
Diffusion Tensor Imaging of Cerebral White Matter: A Pictorial Review of Physics, Fiber Tract...
35
Diffusion Tensor Imaging of Cerebral White Matter Integrity in Cognitive Aging David J. Madden a,b,* , Ilana J. Bennett c , Agnieszka Burzynska d , Guy G. Potter b , Nan-kuei Chen a,e , and Allen W. Song a,e a Brain Imaging and Analysis Center, Duke University Medical Center, Durham, NC 27710 USA b Department of Psychiatry and Behavioral Sciences, Duke University Medical Center, Durham, NC 27710 USA c Center for Brain Health, School of Behavioral and Social Sciences, University of Texas at Dallas, Dallas, TX 75235 USA d Max Planck Institute for Human Development, Lentzeallee 94 D-14195, Berlin, Germany e Department of Radiology, Duke University Medical Center, Durham, NC 27710 USA Abstract In this article we review recent research on diffusion tensor imaging (DTI) of white matter (WM) integrity and the implications for age-related differences in cognition. Neurobiological mechanisms defined from DTI analyses suggest that a primary dimension of age-related decline in WM is a decline in the structural integrity of myelin, particularly in brain regions that myelinate later developmentally. Research integrating behavioral measures with DTI indicates that WM integrity supports the communication among cortical networks, particularly those involving executive function, perceptual speed, and memory (i.e., fluid cognition). In the absence of significant disease, age shares a substantial portion of the variance associated with the relation between WM integrity and fluid cognition. Current data are consistent with one model in which age-related decline in WM integrity contributes to a decreased efficiency of communication among networks for fluid cognitive abilities. Neurocognitive disorders for which older adults are at risk, such as depression, further modulate the relation between WM and cognition, in ways that are not as yet entirely clear. Developments in DTI technology are providing new insight into both the neurobiological mechanisms of aging WM and the potential contribution of DTI to understanding functional measures of brain activity. Keywords Magnetic resonance imaging; Brain; Behavior; Adult development; Neuroaxonal damage © 2011 Elsevier B.V. All rights reserved. Correspondence to: David J. Madden, Ph.D., Brain Imaging and Analysis Center, Box 2737, Hock Plaza, Duke University Medical Center, Durham, NC 27710 USA, [email protected], Ph: (919) 681-9345, FAX: (919) 681-7033. Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. NIH Public Access Author Manuscript Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1. Published in final edited form as: Biochim Biophys Acta. 2012 March ; 1822(3): 386–400. doi:10.1016/j.bbadis.2011.08.003. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
Transcript of Diffusion Tensor Imaging of Cerebral White Matter: A Pictorial Review of Physics, Fiber Tract...

Diffusion Tensor Imaging of Cerebral White Matter Integrity inCognitive Aging
David J. Maddena,b,*, Ilana J. Bennettc, Agnieszka Burzynskad, Guy G. Potterb, Nan-kueiChena,e, and Allen W. Songa,e
aBrain Imaging and Analysis Center, Duke University Medical Center, Durham, NC 27710 USAbDepartment of Psychiatry and Behavioral Sciences, Duke University Medical Center, Durham,NC 27710 USAcCenter for Brain Health, School of Behavioral and Social Sciences, University of Texas at Dallas,Dallas, TX 75235 USAdMax Planck Institute for Human Development, Lentzeallee 94 D-14195, Berlin, GermanyeDepartment of Radiology, Duke University Medical Center, Durham, NC 27710 USA
AbstractIn this article we review recent research on diffusion tensor imaging (DTI) of white matter (WM)integrity and the implications for age-related differences in cognition. Neurobiologicalmechanisms defined from DTI analyses suggest that a primary dimension of age-related decline inWM is a decline in the structural integrity of myelin, particularly in brain regions that myelinatelater developmentally. Research integrating behavioral measures with DTI indicates that WMintegrity supports the communication among cortical networks, particularly those involvingexecutive function, perceptual speed, and memory (i.e., fluid cognition). In the absence ofsignificant disease, age shares a substantial portion of the variance associated with the relationbetween WM integrity and fluid cognition. Current data are consistent with one model in whichage-related decline in WM integrity contributes to a decreased efficiency of communicationamong networks for fluid cognitive abilities. Neurocognitive disorders for which older adults areat risk, such as depression, further modulate the relation between WM and cognition, in ways thatare not as yet entirely clear. Developments in DTI technology are providing new insight into boththe neurobiological mechanisms of aging WM and the potential contribution of DTI tounderstanding functional measures of brain activity.
KeywordsMagnetic resonance imaging; Brain; Behavior; Adult development; Neuroaxonal damage
© 2011 Elsevier B.V. All rights reserved.Correspondence to: David J. Madden, Ph.D., Brain Imaging and Analysis Center, Box 2737, Hock Plaza, Duke University MedicalCenter, Durham, NC 27710 USA, [email protected], Ph: (919) 681-9345, FAX: (919) 681-7033.Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to ourcustomers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review ofthe resulting proof before it is published in its final citable form. Please note that during the production process errors may bediscovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
NIH Public AccessAuthor ManuscriptBiochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
Published in final edited form as:Biochim Biophys Acta. 2012 March ; 1822(3): 386–400. doi:10.1016/j.bbadis.2011.08.003.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

1. IntroductionThe recent decade has seen a rapidly emerging body of scientific investigation in magneticresonance imaging (MRI) of the brain and cognition, including the changes associated withhuman aging. The majority of cognitive and behavioral investigations have focused on eitherstructural MRI measures of brain volume and pathology [1–5] or functional MRI (fMRI)measures of the blood-oxygen-level-dependent (BOLD) changes in cortical gray matter(GM) [6–8]. Research with another form of MRI, diffusion tensor imaging (DTI), hasprovided a new and complementary perspective on age-related differences in brain structureand function, by allowing an in vivo characterization of the microstructural properties ofwhite matter (WM) [9–11]. Measures from DTI exhibit increased sensitivity to age-relateddecline, relative to volumetric measures [12–14], and DTI is beginning to show promise forthe detection of brain disorders such as Alzheimer’s disease and related forms of dementia(see Section 4, below, and Gold, this issue).
The success of this clinical translation, however, rests on the foundation provided by DTI tostudies of aging and WM in the absence of significant disease. Measures from DTI arepromising in this regard because they comprise several indices with a sufficient range ofvalues to permit detailed analyses of normative data. In addition, independently of clinicaltranslation, the empirical measures from DTI can be combined with behavioral measures ofcognitive performance, to further advance understanding of structure-function relations [15].Research with healthy adults has enabled the development of larger sample sizes andincreased statistical power, yielding more reliable estimates of the relations between WMintegrity and cognitive performance, and of age-related effects [16–18].
Most frequently, DTI research on aging is conducted with different groups of individuals atone point in time and thus yields information regarding age-related differences (i.e., cross-sectional differences). The vast majority of the research we consider here is cross-sectional,and thus the estimated effects of age may be influenced by a variety of individualdifferences between the age groups (cohort differences). Measurement of the sameindividuals over time is necessary to assess age-related changes within individuals (i.e.,longitudinal changes). Historically, longitudinal MRI studies have focused on measures ofbrain volume and pathology, such as WM lesions [19–22], but longitudinal studiesconducted with DTI are beginning to emerge [23–25]. Our goal in this article is to reviewthe application of DTI for understanding the changes in both WM integrity and cognitiveperformance associated with aging.
2. Microstructural properties of WM integrity2.1 Measurement of diffusivity in white matter
Measures from DTI allow inferences about WM microstructure in vivo by quantifying thedirectionality and rate of diffusion of water within tissue. At the level of the basic unit of theMRI image (i.e., voxel), this information is represented mathematically as a diffusionellipsoid. Diffusion along the major axis or eigenvector (λ1) of the ellipsoid is termed axialdiffusivity (AD), whereas the average of the second and third minor axes (λ2, λ3), termedradial diffusivity (RD), reflects diffusivity perpendicular to major axis of the tensor [26–29].
The most frequently used DTI summary measures are fractional anisotropy (FA) and meandiffusivity (MD). Fractional anisotropy represents the fraction of the tensor that can beassigned to anisotropic (directional) diffusion [26, 30–31]. Values for FA range between 0and 1, with higher values reflecting increased directionality of diffusion, independently ofthe rate of diffusion. The directionality of diffusion depends on the density of physicalobstructions such as membranes and the distribution of water molecules between different
Madden et al. Page 2
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

cellular compartments. Thus, FA is typically higher in WM, in which diffusion is restrictedby the myelin sheaths of axons, particularly in compact tracts with uniform fiber alignment,such as the corpus callosum, whereas diffusion in GM is less bounded and more isotropic.MD is a mean of all three axes of the diffusion ellipsoid and reflects the rate of waterdiffusion within a voxel, independently of the directionality.
Researchers have used several different methods of post-processing for DTI data andpresenting the results [9–11, 32]; and Figure 1 illustrates four approaches. In region ofinterest (ROI) analysis (panel A), data are extracted from regions that are placed directly onthe DTI image, typically in native (non-normalized) space, for each individual. Voxel-basedmorphometry (VBM; panel B) and tract-based spatial statistics (TBSS; panel C) both rely ongroup-normalized data to estimate either the tissue composition of individual voxels (VBM)or the “skeleton” of WM tracts (TBSS). Fiber tracking of WM pathways (tractography;panel D) is typically conducted in native space, although information from normalizationmay be incorporated into the procedure.
Fiber tracking uses seed and target ROIs to estimate the location of sets of spatially coherentWM fibers, and tractography is emerging as a widely used procedure for DTI analysis.Individual fiber tracts can be color-coded in terms of tract identity, as in Figure 1, butalternatively, color is often used to represent either the diffusion directionality or a relatedproperty of individual voxels within a tract. Figure 2 is an example of WM tractography inwhich seed and target ROIs (panel A) are placed on a DTI image as the basis for estimationof a WM pathway, in this case the genu of the corpus callosum (panel B). The estimatedfiber tract can be simplified by representing a “tube” of the mean values of the fibers withina tract (panel C). Color-coding the diffusion property of individual voxels (typically FA) canillustrate variation in WM integrity within the tract. In the case of the genu, for example, FAis higher in the center (blue-green values), where the fibers are most compact anddirectionally coherent.
The parameters of the ellipsoid reflect distinct aspects of WM microstructure. Animalmodels of nerve injury suggest that different types of WM injury lead to differential effectsin AD and RD. S. K. Song, Sun, and colleagues have reported that axonal damage withoutassociated myelin injury led to a decrease in AD without accompanying change in RD up tothree days after injury, whereas at 14 days following injury RD increased significantly,consistent with myelin damage and secondary (Wallerian) degeneration [33–36]. Theinterpretation of the neurobiological mechanisms of AD and RD, however, is complicatedby many variables (Section 2.4), for example, the local architecture of WM tracts withcrossing fibers, in which case the major axis of the tensor model may not be parallel tomyelinated axons [37–38]. Assaf and Pasternak [39] suggested that conjoint analysis of allmeasures derived from the diffusion tensor should yield a more comprehensive picture ofdifferent elements of WM microstructure. By analyzing FA and MD conjointly with RD andAD, it is possible to distinguish between diffusivity patterns with different neurobiologicalfoundations. In this approach, for example, Wallerian degeneration is characterized bydecreases in both FA and AD, combined with an RD increase and no net difference in MD[36–37, 40–42].
2.2 Adult age differences in WM diffusivity: Neurobiological mechanismsAcross many DTI investigations of aging, a general trend of the findings is a decrease in FAand an increase with measures of diffusivity, with increasing adult age, suggesting an age-related decline in the composition and integrity of WM [16–18]. These DTI findings arevalidated by neurobiological data. Post-mortem histological studies show that advanced agein the absence of neurological disease is linked to alterations of virtually all the WMcomponents. Some fraction of axons degenerate and swell, myelin becomes less compact,
Madden et al. Page 3
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

