i t e 4: Bowling Green Bay I LSD h
im i | 0.02 -
B.P. B.G. R.S. X.G. C.T. L.R.
Species
Fig. 6. Mean leaf succulence for sun (solid), and shade (hatched) leaves of six mangrove species over four sites in north Queensland. B.P., Bru#uiera parviflora ; B.G., B. 9ymnorrhiza ; R.S., Rhizophora stylosa ; C.T., Ceriops tayal ; L.R., Lumnitzera racemosa. LSD is the least significant difference (P<0.05) between any two means
Table 2. The significance (F-ratios and P values) of sun/shade and species in determining the soluble phenolic contents on a leaf-area basis in mangroves at four sites
Site Sun/shade Species
F P F P
1 F~l,ls) = 9.14 0.0165 F s~ ,i5) = 12.38 0.0023 2 F~a.li) = 2.77 0.1470 F(2.11) = 0.900 0.4560 3 F~l.ll) = 5.23 0.0623 F. zt ,11~=24"10 0.0014 4 F(i,1 l) = 48.0 0.0002 F(z.1 l) = 48.53 0.0004
Table 3. The significance (F-ratio and P values) of sun/shade and species in determining the succulence of mangrove leaves at four sites
Site Sun/shade Species
F P F P
1 F(i.15) = 0.00 0.9603 F(3,15j = 14.05 2 F~l.lx) =0.12 0.7412 F(2 ll)= 25.42 3 F~i.li)= 1.99 0.2078 F(2111)= 0.050 4 Fll.ll) = 1.83 0.2253 F(2.ii) = 15.36
0.0015 0.0012 0.9559 0.0044
~'E 005
O90~ 0,04
0.03 Site 1 Site 3 Site 4
Fig. 7a, b. Mean contents of soluble phenolic (a), and leaf suc- culence (b) of sun (solid) and shade (hatched) leaves of Rhizophora stylosa growing at three sites in north Queensland. LSD is the least significant difference (P< 0.05) between any two means
Tha t is, approx. 23 % o f the leaf tissue dry weights are phenolics.
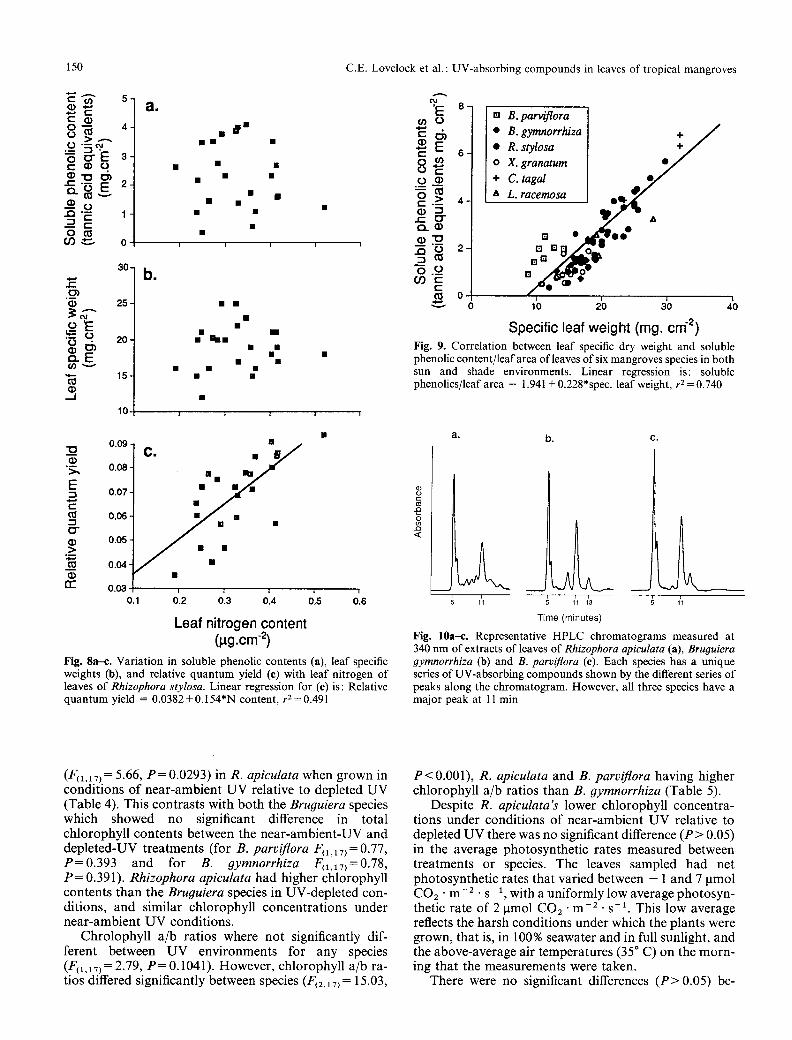
U V and species effects. Representat ive H P L C ch roma to - grams o f extracts o f leaves o f three m a n g r o v e species are shown in Fig. 10. Each species had its own unique series o f UV-abso rb ing compounds . The peak at 11 rain was the largest c o m p o n e n t o f the UV-abso rb ing c o m p o u n d s and was c o m m o n to all three species. Its absorp t ion spect rum