
Human Resource Management Strategic Human Resource Management

1
Quality Over Quantity ‘Dissimilar Fragmentation of Human and Animal Remains. Implications for the Analysis of Commingled Human Bone: A Comparative Study.’ Abstract The point of this paper is to assess methods for recording commingled human remains, identify
weaknesses or flaws within them, and submit modifications aimed at improvement. The human
zonation method is examined and its premise is found to be flawed because human and faunal
material fragment in dissimilar ways. Human culture greatly influences the dissimilar
taphonomic processes which human and animal remains experience, resulting in dissimilar
types of fragmentation and assemblage formation. The zones identified for recording purposes
in the KO method are inconsistently justified and the greater level of detail given to the
postcranial skeleton, over the skull, leads to recording biases. The SDZ method revises the
cranial zones using personal observation of fragmentation trends, relative density, and
likelihood of element survival. The SDZ method is applied to a commingled and fragmented
assemblage along side the KO method to test the difference in accuracy of recording through
the estimation of MNI and MNE.
Acknowledgements Special thanks to Dr. Elizabeth CraigAtkins and Dr. Diana Swales for their input and guidance during this project. Thank you to my personal tutor Dr. Umberto Albarella, my friends and peers from the Osteoarchaeology Msc course, and Louise Loe. To Matthew and my family for all their encouragement. Especially my mother who was my constant cheerleader and proofreader from halfway around the world. Table of Contents i. Title Page and Abstract ii. Acknowledgements iii. Table of contents iv. List of Figures v. List of Tables vi. List of Appendices
1. Introduction

2
1. Aims and Objectives 2. Background
1. Review of Methods 1. Forensic Methods 2. Zooarchaeological Methods 3. Osteological Methods
2. Weaknesses of the KO Method 3. Taphonomy
1. Intrinsic Factors Influencing Survival and Preservation. 1. Chemical Makeup 2. Size and Shape 3. Density
2. Extrinsic Factors Influencing Formation and Fragmentation. 1. Environment and Faunal 2. Consumption Practices 3. Burial and Disposal Practices
4. Developing the SDZ method. 1. Personal Observation of Cranial Fragmentation 2. Element Survivorship
1. Ossuary Examples 3. Correlation between Density and Survivorship
5. Methods and Materials 1. Materials
1. Black Gate Cemetery 2. Commingled Sample
2. Methodology 1. Frontal 2. Temporal 3. Occipital 4. Mandible
6. Results and Discussion 1. Raw data 2. Assessment of the SDZ method 3. Quantitative Techniques 4. MNI and MNE 5. Incomplete Zones 6. Unidentifiable Fragments 7. Demographic Factors 8. Potential bias

3
7. Conclusions and Future Research 1. Summary of Results and Interpretations 2. Implications for Zonation Style Recording 3. Future Research
8. Bibliography 9. Appendix
List of Figures 1. KO anterior view of crania. 2. SDZ, anterior view of frontal. 3. KO, lateral view of crania. 4.1. SDZ, sectioned temporal. 4.2. SDZ, inferior view of left temporal. 5.1. KO, inferior view of the occipital. 5.2. KO, posterior view of the occipital. 6. SDZ, endocranial view of the occipital. 7.1. KO method, mandible. 7.2. SDZ method, mandible. 8. Comparative MNE for Cranial elements. List of Tables 1.1 Skull and Mandible Element Frequencies 1.2 Ossuary I 1.3 Ossuary II 2.1. KO Method Element Frequencies 2.2. SDZ Method Element Frequencies Appendix A. Zone frequencies, SDZ method. B. Raw data from the SDZ method. 1. Introduction
Standard methods of appropriately recording bone are vital for archaeological
interpretations, and forensic cases. In recent years this need has been addressed in a number

4
of ways, but none of which are suitable to the recording of commingled and fragmentary human
remains. The majority of skeletal analysis methodologies in osteoarchaeology are best suited
to articulated remains. Commingled and fragmented assemblages are often considered to be of
little scientific value and the analysis constricted to basic quantification and inventory (Brickley
and Mckinley 2004, 14). However, this prejudice towards disarticulated remains is incorrect. In
fact the analysis of commingled material can give great insight into mortuary practices and
element survivorship studies (Buikstra and Ubelaker 1994, 5; Waldron 1987, 56; Ubelaker and
Rife 2008, 105). This means that methods for the recording of disarticulated and fragmentary
assemblages need to be accurate and critiqued.
The bulk of published material regarding commingled remains is either specific to
zooarchaeology or forensic anthropology. These types of assemblages are important and their
methods considerably reliable, but they can not be directly applied for use with archaeological
assemblages. A review of the available techniques for recording commingled human remains
identifies the human zonation method, as the principal technique. The human zonation method,
henceforth referred to as the KO method, is suited to the inventory of large and fragmentary
commingled assemblages, but it has not been critiqued in the published literature (Knüsel and
Outram, 2004). The KO method will be evaluated for weakness and tested for bias using a
comparative method developed in this paper.
This paper is intended to fill a gap in the osteoarchaeological literature through a critique
of the KO method. The preeminent flaw in the KO method is the assumption that human and
animal remains fragment in similar ways. This assumption is addressed with a review of
taphonomy and the intrinsic and extrinsic differences between the two groups. The KO method
suffers from a recording bias which negatively skews the quantitative assessments of the
cranial data. This paper puts forth a revised technique for recording commingled fragments of
the human crania in the form of the skull diagnostic zone method. The skull diagnostic zone
method will be referred to as the SDZ. The SDZ method is designed to accurately record the
features of the crania which would most likely be the best representative of MNI. The SDZ
method will be comparatively tested against the KO method to assess difference and
improvement. Zones with a greater degree of detail are recorded with the expectation that they

5
will result in greater frequencies for particular features of the crania. Improvement in this
instance is deemed to be the acquisition of more specific data which leads to more accurate
calculation of MNI.
The SDZ method is justified using observed trends of cranial fragmentation, relative
density, and element survivorship data. The revised zones can be used to gain relatively more
specific element frequency data and contribute to commingling analysis as a whole. The use of
a few highly recognizable and high survivorship zones decreases the rate of observer error and
speeds familiarity with the method.
Commingled remains can be quantitatively evaluated by estimating the minimum number
of individuals (MNI) and minimum number of elements (MNE) in an assemblage. Some
demographic and pathological information can also be gleaned from disarticulated remains
depending upon the level of fragmentation present. This paper will demonstrate how
commingled assemblages can be useful as samples to test comparative methodologies and to
establish rates of element survivorship. All of this potential information means that commingled
remains should be viewed as a valuable resource and not excluded from skeletal analysis.
Comparative studies are important for the evaluation and improvement of all
methodologies. Recording systems are the most vital to critique and revise because accurate
quantification is based on accurate and representative recording. Any interpretation of data is
only as reliable as its weakest link. More efficient and accurate recording methods could lead to
an increased use of commingled remains and could open an avenue to unique research
questions.
1.1. Aims and Objectives.
1. Review the available methods for the recording of commingled remains. 2. Evaluate the KO method for strengths and weaknesses. 3. Demonstrate that human skeletal material fragments in predictable ways which are
dissimilar to how faunal bones tend to fragment using taphonomy. 4. Characterize how human cranial material, in particular, tends to fragment. 5. Explore the influence of density on element survivorship. 6. Discuss the implications for the application of commingled assemblage recording
methods. 7. Develop a comparative zonation method using personal observations, relative density,
and element survivorship for the human crania. 8. Test the accuracy of the KO and SDZ comparative methodologies.

6
9. Discuss the results, potential bias, and avenues of future research. 2. Background 2.1. Review of Methods
Establishing MNI is a crucial function of any preliminary skeletal analysis, regardless of
discipline, and is attempted through recording and quantification of elements. The ways in
which an assemblage is recorded vary between disciplines and type, with varying degrees of
success. The need for commingled assemblages to be treated differently from articulated
remains is a recognized but not well addressed challenge. Standard skeletal recording
methods call for the use of inventory sheets and cannot be used with confidence when
assessing disarticulated remains. This disconnect is because inventory sheets are better suited
for use with articulated individuals and associating elements into individuals is difficult to do
reliably (Brickley and McKinley 2004; Kendell and Willey 2014, 86). This difficulty is increased
with scale, in very large archaeological assemblages, reassociating elements may be
impossible (Kendell and Willey 2014, 86). Greater emphasis needs to be placed on recording
and labeling skeletal elements as they are excavated, in this way, the contexts of commingled
remains can be understood with a recreation of past events and understanding of mortuary
behaviors (Roksandic 2002, 109; Saul and Saul 2002, 73). The following sections outline
methods of recording and assessing disarticulated remains from modern forensic study,
zooarchaeology, and osteoarchaeology.
2.1.1. Forensic Methods.
The relationship between forensic and archaeological methods should be symbiotic, with
information and tools of analysis passing both ways, this requires shared knowledge and
cooperation between the disciplines (Saul and Saul 2002, 78). Forensic methods may be
applied to evaluate assemblages in cases of genocide, natural disaster, or conflict and these
assessments usually have much more at stake than archaeological ones. Forensic methods
usually have end goals of personal identification, repatriation, and evidence gathering which are
reached by associating isolated elements to individuals and building biological profiles.
Forensic methods of recording and sorting can be very accurate and lead to accurate estimates
of MNI, but are better suited to small scale assemblages. When recording forensic material
every effort is made to assign disarticulated remains to the correct individual.

7
Smallscale commingling, from forensic contexts is generally assessed with the use of
morphological techniques including; visual pair matching, osteometric and taphonomic sorting,
and a process of elimination (Adams and Byrd 2004, 139; Ubelaker 2002, 332). These
methods aid in the sorting, reassociating, and recording of disarticulated elements. Osteometric
sorting can pairmatch elements by taking the dimensions of the bones, including
measurements of length and width for morphological features, and weight of the element
(Adams and Byrd 2004, 139; Ubelaker 2002, 334). These techniques require relatively whole
elements so can not be applied to very fragmented remains. The use of taphonomic sorting on
a large cemetery collection has the potential to create patterns where none exist through
observer bias. Forensic methods of recording often rely on visually assessing the material all
at once and are not feasible for many archaeological samples. When recording archaeological
collections, attempting to pair match elements while assessing thousands of fragments is time
consuming and little more than guesswork. For this reason, they are not commonly applied to
archaeological assemblages of commingled remains.
2.1.2. Zooarchaeology Methods.
The discipline of zooarchaeology is build upon the analysis of commingled faunal
assemblages and uses a range of recording techniques. The faunal diagnostic zone method for
recording fragmented and commingled remains relies upon the premise that whole bones can
be divided into individual, identifiable morphological zones (Dobney and Rielly 1988, 79). The
strengths of the faunal diagnostic zone method are; flexibility of use, ease of recording features
through the use of standardized zones, and reliance on nonrepeating elements. These
qualities make the diagnostic zone method useful for quantification applications like MNI or
MNE plus this method has the potential for use on a wide range of taxa. The diagnostic zone
premise can be applied to the recording of human assemblages since the use of extremely
detailed morphological features is already widely accepted in human Osteology.
The diagnostic zone method supplies diagrams of the zonation for the mandible and
postcranial skeletons of common archaeological fauna such as bos, sus, equus, and caprines
(Dobney and Rielly 1988, 81). In this method, zones are recorded on a basis of presence,
absence or incompleteness, with incomplete zones being marked as more or less than 50%

