
Conservation of mitotic controls in fission and budding yeasts

Discovering a chemical basis for differentiating wines made byfermentation with ‘wild’ indigenous and inoculated yeasts:
role of yeast volatile compounds
C. VARELA1, T. SIEBERT1, D. COZZOLINO1, L. ROSE2, H. MCLEAN3 and P.A. HENSCHKE1
1 Australian Wine Research Institute, PO Box 197, Glen Osmond,Adelaide, SA 5064,Australia2 The Yalumba Wine Company, Eden Valley Road,Angaston, SA 5353,Australia
3 Orlando Wines, Barossa Valley, Rowland Flat, SA 5352,AustraliaCorresponding author: Dr Paul A. Henschke, fax +61 8 8303 6601, email [email protected]
AbstractBackground and Aims: Winemakers are constantly searching for new techniques to modulate winestyle. Exploiting indigenous yeasts present in grape must is re-emerging as a commercial option in NewWorld wine regions. Wines made with indigenous or ‘wild’ yeasts are perceived to be more complex byshowing a greater diversity of flavours; however, the chemical basis for the flavour characteristics is notyet defined. In order to evaluate techniques for making wine with the ‘wild yeast fermentation’ charactermore reliably, it is necessary to define the salient chemical characteristics of such wines.Methods and Results: Pairs of Chardonnay wines were prepared from the same must and subjected tosimilar fermentation conditions in the wineries of origin, except for the mode of inoculation. Referencewines were made by inoculation with a Saccharomyces cerevisiae starter culture, whereas companion wineswere allowed to undergo fermentation with the indigenous microflora. Of all wine chemicals analysed,only yeast-derived volatile fermentation products showed significant differences between the yeasttreatments.Conclusions: Inoculated wines were associated with the esters ethyl hexanoate and 3-methylbutylacetate and formed a clear cluster by principal component analysis. By comparison with inoculatedwines, ‘wild’ yeast fermented wines showed high variability in volatile compounds that contribute towine aroma, with higher concentrations of 2-methylpropanol, 2-methylbutanoic acid, ethyl2-methylpropanoate, ethyl decanoate and ethyl dodecanoate potentially being sensorially important.Significance of the Study: This study shows that yeast-derived volatile fermentation products are akey difference between inoculated and uninoculated ferments and provides a chemical basis for the ‘wildyeast fermentation’ character.
AbbreviationsPCA principal component analysis; S. Saccharomyces; Uninoculated Fermentation ‘wild yeastfermentation’, indigenous yeast fermentation, natural fermentation, spontaneous fermentation
Keywords: fermentation, indigenous yeast, inoculation, wild yeast, wine composition
IntroductionUninoculated fermentation, often referred to as ‘natural’or ‘spontaneous’ fermentation, is a complex microbialprocess performed by the sequential action of differentnon-Saccharomyces and a diversity of indigenous Saccha-romyces yeasts. These yeasts, often referred to as ‘native’,‘indigenous’ or ‘wild’ yeasts, are found on the grapes,processing equipment, in the must and in the wine, andcan potentially contribute to the sensory characteristicsof that wine (Fleet and Heard 1993, Lambrechtsand Pretorius 2000, Jolly et al. 2006, Ugliano andHenschke 2009). In contrast, inoculation with a starterculture is intended to establish a dominant population
of a selected strain from the beginning of fermentation(Querol and Ramon 1996, Henick-Kling et al. 1998).Although the use of starter cultures can introducea relatively large population of active yeast, manystudies have shown that indigenous strains are notcompletely suppressed or eliminated but can developto a significant extent, especially during the early stagesof fermentation, and might influence the chemicaland sensory profile of the wine (Granchi et al. 1998,Henick-Kling et al. 1998, Fleet 2003, Combina et al.2005).
Grape juices and musts intended for uninoculated fer-mentations are characterised by a variety of indigenous
238 ‘Wild yeast fermentation’ character Australian Journal of Grape and Wine Research 15, 238–248, 2009
doi: 10.1111/j.1755-0238.2009.00054.x© 2009 The Australian Wine Research Institute

non-Saccharomyces species, whereas the population ofSaccharomyces yeasts is highly variable and is usually rela-tively small (Heard and Fleet 1985, Fleet and Heard 1993,Jolly et al. 2006). The ethanol-sensitive apiculate Hans-eniaspora (anamorph Kloeckera) species, which typicallydominate the yeast flora of grape berries, generally domi-nate freshly prepared juices/musts and early stages offermentation. Smaller populations of Candida species, suchas Candida stellata, Candida pulcherrima (teleomorphMetschnikowia pulcherrima), Candida sake and Candida colli-culosa (teleomorph Torulaspora delbrueckii) tend to domi-nate as the apiculate yeasts decline. Finally, low initialpopulations of the ethanol-tolerant Saccharomyces speciesand strains dominate as the non-Saccharomyces speciesdecline or become undetectable; only the more ethanol-tolerant species, such as C. stellata and C. colliculosa, canpersist during fermentation and sometimes complete fer-mentation even when Saccharomyces cerevisiae is present(Gutierrez et al. 1999, Soden et al. 2000, Hierro et al.2006). Other indigenous genera, having principallyoxidative or weak fermentative properties, includingCryptococcus, Debaryomyces, Issatchenkia, Kluyveromyces,Pichia, Rhodotorula, Torulaspora and Zygosaccharomyces, areoften present in initially small populations and tend todecline relatively early in fermentation (Heard 1999, Fleetet al. 2002, Romano et al. 2003b). Dekkera bruxellensis(anamorph Brettanomyces) has also been recently detectedon grape berries in some vineyards and has been shown topossess a strong potential to develop during fermentation(Renouf et al. 2006, Renouf and Lonvaud-Funel 2007).
Population dynamics of indigenous yeasts have beenreported to be affected by a multitude of viticultural aswell as winery factors, including vineyard environmentand management practices, harvest conditions, juice/must processing conditions and fermentation conditions(inoculation, SO2, macro- and micronutrients availability,temperature and microbial interactions) (Fleet and Heard1993, Charoenchai et al. 1997, Epifanio et al. 1999, Torijaet al. 2001, Jemec and Raspor 2005, Sturm et al. 2006).
Indigenous yeasts have been reported to contributeeither positively or negatively to the overall sensory char-acteristics of wine and to wine stability, although fewformal, systematic studies have been published (Herraizet al. 1990, Moreno et al. 1991, Zironi et al. 1993, Lemaet al. 1996, Ciani and Maccarelli 1998, Egli et al. 1998,Henick-Kling et al. 1998, Soden et al. 2000, Nurgel et al.2002a, Jolly et al. 2003, Rojas et al. 2003, Clemente-Jimenez et al. 2004, Ugliano and Henschke 2009). Indig-enous yeast fermentations are often associated with anextended lag phase and fermentation time, higherresidual sugars and with unpredictable by-product andoff-flavour formation. These fermentations are also asso-ciated with greater wine body, unusual or odd aromasand flavours, creamy texture and greater complexity,although little supporting data have been published (seee.g. Lurton et al. 1995, Ramey 1996, Egli et al. 1998,Henick-Kling et al. 1998, Goode 2005, Brajkovich 2008,Ugliano and Henschke 2009). Considering these charac-teristics, many winemakers regard the growth of indig-enous yeasts as contributing to the desirable sensory
properties of wine, whereas wines made by inoculationwith a commercial strain are perceived to be simpler orlacking complexity and less interesting (Herraiz et al.1990, Lema et al. 1996, Henick-Kling et al. 1998).
Most research associated with flavour implications ofuninoculated fermentations has focussed on the meta-bolic diversity of Saccharomyces and non-Saccharomycesisolates (Ciani and Maccarelli 1998, Rojas et al. 2003,Romano et al. 2003a, Clemente-Jimenez et al. 2004,Jolly et al. 2006), but only some studies have reportedsignificant differences in the chemical composition ofwines made by indigenous yeasts when compared tothose made by inoculation with pure cultures (Lemaet al. 1996, Egli et al. 1998, Nurgel et al. 2002b, Jollyet al. 2003). Although such studies have also describeddifferences in some volatile compounds (Romano et al.1997, Nurgel et al. 2002b, Vilanova and Sieiro 2006),the overall wine aroma composition has not beenaddressed.