degenerates, or forms redundant wraps; glial cells accumulate cellular debris, form glialscars, increase in number or show activated morphology [43–47]. At the technical level, asnoted previously, the presence of crossing fibers within a voxel, as well as estimation errorin the primary eigenvector in the tensor model [37–38], lead to difficulties in directlycorrelating specific neurobiological mechanism with individual DTI parameters.
Variability in the pattern of age-related differences in DTI measures, across brain regions,also complicate direct interpretation of the DTI data but do help to provide a link toneurobiology. The first series of DTI studies of age-related effects, using an ROI approach,reported consistently that aging was associated with decreased FA in the genu of the corpuscallosum and associated frontal pericallosal regions, whereas age-related decline was lesspronounced (though detectable) in the splenium of the corpus callosum and parietal WM[48–50]. This anterior-posterior gradient was replicated in subsequent investigations [51–58].
Davis et al. [54] have demonstrated that this gradient is not isomorphic with the concept ofage-related decline in frontal lobe functioning [59–61]. Specifically, tracts traversing thefrontal lobe in an anterior-posterior direction (e.g., the uncinate fasciculus and cingulum)showed a monotonic decrease in the age-related decline in FA in an anterior-posteriordirection, and these age differences in FA did not change abruptly the boundary of thefrontal lobe. This pattern of age-related difference in WM integrity is in line withmylodegeneration as a potential neurobiological mechanism for regional variation in age-related differences in FA. That is, those brain regions that are the latest to myelinate fully,during development, may be those most vulnerable to the negative effects of adult aging.These include frontal regions but also more posterior sites (e.g., occipitotemporal boundary).The assumption is that oligodendrocytes of associative WM pathways are the mostmetabolically active cells and thus correspondingly vulnerable to the accumulation ofmetabolic damage. The anterior-posterior gradient is not an entirely comprehensiveframework, however, and Sullivan and colleagues have recently proposed that age-relateddecline in FA can also be characterized by a superior-inferior gradient, particularly withregard to fibers tracts in the internal capsule (e.g., centrum semiovale and corticospinal tract)[62–64]. Both the anterior-posterior and superior-inferior gradient theories share theassumption that damage within white matter pathways decreases the efficiency ofcommunication among the widely distributed neural systems comprising cognition, thusleading to a disconnection of cognitive networks [4, 65–70].
2.3 Axonal versus myelin degenerationAnalyses of the magnitude of age-related differences in AD and RD suggest that age-relatedRD differences(usually increases) are more prominent than AD differences [54–55, 71–72],which has been interpreted as support for a predominantly myelin-specific effect in aging.Age-related decreases as well as increases in AD, however, have been reported [12, 63–64],and thus differential age-related effects in axonal and myelin-specific variables are not clear.Two recent studies, by Bennett et al. [51] and Burzynska et al. [53] provided a morecomplete account by examining systematically the age-related differences in the diffusivitycomponents, in voxelwise analyses. Both of the studies used TBSS to analyze FA, AD, andRD concurrently, but Burzynska et al. also included MD as a variable.
From the whole-brain analyses of the diffusivity components, Bennett et al. [51] reportedthree main patterns of age-related difference, and Burzynska et al. [53] obtained five distinctpatterns. The two studies agree that the most spatially prominent pattern was a decrease inFA for older adults, relative to younger adults, accompanied by an increase in RD. In asubset of regions (e.g., genu of the corpus callosum), this pattern also included an age-related increase in MD and AD. Figure 3 (panels A and B) illustrates this result in the
Madden et al. Page 4
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Burzynska et al. data. Table 1 provides a direct comparison of the two data sets as well asproposed neurobiological mechanisms for different patterns.
When age-related increases in both RD and AD are present, it is likely that pathology ofboth axon fibers and the surrounding myelin sheaths is involved. Cellular debris clearanceby phagocytic cells limits the restriction of water diffusion and contributes to an increase inisotropic diffusivity. In some regions (e.g., fornix, in Table 1), severe fiber degenerationmay lead to decreased tract volume, and, as a result, to a partial volume effect with thesurrounding cerebrospinal fluid (CSF), which magnifies the effect of the increase in theisotropic diffusivity. This pattern of diffusivity differences likely reflects higher extracellularvolume fraction and lower membrane density as a result of both myelin disruption and axonloss, a smaller volume fraction of axons, an increase in axonal spacing [31], as well as areduction in extracellular tortuosity [73]. A more specific disruption of myelin is impliedwhen an increase in RD occurs without an accompanying increase in AD (e.g., forcepsmajor, dorsal cingulum, in Table 1). Age-related increase in RD is a signature ofdemyelination is consistent with animal models, for example in the loss of myelin due toischemia [33, 35]. In human post-mortem data, from multiple sclerosis patients, RDcorrelated with demyelination severity [74–75].
Bennett et al. [51] and Burzynska et al. [53] both reported that some WM regions in olderadults (e.g., superior corona radiata, in Table 1) were characterized by a pattern in which thedecreased FA and increased RD were accompanied by a decreased AD, resulting in no netdifference in MD. This type of diffusivity pattern has been observed in secondary(Wallerian) degeneration, which is the degeneration, over time, of axon fibers distal to thepoint of transsection or injury [36–37, 40–42]. This process involves initial axonal beading,axolemma breakdown, organelle accumulation, and finally the breakdown of myelin andoligodendrocyte apoptosis. The cellular debris is cleared by activated microglia and glialscar is formed by astrocytes [76]. This glia infiltration, which restricts AD and decreasesMD [77] differentiates Wallerian degeneration from mere loss of fibers and myelin. Localischemic lesions, such as WM lesions typical for the healthy older population [1–2], may bethe primary lesions from which Wallerian degeneration originates in the aging brain [3],although decline in FA associated with healthy aging appears not to be dependent entirelyon WM lesions [53].
Finally, the Bennett et al. [51] and Burzynska et al. [53] studies reported that some WMregions of older adults showed a decrease in FA accompanied by a decrease in AD and MD,without a difference in RD (e.g., cerebral peduncles, in Table 1). Lowered FA driven bydecreased AD is considered a marker of acute and primary axonal damage and wasobserved, in human patients, in the early post-injury phase after corpus callosotomy [78] andin optic neuritis [79], as well as in various animal models of axonal injury [33, 80–85]. Theextent of acute axonal injury in normal aging is not clear, but its origin may be ischemic andrelated to development of WM lesions. Bennett et al. proposed that a decrease in FA and ADwith no significant change in RD and MD may be also a result of disrupted macrostructuralreorganization of the fibers, such as less coherent fiber alignment.
2.4 Limitations in interpreting axial and radial diffusivityAs noted previously (Section 2.1), it is difficult to distinguish components of themicrostructural pathology based on DTI indices alone. Although analyzing multiple DTIindices simultaneously provides the most information, the link to neurobiology remainsincomplete at this time. In the case of chronic WM lesions, for example, two post-mortemstudies of multiple sclerosis patients, combining DTI with histology, reported that myelincontent and axonal count are strongly correlated [74–75]. Specifically, lesions in thesepatients were characterized by lower myelin content, axonal loss, and gliosis, expressed by
Madden et al. Page 5
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

lower FA and higher MD, RD and AD values. Furthermore, in multiple regression analysesconducted on each DTI index, both myelin content an axonal count were concurrentpredictors of all indices except AD. This latter variable was related to myelin content but notto axonal count [74]. Thus, RD is sensitive but not specific to demyelination and may bemore appropriately considered as a marker of overall tissue integrity [74].
Recent findings suggest that partial volume effects (i.e., incorrect assignment of a voxel asWM) may have a significant effect on the estimation of age-related effects. Vos et al. [86]found that estimated diffusivity parameters are differentially affected depending on whetherpartial volume represented CSF, WM with different principal eigenvector orientation, orGM. In addition, the partial volume effect varied as a function of fiber bundle volume,orientation, and curvature. In view of the WM structural changes associated with aging, Voset al. recommend including these structural variables as covariates in analyses of age-relatedeffects. Similarly, Miller et al. [87] compared ultra-high resolution DTI images (0.73 mmisotropic voxels), acquired post-mortem, to data blurred to resemble the more typical scanresolution of 2–3.5 mm3, and found that the lower resolution images led WM tracts toappear thinner than they actually were, due to partial volume effects. Other investigations,however, that have included post-processing methods to specifically address partial volumeeffects, have reported that age-related differences in DTI measures remain detectablefollowing consideration of partial volume effects [71, 88]. Thus, although partial volumingrelated to age-related atrophy clearly affects thin tracts such as fornix (Table 1) [3, 53], age-related differences in diffusivity in other WM structures likely reflect true effects of age onmicrostructure.
3. Cognitive performance and WM integrity in agingDuring healthy aging, behavioral measures of fluid cognitive abilities (those relying onelementary perceptual speed and sensory/motor functioning) tend to decline, whereasmeasures of crystallized cognitive abilities (those relating to knowledge and expertise)exhibit little or no decline until very late in life [89–92]. Imaging studies of WM volume andpathology, particularly studies of the lesions that appear as WM hyperintensities, have longindicated a relation between WM integrity and fluid cognition, particularly in samples ofolder adults [19–20, 93–98]. Building on initial reports by O’Sullivan et al. [69] andSullivan et al. [49], evidence from DTI is rapidly accumulating to indicate thatmicrostructural integrity of normal appearing WM has some role in age-related decline incognitive functioning in healthy adults [16–18].
In a previous review of DTI studies of cognitive aging [16], we discussed severalinterpretive issues that currently limit the characterization of the specific relation betweenWM integrity and cognitive aging. Among these were the focus, in previous research, on theoverall variance in outcome measures rather than the age-related variance. That is, fewstudies have either a) investigated the degree to which individual differences in WMintegrity share age-related variance in cognitive performance; or b) directly compareddifferent age groups in the correlation between WM integrity and cognition. Salthouse [99]has reviewed these and related issues, particularly the failure of previous studies to evaluatecompeting models of the relations among neuroanatomy, age, and cognition. Salthouseproposed that, at the present time, the evidence for concluding that aspects of brain structureare neuroanatomical substrates of age-related cognitive decline is weak. For example, if therelation between a neuroanatomical variable and cognition is reduced when the effect of ageis controlled statistically, different interpretations are possible. This pattern may occurbecause age has a mediating role in the relation between the brain and cognition [100].Alternatively, the neuroanatomical variable and cognition may actually be independent andonly apparently related to each other by virtue of their relations to age.
Madden et al. Page 6
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Recent empirical investigations have taken disparate approaches to characterizing therelation between WM integrity and cognition in aging. In Table 2, we summarize the resultsof 29 published DTI studies addressing this relation; this summary adds 11 published reportsto a corresponding table in our previous review [16]. We identified these studies fromPubMed searches using the terms “white matter integrity AND aging” and “diffusion tensorimaging AND aging AND cognitive.” From this output, we selected articles that wereempirical investigations of WM integrity-cognition effects in healthy older adults. Articlesthat used global rather than regional DTI measures [101–104] or used a measure of WMintegrity other than FA [65, 105] were excluded. In this version of the table, we focusspecifically on the three cognitive domains of executive function (e.g., maintaining arelevant task set and inhibiting distraction), processing speed, and memory, which is adivision that is often used in studies of WM and aging [103–104, 106–109].
Table 2 presents mean effect sizes [110] for relationships between FA and cognitiveperformance across nine brain regions. For each of the cognitive domains, results arepresented separately for studies in which FA-cognition relationships were reported for bothyounger and older adults, or in older adults alone (i.e., when only older adults wereexamined or were reported separately from younger adults). Bold font indicates effect sizesthat are moderate or larger (≥ 0.30). The reported data primarily suggest a stronger relationbetween FA and cognition for executive function and processing speed than for memory.This pattern has been noted previously [109, 111], but few direct comparisons amongcognitive domains have been conducted. The cognitive domains in Table 2 are complex andlikely to rely on multiple brain regions; at the present time, the WM-cognition relation doesnot exhibit a high degree of regional specificity [16, 108]. Within individual studies,however, reliable differentiation of specific information processing components has beenobserved. Madden et al. [55], for example, reported that FA within the genu and spleniumwas correlated with the efficiency of semantic memory retrieval but not with perceptual-motor speed. Bennett et al. [112] found that frontoparietal FA correlated with visual searchspeed, independently of perceptual-motor speed. The anterior-posterior gradient of age-related decline in FA also appears to correlate differentially with tests of speed andexecutive function (anterior) versus memory (posterior) [54, 108].
A new feature of Table 2 is that mean effect sizes are presented separately for: a) the rawbivariate correlations between FA and cognition, without age included in the model; and b)analyses that controlled statistically for the effect of age or age group (i.e., age partialedvalues). Thus, by comparing the raw bivariate and age-partialed values, we can estimate thedegree to which WM integrity and cognition share age-related variance. That is, some formof age dependence is implied when the effect size for the WM-cognition relation decreasesfrom the raw bivariate to the age-partialed version.
The current selection of published research yielded 28 raw bivariate correlations betweenFA and cognition that were both ≥ 0.30 (in bold font) and were accompanied by an age-partialed analysis. For 23 of those correlations, the raw correlation decreased to below 0.30when age was partialed, which was less than half the size of the original raw value for 18 ofthe correlations. That is, in nearly two-thirds of cases with an FA-cognition effect ofmoderate or greater size, age shared more than 50% of the variance in cognition that wasrelated to FA.
Independently of those 28 correlations with both raw and age-partialed values, Table 2 alsocontains 15 correlations from studies that only reported the age-partialed effects. Of these15, eight were of at least moderate size (≥ 0.30) and seven were smaller in magnitude. Thus,studies that reported only age-partialed effects yielded approximately equal numbers ofsmaller and moderate effect sizes for the WM-cognition relations. From this finding, as well
Madden et al. Page 7
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

as from the previously discussed comparison of raw bivariate and age-partialed values, weconclude that age shares a substantial portion of the of the relation between WM integrityand cognition. As Salthouse [99] has noted, however, from this pattern alone we cannotdetermine whether age is directly influencing a relation between WM and cognition. The FAand cognitive variables may be independent but both related to age. In addition, bypartialing age, the contribution of other variables that are associated with age (e.g., healthstatus variables such as aerobic capacity) is also removed [113–115].
Whereas Table 2 illustrates the effect of age in the relation between WM integrity andcognition, several investigations have taken a different approach, using multiple regressionand related techniques to identify the role of WM integrity in the relation between age andcognition. Following the addition of FA to models of the age-cognition relation, theproportion age-related variance is substantially reduced, yielding evidence for WM integrity(particularly within frontoparietal regions) as a mediating variable in age-related decline inthe efficiency of semantic categorization [55], shifting attentional sets [116–117], andimplicit learning [118]. Zahr et al. [64], however, compared these two approaches within thesame data set and reported that the attenuation of the relation between FA and workingmemory, by age, was at least as great in magnitude as the attenuation of the age-relatedvariance by FA. Thus, WM integrity and working memory may have independent relationsto age, without age exhibiting a directly causal role in the WM integrity-working memoryrelation [99].
The identification of specific causal relations is possible in path-analytic and relatedtechniques (e.g., structural equation modeling; SEM) that investigate the direct and indirectpathways of covariation among multiple variables [119]. Charlton et al. [120] have exploredSEM models for the relations among age, cognition, and MD (as a measure of WMintegrity) in a sample of adults 50–90 years of age. The results indicated that WM integrity(from a relatively large ROI: centrum semiovale and periventricular WM) mediated the age-related decline in working memory performance. Burgmans et al. [12] described an SEMmodel in which AD from a large ROI (WM in the parietal, temporal, and occipital lobes)mediated the age-related decline in pattern and letter comparison speed. Voineskos et al.[121] used DTI tractography rather than ROIs and developed an SEM model in which tract-specific FA mediated the relation between age and various forms of cognitive performance.These more sophisticated statistical methods will undoubtedly be valuable for understandingthe role of WM integrity in cognitive aging. Their application, however, will need to betempered by awareness that modeling is more appropriate for comparing different,competing models than for describing or exploring data [122–123], and that measures ofWM from different tracts may not be independent but instead reflect shared variability inWM integrity across tracts [103, 124].
4. White matter integrity in geriatric neuropsychologyAlthough the main focus of this review is healthy aging, WM integrity plays an importantrole in many age-associated neurocognitive disorders, ranging from Alzheimer’s disease andother forms of dementia, to late-life depression, and the effects of stroke and traumatic braininjury [125–126]. Gold (this issue) provides a more detailed review of issues relating toAlzheimer’s disease and mild cognitive impairment. Measures from DTI, includingtractography, are beginning to offer insights into neurological disease diagnosis andtreatment planning [127– 129].
4.1 Alzheimer’s disease and other forms of dementiaA fundamental question is whether Alzheimer’s disease differs qualitatively from normalbrain aging. Several DTI studies suggest that Alzheimer’s-related WM changes are not
Madden et al. Page 8
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

simply an exaggeration of the anterior-posterior gradient of normal aging but instead involveother brain regions (e.g., temporal) [130–133]. Both decreased FA and increased MD in thehippocampus and temporal regions appear to be associated with the episodic memorydeficits that characterize the earliest stages of this disease [134–135]. Multivariate analysesof whole-brain WM have suggested that mean diffusivity maps perform slightly better thanFA maps in classification of individuals with Alzheimer’s disease versus normal elderly[136]. Interestingly, even asymptomatic individuals with a family history of Alzheimer’sdisease show reduced FA in regions typically affected by the disease, which suggests thatWM integrity may be sensitive to Alzheimer’s-related pathology years before disease onset[137]. Data from DTI have also assisted in characterizing the transition from a preclinical tomanifest disease [132, 138–139]. Research with DTI has also clarified some of the clinicaland neuropathological differences between Alzheimer’s disease and other dementias,including frontotemporal dementia [140], vascular dementia [141], and dementia associatedwith Parkinson’s disease [142–144].
4.2. Late-life depressionDepression is the most prevalent psychiatric disorder and is particularly problematic in olderadulthood because of its association with cognitive impairment and other adverse outcomes[145–146]. Region of interest based studies of late-life depression have found lower FA fordepressed individuals relative to controls, particularly in frontal brain regions [147–148];however, whole brain, voxel-based approaches have reported more widely distributeddecrements in WM integrity [149–150]. Some studies have also found higher MDthroughout the brain of older adults relative to controls, but most prominently in frontalregions [151]. The WM differences associated with late-life depression have also beenassociated with worse neuropsychological performance on tests of executive function(Stroop, Tower of Hanoi) and processing speed, although regional localization varies [150–152].
4.3 Plasticity of white matterThe prospect that WM integrity may increase in response to cognitive training has importantimplications for diagnosing and treating age-related cognitive disorders. Lovden et al. [106]compared DTI measures for younger and older adults before and after an intensive trainingregimen (101 1-hr sessions over 180 days), on tests of working memory, episodic memory,and perceptual speed. Relative to a no-training control group, individuals in the cognitivetraining group exhibited greater increase in FA and decrease in MD, in the genu of thecorpus callosum. Further, the magnitude of the training-related improvements in WMintegrity was as great for older adults as for younger adults. This type of plasticity of WMhas also been demonstrated following brain surgery in humans. Yogarajah et al. [153]investigated epilepsy patients before and after temporal lobe resection and noted bothpositive and negative changes in WM integrity following surgery. In particular, largerincreases in AD in the ventro-medial language network after the surgery were associatedwith smaller fall in post-operative verbal fluency, suggestive of structural reorganization.Sidaros et al.[154] and Bendlin et al. [155] examined the utility of DTI measures forassessing recovery following traumatic brain injury. Although the changes in WM integrityfollowing brain injury are not entirely consistent, and a complete model of adult brainreorganization associated with injury is not available, DTI will be valuable for furtheradvances on these issues. The concept of plasticity is particularly important in view of theresults from fMRI, in healthy older adults, which suggest that some patterns of functionalbrain activity represent a direct response to structural changes [153, 156–158].
Madden et al. Page 9
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

