
Dissertacao para impressao 21 02 11
93
SUMÁRIO 1 2 1. INTRODUÇÃO.............................................................................................................17 3 2. REVISÃO DE LITERATURA.....................................................................................19 4 Papilomavírus................................................................................................................19 5 Patologia dos papilomavírus bovino (BPV)................................................................25 6 Resposta imune ao BPV................................................................................................32 7 Imunoprofilaxia para o BPV........................................................................................34 8 Considerações finais......................................................................................................34 9 Referências.....................................................................................................................37 10 3. OBJETIVOS..................................................................................................................46 11 4. ARTIGOS PARA PUBLICAÇÃO...............................................................................47 12 4.1 Teat papillomatosis associated with recent characterized BPV types (BPV-7, 13 -9 and -10) in brazilian cattle………………………………………………………48 14 4.2 Phylogenetic analyses of bovine papillomavirus (BPV) of teat papillomas in 15 Brazilian dairy cattle herds…………………………………………………………63 16 5. CONCLUSÕES………………………….....………………………………………..90 17 APÊNDICES........................................................................................................................91 18 ANEXOS.............................................................................................................................101 19 20
Transcript of Dissertacao para impressao 21 02 11

SUMÁRIO 1
2
1. INTRODUÇÃO.............................................................................................................17 3
2. REVISÃO DE LITERATURA.....................................................................................19 4
Papilomavírus................................................................................................................19 5
Patologia dos papilomavírus bovino (BPV)................................................................25 6
Resposta imune ao BPV................................................................................................32 7
Imunoprofilaxia para o BPV........................................................................................34 8
Considerações finais......................................................................................................34 9
Referências.....................................................................................................................37 10
3. OBJETIVOS..................................................................................................................46 11
4. ARTIGOS PARA PUBLICAÇÃO...............................................................................47 12
4.1 Teat papillomatosis associated with recent characterized BPV types (BPV-7, 13 -9 and -10) in brazilian cattle………………………………………………………48 14
4.2 Phylogenetic analyses of bovine papillomavirus (BPV) of teat papillomas in 15 Brazilian dairy cattle herds…………………………………………………………63 16
5. CONCLUSÕES………………………….....………………………………………..90 17
APÊNDICES........................................................................................................................91 18
ANEXOS.............................................................................................................................101 19
20

17
1. INTRODUÇÃO 1
2
O rebanho bovino brasileiro cresce, em média, 1,5% ao ano e chegou a mais de 3
200 milhões de cabeças em 2010 conferindo ao país os títulos de segundo maior 4
rebanho de bovinos do mundo - atrás apenas da Índia -, de segundo maior produtor de 5
carne bovina - depois dos Estados Unidos, e de maior exportador mundial do produto 6
(FAO). 7
Além da carne, o Brasil também se destaca pela produção anual do leite bovino, 8
a qual registrou recordes de exportação entre 2000 e 2008 e em 2009 foi de, 9
aproximadamente, 30 bilhões de litros, com alta de 5,6% em relação ao ano anterior 10
(IBGE, 2010). De acordo com o Ministério de Desenvolvimento, Indústria e Comércio 11
Exterior (MDIC), esse crescimento anual da produção de leite no Brasil – que em média 12
atingiu 1,95% ao ano - corresponde ao aumento da demanda no mercado internacional, 13
principalmente da China, e a menor oferta dos países produtores de leite. Para Rocha 14
(2007), este cenário torna o Brasil também um forte candidato a grande exportador 15
mundial de leite e de produtos lácteos. 16
Entretanto, do ponto de vista sanitário, os rebanhos bovinos leiteiros brasileiros 17
apresentam-se susceptíveis a várias doenças de caráter infecto-parasitário que podem 18
comprometer consideravelmente a saúde animal, a produção leiteira e a rentabilidade da 19
atividade. Entre as enfermidades tem-se as doenças agudas, que evoluem com sinais 20
clínicos e que são visíveis em uma observação, e as infecções endêmicas com evolução 21
subclínica ou crônica e que são “invisíveis”, o que resulta na tendência de se 22
perpetuarem no rebanho. Como o controle dessas doenças apresenta alto custo 23
financeiro devido o gasto dispendioso com os tratamentos – quando há, pois no caso de 24
certas infecções virais ele não existe - e descarte do animal, resta a alternativa de 25
condutas como o diagnóstico laboratorial e a profilaxia (ALFIERI, 2008). 26
Um exemplo de infecção viral que causa grandes prejuízos aos rebanhos bovinos 27
é a papilomatose cutânea, causada pelo papilomavírus bovino (bovine papillomavirus - 28
BPV) que está amplamente distribuído em rebanhos de todo o mundo e é considerado 29
endêmico em todo o território brasileiro (ALFIERI et al., 2007). Entre seus efeitos 30
destacam-se as infecções secundárias, a dificuldade de amamentação do bezerro, a 31
depreciação do couro e o descarte precoce de animais e ainda, no caso de rebanhos de 32
aptidão leiteira, a papilomatose mamária ocasiona dificuldade de ordenha, retenção de 33
leite e predisposição a mastite. 34

18
Mas se por um lado a papilomatose cutânea caracteriza-se como uma virose que 1
causa prejuízos ao rebanho, por outro ela ainda constitui-se uma doença negligenciada 2
do ponto de vista científico e profissional. Exemplo disso são os resultados 3
contraditórios e deficientes dos vários tratamentos sugeridos, cuja grande maioria não 4
apresenta comprovação de eficiência (SILVA et al., 2004). 5
De acordo com este cenário, desenvolver métodos baseados em evidências 6
científicas para o controle e profilaxia de uma doença negligenciada como a 7
papilomatose, pode trazer reflexos diretos na produção leiteira da grande maioria dos 8
rebanhos nacionais. 9
10

19
2. REVISÃO DE LITERATURA 1
2
Papilomavírus 3
4
O papilomavírus constitui um grupo de vírus DNA altamente diverso com 5
potencial para a indução de lesões hiperproliferativas benignas e malignas, tanto na pele 6
quanto em mucosas de seus hospedeiros naturais. Muito provavelmente, os PVs 7
ocorram na maioria das espécies de mamíferos, aves e répteis (de VILLIERS et al., 8
2004). 9
10
Classificação viral 11
12
A caracterização do genoma dos papilomavírus (PVs) foi sempre o critério 13
utilizado para sua nomenclatura e taxonomia, uma vez que eles não apresentam cultura 14
celular nem marcadores sorológicos. Tradicionalmente, os isolados de PVs que tinham 15
DNA significativamente diferentes de outros isolados eram chamados de “tipos”, e 16
todos os PVs eram agrupados com os poliomavírus na família Papovaviridae. 17
Entretanto, com os estudos moleculares realizados nos últimos 25 anos os PVs 18
foram diferenciados dos poliomavírus. O resultado de comparações filogenéticas de 19
grande número de PVs resultou em sua nova classificação, reconhecida pelo “Comitê 20
Internacional de Taxonomia Viral” (BERNARD, 2007). Atualmente eles são 21
classificados taxonomicamente na família Papillomaviridae, que apresenta várias 22
divisões. 23
Os papilomavírus são identificados pela abreviatura PV, juntamente com uma ou 24
duas letras indicando a espécie hospedeira. Os tipos de PVs seguem a ordem numérica 25
de descrição. Nos últimos 20 anos o Reference Center for Papillomaviruses at the 26
German Cancer Research Center tem sido responsável pelo processo de registro dos 27
HPVs (CHAN et al., 1995; DOOBAR, 2005; BERNARD, 2005; BERNARD, 2007). 28
Nesta família, como a sequência de nucleotídeos mais conservada é a ORF L1, 29
ela é responsável pela identificação de novos tipos virais do genoma do PV. Assim, 30
seguindo a taxonomia atual, o maior ramo taxonômico da família Papillomaviridae é 31
formado pelo “gênero”, que compartilha menos de 60% de identidade na análise de 32
nucleotídeos da ORF L1 e entre 23% e 43% de identidade de nucleotídeos na análise do 33

20
genoma completo. O sub-ramo taxonômico é formado por “espécies” que compartilham 1
no gênero entre 60 e 70% de identidade de nucleotídeos. Um novo “tipo” é reconhecido 2
quando tiver seu genoma completo clonado e o sequenciamento da L1 demonstrar 3
diferença de identidade de nucleotídeos superior a 10%, quando comparado a todos os 4
outros PV já descritos naquela espécie. Diferenças entre 2% e 10% definem um 5
“subtipo” e inferiores a 2% caracterizam uma “variante” viral. Quando somente uma 6
parte do gene L1 é amplificada e avaliada, sugerindo essas diferenças, o isolado é 7
denominado de suposto novo tipo (de VILLIERS et al., 2004; BERNARD 2005). Esse 8
critério, originalmente estabelecido para o HPV, tem sido extrapolado para outros PVs 9
identificados em outras espécies animais como a bovina, uma vez que não existe 10
padronização na taxonomia dos PV isolados em animais (de VILLIERS et al, 2004). 11
Estudos demonstram que o genoma dos PVs evolui lentamente. A árvore 12
filogenética de todos os PVs de origem humana e animal apresenta uma história 13
evolutiva tão antiga quanto a dos seus respectivos hospedeiros naturais, isto é, mais de 14
100 milhões de anos nos casos de PVs de vertebrados e milhares de centenas de anos 15
para os human papilomavirus (HPVs). No nível taxonômico de gênero existe uma 16
correlação limitada entre classificação taxonômica, biologia molecular e propriedades 17
patogênicas, porém, no ramo taxômico que formam uma espécie, os tipos de PV 18
possuem similaridade genômica e propriedades biológicas semelhantes (BERNARD, 19
2007). 20
De acordo com as características clínicas, anatomopatológicas e moleculares, os 21
BPVs -1 a -6 foram distribuídos em subgrupos na antiga classificação. O subgrupo A 22
(BPV-1,-2 e -5) era constituído por vírus que induzem proliferação fibroblástica e 23
subgrupo B (BPV-3,-4 e -6) compreendem os vírus epiteliotrópicos (CAMPO, 1995). 24
Esta distribuição foi baseada na premissa de que cada tipo de BPV tenha especificidade 25
por algum tecido, apresente determinado aspecto macroscópico e tenha localização 26
anatômica específica no corpo do animal afetado. Os estudos moleculares recentes, 27
envolvendo a identificação do BPV presente em lesões cutâneas, sugerem que talvez 28
esse critério de classificação de acordo com aspecto e localização do papiloma não deva 29
ser totalmente seguido. A diversidade dos PVs em seres humanos já está comprovada e 30
alguns tipos virais já foram descritos em outros locais além do primariamente 31
identificado (DOORBAR, 2005; CLAUS et al., 2008c). 32
Atualmente, a família Papillomaviridae, com base na biologia e homologia 33
genômica, é formada por 18 gêneros (Alphapapillomavirus a Sigmapapillomavirus) (de 34

21
VILLIERS et al., 2004). Os tipos de BPV identificados até o momento são classificados 1
em quatro gêneros que incluem o Deltapapillomavirus (BPV-1 e -2), Xipapillomavirus 2
(BPV-3, -4, -6, -9, -10 e -11), Epsilonpapillomavirus (BPV-5 e -8) e um tipo ainda não 3
nominado (BPV-7) (de VILLIERS et al., 2004; OGAWA et al., 2004; TOMITA et al., 4
2007; HATAMA et al., 2008). 5
6
Estrutura e organização genômica dos papilomavírus 7
8
Um aspecto importante que caracteriza os PVs é a dificuldade de propagação in 9
vitro em culturas de monocamada celular, uma vez que in vivo esses vírus se replicam, 10
naturalmente, no núcleo das células do epitélio e requerem o aparato de diferenciação 11
celular para a replicação (CAMPO, 2002). 12
Os PVs são considerados vírus altamente espécie-específicos. O único caso 13
singular de infecção entre espécies é a infecção de equídeos por papilomavírus bovino 14
tipos -1 e -2. Após infecção acidental da derme, principalmente os equídeos podem 15
desenvolver tumores dermais benignos conhecidos como sarcóide equino (CAMPO, 16
2002). 17
Em humanos, os estudos de vírus oncogênicos tem proporcionado importantes 18
descrições de processos celulares como o armazenamento e expressão da informação 19
genética, o controle de ciclo celular, a tradução de sinal, a regulação da resposta imune e 20
carcinogênese. A identificação dos tipos virais envolvidos na patologia de tumores 21
possibilita novas oportunidades de tratamento clínico e o desenvolvimento de 22
estratégias de vacinação. Acredita-se que a ação de imunoprofilaxia com abordagens 23
específicas antivirais, reduza a incidência desses tumores (NASIR e CAMPO, 2008). 24
Os estudos com o BPV em bovinos tem possibilitado a compreensão da história 25
natural do vírus, a ligação direta entre a infecção viral e neoplasia, a correlação entre 26
vírus-hospedeiro-ambiente, o esclarecimento dos mecanismos imunológicos envolvidos 27
na resposta à infecção e o desenvolvimento de vacinas antipapilomavírus (NASIR e 28
CAMPO, 2008). 29
Os PVs são pequenos vírus oncogênicos, não envelopados, com 55-60nm de 30
diâmetro e com genoma constituído por DNA fita dupla. O capsídeo viral é composto 31
por 72 pentâmeros (capsômeros), sendo que 60 capsômeros se ligam de forma 32
hexavalente e 12 de forma pentavalente, arranjados em superfície de triangulação T=7, 33
composto pela proteína principal (L1) do capsídeo e a proteína secundária interna do 34

22
capsídeo (L2). O DNA genômico viral consiste numa molécula fita dupla circular com 1
7.3 a 8 kpb. Nos vírions e nas células hospedeiras o genoma está conjugado à 2
nucleossomos, apresentando aspecto empacotado como cromossomos. A massa 3
molecular do ácido nucléico é de 5.0x106 daltons e representa 12% da massa do vírion. 4
A partícula viral é resistente às condições do meio ambiente e a solventes lipídicos 5
como o éter e clorofórmio (GARCEA e CHEN, 2007; ALFIERI et al., 2007). 6
O genoma do PV possui sequências conservadas, localizadas em uma das fitas 7
do DNA, indicando que somente uma fita é utilizada como molde para a transcrição das 8
proteínas. Não são observadas diferenças na organização entre os gêneros de 9
papilomavírus. A fita codificante contém cerca de 10 sequências abertas de leitura (open 10
reading frames - ORFs), que são classificadas conforme a fase de expressão temporal, 11
sendo que o segmento E (early) contém oito ORFs iniciais, e o segmento L (late) 12
contém duas ORFs tardias. Além das sequências conservadas existe no genoma viral a 13
região LCR (long control region - LCR) que controla os processos de transcrição e 14
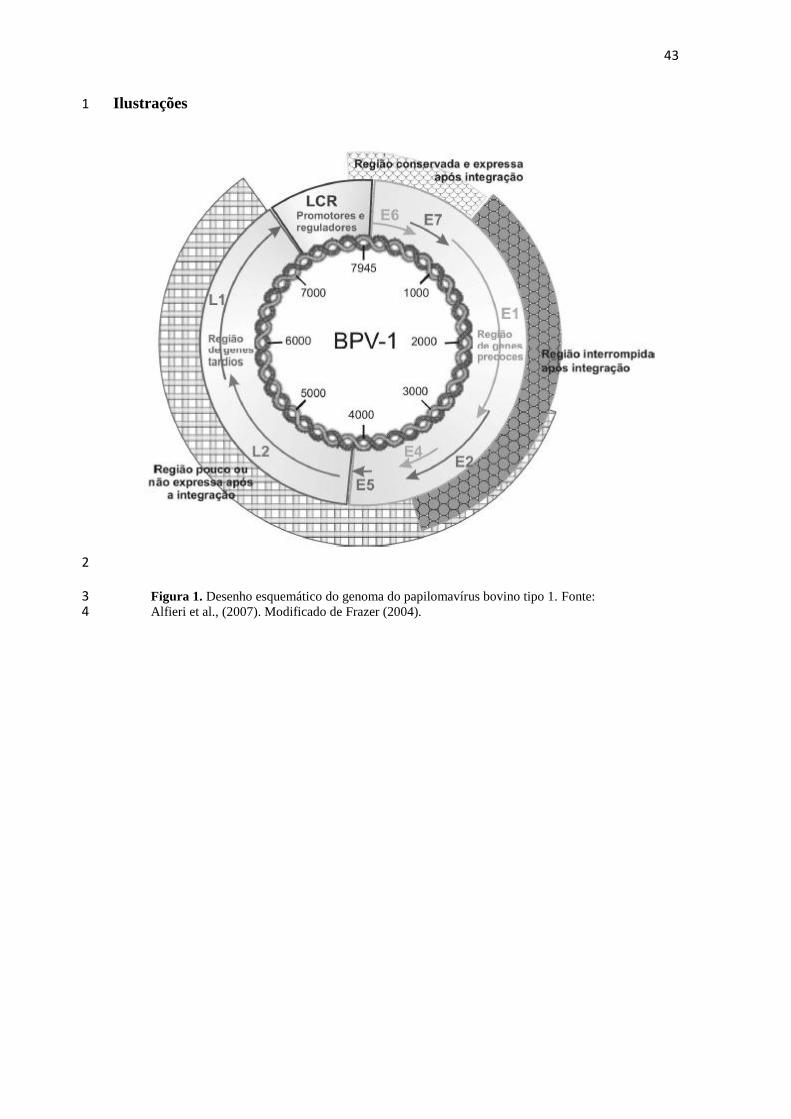
replicação viral. O desenho esquemático do genoma do BPV-1 está representado na 15
figura 1 (ALFIERI et al., 2007). 16
O segmento E representa 45% do genoma viral e codifica proteínas necessárias 17
para as fases de transcrição e de replicação viral. Nesse segmento, estão as ORFs que 18
codificam as proteínas regulatórias e as proteínas oncogênicas dos PVs. As proteínas 19
“E” são expressas em células recém infectadas, em infecções não produtivas, assim 20
como em células transformadas. O segmento L representa 40% do genoma viral e 21
codifica as proteínas do capsídeo (L1 e L2), que são produzidas nas fases tardias da 22
replicação viral e são encontradas somente em células com infecções produtivas. As 23
ORFs dos PVs estão sobrepostas compactando vários genes em uma pequena extensão 24
do genoma. Entre os segmentos E e L está o segmento LCR que representa 15% do 25
genoma viral que não codifica proteínas, mas é constituído por elementos reguladores 26
da replicação e da transcrição e por promotores que são responsáveis pelos fatores 27
celulares, além de conter o ponto de origem (ori) da replicação viral (HOWLEY e 28
LOWY, 2001; ALFIERI et al., 2007). 29

23
Controle da transcrição 1
2
A LCR do BPV contém 12 locais que interagem com a proteína E2. Essa 3
proteína é fundamental no ciclo viral do BPV, sendo que é um fator de replicação do 4
DNA viral e o maior regulador de transcrição viral, que se liga à região LCR ativando 5
ou reprimindo a transcrição dos genes virais. Além disso, vários outros fatores de 6
transcrição celular interagem com a LCR, tanto promovendo quanto inibindo a 7
transcrição dos genes virais. Essas interações resultam em um circuito sincronizado 8
onde elementos de controle, viral e celular, contribuem para regular a expressão dos 9
genes virais Quando essa harmonia é desestabilizada, a transcrição dos genes e 10
expressão das proteínas víricas aumentam, e o resultado é a transformação celular para o 11
estado neoplásico (NASIR e CAMPO, 2008). 12
13
Expressão das proteínas virais 14
15
Os genes E1 e E2 estão envolvidos na regulação da transcrição e na replicação 16
do vírus. Os genes E5, E6 e E7 apresentam regulação negativa dos genes E1 e E2. Na 17
ausência de E1 e E2 os produtos de E6 e E7 são oncogenes potentes. No BPV-5, o gene 18
E5 é o principal gene encontrado em células transformadas. Esse gene é altamente 19
conservado entre os PVs indutores de fibropapilomas nos seus hospedeiros naturais, 20
além de ter a capacidade de induzir tumores fibroblásticos em hamsters. Provavelmente, 21
o gene E5 seja o principal responsável pela proliferação de fibroblastos em 22
fibropapilomas (HOWLEY e LOWY, 2001). 23
Alguns tipos de BPVs não possuem todos os oncogenes virais em seu genoma. 24
No BPV-7, descrito em 2007, o gene E5 não foi identificado. A ORF E6 está ausente 25
nos BPVs-3, -4 e -6, assim como em outros PVs (JACKSON et al.,1991; TERAI et al., 26
2002). Uma hipótese relevante seria a de que um rearranjo genômico, ocorrido em 27
ancestrais virais, pode ter contribuído para a evolução de alguns tipos de PVs sem 28
alguns oncogenes virais. Esses genes também estão envolvidos na adaptação do genoma 29
dos PVs a vários hospedeiros e tecidos (GARCEA e CHEN, 2007). Em contraste, as 30
ORFs E1, E2, L2 e L1 são bem conservadas em todos tipos de PVs e seus produtos são 31
proteínas essenciais para o ciclo de replicação viral (de VILLERS et al., 2004). 32
As proteínas estruturais L1 e L2 são expressas nos núcleos de queratinócitos 33
diferenciados. Durante a morfogênese do virion, a proteína L2 se liga ao DNA viral, 34

24
auxiliando a montagem do vírus. As proteínas L1 e, possivelmente, L2 mediam a 1
ligação do vírus com o receptor de membrana celular na infecção. As duas proteínas 2
codificam epítopos neutralizantes e tem sido empregas com sucesso no 3
desenvolvimento de vacinas (NASIR e CAMPO, 2008). 4
O estado físico do genoma do PV pode se apresentar sob duas formas, conforme 5
o tipo de lesão associada à infecção. Nas lesões benignas, o genoma está presente em 6
múltiplas cópias, não integrado ao genoma celular, isto é, na forma epissomal. Nas 7
lesões malignas, o genoma do PV pode se integrar ao genoma da célula hospedeira 8
formando uma ligação estável. O vírus integrado é incapaz de completar o ciclo 9
replicativo, visto que os genes essenciais para o término da replicação estão 10
interrompidos. Quando o fenômeno da integração acontece, o genoma circular do PV é 11
rompido, geralmente entres os genes E1 e E2. Com a inativação do gene E2, que 12
codifica uma proteína reguladora que reprime a transcrição excessiva das proteínas E6 e 13
E7, há aumento dos produtos desses genes (CULLEN et al., 1991). 14
15
Ciclo replicativo 16
17
Os PVs que induzem uma variedade de lesões hiperproliferativas em tecidos 18
epiteliais. Apesar das diferenças de potencial oncogênico desses vários tipos de PVs, é 19
provável que eles compartilhem mecanismos comuns durante o ciclo de vida produtiva 20
(HOWLEY e LOWY, 2001). 21
O ciclo de vida produtiva do BPV está intimamente ligado ao programa de 22
diferenciação do tecido epitelial do hospedeiro (HOWLEY e LOWY, 2001). O receptor 23
celular de adsorção viral ainda é desconhecido, embora o heparan sulfate pareça 24
facilitar o acesso do vírus em alguns tipos de células. Os alvos da infecção inicial são as 25
células epiteliais basais, que são expostas como resultado de microlesões do epitélio 26
estratificado. Após a ligação e entrada do vírus na célula, os virions migram para o 27
núcleo (LEE e LAIMINS, 2007). A replicação do genoma ocorre na fase S do ciclo 28
celular por meio da ação das proteínas virais E1 e E2, juntamente com proteínas de 29
replicação celular. Após a replicação do genoma viral e divisão celular uma das células 30
filhas migra para longe da camada basal e inicia o programa de diferenciação. 31
Queratinócitos não infectados interrompem o ciclo celular replicativo, após deixar a 32
camada basal, e o núcleo é degradado em muitas dessas células em diferenciação. Em 33
contrapartida, as células infectadas pelo BPV se diferenciam, mas permanecem com o 34