Wine aroma is a unique mixture of volatile com-pounds originating from the grapes, primary and second-ary products formed during fermentation and compoundsproduced during ageing (Lambrechts and Pretorius 2000,Ugliano and Henschke 2009). Yeasts contribute to theoverall wine aroma by activating odourless precursorspresent in the grape must, such as terpenes, long-chainpolyfunctional thiols, C13-norisoprenoids, benzenoids, ali-phatics and volatile phenols (Darriet et al. 1988, Delfiniet al. 2001, Ugliano et al. 2006, Loscos et al. 2007), andsynthesising volatile metabolites, such as volatile sulphurcompounds, higher alcohols, esters, carbonyls and vola-tile fatty acids (Lambrechts and Pretorius 2000, Swiegerset al. 2005, Ferreira et al. 2008). Grape-derived volatilesare mostly responsible for the green (Allen 2008) andfloral characters of wines (Strauss et al. 1986). Hydrogensulphide imparts a rotten egg aroma, while other sulphurcompounds synthesised by yeast are associated withrubber, garlic and cabbage aromas (Bell and Henschke2005, Swiegers et al. 2005). Esters elicit pleasant sweet,fruity and fresh odours, whereas higher alcohols canimpart strong alcohol or solvent odours or floral notes inthe case of 2-phenylethanol. Medium-chain fatty acidshave soapy, unpleasant or rancid odours (Lambrechts andPretorius 2000, Francis and Newton 2005).
In order to devise new strategies to make wine withthe ‘wild yeast fermentation’ character more consistent, itis crucial to identify the most important chemical charac-teristics of ‘wild’ fermented wines. To achieve this aim,pairs of Chardonnay wines were prepared from the samemust and made under similar conditions, excluding theinoculation type; indigenous yeast or inoculation withcommercial wine yeast. The pairs of wines were subjectedto extensive chemical analysis for volatile and non-volatile compounds of grape and yeast origin.
Materials and methods
Yeast strains and fermentation conditionsThe strains used in this study were the commercialactive dry wine yeasts S. cerevisiae EC1118, D47 and
Varela et al. ‘Wild yeast fermentation’ character 239
© 2009 The Australian Wine Research Institute

CY3079 (Lallemand, Ontario, Canada). Chardonnayjuices were prepared in two different wineries located inthe Barossa Valley with grapes sourced from the EdenValley and Adelaide Hills regions (Australia) in 2004using commercial winery conditions (see below). Grapeswere mechanically harvested with no SO2 added, tankpressed; the juice was acid-adjusted and homogenised,and transferred to pairs of barrels (225 L) for fermenta-tion. Sugar concentration ranged between 210 and250 g/L. Two wines were made from each of the ninebatches of juice (carefully homogenised to minimisevariability) by fermentation under similar conditions,except for the mode of inoculation. Therefore, one batchwas inoculated with commercial wine yeast, while theother batch was made by allowing the indigenous yeaststo complete fermentation (‘wild’ yeast fermentation).Eight juices were inoculated with EC1118 and one juiceinoculated with a combination of D47 and CY3079 winestrains, as described by the yeast manufacturer. Juiceswere inoculated at 11°C and then allowed to warm upwith the maximum temperature being maintainedaround 17°C. All ferments were treated in a similarmanner to minimise the impact of fermentation tem-perature as a variable. Juices were fermented in oakbarrels of the same type and age, typically 3 or 4 yearsold, to limit oak flavours that can otherwise introduceconsiderable variability. Juices inoculated with EC1118and their uninoculated companions were fermented atthe same time in one winery, while the juice inoculatedwith D47 and CY3079 and the corresponding uninocu-lated juice were fermented concurrently in a secondwinery. After fermentation had completed and beforemalolactic fermentation had commenced, the wineswere clarified by cold settling with added SO2 (50 mg/L)and ascorbic acid (40 mg/L). No copper fining was madeto avoid loss of thiols. The wines were filtered andbottled without further treatment. The wines werestored at room temperature for 6 months before beingsubjected to extensive chemical analysis.
Analytical methodsAnalytical determinations were carried out in duplicate.Density, specific gravity, pH, volatile acidity, alcohol andtitratable acidity were determined by using a Foss WineS-can FT 120 as described by the manufacturer (Foss, Hill-erød, Denmark). Free and total SO2 were measured by theaspiration method (Rankine and Pocock 1970).
The concentrations of residual sugar, glycerol,malic, tartaric and citric acids were measured by high-performance liquid chromatography using a Bio-Rad(CA, USA) HPX-87H column as described previously(Nissen et al. 1997). Ammonia concentration was quan-tified by using the Glutamate Dehydrogenase EnzymaticBioanalysis UV-method test (Roche, Mannheim,Germany). Free a-amino acid nitrogen was determinedby the o-phthaldialdehyde/N-acetyl-L-cysteine spectro-photometric assay procedure (Dukes and Butzke 1998)using a Roche Cobas FARA spectrophotometric auto-analyser (Roche, Basel, Switzerland).
Brown pigments were estimated by absorbance at420 nm (Skouroumounis et al. 2003). Haze formationwas measured using a heating test (Pocock and Waters2006). Non-hydrolysed glycosylated flavour precursorswere determined by the glycosyl-glucose assay (Williamset al. 1995).
Analysis of volatile compoundsAcetaldehyde was measured using an enzymatic kit asdescribed by the manufacturer (Megazyme, Co. Wicklow,Ireland). Oak flavour compounds (cis-oak lactone, trans-oak lactone, guaiacol, 4-methyl guaiacol and vanillin)were determined using headspace solid-phase micro-extraction coupled with gas chromatography-massspectrometry (HS-SPME-GCMS), with polydeuteratedinternal standards for stable isotope dilution analysis(SIDA) as described previously (Pollnitz et al. 2004).Grape-derived volatile compounds (naphthalene, lina-lool, a-terpineol, nerol, b-damascenone and geraniol)were analysed using HS-SPME-GCMS with SIDA asdescribed elsewhere (Pedersen et al. 2003, Pollnitz et al.2003). Yeast volatile sulphur compounds (hydrogen sul-phide, methanethiol, dimethyl sulphide (DMS), carbondisulphide (CS2), dimethyl disulphide and diethyl disul-phide) were determined by using headspace cool-on-column gas chromatography coupled with an atomicemission detector, with ethyl methyl sulphide and propylthioacetate as internal standards (T.E. Siebert and A.P.Pollnitz, unpubl. data, 2008). Yeast volatile fermentationproducts were analysed using HS-SPME-GCMS, withSIDA as described previously (Siebert et al. 2005). Thirty-one compounds, including ethyl and acetate esters, vola-tile fatty acids and higher alcohols, were quantified.
Statistical analysisStatistical analysis was applied to establish if any mea-surement was associated with the inoculation mode.Thus, all nine inoculated juices were treated as a groupand compared with the group of uninoculated ferments.Statistical differences between inoculated and ‘wild’ferments were determined using analysis of variance(ANOVA). PCA was used to reduce the dimensionalityof data and to identify underlying variables. Cross-validation was used as validation method. ANOVA anddiscriminant analysis were performed using the JMP Soft-ware (SAS Institute, North Carolina, USA). PCA wascarried out with Unscambler 9.5 (CAMO ASA, Oslo,Norway).
Results and discussionTo define chemically the ‘wild yeast fermentation’ char-acter of wines of uninoculated fermentations, pairs ofChardonnay wines were prepared from the same grapejuice and fermented under similar commercial wineryconditions, except for the mode of inoculation. Oneportion of a batch of juice was inoculated with commer-cial S. cerevisiae wine yeast, while the other portion wasfermented with the indigenous yeasts. Inoculated winesfinished fermentation in similar time, while uninoculatedwines showed variable and slower fermentation kinetics.