matched that o f epidermal me thano l extracts (Fig. 4). Because the 11-min peak was the largest and mos t clearly resolved c o m p o n e n t o f the UV-abso rb ing c o m p o u n d s in all species, and because its absorp t ion spectra matched that o f the UV-abso rb ing p igment ex- t racted f rom leaf epidermal peels, the area o f this peak was used to compa re species. The identi ty o f this com- pounds is presently under investigation.
C o m p o u n d s absorb ing U V radia t ion were present in all three species and in bo th U V treatments (Table 4). Unde r near-ambient U V condi t ions all three species con- tained similar amoun t s o f UV-abso rb ing c o m p o u n d s on a leaf-area basis. However , the concen t ra t ion o f UV- absorb ing c o m p o u n d s o f B. parviflora was lower (F(1,17) = 7.66, P = 0.0132) in the deple ted-UV t rea tment than in the nea r -ambien t -UV t rea tment (Table 4). There was no significant difference in the UV-abso rb in g com- pounds between t reatments for B. 9ymnorrhiza (F(1,17)=0.36, P = 0 . 5 5 5 ) or R. apiculata (F(1,iT)=0.06, P = 0 . 8 1 6 ) .
Tota l ch lorophyl l contents were significantly lower
150 C.E. Lovelock et al. : UV-absorbing compounds in leaves of tropical mangroves
E ~ " s 2 E a . c o ~ ~ E 8-
e r e 3 6- ~-" o o �9 �9 �9 o c
o , m roB= I E . _ > 4 - �9 -
o r~O ~,.~ - 0 o ' ~
. . . . " ~ ' ~ 2 -
! "~ ~ 25- ~ o
~ v Fi '
�9 ~ 15 - su pl~ ;
10
a .
"O 0,09-
"--~>~ 0.08- C . �9
0o, " ' / : , Y [] [] �9
O"
o .o , . / _ " " It /
"~ o r e . / �9
B. parviflora B. gymnorrhiza + / R. stylosa + J X. granatum ~ / C. tagal , . y L. racemosa eQ.~ /
[] �9 j ~ P o o-
:o
lo a0 3'o 4;
S p e c i f i c l e a f w e i g h t (mg. cm 2) Fig. 9. Correlation between leaf specific dry weight and soluble phenolic content/leaf area of leaves of six mangroves species in both sun and shade environments. Linear regression is: soluble phenolics/leaf area = 1.941 +0.228*spec. leaf weight, r2=0.740
b. c.