8
complete. Dobney and Rielly assert that the general principles of the diagnostic zone method
should be readily adaptable for other species, and that the results of using this method are
repeatable (1988, 81). The key word in that statement is adaptable, this could be interpreted to
mean either that the method could be directly applied to any group or that adaptational changes
to suit morphologically dissimilar taxa would be required.
Additional methods used to record commingled faunal assemblages rely upon vague
codes for anatomical parts of each bone, such as proximal or distal portions of the element.
Methods like this, provide less detail and accuracy, than the diagnostic zone method, when
recording fragments. A comparative method from Morlan looked at the distinction between two
recording methods, one based on vague element portions and one developed with relatively
more specific ‘portions’. Morlan estimated the MNI for each method to compare the accuracy of
the recording. He used MNI because it was considered to be more accurate for showing
‘abundance’ within data sets than other qualitative techniques (1994, 797). Morlan’s results
supported the need for a specific and detailed recording system and encouraged the use of
specific morphological features in recording. The data was used to examine element
frequency, element survivorship, the correlation between survivorship and volume density, and
percentages of completeness (Morlan 1994, 798). Though the methodologies tested were
developed for use on faunal material the lessons learned can be applied to Osteoarchaeological
recording.
The discipline of zooarchaeology regularly works with commingled assemblages and has
a number of techniques pertaining to the recording of fragmented material. In recent years
osteoarchaeologists have drawn from the field of zooarchaeology and constructed similar
recording methods.
2.1.3. Osteology methods.
A review of the Osteoarchaeological recording techniques best suited to commingled
human assemblages has one standout, the KO method. The KO method, as applied to
fragmentary human remains, addresses the need for a recording system suited to disarticulated
and fragmented human assemblages (Knüsel and Outram 2004, 85). The method was almost
directly adapted from the faunal diagnostic zone method, because it addressed similar problems

9
of analysis. Knüsel and Outram borrowed the zonation for the human mandible and postcrania
from the faunal method, some adjustments were made to elements which are morphologically
distinct from those in faunal biology. Zones were created for elements which lacked a
preexisting model, specifically the crania, fibula, clavicle, and sternum (Knüsel and Outram
2004, 86). The use of standard zones for recording fragmented remains in the KO method
allows infinitely more detail and thus a greater degree of precision than previous methods.
Compared to other Osteological inventory methodologies, the KO method‘s strength is in
its ability to record elements as a series of zones so that the frequency of specific features can
be assessed. Each fragment can be recorded by zone with any additional information required
or observed, and later analyzed in a database. These strengths mean that data can be
collected for sorting and interpretation without the complication of attempting to lay out many
hundreds of elements and fragments at the same time. Earlier attempts at recording
commingled remains in Osteoarchaeology were relatively less specific and usually recorded
fragments using anatomical directional terms, eg: the proximal or distal portions of an element.
The number of zones, in the KO method, ‘enhances resolution’ for studies of element
survivorship and fragmentation (Outram et all. 2010, 1702).
A precedent for modifying the KO method exists in the form of a note in the original
paper. Louise Loe added a zone for the basilar process in the occipital and a zone for each
petrous portion of the temporals, in addition Loe combined some of the zones of the fibula
(Knüsel and Outram 2004, 96). Another mention of the modifications in the 5th edition BABAO
publication are in regards to a procedure for recording specific features (Loe and Cox 2005, 14).
Without any published details or results, discerning the effect of Loe’s modifications on accuracy
of recording is difficult. The lack of published criticism and testing performed on the KO method
is a breach of the scientific process which is only partly addressed in this paper.
2.2. Critiquing the KO method.
Despite its strengths, the KO method has several apparent weaknesses. The KO
method assumes that human and animal remains fragment in similar ways and exists on the
basis of this assumption. At no point does the KO method justify this reasoning. The KO

10
method does not account for the unique taphonomic processes which human and animal
remains are subjected to, or their intrinsically dissimilar morphologies.
The KO method created zones for the elements that were not included in the faunal
diagnostic zone method, but does not justify how or why the zonation is the way it is. The
process used to identify the new zones as diagnostically important needs to be explained.
Without justification there is a possibility that the zones are not representative of how human
remains, or any remains, tend to fragment. The method does not take into account the way in
which the human crania tends to fragment or the likely survivorship of individual elements. The
likely survivorship of isolated cranial elements is not considered. In a fragmented assemblage
most cranial zones will be recorded as incomplete, even if they are greater than 50% complete
or include a non repeating feature.
At first glance the crania seems to have been divided into zones by element, however not
every bone is assigned an individual zone. This is the case for the lacrimal, ethmoid, and
endocranialnasal bones. The KO method calls for the recording of all fragments but, the way in
which this is done is inconsistent. In the skull some elements a not even assigned an individual
zone while others have multiple. Such is the case for the frontal and maxillary bones, which
have zones for the left and right sides respectively. This inconsistency is not explained and
may result in confusion about how to record endocranial, elements like the ethmoid, if they are
recovered in isolation.
Final weaknesses within the KO method relate to its lack of development. The KO
method does not adequately address the morphological dissimilarity of juvenile remains or how
to record zones which have not yet developed. The recording of unfused epiphyses is not
explained, so this is likely to be an area of both intra and interobserver error. In a large sample
there may be hundreds of small, nondiagnostic, incomplete fragments to record, which is time
consuming and ultimately these fragments will not usually factor into quantitative assessments.
The practice of refitting fragments is common in Osteological analysis, but its use is not
specified in the KO method. This could lead to interobserver error, if recorders use inconsistent
refitting practices or have differing levels of skill and patience for the technique.

11
The weaknesses identified within the KO method cannot all be addressed at once. This
paper will begin with an investigation into the assumption that human and animal bones
fragment in similar ways. A review of taphonomy is necessary to establish how morphological
and external factors affect fragmentation within and between groups. Then justification for a
revision of the cranial zonation will be made using element survivorship,observed trends of
fragmentation, and relative density. The resulting SDZ method, with modified zonation, will be
tested against the KO method.
Within human and faunal skeletons there are hundreds of diagnostic features, which
have different rates of survivorship due to intrinsic factors like size, shape, and density (Waldron
1987, 60). This knowledge of differential element survival can be used to predict features which
may be the best representations of the sample’s MNI.
Developing osteological methodologies from existing zooarchaeological techniques is
both useful and practical when the assumption is that human and animal remains are similar.
However, if the materials are dissimilar in nature, then the questions which need to be asked
and the information acquired may change.
3. Taphonomy
Taphonomy is the study of all the potential agents and processes affecting bone, it is the
crux to understanding why the fracture of human and animal bone is dissimilar (Lyman 1994).
Essentially taphonomy is used to identify the factors which cause the loss of data within a
skeletal deposit between the time of death and recovery, this loss is known as attrition. Sources
of attrition include anything which causes a decline in the size and quality of an assemblage
(Lyman 1994). Taphonomic study is necessary to discern the unique processes which act upon
remains and to explain how they reached their current state. Through an examination of the
dispersal, condition, and completeness of bones within an assemblage, archaeologists can
interpret the processes which may have acted upon the material.
Experimental archaeology has been crucial to the understanding of how bone is
changed over time, and in identifying causes of equifinality. Equifinality may cause
misinterpretations about the processes which have acted upon an assemblage, or explain why
two unique samples appear to exhibit similar taphonomy. Equifinality occurs when the same

12
effect is caused by differing factors, and taphonomy is usually a key to decoding this in skeletal
material (Lyman 1994, 38).
In taphonomic study there is a separation between intrinsic and extrinsic factors. Intrinsic
factors are all the morphological variations which can cause dissimilarities within a deposit of a
single taxa or between individuals (Henderson 1987, 44). Extrinsic factors are external to the
remains and act upon them, they include; human culture of burial and consumption practices,
environmental conditions, and faunal activity (Henderson 1987, 45).
3.1. Intrinsic factors.
If all external variables could be controlled for, human and animal remains would still
exhibit dissimilar rates of fragmentation and preservation when recovered. This is due to an
inherent dissimilarity of their intrinsic factors or morphological differences. Size, shape, density,
and chemical composition are intrinsic qualities which vary between and within taxa (Henderson
1987, 44). These intrinsic factors are fluid with age, health, and activity levels, which means
that even a sample of a single taxa contains several variables.
3.1.1. Chemical Makeup.
Human and animal remains will have different rates of survival without the influence of
external factors due to variation in bone composition. The chemical composition of human and
animal bone is dissimilar at a microscopic level Even within individuals bone composition is
varied, elements with greater levels of inorganic material, like tooth enamel, are more durable
and have a higher likelihood of survival (Reitz and Wing 2008, 39). Differences in cellular
structure are why observers are able to discern between human and animal bone at a
microscopic level.
The composition of bone can vary with pathology, poor health of disease may weaken
bone and make it more susceptible to fracture (Buikstra and Ubelaker 1994, 120). The
chemical makeup of bone is related to diet and varies with the type of nutrients and vitamins
which an individual ingests (Reitz and Wing 2008, 8385). Although the degree of influence this
could have on survivorship and fragmentation is likely to be statistically insignificant.
3.1.2. Size and shape.

13
The size and shape of elements influence the way in which bones fragment and are
identified. Bones from large taxa, like bos or equus, can fragment into more pieces than a bone
from a smaller species, like sus or ovis/capra. On one hand larger bones lend themselves to
greater rates of recovery, but on the other, the bones of large taxa can become unidentifiable
fragments at relatively larger sizes than those of smaller taxa (Watson 1972, 224). Fragments
of the same size from the same element of two dissimilar taxa, one large and one small, will
have different rates of identification.
This is because a fragment from a smaller taxa, though exactly the same size as a fragment
from a larger taxa, will have a greater chance of containing a morphologically diagnostic feature
(Watson 1972, 225).
Skeletal elements are usually identified as one of three morphological shape categories:
long, short, and irregular (Schultz 1997, 188). Variation in shape can affect the amount of force
needed to cause fragmentation in a bone; this results in different rates of fragmentation
between elements of dissimilar shapes. Size and shape of skeletal elements can alter
dramatically over the course of an individual’s lifetime, as well as between individuals within a
species (Scheuer and Black 2000; White and Folkens 2005, 31). Dissimilarity in bone
morphology affects how much surface area of the bone is exposed to external factors, which
can have an impact on preservation (Lyman 1994, 187).
3.1.3. Density.
Bone density is judged to be the intrinsic factor most likely to affect element survivorship
in commingled samples (Binford and Bertram 1977, 106; Galloway 1997, 295; Lyman 1994,
249; Waldron 1987, 62). Density is influenced by the sex, age, and health of an individual.
Sexual dimorphism influences the relative density of elements between male and female
individuals in a taxa, in humans greater density has been shown in males (Reitz and Wing
2008, 79; Galloway et all. 1997, 295).
A study comparing two similar faunal taxa; sheep and goat, showed differences in bone
density, but was unclear on the statistical significance of the difference to recovery rates and
element survivorship (Lyman 1994, 237). The density variation may be due to a true
morphological difference or the differences in measurement methods used. Long bone density

14
in a contemporary human sample showed significant differences between male and female
specimens and consistently recorded lower measures of density in the left humerus than the
right (Galloway et all. 1997, 295). These differences could be related to sexual dimorphism and
activity levels, as in modern populations a disproportionate number of people are right arm
dominate.
The exact measurement of bone density is difficult because factors such as length of
storage and climate can affect the density in specimens of study. When speaking about bone
density, it is more accurate to refer to an element as being relatively more or less dense than
another (Binford and Bertram 1977, 107). Additionally, exact measurements of bone density
will not be verbatim for those of the same elements from other individuals and could only be
used to create an ‘expected range’ for very specific samples. Difficulty exists in measuring the
density of specific features, because where features begin and end can be somewhat arbitrary
between observers. Density of complete elements is available in the clinical literature but
variation with age, sex, and pathology make this difficult to apply to an archaeological
assemblage. The majority of published bone density studies in the osteoarchaeological
literature tend to be in regards to the limbs.
The relationship between bone density and age means that over the course of a lifetime
density varies and can influence element survival and patterns of assemblage composition
(Binford and Bertram 1977, 108). A significant correlation of age at death and bone mineral
density influences element survivorship and ultimately MNI estimations (Kendell and Willey
2014, 88). Skeletons transform throughout life with growth and development and changing
conditions, this means that the relative density of elements could be dissimilar at different
growth stages. The variation of density in immature and juvenile remains prompted their
exclusion from the comparative assessment.
In regards to taphonomy, density mediated attrition is the influence of density on the loss
or preservation of elements (Lyman 1994, 252). The correlation between density and survival
occurs because elements with a relatively lower density are likely to be more porous and have a
greater surface area acted upon by destructive environmental factors (Lyman 1994, 239).