240 ‘Wild yeast fermentation’ character Australian Journal of Grape and Wine Research 15, 238–248, 2009
© 2009 The Australian Wine Research Institute

All wines were then minimally processed to preserveall metabolites produced during fermentation, whichinvolved stabilisation with SO2 and ascorbic acid, clarifi-cation by cold settling under protection from oxygen andbottling under roll-on-tamper evident closures. Wineswere subjected to comprehensive chemical analysis fornon-volatile compounds and grape-derived, oak-derivedand yeast-derived volatile compounds.
Although notable differences in non-volatile fermen-tation products have been described between wines madeby inoculated and indigenous yeasts from the samebatches of grape juices (Lema et al. 1996, Egli et al. 1998),
we did not observe any statistically significant differenceswhen using Chardonnay juices sourced from differentviticultural regions. However, trends could be observedfor some compounds when individual pairs of wines areconsidered. Wine specific gravity, residual sugar, volatileacidity and A420 nm tended to be higher in the non-inoculated fermentation wines (data not shown),whereas SO2 concentrations were always higher for theinoculated wines (Figure 1a). This last observation sug-gests that either the dominant indigenous yeast(s) presentin these wineries juices was predominantly a lower SO2
producer than the selected strains of S. cerevisiae used in
Figure 1. (a) Total SO2, (b) hydrogen sulphide, (c) dimethyl sulphide, (d) carbon disulphide, (e) linalool and (f) a-terpineol concentrationsin white wine. White columns, wild ferments; black columns, inoculated ferments.
Varela et al. ‘Wild yeast fermentation’ character 241
© 2009 The Australian Wine Research Institute

the inoculated wines or that when genetically distinctyeasts with different SO2-producing capacities are presenttogether in mixed culture, the final concentration of SO2
is lower than that produced by individual strains inmonoculture. The latter physiological characteristic hasbeen observed for SO2 production by multi-strain culturesof S. cerevisiae (Grossmann et al. 1996). This modulatingeffect was also observed for several other metabolites,such as acetaldehyde, pyruvic acid and acetic acid, and isespecially evident when one of the strains present in amixed culture is characterised as a high-producer strainfor that metabolite in monoculture (see e.g. Mora et al.1990, Ciani and Ferraro 1998, Soden et al. 2000, Cheraitiet al. 2005).
No overall trends were found for wine pH, titratableacidity, alcohol, glycerol, ammonium ion, free aminonitrogen, yeast assimilable nitrogen (YAN) or hazeformed by the heat stability test, although several indi-vidual pairs of wines showed significant trends (data notshown). Several vineyard trends, that is, those associatedwith grape origin and irrespective of mode of inoculation,were also noted for wine density, specific gravity, alcohol,titratable acidity, glycerol, free and total SO2 and A420 nm(data not shown) as would be expected for grapes har-vested from different locations and at different maturities(range 210–250 g/L sugar).
Hydrogen sulphide, DMS and CS2 concentrationspeaked in different wines but did not correlate withresidual YAN concentrations in the respective wines(Figure 1). The concentration of H2S tended to be higherin the uninoculated compared with inoculated fermenta-tion wines, whereas no trend for DMS or CS2 was noted.One of the wine pairs showed a markedly higher con-centration of the two terpenes measured, linalool anda-terpineol, in the uninoculated but not in the inocul-ated fermentation wine (Figure 1e,f), however onlya-terpineol was detectably higher in some of the otheruninoculated fermentation wines.
Glycosidase activity has been described in a varietyof non-Saccharomyces yeasts (Rosi et al. 1994, Charoen-chai et al. 1997, Ferreira et al. 2001), which confers theability to release flavour-active compounds from odour-less precursors. This hydrolytic activity is considered tobe greater in non-Saccharomyces yeasts, and differences inglycosylated flavour precursors and terpenes betweenwines from inoculated and indigenous yeast fermenta-tions have been described (Zoecklein et al. 1997). Somenon-Saccharomyces yeasts have also been characterised ashigh H2S producers (Mendes-Ferreira et al. 2002) andcould potentially deliver wines with high H2S concen-trations when dominating wine fermentations. In thisstudy, however, we did not find any significant overalldifferences in glycosyl-glucose precursors, grape-derivedvolatile compounds, yeast volatile sulphur compounds,oak flavour compounds or acetaldehyde, betweeninoculated and uninoculated fermentation wines,although several pairs of wines showed clear differences(data not shown). Nonetheless, the concentrations ofhydrogen sulphide, DMS, CS2, linalool and a-terpineolwere found to be more associated with a particular
grape juice than to the mode of inoculation, indicatingthat these compounds, although likely to influence thesensory profile of these particular wines, do not contrib-ute to defining the ‘wild yeast fermentation’ character.
These data also suggest that fermentation in 3- to4-year-old barrels has no significant effect on wine com-position, an important source of variability when usingnewly toasted barrels. Fermentation temperature is alsowell known to affect wine chemical composition; there-fore, all ferments were treated in a similar manner tominimise the impact of fermentation temperature as avariable.
Of all the chemical analyses performed, only yeastfermentation products showed statistically significantdifferences between inoculation treatments (Table 1).The higher (fusel) alcohols 2-methylpropanol and2-methylbutanol, the acids 2-methylpropanoic and 2-methylbutanoic, and the ethyl esters ethyl 2-methyl-propanoate, ethyl decanoate and ethyl dodecanoateshowed significantly higher concentrations in uninocu-lated fermentation wines. Overall, wines made by unin-oculated fermentation presented higher concentrationsof higher alcohols and lower concentrations of acetateesters compared with the inoculated fermentation wines.Even though the concentrations of higher alcohols weregreater for uninoculated fermentations, they were belowthe threshold value (300 mg/L) considered by someauthors to be detrimental for wine quality (Rapp andMandery 1986), and therefore might contribute to theoverall sensory complexity of these wines. Increased con-centrations of higher alcohols have also been observed byothers in Chardonnay wines made with indigenous yeasts(Herjavec et al. 2003, Vilanova and Sieiro 2006).
It is interesting to note that the concentrations of2-methylpropanol, 2-methylpropanoic acid, 2-methyl-butanol and 2-methylbutanoic acid were significantlyhigher for indigenous yeast wines. These compounds areformed by three sequential reactions called the Ehrlichpathway; the first two compounds are synthesised fromvaline and the last two from isoleucine. This pathway isalso responsible for the formation of 3-methylbutanoland 3-methylbutanoic acid from leucine (Dickinson et al.1997, Eden et al. 2001). The greater proportion of higheralcohols are however known to be synthesised froma-keto acids, which are derived from glycolysis and des-tined for amino acids biosynthesis (Äyräpää 1971, Chen1978). These observations suggest that in uninoculatedfermentations, some non-Saccharomyces yeasts present avery active synthetic or Ehrlich pathway for valine andisoleucine metabolism and that this pathway is respon-sible for the high concentrations of higher alcohols. Thehigh capability and variability of different non-Saccharomyces species commonly found in uninoculatedfermentations to produce higher alcohols lends furthersupport to this explanation. Furthermore, investigationswith Saccharomyces species suggest that mixed cultures ofnon-Saccharomyces might also tend to produce wines withconcentrations of higher alcohols at the higher end of therange of the individual producer strains (Favale et al.2007).
242 ‘Wild yeast fermentation’ character Australian Journal of Grape and Wine Research 15, 238–248, 2009
© 2009 The Australian Wine Research Institute

Interestingly, some of the volatile compounds asso-ciated with sensory descriptors in Chardonnay wines,such as the alcohols 2-methylpropanol and2-methylbutanol, the ethyl esters ethyl 2-methyl pro-panoate, ethyl hexanoate, ethyl decanoate, the acetateester 3-methylbutyl acetate, and the acids hexanoic andoctanoic acids (Francis and Newton 2005, Smyth et al.2005), were not only significantly different but alsohelped to discriminate between inoculated and uninocu-lated fermentation wines. Indeed, the mode of inocula-
tion imparted different sensory characters in the pairs ofwines, especially in terms of aromatic complexity, asdescribed by informal sensory analysis (data notshown). Nevertheless, further work, such as a quantita-tive descriptive analysis, is needed to show the extent towhich these differences in chemical composition affectthe perceived aroma properties of these wines as theresult of inoculation treatments and to link specificsensory attributes to the ‘wild yeast fermentation’character.