5. Technical developments in DTIAdvances in the technology of DTI can contribute significantly to both defining theneurobiological mechanisms of WM integrity in aging and identifying clinical translation.As discussed previously (Section 2), a difficulty in understanding age-related decline in FAis the presence of crossing fibers within a voxel, which may not always be captured at thelevel of spatial resolution available. Thus, a decline in the structural integrity of WM may bedifficult to distinguish from a change in the local coherence of fibers [37–38, 51]. Similarly,atrophy and related structural changes associated with aging may contribute to estimates ofage-related differences in DTI measures [86–87]. Several types of improvements in DTIresolution, and the integration of DTI and fMRI measures, promise to provide a moreaccurate characterization of age-related WM effects.
5.1. Correction for geometric distortions in single-shot-EPI-based DTI dataMost DTI data in investigations of aging and cognition are acquired with single-shot echo-planar imaging (EPI) pulse sequence, which has superior imaging speed and is relativelyinsensitive to intra-scan motion. However, it is well known that the spatial resolution andspatial accuracy are lower in EPI, as compared with other clinical MRI pulse sequences.Specifically, DTI data obtained with EPI are geometrically distorted, resulting from bothsubject-dependent susceptibility field gradients and subject-independent eddy currents. Oneapproach to reducing distortion is to use a twice-refocused DTI sequence [159], but a moreeffective and complete correction for both susceptibility and eddy current related distortionsrelies on extra reference scans, in the form of field inhomogeneity maps that correspond tothe various diffusion weighting directions [160]. Using this latter approach, Truong et al.[161] demonstrated that, by adding sensitivity encoding (SENSE) DTI acquisition and areconstruction algorithms with higher-order polynomial correction, significantly improvedspatial accuracy and signal-to-noise ratio can be obtained.
Recently, Truong et al. [162] further demonstrated that eddy currents-induced distortionshave a complex time-domain pattern. Truong et al. proposed an alternative dynamic B0mapping and off-resonance correction method that measures the exact spatial, temporal, anddiffusion-weighting direction dependence of the susceptibility- and eddy current-inducedmagnetic fields. As a result, both susceptibility effects and time-varying eddy currentartifacts can be effectively corrected, yielding DTI data with the highest possible spatialfidelity and accuracy. Figure 4 shows color-coded FA maps in three representative sliceswith no correction (A), static correction (B), and dynamic correction (C). The uncorrectedFA maps in three representative slices show severe susceptibility-induced distortions,particularly near the genu of the corpus callosum (Figure 4A, dashed lines), as well as eddycurrent-induced FA errors, most prominently at the anterior and posterior edges of the brain(arrows). The conventional static off-resonance correction can only correct for thesusceptibility-induced distortions, and for the eddy current-induced FA errors at theposterior edge of the brain, but not at the anterior edge (Figure 4B, arrows). In contrast, thedynamic off-resonance correction can more effectively correct for all artifacts (Figure 4C).
5.2. Beyond single-shot EPI acquisition: Alternative imaging pulse sequences for DTI ofhigh spatial-resolution
Several studies have shown that alternative imaging pulse sequences can be used to acquireDTI data with significantly improved spatial-resolution, relative to single-shot EPI basedDTI. First, high-resolution DTI with significantly reduced geometric distortion and motion-related artifact can be achieved through acquiring segmented k-space data along the readoutdirection (i.e., readout-segmented EPI or RS-EPI), in contrast to conventional multi-shot EPIwith k-space segmentation along the phase-encoding direction [163–164]. Second, the self-
Madden et al. Page 10
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

navigated interleaved spiral (SNAILS) sequence for high-resolution DTI, developed by Liuet al. [165] improves spatial resolution by using a multi-shot, fat-saturated diffusion-weighted spin echo sequence that oversamples the center of k-space, providing an inherentmotion compensation capability. Third, Pipe et al. [166] showed that high-resolution andhigh-quality DTI data can be obtained by integrating two sequences: fast spin echo imaging(which reduces distortion), and the periodically rotated overlapping parallel lines withenhanced reconstruction (i.e., PROPELLER), which increases tolerance to intra-scanmotion. It is also feasible to integrate a multi-shot EPI sequence and PROPELLER motion-artifact reduction, producing high-resolution DTI with reduced artifact [167].
5.3. Beyond single- tensor model: Diffusion measurement in q-space and at higherangular-resolution
Within the DTI framework, significant advances in spatial resolution have been achieved byusing extended scanning periods with post-mortem brains [87], and by using the long-rangeinformation contained in the fibers tracts, at higher MRI field strengths [168–169]. Thediffusion tensor model, however, relies on the assumption of Gaussian diffusion [170], witha single diffusion tensor within a voxel, which may not always be valid. To address this,researchers have been using different mathematical frameworks for measuring diffusion toincorporate non-Gaussian diffusion and as a result can better resolve multiple diffusiontensors within the same voxel (angular resolution).
The initial methods for non-Gaussian diffusion measurement include q-space imaging, inwhich the probability density function for molecular diffusion is estimated without theassumption of a Gaussian distribution [171–172]. More recent approaches include diffusionspectrum imaging (DSI) [173–175], high-angular-resolution diffusion imaging [176–177],q-ball imaging [178], and generalized diffusion tensor imaging (GDTI) methods [179],among others. These methods can more accurately resolve crossing fibers, as compared withthe conventional DTI and in some cases can resolve multiple axon directions within a singlevoxel. For example, Figure 5 shows that more fibers can be correctly identified using DSI,in comparison to the conventional DTI model [175]. The patterns of crossing fibersobserved from these advanced imaging methods correspond to those obtained, in other datasets, from autoradiography [174], although validation within the same brain is needed [180].In addition, these new methods tend to require extensive scanning time, and thus moreefficient data acquisition techniques will be needed for applications in vivo.
5.4. Toward tissue specificity: Axon- and myelin-specific DTI measuresAge-related decline in WM integrity tends to be more pronounced in RD than in AD [54–55,71], consistent with a prominent role for decreased myelin integrity in aging [65–66].Multiple DTI indices concomitantly vary with adult age (Table 1), however, and assigningspecific neurobiological interpretation to DTI parameters is problematic (Sections 2.1, 2.4)[37–38].
The MRI pulse sequences may be modified in a way that the derived tensor measuresspecifically reflect physiological information of myelin. A recent study by Avram et al.[181] described a magnetization transfer (MT) prepared stimulated-echo DTI technique thatis sensitive to the microanatomy of myelin tissue. The short echo time enabled by thestimulated-echo acquisition preserves significant signal weighting from myelin water ofshort T2 time constant, and the MT preparation further provides differentiating sensitizationto this signal. This myelin water weighted DTI has the potential to lead to improvedassessment of myelin pathology and detection of degradation in myelin microanatomy.
Madden et al. Page 11
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

One recently developed method, the AxCaliber technique [182–183], will be valuable incharacterizing changes of white-matter microstructure related to aging, particularly withrespect to axonal populations with differing diameters. The AxCaliber technique provides anoninvasive estimate of diameter distribution within each voxel, based on the differences inrestricted diffusion, across populations of axons, at different diffusion times. Therefore, bychanging the diffusion time from short to long one can probe the relative contributions ofdifferent subpopulations, reflecting axonal diameter distribution, within each voxel. Largerdiameter axons generally occur in pathways supporting rapid neural transmission, such asthe pyramidal (corticospinal) tract. Single-cell recording of pyramidal tract neurons in thecat has demonstrated significant reduction in conduction velocity with age [184]. Humanpost-mortem studies, however, are divided as to whether age-related volumetric decline ismore pronounced for larger myelinated axons [45] or more thinly myelinated ones [44, 47,185]. The integrity of smaller, later myelinating fibers within association (e.g., prefrontal)cortex is correlated with maximum motor speed [105]. In view of the central role ofprocessing speed in theories of cognitive aging [186–187], and the relation between FA andspeed (Table 2), AxCaliber is a particularly promising method for investigations of agingand cognition.
5.5. Toward brain activations: Functional DTIIn contrast to functional MRI with BOLD contrast, which measures hemodynamic responsesassociated with neuronal activity, DTI is a structural imaging modality that assesses theintegrity of WM. Although neuroimaging research typically assumes that sensory andperceptual experience derives from neuronal activity, recent evidence suggests thatdetectable changes in WM occur in response to sensory stimulation. Mandl et al.[188]reported that changes occur in FA occur selectively in relevant WM tracts following sensorystimulation, when analyzed as a 200 sec time course. During and following theadministration of a 1 min tactile or visual stimulus, changes occurred in the thalamocorticaltracts for the tactile stimulus and in the splenium and optic radiations for the visual stimulus.The FA changes were relatively small in magnitude (~1%) and comprised both positive andnegative effects, but Mandl et al. proposed that the positive changes (stimulation-relatedincreases in FA) represent local fiber activity, possibly glial cell swelling, although thespecific biophysical mechanisms remain unknown.
5.6. Toward constructing a Brain Connectome: Graph theory representation of DTI basedconnectivity maps
The quantitative maps (e.g., FA maps) derived from DTI data include valuable physiologicalinformation for different brain regions. These data are derived either from an a prioriselection of relevant regions (e.g. tractography) or from an unbiased, voxelwise analysisacross the whole brain (e.g., TBSS). However, the DTI data alone do not define the higher-order structure among WM tracts.
Recently, researchers have been using new mathematical approaches, particularly graph-theory analysis [189–190], to assess the overall topographical network properties (e.g., theglobal efficiency) of FA maps and to estimate the relative strength of the connections amongtracts. Gong et al. [191], for example, analyzed DTI data from 80 healthy young adults andfound that the DTI derived topographical measures of connectivity networks resemble thatof a small-world architecture, which is characterized by greater local interconnectivityrelative to a random network. Iturria-Medina et al. [192] used DTI based graph-theoryanalysis to measure the overall network properties of the right and left hemispheres. Theirresults indicate that the right hemisphere is more efficient and interconnected, whereas theleft hemisphere comprises regions that are more central or indispensable (e.g., language) forthe whole-brain structural network. In a graph-theory study of individual differences in DTI
Madden et al. Page 12
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

tractography data, Li et al. [193] found that individuals with higher intelligence scoresexhibited WM networks with a shorter characteristic path length and a higher globalefficiency, indicating a more efficient parallel information transfer in the brain. The changesin WM integrity associated with normal aging, however, have not as yet been explored froma graph-theory perspective.
Researchers in the area of BOLD resting-state activity have demonstrated reliabledifferences in resting state functional networks associated with aging in healthy adults [194–197]. These differences, predominantly a reduction in connectivity in the default modenetwork of regions, centered in medial prefrontal and posterior cingulate cortical regions,reflect age-related differences in the resting state activity of the brain. Structural WMconnectivity, as assessed from DTI, appears to be an anatomical constraint on this functionalconnectivity [198–200]. In addition, DTI data suggest that age-related changes in cognitionrepresent the combined influences of WM integrity and resting state functional connectivity,not only within the default mode network but also within networks involving prefrontal andsubcortical regions [107, 109, 201]. Further studies that integrate DTI and fMRI measureswill be valuable in characterizing the structure-function relations within the human brainconnectome.
6. ConclusionInformation from DTI provides a unique perspective on cognition in aging by contributingseveral indices of the microstructural integrity of cerebral WM. To date, a wide range ofresearch investigations suggests that, even in the absence of significant disease, individualdifferences in WM integrity share a substantial portion of age-related variance in cognitiveabilities, particularly those related to elementary perceptual speed and executive function.Further, age-related differences in WM integrity vary across brain regions, with older adultsexhibiting relatively greater decline (lower FA, higher diffusivity) in more anterior andsuperior brain regions, which may reflect the later development of myelination in theseregions. Although preliminary findings suggest that the age-related effects derive moredirectly from changes in myelin structure than in axonal integrity, additional technicaldevelopments in DTI resolution and tissue specificity are needed to define theneurobiological mechanisms of WM changes associated with aging. At the interpretivelevel, it will be necessary to move beyond describing individual correlations amongvariables, to testing competing models of the relations among WM integrity, aging, andcognition. Initial evidence supports one model in which decline in cerebral WM integritydirectly contributes to the less efficient performance of fluid cognitive abilities that occursduring healthy aging.
Highlights
> We review diffusion tensor imaging (DTI) of white matter (WM) integrity inaging.
> Even without disease, age-related decline occurs in both cognition and WMintegrity.
> Age-related decline in WM integrity interrupts communication among brainnetworks.
> Disconnection of brain network may contribute to normal cognitive declinewith age.
Madden et al. Page 13
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