15
The small intrinsic differences between two similar taxa may not affect their relative
survival significantly, but the disparity between unique taxa may. Dissimilarity of human and
animal bone morphology at micro and macroscopic levels causes the bone to react in dissimilar
ways to extrinsic taphonomic factors. This means that even under identical circumstances,
human and animal remains could fragment in dissimilar ways. The implications these
differences have for the recording of commingled remains is a clear need for personalized
zonation method systems for humans, animals, and even separate animal taxa. Humans are a
morphologically distinct group from common archaeological fauna so a personalized zonation
method may prevent bias.
3.2. Extrinsic differences
A number of extrinsic factors work on remains and influence fragmentation. Human
culture is the extrinsic factor with the greatest amount of influence due to the dissimilar ways
human beings utilize or dispose of faunal and human remains. In most cultures, humans are
buried near to or immediately following death while animals are butchered and consumed
(Waldron 1987, 56). In addition to consumption and burial practices of human culture
influencing the fragmentation of remains is a number of environmental factors. Environmental
factors effecting fragmentation in human and animal bone include, weathering, climactic events,
animal scavenging, insect activity, and soil quality.
3.2.1. Environment and Fauna
The degree to which bones degrade and fracture after disposal is subject to
environmental conditions. Experimental taphonomy has shown that skeletal material in a
freezing climate will degrade more slowly over time than material in a hot, dry climate (Karr and
Outram 2012, 555). An experimental test on faunal material showed that bones in a hot
environment required more effort to fracture and resulted in a higher degree of small scale
fragmentation than those in cold conditions (Karr and Outram 2012, 556). The data showed
that fracture patterns are influenced by the environment, this dissimilarity may be compounded
in a test which uses dissimilar taxa. Differential fracture properties in dissimilar environments
may explain why different cultures utilize bone in particular ways. As an extrinsic taphonomic
factor, dissimilar environments play a role in constructing dissimilar patterns of fragmentation.

16
If disposed of on the surface or exposed due to environmental factors, skeletal remains
have potential for a wide dispersal pattern. Elements can be displaced from their original
context by fluvial action, animal activity, weather events and more resulting in attrition of an
assemblage (Lyman 1994). The degree to with the bones can become displaced in a burial
context is related to the position of the remains in the deposit and the available space
(Roksandic 2002, 103). Levels of soil acidity, the quality of drainage, and the type of soil matrix
will have an influence on the preservation of buried remains (Henderson 1987, 46). The
chemical makeup of the soil is the extrinsic factor with the greatest influence on the degradation
of undisturbed bone (Garland and Janaway 1989, 26).
The sooner a bone is buried the better its preservation will be, and the less likely that the
remains may be removed from their original position (Lyman 1994, 405; Roksandic 2002, 112).
After just a few years of exposure to the elements a bone can begin to flake apart and splinter.
Weathering acts on the exterior of a bone and degrades the integrity of its surface, this may
make the bone more susceptible to fragmentation (Lyman 1994). This can result in dissimilar
breakage trends between remains disposed of on the surface and those which are intentionally
buried.
The longer an element is exposed the greater a chance there is for a scavenger to take it
away or consume it whole. The taphonomic effects on faunal remains disposed of in an
exposed context are extensively different from those of a burial context; remains on the surface
are more susceptible to degradation than those below (Henderson 1987, 50). The rate of
fragmentation in an archaeological faunal assemblage will be distinctive of its level of use within
a community by people or scavengers.
Animal scavenging can have a great effect on the attrition of an assemblage and may
result in unique patterns of fragmentation. A study by Binford and Bertram showed the
attritional effect of fed dogs on an a faunal assemblage, with complete loss of some elements
and a correlation to the hunting seasons (1977, 78). The degree to which the dogs modified the
assemblage was influenced by human culture which dictated when and what the dogs were fed.
According to Waldron the damage to bone by scavengers has less of an effect on buried human

17
remains than it would on typical faunal material in a midden or rubbish heap (1987, 57). This is
a vital difference between the preservation of human and animal bone.
The activity of insects as an extrinsic factor is vital to the process of disarticulation of both
human and animal remains. Insects eat the connective tissue which articulates the skeleton,
the bones can become disarticulated with ease, on the surface this can result in bones being
removed from the context by animals, water, weather, and people (Lyman 1994, 144). Unlike
surface middens, the soil surrounding a burial protects and holds the remains in place, this
prohibits disarticulation from insect activity alone.
3.2.2. Consumption Practices
The direct human impact on faunal bones in an archaeological context includes
everything from which species are utilized to how the remains are exploited and disposed of.
Effects of consumption include dismemberment for transportation and dispersal, butchery,
cooking, burning, and comminution to extract marrow. Cooking can alter the surface of bone
and damage structural integrity through the application of heat (Buikstra and Ubelaker 1994,
97). Human culture also influences the dissimilar use of taxa, in one study the bones of larger
fauna were found to be more frequently broken by humans for marrow extraction than those of
smaller taxa (Binford and Bertram 1977, 109).
In this way faunal assemblages may be fragmented in dissimilar ways in differing
cultures because the types of bones utilized may vary between and within taxa. Reitz and Wing
separate faunal assemblages by type, these include kill sites, residential or urban deposits, and
intentional burial (2008, 120). These distinctions are made because human behavior influences
how deposits are formed and fragmented.
Observation of fracture types within an assemblage can be used to determine whether
the elements are fragmented due to normal taphonomic factors, or purposely broken open for
consumption of the marrow. Taphonomic study shows that the ways in which intentional bone
fracture and normal sediment pressure fragmentation occur, result in dissimilar trends of
fragmentation within an assemblage (Villa 1991, 27). Intentional bone breakage at or near the
time of death result in ‘greenstick’ fractures; normal taphonomic processes result in ‘dry stick’
types of fragmentation (Buikstra and Ubelaker 1994, 119; Villa 1991, 28). Analysis of fracture

18
patterns in human assemblages is used to identify diagnostic criteria for determining how long
bones tend to fracture as a result of bone marrow extraction. The measures of analysis include:
fracture angle, fracture outline, shaft circumference, and breadth/length ratios (Villa 1991, 45).
This study illustrates that the dissimilar processes human and animal remains experience at the
hands of human culture, results in dissimilar trends of fragmentation.
A comparison of butchery and trauma marks on human and faunal remains from Velim
Skalka, showed very dissimilar patterns. Cattle bones demonstrated a much higher percentage
of butchery/trauma evidence than humans, this will influence the way in which remains are
fractured (Outram et all. 2005, 1705). The analysis of fracture patterns between human and
cattle bones also revealed dissimilar patterns. The presence of ‘helical’ fractures consistent
with consumption practices was very high in cattle bones of high marrow content, but rare in
human bones (Outram et all 2005, 1706). The rare evidence of trauma in the human material
was found to be of a dissimilar type, primarily ‘butterfly’ fractures, which tends to occur when
force is applied to fleshed bone (Outram et all. 2005, 1706). This is strong evidence for the
effect of human culture on dissimilar trends of fragmentation between humans and animals.
The process of butchery and marrow extraction is destructive so it will result in lower
rates of survival and breakage once disposed of. An animal that dies of natural causes, without
being butchered, will experience dissimilar rates of attrition and fragmentation from one which
was killed for human consumption. Preservation depends on how soon the remains are
covered or buried. Human remains generally do not undergo the stresses of dismemberment,
cooking, or comminution and thus experience significantly dissimilar processes from their faunal
counterparts.
3.2.3. Burial and Disposal Practices
How remains are disposed of is dictated by human culture, bones may be discarded in
an exposed midden, buried, or even burned. Faunal bones may be purposefully fed to dogs or
left to be scavenged, animal scavenging and gnawing of bone is a process not usually observed
to affect human remains. The cultural use of middens exposes faunal remains to the range of
environmental factors which act upon the bones. Human cultural tradition commonly dictates
that human remains should be interned underground this will result if very dissimilar rates of

19
preservation between human and faunal assemblages. The sooner a bone is buried the better
it will be preserved and in a cemetery context, individuals are buried before skeletonization
occurs.
Unique burial types, from simple shrouds to resilient crypts, represent another variable
which separates the processes acting on human remains from those acting upon animals
(Henderson 1987, 52; Garland and Janaway, 1989, 18). The cultural act of burial protects
human bone from the most damaging taphonomic effects so that human assemblages form and
fragment in dissimilar ways from faunal. Human activity following burial can still cause
disarticulation and fracture of remains. The most common ways in which human remains can
become commingling occur through human interference after skeletonization.
In antiquity, cemetery remains were rarely left alone for long after burial as people
routinely reused the space for additional burials. This created archaeological features known as
grave cuts. Grave cuts occur when a later burial is dug and cuts into existing graves,
sometimes this results in the disarticulation of earlier burials (Waldron 1994, 13). The long term
use of a cemetery which results in a dense concentration of burials in the same location will be
more likely to exhibit evidence of commingling. The digging of new graves can ‘churn up’
disarticulated elements from previous burials. These elements become commingled in the
cemetery soil and are occasionally exposed on the surface, which may result if some evidence
of weathering.
Human activities which may interfere with the preservation of buried remains can be in
the form of agricultural and architectural projects which disrupt the earth. These can have a
destructive effect on an assemblage and may enable disarticulation and fragmentation of bone
within the cemetery soil or backfill. Secondary burial or relocation of skeletal remains is also
common, and may result in a loss of smaller elements during collection from the primary burial
site. The reasons for removal and ways in which skeletal material may be redeposited to an
ossuary are varied around the world.
This review of taphonomy has discredited the assumption that human and animal
remains fragment in similar ways. Not only do human and animal remains fragment in
dissimilar ways, assemblages form through very different processes, usually dictated by human

20
culture. Intrinsic differences between human and animal remains and the extrinsic factors
acting upon them result in dissimilar trends of fragmentation. No single variable determines
bone preservation alone, rather element survival is influenced by a complex combination of
internal and external factors (Henderson 1987, 42).
Human action greatly affects the postmortem treatment of human and animal remains,
resulting in dissimilar assemblage formations (Ubelaker 1997, 80), and ultimately how bones
tend to fragment. Unique animal species, let alone humans, do not all react to extrinsic
taphonomic factors in similar ways, which is reflected in fragmentation and element
survivorship. This has huge implications for the zonation method because the dissimilarity
between human and animal remains means they require specialized recording processes.
4. Developing the SDZ Method
The quality of information gained depends on the quality of the material studied and in order to make strong interpretations a strong body of theory is necessary (Binford and Bertram 1977, 77; Waldron 1987, 55). General opinion is that commingled remains are of little scientific value, but with the application of well critiqued and well suited methodologies, this can be debunked. Increasing the study of commingled remains begins with the development of recording methods that accurately represent the sample. This chapter examines the observations and data used to inform and justify the development of the SDZ method. 4.1. Personal Observations of Cranial Fragmentation
A visual investigation of the types of fragmentation common in human cranial elements,
both isolated and articulated, is performed to facilitate the identification of features which have a
high likelihood of survival. Trends of fragmentation in the human crania are observed to make
logical sense; they begin with the relatively delicate facial features and proceed in a typical
manner to the robust, relatively dense features. Fragile and thin elements, such as the nasals,
vomer, sphenoid, and the squama of the temporal are rarely observed in the reference material;
when they are present they are fragmented so as to be incomplete. The squama of the
temporal is thin and tends to fragment easily; however, the remainder of the temporal bone is
robust and can withstand greater force. The features of the skull that are observed to remain
intact most frequently are the most robust and are identified to be relatively more dense than
the rest of the skull. This includes portions of the frontal, occipital and temporal.
Following the fragmentation of the most delicate portions of the skull are elements which
can be observed in isolation but fragment indiscriminately. This is particular of the parietal