(3,) I I
n- o.o3 i t i i 0.1 0.2 0.3 0.4 0.5 0,6
L e a f n i t r o g e n c o n t e n t (gg.cm -2)
Fig. 8a---e. Variation in soluble phenolic contents (a), leaf specific weights (b), and relative quantum yield (e) with leaf nitrogen of leaves of Rhizophora stylosa. Linear regression for (e) is: Relative quantum yield = 0.0382 + 0.154"N content, r 2 = 0.491
L l l l 1~3
Time (minutes)
5 1=1
Fig. 10a-e. Representative HPLC chromatograms measured at 340 nm of extracts of leaves of Rhizophora apieulata (a), Bruouiera 9ymnorrhiza (b) and B. parviflora (e). Each species has a unique series of UV-absorbing compounds shown by the different series of peaks along the chromatogram. However, all three species have a major peak at 11 min
(F(1,x7) = 5.66, P = 0.0293) in R. apiculata when grown in conditions of near-ambient UV relative to depleted UV (Table 4). This contrasts with both the Bruouiera species which showed no significant difference in total chlorophyll contents between the near-ambient-UV and depleted-UV treatments (for B. parviflora F(1,17)=0.77, P = 0 . 3 9 3 and for B. 9ymnorrhiza F(m7)=0.78, P = 0.391). Rhizophora apiculata had higher chlorophyll contents than the Bruouiera species in UV-depleted con- ditions, and similar chlorophyll concentrations under near-ambient UV conditions.
Chrolophyll a/b ratios where not significantly dif- ferent between UV environments for any species (F(mv) = 2.79, P = 0.1041). However, chlorophyll a/b ra- tios differed significantly between species (F(z,17)= 15.03,
P<0.001) , R. apiculata and B. parvifiora having higher chlorophyll a/b ratios than B. 9ymnorrhiza (Table 5).
Despite R. apiculata's lower chlorophyll concentra- tions under conditions of near-ambient UV relative to depleted UV there was no significant difference (P > 0.05) in the average photosynthetic rates measured between treatments or species. The leaves sampled had net photosynthetic rates that varied between - 1 and 7 ~tmol COz �9 m - 2 . S - 1 , with a uniformly low average photosyn- thetic rate of 2 lamol CO2" m -2" s -1. This low average reflects the harsh conditions under which the plants were grown, that is, in 100% seawater and in full sunlight, and the above-average air temperatures (35 ~ C) on the morn- ing that the measurements were taken.
There were no significant differences (P>0 .05 ) be-

C.E. Lovelock et al. : UV-absorbing compounds in leaves of tropical mangroves 151
Table 4. Mean and standard errors of means for leaf characteristic of three mangrove species grown in conditions of ambient or depleted UV radioation
B. parviflora R. apiculata B. 9ymnorrhiza
Ambient Depleted Ambient Depleted Ambient Depleted
Relative concn. UV-absorving 2.53• 0.57 1.07• 0.28 2.86• 0.41 3.30• 0.64 3.48• 0.54 2.67• 0.36 pigments - m- 2 Total Chl 131 • 148 • 189 • 305 • 117 +11 135 • (lamol - m- 2)
Chla/b 3.23• 0.28 2.81• 0.16 3.14• 0.17 3.21• 0.15 2.63• 0.30 2.06• 0.12
Table 5. Means (n = 24 leaves) of leaf characteristics of three mangrove species grown in acrylic-sheeting growth cabinets. F-ratio test statistic (df = (2,27)) and probability are shown for the comparison between species
B. parviflora R. apiculata B. 9ymnorrhiza
Leaf area 20.49 (cm -2)
Leaf display 74.33 (% leaf area) Succulence 0.0291 (g H20" cm -2) (V + A + Z)/Chl 215.8 (mmol �9 mol- 1) Total carotenoids/Chl 682.3 (mmol �9 tool- 1)
37.35 39.26 F= 52.05 P<0.001
23.18 63.68 F= 20.66 P<0.001
0.0391 0.0520 F= 26.70 P<0.001
98.40 130.6 F= 11.71 P<0.001
432.5 502.3 F= 15.94 P<0.001
tween UV treatments for leaf size, angle, succulence, total carotenoids/chlorophyll or (V + A + Z)/chlorophyll contents. However, there were significant ( P < 0.05) dif- ferences between the species in these leaf characteristics (Table 5). Bruguiera parviflora had the smallest leaves, the least succulence and the greatest leaf area displayed on a horizontal plane (most horizontal leaf angles). It also had the highest ratios of (V+ A + Z)/chlorophyll and total carotenoids/chlorophyll. Conversely, R. api- culata had the most-vertical leaf angles, lowest (V + A + Z)/chlorophyll ratio and lowest carotenoid/ chlorophyll ratio. Bruguiera 9ymnorrhiza had inter- mediate characteristics for all variables, except for suc- culence levels where it has the highest value with an average of 0.0520 g H 2 0 �9 cm -2.