AcknowledgmentsPreparation of this article was supported by grants R01 AG011622, R21 NS065344 and R01 EB009483, F32AG038299, and K23MH087741 from the National Institutes of Health. We are grateful to Tim Salthouse for hiscomments on a previous version of this article.
References1. de Leeuw FE, de Groot JC, Achten E, Oudkerk M, Ramos LM, Heijboer R, Hofman A, Jolles J, van
Gijn J, Breteler MM. Prevalence of cerebral white matter lesions in elderly people: a populationbased magnetic resonance imaging study. The Rotterdam Scan Study. J Neurol NeurosurgPsychiatry. 2001; 70:9–14. [PubMed: 11118240]
2. Vernooij MW, Ikram MA, Tanghe HL, Vincent AJ, Hofman A, Krestin GP, Niessen WJ, BretelerMM, van der Lugt A. Incidental findings on brain MRI in the general population. N Engl J Med.2007; 357:1821–1828. [PubMed: 17978290]
3. Vernooij MW, de Groot M, van der Lugt A, Ikram MA, Krestin GP, Hofman A, Niessen WJ,Breteler MM. White matter atrophy and lesion formation explain the loss of structural integrity ofwhite matter in aging. Neuroimage. 2008; 43:470–477. [PubMed: 18755279]
4. Raz, N. Aging of the brain and its impact on cognitive performance: Integration of structural andfunctional findings. In: Craik, FIM.; Salthouse, TA., editors. Handbook of aging and cognition, 2nded. Mahwah, NJ: Erlbaum; 2000. p. 1-90.
5. Raz, N. The aging brain observed in vivo: Differential changes and their modifiers. In: Cabeza, R.;Nyberg, L.; Park, D., editors. Cognitive neuroscience of aging: Linking cognitive and cerebralaging. Oxford: Oxford University Press; 2005. p. 19-57.
6. Dennis, NA.; Cabeza, R. Neuroimaging of healthy cognitive aging. In: Craik, FIM.; Salthouse, TA.,editors. The handbook of aging and cognition. 3rd ed.. New York: Psychology Press; 2008. p. 1-54.
7. Grady CL. Cognitive neuroscience of aging. Ann N Y Acad Sci. 2008; 1124:127–144. [PubMed:18400928]
8. Park DC, Reuter-Lorenz P. The adaptive brain: aging and neurocognitive scaffolding. Annu RevPsychol. 2009; 60:173–196. [PubMed: 19035823]
9. Johansen-Berg, H.; Behrens, TE. Diffusion MRI: From quantitative measurement to In vivoneuroanatomy. San Diego, CA: Elsevier; 2009.
10. Jones, DK. Diffusion MRI: Theory, methods, and applications. New York: Oxford UniversityPress; 2011.
11. Mori S. Introduction to diffusion tensor imaging, Elsevier. Amsterdam. 200712. Burgmans S, Gronenschild EH, Fandakova Y, Shing YL, van Boxtel MP, Vuurman EF, Uylings
HB, Jolles J, Raz N. Age differences in speed of processing are partially mediated by differencesin axonal integrity. Neuroimage. 2011
13. Giorgio A, Santelli L, Tomassini V, Bosnell R, Smith S, De Stefano N, Johansen-Berg H. Age-related changes in grey and white matter structure throughout adulthood. Neuroimage. 2010;51:943–951. [PubMed: 20211265]
14. Hugenschmidt CE, Peiffer AM, Kraft RA, Casanova R, Deibler AR, Burdette JH, Maldjian JA,Laurienti PJ. Relating imaging indices of white matter integrity and volume in healthy olderadults. Cereb Cortex. 2008; 18:433–442. [PubMed: 17575289]
15. Johansen-Berg H, Behrens TE. Just pretty pictures? What diffusion tractography can add in clinicalneuroscience. Curr Opin Neurol. 2006; 19:379–385. [PubMed: 16914977]
16. Madden DJ, Bennett IJ, Song AW. Cerebral white matter integrity and cognitive aging:contributions from diffusion tensor imaging. Neuropsychol Rev. 2009; 19:415–435. [PubMed:19705281]
17. Sullivan EV, Pfefferbaum A. Diffusion tensor imaging and aging. Neurosci Biobehav Rev. 2006;30:749–761. [PubMed: 16887187]
18. Sullivan, EV.; Pfefferbaum, A. Diffusion tensor imaging in aging and age-relatedneurodegenerative disorders. In: Jones, DK., editor. Diffusion MRI: Theory, methods, andapplications. New York: Oxford University Press; 2011. p. 624-643.
Madden et al. Page 14
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

19. Raz N, Rodrigue KM, Kennedy KM, Acker JD. Vascular health and longitudinal changes in brainand cognition in middle-aged and older adults. Neuropsychology. 2007; 21:149–157. [PubMed:17402815]
20. Raz N, Yang YQ, Rodrigue KM, Kennedy KM, Lindenberger U, Ghisletta P. White matterdeterioration in 15 months: latent growth curve models in healthy adults. Neurobiol Aging. 2010
21. Resnick SM, Goldszal AF, Davatzikos C, Golski S, Kraut MA, Metter EJ, Bryan RN, ZondermanAB. One-year age changes in MRI brain volumes in older adults. Cereb Cortex. 2000; 10:464–472. [PubMed: 10847596]
22. Resnick SM, Pham DL, Kraut MA, Zonderman AB, Davatzikos C. Longitudinal magneticresonance imaging studies of older adults: a shrinking brain. J Neurosci. 2003; 23:3295–3301.[PubMed: 12716936]
23. Barrick TR, Charlton RA, Clark CA, Markus HS. White matter structural decline in normal ageing:a prospective longitudinal study using tract-based spatial statistics. Neuroimage. 2010; 51:565–577. [PubMed: 20178850]
24. Charlton RA, Schiavone F, Barrick TR, Morris RG, Markus HS. Diffusion tensor imaging detectsage related white matter change over a 2 year follow-up which is associated with working memorydecline. J Neurol Neurosurg Psychiatry. 2010; 81:13–19. [PubMed: 19710050]
25. Sullivan EV, Rohlfing T, Pfefferbaum A. Longitudinal study of callosal microstructure in thenormal adult aging brain using quantitative DTI fiber tracking. Dev Neuropsychol. 2010; 35:233–256. [PubMed: 20446131]
26. Basser PJ. Inferring microstructural features and the physiological state of tissues from diffusion-weighted images. NMR Biomed. 1995; 8:333–344. [PubMed: 8739270]
27. Pierpaoli C, Basser PJ. Toward a quantitative assessment of diffusion anisotropy. Magn ResonMed. 1996; 36:893–906. [PubMed: 8946355]
28. Pierpaoli C, Jezzard P, Basser PJ, Barnett A, Di Chiro G. Diffusion tensor MR imaging of thehuman brain. Radiology. 1996; 201:637–648. [PubMed: 8939209]
29. Song SK, Sun SW, Ramsbottom MJ, Chang C, Russell J, Cross AH. Dysmyelination revealedthrough MRI as increased radial (but unchanged axial) diffusion of water. Neuroimage. 2002;17:1429–1436. [PubMed: 12414282]
30. Beaulieu C. The basis of anisotropic water diffusion in the nervous system - a technical review.NMR Biomed. 2002; 15:435–455. [PubMed: 12489094]
31. Sen PN, Basser PJ. A model for diffusion in white matter in the brain. Biophys J. 2005; 89:2927–2938. [PubMed: 16100258]
32. Smith SM, Jenkinson M, Johansen-Berg H, Rueckert D, Nichols TE, Mackay CE, Watkins KE,Ciccarelli O, Cader MZ, Matthews PM, Behrens TE. Tract-based spatial statistics: voxelwiseanalysis of multi-subject diffusion data. Neuroimage. 2006; 31:1487–1505. [PubMed: 16624579]
33. Song SK, Sun SW, Ju WK, Lin SJ, Cross AH, Neufeld AH. Diffusion tensor imaging detects anddifferentiates axon and myelin degeneration in mouse optic nerve after retinal ischemia.Neuroimage. 2003; 20:1714–1722. [PubMed: 14642481]
34. Song SK, Yoshino J, Le TQ, Lin SJ, Sun SW, Cross AH, Armstrong RC. Demyelination increasesradial diffusivity in corpus callosum of mouse brain. Neuroimage. 2005; 26:132–140. [PubMed:15862213]
35. Sun SW, Liang HF, Le TQ, Armstrong RC, Cross AH, Song SK. Differential sensitivity of in vivoand ex vivo diffusion tensor imaging to evolving optic nerve injury in mice with retinal ischemia.Neuroimage. 2006; 32:1195–1204. [PubMed: 16797189]
36. Sun SW, Liang HF, Cross AH, Song SK. Evolving Wallerian degeneration after transient retinalischemia in mice characterized by diffusion tensor imaging. Neuroimage. 2008; 40:1–10.[PubMed: 18187343]
37. Pierpaoli C, Barnett A, Pajevic S, Chen R, Penix LR, Virta A, Basser P. Water diffusion changes inWallerian degeneration and their dependence on white matter architecture. Neuroimage. 2001;13:1174–1185. [PubMed: 11352623]
38. Wheeler-Kingshott CA, Cercignani M. About "axial" and "radial" diffusivities. Magn Reson Med.2009; 61:1255–1260. [PubMed: 19253405]
Madden et al. Page 15
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

39. Assaf Y, Pasternak O. Diffusion tensor imaging (DTI)-based white matter mapping in brainresearch: a review. J Mol Neurosci. 2008; 34:51–61. [PubMed: 18157658]
40. Cohen-Adad J, El Mendili MM, Lehericy S, Pradat PF, Blancho S, Rossignol S, Benali H.Demyelination and degeneration in the injured human spinal cord detected with diffusion andmagnetization transfer MRI. Neuroimage. 2011; 55:1024–1033. [PubMed: 21232610]
41. Thomalla G, Glauche V, Koch MA, Beaulieu C, Weiller C, Rother J. Diffusion tensor imagingdetects early Wallerian degeneration of the pyramidal tract after ischemic stroke. Neuroimage.2004; 22:1767–1774. [PubMed: 15275932]
42. Thomalla G, Glauche V, Weiller C, Rother J. Time course of wallerian degeneration afterischaemic stroke revealed by diffusion tensor imaging. J Neurol Neurosurg Psychiatry. 2005;76:266–268. [PubMed: 15654048]
43. Aboitiz F, Rodriguez E, Olivares R, Zaidel E. Age-related changes in fibre composition of thehuman corpus callosum: sex differences. Neuroreport. 1996; 7:1761–1764. [PubMed: 8905659]
44. Marner L, Nyengaard JR, Tang Y, Pakkenberg B. Marked loss of myelinated nerve fibers in thehuman brain with age. J Comp Neurol. 2003; 462:144–152. [PubMed: 12794739]
45. Meier-Ruge W, Ulrich J, Bruhlmann M, Meier E. Age-related white matter atrophy in the humanbrain. Ann N Y Acad Sci. 1992; 673:260–269. [PubMed: 1485724]
46. Peters A. The effects of normal aging on myelin and nerve fibers: a review. J Neurocytol. 2002;31:581–593. [PubMed: 14501200]
47. Tang Y, Nyengaard JR, Pakkenberg B, Gundersen HJ. Age-induced white matter changes in thehuman brain: a stereological investigation. Neurobiol Aging. 1997; 18:609–615. [PubMed:9461058]
48. Pfefferbaum A, Sullivan EV, Hedehus M, Lim KO, Adalsteinsson E, Moseley M. Age-relateddecline in brain white matter anisotropy measured with spatially corrected echo-planar diffusiontensor imaging. Magn Reson Med. 2000; 44:259–268. [PubMed: 10918325]
49. Sullivan EV, Adalsteinsson E, Hedehus M, Ju C, Moseley M, Lim KO, Pfefferbaum A. Equivalentdisruption of regional white matter microstructure in ageing healthy men and women. Neuroreport.2001; 12:99–104. [PubMed: 11201100]
50. Sullivan EV, Pfefferbaum A. Diffusion tensor imaging in normal aging and neuropsychiatricdisorders. Eur J Radiol. 2003; 45:244–255. [PubMed: 12595109]
51. Bennett IJ, Madden DJ, Vaidya CJ, Howard DV, Howard JH Jr. Age-related differences inmultiple measures of white matter integrity: A diffusion tensor imaging study of healthy aging.Hum Brain Mapp. 2010; 31:378–390. [PubMed: 19662658]
52. Bucur B, Madden DJ, Spaniol J, Provenzale JM, Cabeza R, White LE, Huettel SA. Age-relatedslowing of memory retrieval: contributions of perceptual speed and cerebral white matter integrity.Neurobiol Aging. 2008; 29:1070–1079. [PubMed: 17383774]
53. Burzynska AZ, Preuschhof C, Backman L, Nyberg L, Li SC, Lindenberger U, Heekeren HR. Age-related differences in white matter microstructure: region-specific patterns of diffusivity.Neuroimage. 2010; 49:2104–2112. [PubMed: 19782758]
54. Davis SW, Dennis NA, Buchler NG, White LE, Madden DJ, Cabeza R. Assessing the effects ofage on long white matter tracts using diffusion tensor tractography. Neuroimage. 2009; 46:530–541. [PubMed: 19385018]
55. Madden DJ, Spaniol J, Costello MC, Bucur B, White LE, Cabeza R, Davis SW, Dennis NA,Provenzale JM, Huettel SA. Cerebral white matter integrity mediates adult age differences incognitive performance. J Cogn Neurosci. 2009; 21:289–302. [PubMed: 18564054]
56. Madden DJ, Spaniol J, Whiting WL, Bucur B, Provenzale JM, Cabeza R, White LE, Huettel SA.Adult age differences in the functional neuroanatomy of visual attention: A combined fMRI andDTI study. Neurobiol Aging. 2007; 28:459–476. [PubMed: 16500004]
57. Madden DJ, Whiting WL, Huettel SA, White LE, MacFall JR, Provenzale JM. Diffusion tensorimaging of adult age differences in cerebral white matter: relation to response time. Neuroimage.2004; 21:1174–1181. [PubMed: 15006684]
58. Salat DH, Tuch DS, Greve DN, van der Kouwe AJ, Hevelone ND, Zaleta AK, Rosen BR, Fischl B,Corkin S, Rosas HD, Dale AM. Age-related alterations in white matter microstructure measuredby diffusion tensor imaging. Neurobiol Aging. 2005; 26:1215–1227. [PubMed: 15917106]
Madden et al. Page 16
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