Table 1. Yeast volatile fermentation products for wild and inoculated ferments.
Compound Wild Inoculated Significance
2-Methylpropanol (mg/L) 42 � 10† 20 � 3 ***
Butanol (mg/L) 1.4 � 1.1 1.0 � 0.3 ns
2-Methylbutanol (mg/L) 36 � 7 21 � 3 ***
3-Methylbutanol (mg/L) 13 � 3 12 � 2 ns
Hexanol (mg/L) 2.5 � 0.6 2.6 � 0.7 ns
2-Phenylethanol (mg/L) 30 � 13† 24 � 11† ns
Acetic acid (mg/L) 406 � 143† 320 � 129† ns
Propanoic acid (mg/L) 1.3 � 0.1 1.9 � 0.5 **
2-Methylpropanoic acid (mg/L) 767 � 331 452 � 92 *
Butanoic acid (mg/L) 1.8 � 0.8 1.8 � 1.1 ns
2-Methylbutanoic acid (mg/L) 5.2 � 2.9† 2.9 � 1.1 *
3-Methylbutanoic acid (mg/L) 203 � 130 279 � 69 ns
Hexanoic acid (mg/L) 4.1 � 0.4 5.1 � 1.2 *
Octanoic acid (mg/L) 7.0 � 0.8 8.1 � 1.3 *
Decanoic acid (mg/L) 3.2 � 0.7 2.9 � 0.6 ns
Ethyl acetate (mg/L) 85 � 22† 70 � 171† ns
Ethyl propanoate (mg/L) 204 � 42 382 � 92 ***
Ethyl 2-methylpropanoate (mg/L) 73 � 28† 43 � 9† **
Ethyl lactate (mg/L) 36 � 32† 24 � 15† ns
Ethyl butanoate (mg/L) 588 � 224† 506 � 114† ns
Ethyl 2-methylbutanoate (mg/L) 8.2 � 5.0† 7.1 � 1.8† ns
Ethyl 3-methylbutanoate (mg/L) 12 � 7† 14 � 5† ns
Ethyl hexanoate (mg/L) 1.0 � 0.1† 1.2 � 0.2† *
Ethyl octanoate (mg/L) 1.8 � 0.3† 1.8 � 0.3† ns
Ethyl decanoate (mg/L) 2.2 � 0.7† 1.2 � 0.4† ***
Ethyl dodecanoate (mg/L) 3.1 � 2.5† 0.5 � 0.5 **
2-Methylpropyl acetate (mg/L) 88 � 54 50 � 26 ns
2-Methylbutyl acetate (mg/L) 55 � 45 65 � 29 ns
3-Methylbutyl acetate (mg/L) 1.2 � 0.7† 2.0 � 0.7† *
Hexyl acetate (mg/L) 42 � 39 139 � 66 **
2-Phenylethyl acetate (mg/L) 115 � 98 174 � 194 ns
Higher alcohols (mg/L) 243 � 58 191 � 21 *
Volatile acids (mg/L)‡ 24 � 5 24 � 5 ns
Esters (mg/L)§ 85 � 46 112 � 28 ns
Ethyl esters (mg/L)§ 83 � 46 111 � 28 ns
Acetate esters (mg/L) 1.5 � 0.8 2.4 � 0.8 *
*, **, ***indicate significance at P < 0.05, P < 0.01, P < 0.001, respectively.†Indicates concentration larger than sensory threshold.‡Volatile acids do not include acetic acid.§Esters and ethyl esters do not include ethyl acetate and ethyl lactate.ns, not significant.
Varela et al. ‘Wild yeast fermentation’ character 243
© 2009 The Australian Wine Research Institute

Although sensory thresholds provide a rough estima-tion of the real contribution of each volatile compound tothe global aroma, they are generally used to evaluate thepotential sensory contribution of a given compound. Ofthe yeast volatile fermentation products measured in allwines, 17 compounds exhibited concentrations over theirreported sensory thresholds (Teranishi et al. 1966, Salo1970, Shinohara and Watanabe 1976, Simpson 1979,Étievant 1991, Guth 1997, Ferreira et al. 2000) in theinoculated fermentation wines. In contrast, 20 com-pounds showed concentrations higher than their sensorythresholds in wines from uninoculated fermentations.Compounds exhibiting concentrations higher than theirrespective sensory thresholds, irrespective of the mode ofinoculation, included ethyl 2-methylpropanoate (associ-ated with fruity characters) and ethyl decanoate (oily,nutlike characters), which was higher in uninoculatedfermentation wines; and 3-methylbutyl acetate (bananacharacters) and ethyl hexanoate (green apple characters),which were higher in inoculated fermentation wines. Thecompounds 2-methylpropanol (fusel, spirituous charac-ters), 2-methylbutanoic acid (cheesy, sweaty characters)and ethyl dodecanoate (soapy, estery characters) showedconcentrations over their sensory thresholds only in uni-noculated fermentation wines. The concentrations of2-methylpropanol and ethyl dodecanoate were, respec-tively, two and five times higher than in wines frominoculated fermentations (Table 1).
Wines made with indigenous yeasts included an addi-tional three volatile compounds not found in inoculatedyeast wines, which exceeded their respective sensorythresholds, indicating a potential role for these com-pounds in aroma complexity and in defining the ‘wildyeast fermentation’ character. Perception of volatile com-pounds is considerably affected by wine composition,which can both mask the odour impact of certain com-pounds present in concentrations above their sensorythresholds and favour the detection of other moleculespresent in concentrations below their detection thresh-olds (Ferreira et al. 2008). Therefore, more work isneeded to link 2-methylpropanol, 2-methylbutanoic acidand ethyl dodecanoate to the ‘wild yeast fermentation’character. This character is not necessarily linked to winequality. Some authors have suggested that better overallquality is achieved by using selected strains of S. cerevisiae(Romano et al. 1998), others have observed good qualitywines with indigenous yeasts (Herjavec et al. 2003, Vil-anova and Sieiro 2006), whereas some reports havedescribed very high or very low quality scores for wildyeast fermented Chardonnay and Riesling wines,depending on the taster (Egli et al. 1998). Nevertheless,atypical flavours not normally achieved with conven-tional S. cerevisiae create wines with new and interestingaroma profiles that differ from each other and frominoculated yeast wines. Even though 2-methylpropanol,2-methylbutanoic acid and ethyl dodecanoate are notnecessarily considered to be desirable odours in pureform, they might, in combination with other aroma com-pounds, contribute to the distinctive aroma of indigenousyeast wines.
PCA was used to reveal the compounds or measure-ments that differentiated best among the inoculationtreatments. Thus, only yeast volatile fermentation prod-ucts enabled a good discrimination between inoculatedand uninoculated fermentation wines (Figure 2a). Whenall yeast volatile fermentation products were included,the first principal component (PC1) accounted for 26% ofthe total variation, while PC2 and PC3 explained 21 and15% of the total variation, respectively. Of these, only thefirst two principal components were statistically signifi-cant in explaining the differences between the pairs ofwines associated with the yeast volatile fermentationproducts. Discriminant analysis for the first two principalcomponents showed that only one sample (wine fer-mented with D47 and CY3079) was not classified cor-rectly by yeast strain; this is most likely due to the lownumber of D47/CY3079 samples analysed.
Positive loadings for PC1 were related to ethyl pro-panoate, hexyl acetate and propanoic acid, whereas2-methylbutanol, 2-methylpropanol and 2-methylpro-panoic acid showed negative loadings. Loadings for PC2were characterised by ethyl hexanoate, hexanoic acid,3-methylbutanoic acid and hexanol with positive values,while ethyl dodecanoate and 2-phenylethyl acetateexhibited negative values (Figure 2b). Wines fermentedwith the strain EC1118 formed a clear cluster, which wasassociated with the esters ethyl propanoate, ethyl hex-anoate and 3-methylbutyl acetate and with propanoic,hexanoic and octanoic acids. The fermentation inoculatedwith the strains D47 and CY3079 was located in a differ-ent quadrant of the PCA plot and was most associatedwith 2-phenylethyl acetate. Conversely, the uninoculatedfermentation wines showed a different pattern by beingwidely scattered in the PCA plot and none were closelyassociated with any of the inoculated fermentation wines(Figure 2a).