Discussion
The distribution of phenolic compounds in the epidermis of leaves of mangrove species (Figs. 1, 2), and the light- absorption characteristics of phenolic compounds in the epidermis of such leaves (Figs. 3, 4) indicate that these compounds, which are probably flavonols, could protect mangrove leaves from the damaging effects of UV radia- tion. However, the occurrence of these compounds in leaf mesophyll tissue, where penetration of UV radiation would be low, indicates other factors may be involved in promoting the production of these compounds (see dis- cussion below).
Sun leaves are exposed to higher levels of UV (Cald-
well et al. 1983) and visible radiation than shade leaves. The higher levels of soluble phenolic compounds and succulence in sun leaves compared with shade leaves in the mangroves surveyed here implicate succulence and phenolic compounds in providing protection form UV radiation (Caldwell et al. 19 83). The correlation observed between phenolic contents per leaf area and dry weights per leaf area across sun and shade conditions (and all mangrove species) (Fig. 9) was probably due to the cor- relation that is known to occur between specific weight of leaves and the light conditions under which they are grown (Givnish 1988). In soybean grown at high visible- light levels, Warner and Caldwell (1983) showed that leaf phenolic contents and leaf thickness were greater than in plants grown at low light levels. Soybean plants grown at high visible-light levels showed no reduction in photosynthetic rates after 5 h of high-level UV-B irra- diance, whereas those grown at low light levels suffered photosynthetic damage. The insensitivity of high-light- grown plants to UV-B radiation was attributed to higher phenolic contents in conjunction with thicker leaves. Tevini et al. (1991) found no indication of photosynthetic damage from UV-B radiation in rye plants in which the accumulation of UV-absorbing compounds had been stimulated by pretreatment with UV-B radiation. Thus, the presence of UV-absorbing compounds and probably leaf succulence could contribute substantially to the at- tenuation of UV radiation and hence protection from UV radiation in mangrove species.
The large effect of site on the accumulation of phenol- ic compounds in sun leaves (Fig. 7), particularly between

152 C.E. Lovelock et al. : UV-absorbing compounds in leaves of tropical mangroves
sites in the same river system (i.e. sites 1 and 3), indicates that factors other than exposure to direct sunlight may also influence the accumulation of UV-absorbing phenolic compounds. The Bowling Green Bay site (Site 4, Table 1, Fig. 5) at which plants had the highest levels of UV-absorbing compounds is the most arid of the sites surveyed. High soil salinities result in highly conservative water use in mangroves, which depresses photosynthetic rates during periods of high solar radia- tion and high leaf temperatures (Ball et al. 1988; Clough and Sim 1989). It is possible that decreased photosyn- thetic rates caused by unfavourable water relations asso- ciated with high soil salinities or other environmental stress could influence the production of UV-absorbing compounds in mangroves. Support for this view comes from the results of experiments in which additional en- vironmental stresses that are known to reduce photosyn- thetic rates were combined with UV-radiation treat- ments. Generally, in experiments in which additional environmental stress was combined with increased UV-B radiation, photosynthetic rates were less affected than when plants were exposed to UV-B radiation alone (Murali and Teramura 1986, 1987; Cen and Bornman 1990; Larson et al. 1990) and this effect was often asso- ciated with increases in the concentration of UV-absorb- ing pigments in leaves (Cen and Bornman 1990; Larson et al. 1990).