21
bones, which are compact and hard, but tend to fragment in no discernible pattern. Identifying
trends of fragmentation for the parietal bones is compounded by their lack of relatively dense
morphological features; no single part of the parietal is more likely to fragment than any other.
This is also observed among the squamal portions of the frontal and occipital bones.
How the temporal bone tends to fragment was already mentioned, but to reiterate; the
temporal is made up of both very dense and very fragile features. The squama of the temporal
is likely to fragment early on with relatively little force; this leaves the bulky thick petrous
pyramid, mastoid process, and auditory canal. The petrous pyramid is most frequently
observed to be complete in the reference material; this is likely related to both its relative
density and endocranial position. The petrous portion and mastoid process are observed to be
among the most dense features of the skull. The petrous pyramid in particular is frequently
isolated but intact and identifiable within the reference collection.
The frontal bone is made up of compact flat bone which has superiorly and irregularly
shaped features of greater relative density and size anteriorly. The frontal eminence is similar
in nature to the parietal and tends to fragment first when the frontal is isolated; this is followed
by the area of the frontozygomatic suture and the supraorbital ridges. The last feature of the
frontal bone which tends to remain intact is the interorbital area, containing the glabella point,
any vestige of the metopic suture, and the posterior frontal crest.
The occipital bone is also observed to fragment in a predictable manner; usually from the
exterior portions in towards the center. The relatively thinner portions nearest to the sutures,
particularly the occipital squama, tend to fragment in separate pieces from their separate
quadrants. The features of the transverse sulci, occipital sulcus, and occipital crest are
relatively more dense; but occasionally break off with fragments of the squama. The inferior
projecting basal portion tends to fracture from the bulk of the occipital, before fragmenting into
smaller features. Occasionally the basilar part is observed complete, but in isolation from the
rest of the occipital. The central features of the occipital, the internal and external
protuberances, and cruciform eminence, are the most frequently observed to remain complete,
along with the the closest portions of the surrounding features. The occipital protuberance is
observed to be one of the most robust and relatively dense area of the skull.

22
Observations of how the human mandible tends to fragment identifies the most durable
feature as the mental eminence. This relatively dense feature is rarely observed to be
fragmented in all of the reference material examined. The superior, posteriorly projecting
features of the mandible seem to fragment the most often; specifically the condyles and
coronoid processes. Following this, the mandible tends to fragment posteriorly to anteriorly;
corresponding with the relative density of the bone.
4.2. Element Survivorship.
A study from Waldron on relative bone survival assessed element frequencies within an
assemblage of human remains from a RomanoBritish site and calculated the percentages
present for each when compared to the estimated MNI (1987, 58). At the site 112 graves were
excavated, 88 of which contained adult individuals, the adult MNI of 88 is used to calculate the
percentage of expected material for individual element types (Waldron 1987, 57). Waldron
showed that elements like the petrous portion, mastoid processes, and mandibular body,
survived and were present in very high amounts. In fact, the petrous portion of the temporal
had a relatively high percentage of elements present with 67% present for the left side and
60.2% on the right, this was surpassed only by the left sciatic notch portion of the pelvis
(Waldron 1987, 5860). The relative survival of cranial elements within Waldron’s sample is
depicted in table 1.1. The results show that the bones of the skull do not survive in equal
amounts. The elements that have a high survivorship are relatively more dense than the rest of
the crania, this seems to correlate with the personal observations of relative density within the
reference material.
Table 1.1 Skull and Mandible Element Frequencies Skeletal elements Side Number
Recovered % of Expected
Skull
Frontal Left Right
36 38
40.9 43.2
Parietal Left Right
29 29
33 33
Occipital Left Right
35 38
39.8 43.2

23
Occipital Condyle Left Right
41 43
46.6 48.9
Petrous Temporal Left Right
59 53
67 60.2
Mastoid Left Right
52 50
59.1 56.8
Zygoma Left Right
42 41
47.7 46.6
Maxilla Left Right
46 50
52.3 56.8
Whole Skull 15 17
Mandible
Head Left Right
46 44
52.3 50
Ramus Left Right
53 51
60.2 58
Body Left Right
57 56
64.8 63.6
Table 1.1. (Waldron, 1987, 5860). Table has been condensed and only presents data from the Skull and Mandible. “Proportion of bones recovered from 88 adults at a RomanoBritish site in London.” Element frequencies and the percentage of expected frequency.
Table 1.1 also illustrates that the way in which bones are recorded can affect the results.
The bones in Waldron’s study were recorded by side, since not all bones tend to fragment into
left and right halves, this may not be a realistic representation. Since observation shows that
the occipital does not tend to fragment into halves but rather around the ‘center’, these results
may be biased. If the portions of the occipital used to record presence are more in line with how
it tends to fragment, the resulting ‘% of expected’ may be significantly changed.
4.2.1. Ossuary examples
Two associated ossuaries dating to the Late Woodland period in North America provide
element frequencies for adult and subadult commingled remains. From the crania only the
temporal, maxillary, and mandibular bone frequencies are provided; for unknown reasons the
other bones of the crania are not included in the assessment (Ubelaker 1974). Ossuary I shows
an adult MNI of 69 based on the right femur, and 62 subadults, estimates from the left temporal
24
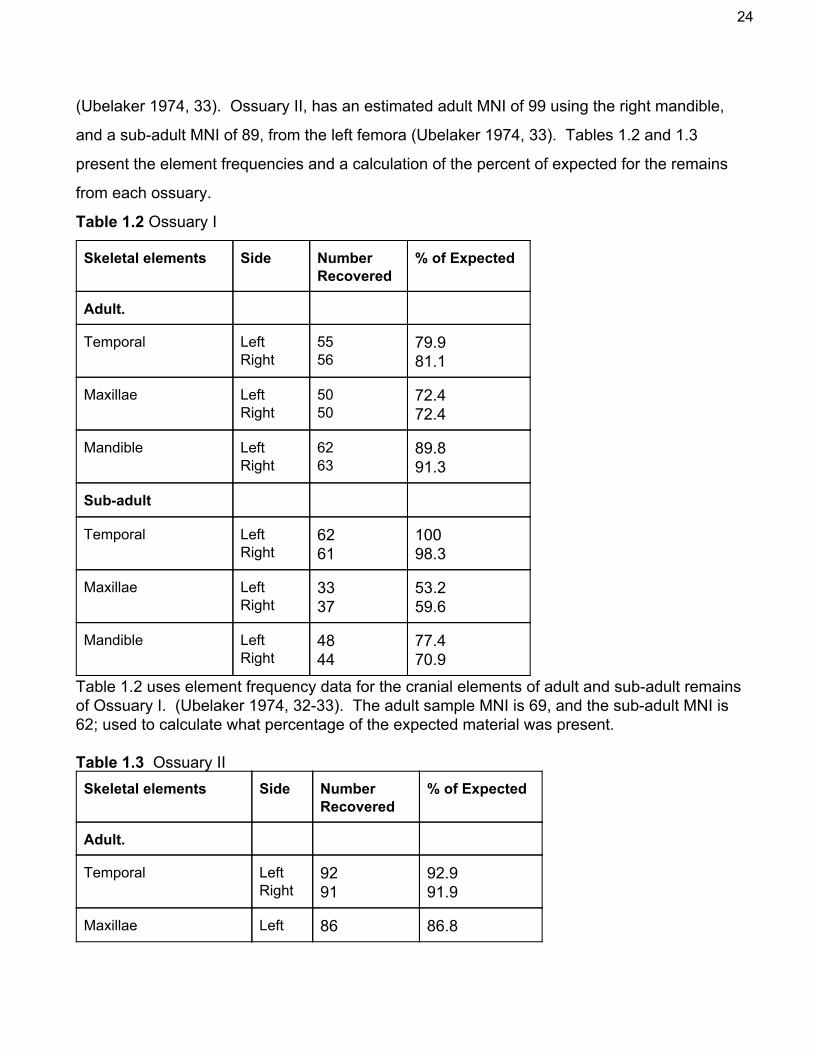
(Ubelaker 1974, 33). Ossuary II, has an estimated adult MNI of 99 using the right mandible,
and a subadult MNI of 89, from the left femora (Ubelaker 1974, 33). Tables 1.2 and 1.3
present the element frequencies and a calculation of the percent of expected for the remains
from each ossuary.
Table 1.2 Ossuary I
Skeletal elements Side Number Recovered
% of Expected
Adult.
Temporal Left Right
55 56
79.9 81.1
Maxillae Left Right
50 50
72.4 72.4
Mandible Left Right
62 63
89.8 91.3
Subadult
Temporal Left Right
62 61
100 98.3
Maxillae Left Right
33 37
53.2 59.6
Mandible Left Right
48 44
77.4 70.9
Table 1.2 uses element frequency data for the cranial elements of adult and subadult remains of Ossuary I. (Ubelaker 1974, 3233). The adult sample MNI is 69, and the subadult MNI is 62; used to calculate what percentage of the expected material was present. Table 1.3 Ossuary II Skeletal elements Side Number
Recovered % of Expected
Adult.
Temporal Left Right
92 91
92.9 91.9
Maxillae Left 86 86.8

25
Right 81 81.8
Mandible Left Right
98 99
98.9 100
Subadult
Temporal Left Right
81 84
91 94.3
Maxillae Left Right
49 49
55 55
Mandible Left Right
52 49
58.4 55
Table 1.3, uses element frequency data for the adult and subadult crania of Ossuary II (Ubelaker 1974, 3334). Frequency data is used to calculate the percentage of expected, with the adult MNI of 99 and the subadult MNI of 89.
The adult material in both ossuaries is recovered in relatively high percentages; more so
than the corresponding elements in table 1.1. The ossuary data compares adult and subadult
individual rates of expected and recovered material; highlighting a difference between the
element survivorship of the two groups. The maxillary bone ‘percentage of expected’ was
significantly decreased from the adult to the subadult samples. This is evidence of dissimilar
preservation and fragmentation between age categories. Element survivorship correlates to
element density; which in turn, is variable with age. The mandible bone is recorded in different
ways in the Waldron and Ubelaker element frequency data; the different recording styles may
relate to the differences in quantification. Waldron records the mandible in six parts, Ubelaker
only records it in two.
The assemblages in this section come from dissimilar contexts and are of dissimilar
ages, but both provide evidence that elements of the crania have relatively high likelihoods of
survivorship. Relative element survivorship for the crania is taken into account during the
development of the SDZ method.
4.3. Correlation between Density and Survivorship
When discussing the relationship between density and likelihood of element survival,
there is no definite answer to which bones will actually survive in higher percentages in any

26
given sample. The presence of a correlation does not denote causality, because a number of
additional factors also influence survivorship (Lyman 1994, 249). Some elements can be
identified as more likely to survive than others based on their density, but the complexity of
interwoven variables makes exact predictions impossible.
A study demonstrated a correlation between contemporary bone density values and
element survival in an archaeological assemblage (Willey et all. 1997, 514). This supports the
supposition that density is the most vital intrinsic factor in potential element survival, since more
dense elements are more likely to withstand the extrinsic taphonomic processes.
The Crow Creek sample, containing victims of human violence, was exposed to a
number of extrinsic taphonomic factors. The sample is unusual because, instead of immediate
burial, the remains were exposed on the surface for an unknown period of time before
collection and burial. The sample underwent a series of attritional processes between death
and discovery similar to those of faunal assemblages which are disposed of and eventually
buried. Despite this, Crow Creek shows a correlation between element density and survival
indicating that, all other things being equal, bone density is a good indicator for potential survival
(Willey et all, 1997, 517). The analysis is focused on postcranial skeleton, without mention of
the effects on cranial elements (Willey et all. 1997, 518&520).
When element survivorship contrasts with the expectations further investigation is
needed. Unexpected results may indicate the presence of an unusual variable skewing element
survival within the assemblage or an unusual process which affected the rates of attrition to be
uncovered by taphonomic study.
The density of bone and specific features is difficult to measure with precision, it is more
accurate to say that a feature is relatively more or less dense than another. The factors
influencing bone density vary between individuals and element s which are more or less dense
may not be constant. Having said that, the correlation between element survivorship and
relative density has been identified in both osteological and zooarchaeological studies. Due to
this, the relative density of cranial features was taken into account in the development of the
SDZ method.