By comparison with inoculated fermentation wines,wild yeast fermentation produced wines with high vari-ability in the composition of volatile compounds thatcontribute to wine aroma. A similar discriminationbetween inoculated and wild yeast fermented wines hasbeen observed previously considering only a few alcoholsand esters (Vilanova and Sieiro 2006). Metabolic interac-tions between yeast species and strains, which deliver adifferent wine profile when yeasts are grown together ascompared with the profile of wine made by blending thecorresponding monocultures (Howell et al. 2006), mightbe responsible for the great variability in volatile compo-sition observed for wild fermented wines. This variabilityis further increased by yeast–must interactions especiallywith musts obtained from different regions (Zamuz andVilanova 2006).
This work should be extended by analysis of yeastpopulations to establish a link with chemical composi-tion. In the present study, although yeast populationswere not studied, the wine composition data of Table 1and Figure 2 clearly show that the wild yeast popula-tions induced very different and diverse chemical com-position from each other and from the inoculated yeast,as is evident from the tight clustering shown by the
244 ‘Wild yeast fermentation’ character Australian Journal of Grape and Wine Research 15, 238–248, 2009
© 2009 The Australian Wine Research Institute
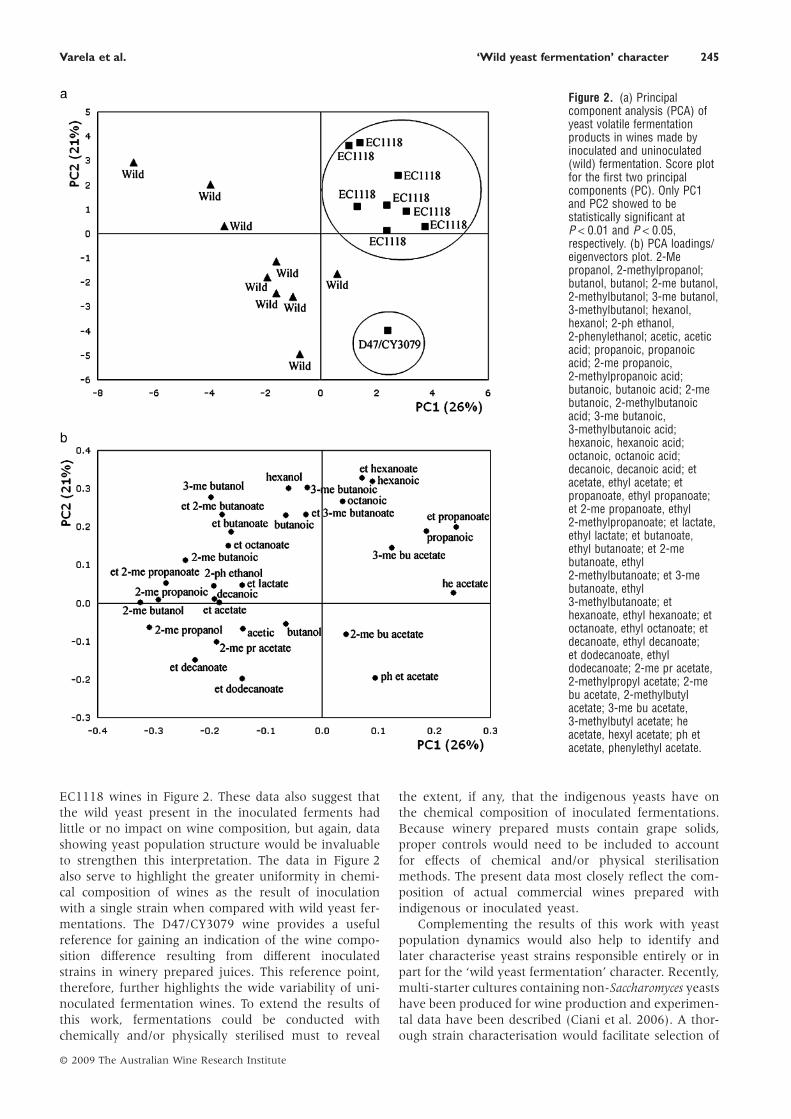
EC1118 wines in Figure 2. These data also suggest thatthe wild yeast present in the inoculated ferments hadlittle or no impact on wine composition, but again, datashowing yeast population structure would be invaluableto strengthen this interpretation. The data in Figure 2also serve to highlight the greater uniformity in chemi-cal composition of wines as the result of inoculationwith a single strain when compared with wild yeast fer-mentations. The D47/CY3079 wine provides a usefulreference for gaining an indication of the wine compo-sition difference resulting from different inoculatedstrains in winery prepared juices. This reference point,therefore, further highlights the wide variability of uni-noculated fermentation wines. To extend the results ofthis work, fermentations could be conducted withchemically and/or physically sterilised must to reveal
the extent, if any, that the indigenous yeasts have onthe chemical composition of inoculated fermentations.Because winery prepared musts contain grape solids,proper controls would need to be included to accountfor effects of chemical and/or physical sterilisationmethods. The present data most closely reflect the com-position of actual commercial wines prepared withindigenous or inoculated yeast.
Complementing the results of this work with yeastpopulation dynamics would also help to identify andlater characterise yeast strains responsible entirely or inpart for the ‘wild yeast fermentation’ character. Recently,multi-starter cultures containing non-Saccharomyces yeastshave been produced for wine production and experimen-tal data have been described (Ciani et al. 2006). A thor-ough strain characterisation would facilitate selection of
Figure 2. (a) Principalcomponent analysis (PCA) ofyeast volatile fermentationproducts in wines made byinoculated and uninoculated(wild) fermentation. Score plotfor the first two principalcomponents (PC). Only PC1and PC2 showed to bestatistically significant atP < 0.01 and P < 0.05,respectively. (b) PCA loadings/eigenvectors plot. 2-Mepropanol, 2-methylpropanol;butanol, butanol; 2-me butanol,2-methylbutanol; 3-me butanol,3-methylbutanol; hexanol,hexanol; 2-ph ethanol,2-phenylethanol; acetic, aceticacid; propanoic, propanoicacid; 2-me propanoic,2-methylpropanoic acid;butanoic, butanoic acid; 2-mebutanoic, 2-methylbutanoicacid; 3-me butanoic,3-methylbutanoic acid;hexanoic, hexanoic acid;octanoic, octanoic acid;decanoic, decanoic acid; etacetate, ethyl acetate; etpropanoate, ethyl propanoate;et 2-me propanoate, ethyl2-methylpropanoate; et lactate,ethyl lactate; et butanoate,ethyl butanoate; et 2-mebutanoate, ethyl2-methylbutanoate; et 3-mebutanoate, ethyl3-methylbutanoate; ethexanoate, ethyl hexanoate; etoctanoate, ethyl octanoate; etdecanoate, ethyl decanoate;et dodecanoate, ethyldodecanoate; 2-me pr acetate,2-methylpropyl acetate; 2-mebu acetate, 2-methylbutylacetate; 3-me bu acetate,3-methylbutyl acetate; heacetate, hexyl acetate; ph etacetate, phenylethyl acetate.
Varela et al. ‘Wild yeast fermentation’ character 245
© 2009 The Australian Wine Research Institute

relevant yeast strains responsible for contributing to the‘wild yeast fermentation’ character. Because yeast volatilefermentation products differentiate wines made withindigenous yeasts, diverse fermentation strategies can betrialed for the more reliable inoculated ferments toachieve similar attributes to those obtained by spontane-ous fermentation.
The observation that the wines made with indigenousyeasts in one of the wineries studied had widely differingprofiles of volatile compounds strongly argues againstcontamination of the juices by a dominant winery resi-dent yeast or yeasts. On the contrary, the highly variedprofiles suggest that populations of yeast species andstrains were highly variable between juice batches, as hasbeen found by others (Jolly et al. 2003, Clemente-Jimenez et al. 2004, Mercado et al. 2007). Indeed, thesedata might represent a vineyard effect being expressedthrough the particular population of yeasts present in theuninoculated fermentations; further studies would beneeded to confirm whether the broad aromatic profilewould be retained over successive vintages, giving a‘microbiological terroir’ as suggested by several recentstudies (Romano et al. 2003a,b).