25
ciclo celular ativo. Isto permite que células suprabasais, altamente diferenciadas, voltem 1
a entrar na fase S e mantenham o alto nível de replicação viral. A capacidade das células 2
diferenciadas de manter a progressão do ciclo celular é mediada, principalmente, pela 3
ação da proteína viral E7. A amplificação do genoma viral dependente da diferenciação 4
em células suprabasais e coincide com a ativação dos promotores virais tardios. As 5
transcrições tardias codificam as proteínas virais do capsídeo (L1 e L2). A progênie dos 6
virions é montada em células altamente diferenciadas e, em seguida, liberada para o 7
meio extracelular (LEE e LAIMINS, 2007). 8
Estudo tem demonstrado que vários tipos de HPV podem ser, aparentemente, 9
não patogênicos e comportarem-se como agentes comensais da pele saudável de seres 10
humanos (FOURSLUND et al., 1999; ANTONSSON et al., 2000). O mesmo ocorre em 11
várias espécies de animais, incluindo a bovina, onde tanto BPVs já caracterizados 12
quanto aqueles ainda descritos como supostos novos tipos virais tem sido isolados a 13
partir de swabs de pele saudável (ANTONSSON e HANSSON, 2002; OGAWA et al., 14
2004; ANTONSSON e MC MILLAN, 2006). 15
16
Patologia do papilomavírus bovino (BPV) 17
18
Os PVs são altamente espécie-específicos e estão relacionados com lesões no 19
epitélio cutâneo e mucoso de várias espécies de mamíferos, aves e répteis (DOOBAR, 20
2005). As infecções entre diferentes espécies hospedeiras são raras, mas não há infecção 21
produtiva na segunda espécie. O sarcóide, que é o tumor de pele mais comumente 22
relatado em equinos e muares, está associado à infecção desses animais pelo BPV-1 e 23
BPV-2 (NASIR e CAMPO, 2008). 24
Os BPVs-1 e -2 pertencem ao gênero Deltapapillomavirus e são, usualmente, 25
denominados fibropapilomavírus. Com frequência, causam infecção no epitélio e na 26
derme induzindo a formação de fibropapilomas. Os BPVs-3, -4, -6, -9, -10 e -11 27
pertencem ao gênero Xipapillomavirus, são vírus exclusivamente epiteliotrópicos e 28
induzem a formação papilomas verdadeiros. Os BPVs-5 e -8 são membros do gênero 29
Episilonpapillomavirus e parecem apresentar patologia dupla, causando fibropapilomas 30
e papilomas epiteliais e seu genoma compartilha homologia tanto com o gênero 31
Deltapapillomavirus quanto com o gênero Xipapillomavirus. Com base em análises 32
filogenéticas foi proposto que o BPV-7 pertença a um novo gênero de BPV. Com base 33
em análises parciais do gene L1, supostos novos tipos de BPVs têm sido descritos e 34

26
aguardam a caracterização genômica completa para a definição taxonômica (BLOCH et 1
al, 1996; OGAWA et al., 2004; CLAUS et al. 2008a; OGAWA et al., 2007; TOMITA 2
et al., 2007; NASIR e CAMPO, 2008). 3
A replicação viral ocorre nos queratinócitos, durante o processo de diferenciação 4
do epitélio escamoso. Nos fibroblastos, onde o genoma do BPV está presente na forma 5
epissomal não há replicação viral. Papilomas e fibropapilomas podem ocorrer em 6
diferentes sítios anatômicos de bovino onde diferentes genotipos de BPV são 7
encontrados. Os BPVs também podem ser encontrados em células polimorfonucleares 8
do sangue periférico, no entanto, não existe evidência da replicação viral nessas células. 9
Essa observação é importante pela implicação na patogênese da infecção, pois sugere 10
que a corrente sanguínea pode carrear o vírus para diferentes tecidos 11
(BORZACCHIELLO e ROPERTO, 2008). 12
13
Tumores do trato digestório e bexiga 14
15
Evidências epidemiológicas demonstram que algumas lesões benignas podem 16
sofrer transformação maligna em resposta a fatores genéticos ou ambientais 17
(HOPKINS, 1986). Nesse contexto, a hematúria enzoótica bovina (HE) e o tumor de 18
trato digestório superior em bovinos são enfermidades relacionadas à associação entre 19
infecção pelo BPV e a ingestão da planta samambaia (CAMPO et al., 1992). 20
A hematúria enzoótica bovina apresenta caráter enzoótico em determinadas 21
regiões geográficas que reúnem condições ideais para o crescimento da samambaia. A 22
samambaia é uma pteridófita do gênero Pteridium, espécie P. aquinilum, de distribuição 23
cosmopolita em todas as regiões tropicais. A samambaia apresenta em sua composição 24
diversas substâncias mutagênicas, carcinogênicas e imunossupressivas. A 25
carcinogenicidade da planta tem sido atribuída a quercetina, ptaquilosídeos e ao ácido 26
shiquímico (ALFIERI et al., 2007). 27
A infecção pelo BPV-2 e a intoxicação crônica pela samambaia foram 28
associadas com lesões vesicais e a sinais cílincos de hematúria enzoótica em bovinos 29
adultos. Vários estudos indicam que a progressão maligna das lesões do aparelho 30
urinário depende de uma inter-relação entre a infecção pelo BPV-2 e os compostos 31
tóxicos presentes na samambaia (CAMPO, 1995; WOSIACHI et al., 2005). 32
O diagnóstico do BPV-2 por meio de técnicas convencionais é de difícil 33
realização, particularmente pela não adaptação do vírus em sistemas de replicação in 34

27
vitro em culturas celulares. Os avanços nos métodos de diagnóstico molecular tornaram 1
possível o diagnóstico da infecção pelo PV partir da identificação do DNA viral por 2
meio de técnicas de hibridação molecular ou pela reação em cadeia pela polimerase 3
(PCR) (WOSIACHI et al., 2005). 4
Em bovinos, a infecção pelo BPV-4 pode resultar em papilomas no trato 5
digestório superior que podem progredir para o câncer quando associada aos compostos 6
tóxicos presentes na samambaia (CAMPO, 2002; BORZACCHIELLO et al., 2003). 7
8
Papilomatose cutânea bovina 9
10
Na espécie bovina, o BPV está amplamente distribuído em rebanhos bovinos de 11
todo mundo, sendo que no Brasil, independentemente do nível de tecnificação da 12
exploração pecuária, a infecção pode ser encontrada tanto em rebanhos bovinos de corte 13
quanto, principalmente, em rebanhos com aptidão leiteira em praticamente todo o país. 14
A maior prevalência da infecção pelo BPV observada nos rebanhos leiteiros, resultando 15
principalmente no quadro da papilomatose cutânea, pode ser justificada pelo emprego 16
de práticas de manejo mais intensivas, comuns a esse tipo de exploração (CLAUS et al., 17
2007). 18
Em rebanhos com papilomatose cutânea, na dependência da extensão das lesões, 19
pode-se observar o comprometimento do desenvolvimento corporal dos animais, 20
predisposição a infecções secundárias, queda na produção de leite e redução do valor de 21
mercado do animal, entre outras consequências que podem acarretar prejuízos 22
econômicos à exploração pecuária de corte e, principalmente, leiteira (CAMPO, 2002). 23
Normalmente, ocorre a regressão espontânea dos papilomas como resultado da resposta 24
imune celular, mas alguns animais são incapazes de combater a infecção e podem 25
sucumbir frente a infecção cutânea generalizada ou ao envolvimento da mucosa. Estas 26
formas de papilomatose são problemáticas e de grande importância econômica 27
(BORZACCHIELLO e ROPERTO, 2008). 28
As lesões cutâneas podem apresentar aspectos variados e são classificadas 29
macroscopicamente em filiformes, na forma típica pedunculada e nas formas atípicas, 30
planas. Os papilomas do tipo pedunculado apresentam aspecto verrucoso, sugerindo a 31
forma de “couve-flor”. A base de inserção pode ser ampla ou estreita, com coloração 32
variando do cinza ao preto e desprovida de pêlos. As lesões, dependendo do número, 33
podem formar uma grande massa e comprometer extensas áreas da pele. Os papilomas 34

28
planos são achatados, com base ampla, de coloração esbranquiçada e com presença de 1
pêlos. Podem se apresentar de forma agrupada ou isolada e são de difícil remoção 2
(GUPTA et al., 1989; SMITH et al., 1995). 3
Alguns tipos de BPVs, ainda não identificados, infectam o epitélio vulvo-4
vaginal e os olhos (HATAMA et al., 2008). Existem algumas formas de apresentação 5
clínica que são relacionadas com o tipo de BPV destacando-se: BPV-1 com 6
fibropapilomas em teto e pênis; BPV-2 com verrugas cutâneas, fibropapilomas do trato 7
digestório e tumores de bexiga; BPV-3 com papilomas cutâneos; BPV-4 com papilomas 8
epiteliais do trato digestório superior; BPV-5 com fibropapilomas de morfologia 9
macroscópica em formato de grão de arroz no úbere; BPV-6 com papilomas de 10
morfologia macroscópica classificada como “frond” presentes nos tetos; BPV-8 com 11
papilomas cutâneos; BPVs-9 e -10 associados com papilomas de teto e úbere 12
(BORZACCHIELLO e ROPERTO, 2008). 13
Embora se tenha correlacionado os genotipos virais com o aspecto morfológico 14
macroscópico das lesões, estudos atuais demonstram resultados que não correspondem a 15
essa correlação (CLAUS et al., 2008a; MAEDA et al., 2007). Fibropapilomas que 16
ocorrem no prepúcio e pênis podem sangrar e, frequentemente, apresentar áreas de 17
necrose. Os fibropapilomas podem se espalhar ao longo do períneo e como os animais 18
infectados apresentam a tendência de lamberem as lesões, as mesmas podem se 19
disseminar para o focinho e a boca. Os fibropapilomas podem ainda causar perda de 20
função reprodutiva e, com isso, promover o descarte precoce dos animais. As lesões 21
cutâneas induzidas pelo BPV-2 são geralmente encontradas na testa, pescoço, tórax e 22
costas. Surtos de papilomatose cutânea têm sido descritos em animais confinados 23
(BORZACCHIELLO e ROPERTO, 2008). 24
O exame histopatológico é uma das formas indiretas de diagnóstico do 25
papilomavírus. Embora permita a identificação de neoplasias intra-epiteliais, essa 26
técnica não possibilita a identificação do tipo viral associado à lesão. Achados 27
histológicos dos papilomas incluem: i) presença de vesículas intracitoplasmáticas 28
(degeneração hidrópica) em células do estrato espinhoso, ii) aumento na quantidade de 29
granulações basofílicas, intracitoplasmáticas e intranucleares no estrato granuloso; e iii) 30
inclusões eosinofílicas intranucleares (HEAD et al., 2002). 31
A observação microscópica dos papilomas cutâneos revela a presença de 32
hiperqueratose e hiperplasia celular da camada espinhosa (acantose). Essas células 33
mostram aumento de tamanho e no número dos desmossomos e tonofibrilas. Outras 34

29
células epiteliais podem apresentar alterações degenerativas como a perda das 1
tonofibrilas, separação dos desmossomos, atipia nuclear focal e vacuolização do 2
citoplasma. Nas camadas superiores do epitélio essas mudanças são mais pronunciadas. 3
Na camada granular a degeneração nuclear é evidente, com marginação e condensação 4
da cromatina (LANCASTER e OLSON, 1982). Em contraste com essas alterações 5
celulares estão os fibromas induzidos pelos fibropapilomavírus. Estes fibromas 6
apresentam extensiva proliferação fibroblástica, com as células arranjadas em círculo e 7
com padrões irregulares. Os fibropapilomas exibem uma combinação desses efeitos, 8
sendo que a primeira reação da pele é a estimulação dos fibroblastos na derme, 9
acompanhada por resposta inflamatória com congestão, edema e infiltração leucocitária. 10
Em torno de uma semana após a infecção a reação inflamatória diminui, porém a 11
estimulação fibroblástica prossegue, seguida pela invasão da camada papilar da derme 12
por fibroblastos (JELÍNEK e TACHEZY, 2005). Nos papilomas pedunculados, as 13
cristas interpapilares penetram profundamente no interior da derme, ao contrário dos 14
papilomas planos onde o tecido da derme sofre pouco ou nenhuma fibroplasia. A 15
literatura ressalta a importância da análise do tipo de lesão, uma vez que estudos 16
demonstram que mesmo com os achados macroscópicos e histológicos compatíveis com 17
a presença do vírus, somente técnicas de diagnóstico etiológico podem comprovar a 18
presença viral (GUPTA et al., 1989; SANTIN e BRITO, 2004; MCGAVIN e 19
ZACHARI, 2007). 20
21
Papilomatose mamária 22
23
A papilomatose mamária é uma forma de manifestação clínica da papilomatose 24
cutânea na ocorrência neste sítio anatômico. As lesões causadas pelos BPVs em tetos, 25
assim como na papilomatose cutânea, são denominadas verrugas, papilomatose 26
mamária ou figueira. Atualmente, foram encontrados oito tipos de BPVs induzindo 27
lesões em tetos e úberes de bovinos, além de outros tipos virais ainda não 28
caracterizados. Desde a década de 80, vários estudos que visavam a identificação de 29
tipos virais associados aos papilomas do aparelho mamário bovino identificaram o 30
BPV-6 como tipo mais prevalente neste sítio anatômico e, a partir desses estudos, a 31
papilomatose mamária bovina tem sido associada principalmente ao BPV-6 32
(LINDHOLM et al., 1984; JARRET et al., 1984; MAEDA et al. 2007). 33

30
Lindholm et al. (1984) em estudo realizado com lesões epiteliais de teto animais 1
provenientes de abatedouro identificaram o BPV-6 em 92% das lesões avaliadas. Maeda 2
et al. (2007) investigaram um surto de papilomatose mamária em bovinos de leite no 3
Japão em que 80% do rebanho de 700 animais foi comprometido e encontraram o BPV-4
6 em, aproximadamente, 86% (12/14) das lesões analisadas. Ogawa et al. (2004) 5
utilizando dois pares de primers genéricos, desenvolvidos para identificação de HPVs 6
cutâneos e mucosos, avaliaram lesões e swabs cutâneos de tetos de bovino de leite de 5 7
properiedades e identificaram o BPV-6 somente em um swab de pele de teto saudável. 8
No Brasil, um estudo de identificação de tipos de BPV em lesões cutâneas, o BPV-6 foi 9
o tipo viral mais prevalente em papilomas de tetos de bovinos de corte (3/3) em 3 10
rebanhos distintos (CLAUS et al, 2007). Outro estudo no Brasil identificou o BPV-6 11
associado em co-infecção ao BPV-1 em lesão de teto de bovino de corte (CLAUS et al., 12
2008b). Atualmente não há estudos no Brasil com objetivo de identificar os tipos virais 13
envolvidos na papilomatose mamária. Entretanto, em todo o mundo, já foram 14
identificados vários tipos de BPVs (BPV-1, -3, -5, -7, -8, -9, -10), além de tipos virais 15
ainda não totalmente caracterizados (supostos novos tipos), a partir de papilomas de 16
aparelho mamário bovino (CAMPO et al., 1981; JARRET et al., 1984; LINDHOLM et 17
al., 1984; CAMPO, 2002; OGAWA et al., 2004; MAEDA et al., 2007; CLAUS et al., 18
2007; NASIR e CAMPO, 2008). 19
A papilomatose mamária é uma doença infecto-contagiosa que compromete 20
animais de todas as idades, incluindo animais adultos. Porém os animais jovens, com até 21
os dois anos de idade, são os mais comprometidos. As lesões têm caráter auto-limitante 22
e tipicamente regridem entre 1-14 meses. Ocasionalmente, alguns tumores podem 23
persistir e progredir para carcinoma de células escamosas (SUNBERG, 1987; CAMPO, 24
1987). Segundo Campo (2002), fibropapilomas e papilomas de tetos e úberes podem ser 25
um grande problema econômico em bovinos uma vez que o papiloma pode atingir a 26
região do períneo e a parte inferior do corpo. Em alguns animais nos quais a regressão 27
espontânea dos papilomas não ocorre, a infecção pode generalizar pelo aparelho 28
mamário, inviabilizando a ordenha. 29
Existe grande variação nos aspectos relacionados à morfologia das lesões e à sua 30
progressão. Também são desconhecidas algumas características específicas da infecção 31
destacando-se entre elas: a correlação do tipo viral com o aspecto morfológico 32
macroscópico da lesão; e a persistência e possível progressão com generalização de 33
alguns casos. Macroscopicamente, a lesão se inicia com a proliferação de uma pequena 34

31
massa abaulada na pele do teto que, com o tempo, aumenta de diâmetro, torna-se áspera 1
e pedunculada até que o papiloma é formado (BORZACCHIELLO e ROPERTO, 2008) 2
Papilomas localizados no úbere e nos tetos de vacas em lactação dificultam a 3
amamentação dos bezerros e a ordenha tanto manual quanto, principalmente, mecânica. 4
Dentre os problemas mais frequentes ocasionados pelos papilomas destacam-se: i) 5
dificuldade do encaixe perfeito das teteiras da ordenhadeira mecânica ocasionada pela 6
irregularidade do tecido epitelial do teto ou distorção do ducto causando vazão do ar e 7
dificuldade na sucção do leite que comprometem a saúde do teto; ii) condenação de 8
novilhas ao abate antes mesmo da primeira lactação devido às extensões das lesões; iii) 9
predisposição à mastite devido à presença de leite residual ocasionada pela sensação 10
dolorosa que inibe a descida do leite; iv) presença de verrugas ao redor do canal do teto 11
que também podem predispor à mastite; v) predisposição aumentada para o 12
desenvolvimento de infecções bacterianas secundárias ocasionadas por lacerações das 13
verrugas presentes nos tetos (BLOWEY e EDMONDSON, 1995; BORZACCHIELLO 14
e ROPERTO, 2008). 15
A característica auto-limite da doença ocasiona negligência por parte dos 16
criadores no que tange ao controle da infecção. Com frequência, a papilomatose entra 17
no rebanho por meio da compra de animais infectados, aparentemente com poucos 18
papilomas, e de baixo valor no mercado, disseminando o vírus no rebanho. O fato dos 19
criadores ignorarem a presença de alguns animais infectados, mantendo-os juntamente 20
com animais sadios também predispõe a disseminação da infecção por meio de 21
equipamentos de ordenha, fômites, mãos dos ordenhadores, entre outras formas diretas e 22
indretas de transmissão do BPV. 23
Na criação de bovinos são reconhecidos vários fatores predisponentes que, 24
atuando de forma isolada ou preferencialmente em associação, reconhecidamente 25
ocasionam distúrbios hormonais que culminam com o desenvolvimento de 26
imunodepressão dos animais. Falhas na resposta imune são um dos principais fatores 27
responsáveis pela manutenção e, principalmente, disseminação da infecção intra-28
rebanho. Dentre as várias condições que ocasionam o desencadeamento de 29
imunodepressão destacam-se aquelas relacionadas ao manejo, nutrição e sanidade. 30
Falhas no manejo dos animais ocasionam condições inapropriadas de bem-estar animal 31
gerando situações de estresse prolongado e constante, tais como temperaturas extremas 32
como calor ou frio excessivos, superlotação, locais inapropriados de criação. Nutrição 33
inadequada, tanto nos aspectos quantitativos quanto qualitativos dos alimentos, diminui 34

32
o potencial de resposta imune dos animais. Infestações por endo e ectoparasitas 1
ocasionam distúrbios relacionados aos aspectos nutricionais e de bem-estar animal 2
contribuindo, consideravelmente, com a redução no potencial de resposta imune dos 3
animais. Ainda é possível citar alguns vírus responsáveis por efeitos imunosupressivos 4
em bovinos como herpesvírus bovino (BoHV-1), vírus da diarréia viral bovina (BVDV) 5
e vírus da leucemia bovina (BLV). 6
A eficiência de diferentes protocolos de tratamentos para papilomatose bovina já 7
foi testada, como a utilização da autovacina, auto-hemoterapia, diaminazina, 8
clorobutanol. Entretanto a eficiência dos resultados dos tratamentos é contraditória e 9
deficiente, levando à necessidade da repetição contínua dos mesmos. Por tratar-se de 10
uma enfermidade autolimitante e que pode variar com cada animal, a avaliação dos 11
tratamentos é relativa. Embora a porcentagem de “cura” demonstrada pelos tratamentos 12
seja bastante variada, a autovacina é o tratamento terapêutico que merece destaque. As 13
variadas taxas de sucesso obtidas com sua utilização podem ser decorrentes de alguns 14
aspectos relacionados à sua produção e características biológicas do BPV (SILVA et al., 15
2004). 16
Várias medidas, específicas e inespecíficas, podem ser implementadas 17
com o objetivo de controle da papilomatose bovina. Todas elas devem ser direcionadas 18
para a redução da circulação viral no rebanho ou para a não geração de situações que 19
proporcionem a manutenção e a disseminação do vírus intra-rebanho. Entre as medidas 20
destacam-se: isolamento dos animais infectados; tratamento dos animais; remoção 21
cirúrgica dos papilomas; emprego de auto-vacinas; anti-sepsia das mãos de 22
ordenhadores; desinfecção de equipamentos e fômites; cuidados especiais em processos 23
que causem ferimentos na pele tais como colocação de brincos e tatuagens; alteração da 24
ordem de ordenha com os animais infectados sendo manejados por último; criação em 25
ambientes adequados que proporcione bem-estar animal; nutrição adequada em termos 26
quantitativos e qualitativos; controle de endoparasitas; controle de carrapatos e moscas. 27
28
Resposta imune ao BPV 29
30
A imunidade para as infecções pelo BPV é considerada tipo-específica e o status 31
imunológico dos animais infectados é considerado fator crucial para a definição da 32
resolução clínica. Enquanto a imunidade humoral tem a habilidade de prevenir novas 33
infecções, a imunidade celular, provavelmente envolvendo linfócitos T, é responsável 34

33
pela regressão espontânea e imunomediada das lesões já estabelecidas (NICHOLLS e 1
STANLEY, 2000). As lesões benignas produzidas na infecção cutânea e mucosa pelo 2
PV apresentam tendência de regressão espontânea. No entanto, algumas infecções com 3
curso clínico prolongado e que determinam lesões extensas podem, ocasionalmente, 4
progredir para o câncer. Infecções pelo BPV, ocasionando lesões benignas podem ser 5
encontradas tanto em animais imunodeprimidos quanto em imunocompetentes. Casos 6
de papilomatose persistente geralmente são observados em animais imunodeprimidos 7
(ALFIERI et al., 2007). 8
O PV deve ser capaz de superar as resposta imune do hospedeiro 9
para se replicar, no entanto, apesar da superação do vírus, eventualmente, o 10
hospedeiro produz resposta imune eficaz e as células infectadas são eliminadas. 11
A resposta imune de bovinos para o BPV é surpreendentemente pobre. Os animais 12
podem apresentar tumores sólidos, produzindo ativamente vírus em grande quantidade, 13
porém não respondem facilmente aos antígenos BPV durante o curso da infecção e 14
anticorpos anti-BPV raramente são detectados. O fracasso do sistema imunológico para 15
reconhecer tanto a adsorção do vírus quanto a progênie viral é devido ao fato de que o 16
ciclo replicativo viral é restrito ao epitélio e, portanto, o vírus não entra em contato com 17
o sistema imune. Essa interpretação é apoiada pelo fato de que os animais a campo que 18
apresentam tumores ulcerados com hemorragia têm altos títulos de anticorpos anti-BPV. 19
Adicionalmente, a resposta humoral pode ser obtida após a inoculação intramuscular de 20
vírus purificado ou de proteínas virais, confirmando que só quando o papiloma está 21
lesionado, ou após a imunização, os antígenos virais entram em contato com as células 22
do sistema imunológico. Podem ser observadas respostas T e B fracas frente às 23
proteínas do capsídeo (L1 e L2) ou à proteínas transformantes (E7) em estádios finais da 24
infecção que parecem estar associadas com a rejeição do papiloma (NASIR e CAMPO, 25
2008). 26
Durante a rejeição dos papilomas, grandes massas de linfócitos ativados se 27
acumulam na derme subjacente. Provavelmente, a resposta imune ineficiente frente ao 28
BPV seja a principal razão para a persistência da infecção, uma vez que mesmo em 29
animais imunocompetentes, os papilomas persistem por muitos meses antes que ocorra 30
a regressão (NASIR e CAMPO, 2008). 31
Recentemente, estudos demonstraram que além do “escape imunológico” em 32
razão do ciclo viral confinado ao epitélio, os PVs podem desenvolver formas de evasão 33