Differences in the concentration of UV-absorbing compounds between sites could also be caused by other factors, for example, soil nutrient status or the degree of tidal innundation. The influence of nitrogen availability on the concentration of UV-absorbing compounds was assessed in R. stylosa growing in a shadehouse environ- ment. Leaf nitrogen contents have been shown to cor- relate with increases in the quantum yield and net photosynthetic rates of plants (Evans and Terashima 1987; Kfippers et al. 1988). Since the relative quantum yield of mangrove leaves increased with no correspond- lng increase in the concentration of UV-absorbing com- pounds (Fig. 8) it seems unlikely the accumulation of these compounds responds to increases in leaf productiv- ity. Thus, although the accumulation of UV-absorbing compounds in mangroves is higher in sun leaves than shade leaves, this is probably not associated with in- creases in photosynthetic rates in sun leaves relative to shade leaves (see review by Givnish 1988), but is more likely the consequence of higher UV radiation levels absorbed by sun leaves compared with shade leaves (Caldwell 1981). Alternatively, it could be related to increases in excess visible solar radiation and decreases in photosynthetic rates resulting from environmental stress such as photoinhibition or water deficiencies. In order to clarify whether UV radiation directly influences the accumulation of UV-absorbing compounds in man- groves, as has been shown in other plant species (e.g. Wellman 1983; Tevini et al. 1991), the effect of UV radiation on three mangrove species was assessed.
Exposure to UV radiation significantly increased the level of UV-absorbing compounds in B. parviflora rela- tive to the UV-depleted treatment (Table 4). This agrees with the results of Wellman (1983) and Tevini et al.
(1991), who found that accumulation of UV-absorbing compounds was correlated with increasing UV-B dose, possibly due to the stimulation of the production of flavonols by activation of key enzymes in their synthetic pathway (Wellman 1983; Tobin and Silverthorne 1985; Jain and Guruprasad 1990). Other factors, like develop- mental stage of tissue, wounding and the presence of pathogens, have also been found to influence the syn- thesis of flavonols (see review by Hahlbrock and Grisebach 1979). Thus, the presence of UV-absorbing compounds within the leaves of all three mangrove spe- cies, in the depleted-UV treatment, indicates that the accumulation of UV-absorbing compounds in mang- roves is not influenced by UV radiation alone, although UV radiation did affect the level of UV-absorbing com- pounds in B. parviflora. This implies a complex response to UV radiation that differs between species, and is in- fluenced by other environmental factors.
The experiment in which the influence of UV radia- tion was directly assessed was conducted at high levels of visible solar radiation. There is a known synergistic in- teraction between UV radiation and visible radiation. Greenberg et al. (1989) found that degradation of the 32-kDa core protein of PSII was greater with exposure to combined UV-B and visible radiation than in response to the added effects of individual UV-B and visible radia- tion treatments, and that UV-B radiation in normal sunlight was responsible for approx. 30% of the turnover of the 32-kDa protein. The authors suggested two photosensitizers are operating, chlorophyll for visible and infrared radiation and plastiquinone for UV-B radiation. There are also other reports that UV radiation may be utilized in photosynthesis (Bornman et al. 1984; Schlicter et al. 1985, 1986). Thus, UV-B and visible radia- tion seem to result in the same outcome although their effect is mitigated through different target molecules. If this is the case then the presence of "protectants" from excess levels of visible solar radiation, like the carote- noids, may be essential in moderating the effect of UV radiation in plants.
Carotenoids are known to be protective against the effects of both UV and visible-radiation damage (Knox and Dodge 1985; Salin 1987). The content of xantho- phyll-cycle components (V + A + Z), a group of the ca- rotenoids which has an energy-dissipating function (Demmig-Adams 1990), and total carotenoids in leaves were not affected by the level of UV radiation in these experiments (Table 5). However, species varied in their (V + A + Z)/chlorophyll and total carotenoid/ chlorophyll ratios. Both B. parviflora and B. 9ymnorrhiza had significantly greater (V + A + Z)/chlorophyll and to- tal carotenoid/chlorophyll ratios than R. apiculata. The species with high carotenoid levels showed no change in total chlorophyll contents in response to UV radiation. This indicates that the levels ofcarotenoids in mangroves may be crucial to avoiding any adverse effects of UV or excess visible radiation.