59. Greenwood PM. The frontal aging hypothesis evaluated. J Int Neuropsychol Soc. 2000; 6:705–726.[PubMed: 11011517]
60. Moscovitch, M.; Winocur, G. The neuropsychology of memory and aging. In: Craik, FIM.;Salthouse, TA., editors. The handbook of aging and cognition. Hillsdale, NJ: Erlbaum; 1992. p.315-372.
61. West RL. An application of prefrontal cortex function theory to cognitive aging. Psychol Bull.1996; 120:272–292. [PubMed: 8831298]
62. Sullivan EV, Rohlfing T, Pfefferbaum A. Quantitative fiber tracking of lateral andinterhemispheric white matter systems in normal aging: relations to timed performance. NeurobiolAging. 2010; 31:464–481. [PubMed: 18495300]
63. Sullivan EV, Zahr NM, Rohlfing T, Pfefferbaum A. Fiber tracking functionally distinctcomponents of the internal capsule. Neuropsychologia. 2010; 48:4155–4163. [PubMed: 20974161]
64. Zahr NM, Rohlfing T, Pfefferbaum A, Sullivan EV. Problem solving, working memory, and motorcorrelates of association and commissural fiber bundles in normal aging: a quantitative fibertracking study. Neuroimage. 2009; 44:1050–1062. [PubMed: 18977450]
65. Bartzokis G, Sultzer D, Lu PH, Nuechterlein KH, Mintz J, Cummings JL. Heterogeneous age-related breakdown of white matter structural integrity: Implications for cortical "disconnection" inaging and Alzheimer's disease. Neurobiol Aging. 2004; 25:843–851. [PubMed: 15212838]
66. Bartzokis G. Age-related myelin breakdown: a developmental model of cognitive decline andAlzheimer's disease. Neurobiol Aging. 2004; 25:5–18. [PubMed: 14675724]
67. Kochunov P, Thompson PM, Lancaster JL, Bartzokis G, Smith S, Coyle T, Royall DR, Laird A,Fox PT. Relationship between white matter fractional anisotropy and other indices of cerebralhealth in normal aging: tract-based spatial statistics study of aging. Neuroimage. 2007; 35:478–487. [PubMed: 17292629]
68. Catani M, ffytche DH. The rises and falls of disconnection syndromes. Brain. 2005; 128:2224–2239. [PubMed: 16141282]
69. O'Sullivan M, Jones DK, Summers PE, Morris RG, Williams SC, Markus HS. Evidence forcortical "disconnection" as a mechanism of age-related cognitive decline. Neurology. 2001;57:632–638. [PubMed: 11524471]
70. Madden DJ, Costello MC, Dennis NA, Davis SW, Shepler AM, Spaniol J, Bucur B, Cabeza R.Adult age differences in functional connectivity during executive control. Neuroimage. 2010;52:643–657. [PubMed: 20434565]
71. Bhagat YA, Beaulieu C. Diffusion anisotropy in subcortical white matter and cortical gray matter:changes with aging and the role of CSF-suppression. J Magn Reson Imaging. 2004; 20:216–227.[PubMed: 15269946]
72. Zhang Y, Du AT, Hayasaka S, Jahng GH, Hlavin J, Zhan W, Weiner MW, Schuff N. Patterns ofage-related water diffusion changes in human brain by concordance and discordance analysis.Neurobiol Aging. 2010; 31:1991–2001. [PubMed: 19036473]
73. Norris DG. The effects of microscopic tissue parameters on the diffusion weighted magneticresonance imaging experiment. NMR in Biomedicine. 2001; 14:77–93. [PubMed: 11320535]
74. Klawiter EC, Schmidt RE, Trinkaus K, Liang HF, Budde MD, Naismith RT, Song SK, Cross AH,Benzinger TL. Radial diffusivity predicts demyelination in ex vivo multiple sclerosis spinal cords.Neuroimage. 2011; 55:1454–1460. [PubMed: 21238597]
75. Schmierer K, Wheeler-Kingshott CA, Tozer DJ, Boulby PA, Parkes HG, Yousry TA, Scaravilli F,Barker GJ, Tofts PS, Miller DH. Quantitative magnetic resonance of postmortem multiplesclerosis brain before and after fixation. Magn Reson Med. 2008; 59:268–277. [PubMed:18228601]
76. Vargas ME, Barres BA. Why is Wallerian degeneration in the CNS so slow? Annu Rev Neurosci.2007; 30:153–179. [PubMed: 17506644]
77. Wang S, Wu EX, Qiu D, Leung LH, Lau HF, Khong PL. Longitudinal diffusion tensor magneticresonance imaging study of radiation-induced white matter damage in a rat model. Cancer Res.2009; 69:1190–1198. [PubMed: 19155304]
Madden et al. Page 17
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

78. Concha L, Gross DW, Wheatley BM, Beaulieu C. Diffusion tensor imaging of time-dependentaxonal and myelin degradation after corpus callosotomy in epilepsy patients. Neuroimage. 2006;32:1090–1099. [PubMed: 16765064]
79. Naismith RT, Xu J, Tutlam NT, Snyder A, Benzinger T, Shimony J, Shepherd J, Trinkaus K, CrossAH, Song SK. Disability in optic neuritis correlates with diffusion tensor-derived directionaldiffusivities. Neurology. 2009; 72:589–594. [PubMed: 19073948]
80. Feng S, Hong Y, Zhou Z, Jinsong Z, Xiaofeng D, Zaizhong W, Yali G, Ying L, Yingjuan C, Yi H.Monitoring of acute axonal injury in the swine spinal cord with EAE by diffusion tensor imaging.J Magn Reson Imaging. 2009; 30:277–285. [PubMed: 19629996]
81. Sun SW, Liang HF, Trinkaus K, Cross AH, Armstrong RC, Song SK. Noninvasive detection ofcuprizone induced axonal damage and demyelination in the mouse corpus callosum. Magn ResonMed. 2006; 55:302–308. [PubMed: 16408263]
82. Thuen M, Olsen O, Berry M, Pedersen TB, Kristoffersen A, Haraldseth O, Sandvig A, Brekken C.Combination of Mn(2+)-enhanced and diffusion tensor MR imaging gives complementaryinformation about injury and regeneration in the adult rat optic nerve. J Magn Reson Imaging.2009; 29:39–51. [PubMed: 19097077]
83. Mac Donald CL, Dikranian K, Bayly P, Holtzman D, Brody D. Diffusion tensor imaging reliablydetects experimental traumatic axonal injury and indicates approximate time of injury. J Neurosci.2007; 27:11869–11876. [PubMed: 17978027]
84. Mac Donald CL, Dikranian K, Song SK, Bayly PV, Holtzman DM, Brody DL. Detection oftraumatic axonal injury with diffusion tensor imaging in a mouse model of traumatic brain injury.Exp Neurol. 2007; 205:116–131. [PubMed: 17368446]
85. Li J, Li XY, Feng DF, Gu L. Quantitative evaluation of microscopic injury with diffusion tensorimaging in a rat model of diffuse axonal injury. Eur J Neurosci. 2011; 33:933–945. [PubMed:21385236]
86. Vos SB, Jones DK, Viergever MA, Leemans A. Partial volume effect as a hidden covariate in DTIanalyses. Neuroimage. 55:1566–1576. [PubMed: 21262366]
87. Miller KL, Stagg CJ, Douaud G, Jbabdi S, Smith SM, Behrens TE, Jenkinson M, Chance SA, EsiriMM, Voets NL, Jenkinson N, Aziz TZ, Turner M, Johansen-Berg H, McNab JA. Diffusionimaging of whole, post-mortem human brains on a clinical MRI scanner. Neuroimage. 2011
88. Pfefferbaum A, Sullivan EV. Increased brain white matter diffusivity in normal adult aging:relationship to anisotropy and partial voluming. Magn Reson Med. 2003; 49:953–961. [PubMed:12704779]
89. Baltes PB, Lindenberger U. Emergence of a powerful connection between sensory and cognitivefunctions across the adult life span: a new window to the study of cognitive aging? Psychol Aging.1997; 12:12–21. [PubMed: 9100264]
90. Craik, FIM.; Salthouse, TA. The handbook of aging and cognition. 3rd ed.. New York: PsychologyPress; 2008.
91. Salthouse TA. The processing-speed theory of adult age differences in cognition. Psychol Rev.1996; 103:403–428. [PubMed: 8759042]
92. Salthouse, TA. Mechanisms of age-cognition relations in adulthood. Hillsdale: NJ: Erlbaum; 1992.93. DeCarli C, Murphy DG, Tranh M, Grady CL, Haxby JV, Gillette JA, Salerno JA, Gonzales-Aviles
A, Horwitz B, Rapoport SI, et al. The effect of white matter hyperintensity volume on brainstructure, cognitive performance, and cerebral metabolism of glucose in 51 healthy adults.Neurology. 1995; 45:2077–2084. [PubMed: 7501162]
94. Leaper SA, Murray AD, Lemmon HA, Staff RT, Deary IJ, Crawford JR, Whalley LJ.Neuropsychologic correlates of brain white matter lesions depicted on MR images: 1921 AberdeenBirth Cohort. Radiology. 2001; 221:51–55. [PubMed: 11568320]
95. Nordahl CW, Ranganath C, Yonelinas AP, Decarli C, Fletcher E, Jagust WJ. White matter changescompromise prefrontal cortex function in healthy elderly individuals. J Cogn Neurosci. 2006;18:418–429. [PubMed: 16513006]
96. Prins ND, van Dijk EJ, den Heijer T, Vermeer SE, Jolles J, Koudstaal PJ, Hofman A, BretelerMM. Cerebral small-vessel disease and decline in information processing speed, executivefunction and memory. Brain. 2005; 128:2034–2041. [PubMed: 15947059]
Madden et al. Page 18
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

97. Rabbitt P, Scott M, Lunn M, Thacker N, Lowe C, Pendleton N, Horan M, Jackson A. White matterlesions account for all age-related declines in speed but not in intelligence. Neuropsychology.2007; 21:363–370. [PubMed: 17484599]
98. van Dijk EJ, Breteler MM, Schmidt R, Berger K, Nilsson LG, Oudkerk M, Pajak A, Sans S, deRidder M, Dufouil C, Fuhrer R, Giampaoli S, Launer LJ, Hofman A. The association betweenblood pressure, hypertension, and cerebral white matter lesions: cardiovascular determinants ofdementia study. Hypertension. 2004; 44:625–630. [PubMed: 15466662]
99. Salthouse TA. Neuroanatomical substrates of age-related cognitive decline. Psychological Bulletin.2011 in press.
100. Baron RM, Kenny DA. The moderator-mediator variable distinction in social psychologicalresearch: conceptual, strategic, and statistical considerations. J Pers Soc Psychol. 1986; 51:1173–1182. [PubMed: 3806354]
101. Kochunov P, Coyle T, Lancaster J, Robin DA, Hardies J, Kochunov V, Bartzokis G, Stanley J,Royall D, Schlosser AE, Null M, Fox PT. Processing speed is correlated with cerebral healthmarkers in the frontal lobes as quantified by neuroimaging. Neuroimage. 2010; 49:1190–1199.[PubMed: 19796691]
102. van Norden AG, de Laat KF, van Dijk EJ, van Uden IW, van Oudheusden LJ, Gons RA, NorrisDG, Zwiers MP, de Leeuw FE. Diffusion tensor imaging and cognition in cerebral small vesseldisease The RUN DMC study. Biochim Biophys Acta. 2011
103. Penke L, Munoz Maniega S, Murray C, Gow AJ, Hernandez MC, Clayden JD, Starr JM,Wardlaw JM, Bastin ME, Deary IJ. A general factor of brain white matter integrity predictsinformation processing speed in healthy older people. J Neurosci. 2010; 30:7569–7574.[PubMed: 20519531]
104. Schiavone F, Charlton RA, Barrick TR, Morris RG, Markus HS. Imaging age-related cognitivedecline: A comparison of diffusion tensor and magnetization transfer MRI. J Magn ResonImaging. 2009; 29:23–30. [PubMed: 19097099]
105. Bartzokis G, Lu PH, Tingus K, Mendez MF, Richard A, Peters DG, Oluwadara B, Barrall KA,Finn JP, Villablanca P, Thompson PM, Mintz J. Lifespan trajectory of myelin integrity andmaximum motor speed. Neurobiol Aging. 2010; 31:1554–1562. [PubMed: 18926601]
106. Lovden M, Bodammer NC, Kuhn S, Kaufmann J, Schutze H, Tempelmann C, Heinze HJ, DuzelE, Schmiedek F, Lindenberger U. Experience-dependent plasticity of white-matter microstructureextends into old age. Neuropsychologia. 2010; 48:3878–3883. [PubMed: 20816877]
107. Andrews-Hanna JR, Snyder AZ, Vincent JL, Lustig C, Head D, Raichle ME, Buckner RL.Disruption of large-scale brain systems in advanced aging. Neuron. 2007; 56:924–935. [PubMed:18054866]
108. Kennedy KM, Raz N. Aging white matter and cognition: differential effects of regional variationsin diffusion properties on memory, executive functions, and speed. Neuropsychologia. 2009;47:916–927. [PubMed: 19166865]
109. Ystad M, Hodneland E, Adolfsdottir S, Haasz J, Lundervold AJ, Eichele T, Lundervold A.Cortico-striatal connectivity and cognition in normal aging: A combined DTI and resting statefMRI study. Neuroimage. 2011; 55:24–31. [PubMed: 21073962]
110. Rosenthal R, DiMatteo MR. Meta-analysis: recent developments in quantitative methods forliterature reviews. Annu Rev Psychol. 2001; 52:59–82. [PubMed: 11148299]
111. Vernooij MW, Ikram MA, Vrooman HA, Wielopolski PA, Krestin GP, Hofman A, Niessen WJ,Van der Lugt A, Breteler MM. White matter microstructural integrity and cognitive function in ageneral elderly population. Arch Gen Psychiatry. 2009; 66:545–553. [PubMed: 19414714]
112. Bennett IJ, Motes MA, Rao NK, Rypma B. White matter tract integrity predicts visual searchperformance in young and older adults. Neurobiol Aging. 2011
113. Colcombe SJ, Kramer AF, McAuley E, Erickson KI, Scalf P. Neurocognitive aging andcardiovascular fitness: recent findings and future directions. J Mol Neurosci. 2004; 24:9–14.[PubMed: 15314244]
114. Colcombe S, Kramer AF. Fitness effects on the cognitive function of older adults: a meta-analyticstudy. Psychol Sci. 2003; 14:125–130. [PubMed: 12661673]
Madden et al. Page 19
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