ConclusionsAs the marketplace for wines becomes more competitive,winemakers are developing and evaluating a wide rangeof tools and practices for modulating wine flavour andstyle. The indigenous yeast flora is known to induce asignificant and usually favourable effect on wine flavourand, consequently, has been exploited to manage winestyle. However, the ‘wild yeast fermentation’ character ofwines produced by indigenous yeasts is not reliable in thehands of many winemakers, and, in addition, uninocu-lated fermentations are associated with a higher risk ofslow and/or stuck fermentations and off-flavours forma-tion (Bisson and Kunkee 1993, Jolly et al. 2006).
In Chardonnay wine, yeast-derived volatile fermenta-tion products have been shown to constitute a significantdifference between inoculated and wild ferments, provid-ing a chemical basis for the ‘wild yeast fermentation’character. Inoculated wines were associated with particu-lar esters forming a distinct cluster by PCA, whereas ‘wild’yeast fermented wines showed high variability in volatilecompounds that contribute to wine aroma; nevertheless,uninoculated wines contained several yeast volatilecompounds that might contribute to its unique sensorycharacteristics.
AcknowledgementsWe thank The Yalumba Wine Company and OrlandoWines for providing the wines studied in this work. Thisproject work carried out at the Australian Wine ResearchInstitute (AWRI) was financially supported by Australia’sgrapegrowers and winemakers through their investmentbody, the Grape and Wine Research and DevelopmentCorporation, with matching funding from the AustralianGovernment. The AWRI is a member of the Wine Inno-vation Cluster located in Adelaide.
ReferencesAllen, M. (2008) Green and mean aromas. Proceedings of the
Thirteenth Australian Wine Industry Technical Conference, Ade-laide, Australia (AWITC) (Australian Wine Industry TechnicalConference Inc.: Adelaide) pp. 56–64.
Äyräpää, T. (1971) Biosynthetic formation of higher alcohols byyeast. Dependence on the nitrogen nutrient level of the medium.Journal of the Institute of Brewing 77, 266–275.
Bell, S.J. and Henschke, P.A. (2005) Implications of nitrogen nutri-tion for grapes, fermentation and wine. Australian Journal ofGrape and Wine Research 11, 242–295.
Bisson, L.F. and Kunkee, R.E. (1993) Microbial interactions duringwine production. In: Mixed cultures in biotechnology. Eds. J.G.Zeikus and E.A. Johnson (McGraw-Hill: New York) pp. 37–68.
Brajkovich, M.J. (2008) Back to the future – A traditional approachto modern winemaking. Proceedings of the Thirteenth AustralianWine Industry Technical Conference, Adelaide, Australia(Australian Wine Industry Technical Conference Inc.: Adelaide)pp. 128–133.
Charoenchai, C., Fleet, G., Henschke, P. and Todd, B. (1997)Screen of non-Saccharomyces wine yeasts for the presence of extra-cellular hydrolytic enzymes. Australian Journal of Grape andWine Research 3, 2–8.
Chen, E.C. (1978) The relative contribution of Ehrlich and biosyn-thetic pathways to the formation of fusel alcohols. Journal of theAmerican Society of Brewing Chemists 36, 39-43.
Cheraiti, N., Guezenec, S. and Salmon, J.M. (2005) Redox inter-actions between Saccharomyces cerevisiae and Saccharomyces uvarumin mixed culture under enological conditions. Applied and Envi-ronmental Microbiology 71, 255–260.
Ciani, M. and Ferraro, L. (1998) Combined use of immobilizedCandida stellata cells and Saccharomyces cerevisiae to improve thequality of wines. Journal of Applied Microbiology 85, 247–254.
Ciani, M. and Maccarelli, F. (1998) Oenological properties of non-Saccharomyces yeasts associated with winemaking. World Journalof Microbiology & Biotechnology 14, 199–203.
Ciani, M., Beco, L. and Comitini, F. (2006) Fermentation behaviourand metabolic interactions of multistarter wine yeast fermenta-tions. International Journal of Food Microbiology 108, 239–245.
Clemente-Jimenez, J.M., Mingorance-Cazorla, L., Martinez-Rodriguez, S., Heras-Vazquez, F.J.L. and Rodriguez-Vico, F.(2004) Molecular characterization and oenological properties ofwine yeasts isolated during spontaneous fermentation of six vari-eties of grape must. Food Microbiology 21, 149–155.
Combina, M., Elia, A., Mercado, L., Catania, C., Ganga, A. andMartinez, C. (2005) Dynamics of indigenous yeast populationsduring spontaneous fermentation of wines from Mendoza, Argen-tina. International Journal of Food Microbiology 99, 237–243.
Darriet, P., Boidron, J. and Dubourdieu, D. (1988) Hydrolysis ofthe terpenic heterosides of small muscat grapes using the periplas-mic enzymes of Saccharomyces cerevisiae. Connaissance de laVigne et du Vin 22, 189–196.
Delfini, C., Cocito, C., Bonino, M., Schellino, R., Gaiaj, P. andBaiocchi, C. (2001) Definitive evidence for the actual contribu-tion of yeast in the transformation of neutral precursors of grapearomas. Journal of Agricultural and Food Chemistry 49, 5397–5408.
Dickinson, J.R., Lanterman, M.M., Danner, D.J., Pearson, B.M.,Sanz, P., Harrison, S.J. and Hewlins, M.J.E. (1997) A 13C nuclearmagnetic resonance investigation of the metabolism of leucine toisoamyl alcohol in Saccharomyces cerevisiae. Journal of BiologicalChemistry 272, 26871–26878.
Dukes, B.C. and Butzke, C.E. (1998) Rapid determination ofprimary amino acids in grape juice using an o-phthaldialdehyde/N-acetyl-L-cysteine spectrophotometric assay. American Journalof Enology and Viticulture 49, 125–134.
Eden, A., Van Nedervelde, L., Drukker, M., Benvenisty, N. andDebourg, A. (2001) Involvement of branched-chain amino acidaminotransferases in the production of fusel alcohols during fer-mentation in yeast. Applied Microbiology and Biotechnology 55,296–300.
246 ‘Wild yeast fermentation’ character Australian Journal of Grape and Wine Research 15, 238–248, 2009
© 2009 The Australian Wine Research Institute

Egli, C.M., Edinger, W.D., Mitrakul, C.M. and Henick-Kling, T.(1998) Dynamics of indigenous and inoculated yeast populationsand their effect on the sensory character of Riesling and Chardon-nay wines. Journal of Applied Microbiology 85, 779–789.
Epifanio, S., Gutierrez, A., Santamaria, M. and Lopez, R. (1999)The influence of enological practices on the selection of wild yeaststrains in spontaneous fermentation. American Journal ofEnology and Viticulture 50, 219–224.
Étievant, P. (1991) Wine. In: Volatile compounds in foods andbeverages. Ed. H. Maarse (Marcel Dekker: New York) pp. 483–546.
Favale, S., Pietromarchi, P. and Ciolfi, G. (2007) Metabolic activityand interactions between two strains, Saccharomyces cerevisiae r.f.bayanus (SBC2) and Saccharomyces cerevisiae r.f. uvarum (S6u), inpure and mixed culture fermentations. Vitis 46, 39–43.
Ferreira, A.C.S., Lopez, R. and Cacho, J.F. (2000) Quantitativedetermination of the odorants of young red wines from differentgrape varieties. Journal of the Science of Food and Agriculture 80,1659–1667.
Ferreira, A.M., Climaco, M.C. and Faia, A.M. (2001) The role ofnon-Saccharomyces species in releasing glycosidic bound fraction ofgrape aroma components – A preliminary study. Journal ofApplied Microbiology 91, 67–71.
Ferreira, V., Escudero, A., Campo, E. and Cacho, J. (2008) Thechemical foundations of wine aroma – A role game aiming at winequality, personality and varietal expression. Proceedings of theThirteenth Australian Wine Industry Technical Conference, Ade-laide, Australia (Australian Wine Industry Technical ConferenceInc.: Adelaide) pp. 142–150.