34
do sistema imune do hospedeiro como a interrupção da expressão do MHC I pela 1
proteína E5 (O’BRIEN e CAMPO, 2002). 2
3
Imunoprofilaxia para o BPV 4
5
A expressão dos genes L1 e L2 em células eucarióticas, ou somente do gene L1, 6
resulta na auto-montagem (empacotamento) das proteínas em partículas semelhantes ao 7
vírus (VLP – virus-like particles), as quais são estruturalmente e antigenicamente 8
similares aos virions naturais (KIRNBAUER et al., 1996). O aproveitamento desta 9
propriedade biológica das proteínas estruturais dos PVs vem sendo empregado com 10
sucesso na elaboração de diversas vacinas profiláticas para diferentes espécies 11
hospedeiras, sendo que VLPs produzidas a partir destes genes tem se mostrado 12
altamente imunogênicas, conferindo proteção quando do desafio com tipo viral 13
(KIRNBAUER et al., 1996). Estudos demonstraram que a vacina elaborada com a 14
proteína L1 do BPV-2 promoveu proteção contra a infecção e que o uso da proteína L2 15
induziu mais rapidamente a regressão de lesões pré-formadas (JARRETT., et al 1991). 16
Em contrapartida, a vacina com a proteína L2 do BPV-4, proporcionou proteção total 17
contra o agente. Curiosamente os epítopos da L2 do BPV-4 são homólogos e têm reação 18
cruzada com epítopos identificados na proteína L2 de vários HPVs, levando a predizer 19
que vacinas baseadas nas proteínas L2 do HPV poderiam induzir reação imune cruzada 20
entre diferentes genotipos de HPVs (NASIR e CAMPO, 2008). 21
Vacinas terapêuticas (curativas), designadas para induzir a regressão das lesões 22
pré-malignas, têm sido desenvolvidas com base na proteína L2 do BPV-2, ou na 23
proteína E7 do BPV-4. Em ambos os casos, a regressão é acompanhada por infiltração 24
de linfócitos e macrófagos, que é indicativo de uma resposta imune mediada por células, 25
semelhante ao observado na regressão natural dos papilomas (NASIR e CAMPO, 26
2008). 27

35
Considerações finais 1
2
Enquanto numerosos tipos HPVs já foram definidos, até o início da década de 80 3
somente seis tipos de BPVs (BPV-1 a -6) haviam sido identificados a partir de casos de 4
papilomatose cutânea e de câncer em bovinos (PFISTER et al., 1979; CAMPO et al., 5
1980, 1981; CAMPO e COGGINS, 1982; CHEN et al., 1982; JARRETT et al., 1984). 6
Entretanto, estudos realizados a partir do início da década atual, com o objetivo de 7
investigar a diversidade do BPV, tem indicado a existência de numerosos tipos de BPV, 8
a exemplo do que ocorre nos seres humanos (ANTONSSON e HANSSON, 2002; 9
OGAWA et al., 2004). 10
O principal impedimento para a caracterização de novos PVs é a ausência de um 11
sistema convencional de cultivo celular para a propagação viral in vitro. Os métodos 12
recentes para a identificação de novos PVs são baseados em reações de PCR 13
empregando primers genéricos (degenerados). Estes primers são desenhados a partir de 14
sequências conservadas do gene que codifica a proteína L1 de tipos de PVs 15
anteriormente caracterizados. Essa estratégia tem sido utilizada com sucesso na 16
detecção de um amplo espectro de tipos virais já conhecidos e de novos tipos de HPV, 17
uma vez que a ORF L1 é a mais conservada no genoma dos PVs. Uma destas 18
metodologias, envolvendo a utilização do par de primers FAP (FAP59 e FAP64), foi 19
originalmente desenvolvida para a amplificação de HPVs cutâneos. Porém, essa 20
estratégia tem também possibilitado a detecção de novos tipos virais tanto em bovinos 21
quanto em outras espécies animais (FORSLUND et al., 1999; ANTONSSON e 22
HANSSON, 2002; OGAWA et al., 2004; ANTOSSON e McMILLAN, 2006; 23
LITERÁK et al., 2006; CLAUS et al., 2008a; CLAUS et al., 2008b; CLAUS et al., 24
2008c). 25
Recentemente, a partir da análise de papilomas cutâneos de bovinos no estado do 26
Paraná, utilizando a técnica de PCR com o par de primers FAP59/FAP64, foram 27
identificados os tipos de BPV já descritos anteriormente (BPV-1, 2, 6 e 8) e quatro 28
prováveis novos tipos de BPV que ainda não haviam sido descritos em todo o mundo 29
(CLAUS et al., 2007; 2008a; 2008b, 2008c). 30
No Brasil, o estudo mais abrangente envolvendo a epidemiologia molecular da 31
papilomatose cutânea, observada em diversos rebanhos bovinos do estado do Paraná, foi 32
realizado por meio do emprego da técnica da PCR com primers genéricos. O BPV-6 foi 33
o tipo viral mais prevalente em papilomas provenientes de tetos e/ou úberes (CLAUS et 34

36
al., 2007). Curiosamente, achado similar foi encontrado em estudo realizado no Japão, 1
que teve como objetivo a identificação dos tipos de BPV mais prevalentes em um surto 2
de papilomatose mamária envolvendo 560 novilhas (MAEDA et al., 2007). 3
Dentre os 11 tipos de BPVs até então caracterizados, tem-se o relato da 4
ocorrência de oito tipos virais em lesões cutâneas dos tetos e úberes de animais 5
acometidos por papilomatose cutânea (NASIR e CAMPO, 2008). Porém, a realização 6
de investigações epidemiológicas em diversas regiões geográficas, as quais venham 7
elucidar quais os tipos mais prevalentes em lesões da glândula mamária de bovinos, 8
ainda podem ser consideradas escassas em todo o mundo. 9
A imunidade para as infecções pelo BPV é considerada tipo-específica e o status 10
imunológico dos animais infectados é considerado fator crucial para definir a resolução 11
clínica. Enquanto a imunidade humoral tem a habilidade de prevenir novas infecções, a 12
imunidade celular é responsável pela regressão espontânea e imunomediada das lesões 13
já estabelecidas (NICHOLLS e STANLEY, 2000). 14
Em bovinos, vacinas preventivas também tem sido desenvolvidas contra os 15
BPVs tipos 2 e 4, sendo que estes dois tipos virais foram, respectivamente, 16
selecionados como representativos para os papilomavírus cutâneos e mucosos, e 17
também porque ambos os tipos estão relacionados com a determinação de câncer em 18
bovinos (CAMPO et al., 1993). 19
O desenvolvimento de vacinas e estratégias imuno-profiláticas contra a 20
papilomatose mamária, requer previamente a identificação dos tipos de BPV mais 21
prevalentes na neste sítio anatômico. O controle e profilaxia de uma doença 22
negligenciada como a papilomatose pode trazer reflexos diretos na produção leiteira da 23
grande maioria dos rebanhos nacionais. 24
.

37
Referências
1
ALFIERI, A.A. Gestão profilática das principais doenças infecto-contagiosas em 2
bovinos leiteiros. In: SANTOS, G.T.; UHLIG, L.; BRANCO, A.F.; JOBIM, C.C.; 3
DAMASCENO, J.C.; CECATO, U. (Edts.) Bovinocultura de Leite. Maringá: 4
UEM, 2008. p.131-178. 5
ALFIERI, A.A; WOSIAKI, S.; ALFIERI, A.F. Papillomavirus. In: FLORES, F. E. 6
Virologia Veterinária. Santa Maria: UFSM, 2007. p.399-412. 7
ANTONSSON, A.; HANSSON, B.G. Healthy skin of many species harbors 8
papilomaviruses which are closely related to their human counterparts. Journal of 9
Virology, v.76, n.24, p.12537-12542, 2002. 10
ANTONSSON, A.; MC MILLAN, N.A. Papillomavirus in healthy skin of Australian 11
animals. Journal of General Virology, v.87, p.3195-3200, 2006. 12
ANTONSSON, A.; FORSLUND, O.; EKBERG, H.; STERNER, G.; HANSSON, B.G. 13
The ubiquity and impressive genomic diversity of human skin papillomaviruses 14
suggest a commensalic nature of these viruses. Journal of Virology, v.74, 2000. 15
BERNARD, H.U. The clinical importance of the nomenclature, evolution and taxonomy 16
of human papillomaviruses. Journal of Clinical Virology, v.32, p.1-6, 2005. 17
BERNARD, H.U. Phylogeny and typing of papillomaviruses. In: GARCEA, L. R. The 18
Papillomavirus. Nova York: Springer Science, 2007. p.69-83. 19
BLOCH, N.; BREEN, M.; IRVIN, Z.V.; SPRADBROW, P.B. Bovine papillomavirus 20
type 4 DNA isolated from a skin lesion in a steer. The Veterinary Record, v.136, 21
p.414-416, 1996. 22
BLOWEY, R.; EDMONDSON, P. Mastitis Control in Dairy Herds. Ipswich United 23
Kingdom: Farming Press, 1995. 196 p. 24
BORZACCHIELLO, G.; ROPERTO, F. Bovine papillomaviruses, papillomas and 25
cancer in cattle. Veterinary Research, v.39, n.5, p.45-64, 2008. 26
BORZACCHIELLO, G.; IOVANE, G.; MARCANTE, M.L.; POGGIALI, F.; 27
ROPERTO, F.; ROPERTO, S.; VENUTI, A. Presence of bovine papillomavirus 28
type 2 DNA and expression of the viral oncoprotein E5 in naturally occuring urinary 29
bladder tumors in cows. Journal of General Virology, v.84, n.11, p.2921-2926, 30
2003. 31
CAMPO, M.S. Papillomas and cancer in cattle. Cancer Surveys, v.06. p. 39-54, 1987. 32

38
CAMPO, M.S. Infection by bovine papillomavirus and prospects for vaccination. 1
Trends in Microbiology, v.3, p.92-97, 1995. 2
CAMPO, M.S. Animal models of papillomavirus pathogenesis. Virus Research, v.89, 3
p.249-261, 2002. 4
CAMPO, M.S.; COGGINS, L.W. Molecular cloning of bovine papillomavirus genomes 5
and comparison of their sequence homologies by heteroduplex mapping. Journal of 6
General Virology, v.63, p.255-264, 1982. 7
CAMPO, M.S.; JARRET, W.F.H.; BARRON, R.; O’NEILL, B.W.; SMITH, K.T. 8
Association of bovine papillomavirus type 2 and bracken fern with bladder cancer in 9
cattle. Cancer Research, v.52, n.24, p.6898-6904, 1992. 10
CAMPO, M.S.; JARRETT, W.F.H.; GRINDLAY, G.J.; CHANDRACHUD, L.M.; 11
MCGARVIE, G.M.; O’NEIL, B.W. Prophylactic and therapeutic vaccination 12
against a mucosal papillomavirus. Journal of General Virology, v.74, p.945-953, 13
1993. 14
CAMPO, M.S.; MOAR, M.H.; JARRETT, W.F.H.; LAIRD, H.M. A new 15
papillomavirus associated with alimentary cancer in cattle. Nature, v.286, p.180-16
182, 1980. 17
CAMPO, M.S.; MOAR, M.H.; LAIRD, H.M.; JARRETT, W.F. Molecular 18
heterogeneity and lesion site specificity of cutaneous bovine papillomaviruses. 19
Virology, v.113, p.323-35, 1981. 20
CHAN, S.Y.; DELIUS, H.; HALPERN, A.L.; BERNARD, H-U. Analysis of genomic 21
sequences of 95 papillomavirus types: uniting typing, phylogeny, and taxonomy. 22
Journal of Virology, v.69, n.5, p.3074-3083, 1995. 23
CHEN, E.Y.; HOWLEY, P.M.; LEVINSON, A.D.; SEEBURG, P.H. The primary 24
structure and genetic organization of the bovine papillomavirus type 1 genome. 25
Nature, v.299, p.529-534, 1982. 26
CLAUS, M.P. Identificação e estudo da diversidade do papilomavírus associado a 27
lesões cutâneas em bovinos. 2008. p. 139. Tese (Doutorado em Saúde Animal) 28
Universidade Estadual de Londrina. 2008c. Disponível em 29
<http://www.bibliotecadigital.uel.br/document/?code=vtls000129203>. Acesso em: 30
13/12/2010. 31
CLAUS, M.P.; LUNARDI, M.; ALFIERI, A.F.; FERRACIN, L.M.; FUNGARO, 32
M.H.P.; ALFIERI, A.A. Identification of unreported putative new bovine 33

39
papillomavirus types in Brazilian cattle herds. Veterinary Microbiology, v.132, 1
n.3-4, p.396-401, 2008a. 2
CLAUS, M.P.; LUNARDI, M.; ALFIERI, A.F.; SARTORI, D.; L.M. FUNGARO, 3
M.H.P; ALFIERI, A.A. Bovine papillomavirus type 8: the new type of BPV, 4
recently described in Asia and Europe, is also present in South America. Pesquisa 5
Veterinária Brasileira, v. 29, p. 25-28, 2008b. 6
CLAUS, M.P.; VIVIAN, D.; LUNARDI, M.; ALFIERI, A.F.; ALFIERI, A.A. 7
Phylogenetic analysis of bovine papillomavirus associated with skin warts in cattle 8
herds from the state of Paraná. Pesquisa Veterinária Brasileira, v.27, p.314-318, 9
2007. 10
CULLEN, A.P.; REID, R.; CAMPION, M.; LÖRINCZ, A.T. Analysis of the physical 11
state of different human papillomavirus DNAs in intraepithelial and invasive 12
cervical neoplasm. Journal of Virology, v.65, n.2, p.606-612, 1991. 13
de VILLIERS, E.M.; FAUQUET, C.; BROKER, T.R.; BERNARD, H.U.; ZUR 14
HAUSEN, H. Classification of papillomaviruses. Virology, v.324, p.17-27, 2004. 15
DOORBAR, J. The papillomavirus life cycle. Jounal of Clinical Virology, v.32, p.7-16
15, 2005. 17
FORSLUND, O.; ANTONSSON, A.; NORDIN, P.; HANSSON, B.G. A broad range of 18
human papillomavirus types detected with a general PCR method suitable for 19
analysis of cutaneous tumours and normal skin. Journal of General Virology, v.80, 20
p.2437-2443, 1999. 21
GARCEA, L.R.; CHEN, X. Papillomavirus Structure and Assembly. In: GARCEA, L. 22
R. The Papillomavirus. Nova York: Springer Science, 2007. p.69-83. 23
GUPTA, M.P.; GUPTA, P. P.; RATHORE, S.S.; GUPTA, A.K. Bovine cutaneous 24
papillomatosis - a case report. Indian Veterinary Jounal, v.66, p.358-359, 1989. 25
HATAMA, S., NOBUMOTO, K., KANNO, T. Genomic and phylogenetic analysis of 26
two novel bovine papillomavirus, BPV-9 e BPV-10. Journal of General Virology, 27
v.89, p.158-163, 2008. 28
HEAD, K.W.; ELSE, R.W.; DUBIELZIG, R.R. Tumours of the alimentary tract. In: 29
MEUTEN, D.J. Tumours of domestic animals. 4ed. Iowa: Ames, 2002. p.401-481. 30
HOPKINS, N.C.G. Aetiology of enzootica haematuria. The Veterinary Record, v.118, 31
n.26, p.715-717, 1986. 32
HOWLEY, P.M.; LOWY, D.R. Papillomaviruses and their replication. In: Fields 33
Virology, 4ed. Philadelphia, PA: Lippincott-Raven, 2001. p. 2197-2229. 34

40
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA, 2010. Abate de 1
animais, produção de leite, couro e ovos. Disponível em 2
<http://www.ibge.gov.br/home/estatistica/indicadores/agropecuaria/producaoagrope3
cuaria/abate-leite-couro-ovos_201003_2.shtm>. Acesso em 03/01/2010. 4
JACKSON, M.E.; PENNIE, W.D.; McCAFFERY, R.E.; SMITH, K.T.; GRINDLAY, 5
G.J.; CAMPO, M.S. The B subgroup bovine papillomavirus lack an identifiable E6 6
open reading frame. Molecular Carcinogenic, v.4, n.5, p.382-387, 1991. 7
JARRETT, W.F.H.; CAMPO, M.S.; O’NEIL, B.W.; LAIRD, H.M; COGGINS, L.W. A 8
novel bovine papillomavirus (BPV-6) causing true epithelial papillomas of the 9
mammary gland skin: a member of a proposed new BPV subgroup. Virology, v.136, 10
p.255-264, 1984. 11
JARRETT, W.F.; SMITH, K.T.; O’NEIL, B.W.; GAUKROGER, J.M.; 12
CHANDRACHUD, L.M.; GRINDLAY, G.J.; McGARVIE, G.M.; CAMPO, M.S. 13
Studies on vaccination against papillomaviruses: prophylatic and therapeutic 14
vaccination with recombinant structural proteins. Virology, v.184, n.1, p.33-42, 15
1991. 16
JELÍNEK, F. and TACHEZY, R. Cutaneous papillomatosis in cattle. Jounal of 17
Comparative Pathology, v.132, p.70-81, 2005. 18
KIRNBAUER, R.; CHANDRACHUD, L.M.; O'NEIL, B.W.; WAGNER, E.R.; 19
GRINDLAY, G.J.; ARMSTRONG, A.; MCGARVIE, G.M.; SCHILLER, J.T.; 20
LOWY, D.R.; CAMPO, M.S. Virus-like particles of bovine papillomavirus type 4 in 21
prophylactic and therapeutic immunization. Virology, v.219, n.1, p.37-44, 1996. 22
LANCASTER, W.D.; OLSON, C. Animal papillomaviruses. Microbiology Research, 23
v.46, n.2, p.191-207, 1982. 24
LEE, C.; LAIMINS, L.A. The life cycle of human papillomaviruses in keratinocytes. In: 25
GARCEA, L. R. The Papillomavirus. Nova York: Springer Science, 2007. p.45-67. 26
LINDHOLM, L.; MURPHY, J.; O’NEIL, B.W; CAMPO, M.S.; JARRETT, W.F.H. 27
Papillomas of the teats and udder of cattle and their causal viruses. Veterinary 28
Record, v.115, n.22, p. 574-577, 1984., 29
LITERÁK, I.; TOMITA, Y.; OGAWA, T.; SHIRASAWA, H.; SMÍD, B.; NOVOTNÝ, 30
L.; ADAMEC, M. Papillomatosis in a European bison. Journal of Wildlife 31
Disease, v.42, p.149-153, 2006. 32
MAEDA, Y.; SHIBAHARA, T.; WADA, Y.; KADOTA, K.; KANNO, T.; UCHIDA, 33
I.; HATAMA, S. An outbreak of teat papillomatosis in cattle caused by bovine 34

41
papillomavirus (BPV) type 6 and unclassified BPVs. Veterinary Microbiology, 1
v.121, p.242-248, 2007. 2
MCGALVIN, M.D.; ZACHARI, J.F. Pathologic Basis of Veterinary Disease. 4ed. St 3
Louis: Mosby, 2007. p.301-461. 4
NASIR, L.; CAMPO, M.S. Bovine papillomaviruses: their role in the aetiology of 5
cutaneous tumours of bovids and equids. Veterinary Dermatology, v.19, p.243-6
254, 2008. 7
NICHOLLS, P.K.; STANLEY, M.A. The immunology of animal papillomaviruses. 8
Veterinary Immunology and Immunopathology, v.73, p.101-127, 2000. 9
O’BRIEN, P.M.; CAMPO, M.S. Evasion of host immunity directed by papillomavirus-10
encoded proteins. Virus Reseach, v.88, p.103-117, 2002. 11
OGAWA, T.; OKADA, M.; SHIRASAWA, H. Complete genomic and phylogenetic 12
position of bovine papillomavius type 7. Jounal of General Virology, v.88, p.1934-13
1938, 2007. 14
OGAWA, T.; TOMITA, Y.; OKADA, M.; SHINOZAKI, H.K.; KAIHO, I.; 15
SHIRASAWA, H. Broad-spectrum detection of papillomaviruses in bovine teat 16
papilomas and health teat skin. Jounal of General Virology, v.85, p.2191-2197, 17
2004. 18
PFISTER, H.; LINZ, U.; GISSMAN, L.; HUCHTHAUSEN, B.; HOFFMAN, D.; ZUR 19
HAUSEN, H. Partial characterization of a new type of bovine papilloma viruses. 20
Virology, v.96, p.1-8, 1979. 21
ROCHA, A.M. Futuro promissor para exportação de leite. Valor Econômico, 2007. 22
Disponível em: 23
<http://www.americalatina.org.br/internas.php?noticias=&interna=22512>. Acesso 24
em 12/01/2011. 25
SANTIN, A.P.I.; BRITO, L.A.B. Caracterização anatomopatológica da papilomatose 26
cutânea em bovinos leiteiros. Revista Brasileira de Ciência Veterinária, v.10, n.3, 27
p.161-165, 2003. 28
SILVA, L.A.F.; VERÍSSIMO, A.C.C.; VIANA FILHO, P.R.L.; FIORANTI, M.S.C.; 29
EURIDES, D.; LINHARES, G.C.F.; ROMANI, A.F.; TRINDADE, B.R. Eficiência 30
da repetição de diferentes protocolos de tratamentos para papilomatose bovina. 31
Revista da Faculdade de Zootecnia, Veterinária e Agronomia, Uruguaiana, 32
v.11, n.1, p.153-165, 2004. 33

42
SMITH, H.A.; JONES, T.C.; HUNT, R.D. Veterinary pathology, v.4, p.508-515, 1
1995. 2
SUNDBERG, J.P. Papillomavirus infections in animals. In: Papillomaviruses and 3
Human disease. Berlin: Springer, 1987. p. 40-103. 4
TERAI, M.; DeSALLE, R.; BURK, R.D. Lack of canonical E6 and E7 open reading 5
frames in bird papillomaviruses: Fringilla coelebs papillomavirus and Psittacus 6
erithacus timneh papillomavirus. Journal of Virology, v.76, n.19, p.10020-10023, 7
2002. 8
TOMITA, Y.; LITERÁK, I.; OGAWA, T.; JIN, Z.; SHIRASAWA, H. Complete 9
genomes and philogenetic positions of bovine papilomavirus type 8 and variant type 10
from European bison. Virus Genes, v.35, p. 243-249, 2007. 11
WOSIACKI, S.R.; BARREIROS, M.A.B.; ALFIERI, A.F.; ALFIERI, A.A. Semi-12
nested-PCR for detection and typing of bovine papillomavirus type 2 in urinary 13
bladder and whole blood from cattle with enzootic haematuria. Journal of 14
Virological Methods, v.126, n.1-2, p.215-219, 2005. 15
43
Ilustrações 1
2
Figura 1. Desenho esquemático do genoma do papilomavírus bovino tipo 1. Fonte: 3 Alfieri et al., (2007). Modificado de Frazer (2004). 4
44
1
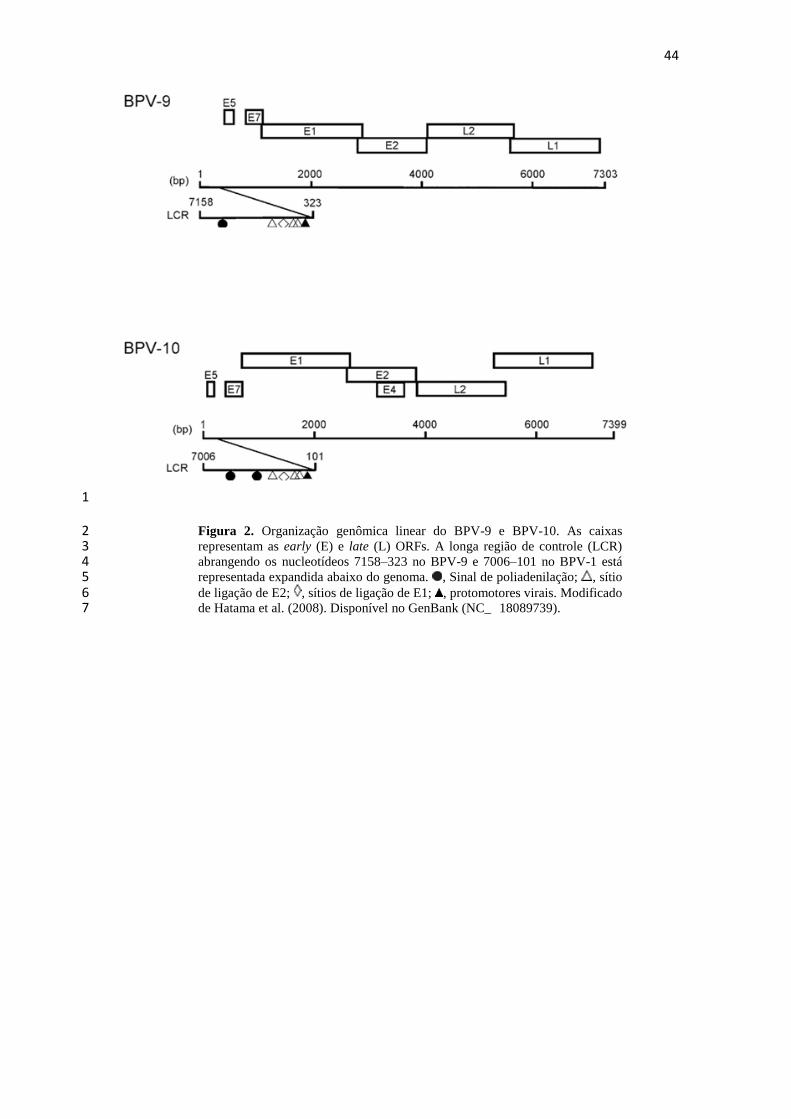
Figura 2. Organização genômica linear do BPV-9 e BPV-10. As caixas 2 representam as early (E) e late (L) ORFs. A longa região de controle (LCR) 3 abrangendo os nucleotídeos 7158–323 no BPV-9 e 7006–101 no BPV-1 está 4 representada expandida abaixo do genoma. , Sinal de poliadenilação; , sítio 5 de ligação de E2; , sítios de ligação de E1; , protomotores virais. Modificado 6 de Hatama et al. (2008). Disponível no GenBank (NC_ 18089739). 7