115. Marks BL, Madden DJ, Bucur B, Provenzale JM, White LE, Cabeza R, Huettel SA. Role ofaerobic fitness and aging on cerebral white matter integrity. Ann N Y Acad Sci. 2007; 1097:171–174. [PubMed: 17413020]
116. Gold BT, Powell DK, Xuan L, Jicha GA, Smith CD. Age-related slowing of task switching isassociated with decreased integrity of frontoparietal white matter. Neurobiol Aging. 2010;31:512–522. [PubMed: 18495298]
117. Perry ME, McDonald CR, Hagler DJ Jr, Gharapetian L, Kuperman JM, Koyama AK, Dale AM,McEvoy LK. White matter tracts associated with set-shifting in healthy aging. Neuropsychologia.2009; 47:2835–2842. [PubMed: 19540862]
118. Bennett IJ, Madden DJ, Vaidya CJ, Howard JH Jr, Howard DV. White matter integrity correlatesof implicit sequence learning in healthy aging. Neurobiol Aging. 2010
119. Salthouse TA, Kausler DH, Saults JS. Utilization of path-analytic procedures to investigate therole of processing resources in cognitive aging. Psychol Aging. 1988; 3:158–166. [PubMed:3268254]
120. Charlton RA, Landau S, Schiavone F, Barrick TR, Clark CA, Markus HS, Morris RG. Astructural equation modeling investigation of age-related variance in executive function and DTImeasured white matter damage. Neurobiol Aging. 2008; 29:1547–1555. [PubMed: 17451845]
121. Voineskos AN, Rajji TK, Lobaugh NJ, Miranda D, Shenton ME, Kennedy JL, Pollock BG,Mulsant BH. Age-related decline in white matter tract integrity and cognitive performance: ADTI tractography and structural equation modeling study. Neurobiol Aging. 2010
122. Charlton RA, Landau S, Schiavone F, Barrick TR, Clark CA, Markus HS, Morris RG, et al. Upthe Garden Path: A critique of Penke and Deary (in press) and further exploration concerning theCharlton et al. (2008) path analysis relating loss of white matter integrity to cognition in normalaging. Neurobiol Aging. 2010
123. Penke L, Deary IJ, et al. Some guidelines for structural equation modelling in cognitiveneuroscience: the case of Charlton et al.'s study on white matter integrity and cognitive ageing.Neurobiol Aging. 2010; 31:1656–1660. [PubMed: 20079555]
124. Wahl M, Li YO, Ng J, Lahue SC, Cooper SR, Sherr EH, Mukherjee P. Microstructuralcorrelations of white matter tracts in the human brain. Neuroimage. 2010; 51:531–541. [PubMed:20206699]
125. Baltan S. Ischemic injury to white matter: an age-dependent process. Neuroscientist. 2009;15:126–133. [PubMed: 19307420]
126. Schonberger M, Ponsford J, Reutens D, Beare R, O'Sullivan R. The relationship between age,injury severity, and MRI findings after traumatic brain injury. J Neurotrauma. 2009; 26:2157–2167. [PubMed: 19624261]
127. Ciccarelli O, Catani M, Johansen-Berg H, Clark C, Thompson A. Diffusion-based tractography inneurological disorders: concepts, applications, and future developments. Lancet Neurol. 2008;7:715–727. [PubMed: 18635020]
128. Catani M. Diffusion tensor magnetic resonance imaging tractography in cognitive disorders. CurrOpin Neurol. 2006; 19:599–606. [PubMed: 17102700]
129. Filley CM. White matter and behavioral neurology. Ann N Y Acad Sci. 2005; 1064:162–183.[PubMed: 16394155]
130. Head D, Buckner RL, Shimony JS, Williams LE, Akbudak E, Conturo TE, McAvoy M, MorrisJC, Snyder AZ. Differential vulnerability of anterior white matter in nondemented aging withminimal acceleration in dementia of the Alzheimer type: evidence from diffusion tensor imaging.Cereb Cortex. 2004; 14:410–423. [PubMed: 15028645]
131. Damoiseaux JS, Smith SM, Witter MP, Sanz-Arigita EJ, Barkhof F, Scheltens P, Stam CJ, ZareiM, Rombouts SA. White matter tract integrity in aging and Alzheimer's disease. Hum BrainMapp. 2009; 30:1051–1059. [PubMed: 18412132]
132. Scola E, Bozzali M, Agosta F, Magnani G, Franceschi M, Sormani MP, Cercignani M, Pagani E,Falautano M, Filippi M, Falini A. A diffusion tensor MRI study of patients with MCI and ADwith a 2-year clinical follow-up. J Neurol Neurosurg Psychiatry. 2010; 81:798–805. [PubMed:20392973]
Madden et al. Page 20
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

133. Stebbins GT, Murphy CM. Diffusion tensor imaging in Alzheimer's disease and mild cognitiveimpairment. Behav Neurol. 2009; 21:39–49. [PubMed: 19847044]
134. Bosch B, Arenaza-Urquijo EM, Rami L, Sala-Llonch R, Junque C, Sole-Padulles C, Pena-GomezC, Bargallo N, Molinuevo JL, Bartres-Faz D. Multiple DTI index analysis in normal aging,amnestic MCI and AD. Relationship with neuropsychological performance. Neurobiol Aging.2010
135. Sexton CE, Mackay CE, Lonie JA, Bastin ME, Terriere E, O'Carroll RE, Ebmeier KP. MRIcorrelates of episodic memory in Alzheimer's disease, mild cognitive impairment, and healthyaging. Psychiatry Res. 2010; 184:57–62. [PubMed: 20832251]
136. Friese U, Meindl T, Herpertz SC, Reiser MF, Hampel H, Teipel SJ. Diagnostic utility of novelMRI-based biomarkers for Alzheimer's disease: diffusion tensor imaging and deformation-basedmorphometry. J Alzheimers Dis. 2010; 20:477–490. [PubMed: 20164559]
137. Bendlin BB, Ries ML, Canu C, Sodhi A, Lazar M, Alexander AL, Carlsson CM, Sager MA,Asthana S, Johnson SC. White matter is altered with parental family history of Alzheimer'sdisease. Alzheimers Dement. 2010; 6:394–403. [PubMed: 20713315]
138. Wang L, Goldstein FC, Veledar E, Levey AI, Lah JJ, Meltzer CC, Holder CA, Mao H.Alterations in cortical thickness and white matter integrity in mild cognitive impairmentmeasured by whole-brain cortical thickness mapping and diffusion tensor imaging. AJNR Am JNeuroradiol. 2009; 30:893–899. [PubMed: 19279272]
139. Wee CY, Yap PT, Li W, Denny K, Browndyke JN, Potter GG, Welsh-Bohmer KA, Wang L,Shen D. Enriched white matter connectivity networks for accurate identification of MCI patients.Neuroimage. 2011; 54:1812–1822. [PubMed: 20970508]
140. Zhang Y, Schuff N, Du AT, Rosen HJ, Kramer JH, Gorno-Tempini ML, Miller BL, Weiner MW.White matter damage in frontotemporal dementia and Alzheimer's disease measured by diffusionMRI. Brain. 2009; 132:2579–2592. [PubMed: 19439421]
141. Zarei M, Damoiseaux JS, Morgese C, Beckmann CF, Smith SM, Matthews PM, Scheltens P,Rombouts SA, Barkhof F. Regional white matter integrity differentiates between vasculardementia and Alzheimer disease. Stroke. 2009; 40:773–779. [PubMed: 19164789]
142. Matsui H, Nishinaka K, Oda M, Niikawa H, Kubori T, Udaka F. Dementia in Parkinson's disease:diffusion tensor imaging. Acta Neurol Scand. 2007; 116:177–181. [PubMed: 17714331]
143. Gattellaro G, Minati L, Grisoli M, Mariani C, Carella F, Osio M, Ciceri E, Albanese A, BruzzoneMG. White matter involvement in idiopathic Parkinson disease: a diffusion tensor imaging study.AJNR Am J Neuroradiol. 2009; 30:1222–1226. [PubMed: 19342541]
144. Matsui H, Nishinaka K, Oda M, Niikawa H, Komatsu K, Kubori T, Udaka F. Wisconsin CardSorting Test in Parkinson's disease: diffusion tensor imaging. Acta Neurol Scand. 2007;116:108–112. [PubMed: 17661796]
145. Potter GG, Steffens DC. Contribution of depression to cognitive impairment and dementia inolder adults. Neurologist. 2007; 13:105–117. [PubMed: 17495754]
146. Potter GG, McQuoid DR, Steffens DC, Welsh-Bohmer KA, Krishnan KR. Neuropsychologicalcorrelates of magnetic resonance imaging-defined subcortical ischemic depression. InternationalJournal of Geriatric Psychiatry. 2008; 24:219–225. [PubMed: 18655212]
147. Alexopoulos GS, Kiosses DN, Choi SJ, Murphy CF, Lim KO. Frontal white mattermicrostructure and treatment response of late-life depression: a preliminary study. Am JPsychiatry. 2002; 159:1929–1932. [PubMed: 12411231]
148. Taylor WD, MacFall JR, Payne ME, McQuoid DR, Provenzale JM, Steffens DC, Krishnan KR.Late-life depression and microstructural abnormalities in dorsolateral prefrontal cortex whitematter. Am J Psychiatry. 2004; 161:1293–1296. [PubMed: 15229065]
149. Yuan Y, Zhang Z, Bai F, Yu H, Shi Y, Qian Y, Zang Y, Zhu C, Liu W, You J. White matterintegrity of the whole brain is disrupted in first-episode remitted geriatric depression.NeuroReport. 2007; 18:1845–1849. [PubMed: 18090324]
150. Murphy CF, Gunning-Dixon FM, Hoptman MJ, Lim KO, Ardekani B, Shields JK, Hrabe J,Kanellopoulos D, Shanmugham BR, Alexopoulos GS. White-matter integrity predicts stroopperformance in patients with geriatric depression. Biol Psychiatry. 2007; 61:1007–1010.[PubMed: 17123478]
Madden et al. Page 21
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

151. Shimony JS, Sheline YI, D'Angelo G, Epstein AA, Benzinger TL, Mintun MA, McKinstry RC,Snyder AZ. Diffuse microstructural abnormalities of normal-appearing white matter in late lifedepression: a diffusion tensor imaging study. Biol Psychiatry. 2009; 66:245–252. [PubMed:19375071]
152. Schermuly I, Fellgiebel A, Wagner S, Yakushev I, Stoeter P, Schmitt R, Knickenberg RJ,Bleichner F, Beutel ME. Association between cingulum bundle structure and cognitiveperformance: an observational study in major depression. Eur Psychiatry. 2010; 25:355–360.[PubMed: 20621455]
153. Yogarajah M, Focke NK, Bonelli SB, Thompson P, Vollmar C, McEvoy AW, Alexander DC,Symms MR, Koepp MJ, Duncan JS. The structural plasticity of white matter networks followinganterior temporal lobe resection. Brain. 2010; 133:2348–2364. [PubMed: 20826432]
154. Sidaros A, Engberg A, Sidaros K, Liptrot MG, Herning M, Petersen P, Paulson OB, Jernigan TL,Rostrup E. Diffusion tensor imaging during recovery from severe traumatic brain injury andrelation to clinical outcome: a longitudinal study. Brain. 2008; 131:559–572. [PubMed:18083753]
155. Bendlin BB, Ries ML, Lazar M, Alexander AL, Dempsey RJ, Rowley HA, Sherman JE, JohnsonSC. Longitudinal changes in patients with traumatic brain injury assessed with diffusion-tensorand volumetric imaging. Neuroimage. 2008; 42:503–514. [PubMed: 18556217]
156. Colcombe SJ, Kramer AF, Erickson KI, Scalf P, McAuley E, Cohen NJ, Webb A, Jerome GJ,Marquez DX, Elavsky S. Cardiovascular fitness, cortical plasticity, and aging. Proc Natl AcadSci U S A. 2004; 101:3316–3321. [PubMed: 14978288]
157. Lustig C, Shah P, Seidler R, Reuter-Lorenz PA. Aging, training, and the brain: a review andfuture directions. Neuropsychol Rev. 2009; 19:504–522. [PubMed: 19876740]
158. Greenwood PM. Functional plasticity in cognitive aging: review and hypothesis.Neuropsychology. 2007; 21:657–673. [PubMed: 17983277]
159. Reese TG, Heid O, Weisskoff RM, Wedeen VJ. Reduction of eddy-current-induced distortion indiffusion MRI using a twice-refocused spin echo. Magn Reson Med. 2003; 49:177–182.[PubMed: 12509835]
160. Chen B, Guo H, Song AW. Correction for direction-dependent distortions in diffusion tensorimaging using matched magnetic field maps. Neuroimage. 2006; 30:121–129. [PubMed:16242966]
161. Truong TK, Chen B, Song AW. Integrated SENSE DTI with correction of susceptibility- andeddy current-induced geometric distortions. Neuroimage. 2008; 40:53–58. [PubMed: 18187344]
162. Truong TK, Chen NK, Song AW. Dynamic correction of artifacts due to susceptibility effects andtime-varying eddy currents in diffusion tensor imaging. Neuroimage. 2011; 57:1343–1347.[PubMed: 21689763]
163. Porter DA, Heidemann RM. High resolution diffusion-weighted imaging using readout-segmented echo-planar imaging, parallel imaging and a two-dimensional navigator-basedreacquisition. Magn Reson Med. 2009; 62:468–475. [PubMed: 19449372]
164. Holdsworth SJ, Skare S, Newbould RD, Bammer R. Robust GRAPPA-accelerated diffusion-weighted readout-segmented (RS)-EPI. Magn Reson Med. 2009; 62:1629–1640. [PubMed:19859974]
165. Liu C, Bammer R, Kim DH, Moseley ME. Self-navigated interleaved spiral (SNAILS):Application to high-resolution diffusion tensor imaging. Magn Reson Med. 2004; 52:1388–1396.[PubMed: 15562493]
166. Pipe JG, Farthing VG, Forbes KP. Multishot diffusion-weighted FSE using PROPELLER MRI.Magn Reson Med. 2002; 47:42–52. [PubMed: 11754441]
167. Wang FN, Huang TY, Lin FH, Chuang TC, Chen NK, Chung HW, Chen CY, Kwong KK.PROPELLER EPI: an MRI technique suitable for diffusion tensor imaging at high field strengthwith reduced geometric distortions. Magn Reson Med. 2005; 54:1232–1240. [PubMed:16206142]
168. Calamante F, Tournier JD, Jackson GD, Connelly A. Track-density imaging (TDI): super-resolution white matter imaging using whole-brain track-density mapping. Neuroimage. 2010;53:1233–1243. [PubMed: 20643215]
Madden et al. Page 22
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