Fleet, G. (2003) Yeast interactions and wine flavour. InternationalJournal of Food Microbiology 86, 11–22.
Fleet, G. and Heard, G. (1993) Yeasts – Growth during fermenta-tion. Taylor & Francis Inc.: New York.
Fleet, G., Prakitchaiwattana, C., Beh, S. and Heard, G. (2002) Theyeast ecology of wine grapes. In: Biodiversity and biotechnologyof wine yeast. Ed. M. Ciani (Research Signpost: Kerala) pp. 1–17.
Francis, I.L. and Newton, J.L. (2005) Determining wine aromafrom compositional data. Australian Journal of Grape and WineResearch 11, 114–126.
Goode, J. (2005) Microbes and wine: Yeasts and lactic-acid bacte-ria. In: Wine science: the application of science in winemaking.(Mitchell Beazley: London) pp. 129–135.
Granchi, L., Ganucci, D., Messini, A., Rosellini, D. and Vincenzini,M. (1998) Dynamics of yeast populations during the early stagesof natural fermentations for the production of Brunello di Mon-talcino wines. Food Technology and Biotechnology 36, 313–318.
Grossmann, M., Muno, H., Lambert, C., Fries, M. and Rapp, A.(1996) Influence of oligo-strain yeast cultures on wine quality.Proceedings of the Eleventh International Oenological Sympo-sium, Sopron, Hungary (International Association for WineryTechnology and Management: Breisach, Germany) pp. 71–82.
Guth, H. (1997) Quantitation and sensory studies of characterimpact odorants of different white wine varieties. Journal of Agri-cultural and Food Chemistry 45, 3027–3032.
Gutierrez, A.R., Santamaria, P., Epifanio, S., Garijo, P. and Lopez,R. (1999) Ecology of spontaneous fermentation in one wineryduring 5 consecutive years. Letters in Applied Microbiology 29,411–415.
Heard, G. (1999) Novel yeasts in winemaking – Looking to thefuture. Food Australia 51, 347–352.
Heard, G.M. and Fleet, G.H. (1985) Growth of natural yeast floraduring the fermentation of inoculated wines. Applied and Envi-ronmental Microbiology 50, 727–728.
Henick-Kling, T., Edinger, W., Daniel, P. and Monk, P. (1998)Selective effects of sulfur dioxide and yeast starter culture additionon indigenous yeast populations and sensory characteristics ofwine. Journal of Applied Microbiology 84, 865–876.
Herjavec, S., Podgorski, V., Redzepovic, S. and Mirosevic, N.(2003) The influence of some commercial Saccharomyces cerevisiaestrains on the quality of Chardonnay wines. Food Technology andBiotechnology 41, 77–81.
Herraiz, T., Reglero, G., Herraiz, M., Martin-Alvarez, P.J. andCabezudo, M.D. (1990) The influence of the yeast and type ofculture on the volatile composition of wines fermented withoutsulfur dioxide. American Journal of Enology and Viticulture 41,313–318.
Hierro, N., Gonzalez, A., Mas, A. and Guillamon, J. (2006) Diver-sity and evolution of non-Saccharomyces yeast populations duringwine fermentation: Effect of grape ripeness and cold maceration.FEMS Yeast Research 6, 102–111.
Howell, K.S., Cozzolino, D., Bartowsky, E.J., Fleet, G.H. andHenschke, P.A. (2006) Metabolic profiling as a tool for revealingSaccharomyces interactions during wine fermentation. FEMS YeastResearch 6, 91–101.
Jemec, K.P. and Raspor, P. (2005) Initial Saccharomyces cerevisiaeconcentration in single or composite cultures dictates bioprocesskinetics. Food Microbiology 22, 293–300.
Jolly, N., Augustyn, O. and Pretorius, I.S. (2003) The occurrenceof non-Saccharomyces cerevisiae yeast species over three vintages infour vineyards and grape musts from four production regions ofthe Western Cape, South Africa. South African Journal of Enologyand Viticulture 24, 35–42.
Jolly, N., Augustyn, O. and Pretorius, I.S. (2006) The role and useof non-Saccharomyces yeasts in wine production. South AfricanJournal of Enology and Viticulture 27, 15–39.
Lambrechts, M.G. and Pretorius, I.S. (2000) Yeast and its impor-tance to wine aroma. South African Journal of Enology andViticulture 21, 97–129.
Lema, C., Garcia-Jares, C., Orriols, I. and Angulo, L. (1996)Contribution of Saccharomyces and non-Saccharomyces populationsto the production of some components of Albarino wine aroma.American Journal of Enology and Viticulture 47, 206–216.
Loscos, N., Hernandez-Orte, P., Cacho, J. and Ferreira, V. (2007)Release and formation of varietal aroma compounds during alco-holic fermentation from nonfloral grape odorless flavor precursorsfractions. Journal of Agricultural and Food Chemistry 55, 6674–6684.
Lurton, L., Snakkers, G., Roulland, C., Galy, B. and Versavaud, A.(1995) Influence of the fermentation yeast strain on the compo-sition of wine spirits. Journal of the Science of Food and Agricul-ture 67, 485–491.
Mendes-Ferreira, A., Mendes-Faia, A. and Leao, C. (2002) Surveyof hydrogen sulphide production by wine yeasts. Journal of FoodProtection 65, 1033–1037.
Mercado, L., Dalcero, A., Masuelli, R. and Combina, M. (2007)Diversity of Saccharomyces strains on grapes and winery surfaces:Analysis of their contribution to fermentative flora of Malbecwine from Mendoza (Argentina) during two consecutive years.Food Microbiology 24, 403–412.
Mora, J., Barbas, J.I. and Mulet, A. (1990) Growth of yeast speciesduring the fermentation of musts inoculated with Kluyveromycesthermotolerans and Saccharomyces cerevisiae. American Journal ofEnology and Viticulture 41, 156–159.
Moreno, J., Millan, C., Ortega, J. and Medina, M. (1991) Ana-lytical differentiation of wine fermentations using pure and mixedyeast cultures. Journal of Industrial Microbiology 7, 181–189.
Nissen, T., Schulze, U., Nielsen, J. and Villadsen, J. (1997) Fluxdistributions in anaerobic, glucose-limited continuous cultures ofSaccharomyces cerevisiae. Microbiology 143, 203–218.
Nurgel, C., Erten, H., Canbas, A., Cabaroglu, T. and Selli, S.(2002a) Influence of Saccharomyces cerevisiae strains on fermenta-tion and flavor compounds of white wines made from cv. Emirgrown in Central Anatolia, Turkey. Journal of Industrial Micro-biology & Biotechnology 29, 28–33.
Nurgel, C., Erten, H., Canbas, A., Cabaroglu, T. and Selli, S.(2002b) Contribution by Saccharomyces cerevisiae yeasts to fermen-tation and flavour compounds in wines from cv. Kalecik karasigrape. Journal of the Institute of Brewing 108, 68–72.
Pedersen, D.S., Capone, D.L., Skouroumounis, G.K., Pollnitz, A.P.and Sefton, M.A. (2003) Quantitative analysis of geraniol, nerol,linalool, and alpha-terpineol in wine. Analytical and BioanalyticalChemistry 375, 517–522.
Varela et al. ‘Wild yeast fermentation’ character 247
© 2009 The Australian Wine Research Institute

Pocock, K.F. and Waters, E.J. (2006) Protein haze in bottled whitewines: How well do stability tests and bentonite fining trialspredict haze formation during storage and transport? AustralianJournal of Grape and Wine Research 12, 212–220.
Pollnitz, A.P., Capone, D.L., Caldersmith, M.C. and Sefton, M.A.(2003) The effect of various wine bottle closures and fining agentson flavour and aroma compounds in wine. Grapegrowing at theedge; Managing the wine business; Impacts on wine flavour, Ade-laide, Australia (Australian Society of Viticulture and Oenology:Adelaide) pp. 59–63.