45
Quadro 1. Funções das proteínas codificadas pelo papilomavírus bovino tipo 1 1
2 Fonte: Alfieri et al. (2007) 3
4
Quadro 2. Primers utilizados na reação em cadeia da polimerase para a identificação e análise 5 filogenética do papilomavírus bovino associado a lesões cutâneas. 6 7 Primer
a Sequência
b nt HPV-8
FAP59 5’TAA CWG TIG GIC AYC CWT ATT 3’ 5981-6001
FAP64 5’CCW ATA TCW VHC ATI TCI CCA TC 3’ 6458-6436
a Fourslund et al. (1999) 8
b Nucleotídeos degenerados: W=T, C; I=Inosina; Y=C, T; V=A, C, G; H=A, T, C. 9

46
3. OBJETIVOS 1
2
Objetivo Geral: 3
4
Identificar os tipos de BPV relacionados com o desenvolvimento de 5
papilomatose mamária em rebanhos bovinos leiteiros brasileiros. 6
7
8
Objetivos Específicos: 9
10
Detectar a presença dos BPVs envolvidos nas lesões cutâneas do aparelho 11
mamário de bovinos leiteiros pertencentes às regiões brasileiras alvo do estudo 12
empregando os primers genéricos (FAP) desenhados a partir da ORF L1; 13
14
Sequenciar os amplicons-FAP e realizar as análises filogenéticas por meio da 15
comparação das sequências obtidas com as disponíveis em bases públicas de dados; 16
17
Verificar as diferentes alterações histopatológicas e correlacionar o tipo de lesão 18
ao genótipo do BPV. 19

47
4. ARTIGOS PARA PUBLICAÇÃO 1

48
4.1 TEAT PAPILLOMATOSIS ASSOCIATED WITH RECENT 1
CHARACTERIZED BPV TYPES (BPV-7, -9 AND -10) IN BRAZILIAN 2
CATTLE* 3
4
Abstract 5 6
Teat papillomatosis has been reported in dairy herds worldwide as a cattle health 7 problem responsible for economic losses. Despite the high number of cattle herds 8
affected by cutaneous papillomatosis, studies evaluating BPV diversity remain sporadic. 9 The aim of the current report was to genotype the BPV types associated with teat 10 papillomas obtained from Brazilian cattle herds. Fifteen teat papilloma lesions 11
underwent papillomavirus DNA detection via the FAP PCR assay and were processed 12 for histopathological evaluation. BPV-6, -7, -9, and -10 were identified in ten (66.7%), 13 two (13.3%), one (6.7%), and two (13.3%) papilloma specimens, respectively. All 14 specimens analyzed were classified as true papillomas. This finding demonstrates the 15
great diversity in BPV types that occurs in benign teat lesions. This study represents the 16 first report of the identification of the recently characterized BPV types 7, 9, and 10 17 associated with cases of teat papillomatosis in cattle from the South American 18 continent. 19
20 Keywords: Bovine papillomavirus; cutaneous papillomatosis; papilloma; warts; 21
histopathology. 22
* Artigo editado de acordo com as normas de publicação do periódico Veterinary Microbiology,
disponível em: http://www.elsevier.com/wps/find/journaldescription.cws_home/503320/authorinstructions

49
1. Introduction 1
2
Papillomaviruses are able to induce cutaneous warts in the skin and mucosal 3
epithelia of most higher vertebrate species examined. Some specific viral types have the 4
potential to cause malignant progression in papillomatous lesions of animals and 5
humans (Antonsson, Hansson, 2002, de Villiers et al., 2004). In cattle, bovine 6
papillomaviruses (BPV) are known as the etiological agents of distressing diseases such 7
as cutaneous and teat papillomatosis, papillomatosis and cancers of the upper 8
gastrointestinal tract and urinary bladder (Campo, 2002, Borzacchiello et al., 2003a, 9
2003b, Wosiacki et al., 2006). 10
Teat papillomatosis has been reported in dairy herds worldwide as a cattle health 11
problem responsible for economic losses, as milking can become difficult in markedly 12
affected individuals. Ulceration and rupture of established cutaneous lesions may 13
predispose dairy cattle to mastitis and distortion of the milk ducts. Additionally, the 14
maintenance of affected heifers with alteration in mammary gland shape and/or even of 15
herds with a high number of affected animals may prevent economic profits in the dairy 16
industry (Campo, 2002, Borzacchiello, Roperto, 2008). 17
Currently, the genomes of ten BPVs (BPV-1 to -10) have been fully 18
characterized. They are classified in four genera based on their biological properties and 19
genome relatedness. These genera are Deltapapillomavirus (BPV-1 and -2), 20
Xipapillomavirus (BPV-3, -4, -6, -9, and -10), Epsilonpapillomavirus (BPV-5 and -8), 21
and a yet unnamed PV genus (BPV-7) (Pfister et al., 1979, Campo et al., 1980, 1981, 22
Campo, Coggins, 1982, Chen et al., 1982, Jarrett et al., 1984, de Villiers, 2004, Ogawa 23
et al., 2007, Tomita et al., 2007, Hatama et al., 2008). Moreover, the occurrence of 24
numerous additional viral types has been proposed based on partial nucleotide sequence 25

50
analysis of the major capsid protein L1, obtained both from benign cutaneous lesions 1
and swab samples of healthy skin from cattle (Antonsson, Hansson, 2002, Ogawa et al., 2
2004, Maeda et al., 2007, Claus et al., 2008, 2009a). Papilloma lesions found in teats 3
and udders can be caused by diverse BPV types. Although these warts can be caused by 4
several viral types, BPV-6 has often been identified in this anatomical location. 5
However, the association of BPV-1, -3, -5, -7, -8, -9, -10, and other types still to be 6
characterized, have indeed been confirmed in mammary glands of cattle (Campo et al., 7
1981, Jarrett et al., 1984, Ogawa et al., 2004, Maeda et al., 2007, Claus et al., 2007, 8
2008). 9
Despite the high number of cattle herds affected by cutaneous papillomatosis in 10
several geographical regions in Brazil, studies evaluating BPV diversity associated with 11
different clinical outcomes remain sporadic (Santos et al., 1998, Freitas et al., 2003, 12
Wosiacki et al., 2005, 2006, Claus et al., 2007, 2009a, 2009b). 13
The aim of the current report was to genotype the BPV types associated with 14
papilloma lesions obtained from cattle herds with teat papillomatosis. To our 15
knowledge, this study represents the first attempt to investigate the BPV diversity 16
related to cases of teat papillomatosis in cattle from Brazil. 17
18
2. Material and Methods 19
20
2.1. Papilloma specimens 21
22
Fifteen teat papilloma lesions were individually collected from dairy cows 23
belonging to two different cattle herds from the northern region of Parana state in 24
southern Brazil. Fragments from each papilloma specimen were ground in phosphate-25

51
buffered saline solution (PBS pH 7.2), and suspensions (10–20%, w/v) were centrifuged 1
for 15 min at 3000 g at 4°C. Aliquots (250 l) from supernatant were treated with 2
lysis buffer [10 mM Tris; 1 mM EDTA; 0.5% Nonidet P40; 1% SDS; and 0.2 mg/ml 3
proteinase K (Invitrogen, Life Technologies, Carlsbad, USA)]. After homogenization, 4
the samples were incubated at 56°C for 30 min. 5
6
2.2. DNA extraction 7
8
For DNA extraction, a combination of phenol/chloroform/isoamyl alcohol and 9
silica/guanidine isothiocyanate methods was performed according to Alfieri et al. 10
(2006). Briefly, fractions from each sample were treated with an equal volume of 11
phenol/ chloroform/isoamyl alcohol (25:24:1), homogenized, and heated at 56°C for 15 12
min (Sambrook et al., 1989). After centrifugation at 10,000 g for 10 min, the aqueous 13
phase was processed according to the silica/guanidine isothiocyanate method (Boom et 14
al., 1990). DNA was eluted in 50 µl of ultrapure (MilliQ®) sterile water and kept at 15
20°C until use. Aliquots of ultrapure sterile water were included as a negative control 16
in the DNA extraction procedures. 17
18
2.3. PCR assay 19
20
The PCR assay was carried out using the primer pair FAP59 (forward: 5’-21
TAACWGTIGGICAYCCWTATT- 3’) and FAP64 (reverse: 5’-22
CCWATATCWVHCATITCICCATC- 3’) according to Forslund et al. (1999), with 23
slight modifications (Claus et al., 2007). The reaction was performed using 5 µl of the 24
extracted DNA and 45 µl of PCR mix consisting of 1 µl (20 pmol) of each primer, 200 25

52
µM of each dNTP (Invitrogen, Life Technologies, Carlsbad, USA), 2.5 units of 1
Platinum Taq DNA polymerase (Invitrogen, Life Technologies, BR), 1 PCR buffer (20 2
mM Tris– HCl pH 8.4 and 50 mM KCl), 1.5 mM MgCl2 and ultrapure sterile water, 3
resulting in a final volume of 50 µl. Amplification was performed in a thermocycler 4
(PTC 200, MJ Research Co., USA) with the following cycling profile: an initial step of 5
10 min at 94°C, followed by 40 cycles of 1 min at 94°C, 1 min at 50°C, 1 min at 72°C, 6
and a final extension step of 10 min at 72°C. Aliquots of 5 µl from the PCR products 7
were analyzed by electrophoresis in a 2% agarose gel in TBE buffer, pH 8.4 (89 mM 8
Tris; 89 mM boric acid; 2 mM EDTA) at constant voltage (90 V) for approximately 45 9
min, stained with ethidium bromide (0.5 µg/ml), and visualized under UV light. 10
11
2.4. Sequence analysis 12
13
Initially, all PCR products were purified using Illustra GFX PCR DNA and the 14
Gel Band Purification kit (GE Healthcare, Little Chalfont, UK). Direct sequencing was 15
then performed using the DYEnamic ET dye terminator cycle sequencing kit (GE 16
Healthcare, Little Chalfont, UK) with FAP59 and FAP64 primers, in a MegaBACE 17
1000/Automated 96 Capillary DNA Sequencer (GE Healthcare, Little Chalfont, UK) 18
according to the manufacturer’s instructions. The obtained sequences were examined 19
with the PHRED software for quality analysis of chromatogram readings. The 20
sequences were accepted if the base quality was equal to or higher than 20. Consensus 21
sequences were determined using CAP3 software, and the sequence identity was 22
verified with all sequences deposited in the GenBank using the BLAST software. The 23
guidelines from the Papillomavirus Nomenclature Committee 1995 (14th International 24

53
Papillomavirus Conference, Quebec City, Quebec, Canada) were followed to identify 1
PV types (de Villiers et al., 2004). 2
3
2.5. Histopathology 4
5
Teat papilloma specimens were fixed in 10% neutral buffered formalin solution, 6
embedded in paraffin and routinely processed for histopathological evaluation. Sections 7
(6 m) were stained with hematoxylin and eosin (HE). 8
9
3. Results 10
11
Fifteen DNA samples extracted from teat papilloma lesions underwent PV DNA 12
detection via the FAP PCR assay, resulting in amplicons of approximately 480 bp in 13
length. The negative control for PCR amplification yielded no amplified product. 14
From the 15 teat cutaneous lesions that were amplified by FAP PCR assay, 15
BPV-6, -7, -9, and -10 were identified in ten (66.7%), two (13.3%), one (6.7%), and two 16
(13.3%) papilloma specimens, respectively. BPV-6 was the most frequent viral type 17
detected in evaluated samples, and this BPV type was present in both investigated 18
herds. Two (F and H) out of five cows with more than one teat papilloma lesion 19
evaluated presented double infection by different BPV types (BPV-7 and -10, and BPV-20
6 and -9, respectively). The different strains identified in this study were designated as 21
BPV6/BR-UEL, BPV7/BR-UEL, BPV9/BR-UEL and BPV10/BR-UEL (GenBank 22
accession numbers: HM245430, HM245431, HM245433 and HM245432, respectively) 23
(Table 1). 24

54
In our analysis, we did not observe any association between the macroscopic 1
characteristics and the viral type harbored in the cutaneous teat lesions. BPV-6 was 2
detected in teat warts grossly presenting frond, cauliflower, filiform and flat-and-round 3
aspects, and BPV-7 could be identified in both filiform and cauliflower lesions. 4
The histopathological features of teat cutaneous lesions showed keratinocytes 5
with viral cytopathic effect. Epidermal hyperplasia differentiation with orthokeratosis or 6
parakeratosis, enlarged keratohyaline granules and the presence of koilocytes were 7
observed in the evaluated histological samples. All 15 specimens analyzed were 8
classified as true papillomas (Figure 1). 9
The gross characteristics of cutaneous lesions as well as the type classification 10
and GenBank accession numbers for the obtained amplicons are indicated in Table 1. 11
12
4. Discussion 13
14
This study involved the analysis of lesions exclusively present in the mammary 15
gland of dairy cows of two Brazilian herds. We detected BPV types 7, 9, and 10, in 16
addition to BPV type 6, as causative agents of warts in the dairy cow teat. This finding 17
demonstrates the great diversity in BPV types that cause this condition. Although BPV-18
6 is the viral type that is classically associated with teat papilloma worldwide, our 19
findings of diverse BPV types are in agreement with the results obtained by other 20
studies investigating viral diversity in Japanese cattle herds affected by teat 21
papillomatosis. Such investigations have confirmed the high BPV diversity that occurs 22
both in benign teat lesions and healthy teat skin, in a commensal form (Campo, 2002, 23
Ogawa et al., 2004, Maeda et al., 2007). 24

55
Even though high viral diversity (BPV-6, -7, -9, and -10) was observed in our 1
study, BPV-6 was the viral type present in most affected individuals. 2
To the authors’ knowledge, this study represents the first report of the 3
identification of the recent characterized BPV types 7, 9 and 10 associated with cases of 4
teat papillomatosis in cattle from the South American continent. The identification of 5
these BPV types in a geographical region not related to the locations where they were 6
first isolated, Japan (BPV-7 and -9) and Sweden (BPV-10), supports the hypothesis that 7
these viral types are widespread and affect herds worldwide (Ogawa et al., 2007, 8
Hatama et al., 2008). 9
Previous studies involving the identification of BPV types 9 and 10 have 10
documented that these viral types are easily isolated from naturally occurring teat 11
papilloma lesions but not from healthy teat skin, highlighting the need for further 12
investigations to confirm their specific association with the etiology of teat 13
papillomatosis (Ogawa et al., 2004, Maeda et al., 2007, Hatama et al., 2008). More 14
recently, the tumorigenic potential of BPV-9 was tested through experimental infection 15
of the teat skin of heifers, resulting in the development of papillomatosis (Hatama et al., 16
2009). Conversely, BPV-7 has been detected mainly from swab samples of healthy teat 17
skin, and has been identified in only a few teat lesions from dairy cattle in Japan 18
(Ogawa et al., 2004). 19
The current taxonomic classification system for the Papillomaviridae family has 20
recently proposed the introduction of the taxonomic terms “species” and “genus” for 21
this viral family, establishing the relationship between phylogenetic assemblages and 22
biological and pathological properties shared by viruses grouped together (de Villiers et 23
al., 2004). The three Xipapillomavirus representatives (BPV-6, -9, and -10) identified in 24
our study were determined to be true papillomas in teat skin. This histological feature is 25

56
in agreement with the biological properties described for this specific genus (de Villiers 1
et al., 2004). Regarding the histological characteristics of BPV-7-induced cutaneous 2
lesions, this viral type gave rise to true epithelial papilloma at the same specific site of 3
the body. The histological characteristics of cutaneous lesions caused by BPV-7 were 4
missing in the work that established its phylogenetic position and characterized its 5
genome. Therefore, we now suggest that future isolates that are to be grouped with 6
BPV-7 in its yet unnamed genus should also be able to induce true papillomas in the 7
skin epithelia of cattle. 8
The limited number of recent studies seeking to identify BPV types associated 9
with lesions of the mammary gland of cattle have reported a considerable diversity of 10
the viral types involved, demonstrating that the viral type that has been classically 11
associated with this anatomical site (BPV-6) is not the only causive of these lesions in 12
affected herds. Therefore, the necessity for epidemiological investigations in different 13
geographical areas is clear, as such studies would facilitate the elaboration of future 14
immunogens while identifying the most prevalent type detected in each region. It is 15
noteworthy that there are no commercially available alternatives for the effective 16
treatment of or prevention against teat papillomatosis. 17
18
5. Acknowledgements 19
20
We thank Brazilian Institutes CNPq, CAPES, FINEP, and Fundação Araucaria 21
(FAP/PR) for financial support. Alfieri, A.A. and Alfieri, A.F. are recipient of CNPq 22
fellowship. 23

57
6. References 1
2
Alfieri, A.A., Parazzi, M.E., Takiuchi, E., Médici, K.C., Alfieri, A.F., 2006. Frequency 3
of group A rotavirus in diarrhoeic calves in Brazilian cattle herds, 1998–2002. Trop. 4
Anim. Health Prod. 38, 521–526. 5
Antonsson, A., Hansson, B.G., 2002. Healthy skin of many animal species harbors 6
papillomaviruses which are closely related to their human counterparts. J. Virol. 76, 7
12537–12542. 8
Boom, R., Sol, C.J.A., Salimans, M.M.M., Jansen, C.L., Wertheim-Van Dillen, P.M.E., 9
Noordaa, J., Van, Der., 1990. Rapid and simple method for purification of nuclic 10
acids. J. Clin. Microbiol. 28, 495–503. 11
Borzacchiello, G., Ambrosio, V., Roperto, S., Poggiali, F., Tsirimonakis, E., Venuti, A., 12
2003b. Bovine papillomavirus type 4 in oesophageal papillomas of cattle from the 13
south of Italy. J. Comp. Pathol. 128, 203–206. 14
Borzacchiello, G., Iovane, G., Marcante, M.L., Poggiali, F., Roperto, F., Roperto, S., 15
Venuti, A., 2003a. Presence of bovine papillomavirus type 2 DNA and expression of 16
the viral oncoprotein E5 in naturally occurring urinary bladder tumours in cows. J. 17
Gen. Virol. 84, 2921-2926. 18
Borzacchiello, G., Roperto, F., 2008. Bovine papillomaviruses, papillomas and cancer 19
in cattle. V. Res. 39, 45-64. 20
Campo, M.S., 2002. Animal model of papillomavirus pathogenesis. Virus Res. 89, 249–21
261. 22
Campo, M.S., Coggins, L.W., 1982. Molecular cloning of bovine papillomavirus 23
genomes and comparison of their sequence homologies by heteroduplex mapping. J. 24
Gen. Virol. 63, 255– 264. 25

58
Campo, M.S., Moar, M.H., Jarret,W.F.H., Laird, H.M., 1980. A new papillomavirus 1
associated with alimentary cancer in cattle. Nature. 286, 180–182. 2
Campo, M.S., Moar, M.H., Laird, H.M., Jarret, W.F.H., 1981. Molecular heterogeneity 3
and lesion site specificity of cutaneous bovine papillomaviruses. Virology. 113, 4
323–335. 5
Chen, E.Y., Howley, P.M., Levinson, A.D., Seeburg, P.H., 1982. The primary structure 6
and genetic organization of the bovine papillomavirus type 1 genome. Nature. 299, 7
529–534. 8
Claus, M.P., Lunardi, M., Alfieri A.F., Ferracin, L.M., Fungaro, M.H.P., Alfieri A.A., 9
2008. Identification of unreported putative new bovine papillomavirus types in 10
Brazilian cattle herds. Vet. Microbiol. 132, 396-401. 11
Claus, M.P., Lunardi, M., Alfieri, A.A., Otonel, R.A.A., Sartori, D., Fungaro, M.H.P., 12
Alfieri, A.F. 2009b. Multiple bovine papillomavirus infections associated with 13
cutaneous papillomatosis in brazilian cattle herds Brazilian. Brazilian Arch. Biol. 14
Technol. 52, 93-98. 15
Claus, M.P., Lunardi, M., Alfieri, A.F., Sartori, D., Fungaro, M.H.P., Alfieri, A. A., 16
2009a. Identification of the recently described new type of bovine papillomavirus 17
(BPV-8) in Brazilian beef cattle herd. Pesqui. Vet. Bras. 29, 25-28. 18
Claus, M.P., Vivian, D., Lunardi, M., Alfieri, A.F., Alfieri A.A., 2007. Phylogenetic 19
analysis of bovine papillomavirus associated with skin warts in cattle herds from the 20
state of Parana. Pesqui .Vet. Bras. 27(7), 314-318. 21
de Villiers, E.M., Fauquet, C., Broker, T.R., Bernard, H.U., zur Hausen, H., 2004. 22
Classification of papillomaviruses. Virology. 324, 17–27. 23

59
Forslund, O., Antonsson, A., Nordin, P., Hansson, B.G., 1999. A broad range of human 1
papillomavirus types detected with a general PCR method suitable for analysis of 2
cutaneous tumours and normal skin. J. Gen. Virol. 80, 2437–2443. 3
Freitas, A.C., Carvalho, C., Brunner, O., Birgel-Jr, E.H., Dellalibera, A.M.M.P., Benesi, 4
F.J., Gregory, L., Beçak, W., Santos, R.C.S., 2003. Viral DNA sequences in 5
peripheral blood and vertical transmission of the virus: a discussion about BPV-1. 6
Braz. J. Microbiol. 34, 76-78. 7
Hatama, S., Nishida, T., Kadota, K., Uchida, I., Kanno, T., 2009. Bovine papillomavirus 8
type 9 induces epithelial papillomas on the teat skin of heifers. Vet. Microbiol. 136, 9
347-351. 10
Hatama, S., Nobumoto, K., Kanno, T., 2008. Genomic and phylogenetic analysis of two 11
novel bovine papillomaviruses, BPV-9 and BPV-10. J. Gen. Virol. 89, 158-163. 12
Jarrett, W.F.H., Campo, M.S., O’Neil, B.W., Laird, H.M., Coggins, L.W., 1984. A 13
novel bovine papillomavirus (BPV-6) causing true epithelial papillomas of the 14
mammary gland skin: a member of a proposed new BPV subgroup. Virology. 136, 15
255–264. 16
Maeda, Y., Shibahara, T., Wada, Y., Kadota, K., Kanno, T., Uchida, I., Hatama, S., 17
2007. An outbreak of teat papillomatosis in cattle caused by bovine papilloma virus 18
(BPV) type 6 and unclassified BPVs. Vet. Microbiol. 121, 242–248. 19
Ogawa, T., Tomita, Y., Okada, M., Shinozaki, K., Kubonoya, H., Kaiho, I., Shirasawa, 20
H., 2004. Broad-spectrum detection of papillomaviruses in bovine teat papillomas 21
and health teat skin. J. Gen. Virol. 85, 2191- 2197. 22
Ogawa, T., Tomita, Y., Okada, M., Shirasawa, H., 2007. Complete genome and 23
phylogenetic position of bovine papillomavirus type 7. J. Gen. Virol. 88, 1934–24
1938. 25