In addition to carotenoids, the photorespiratory path- way and the Mehler reaction may also contribute to the dissipation of excess solar radiation and thus can prob- ably reduce the likelihood of damage to the photosyn-

C.E. Lovelock et al. : UV-absorbing compounds in leaves of tropical mangroves 153
thetic appara tus (Powles 1984). In the Mehler reaction, super-oxide and hydrogen peroxide are produced and then detoxified. The UV-absorbing flavonols (Takahama 1983, 1984) have been found to have a high capacity to eliminate H202 in chloroplasts. Thus, it is plausible that the UV-absorbing compounds within mangrove leaves maybe involved in the detoxification of H202.
In terms of this study, species differences in the capac- ity for dissipation of excess visible solar radiation and removal o f destructive triplet chlorophyll and singlet oxygen may determine the sensitivity of species to UV radiation. Tha t is, B. parviflora may have withstood high radiation in the UV-depleted environment without a large activation of flavonol product ion because of the presence of high concentrations of (V + A + Z) and total carotenoids (Table 5). In the near-ambient-UV treatment the excess visible radiation energy may have reached a level where flavonol product ion was increased to the level of the other species. In R. apiculata, protective processes may have been saturated in the UV-depleted conditions. Thus, an increase in UV radiation asorbed by the leaves under the near-ambient-UV treatment may have resulted in lower chlorophyll levels due to photooxidat ion of chlorophyll. The difference between the response of B. 9ymnorrhiza and R. apiculata (which had similar levels of UV-absorbing compounds in conditions of both am- bient and depleted UV radiation may lie in the greater carotenoid concentrations and greater succulence (which increases the at tenuation of UV radiation) of B. 9ym- norrhiza leaves (Table 4).
Mangrove species varied in their response to UV radiation. Differences in the contents of UV-absorbing compounds between species and UV treatments could be due to the level o f carotenoids, and possibly to leaf succulence. By looking at the effects of visible radiation in the absence of UV radiation it was shown that the accumulat ion of UV-absorbing compounds in mangrove leaves occurs in the absence o f UV radiation, but that UV radiation stimulates the product ion of these com- pounds in some species. It is proposed that the accumula- tion of UV-absorbing compounds may be linked to the amount of UV and visible radiation energy absorbed by leaves.
Thank you to Dr. Bruce Chalker (AIMS) and Dr. Alistar Robert- son who have commented on this manuscript. We wish to acknow- ledge Glenn De'ath (JCUNQ) who helped with the statistics, Ian Ashworth who built the UV chambers and Otto Dalhaus for his expertise in rearing mangrove plants. During this study C.E.L. was supported by an Australian Institute of Marine Science Post Graduate Student Fellowship. This is an Australian Institute of Marine Science publication, No. 570.
References
Allen, S.E., Grimshaw, H.M., Parkinson, J.A., Quarmby, Q. (1974) Chemical analysis of ecological materials. John Wiley and Sons, New York, USA
Ball, M.C., Cowan, I.R., Farquhar, G.D. (1988) Maintenance of leaf temperature and the optimization of carbon gain in relation to water loss in a tropical mangrove forest. Aust. J. Plant Physiol. 15, 263-276
Bjrrkman, O., Demmig, B. (1987) Photon yield of 0 2 evolution and chlorophyll fluorescence characteristics at 77K among vascular plants of diverse origins. Planta 161, 490-504
Bornman J.F., Bjorn, L.O., Akerlund, H.E. (1984) Action spectrum for inhibition by ultraviolet radiation of photosystem II activity in spinach thylakoids. Photobiochem. Photobiophys. 8, 305-313