Pollnitz, A.P., Pardon, K.H., Sykes, M. and Sefton, M.A. (2004)The effects of sample preparation and gas chromatograph injec-tion techniques on the accuracy of measuring guaiacol,4-methylguaiacol and other volatile oak compounds in oakextracts by stable isotope dilution analyses. Journal of Agricul-tural and Food Chemistry 52, 3244–3252.
Querol, A. and Ramon, D. (1996) The application of moleculartechniques in wine microbiology. Trends in Food Science & Tech-nology 7, 73–78.
Ramey, D. (1996) Low input winemaking – Let nature do thework. Proceedings of the Ninth Australian Wine IndustryTechnical Conference, Adelaide, Australia (Winetitles: Adelaide)pp. 26–29.
Rankine, B. and Pocock, K. (1970) Alkalimetric determination ofsulphur dioxide in wine. Australian Wine, Brewing & SpiritReview 88, 40–44.
Rapp, A. and Mandery, H. (1986) Wine aroma. Experientia 42,873–884.
Renouf, V. and Lonvaud-Funel, A. (2007) Development of anenrichment medium to detect Dekkera/Brettanomyces bruxellensis, aspoilage wine yeast, on the surface of grape berries. Microbiologi-cal Research 162, 154–167.
Renouf, V., Falcou, M., Miot-Sertier, C., Perello, M.C., De Revel, G.and Lonvaud-Funel, A. (2006) Interactions between Brettanomy-ces bruxellensis and other yeast species during the initial stages ofwinemaking. Journal of Applied Microbiology 100, 1208–1219.
Rojas, V., Gil, J.V., Pinaga, F. and Manzanares, P. (2003) Acetateester formation in wine by mixed cultures in laboratory fermen-tations. International Journal of Food Microbiology 86, 181–188.
Romano, P., Suzzi, G., Domizio, P. and Fatichenti, F. (1997)Secondary products formation as a tool for discriminatingnon-Saccharomyces wine strains. Antonie Van Leeuwenhoek Inter-national Journal of General and Molecular Microbiology 71, 239–242.
Romano, P., Monteleone, E., Paraggio, M., Marchese, R., Caporale,G. and Carlucci, A. (1998) A methodological approach to theselection of Saccharomyces cerevisiae wine strains. Food Technologyand Biotechnology 36, 69–74.
Romano, P., Caruso, M., Capece, A., Lipani, G., Paraggio, M. andFiore, C. (2003a) Metabolic diversity of Saccharomyces cerevisiaestrains from spontaneously fermented grape musts. World Journalof Microbiology & Biotechnology 19, 311–315.
Romano, P., Fiore, C., Paraggio, M., Caruso, M. and Capece, A.(2003b) Function of yeast species and strains in wine flavour.International Journal of Food Microbiology 86, 169–180.
Rosi, I., Vinella, M. and Domizio, P. (1994) Characterization ofbeta-glucosidase activity in yeasts of enological origin. Journal ofApplied Bacteriology 77, 519–527.
Salo, P. (1970) Determining the odor thresholds for some com-pounds in alcoholic beverages. Journal of Food Science 35,95–99.
Shinohara, T. and Watanabe, M. (1976) Gas chromatographicanalysis of higher alcohols and ethyl acetate in table wines. Agri-cultural and Biological Chemistry 40, 2475–2477.
Siebert, T.E., Smyth, H.E., Capone, D.L., Neuwohner, C., Pardon,K.H., Skouroumounis, G.K., Herderich, M., Sefton, M.A. andPollnitz, A.P. (2005) Stable isotope dilution analysis of wine fer-mentation products by HS-SPME-GC-MS. Analytical and Bioana-lytical Chemistry 381, 937–947.
Simpson, R.F. (1979) Some important aroma compounds of whitewine. Food Technology in Australia 31, 516–522.
Skouroumounis, G.K., Kwiatkowski, M., Sefton, M.A., Gawel, R.and Waters, E.J. (2003) In situ measurement of white wineabsorbance in clear and in coloured bottles using a modifiedlaboratory spectrophotometer. Australian Journal of Grape andWine Research 9, 138–147.
Smyth, H., Cozzolino, D., Herderich, M.J., Sefton, M.A. andFrancis, I.L. (2005) Relating volatile composition to wine aroma:identification of key aroma compounds in Australian white wines.Proceedings of the Twelfth Australian Wine Industry TechnicalConference, Melbourne, Australia (Australian Wine IndustryTechnical Conference Inc.: Adelaide) pp. 31–33.
Soden, A., Francis, I.L., Oakey, H. and Henschke, P.A. (2000)Effects of co-fermentation with Candida stellata and Saccharomycescerevisiae on the aroma and composition of Chardonnay wine.Australian Journal of Grape and Wine Research 6, 21–30.
Strauss, C., Wilson, B., Gooley, P. and Williams, P. (1986) Role ofmonoterpenes in grape and wine flavour. ACS Symposium Series317, 222–242.
Sturm, J., Grossmann, M. and Schnell, S. (2006) Influence ofgrape treatment on the wine yeast populations isolated fromspontaneous fermentations. Journal of Applied Microbiology 101,1241–1248.
Swiegers, J.H., Bartowsky, E.J., Henschke, P.A. and Pretorius, I.S.(2005) Yeast and bacterial modulation of wine aroma and flavour.Australian Journal of Grape and Wine Research 11, 139–173.
Teranishi, R., Flath, R., Guadagni, D., Lundin, R., Mon, T. andStevens, K. (1966) Gas chromatographic, infrared, proton mag-netic resonance, mass spectral, and threshold analyses of all pentylacetates. Journal of Agricultural and Food Chemistry 14, 253–262.
Torija, M.J., Rozes, N., Poblet, M., Guillamon, J.M. and Mas, A.(2001) Yeast population dynamics in spontaneous fermentations:Comparison between two different wine-producing areas over aperiod of three years. Antonie Van Leeuwenhoek InternationalJournal of General and Molecular Microbiology 79, 345–352.
Ugliano, M., Bartowsky, E.J., McCarthy, J., Moio, L. andHenschke, P.A. (2006) Hydrolysis and transformation of grapeglycosidically bound volatile compounds during fermentationwith three Saccharomyces yeast strains. Journal of Agriculturaland Food Chemistry 54, 6322–6331.
Ugliano, M. and Henschke, P.A. (2009) Yeasts and wine flavour.In: Wine Chemistry and Biochemistry. Eds M.V. Moreno-Arribasand M.C. Polo (Springer Verlag: New York) pp. 313–392.
Vilanova, M. and Sieiro, C. (2006) Contribution by Saccharomycescerevisiae yeast to fermentative flavour compounds in wines fromcv. Albarino. Journal of Industrial Microbiology & Biotechnology33, 929–933.
Williams, P.J., Cynkar, W., Francis, I.L., Gray, J.D., Iland, P.G. andCoombe, B.G. (1995) Quantification of glycosides in grapes,juices, and wines through a determination of glycosyl glucose.Journal of Agricultural and Food Chemistry 43, 121–128.
Zamuz, S. and Vilanova, M. (2006) Volatile compounds afterspontaneous fermentation of musts from Vitis vinifera cv. Albarinograpes cultivated in different origins from Rias Baixas AOC, Spain.Flavour and Fragrance Journal 21, 743–748.
Zironi, R., Romano, P., Suzzi, G., Battistutta, F. and Comi, G. (1993)Volatile metabolites produced in wine by mixed and sequentialcultures of Hanseniaspora guilliermondii or Kloeckera Apiculataand Saccharomyces cerevisiae. Biotechnology Letters 15, 235–238.
Zoecklein, B., Marcy, J., Williams, J. and Jasinski, Y. (1997) Effectof native yeasts and selected strains of Saccharomyces cerevisiae onglucosyl glucose, potential volatile terpenes, and selected agly-cones of white Riesling (Vitis vinifera L.) wines. Journal of FoodComposition and Analysis 10, 55–65.
Manuscript received: 16 September 2008
Revised manuscript received: 16 January 2009
Accepted: 5 February 2009
248 ‘Wild yeast fermentation’ character Australian Journal of Grape and Wine Research 15, 238–248, 2009
© 2009 The Australian Wine Research Institute
Copyright © 2022 FDOKUMEN




