60
Pfister, H., Linz, U., Gissmann, L., Huchthausen, B., Hoffman, D., Zur Hausen, H., 1
1979. Partial characterization of a new type of bovine papillomavirus. Virology. 96, 2
1–8. 3
Sambrook, J., Fritsch, E.F., Maniatis, T., 1989. Molecular Cloning: A Laboratory 4
Manual, second ed. Cold Spring Harbor Laboratory Press, NY, pp. 1659. 5
Santos, R.C.S., Lindsey, C.J., Ferraz, O.P., Pinto, J.R., Mirandola, R.S., Benesi, E.J., 6
Birgel-Jr, E.H., Pereira, C.A.B., Beçak, W., 1998. Bovine papillomavirus 7
transmission and chromosomal aberrations: an experimental model. J. Gen. Virol. 8
79, 2127-2135. 9
Tomita, Y., Literak, I., Ogawa, T., Jin, Z., Shirasawa, H., 2007. Complete genomes and 10
phylogenetic positions of bovine papillomavirus type 8 and a variant type from 11
European bison. Virus Genes. 35, 243–249. 12
Wosiacki, S.R., Barreiros, M.A.B., Alfieri, A.F., Alfieri, A.A., 2005. Seminested- PCR 13
for detection and typing of bovine papillomavirus type 2 in urinary bladder and 14
whole blood from cattle with enzootic haematuria. J. Virol. Methods. 126, 215-219. 15
Wosiacki, S.R., Claus, M.C., Alfieri, A.F., Alfieri, A.A., 2006. Bovine papillomavirus 16
type 2 detection in the urinary bladder of cattle with chronic enzootic haematuria. 17
Mem. Inst. Oswaldo Cruz. 101, 635-638. 18
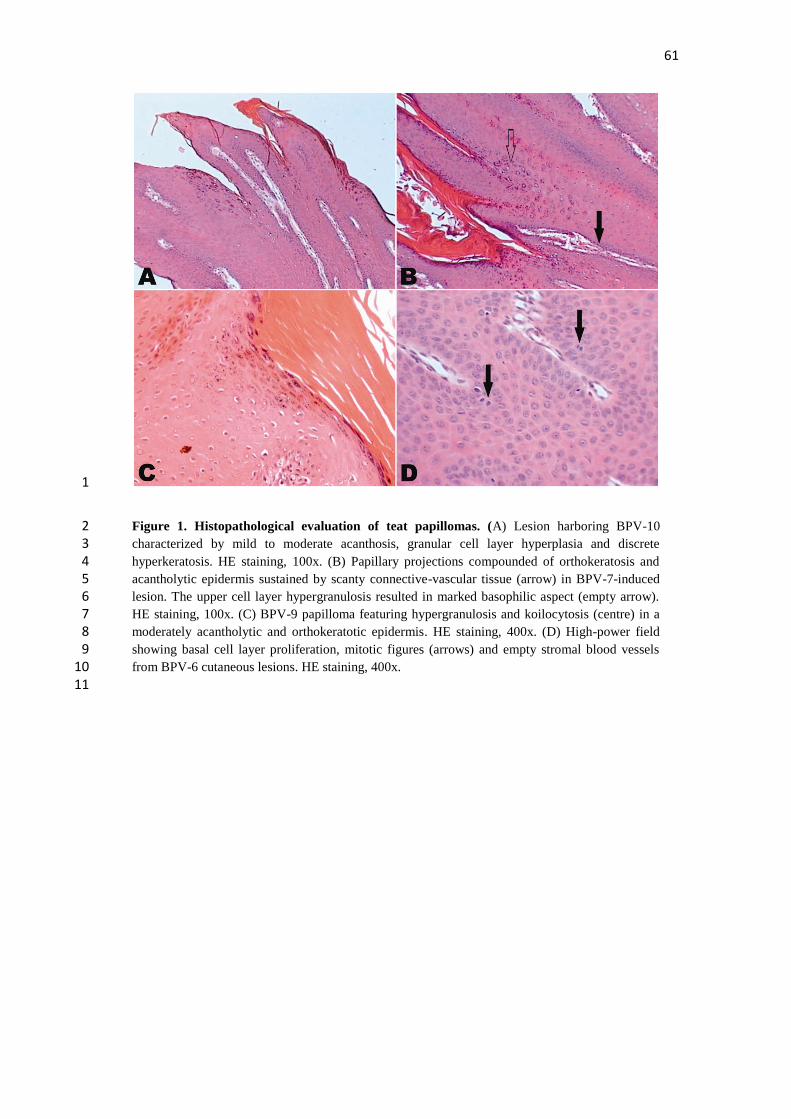
61
1
Figure 1. Histopathological evaluation of teat papillomas. (A) Lesion harboring BPV-10 2 characterized by mild to moderate acanthosis, granular cell layer hyperplasia and discrete 3 hyperkeratosis. HE staining, 100x. (B) Papillary projections compounded of orthokeratosis and 4 acantholytic epidermis sustained by scanty connective-vascular tissue (arrow) in BPV-7-induced 5 lesion. The upper cell layer hypergranulosis resulted in marked basophilic aspect (empty arrow). 6 HE staining, 100x. (C) BPV-9 papilloma featuring hypergranulosis and koilocytosis (centre) in a 7 moderately acantholytic and orthokeratotic epidermis. HE staining, 400x. (D) High-power field 8 showing basal cell layer proliferation, mitotic figures (arrows) and empty stromal blood vessels 9 from BPV-6 cutaneous lesions. HE staining, 400x. 10
11
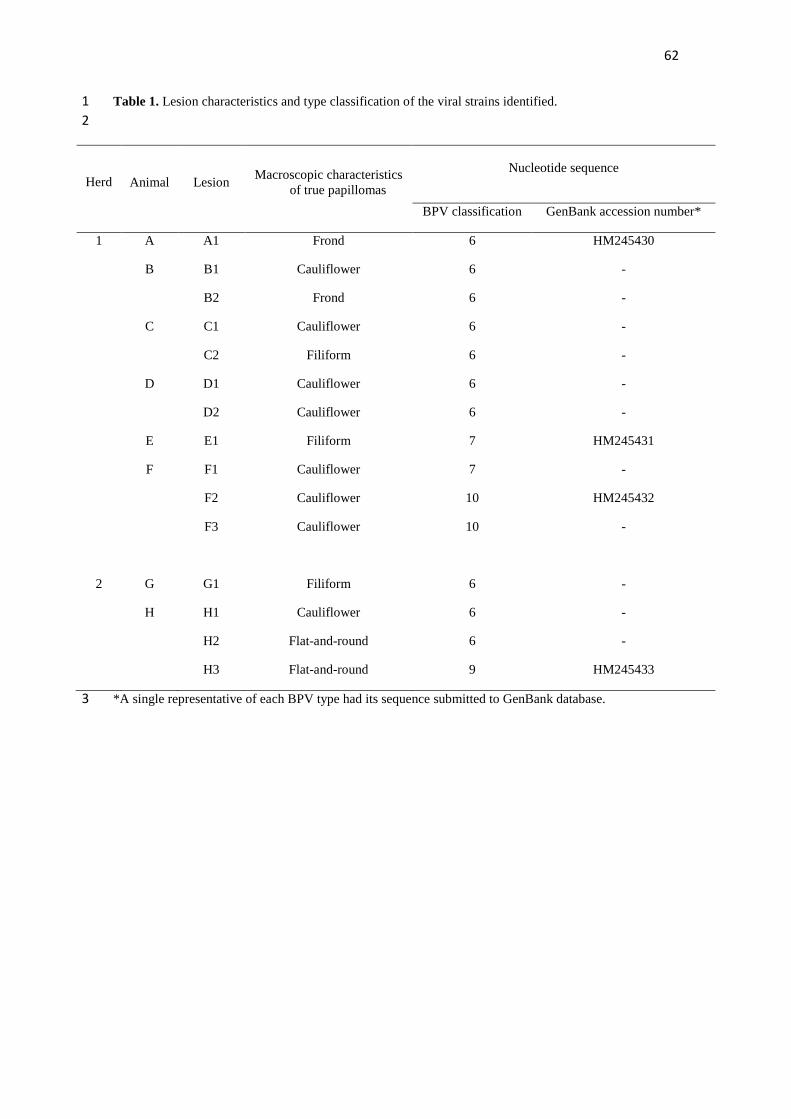
62
Table 1. Lesion characteristics and type classification of the viral strains identified. 1
2
Herd Animal Lesion Macroscopic characteristics
of true papillomas
Nucleotide sequence
BPV classification GenBank accession number*
1 A A1 Frond 6 HM245430
B B1 Cauliflower 6 -
B2 Frond 6 -
C C1 Cauliflower 6 -
C2 Filiform 6 -
D D1 Cauliflower 6 -
D2 Cauliflower 6 -
E E1 Filiform 7 HM245431
F F1 Cauliflower 7 -
F2 Cauliflower 10 HM245432
F3 Cauliflower 10 -
2 G G1 Filiform 6 -
H H1 Cauliflower 6 -
H2 Flat-and-round 6 -
H3 Flat-and-round 9 HM245433
*A single representative of each BPV type had its sequence submitted to GenBank database. 3

63
4.2. PHYLOGENETIC ANALYSES OF BOVINE PAPILLOMAVIRUS (BPV) 1
OF TEAT PAPILLOMAS IN BRAZILIAN DAIRY CATTLE HERDS 2
3
Abstract 4
5
Teat papillomatosis is an infectious disease that has been reported in dairy herds 6 worldwide as a cattle health problem responsible for economic losses. Despite the high 7 number of cattle herds affected by cutaneous papillomatosis, studies evaluating BPV 8
diversity affecting the mammary gland remain sporadic. The aim of this study is 9 identify the prevalence of BPV types involved in teat papilloma. Were collected 40 teat 10 papillomas of 25 dairy cows, in 13 dairy cattle herds of tree Brazilian state (Paraná, São 11 Paulo e Santa Catarina). The use of FAP59/64 primers pair assay amplified a fragment 12
of approximately 480 pb in all sample analyzed. Twenty nine teat papillomas specimens 13 processed for histopathological evaluation and two papillomas specimens were 14 submitted to the histochemical technique (Argyrophil Nucleolar Organizer Regions) 15 aiming visualization of NORs. The sequence analysis of the amplicons identified eight 16
different BPVs types: BPV-6, -7, -8, -9, -10, a previously described putative type 17 BAPV9 and two putative new types never described in the world (BPV/BR-UEL6 e -18 7). BPV-6 was the main viral type identified in teat lesions analyzed (45% -18/40), 19 followed by BPV-10 (32.5% -13/40). More than one BPV type were identified in 61.5% 20
(8/13) of the herds evaluated in this study. Multiple infections were identified in twelve 21 animals (48%) and of these, seven presented mixed infection (58.3%). The animal with 22
multiple papillomas that presented single infection were caused by BPV-6 (5/12). The 23 histological features revealed benign proliferation and viral citopathic effects in the 24
epidermis. All papillomas analysed were classified as true papilloma. The AgNOR 25 stained showed de intense proliferation of spinous layer of epidermis. The BPV-6 was 26
the prevalent BPV type causative teat papillomatosis identified, followed by BPV-10. 27 Despite, this study demonstrated the great diversity of BPV that associated with this 28 anatomical site. 29
30
Key-words: Bovine papillomavirus; teat papillomatosis; papilloma; histopathology. 31
Artigo editado de acordo com as normas de publicação do periódico Veterinary Microbiology,
disponível em: http://www.elsevier.com/wps/find/journaldescription.cws_home/503320/authorinstructions

64
1. Introduction 1
2
Papillomaviruses (PVs) are a group of non-enveloped, double-stranded and 3
circular DNA viruses that are strictly species and tissue specific and occur in a broad 4
range of distantly related animals. The only case of cross-species infection is for horses 5
and other equids by bovine papillomaviruses (BPVs) types 1 (BPV-1) or -2 (BPV-2) 6
(Antonsson, Hansson, 2002, de Villiers et al., 2004, Campo, 2002). 7
In cattle, BPV is widespread and induces papillomas and fibropapillomas in both 8
mucosal and cutaneous epithelia. BPV can induce the vulvo-vaginal, penis, and eye 9
epithelium lesions, skin and teat warts, papillomatosis and cancer of the upper 10
gastrointestinal tract and urinary bladder (Borzacchiello, Roperto, 2008). The bovine 11
cutaneous papillomatosis determines economic losses due to secondary infections, 12
depreciation of leather and may interfere in the bulls’ function (Campo, 2002). 13
Teat papillomatosis is an infectious disease that affects two years animals, but 14
can also affect adults. The lesions are characterized as self-limiting and most of them 15
underwent spontaneous regression, however, occasionally, some tumors may persist and 16
induce severe and generalized infection affecting various body sites (Sundeberg et al., 17
1987, Borzacchiello, Roperto, 2008). In dairy cows, teat papillomatosis can difficult 18
both manual and mechanical milking. The most common problems caused by 19
papillomas are: i) the difficulty of perfect coupling of liners caused by the irregularity of 20
the teat epithelial tissue or duct distortion causing air leaking and difficulty in sucking, 21
that endanger the teat health; ii) predisposition to mastitis due to the presence of residual 22
milk caused by the painful sensation that inhibits milk “let-down”; iii) the presence of 23
warts around the teat canal which may also predispose to mastitis; iv) an increased 24
predisposition to the development of secondary bacterial infections caused by teat warts 25

65
damage, and v) slaughter of heifers before the first lactation due to the lesion severity. 1
Therefore, is subject of the severity of the lesions and the number of affected animals, 2
the milk production can be unviable and the entire herd be slaughtered (Campo, 2002, 3
Blowey, Edmondson, 1995). 4
The BPVs are classified in four genera based on their biological properties and 5
genome relatedness. These genera are Deltapapillomavirus (BPV-1 and -2), 6
Xipapillomavirus (BPV-3, -4, -6, -9, -10 and -11), Epsilonpapillomavirus (BPV-5 and -7
8), and a yet unnamed PV genus (BPV-7) (de Villiers et al, 2004). 8
The L1 gene is the most conserved in genome and the sequence identity of this 9
gene has been used in the identification of new PVs types (de Villiers et al., 2004). The 10
partial amplification of L1 ORF by generic primers FAP59/FAP64 followed by 11
sequencing has allowed the identification of PV types in human and other animal hosts 12
(Forslund et al., 1999, Antonsson, Hansson, 2002, Ogawa et al., 2004, Claus et al., 13
2008). Currently had been complete sequenced 11 BPV types (BPV-1 to -11) and had 14
been partially described 15 putative new types, based on the highly conserved region 15
analyses (Ogawa et al., 2007, Tomita et al., 2007, Hatama et al., 2008, Hatama et al., 16
unpublished). The complete characterization of genome sequences of some putative 17
BPV types lead the identification of four most recently described BPV types (BPV-7, -18
8, -9, -10) (Ogawa et al., 2007, Tomita et al., 2007, Hatama et al., 2008). 19
The diagnosis of bovine papillomatosis is usually made from the presenting 20
clinical signs, because the structure of papillomas and fibropapillomas on the skin or 21
mucosal membranes is easily observed and identified. The diagnosis can be confirmed 22
by biopsy and histopathological examination. However these diagnostic methods do 23
not allow the identification of the causative BPV types (Alfieri et al., 2007). 24

66
A variety of treatments for cutaneous papillomatosis has been suggested but 1
currently there is no effective science-based treatment to bovine papillomatosis (Santin, 2
Brito, 2004). Immunity to BPV infection is considered type-specific and immune status 3
of infected animals is considered critical to define the clinical resolution. While humoral 4
immunity confers the ability to prevent new infections, cellular immunity, probably 5
involving T lymphocytes, is responsible for immune-mediated and spontaneous 6
regression of established lesions (Nicholls, Stanley, 2000). In cattle, preventive vaccines 7
have been developed against the BPV types 2 and 4. These two viral types were selected 8
as representative for the cutaneous and mucosal papillomaviruses, and also because both 9
types are related to determination of cancer in bovines (Campo et al., 1993). 10
Until now, eight BPV types were described as causative agents for udder and 11
teat papillomas (BPV-1, -3, -5, -6, -7, -8, -9, -10). Other BPV putative new types have 12
been reported from bovine teat papillomas and healthy skin swabs (Ogawa et al., 2004, 13
Maeda et al., 2007, Claus et al., 2007, Claus et al., 2008). Several studies aiming for 14
identification of BPVs in teat papillomatosis had identified BPV-6 as being the 15
prevalent type in this anatomical site (Lindholm et al., 1984, Jarret et al., 1984, Campo, 16
2002, Maeda et al., 2007). 17
As regards a viral infectious disease, the teat papillomatosis causes great losses 18
due spreading and lack of effective treatment. At present there are few epidemiological 19
studies that define the type of BPV teat lesions. The development of type-specific 20
vaccines can be an alternative to control of this disease. 21
The aim of this study was to identify the prevalence of BPV types involved in 22
teat papillomas collected from Brazilian dairy cattle herds of tree Brazilian states. 23

67
2. Materials and Methods 1
2
2.1. Papilloma specimens 3
4
Forty teat lesions were collected from 25 animals (16 cows and nine heifers) of 5
13 dairy cattle herds in three Brazilian states (Paraná, São Paulo and Santa Catarina). 6
The samples from Santa Catarina state were collected in an abattoir receiving animals 7
from the state. Twelve animals had multiples lesions and had over a sample. All dairy 8
cattle herds included in this study were small or medium milking producer and the total 9
number of animals among herds varied between 19 and 190 animals. The teat 10
papillomas collected had size ranging from 0.5 to 3 cm. The teat papillomas were 11
macroscopically classified as “frond, “caulyflower, “filiform” and “flat-and-round”. 12
13
2.2. DNA extraction 14
15
Fragments from each papilloma specimen were ground in phosphate-buffered 16
saline solution (PBS pH 7.2), and suspensions (10-20% w/v) were centrifuged for 15 17
min at 3000 g at 4 °C. Aliquots (250 μL) from supernatant were treated with lysis buffer 18
[10mM Tris; 1mM EDTA; 0.5% Nonidet P40; 1% SDS; and 0.2 mg/ml proteinase K 19
(Invitrogen, Life Technologies, Carlsbad, USA)]. After homogenization, samples were 20
incubated at 56°C for 30 min. 21
For DNA extraction, a combination of phenol/chloroform/isoamyl alcohol and 22
silica/guanidine isothiocyanate methods was carried out according to Alfieri et al. 23
(2006). Fractions from each sample were treated with an equal volume of 24
phenol/chloroform/isoamyl alcohol (25:24:1), homogenized and heated at 56 °C for 15 25

68
min (Sambrook, Russell, 2001). After centrifugation at 10,000 g for 10 min, the 1
aqueous phase was processed according to silica/guanidine isothiocyanate method 2
(Boom et al., 1990). DNA was eluted in 50 μL of ultrapure sterile water and kept at -20 3
°C until use. Aliquots of ultrapure sterile water were included as negative control in the 4
DNA extraction procedures. 5
6
2.3. PCR assay 7
8
The PCR assay was carried out using the primer pair FAP59 (forward: 5’- 9
TAACWGTIGGICAYCCWTATT-3’) and FAP64 (reverse: 5’- 10
CCWATATCWVHCATITCICCATC-3’) according to Forslund et al. (1999), with 11
slight modifications (Claus et al., 2007). Reaction was performed using 5 µL of the 12
extracted DNA and 45 µL of PCR-mix consisting of 1 µL (20 pmol) of each primer; 13
200 mM of each dNTP (Invitrogen, Life Technologies, Carlsbad, CA, USA); 2.5 units 14
of Platinum Taq DNA polymerase (Invitrogen, Life Technologies, São Paulo, SP, BR); 15
1x PCR buffer (20 mM Tris-HCl pH 8.4 and 50 mM KCl); 1.5 mM of MgCl2 and 16
ultrapure sterile water to a final volume of 50 μL. Amplification was performed in a 17
thermocycler (PTC 200, MJ Research Co., Water Town, Ma, USA) with the following 18
cycling profile: an initial step of 10 min at 94ºC, followed by 40 cycles of 1 min/94ºC, 1 19
min/50ºC, 1 min/72ºC, and a final extension step of 10 min/72ºC. Aliquots of 5 μL from 20
the PCR products were analyzed by electrophoresis in 2% agarose gel in TBE buffer pH 21
8.4 (89 mM Tris; 89 mM boric acid; 2 mM EDTA) at constant voltage (90V) for 22
aproximately 45 min. The agarose gel was stained with ethidium bromide (0.5 mg/mL), 23
and visualized under UV ligth. 24

69
2.4. Sequence analysis 1
2
Initially, all PCR products were purified using PureLink Quick Gel Extraction 3
Kit (Invitrogen, Life Technologies, Carlsbad, CA, USA) and quantified using Quant-iT 4
TM dsDNA BR Assay kit (Invitrogen, Molecular Probes, Eugene, OR, USA) in the 5
Qubit TM Fluorometer (Invitrogen, Molecular Probes, Eugene, OR). The direct 6
sequencing was performed by using the DYEnamic ET dye terminator cycle sequencing 7
kit (DYEnamic ET dye terminator cycle sequencing kit (Amersham Biosciences, 8
Piscataway, NJ, USA) with FAP59 and FAP64 primers, in a MegaBACE 9
1000/Automated 96 Capillary DNA Sequencer (GE Healthcare, Little Chalfont, UK), 10
according to the manufacturer’s instructions. The obtained sequences were examined 11
with the PHRED software (http://asparagin.cenargen.embrapa.br/phph) for quality 12
analysis of chromatogram readings. The sequences were accepted if base quality was 13
equal to or higher than 20. Consensus sequences were determined by CAP3 software 14
(http://asparagin.cenargen.embrapa.br/cgi-bin/phph/cap3.pl) and the sequence identity 15
was verified with all sequences deposited in the GenBank using the BLAST software 16
(http://blast.ncbi.nlm.nih.gov/Blast.cgi). The guidelines from the Papillomavirus 17
Nomenclature Committee 1995 (14th
International Papillomavirus Conference, Quebec 18
City, Quebec, Canada) were followed to identify putative new PV types. A new PV 19
isolate is defined as such if its complete genome has been cloned and the L1 nucleotide 20
sequence is more than 10% dissimilar from the closest known PV type. Differences 21
between 2% and 10% identity define a subtype and less than 2% a variant. Since 22
FAP59/FAP64 products represent only part of the L1 gene, the sequences obtained were 23
denominated putative new PV types instead of new PV types (Antonsson and Hansson, 24
2002; de Villiers et al., 2004; Ogawa et al., 2004). 25

70
2.5. Phylogenetic analysis 1
2
Two phylogenetic analysis were performed, with and without the BPV putative 3
BAA3 because of the minor sequence length. 4
The alignment and degree of similarity among sequences, at both nucleotide and 5
amino acid levels, were obtained using BIOEDIT version 5.0.9 software (Hall, 1999). 6
Phylogenetic trees were obtained by the Neighbour-joining method with the Kimura 7
two parameter distance estimate (Kimura, 1980), using MEGA version 3.1 program 8
(Kumar et al., 2004). Statistical analysis of phylogenetic trees were determined by 9
bootstrap method on 1000 replicates. The completely sequenced and putative viral types 10
of BPVs were included in the phylogenetic analysis. The putative BPV types included 11
in the study are denominated BAA1 to -4 and BAPV-4, -5, -7, -8, -9, and -10, BPV/BR-12
UEL2, -to -5 described by Antonsson, Hansson (2002), Ogawa et al. (2004), and Claus 13
et al. (2008), respectively. The GenBank accession numbers of BPV and putative new 14
BPV types sequences reported in this paper are: AF4885375 (BAA1); AF4885376 15
(BAA2); AF4885377 (BAA3); AF4885378 (BAA4); AY300819 (BAPV3); AY426550 16
(BAPV4); AY426551 (BAPV5); AY426553 (BAPV7); AY426554 (BAPV8); 17
AY426555 (BAPV9); AY426556 (BAPV10); XO2346 (BPV-1); M20219 (BPV-2); 18
AF486184 (BPV-3); XO5817 (BPV-4); AJ620206 (BPV-5); AJ620208 (BPV-6); 19
DQ217793 (BPV-7); DQ098913 (BPV-8); AB331650 (BPV-9); AB331651 (BPV- 10), 20
AB543507 (BPV-11). 21

71
2.6. Histopathology 1
2
Twenty nine teat papillomas specimens were fixed in 10% neutral buffered 3
formalin solution, embedded in paraffin and routinely processed for histopathological 4
evaluation. Sections (5 m) were stained with hematoxylin and eosin (HE). 5
Two papillomas (F2 and F3) specimens were submitted to the histochemical 6
technique (Argyrophil Nucleolar Organizer Regions) that allows visualization of NORs, 7
based on the technique described by Crocker, Skilbek (1987) with modifications. 8
9
3. Results 10
11
3.1 PCR and sequence analysis 12
13
The FAP59/64 primer pair assay amplified a fragment of approximately 480 bp 14
in all teat papillomas included in this study. The sequence analysis of the amplicons 15
identified eight different BPV types: BPV-6, -7, -8, -9, and -10; the putative BAPV9 16
and two putative new type never described in the world. BPV-6 was the main viral type 17
identified in the teat lesions analyzed (45% -18/40), followed by BPV-10 (32.5% -18
13/40), other BPV types and BAPV9 (17.5%- 7/40), and putative new BPV types (5%- 19
2/40) (Table 1). 20
Multiple infections were identified in twelve animals (48%- 12/25) and 58,3% 21
(7/12) had mixed infection, characterized by infections with more than one type of 22
BPV. The animals that had single infection were exclusively caused by BPV-6 (5/12), 23
and 75% (9/12) had papillomas harboring BPV-6 sequences, associated or not with 24