Caldwell, M.M. (1981) Plant response to solar ultraviolet radiation. In: Encyclopedia of Plant Physiology, N. S., vol. 12A; Physio- logical plant ecology. I. Responses to the physical environment, pp. 169-198, Lange, O.L., Nobel, P.S., Osmond, C.B. and Zieg- ler, H. eds. Springer-Verlag, New York
Caldwell, M.M., Robberecht, R. Flint, S.D. (1983) Internal filters, prospects for UV-B acclimation in higher plants. Physiol. Plant. 58, 445-450
Caldwell, M.M., Teramura, A.H., Tevini, M. (1989) The changing solar ultraviolet climate and the ecological consequences for higher plants. Trees 4, 363-367
Cen, Y.P., Bornman, J.F. (1990) The response of bean plants to UV-B radiation under different irradiances of background vis- ible light. J. Exp. Bot. 41, 1489-1495
Clough, B.F. (1984) Growth and salt balance of the mangroves Avicennia marina (Forsk.) Vierh. and Rhizophora stylosa Griff. in relation to salinity. Aust. J. Plant Physiol. 11, 419-430
Clough, B.F., Sim, R.G. (1989) Changes in gas exchange charac- teristics and water use efficiency of mangroves in response to salinity and vapour pressure deficit. Oecologia 79, 38--44
Demmig-Adams, B. (1990) Carotenoids and photoprotection in plants: A role for the xanthophyll zeaxanthin. Biochim. Biophys. Acta 1020, 1-24
Evans, J.R., Terashima, I. (1987) Effects of nitrogen nutrition on electron transport components and photosynthesis in spinach. Aust. J. Plant Physiol. 14, 59-68
Flint, S.D., Jordan, P.W., Caldwell, M.M. (1985) Plant protective response to enhanced UV-B radiation under field conditions, leaf optical properties and photosynthesis. Photochem. Photo- biol. 41, 95-99
Frederick, J.E., Snell, H.E., Haywood, E.K. (1989) Solar ultraviolet radiation at the earths surface. Photochem. Photobiol. 50, 443-450
Givnish, T.J. (1988) Adaption of sun and shade: a whole-plant perspective. Aust. J. Plant Physiol. 15, 63-92
Greenberg, B.M., Gaba, V., Canaani, O., Malkin, S., Mattoo, A.K., Edelman, M. (1989) Separate photosensitizers mediate degradation of the 32-kDa photosystem II reaction center pro- tein in the visible and UV spectral regions. Proc. Natl. Acad. Sci. USA 88, 6617-6620
Hahlbrock, K., Grisebach, H. (1979) Enzymic controls in the bio- synthesis oflignin and flavonoids. Annu. Rev. Plant Physiol. 30, 105-130
Jain, V.K., Guruprasad, K.N. (1990) Involvement of riboflavin in UV-A and white light-induced synthesis of anthocyanin in Sorohum bicolor. J. Exp. Bot. 41, 53-58
Johansen, D.A. (1940) Plant microtechniques. McGraw-Hill, New York
Juhler, R.K., Cox, R.P. (1990) High performance liquid chromato- graphic determination of chloroplast pigments with optimized separations of lutein and zeaxanthin. J. Chromatogr. 508, 232-235
Knox, J.P., Dodge, A.D. (1985) Singlet oxygen in plants. Phyto- chemistry 24, 889-896
K/ippers, M., Koch, G., Mooney, H.L. (1988) Compensating effects to growth of changes in dry matter allocation in response to variation in photosynthetic characteristics induced by photoperiod, light and nitrogen. Aust. J. Plant Physiol. 15, 287-298
Larson, R.A., Garrison, W.J., Carlson R.W. (1990) Differential responses of alpine and non-alpine Aquilegia species to increases ultraviolet-B radiation. Plant Cell Environ. 13, 983-987
Les, D.H., Sheridan D.J. (1990) Biochemical heterophylly and flavonoid evolution in north American Potamogeton (Pota- mogetonaceae). Am. J. Bot. 77, 453-465

154 C.E. Lovelock et al. : UV-absorbing compounds in leaves of tropical mangroves
Lovelock, C.E. (1992) Influence of solar radiation and leaf angle on the xanthophyll concentration in leaves of mangroves. Oecologia, in press