169. Calamante F, Tournier JD, Heidemann RM, Anwander A, Jackson GD, Connelly A. Trackdensity imaging (TDI): Validation of super resolution property. Neuroimage. 2011; 56:1259–1266. [PubMed: 21354314]
170. Basser PJ. Relationships between diffusion tensor and q-space MRI. Magn Reson Med. 2002;47:392–397. [PubMed: 11810685]
171. Assaf Y, Cohen Y. Structural information in neuronal tissue as revealed by q-space diffusionNMR spectroscopy of metabolites in bovine optic nerve. NMR Biomed. 1999; 12:335–344.[PubMed: 10516615]
172. King MD, Houseman J, Roussel SA, van Bruggen N, Williams SR, Gadian DG. q-Space imagingof the brain. Magn Reson Med. 1994; 32:707–713. [PubMed: 7869892]
173. Gilbert RJ, Wedeen VJ, Magnusson LH, Benner T, Wang R, Dai G, Napadow VJ, Roche KK.Three-dimensional myoarchitecture of the bovine tongue demonstrated by diffusion spectrummagnetic resonance imaging with tractography. Anat Rec A Discov Mol Cell Evol Biol. 2006;288:1173–1182. [PubMed: 17031810]
174. Schmahmann JD, Pandya DN, Wang R, Dai G, D'Arceuil HE, de Crespigny AJ, Wedeen VJ.Association fibre pathways of the brain: parallel observations from diffusion spectrum imagingand autoradiography. Brain. 2007; 130:630–653. [PubMed: 17293361]
175. Wedeen VJ, Wang RP, Schmahmann JD, Benner T, Tseng WY, Dai G, Pandya DN, Hagmann P,D'Arceuil H, de Crespigny AJ. Diffusion spectrum magnetic resonance imaging (DSI)tractography of crossing fibers. Neuroimage. 2008; 41:1267–1277. [PubMed: 18495497]
176. Frank LR. Anisotropy in high angular resolution diffusion-weighted MRI. Magn Reson Med.2001; 45:935–939. [PubMed: 11378869]
177. Frank LR. Characterization of anisotropy in high angular resolution diffusion-weighted MRI.Magn Reson Med. 2002; 47:1083–1099. [PubMed: 12111955]
178. Tuch DS, Reese TG, Wiegell MR, Wedeen VJ. Diffusion MRI of complex neural architecture.Neuron. 2003; 40:885–895. [PubMed: 14659088]
179. Liu C, Mang SC, Moseley ME. In vivo generalized diffusion tensor imaging (GDTI) usinghigher-order tensors (HOT). Magn Reson Med. 2010; 63:243–252. [PubMed: 19953513]
180. Catani M. From hodology to function. Brain. 2007; 130:602–605. [PubMed: 17322561]181. Avram AV, Guidon A, Song AW. Myelin water weighted diffusion tensor imaging. Neuroimage.
2010; 53:132–138. [PubMed: 20587369]182. Assaf Y, Blumenfeld-Katzir T, Yovel Y, Basser PJ. AxCaliber: a method for measuring axon
diameter distribution from diffusion MRI. Magn Reson Med. 2008; 59:1347–1354. [PubMed:18506799]
183. Barazany D, Basser PJ, Assaf Y. In vivo measurement of axon diameter distribution in the corpuscallosum of rat brain. Brain. 2009; 132:1210–1220. [PubMed: 19403788]
184. Xi MC, Liu RH, Engelhardt JK, Morales FR, Chase MH. Changes in the axonal conductionvelocity of pyramidal tract neurons in the aged cat. Neuroscience. 1999; 92:219–225. [PubMed:10392844]
185. Terao S, Sobue G, Hashizume Y, Shimada N, Mitsuma T. Age-related changes of the myelinatedfibers in the human corticospinal tract: a quantitative analysis. Acta Neuropathologica. 1994;88:137–142. [PubMed: 7985494]
186. Madden, DJ. Speed and timing of behavioral processes. In: Birren, JE.; Schaie, KW., editors.Handbook of the psychology of aging. 5th ed.. San Diego, CA: Academic Press; 2001. p.288-312.
187. Salthouse, TA.; Madden, DJ. Information processing speed and aging. In: Deluca, J.; Kalmar, J.,editors. Information processing speed in clinical populations. New York: Psychology Press;2007. p. 221-241.
188. Mandl RC, Schnack HG, Zwiers MP, van der Schaaf A, Kahn RS, Hulshoff Pol HE. Functionaldiffusion tensor imaging: measuring task-related fractional anisotropy changes in the humanbrain along white matter tracts. PLoS ONE. 2008; 3:e3631. [PubMed: 18982065]
189. Sporns O, Chialvo DR, Kaiser M, Hilgetag CC. Organization, development and function ofcomplex brain networks. Trends Cogn Sci. 2004; 8:418–425. [PubMed: 15350243]
Madden et al. Page 23
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