27
5. Materials and Method
5.1. Materials
Reference material used in this assessment includes the skeletal reference collection at
the University of Sheffield, which was observed to record the way in which human skulls ‘tend’
to fragment. The collection contains a large number of isolated cranial elements in various
stages of fragmentation, so identifying trends of fragmentation was simplified. There is a
possibility of inclusion bias in the reference collection, potentially from a preference towards
well preserved and complete fragments.
5.1.1. Black Gate Cemetery
The skeletal material used for the comparative assessment is a sample of commingled
remains from the Black Gate cemetery collection. Dated to the early medieval period and
located in Newcastle upon Tyne, the cemetery was in use for an extended period of time. The
earliest cemetery burials dated to the 8th century C.E. and the latest to the 12th century, as
many as 11 ‘burial generations’ were identified stratigraphically (Nolan 2010, 159). The site
suffered several disruptions from construction activity, which may have contributed to
disarticulation, before being archaeologically excavated between 1973 and 1992.
The cultural context of Black Gate and its founding as an AngloSaxon cemetery
happened at about the same time as the transition towards churchyard graves instead of field
burial (Cherryson 2010, 54). During the cemetery’s early period of use, burial in consecrated
ground was becoming culturally important which accounts for the perpetual reuse of the same
space (Cherryson 2010, 54). These burials are associated with and appear to be oriented with
a small building which may be a chapel. This supporting the churchyard burial hypothesis
(Nolan 2010, 159). The most dense areas of the cemetery, where the sample comes from, may
have been considered a high status area, which would account for the grave cuts of later burials
and the large amount of disarticulated remains.
The skeletal results of the excavation were 660 numbered individuals with a total
estimate of 679 individuals (Nolan 2010, 148). Multiple cross cut burials were identified along
with an abundance of disarticulated remains found in the ‘cemetery soil’ backfill which indicates
long term useage of the same space (Nolan 2010, 159). A cemetery report, from 1993,

28
indicates that even though the scientific value of disarticulated remains is questionable, they
can be used to give Osteologists an idea of the missing or ‘vanished’ cemetery dwellers (Nolan
2010, 151). The disarticulated remains found in cemetery backfill represent the earlier
generations of cemetery residents, destroyed and dispersed by the digging of later graves.
Three main types of burial identified at Black Gate may have had differing effects on
element survivorship and recovery. The majority of the Black Gate burials were ‘plain graves’
without any associated coffenry or stonework, but there is evidence that many individuals were
shrouded (Nolan 2010, 204). ‘Earmuffs’ and ‘HeadBoxes’ were grave types meant to support
and protect the head using stones or wood (Nolan 2010, 204205). The final burial type found
at Black Gate is enclosed and includes; rubble lined graves, coffins, and chests (Nolan 2010,
205206). The enclosed burial type was the most unusual and may have protected skeletons
from disarticulation to some extent, but there is no way of knowing if the commingled remains
were not originally buried in this manner before disturbance.
The 19901992 excavated material, from which this sample comes, was examined in its
entirety by Boulter and Rega, they assessed both individual burials and the disarticulated
material (1993, 6674). The authors choose to attempt assessments of MNI, age, sex, and
pathology. At the time of assessment there was no standard method for recording commingled
remains so Boulter and Rega modified a methodology from Zooarchaeology. Their method
used codes to identify and record elements and fragments, as distal and proximal portions,
assessing the material in the same manner they would have a faunal assemblage. To avoid
over recording the presence of the parietal, only fragments with the anterior meningeal groove
present were recorded (Boulter and Rega 1993, 67). This use of a diagnostic feature to prevent
overestimation is similar to the modifications suggested in the later half of this paper and in
fact, sets a precedent. In the 1993 Black Gate skeletal assessment the modified method was
used to record every fragment, but since the ‘codes’ used were nonspecific, less detail and
accuracy can be expected.
5.1.2. Commingled Sample
The sample used for this project was excavated between 1990 and 1992 from the
densest burial areas of the cemetery; RA29, RA28, and Area C, and consists of a large sample

29
of the disarticulated finds. The sample is stored in nine large plastic crates, with 30 identified
contexts separated into bags. Most of the bones are labeled with context or are in labeled
bags, some unlabeled material in an unlabeled bag was recorded as a single context: ‘BG?’.
The contexts range in size from a single bone to over 100. The sample is for the most part well
preserved, with an average score of 12 on the McKinley scale (2004). Despite good
preservation the sample is badly fragmented, which made it an ideal candidate for a test of the
difference between the KO and SDZ methodologies. Some remains show signs of advanced
weathering, possibly caused by a period of time exposed to the elements after becoming
disarticulated and churned up in the cemetery soil. The majority of the remains are mature, less
than 25% are juvenile, and less than 2% are fetal or infant. A few elements within the sample
have evidence of postmortem damage, from what appears to be contemporary tools, and fewer
still have evidence of rodent gnawing. A small range of pathological traits are present as well
as some evidence of healed trauma. This material resides at the University of Sheffield and
was selected as a representation of the average commingled, open, human assemblage.
5.2. Methodology
Having established that human and animal remains fragment in dissimilar ways, the KO
method weaknesses regarding the crania can be addressed. This section presents a series of
modifications to the zonation of the frontal, occipital, temporal and mandible bones, which make
up the SDZ method. The location and form of the new zones are justified using observed
trends of fragmentation, relative density, and element survivorship data. Each element included
in the SDZ method contains one or more relatively dense features with a high likelihood of
survival in human assemblages. The zones are recorded with the hypothesis that they would
provide a greater degree of accuracy in MNI estimations. To identify new zones
morphologically and place emphasis on particular features standard anatomical names and
terminology are used (White and Folkens 2005).
In order to assess the difference between the KO method and the SDZ method the
Black Gate sample was recorded twice. First with the KO method and second with the SDZ
method. Recording was performed by a single observer, and the comparative methods were
applied at separate times. When applying the KO method refitting was performed, within

30
contexts, where an obvious, postmortem break had occurred. When recording the SDZ
method refitting was not used. In the SDZ method when the cranial vault was intact and one or
more of the elements being recorded were fused they were recorded separately to ensure
consistency, but a note to record the event was made in the data. During recording of the SDZ
method, when multiple elements were articulated in the same specimen, they were recorded
independently to retain consistency. Each fragment was recorded as present, incomplete, or
absent in the required zones. The presence of isolated teeth was noted once, when first
recording the sample.
Adult and juvenile material was separated into broad categories during the recording
process using size, morphology, suture closure, and epiphyseal fusion (Scheuer and Black
2000). Distinguishing between immature and adult material is often the limit of specificity when
estimating age in a commingled assemblage because of the lack of reliability when working with
isolated elements (Buikstra and Ubelaker 1994, 9). Estimation of sex was not attempted during
the recording process due to the fragmented nature of the remains. The use of isolated
fragments is not considered reliable in estimations of sex (Buikstra and Ubelaker 1994, 20).
The presence of pathology was recorded when observed.
During the recording process diagrams for the zonation of each method were carefully
referenced to speed familiarity with the zones and reduce observer error. A number of
fragments in the Black Gate sample were so badly fragmented that they were unidentifiable.
These fragments were simply recorded by size and type. Care was taken during both recording
sessions to accurately record the presence, absence or incompletion of each required zone.
The following sections describe the SDZ method by element and zone.
5.2.1. Frontal
In the KO method the frontal bone consisted of two zones out of 15 within the skull. The
frontal zones divided the bone into left and right sides. This could be a useful way of recording
immature material before the metopic suture has fused, but mature frontal bones rarely
fragment in that way. The SDZ method divides the frontal into five zones which more accurately

31
represent how the bone tends to fragment. Figures 1. and 2. depict the frontal zonation of the
KO method in its original form and the modified SDZ method.
Figure 1. KO zonation, view of frontal. Unmodified diagram of the KO method zonation. Anterior view or the crania. (Knüsel and Outram 2004, 93). Figure 2. SDZ frontal zonation. Image modified from fig 1. zones altered to depict the SDZ method and image cropped.
Zone 1 consists of the dense interorbital area, this zone includes the frontal crest on the
posterior surface. Zone 1 is easy to identify, dense, and a nonrepeating feature. Zones 2 and
3 of the frontal are the supraorbital margins and superciliary arches on the left and right sides of
zone 1. The brow ridges are also relatively dense features. Zones 4 and 5 encompass the
whole of the frontal squama, and represent the left and right sides respectively. Accurate
identification of zones can be difficult for small fragments of the frontal squama. If a portion of
parietal bone is articulated, the orientation of the meningeal grooves can aid in the
determination of side.
5.2.2. Temporal
In the KO method the left and right temporal bones were each assigned a single zone,
zones 6 and 7 out of 15 in the skull. In the SDZ method each temporal bone consists of two
zones, the bones are identified by side before recording the presence or absence of zones.

32
Figure 3 depicts the diminished place of the right temporal in the KO method, as a part of a
whole. Figures 4 and 5 illustrate the temporal bone zonation for the SDZ method. In the SDZ
method, zone 1 of the temporal bone is the dense petrous pyramid, this feature was frequently
observed in isolation in the reference material. The remainder of the temporal is assigned to
zone 2 and specifically includes the fragile temporal squama, external acoustic meatus,
zygomatic process, and mastoid process.
Figure 3. KO zonation, view of temporal. A view of the temporal bone from the right side as it appears in the KO method, unmodified image (Knüsel and Outram 2004, 94). Figure 4.1 SDZ method, temporal. A view of the left temporal bone, sectioned, depicts the SDZ method zonation, image modified with the addition of zones. Figure 4.2. Inferior view of the left temporal, depicts SDZ zonation, image modified with the addition of zones (Gray’s Anatomy, 1980, 327329). 5.2.3. Occipital
Modifications transform the occipital from a single bone out of 15 in the KO method to an
individual element with three zones in the SDZ method. Figures 6 and 7 show the occipital as it
is in the KO method and figure 8 illustrates the zones applied in the SDZ method.