Mole, S., Waterman, P.G. (1987) A critical analysis of techniques for measuring tannins in ecological studies. I. Techniques for chemically defining tannins. Oecologia 72, 137-147
Murali, N.S., Teramura, A.H. (1986) Effectiveness of UV-B radia- tion on the growth and physiology of field grown soybean modified by water stress. Photochem. Photobiol. 44, 215-219
Murali, N.S., Teramura, A.H. (1987) Sensitivity of soybean photosynthesis to ultraviolet-B radiation under phosphorous deficiency. J. Plant. Nutr. 10, 501-515
Murali, N.S., Teramura, A.H., Randall, S.K. (1988) Response differences between two soybean eultivars with contrasting UV- B radiation sensitivities. Photochem. Photobiol. 48, 653-657
Negash, L. (1987) Wavelength-dependence of stomatal closure by ultraviolet radiation in attached leaves of Eraorostis tef, action spectra under backgrounds of red and blue lights. Plant Physiol. Biochem. 25, 753-760
Poovachiranon, S., Boto, K.G., Duke, N.C. (1986) Food preference studies and ingestion rate measurements of the mangrove amphipod Parhyale hawaiensis (Dana). J. Exp. Mar. Biol. Ecol. 98, 129-140
Powles, S.B. (1984) Photoinhibition of photosynthesis induced by visible light. Annu. Rev. Plant Physiol. 35, 15-44
Robberecht, R., Caldwell, M.M. (1978) Leaf epidermal transmit- tance of ultraviolet radiation and its implications for plant sensitivity to ultraviolet-radiation induced injury. Oecologia 32, 277-287
Robertson, A.I. (1988) Decomposition of mangrove leaf litter in tropical Australia. J. Exp. Mar. Biol. Ecol. 116, 235-247
Salin, M.L. (1987) Toxic oxygen species and protective systems of the chloroplast. Physiol. Plant. 72, 681-689
Schlicter, D., Weber, W., Fricke, H.W. (1985) A chromatophore system in the hermatypic deep-water coral Leptoseris fragilis (Anthozoa: Hexacorallia). Mar. Biol. 89, 143-148
Schlicter, D., Fricke, H.W., Weber, W. (1986) Light harvesting by wavelength transformation in a symbiotic coral of the Red Sea twilight zone. Mar. Biol. 91, 403-408
Strid, A., Chow, W.S., Anderson, J.M. (1990) Effects of supplemen- tary ultraviolet-B radiation on photosynthesis in Pisum sativum. Biochem. Biophys. Acta 1020, 260-268
Swain, T. (1976) Flavonoids. In: Chemistry and biochemistry of plant pigments, vol. 2, 2nd edn. pp. 166-207, Goodwin, T.W., ed. Academic Press, London
Takahama, U. (1983) Redox reactions between kaempferol and illuminated chloroplasts. Plant Physiol. 71, 598-601
Takahama, U. (1984) Hydrogen peroxide-dependent oxidation of flavonols by intact chloroplasts. Plant Physiol. 74, 852-855
Teramura, A.H. (1983) Effects of ultraviolet-B radiation on the growth and yield of crop plants. Physiol. Plant. 58, 415-427
Teramura, A.H., Biggs, R.H., Kossuth, S. (1990) Effects of ul- traviolet-B irradiances on soybean. Plant Physiol. 65, 483-488
Tevini, M., Braun, J., Fieser, G. (1991) The protective function of the epidermal layer of rye seedlings against ultraviolet-B radia- tion. Photochem. Photobiol. 53, 329-333
Tevini, M., Teramura, A.H. (1989) UV-B effects on terrestrial plants. Photochem. Photobiol. 50, 479-487
Tobin, E.M., Silverthorne, J. (1985) Light regulation of gene ex- pression in higher plants. Annu. Rev. Plant Physiol. 36, 569-593
Warner, C.W., Caldwell, M.M. (1983) Influence of photon flux density in the 400-700 nm waveband on the inhibition of photosynthesis by UV-B (280-320 nm) irradiation in soybean leaves, separation of indirect and immediate effects. Photochem. Photobiol. 38, 341-346
Wellman, E. (1983) UV radiation in photomorphogenesis. In: Encyclopedia of Plant Physiology, N. S., vol. 16B: Photomor- phogenesis, pp. 745-756, Shropshire, W., Mohr, H., eds. Springer-Verlag, New York
Copyright © 2022 FDOKUMEN




