190. Bullmore E, Sporns O. Complex brain networks: graph theoretical analysis of structural andfunctional systems. Nat Rev Neurosci. 2009; 10:186–198. [PubMed: 19190637]
191. Gong G, He Y, Concha L, Lebel C, Gross DW, Evans AC, Beaulieu C. Mapping anatomicalconnectivity patterns of human cerebral cortex using in vivo diffusion tensor imagingtractography. Cereb Cortex. 2009; 19:524–536. [PubMed: 18567609]
192. Iturria-Medina Y, Perez Fernandez A, Morris DM, Canales-Rodriguez EJ, Haroon HA, GarciaPenton L, Augath M, Galan Garcia L, Logothetis N, Parker GJ, Melie-Garcia L. Brainhemispheric structural efficiency and interconnectivity rightward asymmetry in human andnonhuman primates. Cereb Cortex. 2011; 21:56–67. [PubMed: 20382642]
193. Li Y, Liu Y, Li J, Qin W, Li K, Yu C, Jiang T. Brain anatomical network and intelligence. PLoSComput Biol. 2009; 5 e1000395.
194. Biswal BB, Mennes M, Zuo XN, Gohel S, Kelly C, Smith SM, Beckmann CF, Adelstein JS,Buckner RL, Colcombe S, Dogonowski AM, Ernst M, Fair D, Hampson M, Hoptman MJ, HydeJS, Kiviniemi VJ, Kotter R, Li SJ, Lin CP, Lowe MJ, Mackay C, Madden DJ, Madsen KH,Margulies DS, Mayberg HS, McMahon K, Monk CS, Mostofsky SH, Nagel BJ, Pekar JJ, PeltierSJ, Petersen SE, Riedl V, Rombouts SA, Rypma B, Schlaggar BL, Schmidt S, Seidler RD, SiegleGJ, Sorg C, Teng GJ, Veijola J, Villringer A, Walter M, Wang L, Weng XC, Whitfield-GabrieliS, Williamson P, Windischberger C, Zang YF, Zhang HY, Castellanos FX, Milham MP. Towarddiscovery science of human brain function. Proc Natl Acad Sci U S A. 2010; 107:4734–4739.[PubMed: 20176931]
195. Damoiseaux JS, Beckmann CF, Arigita EJ, Barkhof F, Scheltens P, Stam CJ, Smith SM,Rombouts SA. Reduced resting-state brain activity in the "default network" in normal aging.Cereb Cortex. 2008; 18:1856–1864. [PubMed: 18063564]
196. Fox MD, Greicius M. Clinical applications of resting state functional connectivity. Front SystNeurosci. 2010; 4:1–13. [PubMed: 20204156]
197. Greicius MD, Srivastava G, Reiss AL, Menon V. Default-mode network activity distinguishesAlzheimer's disease from healthy aging: evidence from functional MRI. Proc Natl Acad Sci U SA. 2004 a.101:4637–4642. [PubMed: 15070770]
198. Greicius MD, Supekar K, Menon V, Dougherty RF. Resting-state functional connectivity reflectsstructural connectivity in the default mode network. Cereb Cortex. 2009; 19:72–78. [PubMed:18403396]
199. Teipel SJ, Bokde AL, Meindl T, Amaro E Jr, Soldner J, Reiser MF, Herpertz SC, Moller HJ,Hampel H. White matter microstructure underlying default mode network connectivity in thehuman brain. Neuroimage. 2010; 49:2021–2032. [PubMed: 19878723]
200. van den Heuvel MP, Mandl RC, Kahn RS, Hulshoff Pol HE. Functionally linked resting-statenetworks reflect the underlying structural connectivity architecture of the human brain. HumBrain Mapp. 2009; 30:3127–3141. [PubMed: 19235882]
201. Chen NK, Chou YH, Song AW, Madden DJ. Measurement of spontaneous signal fluctuations infMRI: adult age differences in intrinsic functional connectivity. Brain Struct Funct. 2009;213:571–585. [PubMed: 19727810]
202. Grieve SM, Williams LM, Paul RH, Clark CR, Gordon E. Cognitive aging, executive function,and fractional anisotropy: a diffusion tensor MR imaging study. AJNR Am J Neuroradiol. 2007;28:226–235. [PubMed: 17296985]
203. Charlton RA, Barrick TR, McIntyre DJ, Shen Y, O'Sullivan M, Howe FA, Clark CA, Morris RG,Markus HS. White matter damage on diffusion tensor imaging correlates with age-relatedcognitive decline. Neurology. 2006; 66:217–222. [PubMed: 16434657]
204. Charlton RA, Barrick TR, Lawes IN, Markus HS, Morris RG. White matter pathways associatedwith working memory in normal aging. Cortex. 2010; 46:474–489. [PubMed: 19666169]
205. Ziegler DA, Piguet O, Salat DH, Prince K, Connally E, Corkin S. Cognition in healthy aging isrelated to regional white matter integrity, but not cortical thickness. Neurobiol Aging. 2010;31:1912–1926. [PubMed: 19091444]
206. Kantarci K, Senjem ML, Avula R, Zhang B, Samikoglu AR, Weigand SD, Przybelski SA,Edmonson HA, Vemuri P, Knopman DS, Boeve BF, Ivnik RJ, Smith GE, Petersen RC, Jack CR
Madden et al. Page 24
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Jr. Diffusion tensor imaging and cognitive function in older adults with no dementia. Neurology.2011
207. Deary IJ, Bastin ME, Pattie A, Clayden JD, Whalley LJ, Starr JM, Wardlaw JM. White matterintegrity and cognition in childhood and old age. Neurology. 2006; 66:505–512. [PubMed:16505302]
208. Shenkin SD, Bastin ME, MacGillivray TJ, Deary IJ, Starr JM, Wardlaw JM. Childhood andcurrent cognitive function in healthy 80-year-olds: a DT-MRI study. Neuroreport. 2003; 14:345–349. [PubMed: 12634481]
209. Shenkin SD, Bastin ME, Macgillivray TJ, Deary IJ, Starr JM, Rivers CS, Wardlaw JM. Cognitivecorrelates of cerebral white matter lesions and water diffusion tensor parameters in community-dwelling older people. Cerebrovasc Dis. 2005; 20:310–318. [PubMed: 16141715]
210. Sullivan EV, Adalsteinsson E, Pfefferbaum A. Selective age-related degradation of anteriorcallosal fiber bundles quantified in vivo with fiber tracking. Cereb Cortex. 2006; 16:1030–1039.[PubMed: 16207932]
211. Schulte T, Sullivan EV, Muller-Oehring EM, Adalsteinsson E, Pfefferbaum A. Corpus callosalmicrostructural integrity influences interhemispheric processing: a diffusion tensor imagingstudy. Cereb Cortex. 2005; 15:1384–1392. [PubMed: 15635059]
212. Davis SW, Kragel J, Madden DJ, Cabeza R. The architecture of cross-hemisphericcommunication in the aging brain: Linking behavior to functional and structural connectivity.Cereb Cortex. 2011 in press.
213. Fling BW, Walsh CM, Bangert AS, Reuter-Lorenz PA, Welsh RC, Seidler RD. Differentialcallosal contributions to bimanual control in young and older adults. J Cogn Neurosci. 2010
214. Charlton RA, Barrick TR, Markus HS, Morris RG. The relationship between episodic long-termmemory and white matter integrity in normal aging. Neuropsychologia. 2010; 48:114–122.[PubMed: 19699215]
Madden et al. Page 25
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1.Methods for representing diffusion tensor imaging (DTI) data. A = regions of interest (incolor) placed directly on DTI image; B = voxel-based morphometry (VBM); C = mean“skeleton” of white matter tracts from tract-based spatial statistics (TBSS); D = fibertracking of white matter pathways. Image courtesy of Simon Davis, University ofCambridge.
Madden et al. Page 26
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 2.Fiber tracking methodology. A = seed and target regions placed on DTI raw image; B =estimated fibers of the genu of the corpus callosum passing through the seed and targetregions; C = “tube” of averaged fibers within the genu; D = voxels within averaged fiberscolor coded by age group differences in fractional anisotropy (FA), with the magnitude ofage-related decline in FA scaled from lower (blue) to higher (red) values [54]. Adapted withpermission from Neuroimage, Volume 46/Issue 2, S. W. Davis, N. A. Dennis, N. G.Buchler, L. E. White, D. J Madden, & R. Cabeza, Assessing the effects of age on long whitematter tracts using diffusion tensor tractography, pp. 530–541, Copyright 2009, withpermission from Elsevier.
Madden et al. Page 27
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3.A = The spatial extent of five patterns of age-related difference in diffusivity, relative to thetotal volume of age-related decrease in FA values across regions (55,973 mm2), fromBurzynska et al. [53]; B = spatial localization of the main diffusivity pattern: age-relateddecrease in FA, increase in RD and MD (in red) and additionally an increase in AD (inorange); C = WMH volume accounts for age-related decrease only in a few WM regions.Age-related FA decrease (in yellow) is contrasted with age-related FA decrease afteraccounting for WMH volume (in red). FA = fractional anisotropy; RD = radial diffusivity;AD = axial diffusivity; MD = mean diffusivity; WMH = white matter Hyperintensity; ALIC= anterior limb of the internal capsule; BCC = body of the corpus callosum; EC = externalcapsule; FX = fornix; GCC = genu of the corpus callosum; MFG = middle frontal gyrus;SCC = splenium of the corpus callosum. Adapted with permission from Neuroimage,Volume 49/Issue 3, A.Z. Burzynska, C. Preuschhof, L. Backman, L. Nyberg, S. C. Li, U.Lindenberger, & H. R. Heekeren, Age-related differences in white matter microstructure:region-specific patterns of diffusivity, pp. 2104–2112, Copyright 2010, with permissionfrom Elsevier.
Madden et al. Page 28
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
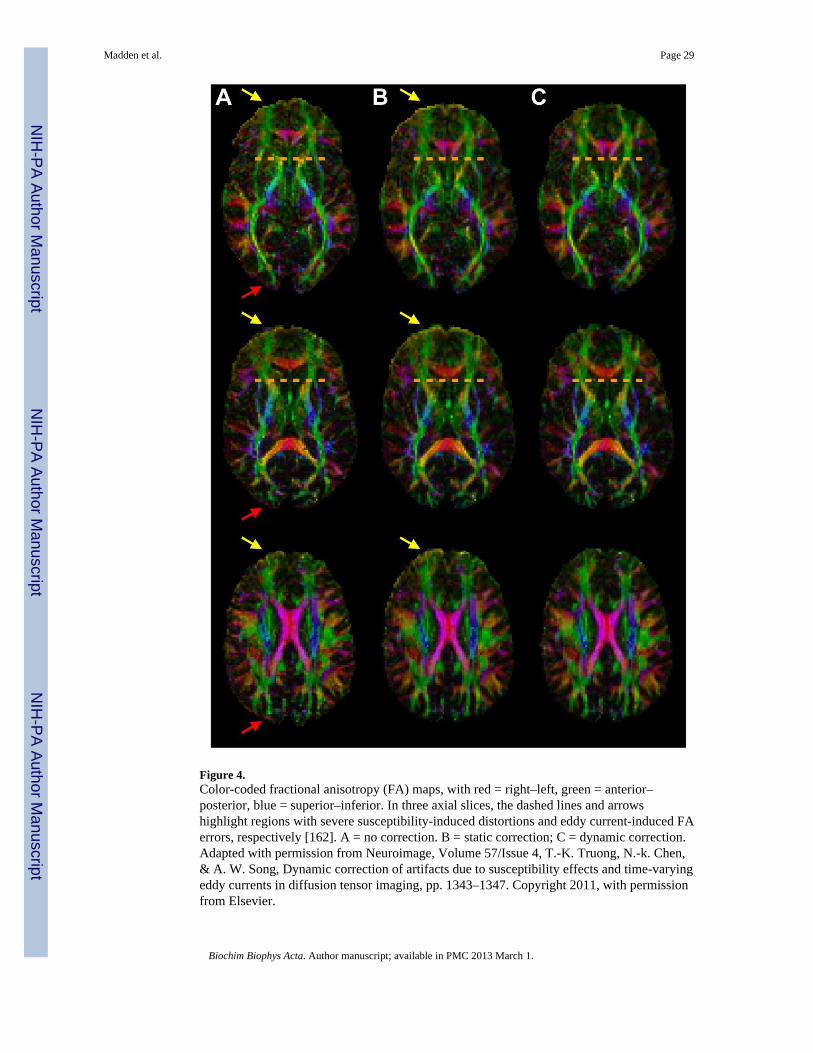
Figure 4.Color-coded fractional anisotropy (FA) maps, with red = right–left, green = anterior–posterior, blue = superior–inferior. In three axial slices, the dashed lines and arrowshighlight regions with severe susceptibility-induced distortions and eddy current-induced FAerrors, respectively [162]. A = no correction. B = static correction; C = dynamic correction.Adapted with permission from Neuroimage, Volume 57/Issue 4, T.-K. Truong, N.-k. Chen,& A. W. Song, Dynamic correction of artifacts due to susceptibility effects and time-varyingeddy currents in diffusion tensor imaging, pp. 1343–1347. Copyright 2011, with permissionfrom Elsevier.
Madden et al. Page 29
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 5.A = Diffusion spectrum imaging (DSI) tractography of monkey cerebral cortex showingfibers within the white matter of the cortical gyrus, and radiate fibers within adjacent gyri ofthe cerebral cortex (arrows); B = DTI tractography of a corresponding region shows fewerfibers within the white matter of the gyrus and no radiate fibers within the cortex [175].Reprinted from Neuroimage, Volume 41/Issue 4, V.J. Wedeen, R.P. Wang, J.D.Schmahmann, T. Benner, W.Y.I. Tseng, G. Dai, D.N. Pandya, P. Hagmann, H. D'Arceuil, &A.J. de Crespigny, Diffusion spectrum magnetic resonance imaging (DSI) tractography ofcrossing fibers, pp. 1267–1277, Copyright 2008, with permission from Elsevier.
Madden et al. Page 30
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Madden et al. Page 31
Table 1
Patterns of adult age differences in white matter diffusivity properties
Anatomical location Pattern for older adults,relative to younger adults
Biologicalinterpretation
Genu of the corpus callosum FA ↓ RD↑ AD↑ (MD↑)* Loss/disruption of both axons and myelin
Body of the corpus callosum (FA ↓ RD↑ AD↑ MD↑)*
External capsule FA ↓ RD↑ AD↑ (MD↑)*
Anterior dorsal cingulum (FA ↓ RD↑ AD↑ MD↑)*
Fornix FA ↓ RD↑ AD↑ (MD↑)* Loss/disruption of both axons and myelin; partial volume with CSF
Anterior limb of internal capsule (FA↓ RD↑ MD↑)* Predominant myelin disruption or loss
Middle frontal gyrus (SLF) FA ↓ RD↑ (AD↑ MD↑)*
WM of the straight gyrus FA↓ RD↑
Forceps major FA↓ RD↑
Sagittal stratum FA↓ RD↑ (MD↑)*
Posterior dorsal cingulum (FA ↓ RD↑)*
Superior corona radiata FA↓ RD↑ AD↓ Fiber loss with glial infiltration
Forceps minor FA↓ RD↑ AD↓
Retrolenticular internal capsule FA↓ RD↑ AD↓
Parahippocampal WM FA↓ RD↑ (AD↓)*
Posterior limb of internal capsule (FA↓ AD↓ MD↓)* Gliosis or early axonal injury/Less coherent tract organization
Cerebral peduncles FA↓ AD↓
Note. The patterns of age differences (second column) are expressed as the direction of significant diffusivity effects, for older adults, relative toyounger adults, reported by Bennett et al. [51] and Burzynska et al. [53]. FA = fractional anisotropy; AD = axial diffusivity; RD = radialdiffusivity; MD = mean diffusivity; CSF = cerebrospinal fluid; SLF = superior longitudinal fasciculus; WM = white matter; ↓ = decrease; ↑ =increase.
*These effects (within parentheses) were specific to Burzynska et al. [53]. The remaining effects were common to both Bennett et al. (2010) and
Burzynska et al. [53]. None of the effects was specific to Bennett et al. [51].
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Madden et al. Page 32
Tabl
e 2
Sum
mar
y of
agi
ng st
udie
s exa
min
ing
rela
tions
hips
bet
wee
n fr
actio
nal a
niso
tropy
(FA
) and
cog
nitiv
e pe
rfor
man
ce
Cog
nitiv
e D
omai
nB
rain
Reg
ion
Fron
tal
CC
gIn
f Fro
ntIC
Mid
Sup
CC
sPa
rO
cc
Exec
utiv
e Fu
nctio
n
You
nger
and
Old
er
C
ompo
site
mea
sure
s [63
–64,
121
]
Raw
biv
aria
te—
0.62
0.34
0.53
0.65
0.28
0.24
——
Age
par
tiale
d—
0.06
——
0.06
——
——
S
et sh
ift/ta
sk sw
itch
[108
, 116
–117
, 202
]
Raw
biv
aria
te0.
400.
650.
61—
0.22
0.45
—0.
400.
26
Age
par
tiale
d0.
25—
0.52
—0.
230.
46—
0.29
0.23
W
orki
ng m
emor
y [1
08, 2
02]
Raw
biv
aria
te0.
420.
30—
—0.
18—
—0.
470.
04
Age
par
tiale
d0.
14—
——
−0.04
——
0.18
0.03
M
emor
y sp
an [1
08, 2
02]
Raw
biv
aria
te0.
130.
28—
—0.
02—
—0.
060.
02
Age
par
tiale
d−0.02
——
—0.
00—
—0.
020.
06
Old
er o
nly
C
ompo
site
mea
sure
s [20
3–20
6]
Raw
biv
aria
te0.
380.
30—
—0.
330.
29—
0.30
—
Age
par
tiale
d0.
260.
020.
15—
0.07
0.20
—0.
05—
S
et sh
ift/ta
sk sw
itch
[54,
69,
116
]
Raw
biv
aria
te0.
41—
0.63
—0.
090.
47—
0.32
—
Age
par
tiale
d0.
09—
——
−0.34
——
−0.04
—
W
orki
ng m
emor
y [5
4, 2
07]
Raw
biv
aria
te0.
09—
0.57
——
0.41
——
0.28
Age
par
tiale
d—
——
——
——
——
M
emor
y sp
an [5
4]
Raw
biv
aria
te—
0.57
——
——
——
—
Age
par
tiale
d—
——
——
——
——
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Madden et al. Page 33
Cog
nitiv
e D
omai
nB
rain
Reg
ion
Fron
tal
CC
gIn
f Fro
ntIC
Mid
Sup
CC
sPa
rO
cc
F
luen
cy [6
9, 2
07–2
09]
Raw
biv
aria
te0.
09—
——
0.69
0.20
—0.
370.
20
Age
par
tiale
d0.
04—
——
0.61
——
0.28
—
R
easo
ning
[207
–209
]
Raw
biv
aria
te0.
13—
——
—0.
16—
0.23
0.10
Age
par
tiale
d—
——
——
——
——
S
troop
Inte
rfer
ence
[210
]
Raw
biv
aria
te—
——
—0.
64—
——
—
Age
par
tiale
d—
——
——
——
——
Proc
essi
ng S
peed
You
nger
and
Old
er
C
ompo
site
mea
sure
s [63
–64,
121
]
Raw
biv
aria
te—
0.64
0.60
0.54
0.66
0.42
0.35
——
Age
par
tiale
d—
0.13
0.34
—0.
19—
——
—
D
igit
Sym
bol [
62]
Raw
biv
aria
te0.
300.
19−0.01
0.14
0.15
0.17
0.24
—0.
25
Age
par
tiale
d—
——
——
——
——
S
impl
e/ch
oice
reac
tion
time
[57,
202
, 211
]
Raw
biv
aria
te0.
350.
51—
0.31
0.16
—0.
510.
350.
05
Age
par
tiale
d0.
17—
——
0.05
——
0.19
0.05
F
inge
r tap
ping
/mov
emen
ts [4
9, 6
2, 2
02]
Raw
biv
aria
te0.
22—
——
0.42
——
0.41
0.18
Age
par
tiale
d0.
190.
100.
030.
350.
110.
120.
090.
170.
20
V
isua
l sea
rch
[112
]
Raw
biv
aria
te—
0.68
0.41
——
0.40
0.38
——
Age
par
tiale
d—
——
——
——
——
Old
er o
nly
C
ompo
site
mea
sure
s [20
3]
Raw
biv
aria
te0.
32—
——
0.42
——
0.34
—
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Madden et al. Page 34
Cog
nitiv
e D
omai
nB
rain
Reg
ion
Fron
tal
CC
gIn
f Fro
ntIC
Mid
Sup
CC
sPa
rO
cc
Age
par
tiale
d0.
01—
——
0.02
——
−0.06
—
D
igit
sym
bol [
207]
Raw
biv
aria
te0.
14—
——
—0.
36—
—0.
10
Age
par
tiale
d—
——
——
——
——
S
impl
e/ch
oice
reac
tion
time
[57,
207
, 212
]
Raw
biv
aria
te0.
200.
63—
0.55
—0.
43—
—0.
01
Age
par
tiale
d—
——
——
——
——
F
inge
r tap
ping
/mov
emen
ts [2
13]
Raw
biv
aria
te—
——
——
—0.
71—
—
Age
par
tiale
d—
——
——
——
——
Mem
ory
You
nger
and
Old
er
F
ree
reca
ll [1
08]
Raw
biv
aria
te—
——
——
——
——
Age
par
tiale
d—
——
0.36
0.30
——
——
P
aire
d-as
soci
ate
lear
ning
[108
]
Raw
biv
aria
te—
——
——
——
——
Age
par
tiale
d—
——
0.43
——
——
—
Im
plic
it le
arni
ng [1
18]
Raw
biv
aria
te0.
46—
——
——
——
—
Age
par
tiale
d—
——
——
——
——
S
eman
tic c
ateg
oriz
atio
n [1
6]
Raw
biv
aria
te—
——
——
——
——
Age
par
tiale
d—
0.46
——
——
0.52
——
Old
er o
nly
F
ree
reca
ll [2
06, 2
08–2
09, 2
14]
Raw
biv
aria
te0.
14—
——
0.35
0.12
—0.
46−0.08
Age
par
tiale
d—
−0.01
0.26
—0.
090.
18—
0.58
—
R
ecog
nitio
n [5
2, 5
4]
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Madden et al. Page 35
Cog
nitiv
e D
omai
nB
rain
Reg
ion
Fron
tal
CC
gIn
f Fro
ntIC
Mid
Sup
CC
sPa
rO
cc
Raw
biv
aria
te—
—0.
27—
—0.
020.
29—
—
Age
par
tiale
d0.
490.
49—
——
——
——
P
aire
d-as
soci
ate
lear
ning
[54]
Raw
biv
aria
te—
—0.
43—
—0.
47—
——
Age
par
tiale
d—
——
——
——
——
Not
e. V
alue
s are
mea
n ef
fect
size
s (Pe
arso
n r)
for e
ffec
ts re
porte
d in
the
refe
renc
ed st
udie
s in
the
cogn
itive
dom
ains
of E
xecu
tive
Func
tion,
Pro
cess
ing
Spee
d, a
nd M
emor
y. R
esul
ts a
re se
para
ted
acco
rdin
gto
whe
ther
the
FA-c
ogni
tion
rela
tions
hips
wer
e ex
amin
ed a
cros
s bot
h yo
unge
r and
old
er a
dults
com
bine
d or
onl
y am
ong
olde
r adu
lts (d
ata
are
not p
rese
nted
from
the
few
stud
ies t
hat a
lso
exam
ined
the
FA-
cogn
ition
rela
tions
hips
for y
oung
er a
dults
sepa
rate
ly).
The
mea
n ef
fect
size
s obt
aine
d w
ith a
ge-r
elat
ed v
aria
nce
incl
uded
in th
e m
odel
(i.e
., ra
w b
ivar
iate
val
ues)
are
pre
sent
ed se
para
tely
from
thos
e th
atw
ere
parti
aled
for t
he e
ffec
t of a
ge. M
oder
ate
effe
ct si
zes a
nd la
rger
(> 0
.30)
are
list
ed in
bol
d fo
nt.
Mea
n ef
fect
size
s wer
e co
mpu
ted
follo
win
g th
e m
etho
d of
Ros
enth
al a
nd D
imat
teo
[110
]. Pe
arso
n r v
alue
s wer
e ob
tain
ed fr
om F
and
Spe
arm
an p
stat
istic
s (in
the
latte
r cas
e vi
a co
nver
sion
to st
anda
rdno
rmal
dev
iate
Z) w
hen
nece
ssar
y, a
nd r
valu
es w
ere
aver
aged
afte
r tra
nsfo
rmin
g th
em to
wei
ghte
d Fi
sher
z sc
ores
. In
som
e ca
ses,
r val
ues w
ere
adju
sted
such
that
pos
itive
rela
tions
hips
indi
cate
whe
rebe
tter p
erfo
rman
ce is
ass
ocia
ted
with
hig
her F
A.
Fron
t = fr
onta
l whi
te m
atte
r (in
clud
ing
forc
eps m
inor
, ant
erio
r per
ical
losa
l whi
te m
atte
r, an
d fr
onto
-sub
corti
cal t
ract
s); C
Cg
= ge
nu o
f the
cor
pus c
allo
sum
(inc
ludi
ng to
tal c
orpu
s cal
losu
m);
Inf F
ront
=in
ferio
r fro
ntal
whi
te m
atte
r (in
clud
ing
infe
rior l
ongi
tudi
nal f
asci
culu
s, in
ferio
r fro
nto-
occi
pita
l fas
cicu
lus,
and
unci
nate
fasc
icul
us);
IC =
inte
rnal
cap
sule
(inc
ludi
ng a
nter
ior,
genu
, and
pos
terio
rsu
breg
ions
); M
id =
mid
dle
whi
te m
atte
r (in
clud
ing
the
body
of t
he c
orpu
s cal
losu
m, f
orni
x, e
xter
nal c
apsu
le, c
ortic
ospi
nal t
ract
, and
tem
pora
l whi
te m
atte
r); S
up =
supe
rior w
hite
mat
ter (
incl
udin
gci
ngul
um b
undl
e, su
perio
r lon
gitu
dina
l fas
cicu
lus,
and
cent
rum
sem
iova
le);
CC
s = sp
leni
um o
f the
cor
pus c
allo
sum
(inc
ludi
ng te
mpo
ral s
ubre
gion
s of t
he c
orpu
s cal
losu
m);
Par =
par
ieta
l whi
te m
atte
r(in
clud
ing
post
erio
r per
ical
losa
l whi
te m
atte
r); a
nd O
cc =
occ
ipita
l whi
te m
atte
r (in
clud
ing
forc
eps m
ajor
).
Biochim Biophys Acta. Author manuscript; available in PMC 2013 March 1.




