72
other BPV types. The distribution of BPV viral types in herds, animals and lesions are 1
showed in table 2. 2
More than one BPV type were identified in 61.5% (8/13) of the herds evaluated 3
in this study. Whereas of the five herds that had only one type identified, three of them 4
had only a sample collected, the number of herds infected by multiples BPV types is 5
noteworthy prevalent. 6
Association between the macroscopic characteristics and the viral type detected 7
in the cutaneous teat lesions was not found. BPV-6 was detected in teat warts presenting 8
frond, cauliflower, filiform and flat-and-round gross aspects; BPV-7 in both filiform 9
and cauliflower; BPV-9 in flat-and-round and cauliflower, BPV-10 in cauliflower, flat-10
and-round and frond macroscopic classified lesions. 11
12
3.2. Phylogenetic analyses 13
14
A representative of each BPV type and the two Brazilian putative new BPV types 15
identified in this study were grouped with BPV types and putative BPV types, 16
representing three previously defined PV genera (Deltapapillomavirus, 17
Epsilonpapillomavirus and Xipapillomavirus) (de Villiers et al., 2004). The Brazilian 18
putative new BPV BPV/BR-UEL7 was gouped with BPV types and putative types 19
(BAA1, BAPV-3, -9, -10, BPV/BR-UEL-2 e -3) grouped in Xipapillomavirus genus 20
whereas BPV/BR-UEL6 did not group in any genus (Figure 1). In a second analysis, 21
considering in a small length alignment with the BPV putative BAA3, the BPV/BR-22
UEL6 clustered with this BPV putative type (Figure 2). 23
In table 3 and 4, the two lesion characteristics are indicated as well as the highest 24
similarity percentage of Brazilian putative new BPV types with BPV types 1 to 10. 25

73
3.3. Histology 1
2
The histological features of 29 teat papillomas revealed benign exophytic 3
proliferation of epidermis, epidermal differentiation showing orthokeratosis or 4
parakeratosis; hyperplasia of granule cells in the esquamous layer; enlarge keratohyaline 5
granules; koilocitose; and mitotic figures in basal layer. In the histochemical AgNOR 6
stained, the intense cellular proliferation activity in the zone of spinous layer of 7
epidermis was detected (Figure 3). All 29 specimens analyzed were classified as true 8
papillomas. The histological features of macroscopic classification of papillomas are 9
shown in figure 4. 10
The new BPV putative type BPV/BR-UEL7 described in this study showed in 11
histopathology papillary hyperplasia, large keratohyalin granules in moderate amount, 12
intense koilocytosis and was classified as true papilloma. The histopathology of the 13
other putative new type (BPV/BR-UEL6) described in this study was not undertaken. 14
1. 15
2. 4. Discussion 16
3. 17
The bovine teat papillomatosis is a contagious infectious viral disease spread 18
worldwide. Most animals eventually undergo self-cure and by the second or third 19
lactation the warts are gone. Despite this, teat papillomatosis may persist in some cows, 20
can lead to mastitis, causing distortion of the milk canals, prevent the suckling of calves 21
and make milking impossible. Among the economic problems caused by teat 22
papillomatosis is the culled of heavily affected animals and so occasionally are whole 23
herds. In this study, eight heifers were severe affected and probably would be culled 24
whether papillomas not recede. Despite the high frequency of infection, there are few 25

74
studies involving teat papillomatosis and the determination of viral types is still 1
sporadic. Whereas, the elucidation of the epidemiology and pathology of the disease is 2
critical for the development of control strategies. This is the largest study to determine 3
the prevalence of different types of BPV involved in teat papillomatosis in different 4
Brazilian geographic regions. 5
The identification of teat papillomas harboring the BPV types -6, -7, -8, -9, -10 and 6
BAPV9 demonstrates the high BPV viral diversity that affects cattle in this anatomical 7
site. Furthermore, the strategy of using the primer pair FAP59/64 enabled the 8
identification of two putative new BPV types, not before described. 9
In this study the most prevalent BPV type identified in teat papillomas was the 10
BPV-6 followed by BPV-10. The BPV-6 is the viral type classically associated with 11
teat papillomatosis lesions and several studies in different countries have identified the 12
BPV-6 as the prevalent viral type involved in teat warts (Lindholm et al., 1984, Jarret et 13
al., 1984, Maeda et al., 2006, Claus et al., 2007). In Japan, in an outbreak of teat 14
papillomatosis, the BPV-6 was identified in 64% of the lesions analyzed, in addition to 15
BPV-9 and BPV-10. In another study in Japan, including five farms, the BPV-6 was 16
identify only in swabs of healthy skin of dairy cattle, suggesting that the animal may 17
harbor the virus commensally, or may harbor a latent or subclinical infection. In the 18
same study, the BPV-10 was the prevalent viral type identified in teat lesions (7 / 15) 19
and was not frequently identified from swabs of healthy skin. The BPV-6 was identified 20
in all investigations that were related to teat papillomatosis, despite this, the prevalence 21
of particular BPV types highly pathogenic for the mammary gland may differ by 22
geographic region. 23
Despite the frequent description of multiple human papillomavirus (HPV) infections 24
in human’s warts, studies with the aim to evaluate the occurrence of mixed BPV 25

75
infections and identifying BPV types associated with different skin warts from 1
individual animals are still sporadic. Claus et al. (2009) demonstrates the high frequency 2
of multiple infections in different bovine anatomical sites in a study performed in 3
Brazil. In addition, the present study demonstrates the high frequency which the animal 4
may harbor mixed BPVs infections in one anatomical site. 5
In addition to BPV types completely sequenced, there are several types identified 6
through analysis of partial L1 segments. This is the first description of the putative BPV 7
type BAPV9 outside of Japan, whitch was identified from swabs of teat healthy skin. In 8
our study, the BAPV9 was identified from a teat papilloma, suggesting that, despite of 9
this BPV type having been identified only in swabs of healthy skin, such putative 10
pathogen may have potential to induce proliferative lesions. 11
Currently, more than 200 human papillomavirus were described, showing the high 12
diversity of this viral family. In cattle 11 types of BPV were completely sequenced, but 13
this small number probably reflects the lack of determination of BPV types in 14
infections. The use of the generic primer pair FAP59/64, originally developed to 15
identify epithelial human papillomaviruses, followed by sequencing and phylogenetic 16
analysis has allowed the identification of BPV types completed sequenced and 17
characterized putative new types from partial analysis of the L1 gene (Forslund et al., 18
1999, Antosson, Hansson, 2002, Ogawa et al., 2004, Claus et al., 2008). Many new 19
BPV types have been identified using this strategy and in our study two new putative 20
BPVs were described. 21
In current studies with HPVs, the way that specific HPV genotypes are associated 22
with distinct clinical and histological morphologies, and the marked preference of each 23
HPV genotype for specific tissues and sites have been considered as important aspects 24
of the nature of this group of heterogeneous viruses (Egawa et al., 1998). In order to 25

76
correlate microscopic findings with the viral type identified in teat lesions, the 1
prevalence of type BPV-6 and -10 in bovine teat papillomas is notable. However, the 2
viral cytopathic effects displayed in histopathology were similar and there was no 3
correlation with viral types, once samples that had the same BPV type identified 4
determined histopathological alterations in different intensities. According to Hatama et 5
al., (1990), the histopathological alterations may vary in effect and intensity with the 6
stage of lesion development. 7
The microscopic features of lesions showed characteristics of viral cytopathic 8
alterations set at different intensity among the samples. Benign exophytic proliferation 9
of epidermis, epidermal differentiation showing orthokeratosis or parakeratosis; 10
enlargement of granule cells in the stratum granulosum; basophilic and enlarged 11
keratohyaline granules; koilocitose and acidophilic intranuclear inclusions. All teat 12
papilloma samples had only involvement of the epidermic tissue and were classified as 13
true papillomas. 14
The lesions that were from the abattoir of Santa Catarina state contained the adjacent 15
derma, which allowed the identification of lymphoplasmacytic perivascular dermatitis 16
in the histopathology. This finding probably indicates the early stages of papilloma 17
regression. This finding does not mean that the other lesions did not have this alteration 18
because that did not contain the adjacent part of the dermis. 19
The intense cellular proliferation activity in spinous layer of epidermis was showed 20
by the technique of AgNOR that is considered a good marker of cellular proliferation 21
and used to evaluate progress of the malignant transformation (Carneiro et al., 2000). 22
In 2004, there was a reclassification of the family Papillomaviridae and currently 23
the BPVs are classified into three different genera according to the sequence homology 24
and biological features (de Villiers et al., 2004). BPV-1 and -2, grouped in 25

77
Deltapapillomavirus genus, are etiologic agents of infection in epithelium and dermis 1
by inducing the formation of fibropapillomas. BPV-3, -4, -6, -9 -10, and -11, belong to 2
the Xipapillomavirus genus and are exclusively epitheliotropic viruses that infect only 3
the epithelium and induce true papillomas formation. BPV-5 and -8 are members of the 4
genus Epsilonpapillomavirus and seem to have dual pathology, inducing 5
fibropapillomas and epithelial papillomas, and its genome shares homology with both 6
Deltapapillomavirus and Xipapillomavirus. Based on phylogenetic analysis, it was 7
proposed that BPV-7 belongs to a new genus of BPV. All completely described BPVs 8
identified in this study belong to the Xipapillomavirus (BPV-6, -9 e -10), 9
Epsilonpapillomavirus (BPV-8), and a not classified genera (BPV-7). In the 10
histopathological evaluation all samples were classified as true papillomas, reinforcing 11
that the current taxonomic classification of genus share genomic homology and 12
pathologial characteristics. 13
The causative viral type infecting lesions did not showed correlation with the 14
respectively macroscopic morphology. This result corroborates with other authors and 15
suggests that the viral type involved in the pathology should not be the only factor that 16
plays a role in determination of the lesion gross aspect. 17
The BPV-6 is the viral type classically associated with teat papillomas and was the 18
prevalent viral type identified in lesions in this study. It is unclear the reason for the 19
predilection for this anatomical site. This study demonstrated the great diversity of BPV 20
of bovine teat papillomatosis in three states of Brazil and the great diversity of viruses 21
in the same herd and in animals, since animals with multiple lesions was found to be 22
harbor different viral types. No data is available about the prevalence of different types 23
of BPV involved in mammary papillomatosis in dairy cattle in Brazil. This is the first 24
epidemiological study that identifies BPV types in teat papillomatosis of dairy cows in 25

78
different geographic regions, and correlates with the histopathological finds of the 1
lesion. The molecular epidemiology of BPV knowledge is important because the 2
immunity developed is type-specific. Vaccination strategies should be developed in 3
order to prevent the infection and spread of disease. 4
5
5. Acknowledgements 6
7
We thank Brazilian Institutes CNPq, CAPES, FINEP, and Fundação Araucaria 8
(FAP/PR) for financial support. Alfieri, A.A. and Alfieri, A.F. are recipient of CNPq 9
fellowship. 10

79
6. References 1
2
Alfieri, A.A., Parazzi, M.E., Takiuchi, E., Médici, K.C., Alfieri, A.F., 2006. Frequency 3
of group A rotavirus in diarrhoeic calves in Brazilian cattle herds, 1998–2002. Trop. 4
Anim. Health Prod. 38, 521–526. 5
Alfieri, A.A, Wosiaki, S., Alfieri, A.F., 2007. Papillomavirus. In: FLORES, F. E. 6
Virologia Veterinária. UFSM, Santa Maria, pp. 399-412. 7
Antonsson, A., Hansson, B.G., 2002. Healthy skin of many animal species harbors 8
papillomaviruses which are closely related to their human counterparts. J. Virol. 76, 9
12537–12542. 10
Blowey, R., Edmondson P., 1995. Mastitis Control in Dairy Herds. Farming Press, 11
United Kingdom, 196p. 12
Boom, R., Sol, C.J.A., Salimans, M.M.M., Jansen, C.L., Wertheim-Van Dillen, P.M.E., 13
Noordaa, J., Van, Der., 1990. Rapid and simple method for purification of nuclic 14
acids. J. Clin. Microbiol. 28, 495–503. 15
Borzacchiello, G., Roperto, F., 2008. Bovine papillomaviruses, papillomas and cancer 16
in cattle. V. Res. 39, 45-64. 17
Campo, M.S., 2002. Animal model of papillomavirus pathogenesis. Virus Res. 89, 249–18
261. 19
Campo, M.S., Jarret, W.F.H., Grindlay, G.J., Chandrachud, L.M., Mcgarvie, G.M., 20
O’neil, B.W., 1993. Prophylactic and therapeutic vaccination against a mucosal 21
papillomavirus. J. Gen. Virol. 74, 945-953. 22
Carneiro, S. S., Moreira, M.A.R., Almeida Netto, J.C., 2004. HPV e câncer 23
do colo uterino. Rev. Patol. Trop., 33, 01-20. 24

80
Claus, M.P., Vivian, D., Lunardi, M., Alfieri, A.F., Alfieri A.A., 2007. Phylogenetic 1
analysis of bovine papillomavirus associated with skin warts in cattle herds from the 2
state of Parana. Pesq.Vet. Bras. 27(7), 314-318. 3
Claus, M.P., Lunardi, M., Alfieri A.F., Ferracin, L.M., Fungaro, M.H.P., Alfieri A.A., 4
2008. Identification of unreported putative new bovine papillomavirus types in 5
Brazilian cattle herds. Vet. Microbiol. 132, 396-401. 6
Claus, M.P., Lunardi, M., Alfieri, A.A., Otonel, R.A.A., Sartori, D., Fungaro, M.H.P., 7
Alfieri, A.F., 2009. Multiple bovine papillomavirus infections associated with 8
cutaneous papillomatosis in brazilian cattle herds Brazilian. Braz. Arch. Biol. 9
Technol. 52, 93-98. 10
Crocker, J., Silbeck, N., 1987. Nucleolar organiser region associated proteins I 11
cutaneous melanotic lesions: a quantitative study. J. Clin. Pathol. 40, 885–889. 12
de Villiers, E.M., Fauquet, C., Broker, T.R., Bernard, H.U., Zur Hausen, H., 2004. 13
Classification of papillomaviruses. Virology. 324, 17–27. 14
Egawa, K., Honda, Y., Inaba Y., Ono T., 1998. Pigmented viral warts: a clinical and 15
histopathological study including human papillomavirus typing. Br. J. Dermatol. 16
138, 381-389. 17
Forslund, O., Antonsson, A., Nordin, P., Hansson, B.G., 1999. A broad range of human 18
papillomavirus types detected with a general PCR method suitable for analysis of 19
cutaneous tumours and normal skin. J. Gen. Virol. 80, 2437–2443. 20
Hall, T.A., 1999. BioEdit: a user-friendly biological sequence alignment editor and 21
analysis program for Windows 95/98/NT. Nucleic Acids Symposium. 41, 95-98. 22
Hamada, M., Oyamada, T., Yoshikawa, H., Yoshikawa, T., Itakura, C., 1990. 23
Histopathological development of equine cutaneous papillomas. J. Comp. Pathol. 24
102, 393-403. 25

81
Hatama, S., Ishihara, R., Kanno, T., Uchida, I. Unpublished (GenBank: AB543507.1). 1
Identification of a new bovine papillomavirus type, BPV-11, in the genus Xi 2
papillomavirus from a Holstein heifer. Acesso em 19/12/2010. 3
Hatama, S., Nobumoto, K., Kanno, T., 2008. Genomic and phylogenetic analysis of two 4
novel bovine papillomaviruses, BPV-9 and BPV-10. J. Gen. Virol. 89, 158-163. 5
Jarrett, W.F.H., Campo, M.S., O’Neil, B.W., Laird, H.M., Coggins, L.W., 1984. A 6
novel bovine papillomavirus (BPV-6) causing true epithelial papillomas of the 7
mammary gland skin: a member of a proposed new BPV subgroup. Virology. 136, 8
255–264. 9
Kimura, M., 1980. A simple method for estimating evolutionary rates of base 10
substitution through comparative studies of nucleotide sequences. J. Mol. Evol. 16, 11
111–120. 12
Kumar, S., Tamura, K., Nei, M., 2004. MEGA3: integrated software for molecular 13
evolutionary genetics analysis and sequence alignment. Brief. Bioinform. 5, 150–14
163. 15
Lindholm, L. Murphy, J., O’Neil, B.W, Campo, M.S., Jarrett, W.F.H., 1984. 16
Papillomas of the teats and udder of cattle and their causal viruses Veterinary 17
Record, 115(22), 574-577. 18
Maeda, Y., Shibahara, T., Wada, Y., Kadota, K., Kanno, T., Uchida, I., Hatama, S., 19
2007. An outbreak of teat papillomatosis in cattle caused by bovine papilloma virus 20
(BPV) type 6 and unclassified BPVs. Vet. Microbiol. 121, 242–248. 21
Nicholls, P.K., Stanley, M.A., 2000. The immunology of animal papillomaviruses. Vet. 22
Immun. and Immunop. 73, 101-127. 23