33
Figure 5.1. KO inferior view of the occipital. Unmodified image. Figure 5.2. KO posterior view of the occipital. Posterior view of the occipital. Unmodified image (Knüsel and Outram 2004, 95). Figure 6. SDZ, endocranial view of the occipital. Image modified by addition of zones (Gray’s Anatomy 1980, 321).
Zone 1 is the densest area of the occipital, it is made up of the central part of the
cruciform eminence, the internal occipital protuberance and the center most portions of the
transverse sulci, occipital sulcus, and internal occipital crest. The occipital squama and portions
of the transverse sulci, occipital sulcus, and internal occipital crest most distal from the internal
occipital protuberance make up zone 2, along with the superior portion of the foramen magnum.

34
Zone 3 consists of the basilar portion of the occipital, including the inferior portion of the
foramen magnum, occipital condyles and inferior most features.
5.2.4. Mandible
In the KO method the mandible is represented by six zones, each with a left and right
portion. The SDZ method modifies this and assigns individual zone numbers to each side, with
a total of nine zones. Figure 9, depicts the KO mandible zonation of the left side, the zones are
mirrored on the right half. Figure 10 shows the nine zones of the SDZ method in a medial view
of each side.
Figure 7.1. KO method, mandible. KO method zonation for the left half of the mandible, top: medial view, bottom: lateral view. Unmodified image (Knüsel and Outram 2004, 88). Figure 7.2. SDZ method, mandible. the SDZ method zonation for the mandible, top: right half, medial view, bottom: left half, medial view. Original image.
Zone 1 of the mandible is the mental eminence portion including the incisors and
canines. The portions of the mandibular body including the premolars and molars are zone 2
on the right and zone 3 on the left. On the right side of the mandible zone 4 is the gonial angle
and the space from the oblique line to the ramus. Zone 5 is the right coronoid process and

35
zone 6 is the right mandibular condyle. In the name of consistency the right side’s zone division
is mirrored on the left side of the mandible in zones 79. On the left zone 7 is the gonial angle
and all the space between the oblique line and the ascending ramus. Zone 8 is the left
mandibular condyle, and zone 9 is the left coronoid process. When recording the mandible, the
presence of individual teeth should be noted.
6. Results and Discussion
6.1 Raw Data
The KO method is used to recorded the assemblage in its entirety, including 29 contexts,
with over 1500 identified bone entries. Less than 300 unidentifiable fragments are recorded,
about 50% of which are flat skull bone fragments of less than 40mm in length. Based on the
data from zones recorded as present in the KO method, the MNI of the adult material is
estimated to be 25. The MNI is established with zone 6 of the left femur, which is the most
frequent, nonrepeating feature. Assessment of the most frequently recorded element zones in
the KO method are made, and include the femur, tibia, and humerus. In order to compare the
KO method to the SDZ results MNE is calculated for the cranial zones corresponding to the
frontal, occipital, temporal and mandible bones. See table 2.1 for the MNE’s as they are
represented by the KO method data. There is a disparity between the element frequencies of
the cranial and postcranial skeleton.
Table 2.1 KO Method Element Frequencies Element Most Frequent
Zone # Recorded as Present
# Recorded as Incomplete
MNE
Femur (Left) 6 25 19 25
Tibia (Right) 8 21 6 21
Humerus (Left) 5 14 5 14
Frontal 1 9 40 9
Temporal (Left) 6 11 11 11
Temporal (Right) 7 9 24 11

36
Occipital 5 2 61 2
Mandible 7 14 2 14
Table 2.1. Presents the raw quantitative data for elements of interest as recorded with the KO method. Identifies the number of instances of ‘present’ and ‘incomplete’ zones. MNE is estimated from the zone with the highest number of specimens recorded as present Table 2.2 SDZ Method Element Frequencies Element Most frequent
zone # Recorded as Present
# Recorded as Incomplete
MNE
Frontal 1 17 12 17
Temporal (Left)
1 20 3 20
Temporal (Right)
1 22 5 22
Occipital 1 30 6 30
Mandible 3 14 5 14
Table 2.2. Raw data quantifying the number of present and incomplete zones as recorded by the SDZ method. MNE is estimated from the zone with the highest number of specimens recorded as present.
Data recorded for the SDZ method includes zone frequencies for the frontal, temporal,
occipital and mandible bones of mature individuals. The data comes from 20 contexts, with a
mere 197 entries. No data is recorded for unidentifiable fragments. The MNI of the
commingled adult cranial material, as recorded by the SDZ method, comes to 30 and is based
on zone 1 of the occipital. The MNE is calculated for each of the four element types included in
analysis and is compared to the corresponding data from the KO method. Table 2.2 depicts the
number of fragments recorded as present and incomplete for the most frequent zone of each
element in the SDZ method and the MNe as calculated using the most frequently present
zones. This is fed into table 2.3 which shows the percentage of expected for each element
MNE.

37
Table 2.3. Percent of expected.
KO Method SDZ Method
Skeletal Elements
Number Recovered
% of Expected
Number Recovered
% of Expected
Frontal 9 30 17 56.6
Left Temporal 11 36.6 20 66.6
Right Temporal 11 36.6 22 73.3
Occipital 2 6.6 30 100
Mandible 14 46.6 14 46.6 Table 2.3. Demonstrates what percent of the expected material is present according to the MNE estimations from both the SDZ and KO method. Percentages of expected are calculated from the SDZ method MNI of 30.
Table 2.3 is interesting because it represents a clear improvement in the accuracy of
recording through the SDZ method. The percentages of expected are much higher in the SDZ
data than the KO data. While they do not correlate exactly to the element survivorship data of
the RomanoBritish or Ossuary sites mentioned in chapter 4.2 they do demonstrate differential
rates of element survivorship among the Black Gate sample of commingled remains
6.2 Assessment of Results
The cranial zonation is a distinct weakness within the KO method. Results of the
comparative method illustrate that the potential of cranial elements for quantitative methods is
neglected. The analysis of postcranial elements in this discussion is minimal, merely discussing
the gulf between MNE calculations for limb and skull bones in the KO method. The SDZ
method applies a greater number of zones to some elements of the skull, with careful
justification for the additions.
Figure 8. is a sidebyside comparison of the reported element frequencies for the KO
and SDZ comparative methodologies. The figure illustrates how changing the way in which

38
bones are recorded can influence element frequencies within a sample. The MNE for every
bone except the mandible increases, which is indicative of the ways in which zonation between
the KO and SDZ method is altered. The mandible is the only bone where the location and
distribution of the zones are largely unchanged. During revision zone 2 and 7 are combined to
make what is now zone 1 in the SDZ method, and the number of zones in the ascending ramus
portion decreases from three to two, but these changes appear to be insignificant in estimating
the MNI of the element.
Figure 8. Comparative MNE for Cranial elements.
Fig. 8. The MNE for the bones used in the SDZ method compared to the MNE for the corresponding zones from the KO method. Only zones recorded as ‘present’, were used for the estimations shown in this figure.
The zonation of the mandible in the KO method corresponds to its use in the faunal
diagnostic zone method. The authors treat the mandible as though the two sides are separate,
when in fact the mandible is a single bone that is recovered in varying degrees of completion.

39
This makes sense when recording faunal mandibles, which are often in two halves, but does not
work as well with human material, because the mandibles fragment in dissimilar ways. The
zonation of the mandible in the SDZ method is intended for greater clarity, which it seems to
achieve. The deletion of KO mandible zone 2, which represents separate zones for the canine
teeth, results in a loss of specificity out of keeping with the rest of the SDZ method, but it does
not seem to have greatly influenced the results
The changes to the zonation of the frontal bone appear to be successful in that the MNE
of the frontal increases. The focus on the dense morphological features results in greater
specificity. Zones 4 and 5 of the frontal still result in a large amount of incomplete fragments,
but this may be unavoidable, as this portion of the frontal tends to fragment indiscriminately. An
increase in the number of zones would likely not make a significant difference as this part of the
frontal lacks highly recognisable diagnostic features.
In the KO method the temporal bones are each represented by a single zone which does
not take into account the duality of this bone. The temporal bone contains features that tend to
fragment with either, relative ease, or not at all. In the KO method data set, the temporal is
usually recorded as incomplete due to the nature of the temporal squama to fragment easily,
even though the petrous portion remains intact long after. The SDZ method separates the
temporal into sections, one which is relatively dense and one which is not. Adding a zone to
represent the petrous pyramid on the endocranial side of the bone is a successful change which
increases temporal MNE for both the left and right sides.
The modifications to the occipital bone zonation are the most successful in this
comparative assessment. The MNE for the occipital increases from two to 30 with the addition
of zones in the SDZ method. This change also increases the MNI estimate for the sample from
25 to 30. In the KO method the occipital is vastly under represented due to poor zonation. In
the SDZ method the bias appears to be corrected for. Zone 2 of the occipital is likely to be
represented as several fragments from the separate quadrants around zone 1 unless the
occipital is relatively complete. The decision not to create unique zones for each quadrant is

40
made in an attempt to not over complicate the method. Now with the additional zone
frequencies future revisions may be made.
The inclusion and exclusion of specific elements in the SDZ method is based on the
observed relative density of elements and survivorship data. The delicate maxillaryfacial bones
are not included as they tend to survive at much lower rates and are observed to fragment
indiscriminately. The parietal bones are excluded due to the lack of relatively dense diagnostic
features and random fragmentation. Without the presence of exceptionally diagnostic features
which survive and can be identified with ease, the average Osteologist could spend weeks
siding and fitting these fragments into the correct zones. Most of these fragments are ultimately
recorded as incomplete, and potentially skew MNI calculations with their inclusion or exclusion,
so they are not relied upon for accuracy. The interpretations that can be made from these types
of fragments are limited due to their low rate of survival and poor condition.
The SDZ method in isolation can not be used for inventory purposes, as it only records
the zonation of four elements. Complete inventory of remains, as applied in the KO method, is
necessary to record any pathological traits and gain a more inclusive representation of the
sample.
6.3. Quantitative Techniques
Multiple techniques for estimating the size of a population using skeletal remains exist.
Not every quantitative method is suited to every assemblage. There is a difference between
estimating how many individuals are physically represented within the sample (recovered), and
estimating how many are present in the original burial or living population. These questions
have vastly different answers due to the loss of material and data over time from death to
recovery.
Any preliminary investigation into any skeletal material, be it human or animal, modern or
ancient, should establish the MNI. Morlan uses MNI to test comparative methods because he
considers MNI estimations to be more accurate for recording ‘abundance’ than other available
techniques (1994, 797). Accurate estimation of MNI is dependant upon the precision of the
recording techniques applied. This makes MNI ideal for gauging the level of improvement to the
zonation recording method through the development of the SDZ system.