82
Ogawa, T., Tomita, Y., Okada, M., Shinozaki, K., Kubonoya, H., Kaiho, I., Shirasawa, 1
H., 2004. Broad-spectrum detection of papillomaviruses in bovine teat papillomas 2
and health teat skin. J. Gen. Virol. 85, 2191- 2197. 3
Ogawa, T., Tomita, Y., Okada, M., Shirasawa, H., 2007. Complete genome and 4
phylogenetic position of bovine papillomavirus type 7. J. Gen. Virol. 88, 1934–5
1938. 6
Sambrook, J., Russell, D.W., 2001. Molecular Cloning: A Laboratory Manual. 3 ed. 7
New York: Cold Spring Harbor Laboratory. 8
Santin, A.P.I., Brito, L.A.B. Estudo da papilomatose cutânea em bovinos leiteiros: 9
comparação de diferentes tratamentos. Ciência Animal Brasileira, v.5, n.1, p.39-45, 10
2004. 11
Sundeberg, J.P., 1987. Papillomavirus infections in animals. In Papillomaviruses and 12
Human disease. In: Syrjänen, L.G., Koss, L.G. Springer, Berlin. 13
Tomita, Y., Literák, I., Ogawa, T., Jin, Z., Shirasawa, H., 2007. Complete genomes and 14
philogenetic positions of bovine papilomavirus type 8 and variant type from 15
European bison. Virus Genes. 35, 243-249. 16
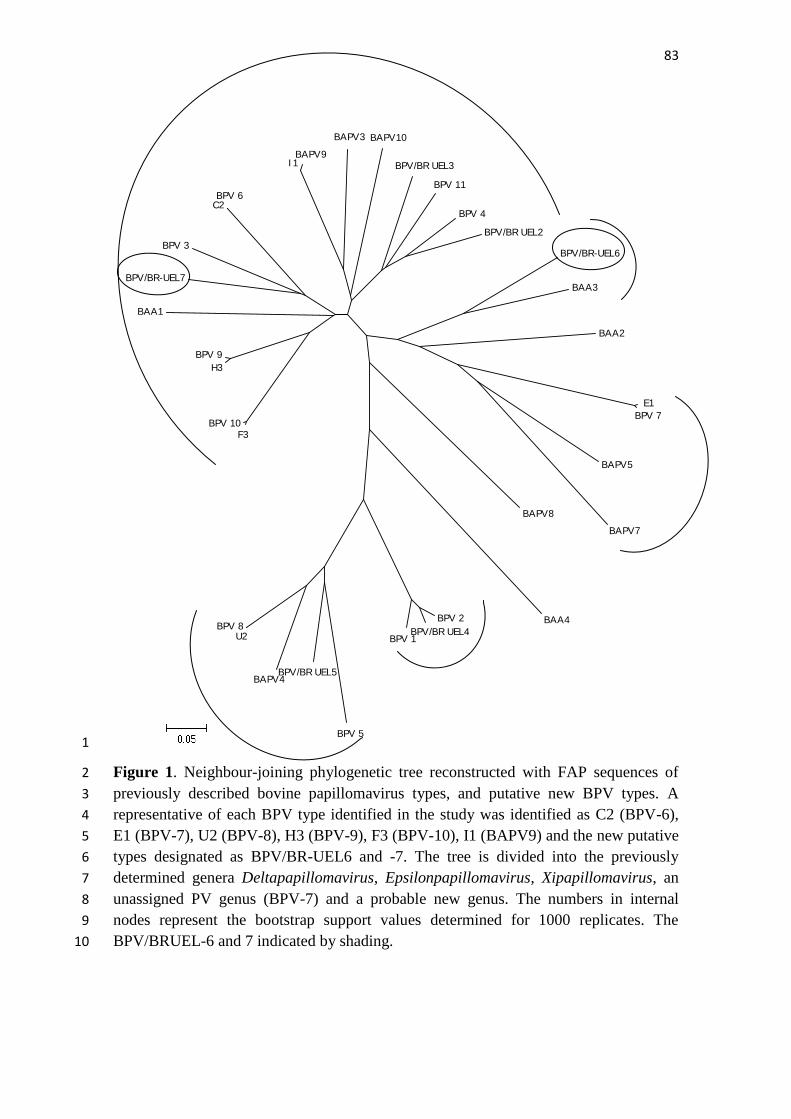
83
F3BPV 10
H3
BPV 9
BAA1
BPV/BR-UEL7
BPV 3
C2 BPV 6
I 1 BAPV9
BAPV3 BAPV10
BPV/BR UEL3
BPV 11
BPV 4
BPV/BR UEL2
BPV/BR-UEL6
BAA3
BAA2
E1
BPV 7
BAPV5
BAPV7
BAPV8
BAA4 BPV 2
BPV/BR UEL4BPV 1
BPV 5
BPV/BR UEL5BAPV4
U2BPV 8
1
Figure 1. Neighbour-joining phylogenetic tree reconstructed with FAP sequences of 2
previously described bovine papillomavirus types, and putative new BPV types. A 3
representative of each BPV type identified in the study was identified as C2 (BPV-6), 4
E1 (BPV-7), U2 (BPV-8), H3 (BPV-9), F3 (BPV-10), I1 (BAPV9) and the new putative 5
types designated as BPV/BR-UEL6 and -7. The tree is divided into the previously 6
determined genera Deltapapillomavirus, Epsilonpapillomavirus, Xipapillomavirus, an 7
unassigned PV genus (BPV-7) and a probable new genus. The numbers in internal 8
nodes represent the bootstrap support values determined for 1000 replicates. The 9
BPV/BRUEL-6 and 7 indicated by shading. 10
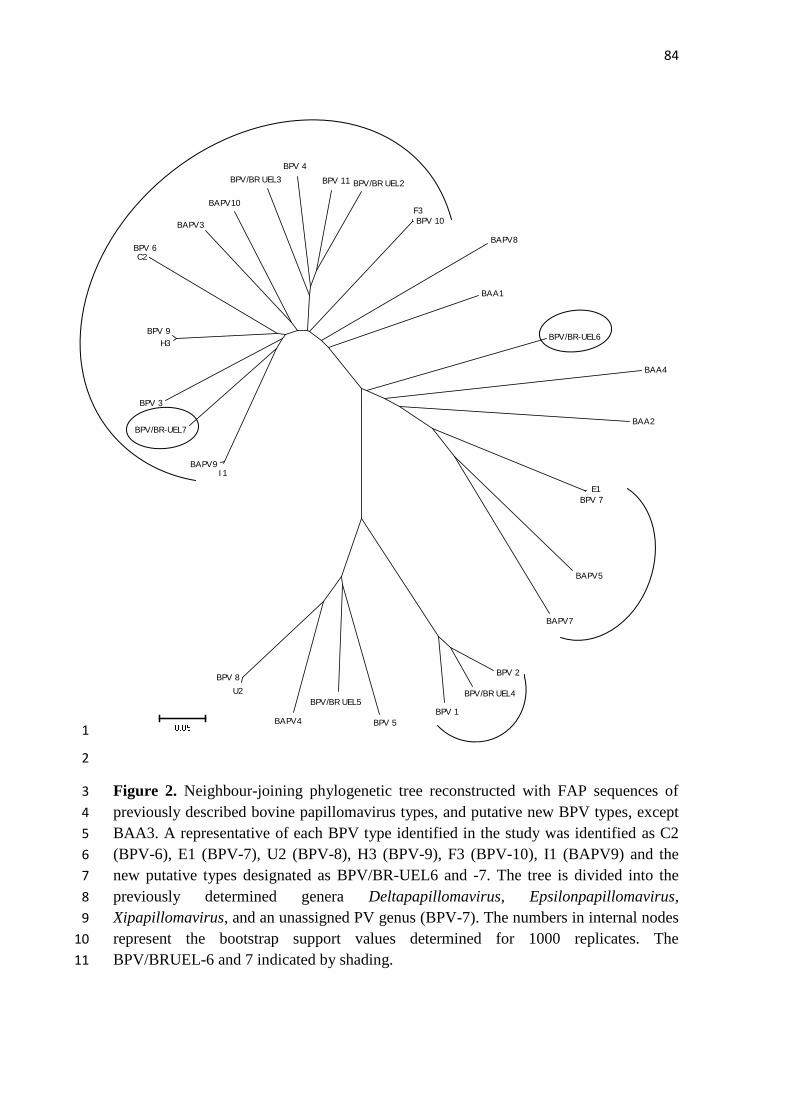
84
I 1BAPV9
BPV/BR-UEL7
BPV 3
H3
BPV 9
C2BPV 6
BAPV3
BAPV10
BPV/BR UEL3
BPV 4
BPV 11 BPV/BR UEL2
F3
BPV 10
BAPV8
BAA1
BPV/BR-UEL6
BAA4
BAA2
E1
BPV 7
BAPV5
BAPV7
BPV 2
BPV/BR UEL4
BPV 1
BPV 5
BPV/BR UEL5
BAPV4
U2
BPV 8
1
2
Figure 2. Neighbour-joining phylogenetic tree reconstructed with FAP sequences of 3
previously described bovine papillomavirus types, and putative new BPV types, except 4
BAA3. A representative of each BPV type identified in the study was identified as C2 5
(BPV-6), E1 (BPV-7), U2 (BPV-8), H3 (BPV-9), F3 (BPV-10), I1 (BAPV9) and the 6
new putative types designated as BPV/BR-UEL6 and -7. The tree is divided into the 7
previously determined genera Deltapapillomavirus, Epsilonpapillomavirus, 8
Xipapillomavirus, and an unassigned PV genus (BPV-7). The numbers in internal nodes 9
represent the bootstrap support values determined for 1000 replicates. The 10
BPV/BRUEL-6 and 7 indicated by shading. 11
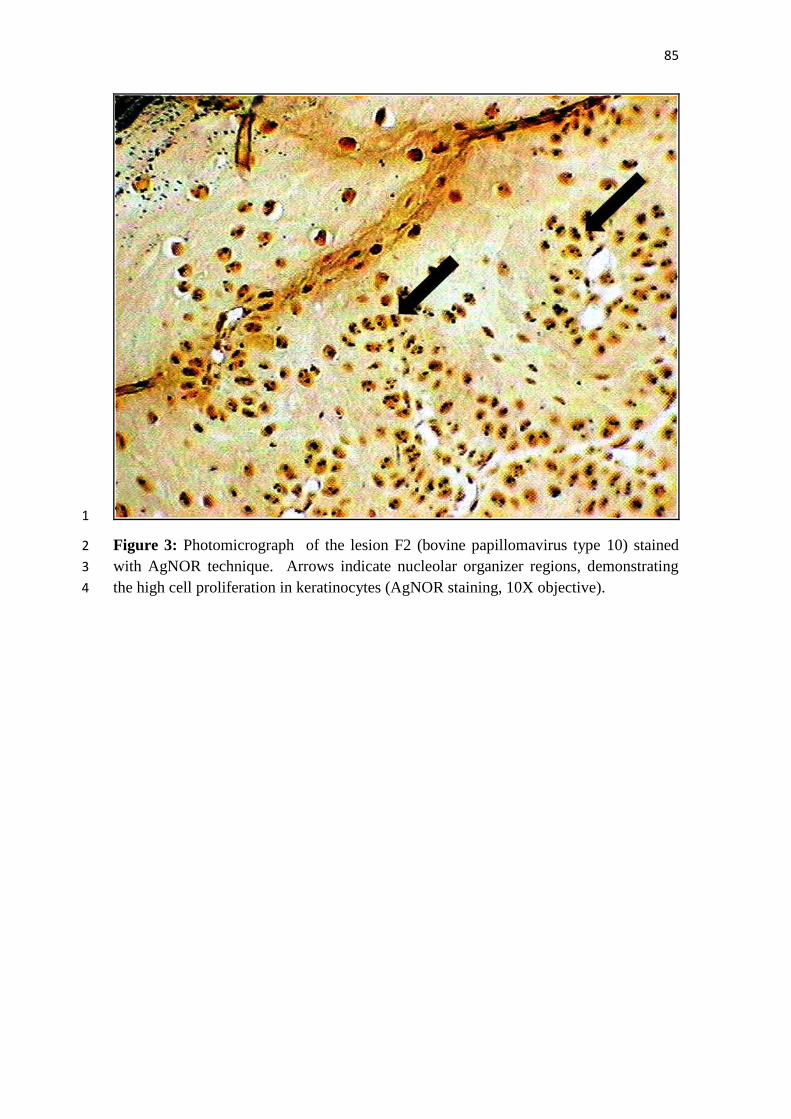
85
1
Figure 3: Photomicrograph of the lesion F2 (bovine papillomavirus type 10) stained 2
with AgNOR technique. Arrows indicate nucleolar organizer regions, demonstrating 3
the high cell proliferation in keratinocytes (AgNOR staining, 10X objective). 4
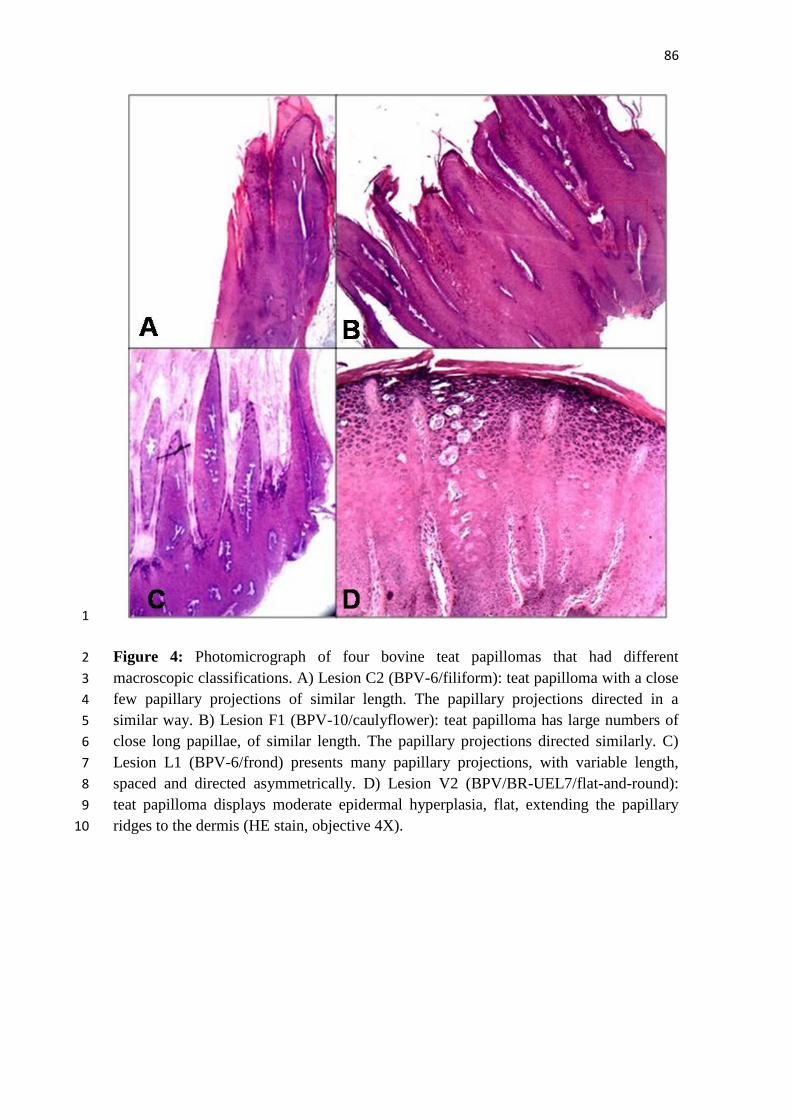
86
1
Figure 4: Photomicrograph of four bovine teat papillomas that had different 2
macroscopic classifications. A) Lesion C2 (BPV-6/filiform): teat papilloma with a close 3
few papillary projections of similar length. The papillary projections directed in a 4
similar way. B) Lesion F1 (BPV-10/caulyflower): teat papilloma has large numbers of 5
close long papillae, of similar length. The papillary projections directed similarly. C) 6
Lesion L1 (BPV-6/frond) presents many papillary projections, with variable length, 7
spaced and directed asymmetrically. D) Lesion V2 (BPV/BR-UEL7/flat-and-round): 8
teat papilloma displays moderate epidermal hyperplasia, flat, extending the papillary 9
ridges to the dermis (HE stain, objective 4X). 10
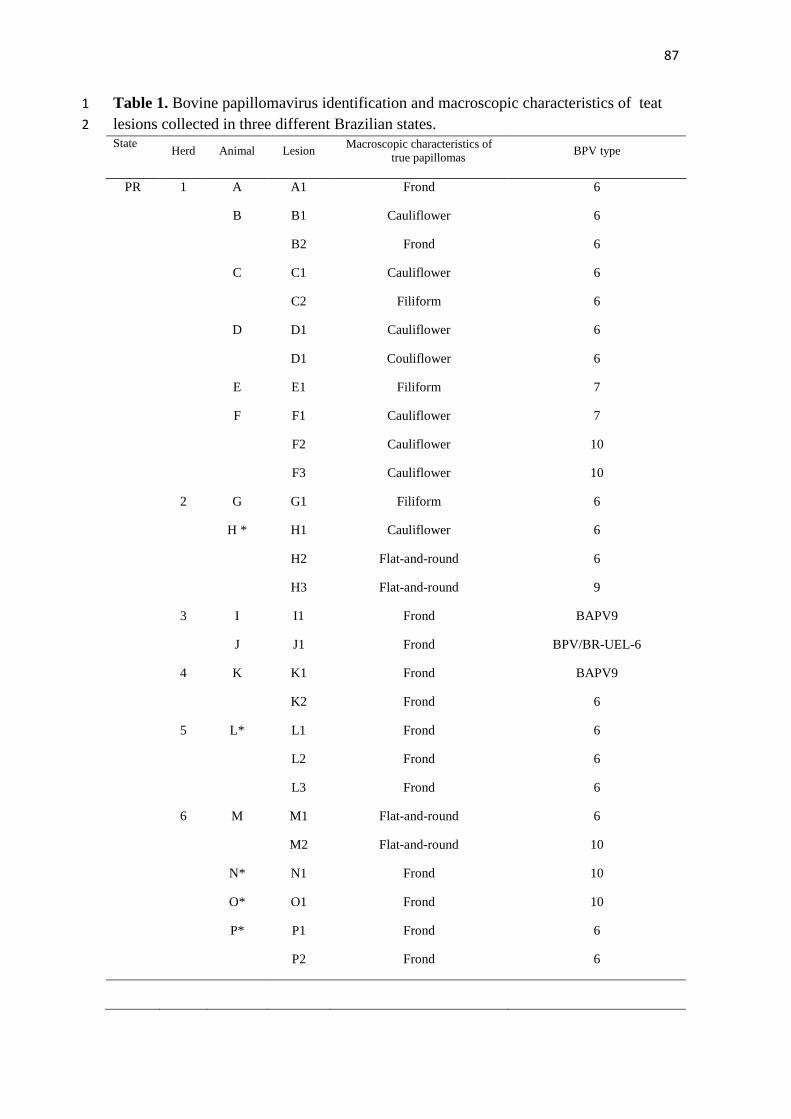
87
Table 1. Bovine papillomavirus identification and macroscopic characteristics of teat 1
lesions collected in three different Brazilian states. 2 State
Herd Animal Lesion Macroscopic characteristics of
true papillomas BPV type
PR 1 A A1 Frond 6
B B1 Cauliflower 6
B2 Frond 6
C C1 Cauliflower 6
C2 Filiform 6
D D1 Cauliflower 6
D1 Couliflower 6
E E1 Filiform 7
F F1 Cauliflower 7
F2 Cauliflower 10
F3 Cauliflower 10
2 G G1 Filiform 6
H * H1 Cauliflower 6
H2 Flat-and-round 6
H3 Flat-and-round 9
3 I I1 Frond BAPV9
J J1 Frond BPV/BR-UEL-6
4 K K1 Frond BAPV9
K2 Frond 6
5 L* L1 Frond 6
L2 Frond 6
L3 Frond 6
6 M M1 Flat-and-round 6
M2 Flat-and-round 10
N* N1 Frond 10
O* O1 Frond 10
P* P1 Frond 6
P2 Frond 6
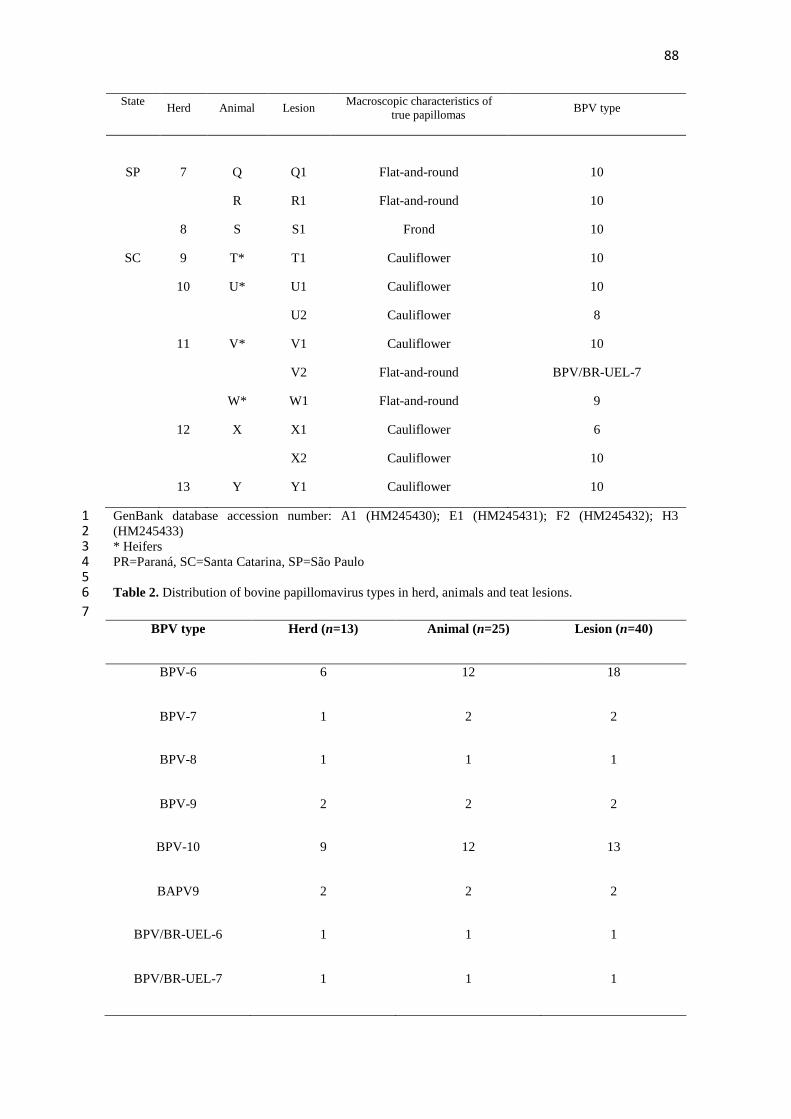
88
State Herd Animal Lesion
Macroscopic characteristics of
true papillomas BPV type
SP 7 Q Q1 Flat-and-round 10
R R1 Flat-and-round 10
8 S S1 Frond 10
SC 9 T* T1 Cauliflower 10
10 U* U1 Cauliflower 10
U2 Cauliflower 8
11 V* V1 Cauliflower 10
V2 Flat-and-round BPV/BR-UEL-7
W* W1 Flat-and-round 9
12 X X1 Cauliflower 6
X2 Cauliflower 10
13 Y Y1 Cauliflower 10
GenBank database accession number: A1 (HM245430); E1 (HM245431); F2 (HM245432); H3 1 (HM245433) 2 * Heifers 3 PR=Paraná, SC=Santa Catarina, SP=São Paulo 4 5 Table 2. Distribution of bovine papillomavirus types in herd, animals and teat lesions. 6
7 BPV type Herd (n=13) Animal (n=25) Lesion (n=40)
BPV-6 6 12 18
BPV-7 1 2 2
BPV-8 1 1 1
BPV-9 2 2 2
BPV-10 9 12 13
BAPV9 2 2 2
BPV/BR-UEL-6 1 1 1
BPV/BR-UEL-7 1 1 1
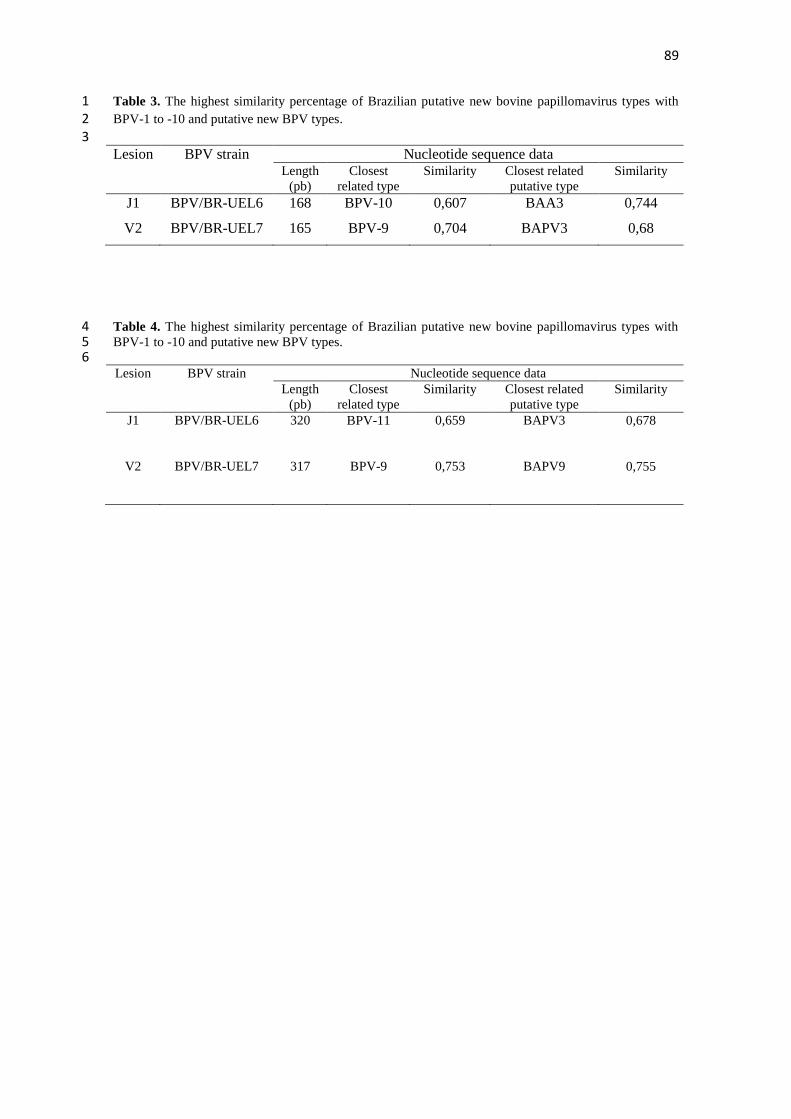
89
Table 3. The highest similarity percentage of Brazilian putative new bovine papillomavirus types with 1 BPV-1 to -10 and putative new BPV types. 2 3 Lesion BPV strain Nucleotide sequence data
Length
(pb)
Closest
related type
Similarity Closest related
putative type
Similarity
J1 BPV/BR-UEL6 168 BPV-10 0,607 BAA3 0,744
V2 BPV/BR-UEL7 165 BPV-9 0,704 BAPV3 0,68
Table 4. The highest similarity percentage of Brazilian putative new bovine papillomavirus types with 4 BPV-1 to -10 and putative new BPV types. 5 6 Lesion BPV strain Nucleotide sequence data
Length
(pb)
Closest
related type
Similarity Closest related
putative type
Similarity
J1 BPV/BR-UEL6 320 BPV-11 0,659
BAPV3 0,678
V2 BPV/BR-UEL7 317 BPV-9 0,753
BAPV9 0,755

90
5. CONCLUSÕES 1
2
A PCR, realizada com os primers genéricos FAP59/FAP64, foi eficaz na 3
amplificação de um produto com aproximadamente 480 pb do gene L1 do BPV 4
em todos os papilomas de tetos bovinos; 5
6
O estudo dos tipos de BPVs provenientes de lesões de teto de gado leiteiro, 7
realizado a partir das seqüencias de nucleotídeos dos produtos amplificados pela 8
PCR, proporcionaram a identificação dos tipos papilomavírus bovino BPV-6, -7, 9
-8, -9 e 10, do BAPV9 ( provável novo tipo) e dois prováveis novos tipos nunca 10
descritos; 11
12
O tipo viral prevalente identificado em de lesões de teto no estudo foi o BPV-6, 13
seguido do BPV-10; 14
15
A identificação pela primeira vez no continente Americano dos tipos virais 16
BPV-7, -9 e -10 recentemente descrito na Europa (BPV-10) e Japão (BPV-7, -9 17
e -10), sugere a distribuição mundial desses tipos virais; 18
19
A primeira identificação do suposto novo tipo viral BAPV9 em lesões de teto 20
demonstra o potencial patogênico desse vírus, que foi somente descrito 21
anteriormente em swab de pele de teto saudável de bovinos no Japão; 22
23
Nos rebanhos bovinos da regiões estudadas, as formas de infecção pelo BPV 24
foram identificadas como infecções simples (apenas um tipo viral no mesmo 25
animal) e mistas (mais de um tipo viral no mesmo animal); 26
27
A histopatologia permitiu a identificação dos efeitos citopáticos virais clássicos 28
associados à formação de papilomas verdadeiros. 29
30
Não houve relação do tipo de alteração histopatológica com o tipo de BPV 31
identificado. 32
33

91
APÊNDICES

92
APÊNDICE A 1
2
Lista de Reagentes 3
1. 100 mM dNTP Set, 4 x 250 mL; 25mmol each (100 mM dATP Solution, 100 4
mM dCTP Solution, 100 mM dGTP Solution, 100 mM dTTP Solution) (Invitrogen Life 5
Technologies®) 6
2. 10 x PCR-Buffer (200 mM Tris-HCl, pH 8.4, 500 mM KCl) (Invitrogen Life 7
Technologies®) 8
3. 123 bp DNA Ladder (Invitrogen Life Technologies®) 9
4. 2-Mercapto-ethanol (C2H6O5) P.M. 78,13 (Fluka®) 10
5. Acetona, P.A. (CH3COCH3) P.M. 58,08 (Dinâmica®) 11
6. Ácido acético glacial, P.A. (CH3COOH) P.M. 60,05 (Nuclear®) 12
7. Ácido bórico (H3BO3) P.M. 61,83 13
8. Ácido clorídrico (HCl) P.M. 36,46 (Reagen®) 14
9. Ácido etilenodiaminotetraácido Sal di-sódico – EDTA, P.A. 15
(C10H14N2O8Na22H2O) P.M. 372,24 (Reagen®) 16
10. Agarose (Gibco BRL®) 17
11. Ágar - Himedia Laboratories, India 18
12. Álcool etílico absoluto (C2H2OH) P.M. 46,07 (Nuclear®) 19
13. Álcool isoamílico ((CH3)2CHCH2CH2OH) P.M. 88,15 (Synth®) 20
14. Ampicilina trihidratada - USB, Cleveland, Ohio 21
15. Azul de bromofenol (Sigma®) 22
16. Cloreto de Potássio, P.A. (KCl) P.M. 74,56 (Reagen®) 23
17. Cloreto de Sódio, P.A. (NaCl) P.M. 58,45 (Reagen®) 24
18. Clorofórmio, P.A. (CHCl3) P.M. 119,38 (Dinâmica®) 25

93
19. Dodecil Sulfato de Sódio – Lauril Sulfato de Sódio – SDS (C12H25NaO4S) 1
P.M. 288,38 (Synth®) 2
20. Ethidium bromide (C21H20N3Br) P.M. 394,3 (Sigma®) 3
21. Extrato de Levedura - USB, Cleveland Ohio 4
22. Gibco BRL – Concert™ Rapid Plasmid purification System - Invitrogen Life 5
Technologies® 6
23. Glicina, P.A. (Nuclear®) 7
24. Guanidine isothiocyanate P.M. 118,16 (Gibco BRL®) 8
25. Hidróxido de sódio, P.A. (NaOH) P.M. 40,00 (Dinâmica®) 9
26. Hidroximetil amino metano – TRIS 99% P.M. 121,14 (Inlab®) 10
27. Lambda DNA - (Invitrogen Life Technologies®) 11
28. Metanol, P.A. (CH3OH) P.M. 32,04 (Allkimia®) 12
29. Platinum Taq DNA Polymerase recombinant 500 units (Invitrogen Life 13
Technologies®) 14
30. PureLink Quick Gel Extraction Kit (Invitrogen Life Technologies®) 15
31. Silicon dioxide (SiO2) P.M. 60,08 (Sigma®) 16