41
To evaluate the effect of the ‘zone revision’ on recording accuracy, MNI and MNE are
considered the best techniques for showing quantitative difference. MNI identifies the most
frequent, nonrepeating element or zone in a sample as representing the minimum number of
people who can be physically represented in the sample. This means that by necessity MNI
underestimates the size of the original population, as MNI does not account for attrition (Adams
and Konigsberg 2004, 139). This weakness is not detrimental to the use of MNI in this analysis
because it is used to test which recording method most accurately represents the physical
sample present.
MNE compares element frequencies in the data to assess difference. MNE is
established by counting the number of bones present, the technique can be more or less
specific as the number of variables are adjusted. In this comparison MNE is calculated for
individual elements by side and zone. MNE relates to the calculation of MNI and is useful for
calculations of element frequency within the sample. Calculating the MNI for a sample is as
simple as identifying the sided element with the highest frequency, with the use of appropriate
zones these estimates become more specific (Adams and Konigsberg 2004, 139).
A variety of quantification techniques attempt to correct for the bias which occurs from
taphonomic attrition on an assemblage, by estimating the size of the original living population.
The Lincoln Index (LI) or Most Likely Number of Individuals (MLNI), are both used to provide an
estimate of the original population size at the time of burial (Adams and Konigsberg 2004,
138149). The MLNI and LI techniques involve pairmatching and are considered
overlycomplex and time consuming for this analysis. Additionally MLNI is applied to the
postcranial elements and is thus irrelevant to the data collected by the SDZ method.
Adams and Konigsberg stress that if a sample is extremely fragmented then LI and MLNI
are prone to miscalculation and with bad fragmentation no method is reliable for estimating the
original population (2004, 150). Another technique is the Number of Identified Specimens
(NISP) which views each fragment or element as a unique specimen. NISP is not used
because in a fragmented assemblage the results may be badly skewed by the breakage of
large bones into many pieces. The different levels of fragmentation between elements cause

42
bias in NISP estimations because larger bones are able to fragment into more pieces than small
bones (Outram et all. 2010, 1702).
6.4. MNI and MNE
MNE calculations, using the cranial material recorded with the KO method, under
represent the sample and illustrate a bias. This is especially true when only the data for zones
recorded as ‘present’ are included. Using the SDZ method, the MNI of the Black Gate sample
is calculated with a higher degree of accuracy because the data is much more detailed. In
addition to applying a greater degree of detail, the SDZ method excludes some of the ‘white
noise’ which comes from having too many vague zones. The MNE estimations from the SDZ
data are in most cases significantly greater than those of their corresponding zones in the KO
data. The only instance where this is not true is in the zonation of the mandible; the MNE for
the mandible is the same in both data sets. This is likely because the zonation of the mandible
is only slightly altered.
The difference between MNI estimates in these two zonation methodologies is down to
the difference in zone type. In the KO method, the skull consists of 15 zones within the crania,
and is treated as a single bone. When recording isolated and damaged fragments this results in
the majority of material being recorded as incomplete. The MNI, of the KO data set, therefore
comes from the postcranial skeleton. The femur, which is divided into 11 zones, is recorded in
much greater detail in the KO method than most of the cranial elements. This disproportionate
level of detail is probably originally applied to the femur in zooarchaeological methods because
it is very dense and has a high likelihood of survival. This results in a recording bias in the KO
method and may be an example of a self fulfilling prophecy, eg: if the most frequent element is
expected to be the femur, then the method will reflect that expectation and result in a skewed
data set.
6.5 Incomplete Zones.
The KO method records high numbers of incomplete fragments because the zonation
used is not specific to how the bones tend to fragment. In a commingled assemblage, perfectly
complete elements are in the minority. Only two complete occipitals are recorded in the KO
method dataset, with a huge number of incompletes. In the SDZ method the number of

43
complete occipital bones is the same, but the number of fragments representing a complete
zone 1 is much higher.
In the future incomplete fragments should be recorded as more or less than 50%
complete, this would allow for more accuracy in MNI calculations. If a number of incomplete,
nonrepeating zones are recorded, with a portion recorded as greater than 50% complete then
those can be assumed to represent an individual. As it stands now the number of incomplete
zones are useful in assessing the usefulness of the zonation for different elements.
A zonation method which accurately represents how the material tends to fragment will
have a low percentage of incomplete zones. A method which is a poor representation is
expected to have large numbers of incomplete zones because the zonation applied is not suited
to trends of fragmentation within an element or elements. This is what occurs in the KO
method, a high number of incomplete cranial zones and low number of completes. The SDZ
method, which is specifically developed using fragmentation trends, has a much lower number
of incomplete zones (Appendix B). The decrease in the number of incomplete zones in the SDZ
method is related to both the smaller zone sizes and the more accurate representation of how
cranial elements will tend to fragment.
6.6. Unidentified Fragments
The number of unidentified specimens in the KO method is related to the difficulty of
accurately identifying small, nonmorphological fragments to element. For instance, over 100
flat, compact, skull fragments measuring less than 40mm at their widest part, are recorded
during the KO method. These fragments most likely belong to the parietal bones but are not
sided accurately due to their diminutive size and lack of morphological features. The use of
specific diagnostic zones in the SDZ method means that only those features are being
recorded. This greatly decreases the time needed for recording and is overall much more
efficient.
Many osteologists assert that with enough time no fragment is ever truly ‘unidentifiable’,
but the degree of fragmentation in this sample is staggering and time is limited. The analysis of
‘unidentifiable’ fragments by type eg: shaft, epiphysis, cortical, or trabecular bone, can

44
contribute to statements about the causes of fragmentation within an assemblage (Outram et
all. 2010, 1704).
6.7. Demographic Factors
The value of disarticulated remains does not only extend to taphonomic studies of
survival and fragmentation; there is also some limited potential for contribution to demography
and pathology. Although any information gleaned in either respect is purely additional data of a
vague nature, because estimating sex, or age, or diagnosing an ailment is little better than
making a guess when working with an isolated element. For this reason estimation of sex is not
included in analysis and the estimation of age is performed only on a basic level.
Adult materials are included in this assessment because the SDZ method is constructed
around the relative density of cranial features, and density is highly variable with age. Once an
individual reaches maturity, age estimation becomes increasingly difficult and even with the use
of an articulated skeleton, age can only be accurately estimated to a range of about 10 years
(Buikstra and Ubelaker 1994, 36). Since the comparative method only includes adult materials,
bones are lumped into broad categories of ‘adult’ and included or ‘juvenile’ and excluded.
The estimation of age requires a multifactorial assessment of several features which are
combined to give an ‘age range’, in a fragmented and commingled assemblage this is
impossible to achieve. Problems of age estimation are compounded in cranial samples
because methods for the crania (eg; suture closure) are considered less reliable than those of
the postcranial skeleton (Brickley and McKinley 2004, 18; Buikstra and Ubelaker 1994, 36).
There is no significant amount of pathology recorded within the Black Gate sample
outside a small variety of common ailments. This includes evidence of osteoarthritis, which is to
be expected in an adult sample, a case of rickets, and evidence of osteolytic lesions. The lack
of significant pathological traits in the sample may be due to the osteological paradox. The
osteological paradox is the concept that a disease which kills quickly may not affect the skeleton
enough to be detectable.
Pathology can be observed and noted by type, but a diagnosis cannot be made with an
isolated element. By considering how difficult a correct diagnosis is to achieve with a living
individual and an array of information, it is understandable that disarticulated remains may not

45
be diagnosed with any accuracy (Brickley and McKinley 2004, 36; Waldron 1994, 36).
Commingled remains can be very useful in increasing the rates of incidence with in a sample,
because unique pathological traits not exhibited in the articulated remains may be present
(Waldron 1994, 42).
6.8. Potential bias.
The zonation methodologies are designed to record disarticulated and fragmentary
remains, but, if the methods are biased, they do not provide reliable data. The KO method is
biased towards the postcranial skeleton, which is apparent in the comparative data. Compared
to the postcranial element frequencies, the skull bones are vastly underrepresented. This could
stem from the lack of a skull zonation precedent in the faunal diagnostic method, but can no
longer be excused. The SDZ method is intended to correct for this bias, with a detailed
recording of cranial elements expected to have a high likelihood of survival. The ability to
predict which skeletal features are more likely to survive is not the same as predicting what will
actually be recovered in a sample. Both recovery bias and recording bias must be accounted
for in studies of element survival.
An array of variables could have influenced the data and acted as a bias, skewing
results. The most likely bias within the sample is variation in the relative density of features
from age, sex, and pathology. The bias cannot be completely accounted for because the
estimation of age and sex in commingled remains is not reliable. Pathology can also be a
source of bias within the sample because pathological changes to bone may alter density and
surface area. Due to the variation in bone density caused by disease, the potential for bias
from pathology in this sample exists.
External bias reflect the processes of selection of the sample, including taphonomic
attrition and recovery bias. Selective bias towards particular elements may be present because
the sample is used as a teaching collection, meaning it may not be highly representative of a
normal assemblage. The results could be biased due to inadequate sample size or the
presence of observer bias.
The KO method does not specify whether the refitting of fragments to form a complete
element should be performed, which could result in interobserver recording biases or error. The

46
error would be caused from differing levels of interest and skill in refitting fragments between
observers. Dissimilar forms of refitting may cause inconsistency in the recording of fragmented
remains. For these reasons the SDZ method specifies that no refitting should be done, every
fragment should be recorded as it is. This aids in consistency between observers and is a more
efficient way to record assemblages. Experience shows that it is easy to get swept up in
reconstructing a skull from small fragments and in order to record skull zones as complete in the
KO method this may be useful. The detail of zones in the SDZ method however, make
identifying skull fragments as complete a simple endeavor.
Relative bone density for cranial elements is established using personal observations
and therefore a potential for observer bias exists. The trends of fragmentation are observed
using reference material from a similar context to the Black Gate cemetery assemblage. This
could cause a bias towards how elements fragment in dissimilar contexts. The analysis of
material from different environments or contexts could show that the SDZ zones are inaccurate
when removed from their context. The likelihood of survival for elements in assemblages from
unique environments would need to be assessed before applying these methods, and
adjustments made as necessary. The analysis of element survivorship attempts to negate the
possibility of observation bias in the developmental phase of the SDZ method.
Unusual results may simply reflect that a sample is biased due to an unusual
taphonomic factor and may not indicate a flawed method. However, the possibility of observer
error in estimations and recording can not be ruled out without independant testing. No method
will ever be completely free of bias, but through regular revision and comparative tests, they can
be improved.
7. Conclusions and Future Research.
In zonation style recording, the precision and value of the zones used are what matter
not how many zones exist, in essence: quality over quantity. Zones should be identifiable and
analytically useful, recording an infinite number of zones is unlikely to tell you more than
recording a smaller number of very morphologically specific zones.
7.1. Summary of results and interpretations

47
This paper addresses the lack of critique to the KO method, as applied to fragmentary
remains. The KO method is currently the foremost recording technique for use with human
assemblages of commingled remains in archaeological contexts. Evaluation of the KO method
revealed it to have uncritiqued weaknesses which affect the accuracy of quantification. In this
paper the assumption that human and animal remains fragment in similar ways, is shown to be
flawed. This is the premise of the KO method, so it cannot be considered reliable for
quantitative assessments at this time. Human and animal remains, in most contexts, fragment
and become commingled in dissimilar ways because they experience dissimilar taphonomic
processes.
The KO method does not justify how the human crania is divided into zones, so an
investigation into the survivorship of the skull is made. This investigation assesses trends of
fragmentation, relative density, and element survival data to show that a more representative
zonation of the skull is possible. A revised zonation for the crania is developed, which takes
into account trends of fragmentation, relative density, and element survival data, to record
zones of the frontal, occipital, temporal, and mandible bones. The comparative analysis shows
that the SDZ method increases the accuracy of quantitative techniques including MNI and MNE
on the frontal, temporal, and occipital bones.
The expectations of greater accuracy in recording for quantitative purposes with the SDZ
method are, for the most part, fulfilled, which suggests that the method is valid. The use of very
specific crania zones in the SDZ method results in an increase of the Black Gate sample MNI,
from 25 to 30.
In the Black Gate sample the cranial data is underrepresented by the KO zonation which
results in a negative skew to the MNE estimations. Recording bias in the KO method leads to
the underrepresentation of most elements of the skull, but is not found to significantly affect the
representation of the mandible. This disparity in detail leads to vast discrepancies in MNE
estimation between the KO and SDZ methods when analysing the data. Hopefully, in the
future, the same levels of detail and attention will be applied to all aspects of the skeleton as
more complete survivorship data is gathered.

48
The results show that the KO method has a bias in the form of preferential recording of
the postcranial skeleton. Preferential recording may have resulted in a selffulfilling prophecy
regarding the elements which are expected to be, and recorded as, the most frequent. The
expectation of a greater degree of element survival for the postcranial skeleton may stem back
to the faunal diagnostic method which does not provide a method of zonation for the skull.
7.2. Implications for Zonation Style Recording
Due to dissimilarity between human and animal taxa, recording methods should be
personalized to individual groups. Zonation specificity can be to specific taxa, or general
morphological types. By treating cranial elements as individual bones with unique diagnostic
zones, rather than just a zone within the skull, the data representing element frequency is
greatly altered.
In future applications of the KO method, the SDZ method should be applied to record
fragments of the frontal, temporal, and occipital. The recording of postcranial bones should not
be neglected and ideally should be used in conjunction with the SDZ crania zone modifications
to provide an accurate estimate of MNI within commingled assemblages. The SDZ method
would be useful in a fragmented and disarticulated, open collection or ossuary situation.
Hopefully, the simplification of the method will inspire an increase in the analysis of commingled
remains prompting further revision and critique.
A balance is needed between recording as much information as possible and using the
available time efficiently because not every situation allows for extended study. Osteologists
must gauge how to make the most of the time allotted for study and gather reliable data to make
meaningful interpretations, this is particularly important for commingled remains. In some cases
this may mean assessing a smaller amount of the material in more detail rather than assessing
all of the material in little detail.
7.3. Future Research
The SDZ modifications should be independently tested for accuracy and bias when
applied to dissimilar contexts. Element survivorship data should be acquired from contexts
similar to the assemblage being recorded and used with relative density observations to assure
accurate representation of elements within the sample. Survivorship variation in contexts

49
around the world could be taken into account when building a standardized recording method to
try and prevent recording biases. Independant testing should be performed to account for the
level of interobserver error.
Future research into recording methodologies for commingled remains should include a
statistical assessment of difference. This could be achieved using a nullhypothesis test to
identify the potential for error. Statistical significance of the difference between the KO and the
SDZ recording methodologies could be tested by taking random samples of a population,
calculating the MNE for various elements using the data from each method and inputting the
results into a statistic program. Using a paired ttest or a nonparametric test, random samples
could be selected, representing various real world scenarios to which the zonation methods
may be applied. The results would show the statistical significance of any difference in the
quantitative data between the KO and SDZ methodologies.
Use of the SDZ method could be adopted into the KO method by substituting the
modified zonation for the corresponding zones in the skull zonation. This would most likely
result in more accurate representations of cranial element frequencies within assemblages.
When estimating the MNI of assemblages where cranial bones are the majority, inclusion of the
SDZ method is vital to prevent the under representation of elements. In the case of an
assemblage where cranial fragments are a minority, the SDZ modifications may be irrelevant.
Other avenues for further research in the field of commingled remains should include the
weaknesses which could not be addressed at length in this paper. This includes the
development of standard methods for the recording of juvenile material at different points of
maturity. Juvenile remains are morphologically distinct at different age stages so, this may
require the creation of zonation personalized to particular stages of development. This could be
approached through a study of juvenile development to identify which elements have a higher
likelihood of survival at different ages and contexts. The identification of juvenile zones could be
performed using similar methods to those applied in the SDZ method including relative density,
survivorship, and trends of fragmentation.
In regards to the crania excluded from the SDZ method, future research may consider
weighing fragments and developing a database with a range of potential element and crania

50
weights. Or calculating the surface area of fragments, like those from the parietal, and from that
working out a MNE which would have contributed to create the amount of material recovered.
This may address the difficulty in estimating frequency of fragmented elements, like the parietal,
which lack dense morphological features. A study could begin by taking the weights or surface
areas of complete elements within the sample to build a range of the average element weight,
to estimate how many of a particular element are represented in a fragmented set. This could
be useful for seeding the recording process of fragmented assemblages. Both of these ideas
have potential in the field of assessing commingled remains, but involve a staggering number of
variables.
8. Bibliography Adams, B.J., and Konigsberg, L.W. 2004. Estimation of the Most Likely Number of Individuals from Commingled Human Skeletal Remains. American Journal of Physical Anthropology 125: 138151.
Binford, L.R. and Bertram, J.B. 1977. Bone Frequencies And Attritional Processes. In Studies in Archaeology. ed. L.R. Binford. pp.77152. London: Academic Press Inc.
Boulter, S. and Rega, E. 0ctober 1993. Report of Human Remains from Black Gate, NewcastleUpon Tyne. Unpublished. University of Sheffield. pp.6674.
Brickley, M. and McKinley, J.I. (eds.) 2004. Guidelines to the Standards for Recording Human Remains. Southampton: BABAO.
Buikstra, J.E. and Ubelaker, D.H. (eds.) 1994. Standards for Data Collection from Human Skeletal Remains. Fayetteville, Arkansas: Arkansas Archaeological Survey.
Cherryson, A.K. 2010. “Such a restingplace as is necessary for us in God’s sight and fitting in the eyes of the world.”: Saxon Southampton and the Development of Churchyard Burial. In Burial in Later AngloSaxon England c.6501100AD. ed. J. Buckberry and A.K. Cherryson. pp.5472. Oxford: Oxbow Books.

51
Dobney, K.M. and Rielly, K. 1988. A Method for Recording Archaeological Animal Bones: The Use of Diagnostic Zones. Circaea 5: 7996.
Galloway, A., Willey, P. and Snyder, L. 1997. Human Bone Mineral Densities and Survival of Bone Elements: A Contemporary Sample. In Forensic Taphonomy: The Postmortem Fate of Human Remains. eds. W.D. Haglund and M.H. Sorg. 295317. London: CRC Press.
Garland, A.N. and Janaway, R.C. 1989. The taphonomy of inhumation burials. In. Burial Archaeology: Current Research Methods and Developments. eds. C. Roberts, F. Lee, and J. Bintliff. BAR British Series 211: 1537.
Henderson, J. 1987. Factors Determining the State of Preservation of Human Remains. In Death, Decay, and Reconstruction: Approaches to Archaeology and Forensic Science. ed. A. Boddington, A.N. Garland, and R.C. Janaway, pp.4353. Manchester: Manchester University Press.
Karr, L.P. and Outram, A.K. 2012. Tracking changes in bone fracture morphology over time: environment, taphonomy, and the archaeological record. Journal of Archaeological Science. 39: 555559.
Kendell, A. and Willey, P. 2014. Crow Creek Bone Bed Commingling: Relationship Between Bone Mineral Density and Minimum Number of Individuals and its Effect on Paleodemographic Analysis. In Commingled and Disarticulated Human Remains: Working Towards Improved Theory, Method, and Data. eds. A.J. Osterholtz, K.M. Baustial, and D.L. Martin. pp. 85104. New York: Springer Science.
Knüsel, C.J. and Outram, A.K. 2004. Fragmentation: The Zonation Method Applied to Fragmented Human Remains from Archaeological and Forensic Contexts. Environmental Archaeology 9: 8597.
Loe, L. and Cox, M. 2005. Peri and post mortem surface features on archaeological human bone: why they should not be ignored and a protocol for their identification and interpretation. Paper presented at The Fifth Annual Conference of the British Association for Biological Anthropology and Osteoarchaeology. eds. S.R. Zakrezewski and M. Clegg. pp. 1122. Oxford: Archaeopress.

52
Lyman, R. L. 1994. Vertebrate Taphonomy. Cambridge: Cambridge University Press.
Morlan, R.E. 1994. Bison Bone Fragmentation and Survivorship: a Comparative Method. Journal of Archaeological Science 21: 797807.
Nolan, J. 2010. The early medieval cemetery at the Castle, NewcastleuponTyne. Archaeologia Aeliana 39: 147287.
Outram, A.K., C.J. Knüsel, S. Knight, and A.F. Harding. 2005. Understanding Complex Fragmented Assemblages of Human and Animal Remains: A Fully Integrated Approach. Journal of Archaeological Science 32: 16991710.
Reitz, E.J. and Wing, E.S. 2008. Zooarchaeology. Cambridge: Cambridge University Press.
Roksandic, M. 2002. Position of Skeletal Remains as a Key to Understanding Mortuary Behavior. In Advances in Forensic Taphonomy: Method, Theory, and Archaeological perspectives. eds. W.D. Haglund and M.H. Sorg. 100133. London: CRC Press.
Saul, J.M. and Saul, F.P. 2002. Forensics, Archaeology, and Taphonomy: The Symbiotic Relationship. In Advances in Forensic Taphonomy: Method, Theory, and Archaeological perspectives. eds. W.D. Haglund and M.H. Sorg. 7197. London: CRC Press.
Scheuer, L. and Black, S. 2000. Developmental Juvenile Osteology. London: Academic Press.
Schultz, M. 1997. Microscopic Structure of Bone. In Forensic Taphonomy: The Postmortem Fate of Human Remains. eds. W.D. Haglund and M.H. Sorg. 187199. London: CRC Press.
Ubelaker, D.H. 1974. Reconstruction of Demographic Profiles from Ossuary Skeletal Samples: A Case Study from the Tidewater Potomac. Smithsonian Contributions to Anthropology 18. Washington DC: Smithsonian Institution Press. 1997. Taphonomic Applications in Forensic Anthropology. In Forensic Taphonomy: The Postmortem Fate of Human Remains. eds. W.D. Haglund and M.H. Sorg. 7790. London: CRC Press. 2002. Approaches to the study of Commingling is Human Skeletal Biology. In Advances in Forensic Taphonomy: Method, Theory, and Archaeological perspectives. eds. W.D. Haglund and M.H. Sorg. 332346. London: CRC Press.

53
Ubelaker, D.H. and Rife, J.L. 2008. Approaches to Commingling Issues in Archaeological Samples: A Case Study From Roman Era Tombs in Greece. In Recovery, Analysis, and Identification of Commingled Human Remains. eds. B.J. Adams and L.W. Konigsberg. pp.97122. Totowa: Humana Press.
Villa, P. 1991, Breakage Patterns of Human Long Bones. Journal of Human Evolution. 21: 2748.
Waldron, T. 1987. The Relative Survival of the Human Skeleton: Implications for Paleopathology. In Death, Decay, and Reconstruction: Approaches to Archaeology and Forensic Science. ed. A. Boddington, A.N. Garland, and R.C. Janaway, pp.5564. Manchester: Manchester University Press. 1994. Counting the Dead: The Epidemiology of Skeletal Populations. Chichester: Wiley & Sons Ltd.
Watson, J.P.N. 1972. Fragmentation Analysis of Animal Bone Samples From Archaeological Sites. Archaeometry. 14(2) 221228.
White, T.D., and Folkens, P.A. 2005. The Human Bone Manual. Burlington: Elsevier Academic Press.
Willey, P., A. Galloway, and L. Snyder. 1997. Bone Mineral Density and Survival of Elements and Element Portions in the Bones of the Crow Creek Massacre Victims. American Journal of Physical Anthropology. 104: 513528.
Williams, P.L. and Warwick, R. eds. 1980. Gray’s Anatomy 36th ed. London: Churchill Livingstone.
Appendix A. Zone frequencies, SDZ method.
Frontal Zone 1 Zone 2 Zone 3 Zone 4 Zone 5
# Present 17 17 9 18 6

54
# Incomplete 12 9 12 7 15
Left Temporal
Zone 1 Zone 2 Right Temporal
Zone 1 Zone 2
# Present 20 9 22 8
# Incomplete 3 9 5 13
Occipital Zone 1 Zone 2 Zone 3
# Present 30 8 4
# Incomplete 6 26 14
Mandible # Present # Incomplete
Zone 1 14 7
Zone 2 8 5
Zone 3 14 5
Zone 4 8 3
Zone 5 4 1
Zone 6 5 1
Zone 7 12 4
Zone 8 10 1
Zone 9 8 1
Appendix B. Raw data from the SDZ method.
Copyright © 2022 FDOKUMEN




