94
APÊNDICE B 1
2
Soluções e Tampões 3
• Hidratação da sílica 4
- 60 g de sílica (SIGMA®) 5
- Adicionar 500 mL de água MilliQ autoclavada 6
- Agitar lentamente e manter em repouso durante 24 horas 7
- Por sucção, desprezar 430 mL do sobrenadante 8
- Ressuspender a sílica em 500 mL de água bidestilada 9
- Manter em repouso durante 5 horas para sedimentar 10
- Desprezar 440 mL do sobrenadante 11
- Adicionar 600 μL de HCl (32% w/v) para ajustar o pH (pH=2,0) 12
- Aliquotar e autoclavar 13
14
• Solução L6 15
- 120 g de tiocianato de guanidina (GUSCN) 16
- 100 mL de TRIS-HCl 0,1 M pH 6,4 17
- 22 mL de EDTA 0,2 M pH 8,0 18
- 2,6 mL de Triton x 100 19
20
• Solução L2 21
- 120 g de tiocianato de guanidina (GUSCN) 22
- 100 mL de TRIS-HCl 0,1 M pH 6,4 23

95
• Tampão de Amostra 1
- Azul de bromofenol 0,25% 2
- Sacarose – sucrose (C12H22O11) 45% 3
4
Tampão de corrida: TBE (TRIS – Ácido bórico – EDTA) 10 x [ ] 5
- 0,89 M TRIS 6
- 0,89 M ácido bórico 7
- 0,02 M EDTA dissodium 8
- Água bidestilada qsp. 1 L 9
pH = 8,4 10
11
• Tampão fosfato salina (PBS) 12
-137 mM cloreto de sódio (NaCl) 13
- 3 mM cloreto de potássio (KCl) 14
- 8 mM sódio fosfato dibásico anidro (Na2HPO4) 15
- 15 mM potássio fosfato monobásico (K2H2PO4) 16
- Água MilliQ autoclavada q.s.p. 1 L 17
pH = 7,2 18
19
• Fenol / clorofórmio – álcool isoamílico (24:24:1) 20
- 24 mL fenol saturado 21
- 24 mL clorofórmio 22
- 1 mL álcool isoamílico 23

96
Tampão de Amostra 1
- Azul de bromofenol 0,25% 2
- Sacarose – sucrose (C12H22O11) 45% 3
4
Triton X -100 a 0.5% (Técnica de AgNOR) 5
- Acrescentar 0,5 mL de Triton X -100 em 99,5 mL de PBS 6
7
Solução A – Solução Colóide (Técnica de AgNOR) 8
- Aquecer 50 mL de água deionizada a 60°C 9
- Dissolver 1 g de gelatina 10
- Quando solução à temperatura ambiente, adicionar 0,5 mL de ácido fórmico 11
12
Solução B – Solução de Prata (Técnica de AgNOR) 13
- Dissolver 50 g de nitrato de prata em 100 mL de água destilada 14

97
APÊNDICE C 1
2
Protocolo de Técnicas 3
• Macerado de Papilomas de teto 4
- Triturar o papiloma 5
- Macerar o papiloma com pistilo em um gral 6
- Adicionar PBS 1x 7
- Homogeneizar em vórtex 8
- Centrifugar a 4.000 x g durante15 min 9
- Recolher 250 μL do sobrenadante em eppendorf para extração de DNA 10
11
• Lise Celular 12
- 250 μL do sobrenadante do macerado 13
- 250 μL de PBS 1x 14
- 1% de SDS (50 μL) 15
- 0,2 mg/mL de proteinase K (10 μL) 16
- Homogeneizar em vórtex 10/s 17
- Incubar em banho-maria à 56oC/30 min 18
19
• Extração do DNA pela técnica fenol / clorofórmio – álcool isoamílico / 20
sílica / tiocianato de guanidina 21
- Adicionar 500 μL de fenol clorofórmio álcool isoamílico após a lise celular 22
- Homogeneizar em vórtex 23
- Incubar em banho-maria à 56oC/15 min 24

98
- Homogeneizar em vórtex 1
- Centrifugar a 10.000 x g/12 min 2
- Recolher a fase aquosa em outro eppendorf 3
- Adicionar 25 μL de sílica hidratada 4
- Adicionar 1.000 μL de solução L6 5
- Homogeneizar em vórtex 6
- Agitar durante 30 min em temperatura ambiente 7
- Centrifugar a 10.000 x g 30/s 8
- Desprezar o sobrenadante em solução contendo NaOH 10 M 9
- Adicionar 500 μL de solução L2 10
- Homogeneizar em vórtex 11
- Centrifugar a 10.000 x g 30/s 12
- Desprezar o sobrenadante em solução contendo NaOH 10M 13
- Adicionar 500 μL de solução L2 14
- Homogeneizar em vórtex 15
- Centrifugar a 10.000 x g 30/s 16
- Desprezar o sobrenadante em solução contendo NaOH 10M 17
- Adicionar 1.000 μL de etanol 70% gelado 18
- Homogeneizar em vórtex 19
- Centrifugar a 10.000 x g 30/s 20
- Desprezar o sobrenadante 21
- Adicionar 1.000 μL de etanol 70% gelado 22
- Homogeneizar em vórtex 23

99
- Centrifugar a 10.000 x g 30/s 1
- Desprezar o sobrenadante 2
- Adicionar 1.000 μL de acetona PA gelada 3
- Homogeneizar em vórtex 4
- Centrifugar a 10.000 x g /2 min 5
- Desprezar o sobrenadante 6
- Secar o pellet em banho-maria a 56oC/15 min 7
- Adicionar 100μL de água miliQ autoclavada 8
- Homogeneizar em vórtex 9
- Descansar em banho-maria à 56oC/15 min 10
- Homogeneizar em vórtex 11
- Centrifugar a 10.000 x g /2 min 12
- Recolher o sobrenadante 13
- Estocar a –20ºC 14
15
• Gel de agarose a 2% 16
- 1 g de agarose 17
- 50 mL TEB buffer (Tris 89 mM; ácido bórico 89 mM; EDTA 2 mM) pH 8,4 18
- 30 µL de brometo de etídeo (0,5 μg/mL) 19

100
AgNOR 1
- Desparafinizar a lâmina à temperatura de 80°C 2
- Reidratar a partir de xilol a álcool 70° GL 3
- Colocar em água destilada por 10 min 4
- Colocar em imersão na solução de Triton X 100 a 0.5% por 15 min 5
- Lavar em água corrente por 20 min 6
- Colocar a lâmina invertida no laminário 7
- Preparar solução de A+B na proporção de 1A:2B 8
- Adicionar para cada lâmina, 3 mL da solução A+B 9
- Incubar em estufa a 37°C por 25 min 10
- Lavar em água corrente por 20 min 11
- Secar e montar 12

101
ANEXOS 1
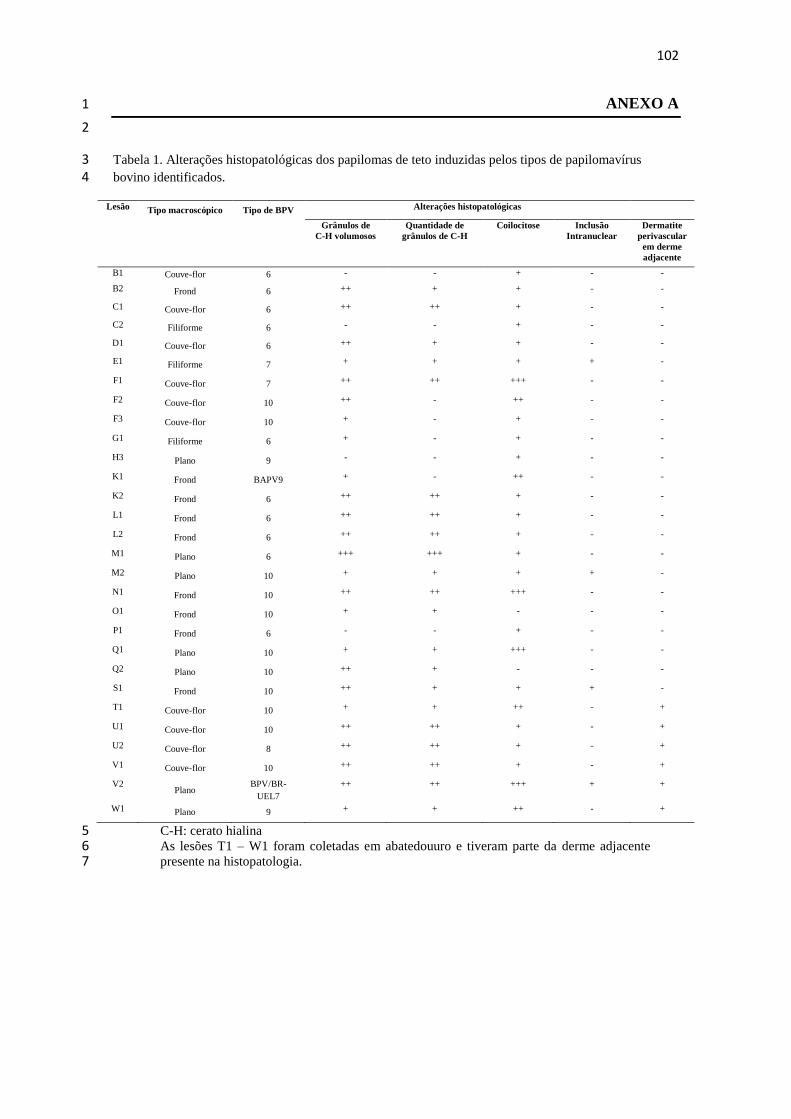
102
ANEXO A 1
2
Tabela 1. Alterações histopatológicas dos papilomas de teto induzidas pelos tipos de papilomavírus 3 bovino identificados. 4
Lesão Tipo macroscópico Tipo de BPV Alterações histopatológicas
Grânulos de
C-H volumosos
Quantidade de
grânulos de C-H
Coilocitose Inclusão
Intranuclear
Dermatite
perivascular
em derme
adjacente
B1 Couve-flor 6 - - + - -
B2 Frond 6 ++ + + - -
C1 Couve-flor 6 ++ ++ + - -
C2 Filiforme 6 - - + - -
D1 Couve-flor 6 ++ + + - -
E1 Filiforme 7 + + + + -
F1 Couve-flor 7 ++ ++ +++ - -
F2 Couve-flor 10 ++ - ++ - -
F3 Couve-flor 10 + - + - -
G1 Filiforme 6 + - + - -
H3 Plano 9 - - + - -
K1 Frond BAPV9 + - ++ - -
K2 Frond 6 ++ ++ + - -
L1 Frond 6 ++ ++ + - -
L2 Frond 6 ++ ++ + - -
M1 Plano 6 +++ +++ + - -
M2 Plano 10 + + + + -
N1 Frond 10 ++ ++ +++ - -
O1 Frond 10 + + - - -
P1 Frond 6 - - + - -
Q1 Plano 10 + + +++ - -
Q2 Plano 10 ++ + - - -
S1 Frond 10 ++ + + + -
T1 Couve-flor 10 + + ++ - +
U1 Couve-flor 10 ++ ++ + - +
U2 Couve-flor 8 ++ ++ + - +
V1 Couve-flor 10 ++ ++ + - +
V2 Plano
BPV/BR-
UEL7
++ ++ +++ + +
W1 Plano 9 + + ++ - +
C-H: cerato hialina 5 As lesões T1 – W1 foram coletadas em abatedouuro e tiveram parte da derme adjacente 6 presente na histopatologia. 7
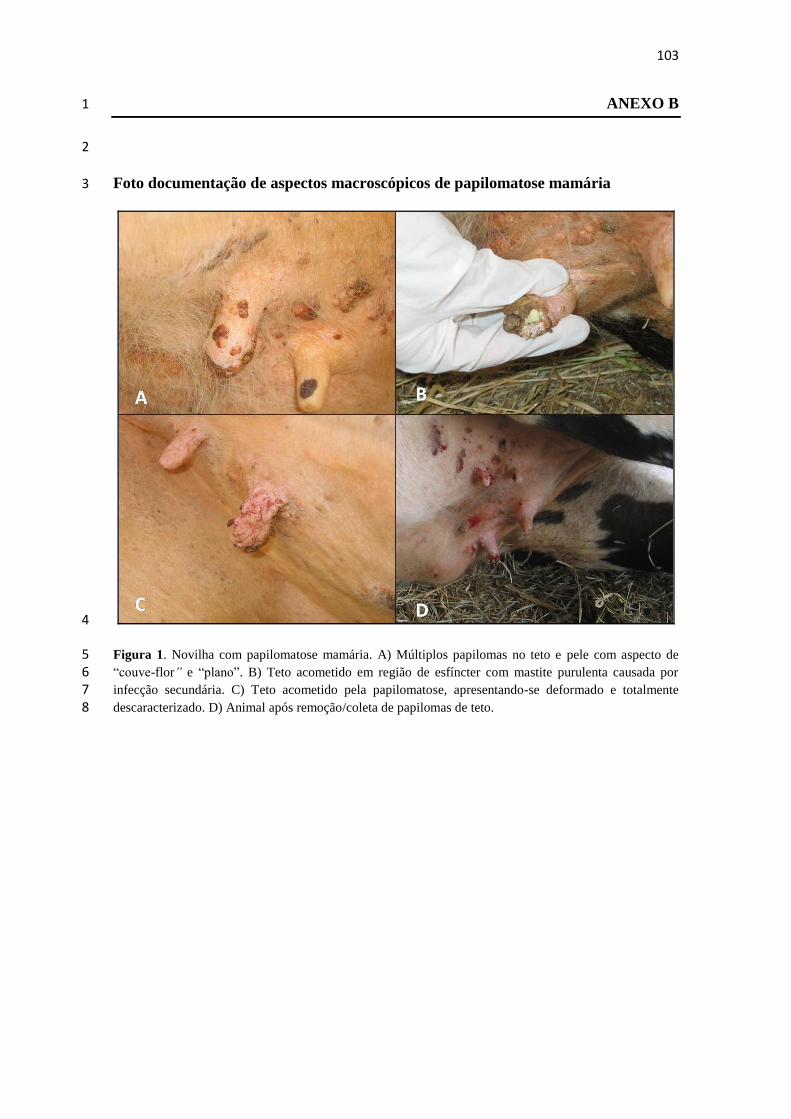
103
ANEXO B 1
2
Foto documentação de aspectos macroscópicos de papilomatose mamária 3
4
Figura 1. Novilha com papilomatose mamária. A) Múltiplos papilomas no teto e pele com aspecto de 5 “couve-flor” e “plano”. B) Teto acometido em região de esfíncter com mastite purulenta causada por 6 infecção secundária. C) Teto acometido pela papilomatose, apresentando-se deformado e totalmente 7 descaracterizado. D) Animal após remoção/coleta de papilomas de teto. 8
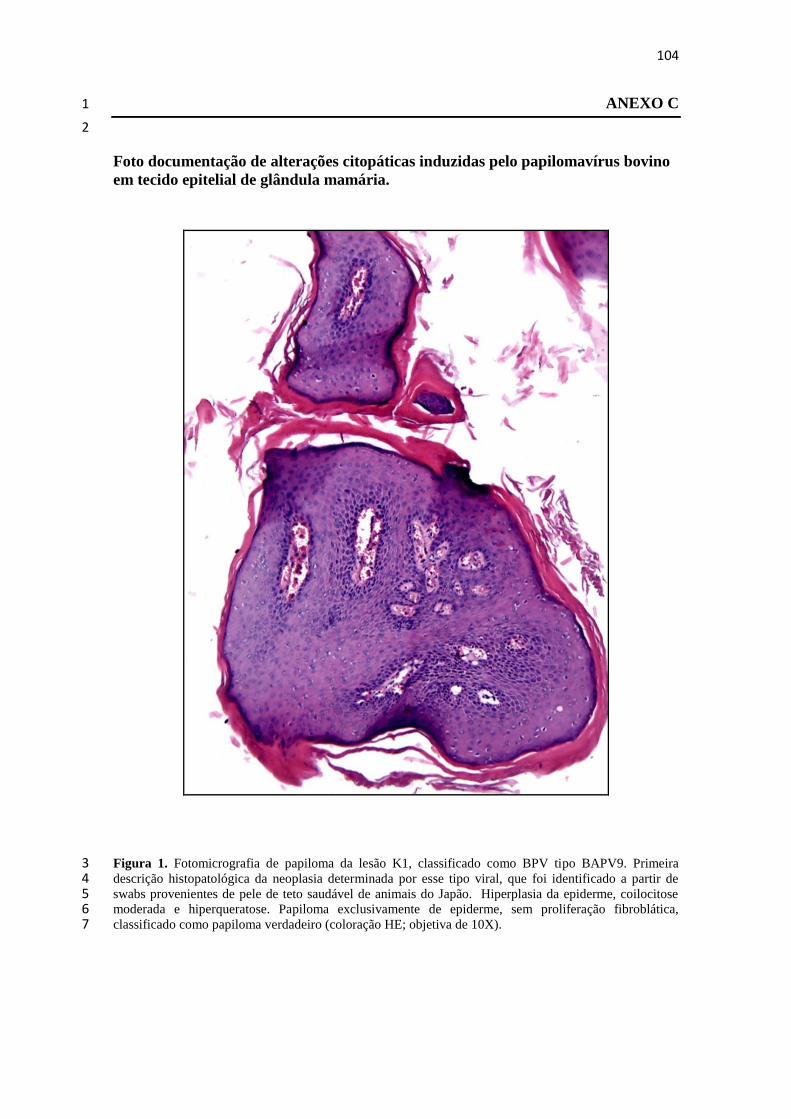
104
ANEXO C 1
2
Foto documentação de alterações citopáticas induzidas pelo papilomavírus bovino
em tecido epitelial de glândula mamária.
Figura 1. Fotomicrografia de papiloma da lesão K1, classificado como BPV tipo BAPV9. Primeira 3 descrição histopatológica da neoplasia determinada por esse tipo viral, que foi identificado a partir de 4 swabs provenientes de pele de teto saudável de animais do Japão. Hiperplasia da epiderme, coilocitose 5 moderada e hiperqueratose. Papiloma exclusivamente de epiderme, sem proliferação fibroblática, 6 classificado como papiloma verdadeiro (coloração HE; objetiva de 10X). 7
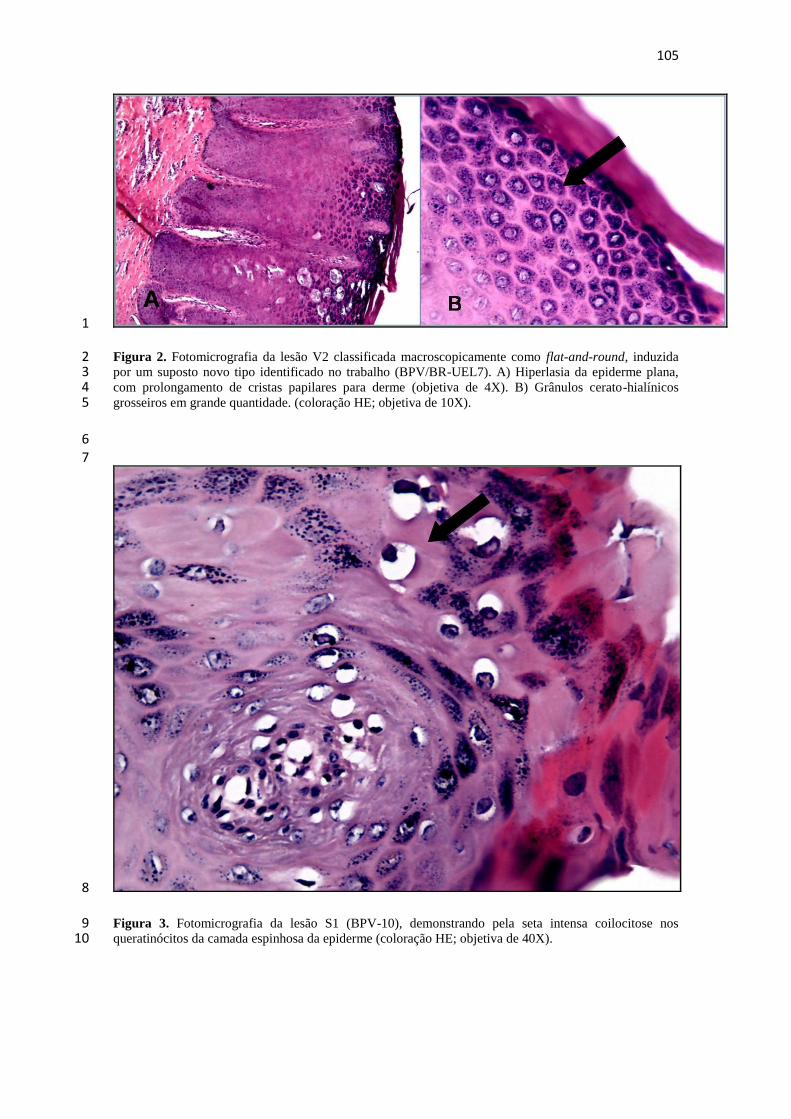
105
1
Figura 2. Fotomicrografia da lesão V2 classificada macroscopicamente como flat-and-round, induzida 2 por um suposto novo tipo identificado no trabalho (BPV/BR-UEL7). A) Hiperlasia da epiderme plana, 3 com prolongamento de cristas papilares para derme (objetiva de 4X). B) Grânulos cerato-hialínicos 4 grosseiros em grande quantidade. (coloração HE; objetiva de 10X). 5
6
7
8
Figura 3. Fotomicrografia da lesão S1 (BPV-10), demonstrando pela seta intensa coilocitose nos 9 queratinócitos da camada espinhosa da epiderme (coloração HE; objetiva de 40X). 10
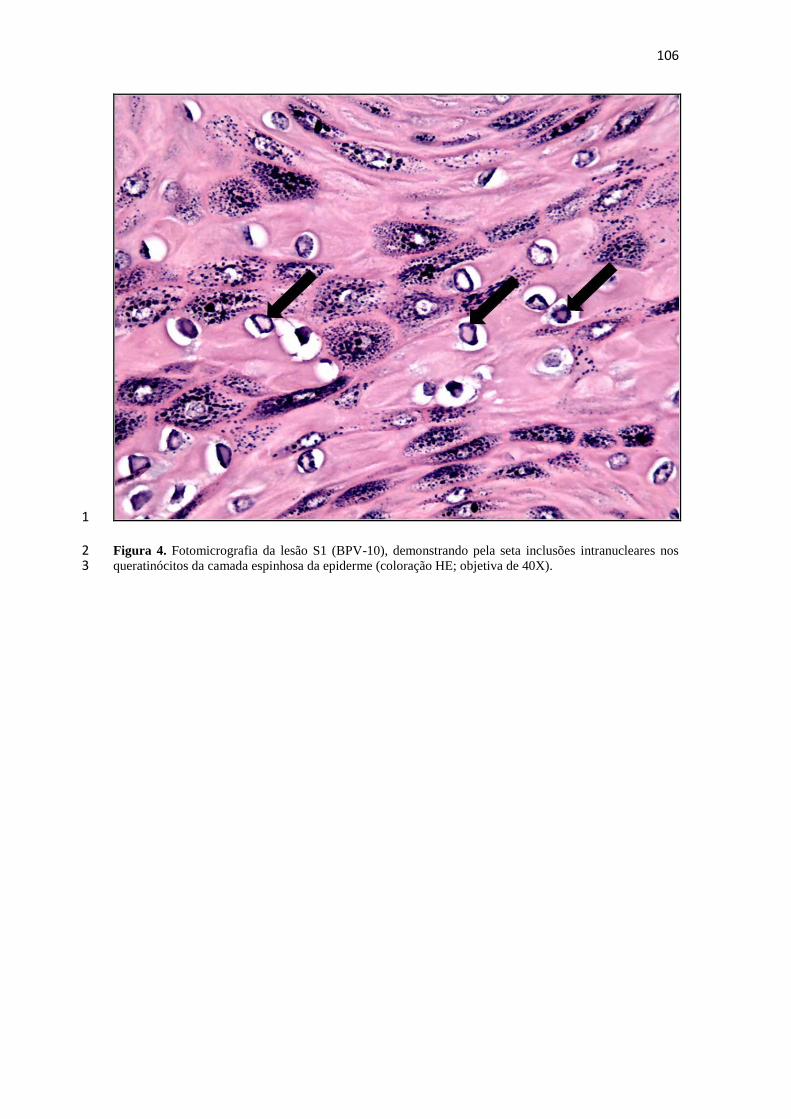
106
1
Figura 4. Fotomicrografia da lesão S1 (BPV-10), demonstrando pela seta inclusões intranucleares nos 2 queratinócitos da camada espinhosa da epiderme (coloração HE; objetiva de 40X). 3

107
1
Figura 5. Fotomicrografia da lesão U2 (BPV-8). A seta indica região de dermatite perivascular 2 linfoplasmocitária com predomínio de linfócitos em derme adjacente, sugerindo provavelmente um 3 estágio inicial de regressão do papiloma (coloração HE; objetiva de 10X). 4

108




















